speciation rate and character evolution - the
TRANSCRIPT

PRELIMINARY PHYLOGENY OF DIPLOSTEPHIUM (ASTERACEAE):SPECIATION RATE AND CHARACTER EVOLUTION
Oscar M. Vargas1,2 and Santiago Madrinan2
1Section of Integrative Biology and the Plant Resources Center, The University of Texas, 205 W 24th St.,Stop CO930, Austin, Texas 78712 U.S.A., email: [email protected]
2Laboratorio de Botanica y Sistematica Universidad de los Andes, Apartado Aereo 4976, Bogota, D. C., Colombia
Abstract: Diplostephium comprises 111 neotropical species that live in high elevationhabitats from Costa Rica to Chile. Primarily Andean, the genus seems to haveundergone an adaptive radiation indicated by its high number of species, broadmorphological variation, and diversification primarily in an ecosystem (paramo) thatformed within the last 2–5 my. Internal transcriber spacer (ITS) sequences and severalchloroplast markers, rpoB, rpoC1, and psbA-trnH were sequenced in order to infer apreliminary phylogeny of the genus. The chloroplast regions showed no significantvariation within the genus. New ITS data were therefore analyzed together withpublished sequences for generating a topology. Results suggest that Diplostephium andother South American genera comprise a polytomy within which a previously describedNorth American clade is nested. Monophyly of Diplostephium was neither supportednor rejected, but the formation of a main crown clade using different methods ofanalysis suggests that at least a good portion of the genus is monophyletic. AShimodaira-Hasegawa test comparing the topology obtained and a constrained oneforcing Diplostephium to be monophyletic showed no significant difference betweenthem. Monophyly of some of the previously proposed series of the genus was notsupported by the phylogenetic tree. Morphological character mapping results suggestthat the high Andean forest tree species are derived from shrubby paramo-punaancestors, contradicting previous hypotheses about morphological evolution of thegenus and documenting an atypical trend of downslope diversification in paramoplants.
Keywords: Adaptive radiation, Andes Cordillera, Astereae, Diplostephium, high Andeanforest, ITS, morphological evolution, paramo, psbA-trnH, puna, rpoB, rpoC1.
Diplostephium Kunth is a genus of 111known species (Vargas, 2011), distributedfrom Costa Rica to northern Chile in highelevation cloud forests (2500–3000 m), para-mos (3000–4500 m), and puna habitats(3800–4200 m). The genus is characterizedby alternate leaves, white to purplish or bluishligules, and a pappus formed by two rows ofbristles, the outermost one reduced. Mor-phologically diverse, growth forms in thegenus vary from decumbent subshrubs tosmall trees up to 6 m tall, with leaves thatrange from 3 mm to 12 cm long. Arrange-ments of capitula vary from single heads topaniculiform or umbelliform sinflorescences.
Sydney F. Blake compiled the first revisionof the genus in 1922, recognizing 40 species.He divided the genus into five series (DEN-
TICULATA, FLORIBUNDA, LAVANDULIFOLIA5
DIPLOSTEPHIUM, ROSMARINIFOLIA, and RUPES-
TRIA) based on leaf and sinflorescence varia-tion. In 1928 he added 15 more species andeliminated the series DENTICULATA and FLOR-
IBUNDA. Later, Jose Cuatrecasas studied thegenus, describing several species and publish-ing two revisions (Cuatrecasas, 1943, 1969).In the second revision, in which 53 speciesknown from Colombia were treated, hesubdivided the genus even further, reinstatingseries DENTICULATA and FLORIBUNDA, andincorporating seven new series: ANACTINOTA,CORIACEA, CRASSIFOLIA, HUERTASINA, PHYLI-
COIDEA, SAXATILIA, and SCHULTZIANA. Cua-trecasas (1969: 92) proposed a ‘‘phylogenetic’’order to these 12 series (Table 1), intuitivelypolarizing arborescent forms, large leaves,multi-capitulate sinflorescences, and smallheads as ‘‘atavistic’’ (i.e., ancestral). These
NUMBER 15 VARGAS AND MADRINAN: SPECIATION AND CHARACTER EVOLUTION OF DIPLOSTEPHIUM 1
LUNDELLIA 15:1–15. 2012

character-states are found in series DENTICU-
LATA, CORIACEA, and CRASSIFOLIA, whichgrow mostly in montane forests. In contrast,character-states such as a shrubby habit, smallthick revolute leaves, single capitula, and largeheads were considered derived and occur inthe paramo and puna species placed in seriesRUPESTRIA, ANACTINOTA, and DIPLOSTEPH-
IUM. This hypothesis suggests that extantparamo-puna species were derived from highAndean forest-dwelling ancestors, comparableto other paramo plant radiations such as theEspeletiinae intensely studied by Cuatrecasas(1986). Cuatrecasas’ revision of the Colom-bian species (1969) is considered the lastcomprehensive study of the genus because ofthe number of species included and therearrangement and description of new series.Additionally, Cuatrecasas (1986) hypothesizedthat the main center of speciation and theorigin of the genus were in Colombia probablybased this on the fact that more than half ofthe species of the genus from all the seriesdescribed including the presumed ‘‘ancestralmorphotypes’’ are found in that country.Colombia now has 63 reported speciesaccounting for ca. 57% of the currently
recognized species (Vargas and Madrinan,2006).
Within the Asteraceae, Diplostephiumhas been placed in the tribe Astereae and thesubtribe Hinterhuberinae based on mor-phology (Nesom, 1994; Nesom and Robin-son, 2007), along with other South Ameri-can genera such as Floscaldasia Cuatrec.,Flosmutisia Cuatrec., Hinterhubera Sch. Bip.ex Wedd., Llerasia Triana, Laestadia Kunthex Less., and Oritrophium (Kunth) Cuatrec.Bonifacino and Sancho (2004) also hypoth-esized that Diplostephium is closely related tothe genus Guynesomia Bonifacino & Sancho.At the molecular level, there are threepublished sequences of the genus represent-ing two species (two for the internaltranscriber spacer nrDNA marker ITS, andone for the external transcriber spacer ETS).These have been used in studies focusing onthe generic relationships within tribe Aster-eae (Noyes and Rieseberg, 1999; Brouilletet al., 2008; Karaman-Castro and Urbatsch,2009). Noyes and Rieseberg’s study (1999)included one ITS sequence of Diplostephiumrupestre. Using ITS, Noyes and Rieseberg(1999) revealed a North American crown
TABLE 1. Cuatrecasas’ circumscription of Diplostephium species for Colombia (1963). Series areordered by morphological polarization of series that contain atavistic (i.e., ancestral) characteristics(top), to series that present derived characteristics (bottom).
Series Species
DENTICULATA D. ochraceum, D. bicolor, D. ellipticum, D. dentatum, D. oblongifolium, D.antioquense, D. tenuifolium, D. tamanum, D. mutiscuanum, D. fosbergii, D.grantii, D. ocanense, D. tachirense, D. leiocladum.
CRASSIFOLIA D. crassifolium.CORIACEA D. coriaceum.HUERTASINA D. huertasii, D. juliani.FLORIBUNDA D. floribundum, D. tolimense, D.farallonense, D. pittierii, D. cinerascens, D.
cayambense.SCHULTZIANA D. juabioyi, D. alveolatum, D schultzii, D. rhododendroides, D. romboidale.ROSMARINIFOLIA D. lacunosum, D. revolutum, D. violaceum, D. heterophyllum, D. cyparissias, D.
rosmarinifolium.PHYLICOIDEA D. phylicoides.RUPESTRIA D. rupestre, D. eriophorum, D. weddellii.SAXATILIA D. saxatile, D. romeroi, D. tergocanum.DIPLOSTEPHIUM D. micradenium, D. glutinosum, D. apiculatum, D. nevadense, D. glandulosum,
D. spinulosum, D. colombianum, D. parvifolium, D. hartwegii.ANACTINOTA D. anactinotum, D. inesianum.
2 LUNDELLIA DECEMBER, 2012

clade nested in a Southern Hemisphere grade,suggesting a single origin for the NorthAmerican Astereae. Diplostephium appearedin a South American grade close to the NorthAmerican clade with low support. Noyes andRieseberg suggested that the putative ances-tors of the North American Astereae wereSouth American taxa, and that the SouthernHemisphere basal grade contains the moreancient lineages for the tribe.
Brouillet et al.’s (2008) study compriseda comprehensive ITS phylogenetic analysisof the Astereae tribe using a considerablesample size of genera. The authors of thispaper concluded that the origin of the tribewas probably African, due to the biogeo-graphic pattern of the basal lineages in thetopology. Based on these results, theyhypothesized that South American speciesof the tribe are split between the ‘‘PaleoSouth American clade’’ and the ‘‘SouthAmerican lineages.’’ The Paleo South Amer-ican clade is nested between the basallineages and a New Zealand clade, whilethe South American lineages appear in amore derived clade forming a polytomy withsome Australasian lineages and a NorthAmerican clade. Brouillet et al. used twoDiplostephium sequences in their study, D.rupestre and D. ericoides, which were part ofa polytomy in the topology along with otherSouth America lineages. Their result sug-gested that Diplostephium is not monophy-letic, but phylogenetic support was notshown and was simply mentioned as beingnon-significant (Brouillet et al., 2008).
Karaman-Castro and Urbatsch (2009)used a subset of the taxa sampled byBrouillet et al. (2008), focusing their studyin the Hinterhubera group, seven genera ofthe Hinterhuberinae subtribe in whichDiplostephium is not included. The ITSregion and part of the ETS were used asmarkers. These authors concluded that thesemarkers do not support the monophyly ofthe Hinterhubera group, nor the monophylyof the Hinteruberinae, a result that agreedwith previous phylogenetic studies (Noyesand Rieseberg, 1999; Brouillet et al., 2008).
Here, however, the Diplostephium samples,the same used by Brouillet et al. (2008),formed a monophyletic clade in the ITS treewith moderate support. For the ETSphylogenetic analysis, Karaman-Castro andUrbatsch (2009) sequenced one species ofDiplostephium yielding no evidence aboutthe monophyly of the genus. Additionally,the authors concluded that the ETS topol-ogy was poorly resolved in comparison tothe ITS.
Diplostephium appears to be a rapidlyevolving genus. It is morphologically diverseand has more than 90 species living in theAndean paramos and punas, habitats thatwere not available until 2–4 mya (van derHammer and Cleef, 1986). Thus, in order toattempt to reconstruct a molecular phylo-gentic tree of the genus and test if the genusis monophyletic it is necessary to usemarkers with high levels of sequence varia-tion. The ITS, the region between the 18S-26S nuclear ribosomal DNA (Baldwin et al.,1995), has been proven to be a valuablemarker for phylogenetic reconstruction inthe Asteraceae, particularly within the Aster-eae tribe (Noyes and Rieseberg, 1999; Low-rey et al., 2001; Cross et al., 2002; Brouilletet al., 2008; Karaman-Castro and Urbatsch,2009). We also tested the chloroplastmarkers psbA-trnH, rpoB, and rpoC1 to seeif they might also provide phylogeneticallyinformative variation; those markers havebeen proposed as DNA barcodes becausethey have been shown to be variable at thespecies (Holligsworth et al., 2009).
Our aims in this research were: 1) testadditional molecular markers of potentialuse to reconstruct a phylogeny of Diplo-stephium; 2) generate a preliminary phylo-genetic tree of various species currentlyrecognized in Diplostephium; 3) analyze thesister group relationships of Diplostephiumwithin the Astereae by including a represen-tative sample on the South Americanlineages; 4) test the monophyly of the genus;and 5) assess Cuatrecasas’ evolutionaryhypotheses of the genus and the monophylyof his circumscriptions of the series.
NUMBER 15 VARGAS AND MADRINAN: SPECIATION AND CHARACTER EVOLUTION OF DIPLOSTEPHIUM 3

MATERIALS AND METHODS
TAXON SAMPLING. The chloroplast re-gions, psbA-trnH, rpoB, and rpoC1 weresequenced using nine, five, and four mor-phologically divergent species of Diploste-phium respectively (see Appendix 1 forspecies names, voucher specimens, andGenBank accession numbers). For the ITSdata set we newly sequenced 28 samples ofDiplostephium and one each of Laestadia,Llerasia, and Oritrophium. These genera weresequenced due to their placement in theHinterhuberinae (Nesom, 1994; Nesom andRobinson, 2007). Seventy-five additionalAstereae sequences from previous studies(Cross et al., 2002; Noyes and Rieseberg,1999; Karaman-Castro and Urbatsch, 2009)were downloaded from GenBank and includ-ed in the analysis. Three of these 70 sequenceswere from Diplostephium. The taxon sam-pling focused on Diplostephium and particu-larly the South American lineages describedin Brouillet et al. (2008). Taking into accountthis focus, just two samples representing theNorth American clade (Noyes and Rieseberg,1999, Brouillet et al., 2008) were included inthe analysis. The Genbank sequences gener-ated in this study are listed in Appendix 1,and the sequences downloaded are listed inAppendix 2. The complete ITS matrix (newsequences plus those downloaded fromGenBank) contained a total of 101 samplesof the tribe Astereae, including 31 samplesof Diplostephium, which represent 27 species,and 11 of the 12 series proposed byCuatrecasas. Outgroup taxa were selectedtaking into account the general phylogenetictree of the tribe published by Brouillet et al.(2008). Llerasia and Chiliotrichum Cass. wereused as outgroups due to their position in thepaleo South American clade (Brouillet et al.,2008), which is basal to the New Zealandclade, the Australasian lineages, the NorthAmerican clade, and the South Americanlineages.
DNA EXTRACTION, AMPLIFICATION, AND
SEQUENCING. Total DNA extractions of alltaxa were performed using the Dneasy Plant
Mini Kit (Qiagen) from silica gel driedleaves or herbarium specimens. The chloro-plast regions were amplified using thepolymerase chain reaction (PCR) in a finalvolume of 25mL, with total DNA andGoTaqH Green Master Mix (Promega)following manufacture’s protocols, withaddition of BSA at a final concentration of0.1mg/ml. The thermal profile used for thechloroplast regions was 94uC for 1 min then40 cycles of 94uC for 30 sec, 50uC for 40 sec,72uC for 40 sec and a final extension of 72uCfor 5 min. The primers used to amplify thechloroplast regions where those of Sang et al.(1997) for psbA-trnH and those described inthe barcoding protocols from Plant DNABarcoding working group led by KewGardens (http: //www.kew.org/barcoding/protocols.html) for rpoB (2 and 4) andrpoC1 (1 and 4).
Amplification for the ITS marker usingthe PCR technique was made in a finalvolume of 25 ml, with total DNA and GoTaqHGreen Master Mix (Promega) followingmanufacturer’s protocols, 0.3 ml of pureDimethyl Sulfoxide (DMSO), and bovineserum albumen (BSA) at a final concentra-tion of 0.1 mg/ml. The thermal profile usedfor ITS was 94uC for 1 min, then 30 cycles of94uC for 1 min, 54u for 1 min, and a finalextension 72uC for 1 min. Amplifications forDNA obtained from fresh material wereperformed using the primers ITS1 and ITS4in a single reaction (White et al., 1990).Herbarium specimen material was amplifiedusing two reactions with the internal primersITS2 and ITS3 (White et al., 1990).
PCR products were cleaned using theWizardH SV Gel and PCR Clean-Up System(Promega). Direct sequencing using Big DyeTerminator Ready Reaction Mix (AppliedBiosystems Inc.) was performed in an ABIPRISM 310 sequencer (Applied BiosystemsInc.) at the sequencing facility of theUniversidad de los Andes, Bogota, Colombia.Electropherograms were edited in Geneious3.5.4 (Drummond et al., 2007). Sequenceswere aligned in MacClade 4 (Maddison andMaddison, 2000) using pair-wise alignment
4 LUNDELLIA DECEMBER, 2012

and then manually edited. Aligned matricescan be accessed from TreeBase (http://purl.org/phylo/treebase/phylows/study/TB2:S11212).
PHYLOGENETIC ANALYSIS. MaximumParsimony (MP) analysis of the ITS datasetwas initiated using PAUPRat (Sikes andLewis, 2001) along with PAUP* 4.0b10(Swofford, 1998) on the CIPRES onlineportal (Miller et al., 2009). Two thousanditerations were made using random addi-tion, and tree-bisection-reconnection (TBR)branch swapping, saving one tree per itera-tion. The set of trees recovered from theparsimony ratchet was used as the startingpoint for a heuristic search performed inPAUP* 4.0b10 (Swofford, 1998) with thefollowing options: multistate taxa interpretedas polymorphism, all characters equallyweighted and unordered, gaps treated asmissing, 100 replicates with random sequenceaddition, TBR branch swapping, and branch-es collapsed if maximum branch length waszero. Bootstrap support (BS) for MP wasperformed in PAUP* 4.0b10 (Swofford,1998) using 1,000 replicates and the sameparameters used in the heuristic search.
For the maximum likelihood analysis(ML) the model chosen was calculated inModeltest 3.7 (Posada and Crandall, 1998).The ITS ML analysis was performed inRAxML 7.0.4 (Stamatakis, 2006) with theGeneral Time Reversible (GTR) model, andrates inv-gamma. ML support was assessedusing BS with 100 replicates in the sameprogram with the same parameters.
Bayesian Inference (BI) analysis wasperformed with Mr. Bayes 1.3.2 (Huelsen-beck and Ronquist, 2001) using 10,000,000generations, four chains, sample frequency5 1,000 generations, burn-in 5 2,500,000with GTR model, and rates inv-gamma.
A Shimodaira-Hasegawa-test (SH-test,Shimodaira and Hasegawa, 1999) was per-formed on PAUP* 4.0b10 (Swofford, 1998)to compare statistically the ITS ML treeagainst a constraint topology where Diplo-stephium was forced to be monophyletic.The constraint tree was obtained by forcingDiplostephium to be monophyletic under a
ML analysis with the General Time Revers-ible (GTR) model, and rates inv-gamma inRAxML 7.0.4 (Stamatakis, 2006). This con-straint tree was compared with the phyloge-netic tree obtained initially by ML. Treetopologies were considered to be significant-ly different if p , 0.05. All the aboveanalyses, with the exception of the parsimo-ny ratchet, were performed on the phy-locluster computer facility at The Universityof Texas at Austin.
CHARACTER MAPPING. Character stateand habitat reconstruction was performedin MacClade 4 using implicit examination(Maddison and Maddison, 2000), based in areduced ITS ML topology limited to theclade comprising Diplostephium species(clade A). Four characters were reconstruct-ed: habit, leaf size, sinflorescence type, andhabitat. In addition, Cuatrecasas’ series werecoded in order to evaluate their monophyly.For the habit character, the states consideredwere: subshrub (0–30 cm tall), shrub (0.3–2 m tall), and tree (.2 m tall); for leaf size:small (0–2 cm long), medium (2–5 cmlong), and large (.5 cm long); for sinflor-escence type: solitary capitula, panicle,raceme, and umbel; finally for the habitat:high Andean forest and paramo (puna wasnot mapped because species sampled fromclade A are not present in this habitat). Datawere taken from herbarium specimensdeposited in US and COL herbaria.
RESULTS
CHLOROPLAST MARKERS. Sequencing ofthe chloroplast markers selected was aban-doned early in the study due to their lowlevels of variation. The rpoC1 region showedtwo variable sites in the alignment for thefour species sequenced, while rpoB showedonly one mutation in one species from thefive sequenced. Finally, psbA-trnH showedfour variable sites in the matrix of ninespecies sampled (see Appendix 1 for speciessequenced).
ITS PHYLOGENETIC ANALYSIS. As inprevious studies (Noyes and Rieseberg,
NUMBER 15 VARGAS AND MADRINAN: SPECIATION AND CHARACTER EVOLUTION OF DIPLOSTEPHIUM 5

1999; Brouillet et al., 2008; Karaman-Castroand Urbatsch, 2009), the ITS marker showedsufficient variation to assess the partialbackbone phylogenetic tree of the tribe withrespect to the group of interest; in contrast,it failed in revealing enough resolution andsupport for elucidating the relationshipsbetween the South American lineages. Thealigned matrix of the complete datasetconsisted of 559 bp for 101 samples,including the outgroup. Of these, 269characters were constant, 84 were variableand parsimony-uninformative, and 206 wereparsimony-informative. The MP analysisfound 42,568 shortest trees of 1,098 steps(see Fig. 1 for strict consensus). The treeshad a consistency index (CI) of 0.4487, aretention index (RI) of 0.6501, and arescaled consistency index (RC) of 0.2919.The ML analysis found one tree of score25893.165595 (Fig. 2). The topology ob-tained by the BI analysis 50 percent majorityrule consensus is shown in Fig. 3.
Those clades where BS and PosteriorProbability (PP) have significant values areconsistent among the three topologies ob-tained by the different analysis methodsused. Contradictory positions of taxa be-tween the topologies were the position of theAustralasian lineages and the position ofsome species on the South American line-ages. Nonetheless, BS and PP do not supportthese contradictory positions across thetopologies obtained (Figs. 1–3). The MPstrict consensus tree (Fig. 1) differs fromthe ML and BI topologies (Figs. 2 and 3) inthe position of the Australasian lineages.While in the MP strict consensus theAustralasian lineages appear in a polytomywith South American lineages, in ML theyappear as the sister clade of a large polytomyformed by the South American lineages andthe North American clade; in BI they appearas a grade in which the large polytomyformed by the South American lineages andthe North American clade is nested. None ofthese three different positions is supportedby significant BS values in MP and ML, norby the PP inferred by BI.
All the species of Diplostephium arenested in the large polytomy formed by theSouth American lineages and the NorthAmerican clade in ML and BI, while in theMP tree they are nested in the largepolytomy formed by the South Americanlineages, the North American clade, and theAustralasian lineages. None of the topologiesshowed all the species of Diplostephiumsampled forming a monophyletic group. InMP D. azureum is the sister species of a cladeformed by Blakiella bartsiifolia and Aztec-aster G.L. Nesom species with a BS less than50%. The remaining 26 species of Diplo-stephium (30 samples) form a monophyleticgroup with a BS less than 50%. Thismonophyletic group includes a clade labeledas A (Fig. 1), recovered also in ML and BIanalyses, representing 22 species of Diplo-stephium (24 samples). Nested in clade A isthe largest clade of the genus recovered byMP with a BS more than 50%, this group ofDiplostephium contains 22 samples repre-senting 20 species (BS558).
In the ML topology, Diplostephiumazureum is sister to a clade of ParastrephiaNutt. species. Again, BS does not supportthe position of this species. The rest of theDiplostephium species form a monophyleticgroup similar to one described for MP butwith the inclusion of Sommerfeltia spinulosaas sister species of clade A. The position ofS. spinulosa on the ML topology is notsupported by ML-BS, nor found in theMP and BI topologies. This large cladeformed by 30 samples of Diplostephium andS. spinulosa has a BS support less than50%. The largest Diplostephium monophy-letic group with support is clade A withBS570%.
The BI tree also recovers clade A with aPP50.95 but does not recover a larger cladewith species of Diplostephium like MP andML. As in ML, D. azureum appears sister tothe clade formed by Parastrephia species inthe BI topology with a posterior PP50.59.Diplostephium glandulosum appears in thelarge polytomy with the other South Amer-ican lineages. Diplostephium haenkei also
6 LUNDELLIA DECEMBER, 2012
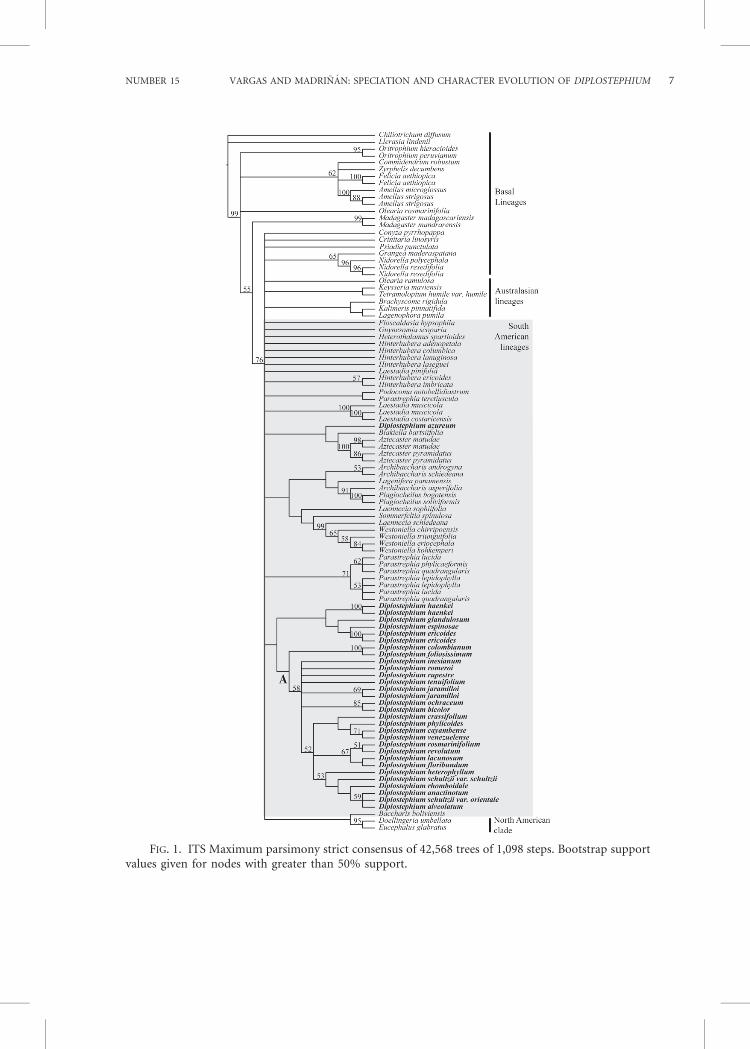
FIG. 1. ITS Maximum parsimony strict consensus of 42,568 trees of 1,098 steps. Bootstrap supportvalues given for nodes with greater than 50% support.
NUMBER 15 VARGAS AND MADRINAN: SPECIATION AND CHARACTER EVOLUTION OF DIPLOSTEPHIUM 7

FIG. 2. ITS Single phylogram obtained under maximum likelihood criterion. Bootstrap supportvalues given for nodes with greater than 50% support.
8 LUNDELLIA DECEMBER, 2012
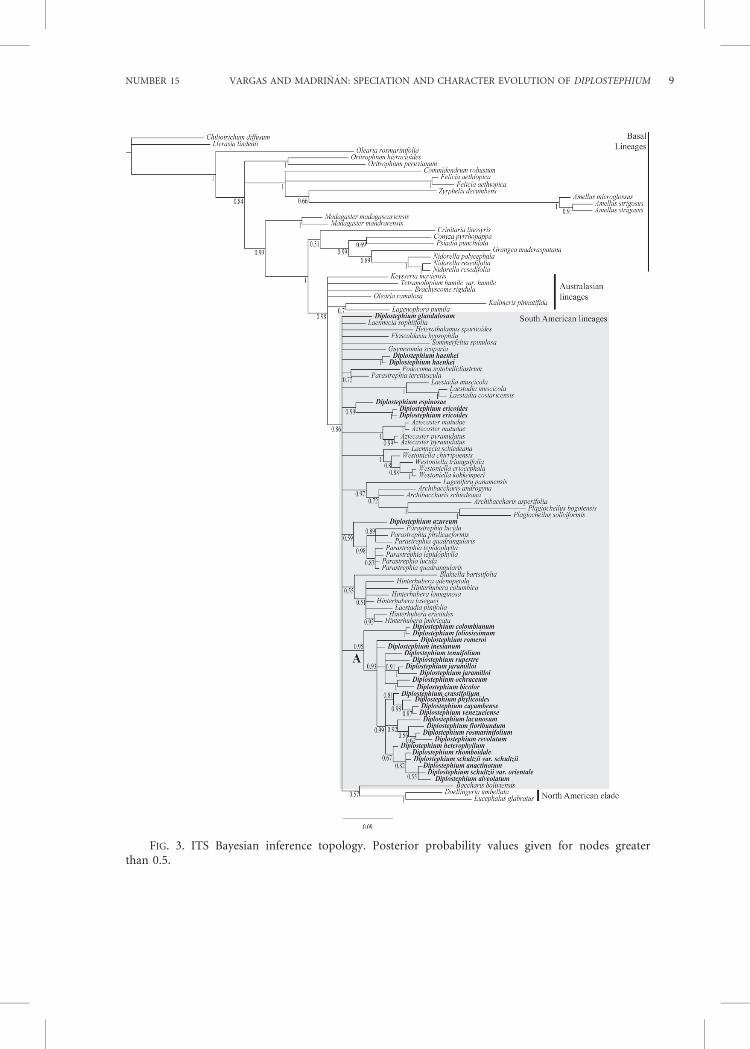
FIG. 3. ITS Bayesian inference topology. Posterior probability values given for nodes greaterthan 0.5.
NUMBER 15 VARGAS AND MADRINAN: SPECIATION AND CHARACTER EVOLUTION OF DIPLOSTEPHIUM 9

appears in the large polytomy with the otherSouth American lineages in which its twosamples form a monophyletic clade. Diplo-stephium espinosae and D. ericoides form aclade with PP50.99, a clade that also formspart of the grand polytomy in the topology.Additionally, none of our topologies sup-ports the monophyly of Laestadia muscicolaand Diplostephium shultzii as monophyleticspecies. Laestadia muscicola is paraphyletic,with L. costaricensis nested within it, aposition with strong support in all thetopologies obtained. Diplostephium schultziiis polyphyletic, with the two varieties beingin different branches inside clade A, bothpositions have moderate or low support.
CONSTRAINT ANALYSIS. Maximum like-lihood analysis with Diplostephium forced tobe monophyletic obtained a likelihood valueof 25895.207338. This topology comparedwith the initial tree generated by ML (score25893.165595) was not significantly differ-ent from that inferred by the SH-test(p50.439). This result fails to reject the nullhypothesis of the topologies not beingsignificantly different, leading to the premisethat the monophyly of Diplostephium cannotbe rejected by these data.
CHARACTER STATE RECONSTRUCTION TO
TEST CUATRECASAS’ HYPOTHESES AND SERIES.Because clade A was consistently recoveredin all the phylogenetic methods used andpresented moderate support in ML and BI, itwas used to test the hypotheses proposed byCuatrecasas about the morphological evolu-tion of Diplostephium and its subgenericsubdivision. The morphological reconstruc-tion of characters (Fig. 4a) showed that thehypothetical ancestor of early divergingspecies of Diplostephium in clade A wasprobably a shrub, and that the tree habitevolved more than once in the genus.Character mapping of leaf size (Fig. 4b), ahighly valuable character in the genusassociated with habit, suggests that smallleaves were ancestral and that medium sizedand large leaves evolved at different times inclade A. Likewise solitary capitula (Fig. 4c) canbe interpreted as ancestral in the clade A. The
reconstruction of habitat (Fig. 4d) showed ahigh elevation mountain-living morphotype(paramo or proto-paramo) hypothetical an-cestor, rather than a high-mountain forestmorphotype for clade A. Taking into accountthat paramo and puna habitats are correlatedwith shrubby and sub-shrubby growth formsand that Diplostephium species not grouped inclade A are not arborescent, the habitatcharacter mapping suggests that modernarborescent forest species were derived fromhigh elevation ancestors in the genus.
Our results suggest that the seriesANACTINOTA, DENTICULATA, FLORIBUNDA,ROSMARINIFOLIA and SCHULTZIANA arenot monophyletic when mapped on clade A(Fig. 4a). Even though the series DIPLO-
STEPHIUM and RUPESTRIA appear to bemonophyletic in clade A, they have represen-tatives in our topologies that are not part ofthis clade suggesting their non-monophyly.For the series SAXATILIA and HUERTASINA,just one species was sampled for each one andno information about their monophyly wasgathered. The series CRASSIFOLIA and PHYLI-
COIDEA sampled in the study are monotypic,and the series CORIACEA was not sampled.This initial evidence does not support thegrouping of species proposed by Cuatrecasas,but the low resolution and support suggestthat in order to prove this, a more reliablephylogenetic analysis is needed.
DISCUSSION
The phylogenetic utility of the chloro-plast markers used in this study (psbA-trnH,rpoB, and rpoC1) is extremely low with mostof the sequences almost identical. For thisreason, we do not recommend these makersfor the study of recently evolved taxa in theAstereae at the infrageneric level and maynot be useful as DNA barcode markers.
The backbone topology obtained in thisstudy in relation to the main groupsformerly described within the Astereaeagreed with previously published phyloge-nies (Noyes and Rieseberg, 1999; Brouilletet al., 2008; Karaman-Castro and Urbatsch,
10 LUNDELLIA DECEMBER, 2012

2009). All the newly sequenced samples ofDiplostephium make up part of the polytomyformed by South American lineages and theNorth American clade. This result confirms
the position of Diplostephium in the SouthAmerican lineages as described by Brouilletet al. (2008), and Karaman-Castro andUrbatsch (2009). The genus Guynesomia
FIG. 4. Clade A character reconstructions. A. Habit and Cuatrecasas’ series in parenthesis.Dip: Diplostephium, Rup: RUPESTRIA, Sax: SAXATILIA, Ana: ANACTINOTA, Den: DENTICULATA,Hue: HERTASINA, Cra: CRASSIFOLIA, Phy: PHYLICOIDEA, Flo: FLORIBUNDA, Ros: ROSMARINIFOLIA,Sch: SCHULTZIANA. B. Leaf size. C. Sinflorescence type. D. Habitat.
NUMBER 15 VARGAS AND MADRINAN: SPECIATION AND CHARACTER EVOLUTION OF DIPLOSTEPHIUM 11

also appears in this polytomy along withDiplostephium and the other South Ameri-can lineages, which does not allow accep-tance or rejection of the hypothesis ofBonifacino and Sancho (2004) that the twogenera are closely related. In this polytomy,the majority of Diplostephium samples (24)formed a monophyletic group (clade A,Figs. 1–3) composed of 22 species with amoderate support in ML and BI. Theremaining Diplostephium samples have con-tradictory positions between the analysesand no support on the topology. Theseresults do not allow the determination of themonophyly or non-monophyly of Diplo-stephium as a whole. Additionally, the SH-test showed no statistical difference betweenthe topology obtained in the analysis and thetopology obtained forcing the genus to bemonophyletic under ML.
Even though our results do not allowconclusions about the monophyly of theentire genus, the data suggest that at leastpart of the genus is monophyletic. Clade Awas recovered in all the topologies withBS570% in ML, and a PP50.95. This claderepresents 80% of the species sampled in thisstudy and contains species from 11 of the 12series treated by Cuatrecasas (1969) (11series were sampled). This means that cladeA is formed by different morphotype groupsdescribed in the genus, and could represent asignificant sample of it. If this assumption iscorrect, we could conclude that even thoughthe traditional genus may not be monophy-letic there is a good probability that at least agood portion of it is.
The large number of species in the genusand low molecular variability of ITS presentwithin Diplostephium in comparison withthat found in other South American taxa,(e.g. Parastrephia with four species, Laesta-dia with five, Blakiella with one, andHinterhubera with eight) support the hy-pothesis of a recent adaptive radiation ofDiplostephium in the Andes. The largemorphological variation that evidence char-acter diversification also supports the idea ofan adaptive radiation. With 70 species of
Diplostephium found in the paramo, onewould expect that the elements that occupythis recently formed habitat would have asimilar or younger age. Paramo vegetation isfound above 3,000 m and has been shown toappear with the emergence of the Andeancordillera 2–5 mya (van der Hammen andCleef, 1986). This recent origin, combinedwith many available niches, explains theradiations seen in many plant taxa nowfound in these extreme habitats, such asEspeletia Mutis ex Humb. and Bonpl.(Rausher, 2002), Jamesonia Hook. & Grev.(Sanchez-Baracaldo, 2004), Valeriana L.(Bell and Donoghue, 2005), and Lupinus L.(Hughes and Eastwood, 2006). Given thesignal obtained in this study it is clear that amore powerful phylogenetic tool like the useof high-throughput DNA sequencing, wherewhole genomes can be obtained in a shortperiod of time, should be implemented toelucidate the phylogenetic relationships ofrecently evolved taxa.
It is significant that members of theseries DIPLOSTEPHIUM (D. colombianum andD. foliosissimum), characterized by beingshrubby and having solitary capitula, arethe sister group to the rest of clade A(Fig. 4a). This result, along with the out-come of Diplostephium arborescent speciesbeing nested in clade A, contradicts thephylogenetic hypothesis proposed by Cua-trecasas (1969) of a forest ancestral mor-photype. Additionally, the character recon-structions also indicate that hypotheticalproto-paramo dwelling species representancestral lineages in clade A. It is importantto note that all Diplostephium species outsideof clade A are predominately shrubby, andtheir uncertain position does not contradictour previous statements. From the biogeo-graphical point of view, no further evalua-tion can be made about the Colombianorigin hypothesis proposed by Cuatrecasas’(1986) due to the low resolution of ourresults. While Diplostephium haenkei, fromsouthern Peru, appears at the base ofDiplostephium non-supported clades in MPand ML, a clade formed by D. colombianum
12 LUNDELLIA DECEMBER, 2012

(western Colombian Cordillera) and D.foliosissimum (northern Peru and Ecuador)is sister to the remainder species in clade A(mostly Colombian) on MP, ML, and BI.These contradictory outcomes leave theformer hypothesis about area of origin tobe tested in the future.
Taking into account the evidence given,we can conclude that the character mappingdoes not support Cuatrecasas’ hypothesisabout the morphological evolution of thegenus. Furthermore, we hypothesize that theancestor of the genus was more likely aparamo-puna-shrub morphotype that orig-inated along the high Andes, with subse-quent colonizations to the Andean forest.Evolution of forest tree species derived fromparamo ancestors has not been previouslydescribed (see van der Hammen and Cleef,1986). This finding adds a new componentto the already complex evolutionary inter-play between the paramo flora and theAndean forest.
ACKNOWLEDGEMENTS
We would like to dedicate this paper tothe memory of Margarita Gomez (1987–2011), friend and botanist murdered inColombia while doing research. We thankthe two anonymous reviewers for theirvaluable comments during the preparationof this manuscript. We are grateful to BerylSimpson, Santiago Dıaz Piedrahita, JulianaChacon, Carlos Mora, Juan A. Sanchez,Vicky Funk, Harold Robinson, MichaelDillon, Carol Kellof, and Mateo Hernan-dez-Smith for their help in different stages ofthis study. Vicki Funk (US), Harold Robin-son (US), Julio Betancur (COL), Luis CarlosJimenez (COL), Adriana Prieto (FMB), andDiego Perico (FMB) facilitated access totheir respective herbarium collections. TheFaculty of Sciences of the Universidad de losAndes (Bogota, Colombia) Seed Grant(2007) and the Smithsonian InstitutionCuatrecasas Award (2006) of the US Na-tional Herbarium provided partial supportfor this study.
LITERATURE CITED
Baldwin, B. G., M. J. Sanderson, J. M. Porter, M. F.
Wojciechowski, C. S. Campbell, and M. J.
Donoghue. 1995. The ITS region of nuclear
ribosomal DNA: a valuable source of evidence on
angiosperm phylogeny. Annals of the Missouri
Botanical Garden 82: 247–277.
Bell, C. D. and M. J. Donoghue. 2005. Phylogeny and
biogeography of Valerianaceae (Dipsacales) with
special reference to the South American valerians.
Organisms Diversity and Evolution 5: 147–159.
Blake, S. F. 1922. Key to the genus Diplostephium, with
descriptions of new species. Contributions from the
United States National Herbarium 24: 65–86.
Blake, S. F. 1928. Review of the genus Diplostephium.American Journal of Botany 15: 43–64.
Bonifacino, J. M. and G. Sancho. 2004. Guynesomia
(Asteraceae: Astereae), a new genus from central
Chile. Taxon 53: 673–678.
Brouillet, L., T. K. Lowrey, L. E. Urbatsch, V.
Karaman-Castro, G. Sancho, S. J. Wagstaff, and J.
C. Semple. 2008. Phylogeny and evolution of the
Astereae (Compositae). In Funk, V. A., T. Stuessy, &
R. Bayer, eds. Systematics, Evolution and Biogeogra-phy of the Compositae. International Association of
Plant Taxonomists, Vienna, pp. 484–523.
Cross, C. W., J. J. Quinn, and S. J. Wagstaff. 2002.
Molecular evidence for the polyphyly of Olearia
(Astereae: Asteraceae). Plant Systematics and Evo-
lution 235: 99–120.
Cuatrecasas, J. 1943. Estudios sobre plantas andinas—
V. Caldasia 2: 209–240.
Cuatrecasas, J. 1969. Prima Flora Colombiana—3
Compositae–Astereae. Webbia 3: 90–194.
Cuatrecasas, J. 1986. Development of the High Andean
Paramo Flora and Vegetation. In: Vuilleumier, F.
and M. Monasterio, eds. High altitude tropical
biogeography. Oxford University Press, Oxford,
pp. 153–201.
Drummond, A. J., B. Ashton, M. Cheung, J. Heled,
M. Kearse, R. Moir, S. Stones-Havas, T. Thierer,
and A. Wilson. 2007. Geneious v3.5. Available at
http://www.geneious.com
Hollingsworth, P. M., L. L. Forrest, J. L. Spouge, M.
Hajibabaei, S. Ratnasingham, M. van der Bank,
M. W. Chase, R. S. Cowan, D. L. Erickson, A. J.
Fazekas, S. W. Graham, K. E. James, K-J. Kim,
W. J. Kress, H. Schneider, J. van Alphen Stahl, S.
C. H. Barrett, C. van den Berg, D. Bogarin, K. S.
Burgess, K. M. Cameron, M. Carine, J. Chacon, A.Clark, F. Conrad, D. S. Devey, J. J. Clarkson, C. S.
Ford, T. A. J. Hedderson, M. L. Hollingsworth,
B. C. Husband, L. J. Kelly, P. R. Kesanakurti, J. S.
Kim, Y. D. Kim, R. Lahaye, H-L. Lee, D. G. Long,
S. Madrinan, O. Maurin, I. Meusnier, S. G.
Newmaster, C-W. Park, D. M. Percy, G. Petersen,
J. E. Richardson, G. A. Salazar, V. Savolainen, O.
NUMBER 15 VARGAS AND MADRINAN: SPECIATION AND CHARACTER EVOLUTION OF DIPLOSTEPHIUM 13

Seberg, M. J. Wilkinson, D-K. Yi, and D. P. Little.2009. A DNA Barcode for Land Plants. Proceedingsof the National Academy of Sciences 106: 12794–12797.
Huelsenbeck, J. P. and F. Ronquist. 2001. MrBayes:Bayesian inference of phylogeny. Bioinformatics 17:754–755.
Hughes, C. and R. Eastwood. 2006. Island radiationon a continental scale: exceptional rates of plantdiversification alter uplift of the Andes. Proceedingsof the National Academy of Sciences 103:10334–10339.
Karaman-Castro, V. and L. E. Urbatsch. 2009.Phylogeny of Hinterhubera group and relatedgenera (Hinterhuberinae: Astereae) based on thenrDNA ITS and ETS sequences. Systematic Botany34: 805–817.
Lowrey, T. K., C. J. Quinn, R. K. Taylor, R. Chan, R. T.Kimball, and J. C. De Nardi JC. 2001. Molecularand Morphological Reassessment of relationshipswithin the Vittadinia Group of Astereae (Astear-aceae). American Journal of Botany 88: 1279–1299.
Maddison, D. R. and W. P. Maddison. 2000.MacClade 4: analysis of phylogeny and characterevolution. Sinauer, Suderland.
Miller, M. A., M. T. Holder, R. Vos R, P. E. Midford,T. Liebowitz, L. Chan, P. Hoover, and T. Warnow.2009. The CIPRES Portals http: //wwwphyloorg/sub_sections/portal.
Nesom, G. 1994. Subtribal classification of the Astereae(Asteraceae). Phytologia 76: 193–274.
Nesom, G. and H. Robinson. 2007. Tribe Astereae, inKadereit, J. W. and C. Jeffrey, eds. The Families andGenera of Vascular Plants vol 8. Springer, Berlin,pp. 284–342.
Noyes, R. and L. Rieseberg. 1999. ITS sequence datasupport a single origin for North AmericanAstereae (Asteraceae) and reflect deep geographicdivisions in Aster s.l. American Journal of Botany86: 398–412.
Posada, D. and K. A. Crandall. 1998. Modeltest:testing the model of DNA substitution. Bioinfor-matics 14: 817–818.
Rauscher, J. 2002. Molecular phylogenetics of theEspeletia complex (Asteraceae): evidence from
nrDNA ITS sequences on the closest relatives of
an Andean adaptive radiation. American Journal of
Botany 89: 1074–1084.
Sanchez-Baracaldo, P. 2004. Phylogenetics and bioge-
ography of the neotropical fern genera Jamesonia
and Eriosorus (Pteridaceae). American Journal of
Botany 91: 274–284.
Sang, T., D. J. Crawford, and T. F. Stuessy. 1997.
Chloroplast DNA phylogeny, reticulate evolution,
and biogeography of Paeonia (Paeoniaceae). Amer-
ican Journal of Botany 84: 1120–1136.
Shimodaira, H. and M. Hasegawa. 1999. Multiple
comparisons of log-likelihoods with applications
to phylogenetic inference. Molecular Biology and
Evolution 16: 1114–1116.
Sikes, D. S. and P. O. Lewis. 2001. PAUPRat: A tool
to implement Parsimony Ratchet searches using
PAUP*. Storrs, Connecticut.
Stamatakis, A. 2006. RAxML-VI-HPC: Maximum
Likelihood-based Phylogenetic Analyses with
Thousands of Taxa and Mixed Models. Bioinfor-
matics 22: 2688–2690.
Swofford, D. L. 1998. PAUP* Phylogenetic analysis
using parsimony (*and other methods), v 4.0 beta
10. Sinauer, Sunderland.
van der Hammen, T. and A. M. Cleef. 1986.
Development of the high Andean paramo flora
and vegetation. In Vuilleumier, F. and M. Mon-
asterio, eds. High Altitude Tropical Biogeography.
Oxford University Press, Oxford, pp. 153–201.
Vargas, O. M. 2011. A nomenclator of Diplostephium
(Asteraceae: Astereae): list of species with their synon-
ymy and distribution by country. Lundellia 14: 32–51.
Vargas, O. M. and S. Madrinan. 2006. Clave para la
identificacion de las especies de Diplostephium
(Asteraceae, Astereae) en Colombia. Revista de la
Academia Colombiana de Ciencias Exactas,
Fısicas y Naturales 30: 489–494.
White, T. J., T. S. Bruns, T. Lee, and J. Taylor. 1990.
Amplification and direct sequencing of fungal
ribosomal RNA genes for phylogenetics. In Innis,
M. A., D. H. Gelfand, J. J. Sninsky, & T. J. White, eds.
PCR protocols: a guide to methods and application.
Academic Press, New York, pp. 315–322.
APPENDICES
APPENDIX 1. List of sequences generated in this study. Data is presented in the following order: taxonname, voucher, herbarium and GenBank accession numbers. Accession numbers appear in following orderwhere more than one sequence were produced: ITS, psbA-trnH, rpoB, and rpoC1. Herbaria: ANDES,Herbario Andes, Universidad de los Andes, Bogota, Colombia; COL, Herbario Nacional Colombiano,Universidad Nacional de Colombia, Bogota, Colombia; FMB, Herbario Federico Medem Bogota, Institutode Investigacion de Recursos Biologicos Alexander von Humboldt, Villa de Leyva, Colombia; US, UnitedStates National Herbarium, Smithsonian Institution, Washington D.C., U.S.A.
Diplostephium alveolatum Cuatrec.,Vargas 183 (ANDES), FJ423515;D. anactinotumWedd.,Cuatrecasas 24557(US), FJ423516, FJ423506;D. azureumCuatrec., Loukuy-Lopez 5295 (US), FJ423517;D. bicolor S.F. Blake, Sarria
14 LUNDELLIA DECEMBER, 2012

462 (COL), FJ423518; D. cayambense Cuatrec., Quenguan 223 (FMB), FJ423519; D. colombianum (Cuatrec.)Cuatrec., Vargas 48 (ANDES), FJ423520, FJ423507; D. crassifolium Cuatrec., Rangel 11293 (COL), FJ423521; D.espinosae Cuatrec., Keating 413 (US), FJ423522;D. floribundum (Benth.) Wedd., Vargas 177 (ANDES), FJ423523;D. foliosissimum S.F. Blake, Smith 12353 (US), FJ423524; D. glandulosum Hieron., Vela 69 (FMB), FJ423525; D.haenkeiWedd., Lewis 88333 (US), FJ423526; Smith 12525 (US), FJ423527;D. heterophyllum Cuatrec., Vargas 162(ANDES), FJ423528; D. inesianum Cuatrec., Barclay 6546 (US), FJ423529; D. jaramilloi Cuatrec., Galvis 356(COL), FJ423530;Galvis 365 (COL), FJ423531;D. lacunosumCuatrec.,Vargas 46 (ANDES), FJ423532, FJ423508;D. ochraceum (HBK) Nees, Vargas 161 (ANDES), FJ423533, FJ423509, FJ423501, FJ423497; D. phylicoidesWedd., Vargas 166 (ANDES), FJ423534, FJ423510, FJ423502, FJ423498; D. revolutum S.F. Blake, Vargas 45(ANDES), FJ423535, FJ423511, FJ423503, FJ423499; D. rhomboidale Cuatrec., Vargas 47 (ANDES), FJ423536,FJ423512; D. romeroi Cuatrec., Barclay 6775 (US), FJ423537; D. rosmarinifolium Wedd., Vargas 165 (ANDES),FJ423538, FJ423513, FJ423504, FJ423500; D. schultzii Wedd. var. orientale Cuatrec., Vargas 164 (ANDES),FJ423539, FJ423514, FJ423505; D. schultzii Wedd. var. schultzii Wedd., Vargas 179 (ANDES), FJ423540; D.tenuifolium Cuatrec., Vargas 184 (ANDES), FJ423541; D. venezuelense Cuatrec., Dorr 7367 (US), FJ423542;Laestadia muscicola Wedd., Zapata 79 (ANDES), FJ423543; Llerasia lindenii Triana, Vargas 185 (ANDES),FJ423544; Oritrophium peruvianum (Lam.) Cuatrec. subsp. lineatum (Cuatrec.) Cuatrec., Zapata 65 (ANDES),FJ423545.
APPENDIX 2. List of previously published GenBank ITS sequences used in this paper.Amellus microglossus DC., DQ478995; Amellus strigosus (Thumb.) Less, AF046942, DQ478996;
Archibaccharis androgyna Blake, DQ478989; Archibaccharis asperifolia (Benth.) Blake, DQ478990;Archibaccharis schiedeana (Benth) J. D. Jackson, DQ478991; Aztecaster matudae (Rzed.) Nesom,DQ479005, DQ479006; Aztecaster pyramidatus (B.L.Rob. and Greenm.) Nesom, DQ479007, DQ479008;Baccharis boliviensis (Wedd.) Cabrera, DQ478992; Blakiella bartsiaefolia (Blake) Cuatrec., DQ479034;Brachyscome rigidula (DC.) G. L. Davis, DQ478994; Chiliotrichum diffusum (G. Forst.) Kuntze, AF046945;Commidendrum robustum DC., AF046943; Conyza pyrrhopappa Schultz-Bip. ex A. Rich., AF046953; Crinitarialinosyris Less., DQ479043; Diplostephium ericoides (Lam.) Cabrera, DQ479003, DQ479004; Diplostephiumrupestre (Kunth) Wedd., AF046962; Doellingeria umbellata (Mill.) Nees, AF046966; Eucephalus glabratus(Greene) Greene, DQ479041; Felicia aethiopica (Burm.f.) Bol. and W.Dod ex Adams and Salt., AF046941,DQ478997; Floscaldasia hypsophila Cuatrec., DQ479009; Grangea maderaspatana (L.) Poir., AF046951;Guynesomia scoparia (Phil.) Bonif. and G. Sancho, DQ479035; Heterothalamus spartioides Hook. and Arn.,DQ478993; Hinterhubera adenopetala Cuatrec. and Aristeg., DQ479010; Hinterhubera columbica Sch. Bip. exWedd., DQ479011; Hinterhubera ericoides Wedd., DQ479012; Hinterhubera imbricata Cuatrec. and Aristeg.,DQ479013;Hinterhubera lanuginosaCuatrec. and Aristeg., DQ479014;Hinterhubera lasegueiWedd., DQ479015;Kalimeris pinnatifida (Maxim.) Kitam., DQ478988; Keysseria maviensis (H. Mann) Cabrera, DQ479036;Laennecia schiedeana (Less.) Nesom, DQ479038; Laennecia sophiifolia (Kunth) Nesom, AF046964; Laestadiacostaricensis S.F.Blake, DQ479016; Laestadia muscicola (Sch. Bip.) Wedd., DQ479017; Laestadia pinifolia Kunth,DQ479018; Lagenifera panamensis Blake, AF046965; Lagenophora pumila Cheeseman, DQ479037; Madagastermadagascariensis (Humbert) Nesom, DQ479031; Madagaster mandrarensis (Humbert) Nesom, DQ479032;Nidorella polycephala DC., DQ478999; Nidorella resedifolia DC., AF046952, DQ479000; Olearia ramulosa(Labill.) Benth., DQ479033; Olearia rosmarinifolia (DC.) Benth., AF497706; Oritrophium hieracioides(Wedd.) Cuatrec., AF046946; Parastrephia lepidophylla (Wedd.) Cabrera, DQ479019, DQ479020;Parastrephia lucida (Meyen) Cabrera, DQ479021, DQ479022; Parastrephia phylicaeformis (Meyen) Cabrera,DQ479023; Parastrephia quadrangularis (Meyen) Cabrera, DQ479024, DQ479025; Parastrephia teretiuscula(Kuntze) Cabrera, DQ479026; Plagiocheilus bogotensis (Kunth) Wedd., DQ479001; Plagiocheilus solivaeformisDC., DQ479002; Podocoma notobellidiastrum (Griseb.) Nesom, AF046963; Psiadia punctulata (DC.) Vatke,AF046954; Sommerfeltia spinulosa Less., DQ479039; Tetramolopium humile (A. Gray) Hillebr. ssp. humile var.humile, DQ479040; Westoniella chirripoensis Cuatrec., DQ479027; Westoniella eriocephala (Klatt) Cuatrec.,DQ479028; Westoniella kohkemperi Cuatrec., DQ479029; Westoniella triunguifolia Cuatrec., DQ479030;Zyrphelis decumbes (Schltr.) Nesom, DQ478998.
NUMBER 15 VARGAS AND MADRINAN: SPECIATION AND CHARACTER EVOLUTION OF DIPLOSTEPHIUM 15