structural and functional analyses of activating amino acid substitutions in the receiver domain of...
TRANSCRIPT

J. Mol. Biol. (1997) 273, 299±316
Structural and Functional Analyses of ActivatingAmino Acid Substitutions in the Receiver Domain ofNtrC: Evidence for an Activating Surface
Michael Nohaile1, Dorothee Kern1, David Wemmer1*, Kenneth Stedman2
and Sydney Kustu2
1Department of ChemistryUniversity of CaliforniaBerkeley & Structural BiologyDivision, Lawrence BerkeleyNational Laboratory, BerkeleyCA 94720, USA2Department of Molecular andCell Biology, University ofCalifornia, Berkeley, CA 94720USA
Present address: K. Stedman, MaBiochemistry, D-82152 Martinsried
Abbreviations used: HMQC, hetquantum coherence; HSQC, heteroquantum coherence; MBP, maltoseNMR, nuclear magnetic resonanceOverhauser and exchange spectrosregulatory protein C; NtrCN-term, NNtrC; PCR, polymerase chain reaccorrelated spectroscopy.
0022±2836/97/410299±18 $25.00/0/mb
The bacterial enhancer-binding protein NtrC activates transcription whenphosphorylated on aspartate 54 in its amino (N)-terminal regulatorydomain or when altered by constitutively activating amino acid substi-tutions. The N-terminal domain of NtrC, which acts positively on theremainder of the protein, is homologous to a large family of signal trans-duction domains called receiver domains. Phosphorylation of an aspar-tate residue in a receiver domain modulates the function of adownstream target, but the accompanying structural changes are notclear. In the present work we examine structural and functional differ-ences between the wild-type receiver domain of NtrC and mutant formscarrying constitutively activating substitutions. Combinations of suchsubstitutions resulted in both increased structural changes in the N-term-inal domain, monitored by NMR chemical shift differences, and increasedtranscriptional activation by the full-length protein. Structural changescaused by substitutions outside the active site (D86N and A89T) werenot only local but extended over a substantial portion of the N-terminaldomain including the region from a-helix 3 to b-strand 5 (``3445 face'')and propagating to the active site. Interestingly, the activating substi-tution of glutamate for aspartate at the site of phosphorylation (D54E)also triggered structural changes in the 3445 face. Thus, the active siteand the 3445 face appear to interact. Implications with respect to howphosphorylation may affect the structure of receiver domains and howstructural changes may be communicated to the remainder of NtrC arediscussed.
# 1997 Academic Press Limited
Keywords: two-component systems; constitutive mutant forms of NtrC;phosphorylation; signal transduction; NMR chemical shift analysis
*Corresponding authorIntroduction
Two-component systems are a large family ofsignal transduction proteins that have been foundin all domains of life (Woese et al., 1990): bacteria,archaea and eukarya, formerly eubacteria, archae-
x Planck Institute for, Germany.eronuclear multiplenuclear singlebinding protein;
; NOESY, nuclearcopy; NtrC, nitrogen-terminal domain of
tion; TOCSY, total
971296
bacteria and eukaryotes (Albright et al., 1989;Chang et al., 1993; Hoch & Silhavy, 1995; Maedaet al., 1994; Nixon et al., 1986; Ota & Varshavsky,1993; Rudolph & Oesterhelt, 1995; Stock et al.,1989). They are characterized by two highly con-served protein domains, which are usually carriedon separate multi-domain proteins. In bacteria, thesensor or histidine kinase domain autophosphory-lates on a conserved histidine residue under appro-priate environmental or physiological conditions.The phosphate group is then transferred to anaspartate residue in the receiver domain (reviewedby Parkinson & Kofoid, 1992). This phosphotrans-fer event, which requires a divalent cation (Stocket al., 1993), activates the receiver domain, oftencalled a response regulator.
# 1997 Academic Press Limited

Figure 1. Ribbon diagram of the receiver domain ofNtrC. Side-chains of active site residues (D10, D11, D54,T82 and K104) and activating amino acid substitutions(D86N, A89T and V115I) are shown.
300 Receiver Domain of NtrC
The system that controls nitrogen-regulated tran-scription in enteric bacteria is a paradigm for two-component systems and consists of the sensorprotein NtrB and the response regulator NtrC(reviewed by Reitzer & Magasanik, 1987). In vivothe NtrC protein is activated under conditions ofnitrogen limitation (Keener & Kustu, 1988; Ninfa &Magasanik, 1986; Weiss & Magasanik, 1988) as aconsequence of phosphorylation of the aspartateresidue at position 54 (Sanders et al., 1992) in itsamino (N)-terminal receiver domain (Drummondet al., 1986; Keener & Kustu, 1988; Ninfa &Magasanik, 1986; Weiss & Magasanik, 1988). NtrCis also a well-studied member of a class of bacterialenhancer-binding proteins that activate transcrip-tion by s54-holoenzyme (RNA polymerase contain-ing the alternative sigma factor, s54: reviewed byKustu et al. (1989); Thony & Hennecke (1989)).These enhancer-binding proteins catalyze for-mation of transcriptionally productive open com-plexes by the polymerase in a reaction thatdepends upon hydrolysis of ATP (Weiss et al.,1991). Phosphorylation of D54 results in oligomeri-zation of NtrC (Porter et al., 1993; V. Weiss et al.,1992; Wyman et al., 1997) and activates its essentialATPase activity. The receiver domain of NtrC actspositively; that is, deleting it does not substitutefor phosphorylation. However, other receiverdomains, even those of other activators of s54-holoenzyme, act negatively (Gu et al., 1994; Hualaet al., 1992; Ireton et al., 1993; Kahn & Ditta, 1991;Simms et al., 1985).
Receiver domains can autophosphorylate (Fenget al., 1992; Lukat et al., 1992; McCleary & Stock,1994; Reyrat et al., 1993; Schroder et al., 1994), i.e.they can accept phosphate from low molecularmass donors as well as phosphoprotein donors,and most have an autophosphatase activity thatrapidly hydrolyses the aspartyl phosphate (Hesset al., 1988; Keener & Kustu, 1988; Weiss &Magasanik, 1988). The half-life for the aspartylphosphate residue in folded receiver domains var-ies from a few seconds to hours and is typically ofthe order of a few minutes (reviewed by Stocket al., 1995). Thus, the active, phosphorylated formof the receiver domain is usually transient, due tothe autophosphatase activity, and must be main-tained in situ.
Homogeneously active response regulator pro-teins have been obtained genetically. Many ofthese mutant forms, which are called constitutive,are active in vivo in the absence of their cognatesensor kinases and carry amino acid substitutionsor deletions in their receiver domains (Bourret et al.,1990; Bowrin et al., 1992; Flashner et al., 1995;Hirschman et al., 1985; Ireton et al., 1993; Jin et al.,1993; Kahn & Ditta, 1991; Pazour et al., 1992; Shojiet al., 1988; Speigelman et al., 1990; Stewart, 1993;Stewart et al., 1990). However, with the exceptionof proteins carrying amino acid substitutions in theactive site (Bowrin et al., 1992; Da Re et al., 1994;Gu et al., 1994; Han & Winans, 1994; Klose et al.,1993; Moore et al., 1993; Stewart, 1993; Welch et al.,
1994; Zhu et al., 1996), relatively few constitutiveproteins have been studied biochemically.
Constitutive forms of NtrC were isolated byselecting for mutations that suppressed the growthdefects caused by loss of function of NtrB(Flashner et al., 1995; Hirschman et al., 1985) (J. Lee& G. Oldroyd, personal communication). Whenpuri®ed and analyzed in vitro, the NtrC constitu-tive proteins activated transcription without beingphosphorylated (Flashner et al., 1995). Three ofthem were found to carry amino acid substitutionsin the N-terminal domain of NtrC; aspartate 86 toasparagine (D86N), alanine 89 to threonine (A89T)and valine 115 to isoleucine (V115I). Site-directedsubstitution of glutamate for the aspartate that isphosphorylated (D54E) also yielded a constitutiveform of NtrC (Klose et al., 1993).
The three-dimensional structures of four receiverdomains have been determined. These are CheY(Bellsolell et al., 1994; Moy et al., 1994; Stock et al.,1989, 1993; Volz & Matsumura, 1991) and Spo0F(Feher et al., 1995; Madhusudan et al., 1996), whichare isolated receiver domains, and the receiverdomains of NtrC (Figure 1: Volkman et al., 1995)and NarL (Baikalov et al., 1996). In the case ofNarL, the structure of the intact protein, receiverand output domains, was determined. All of thesestructures have an a5/b5 fold, with the active sitefor phosphorylation-dephosphorylation formed byloops at the carboxy termini of the strands thatcompose the parallel b-sheet. This overall fold andthe active site residues of receiver domains appearto be highly conserved (Volz, 1993). In the N-term-
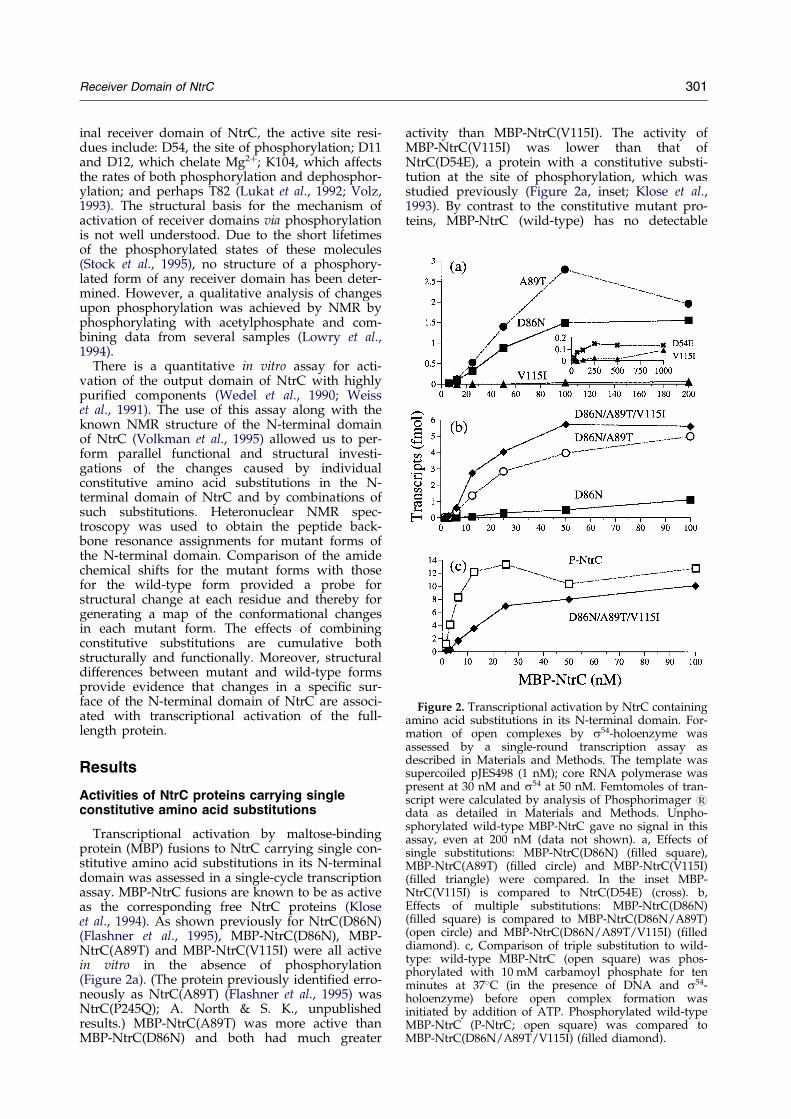
Figure 2. Transcriptional activation by NtrC containingamino acid substitutions in its N-terminal domain. For-mation of open complexes by s54-holoenzyme wasassessed by a single-round transcription assay asdescribed in Materials and Methods. The template wassupercoiled pJES498 (1 nM); core RNA polymerase waspresent at 30 nM and s54 at 50 nM. Femtomoles of tran-script were calculated by analysis of Phosphorimager 1data as detailed in Materials and Methods. Unpho-sphorylated wild-type MBP-NtrC gave no signal in thisassay, even at 200 nM (data not shown). a, Effects ofsingle substitutions: MBP-NtrC(D86N) (®lled square),MBP-NtrC(A89T) (®lled circle) and MBP-NtrC(V115I)(®lled triangle) were compared. In the inset MBP-NtrC(V115I) is compared to NtrC(D54E) (cross). b,Effects of multiple substitutions: MBP-NtrC(D86N)(®lled square) is compared to MBP-NtrC(D86N/A89T)(open circle) and MBP-NtrC(D86N/A89T/V115I) (®lleddiamond). c, Comparison of triple substitution to wild-type: wild-type MBP-NtrC (open square) was phos-phorylated with 10 mM carbamoyl phosphate for tenminutes at 37�C (in the presence of DNA and s54-holoenzyme) before open complex formation wasinitiated by addition of ATP. Phosphorylated wild-typeMBP-NtrC (P-NtrC; open square) was compared toMBP-NtrC(D86N/A89T/V115I) (®lled diamond).
Receiver Domain of NtrC 301
inal receiver domain of NtrC, the active site resi-dues include: D54, the site of phosphorylation; D11and D12, which chelate Mg2�; K104, which affectsthe rates of both phosphorylation and dephosphor-ylation; and perhaps T82 (Lukat et al., 1992; Volz,1993). The structural basis for the mechanism ofactivation of receiver domains via phosphorylationis not well understood. Due to the short lifetimesof the phosphorylated states of these molecules(Stock et al., 1995), no structure of a phosphory-lated form of any receiver domain has been deter-mined. However, a qualitative analysis of changesupon phosphorylation was achieved by NMR byphosphorylating with acetylphosphate and com-bining data from several samples (Lowry et al.,1994).
There is a quantitative in vitro assay for acti-vation of the output domain of NtrC with highlypuri®ed components (Wedel et al., 1990; Weisset al., 1991). The use of this assay along with theknown NMR structure of the N-terminal domainof NtrC (Volkman et al., 1995) allowed us to per-form parallel functional and structural investi-gations of the changes caused by individualconstitutive amino acid substitutions in the N-terminal domain of NtrC and by combinations ofsuch substitutions. Heteronuclear NMR spec-troscopy was used to obtain the peptide back-bone resonance assignments for mutant forms ofthe N-terminal domain. Comparison of the amidechemical shifts for the mutant forms with thosefor the wild-type form provided a probe forstructural change at each residue and thereby forgenerating a map of the conformational changesin each mutant form. The effects of combiningconstitutive substitutions are cumulative bothstructurally and functionally. Moreover, structuraldifferences between mutant and wild-type formsprovide evidence that changes in a speci®c sur-face of the N-terminal domain of NtrC are associ-ated with transcriptional activation of the full-length protein.
Results
Activities of NtrC proteins carrying singleconstitutive amino acid substitutions
Transcriptional activation by maltose-bindingprotein (MBP) fusions to NtrC carrying single con-stitutive amino acid substitutions in its N-terminaldomain was assessed in a single-cycle transcriptionassay. MBP-NtrC fusions are known to be as activeas the corresponding free NtrC proteins (Kloseet al., 1994). As shown previously for NtrC(D86N)(Flashner et al., 1995), MBP-NtrC(D86N), MBP-NtrC(A89T) and MBP-NtrC(V115I) were all activein vitro in the absence of phosphorylation(Figure 2a). (The protein previously identi®ed erro-neously as NtrC(A89T) (Flashner et al., 1995) wasNtrC(P245Q); A. North & S. K., unpublishedresults.) MBP-NtrC(A89T) was more active thanMBP-NtrC(D86N) and both had much greater
activity than MBP-NtrC(V115I). The activity ofMBP-NtrC(V115I) was lower than that ofNtrC(D54E), a protein with a constitutive substi-tution at the site of phosphorylation, which wasstudied previously (Figure 2a, inset; Klose et al.,1993). By contrast to the constitutive mutant pro-teins, MBP-NtrC (wild-type) has no detectable
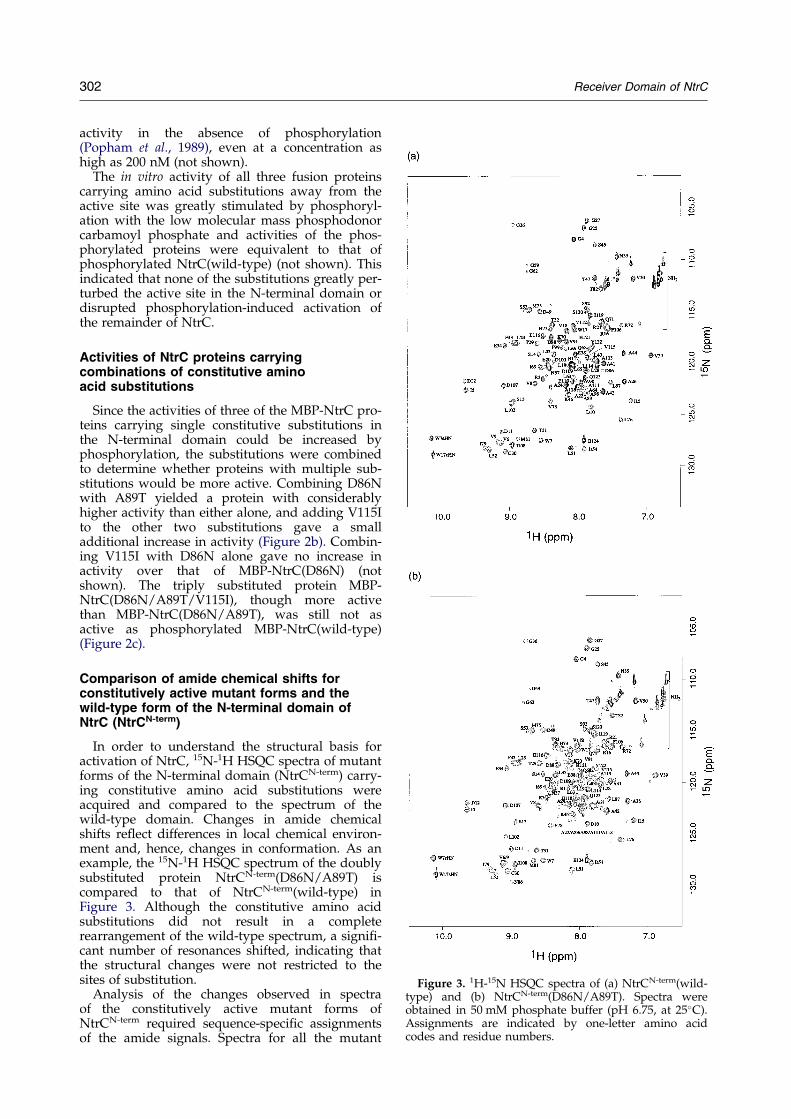
Figure 3. 1H-15N HSQC spectra of (a) NtrCN-term(wild-type) and (b) NtrCN-term(D86N/A89T). Spectra wereobtained in 50 mM phosphate buffer (pH 6.75, at 25�C).Assignments are indicated by one-letter amino acidcodes and residue numbers.
302 Receiver Domain of NtrC
activity in the absence of phosphorylation(Popham et al., 1989), even at a concentration ashigh as 200 nM (not shown).
The in vitro activity of all three fusion proteinscarrying amino acid substitutions away from theactive site was greatly stimulated by phosphoryl-ation with the low molecular mass phosphodonorcarbamoyl phosphate and activities of the phos-phorylated proteins were equivalent to that ofphosphorylated NtrC(wild-type) (not shown). Thisindicated that none of the substitutions greatly per-turbed the active site in the N-terminal domain ordisrupted phosphorylation-induced activation ofthe remainder of NtrC.
Activities of NtrC proteins carryingcombinations of constitutive aminoacid substitutions
Since the activities of three of the MBP-NtrC pro-teins carrying single constitutive substitutions inthe N-terminal domain could be increased byphosphorylation, the substitutions were combinedto determine whether proteins with multiple sub-stitutions would be more active. Combining D86Nwith A89T yielded a protein with considerablyhigher activity than either alone, and adding V115Ito the other two substitutions gave a smalladditional increase in activity (Figure 2b). Combin-ing V115I with D86N alone gave no increase inactivity over that of MBP-NtrC(D86N) (notshown). The triply substituted protein MBP-NtrC(D86N/A89T/V115I), though more activethan MBP-NtrC(D86N/A89T), was still not asactive as phosphorylated MBP-NtrC(wild-type)(Figure 2c).
Comparison of amide chemical shifts forconstitutively active mutant forms and thewild-type form of the N-terminal domain ofNtrC (NtrCN-term)
In order to understand the structural basis foractivation of NtrC, 15N-1H HSQC spectra of mutantforms of the N-terminal domain (NtrCN-term) carry-ing constitutive amino acid substitutions wereacquired and compared to the spectrum of thewild-type domain. Changes in amide chemicalshifts re¯ect differences in local chemical environ-ment and, hence, changes in conformation. As anexample, the 15N-1H HSQC spectrum of the doublysubstituted protein NtrCN-term(D86N/A89T) iscompared to that of NtrCN-term(wild-type) inFigure 3. Although the constitutive amino acidsubstitutions did not result in a completerearrangement of the wild-type spectrum, a signi®-cant number of resonances shifted, indicating thatthe structural changes were not restricted to thesites of substitution.
Analysis of the changes observed in spectraof the constitutively active mutant forms ofNtrCN-term required sequence-speci®c assignmentsof the amide signals. Spectra for all the mutant

Receiver Domain of NtrC 303
forms were ®rst tentatively assigned by assuminga minimum change from the spectrum ofNtrCN-term (wild-type). To con®rm the initialassignments, direct assignments were made forone of the mutant forms with large changes inspectrum, NtrCN-term(D86N/A89T). Most assign-ments were determined from 15N edited 3DNOESY-HMQC and 15N edited 3D TOCSY-HSQCexperiments. For weak and overlapped peaks,additional assignments were obtained from 3DCBCA(CO)NH and 3D HNCA experiments. Ingeneral, the assumption of minimal change fromthe wild-type spectrum was validated.
Since the spectrum of NtrCN-term(D86N) showedchanges similar to but smaller than those ofNtrCN-term(D86N/A89T), assignments for the latteralso supported assignments for NtrCN-term(D86N).Given the assignments for the doubly substitutedform, an 15N edited 3D NOESY-HMQC experimentwas suf®cient to verify the assignments for thetriply substituted NtrCN-term(D86N/A89T/V115I).The spectra for NtrCN-term(D54E) and NtrCN-term
(V115I) showed the smallest changes from thewild-type spectrum and were therefore assignedsolely on the basis of the assumption of minimumchange.
Based on the NOE patterns observed in the 15Nedited 3D NOESY-HMQC spectra for the doublyand triply substituted forms NtrCN-term(D86N/A89T) and NtrCN-term(D86N/A89T/V115I), thesecondary structural elements observed forNtrCN-term(wild-type) were unchanged, includingall of the NMR connectivities. Thus, the changes inchemical shift in spectra of mutant forms appear tore¯ect some reorganization of secondary structureelements. The effects on chemical shift may propo-gate from the site of reorganization, and hencechemical shift data are not suf®cient to reallyde®ne the structural change.
Regions involved in conformational change inmutant forms of NtrCN-term carryingconstitutive substitutions
The chemical shift differences between spectra ofthe mutant and wild-type forms of NtrCN-term werequanti®ed as described in Materials and Methods,and are displayed for each mutant form in Figure 4.For some residues, quanti®cation of the chemicalshift difference was not possible because the amideresonance was not observed or it overlappedseverely with other resonances in either mutant orwild-type spectra. For these residues an arbitraryvalue of ÿ10 Hz is used in the bar graphs.
As expected, residues showing signi®cant chemi-cal shift changes in NtrCN-term(D86N) were foundin helix 4 near the site of the amino acid substi-tution (Figure 4a). However, the changes were notcon®ned to this site, but extended over a largerregion of the molecule encompassing a-helix 3, b-strand 4, a-helix 4, b-strand 5, and the C-terminalend of a-helix 5. These secondary structural
elements are clustered together and form a face ofthe N-terminal domain, the ``3445 face'' (Figure 5a).
Interestingly, the D86N substitution, which liesoutside the active site, apparently also causedrearrangement in the active site. This is shown bychemical shift changes for residues D11, D12 andD54, all of which lie outside the 3445 face, andT82. Unfortunately, no peak was identi®ed for theamide group of the conserved active site residueK104 in spectra of either NtrCN-term(wild-type)(Volkman et al., 1995) or mutant forms carryingconstitutive amino acid substitutions. However,the signal for the amide group of the nearby resi-due L102 changed signi®cantly (Figure 4a).
The residues showing the largest chemical shiftchanges in the doubly substituted NtrCN-term
(D86N/A89T) (Figure 4b), lie in essentially thesame regions of the molecule as those in the singlysubstituted NtrCN-term(D86N). However, most ofthe changes in the doubly substituted form werelarger, compatible with the increased activity ofintact MBP-NtrC(D86N/A89T) over that of MBP-NtrC(D86N).
Chemical shift changes in the triply substitutedNtrCN-term(D86N/A89T/V115I) were similar to butlarger than those in the singly and doubly substi-tuted forms discussed above (Figure 4c), parallel-ing the increased activity of the triply substitutedNtrC protein. However, there are additional resi-dues that showed chemical shift changes in the tri-ply substituted domain: namely, residues in a-helix1, b-strand 1 and a-helix 5. The same residuesshowed chemical shift changes in NtrCN-term(V115I)(Figure 4d), which also showed changes in theactive site and in the 3445 face of the molecule.However, compatible with the low activity ofNtrC(V115I), the changes in the latter two regionsaffected fewer residues and were smaller than thechanges seen in the singly and doubly substitutedN termini discussed above.
Chemical shift changes were investigated notonly in mutant forms of NtrCN-term carrying consti-tutive substitutions removed from the active sitebut also in NtrCN-term(D54E), which carries an acti-vating substitution at the site of phosphorylation.In addition to the expected chemical shift changesfor residues in the active site, signi®cant changeswere observed for residues in the 3445 face of themolecule (Figure 4e). The magnitude of the chemi-cal shift changes for residues in the 3445 faceof NtrCN-term(D54E) was greater than that ofNtrCN-term(V115I) and less than that of NtrCN-term
(D86N) and NtrCN-term(A89T), compatible with therelative transcriptional activities of the intact pro-teins carrying these single amino acid substitutions(Figure 2a).
Mapping structural changes in NtrCN-term
carrying constitutive amino acid substitutions
An overview of the spatial distribution of thechemical shift differences plotted in Figure 4 isgiven in Figure 5. Differences in chemical shift of
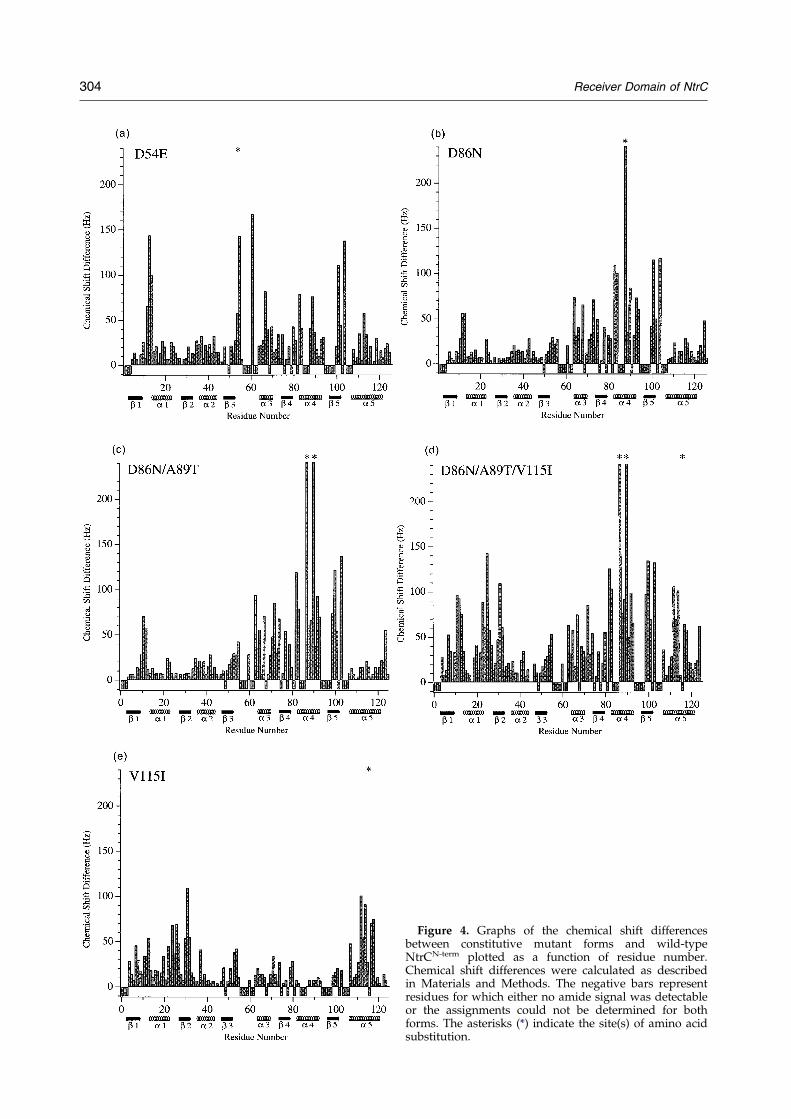
Figure 4. Graphs of the chemical shift differencesbetween constitutive mutant forms and wild-typeNtrCN-term plotted as a function of residue number.Chemical shift differences were calculated as describedin Materials and Methods. The negative bars representresidues for which either no amide signal was detectableor the assignments could not be determined for bothforms. The asterisks (*) indicate the site(s) of amino acidsubstitution.
304 Receiver Domain of NtrC
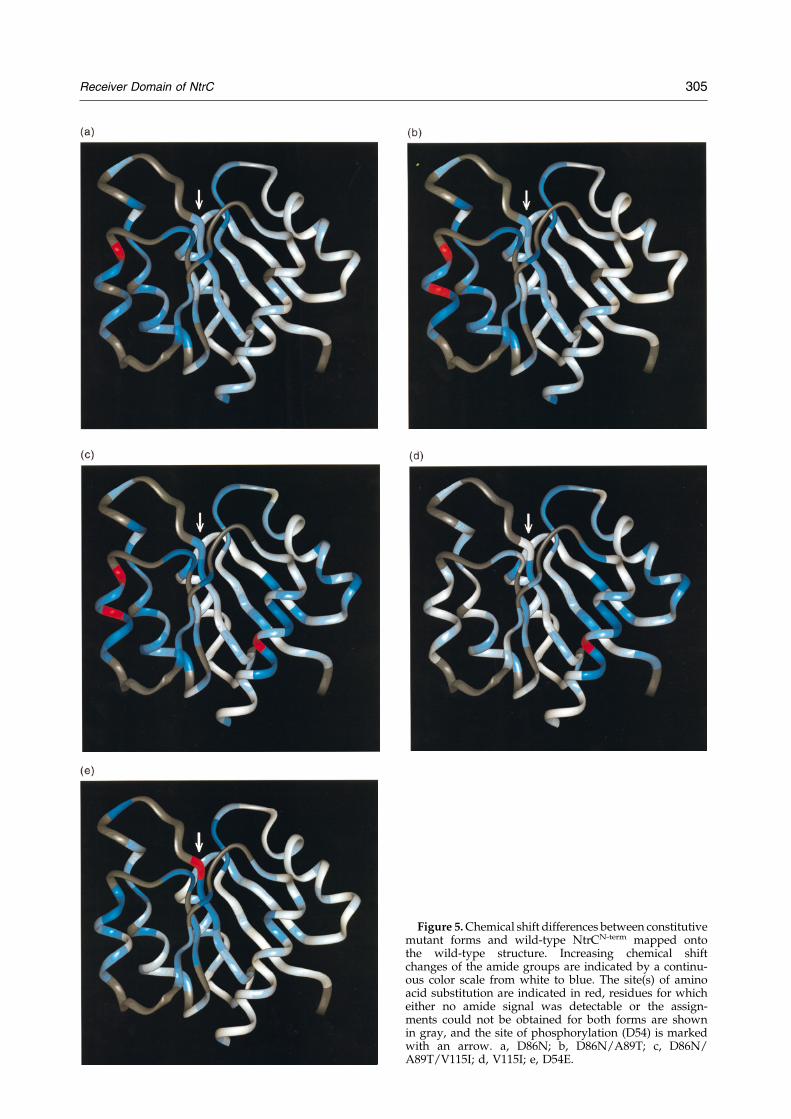
Figure 5. Chemical shift differences between constitutivemutant forms and wild-type NtrCN-term mapped ontothe wild-type structure. Increasing chemical shiftchanges of the amide groups are indicated by a continu-ous color scale from white to blue. The site(s) of aminoacid substitution are indicated in red, residues for whicheither no amide signal was detectable or the assign-ments could not be obtained for both forms are shownin gray, and the site of phosphorylation (D54) is markedwith an arrow. a, D86N; b, D86N/A89T; c, D86N/A89T/V115I; d, V115I; e, D54E.
Receiver Domain of NtrC 305

306 Receiver Domain of NtrC
the backbone amide for each residue in mutantforms of NtrCN-term are mapped on a ribbon dia-gram of NtrCN-term(wild-type) (Volkman et al.,1995). The amount of chemical shift difference isindicated by a continuous color scale from whitefor no change to blue for the largest changes. Redis used to mark the sites of amino acid substi-tutions and gray to designate residues for whichthe chemical shift difference could not be deter-mined.
Effects of combining substitutions in helices 4and 5 of NtrCN-term on the chemical shifts ofresidues in the active site
To evaluate global structural changes in mutantforms of NtrCN-term carrying single and multipleamino acid substitutions in the 3445 face, wecompared chemical shift changes of the activesite residues D11, D12 and D54, which shouldnot be subject to direct effects of the mutations.Surprisingly, as seen in overlays of a small partof HSQC spectra showing only the peak arisingfrom residue D11 (Figure 6), the chemical shiftsbetween NtrCN-term (wild-type) and the singly,doubly and triply mutant forms of NtrCN-term dis-cussed above lie on a straight line. Moreover, themagnitude of the shift increased with the numberof amino acid substitutions, which correlates withincreasing activities of the corresponding intactproteins. Similar behavior was observed forchemical shift changes of the other active siteresidues D12 and D54, and for residues G36, G62and M81 (data not shown).
Magnesium binding and structural changes inNtrCN-term carrying constitutive aminoacid substitutions
Mg2� is required for the in vitro transcriptionassay used to measure the activity of intact NtrC
Figure 6. An overlay of a region of 15N-1H HSQCspectra showing residue D11 for NtrCN-term(wild-type), NtrCN-term(D86N), NtrCN-term(D86N/A89T) andNtrCN-term(D86N/A89T/V115I).
and is absolutely required for phosphorylation ofD54 (Stock et al., 1993; A. North, personal com-munication). Since Mg2� is required functionallyand was not present in the NMR experiments, weneeded to determine whether the structuralchanges in NtrCN-term in the absence of Mg2�
detected via chemical shift were suf®cient for acti-vation of the protein. Therefore, we comparedHSQC spectra for NtrCN-term(D54E) and NtrCN-term
(wild-type) obtained in the presence and absenceof 20 mM Mg2�. Whereas signals for the backboneamide groups of active site residues D11, D12and D54 disappeared upon addition of Mg2� toNtrCN-term(wild-type), no differences wereobserved for NtrCN-term(D54E). Disappearance ofsignals for NtrCN-term(wild-type) was observedeven at Mg2� concentrations as low as 1.5 mM.These results indicate that the mutantNtrCN-term(D54E) fails to bind Mg2� at the activesite but is nonetheless active (Figure 2a) and there-fore that Mg2� is not required for structuralchanges in NtrCN-term that result in activation ofthe remainder of the protein.
Discussion
Constitutive substitutions in the receiverdomain of NtrC create an active state
Like forms of NtrC carrying the D54E substi-tution at the site of phosphoryation (Klose et al.,1993), mutant forms carrying the D86N (Flashneret al., 1995), A89T or V115I substitutions activatetranscription in vitro without being phosphory-lated, indicating that they are in an active state.This is an important observation because the selec-tion used to isolate such mutant forms, suppres-sion of growth defects caused by the absence ofthe physiological phosphodonor NtrB (see Flashneret al., 1995), could also give rise to receiverdomains with increased ability to use other phos-phodonor proteins (e.g. see Olmedo et al., 1990;Speigelman et al., 1990) or low molecular massphosphodonors such as acetyl or carbamoyl phos-phate (Feng et al., 1992). Although similar selec-tions have been employed to isolate mutant formsof many other proteins containing receiverdomains (Bourret et al., 1990; Bowrin et al., 1992;Flashner et al., 1995; Hirschman et al., 1985; Iretonet al., 1993; Jin et al., 1993; Kahn & Ditta, 1991;Pazour et al., 1992; Shoji et al., 1988; Speigelmanet al., 1990; Stewart, 1993; Stewart et al., 1990), rela-tively few have been studied in vitro to identifythose that mimic activation in the absence of phos-phorylation (Bowrin et al., 1992; Da Re et al., 1994;Gu et al., 1994; Han & Winans, 1994; Klose et al.,1993; Moore et al., 1993; Stewart, 1993; Welch et al.,1994; Zhu et al., 1996).
Effects of amino acid substitutions in the receiverdomain of NtrC can be assessed quantitatively in asingle-cycle transcription assay (Popham et al.,1989; Weiss et al., 1991). This allowed us to assessboth the relative activities of proteins carrying

Receiver Domain of NtrC 307
single amino acid substitutions and those of pro-teins carrying multiple substitutions.
Constitutive amino acid substitutions in NtrCact positively
NtrCN-term is required for transcriptional acti-vation by the remainder of the protein(Drummond et al., 1990; Flashner et al., 1995; D. S.Weiss et al., 1992) and hence constitutively activat-ing amino acid substitutions in NtrCN-term mustrepresent a gain of function. Interestingly, the con-stitutive substitutions in NtrCN-term that areremoved from the active site lie in two distinct sec-ondary structural elements, a-helix 4 (D86N andA89T) and a-helix 5 (V115I). These relatively con-servative substitutions affect residues that are notidentical either among activators of s54-holoen-zyme that contain receiver domains (e.g. DctD,AlgB; see Pao & Saier, 1995) or even among NtrCproteins. In fact, amino acid residues 89 and 115 ofNtrC proteins from several alpha-proteobacteriaare naturally threonine or isoleucine, respectively.
By contrast to the case for NtrC, some otherreceiver domains (e.g. that of DctD, which is alsoan activator of s54-holoenzyme, and those of FixJ,SpoOA and CheB) apparently work as intramole-cular repressors, i.e. the removal of such domainsactivates the remainder of the protein (Gu et al.,1994; Huala et al., 1992; Ireton et al., 1993; Kahn &Ditta, 1991; Simms et al., 1985). In these lattercases, phosphorylation presumably displaces thereceiver domain from the rest of the protein to acti-vate it (see Baikalov et al., 1996); hence, amino acidsubstitutions in the receiver domain that disruptedits repressive interaction with the rest of the pro-tein or simply disrupted its structure would beamong those that allowed these proteins to beactive in vitro in the absence of phosphorylation.Such substitutions would be unlikely to activateNtrC.
Combining constitutive amino acidsubstitutions in the N-terminal domain of NtrCincreases activity
Combinations of constitutive amino acid substi-tutions in helices 4 and 5 of the receiver domain ofNtrC gave increased activity (Figure 2b), as didphosphorylation of proteins carrying the individ-ual substitutions (not shown). The relative activi-ties of the NtrC proteins studied were:NtrC(V115I) < D54E < D86N < D86N/A89T < D86-N/A89T/V115I < phosphorylated NtrC(wild-type).It will be of interest to see whether other positivelyacting receiver domains can also be increasinglyactivated by multiple amino acid substitutions.Multistep models for activation of CheY suggestthat in this case, too, combinations of substitutionsmight yield a more active protein (Bourret et al.,1993; Welch et al., 1994).
NMR chemical shift differences as a probe ofconformational change
Chemical shift differences have long been usedas a probe of changes in structure associated withligand or macromolecular binding. In the case ofCheY, changes in chemical shift have been used formapping the regions involved in binding CheA(Swanson et al., 1995) and for de®ning areas under-going rearrangement due to phosphorylation oramino acid substitutions (Bourret et al., 1993;Drake et al., 1993; Lowry et al., 1994). In general, achange in chemical shift arises from a difference inthe magnetic environment which, in turn, is causedby a conformational rearrangement. Althoughthere is no simple relationship between the magni-tude of the chemical shift difference and degree ofconformational change, chemical shift differencesare nonetheless useful for de®ning the regions thatare involved in a conformational change. In thisstudy we have used the differences between amidechemical shifts of mutant and wild-type forms ofNtrCN-term as a probe of conformational changescaused by activating amino acid substitutions.
The magnitude of the amide chemical shiftdifference was obtained for about 80% of the resi-dues in mutant forms of NtrCN-term. Since theseresidues are well distributed throughout the mol-ecule, a good overall picture of the spatial locationof conformational change was obtained. Theremaining 20% of the residues were those forwhich no amide signal could be detected due to ahigh exchange rate or, in a few cases, due to severeoverlap with other signals. As seen for NtrCN-term
(wild-type), most of the residues for which signalswere undetectable are clustered in the loopsbetween b3 and a3, a4 and b5, and b5 and a5(Volkman et al., 1995).
Structural changes in the 3445 face ofNtrCN-term are correlated with activation of theintact protein
The results in Figure 4 clearly show that consti-tutive substitutions in NtrCN-term cause more thana local perturbation in its structure, i.e. one inwhich the contact interface between NtrCN-term andthe neighboring domain is locally remodeled.Rather, such constitutive substitutions cause globalchemical shift changes in the entire 3445 face of theN-terminal domain and in the active site. Althoughthis is a substantial portion of the molecule,changes are not scattered randomly throughout thedomain. Moreover, secondary structural elementsare conserved and, thus, most of the observedchemical shift differences appear to arise fromchanges in tertiary structure due to differences inthe relative orientations of these elements.
A comparison of the magnitudes of chemical shiftchanges in all of the mutant forms of NtrCN-term
that we examined indicated the followingorder: NtrCN-term(V115I) < D54E < D86N < D86N/A89T < D86N/A89T/V115I. This revealed a strong

308 Receiver Domain of NtrC
correlation between the magnitude of chemical shiftchanges in the 3445 face and the active site of NtrCN-
term and the increase in transcriptional activity of theintact protein carrying the corresponding substi-tutions.
Notably, NtrCN-term(V115I) shows only smallchemical shift changes in the 3445 face and theactive site, commensurate with the low in vitroactivity of intact NtrC(V115I). Similarly, combiningV115I with both the D86N and A89T substitutionsin helix 4 resulted in a relatively modest increasein chemical shift changes in the 3445 face andactive site, and a correspondingly modest increasein activity of the intact protein. NtrCN-term proteinscarrying the V115I substitution also showed chemi-cal shift changes in additional residues (Figure 5cand d). These changes appear to be due to localstructural perturbations caused directly by theV115I substitution.
The parallel examination of constitutive aminoacid substitutions removed from the active site anda substitution at the site of phosphorylation offeredthe possibility of investigating the link between the3445 face and the active site of NtrCN-term. Interest-ingly, there is communication between these tworegions: the D54E substitution causes structuralchanges in the 3445 face of the molecule, whereasthe substitutions in helices 4 and 5 cause changesin the active site. Moreover, changes in the chemi-cal shifts of the active site residues D11, D12 andD54 and of residues G36, G62 and M81 werecumulative in N termini carrying one, two or threesubstitutions in helices 4 and 5, paralleling theincreasing activity of the intact NtrC proteins car-rying these substitutions. Surprisingly, the shift ofthe signals follows a straight line, which suggeststhat the activating amino acid substitutions shiftthe equilibrium between an inactive and an acti-vated state (two-state model). The chemical shiftchanges can be explained by a conformationalchange between inactive and active forms ofNtrCN-term that is fast on the NMR timescale. Sucha model is in contrast to one in which the activat-ing amino acid substitutions result in multiple, par-tially active conformations.
Two observations support the hypothesis thatactivating amino acid substitutions cause a shift inthe equilibrium between two states of NtrCN-term.First, larger chemical shift changes in mutant formsof NtrCN-term carrying multiple amino acid substi-tutions correlate with greater activity of the corre-sponding intact NtrC proteins. Second, the HSQCspectrum of phosphorylated NtrCN-term(D86N/A89T) is nearly indistinguishable from that ofphosphorylated wild-type NtrCN-term (bothobtained with an approach similar to that used byLowry et al., 1994; unpublished results), whichwould indicate that phosphorylation drives theequilibrium completely towards the active state,whereas the activating amino acid substitutionsresult in a mixture of active and inactive forms.
Constitutively activating amino acidsubstitutions allow definition of an activatingsurface of NtrCN-term
The approach of using constitutively activatingamino acid substitutions to investigate structuralchanges involved in activation of receiver domains,such as that of NtrC, is attractive for severalreasons. First, such substitutions yield stable, acti-vated forms of response regulator proteins, in con-trast to the phosphorylated wild-type form, whichis often labile; second, studies of mutant forms car-rying combinations of such substitutions providemultiple points for comparison between structuralchanges and the amount of activity; third, as elabo-rated below, such mutant forms allow discrimi-nation between chemical shift changes that areassociated with activation and those that arisefrom the introduction of Mg2� and phosphate. Itremains to be determined whether the active con-formational state achieved by mutations and phos-phorylation are the same.
Although chemical shift changes in mutantforms of NtrCN-term are, in general, congruent withchanges observed in the phosphorylated forms ofNtrCN-term(wild-type) (unpublished results) andCheY (Lowry et al., 1994), the residues whose shiftsare perturbed by phosphorylation of these receiverdomains are distributed over a larger area. Someof the large chemical shifts in the phosphorylateddomains can be attributed to the presence of Mg2�
and a phosphate group in the active site, both ofwhich are known to cause large electrostatic per-turbations. Use of constitutive amino acid substi-tutions allows discrimination between localchanges directly due to the phosphorylation eventand those essential for activation of the cognateoutput domain.
Comparison of NtrCN-term with CheY
An 19F NMR study of structural changes causedby constitutive substitutions in CheY (D13K andD13R, which correspond to changes in D12 ofNtrCN-term) detected large changes in chemicalshifts of phenylalanine probes near the active site(Bourret et al., 1993), a result analogous to ours forthe receiver domain of NtrC. Unfortunately, therewas only one phenylalanine probe in the 3445 faceof CheY. This probe, at the C-terminal end of helix5, did show a signi®cant change in chemicalenvironment, commensurate with our results forNtrCN-term.
Regions of CheY involved in mediating inter-actions with its downstream targets FliM and FliGhave been de®ned genetically by suppression anal-ysis (Roman et al., 1992; Sockett et al., 1992). Manyof the amino acid substitutions in CheY that sup-press defects in its target ¯agellar switch proteinsare found in either the 3445 face of the molecule orthe active site. Overall, the regions of NtrCN-term
that appear to undergo structural change in consti-

Receiver Domain of NtrC 309
tutively active mutant forms overlap with regionsof CheY de®ned by suppression analysis.
Mechanism of activation of NtrC and otherresponse regulator proteins
Mutant forms of NtrCN-term carrying constitutiveamino acid substitutions have been used to de®nethe 3445 face not only as a region of the moleculethat undergoes structural changes in the activatedstate but as a region that is likely to play animportant role in signal transduction to the remain-der of the protein. At least two lines of evidencesupport the latter hypothesis. First, there is a directcorrelation between increasing activity of full-length NtrC and larger structural changes in the3445 face of NtrCN-term in a series of mutant formscarrying combinations of constitutive substitutionsin NtrCN-term. Second, chemical shift changes in the3445 face of the N-terminal domain parallel thosethat are induced by alterations at the active site:substitution of glutamate for aspartate at the site ofphosphorylation causes changes in the 3445 face(Figure 5e), and phosphorylation of NtrCN-term
causes similar spectral changes. Conversely, consti-tutive amino acid substitutions in this face causechemical shift changes in the active site.
The loops between b-strand 4 and a-helix 4, andbetween b-strand 5 and a-helix 5 probably havemultiple roles: they contain residues that form partof the active site and they link together secondarystructural elements in the 3445 face. These loopsare poised for communication between the site ofphosphorylation and the 3445 face and amidebackbone signals from residues in these loops (I80,M81 and L102) undergo large chemical shiftchanges in mutant forms of NtrCN-term carryingconstitutive substitutions in either place. Further-more, substitution of isoleucine for the highlyconserved threonine residue in the loop betweenb-strand 4 and a-helix 4 in CheY (T87, which ishomologous to T82 in NtrCN-term), results in amutant form of the protein that is defective in sig-nal transduction despite the fact that it can bephosphorylated (Ganguli et al., 1995).
In response regulator proteins of two-componentsystems, activation of the receiver domain by phos-phorylation of an aspartic acid residue in a highlyconserved active site is a general feature. Based onthe conserved structure of receiver domains (Volz,1993), data on the structure of activated forms ofCheY and NtrC may be relevant to the structuralbasis for activation of receiver domains generally.However, different receiver domains in the super-family regulate output domains of very differentfunction, as well as regulating output domains ofsimilar function differently (e.g. positive versusnegative regulation of s54 activators by the receiverdomains of NtrC and DctD, respectively:Drummond et al., 1990; Gu et al., 1994; Huala et al.,1992; V. Weiss et al., 1992). It is possible that phos-phorylation-induced structural changes in differentparts of the domain are important for triggering
signal transduction to diverse downstream targetsor for controlling similar targets differently. More-over, some phosphorylation-induced structuralchanges are likely to be involved in modulatinginteractions with sensor kinases (Swanson et al.,1995). Our data on mutant forms of NtrCN-term car-rying constitutive substitutions provide infor-mation about the conformational changes in thereceiver domain of this protein that are crucial foractivating the ATPase activity of the remainder ofthe protein. It will be interesting to determinewhether phosphorylation-induced structuralchanges in the 3445 face of receiver domains aregenerally important for triggering signal transduc-tion to diverse output domains and/or for control-ling s54 activation domains differently. B.T. Nixonand his colleagues have provided evidence for thelatter (Gu et al., 1994; B. T. Nixon, personal com-munication).
Materials and Methods
Identification and cloning of mutant forms of NtrC(Salmonella typhimurium) carrying constitutiveamino acid substitutions
The lesion causing the aspartate to asparagine substi-tution at position 86 of NtrC, D86N, was identi®edand cloned as described (Flashner et al., 1995). Thelesions causing the alanine to threonine substitution atposition 89, A89T, and the valine to isoleucine substi-tution at position 115, V115I, were identi®ed in thesame manner but were cloned by PCR-mediated ampli-®cation from the mutant chromosomes into thepNoTA/T7 vector (50 to 30 Inc.) to yield plasmidspJES965 and pJES964, respectively; sequence changeswere con®rmed in the latter plasmids (Jonghui Lee,personal communication). DNA encoding the A89Tsubstitution was recloned in this way because the pro-tein previously believed to carry this substitution(Flashner et al., 1995) was found to be NtrC(P245Q)(A. North & S. K., unpublished results).
Construction of plasmids for overexpression ofmaltose binding protein (MBP) fusions to mutantforms of NtrC
The original plasmid used as a template for oligonu-cleotide-mediated site-directed mutagenesis (Kunkel et al.,1987) was pJES608 (Klose, 1993), a pTZ18U (Biorad)derivative that contains a portion of the malE (maltosebinding protein) gene and all of the ntrC gene (Figure 7and Table 1). Plasmid pJES779 (K. Klose, personal com-munication), which was used for the construction of allmutant forms of NtrC carrying multiple amino acidsubstitutions in its N-terminal domain, was made bysite-directed mutagenesis of pJES608 using the oligonu-cleotide: 50-GCGCATTCCAATCTAGATGCC-30, whichintroduces the D86N substitution and an XbaI site intopJES608. For overexpression of the MBP-NtrC(D86N)protein, a 1.1 kb AatII-CspI fragment from pJES779 wasligated into similarly digested pJES559 (Klose et al.,1994) to create pJES862. Plasmid pJES946 was made bysite-directed mutagenesis of pJES779 using the oligonu-cleotide: 50-TCCAATCTAGATACGGCCGTGAGCGCC-30, which adds the A89T substitution and an EagI site topJES779. For overexpression of MBP-NtrC(D86N/A89T),

Figure 7. Plasmid constructions: strategy for construction of plasmids encoding mutant forms of NtrC containing con-stitutive amino acid substitutions in its N-terminal domain. In general, oligonucleotide-mediated, site-directed muta-genesis was performed as described (Kunkel et al., 1987) to introduce appropriate base-pair substitutions into thentrC gene of S. typhimurium in the plasmid labeled mutagenesis. This vector contained a small portion of the geneencoding the maltose binding protein malE fused to the ntrC gene. After mutagenesis, the portion of the DNA con-taining the substitutions was subcloned into vectors for expression of maltose binding protein fusions to NtrC, MBP-fusion, or for expression of the isolated N-terminal domain, N-terminus. Transcription of the N-terminal domain wasunder control of phage T7 RNA polymerase, as described (Studier et al., 1990). A few restriction endonuclease sitesare listed to indicate which endonucleases can be used for subcloning purposes and which to avoid. Generally, muta-genized ntrC was subcloned into the MBP-fusion vector using KpnI and CspI, and into the N terminus vector usingP¯MI and BssHII as indicated by the arrows. For details, see Materials and Methods, and for plasmid numbers andreferences see Table 1.
310 Receiver Domain of NtrC
a 1.27 kb KpnI-CspI fragment from pJES946 was ligatedinto similarly digested pJES623 to yield pJES951. (Plas-mid pJES623 is identical with pJES559, except that an800 bp BspEI-EcoRI fragment from pJES608 was used toreplace the HpaI site at nucleotide 1040 of ntrC with anEco47-III site for screening of transformants. The pre-sence or absence of the second HpaI site was used toscreen potential recombinant plasmids for the appropri-ate integration of donor DNA, which contained eitherone or two HpaI sites. (K. Klose, personal communi-cation).) Plasmid pJES947 was made by site directedmutagenesis of pJES779 using the oligonucleotides50-GATATTGATGAAGCCGTAGCGCTGATCGAGCGC-GCG-30 and 50-TCCAATCTAGATACGGCCGT-GAGCGCC-30, which add both the V115I and A89T sub-stitutions to D86N. For overexpression of MBP-NtrC(D86N/A89T/V115I) a 1.27 kb KpnI-CspI fragment
from pJES947 was ligated into similarly digestedpJES623 to yield pJES953.
Plasmids pJES971 and pJES970 for overexpression ofMBP-NtrC(V115I) and MBP-NtrC(A89T), respectively,were constructed as follows. Plasmid pJES966, an inter-mediate in construction of pJES971, was made by ligat-ing a 356 bp P¯MI-HpaI fragment from pJES964, whichencodes the V115I substitution, into similarly digestedpJES608. For overexpression of MBP-NtrC(V115I), thisplasmid was then digested with KpnI and CspI and a1.27 kb fragment was ligated into similarly digestedpJES559 to create pJES971. Plasmid pJES967, an inter-mediate in the construction of pJES970, was made byligating a 356 bp P¯MI-HpaI fragment from pJES965into similarly digested pJES608. For overexpression ofMBP-NtrC(A89T), this plasmid was then digested withKpnI and CspI and a 1.27 kb fragment was ligated into

Table 1. Plasmids
Plasmid Amino acid substitution Typea Source
pJES311 None Expression D. Weiss, 1991; Weiss et al. (1991)pJES454 D54E Expression Klose et al. (1993)pJES498 GlnA promoter Transcription template Porter et al. (1993)pJES557 None N terminus Klose et al. (1994)pJES559 None MBP-fusion Klose et al. (1994)pJES608 None Mutagenesis Klose (1993)pJES623 None MBP-fusion Klose (1993)pJES779 D86N Mutagenesis Klose (1993)pJES820 None N terminus This workpJES862 D86N MBP-fusion This workpJES941 D54E N terminus This workpJES946 D86N/A89T Mutagenesis This workpJES947 D86N/A89T/V115I Mutagenesis This workpJES951 D86N/A89T MBP-fusion This workpJES953 D86N/A89T/V115I MBP-fusion This workpJES956 D86N N terminus This workpJES958 D86N/A89T/V115I N terminus This workpJES959 D86N/A89T N terminus This workpJES964 V115I PCR clone J. Lee, personal communicationpJES965 A89T PCR clone J. Lee, personal communicationpJES966 V115I Cloning intermediate This workpJES967 A89T Cloning intermediate This workpJES970 A89T MBP-fusion This workpJES971 V115I MBP-fusion This workpJES972 V115I N terminus This workpJES973 A89T N terminus This work
a See Figure 7.
Receiver Domain of NtrC 311
similarly digested pJES559 to create pJES970. Allmutations created by site-directed mutagenesis orcloned by PCR were con®rmed by DNA sequencing ofthe mutagenized or ampli®ed region and all subcloningwas checked by analysis with restriction endonucleases.
Construction of plasmids for overexpression ofmutant forms of the N-terminal domain ofNtrC (NtrCN-term)
Plasmid pJES820, which was used to overexpressNtrCN-term, was made by digesting pJES557 (Klose et al.,1994), which carries ntrC with a stop codon at a positioncorresponding to amino acid residue 125, with HpaI andreligating to delete the bulk of ntrC (Figure 7 andTable 1). (Plasmid pJES557 was derived from pJES311,which was used to overexpress intact NtrC (Weiss et al.,1991).) For expression of mutant forms of NtrCN-term, a307 bp P¯MI-BssHII fragment from pJES820 wasreplaced by the corresponding fragment from pJES454(Klose et al., 1993), pJES779, pJES953, pJES951, pJES966or pJES967, respectively (see above), to yield the plas-mids containing the substitutions listed in parentheses:pJES941(D54E), pJES956(D86N), pJES958(D86N/A89T/V115I), pJES959(D86N/A89T), pJES972(V115I) and pJE-S973(A89T).
Expression and purification of full-length NtrC
MBP-NtrC fusion proteins were puri®ed essentially asdescribed (Klose et al., 1994). Brie¯y, DH5a cells harbor-ing the appropriate expression plasmid (Table 1) wereallowed to grow to an A600 of 0.4 in maximal inductionmedium (MIM; Mott et al., 1985), plus glutamine (2 mM®nal concentration) and ampicillin (200 mg/ml ®nal con-centration) and then induced with 0.4 mM isopropyl-b-D-thiogalactopyranoside (IPTG) and allowed to grow fortwo hours at 37�C with vigorous shaking. The cells were
harvested by centrifugation and resuspended in 10 ml ofbreakage buffer (50 mM Tris-acetate (pH 8.2), 200 mMpotassium chloride, 1 mM EDTA, 1 mM DTT) per liter ofcell culture (Keener, 1988). The cells were lysed by pas-sage through a French Pressure Cell (SLM-Aminco) at8000 psi. The resulting lysate was spun at 23,000 g for45 minutes and the supernatant was fractionated withammonium sulfate. The material that precipitatedbetween 40 and 70% saturation was dissolved in andthen dialyzed against amylose column buffer (20 mMsodium phosphate (pH 7.0), 500 mM sodium chloride,1 mM sodium azide, 1 mM DTT, 1 mM EDTA) andapplied to an amylose af®nity column (New EnglandBiolabs). The fusion protein was eluted with 10 mM mal-tose in amylose column buffer. The protein was dialyzedinto B-buffer (50 mM Tris-acetate (pH 8.3), 50 mM pot-assium chloride, 1 mM DTT, 5% (v/v) glycerol, 0.1 mMEDTA), loaded onto a strong anion-exchange column(Mono-Q HR5/5, Pharmacia), and eluted with a gradientof potassium chloride to 500 mM. The peak fractions (asmonitored at 280 nm) were pooled, dialyzed into B-buf-fer, quickly frozen in solid CO2 and stored at ÿ80�C.Protein concentrations were determined from absorptionat 280 nm in the presence of 6 M guanidine hydrochlo-ride (Pierce) using an extinction coef®cient of 111,190Mÿ1 cmÿ1 for MBP-NtrC (Edelhoch, 1967). Proteins wereassessed to be >95% pure (data not shown) by visualinspection of sodium dodecyl sulfate/polyacrylamidegels (Laemmli, 1970) stained with Coomassie brilliantblue R (Sigma).
Single-cycle transcription assay
In vitro transcription was performed basically asdescribed (Popham et al., 1989; Wedel et al., 1990);exceptions are noted in the legend to Figure 2. TheDNA template was supercoiled pJES498. This templatecarries a strong enhancer (two strong NtrC binding

312 Receiver Domain of NtrC
sites with the spacing found in the glnA enhancer of S.typhimurium) located approximately 400 bp upstream ofthe site of transcriptional initiation (Porter et al., 1993)and encodes a 152 nucleotide transcript lacking uracilresidues. The template was present at 1 nM, core RNApolymerase (from E. coli) was present at a ®nal concen-tration of 30 nM, and s54 puri®ed as described(Popham et al., 1991) was at 50 nM. Core RNA poly-merase and s54 were incubated together for at least®ve minutes on ice to allow their association beforethey were added to other components. NtrC concen-trations were varied and proteins were diluted inB-buffer (see above) containing 0.1 mg/ml bovine serumalbumin. The buffer used for transcription was 50 mMTris-acetate (pH 8.0), 100 mM potassium acetate, 8 mMmagnesium acetate, 27 mM ammonium acetate, 3.5%(w/v) polyethylene glycol 6000 and 1 mM DTT. Inexperiments with phosphorylated NtrC, carbamoylphosphate was added to 10 mM. Proteins were incu-bated with DNA for ten minutes at 37�C to allowenhancer-binding by NtrC, closed complex formation bypolymerase, and phosphorylation of NtrC where appli-cable. ATP was then added to 4 mM and the mixturewas incubated for ten minutes at 37�C to allow opencomplex formation. Open complex formation was moni-tored by adding GTP (400 mM), CTP (100 mM) and[a-32P]CTP (3000 Ci/ mmol) to allow synthesis of tran-scripts and heparin (to 0.1 mg/ml) to prevent furtheropen complex formation. After ten minutes at 37�C,transcripts were precipitated and electrophoresed on a6% denaturing polyacrylamide gel, which was thendried without ®xation and exposed to X-ray ®lm andPhosphorimager1 (Molecular Dynamics) screens. Forphosphorimager analysis, rectangles slightly larger thantranscript bands were drawn around each transcript andrectangles of equal size were drawn immediately belowthese. Signals (``volumes'') from the latter were sub-tracted from those for the former to correct for back-ground. Transcripts were quanti®ed by comparingcorrected signals for transcript bands to a diluted ali-quot of the same nucleotide mixture used for their syn-thesis.
Expression, purification and sample preparation ofNtrCN-term for NMR experiments
The expression vectors were transformed into E. coliBL21 (DE3) cells carrying the pLysS plasmid (Studieret al., 1990). To obtain uniform labeling of proteinsamples, cells were grown on M9 minimal medium(Sambrook et al., 1989) at 37�C with 15NH4Cl and(13C6)-D-glucose as the sole sources of nitrogen and car-bon, respectively. After the cell density had reached anA595 of 0.9, production of NtrCN-term was induced bythe addition of IPTG to 1 mM. The cells were grownfor six to eight hours after induction, whereupon theywere harvested by centrifugation. The cells from 1 to 2l of culture were lysed by sonication in lysis buffer(50 mM Tris-acetate (pH 8.2), 50 mM KCl, 10% glycer-ol) and a crude extract was prepared by centrifugationat 40,000 g for 20 minutes. The supernatant wasapplied to a Q-Sepharose fast-¯ow column(15 cm � 3 cm; Pharmacia) pre-equilibrated with 50 mMsodium phosphate buffer (pH 6.9). The column waswashed with the same buffer until no absorbance wasdetected at 280 nm. NtrCN-term was eluted with a400 ml linear NaCl gradient (0 to 400 mM) in the samebuffer. The fractions containing NtrCN-term were pooledand concentrated by ultra®ltration (Amicon, 3 kDa cut-
off). The concentrated protein solution was thenapplied to a Superdex 75 preparative grade column(90 cm � 6 cm, Pharmacia Biotech) to remove highmolecular mass contaminants. Fractions containingNtrCN-term were combined and concentrated by ultra®l-tration (Amicon, 3 kDa cutoff). The sample was thendesalted in 50 mM sodium phosphate buffer (pH 6.75),using a PD-10 gel-®ltration column (Pharmacia Bio-tech).
In order to match the conditions, all samples were¯ow-dialyzed at the same time against 50 mM sodiumphosphate buffer (pH 6.75). The ®nal concentration ofthe samples was 1.5 mM NtrCN-term containing 10%2H2O. The molar protein concentration was determinedby monitoring A280 using an extinction coef®cient of14,060 Mÿ1 cmÿ1 (Gill & von Hippel, 1989).
NMR experiments
NMR experiments were performed at 600 MHz on aBruker AMX-600 spectrometer at 25�C. Chemical shiftvalues were externally referenced to TSP (1H and 13C;Driscoll et al., 1990) and liquid ammonia (15N; Live et al.,1984). The States-TPPI method was used for quadraturedetection in the indirect dimensions of all multidimen-sional experiments (Marion et al., 1989a). All data wereprocessed with FELIX version 950 (Biosym). Shifted,skewed sine-bell functions were used for apodization ofthe free induction decays.
15N-1H 2D PEP-Z HSQC (Akke et al., 1994) exper-iments were collected with spectral widths of 6944 Hzand 2102 Hz in the 1H and 15N dimensions, respectively.The 1H carrier was placed on the H2O resonance at 4.78ppm, and the 15N carrier set to 119.1 ppm. A total of128 � 2048 complex points were collected in the t1 and t2
dimensions, respectively. Data were apodized in eachdimension with a shifted, skewed sine-bell. A shift of 85�was used in each dimension, with a skew of 1.0 and 0.75in the t1 and t2 dimensions, respectively. Data were zero-®lled to yield a 512 � 512 real matrix upon Fourier trans-formation.
15N-edited 3D NOESY-HMQC (Kay et al., 1989;Marion et al., 1989b) and 3D TOCSY-HSQC (Cavanaugh& Race, 1992) experiments were collected with spectralwidths of 6944 Hz for the 1H dimensions and 1861 Hzfor the 15N dimension. The 1H carrier was placed on theH2O resonance at 4.78 ppm, and the 15N carrier was setto 119.1 ppm. The NOESY mixing time was 100 ms, andthe TOCSY spin-lock period was 75 ms. A total of128 � 30 � 1024 complex points were collected in the t1,t2 and t3 dimensions, respectively. Data were apodizedin each dimension with a shifted, skewed sine-bell. Ashift of 75� was used in each dimension, with a skew of1.0, 0.8 and 0.5 in the t1, t2 and t3 dimensions, respect-ively. Data were zero-®lled to yield a 512 � 64 � 512 realmatrix upon Fourier transformation.
The 3D CBCA(CO)NH (Grzesiek & Bax, 1992a) andthe 3D HNCA (Grzesiek & Bax, 1992b) experimentswere collected with 1H and 15N parameters identicalwith the 3D 15N experiments described above. The 13Cdimension was centered at 43.16 ppm with a spectralwidth of 8446 Hz for the CBCA(CO)NH. The 13C dimen-sion was centered at 58.0 ppm with a spectral width of4528 Hz for the HNCA. In both experiments, 50 complexpoints were collected in the 13C dimension. A shifted,skewed sine-bell or sine-bell squared with a shift of 85�and skew of 0.8, 0.8 and 1.0 in the t1, t2, and t3 dimen-sions, respectively, was used for apodization. Time-

Receiver Domain of NtrC 313
domain data were zero-®lled to yield a 256 � 64 � 256real matrix.
Chemical shift difference analysis
The chemical shift differences between the amidesignals of mutant and wild-type forms of NtrCN-term
were calculated as follows:
�d �����������������������������������������1H�2 � ��15N�2
qwhere �1H and �15N are the chemical shift differencesin the proton and nitrogen dimensions, respectively.
Acknowledgments
We thank Jonghui Lee for isolation of the V115I sub-stitution and for construction of plasmids pJES964 andpJES965, Anne North for plasmid pJES498, and D.Hager, R. Burgess, D. Greiner, J. Owens and C. Mearesfor RNA polymerase core enzyme. This work (D.E.W.)was supported by the Of®ce of Energy Research, Of®ceof Health and Environmental Research, Health EffectsResearch Division of the U.S. Department of Energyunder contract no. DE-AC03-76SF00098, and throughinstrumentation grants from the U.S. Department ofEnergy, DE FG05-86ER75281 and the National ScienceFoundation, DMB 86-09035 and BBS 87-20134. This workwas also supported by NIH grant GM38361 to S.K.
References
Akke, M., Carr, P. A. & Palmer, A. G. (1994). Hetero-nuclear-correlation NMR spectroscopy with simul-taneous isotope ®ltration, quadrature detection, andsensitivity enhancement using Z rotations. J. Magn.Reson. ser. B, 104, 298±302.
Albright, L. M., Huala, E. & Ausubel, F. M. (1989). Pro-karyotic signal transduction mediated by sensorand regulator protein pairs. Annu. Rev. Genet. 23,311±336.
Baikalov, I., Schroder, I., Kaczor-Grzeskowiak, M.,Grzeskowiak, K., Gunsalus, R. P. & Dickerson, R. E.(1996). Structure of the Escherichia coli response reg-ulator NarL. Biochemistry, 35, 11053±11061.
Bellsolell, L., Prieto, J., Serrano, L. & Coll, M. (1994).Magnesium binding to the bacterial chemotaxis pro-tein CheY results in large conformational changesinvolving its functional surface. J. Mol. Biol. 238,489±495.
Bourret, R. B., Hess, J. F. & Simon, M. I. (1990). Con-served aspartate residues and phosphorylation insignal transduction by the chemotaxis proteinCheY. Proc. Natl Acad. Sci. USA, 87, 41±45.
Bourret, R. B., Drake, S. K., Chervitz, S. A., Simon,M. I. & Falke, J. J. (1993). Activation of the phos-phosignaling protein CheY. II. Analysis of activatedmutants by 19F NMR and protein engineering.J. Biol. Chem. 268, 13089±13096.
Bowrin, V., Brissette, R. & Inouye, M. (1992). Two tran-scriptionally active OmpR mutants that do notrequire phosphorylation by EnvZ in an Escherichiacoli cell-free system. J. Bacteriol. 174, 6685±6687.
Cavanaugh, J. & Race, M. (1992). Suppression of crossrelaxation effects in TOCSY spectra via a modi®ed
DIPSI-2 mixing sequence. J. Magn. Reson. 96, 670±678.
Chang, C., Kwok, S. F., Bleecker, A. B. & Meyerowitz,E. M. (1993). Arabidopsis ethylene-response geneETR1: similarity of product to two-componentregulators. Science, 262, 539±544.
Da Re, S., Bertagnoli, S., Fourment, J., Reyrat, J. M. &Kahn, D. (1994). Intramolecular signal transductionwithin the FixJ transcriptional activator: in vitro evi-dence for the inhibitory effect of the phosphorylata-ble regulatory domain. Nucl. Acids Res. 22, 1555±1561.
Drake, S. K., Bourret, R. B., Luck, L. A., Simon, M. I. &Falke, J. J. (1993). Activation of the phosphosignal-ing protein CheY. I. Analysis of the phosphorylatedconformation by 19F NMR and protein engineering.J. Biol. Chem. 268, 13081±13088.
Driscoll, P. C., Clore, G. M., Marion, D., Wing®eld,P. T. & Gronenborn, A. M. (1990). Complete reson-ance assignment for the polypeptide backbone ofinterleukin 1b using three-dimensional heteronuc-lear NMR spectroscopy. Biochemistry, 29, 3542±3556.
Drummond, M., Whitty, P. & Wootton, J. (1986).Sequence and domain relationships of ntrC andnifA from Klebsiella pneumoniae: homologies to otherregulatory proteins. EMBO J. 5, 441±447.
Drummond, M. H., Contreras, A. & Mitchenall, L. A.(1990). The function of isolated domains and chi-maeric proteins constructed from the transcriptionalactivators NifA and NtrC of Klebsiella pneumoniae.Mol. Microbiol. 4, 29±37.
Edelhoch, H. (1967). Spectroscopic determination oftryptophan and tyrosine in proteins. Biochemistry, 6,1948±1954.
Feher, V. A., Zapf, J. W., Hoch, J. A., Dahlquist, F. W.,Whiteley, J. M. & Cavanagh, J. (1995). 1H, 15N, and13C backbone chemical shift assignments, secondarystructure, and magnesium-binding characteristics ofthe Bacillus subtilis response regulator, SpoOF,determined by heteronuclear high-resolution NMR.Protein Sci. 4, 1801±1814.
Feng, J., Atkinson, M. R., McCleary, W., Stock, J. B.,Wanner, B. L. & Ninfa, A. J. (1992). Role of phos-phorylated metabolic intermediates in the regu-lation of glutamine synthetase synthesis inEscherichia coli. J. Bacteriol. 174, 6061±6070.
Flashner, Y., Weiss, D. S., Keener, J. & Kustu, S. (1995).Constitutive forms of the enhancer-binding proteinNtrC: evidence that essential oligomerization deter-minants lie in the central activation domain. J. Mol.Biol. 249, 700±713.
Ganguli, S., Wang, H., Matsumura, P. & Volz, K. (1995).Uncoupled phosphorylation and activation in bac-terial chemotaxis. The 2.1-AÊ structure of a threonineto isoleucine mutant at position 87 of CheY. J. Biol.Chem. 270, 17386±17393.
Gill, S. C. & von Hippel, P. H. (1989). Calculation ofprotein extinction coef®cients from amino acidsequence data. Anal. Biochem. 182, 319±326.
Grzesiek, S. & Bax, A. (1992a). An ef®cient experimentfor sequential backbone assignment of medium-sized isotopically enriched proteins. J. Magn. Reson.99, 201±207.
Grzesiek, S. & Bax, A. (1992b). Improved 3D triple-res-onance NMR techniques applied to a 31-kDaprotein. J. Magn. Reson. 96, 432±440.
Gu, B., Lee, J. H., Hoover, T. R., Scholl, D. & Nixon,B. T. (1994). Rhizobium meliloti DctD, a sigma 54-

314 Receiver Domain of NtrC
dependent transcriptional activator, may be nega-tively controlled by a subdomain in the C-terminalend of its two-component receiver module. Mol.Microbiol. 13, 51±66.
Han, D. C. & Winans, S. C. (1994). A mutation in thereceiver domain of the Agrobacterium tumefacienstranscriptional regulator VirG increases its af®nityfor operator DNA. Mol. Microbiol. 12, 23±30.
Hess, J. F., Oosawa, K., Kaplan, N. & Simon, M. I.(1988). Phosphorylation of three proteins in the sig-naling pathway of bacterial chemotaxis. Cell, 53,79±87.
Hirschman, J., Wong, P.-K., Sei, K., Keener, J. & Kustu,S. (1985). Products of nitrogen regulatory genesntrA and ntrC of enteric bacteria activate glnA tran-scription in vitro: evidence that the ntrA product isa sigma factor. Proc. Natl Acad. Sci. USA, 82, 7525±7529.
Hoch, J. A. & Silhavy, T. J. (1995). Two-component SignalTransduction, American Society for Microbiology(ASM), Washington, DC.
Huala, E., Stigter, J. & Ausubel, F. M. (1992). The centraldomain of Rhizobium leguminosarum DctD functionsindependently to activate transcription. J. Bacteriol.174, 1428±1431.
Ireton, K., Rudner, D. Z., Siranosian, K. J. & Grossman,A. D. (1993). Integration of multiple developmentalsignals in Bacillus subtilis through the Spo0A tran-scription factor. Genes Dev. 7, 283±294.
Jin, S., Song, Y., Pan, S. Q. & Nester, E. W. (1993).Characterization of a virG mutation that confersconstitutive virulence gene expression inAgrobacterium. Mol. Microbiol. 7, 555±562.
Kahn, D. & Ditta, G. (1991). Modular structure of FixJ:homology of the transcriptional activator domainwith the ÿ35 binding domain of sigma factors. Mol.Microbiol. 5, 987±997.
Kay, L. E., Marion, D. & Bax, A. (1989). Practical aspectsof 3D heteronuclear NMR of proteins. J. Magn.Reson. 84, 72±84.
Keener, J. W. (1988). Nitrogen regulation in enteric bac-teria: protein kinase and phosphoprotein phospha-tase activities of the NTRB and NTRC proteins.PhD thesis, University of California.
Keener, J. & Kustu, S. (1988). Protein kinase and phos-phoprotein phosphatase activities of nitrogen regu-latory proteins NTRB and NTRC of enteric bacteria:roles of the conserved amino-terminal domain ofNTRC. Proc. Natl Acad. Sci. USA, 85, 4976±4980.
Klose, K. E. (1993). Studies in the structure and functionof the nitrogen-regulatory protein NTRC from Sal-monella typhimurium. PhD thesis, University of Cali-fornia.
Klose, K. E., Weiss, D. S. & Kustu, S. (1993). Glutamateat the site of phosphorylation of nitrogen-regulatoryprotein NTRC mimics aspartyl-phosphate and acti-vates the protein. J. Mol. Biol. 232, 67±78.
Klose, K. E., North, A. K., Stedman, K. M. & Kustu, S.(1994). The major dimerization determinants of thenitrogen regulatory protein NTRC from enteric bac-teria lie in its carboxy-terminal domain. J. Mol. Biol.241, 233±245.
Kunkel, T. A., Roberts, J. D. & Zakour, R. A. (1987).Rapid and ef®cient site-speci®c mutagenesis with-out phenotypic selection. Methods Enzymol. 154,367±382.
Kustu, S., Santero, E., Keener, J., Popham, D. & Weiss,D. (1989). Expression of sigma 54 (ntrA)-dependent
genes is probably united by a common mechanism.Microbiol. Rev. 53, 367±376.
Laemmli, U. K. (1970). Cleavage of structural proteinsduring the assembly of the head of bacteriophageT4. Nature, 227, 680±685.
Live, D. H., Davis, D. G., Agosta, W. C. & Cowburn, D.(1984). Long-range hydrogen bond mediated effectsin peptides: 15N NMR study of gramicidin S inwater and organic solvents. J. Am. Chem. Soc. 106,1934±1941.
Lowry, D. F., Roth, A. F., Rupert, P. B., Dahlquist, F. W.,Moy, F. J., Domaille, P. J. & Matsumura, P. (1994).Signal transduction in chemotaxis. A propagatingconformation change upon phosphorylation ofCheY. J. Biol. Chem. 269, 26358±26362.
Lukat, G. S., McCleary, W. R., Stock, A. M. & Stock, J. B.(1992). Phosphorylation of bacterial response regu-lator proteins by low molecular weight phospho-donors. Proc. Natl Acad. Sci. USA, 89, 718±722.
Madhusudan, Zapf J., Whiteley, J. M., Hoch, J. A.,Xuong, N. H. & Varughese, K. I. (1996). Crystalstructure of a phosphatase-resistant mutant of spor-ulation response regulator SpoOF from Bacillussubtilis. Structure, 4, 679±690.
Maeda, T., Wurgler-Murphy, S. M. & Saito, H. (1994). Atwo-component system that regulates an osmosen-sing MAP kinase cascade in yeast. Nature, 369,242±245.
Marion, D., Ikura, M., Tschudin, R. & Bax, A. (1989a).Rapid recording of 2D NMR spectra without phasecycling - application to the study of hydrogenexchange in proteins. J. Magn. Reson. 85, 393±399.
Marion, D., Driscoll, P. C., Kay, L. E., Wing®eld, P. T.,Bax, A., Gronenborn, A. M. & Clore, G. M. (1989b).Overcoming the overlap problem in the assignmentof 1H NMR spectra of larger proteins by use ofthree-dimensional heteronuclear 1H-15N Hartmann-Hahn-multiple quantum coherence and nuclearOverhauser-multiple quantum coherence spec-troscopy: application to interleukin 1 beta. Biochem-istry, 28, 6150±6156.
McCleary, W. R. & Stock, J. B. (1994). Acetyl phosphateand the activation of two-component responseregulators. J. Biol. Chem. 269, 31567±31572.
Moore, J. B., Shiau, S. P. & Reitzer, L. J. (1993). Altera-tions of highly conserved residues in the regulatorydomain of nitrogen regulator I (NtrC) of Escherichiacoli. J. Bacteriol. 175, 2672±2701.
Mott, J. E., Grant, R. A., Ho, Y.-S. T. & Platt, T. (1985).Maximizing gene expression from plasmid vectorscontaining the lambda PL promoter: strategies foroverproducing transcription termination factor-RHO. Proc. Natl Acad. Sci. USA, 82, 88±92.
Moy, F. J., Lowry, D. F., Matsumura, P., Dahlquist,F. W., Krywko, J. E. & Domaille, P. J. (1994).Assignments, secondary structure, global fold, anddynamics of chemotaxis Y protein using three- andfour-dimensional heteronuclear (13C,15N) NMRspectroscopy. Biochemistry, 33, 10731±10742.
Ninfa, A. J. & Magasanik, B. (1986). Covalent modi®-cation of the glnG product NRI, by the glnL pro-duct, NRII, regulates the transcription of theglnALG operon in Escherichia coli. Proc. Natl Acad.Sci. USA, 83, 5909±5913.
Nixon, B. T., Ronson, C. W. & Ausubel, F. M. (1986).Two-component regulatory systems responsive toenvironmental stimuli share strongly conserveddomains with the nitrogen assimilation regulatory

Receiver Domain of NtrC 315
genes ntrB and ntrC. Proc. Natl Acad. Sci. USA, 83,7850±7854.
Olmedo, G., Ninfa, E. G., Stock, J. & Youngman, P.(1990). Novel mutations that alter the regulation ofsporulation in Bacillus subtilis. Evidence that phos-phorylation of regulatory protein SpoOA controlsthe initiation of sporulation. J. Mol. Biol. 215, 359±372.
Ota, I. M. & Varshavsky, A. (1993). A yeast proteinsimilar to bacterial two-component regulators.Science, 262, 566±569.
Pao, G. M. & Saier, M. H. J. (1995). Response regulatorsof bacterial signal transduction systems: selectivedomain shuf¯ing during evolution. J. Mol. Evol. 40,136±154.
Parkinson, J. S. & Kofoid, E. C. (1992). Communicationmodules in bacterial signaling proteins. Annu. Rev.Genet. 26, 71±112.
Pazour, G. J., Ta, C. N. & Das, A. (1992). Constitutivemutations of Agrobacterium tumefaciens transcrip-tional activator virG. J. Bacteriol. 174, 4169±4174.
Popham, D. L., Szeto, D., Keener, J. & Kustu, S. (1989).Function of a bacterial activator protein that bindsto transcriptional enhancers. Science, 243, 629±635.
Popham, D., Keener, J. & Kustu, S. (1991). Puri®cationof the alternative s factor, s54, from Salmonellatyphimurium and characterization of s54-holo-enzyme. J. Biol. Chem. 266, 19510±19518.
Porter, S. C., North, A. K., Wedel, A. B. & Kustu, S.(1993). Oligomerization of NTRC at the glnA enhan-cer is required for transcriptional activation. GenesDev. 7, 2258±2273.
Reitzer, L. J. & Magasanik, B. (1987). Ammonia assimila-tion and the biosynthesis of glutamine, glutamate,aspartate, asparagine, L-alanine, and D-alanine. InEscherichia coli and Salmonella typhimurium: Cellu-lar and Molecular Biology (Neidhardt, F. C., ed.), vol.1, pp. 302±320, American Society for Microbiology,Washington, DC.
Reyrat, J. M., David, M., Blonski, C., Boistard, P. &Batut, J. (1993). Oxygen-regulated in vitro transcrip-tion of Rhizobium meliloti nifA and ®xK genes.J. Bacteriol. 175, 6867±6872.
Roman, S. J., Meyers, M., Volz, K. & Matsumura, P.(1992). A chemotactic signaling surface on CheYde®ned by suppressors of ¯agellar switchmutations. J. Bacteriol. 174, 6247±6255.
Rudolph, J. & Oesterhelt, D. (1995). Chemotaxis andphototaxis require a CheA histidine kinase in thearchaeon Halobacterium salinarium. EMBO J. 14,667±673.
Sambrook, J., Fritsch, E. F. & Maniatis, T. (1989). Molecu-lar Cloning: A Laboratory Manual, 2nd edit., ColdSpring Harbor Laboratory Press, Cold SpringHarbor, NY.
Sanders, D. A., Gillece-Castro, B. L., Burlingame, A. L. &Koshland, D. E., Jr (1992). Phosphorylation site ofNtrC, a protein phosphatase whose covalent inter-mediate activates transcription. J. Bacteriol. 174,5117±5122.
Schroder, I., Wolin, C. D., Cavicchioli, R. & Gunsalus,R. P. (1994). Phosphorylation and dephosphoryla-tion of the NarQ, NarX, and NarL proteins of thenitrate-dependent two-component regulatory sys-tem of Escherichia coli. J. Bacteriol. 176, 4985±4992.
Shoji, K., Hiratsuka, S., Kawamura, F. & Kobayashi, Y.(1988). New suppressor mutation sur0B of spo0Band spo0F mutations in Bacillus subtilis. J. Gen.Microbiol. 134, 3249±3257.
Simms, S. A., Keane, M. G. & Stock, J. (1985). Multipleforms of the CheB methylesterase in bacterialchemosensing. J. Biol. Chem. 260, 10161±10168.
Sockett, H., Yamaguchi, S., Kihara, M., Irikura, V. M. &Macnab, R. M. (1992). Molecular analysis of the¯agellar switch protein FliM of Salmonellatyphimurium. J. Bacteriol. 174, 793±806.
Spiegelman, G., Van Hoy, B., Perego, M., Day, J., Trach,K. & Hoch, H. J. (1990). Structural alterations inthe Bacillus subtilis Spo0A regulatory protein whichsuppress mutations at several spo0 loci. J. Bacteriol.172, 5011±5019.
Stewart, R. C. (1993). Activating and inhibitorymutations in the regulatory domain of CheB, themethylesterase in bacterial chemotaxis. J. Biol. Chem.268, 1921±1930.
Stewart, R. C., Roth, A. F. & Dahlquist, F. W. (1990).Mutations that affect control of the methylesteraseactivity of CheB, a component of the chemotaxisadaptation system in Escherichia coli. J. Bacteriol. 172,3388±3399.
Stock, A. M., Mottonen, J. M., Stock, J. B. & Schutt, C. E.(1989). Three-dimensional structure of CheY, theresponse regulator of bacterial chemotaxis. Nature,337, 745±749.
Stock, A. M., Martinez-Hackert, E., Rasmussen, B. F.,West, A. H., Stock, J. B., Ringe, D. & Petsko, G. A.(1993). Structure of the Mg(2�)-bound form ofCheY and mechanism of phosphoryl transfer in bac-terial chemotaxis. Biochemistry, 32, 13375±13380.
Stock, J. B., Surette, M. G., Levit, M. & Park, P. (1995).Two-component signal transduction systems: struc-ture-function relationships and mechanisms ofcatalysis. In Two-Component Signal Transduction(Hoch, J. A. & Silhavy, T. J., eds), pp. 25±51,American Society for Microbiology, Washington,DC.
Studier, F. W., Rosenberg, A. H., Dunn, J. J. &Dubendorff, J. W. (1990). Use of T7 RNA polymer-ase to direct expression of cloned genes. MethodsEnzymol. 185, 60±89.
Swanson, R. V., Lowry, D. F., Matsumura, P., McEvoy,M. M., Simon, M. I. & Dahlquist, F. W. (1995). Loca-lized perturbations in CheY structure monitored byNMR identify a CheA binding interface. NatureStruct. Biol. 2, 906±910.
Thony, B. & Hennecke, H. (1989). The -24/-12 promotercomes of age. FEMS Microbiol. Rev. 63, 341±358.
Volkman, B. F., Nohaile, M. J., Amy, N. K., Kustu, S. &Wemmer, D. E. (1995). Three-dimensional solutionstructure of the N-terminal receiver domain ofNTRC. Biochemistry, 34, 1413±1424.
Volz, K. (1993). Structural conservation in the CheYsuperfamily. Biochemistry, 32, 11741±11753.
Volz, K. & Matsumura, P. (1991). Crystal structure ofEscherichia coli CheY re®ned at 1.7-AÊ resolution.J. Biol. Chem. 266, 15511±15519.
Wedel, A., Weiss, D. S., Popham, D., Droge, P. & Kustu,S. (1990). A bacterial enhancer functions to tether atranscriptional activator near a promoter. Science,248, 486±490.
Weiss, D. S., Batut, J., Klose, K. E., Keener, J. & Kustu,S. (1991). The phosphorylated form of the enhancer-binding protein NTRC has an ATPase activity thatis essential for activation of transcription. Cell, 67,155±167.
Weiss, D. S., Klose, K. E., Hoover, T. R., North, A. K.,Porter, S. C., Wedel, A. B. & Kustu, S. (1992). Pro-karyotic transcriptional enhancers. In Transcriptional

316 Receiver Domain of NtrC
Regulation (McKnight, S. L. & Yamamoto, K. R.,eds), pp. 667±694, Cold Spring Harbor LaboratoryPress, Cold Spring Harbor, NY.
Weiss, V. & Magasanik, B. (1988). Phosphorylation ofnitrogen regulator I (NRI) of Escherichia coli. Proc.Natl Acad. Sci. USA, 85, 8919±8923.
Weiss, V., Claverie-Martin, F. & Magasanik, B. (1992).Phosphorylation of nitrogen regulator I of Escheri-chia coli induces strong cooperative binding to DNAessential for activation of transcription. Proc. NatlAcad. Sci. USA, 89, 5088±5092.
Welch, M., Oosawa, K., Aizawa, S. I. & Eisenbach, M.(1994). Effects of phosphorylation, Mg2�, and con-formation of the chemotaxis protein CheY on its
binding to the ¯agellar switch protein FliM. Bio-chemistry, 33, 10470±10476.
Woese, K., Kandler, O. & Wheelis, M. L. (1990).Towards a natural system of organisms: proposalfor the domains Archaea, Bacteria, and Eukarya.Proc. Natl Acad. Sci. USA, 87, 4576±4579.
Wyman, C., Rombel, I., North, A. K., Bustamante, C. &Kustu, S. (1997). Unusual oligomerization requiredfor activity of NtrC, a bacterial enhancer-bindingprotein. Science, 275, 1658±1661.
Zhu, X., Amsler, C. D., Volz, K. & Matsumura, P.(1996). Tyrosine 106 of CheY plays an importantrole in chemotaxis signal transduction in Escherichiacoli. J. Bacteriol. 178, 4208±4215.
(Received 28 March 1997; received in revised form 17 July 1997; accepted 18 July 1997)
Edited by P. E. Wright