selective activation of peroxisome proliferator-activated receptor (ppar) and ppar induces...
TRANSCRIPT

Selective Activation of PeroxisomeProliferator–Activated Receptor (PPAR)� and PPAR�Induces Neoangiogenesis Through a Vascular EndothelialGrowth Factor–Dependent MechanismFederico Biscetti,
1Eleonora Gaetani,
1Andrea Flex,
1Tamar Aprahamian,
2Teresa Hopkins,
2
Giuseppe Straface,1
Giovanni Pecorini,1
Egidio Stigliano,3
Roy C. Smith,4
Flavia Angelini,1
John J. Castellot, Jr.,4
and Roberto Pola1,4,5
OBJECTIVE—Peroxisome proliferator–activated receptors(PPARs) are therapeutic targets for fibrates and thiazolidinedio-nes, which are commonly used to ameliorate hyperlipidemia andhyperglycemia in type 2 diabetes. In this study, we evaluatedwhether activation of PPAR� and PPAR� stimulates neoangio-genesis.
RESEARCH DESIGN AND METHODS—We used selectivesynthetic PPAR� and PPAR� agonists and investigated theirangiogenic potentials in vitro and in vivo.
RESULTS—Activation of PPAR� and PPAR� leads to endothe-lial tube formation in an endothelial/interstitial cell co-cultureassay. This effect is associated with increased production of theangiogenic cytokine vascular endothelial growth factor (VEGF).Neovascularization also occurs in vivo, when PPAR� and PPAR�agonists are used in the murine corneal angiogenic model. Novascular growth is detectable when PPAR� and PPAR� agonistsare respectively used in PPAR� knockout mice and mice treatedwith a specific PPAR� inhibitor, demonstrating that this angio-genic response is PPAR mediated. PPAR�- and PPAR�-inducedangiogenesis is associated with local VEGF production and doesnot differ in extent and morphology from that induced by VEGF.In addition, PPAR�- and PPAR�-induced in vitro and in vivoangiogenesis may be significantly decreased by inhibiting VEGFactivity. Finally, in corneas treated with PPAR� and PPAR�agonists, there is increased phosphorylation of endothelial nitricoxide synthase and Akt.
CONCLUSIONS—These findings demonstrate that PPAR� andPPAR� activation stimulates neoangiogenesis through a VEGF-dependent mechanism. Neoangiogenesis is a crucial pathological
event in type 2 diabetes. The ability of PPAR� and PPAR�agonists to induce neoangiogenesis might have important impli-cations for the clinical and therapeutic management of type 2diabetes. Diabetes 57:1394–1404, 2008
Peroxisome proliferator-activated receptors(PPARs) are ligand-inducible transcription fac-tors that belong to the nuclear hormone receptorsuperfamily (1). The clinical importance of
PPARs originates with fibrates and thiazolidinediones(TZDs), which respectively act on PPAR� and PPAR� andare used to ameliorate hyperlipidemia and hyperglycemiain subjects with type 2 diabetes. Fibrates (gemfibrozil,clofibrate, fenofibrate, and bezofibrate) are drugs thateffectively reduce triglycerides (TGs) and free fatty acids(FFAs) and increase HDL cholesterol (2–5). Fibrates alsoimprove glucose tolerance in type 2 diabetic patients,although this activity might be attributable to the fact thatsome of these compounds also have potential PPAR�activity (6). TZDs (such as rosiglitazone, troglitazone,pioglitazone, and ciglitazone) are insulin-sensitizing drugsand have constituted a major advance in the recenttherapeutic management of type 2 diabetes (7–9). Inaddition to improving insulin sensitivity, TZDs have alsoeffects on TG, FFA, and ketone body level in severalanimal models of type 2 diabetes. Recently, PPAR�/� dualagonists have also been produced, hypothesizing that thesimultaneous activation of these nuclear receptors mightprovide better glucose and lipid control than single sub-type selective agents (10–13).
In recent years, there has been increasing appreciationof the fact that PPAR� and PPAR� might be involved in themolecular mechanisms that regulate neoangiogenesis, de-fined as the growth of new blood vessels from preexistingvascular networks, through the action of growth factorsand cytokines that stimulate migration, proliferation, andsurvival of endothelial cells. Neoangiogenesis plays a dualrole in type 2 diabetes. On one hand, it is involved in thepathogenesis of diabetic retinopathy and, according tosome studies, contributes to rupture of atheroscleroticplaques in coronary and carotid arteries. On the otherhand, neoangiogenesis is important to promote revascu-larization and contrast ischemia in tissues affected bydiabetic microangiopathy. Therefore, the effects of PPAR�and PPAR� in angiogenesis merit careful evaluation. Sev-eral studies have analyzed the angiogenic properties ofPPAR� and PPAR� using fibrates and TZDs, respectively.
From the 1Laboratory of Vascular Biology and Genetics, Department ofMedicine, A. Gemelli University Hospital, Catholic University School ofMedicine, Rome, Italy; 2Molecular Cardiology, Whitaker Cardiovascular Insti-tute, Boston University School of Medicine, Boston, Massachusetts; the3Department of Pathology, Catholic University School of Medicine, Rome,Italy; the 4Department of Anatomy and Cell Biology, Tufts University Schoolof Medicine, Boston, Massachusetts; and 5Istituto di Ricovero e Cura aCarattere Scientifico Oasi, Troina, Italy.
Corresponding author: Roberto Pola, MD, PhD, Department of Anatomy andCell Biology, Tufts University School of Medicine, 136 Harrison Ave., Boston,MA 02111. E-mail: [email protected].
Received for publication 6 June 2007 and accepted in revised form 24January 2008.
Published ahead of print at http://diabetes.diabetesjournals.org on 11 Feb-ruary 2008. DOI: 10.2337/db07-0765.
eNOS, endothelial nitric oxide synthase; FBS, fetal bovine serum; FFA, freefatty acid; HDMVEC, human dermal microvascular endothelial cell; HUVEC,human umbilical vein endothelial cell; PPAR, peroxisome proliferator–acti-vated receptor; TG, triglyceride; TZD, thiazolidinedione; VEGF, vascularendothelial growth factor.
© 2008 by the American Diabetes Association.The costs of publication of this article were defrayed in part by the payment of page
charges. This article must therefore be hereby marked “advertisement” in accordance
with 18 U.S.C. Section 1734 solely to indicate this fact.
ORIGINAL ARTICLE
1394 DIABETES, VOL. 57, MAY 2008

However, the results of these investigations have beenvery controversial (14–22). In this regard, it is important topoint out that both fibrates and TZDs do not selectively acton PPARs but have pleiotropic activities that occurthrough PPAR-independent pathways. For instance, fi-brates activate other nuclear and superficial receptors(23,24) and also stimulate pathways that do not depend onPPAR� (25,26). Similarly, TZDs act on intracellular mech-anisms regulated by pERK, p38 mitogen-activated proteinkinase, plasminogen activator inhibitor 1, matrix metallo-proteinase-2, Cyclin D1, Bcl-xL/Bcl-2, and tumor necrosisfactor-� in a PPAR�-independent way (27–33). In addition,TZDs are not PPAR� specific, because high concentrationsof these compounds may also activate PPAR� and PPAR�(34). A clear demonstration of the PPAR�-independentproperties of TZDs is provided by the fact that thesemolecules are able to inhibit the release of pro-inflamma-tory mediators in cells that lack the PPAR� gene (35).
In the present study, we used selective PPAR� andPPAR� synthetic agonists and tested their potential abilityto stimulate neoangiogenesis in well-established in vitroand in vivo assays. We found that specific and selectiveactivation of PPAR� and PPAR� leads to increased pro-duction of vascular endothelial growth factor (VEGF), aprototypical angiogenic agent, and formation of endothe-lial tubules when endothelial cells are co-cultured withinterstitial cells. In vivo, PPAR� and PPAR� syntheticagonists stimulate angiogenesis in the mouse cornealneovascularization assay, whereas fibrates and TZDs areunable to induce angiogenesis in the same experimentalsetting. PPAR�- and PPAR�-angiogenic process is associ-ated with increased expression of VEGF and increasedphosphorylation of endothelial nitric oxide (NO) synthase(eNOS) and Akt. Finally, it may be inhibited by blockingVEGF activity.
RESEARCH DESIGN AND METHODS
PPAR-selective agonists. The WY14643 and the GW1929 compounds wereused to specifically activate PPAR� and PPAR�, respectively. GW9662 wasused to inhibit PPAR�. VEGF was used as positive control. All molecules werefrom Sigma.Endothelial cell migration and proliferation assays. Human umbilicalvein endothelial cells (HUVECs) and human dermal microvascular endothelialcells (HDMVECs) (Cambrex) were cultured in EGM2. Migration of endothelialcells was determined using a monolayer denudation assay. Eighty to 90%confluent endothelial cells were scraped with a 1,000-�l pipette tip. Cultureswere washed three times with Hanks’ balanced salt solution and incubatedwith EBM2/0.5% fetal bovine serum (FBS). Control cultures received mediumalone, whereas experimental groups received PPAR agonists at concentra-tions of 1 nmol/l, 10 nmol/l, 100 nmol/l, 1 �mol/l, 5 �mol/l, 10, and 20 �mol/l.VEGF was used as positive control at a concentration of 10 nmol/l. The rateof wound closure was determined by photographing cells at three premarkedareas at time 0 and at 6 and 12 h. Using a digital imaging system, the rate ofcell migration was determined by calculating the difference of the wound areaafter 6 and 12 h, divided by 2. For cell proliferation experiments, endothelialcells were plated into 96-well plates at 1.0 � 105 or 0.5 � 105 cells/well andallowed to attach overnight. Cells were incubated in EBM2/0.5% FBS withPPAR agonists at concentrations of 1 nmol/l, 10 nmol/l, 100 nmol/l, 1 �mol/l,5 �mol/l, 10, and 20 �mol/l. VEGF was used as positive control at aconcentration of 10 nmol/l. After 24, 36, and 48 h, the amount of proliferationwas compared by an MTT assay performed according to manufacturer’sinstructions (American Type Culture Collection). We also evaluated whetherWY14643 and GW1929 were able to increase VEGF levels in endothelial cellsusing a commercially available ELISA kit (R&D Systems). All of the experi-ments were performed in triplicate.Endothelial cell/interstitial cell coculture assay. We used a commerciallyavailable assay (TCS CellWorks), in which proliferating early passage normalhuman endothelial cells are co-cultured with early passage normal humaninterstitial cells in 24-well plates, in a specially formulated culture medium. Inthis assay, cells spontaneously proliferate and then enter a migratory phase,
during which they move through the matrix to form, after 11 days in culture,a network of capillary-like tubules (36). This degree of spontaneous angio-genic activity was evaluated and used as a control to quantify the angiogeniceffects induced by the addition of PPAR agonists. The following concentra-tions of WY14643 and GW1929 were used: 1 nmol/l, 10 nmol/l, 100 nmol/l, and1 �mol/l. VEGF and suramin were used as positive and negative controls,respectively, at the concentration suggested my the manufacturer (TCSCellWorks). On day 11, cells were fixed with ice-cold 70% ethanol, and tubuleformation was visualized by immunostaining for the endothelial cell markerCD31 and quantified by image analysis (TCS AngioSys software), as previouslydescribed (37,38). Number of tubules, total tubule length, and number ofjunctions were calculated in untreated cells and in cells stimulated withPPAR-selective agonists, VEGF, and suramin. The ability of WY14643 andGW1929 to increase VEGF levels was evaluated using a commercially avail-able ELISA kit (R&D Systems). Activity of VEGF was inhibited by adding tothe culture medium a goat anti-human VEGF-neutralizing antibody (R&DSystems) at a concentration of 500 ng/ml, as previously described (39). All ofthe experiments were performed in triplicate. Results were expressed as ratiobetween the angiogenic effect induced by the abovementioned compoundsand that observed in untreated cells. All quantifications were performed in ablinded fashion by two independent operators.Mouse cornea neovascularization assay. Male 8- to 12-week-old C57BL/6Jmice were used for these experiments. Mice were anesthetized with anintramuscular injection of ketamine and killed with an overdose of ketamine.A single corneal pocket was created, and pellets containing 0.3 �g WY14643 orGW1929 were prepared and placed in each corneal pocket, as previouslydescribed (40). Pellets containing 0.3 �g VEGF were used as positive controls.We also implanted pellets containing 0.03, 0.3, and 3 �g gemfibrozil (TevaPharma Italia) or rosiglitazone (GlaxoSmithKline) to perform a direct com-parative analysis between the angiogenic properties of these two classicalPPAR agonists and those of WY14643 and GW1929. Doses were calculated onthe basis of the established ability of gemfibrozil and rosiglitazone to bind,respectively, PPAR� and PPAR� in vitro, compared with WY14643 andGW1929 (41,42). Pellets containing control buffer were used as negativecontrols. Eight to 10 eyes were studied in each group. The investigation wasin accordance with the A. Gemelli University Hospital Institutional AnimalCare and Use Committee. Corneal neoangiogenesis was quantified 6 days afterpellet implantation, as established and described in previous reports (40).Vessel length, circumferential extent of neovascularity, number of vessels percross section, and lumen diameter were evaluated using fluorescence micros-copy. For these analyses, mice received an intracardiac injection of 500 �gendothelial cell marker BS-1 lectin, conjugated to fluorescein isothiocyanate(Vector Laboratories). Thirty minutes later, mice were killed, and each eyewas enucleated and fixed in 1% paraformaldehyde solution for 1 h. Cornealhemispheres were prepared under the dissecting microscope, placed on glassslides, and analyzed using a fluorescent microscope. To verify that theneoangiogenic effects of WY14643 and GW1929 were specifically mediated byPPAR� and PPAR�, respectively, the experiments were repeated in micelacking the PPAR� gene (PPAR��/� mice, homozygous for the Pparatm1Gonzgene) (Jackson Laboratories) and in mice in which PPAR� activity wasblocked by the specific inhibitor GW9662 (Sigma) (43,44). In addition, theability of PPAR��/� mice and mice treated with the PPAR� inhibitor GW9662to respond normally to a prototypical angiogenic stimulus were to tested byimplanting pellets containing VEGF. All quantifications were performed in ablinded fashion, by two independent operators.VEGF expression in mouse cornea. We analyzed whether PPAR�- andPPAR�-induced corneal neoangiogenesis occurred in association with produc-tion of VEGF. Levels of VEGF were studied at the RNA level by RT-PCR andat the protein level by ELISA and immunohistochemistry. For this set ofexperiments, the right corneas were implanted with pellets containingWY14643 or GW1929, whereas the left corneas of the same animals wereimplanted with pellets containing control buffer and served as internalcontrol. For RT-PCR analyses, corneas were harvested 4 days after pelletimplantation, and RNA was extracted as previously established (45). cDNAwas obtained and amplified using the SuperScript preamplification system(Gibco-BRL). Signals were normalized to 18s rRNA using optimal 18S primer–to–competimer ratios as determined for the target gene following the manu-facturer’s recommendations (Ambion). The following primer pairs and PCRconditions were used: 5�-CACATCTGCAAGTACGTTCGTTT-3� and 5�-GTTCA-GAGCGGAGAAAGCATTTG-3� with 30 cycles of 94 (30 s), 62 (1 min), and 72°C(1 min). Results were expressed as mRNA fold increase, calculated as theratio between VEGF mRNA expression in right and left (control) corneas. ForELISA, corneas were harvested 6 days after pellet implantation. VEGF165
levels were measured using a commercially available kit (R&D Systems), aspreviously described (45). Results were expressed as protein fold increase,calculated as the ratio between VEGF protein levels in right and left (control)corneas. For immunohistochemical staining, eyes treated with pellets contain-
F. BISCETTI AND ASSOCIATES
DIABETES, VOL. 57, MAY 2008 1395

ing PPAR� and PPAR� synthetic agonists were excised 6 days after pelletimplantation and fixed in 1% paraformaldehyde solution for 1 h. After fixation,corneas were embedded in paraffin and cut in cross sections. VEGF immuno-staining was performed using a rabbit polyclonal anti-mouse VEGF antibody(Santa Cruz Biotechnology) followed by a biotinylated goat anti-rabbit immu-noglobulin secondary antibody (Signet Labs). Negative control slides wereprepared by substituting preimmune rat serum.Western blotting for phosphorylated eNOS and Akt. For immunoblotting,homogenates of corneal tissues were analyzed. Proteins (40 �g/lane) wereseparated in 10% SDS-polyacrylamide gels and transferred onto polyvinylidinedifluoride membranes. Membranes were incubated with antibodies againsteNOS (1:1,000) (BD Biosciences Pharmingen), phospho-eNOS (Ser1177,1:1,000) (Cell Signaling Technology Company), phospho-Akt (Ser473, 1:500)(Cell Signaling Technology Company), and Akt (1:1,000) (New EnglandBiolabs). Antibody binding was detected with horseradish peroxidase–conju-gated secondary antibodies (1:2,000) (Chemicon) and enhanced chemilumi-nescence system (GE Healthcare Bioscience). Then, the blots were reprobedwith total eNOS (1:1,000) (Transduction Laboratories), Akt (1:5,000) (Sigma),or actin (1:5,000) (Sigma).In vivo inhibition of VEGF activity. Activity of VEGF was systemicallyinhibited in vivo by transfection of mice thigh muscles with a plasmid DNAencoding a soluble form of the murine VEGF receptor Flt-1, as previouslydescribed (45). The plasmid was provided by Dr. Kensuke Egashira. SolubleFlt-1 can suppress VEGF activity both by sequestering VEGF and by function-ing as a dominant-negative inhibitor of VEGF receptors. Control mice receivedan equal amount of empty plasmid with an intramuscular injection on thesame time schedule. Eight eyes were analyzed in each group.Statistical analysis. All results are expressed as means � SE. Differenceswere analyzed by Student’s t test and considered statistically significant at P 0.05.
RESULTS
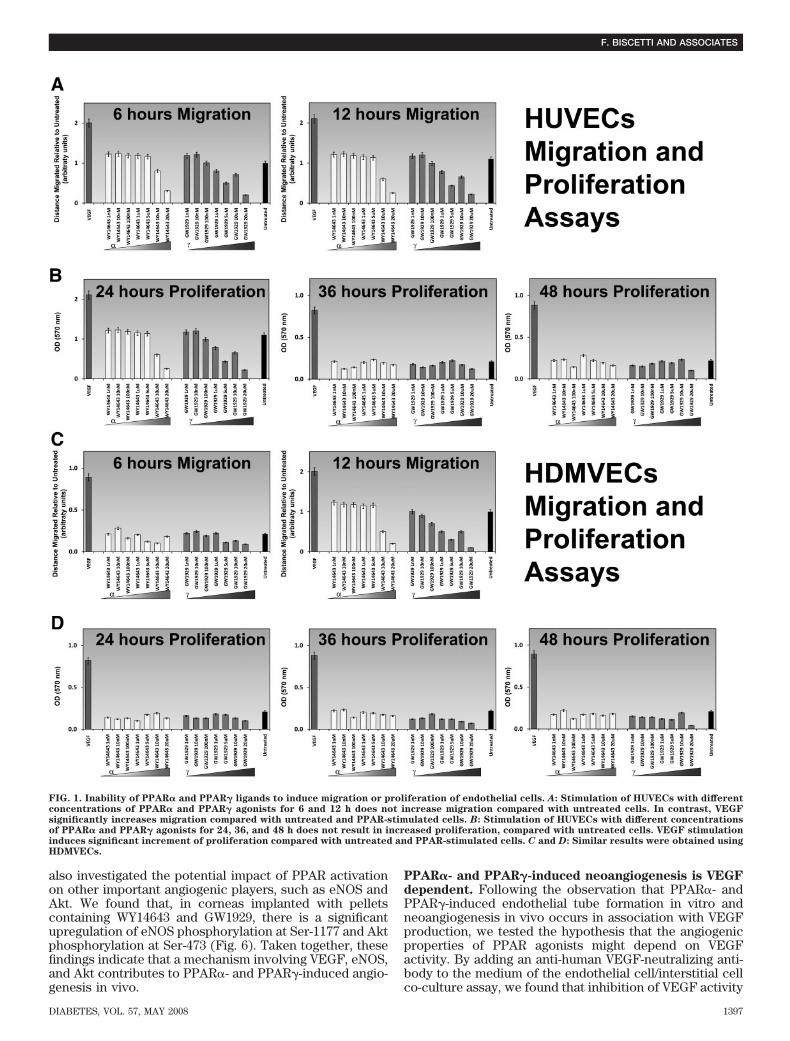
Effects of PPAR� and PPAR� selective agonists onendothelial cell migration and proliferation. We firsttested whether PPAR� and PPAR� activation stimulatesendothelial cell migration or proliferation, two eventsthat are crucial for angiogenesis. No increased migra-tion was seen in HUVECs treated with PPAR agonists(Fig. 1A). Likewise, PPAR agonists were unable toinduce proliferation of HUVECs, compared with control(Fig. 1B). Similar results were obtained using a micro-vascular cell line (HDMVECs) (Fig. 1C and D). PPAR�and PPAR� agonists were also unable to increase VEGFproduction, measured by ELISA, in endothelial cells(data not shown).Effects of PPAR� and PPAR� selective agonists in anendothelial cell/interstitial cell co-culture assay. Forthese experiments, we used a co-culture system contain-ing both endothelial cells and interstitial cells (TCS Cell-Works). In this assay, endothelial cells spontaneouslymigrate and proliferate to form tubule-like CD31-positivestructures (Fig. 2A). Treatment with the anti-angiogenicmolecule suramin resulted in abolishment of tubule for-mation (Fig. 2B). In contrast, treatment with the angio-genic cytokine VEGF led to significant increase of tubuleformation (Fig. 2C). Interestingly, both PPAR� and PPAR�agonists exhibited the ability to strongly increase tubuleformation (Fig. 2D and E). The neoangiogenic propertiesof these compounds were quantified in terms of number oftubules, tubule length, and number of junctions and com-pared with those displayed by untreated cells and VEGF-treated cells (Fig. 2F). PPAR� and PPAR� agonistsinduced significant increase of tubule number, tubulelength, and number of junctions, with the maximum effectbeing observed at the dose of 100 and 10 nmol/l, respec-tively. Protein analyses also demonstrated that, in culturemedia of cells treated with PPAR� and PPAR� agonists,VEGF levels were significantly higher than in untreatedcells (data not shown).
In vivo neongiogenic properties of PPAR� and PPAR�selective agonists. The murine corneal model of angio-genesis was used for these experiments. Pellets containingselective PPAR agonists were implanted in the corneas ofmice, and the resulting neoangiogenic response was ana-lyzed 6 days later. BS1-lectin fluorescent staining demon-strated an important neoangiogenic response in eyestreated with pellets containing 0.3 �g WY14643 or GW1929(Fig. 3A and B). In contrast, no angiogenic response wasobserved using pellets containing gemfibrozil or rosiglita-zone (Fig. 3C and D). As expected, VEGF pellets inducedstrong neoangiogenic growth (Fig. 3E), whereas no angio-genic response was obtained in corneas implanted withpellets containing control buffer (Fig. 3F). To confirm thatthe neoangiogenic effect of WY14643 was mediated byPPAR�, WY14643-containing pellets were implanted in thecorneas of PPAR��/� mice. No angiogenic response wasdetected in these animals (Fig. 3G). Similarly, the PPAR�-specific activity of GW1929 was confirmed by implantingGW1929-containing pellets in the corneas of mice that hadbeen pretreated with the specific PPAR� inhibitorGW9662. No angiogenic response was detected in theseanimals (Fig. 3H). Importantly, both PPAR��/� mice andmice treated with the specific PPAR� antibody respondednormally to stimulation with pellets containing VEGF (Fig.3I and J), demonstrating that these animals do not have anendogenous impairment of angiogenesis.
To quantify the extent of PPAR�- and PPAR�-inducedcorneal angiogenesis, measurements of vessel length, cir-cumferential neovascularity, number of vessels per crosssection, and lumen diameter were carried out. Theseanalyses demonstrated that stimulation of the PPAR� andPPAR� pathways resulted in a neoangiogenic process thatwas statistically similar, in terms of morphological char-acteristics, to that induced by VEGF (Fig. 4). The averagelength of neovessels was 0.73 � 0.11 mm in PPAR�-induced neovascularization, 0.71 � 0.12 mm in PPAR�-induced angiogenesis, and 0.75 � 0.09 mm in the VEGFgroup. Mean lumen diameter was 8.02 � 1.20 �m inPPAR�-induced neovascularization, 7.73 � 1.10 �m in thePPAR� group, and 8.33 � 1.10 �m in the VEGF group.Circumferential extent of corneal neovascularity was102.30 � 7.50° in PPAR�-induced neovascularization,99.10 � 3.90° in the PPAR� group, and 107.20 � 4.40° inthe VEGF group. The number of vessels per cross sectionwas 121.30 � 6.90 in PPAR�-induced neovascularization,112.30 � 7.20 in PPAR�-induced angiogenesis, and131.21 � 7.40 in the VEGF group.PPAR�- and PPAR�-induced neoangiogenesis occursin association with VEGF production and increasedphosphorylation of eNOS and Akt. First, we evaluatedwhether VEGF is expressed in association with PPAR�-and PPAR�-induced neovascularization. RT-PCR andELISA analyses demonstrated that VEGF RNA and proteinlevels are significantly increased in corneas treated withWY14643 and GW1929 (Fig. 5A and B). Second, we per-formed immunohistochemical analysis that showed thatcorneas treated with PPAR agonists were strikingly immu-nopositive for VEGF (Fig. 5C and D). VEGF-producingcells during corneal neovascularization were immunopo-sitive for vimentin (data not shown), a fibroblast mesen-chymal marker, suggesting that PPAR agonists stimulateexpression of endogenous VEGF in interstitial fibroblastswithin the neovascular foci. No VEGF-positive stainingwas seen in corneas implanted with control pellets orwhere control staining was performed (Fig. 5E and F). We
PPARS AND ANGIOGENESIS
1396 DIABETES, VOL. 57, MAY 2008
also investigated the potential impact of PPAR activationon other important angiogenic players, such as eNOS andAkt. We found that, in corneas implanted with pelletscontaining WY14643 and GW1929, there is a significantupregulation of eNOS phosphorylation at Ser-1177 and Aktphosphorylation at Ser-473 (Fig. 6). Taken together, thesefindings indicate that a mechanism involving VEGF, eNOS,and Akt contributes to PPAR�- and PPAR�-induced angio-genesis in vivo.
PPAR�- and PPAR�-induced neoangiogenesis is VEGFdependent. Following the observation that PPAR�- andPPAR�-induced endothelial tube formation in vitro andneoangiogenesis in vivo occurs in association with VEGFproduction, we tested the hypothesis that the angiogenicproperties of PPAR agonists might depend on VEGFactivity. By adding an anti-human VEGF-neutralizing anti-body to the medium of the endothelial cell/interstitial cellco-culture assay, we found that inhibition of VEGF activity
FIG. 1. Inability of PPAR� and PPAR� ligands to induce migration or proliferation of endothelial cells. A: Stimulation of HUVECs with differentconcentrations of PPAR� and PPAR� agonists for 6 and 12 h does not increase migration compared with untreated cells. In contrast, VEGFsignificantly increases migration compared with untreated and PPAR-stimulated cells. B: Stimulation of HUVECs with different concentrationsof PPAR� and PPAR� agonists for 24, 36, and 48 h does not result in increased proliferation, compared with untreated cells. VEGF stimulationinduces significant increment of proliferation compared with untreated and PPAR-stimulated cells. C and D: Similar results were obtained usingHDMVECs.
F. BISCETTI AND ASSOCIATES
DIABETES, VOL. 57, MAY 2008 1397

was sufficient to significantly decrease PPAR�- andPPAR�-induced tubulogenesis (Fig. 7A and B). In addition,we suppressed VEGF activity in vivo and evaluatedwhether pellets containing WY14643 or GW1929 were stillable to induce corneal angiogenesis. In vivo inhibition ofVEGF was accomplished using the soluble Flt1 plasmid,which suppresses VEGF activity both by sequestering
VEGF and by functioning as a dominant-negative inhibitorof VEGF receptors (45). Mice transfected with the emptyplasmid were used as controls. A dramatic reduction ofPPAR�- and PPAR�-induced corneal neoangiogenesis wasobserved when VEGF activity was suppressed (Fig. 7C–F).Quantification analyses demonstrated that inhibition ofVEGF activity resulted in statistically significant reduction
FIG. 2. PPAR� and PPAR� ligands induce tube formation in an endothelial cell/interstitial cell co-culture assay. A: Immunostaining with theendothelial cell marker CD31 demonstrates the ability of untreated cells to spontaneously form tubules. B: In cells treated with suramin, tubuleformation is greatly reduced. C: In contrast, tubule formation is importantly increased in cells stimulated with VEGF. Similarly, treatment of cellswith WY14643 (100 nmol/l) (D) and GW1929 (10 nmol/l) (E) results in conspicuous enhancement of tubule formation. F: Quantification of tubeformation shows that compared with untreated cells, number of tubules, total tubule length, and number of junctions are significantly increasedin cells treated with VEGF, WY14643, and GW1929. The ratio between the number of tubules detectable in the VEGF group and that observed inuntreated cells was 10.40 � 0.52 (P < 0.001). The ratio between the total tubule length in the VEGF group and in untreated cells was 16.22 �0.81 (P < 0.001). The ratio between the number of junctions in the VEGF group and in untreated cells was 29.71 � 1.48 (P < 0.001). Also cellsstimulated with WY14643 showed significant increments of tubule number, tubule length, and junction number, compared with untreated cells.WY14643 exhibited its maximum pro-angiogenic effect at the concentration of 100 nmol/l with ratios of 18.75 � 0.93, 9.43 � 0.47, and 23.89 � 1.19for tubule number, tubule length, and number of junctions, respectively (P < 0.001). Likewise, cells stimulated with GW1929 showed significantincrements of all of the investigated parameters, compared with untreated cells. The maximum effect was observed at the concentration of 10nmol/l, with a tubule ratio of 21.28 � 1.06 (P < 0.001), a length ratio of 16.00 � 0.8 (P < 0.001), and a junction ratio of 70.00 � 3.5 (P < 0.001).
PPARS AND ANGIOGENESIS
1398 DIABETES, VOL. 57, MAY 2008

of PPAR�- and PPAR�-induced neoangiogenesis, mea-sured in terms of vessel length, circumferential extent ofneovascularity, and number of vessels per cross section(Fig. 7G). These findings demonstrate that selectivePPAR� and PPAR� agonists induce angiogenesis via aVEGF-dependent mechanism.
DISCUSSION
In recent years, the angiogenic potentials of TZDs havebeen intensively investigated. Some studies have reportedthat ciglitazone, troglitazone, and rosiglitazone suppressmigration, proliferation, and differentiation of endothelialcells and inhibit angiogenesis in a number of experimentalmodels (17–21). Other studies have instead provided evi-dence that the same molecules may have important pro-angiogenic effects. In particular, it has been shown thatTZDs are able to increase VEGF expression in smoothmuscle vascular cells and macrophages (14,15), that ros-iglitazone increases number and migratory activity ofendothelial progenitor cells (16) and promotes angiogen-esis after focal cerebral ischemia in rats (22), and thattroglitazone induces the expression of VEGF and itsreceptors in cultured cardiac myofibroblasts (35). In all ofthese studies, the authors have assumed that the effectsinduced by TZDs were mediated by the activation ofPPAR� without considering that TZDs are nonselectiveand nonspecific ligands of this nuclear receptor, becausethey are able to stimulate several PPAR�-independentpathways that are potentially important in angiogenesis(27–33,46). Similarly, the role of PPAR� in angiogenesishas been investigated using fibrates, which not only acti-vate PPAR�, but also bind other nuclear and superficialreceptors and regulate important PPAR�-independentpathways (23–26). Therefore, to conclude that the effectsof TZDs and fibrates on angiogenesis specifically dependon PPAR, all of the potential activities that these mole-cules exert on pathways that are PPAR independentshould be excluded.
In this study, we used selective PPAR� and PPAR�
synthetic agonists and found that they were able toenhance endothelial tube formation in vitro and induceneovascularization in vivo. We also demonstrated thatthese effects were specifically PPAR mediated, by showingthat the PPAR� activator WY14643 was unable to induceangiogenesis in mice lacking the PPAR� gene and that thePPAR� activator GW1929 did not stimulate angiogenesis inmice treated with a PPAR�-specific inhibitor. Importantly,PPAR��/� mice and mice treated with the PPAR� inhibi-tor showed a normal response on stimulation with VEGF,demonstrating that these animals do not have an endoge-nous impairment of angiogenesis and providing additionalconfirmation that WY14643 and GW1929 inability to induceangiogenesis in these mice depends on PPAR lack offunction. Another interesting finding was that gemfibroziland rosiglitazone, two prototypical fibrates and TZDs thatare generally viewed as classical PPAR agonists and havebeen widely used to respectively test the biological effectsof PPAR� and PPAR�, were unable to induce angiogenesisin our experimental setting. This finding might help us tounderstand the controversial results in the literature aboutthe angiogenic properties of PPARs. In this respect, it isimportant to point out that this is the first time that theangiogenic effects of TZDs and fibrates are directly com-pared with those of synthetic PPAR-specific agonists, suchas WY14643 and GW1929. These data clearly eliminate thepossibility that our observations differ from those of otherresearch groups as a result of different experimentalsettings and/or surgical procedures. They instead supportthe concept that fibrates and TZDs cannot be consideredspecific and selective PPAR agonists and that not all oftheir biological activities can be attributed to activation ofPPARs.
Another important finding of this study is the demon-stration that PPAR�- and PPAR�-induced angiogenesisdoes not occur through direct stimulation of endothelialcell migration or proliferation. In contrast, PPAR� andPPAR� agonists stimulate angiogenesis indirectly, throughupregulation of the angiogenic cytokine VEGF. The in vivo
FIG. 3. PPAR� and PPAR� agonists induce angiogenesis in vivo. BS1 lectin whole-mount staining in corneas in which pellets containing 0.3 �gWY14643 (A), 0.3 �g GW1929 (B), 3 �g gemfibrozil (C), 3 �g rosiglitazone (D), 0.3 �g VEGF (E), or control buffer (F) were implanted. G: Pelletscontaining WY14643 are unable to induce corneal angiogenesis in PPAR��/� mice. H: Pellets containing GW1929 are unable to induce cornealangiogenesis in mice pretreated with the PPAR� inhibitor GW9662. VEGF induces a normal angiogenic response in PPAR��/� mice (I) and in micepretreated with the PPAR� inhibitor GW9662 (J). (Please see http://dx.doi.org/10.2337/db07-0765 for a high-quality digital representation of thisimage.)
F. BISCETTI AND ASSOCIATES
DIABETES, VOL. 57, MAY 2008 1399

neovascularization resulting from the activation of PPAR�and PPAR� is morphologically similar to that induced byVEGF and is associated with local VEGF production. Mostimportantly, suppression of VEGF activity is sufficient toinhibit PPAR�- and PPAR�-induced angiogenesis both invitro and in vivo. However, it is important to note thatVEGF inhibition does not completely suppress PPAR�-and PPAR�-induced neovascularization. Therefore, it ispossible to speculate that additional cytokines might con-tribute to the angiogenic process stimulated by PPARs. Inthis scenario, we also show that PPAR�- and PPAR�-induced neovascularization occurs in association withenhanced phosphorylation of eNOS and Akt. This findingis important because NO and VEGF display reciprocalregulatory activities that are crucial for angiogenesis, withthe fundamental contribution of Akt. On one hand, VEGFmay activate eNOS by activating Akt. On the other hand,NO can mediate VEGF expression by stimulating hypoxia-inducible factors and heme oxygenase-1 activity. Althoughthe precise molecular mechanisms responsible for PPAR-induced angiogenesis still merit further investigation, ourdata provide new insights to understanding the ability ofPPARs to modulate the angiogenic process in vivo.
The discovery that both PPAR� and PPAR� are ableto induce angiogenesis also suggests the possible exis-
tence of a synergistic interaction between the tworeceptors, although we have no data regarding theirpossible relative roles in the activation of VEGF and/orother angiogenic molecules. This concept might beparticularly important when considering the biologicalactivities of PPAR�/� dual agonists. In addition toreducing glucose levels and improving the lipid profile,these molecules also display beneficial activities oncardiac and endothelial function, oxidative stress, andatherosclerosis (47). It is possible to hypothesize thatthese effects might partially result from the ability ofPPAR�/� dual agonists to induce angiogenesis andVEGF production. Similar mechanisms might be hypoth-esized to explain the potential carcinogenic effects ofsome PPAR�/� dual agonists (48) and their ability toinduce edema and precipitate heart failure (49,50).
Although our data need to be confirmed in other exper-imental models and in humans, the observation that neo-angiogenesis is a potential biological result of PPARactivation might help to understand, at least in part, someof the biological and clinical effects of TZDs and fibrates.For instance, PPAR�-induced VEGF upregulation might beresponsible for the recently reported ability of rosiglita-zone to ameliorate endothelial dysfunction in type 2diabetes (51) and increase number and migratory activity
0.0
0.3
0.6
0.9
VEGF
mm
Vessel Length
p = n. s.
0
30
60
90
120
deg
rees
Circumferential Neovascularity
p = n. s.
0
30
60
90
120
150
vess
els/
cro
ss s
ecti
on
Number of Vessels
p = n. s.
0
3
6
9
12m
icro
met
ers
Lumen Diameter
p = n. s.
VEGF PPAR- PPAR-
VEGF PPAR- PPAR- VEGF PPAR- PPAR-
PPAR- PPAR-
FIG. 4. Quantification of corneal angiogenesis induced by PPAR agonists. Vessel length, lumen diameter, circumferential extent of neovascularity,and number of vessels per cross section are not statistically different between corneal angiogenesis induced by WY14643, GW1929, and VEGF.
PPARS AND ANGIOGENESIS
1400 DIABETES, VOL. 57, MAY 2008

of endothelial progenitor cells (16). Because VEGF aug-ments vascular permeability (52), increased production ofthis cytokine might also contribute to edema, a commonside effects of TZDs. It is also possible that PPAR�-inducedneoangiogenesis has deleterious effects in subjects withtype 2 diabetes. Although angiogenic therapy has beenwidely regarded as an attractive approach both for treatingischemic heart disease and for enhancing arterioprotectivefunctions of the endothelium, a variety of studies have alsosuggested that neovascularization contributes to thegrowth of atherosclerotic lesions and is a key factor inplaque destabilization and rupture (53). These negativeeffects might contribute to the recently reported poten-tial adverse cardiovascular effects of rosiglitazone intype 2 diabetes (54). Similarly, some clinical activities offibrates, such as preservation of renal function and ame-lioration of endothelial dysfunction (55,56), might be par-tially explained by PPAR�-induced angiogenesis andVEGF production.
FIG. 5. Angiogenesis induced by PPAR agonists is associated with local VEGF production. A: RT-PCR for mouse VEGF shows significantlyincreased RNA levels in corneas stimulated with pellets containing WY14643 and GW1929. B: VEGF protein levels, measured by ELISA, aresignificantly increased in corneas stimulated with pellets containing WY14643 and GW1929. Immunohistochemistry for mouse VEGF in 5-�msections shows VEGF-positive staining (red staining) in the neovascular area of corneas stimulated with pellets containing WY14643 (C) andGW1929 (D). No staining is present in corneas in which control pellets were implanted (E) and in sections in which control staining wasperformed (F). (Please see http://dx.doi.org/10.2337/db07-0765 for a high-quality digital representation of this image.)
mouse 1
contro
l
contro
l
contro
l
WY14
643
WY14
643
GW19
29
contro
l
GW19
29
mouse 2 mouse 3 mouse 4
eNOS
eNOS Ser1177
Akt
p-Akt Ser473
actin
FIG. 6. Angiogenesis induced by PPAR agonists is associated withincreased phosphorylation of eNOS and Akt. Western blotting analysisshows increased phosphorylation of eNOS Ser1177 and Akt Ser473 incorneas implanted with pellets containing WY14643 and GW1929.
F. BISCETTI AND ASSOCIATES
DIABETES, VOL. 57, MAY 2008 1401

In conclusion, we used selective synthetic agonists ofPPAR� and PPAR� and demonstrated that the stimula-tion of these nuclear receptors results in the activationof a strong neoangiogenic process in vitro and in vivo.This angiogenic response does not occur through directstimulation of endothelial cells but is instead dependent
on VEGF activity. These findings provide new informa-tion to understand the biological, clinical, and therapeu-tic effects of drugs that stimulate the activity of PPAR�and PPAR�, with potentially important implicationsfor the management of subjects affected by type 2diabetes.
FE
C D
BA
G
FIG. 7. Angiogenesis induced by PPAR agonists is inhibited by suppression of VEGF activity. CD31 immunostaining shows that treatment with aVEGF-neutralizing antibody significantly reduces WY14643-induced (A) and GW1929-induced (B) tubulogenesis in the endothelial cell/interstitial cell co-culture assay. BS-1 lectin fluorescent staining shows that compared with mice transfected with empty plasmid (C and D),corneal angiogenesis induced by PPAR� and PPAR� ligands is inhibited in mice transfected with soluble Flt-1 vector (E and F). G: Quantificationof corneal vascularization in the two groups. (Please see http://dx.doi.org/10.2337/db07-0765 for a high-quality digital representation of thisimage.)
PPARS AND ANGIOGENESIS
1402 DIABETES, VOL. 57, MAY 2008

REFERENCES
1. Kota BP, Huang THW, Roufgalis BD: An overview on biological mecha-nisms of PPARs. Pharm Res 51:85–94, 2005
2. Lefebvre AM, Peinado-Onsurbe J, Leitersdorf I, Briggs MR, Paterniti JR,Fruchart JC, Fievet C, Auwerx J, Staels B: Regulation of lipoproteinmetabolism by thiazolidinediones occurs through a distinct but comple-mentary mechanism relative to fibrates. Arterioscler Thromb Vasc Biol
17:1756–1764, 19973. Staels B, Dallongeville J, Auwerx J, Schoonjans K, Leitersdorf E, Fruchart
JC: Mechanism of action of fibrates on lipid and lipoprotein metabolism.Circulation 98:2088–2093, 1998
4. Elisaf M: Effects of fibrates on serum metabolic parameters. Curr Med Res
Opin 18:269–276, 20025. Devroey D, Velkeniers B, Duquet W, Betz W: The benefit of fibrates in the
treatment of ‘bad HDL-C responders to statins’. Int J Cardiol 101:231–235,2005
6. Jones AB: Peroxisome proliferator-activated receptor (PPAR) modulators:diabetes and beyond. Med Res Rev 21:540–552, 2001
7. Saltiel AR, Olefsky JM: Thiazolidinediones in the treatment of insulinresistance and type II diabetes. Diabetes 45:1661–1669, 1996
8. Takano H, Hasegawa H, Zou Y, Komuro I: Pleiotropic actions of PPARgamma activators thiazolidinediones in cardiovascular diseases. Curr
Pharm Des 10:2779–2786, 20049. Dumasia R, Eagle KA, Kline-Rogers E, May N, Cho L, Mukherjee D: Role of
PPARgamma agonist thiazolidinediones in treatment of pre-diabetic anddiabetic individuals: a cardiovascular perspective. Curr Drug Targets
Cardiovasc Haematol Disord 5:377–386, 200510. Desai RC, Han W, Metzger EJ, Bergman JP, Gratale DF, MacNaul KL,
Berger JP, Doebber TW, Leung K, Moller DE, Heck JV, Sahoo SP: 5-Arylthiazolidine-2,4-diones: discovery of PPAR dual alpha/gamma agonists asantidiabetic agents. Bioorg Med Chem Lett 13:2795–2798, 2003
11. Desai RC, Gratale DF, Han W, Koyama H, Metzger E, Lombardo VK,MacNaul KL, Doebber TW, Berger JP, Leung K, Franklin R, Moller DE,Heck JV, Sahoo SP: Aryloxazolidinediones: identification of potent orallyactive PPAR dual alpha/gamma agonists. Bioorg Med Chem Lett 13:3541–3544, 2003
12. Binggell A, Boehringer M, Grether U. The use of PPAR-alpha/gamma dualagonists for the treatment of metabolic syndromes: Expert Opin Ther
Patents 14:1797–1801, 200413. Nissen SE, Wolski K, Topol EJ: Effect of muraglitazar on death and major
adverse cardiovascular events in patients with type 2 diabetes mellitus.JAMA 294:2581–2586, 2005
14. Yamakawa K, Hosoi M, Koyama H, Tanaka S, Fukumoto S, Morii H,Nishizawa Y: Peroxisome proliferator-activated receptor-gamma agonistsincrease vascular endothelial growth factor expression in human vascularsmooth muscle cells. Biochem Biophys Res Commun 271:571–574, 2000
15. Jozkowicz A, Dulak J, Piatkowska E, Placha W, Dembinska-Kiec A:Ligands of peroxisome proliferator-activated receptor-gamma increase thegeneration of vascular endothelial growth factor in vascular smoothmuscle cells and in macrophages. Acta Biochim Pol 47:1147–1157,2000
16. Pistrosch F, Herbrig K, Oelschlaegel U, Richter S, Passauer J, Fischer S,Gross P: PPARgamma-agonist rosiglitazone increases number and migra-tory activity of endothelial progenitor cells. Atherosclerosis 183:163–167,2005
17. Xin X, Yang S, Kowalski J, Gerritsen ME: Peroxisome proliferator-acti-vated receptor gamma ligands are potent inhibitors of angiogenesis in vitroand in vivo. J Biol Chem 274:9116–9121, 1999
18. Panigrahy D, Singer S, Shen LQ, Butterfield CE, Freedman DA, Chen EJ,Moses MA, Kilroy S, Duensing S, Fletcher C, Fletcher JA, Hlatky L,Hahnfeldt P, Folkman J, Kaipainen A: PPARgamma ligands inhibit primarytumor growth and metastasis by inhibiting angiogenesis. J Clin Invest
110:923–932, 200219. Murata T, He S, Hangai M, Ishibashi T, Xi XP, Kim S, Hsueh WA, Ryan SJ,
Law RE, Hinton DR: Peroxisome proliferator-activated receptor-gammaligands inhibit choroidal neovascularization. Invest Ophthalmol Vis Sci
41:2309–2317, 200020. Peeters LL, Vigne JL, Tee MK, Zhao D, Waite LL, Taylor RN: PPARgamma
represses VEGF expression in human endometrial cells: implications foruterine angiogenesis. Angiogenesis 8:373–379, 2005
21. Sheu WH, Ou HC, Chou FP, Lin TM, Yang CH: Rosiglitazone inhibitsendothelial proliferation and angiogenesis. Life Sci 78:1520–1528, 2006
22. Chu K, Lee ST, Koo JS, Jung KH, Kim EH, Sinn DI, Kim JM, Ko SY, Kim SJ,Song EC, Kim M, Roh JK: Peroxisome proliferator-activated receptor-gamma-agonist, rosiglitazone, promotes angiogenesis after focal cerebralischemia. Brain Res 1093:208–218, 2006
23. Vu-Dac N, Schoonjans K, Laine B, Fruchart JC, Auwerx J, Staels B:Negative regulation of the human apolipoprotein A-I promoter by fibratescan be attenuated by the interaction of the peroxisome proliferator-activated receptor with its response element. J Biol Chem 269:31012–31018, 1994
24. Scatena R, Bottoni P, Vincenzoni F, Messana I, Martorana GE, Nocca G, DeSole P, Maggiano N, Castagnola M, Giardina B: Bezafibrate induces amitochondrial derangement in human cell lines: a PPAR-independentmechanism for a peroxisome proliferator. Chem Res Toxicol 16:1440–1447, 2003
25. Selim E, Frkanec JT, Cunard R: Fibrates upregulate TRB3 in lymphocytesindependent of PPARalpha by augmenting CCAAT/enhancer-binding pro-teinbeta (C/EBPbeta) expression. Mol Immunol 44:1218–1229, 2007
26. Zhang X, Tanaka N, Nakajima T, Kamijo Y, Gonzalez FJ, Aoyama T:Peroxisome proliferator-activated receptor alpha-independent peroxisomeproliferation. Biochem Biophys Res Commun 346:1307–1311, 2006
27. Liu HB, Hu YS, Medcalf RL, Simpson RW, Dear AE: Thiazolidinedionesinhibit TNFalpha induction of PAI-1 independent of PPARgamma activa-tion. Biochem Biophys Res Commun 334:30–37, 2005
28. Turturro F, Oliver R, 3rd, Friday E, Nissim I, Welbourne T: Troglitazoneand pioglitazone interactions via PPAR-gamma-independent and -depen-dent pathways in regulating physiological responses in renal tubule-derived cell lines. Am J Physiol Cell Physiol 292:C1137–C1146, 2007
29. Shiau CW, Yang CC, Kulp SK, Chen KF, Chen CS, Huang JW, Chen CS:Thiazolidenediones mediate apoptosis in prostate cancer cells in partthrough inhibition of Bcl-xL/Bcl-2 functions independently of PPAR-gamma. Cancer Res 65:1561–1569, 2005
30. Galli A, Ceni E, Crabb DW, Mello T, Salzano R, Grappone C, Dilani S,Surreni E, Surreni C, Casini A Antidiabetic thiazolidinediones inhibitinvasiveness of pancreatic cancer cells via PPARgamma independentmechanisms. Gut 53:1688–1697, 2004
31. Gardner OS, Shiau CW, Chen CS, Graves LM: Peroxisome proliferator-activated receptor gamma-independent activation of p38 MAPK by thiazo-lidinediones involves calcium/calmodulin-dependent protein kinase II andprotein kinase R. J Biol Chem 280:10109–10118, 2005
32. He G, Thuillier P, Fischer W, Fischer S: Troglitazone inhibits Cyclin D1expression and cell cycling independently of PPARgamma in normalmouse skin keratinocytes. J Invest Dermatol 123:1110–1119, 2004
33. Huang JW, Shiau CW, Yang YT, Kulp SK, Chen KF, Brueggemeier RW,Shapiro CL, Chen CS: Peroxisome proliferator-activated receptor gamma-independent ablation of cyclin D1 by thiazolidinediones and their deriva-tives in breast cancer cells. Mol Pharmacol 67:1342–1348, 2005
34. Kim EH, Na HK, Surh YJ: Upregulation of VEGF by 15-deoxy-Delta12,14-prostaglandin J2 via heme oxygenase-1 and ERK1/2 signaling in MCF-7cells. Ann N Y Acad Sci 1090:375–384, 2006
35. Chintalgattu V, Harris GS, Akula SM, Katwa LC: PPAR-gamma agonistsinduce the expression of VEGF and its receptors in cultured cardiacmyofibroblasts. Cardiovasc Res 74:140–150, 2007
36. Donovan D, Brown NJ, Bishop ET, Lewis CE: Comparison of three in vitrohuman angiogenesis assays with capillaries formed in vivo. Angiogenesis
4:113–121, 200137. Patel HB, Nasir FA, Nash GF, Scully MF, Kakkar AK: Enhanced angiogen-
esis following allogeneic blood transfusion. Clin Lab Haematol 26:129–135, 2004
38. Bingle L, Lewis CE, Corke KP, Reed MW, Brown NJ: Macrophages promoteangiogenesis in human breast tumour spheroids in vivo. Br J Cancer
94:101–107, 200639. Saito M, Hamasaki M, Shibuya M: Induction of tube formation by angio-
poietin-1 in endothelial cell/fibroblast co-culture is dependent on endoge-nous VEGF. Cancer Sci 94:782–790, 2003
40. Pola R, Ling LE, Silver M, Corbley MJ, Kearney M, Blake Pepinsky R,Shapiro R, Taylor FR, Baker DP, Asahara T, Isner JM: The morphogenSonic hedgehog is an indirect angiogenic agent upregulating two familiesof angiogenic growth factors. Nat Med 7:706–711, 2001
41. Mukherjee R, Sun S, Santomenna L, Miao B, Walton H, Liao B, Locke K,Zhang JH, Nguyen SH, Zhang LT, Murphy K, Ross HO, Xia MX, Teleha C,Chen SY, Selling B, Wynn R, Burn T, Young PR: Ligand and coactivatorrecruitment preferences of peroxisome proliferator activated receptoralpha. J Steroid Biochem Mol Biol 81:217–225, 2002
42. Way JM, Gorgun CZ, Tong Q, Uysal KT, Brown KK, Harrington WW, OliverWR Jr, Willson TM, Kliewer SA, Hotamisligil GS: Adipose tissue resistinexpression is severely suppressed in obesity and stimulated by peroxisomeproliferator-activated receptor gamma agonists. J Biol Chem 276:25651–25653, 2001
43. Seargent JM, Yates EA, Gill JH: GW9662, a potent antagonist of PPAR-gamma, inhibits growth of breast tumour cells and promotes the antican-
F. BISCETTI AND ASSOCIATES
DIABETES, VOL. 57, MAY 2008 1403

cer effects of the PPARgamma agonist rosiglitazone, independently ofPPARgamma activation. Br J Pharmacol 143:933–937, 2004
44. Collino M, Patel NS, Lawrence KM, Collin M, Latchman DS, Yaqoob MM,Thiemermann C: The selective PPARgamma antagonist GW9662 reversesthe protection of LPS in a model of renal ischemia-reperfusion. Kidney Int
68:529–536, 200545. Pola R, Gaetani E, Flex A, Aprahamian TR, Bosch-Marce M, Losordo DW,
Smith RC, Pola P: Comparative analysis of the in vivo angiogenic proper-ties of stable prostacyclin analogs: a possible role for peroxisome prolif-erator-activated receptors. J Mol Cell Cardiol 36:363–370, 2004
46. Phulwani NK, Feinstein DL, Gavrilyuk V, Akar C, Kielian T: 15-deoxy-Delta12,14-prostaglandin J2 (15d-PGJ2) and ciglitazone modulate Staphy-
lococcus aureus dependent astrocyte activation primarily through a PPAR-gamma-independent pathway. J Neurochem 99:1389–1402, 2006
47. Balakumar P, Rose M, Ganti SS, Krishan P, Singh M: PPAR dual agonists:are they opening Pandora’s Box? Pharmacol Res 56:91–98, 2007
48. Hellmold H, Zhang H, Andersson U, Blomgren B, Holland T, Berg AL,Elebring M, Sjogren N, Bamberg K, Dahl B, Westerberg R, Dillner B,Tugwood J, Roberts R, Lundholm E, Camejo G, Skanberg I, Evans J:Tesaglitazar, a PPAR{alpha}/{gamma} agonist, induces interstitial mesen-chymal cell DNA synthesis and fibrosarcomas in subcutaneous tissues inrats. Toxicol Sci 98:63–74, 2007
49. Kendall DM, Rubin CJ, Mohideen P, Ledeine JM, Belder R, Gross J,Norwood P, O’Mahony M, Sall K, Sloan G, Roberts A, Fiedorek FT,DeFronzo RA: Improvement of glycemic control, triglycerides, and HDLcholesterol levels with muraglitazar, a dual (�/�) peroxisome proliferator-activated receptor activator, in patients with type 2 diabetes inadequately
controlled with metformin monotherapy: a double-blind, randomized,pioglitazone-comparative study. Diabetes Care 29:1016–1023, 2006
50. Mittra S, Sangle G, Tandon R, Sharma S, Roy S, Khanna V, Gupta A,Sattigeri J, Sharma L, Priyadarsiny P, Khattar SK, Bora RS, Saini KS, BansalVS: Increase in weight induced by muraglitazar, a dual PPARalpha/gammaagonist, in db/db mice: adipogenesis/or oedema? Br J Pharmacol 150:480–487, 2007
51. Pistrosch F, Passauer J, Fischer S, Fuecker K, Hanefeld M, Gross P: In type2 diabetes, rosiglitazone therapy for insulin resistance ameliorates endo-thelial dysfunction independent of glucose control. Diabetes Care 27:484–490, 2004
52. Senger DR, Van de Water L, Brown LF, Nagy JA, Yeo KT, Yeo TK, Berse B,Jackman RW, Dvorak AM, Dvorak HF: Vascular permeability factor (VPF,VEGF) in tumor biology. Cancer Metastasis Rev 12:303–324, 1993
53. Khurana R, Simons M, Martin JF, Zachary IC: Role of angiogenesis incardiovascular disease: a critical appraisal. Circulation 112:1813–1824,2005
54. Nissen SE, Wolski K: Effect of rosiglitazone on the risk of myocardialinfarction and death from cardiovascular causes. N Engl J Med 356:2457–2471, 2007
55. Jackson TC, Mi Z, Bastacky SI, McHale T, Melhem MF, Sonalker PA,Tofovic SP, Jackson EK: PPAR alpha agonists improve renal preservationin kidneys subjected to chronic in vitro perfusion: interaction withmannitol. Transpl Int 20:277–290, 2007
56. Skrha J, Stulc T, Hilgertova J, Weiserova H, Kvasnicka J, Ceska R: Effect ofsimvastatin and fenofibrate on endothelium in type 2 diabetes. Eur
J Pharmacol 493:183–189, 2004
PPARS AND ANGIOGENESIS
1404 DIABETES, VOL. 57, MAY 2008