silencing the expression of mitochondrial acyl-coa thioesterase i and acyl-coa synthetase 4 inhibits...
TRANSCRIPT

Silencing the expression of mitochondrial acyl-CoAthioesterase I and acyl-CoA synthetase 4 inhibitshormone-induced steroidogenesisPaula Maloberti*, Rocıo Castilla*, Fernanda Castillo, Fabiana Cornejo Maciel, Carlos F. Mendez,Cristina Paz and Ernesto J. Podesta
Department of Biochemistry, School of Medicine, University of Buenos Aires, Argentina
Steroid hormones are synthesized in specialized steroido-
genic cells in the adrenal gland, ovary, testis, placenta
and brain and are essential for maintaining normal
body homeostasis and reproductive capacity. The bio-
synthesis of all steroid hormones begins at the mito-
chondria with the conversion of cholesterol into
pregnenolone by the cholesterol side-chain cleavage
cytochrome P-450 enzyme (P450scc) [1,2]. The trans-
port of cholesterol to the inner mitochondrial mem-
brane and the availability of cholesterol for P450scc
constitutes the rate-limiting step of steroidogenesis, a
step controlled by the steroidogenic acute regulatory
protein (StAR) [3,4] and the peripheral benzodiazepine
receptor (PBR) [5,6].
Arachidonic acid (AA) can act as a signaling
messenger itself or through its metabolites exerting
numerous effects on different cellular processes [7–9].
Several reports have shown that AA and lipoxyge-
nase metabolites play an essential role in the regula-
tion of steroidogenesis. In steroidogenic cells, AA
participates in hormone-stimulated induction of StAR
expression [10,11]. However, the manner in which
Keywords
ACS4; acyl-CoA; arachidonic acid;
mitochondrial acyl-CoA thioesterase I
(MTE-I); steroidogenesis
Correspondence
E. J. Podesta, Depto. de Bioquımica,
Facultad de Medicina, Paraguay 2155,
piso 5, C1121ABG Buenos Aires, Argentina
Tel ⁄ Fax: +54 11 4508 3672, ext. 31
E-mail: [email protected]
*Note
These authors contributed equally to this
work
(Received 27 October 2004, revised 9
February 2005, accepted 16 February 2005)
doi:10.1111/j.1742-4658.2005.04616.x
Arachidonic acid and its lypoxygenated metabolites play a fundamental
role in the hormonal regulation of steroidogenesis. Reduction in the expres-
sion of the mitochondrial acyl-CoA thioesterase (MTE-I) by antisense or
small interfering RNA (siRNA) and of the arachidonic acid-preferring
acyl-CoA synthetase (ACS4) by siRNA produced a marked reduction in
steroid output of cAMP-stimulated Leydig cells. This effect was blunted
by a permeable analog of cholesterol that bypasses the rate-limiting step in
steroidogenesis, the transport of cholesterol from the outer to the inner
mitochondrial membrane. The inhibition of steroidogenesis was overcome
by addition of exogenous arachidonic acid, indicating that the enzymes are
part of the mechanism responsible for arachidonic acid release involved in
steroidogenesis. Knocking down the expression of MTE-I leads to a signifi-
cant reduction in the expression of steroidogenic acute regulatory protein.
This protein is induced by arachidonic acid and controls the rate-limiting
step. Overexpression of MTE-I resulted in an increase in cAMP-induced
steroidogenesis. In summary, our results demonstrate a critical role for
ACS4 and MTE-I in the hormonal regulation of steroidogenesis as a new
pathway of arachidonic acid release different from the classical phospho-
lipase A2 cascade.
Abbreviations
AA, arachidonic acid; ACS4, arachidonic acid-preferring acyl-CoA synthetase 4; ACTH, adrenocorticotrophin; 8Br-cAMP, 8-bromo-3¢,5¢-cAMP;
22(R)-OH-cholesterol, 22a-hydroxycholesterol; DAPI, 4¢,6-diamidino-2-phenylindole; DBI, diazepam binding inhibitor; EGFP, enhanced green
fluorescent protein; MTE-I, mitochondrial acyl-CoA thioesterase I; MTT, 3-(4,5-dimethyl-2-thiazolyl)-2,5-diphenyl-2H-tetrazolium bromide;
P450scc, cholesterol side-chain cleavage cytochrome P-450 enzyme; PBR, peripheral benzodiazepine receptor; siRNA, small interfering RNA;
StAR protein, steroidogenic acute regulatory protein.
1804 FEBS Journal 272 (2005) 1804–1814 ª 2005 FEBS

trophic hormones, such as adrenocorticotrophin
(ACTH) and luteinizing hormone, regulate AA release
is not entirely clear.
We have proposed the involvement of acyl-CoA
synthetase 4 (ACS4) and mitochondrial acyl-CoA thio-
esterase I (MTE-I) as important regulators of AA release
in the mechanism of action of trophic hormone-stimu-
lated cholesterol metabolism [12]. ACS4 has been des-
cribed as an AA-preferring acyl-CoA synthetase that is
preferentially expressed in steroidogenic tissues such
as adrenal cortex, luteal and stromal cells of the ovary
and Leydig cells of the testis [13]. Moreover, it has
been demonstrated that ACS4 expression in the murine
adrenocortical tumor cell line Y1 is induced by ACTH
and suppressed by glucocorticoids [14].
The acyl-CoA thioesterase was first identified as a
43-kDa phosphoprotein by its capacity to increase mito-
chondrial steroidogenesis in a cell-free assay [15]. The
protein was then purified to homogeneity [15]. Further
cloning and sequencing of its cDNA revealed that it is
member of a thioesterase family with long-chain acyl-
CoA thioesterase activity [16] which includes four iso-
forms with different subcellular localization and a high
degree of homology [17,18]. In particular, MTE-I was
shown by immunoelectron microscopy to associate with
the matrix face of mitochondrial cristae [17]. In accord-
ance with the postulated role of MTE-I in steroido-
genesis, we detected the protein and its mRNA in the
adrenal gland, ovary, testis, placenta and brain [16].
Although it is known that acyl-CoA thioesterases
are a group of enzymes that catalyze the hydrolysis of
acyl-CoA to the nonesterified fatty acid and CoA [19]
and that they can release AA from the arachidonoyl-
CoA, so far, the phospholipase A2 pathway is the most
commonly accepted mechanism operating to produce
lipoxygenated products from plasma membrane signa-
ling [7].
Using inhibitors of the acyl-CoA synthetase and the
acyl-CoA thioesterase, we have previously postulated
the existence of a new pathway for AA release that
operates in the regulation of steroid synthesis in adre-
nal cells [12]. Here we address the question of whether
MTE-I and ACS4 are indeed essential for AA release
and cholesterol metabolism in steroidogenic cells. We
show for the first time that silencing the expression of
MTE-I by antisense cDNA or small interfering RNA
(siRNA) or of ACS4 by siRNA results in the inhibi-
tion of steroid synthesis, an effect that is overcome by
the addition of exogenous AA. In summary, our
results demonstrate a critical role for both ACS4 and
MTE-I in the hormonal regulation of steroidogenesis
as a new pathway of AA release different from the
classical phospholipase A2.
Results
MTE-I knock down and overexpression
in MA-10 cells
To provide definitive proof about the role played by
the mitochondrial acyl-CoA thioesterase in steroido-
genesis, we performed experiments aimed at silencing
the expression of MTE-I or at overexpressing the pro-
tein in steroidogenic cells. For this purpose, we transi-
ently transfected MA-10 cells with pRc ⁄CMVi plasmid
containing either an antisense or the full-sense MTE-I
cDNA (accession No. Y09333). The efficiency of trans-
fection using the protocol described in Experimental
Procedures was 50–70%.
The effect of sense and antisense plasmid transfec-
tion on MTE-I protein concentrations was studied
by immunocytochemistry, using a specific antibody
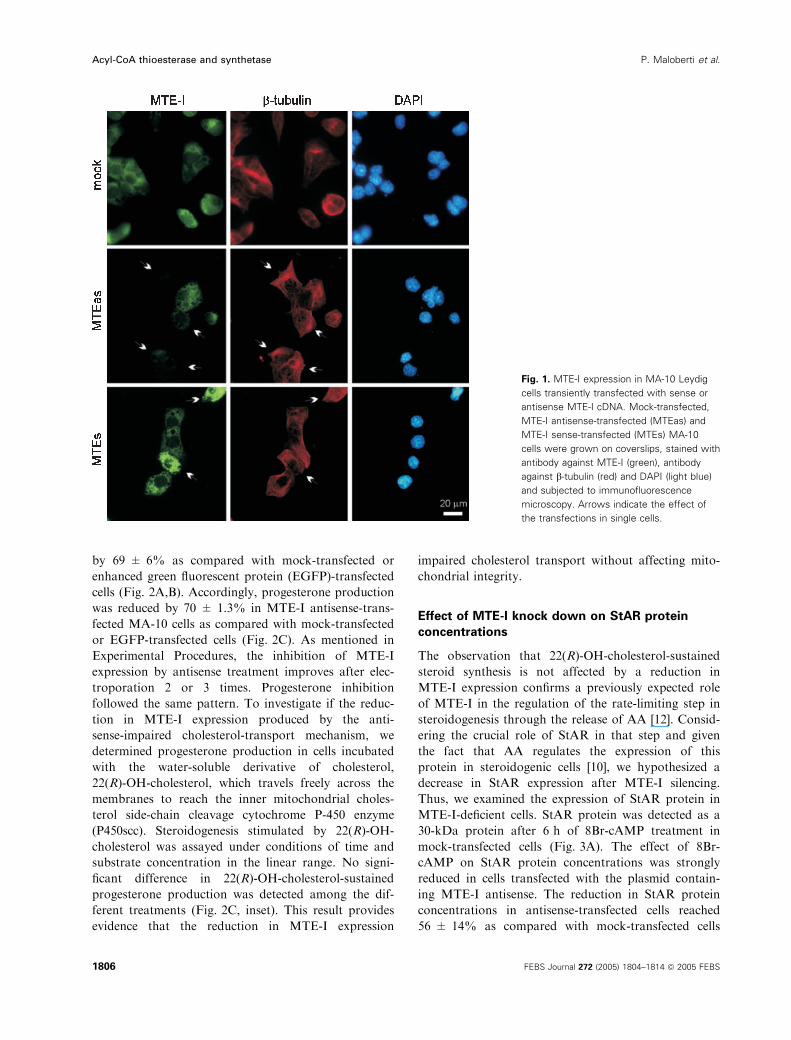
against the MTE-I and b-tubulin as control (Fig. 1).
As expected, antisense-transfected cells showed a
strong reduction in MTE-I protein concentrations,
whereas cells transfected with full MTE-I sense cDNA
showed a clear increase compared with cells trans-
fected with vector alone. The expression of b-tubulinremained unchanged in spite of the treatments used.
Neither MTE-I antisense or sense expression affected
cell viability, as assessed by the trypan blue exclusion
method and 3-(4,5-dimethyl-2-thiazolyl)-2,5-diphenyl-
2H-tetrazolium bromide (MTT) assay (0.68 ± 0.02,
0.66 ± 0.01 and 0.65 ± 0.01 absorbance units for
antisense-transfected, sense-transfected and mock-trans-
fected cells, respectively). As the MTT assay is based
on MTT reduction by the mitochondrial diaphorase
enzyme, an index of mitochondrial integrity [20],
these results indicate that neither antisense nor sense
transfection affect mitochondrial function. Moreover,
staining of the cells with the nuclear dye 4¢,6-diami-
dino-2-phenylindole (DAPI) showed no changes in
nuclear morphology, indicating that neither of the
treatments used induced apoptosis in MA-10 cells
(Fig. 1).
Effect of MTE-I knock down on steroidogenesis
Steroid hormone production in Leydig cells involves
increases in intracellular cAMP concentrations. Thus,
we studied the effect of MTE-I antisense on 8-bromo-
3¢,5¢-cAMP (8Br-cAMP)-stimulated progesterone pro-
duction (the major steroid produced by the MA-10 cell
line) and correlated the steroid-producing capability
with the level of protein expression as assessed by
western blot. The expression of MTE-I protein in
MTE-I antisense-transfected MA-10 cells was reduced
P. Maloberti et al. Acyl-CoA thioesterase and synthetase
FEBS Journal 272 (2005) 1804–1814 ª 2005 FEBS 1805
by 69 ± 6% as compared with mock-transfected or
enhanced green fluorescent protein (EGFP)-transfected
cells (Fig. 2A,B). Accordingly, progesterone production
was reduced by 70 ± 1.3% in MTE-I antisense-trans-
fected MA-10 cells as compared with mock-transfected
or EGFP-transfected cells (Fig. 2C). As mentioned in
Experimental Procedures, the inhibition of MTE-I
expression by antisense treatment improves after elec-
troporation 2 or 3 times. Progesterone inhibition
followed the same pattern. To investigate if the reduc-
tion in MTE-I expression produced by the anti-
sense-impaired cholesterol-transport mechanism, we
determined progesterone production in cells incubated
with the water-soluble derivative of cholesterol,
22(R)-OH-cholesterol, which travels freely across the
membranes to reach the inner mitochondrial choles-
terol side-chain cleavage cytochrome P-450 enzyme
(P450scc). Steroidogenesis stimulated by 22(R)-OH-
cholesterol was assayed under conditions of time and
substrate concentration in the linear range. No signi-
ficant difference in 22(R)-OH-cholesterol-sustained
progesterone production was detected among the dif-
ferent treatments (Fig. 2C, inset). This result provides
evidence that the reduction in MTE-I expression
impaired cholesterol transport without affecting mito-
chondrial integrity.
Effect of MTE-I knock down on StAR protein
concentrations
The observation that 22(R)-OH-cholesterol-sustained
steroid synthesis is not affected by a reduction in
MTE-I expression confirms a previously expected role
of MTE-I in the regulation of the rate-limiting step in
steroidogenesis through the release of AA [12]. Consid-
ering the crucial role of StAR in that step and given
the fact that AA regulates the expression of this
protein in steroidogenic cells [10], we hypothesized a
decrease in StAR expression after MTE-I silencing.
Thus, we examined the expression of StAR protein in
MTE-I-deficient cells. StAR protein was detected as a
30-kDa protein after 6 h of 8Br-cAMP treatment in
mock-transfected cells (Fig. 3A). The effect of 8Br-
cAMP on StAR protein concentrations was strongly
reduced in cells transfected with the plasmid contain-
ing MTE-I antisense. The reduction in StAR protein
concentrations in antisense-transfected cells reached
56 ± 14% as compared with mock-transfected cells
Fig. 1. MTE-I expression in MA-10 Leydig
cells transiently transfected with sense or
antisense MTE-I cDNA. Mock-transfected,
MTE-I antisense-transfected (MTEas) and
MTE-I sense-transfected (MTEs) MA-10
cells were grown on coverslips, stained with
antibody against MTE-I (green), antibody
against b-tubulin (red) and DAPI (light blue)
and subjected to immunofluorescence
microscopy. Arrows indicate the effect of
the transfections in single cells.
Acyl-CoA thioesterase and synthetase P. Maloberti et al.
1806 FEBS Journal 272 (2005) 1804–1814 ª 2005 FEBS

when quantified by densitometry related to the b-tubu-lin signal.
As was the case for StAR protein concentrations,
knock down of MTE-I resulted in a significant reduction
in progesterone production in cells that had been stimu-
lated for 6 h with 8Br-cAMP (data not shown). This
result demonstrates the participation of MTE-I in the
regulation of the rate-limiting step in steroidogenesis.
To provide additional insights into the sequential
steps involved in down-regulation of StAR protein
concentrations, we investigated whether treatment with
MTE-I antisense had any effect on StAR mRNA con-
centrations. Down-regulation of MTE-I produced a
marked inhibition of StAR mRNA concentrations,
which was already detectable after 1 h of 8Br-cAMP
stimulation (Fig. 3B). These results are in agreement
with the inhibition of steroidogenesis produced by
actinomycin D in this cell type, as this drug inhibits
only 30% of steroid synthesis induced by 8Br-cAMP
or AA for 1 h (data not shown). However, the effect
of cycloheximide on steroidogenesis stimulated with
8Br-cAMP or AA produced inhibition before 1 h of
stimulation. These results suggest that AA regulates
StAR protein at transcriptional and protein levels.
Effect of MTE-I overexpression on steroidogenesis
To further explore the role of MTE-I in the regulation
of steroidogenesis, we examined the effect of MTE-I
A
B
C
Fig. 2. Effect of MTE-I knock down on steroidogenesis in MA-10
cells. Mock-transfected (h), EGFP-transfected ( ) or MTE-I anti-
sense-transfected (MTEas, ) MA-10 cells were incubated for 1 h
in serum-free medium in the presence or absence of 8Br-cAMP
(0.5 mM). (A) MTE-I expression was assayed by immunoblotting.
The membrane was incubated sequentially with anti-MTE-I and
anti-(b-tubulin) sera. (B) Western blot quantification by densitome-
try. Bars denote relative levels of MTE-I expression. (C) Determin-
ation of progesterone production by RIA. Inset: MA-10-transfected
cells were incubated for 1 h in serum-free medium in the presence
of 22(R)-OH-cholesterol (5 lM). Results are expressed as the mean
± SD from one representative (n ¼ 3) experiment performed in
triplicate. ***P < 0.001 vs. mock-transfected or EGFP-transfected
8Br-cAMP-treated cells.
A
B
Fig. 3. Effect of MTE-I knock down on StAR expression in steroido-
genic cells. Mock-transfected or MTE-I antisense-transfected
(MTEas) MA-10 cells were incubated in the presence or absence of
8Br-cAMP (0.5 mM). (A) Representative western blot of MA-10
transfected cells stimulated for 6 h with 8Br-cAMP. The membrane
was blotted sequentially with anti-MTE-I, anti-StAR and antib-tubu-
lin sera. (B) Representative northern blot analysis of total RNA from
MA-10 transfected cells stimulated for 1 and 6 h with 8Br-cAMP.
The membrane was developed using probes against StAR and 28S.
P. Maloberti et al. Acyl-CoA thioesterase and synthetase
FEBS Journal 272 (2005) 1804–1814 ª 2005 FEBS 1807
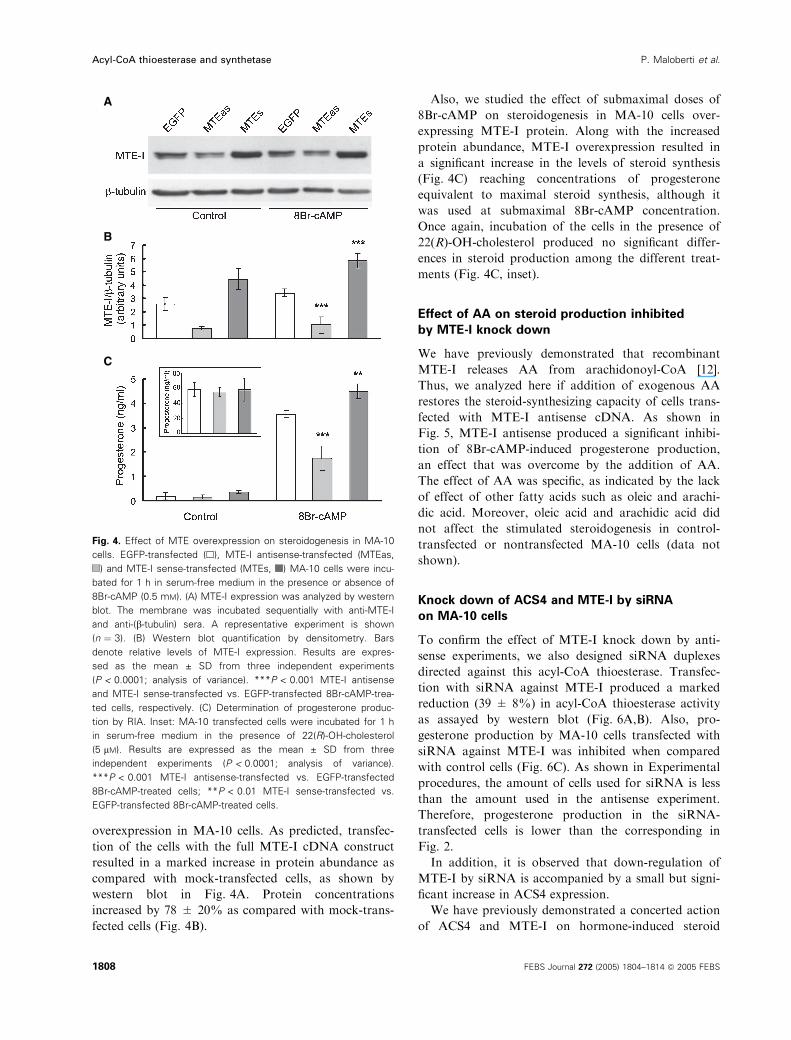
overexpression in MA-10 cells. As predicted, transfec-
tion of the cells with the full MTE-I cDNA construct
resulted in a marked increase in protein abundance as
compared with mock-transfected cells, as shown by
western blot in Fig. 4A. Protein concentrations
increased by 78 ± 20% as compared with mock-trans-
fected cells (Fig. 4B).
Also, we studied the effect of submaximal doses of
8Br-cAMP on steroidogenesis in MA-10 cells over-
expressing MTE-I protein. Along with the increased
protein abundance, MTE-I overexpression resulted in
a significant increase in the levels of steroid synthesis
(Fig. 4C) reaching concentrations of progesterone
equivalent to maximal steroid synthesis, although it
was used at submaximal 8Br-cAMP concentration.
Once again, incubation of the cells in the presence of
22(R)-OH-cholesterol produced no significant differ-
ences in steroid production among the different treat-
ments (Fig. 4C, inset).
Effect of AA on steroid production inhibited
by MTE-I knock down
We have previously demonstrated that recombinant
MTE-I releases AA from arachidonoyl-CoA [12].
Thus, we analyzed here if addition of exogenous AA
restores the steroid-synthesizing capacity of cells trans-
fected with MTE-I antisense cDNA. As shown in
Fig. 5, MTE-I antisense produced a significant inhibi-
tion of 8Br-cAMP-induced progesterone production,
an effect that was overcome by the addition of AA.
The effect of AA was specific, as indicated by the lack
of effect of other fatty acids such as oleic and arachi-
dic acid. Moreover, oleic acid and arachidic acid did
not affect the stimulated steroidogenesis in control-
transfected or nontransfected MA-10 cells (data not
shown).
Knock down of ACS4 and MTE-I by siRNA
on MA-10 cells
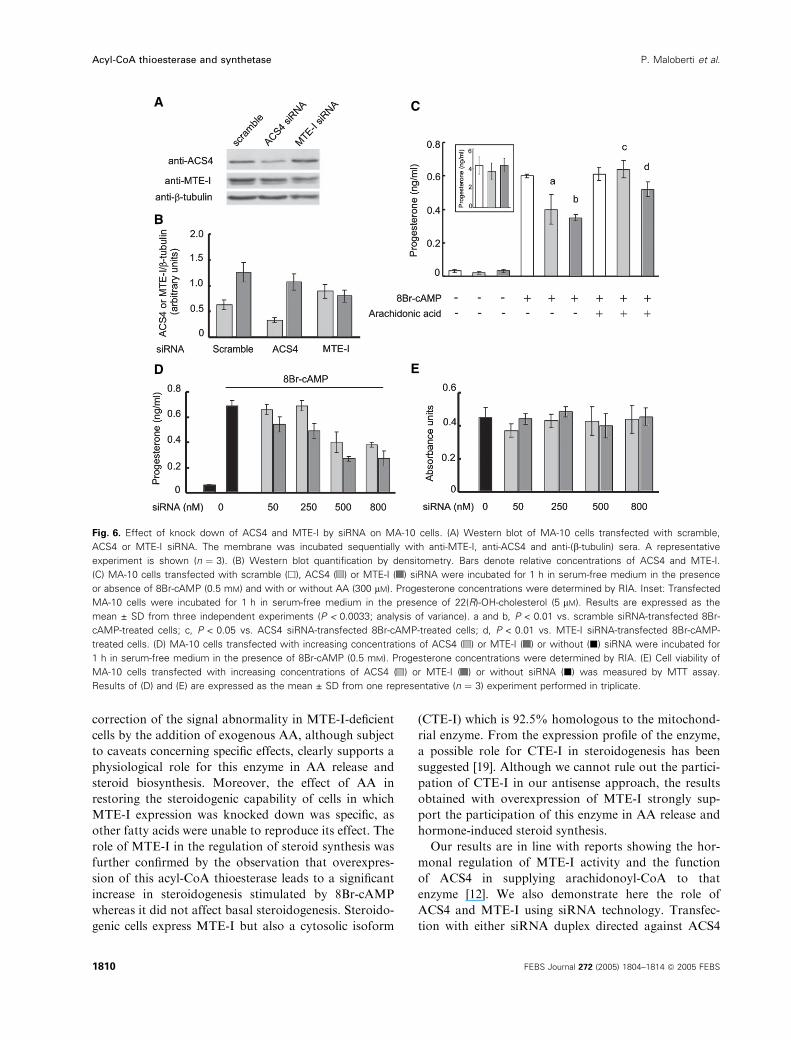
To confirm the effect of MTE-I knock down by anti-
sense experiments, we also designed siRNA duplexes
directed against this acyl-CoA thioesterase. Transfec-
tion with siRNA against MTE-I produced a marked
reduction (39 ± 8%) in acyl-CoA thioesterase activity
as assayed by western blot (Fig. 6A,B). Also, pro-
gesterone production by MA-10 cells transfected with
siRNA against MTE-I was inhibited when compared
with control cells (Fig. 6C). As shown in Experimental
procedures, the amount of cells used for siRNA is less
than the amount used in the antisense experiment.
Therefore, progesterone production in the siRNA-
transfected cells is lower than the corresponding in
Fig. 2.
In addition, it is observed that down-regulation of
MTE-I by siRNA is accompanied by a small but signi-
ficant increase in ACS4 expression.
We have previously demonstrated a concerted action
of ACS4 and MTE-I on hormone-induced steroid
A
B
C
Fig. 4. Effect of MTE overexpression on steroidogenesis in MA-10
cells. EGFP-transfected (h), MTE-I antisense-transfected (MTEas,
) and MTE-I sense-transfected (MTEs, ) MA-10 cells were incu-
bated for 1 h in serum-free medium in the presence or absence of
8Br-cAMP (0.5 mM). (A) MTE-I expression was analyzed by western
blot. The membrane was incubated sequentially with anti-MTE-I
and anti-(b-tubulin) sera. A representative experiment is shown
(n ¼ 3). (B) Western blot quantification by densitometry. Bars
denote relative levels of MTE-I expression. Results are expres-
sed as the mean ± SD from three independent experiments
(P < 0.0001; analysis of variance). ***P < 0.001 MTE-I antisense
and MTE-I sense-transfected vs. EGFP-transfected 8Br-cAMP-trea-
ted cells, respectively. (C) Determination of progesterone produc-
tion by RIA. Inset: MA-10 transfected cells were incubated for 1 h
in serum-free medium in the presence of 22(R)-OH-cholesterol
(5 lM). Results are expressed as the mean ± SD from three
independent experiments (P < 0.0001; analysis of variance).
***P < 0.001 MTE-I antisense-transfected vs. EGFP-transfected
8Br-cAMP-treated cells; **P < 0.01 MTE-I sense-transfected vs.
EGFP-transfected 8Br-cAMP-treated cells.
Acyl-CoA thioesterase and synthetase P. Maloberti et al.
1808 FEBS Journal 272 (2005) 1804–1814 ª 2005 FEBS

synthesis [12]. On the basis of these results, silencing
the expression of ACS4 should also result in inhibition
of hormone-induced steroid synthesis. siRNA duplexes
directed against ACS4 were also produced for transfec-
tion of MA-10 cells. siRNA transfection resulted in a
marked reduction (56 ± 6%) in ACS4 protein concen-
trations and steroidogenesis as detected by western
blot and RIA (Fig. 6A,B,C, respectively).
As demonstrated for MTE-I antisense cDNA effect,
addition of exogenous AA blunted the inhibition of
progesterone production produced by both siRNAs
(Fig. 6B). As shown in Fig. 2, the reduction in protein
expression correlates with the inhibition of progester-
one production. As was the case in previous experi-
ments, the 22(R)-OH-cholesterol effect on steroid
synthesis was not modified by ACS4 or MTE-I siRNA
transfection (Fig. 6B, inset). Dose–response curves of
progesterone production and cell viability for both
siRNAs are shown in Fig. 6D,E. The maximal inhi-
bition of progesterone production was observed at
500 nm siRNA. Importantly, cell viability measured by
the MTT assay remained unchanged after siRNA
transfection independently of the concentrations of
siRNA used (Fig. 6E). Transfection with control
siRNA remained unchanged for progesterone produc-
tion and cell viability (data not shown).
Discussion
We have previously reported that recombinant MTE-I
releases AA from arachidonoyl-CoA in vitro and that
ACTH increases the activity of the endogenous enzyme
in adrenal cells [12]. Moreover, inhibitors of MTE-I
and ACS4 inhibited ACTH-induced steroidogenesis in
adrenal cells [12]. These results are consistent with the
involvement of ACS4 and MTE-I as important regula-
tors in the mechanism of action of trophic hormone-
stimulated cholesterol metabolism.
To provide definitive proof about the role of MTE-I
and ACS4 in the regulation of hormone-stimulated
steroid synthesis, we performed experiments aimed at
studying the effect of suppressing the expression of
both enzymes. Immunocytochemistry and western blot
experiments demonstrate that the expression of MTE-I
was strongly reduced when MA-10 cells were transfect-
ed with a vector containing an MTE-I antisense
cDNA. Importantly, the transfection procedure devel-
oped and used in this study did not affect cell viability
and ⁄or morphology of sense-transfected or antisense-
transfected cells as compared with mock-transfected
cells. Moreover, a possible negative effect of the treat-
ments on mitochondrial activity was ruled out by using
the MTT assay. Finally, a possible apoptotic effect
produced by the manipulation of MTE-I protein con-
centrations was also discarded after staining of the
cells with DAPI showed no changes in nuclear mor-
phology.
Knocking down MTE-I expression levels leads to a
strong reduction in 8Br-cAMP-stimulated steroidogen-
esis. The reduction in protein expression does not
affect steroid biosynthesis when it is induced by a per-
meable analog of cholesterol, 22(R)-OH-cholesterol,
which bypasses the rate-limiting step of steroidogene-
sis. Given the role of StAR in cholesterol transport
and the fact that AA induces StAR expression, we
studied here whether the reduction in MTE-I expres-
sion could affect steroid hormone synthesis by affect-
ing StAR protein concentrations. We demonstrate that
a reduction in MTE-I expression in MA-10 cells leads
to a marked reduction in 8Br-cAMP-mediated StAR
induction at the RNA and protein level. Together,
these results provide evidence that the reduction in
MTE-I expression impairs cholesterol transport with-
out affecting mitochondrial integrity and are in line
with previous reports that AA regulates the rate-limit-
ing step in steroid biosynthesis [10].
Importantly, the reduction in steroid biosynthesis was
restored by addition of exogenous AA, clearly demons-
trating that the effect produced by MTE-I protein
reduction is based on decreased AA concentrations. The
Fig. 5. Effect of AA on steroid production inhibited by MTE-I knock
down. EGFP-transfected (h) or MTE-I antisense-transfected
(MTEas, ) MA-10 cells were incubated for 1 h in the presence or
absence of 8Br-cAMP (0.5 mM) and with or without 300 lM differ-
ent fatty acids (arachidonic, arachidic and oleic acid) as indicated.
Progesterone concentrations were determined by RIA. Results are
expressed as the mean ± SD from three independent experiments
(P < 0.0019; analysis of variance). **P < 0.01 MTE-I antisense-
transfected vs. EGFP-transfected 8Br-cAMP-treated cells; *P < 0.05
MTE-I antisense-transfected 8Br-cAMP-treated cells vs. MTE-I
antisense-transfected cells treated with 8Br-cAMP and arachidonic
acid.
P. Maloberti et al. Acyl-CoA thioesterase and synthetase
FEBS Journal 272 (2005) 1804–1814 ª 2005 FEBS 1809
correction of the signal abnormality in MTE-I-deficient
cells by the addition of exogenous AA, although subject
to caveats concerning specific effects, clearly supports a
physiological role for this enzyme in AA release and
steroid biosynthesis. Moreover, the effect of AA in
restoring the steroidogenic capability of cells in which
MTE-I expression was knocked down was specific, as
other fatty acids were unable to reproduce its effect. The
role of MTE-I in the regulation of steroid synthesis was
further confirmed by the observation that overexpres-
sion of this acyl-CoA thioesterase leads to a significant
increase in steroidogenesis stimulated by 8Br-cAMP
whereas it did not affect basal steroidogenesis. Steroido-
genic cells express MTE-I but also a cytosolic isoform
(CTE-I) which is 92.5% homologous to the mitochond-
rial enzyme. From the expression profile of the enzyme,
a possible role for CTE-I in steroidogenesis has been
suggested [19]. Although we cannot rule out the partici-
pation of CTE-I in our antisense approach, the results
obtained with overexpression of MTE-I strongly sup-
port the participation of this enzyme in AA release and
hormone-induced steroid synthesis.
Our results are in line with reports showing the hor-
monal regulation of MTE-I activity and the function
of ACS4 in supplying arachidonoyl-CoA to that
enzyme [12]. We also demonstrate here the role of
ACS4 and MTE-I using siRNA technology. Transfec-
tion with either siRNA duplex directed against ACS4
A
B
D
C
E
Fig. 6. Effect of knock down of ACS4 and MTE-I by siRNA on MA-10 cells. (A) Western blot of MA-10 cells transfected with scramble,
ACS4 or MTE-I siRNA. The membrane was incubated sequentially with anti-MTE-I, anti-ACS4 and anti-(b-tubulin) sera. A representative
experiment is shown (n ¼ 3). (B) Western blot quantification by densitometry. Bars denote relative concentrations of ACS4 and MTE-I.
(C) MA-10 cells transfected with scramble (h), ACS4 ( ) or MTE-I ( ) siRNA were incubated for 1 h in serum-free medium in the presence
or absence of 8Br-cAMP (0.5 mM) and with or without AA (300 lM). Progesterone concentrations were determined by RIA. Inset: Transfected
MA-10 cells were incubated for 1 h in serum-free medium in the presence of 22(R)-OH-cholesterol (5 lM). Results are expressed as the
mean ± SD from three independent experiments (P < 0.0033; analysis of variance). a and b, P < 0.01 vs. scramble siRNA-transfected 8Br-
cAMP-treated cells; c, P < 0.05 vs. ACS4 siRNA-transfected 8Br-cAMP-treated cells; d, P < 0.01 vs. MTE-I siRNA-transfected 8Br-cAMP-
treated cells. (D) MA-10 cells transfected with increasing concentrations of ACS4 ( ) or MTE-I ( ) or without (j) siRNA were incubated for
1 h in serum-free medium in the presence of 8Br-cAMP (0.5 mM). Progesterone concentrations were determined by RIA. (E) Cell viability of
MA-10 cells transfected with increasing concentrations of ACS4 ( ) or MTE-I ( ) or without siRNA (j) was measured by MTT assay.
Results of (D) and (E) are expressed as the mean ± SD from one representative (n ¼ 3) experiment performed in triplicate.
Acyl-CoA thioesterase and synthetase P. Maloberti et al.
1810 FEBS Journal 272 (2005) 1804–1814 ª 2005 FEBS

or MTE-I produced a significant inhibition in 8-Br-
cAMP-induced steroid synthesis. These results together
with our finding that AA or 22(R)-OH-cholesterol can
bypass the effect of siRNA strongly indicate that
ACS4 and MTE-I act in the same signaling pathway
at a step before the rate-limiting passage of cholesterol
from the outer to inner mitochondrial. Therefore, a
concerted action of ACS4 and MTE-I in regulating
the concentrations of AA during steroidogenesis seems
plausible.
Down-regulation of MTE-I by siRNA is accompan-
ied by an increase in ACS4 expression. We do not
know the mechanisms involved in this regulation. It
can be speculated that either the reduction in steroid
synthesis or the reduction in arachidonic acid release
may regulate the expression of ACS4.
The human gene encoding ACS4 is located in the
chromosome Xq 22–23 region close to the a5 chain of
the type 4 collagen gene which is related to X-linked
Alport syndrome. Genetic analysis of the gene revealed
that it was deleted in a family with Alport syndrome,
ellipoptocytosis and mental retardation [21].
ACS4-deficient mice have been generated [22].
Female mice heterozygous for ACS4 deficiency became
pregnant less frequently and produced small litters
with extremely low transmission of the disrupted alle-
les with a high frequency of uterus embryonic death.
ACS4+ females showed marked accumulation of pros-
taglandin in the uteruses of the heterozygous females,
suggesting that ACS4 modulates female fertility and
uterus prostaglandin production. ACS4 - ⁄Y hemizy-
gous males presented an apparently normal phenotype.
The authors suggest that ACS3, an isoform of the
enzyme that also prefers arachidonate [23], may com-
pensate for the ACS4 deficiency.
Under the experimental conditions used in this study,
the knock down of ACS4 expression was performed in
isolated cells. Under these conditions, cells respond to
the acute hormonal stimulus in a time frame in which no
compensatory mechanisms may occur. Thus, our system
seems more appropriate to exploration of the actual
physiological role of ACS4 in AA release, leukotriene
formation and steroid synthesis.
It is known that AA has to be metabolized through
the lipoxygenase pathway in order to stimulate StAR
expression and steroidogenesis [11,24]. This raises the
question of whether the existence of a different
AA-releasing pathway may provide AA metabolites in
a special compartment of the cell (e.g. mitochondria).
The reason why the action of the two opposite
enzymes, namely the arachidonoyl-CoA synthetase and
the arachidonoyl-CoA thioesterase, is needed may be
the need to sequester AA from the free pool in the
form of arachidonoyl-CoA to bring this substrate to a
special compartment of the cells. The compartmental-
ization of long-chain acyl-CoA esters is an important
unsolved problem. The high degree of sequestration
of CoA into long-chain acyl-CoA suggests that AA is
limiting for diverse roles in specific compartments of
the cells.
The role of long-chain acyl-CoA esters in signal-
transduction pathways is an important issue [25]. It is
known that an acyl-CoA-binding protein known also
as DBI (diazepam-binding inhibitor) is expressed at
high concentrations in steroidogenic cells [26,27] and
interacts with the PBR in the mitochondria [5]. Both
DBI and PBR have been reported to be essential for
normal steroid biosynthesis [28,29]. Moreover, the site
of action of these two proteins is the rate-limiting step
in steroidogenesis [6]. Fatty acyl-CoAs bind to the
acyl-CoA-binding protein, which can bind and may
thereby activate PBR. This interaction favors the
accumulation of fatty acids near StAR, perhaps by
promoting fatty acyl-CoA transfer into the inner
mitochondria.
Although it is clear from our results that the acyl-
CoA thioesterase is critical in steroidogenesis and that
the AA produced promotes, at least, the stimulation of
StAR protein expression, a possible action of the fatty
acid at a different level of the steroidogenic pathway
cannot be ruled out. In accordance with this sugges-
tion, a positive action on cholesterol metabolism of
nonesterified fatty acids in the mitochondrial mem-
brane has been suggested [30]. It was demonstrated
that cholesterol binding to P450scc in lipid vesicles is
greatly potentiated when the local membrane is ren-
dered more fluid by the addition of nonesterified fatty
acids [31]. P450scc interactions with cholesterol, but
not with hydroxycholesterol, are strongly affected by
the lipid environment of the inner mitochondrial mem-
brane. Cholesterol interacts strongly with the fatty acid
chains of many phospholipids and is thereby con-
strained from interacting with P450scc. The increase in
membrane fluidity in the presence of these fatty acids
possibly favors the interaction of cholesterol with
StAR or P450scc [30,31]. All these observations may
explain why the free cytosolic AA released from cho-
lesterol esters or phospholipids should be re-esterified
by the AA-preferring acyl-CoA synthetase in order to
be released in mitochondria by the specific acyl-CoA
thioesterase.
In summary, the present work shows for the first
time that knocking down MTE-I and ACS4 mRNA
by antisense or siRNA in steroidogenic cells results in
a reduction in steroid biosynthesis. This provides evi-
dence for the pivotal role played by ACS4 and MTE-I
P. Maloberti et al. Acyl-CoA thioesterase and synthetase
FEBS Journal 272 (2005) 1804–1814 ª 2005 FEBS 1811

in AA release, StAR protein expression, and steroido-
genesis.
Experimental procedures
Materials
8Br-cAMP, 22(R)-OH-cholesterol, fatty acid-free BSA,
arachidonic, arachidic and oleic acids were purchased from
Sigma Chemical Co. (St Louis, MO, USA). Ham-F10 and
Waymouth MB752 ⁄ 1 cell culture media were from Life
Technologies Inc. (Gaithersburg, MD, USA). All other rea-
gents were of the highest grade available.
Cell culture
The MA-10 cell line is a clonal strain of mouse Leydig
tumor cells that produce progesterone rather than testoster-
one as the major steroid [32]. MA-10 cells were generously
provided by Mario Ascoli, University of Iowa, College of
Medicine (Iowa City, IA, USA) and were handled as ori-
ginally described [30]. The growth medium consisted of
Waymouth MB752 ⁄ 1 containing 1.1 gÆL)1 NaHCO3, 20 mm
Hepes, 50 lgÆmL)1 gentamicin, and 15% horse serum.
Flasks and multiwell plates were maintained at 36 �C in a
humidified atmosphere containing 5% CO2.
Plasmid transfection
To develop optimized conditions for plasmid delivery to
MA-10 cells, several electroporation variables were tested.
Electroporation experiments were performed using a Gene
Pulser� II Electroporation System (Gibco, Grand Island,
NY, USA). MA-10 cells were routinely subcultured, and
cells were harvested and resuspended in NaCl ⁄Pi at a den-
sity of 1.6 · 107 cellsÆmL)1. pRc ⁄CMVi plasmid [33] con-
taining the enhanced form of green fluorescent protein
(EGFP) (10 lg) was added to a 0.4-cm gap cuvette (Gibco)
containing 600 lL cell suspension. This suspension was
electroporated with one pulse of 0.25 V and 600 lF. Cellswere then cultured in complete fresh medium for 24 h and
electroporated under the same conditions twice more. The
transfection efficiency of electroporation was estimated to
reach 50–70% by counting EGFP-transfected fluorescent
cells. Approximately 24 h after transfection, cells were
stimulated with 8Br-cAMP (a permeable analog of cAMP)
in culture medium containing 0.1% fatty acid-free BSA.
Progesterone produced was measured by RIA as previously
described [12,15].
SiRNA transfection
Two siRNAs were designed to target MTE-I (accession No.
AF180798) and the ACS4 (accession No. NM_207625)
coding sequence. siRNA sequences 5¢-AAGAGCGAGT
TCTATGCTGAT (nucleotides 322–342 of MTE-I cDNA),
5¢-AAATGACAGGCCAGTGTGAAC (nucleotides 1124–1143
of ACS4 cDNA), and 5¢-CGAGAAGACGTAAAGC
(scramble siRNA of MTE-I) were custom-designed by
Dharmacon (Lafayette, CO, USA). One day before trans-
fection, MA-10 cells (5 · 105 cells per well) were grown up
to 80% confluence on 24-well plates. Transfection was per-
formed using siRNA (800 nm) in Opti-MEM medium and
2 lL Lipofectamine 2000 reagent (Invitrogen, Carlsbad,
CA, USA) according to the instructions of the manufac-
turer. Cells were placed in normal culture medium 6 h after
transfection and further grown for 48 h. MA-10 cells were
stimulated with 8Br-cAMP in culture medium containing
0.1% fatty acid-free BSA. Progesterone production was
measured by RIA, and data are shown as progesterone pro-
duction (ngÆmL)1) in the incubation medium.
Cell viability and mitochondrial integrity
Cell viability was analyzed using the trypan blue exclusion
method. At the end of the incubation, the cells were washed
three times with NaCl ⁄Pi and incubated for 15 min with
0.1% trypan blue stain. After being washed, stained (dead)
cells were counted by light microscopy. Cell viability and
mitochondrial integrity were also evaluated by measuring
the levels of cellular MTT reduction [20].
SDS/PAGE and immunoblot assay
Proteins were separated by SDS ⁄PAGE (12% gel) and
electrophoretically transferred to poly(vinylidene difluoride)
membrane (Bio-Rad Laboratories Inc, Hercules, CA, USA)
in buffer (25 mm Tris ⁄HCl, 192 mm glycine, pH 8.3, 20%
methanol) at a constant voltage of 2.4 mAÆcm)2 for 90 min.
Membranes were then incubated with 5% fat-free powdered
milk in NaCl ⁄Tris ⁄Tween (500 mm NaCl, 20 mm Tris ⁄HCl,
pH 7.5, 0.5% Tween-20) for 60 min at room temperature
with gentle shaking. The membranes were then rinsed twice
in NaCl ⁄Tris ⁄Tween and incubated overnight with the
appropriate dilutions of primary antibody at 4 �C. Boundantibodies were detected by chemiluminescence using the
ECL kit (Amersham Pharmacia Biotech, Buenos Aires,
Argentina).
Northern blot
Total RNA from MA-10 cells was prepared by homogeni-
zation in TRIzol reagent according to the manufacturer’s
instructions. Samples of RNA (24 lg) were resolved on
1.2% agarose ⁄ 2.2 m formaldehyde gels and transferred on
to Hybond-N+ nylon membranes (Amersham Pharmacia
Biotech). A cDNA probe for StAR was prepared by
RT-PCR from total RNA from MA-10 cells. Primers were
Acyl-CoA thioesterase and synthetase P. Maloberti et al.
1812 FEBS Journal 272 (2005) 1804–1814 ª 2005 FEBS

designed according to the published sequence of mouse
StAR. The forward (5¢-AAAGGATTAAGGCACCAA
GCTGTGC-3¢) and reverse (5¢-CTCTGATGACACCA
CTCTGCTCCGG-3¢) primers were used to amplify a
588-bp fragment. The PCR product was sequenced to con-
firm its identity. After prehybridization for 8 h at 42 �C,blots were hybridized overnight with the [32P]dCTP[aP]-radiolabeled cDNA probe at 42 �C. The hybridization
solution contained 6· SCC, 5· Denhardt’s solution, 0.5%
formamide, and 100 lgÆmL)1 denatured salmon sperm
DNA. Blots were subsequently washed twice with 2· SSPE
(150 mm NaCl, 10 mm NaH2PO4, 1 mm EDTA) ⁄ 0.5%SDS at room temperature and twice with 1· SSPE ⁄ 0.1%SDS at 65 �C. StAR hybridization signals were revelead
using a Storm PhosphorImager (Molecular Dynamics, Inc,
Sunnyvale, CA, USA). The membranes were then stripped
and rehybridized with 28S rRNA probe as loading control.
Immunofluorescence and microscopy
MA-10 cells grown on poly(l-lysine) glass coverslips were
washed once with NaCl ⁄Pi and then fixed for 10 min at
room temperature with 4% (w ⁄ v) paraformaldehyde in
NaCl ⁄Pi. Briefly, MA-10 fixed cells were rinsed in NaCl ⁄Pi
and incubated with blocking solution (1.5% goat serum in
0.3% Triton X-100 ⁄NaCl ⁄Pi) for 1 h at room temperature
and incubated with rabbit polyclonal antibody against
recombinant acyl-CoA thioesterase I [12] and mouse mono-
clonal antibody against b-tubulin in a humidified chamber
for 24 h at 4 �C. Primary antibodies were detected by cy2-
conjugated goat anti-(rabbit IgG) Ig or cy3-conjugated goat
anti-(mouse IgG) Ig (Molecular Probes, Eugene, OR,
USA). DNA was stained with DAPI. The glass coverslips
were mounted in FluorSave reagent (Calbiochem) and
examined in an Olympus BX 50 epifluorescence micro-
scope.
Statistical analysis
Data from the progesterone assay were analyzed for statis-
tical significance using analysis of variance followed by the
Student–Newman–Kuels test.
Acknowledgements
Thanks are due to Douglas Stocco for the StAR anti-
body (Department of Cell Biology and Biochemistry,
Texas Tech University, Lubbock, TX, USA). We also
thank Ingo Leibiger (Department of Molecular Medi-
cine, Karolinska Hospital L3, Karolinska Institutet,
Stockholm, Sweden) for the pRc ⁄CMVi plasmid. This
work was supported by grants from Agencia Nacional
de Promocion Cientıfica y Tecnologica (PICT6738, to
E.J.P.), Universidad de Buenos Aires (M034, to E.J.P.),
Fundacion Antorchas to E.J.P. and Consejo Nacional de
Investigaciones Cientıficas y Tecnicas (PEI02535, to
C.P.).
References
1 Privalle CT, Crivello JF & Jefcoate CR (1983) Regula-
tion of intramitochondrial cholesterol transfer to side-
chain cleavage cytochrome P-450 in rat adrenal gland.
Proc Natl Acad Sci USA 80, 702–706.
2 Crivello JF & Jefcoate CR (1980) Intracellular move-
ment of cholesterol in rat adrenal cells. Kinetics and
effects of inhibitors. J Biol Chem 255, 8144–8151.
3 Stocco DM (2000) The role of the StAR protein in ste-
roidogenesis: challenges for the future. J Endocrinol 164,
247–253.
4 Stocco DM (2001) Tracking the role of a star in the
sky of the new millennium. Mol Endocrinol 15, 1245–
1254.
5 Papadopoulos V, Amri H, Li H, Yao Z, Brown RC,
Vidic B & Culty M (2001) Structure, function and regu-
lation of the mitochondrial peripheral-type benzodiaze-
pine receptor. Therapie 56, 549–556.
6 Lacapere JJ & Papadopoulos V (2003) Peripheral-type
benzodiazepine receptor: structure and function of a
cholesterol-binding protein in steroid and bile acid
biosynthesis. Steroids 68, 569–585.
7 Khan WA, Blobe GC & Hannun YA (1995) Arachido-
nic acid and free fatty acids as second messengers and
the role of protein kinase C. Cell Signal 7, 171–184.
8 Hartl WH & Wolfe RR (1990) The phospholipid ⁄arachidonic acid second messenger system: its possible
role in physiology and pathophysiology of metabolism.
JPEN J Parenter Enteral Nutr 14, 416–427.
9 Whatley RE, Zimmerman GA, McIntyre TM &
Prescott SM (1990) Lipid metabolism and signal trans-
duction in endothelial cells. Prog Lipid Res 29, 45–63.
10 Wang X & Stocco DM (1999) Cyclic AMP and arachi-
donic acid: a tale of two pathways. Mol Cell Endocrinol
158, 7–12.
11 Wang XJ, Dyson MT, Jo Y, Eubank DW & Stocco
DM (2003) Involvement of 5-lipoxygenase metabolites
of arachidonic acid in cyclic AMP-stimulated steroido-
genesis and steroidogenic acute regulatory protein gene
expression. J Steroid Biochem Mol Biol 85, 159–166.
12 Maloberti P, Lozano RC, Mele PG, Cano F, Colonna
C, Mendez CF, Paz C & Podesta EJ (2002) Concerted
regulation of free arachidonic acid and hormone-
induced steroid synthesis by acyl-CoA thioesterases and
acyl-CoA synthetases in adrenal cells. Eur J Biochem
269, 5599–5607.
13 Kang MJ, Fujino T, Sasano H, Minekura H, Yabuki
N, Nagura H, Iijima H & Yamamoto TT (1997) A
novel arachidonate-preferring acyl-CoA synthetase is
P. Maloberti et al. Acyl-CoA thioesterase and synthetase
FEBS Journal 272 (2005) 1804–1814 ª 2005 FEBS 1813

present in steroidogenic cells of the rat adrenal, ovary,
and testis. Proc Natl Acad Sci USA 94, 2880–2884.
14 Cho YY, Kang MJ, Ogawa S, Yamashita Y, Fujino T
& Yamamoto TT (2000) Regulation by adrenocortico-
tropic hormone and arachidonate of the expression of
acyl-CoA synthetase 4, an arachidonate-preferring
enzyme expressed in steroidogenic tissues. Biochem
Biophys Res Commun 274, 741–745.
15 Paz C, Dada LA, Cornejo Maciel MF, Mele PG,
Cymeryng CB, Neuman I, Mendez CF, Finkielstein CV,
Solano AR, Park M et al. (1994) Purification of a novel
43-kDa protein (p43) intermediary in the activation of
steroidogenesis from rat adrenal gland. Eur J Biochem
224, 709–716.
16 Finkielstein C, Maloberti P, Mendez CF, Paz C,
Cornejo Maciel F, Cymeryng C, Neuman I, Dada L,
Mele PG et al. (1998) An adrenocorticotropin-regulated
phosphoprotein intermediary in steroid synthesis is
similar to an acyl-CoA thioesterase enzyme. Eur J
Biochem 256, 60–66.
17 Svensson LT, Engberg ST, Aoyama T, Usuda N,
Alexson SE & Hashimoto T (1998) Molecular cloning
and characterization of a mitochondrial peroxisome
proliferator-induced acyl-CoA thioesterase from rat
liver. Biochem J 329, 601–608.
18 Hunt MC, Nousiainen SE, Huttunen MK, Orii KE,
Svensson LT & Alexson SE (1999) Peroxisome prolif-
erator-induced long chain acyl-CoA thioesterases
comprise a highly conserved novel multi-gene family
involved in lipid metabolism. J Biol Chem 274, 34317–
34326.
19 Hunt MC & Alexson SE (2002) The role acyl-CoA
thioesterases play in mediating intracellular lipid meta-
bolism. Prog Lipid Res 41, 99–130.
20 Mosmann T (1983) Rapid colorimetric assay for cellular
growth and survival: application to proliferation and
cytotoxicity assays. J Immunol Methods 65, 55–63.
21 Piccini M, Vitelli F, Bruttini M, Pober BR, Jonsson JJ,
Villanova M, Zollo M, Borsani G, Ballabio A & Renieri
A (1998) FACL4, a new gene encoding long-chain acyl-
CoA synthetase 4, is deleted in a family with Alport
syndrome, elliptocytosis, and mental retardation. Geno-
mics 47, 350–358.
22 Cho YY, Kang MJ, Sone H, Suzuki T, Abe M, Igarashi
M, Tokunaga T, Ogawa S, Takei YA, Miyazawa T
et al. (2001) Abnormal uterus with polycysts, accumula-
tion of uterine prostaglandins, and reduced fertility in
mice heterozygous for acyl-CoA synthetase 4 deficiency.
Biochem Biophys Res Commun 284, 993–997.
23 Fujino T, Kang MJ, Suzuki H, Iijima H & Yamamoto T
(1996) Molecular characterization and expression of rat
acyl-CoA synthetase 3. J Biol Chem. 271, 16748–16752.
24 Wang X, Walsh LP, Reinhart AJ & Stocco DM (2000)
The role of arachidonic acid in steroidogenesis and ster-
oidogenic acute regulatory (StAR) gene and protein
expression. J Biol Chem 275, 20204–20209.
25 Faergeman NJ & Knudsen J (1997) Role of long-chain
fatty acyl-CoA esters in the regulation of metabolism
and in cell signalling. Biochem J 323, 1–12.
26 Knudsen J, Mandrup S, Rasmussen JT, Andreasen PH,
Poulsen F & Kristiansen K (1993) The function of acyl-
CoA-binding protein (ACBP) ⁄ diazepam binding inhib-
itor (DBI). Mol Cell Biochem 123, 129–138.
27 Papadopoulos V, Amri H, Boujrad N, Cascio C, Culty
M, Garnier M, Hardwick M, Li H, Vidic B, Brown AS
et al. (1997) Peripheral benzodiazepine receptor in
cholesterol transport and steroidogenesis. Steroids 62,
21–28.
28 Boujrad N, Hudson JR Jr & Papadopoulos V (1993)
Inhibition of hormone-stimulated steroidogenesis in
cultured Leydig tumor cells by a cholesterol-linked
phosphorothioate oligodeoxynucleotide antisense to
diazepam-binding inhibitor. Proc Natl Acad Sci USA
90, 5728–5731.
29 Papadopoulos V, Amri H, Li H, Boujrad N, Vidic B &
Garnier M (1997) Targeted disruption of the peripheral-
type benzodiazepine receptor gene inhibits steroidogen-
esis in the R2C Leydig tumor cell line. J Biol Chem 272,
32129–32135.
30 Jefcoate C (2002) High-flux mitochondrial cholesterol
trafficking, a specialized function of the adrenal cortex.
J Clin Invest 110, 881–890.
31 Dhariwal MS & Jefcoate CR (1989) Cholesterol meta-
bolism by purified cytochrome P-450scc is highly stimu-
lated by octyl glucoside and stearic acid exclusively in
large unilamellar phospholipid vesicles. Biochemistry 28,
8397–8402.
32 Ascoli M (1981) Characterization of several clonal lines
of cultured Leydig tumor cells: gonadotropin receptors
and steroidogenic responses. Endocrinology 108, 88–95.
33 Leibiger B, Moede T, Schwarz T, Brown GR, Kohler
M, Leibiger IB & Berggren PO (1998) Short-term regu-
lation of insulin gene transcription by glucose. Proc
Natl Acad Sci USA 95, 9307–9312.
Acyl-CoA thioesterase and synthetase P. Maloberti et al.
1814 FEBS Journal 272 (2005) 1804–1814 ª 2005 FEBS