sine wave extremely low frequency magnetic fields protect chick embryos against uv-induced death
TRANSCRIPT

ELECTROMAGNETIC BIOLOGY AND MEDICINE
Vol. 23, No. 2, pp. 113–124, 2004
Sine Wave Extremely Low Frequency Magnetic
Fields Protect Chick Embryos Against
UV-Induced Death
Anna-Maria Bjorasen,1 Anna Sjodin,1 Kjell Hansson Mild,1,2
and Mats-Olof Mattsson1,*
1Department of Natural Sciences, Orebro University, Orebro, Sweden2National Institute for Working Life, Umea, Sweden
ABSTRACT
Four-day-old chicken embryos were exposed to extremely low frequency (ELF)
magnetic fields (MF) prior to UV exposure (75min, predominantly UV-C,
0.4mW/cm2) to investigate possible MF-mediated protection against lethal effects
of UV. The UV exposure typically resulted in a 20% survival rate (as judged by
beating hearts) in sham-exposed embryos 3 h postexposure. In contrast, exposure
to a 50 (10, 50, or 100 mT) or 60Hz (10mT) vertical MF caused a significant
increase in survival rate, observed only 30min after UV exposure. No difference
in protection levels was seen between these exposure intensities. A horizontal
50Hz MF (10, 50, or 100 mT) did not result in the general protection against
UV-induced death observed for vertical fields, suggesting that the size of the
induced electric field (which differs between horizontal and vertical exposure) is
important for the MF-induced protection. To explore the molecular mechanisms
involved in this effect, immunoblotting experiments with an antibody against the
inducible form of hsp70 were performed. These showed that application of MF
(50Hz, 200mT, 1 h) induced hsp70 expression in human K562 cells.
*Correspondence: Dr. Mats-Olof Mattsson, Department of Natural Sciences, Fakultetsgatan
1, Orebro University, S-70182 Orebro, Sweden; Fax: +46 19 303566; E-mail: mats-olof.
113
DOI: 10.1081/LEBM-200037573 1536-8378 (Print); 1536-8386 (Online)
Copyright & 2004 by Marcel Dekker, Inc. www.dekker.com

Key Words: ELF magnetic field; Chicken embryo; UV-protection; hsp70.
INTRODUCTION
Recently, a number of reports from a single laboratory showed protective effectsby extremely low frequency (ELF) magnetic fields (MF) on the lethal outcomeof several stressors on chick embryos. The experiments included a short MFexposure prior to the stressor (hypoxia, UV irradiation) and subsequent observa-tions of the presence of heart beat in the embryo (Di Carlo et al., 1998, 1999).Irrespective of the stressor, the MF exposure seemed to induce a stress response,which at least for some time provided protection against otherwise lethalenvironmental factors. If correct, these observations imply that use of at leastcertain ELF MFs, at comparably low field strengths, can have beneficial effects.
Pure biological effects by MFs on cells and animals have frequently beenreported, although their physiological implications as well the physical andbiological interaction mechanisms are unclear. Health effects, both negative(e.g., childhood leukemia; Portier and Wolfe, 1998) and positive (bone healing;Quittan et al., 2000) are associated with MF exposure. However, the mentionedchick embryo experiments, elegant in their simplicity, show promise for under-standing several fundamental questions regarding biological effects of MF. Sincethey are performed on organisms rather than on cells, the model can be used formolecular analyses in a physiological setting, without performing expensive andcumbersome experiments on mammals. The chick is a vertebrate, and thus a suitablygood model for more medically relevant experiments on embryo development. Thevarious aspects of what constitutes dose in MF exposure can also be addressed,including experiments on time dependence, frequency effects, possible dose–responserelationships between field strength and experimental outcome, the interdependenceof time and flux density, etc. This depends, however, on whether the original findingsby Di Carlo et al. (1998, 1999) are replicable in other laboratories.
The present study was undertaken to see first if reported protection by a 60Hzsine wave MF on UV-induced death in 96-h-old chick embryos are reproducible inanother laboratory, with a different flock of hens with a different genetic makeup.Second, possible frequency dependence was tested, as well as effects of various fluxdensities and of different polarizations of the applied field. The outcome of theeffects is discussed in terms of possible molecular mechanisms involved in theinduced MF protection.
MATERIALS AND METHODS
Embryos and Embryo Handling
Fertilized hen eggs (White Leghorn, inbred since 1981) were obtained from OvaProduction AB (Morgongava, Sweden). Prior to any experiments, freshly laid eggs(delivered from breeder to laboratory by van) were stored at 18�C, for 1 or 2 days.Subsequently, the eggs were incubated vertically with pointed ends down at 37.7�C in
114 Bjorasen et al.

a Masalles egg incubator for 96 h. The development corresponded to Hamburgerand Hamilton stage 24 (Hamburger and Hamilton, 1992). On average, each eggweighed 60.2 g. The AC magnetic field in the egg incubator was 0.5 mT rms or lower.
Magnetic Field Exposure
All experiments were performed in a matched pair of conventional cell cultureincubators (Forma Scientific model 3336 CO2 incubator), in normal air at constanttemperature (37.6�C) and humidity (75%). The AC magnetic field in the incubatorswas created by a double Helmholtz coil arrangement (made in-house), with twohorizontal and two vertical coils (ø 40 and 42 cm, respectively). The coils were woundwith 30 turns each of a twisted set of three Litz threads, where the twist was aboutthree turns per meter of length of the thread. Thus each coil had three separatewindings, each with 30 turns. Two of the windings were used for the AC current andone for DC current (this option was not used in this study). The two AC windingswere coupled through a switch, which allowed us to control the current in the twowindings as either parallel or anti-parallel. The latter case then served as a ‘‘sham’’exposure, since the AC fields canceled each other. The AC coils were fed froma signal generator (Kenwood FG-273 or Metrix Gx240) and an amplifier (SentecPA9), through a 4 Ohm resistor (Arcol). The current and thus the B-field strength(all flux density values given are rms values) were constantly monitored witha commercial multimeter (Unimer 45). Furthermore, a handheld instrument(a triaxial ELF microteslameter, MFD-III, SMPS Electronics) was used to measurethe 50 or 60Hz magnetic field within the exposure chamber before and after eachexperimental run. The ambient DC field within the incubators was found to be 30.5and 41.8 mT, respectively, with a predominantly vertical inclination (field vectorsmeasured in three dimensions with a mMAG handheld digital magnetometer,MEDA Inc).
Both incubators served as ‘‘sham’’ or ‘‘exposure’’ incubator in a randomfashion. The background ELF magnetic field during sham conditions was measuredbefore and after each experiment and was 0.5 mT rms or lower.
UV Exposure
UV irradiation was obtained by employing an UV-C source (Phillips TUV 30W/630T8, The Netherlands), which emits predominantly UV-C (i.e., 200–290 nm;as measured by an Optronic model 754-0-PMT spectroradiometer; see Fig. 1 forspectral analysis). Embryos typically received 0.4mW/cm2 at 254 nm. Exposure tookplace at room temperature (25�C) for 75min.
Experimental Design
During each experiment, 60 eggs were randomly divided into sham or exposuregroups (30 eggs each). Experiments began with placing the eggs within the
Magnetic Fields and UV-Protection 115

incubators (sham or exposure, unknown to the operator) for 20min. The eggs werethen maintained for 1 h at room temperature (to allow for response systems to beactivated). During this time, the operator ‘‘windowed’’ the eggs (removal of the shelland outer and inner shell membranes at the top of the egg, ca 40 s per egg), anddiscarded undeveloped, damaged, or dead embryos. Subsequently, embryos wereUV-irradiated for 75min after which vitality (as judged by the beating heart) waschecked every 30min beginning immediately after UV treatment. Whether aparticular embryo was sham or magnetic field-exposed was not known by theoperator during the procedure.
Protein Preparation and Immunoblotting
Embryos, or cultured cells (human K562 chronic myelogenous leukemia cells;ATCC #CCL-243, ATCC, Rockville, MD) that served as control material, werehomogenized with a pipette in ice-cold RIPA buffer (1% Igepal, 0.5%Na-deoxycholate, 0.1% SDS [sodium dodecyl sulphate], dissolved in PBS, pH 7.4)with 1% PMSF addded, incubated on ice for 30min, and further homogenized in aglass homogenizer with additional PMSF (1% final concentration) added. After30min incubation on ice, the homogenate was centrifuged (10,000 g, 10min). Thesupernatant was recovered and stored at �20�C until further analysis. A total of20-mg protein (as determined by the method of Bradford, 1976, with bovine serumalbumin as standard, Bio-Rad) was dissolved in 4x SDS electrophoresis buffer with2% beta-mercaptoethanol and boiled (microwave oven, 850W, 2–3min), andsubsequently separated by SDS-polyacrylamide gel electrophoresis (200V, 35min)together with a molecular weight marker (Kaleidoscope, Bio-Rad) and a positivecontrol for hsp70 expression (heat shocked HeLa cell extract, Santa CruzBiotechnology Inc.). Prior to transfer, the gels were equilibrated in buffer and themembranes (Hybond N, Amersham) were handled according to the manufacturer’sinstructions. Subsequently, the gels were transferred by electroblotting (Trans Blot
Figure 1. Spectral analysis of the UV radiation used during exposure of 96-h chicken
embryos. For details, see Materials and Methods.
116 Bjorasen et al.

unit, Bio-Rad) in Towbin buffer (25mM Tris, 192mM glycine, 20% vol/volmethanol) for 1 h at 100V. The membranes were blocked (30min) in TBS-T buffer(20mM Tris-Cl, 137mM NaCl, 0.1% Tween 20, pH 7.6) with 5% milk powder andstained with antibodies against hsp70 (mouse anti-human W27 antibody, Santa CruzBiotechnology Inc., diluted 1:500; or mouse anti-human SPA-810 antibody,Stressgen Biotechnologies [Victoria, Canada], diluted 1:1000) for 45min. Afterwashing, the membranes were incubated with a secondary antibody (goat anti-mouseIgG2a - HRP antibody, Vector) diluted 1:1000. The formed complex was finallydetected by means of Vector Elite DAB-kit, according to the manufacturer’sinstructions. All chemicals were obtained from Sigma unless otherwise stated.
Statistical Analysis
For each experimental run, the percentage surviving embryos at each time pointafter UV exposure was calculated. Average and standard error for 4–6 runs weredetermined, after which the two conditions were analyzed by chi-squared tests forsignificant differences. All data handling and statistical calculations were performedwith a Microsoft Excel software package.
RESULTS
Initial experiments were performed to ascertain that the described windowingprocedure, and the application of MF exposure prior to windowing and subsequentobservation in itself did not influence viability of embryos. Observations were doneup to 5 h after windowing, with or without prior MF exposure, and no dead embryosappeared to be due to this treatment (data not shown). Subsequently, pilot studieswere performed to determine at what UV irradiation intensity we would find that50% or less of embryos survived 3 h postexposure. This level was seen to differbetween flocks of hens, ranging from 0.4 mW/cm2 in the first flock to >0.8mW/cm2
in a second flock. Experiments with MF and UV were then performed from June toAugust 2002, with the two different flocks.
We documented a protective effect from a vertical 60Hz, 10 mT, MF appliedfor 20min prior to UV exposure (Table 1). A 50% survival rate after 3 h (n¼ 50)
Table 1. Magnetic field effect on chick embryo survival after UV exposure. Survival rate was
monitored among embryos exposed to UV radiation at 0.4mW/cm2, pretreated (exposed) or
not (sham) with a vertical, 60Hz, 10 mT magnetic field for 20min. Survival was monitored at
noted times after UV exposure. Results are given as mean from two separate experiments.
(n¼ 50 in the exposed group; n¼ 46 in the sham-exposed group).
Time (min) 0 30 60 90 120 150 180
Survival of exposed embryos (%) 100 96 88 82 70 61 50
Survival of sham embryos (%) 95 81 66 53 37 29 18
Magnetic Fields and UV-Protection 117

was seen in the exposed group, whereas only 18% (n¼ 46) survived amongsham-exposed embryos.
Effects of a vertical 50Hz MF were studied at three different flux densities: 10,50, and 100 mT (Fig. 2a–c). At all exposure levels, survival among exposed embryossignificantly differed from sham-exposed. The difference was apparent only 30minpostUV, increasing until the last observation time (3 hr postUV). However, no dose–response relationship was seen, all three intensity levels causing comparable levelsof protection.
In contrast, exposure to a horizontal 50-Hz MF, at the same flux densities, didnot provide the same general protection against the UV radiation (Fig. 3a–c). Onlyat 10-mT, after 2.5 h, was significant protection observed. These experiments wereperformed with embryos from two flocks of hens, where the 10-mT study wasperformed on one flock, and embryos from a different flock was used for theremaining runs. Notably, to obtain the desired stressful effects of UV, we hadto increase UV levels for the second flock, as mentioned previously.
As a final test, embryos from the second flock were also exposed to the vertical10-mT field, to see if our initial observations could be replicated. Once again,a protective effect of the MF treatment was seen, although the levels differedsomewhat compared with the initial experiments (41% survival among exposed 3 hafter UV exposure (n¼ 53), 12% among sham-exposed (n¼ 45).
To investigate the possible induction of heat shock protein 70 (hsp70) as aconsequence of MF-induced protection against UV effects, immunoblotting wasperformed on material obtained from human K562 cells (as controls) and fromMF-exposed chicken embryos. The cells responded both to traditional heat shock(42�C, 1 h, and subsequent culture at 37�C for 4 or 24 h) and to 50Hz, 200 mT,vertical MF (20min, followed by 4 or 24 h at 37�C in sham condition) witha marked increase in hsp70 levels (Fig. 4a). However, no hsp70 expression was seenin the chicken material, irrespective of treatment conditions [heat shock asmentioned above or MF exposure (50Hz, vertical, 10 mT, 20min or 200 mT, 1 h)(Fig. 4b)]. These experiments were repeated several times, and with two differentanti-hsp70 antibodies. In no single experiment could we find any trace of hsp70expression in the chicken material.
DISCUSSION
The main observation in the present study is that a short-term exposure to MF,at 50 or 60Hz, can protect chicken embryos against detrimental effects of subsequentUV exposure. This is furthermore a confirmation of the results reported by Di Carloet al. (1998, 1999, 2000), who originally performed similar studies with 60Hz MF.Their observations reported that UV protection is obtained at 8-mT rms, comparableto our setting of 10 mT. Due to circumstances beyond our control, the egg supplierchanged flocks of hens between a series of experiments. This forced us to employ UVexposure at higher intensities to obtain a desired fraction of dead embryos after a 3-hobservation period. The sensitivity to UV thus differs between biological specimens,probably indicating that the sensitivity to other environmental factors also differs.Indeed, our later experiments revealed a consistent protection by the applied MF,
118 Bjorasen et al.
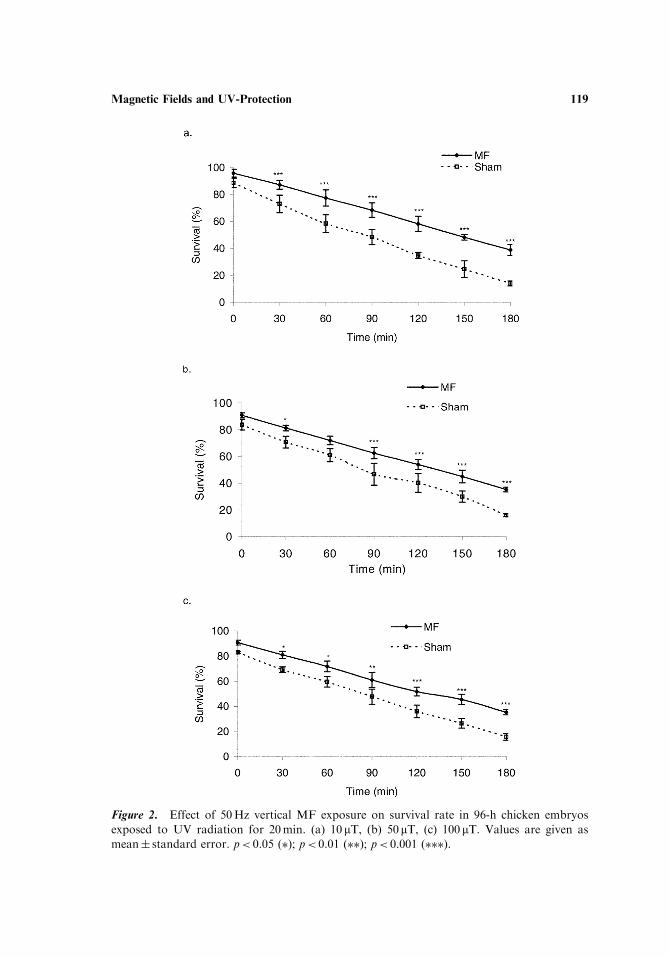
Figure 2. Effect of 50Hz vertical MF exposure on survival rate in 96-h chicken embryos
exposed to UV radiation for 20min. (a) 10mT, (b) 50mT, (c) 100 mT. Values are given as
mean� standard error. p< 0.05 (�); p< 0.01 (��); p< 0.001 (���).
Magnetic Fields and UV-Protection 119
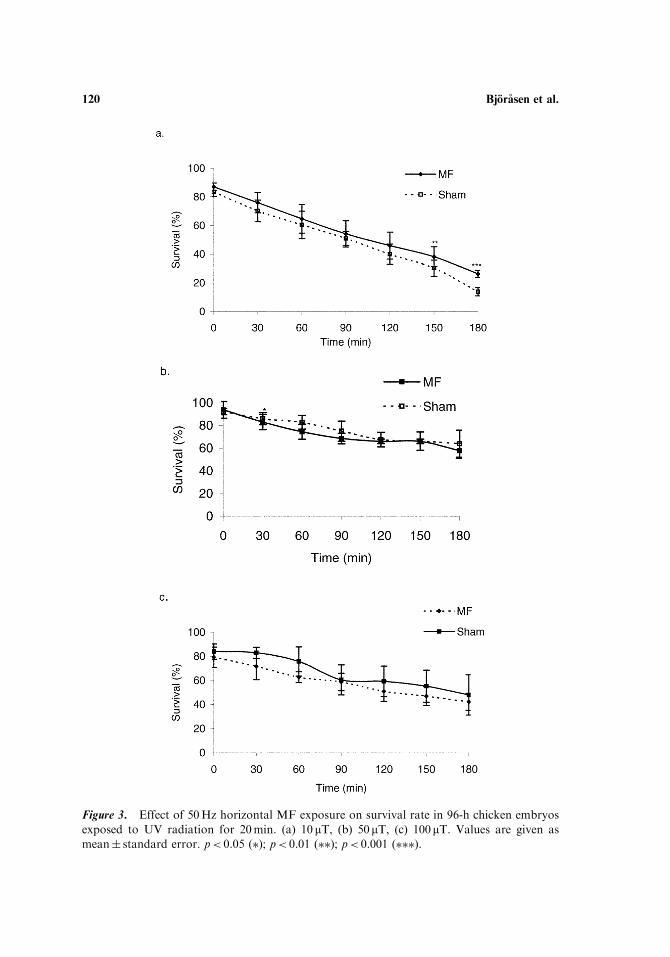
Figure 3. Effect of 50Hz horizontal MF exposure on survival rate in 96-h chicken embryos
exposed to UV radiation for 20min. (a) 10 mT, (b) 50mT, (c) 100mT. Values are given as
mean� standard error. p< 0.05 (�); p< 0.01 (��); p< 0.001 (���).
120 Bjorasen et al.

although at a different level than in earlier experiments. That genetic makeupconstitutes an important parameter in bio-effect research in general is presentlyaccepted, and also substantiated in bioelectromagnetics research. This was thusobserved in 1999, when Di Carlo et al. studied survival in two flocks of henssubjected to hypoxia with or without prior MF treatment. The survival rate to agiven MF exposure differed considerably between the two flocks. Recently, a similarobservation was presented by Fedrowitz and Loscher (2002), who employed twodifferent substrains of Sprague-Dawley rats. These substrains were seen to responddifferently to the carcinogen DMBA, as well as to 50-Hz MF.
Vertical 50-Hz MF exposure provides similar degrees of protection against UVeffects irrespective of flux densities (10, 50, or 100 mT). Still, there are a substantialnumber of dead embryos also in the MF exposed group. The nature of the protectionis unknown, but one can envisage that the MF levels triggered the protection systemin the same manner at 10 as at 100 mT. The used range of flux densities might thenrepresent a plateau phase in the response curve. According to Di Carlo et al. (1998),protection by 60-Hz MF against hypoxia was seen at 3 mT and higher, suggesting athreshold for the protective effect to occur close to that figure. If such a comparablylow threshold would be present in this study is however not investigated. Wouldhigher levels than 100 mT confer better protection (the ultimate protection being thatall embryos survive UV treatment)? Not necessarily, since other systems couldbe triggered, that in themselves are less beneficial to the embryos, or the increasedputative production of protecting molecules could be more damaging at higherlevels. Obviously, further experiments into the importance of flux density levels areneeded.
Interestingly, application of a horizontal, rather than vertical MF promotes onlya weak, or nonexistent, protection against UV. At 10 mT, there is a small butsignificant difference between exposed and sham-exposed embryos 2.5 and 3 h afterUV exposure, but not earlier. A 50-mTMF could not protect the embryos against theUV, also seen when 100-mT MF was applied. Rather, the horizontal exposure
Figure 4. Immunoblots of representative experiments on (a) K562 cells and (b) 96-h old
chicken embryo material analyzed with an antibody (W27) against human hsp70. C¼ control
(sham) material; MF¼magnetic field exposed (200 mT, 1 h, followed by postexposure for
given time); HS¼ heat shock treatment (42�C, 1 h, followed by posttreatment for given time);
HeLa¼ extract from HeLa cells treated with heat shock (positive control).
Magnetic Fields and UV-Protection 121

seemed to aggravate the damaging effect of UV at these levels. When comparing thetwo directions of the MF, one must be aware that the applied MF will inducedifferently sized electric fields. Due to the configuration of the egg, a horizontal MF,which in this study will be perpendicular to the main axis of the egg, gives rise toa larger induced electric field than an MF parallel to the egg axis (vertical exposure).The importance of the size of the induced E-field has also been documented in earlierstudies (Valtersson et al., 1997), who studied ODC activity in Jurkat cells. There isthus a possibility that the primary reason for this effect is the induced electric fieldwhen it is above or below a certain threshold. However, additional studies areneeded to explore why there seems to be a difference between these two kindsof exposure.
What is then the mechanism by which MF exposure protects against UVradiation? DNA damage is known to occur as a consequence of the DNA moleculeabsorbing photon energy directly for wavelengths below 320 nm, i.e., in the UV partof the spectrum (Ravanat et al., 2001). This is however probably not the cause ofdeath in the UV-exposed chicken embryos in this study, since such an effect on DNAwould appear much later than the observation time (0–3 h postUV) employed hereallows. On the other hand, UV exposure leads to formation of reactive oxygenspecies (ROS), which in turn affect vital cellular constituents (Meewes et al., 2001),that can be lethal to the organism. ELF MF’s at field strengths comparable to theones used in this study have also been seen to stimulate free radical production(Katsir and Parola, 1998; Simko et al., 2001). Hypothetically, the stress exerted bythe MFs is inducing a general stress response, which then to some extent is functionalalready when the second stressor, UV, is applied to the embryos (‘‘cross-protection’’is initiated). A well-studied and general stress response system is the heat shockprotein (hsp) system, which is induced by a plethora of environmental factors(Latchman, 2001). Among these proteins, the inducible form of hsp70 has receivedconsiderable attention, due to its versatile role in protection against a multitudeof agents. Interestingly, this molecule has been shown to appear as a function of ELFMF exposure, both in cells (Pipkin et al., 1999), and in chick embryos (Di Carloet al., 2002). Studies of hsp70 mRNA expression and hsp70 promoter studies alsosuggest that power frequency MFs have a general effect on the cells stress responsesystems (reviewed in Goodman and Blank, 2002). However, it remains to analyzespecific molecular mechanisms behind the MF-induced UV protection in the chickembryo system. The experiments performed in this study with antibodies against theinducible form of hsp70 were inconclusive. Interestingly, MF exposure indeedproduced higher levels of hsp70 in human cells, comparable with heat shocktreatment, supporting the notion of a general stress effect of MF. Unfortunately, theused antibodies and protocols failed to detect any hsp70 expression whatsoeverin the chicken. Obviously, this can be due to unsuitable antibodies, unable to detectthe avian form of hsp70, or other, unknown, experimental errors. The issue isimportant and further experiments are underway to solve the issue.
The chick embryo system is well suited for studies of a neglected issue inbioelectromagnetics research, i.e., the question of dose. We are presently more or lessignorant about what constitutes dose in this context. The model is also appropriatefor studies of molecular constituents of biological interaction mechanisms, of whichwe will report in the near future.
122 Bjorasen et al.

REFERENCES
Di Carlo, A. L., Litovitz, T. A. (1999). Is genetics the unrecognized confounding
factor in bioelectromagnetics? Flock-dependence of field-induced anoxia
protection in chick embryos. Bioelectrochem. Bioener. 48:209–215.Di Carlo, A. L., Farrell, J. M., Litovitz, T. A. (1998). A simple experiment to study
electromagnetic field effects: protection induced by short-term exposures to
60 hz magnetic fields. Bioelectromagnetics 19:498–500.Di Carlo, A. L., Mullins, J. M., Litovitz, T. A. (2000). Thresholds for
electromagnetic field-induced hypoxia protection: evidence for a primary
electric field effect. Bioelectrochemistry 52:9–16.Di Carlo, A. L., Hargis, M. T., Penafiel, M. L., Litovitz, T. A. (1999). Short-term
magnetic field exposures (60 hz) induce protection against ultraviolet radiation
damage. Int. J. Rad. Biol. 75(12):1541–1549.Di Carlo, A. L., White, N., Fuling, G., Garrett, P., Litovitz, T. A. (2002).
Chronic electromagnetic field exposure decreases hsp70 levels and lowers
cytoprotection. J. Cell. Biochem. 84:447–454.Fedrowitz, M., Loscher, W. (2002). Effects of magnetic field exposure in the DMBA
model of breast cancer in different substrains of Sprague-Dawley rats.
Abstract, 24th Annual Meeting of the Bioelectromagnetics Society.
Quebec City, Quebec, Canada, June 23–27, 2002. Washington, DC:
Bioelectromagnetics Society, BEMS 5–6.Goodman, R., Blank, M. (2002). Insight into electromagnetic interaction
mechanisms. J. Cell. Physiol. 192:16–22.Hamburger, V., Hamilton, H. L. (1992). A series of normal stages in the
development of the chick embryo. Dev. Dyn. 195:231–272.Katsir, G., Parola, A. H. (1998). Enhanced proliferation caused by a low frequency
weak magnetic field in chick embryo fibroblasts is suppressed by radical
scavengers. Biochem. Biophys. Res. Commun. 252(3):753–756.Latchman, D. S. (2001). Heatshock proteins and cardial protection. Cardiovasc. Res.
51:637–646.Meewes, C., Brenneisen, P., Wenk, J., Kuhr, L., Wenjian, M., Alikoski, A., Poswig,
A., Krieg, T., Sharfetter-Kochanek, K. (2001). Adaptive antioxidant response
protects dermal fibroblasts from uva-induced phototoxicity. Free radic. Biol. &
Med. 30(3):238–247.Pipkin, J. L., Hinson, W. G., Young, J. F., Rowland, J. G., Shaddock, W. H.,
Tolleson, W. H., Duffy, P. H., Casciano, D. A. (1999). Induction of stress
proteins by electromagnetic fields in cultured HL-60 cells. Bioelectromagnetics
20:347–357.Portier, C. J., Wolfe, M. S., eds. (1998). Assessment of Health Effects from Exposure
to Power-Line Frequency Electric and Magnetic Fields, NIH Publication
No. 98–3981; National Institute of Environmental Health Sciences, U.S.
National Institutes of Health: Research Triangle Park.Quittan, M., Schufried, O., Wiesinger, G. F., Fialka-Moser, V. (2000). Clinical
effectiveness of magnetic field therapy—a review of the literature. Acta Med.
Austriaca 27(3):61–68.
Magnetic Fields and UV-Protection 123

Ravanat, J.-L., Douki, T., Cadet, J. (2001). Direct and indirect effects of UVradiation on DNA and its components. J. Photochem. Photobiol. 63:88–102.
Simko, M., Droste, S., Kreihuber, R., Weiss, D. G. (2001). Stimulation ofphagocytosis and free radical production in murine macrophages by 50Hzelectromagnetic fields. Eur. J. Cell. Biol. 80(8):562–566.
Valtersson, U., Mild, K. H., Mattsson, M.-O. (1997). Ornithine decarboxylaseactivity and polyamine levels are different in Jurkat and CEM-CM3 cells afterexposure to a 50Hz magnetic field. Bioelectrochem. Bioenerg. 43:169–172.
124 Bjorasen et al.