sinorhizobium medicae genes whose regulation involves the acts and/or actr signal transduction...
TRANSCRIPT

FEMS Microbiology Letters 236 (2004) 21–31
www.fems-microbiology.org
Sinorhizobium medicae genes whose regulation involves the ActSand/or ActR signal transduction proteins
Beau J. Fenner a, Ravi P. Tiwari b,*, Wayne G. Reeve b, Michael J. Dilworth b,Andrew R. Glenn c
a Animal Health Biotechnology, Temasek Life Science Laboratory, National University of Singapore, Singapore 117604, Singaporeb Centre for Rhizobium Studies, School of Biological Sciences and Biotechnology, Murdoch University, Perth, WA 6150, Australia
c Office of the Pro Vice Chancellor (Research), University of Tasmania, Hobart, Tasmania 7001, Australia
Received 27 January 2004; received in revised form 7 May 2004; accepted 12 May 2004
First published online 25 May 2004
Abstract
ActS–ActR proteins belong to a highly conserved family of two-component signal transduction systems involved in global
regulation in the a-proteobacteria; they were first identified in Sinorhizobium medicae (previously Sinorhizobium meliloti) as essential
for acid-tolerance. This paper reports on the identification of genes regulated by ActS and/or ActR in S. medicae. To do this,
random gusA fusions were created in S. medicae to follow gene transcription in an actS chromosomal knockout mutant containing
plasmid-borne actS. Plasmid borne actS was cured from the mutants and b-glucuronidase (GUS) activity compared between the
different genetic backgrounds. We detected actS-dependent regulation of the genes gst1 (detoxification), hyuA (hydantoin utiliza-
tion) and fixN2 (microaerobic respiration). We show that ActR is involved in regulating cbbS (CO2 fixation), narB (nitrate as-
similation) and required for low pH and microaerobic induction of the nitrogen fixation regulators fixK and nifA. In particular, we
demonstrate that the transcriptional activation of fixN2 is regulated by ActR through FixK.
� 2004 Federation of European Microbiological Societies. Published by Elsevier B.V. All rights reserved.
Keywords: ActS; ActR; Two-component signal transduction; mTn5 mutagenesis; Low pH induction; General regulation; Redox sensing; Nitrogen
fixation; FixL; FixJ; FixK
1. Introduction
In recent years a new family of closely related two-
component signal transduction systems has been de-scribed in the a-subgroup of Proteobacteria. The first
described members were the RegB histidine kinase and
the RegA response regulator that are required for the
anaerobic induction of the photosynthetic apparatus of
the purple nonsulfur bacterium Rhodobacter capsulatus
[1,2]. RegBA act as a typical two-component system,
with the membrane-spanning RegB histidine kinase
phosphorylating RegA and allowing the activated reg-ulator to modulate transcription by interacting with
specific DNA targets [3–5]. RegBA homologs have been
* Corresponding author. Tel.: +618-9360-2202; fax: +618-9360-6486.
E-mail address: [email protected] (R.P. Tiwari).
0378-1097/$22.00 � 2004 Federation of European Microbiological Societies
doi:10.1016/j.femsle.2004.05.016
discovered in other closely related anoxygenic photo-
trophs [6–9], plant-associated and soil bacteria [10–15]
and more recently in pathogens like Brucella melitensis
[16] and Mycobacterim tuberculosis [17].RegBA homologs regulate a diverse range of meta-
bolic processes including photosystem formation [9,18–
20], CO2 assimilation [21], nitrogen assimilation and
hydrogen uptake [22,23], respiration and electron
transport [24,25] and formaldehyde assimilation [26]. In
the root nodule bacteria, the RegSR system from
Bradyrhizobium japonicum is responsible for microaer-
obic induction of the nitrogen fixation regulator NifA,and regulation of CO2 assimilation [11,27]. RegBA
homologs from Rhodobacter sphaeroides regulate redox-
sensitive processes in response to perturbations in
reductant flow through the electron transport chain
[20,28,29]. DNA target sites for RegA homologs have
. Published by Elsevier B.V. All rights reserved.

22 B.J. Fenner et al. / FEMS Microbiology Letters 236 (2004) 21–31
been identified using footprinting studies and tran-
scriptional analysis [4,23,30–33].
Previously, we described the isolation, sequencing
and characterization of the RegBA homologs ActS and
ActR from the root nodule microsymbiont Sinorhizo-
bium medicae, both of which are essential for growth at
low pH [10]. ActS and ActR have been identified in
Rhizobium leguminosarum; knockout mutations in actS
and actR create acid sensitivity but impart slower
growth in neutral pH conditions [12].
The genetic targets for ActSR in Sinorhizobium have
not yet been elucidated. To initiate our investigation of
genetic regulation by the ActSR system we have created
Table 1
Bacterial strains and plasmids
Strain/plasmid Genotype/phenotype
E. coli
BW20767 RP4-2-tet:Mu-1kan::Tn7 integrant leu-63::IS10 recA1 c
DH5a supE44 DlacU169 (/80 lacZ DM15) hsdR17 recA1 end
Sinorhizobium strains
BF1212S Cmr Kmr Smr; mTn5-GNm mutant of RT295S
BFL11 Cmr Gmr; fixL:pJQ200SK single crossover knockout
BFL11R Cmr Gmr Kmr; fixL:pJQ200SK single crossover knoc
GMI5630 Kmr Smr; D(nod fix)JB16 fixK1::Tn5 recA::Tn5-233 m
Rm1021 Smr derivative of Rm2011
RT295S Cmr Smr; actS::XSmr mutant of WSM419, acids
RTA6S Cmr Kmr Smr; mTn5-GNm mutant of RT295S
RTA15S Cmr Kmr Smr; mTn5-GNm mutant of RT295S
RTG47S Cmr Kmr Smr; mTn5-GNm mutant of RT295S
RTL19S Cmr Kmr Smr; mTn5-GNm mutant of RT295S
RTM11S Cmr Kmr Smr; mTn5-GNm mutant of RT295S
RTN37S Cmr Kmr Smr; mTn5-GNm mutant of RT295S
RTO33S Cmr Kmr Smr; mTn5-GNm mutant of RT295S
TG5-46 Cmr Kmr; actR:Tn5 mutant of WSM419, acids
WSM419 Cmr; wild type, acidt
Plasmids
pBF1212 14.1 kb HindIII fragment containing the BF1212S mT
pBFA15 20.5 kb HindIII fragment containing the RTA15S mT
pBFL11GS 1.1 kb ApaI–XbaI fixL fragment from pDP3 in pJQ2
pBFL19 11.6 kb HindIII fragment containing the RTL19S mT
pBFO33 6.6 kb EcoRI fragment containing the RTO33S mTn5
pCHK57 Tcr; pGD926 containing a 0.3 kb nifA fragment from
pCRS487 Apr Kmr; pUT::mTn5-GNm
pDP3 Gmr; pML133 containing a 2.9 kb blunted SnaI–Bam
pFUS1 Tcr; broad-host-range gusA transcriptional fusion vec
pGEM-7Zf()) Apr; cloning vector
pJJ5 Tcr; pIJ1363 containing a 0.8 kb fixJTK fragment fro
pJQ200SK Gmr; suicide delivery vector with sacB counter-selecta
pKW118 Apr; gusA translational fusion vector with trpA termi
pPFIX1 pFUS1 containing a 1.9-kb blunted BamHI–NotI fixN
pPH1JI Gmr Smr Spr; broad-host-range IncP plasmid
pREG118L pREG750S containing gusA from pKW118 cloned at
pREG118N pBRO1 containing the actR gusA fusion of pREG118
pREG750S S. medicae actR on a 0.8 kb PstI(blunted)–XbaI fragm
pRT546-20 S. medicae actSR on a 4.0 kb BglII–KpnI fragment in
pRT546-6 S. medicae actS on a 3.8 kb BglII fragment in pSW21
pRTA6 7.9 kb EcoRI fragment containing the RTA6S mTn5-
pRTG47 7.6 kb EcoRI fragment containing the RTG47S mTn
pRTM11 5.1 kb EcoRI fragment containing the RTM11S mTn
pRTN37 15.9 kb EcoRI fragment containing the RTN37S mTn
pSW172 Tcr; broad-host-range cloning vector
and screened a library of reporter gene fusions in
S. medicae. By identifying gene fusions regulated by
these proteins we provide insight into the types of pro-
cesses controlled by ActSR.
2. Materials and methods
2.1. Bacterial strains and growth conditions
Strains and plasmids used in this work are listed in
Table 1. Strains of Sinorhizobium were grown at 28 �C in
TY medium or minimal salts medium [34] containing 20
Source/reference
reC510 hsdR17 endA1 zbf-5 gusA (DMluI)::pirþ thi [58]
A1 gyrA96 thi-1 relA1k� [59]
This work
mutant of WSM419 This work
kout mutant of TG5-46 This work
utant of Rm2011 [44]
[60]
This work
This work
This work
This work
This work
This work
This work
This work
[34]
[36]
n5-GNm fusion in pGEM-7Zf()) This work
n5-GNm fusion in pGEM-7Zf()) This work
00SK This work
n5-GNm fusion in pGEM-7Zf()) This work
-GNm fusion in pGEM-7Zf()) This work
S. meliloti Rm2011; nifA lacZ [45]
[36]
HI fixLJT fragment from Rm2011 [61]
tor [36]
Promega
m S. meliloti Rm2011; fixK-lacZ [44]
ble marker [62]
nator [38]
fragment from pBF1212; fixN-gusA This work
[63]
a SalI site in actR; (Plac)actR-gusA This work
L; (PactR)actR-gusA This work
ent in pSW172 This work
pMP220 [10]
3 [10]
GNm fusion in pGEM-7Zf()) This work
5-GNm fusion in pGEM-7Zf()) This work
5-GNm fusion in pGEM-7Zf()) This work
5-GNm fusion in pGEM-7Zf()) This work
[42]

B.J. Fenner et al. / FEMS Microbiology Letters 236 (2004) 21–31 23
mM DLDL-malate and 10 mM NH4Cl as the sole carbon
and nitrogen sources, respectively. Minimal media were
buffered with HEPES (pH 7.0) or MES (pH 5.7) at 40
mM. The final pH of liquid minimal media following
culturing did not vary by more than 0.2 units from theinitial pH. Cultures were routinely grown in 30 ml
McCartney bottles containing five ml of growth me-
dium. Where aerobic growth is specified, cultures were
grown in 100 ml Erlenmeyer flasks containing 10 ml of
growth medium. For microaerobic growth, cultures
were grown in seven ml Bijou bottles containing six ml
of growth medium. Both aerobic and microaerobic
cultures were shaken at 200 rpm on a gyratory platformshaker. Escherichia coli strains were grown at 37 �C in
Luria–Bertani medium [35]. When necessary, antibiotics
were added to cultures at the following concentrations
(lg mL�1): ampicillin, 50; chloramphenicol, 20; genta-
mycin, 10 (E. coli) or 40 (Sinorhizobium); kanamycin, 50;
streptomycin, 50 (E. coli) or 200 (Sinorhizobium); and
tetracycline, 12.5 (E. coli) or 20 (Sinorhizobium).
2.2. Genetic techniques and DNA analysis
Mutagenesis of S. medicae with mTn5-GNm was
performed as described previously [36]. Plasmids were
transferred from E. coli BW20767 hosts into S. medicae
recipients by conjugation [36]. Plasmid or genomic
DNA isolation, manipulation, modification and trans-
pH 7.0 pH 5.7
low pH repressed
low pH induced
no pH effect
curinpPH
p
actS actR
pRT546-6
mTn5-GNm
actS
pREG7
mTn5-G
pRT54
mTn5-G
mutagenesis with mTn5-GNm1
Sinorhizobium medicae RT295S (pRT546-6)
actS actR
pRT546-6
genome
actS
Fig. 1. Schematic diagram of the genetic scree
formation techniques used have been described else-
where [37]. Kanamycin resistant EcoRI fragments were
cloned from each mutant and the DNA sequence
flanking the site of a mTn5-GNm insertion was gener-
ated using the WIL3 [38] and TAC-105F primers [36]and queried against the Sinorhizobium meliloti Rm1021
genomic database [39] using the BLAST algorithm.
2.3. Plasmid mobilization and curing
Plasmids were transferred into Sinorhizobium strains
by conjugation as described previously [40]. Transfer of
pPH1JI and simultaneous loss of pRT546-6 was con-firmed by replica plating and isolating the GmR and TcS
clones (Fig. 1 step 2). Transfer of pMP220, pSW213
(and derivatives) with concomitant loss of pPH1J1 was
confirmed by replica plating and selecting for GmS and
TcR clones (Fig. 1 step 3).
2.4. Enzyme assays
For quantitative measurement of b-glucuronidase(GUS) activity, strains of Sinorhizobium were inoculated
into liquid minimal medium and grown to late log phase
(A600 0.6–0.8). Cells were pelleted, resuspended in saline,
and inoculated into fresh minimal medium at an A600 of
either 0.005–0.01 for pH 7.0 aerobic, 0.1 for pH 7.0
microaerobic, or 0.2 for pH 5.7 to compensate for the
g with1JI
2
3lasmid replacement &quantitative screening
actS actR
pPH1JI
mTn5-GNm
actS+ actS–
actS actR
50S
Nm
Plac
actS actR
pSW172
mTn5-GNm
actR
actS actR
6-20
Nm
actS
actS actR
pMP220
mTn5-GNm
actR
Plac
n used to identify ActR regulated genes.

24 B.J. Fenner et al. / FEMS Microbiology Letters 236 (2004) 21–31
differences in growth rate at neutral pH, microaerobic
and low pH environments. Cultures were then harvested
at A600 0.2–0.6 after 20–25 h of growth. GUS activity
was measured as described previously [40]. b-Galacto-
sidase activities were determined in essentially the sameway except that reactions were carried out in modified
Z-buffer (0.1 M sodium phosphate, pH 7.0, 10 mM KCl,
1 mM MgSO4, 5 mM DTT) using o-nitrophenyl-b-DD-galactopyranoside at a final concentration of 0.4 mg
mL�1 as the substrate. Endogenous b-galactosidase ac-
tivity in Sinorhizobium cells was inactivated as described
previously [41]. Protein concentrations in cell prepara-
tions were determined with a Bio-Rad protein assay kitusing BSA as the protein standard.
3. Results
3.1. Genetic screen to identify ActS-responsive genes
The ActSR orthologs are involved in global gene reg-ulation in the a-proteobacteria. A genetic screen
was conducted to identify the types of genes regulated in
S. medicae through ActS (histidine kinase ‘sensor’ pro-
tein). The first step in this approach was to construct a
library of mTn5-GNm induced mutants of RT295S
(actS::XSmr) containing pRT546-6 (an actS-comple-
menting plasmid), thereby generating a pool of strains
containing transcriptional fusions to gusA (Fig. 1, step 1).This approach enabled us to later eliminate actS by
plasmid curing (Fig. 1, step 2). Approximately 5000
mutants were patched onto pH 7.0 or 5.7 minimal me-
dium plates containing the chromogenic GUS substrate
X-Glc and inspected for differences in color development
to identify pH-responsive gusA transcriptional fusions.
Transconjugants showing differences in the intensity of
blue color at low and neutral pH buffered plates wereselected for further screening. The selected strains were
Table 2
Effect of multicopy actSR or overexpressed actR on expression of gusA tran
Mutant b-Glucuronidase specific activity (nmol pNP min�1 [mg
Comparison 1 Difference (fold)b
pMP220a
(control)
pRT546-20
(multicopy actSR)
RTA6S 17.0� 5.1 4.4� 0.9 # 3.9�RTA15S 41.5� 0.4 35.8� 4.9
RTG47S 58.9� 7.7 38.1� 2.1
RTL19S 79.2� 4.5 329� 28.5 " 4.2�RTM11S 20.1� 1.4 8.5� 0.1 # 2.4�RTN37S 127� 13.6 238� 1.2 " 1.9
RTO33S 352� 69.3 20.7� 0.1 # 17�BF1212S 3.3� 1.0 12.3� 3.0 " 3.7�a The low copy number parent plasmids pMP220 and pSW172 are maintabGUS activity was either induced ð"Þ or repressed ð#Þ in mutants contain
pMP220 or pSW172, respectively. Values shown are the average of at least tw
first cured of the existing actS plasmid (pRT546-6) by
mobilizing the incompatible plasmid pPH1J1. Another
plasmid construct (pRT546-20, pMP220, pREG750S or
pSW172) was then reintroduced using conjugational
transfer and transconjugants devoid of pPH1J1 wereisolated. To determine if ActS regulated the transcription
of selected fusions, GUS activity was compared between
isogenic strains either containing, or devoid of, actS
(Fig. 1, step 3). Quantitation of GUS revealed that the
expression of six transcriptional fusions differed by �2-
fold or more between these two genetic backgrounds
(Table 2). Expression of GUS decreased in three mutants
(RTA6S, RTM11S, RTO33S) but increased in anotherthree (RTL19S, RTN37S, BF1212S) in the presence of
multicopy actSR (Table 2). Transcriptional fusions
whose expression did not differ by more than 20% be-
tween the different actS genetic backgrounds were con-
sidered as being actS unresponsive (data not shown).
3.2. Isolation of ActR regulated genes
Previously, we showed that multiple copies of actR
rescue an actS mutant from acid-sensitivity [10] sug-
gesting that the presence of multiple copies of actR
overrides the requirement for activation of ActR by
ActS. We therefore reasoned that additional genetic
targets could be identified from the pool of pH-re-
sponsive gusA fusions by screening cells in the presence
and absence of an excess dosage of ActR. Mobilizationof pREG750S into S. medicae mutants allowed for high-
level expression of actR from Plac in pSW172 [42] since
the Plac promoter in this plasmid yields approximately
13 times more ActR than the native PactR promoter
(data not shown). By comparing GUS activities between
isogenic strains containing pREG750S or pSW172, two
mutants (RTA15S and RTG47S) were identified that
contained fusions that were down-regulated by overex-pressed actR (Table 2). Comparisons of GUS activities
scriptional fusions in actS mTn5-GNm mutants
protein]�1)
Comparison 2 Difference (fold)b
pSW172a
(control)
pREG750S
(multicopy Plac-actR)
15.7� 1.6 14.7� 0.4
57.7� 8.5 21.6� 0.1 # 2.7�49.1� 6.1 13.7� 1.6 # 3.6�66.2� 6.7 34.3� 0.2 # 1.9�23.1� 5.7 19.1� 0.2
138� 2.5 169� 5.1
281� 39.5 552� 35.8 " 2.0�3.4� 0.7 3.2� 0.2
ined at 5–10 copies per cell [42,64].
ing pRT546-20 or pREG750S in comparison to the parent plasmids
o independent cultures grown at pH 7.0 and assayed in quadruplicate.

B.J. Fenner et al. / FEMS Microbiology Letters 236 (2004) 21–31 25
for the six initially identified mutants revealed that in the
case of RTL19S and RTO33S, the regulation of GUS
activity was dependent on both the presence of actS and
the dosage of actR (Table 2).
3.3. Effect of pH on actS or actR responsive fusions
Comparisons of GUS activity between mTn5 mu-
tants complemented with plasmid-borne actSR grown at
either pH 7.0 or 5.7 showed either no effect on expres-
sion or a difference of up to 7-fold induction (Fig. 2).
Although initial selection of mutants was based on dif-
ferences in colour development on neutral and low pHmedia plates containing the substrate X-Glc, surpris-
ingly two of the mutants RTM11S and RTN37S did not
Fig. 2. Low-pH regulation of ActSR-regulated gusA fusions in mTn5-
GNm mutants. GUS activities of mTn5-GNm mutants containing
either pRT546-20 (multicopy-actSR; filled bars) or pMP220 in an actS
background (unfilled bars) and grown at pH 7.0 have been compared
with mutants grown at pH 5.7. Values of approximately 1 (indicated
by a dotted line) represent no significant difference in GUS activity
between the different pH values. GUS activities (standard error <15%)
were measured from at least two independent cultures assayed in
quadruplicate.
Table 3
Sequence homologies of DNA regions flanking the sites of mTn5-GNm inse
Mutant Homologous gene(s) Putative gene function
RTA6Sb SMc04059/glgP Hypothetical protein/glycogen phosp
RTA15S cbbS Ribulose-1,5-bisphosphate carboxyla
subunit
RTG47S narB Assimilatory nitrate reductase large
RTL19Sb SMc00795 Hypothetical protein
RTM11S hyuA Hydantoin utilization protein
RTN37Sb gst1 Glutathione S-transferase
RTO33S SMc00888 PAS two-component receiver protein
BF1212S fixN2O2 cbb3 Cytochrome oxidase componen
Sequences were compared to the S. meliloti Rm1021 genomic database [a Significance of homology refers to the probability that such a match woubThe mTn5-GNm insertion in RTN37S is 10 bp downstream from the puta
the putative stop codon of SMc4059 and 121 bp upstream of the putative s
putative start codon of SMc00795.
show pH dependent regulation when GUS activity was
quantified in broth cultures. It demonstrates the im-
portance of GUS quantification in broth cultures to
establish real differences in gene expression.
When the mTn5 mutants devoid of plasmid-borneactSR were grown in the same conditions, a similar level
of pH response was observed with all mutants with the
exception of BF1212S. In this strain complemented with
plasmid-borne actSR the gusA fusion was induced ap-
proximately 4-fold at low pH (Fig. 2) with the activity
rising from approximately 14 to 60 nmol pNP min�1
(mg protein)�1 at pH 7.0 versus pH 5.7 respectively.
However, in an actS background, GUS activity at bothpH 7.0 and 5.7 fell to approximately 3.0 nmol pNP
min�1 (mg protein)�1.
3.4. Sequence analysis of DNA flanking mTn5-GNm
insertions
Southern hybridization experiments revealed that
each of the selected mutants was the result of a singlegenomic mTn5-GNm insertion event (data not shown).
Sequence comparison with the homologous S. meliloti
Rm1021 sequence showed that insertions occurred in
eight unique positions (Table 3) with four located in
intergenic regions and four intragenic. The mTn5-GNm
insertion in RTN37S occurred downstream from a
putative stop codon of a gene encoding glutathione S-
transferase. The insertion in RTL19S was 47 bp up-stream of the putative start codon of a gene encoding a
hypothetical protein showing similarity to OsmB (os-
motically induced lipoprotein B precursor) from E. coli.
The mutant RTA6S contains mTn5-GNm between the
stop codon of a gene encoding a hypothetical protein
and the start codon of glgP, encoding glycogen phos-
phorylase. The BF1212S insertion occurred within a 13
bp intergenic sequence between the fixN2 and fixO2
genes. Intragenic insertions occurred in the mutants
RTA15S, RTM11S, RTO33S and RTG47S, disrupting
rtion in ActSR-responsive mutants
Genome location in Rm1021 Significance of homologya
horylase Chromosome 9� 10�37/6� 10�67
se small pSymB 0
subunit pSymB 0
Chromosome 4� 10�8
pSymB 0
Chromosome 2� 10�69
Chromosome 6� 10�81
ts pSymA 0
39].
ld occur by chance as determined by the BLASTX algorithm [65].
tive stop codon of gst1; the insertion in RTA6S is 77 bp downstream of
tart codon of glgP; the insertion of RTL19S is 47 bp upstream of the

26 B.J. Fenner et al. / FEMS Microbiology Letters 236 (2004) 21–31
genes encoding the small subunit of RuBisCO, a hy-
dantoin utilization protein, a PAS protein containing a
two-component receiver domain and the large subunit
of an assimilatory nitrate reductase (Table 3). Sequence
comparisons made with the S. meliloti Rm1021 genomicdatabase showed that the ActSR-regulated genes are
distributed throughout each of the three replicons of S.
meliloti (chromosome, pSymA, and pSymB).
3.5. ActSR and FixLJ coordinately regulate fixN2
expression in S. medicae
Expression of fixN in S. meliloti is dependent on atleast two regulators; the FixJ response regulator, which
is activated by the FixL histidine kinase, and the FNR-
family FixK regulator [43]. Though a functional copy of
actS is clearly required for the low pH induction of
fixN2 in S. medicaeWSM419, this effect could have been
mediated through these regulators. We therefore created
a single crossover fixL knockout mutation in parental
and actR genetic backgrounds. A fixN2-gusA tran-scriptional reporter plasmid, pPFIX1, was then mobi-
lized into both of these strains and also into parental,
actS and actR strains. In the wild type background,
expression of fixN2 was induced 16-fold by microaero-
biosis at pH 7.0 and 2-fold by low pH (pH 5.7) under
aerobic conditions (Table 4). In actS and actR back-
grounds, expression of fixN2 was reduced to approxi-
mately half that observed in the wild type under bothaerobic and microaerobic conditions, while the low pH
induction of fixN2 was essentially abolished. The mi-
croaerobic induction of fixN2 at low pH was similar to
that observed at neutral pH, with GUS activities for
microaerobically grown wild type cells of 605� 87.2 and
490� 67.1 nmol pNP min�1 (mg protein)�1 at pH 7.0
and 5.7, respectively. Inactivation of fixL reduced the
microaerobic induction of fixN2 from 16-fold to 2-fold,but did not affect the observed low pH induction (Table
Table 4
Regulation of fixN 2 by ActSR, FixLJ and FixK. GUS assays were performed
gusA reporter plasmid pPFIX1
b-Glucuronidase specific activity
Strain and genotype pH 7.0 aerobic
S. medicae strains
WSM419 (wild type) 36.6� 0.5
RT295S (actS) 16.3� 3.2
TG5-46 (actR) 19.5� 0.9
BFL11 (fixL) 32.1� 1.9
BFL11R (actR-fixL) 15.1� 1.2
S. meliloti strains
Rm1021 (wild type) 38.3� 1.9
GMI5630 (fixK1-fixK2) 30.4� 1.3
Values shown are the average of at least two independent cultures assay
4). In an actR-fixL double mutant the 2-fold microaer-
obic and low pH induction of fixN2 was undetectable,
indicating that both ActR and FixL positively regulate
fixN2 during microaerobiosis but that only ActR directs
a low pH transcriptional response.
3.6. FixK is required for low pH induction of fixN2 in
S. lmeliloti
Microaerobic activation of fixN by the FixLJ system
is channelled through the FixK regulator in S. meliloti
[44]. The possibility of ActR activating fixN expression
through FixK in S. meliloti was explored by examiningexpression of the fixN2-gusA fusion on plasmid pPFIX1
in the mutant GMI5630 [44] and the wild type S. meliloti
1021; GMI5630 does not carry a functional fixK gene as
only two copies of fixK are present in the S. meliloti 1021
genome [39]. The expression of fixN2 fusion derived
from S. medicae was increased 3-fold in strain S. meliloti
1021 by low pH, whereas in the S. meliloti fixK mutant
there was no significant change at low pH (Table 4).
3.7. fixK is regulated through ActR in S. medicae
It was reasoned that if FixK is required for both the
microaerobic and low pH induction of fixN2, then ActR
should regulate the expression of fixK in a microaerobic
and low pH-dependent manner. This possibility was
investigated by assaying S. medicae parental and actR
strains carrying the S. meliloti fixK-lacZ translational
fusion on the plasmid pJJ5 [44]. In both strains of S.
medicae WSM419 the fixK fusion was induced 30- to 40-
fold during microaerobic growth, though the overall
level of expression in the actR strain was only half that
of the parental strain. The fusion was also induced 13-
fold in the parental strain when exposed to acidity, but
only 3-fold in the actR strain exposed to the same con-ditions (Fig. 3).
on the indicated strains of S. medicae and S. meliloti carrying the fixN2
(nmol pNP�1 min�1 [mg protein]�1)
pH 7.0 microaerobic pH 5.7 aerobic
605� 87.2 66.8� 5.4
366� 20.4 17.8� 0.2
358� 15.0 26.8� 2.8
60.3� 6.1 64.9� 1.8
17.3� 0.4 14.5� 1.1
409� 36.2 106� 9.4
34.3� 1.6 37.7� 2.9
ed at quadruplicate.
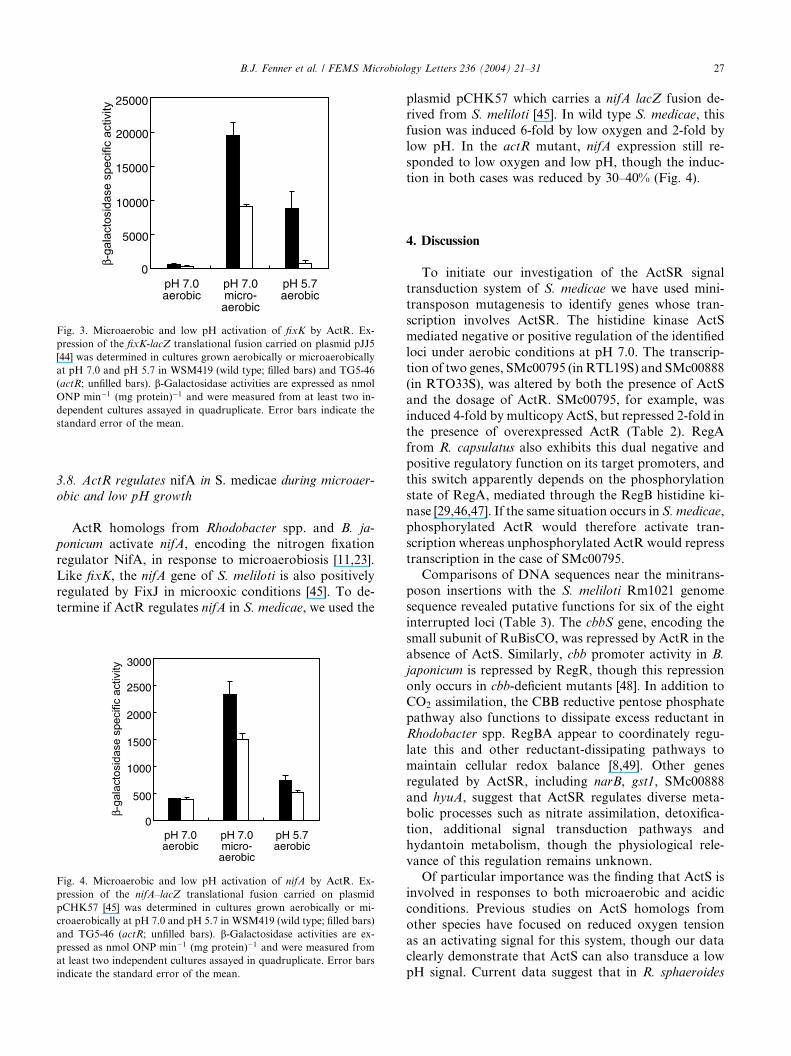
Fig. 3. Microaerobic and low pH activation of fixK by ActR. Ex-
pression of the fixK-lacZ translational fusion carried on plasmid pJJ5
[44] was determined in cultures grown aerobically or microaerobically
at pH 7.0 and pH 5.7 in WSM419 (wild type; filled bars) and TG5-46
(actR; unfilled bars). b-Galactosidase activities are expressed as nmol
ONP min�1 (mg protein)�1 and were measured from at least two in-
dependent cultures assayed in quadruplicate. Error bars indicate the
standard error of the mean.
B.J. Fenner et al. / FEMS Microbiology Letters 236 (2004) 21–31 27
3.8. ActR regulates nifA in S. medicae during microaer-
obic and low pH growth
ActR homologs from Rhodobacter spp. and B. ja-
ponicum activate nifA, encoding the nitrogen fixation
regulator NifA, in response to microaerobiosis [11,23].
Like fixK, the nifA gene of S. meliloti is also positively
regulated by FixJ in microoxic conditions [45]. To de-
termine if ActR regulates nifA in S. medicae, we used the
500
1000
1500
2000
2500
0pH 7.0aerobic
pH 7.0micro-aerobic
pH 5.7aerobic
3000
Fig. 4. Microaerobic and low pH activation of nifA by ActR. Ex-
pression of the nifA–lacZ translational fusion carried on plasmid
pCHK57 [45] was determined in cultures grown aerobically or mi-
croaerobically at pH 7.0 and pH 5.7 in WSM419 (wild type; filled bars)
and TG5-46 (actR; unfilled bars). b-Galactosidase activities are ex-
pressed as nmol ONP min�1 (mg protein)�1 and were measured from
at least two independent cultures assayed in quadruplicate. Error bars
indicate the standard error of the mean.
plasmid pCHK57 which carries a nifA lacZ fusion de-
rived from S. meliloti [45]. In wild type S. medicae, this
fusion was induced 6-fold by low oxygen and 2-fold by
low pH. In the actR mutant, nifA expression still re-
sponded to low oxygen and low pH, though the induc-tion in both cases was reduced by 30–40% (Fig. 4).
4. Discussion
To initiate our investigation of the ActSR signal
transduction system of S. medicae we have used mini-
transposon mutagenesis to identify genes whose tran-scription involves ActSR. The histidine kinase ActS
mediated negative or positive regulation of the identified
loci under aerobic conditions at pH 7.0. The transcrip-
tion of two genes, SMc00795 (in RTL19S) and SMc00888
(in RTO33S), was altered by both the presence of ActS
and the dosage of ActR. SMc00795, for example, was
induced 4-fold by multicopy ActS, but repressed 2-fold in
the presence of overexpressed ActR (Table 2). RegAfrom R. capsulatus also exhibits this dual negative and
positive regulatory function on its target promoters, and
this switch apparently depends on the phosphorylation
state of RegA, mediated through the RegB histidine ki-
nase [29,46,47]. If the same situation occurs in S. medicae,
phosphorylated ActR would therefore activate tran-
scription whereas unphosphorylated ActR would repress
transcription in the case of SMc00795.Comparisons of DNA sequences near the minitrans-
poson insertions with the S. meliloti Rm1021 genome
sequence revealed putative functions for six of the eight
interrupted loci (Table 3). The cbbS gene, encoding the
small subunit of RuBisCO, was repressed by ActR in the
absence of ActS. Similarly, cbb promoter activity in B.
japonicum is repressed by RegR, though this repression
only occurs in cbb-deficient mutants [48]. In addition toCO2 assimilation, the CBB reductive pentose phosphate
pathway also functions to dissipate excess reductant in
Rhodobacter spp. RegBA appear to coordinately regu-
late this and other reductant-dissipating pathways to
maintain cellular redox balance [8,49]. Other genes
regulated by ActSR, including narB, gst1, SMc00888
and hyuA, suggest that ActSR regulates diverse meta-
bolic processes such as nitrate assimilation, detoxifica-tion, additional signal transduction pathways and
hydantoin metabolism, though the physiological rele-
vance of this regulation remains unknown.
Of particular importance was the finding that ActS is
involved in responses to both microaerobic and acidic
conditions. Previous studies on ActS homologs from
other species have focused on reduced oxygen tension
as an activating signal for this system, though our dataclearly demonstrate that ActS can also transduce a low
pH signal. Current data suggest that in R. sphaeroides

Fig. 5. Model comparing low oxygen and/or low pH regulation of fix and nif genes by the FixLJ and ActSR (RegSR) two-component systems of
(a) Sinorhizobium and (b) B. japonicum. During growth under low oxygen and/or low pH conditions, the membrane-bound histidine kinases FixL and
ActS/RegS activate their respective response regulator partners FixJ and ActR/RegR. The active response regulators then regulate the expression
target genes, presumably by binding to upstream promoter elements. The symbol � has been used to indicate either direct or indirect involvement.
28 B.J. Fenner et al. / FEMS Microbiology Letters 236 (2004) 21–31
the flow of reductant through the microaerobically
inducible cbb3 cytochrome oxidase generates an inhib-
itory signal under aerobic conditions to maintain the
ActS homolog, PrrB, in the phosphatase-dominant
state, thus preventing activation of the PrrA response
regulator [20,28,50,51]. Similarly, oxidized electroncarriers generated by terminal oxidases inhibit the ki-
nase activity of the ArcB histidine kinase of E. coli,
preventing phosphorylation of the global response
regulator ArcA under aerobic conditions [52]. The
finding that both ActR (this study) and ArcA [53] can
transduce a low pH signal in addition to a microaerobic
signal suggests that low pH stimulates both ArcB and
the ActS homologs. It is possible that at low pH theproton pumping activity [54–56] of the inhibiting ter-
minal oxidases is retarded, leading to a reduced re-
ductant flow and therefore activation of the histidine
kinase. Interestingly, defects in cytochrome bd oxidase
expression caused by the interruption of cydB in Bru-
cella abortus lead to extreme acid sensitivity [57]. This
sensitivity is postulated to occur due to an accumula-
tion of oxidative radicals at low pH that overruns thecell’s protective enzymes such as superoxide dismutase
and catalase [57]. In light of these findings, future work
will focus on the potential involvement of electron
transport components such as cbb3 oxidase in the acid
tolerance of S. medicae.
To our knowledge this is the first report of a second
two-component system, in addition to FixLJ, that mod-
ulates expression of the fixK-fixNOQP and nifA genes inSinorhizobium. Unlike Sinorhizobium, the microaerobic
induction of nifA in B. japonicum is independent of
FixLJ, but absolutely requires the ActSR homologs,
RegSR [11]. In B. japonicum, therefore, an intact RegSR
system is essential for symbiotic N2fixation [11], while the
FixLJ system is also essential through its induction of the
FixNOQP system through FixK2 (Fig. 5(b)).
The situation is markedly different in S. medicae,
where the actR mutant, TG5-46, fixes nitrogen symbi-otically at rates comparable to the wild type (unpub-
lished observations). To integrate our observations of
the regulation of fixK-fixNOQP and nifA by ActSR and
FixLJ in S. medicae we propose a working model of
this regulatory circuit, illustrated in Fig. 5(a). Low O2
tension and low pH signals are transmitted via ActS
and ActR to positively affect transcription of fixK, fixN
and nifA, though it is unclear whether these effects ofActR are directly on the relevant promoters or indirect.
Though an intact ActSR system is required for maxi-
mum microaerobic induction of nifA, microaerobic in-
duction of nifA by the FixLJ system in S. medicae is
apparently fully sufficient for symbiotic nitrogen fixa-
tion even in the absence of ActSR, thus explaining the
difference in symbiotic behaviour of mutants in
RegSR and ActSR in B. japonicum and S. medicae,respectively.
Acknowledgements
We thank Jacques Batut, Daniel Kahn, Gary Ditta
and Stephen Winans for supplying strains and plasmids
used in this study. B. Fenner was supported by anAustralian Postgraduate Award. Additional support
was provided by the Australian Research Council.

B.J. Fenner et al. / FEMS Microbiology Letters 236 (2004) 21–31 29
References
[1] Sganga, M.W. and Bauer, C.E. (1992) Regulatory factors
controlling photosynthetic reaction center and light-harvesting
gene expression in Rhodobacter capsulatus. Cell 68, 945–954.
[2] Mosley, C.S., Suzuki, J.Y. and Bauer, C.E. (1994) Identification
and molecular genetic characterization of a sensor kinase respon-
sible for coordinately regulating light harvesting and reaction
center gene expression in response to anaerobiosis. J. Bacteriol.
176, 7566–7573.
[3] Ouchane, S. and Kaplan, S. (1999) Topological analysis of the
membrane-localized redox-responsive sensor kinase PrrB from
Rhodobacter sphaeroides 2.4.1. J. Biol. Chem. 274, 17290–17296.
[4] Du, S., Bird, T.H. and Bauer, C.E. (1998) DNA binding
characteristics of RegA*: a constitutively active anaerobic activa-
tor of photosynthesis gene expression in Rhodobacter capsulatus.
J. Biol. Chem. 273, 18509–18513.
[5] Du, S., Kouadio, J.L. and Bauer, C.E. (1999) Regulated expres-
sion of a highly conserved regulatory gene cluster is necessary for
controlling photosynthesis gene expression in response to anaero-
biosis in Rhodobacter capsulatus. J. Bacteriol. 181, 4334–4341.
[6] Eraso, J.M. and Kaplan, S. (1994) prrA, a putative response
regulator involved in oxygen regulation of photosynthesis gene
expression in Rhodobacter sphaeroides. J. Bacteriol. 176, 32–43.
[7] Eraso, J.M. and Kaplan, S. (1995) Oxygen-insensitive synthesis of
the photosynthetic membranes of Rhodobacter sphaeroides: a
mutant histidine kinase. J. Bacteriol. 177, 2695–2706.
[8] Joshi, H.M. and Tabita, F.R. (1996) A global two-component
signal transduction system that integrates the control of photo-
synthesis, carbon dioxide assimilation, and nitrogen fixation.
Proc. Natl. Acad. Sci. USA 93, 14515–14520.
[9] Masuda, S., Matsumoto, Y., Nagashima, K.V., Shimada, K.,
Inoue, K., Bauer, C.E. and Matsuura, K. (1999) Structural and
functional analyses of photosynthetic regulatory genes regA and
regB from Rhodovulum sulfidophilum, Roseobacter denitrificans,
and Rhodobacter capsulatus. J. Bacteriol. 181, 4205–4215.
[10] Tiwari, R.P., Reeve, W.G., Dilworth, M.J. and Glenn, A.R.
(1996) Acid tolerance in Rhizobium meliloti strain WSM419
involves a two-component sensor-regulator system. Microbiology
142, 1693–1704.
[11] Bauer, E., Kaspar, T., Fischer, H.M. and Hennecke, H. (1998)
Expression of the fixR-nifA operon in Bradyrhizobium japonicum
depends on a new response regulator. RegR. J. Bacteriol. 180,
3853–3863.
[12] Boesten, B., Schl€uter, A., Prell, J., Kr€amer, M., Lipka, V. and
Priefer, U.B. (2000) Identifying genes suitable for constructing pH
and salt tolerant Rhizobium inoculants for improving French bean
cultivation under semiarid conditions. In: Proceedings of the
Fourth European Nitrogen Fixation Conference (Olivares, J.,
Palomares, A.J., Eds.), Vol., p. 319. Viceconsejer�ıa, Seville.
[13] Kaneko, T., Nakamura, Y., Sato, S., Asamizu, E., Kato, T.,
Sasamoto, S., Watanabe, A., Idesawa, K., Ishikawa, A., Kawa-
shima, K., Kimura, T., Kishida, Y., Kiyokawa, C., Kohara, M.,
Matsumoto, M., Matsuno, A., Mochizuki, Y., Nakayama, S.,
Nakazaki, N., Shimpo, S., Sugimoto, M., Takeuchi, C., Yamada,
M. and Tabata, S. (2000) Complete genome structure of the
nitrogen-fixing symbiotic bacterium Mesorhizobium loti. DNA
Res. 7, 381–406.
[14] Wood, D.W., Setubal, J.C., Kaul, R., Monks, D.E., Kitajima,
J.P., Okura, V.K., Zhou, Y., Chen, L., Wood, G.E., Almeida Jr.,
N.F., Woo, L., Chen, Y., Paulsen, I.T., Eisen, J.A., Karp, P.D.,
Bovee Sr., D., Chapman, P., Clendenning, J., Deatherage, G.,
Gillet, W., Grant, C., Kutyavin, T., Levy, R., Li, M.J., McClel-
land, E., Palmieri, A., Raymond, C., Rouse, G., Saenphimma-
chak, C., Wu, Z., Romero, P., Gordon, D., Zhang, S., Yoo, H.,
Tao, Y., Biddle, P., Jung, M., Krespan, W., Perry, M., Gordon-
Kamm, B., Liao, L., Kim, S., Hendrick, C., Zhao, Z.Y., Dolan,
M., Chumley, F., Tingey, S.V., Tomb, J.F., Gordon, M.P., Olson,
M.V. and Nester, E.W. (2001) The genome of the natural genetic
engineer Agrobacterium tumefaciens C58. Science 294, 2317–2323.
[15] Nierman, W.C., Feldblyum, T.V., Laub, M.T., Paulsen, I.T.,
Nelson, K.E., Eisen, J., Heidelberg, J.F., Alley, M.R., Ohta, N.,
Maddock, J.R., Potocka, I., Nelson, W.C., Newton, A., Stephens,
C., Phadke, N.D., Ely, B., DeBoy, R.T., Dodson, R.J., Durkin,
A.S., Gwinn, M.L., Haft, D.H., Kolonay, J.F., Smit, J., Craven,
M.B., Khouri, H., Shetty, J., Berry, K., Utterback, T., Tran, K.,
Wolf, A., Vamathevan, J., Ermolaeva, M., White, O., Salzberg,
S.L., Venter, J.C., Shapiro, L. and Fraser, C.M. (2001) Complete
genome sequence of Caulobacter crescentus. Proc. Natl. Acad. Sci.
USA 98, 4136–4141.
[16] DelVecchio, V.G., Kapatral, V., Redkar, R.J., Patra, G., Mujer,
C., Los, T., Ivanova, N., Anderson, I., Bhattacharyya, A.,
Lykidis, A., Reznik, G., Jablonski, L., Larsen, N., D’Souza, M.,
Bernal, A., Mazur, M., Goltsman, E., Selkov, E., Elzer, P.H.,
Hagius, S., O’Callaghan, D., Letesson, J.J., Haselkorn, R.,
Kyrpides, N. and Overbeek, R. (2002) The genome sequence of
the facultative intracellular pathogen Brucella melitensis. Proc.
Natl. Acad. Sci. USA 99, 443–448.
[17] Ewann, F., Locht, C. and Supply, P. (2004) Intracellular
autoregulation of the Mycobacterium tuberculosis PrrA response
regulator. Microbiology 150, 241–246.
[18] Bauer, C.E. and Bird, T.H. (1996) Regulatory circuits controlling
photosynthesis gene expression. Cell 85, 5–8.
[19] O’Gara, J.P., Eraso, J.M. andKaplan, S. (1998)A redox-responsive
pathway for aerobic regulationof photosynthesis gene expression in
Rhodobacter sphaeroides 2.4.1. J. Bacteriol. 180, 4044–4050.
[20] Oh, J.I., Eraso, J.M. and Kaplan, S. (2000) Interacting regulatory
circuits involved in orderly control of photosynthesis gene
expression in Rhodobacter sphaeroides 2.4.1. J. Bacteriol. 182,
3081–3087.
[21] Vichivanives, P., Bird, T.H., Bauer, C.E. and Tabita, F.R. (2000)
Multiple regulators and their interactions in vivo and in vitro with
the cbb regulons of Rhodobacter capsulatus. J. Mol. Biol. 300,
1079–1099.
[22] Laratta, W.P., Choi, P.S., Tosques, I.E. and Shapleigh, J.P. (2002)
Involvement of the PrrB/PrrA two-component system in nitrite
respiration in Rhodobacter sphaeroides 2.4.3: evidence for tran-
scriptional regulation. J. Bacteriol. 184, 3521–3529.
[23] Elsen, S., Dischert, W., Colbeau, A. and Bauer, C.E. (2000)
Expression of uptake hydrogenase and molybdenum nitrogenase
in Rhodobacter capsulatus is coregulated by the RegB–RegA two-
component regulatory system. J. Bacteriol. 182, 2831–2837.
[24] Swem, L.R., Elsen, S., Bird, T.H., Swem, D.L., Koch, H.G.,
Myllykallio, H., Daldal, F. and Bauer, C.E. (2001) The RegB/
RegA two-component regulatory system controls synthesis of
photosynthesis and respiratory electron transfer components in
Rhodobacter capsulatus. J. Mol. Biol. 309, 121–138.
[25] Kappler, U., Huston, W.M. and McEwan, A.G. (2002) Control of
dimethylsulfoxide reductase expression in Rhodobacter capsulatus:
the role of carbon metabolites and the response regulators DorR
and RegA. Microbiology 148, 605–614.
[26] Barber, R.D. and Donohue, T.J. (1998) Pathways for transcrip-
tional activation of a glutathione-dependent formaldehyde dehy-
drogenase gene. J. Mol. Biol. 280, 775–784.
[27] Fischer, H.M., Emmerich, R., Chanfon, A., and Hennecke, H.
(2000) Characterization of the global two-component regulatory
system RegSR in Bradyrhizobium japonicum. In: Proceedings of
the Fourth European Nitrogen Fixation Conference, Viceconse-
jer�ia, Seville (Olivares, J., Palomares, A.J., Eds.), Vol., p. 108.
[28] Eraso, J.M. and Kaplan, S. (2000) From redox flow to gene
regulation: role of the PrrC protein of Rhodobacter sphaeroides
2.4.1. Biochemistry 39, 2052–2062.

30 B.J. Fenner et al. / FEMS Microbiology Letters 236 (2004) 21–31
[29] Swem, L.R., Kraft, B.J., Swem, D.L., Setterdahl, A.T., Masuda,
S., Knaff, D.B., Zaleski, J.M. and Bauer, C.E. (2003) Signal
transduction by the global regulator RegB is mediated by a redox-
active cysteine. EMBO J. 22, 4699–4708.
[30] Karls, R.K., Wolf, J.R. and Donohue, T.J. (1999) Activation of
the cycA P2 promoter for the Rhodobacter sphaeroides cytochrome
c2 gene by the photosynthesis response regulator. Mol. Microbiol.
34, 822–835.
[31] Bird, T.H., Du, S. and Bauer, C.E. (1999) Autophosphorylation,
phosphotransfer, and DNA-binding properties of the RegB/RegA
two-component regulatory system in Rhodobacter capsulatus. J.
Biol. Chem. 274, 16343–16348.
[32] Emmerich, R., Panglungtshang, K., Strehler, P., Hennecke, H.
and Fischer, H.M. (1999) Phosphorylation, dephosphorylation
and DNA-binding of the Bradyrhizobium japonicum RegSR two-
component regulatory proteins. Eur. J. Biochem. 263, 455–463.
[33] Dubbs, J.M., Bird, T.H., Bauer, C.E. and Tabita, F.R. (2000)
Interaction of CbbR and RegA* transcription regulators with the
Rhodobacter sphaeroides cbbI promoter-operator region. J. Biol.
Chem. 275, 19224–19230.
[34] O’Hara, G.W., Goss, T.J., Dilworth, M.J. and Glenn, A.R. (1989)
Maintenance of intracellular pH and acid-tolerance in Rhizobium
meliloti. Appl. Environ. Microbiol. 55, 1870–1876.
[35] Miller, J.H. (1972) In: Experiments in Molecular Biology, Vol.
Cold Spring Harbor Laboratory, Cold Spring Harbor, NY.
[36] Reeve, W.G., Tiwari, R.P., Worsley, P.S., Dilworth, M.J., Glenn,
A.R. and Howieson, J.G. (1999) Constructs for insertional
mutagenesis, transcriptional signal localization and gene regula-
tion studies in root nodule and other bacteria. Microbiology 145,
1307–1316.
[37] Reeve, W.G., Tiwari, R.P., Dilworth, M.J. and Glenn, A.R.
(1997) A helicase gene (helO) in Rhizobium meliloti WSM419.
FEMS Microbiol. Lett. 153, 43–49.
[38] Wilson, K.J., Sessitsch, A., Corbo, J.C., Giller, K.E., Akkermans,
A.D. and Jefferson, R.A. (1995) b-Glucuronidase (GUS) trans-
posons for ecological and genetic studies of rhizobia and other
Gram-negative bacteria. Microbiology 141, 1691–1705.
[39] Galibert, F., Finan, T.M., Long, S.R., Puhler, A., Abola, P.,
Ampe, F., Barloy-Hubler, F., Barnett, M.J., Becker, A., Boistard,
P., Bothe, G., Boutry, M., Bowser, L., Buhrmester, J., Cadieu, E.,
Capela, D., Chain, P., Cowie, A., Davis, R.W., Dreano, S.,
Federspiel, N.A., Fisher, R.F., Gloux, S., Godrie, T., Goffeau, A.,
Golding, B., Gouzy, J., Gurjal, M., Hernandez-Lucas, I., Hong,
A., Huizar, L., Hyman, R.W., Jones, T., Kahn, D., Kahn, M.L.,
Kalman, S., Keating, D.H., Kiss, E., Komp, C., Lelaure, V.,
Masuy, D., Palm, C., Peck, M.C., Pohl, T.M., Portetelle, D.,
Purnelle, B., Ramsperger, U., Surzycki, R., Thebault, P., Van-
denbol, M., Vorholter, F.J., Weidner, S., Wells, D.H., Wong, K.,
Yeh, K.C. and Batut, J. (2001) The composite genome of the
legume symbiont Sinorhizobium meliloti. Science 293,
668–672.
[40] Reeve, W.G., Tiwari, R.P., Kale, N.B., Dilworth, M.J. and Glenn,
A.R. (2002) ActP controls copper homeostasis in Rhizobium
leguminosarum bv. viciae and Sinorhizobium meliloti preventing
low pH-induced copper toxicity. Mol. Microbiol. 43, 981–991.
[41] David, M., Daveran, M.L., Batut, J., Dedieu, A., Domergue, O.,
Ghai, J., Hertig, C., Boistard, P. and Kahn, D. (1988) Cascade
regulation of nif gene expression in Rhizobium meliloti. Cell 54,
671–683.
[42] Chen, C.Y. and Winans, S.C. (1991) Controlled expression of the
transcriptional activator gene virG in Agrobacterium tumefaciens
by using the Escherichia coli lac promoter. J. Bacteriol. 173, 1139–
1144.
[43] Fischer, H.M. (1994) Genetic regulation of nitrogen fixation in
rhizobia. Microbiol. Rev. 58, 352–386.
[44] Batut, J., Daveran-Mingot, M.L., David, M., Jacobs, J., Garn-
erone, A.M. and Kahn, D. (1989) fixK, a gene homologous with
fnr and crp from Escherichia coli, regulates nitrogen fixation genes
both positively and negatively in Rhizobium meliloti. EMBO J. 8,
1279–1286.
[45] Ditta, G., Virts, E., Palomares, A. and Kim, C.H. (1987) The nifA
gene of Rhizobium meliloti is oxygen regulated. J. Bacteriol. 169,
3217–3223.
[46] Swem, D.L. and Bauer, C.E. (2002) Coordination of ubiquinol
oxidase and cytochrome cbb3 oxidase expression by multiple
regulators in Rhodobacter capsulatus. J. Bacteriol. 184,
2815–2820.
[47] Comolli, J.C., Carl, A.J., Hall, C. and Donohue, T. (2002)
Transcriptional activation of the Rhodobacter sphaeroides cyto-
chrome c2 gene P2 promoter by the response regulator PrrA. J.
Bacteriol. 184, 390–399.
[48] Fischer, H.M., Sciotti, H. and Hennecke, H. (2001) The network
controlling symbiotic nitrogen fixation genes in Bradyrhizobium
japonicum. In: Proceedings of the 13th International Congress on
Nitrogen Fixation, CABI Publishing, Hamilton, Ontario (Finan,
T.M., O’Brian, M., Layzell, D.B., Vessey, K., Newton, W., Eds.),
Vol., pp. 213–217.
[49] Tichi, M.A. and Tabita, F.R. (2001) Interactive control of
Rhodobacter capsulatus redox-balancing systems during photo-
trophic metabolism. J. Bacteriol. 183, 6344–6354.
[50] Oh, J.I., Ko, I.J. and Kaplan, S. (2001) The default state of the
membrane-localized histidine kinase PrrB of Rhodobacter sph-
aeroides 2.4.1 is in the kinase-positive mode. J. Bacteriol. 183,
6807–6814.
[51] Oh, J.I. and Kaplan, S. (2000) Redox signaling: globalization of
gene expression. EMBO J. 19, 4237–4247.
[52] Georgellis, D., Kwon, O. and Lin, E.C. (2001) Quinones as the
redox signal for the arc two-component system of bacteria.
Science 292, 2314–2316.
[53] King, P.W. and Przybyla, A.E. (1999) Response of hya expression
to external pH in Escherichia coli. J. Bacteriol. 181, 5250–5256.
[54] Poole, R.K. and Cook, G.M. (2000) Redundancy of aerobic
respiratory chains in bacteria. Routes, reasons and regulation.
Adv. Microb. Physiol. 43, 165–224.
[55] Toledo-Cuevas, M., Barquera, B., Gennis, R.B., Wikstrom, M.
and Garcia-Horsman, J.A. (1998) The cbb3-type cytochrome c
oxidase from Rhodobacter sphaeroides, a proton-pumping heme-
copper oxidase. Biochim. Biophys. Acta 1365, 421–434.
[56] Arslan, E., Kannt, A., Thony-Meyer, L. and Hennecke, H. (2000)
The symbiotically essential cbb3-type oxidase of Bradyrhizobium
japonicum is a proton pump. FEBS Lett. 470, 7–10.
[57] Endley, S., McMurray, D. and Ficht, T.A. (2001) Interruption of
the cydB locus in Brucella abortus attenuates intracellular survival
and virulence in the mouse model of infection. J. Bacteriol. 183,
2454–2462.
[58] Metcalf, W.W., Jiang, W., Daniels, L.L., Kim, S.K., Haldimann,
A. and Wanner, B.L. (1996) Conditionally replicative and
conjugative plasmids carrying lacZa for cloning, mutagenesis,
and allele replacement in bacteria. Plasmid 35, 1–13.
[59] Hanahan, D. (1983) Studies on transformation of Escherichia coli
with plasmids. J. Mol. Biol. 166, 557–580.
[60] Meade, H.M., Long, S.R., Ruvkun, G.B., Brown, S.E. and
Ausubel, F.M. (1982) Physical and genetic characterization of
symbiotic and auxotrophic mutants of Rhizobium meliloti induced
by transposon Tn5 mutagenesis. J. Bacteriol. 149, 114–122.
[61] De Philip, P., Batut, J. and Boistard, P. (1990) Rhizobium meliloti
FixL is an oxygen sensor and regulates R. meliloti nifA and
fixK genes differently in Escherichia coli. J. Bacteriol. 172,
4255–4262.
[62] Quandt, J. and Hynes, M.F. (1993) Versatile suicide vectors which
allow direct selection for gene replacement in gram-negative
bacteria. Gene 127, 15–21.
[63] Hirsch, P.R. and Beringer, J.E. (1984) A physical map of pPH1JI
and pJB4JI. Plasmid 12, 139–141.
B.J. Fenner et al. / FEMS Microbiology Letters 236 (2004) 21–31 31
[64] Spaink, H.P., Robert, J.H., Okker, C.A., Wijffelman, E.P. and
Lugtenberg, B.J.J. (1987) Promoters in the nodulation region of
the Rhizobium leguminosarum Sym plasmid pRL1JI. Plant Mol.
Biol. 9, 27–39.
[65] Altschul, S.F., Madden, T.L., Schaffer, A.A., Zhang, J., Zhang,
Z., Miller, W. and Lipman, D.J. (1997) Gapped BLAST and PSI-
BLAST: a new generation of protein database search programs.
Nucleic Acids Res. 25, 3389–3402.