rôle des interneurones corticaux parvalbuminergiques dans
TRANSCRIPT

Année 2013 Thèse n°2045
THÈSE
pour le
DOCTORAT DE L’UNIVERSITÉ BORDEAUX 2
Ecole doctorale Sciences de la Vie et de la Santé
Mention : Sciences, Technologie, Santé
Spécialité : Neurosciences
Présentée et soutenue publiquement
Le 13 Décembre 2013
Par Julien Courtin
Rôle des interneurones corticaux
parvalbuminergiques dans les
comportements de peur
Membres du Jury
M. Thomas Boraud (DR-CNRS Bordeaux) ............................................ Président
Mme Thérèse Jay (DR-INSERM Paris) .................................................. Rapporteur
M. Bruno Poucet (DR-CNRS Aix-Marseille) ......................................... Rapporteur
M. Karim Benchenane (CR1-CNRS Paris) ............................................. Examinateur
M. François Georges (CR1-CNRS Bordeaux) ........................................ Examinateur
M. Cyril Herry (CR1-INSERM Bordeaux) ............................................. Directeur de thèse

Année 2013 Thèse n°2045
THÈSE
pour le
DOCTORAT DE L’UNIVERSITÉ BORDEAUX 2
Ecole doctorale Sciences de la Vie et de la Santé
Mention : Sciences, Technologie, Santé
Spécialité : Neurosciences
Présentée et soutenue publiquement
Le 13 Décembre 2013
Par Julien Courtin
Role of cortical parvalbumin interneurons
in fear behaviour
Membres du Jury
M. Thomas Boraud (DR-CNRS Bordeaux) ............................................ Président
Mme Thérèse Jay (DR-INSERM Paris) .................................................. Rapporteur
M. Bruno Poucet (DR-CNRS Aix-Marseille) ......................................... Rapporteur
M. Karim Benchenane (CR1-CNRS Paris) ............................................. Examinateur
M. François Georges (CR1-CNRS Bordeaux) ........................................ Examinateur
M. Cyril Herry (CR1-INSERM Bordeaux) ............................................. Directeur de thèse

REMERCIEMENTS
L’ensemble des travaux de thèse présentés dans ce manuscrit ont été réalisés au sein
du Neurocentre Magendie, dans le laboratoire INSERM U862 du docteur Pier-Vincenzo
Piazza sous la direction du docteur Cyril Herry.
Il m’est tout naturel de commencer ces remerciements en exprimant toute ma gratitude
à Cyril Herry. Je souhaite que le travail présenté ici soit à la hauteur des qualités
incontestables de la formation que j’ai reçue à tes côtés. Je te remercie pour ta grande
disponibilité et pour la grande part de liberté que tu m’as laissée au cours de ces trois années
d’encadrement. J’ai pu ainsi bénéficier de ta confiance et de tes qualités scientifiques et
humaines. Travailler avec toi a été un plaisir et j’espère que nos relations perdureront.
Je remercie également le docteur Pier-Vincenzo Piazza de m’avoir accueilli au sein de
l’institut qui a été pour moi un lieu propice à la réalisation de cette thèse.
Je souhaiterais exprimer ma reconnaissance à ceux qui ont accepté de consacrer du
temps pour étudier ce manuscrit. Je remercie sincèrement les docteurs Thérèse Jay et Bruno
Poucet d’avoir accepté d’évaluer mon travail en tant que rapporteurs. Je remercie le docteur
Thomas Boraud de présider le jury de cette thèse ainsi que les docteurs Karim Benchenane et
François Georges qui ont bien voulu examiner mon travail.
Je remercie également l’ensemble des membres de notre équipe, tant pour leurs
qualités personnelles que professionnelles. Plus que des collègues, certains sont mêmes
devenus des amis. Je remercie particulièrement Cécilia, Fabrice, Robert, Nikolas, Hélène et
Thomas pour leur aide. Je n’aurais jamais pu publier aussi bien mes recherches sans vous.
J’ai une pensée toute particulière pour le docteur François Georges qui m’a accueilli
dans son équipe pour mon stage de Master 1. Malgré le peu de temps que nous avons passé
ensemble, tu m’as permis de confirmer mon intérêt pour la recherche et surtout ma passion
pour l’électrophysiologie. Je tiens à vous remercier, Marion et toi, pour la disponibilité et le
soutien que vous m’avez accordés au cours de ce stage.

Je remercie les docteurs Andreas Lüthi et Johannes Letzkus de m’avoir proposé de
collaborer avec eux. Cette collaboration fructueuse m’a permis de bien commencer ma thèse
et m’a donné la motivation nécessaire pour la suite.
J’adresse mes remerciements à l’ensemble des membres de l’institut Magendie qui ont
participé de près ou de loin à la réalisation de ces travaux de thèse.
Je souhaite remercier mes amis Jonathan, Wilfried, Pierrick, Medhi et Amandine.
Merci pour votre soutien et pour les moments passés ensemble.
Je tiens à remercier ma famille, notamment mes parents. Merci de m’avoir poussé à
faire des études et de m’en avoir donné les moyens.
Pour finir, cette thèse n'aurait pas vu le jour sans le soutien de ma fiancée. Ce travail
de thèse m’a demandé beaucoup de fidélité et m’a souvent éloigné de toi. Tu as su
m’encourager quand j’en avais besoin, passer outre les nombreuses soirées et week-ends l’un
sans l’autre. Je tiens à te dédier ce travail qui, je l’espère, te rendra fière.

RESUME
Les processus d'apprentissage et de mémoire sont contrôlés par des circuits et éléments
neuronaux spécifiques. De nombreuses études ont récemment mis en évidence que les circuits
corticaux jouent un rôle important dans la régulation des comportements de peur, cependant,
leurs caractéristiques anatomiques et fonctionnelles restent encore largement inconnues. Au
cours de ma thèse, en utilisant des enregistrements unitaires et des approches optogénétiques
chez la souris libre de se comporter, nous avons pu montrer que les interneurones inhibiteurs
du cortex auditif et du cortex préfrontal médian forment un microcircuit désinhibiteur
permettant respectivement l'acquisition et l'expression de la mémoire de peur conditionnée.
Dans les deux cas, les interneurones parvalbuminergiques constituent l'élément central du
circuit et sont inhibés de façon phasique. D’un point de vue fonctionnel, nous avons démontré
que cette inhibition était associée à la désinhibition des neurones pyramidaux par un
mécanisme de réduction de l'inhibition continue exercée par les interneurones
parvalbuminergiques. Ainsi, les interneurones parvalbuminergiques peuvent contrôler
temporellement l'excitabilité des neurones pyramidaux. En particulier, nous avons montré que
l'acquisition de la mémoire de peur conditionnée dépend du recrutement d'un microcircuit
désinhibiteur localisé dans le cortex auditif. En effet, au cours du conditionnement de peur, la
présentation du choc électrique induit l'inhibition des interneurones parvalbuminergiques, ce
qui a pour conséquence de désinhiber les neurones pyramidaux du cortex auditif et de
permettre l’apprentissage du conditionnement de peur. Dans leur ensemble, ces données
suggèrent que la désinhibition est un mécanisme important dans l'apprentissage et le
traitement de l'information dans les circuits corticaux. Dans un second temps, nous avons
montré que l'expression de la peur conditionnée requière l'inhibition phasique des
interneurones parvalbuminergiques du cortex préfrontal médian. En effet, leur inhibition
désinhibe les cellules pyramidales préfrontales et synchronise leur activité en réinitialisant les
oscillations thêta locales. Ces résultats mettent en évidence deux mécanismes neuronaux
complémentaires induits par les interneurones parvalbuminergiques qui coordonnent et
organisent avec précision l’activité neuronale des neurones pyramidaux du cortex préfrontal
pour contrôler l'expression de la peur conditionnée. Ensemble, nos données montrent que la
désinhibition joue un rôle important dans les comportements de peur en permettant
l’association entre des informations comportementalement pertinentes, en sélectionnant les
éléments spécifiques du circuit et en orchestrant l'activité neuronale des cellules pyramidales.

ABSTRACT
Learning and memory processes are controlled by specific neuronal circuits and elements.
Numerous recent reports highlighted the important role of cortical circuits in the regulation of
fear behaviour, however, the anatomical and functional characteristics of their neuronal
components remain largely unknown. During my thesis, we used single unit recordings and
optogenetic manipulations of specific neuronal elements in behaving mice, to show that both
the auditory cortex and the medial prefrontal cortex contain a disinhibitory microcircuit
required respectively for the acquisition and the expression of conditioned fear memory. In
both cases, parvalbumin-expressing interneurons constitute the central element of the circuit
and are phasically inhibited during the presentation of the conditioned tone. From a functional
point of view, we demonstrated that this inhibition induced the disinhibition of cortical
pyramidal neurons by releasing the ongoing perisomatic inhibition mediated by parvalbumin-
expressing interneurons onto pyramidal neurons. Thereby, this disinhibition allows the precise
temporal regulation of pyramidal neurons excitability. In particular, we showed that the
acquisition of associative fear memories depend on the recruitment of a disinhibitory
microcircuit in the auditory cortex. Fear-conditioning-associated disinhibition in auditory
cortex is driven by foot-shock-mediated inhibition of parvalbumin-expressing interneurons.
Importantly, pharmacological or optogenetic blockade of pyramidal neuron disinhibition
abolishes fear learning. Together, these data suggest that disinhibition is an important
mechanism underlying learning and information processing in cortical circuits. Secondly, in
the medial prefrontal cortex, we demonstrated that expression of fear behaviour is causally
related to the phasic inhibition of prefrontal parvalbumin-expressing interneurons. Inhibition
of parvalbumin-expressing interneuron activity disinhibits prefrontal pyramidal neurons and
synchronizes their firing by resetting local theta oscillations, leading to fear expression. These
results identify two complementary neuronal mechanisms both mediated by prefrontal
parvalbumin-expressing interneurons that precisely coordinate and enhance the neuronal
efficiency of prefrontal pyramidal neurons to drive fear expression. Together these data
highlighted the important role played by neuronal disinhibition in fear behaviour by binding
behavioural relevant information, selecting specific circuit elements and orchestrating
pyramidal neurons activity.

TABLE OF CONTENTS
INTRODUCTION ..................................................................................................... - 1 -
I/ Cortical circuits of conditioned fear behaviour .................................................... - 2 -
A/ Anatomical description of the auditory and medial prefrontal cortices ...................... - 2 -
1/ Gross anatomy of the auditory cortex ................................................................. - 2 -
2/ Gross anatomy of the medial prefrontal cortex .................................................. - 4 -
B/ Fine anatomical description of the neuronal elements of the cortex ........................... - 5 -
1/ Pyramidal neurons .............................................................................................. - 7 -
2/ Interneurons ........................................................................................................ - 9 -
a/ Somatodendritic profile and target selectivity ........................................... - 10 -
b/ Neurochemical expression profile ............................................................. - 12 -
c/ Electrophysiological profile ....................................................................... - 14 -
d/ Function of cortical interneurons .............................................................. - 15 -
II/ Role of cortical circuits in conditioned fear behaviour ................................... - 17 -
A/ Role of the AC in the acquisition of conditioned fear behaviour .............................. - 17 -
B/ Role of the mPFC in the expression/extinction of conditioned fear behaviour ......... - 19 -
1/ mPFC circuits modulating fear behaviour ....................................................... - 26 -
2/ mPFC elements encoding fear behaviour ......................................................... - 28 -
III/ Neuronal encoding of fear behaviour in the cortex ........................................ - 31 -
AIM OF THE THESIS ......................................................................................... - 34 -
MATERIAL AND METHODS ....................................................................... - 35 -
I/ Animals ......................................................................................................................... - 35 -
II/ Behaviour .................................................................................................................... - 35 -
A/ Fear conditioning and extinction paradigm ............................................................... - 35 -
B/ Fear conditioning to complex tone paradigm ............................................................ - 36 -

C/ Place avoidance paradigm ......................................................................................... - 37 -
III/ Surgery and recordings .......................................................................................... - 37 -
IV/ Single-unit analyses ................................................................................................ - 38 -
V/ Field potential analyses ........................................................................................... - 40 -
VI/ Virus injections and optogenetics ....................................................................... - 42 -
VII/ Anatomical analysis .............................................................................................. - 42 -
VIII/ Muscimol inactivation ........................................................................................ - 45 -
IX/ Extracellular stimulation ....................................................................................... - 45 -
RESULTS...................................................................................................................... - 46 -
I/ Results Part 1 ............................................................................................................... - 46 -
A/ Introduction ............................................................................................................... - 46 -
B/ Fast spiking interneurons disinhibited pyramidal neurons in the auditory cortex during
fear acquisition ............................................................................................................... - 47 -
C/ Article 1: “A disinhibitory microcircuit for associative fear learning in the auditory
cortex” ............................................................................................................................ - 48 -
II/ Results Part 2 ............................................................................................................. - 49 -
A/ Introduction ............................................................................................................... - 49 -
B/ Article 2: “Prefrontal parvalbumin interneurons shape neuronal activity to drive fear
expression” ..................................................................................................................... - 49 -
GENERAL DISCUSSION.................................................................................. - 74 -
PUBLICATIONS AND COMMUNICATIONS ................................... - 78 -
REFERENCES .......................................................................................................... - 79 -

LIST OF ABBREVIATIONS
AAV Adeno-associated virus
AC Auditory cortex
ACC Anterior cingulate cortex
AHP After hyperpolarization
ArchT Archeorhodopsin
Au1 Primary area of the auditory cortex
AuD Dorsal auditory areas
AuV Ventral auditory areas
BA Basal amygdala
BDNF Brain-derived neurotrophic factor
BLA Basolateral amygdala
CB Calbindin
CCK Cholecystokinin
ChR2 Channelrhodopsin
CR Calretinin
CeA Central amygdala
CEm Medial division of the central amygdala
CS Conditioned stimulus
dmPFC Dorsal medial prefrontal cortex
ERK Extracellular-regulated kinase
Fr2 Frontal cortex area 2
GABA Gamma aminobutyric acid
IB Intrinsic bursting or low-threshold spiking
IL Infralimbic cortex
IN Interneuron
ITC Intercalated cell
LA Lateral amygdala
LTD Long term depression
LTP Long term potentiation
LFP Local field potential
MD Medio-dorsal thalamus
MG Medial geniculate body of the thalamus

mPFC Medial prefrontal cortex
MRL Mean resultant length
MS Medial septum
MUS Muscimol
NBM Nucleus Basalis of Meynert
NIB Non-inactivating bursting
NPY Neuropeptide Y
PAG Periaqueductal grey area
PC Principal component
PCA Principal component analysis
PFA Paraformaldehyde
PL Prelimbic cortex
PN Pyramidal neuron
PV Parvalbumin
RS Regular spiking
SOM Somatostatin
US Unconditioned stimulus
vHPC Ventral hippocampus
VIP Vasoactive intestinal peptide

- 1 -
INTRODUCTION
An important question in neuroscience is how the brain controls behaviour. One of the
main functions of the brain is its capacity to promote behavioural adaptation according to
internal or environmental cues. This function is, in part, sustained by learning and memory
processes that are supported by specific cellular and molecular mechanisms within dedicated
neuronal circuits. In contrast to our detailed understanding of these mechanisms, the
involvement of specific neuronal circuits and elements controlling neuronal activity during
learning and memory in the intact animal remains poorly understood.
Since many years, classical Pavlovian auditory fear conditioning has been intensively
used in the laboratory to study the cellular mechanisms leading to associative learning and
memory. Auditory fear conditioning is a rapid and robust learning paradigm during which an
animal learns to associate a previously neutral tone (the conditioned stimulus, or CS) with a
coincident aversive stimulus (the unconditioned stimulus, or US, typically a footshock).
Following auditory fear conditioning, re-exposure to the CS induces the expression of a broad
range of conditioned fear responses, including an innate immobilization reaction called
freezing, a behavioural measure of fear memory. Although a single association between the
CS and the US can lead to long-lasting fear memories (Gale et al., 2004; LeDoux, 2000), the
repeated presentation of the CS in the absence of the US can transiently inhibit conditioned
fear responses, a phenomenon labeled fear extinction (Myers & Davis, 2007). Fear extinction
is thought to be sustained by new learning that promotes the development of an inhibitory
association between the CS and the US. Subsequently, this inhibitory association competes
with the original fear memory rather than erasing it (Pavlov, 1927). This hypothesis is
supported by the fact that following extinction, conditioned responses can recover with the
passage of time (spontaneous recovery) (Quirk, 2002), a contextual shift (renewal) (Bouton &
King, 1983), or an exposure to the original US (reinstatement) (Rescorla & Heth, 1975).
Extinction learning can lead to the formation of short- and long-lasting forms of extinction
memory that can be retrieved in a context-dependent manner (Bouton et al., 2006; Herry et
al., 2010). Thus, fear and extinction memories co-exist and interact to regulate fear behaviour
(Herry et al., 2008).
In the past decades, numerous studies have attempted to define the neuronal substrates
and the cellular and molecular mechanisms governing the different phases of fear behaviour.
Most of these studies pointed toward the amygdala, a neuronal structure of the medial
temporal lobe, as a key neuronal element involved in the formation, extinction and expression

- 2 -
of associative fear memories (Herry et al., 2010; Johansen et al, 2011; LeDoux, 2000; Maren,
2001; Maren & Quirk, 2004; Pape & Pare 2010). The amygdala comprises several
anatomically and functionally distinct nuclei including the lateral (LA), basal (BA) (together
referred as the basolateral complex of the amygdala (BLA)) and central nuclei (CeA). During
fear conditioning, the LA and the CeA are supposed to be the likely sites of the plasticity
underlying acquisition of fear conditioning (Ciocchi et al, 2010; Maren, 2001; Mckernan &
Shinnick-Gallagher, 1997; Rogan et al., 1997; Schafe & LeDoux, 2008; Wilensky et al,
2006). Following fear acquisition, several lines of evidence indicate that conditioned fear
responses are controlled by CeA circuits and that the BLA is required for extinction learning
(Cassell & Wright, 1986; Ciocchi et al, 2010; Herry et al., 2006, 2008, 2010; Hopkins &
Holstege, 1978; Sotres-Bayon et al., 2007; Veening et al., 1984).
Although most of the research has concentrated on the role played by amygdala
neuronal circuits in fear behaviour, some reports have also highlighted the critical role played
by the cortex in the regulation of amygdala-dependent fear memories (Herry et al., 2010;
Sotres-Bayon & Quirk, 2010; Quirk & Mueller, 2008; Weinberger, 2004). Indeed, over the
past decade, a substantial amount of studies have identified the auditory cortex (AC) and the
medial prefrontal cortex (mPFC) as two main cortical structures involved in fear behaviour. In
particular, the AC is hypothesized to play a role in the acquisition of auditory fear
conditioning, whereas the mPFC is thought to be more implicated in the consolidation phase
of extinction learning and the expression of fear and extinction memories (Sotres-Bayon &
Quirk, 2010; Suga & Ma, 2003; Weinberger, 2004). In the following sections, I will first
review the gross and fine cellular anatomy of both the AC and mPFC. In a second step, I will
describe the available functional data on the role played by the AC and mPFC in fear
behaviour. Finally, in the last section of the introduction, I will discuss the different neuronal
mechanisms allowing for the encoding of fear behaviour in dedicated neuronal structures and
circuits.
I/ Cortical circuits of conditioned fear behaviour
A/ Anatomical description of the auditory and medial prefrontal cortices
1/ Gross anatomy of the auditory cortex
The AC is located in the temporal lobe and in rodents, is generally divided in several
regions: the primary area or core auditory cortex (Au1) and the secondary areas or belt

- 3 -
auditory cortex (Caviness, 1975; Frisina & Walton, 2001; Willard & Ryugo, 1983)
surrounding the Au1 and segregated in dorsal and ventral auditory areas (AuD and AuV,
respectively). The distinction between the different auditory regions depends mainly on
cytoarchitectonic organization and functional connectivity. The Au1 is characterized by a
clear differentiation of six cellular layers with prominence of layers IV and V (Willard &
Ryugo, 1983). The Au1 is considered as the main terminus of the auditory system forming the
ascending auditory pathway. In particular, it receives inputs from the ventral subdivision of
the lemniscal nucleus of the medial geniculate body of the thalamus (MG), (Romanski &
LeDoux, 1993a). These auditory inputs target the layer 4 of the Au1 and convey information
relative to the perception of auditory stimuli with a precise rostro-caudal tonotopic
organization (Ehret, 1997; Schreiner & Winer, 2007). Furthermore, layers 2 and 3 of the Au1
also receive non tonotopic inputs originating from the medial sub-division of the MG and the
posterior intralaminar nucleus of the thalamus (Lee & Sherman, 2011). Besides these thalamic
afferents, the Au1 receives inputs from other secondary auditory areas and cortices, including
the contralateral AC via commissural fibers, the ventral tegmental area, and the nucleus
basalis of Meynert (NBM; Suga & Ma 2003; Willard & Ryugo 1983). The Au1 projects back
to most of the sub-cortical structures of the auditory system giving rise to the descending
auditory pathway (Doucet et al., 2003; Hazama et al., 2004; Kimura et al., 2004). The Au1
emits also numerous corticocortical projections emanating from all layers and representing
approximately half of Au1 outputs (Lee & Sherman, 2011; Romanski & Ledoux, 1993b; Suga
& Ma, 2003).
Concerning, auditory secondary areas, both AuD and AuV are characterized by a
decrease of cortical thickness and an increase of neuronal density (Willard & Ryugo, 1983).
Besides inputs from the Au1, they receive auditory inputs that are typically non tonotopic
from extralemniscal thalamic nuclei, such as the dorsal and medial divisions of the MG, and
other thalamic and cortical sensory inputs from different modalities (Hu et al., 1994; Kimura
et al., 2004; Lee & Sherman, 2011; Llano & Sherman, 2008). The AuD project back to
structures involved in directed attention and visual processing (Kimura et al., 2007), whereas
the AuV send projections to structures such as the amygdala and the perirhinal cortex
(LeDoux et al. 1991; Mascagni et al., 1993; Romanski & LeDoux, 1993b; Shi & Cassell,
1997).

- 4 -
2/ Gross anatomy of the medial prefrontal cortex
In rodents, the mPFC can be separated based on cytoarchitectonic and hodologic
criteria in four distinct areas which, from dorsal to ventral, are the frontal cortex area 2 (Fr2;
Heidbreder & Groenewegen, 2003; Uylings et al., 2003), the anterior cingulate cortex (ACC),
the prelimbic (PL) and the infralimbic (IL) cortices (Guldin et al., 1981; Krettek & Price,
1977; Leonard, 1969; Ongur & Price, 2000; Ray & Price, 1992; Van Eden & Uylings, 1985).
The different regions of the mPFC are agranular cortex (lacking layer 4) and present distinct
organization on their cellular lamination. The Fr2 also called medial precentral cortex (Van
Eden & Uylings, 1985) or medial agranular cortex (Donoghue & Wise, 1982) or secondary
motor area (Franklin et al., 1997) and the ACC divided in dorsal and ventral parts (Uylings et
al., 2003) are characterized by a well-defined laminar organization with two distinguishable
pyramidal layers 2 and 5 and a large layer 6b. The PL is characterized by a weak
disorganization of the layer 5 and a layer 6 divided in two sub-layers 6a and 6b. The IL,
bordered rostraly by the medial orbital cortex and posteriorly by the dorsal peduncular cortex,
is characterized by homogeneity of its organization with no remarkable frontiers between the
different layers. The incorporation of the Fr2 and ACC to the mPFC has long been debated,
but their reciprocal connections with the medio-dorsal thalamus (MD) lead to the consensual
view that both Fr2 and ACC belong to the mPFC. Indeed, in the prefrontal cortex, the mPFC
differs from the lateral prefrontal cortex and the orbital prefrontal cortex by its agranular
nature but also by its strong connectivity with the MD. In particular, the mPFC receives a
strong input from the MD, with the medial segment of the MD projecting to the PL and IL,
the lateral segment to both the PL and dorsal ACC and the paralamellar segment contacting
mainly the Fr2 (Uylings & van Eden, 1990). These thalamic projections are mostly ipsilateral
and terminate in cellular layers 1 and 3 (Groenewegen, 1988; Krettek & Price, 1977; Kuroda
et al., 1993; Minciacchi & Granato, 1989). Besides these thalamic afferents, the mPFC
receives inputs from numerous sub-cortical neuronal structures including the basal ganglia
(Groenewegen et al., 1997), the amygdala (Krettek & Price, 1977; McDonald, 1987, 1991;
Shinonaga et al., 1994) and the hippocampus (Jay et al., 1989; Swanson, 1981). Notably,
hippocampal glutamatergic inputs from the ventral CA1 and the subiculum terminate in deep
mPFC cellular layers and projections from the basolateral amygdala (BLA) preferentially
contact PL and IL regions (Gigg et al., 1994; Jay et al., 1989; McDonald, 1987, 1991;
Swanson, 1981). In terms of neuromodulatory system, the mPFC received dopaminergic
innervation from the ventral tegmental area (Thierry et al., 1973), noradrenergic innervations

- 5 -
from the locus coeruleus, cholinergic innervations from the basal forebrain and serotoninergic
innervations from the raphe nuclei (Chandler et al., 2013; Uylings et al., 2003). The mPFC
also receives cortical projections originating from the paralimbic cortex (enthorinal and
perirhinal cortices) that target the PL and IL, and from somatosensory and motor cortices that
terminate in the dorsal prefrontal regions. The mPFC contains reciprocal projections to the
MD, hippocampus, BLA, and basal ganglia where it participates in several cortico-striato-
pallido-thalamo-cortical loops (Alexander et al., 1986, 1990a, 1990b, 1994; Berendse et al.,
1992; Floyd et al., 2000, 2001; Groenewegen, 1988; Groenewegen et al., 1990; Hirel et al.,
2013; Krettek & Price, 1977; McDonald et al., 1996; McDonald, 1998; Sesack et al., 1989;
Takagishi & Chiba, 1991; Terreberry & Neafsey, 1987; Vertes, 2004). The mPFC also
projects directly to the periaqueductal gray area (PAG), the septum and some hypothalamic
areas (Gabbott et al., 2005; Heidbreder & Groenewegen, 2003; Vianna et al., 2001). Finally,
the mPFC contains important intrinsic ipsilateral connectivity. Namely, the PL region projects
to ACC, the IL projects to both the PL and the dorsal part of ACC. The mPFC also contains
an overall homotopic contralateral connectivity (Audinat et al., 1988; Beckstead, 1979;
Sesack et al., 1989).
B/ Fine anatomical description of the neuronal elements of the cortex
The cortex is a complex, but relatively stereotypic organized network, composed of
multiple cell types forming micro to large scale neuronal circuits (DeFelipe & Farinas, 1992;
Silberberg et al., 2002; Varela et al., 2001). The cortical functions that sustain fear behaviour
cannot be understood outside their structural frame. What kind of neurons forms cell
assemblies within the cortex? What remote or local neurons do they target? How does firing
translate into synaptic activity and finally how do individual cortical neurons integrate
multiple inputs to produce their firing? Addressing these questions requires circuit-level
analysis, taking into account the diversity of the multiple cell types and understanding their
anatomical and functional interactions. The cortex is composed of two main neuronal groups,
glutamatergic pyramidal neurons (PN) and GABAergic interneurons (IN), which are
distributed across different cortical layers. The laminar organization of the cortex, largely
conserved across mammalian species, is based on cytoarchitectonic criterions that define,
horizontally to the cortical surface, six layers numbered from the most superficial to the
deepest (Fig. 1; Douglas & Martin, 2004; Nieuwenhuys, 1994). The layer 1 (molecular layer)
constitutes the most superficial layer, situated directly below to the pial surface it is
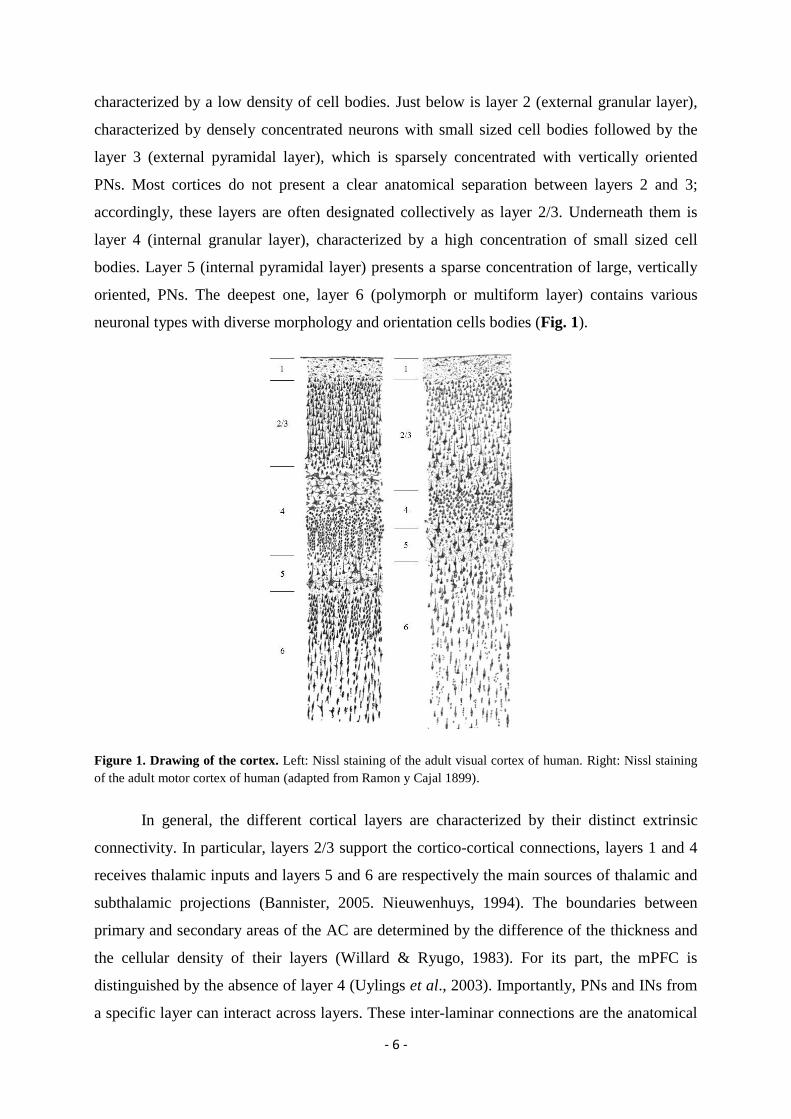
- 6 -
characterized by a low density of cell bodies. Just below is layer 2 (external granular layer),
characterized by densely concentrated neurons with small sized cell bodies followed by the
layer 3 (external pyramidal layer), which is sparsely concentrated with vertically oriented
PNs. Most cortices do not present a clear anatomical separation between layers 2 and 3;
accordingly, these layers are often designated collectively as layer 2/3. Underneath them is
layer 4 (internal granular layer), characterized by a high concentration of small sized cell
bodies. Layer 5 (internal pyramidal layer) presents a sparse concentration of large, vertically
oriented, PNs. The deepest one, layer 6 (polymorph or multiform layer) contains various
neuronal types with diverse morphology and orientation cells bodies (Fig. 1).
Figure 1. Drawing of the cortex. Left: Nissl staining of the adult visual cortex of human. Right: Nissl staining
of the adult motor cortex of human (adapted from Ramon y Cajal 1899).
In general, the different cortical layers are characterized by their distinct extrinsic
connectivity. In particular, layers 2/3 support the cortico-cortical connections, layers 1 and 4
receives thalamic inputs and layers 5 and 6 are respectively the main sources of thalamic and
subthalamic projections (Bannister, 2005. Nieuwenhuys, 1994). The boundaries between
primary and secondary areas of the AC are determined by the difference of the thickness and
the cellular density of their layers (Willard & Ryugo, 1983). For its part, the mPFC is
distinguished by the absence of layer 4 (Uylings et al., 2003). Importantly, PNs and INs from
a specific layer can interact across layers. These inter-laminar connections are the anatomical

- 7 -
framework of the cortical columns that form functional entities of interconnected neurons
(Adesnik & Scanziani, 2010; Hubel & Wiesel, 1962; Mountcastle, 1957).
To understand the anatomical and functional organization of the cortex, it is important
to have a detailed knowledge of the characteristics and the specificity of the different PNs and
INs that constitute cortical circuits. The two next sections reviewed the literature concerning
the PNs and INs.
1/ Pyramidal neurons
Pyramidal excitatory neurons, also named principal or projection neurons represent the
vast majority of cortical neurons (70-80%). They used glutamate as a neurotransmitter and are
located in all six cortical layers, except layer 1 (DeFelipe & Farinas, 1992). PNs are spiny
neurons and their dendritic arbors typically display two different subregions containing short
basal dendrites emerging from the base of the soma and apical dendrites arising from the top
of the soma and running perpendicularly to the pia matter, terminating in a dendritic tuft in
layer 1 (DeFelipe & Farinas, 1992, Spruston, 2008). Their axon terminals or axon collaterals
form asymmetrical synapses, corresponding to Gray’s type I (Gray, 1959). They receive
synaptic inputs targeting distinct dendritic, somatic and axonal sub-cellular compartments
with a regional distribution that depends on the nature of the input. In particular, excitatory
synaptic inputs preferentially contact dendritic spines (DeFelipe & Farinas, 1992,
Nieuwenhuys, 1994, Spruston, 2008), whereas local inhibitory inputs emanating from specific
subpopulations of INs target different PN sub-cellular compartments (see section interneurons
below).
Cortical PNs are often considered as a homogeneous cell population. However, based
on the characterization of their morphology, connectivity, electrophysiological properties and
gene expression, it is becoming clear that various PN types may exist (Krook-Magnuson et
al., 2012). These accumulating evidence strongly suggests that different PN subpopulations
exist and sustain specific functions (Bannister, 2005; Mizuseki et al., 2011; Watakabe, 2009).
The morphology of the PNs can differ between layers and cortical regions. Indeed, the cell
body shape, the dendritic orientation, the number of dendrite branches and the distribution of
spines are variable (DeFelipe & Farinas, 1992; Nieuwenhuys, 1994). For example, spiny
stellates cells are glutamatergic cells located in layer 4 of primary sensory cortex which lack
apical dendrites. The synaptic target of the PNs seems to be dependent on their cell body layer
position. In general, PNs located in superficial layers (supragranular layers: layers 1 to 3)

- 8 -
project to cortical areas, those in deep layers (infragranular layers: layers 5 to 6) have long
range projections and form the corticothalamic or corticosubcortical pathway (Bannister,
2005. Nieuwenhuys, 1994). PNs received intra- and extra-cortical excitatory inputs that
globally reach the cortex via supragranular layers. For example, mPFC inputs from the BLA,
vHIP and the MD have been shown to make excitatory synapses mostly with dendritic spines
of PNs in layer 2 (Bacon et al., 1996; Carr & Sesack, 1996; Gabbott et al., 2012; Kuroda et
al., 1995a, b; Orozco-Cabal et al., 2006; Parent et al., 2010). Moreover, the selective targeting
of specific dendritic compartments of layer 2 PNs was shown to differentially affect synaptic
responses (Little & Carter, 2012). For primary sensory cortices, such as the AC, thalamic
inputs terminate on the internal granular layer (Bannister, 2005). Accumulating evidence
indicates that local excitatory and inhibitory inputs onto PNs are highly organized, forming
local microcircuits. Thereby, PNs having the same synaptic target share inputs from the same
cell types (Bannister, 2005; Brown & Hestrin, 2009; Krook-Magnuson et al., 2012; Markram
et al., 2004; Morishima et al., 2006, 2011). Based on their spiking characteristics in response
to depolarizing current pulses, PNs present different electrophysiological profiles (Fig. 2).
Except specific differences between species, in vitro or in vivo preparations, and cellular
location, PNs have been divided into three main classes: regular spiking (RS), low-threshold
spiking or intrinsic bursting (IB) and non-inactivating bursting (NIB) neurons (Chang &
Luebke, 2007; Degenetais et al., 2002; Huggenberger at al., 2009; Sun et al., 2013; Yang et
al., 1996; Zaitsev et al., 2012). RS PNs display a sustained firing pattern in response to
depolarizing current pulses and can be further divided into three subgroups based on
frequency adaptation responses: RS slow adapting-group I, RS slow adapting-group II and RS
fast adapting PNs. IB PNs emit bursts characterized by an intra-burst decrease of spike
amplitude and duration, whereas NIB PNs exhibit bursts without spike amplitude
modifications (Chang & Luebke, 2007; Degenetais et al., 2002; Huggenberger at al., 2009;
Yang et al., 1996; Zaitsev et al., 2012). Despite these different firing characteristics, it is still
unknown whether these PN classes correspond to distinct functional entities. Finally, some
researchers have attempted to define cortical PN populations based on their gene expression
profile. Using this strategy, strong correlations were established between gene expression
profiles in PNs and projection patterns (Hevner et al., 2003; Molnar & Cheung, 2006; Sugino
et al., 2006; Watakabe et al., 2007). For example, Sugino et al., (2006) demonstrated that two
populations of PNs in the ACC with distinct morpho-functional characteristics displayed
different gene expression profiles. The first population was located in layer 5 and belonged to
non-adapting PNs whereas the second population was located in layer 6 and corresponded to

- 9 -
adapting corticothalamic neurons (Sugino et al., 2006). PNs represent both the main receptive
surface for extra-cortical inputs and quantitatively the main cortical output. In light of their
apparent heterogeneity, it will be important to consider in the future which and how specific
types of PNs regulate fear behaviour.
Figure 2. Characteristics of the three main electrophysiological classes of prefrontal pyramidal neurons.
Top, left: schematic representation of the orientation of a PN within the prelimbic/medial orbital area in a frontal
section. The arrow indicates the caudo-rostral (C-R) and the ventro-dorsal (V-D) axes of the brain section and of
the reconstructed cells (upper left). Upper right panel: two slow-adapting RS cells (group I) with soma in deep
and superficial layers, respectively. Bottom left panel: two NIB cells with soma in deep and superficial layers,
respectively. Bottom right panel: an IB cell with soma in layer V. Note that NIB cells have a smaller soma and
less developed dendritic fields than RS and IB cells and that the IB cell presents thick dendrites and a large
dendritic extent. Insets: characteristic firing patterns of RS, NIB and IB cells (from Degenetais et al., 2002).
2/ Interneurons
The cortex also contains GABAergic INs that are located in all cortical layers and
represent about 20-30% of cortical neurons (DeFelipe & Farinas, 1992). INs, also called non-
pyramidal, aspiny, inhibitory, or local neurons are mainly aspiny neurons and release gamma
aminobutyric acid (GABA). Apart from some exceptions, IN projections do not leave the

- 10 -
cortex and are restricted to a local environment (Gonchar et al., 1995; Letinic et al., 2002).
Their axon terminals form symmetrical synapses, corresponding to Gray’s type II (Gray,
1959). Some INs are electrically coupled through Gap junctions forming a direct link between
the cytoplasm of neighboring INs (Druga, 2009). Cortical inhibitory INs are highly diverse
and are comprised of many types. Interneuronal cell types can be identified on the basis of
anatomical, neurochemical and electrophysiological criteria (Fig. 3; Ascoli et al., 2008;
DeFelipe et al., 2013; Gupta et al., 2000; Markram et al., 2004). Using anatomical techniques,
INs can be described according to their somatodendritic profiles (Ascoli et al., 2008;
Markram et al., 2004). Remarkably, individual INs selectively target distinct compartments of
PNs (dendrites, soma or axon initial segment) or other IN types. At the neurochemical level,
several categories of molecules have been identified to help distinguish IN subtypes (Ascoli et
al., 2008; Burkhalter, 2008; DeFelipe, 1997; Markram et al., 2004). Next, cortical inhibitory
INs can be distinguished based on their electrophysiological properties (Cauli et al., 1997;
Gupta et al., 2000; Markram et al., 2004). Importantly, none of the anatomical, neurochemical
or electrophysiological criteria alone can reliably classify cortical inhibitory INs. Some INs
with distinct morphologies can display identical neurochemical or electrophysiological
profiles. Therefore, the combination of multiple criteria represents the preferred solution to
identify specific IN subtypes (Cauli et al., 1997; Markram et al., 2004; Sugino et al., 2006;
Toledo-Rodriguez et al., 2004). Because of their morphological, electrophysiological and
molecular diversity, INs are believed to differentially sculpt cortical activity. Thus, it is
widely accepted that a detailed understanding of IN functions is a prerequisite to understand
the functional organization of cortical neuronal circuits.
a/ Somatodendritic profile and target selectivity
Cortical INs display variable layer locations as well as distinct somatic, dendritic and
axonal morphological features (Fig. 3). Accumulating anatomical data suggests that distinct
INs target specific subcellular regions of the postsynaptic PN. In particular, some INs
preferentially innervate perisomatic regions, proximal dendrites or axon initial segments,
whereas others target more distal dendrites (Freund, 2003; Markram et al., 2004).
Furthermore, other INs have been described as specialized in targeting other cortical INs
(Somogyi et al., 1998; Staiger et al., 1997). The specialization of IN connectivity is thought to
contribute largely to their functional specificities (Gentet, 2012; Isaacson & Scanziani, 2011).
The soma and proximal dendrite-targeting cells are composed by the basket cells that

- 11 -
represent the major INs subpopulation (Druga, 2009; Markram et al., 2004). They are called
basket cells because they collectively form basket-like structures around PN somata (Druga,
2009; Markram et al., 2004). Basket cells are characterized by their high percentage of
axosomatic boutons (higher than 14%: Karube et al., 2004). Basket cells can be further
subdivided into several classes based on their soma size, frequency of axonal branching, axo-
dendritic morphology and firing properties (i.e., large or small basket cells, descending and
nest basket cells) (Karube et al., 2004; Krimer et al., 2005; Markram et al., 2004; Uematsu et
al., 2008; Wang et al., 2002). Basket cells are mutually interconnected via chemical and
electrical synapses (Cobb et al., 1995; Fukuda, 2007; Somogyi et al., 1998). The axo-axonic
cells (Somogyi, 1977; Somogyi et al., 1982) are also called chandelier cells in the cortex
because their axon terminals form vertical rows resembling candle sticks on a chandelier.
These INs target specifically the initial segment of PNs (Karube et al., 2004; Krimer et al.,
2005; Markram et al., 2004). The dendrite-targeting cells can be subdivided into several cell
types: bipolar, double bouquet, bitufted and neurogliaform cells. The bipolar, double bouquet
and bitufted cells present bipolar and bitufted dendrites but are differentiated by their axonal
orientation or morphology (Markram et al., 2004). Some bipolar and double bouquet cells are
thought to specifically target other INs types (Gentet, 2012; Somogyi et al., 1998). The
neurogliaform cells present a spherical dendritic field and form electrical synapses with INs
from the same or different type (Druga, 2012). The dendritic and tuft-targeting cells are
divided in Martinotti and layer 1 cells. The Martinotti cells project on the apical tuft of PNs in
layer 1 (Markram et al., 2004). The layer 1 cells are confined to layer 1 and are divided in
heterogeneous group of INs and Cajal-Retzius cells (Druga, 2012).
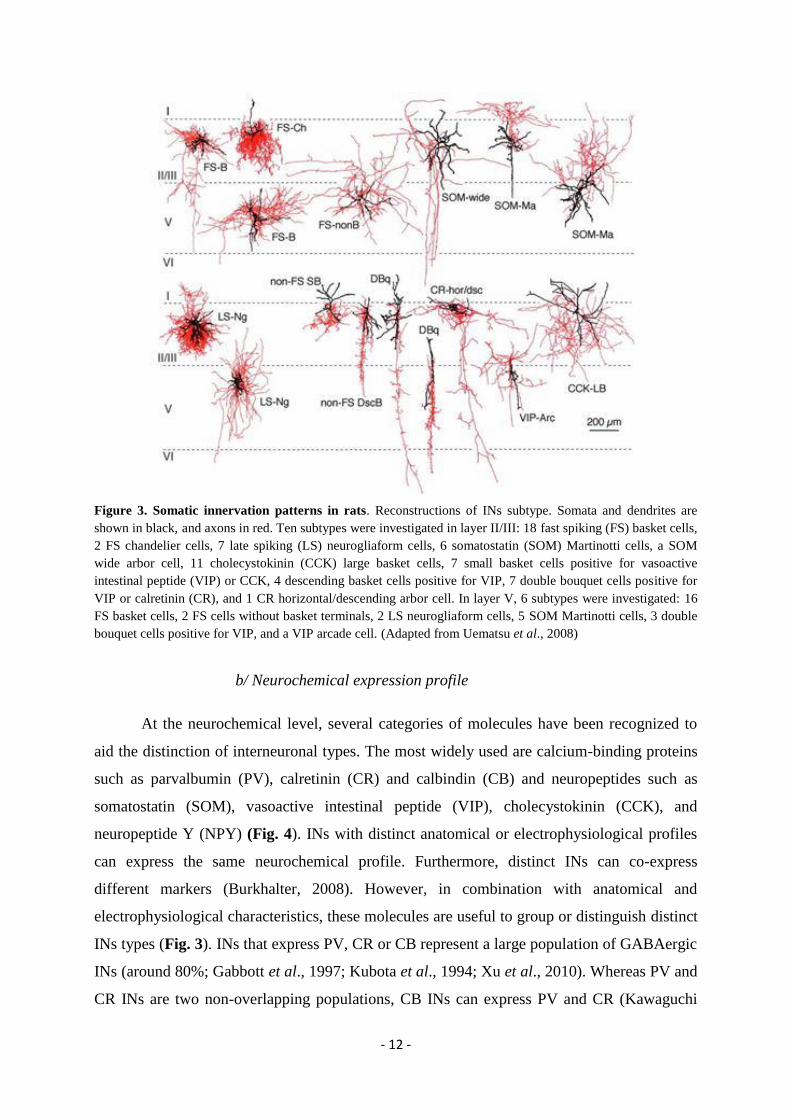
- 12 -
Figure 3. Somatic innervation patterns in rats. Reconstructions of INs subtype. Somata and dendrites are
shown in black, and axons in red. Ten subtypes were investigated in layer II/III: 18 fast spiking (FS) basket cells,
2 FS chandelier cells, 7 late spiking (LS) neurogliaform cells, 6 somatostatin (SOM) Martinotti cells, a SOM wide arbor cell, 11 cholecystokinin (CCK) large basket cells, 7 small basket cells positive for vasoactive
intestinal peptide (VIP) or CCK, 4 descending basket cells positive for VIP, 7 double bouquet cells positive for
VIP or calretinin (CR), and 1 CR horizontal/descending arbor cell. In layer V, 6 subtypes were investigated: 16
FS basket cells, 2 FS cells without basket terminals, 2 LS neurogliaform cells, 5 SOM Martinotti cells, 3 double
bouquet cells positive for VIP, and a VIP arcade cell. (Adapted from Uematsu et al., 2008)
b/ Neurochemical expression profile
At the neurochemical level, several categories of molecules have been recognized to
aid the distinction of interneuronal types. The most widely used are calcium-binding proteins
such as parvalbumin (PV), calretinin (CR) and calbindin (CB) and neuropeptides such as
somatostatin (SOM), vasoactive intestinal peptide (VIP), cholecystokinin (CCK), and
neuropeptide Y (NPY) (Fig. 4). INs with distinct anatomical or electrophysiological profiles
can express the same neurochemical profile. Furthermore, distinct INs can co-express
different markers (Burkhalter, 2008). However, in combination with anatomical and
electrophysiological characteristics, these molecules are useful to group or distinguish distinct
INs types (Fig. 3). INs that express PV, CR or CB represent a large population of GABAergic
INs (around 80%; Gabbott et al., 1997; Kubota et al., 1994; Xu et al., 2010). Whereas PV and
CR INs are two non-overlapping populations, CB INs can express PV and CR (Kawaguchi

- 13 -
and Kubota, 1997; Xu et al., 2010). PV INs represent about 40% of GABAergic neurons in
the rodent cortex (Gonchar et al., 2008; Uematsu et al., 2008; Xu et al., 2010). Co-
localization experiments indicate that PV INs did not co-express CR, SOM, VIP, CCK and
NPY (Gonchar et al., 2008; Kawaguchi and Kubota, 1997; Uematsu et al., 2008; Xu et al.,
2010). PV INs present basket and chandelier cells morphologies (DeFelipe et al., 1989;
Karube et al., 2004; Massi et al., 2012; Taniguchi et al., 2013). Two types of basket cells are
classically recognized: those expressing PV and CB (Hartwich et al., 2009; Kubota and
Kawaguchi, 1997) and those expressing CCK (Freund et al., 1986; Karube et al., 2004;
Kubota and Kawaguchi, 1997; Somogyi et al., 2004). CR INs represent about 20% of
GABAergic neurons in the rodent cortex (Gonchar et al., 2008; Uematsu et al., 2008; Xu et
al., 2010). CR INs can co-express different neuropeptides such as NPY, VIP and CCK, and
present dendrite-targeting and layer 1 cell morphologies. CR INs interact preferentially with
other INs in superficially layers and with PNs in deep layers (Druga, 2012; Meskenaite, 1997;
Somogyi et al., 1998). Neuropeptides are expressed in different INs populations: bipolar cells
express CR and VIP; double bouquet cells express CB/CR/VIP/CCK but not PV/SOM/NPY;
bitufted cells express CB/CR/NPY/VIP/SOM/CCK but not PV and martinotti cells express
SOM (Gonchar et al., 2007; Markram et al., 2004; Xu et al., 2010). SOM- and VIP-
expressing INs represent respectively around 20% and 10% of GABAergic cells of the rodent
cortex (Gonchar et al., 2007; Uematsu et al., 2008; Xu et al., 2010). Among the different
neurochemical expression profiles, PV, SOM and VIP INs are described as three non-
overlapping subpopulations that accounted for the majority of GABAergic cells (Kawaguchi
and Kubota 1997; Xu et al., 2010). Recently, the Petilla Interneuron Nomenclature Group
proposed to distinguish five different main groups of INs: (1) the PV INs, including
chandelier and basket cells; (2) the SOM INs, such as Martinotti cells; (3) those expressing
NPY but not SOM; (4) those expressing VIP; and (5) those expressing CCK but not SOM or
VIP. The molecular classification of INs is continually re-defined and new molecular markers
are discovered (Ascoli et al., 2008). The identification of different neurochemical markers
offer relevant tools from simple immunochemistry to optogenetic to study and manipulate the
genetically define INs in wild type or transgenic mouse lines (Cardin et al., 2009; Kawaguchi
and Kubota, 1997; Kvitsiani et al., 2013; Pfeffer et al., 2013; Xu et al., 2010).

- 14 -
Figure 4. Immunohistochemical profile of GABAergic cells in rat frontal cortex. Shared chemical
composition of INs. CB, calbindin; CCK, cholecystokinin; CR, calretinin; NPY, neuropeptide Y; PV,
parvalbumin; SOM, somatostatin; VIP, vasoactive intestinal polypeptide. The relative number of cells
immunoreactive for a substance is proportional to the size of the box (adapted from Kawaguchi & Kubota 1997).
c/ Electrophysiological profile
Accumulating evidence shows that cortical INs display various electrophysiological
properties. Kawaguchi and Kubota identified in the rat frontal cortex four main INs
electrophysiological profiles: i) fast spiking, ii) late spiking, iii) regular spiking and iv) burst
spiking or low threshold spiking (Kawaguchi and Kubota, 1993, 1996, 1997). Fast spiking
INs present short duration spikes, brief afterhyperpolarisation (AHP) and the ability to sustain
high frequency of discharge (Kawaguchi, 1995). The fast spiking profile has been linked to
PV chandelier and basket cells, however, not all basket cells are fast spiking INs (Karube et
al., 2004; Kawaguchi and Kubota, 1997; Krimer et al., 2005; Uematsu et al., 2008; Wang et
al., 2002). Late spiking INs showed a slow depolarizing ramp occurring around the spike
threshold and retarding the onset of action potentials. This profile corresponds to
neurogliaform or some basket INs (Karube et al., 2004; Kawaguchi and Kubota, 1997;
Krimer et al., 2005; Uematsu et al., 2008). Finally, regular spiking and burst spiking INs,
characterized by two or more spikes on slow depolarizing humps emitted in burst or not,
correspond to a range of INs morphologies and include SOM Martinotti cells, large basket

- 15 -
cells positive for CCK and VIP-containing double bouquet cells (Kawaguchi and Kubota,
1996, 1997). Recently, the Petilla Interneuron Nomenclature Group proposed to distinguish13
firing patterns depending on firing rate (fast or, non-fast spiking), regularity (facilitating,
regular, and accelerating) and bursting activity (Fig. 5; Ascoli et al., 2008).
Figure 5. Petilla terminology: types of firing patterns. Cortical GABAergic INs display a vast repertoire of
discharge responses. These samples are representative of the most common responses to standardized
intrasomatic step-current injections in the rat neocortex. The features of firing patterns in response to step-onset,
organized in columns, include bursts, delays and continuous firing, which is neither burst nor delayed. Steady-
state patterns, displayed in rows, can be fast spiking, non-adapting non-fast spiking, adapting, irregular spiking,
intrinsic burst firing or accelerating. Fast spiking neurons can also display a stuttering or ‘Morse-code-like’
discharge that is characterized by high-frequency spike clusters that are intermingled with unpredictable periods
of silence for a wide range of long, sustained, somatic-current injections. Blank areas of the table and boxes
containing only scale bars correspond to firing patterns that have not yet been characterized in neocortical INs.
The scale bar at the top left refers to the traces in the first four rows; the scale bars in the fifth and sixth rows
refers to the traces in the fifth and sixth rows, respectively (from Ascoli et al., 2008).
d/ Function of cortical interneurons
INs have been described as the orchestrator of cortical processes. The general principle
is that distinct interneuronal subpopulations differentially control, sculpt, synchronize or pace
cortical activity (Ascoli et al., 2008; Atallah et al., 2012; Burkhalter, 2008; Cardin et al.,

- 16 -
2010; Gentet, 2012; Issacson & Scanziani, 2011; Markram et al., 2004; Massi et al., 2012;
Merchant et al., 2012; Moore et al., 2010; Royer et al., 2012; Sohal et al., 2009). The
diversity of INs, in particular their distinct wiring layout are supposed to be the anatomical
basis of their distinct roles in the control of the inflow and outflow of activity inside cortical
microcircuits (Buzsaki et al., 2004; Merchant et al., 2012; Moore et al., 2010). Dendritic
inhibition likely controls the efficacy and plasticity of excitatory synaptic inputs onto PNs
dendrites (Chiu et al., 2013; Freund & Buzsaki, 1996; Gentet et al., 2012; Lovett-Barron et
al., 2012). For example, in primary visual cortex, dendrite-targeting, SOM INs sharpen visual
stimulus selectivity of PNs via their inhibition of dendritic excitatory responses (Wilson et al.,
2012). Somata and proximal dendrite-targeting INs effectively control the gain of summated
synaptic potential and therefore the local generation of Na+-dependent action potentials (Miles
et al., 1996). By targeting the site of action potential generation, axo-axonic cells are thought
to influence the output activity of PNs (Howard et al., 2005). It has been calculated that each
axo-axonic cell may give rise to 200 to 300 cartridges capable of making synaptic contacts
with as many PNs (Somogyi et al., 1985). Axo –axonic INs may therefore shunt the activity
of large groups of PNs. Other IN types present specific connectivity that gives them unique
functions. For example neurogliaform cells are believed to control the excitability of local
cortical microcircuit via extrasynaptic GABA release (Olah et al., 2009). Besides their
connectivity with PNs, INs are interconnected through electrical and chemical synapses.
Electrical coupling is bidirectional and occurs almost exclusively between INs of the same
type and are thought to shape synchronization of cortical activity (Druga, 2009; Galarreta et
al., 2002; Gibson et al., 1999; Hestrin and Galarreta, 2005). INs also make chemical synapses
onto INs, in particular, accumulating evidence indicate that some INs preferentially target INs
from the same and/or different types, For instance, fast spiking PV-expressing INs mainly
project onto INs from the same type, often via reciprocal connections (Galarreta et al., 2002;
Pfeffer et al., 2013; Tamas et al., 1998). This mutual connectivity is supposed to play a key
role in the genesis of gamma oscillations (Bartos et al., 2007). The functional consequence of
inhibition of INs is the disinhibition of the targeted cells. For example, by targeting fast
spiking PV-expressing INs, CR- or VIP-expressing INs can inhibit the powerful perisomatic
inhibition onto PNs and consequently disinhibit their somatic compartment (see Discussion).
An important aspect concerning cortical processing is that excitatory activity is permanently
balanced by inhibitory inputs (Isaacson & Scanziani, 2011; Yizhar et al., 2011). This is due to
the fact that INs and PNs are highly interconnected and form feedback or feedforward
inhibitory circuits. These circuits serve to control the duration of PNs excitability and the

- 17 -
strength of oncoming excitatory input allowing a fine spatial and temporal tuning of PNs
activity (Pouille & Scanziani, 2001). A second prominent aspect concerning cortical
processing is the presence of strong oscillatory activity. Oscillations are thought to be
essential for cortical function, especially because oscillations can temporally organize cortical
activity (Buzsaki & Draguhn, 2004). Accumulating evidence indicates that neuronal
inhibition is necessary for the generation and maintenance of cortical oscillations (Buzsaki &
Chrobak, 1995; Buzsaki et al., 2004, 2012; Issacson & Scanziani, 2011; Moore et al., 2010).
In particular, among different INs type, it has been shown that fast spiking PV basket cells
play a key role, at least in the genesis of gamma oscillations (30 to 80 Hz, Cardin et al., 2010;
Cobb et al., 1995; Sohal et al., 2009).
II/ Role of cortical circuits in conditioned fear behaviour
A/ Role of the AC in the acquisition of conditioned fear behaviour
The main function of the auditory system is to convert external auditory stimuli into
behaviorally relevant internal information. The AC is considered at the higher order auditory
areas. It is involved in the processing, and the fine tuning of auditory information allowing
their sensation and perception (Carlyon, 2004; Malmierca, 2003; Micheyl et al., 2007).
Besides its role in sensory processes, the AC is thought to be involved in learning and
memory processes such as the acquisition of auditory fear conditioning. The acquisition of
auditory fear conditioning relies on the formation of CS-US associations (Rescorla, 1988;
Weinberger, 2011). Cortical and thalamic sensory inputs convey information relative to the
CS and the US to different brain structures. The convergence of auditory and nociceptive
sensory inputs onto the same neuron is a prerequisite for associative fear memories formation
via the induction of plasticity mechanisms such as long term potentiation (LTP), one of the
most prevalent cellular model of associative learning (Blair et al., 2001; Bliss & Lomo, 1973;
Fanselow & LeDoux 1999; Madison et al., 1991; Hebb, 1949). During auditory fear
conditioning, CS sensory inputs can reach the amygdala, specifically the LA, directly via the
auditory thalamus or indirectly via the AC (LeDoux et al., 1991; Romanski & LeDoux, 1992).
Whereas, the direct thalamic pathway has been demonstrated to be involved in the formation
of fear memories (Johansen et al., 2010; LeDoux, 2000; Rogan et al., 1997), the contribution
of the AC in the acquisition of fear behaviour has been intensively debated over past years
(Weinberger, 2011).

- 18 -
For instance, numerous studies have directly evaluated using classical electrolytic
lesions the role of AC in auditory fear conditioning. Pre-training lesions of the MG, the main
source of auditory inputs to the LA and the AC, prevents fear conditioning (Campeau &
Davis, 1995; LeDoux et al., 1984; Romanski & LeDoux, 1992). In contrast, a lesion
encompassing the medial division of the MG, the posterior intralaminar nucleus, and the
suprageniculate nucleus that project to the BLA and Au1 or a lesion encompassing the ventral
and dorsal divisions of the MG that project to the AuV, did not disrupt auditory fear
conditioning (Antunes & Moita, 2010; Campeau & Davis, 1995; Romanski & LeDoux, 1992).
Furthermore, pre-training lesions of the AC failed to block auditory fear conditioning
(Campeau & Davis, 1995; LeDoux et al., 1984; Romanski & LeDoux, 1992; Teich et al,.
1988). Interestingly, post-training lesions of the AC indicated that this structure is an essential
relay for CS inputs to produce conditioned fear expression (Boatman and Kim, 2006; Jarrell et
al., 1987; Quirk et al., 1997). Additional data support the notion that the AC is necessary for
fear acquisition under specific conditions, in particular when the CS used are complex tones
(Jarrell et al, 1987; Ledoux, 2000), or when the training procedure requires CSs
discrimination (Teich et al., 1988; Weinberger, 2007). Beside the role of AC in the
transmission of CS inputs to the amygdala, accumulating evidence indicated that auditory
conditioning produces plasticity in AC, an observation strongly suggesting that AC plays an
important role in the encoding of CS-US associations (Quirk et al., 1997; Suga and Ma, 2003;
Weinberger, 1998, 2004, 2007). In particular, fear conditioning produces long-lasting changes
in CS-evoked firing responses of AC neurons. Indeed, Quirk et al., 1997 demonstrated using
single unit recordings in behaving animals that AuV neurons also undergo plasticity during
auditory fear conditioning (Quirk et al., 1997). More recently, fear acquisition was shown to
retune the receptive field of AC neurons causing a shift of their “preferred tone frequency”.
This retuning produced a tonotopic remapping that consist in an over representation of the
neurons responding to the CS frequency associated with the US (Rutkowski & Weinberger,
2005; Weinberger, 2004). Interestingly, AC neurons also respond to the US during auditory
fear conditioning, an observation suggesting that they could be involved in the formation of
CS-US associations (Peter et al., 2012). Importantly, several studies have pointed out the
importance of Au1 cholinergic inputs emanating from the NBM to induce plastic changes
during fear acquisition (Weinberger, 2004). In particular, it has been shown that direct
application of cholinergic agonists into the Au1 during conditioning promoted AC neuronal
retuning, whereas application of antagonists prevented it (Ji et al., 2001; Ji & Suga, 2003).
Moreover, whereas pairing of tones with NBM electrical stimulation elicited plasticity in AC,

- 19 -
NBM lesion disrupted it (Bakin & Weinberger 1996; Edeline et al., 1994; Froemke et al.,
2007; Hars et al., 1993; Ma & Suga, 2003; Metherate & Ashe, 1991). Furthermore, it has
been shown that AC inputs to LA neurons could display plastic changes following auditory
fear conditioning in vitro and in vivo (Doyère et al., 2003; Shin et al., 2006; Tsvetkov et al.,
2002; Weisskopf & LeDoux, 1999). Taken together, all these studies strongly suggest that the
AC (comprising the AU1 and AuV) could be an important brain area involved in the
development of associative plasticity during fear acquisition although a direct demonstration
of this hypothesis is still lacking (Fig. 6).
Figure 6. Neuronal circuits involved in fear conditioning. During auditory fear conditioning, information
relative to the tone (CS) reaches the medial geniculate body of the thalamus (MG) and the posterior intralaminar
nucleus of the thalamus (PIN). Then CS and electrical shock (US) converge to the amygdala (BLA) and both
Au1 and AuV. In Au1, plastic changes related to fear acquisition require a release of acetylcholine from the
NBM. Afterwards, Au1 neurons influence fear responses via their projections to the AuV that in turn target the
BLA. Finally, the BLA through its central nucleus control fear responses.
B/ Role of the mPFC in the expression/extinction of conditioned fear behaviour
Following auditory fear conditioning, presentation of the CS evokes a broad range of
conditioned fear responses including defensive behaviour (freezing, avoidance), autonomic
(heart rate, blood pressure), and endocrine (stress hormone release) responses. Expression of
these conditioned fear responses is thought to be controlled by specific neuronal circuits
within the medial part of the central nucleus of the amygdala (CEm), which projects to the
hypothalamus and different brainstem areas (Cassell & Wright, 1986; Hopkins & Holstege,
1978; Veening et al., 1984). In particular, the CEm projects to the autonomic nervous system
(autonomic fear responses), the hypothalamic-pituitary axis (hormonal fear responses) and the

- 20 -
PAG (defensive fear responses). Recent studies demonstrated that additional neuronal
structures could also be involved in the expression of conditioned fear responses. Indeed,
recent data have demonstrated that the MD the ventral hippocampus (vHPC) and the mPFC
could play a role in the expression of conditioned fear behaviour (Sierra-Mercado et al., 2010;
Padilla-Coreano et al., 2012). The mPFC is involved in the regulation of a broad range of
brain functions related to attention, executive control or working memory. The capacity of the
mPFC to sustain such complex behaviours is thought to reside in its numerous connections
with a wide range of brain structures (Miller & Cohen, 2001; Benchenane et al., 2011; Hok et
al., 2013 Hirel et al., 2013). Besides these functions, the mPFC is known to regulate
emotional behaviour (Fuster, 2008), and dysfunction of the mPFC has been related to
psychiatric conditions such as post-traumatic stress disorder (Pitman et al., 2012; Shin &
Liberzon, 2010). Because of the potential clinical implications of these findings, numerous
animal studies have been conducted over the past decades in order to reveal the precise role of
mPFC in modulating fear behaviour. This literature is reviewed in the next section.
The role of the mPFC in the modulation of fear behaviour has long been discussed. An
early evidence of mPFC involvement in learned fear can be traced back more than 50 years
ago with experimental data showing that post-conditioning frontal lobotomy eliminates
conditioned fear responses in rats and monkeys (Maher & McIntire, 1960; Streb & Smith,
1955; Waterhouse, 1957). More recently, lesions and inactivation have been used to evaluate
the role of mPFC in cued fear conditioning in rodents. Because of conflicting results gathered
using these techniques and for the sake of clarity, the main findings are first described in the
text below and further summarized in Table 1. Pre- and post-training lesions of the dorsal
mPFC, including ACC and dorsal PL, enhanced fear conditioning (Morgan & LeDoux, 1995,
Vouimba et al., 2000, but see Bissiere et al., 2008) and blocked fear extinction (Morgan &
LeDoux, 1995). In addition, pre-training electrolytic or pharmacological lesions of the ventral
mPFC, including the ventral PL and IL, had no effect on fear conditioning but selectively
blocked extinction of fear conditioning (Morgan et al., 1993; Morrow et al., 1999b, but see
Fernandez Espejo, 2003 and Lacroix et al., 2000). Post-conditioning lesions of the ventral
mPFC including the ventral PL and IL produced somewhat inconsistent results as some
studies reported blockade of fear expression or extinction (Frysztak & Neafsey, 1991;
Morrow et al., 1999b), whereas another did not (Morgan et al., 2003). Later on, several
groups extended these findings by showing that pre-training ventral mPFC lesions blocked the
consolidation but not the original formation of fear extinction memories (Quirk et al., 2000;
Lebron et al., 2004; Tian et al., 2011). Although these data suggest that distinct mPFC

- 21 -
subregions might play a differential role in fear behaviour, it is worth noting that a number of
studies did not find any effect of mPFC lesions on fear conditioning or extinction behaviour
(Farinelli et al., 2006; Garcia et al., 2006; Gewirtz et al., 1997; Holson, 1986; Rosen et al.,
1992). The use of local, reversible, pharmacological inactivation has also yielded contrasting
results as inactivation of the ventral PL and IL impaired between session extinction, prevented
discrimination of a non-conditioned tone, increased, decreased, or did not change fear
expression during a test performed after extinction (Lee & Choi, 2012; Morawska & Fendt,
2012; Resstel et al., 2006; Sierra-Mercado et al., 2006). The general lack of consistency of
prefrontal lesions and inactivation might be explained by several factors including the time at
which the lesion was performed related to fear acquisition, the precise region targeted, the
training procedure, or species and sex differences as recently suggested (Baran et al., 2010;
Chang & Maren, 2010). Restricted pre-training or post-training inactivation of ACC or PL
provided more consistent results. Namely, pre-training inactivation of the ACC blocked fear
acquisition (Bissiere et al., 2008; Sacchetti et al., 2002; Tang et al., 2005). Furthermore,
restricted pre- or post-inactivation of the PL consistently reduced fear expression without
altering extinction learning (Corcoran & Quirk, 2007; Laurent & Westbrook, 2009; Sierra-
Mercado et al., 2011; Stevenson, 2011). In contrast, when restricted to the IL, the same
manipulations had no effect on fear expression but impaired within session extinction
(Laurent & Westbrook, 2009; Sierra-Mercado et al., 2011), although one study reported a
facilitating effect on extinction (Akirav et al., 2006). Interestingly, post-training activation of
IL using the GABAA receptor antagonist picrotoxin facilitates between session extinction of
fear, further supporting a role of IL in consolidation of fear extinction (Chang & Maren, 2011;
Thompson et al., 2010). Finally, in accordance with previous lesion studies (Morgan &
LeDoux, 1995; Vouimba et al., 2000), targeted pre-training activation of the ACC using the
mGluR agonist tACPD or the GABAA receptor antagonist bicuculline revealed an
enhancement of fear behaviour, suggesting an involvement of the ACC in the acquisition of
fear behaviour (Bissiere et al., 2008; Tang et al., 2005). In summary, studies using massive
lesions or inactivation of overlapping mPFC regions have produced inconsistent results
whereas more delimited manipulations of, ACC, PL and IL have produced consistent and
specific effects. In particular, these data strongly suggest that (i) ACC plays a key role in the
formation of aversive memories, (ii) PL mediates fear expression and (iii) IL is involved in
the consolidation of fear extinction memories (Fig. 7).
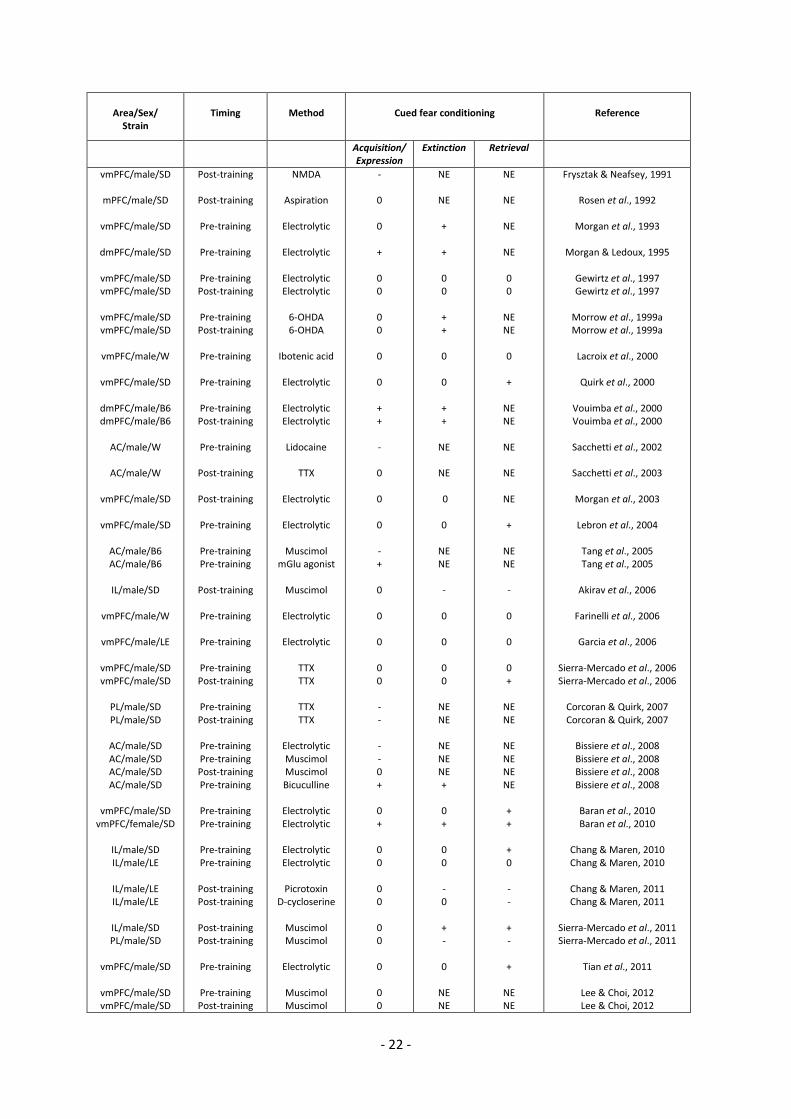
- 22 -
Area/Sex/
Strain
Timing
Method
Cued fear conditioning
Reference
Acquisition/Expression
Extinction
Retrieval
vmPFC/male/SD
mPFC/male/SD
vmPFC/male/SD
dmPFC/male/SD
vmPFC/male/SD vmPFC/male/SD
vmPFC/male/SD vmPFC/male/SD
vmPFC/male/W
vmPFC/male/SD
dmPFC/male/B6 dmPFC/male/B6
AC/male/W
AC/male/W
vmPFC/male/SD
vmPFC/male/SD
AC/male/B6 AC/male/B6
IL/male/SD
vmPFC/male/W
vmPFC/male/LE
vmPFC/male/SD vmPFC/male/SD
PL/male/SD PL/male/SD
AC/male/SD AC/male/SD AC/male/SD AC/male/SD
vmPFC/male/SD
vmPFC/female/SD
IL/male/SD IL/male/LE
IL/male/LE IL/male/LE
IL/male/SD PL/male/SD
vmPFC/male/SD
vmPFC/male/SD vmPFC/male/SD
Post-training
Post-training
Pre-training
Pre-training
Pre-training Post-training
Pre-training Post-training
Pre-training
Pre-training
Pre-training Post-training
Pre-training
Post-training
Post-training
Pre-training
Pre-training Pre-training
Post-training
Pre-training
Pre-training
Pre-training Post-training
Pre-training Post-training
Pre-training Pre-training Post-training Pre-training
Pre-training Pre-training
Pre-training Pre-training
Post-training Post-training
Post-training Post-training
Pre-training
Pre-training Post-training
NMDA
Aspiration
Electrolytic
Electrolytic
Electrolytic Electrolytic
6-OHDA 6-OHDA
Ibotenic acid
Electrolytic
Electrolytic Electrolytic
Lidocaine
TTX
Electrolytic
Electrolytic
Muscimol
mGlu agonist
Muscimol
Electrolytic
Electrolytic
TTX TTX
TTX TTX
Electrolytic Muscimol Muscimol Bicuculline
Electrolytic Electrolytic
Electrolytic Electrolytic
Picrotoxin
D-cycloserine
Muscimol Muscimol
Electrolytic
Muscimol Muscimol
-
0
0
+
0 0
0 0
0
0
+ + -
0
0
0 - +
0
0
0
0 0 - - - - 0 +
0 +
0 0
0 0
0 0
0
0 0
NE
NE
+
+
0 0
+ +
0
0
+ +
NE
NE
0
0
NE NE
-
0
0
0 0
NE NE
NE NE NE +
0 +
0 0 - 0
+ -
0
NE NE
NE
NE
NE
NE
0 0
NE NE
0
+
NE NE
NE
NE
NE
+
NE NE
-
0
0
0 +
NE NE
NE NE NE NE
+ +
+ 0 - -
+ -
+
NE NE
Frysztak & Neafsey, 1991
Rosen et al., 1992
Morgan et al., 1993
Morgan & Ledoux, 1995
Gewirtz et al., 1997 Gewirtz et al., 1997
Morrow et al., 1999a Morrow et al., 1999a
Lacroix et al., 2000
Quirk et al., 2000
Vouimba et al., 2000 Vouimba et al., 2000
Sacchetti et al., 2002
Sacchetti et al., 2003
Morgan et al., 2003
Lebron et al., 2004
Tang et al., 2005 Tang et al., 2005
Akirav et al., 2006
Farinelli et al., 2006
Garcia et al., 2006
Sierra-Mercado et al., 2006 Sierra-Mercado et al., 2006
Corcoran & Quirk, 2007 Corcoran & Quirk, 2007
Bissiere et al., 2008 Bissiere et al., 2008 Bissiere et al., 2008 Bissiere et al., 2008
Baran et al., 2010 Baran et al., 2010
Chang & Maren, 2010 Chang & Maren, 2010
Chang & Maren, 2011 Chang & Maren, 2011
Sierra-Mercado et al., 2011 Sierra-Mercado et al., 2011
Tian et al., 2011
Lee & Choi, 2012 Lee & Choi, 2012

- 23 -
vmPFC/male/B6
Pre-training
Muscimol
0
+
+
Morawska & Fendt, 2012
Table 1. Effect of mPFC lesions, inactivations and activations on cued fear conditioning. D: Sprague
Dawley rats; W: Wistar rats; LE: Long Evans rats; LH: Lister Hooded rats; B6: C57BL6 mice; dmPFC: dorsal
medial prefrontal cortex; vmPFC: ventral medial prefrontal cortex; AC: cingulate cortex; PL: prelimbic area; IL:
infralimbic area; 6-OHDA: 6-hydroxydopamine; NMDA: N-Methyl-D-Aspartate; TTX: tetrodotoxin; CoCl2:
Cobalt chloride; +: Fear increased in comparison to control animals; -:Fear decreased in comparison to control
animals; 0: no change relative to control; NE: not evaluated.
The identification of a functional role for the mPFC in mediating fear behaviour has
also gained support from histochemical studies using immediate early genes (IEG), such as c-
fos and zif-268, as markers of neuronal activity. Essentially, this approach assumes that the
formation of fear memories depends on specific signaling cascades that induce IEG
transcription and ultimately de novo protein synthesis to stabilize consolidation of fear
memories. Following behavioural training, detection of IEGs in the brain is thought to
indicate where and when neurons were activated during learning. Although this approach is
widely accepted and validated in neuroscience research, it is still not clear if all neuronal
types and neuronal structures express IEGs, whether some brain areas show constitutive IEGs
expression, and if the lack of IEG expression necessarily precludes the involvement of a
subset of neurons in a particular functional circuit. An early observation of IEGs expression in
the mPFC following conditioned fear behaviour was reported by Smith et al. (1992) who
observed increased c-fos expression relative to controls in the ACC of rats exposed to a CS
previously paired with footshock (Smith et al., 1992). Subsequent studies revealed increased
c-fos and zif-268 levels in PL and IL following fear conditioning and extinction learning
(Herry & Mons, 2004; Kim et al., 2010; Morrow et al., 1999a; Santini et al., 2004). However,
examination of c-fos expression in more dorsal prefrontal areas, including ACC, following
extinction learning is less clear (Morrow et al., 1999a; Plendl & Wotjak, 2010; Santini et al.,
2004). It is worth noting that one study using functional metabolic mapping observed elevated
activity in ACC and IL during fear extinction (Barrett et al., 2003). Interestingly, suppressing
c-fos expression in PL and IL using injections of a DNA antisense oligonucleotide during fear
acquisition reduced subsequent fear expression, suggesting that c-fos expression in PL and IL
might be necessary for the consolidation or expression of fear behaviour (Morrow et al.,
1999a). Recently, a number of studies have examined IEG expression following retrieval after
extinction. The lack of extinction memory consolidation, as assessed by the amount of
freezing during post-extinction fear memory retrieval was correlated with reduced (c-fos) or
no changes (zif268) in IEG expression within PL and IL (Herry & Mons, 2004). Alternatively,
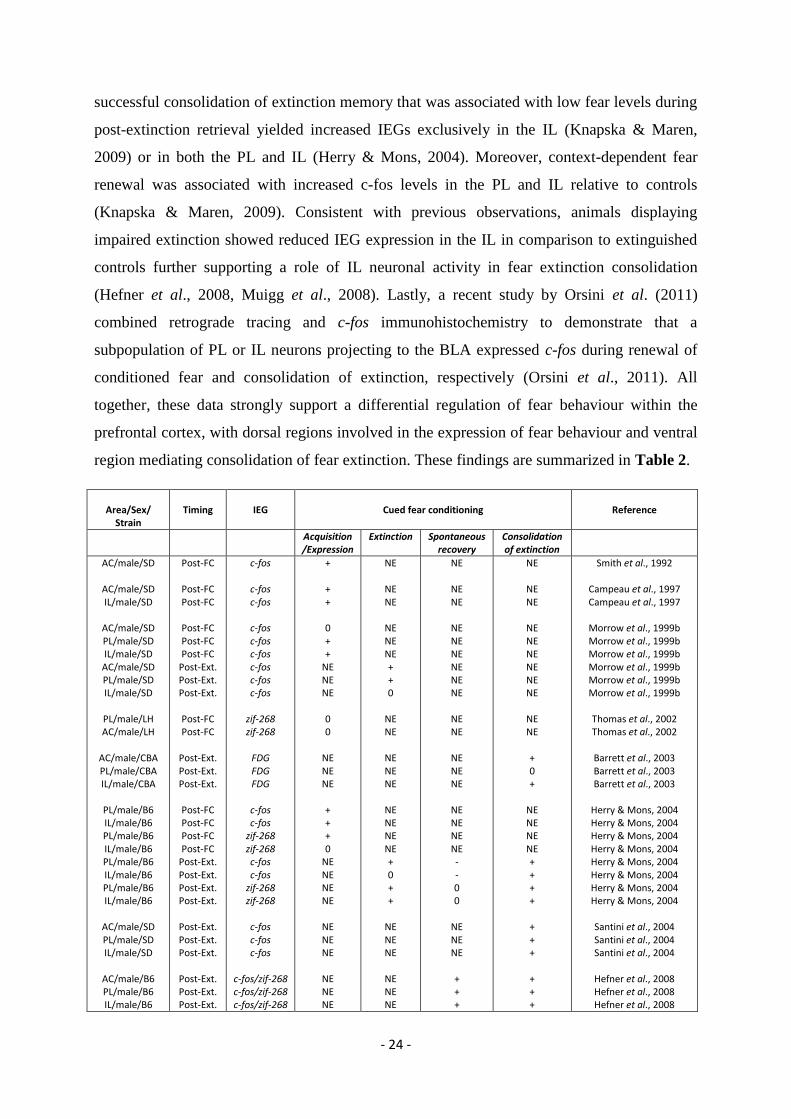
- 24 -
successful consolidation of extinction memory that was associated with low fear levels during
post-extinction retrieval yielded increased IEGs exclusively in the IL (Knapska & Maren,
2009) or in both the PL and IL (Herry & Mons, 2004). Moreover, context-dependent fear
renewal was associated with increased c-fos levels in the PL and IL relative to controls
(Knapska & Maren, 2009). Consistent with previous observations, animals displaying
impaired extinction showed reduced IEG expression in the IL in comparison to extinguished
controls further supporting a role of IL neuronal activity in fear extinction consolidation
(Hefner et al., 2008, Muigg et al., 2008). Lastly, a recent study by Orsini et al. (2011)
combined retrograde tracing and c-fos immunohistochemistry to demonstrate that a
subpopulation of PL or IL neurons projecting to the BLA expressed c-fos during renewal of
conditioned fear and consolidation of extinction, respectively (Orsini et al., 2011). All
together, these data strongly support a differential regulation of fear behaviour within the
prefrontal cortex, with dorsal regions involved in the expression of fear behaviour and ventral
region mediating consolidation of fear extinction. These findings are summarized in Table 2.
Area/Sex/
Strain
Timing
IEG
Cued fear conditioning
Reference
Acquisition/Expression
Extinction
Spontaneous recovery
Consolidation of extinction
AC/male/SD
AC/male/SD IL/male/SD
AC/male/SD PL/male/SD IL/male/SD AC/male/SD PL/male/SD IL/male/SD
PL/male/LH AC/male/LH
AC/male/CBA PL/male/CBA IL/male/CBA
PL/male/B6 IL/male/B6 PL/male/B6 IL/male/B6 PL/male/B6 IL/male/B6 PL/male/B6 IL/male/B6
AC/male/SD PL/male/SD IL/male/SD
AC/male/B6 PL/male/B6 IL/male/B6
Post-FC
Post-FC Post-FC
Post-FC Post-FC Post-FC
Post-Ext. Post-Ext. Post-Ext.
Post-FC Post-FC
Post-Ext. Post-Ext. Post-Ext.
Post-FC Post-FC Post-FC Post-FC
Post-Ext. Post-Ext. Post-Ext. Post-Ext.
Post-Ext. Post-Ext. Post-Ext.
Post-Ext. Post-Ext. Post-Ext.
c-fos
c-fos c-fos
c-fos c-fos c-fos c-fos c-fos c-fos
zif-268 zif-268
FDG FDG FDG
c-fos c-fos
zif-268 zif-268 c-fos c-fos
zif-268 zif-268
c-fos c-fos c-fos
c-fos/zif-268 c-fos/zif-268 c-fos/zif-268
+
+ +
0 + +
NE NE NE
0 0
NE NE NE
+ + + 0
NE NE NE NE
NE NE NE
NE NE NE
NE
NE NE
NE NE NE + + 0
NE NE
NE NE NE
NE NE NE NE + 0 + +
NE NE NE
NE NE NE
NE
NE NE
NE NE NE NE NE NE
NE NE
NE NE NE
NE NE NE NE - - 0 0
NE NE NE
+ + +
NE
NE NE
NE NE NE NE NE NE
NE NE
+ 0 +
NE NE NE NE + + + +
+ + +
+ + +
Smith et al., 1992
Campeau et al., 1997 Campeau et al., 1997
Morrow et al., 1999b Morrow et al., 1999b Morrow et al., 1999b Morrow et al., 1999b Morrow et al., 1999b Morrow et al., 1999b
Thomas et al., 2002 Thomas et al., 2002
Barrett et al., 2003 Barrett et al., 2003 Barrett et al., 2003
Herry & Mons, 2004 Herry & Mons, 2004 Herry & Mons, 2004 Herry & Mons, 2004 Herry & Mons, 2004 Herry & Mons, 2004 Herry & Mons, 2004 Herry & Mons, 2004
Santini et al., 2004 Santini et al., 2004 Santini et al., 2004
Hefner et al., 2008 Hefner et al., 2008 Hefner et al., 2008
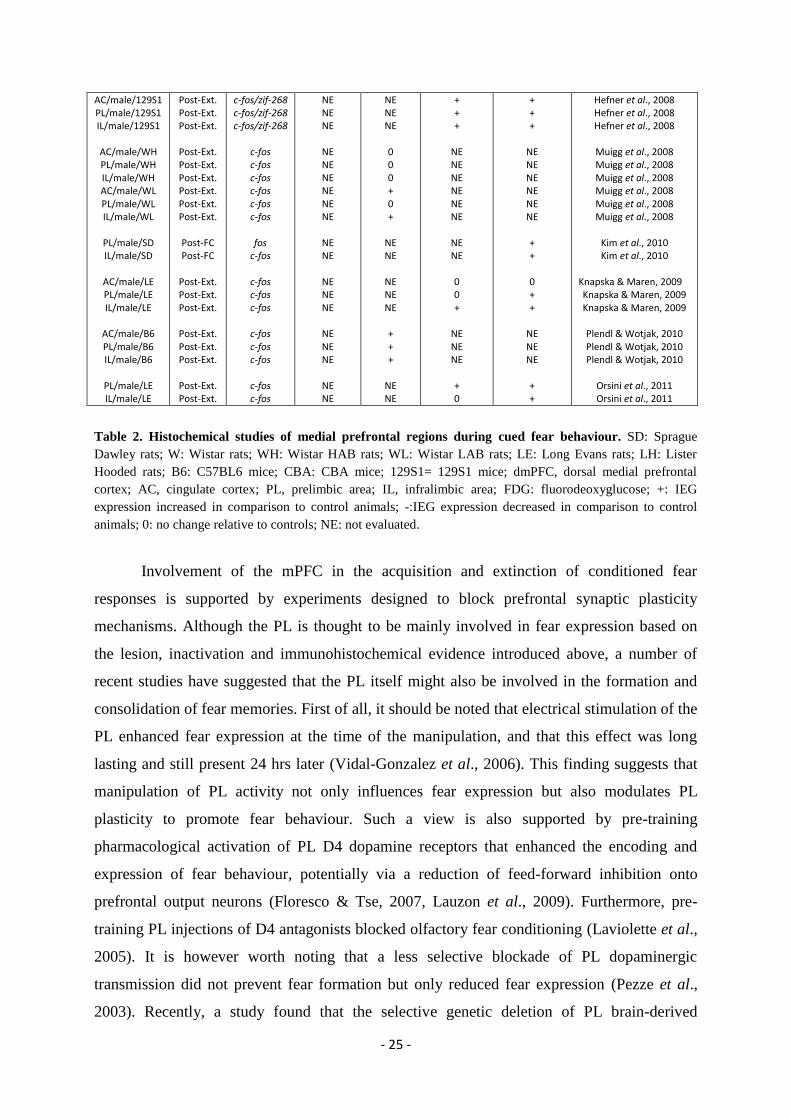
- 25 -
AC/male/129S1 PL/male/129S1 IL/male/129S1
AC/male/WH PL/male/WH IL/male/WH AC/male/WL PL/male/WL IL/male/WL
PL/male/SD IL/male/SD
AC/male/LE PL/male/LE IL/male/LE
AC/male/B6 PL/male/B6 IL/male/B6
PL/male/LE IL/male/LE
Post-Ext. Post-Ext. Post-Ext.
Post-Ext. Post-Ext. Post-Ext. Post-Ext. Post-Ext. Post-Ext.
Post-FC Post-FC
Post-Ext. Post-Ext. Post-Ext.
Post-Ext. Post-Ext. Post-Ext.
Post-Ext. Post-Ext.
c-fos/zif-268 c-fos/zif-268 c-fos/zif-268
c-fos c-fos c-fos c-fos c-fos c-fos
fos
c-fos
c-fos c-fos c-fos
c-fos c-fos c-fos
c-fos c-fos
NE NE NE
NE NE NE NE NE NE
NE NE
NE NE NE
NE NE NE
NE NE
NE NE NE
0 0 0 + 0 +
NE NE
NE NE NE
+ + +
NE NE
+ + +
NE NE NE NE NE NE
NE NE
0 0 +
NE NE NE
+ 0
+ + +
NE NE NE NE NE NE
+ +
0 + +
NE NE NE
+ +
Hefner et al., 2008 Hefner et al., 2008 Hefner et al., 2008
Muigg et al., 2008 Muigg et al., 2008 Muigg et al., 2008 Muigg et al., 2008 Muigg et al., 2008 Muigg et al., 2008
Kim et al., 2010 Kim et al., 2010
Knapska & Maren, 2009 Knapska & Maren, 2009 Knapska & Maren, 2009
Plendl & Wotjak, 2010 Plendl & Wotjak, 2010 Plendl & Wotjak, 2010
Orsini et al., 2011 Orsini et al., 2011
Table 2. Histochemical studies of medial prefrontal regions during cued fear behaviour. SD: Sprague
Dawley rats; W: Wistar rats; WH: Wistar HAB rats; WL: Wistar LAB rats; LE: Long Evans rats; LH: Lister
Hooded rats; B6: C57BL6 mice; CBA: CBA mice; 129S1= 129S1 mice; dmPFC, dorsal medial prefrontal
cortex; AC, cingulate cortex; PL, prelimbic area; IL, infralimbic area; FDG: fluorodeoxyglucose; +: IEG
expression increased in comparison to control animals; -:IEG expression decreased in comparison to control
animals; 0: no change relative to controls; NE: not evaluated.
Involvement of the mPFC in the acquisition and extinction of conditioned fear
responses is supported by experiments designed to block prefrontal synaptic plasticity
mechanisms. Although the PL is thought to be mainly involved in fear expression based on
the lesion, inactivation and immunohistochemical evidence introduced above, a number of
recent studies have suggested that the PL itself might also be involved in the formation and
consolidation of fear memories. First of all, it should be noted that electrical stimulation of the
PL enhanced fear expression at the time of the manipulation, and that this effect was long
lasting and still present 24 hrs later (Vidal-Gonzalez et al., 2006). This finding suggests that
manipulation of PL activity not only influences fear expression but also modulates PL
plasticity to promote fear behaviour. Such a view is also supported by pre-training
pharmacological activation of PL D4 dopamine receptors that enhanced the encoding and
expression of fear behaviour, potentially via a reduction of feed-forward inhibition onto
prefrontal output neurons (Floresco & Tse, 2007, Lauzon et al., 2009). Furthermore, pre-
training PL injections of D4 antagonists blocked olfactory fear conditioning (Laviolette et al.,
2005). It is however worth noting that a less selective blockade of PL dopaminergic
transmission did not prevent fear formation but only reduced fear expression (Pezze et al.,
2003). Recently, a study found that the selective genetic deletion of PL brain-derived

- 26 -
neurotrophic factor (BDNF) expression spared auditory fear conditioning acquisition and
extinction but impaired fear consolidation, an effect rescued with systemic injections of a
TrkB receptor agonist (Choi et al., 2010). Lastly, PL infusions of cannabinoid receptor CB1
antagonists and agonists blocked and potentiated olfactory fear conditioning, respectively
(Laviolette & Grace, 2006). A number of studies have consistently revealed that plasticity
blockade within the ventral mPFC does not impair the formation of extinction memories, but
selectively blocks consolidation of extinction. For instance, blockade of NMDA receptors in
the ventral mPFC impaired post-extinction retrieval (Burgos-Robles et al., 2007; Sotres-
Bayon et al., 2009). The same effect was observed upon blockade of noradrenergic and
dopaminergic receptors in IL or when anisomycin was used (Hikind & Maroun, 2008;
Mamiya et al., 2009; Mueller et al., 2008, 2009; Pfeiffer & Fendt, 2006; Santini et al., 2004).
Recent studies have investigated the role of prefrontal cannabinoids in fear extinction mainly
because activation of CB1 receptors is known to phosphorylate extracellular-regulated kinase
(ERK) during extinction. Moreover, inhibition of the ERK/MAPK signaling pathway in the
ventral mPFC impaired post-extinction retrieval (Cannich et al., 2004; Hugues et al., 2004,
2006). Although extinction of cued fear conditioning does not increase levels of the two major
endocannabinoids, anandamide and 2-arachidonoyglycerol in the ventral mPFC (Marsicano et
al., 2002), infusion of the CB1 antagonist AM251 within the IL blocked, whereas IL infusion
of the CB1 agonist WIN enhanced, extinction of fear potentiated startle (Lin et al., 2009).
However, it should be noted that the facilitation of extinction behaviour by the CB1 agonist
WIN was observed upon exposure to the conditioned or the control stimulus, suggesting that
WIN produced a general reduction of fear responses independent of the conditioned stimulus
(Lin et al., 2009).
1/ mPFC circuits modulating fear behaviour
Mounting evidence suggests that fear behaviour is differentially modulated by
neuronal circuits involving IL and PL subregions. It is thought that the opposite influence
these structures have on fear responding is orchestrated by differential connectivity with the
amygdala. The IL is thought to inhibit fear behaviour via its projections to the amygdala,
although the exact mechanism of this circuit remains the focus of some debate. Support for
this idea comes from findings showing that electrical stimulation of the IL inhibits the output
activity of the CEm, a subregion known to be necessary for the expression of specific fear
responses (Cassell & Wright, 1986; Hopkins & Holstege, 1978, Veening et al., 1984). This

- 27 -
IL-evoked inhibition is supposed to be indirect as the IL has very sparse direct projections to
CEm output neurons (McDonald et al., 1996). Accumulating evidence supports a model of
inhibitory circuits from the IL to the CEm, via the activation of intercalated (ITC) amygdala
neurons (Pare et al., 2004), a population of GABAergic cells surrounding the BLA
(Millhouse, 1986). Selective ablation of ITC cells after extinction training resulted in recovery
of fear responses, suggesting that ITC neurons are required for the expression of extinction
memory (Likhtik et al., 2008). Tracing studies have shown projections from the IL to ITC
clusters and to the capsular division of the CeA (Amir et al., 2011; McDonald et al., 1996;
Pinard et al., 2012; Vertes, 2004), some of which project to the CEm (Amir et al., 2011; Busti
et al., 2011; Geracitano et al., 2007). Pharmacological disinhibition of the IL increases the
number of c-fos immunoreactive ITC cells, indicating that their activity can be driven by
inputs from the IL (Berretta et al., 2005). Moreover, a recent study using juxtacellular
recordings and labeling of amygdala neurons in anesthetized rats showed that ITC cells were
uniformly responsive to IL stimulation, and displayed unique response patterns not found in
neighboring CeA or BA nuclei (Amir et al., 2011). Together, these findings support the idea
that IL excitatory outputs activate ITC cells, which leads to feedforward inhibition of CEm
neurons (Pare et al., 2004). Thus there is strong evidence for ITC cells involvement in the
expression of extinction memory, although it remains to be established whether or not ITC
cells are required for encoding extinction memory. Furthermore, it is still possible that ITC
cells integrate additional inputs from the BLA to set the level of inhibition of CEm neurons
during fear behaviour (Amano et al., 2011). In addition to an IL-ITC inhibitory circuit, it has
been suggested that the mPFC supports expression of extinction memory expression via IL
afferents to local BLA INs (Rosenkranz et al., 2003). Such an inhibitory pathway is unlikely
to be mediated by the BA, as IL outputs predominantly synapse onto PNs in the BA (Likhtik
et al., 2005). Additionally, BA activation is not necessary for the expression of extinction
memory (Anglada-Figueroa & Quirk, 2005; Herry et al., 2008). One possible pathway would
involve mPFC efferents to a subpopulation of LA PNs that show context-dependent inhibition
of CS-evoked activity (Hobin et al., 2003; Maren & Hobin, 2007; Repa et al., 2001).
In contrast to the inhibitory role of IL in fear memory, the PL has been hypothesized
to promote fear expression through its activation of BLA neurons projecting to the CeA (Pape
& Pare, 2010), although this possibility remains to be demonstrated. In line with the findings
showing that PL is involved in fear memory expression, but not fear extinction (Burgos-
Robles et al., 2009; Sierra-Mercado et al., 2011), it has been suggested that PL neuronal
circuits modulate renewal of extinguished conditioned fear responses (Knapska & Maren,

- 28 -
2009). Consistent with this hypothesis, a recent tracing study showed that during the retrieval
of extinction memory the dominant inputs to c-fos active neurons in the LA were from the IL,
whereas renewal of fear memory was associated with c-fos labeled neurons receiving inputs
from the PL and the vHPC (Knapska et al., 2012). Moreover, BA neurons that are
preferentially active during context-dependent fear renewal receive strong hippocampal
inputs, while BA neurons preferentially active during extinction do not (Herry et al., 2008).
These studies suggest that the hippocampus regulates neuronal activity in the BLA via direct
projections, although indirect regulation through the PL is also possible (Vertes, 2006).
Finally, the mPFC projects directly to the PAG, a neuronal structure involved in the
genesis of freezing responses and conditioned fear behaviour (Gabbott et al., 2005; Vianna et
al., 2001) and therefore may directly control fear behaviour at this level. To our knowledge,
the boundary conditions during which this direct mPFC-PAG pathway might be involved to
regulate fear responses are still unclear and will require further work.
Figure 7. Prefrontal control of fear expression and extinction. During fear expression (left) and extinction
(right) of auditory fear conditioning, tone responses from the amygdala (AMYG) get integrated by the prelimbic
(PL) or infralimbic (IL) prefrontal cortex with converging information from diverse sources such as
hippocampus (Hipp), brainstem monoamines (Bstm), mediodorsal thalamus (MD), and orbital prefrontal cortex
(OFC) to determine whether or not to produce a fear response. Fear excitation involves PL projections back to
basal amygdala (BA), whereas fear inhibition involves IL projections to amygdala-intercalated cells (ITC). In
turn, BA excites neurons in the medial division of the central nucleus of the amygdala (CeM) to produce fear
responses, while ITCs inhibit these amygdala output neurons thereby inhibiting fear responses. Thus, the same
conditioned stimulus (e.g. a tone) signals either high fear (red) or low fear (green) states in the appropriate
circumstances (from Sotres-Bayon & Quirk, 2010).
2/ mPFC elements encoding fear behaviour
To understand emotional processing in the cortex, the precise role of distinct neuronal
elements must be identified. Only a few studies have addressed this question by using single

- 29 -
neuron (unit) recordings in behaving animals submitted to fear conditioning and extinction.
For instance, it has been shown that IL, but not PL, neurons selectively increased their firing
rate to extinguished CSs (Milad & Quirk, 2002). In addition, bursting activity of IL neurons
during extinction learning predicts subsequent consolidation of extinction. Additionally, IL
stimulation during extinction learning enhanced extinction retrieval (Milad & Quirk, 2002,
Milad et al., 2004, Vidal-Gonzalez et al., 2006, Burgos-Robles et al., 2007, Kim et al., 2010).
A similar result was obtained when IL high frequency stimulation was applied before
extinction training (Maroun et al., 2012). In contrast IL low frequency stimulation impaired
consolidation of extinction (Shehadi & Maroun, 2012). In addition, ex-vivo experiments
performed on mPFC slices from previously conditioned and extinguished animals revealed
that fear conditioning and extinction training was associated with a decrease and increase of
IL neurons intrinsic excitability and bursting activity, respectively (Santini et al., 2008). This
increase in intrinsic excitability and bursting activity of IL neurons associated with
consolidation of extinction was dependent on noradrenaline signaling and mGluR5 receptors
activation and could be modulated by M-type potassium channels and cholinergic inputs
(Mueller et al., 2008, Santini & Porter, 2010, Fontanez-Nuin et al., 2011, Santini et al., 2012).
Together these results strongly suggest that the changes in the firing rate of IL pyramidal
neurons encode long-term extinction memories.
The opposite pattern was found with recordings targeting the PL area. Indeed,
neuronal activity of PL neurons did not correlate with extinction memories. Instead, ~15% PL
neurons acquired CS+-evoked responses with fear conditioning and ~25% PL neurons
displayed sustained conditioned tone-evoked responses during fear behaviour (Milad &
Quirk, 2002, Burgos-Robles et al., 2009). In addition, Burgos-Robles and colleagues (2009)
proposed that sustained activation of PL neurons confers resistance to extinction (Burgos-
Robles et al., 2009). The observations that low frequency stimulation-mediated long term
depression (LTD) of the monkey dACC, (analogous of the rodent PL) facilitated extinction
learning (Klavir et al., 2012), or that electrical stimulation of the rodent ACC or PL facilitated
fear expression (Vidal-Gonzalez et al., 2006) are also in line with the general view that
neuronal activity in the PL is associated with the expression of fear behaviour, although a
detailed understanding of the specific neuronal types and mechanisms mediating this effect is
still largely missing.
Concerning INs, only few studies have evaluated directly their role during fear
behaviour. In particular, one study using extracellular single unit recordings in behaving
rodents described transient modifications of tone-evoked neuronal responses in putative

- 30 -
prefrontal INs following fear learning (Baeg et al., 2001). More recently, local inactivation of
the vHPC that monosynaptically contact prefrontal INs was found to reduce the basal activity
of prefrontal INs and to increase fear behaviour following extinction learning as evidenced by
increased lever pressing to receive food during CS presentations (Sotres-Bayon et al., 2012;
Tierney et al., 2004). However, an unambiguous identification of INs with extracellular
recordings cannot be possible. These results are in line with the findings that genetic ablation
of NMDA receptors on prefrontal PV-expressing INs impaired cued and contextual fear
behaviour (Carlen et al., 2012). Nevertheless, the development of recent optogenetic
techniques that allow the identification and manipulation of excitatory and inhibitory neuronal
elements in combination with imaging or electrophysiological approaches will certainly
become instrumental to address the role of cortical INs in fear behaviour.
Nevertheless, several studies have reported findings partially inconsistent with the
above-mentioned studies. In particular, one study reported heterogeneous response patterns to
conditioned tones in the mPFC ranging from prolonged decrease, to increase or transient
increase in firing activity that were not different between IL and PL (Baeg et al., 2001). Two
other studies reported increased tone-evoked activity in PL and IL neurons during extinction
learning and later during post-extinction retrieval (Chang et al., 2010, Holmes et al., 2012),
whereas Milad and Quirk found this to be specific to the IL (Milad & Quirk, 2002). These
differences might be due to the task or a different spatial sampling of units in mPFC (e.g.
layer) or species in the last case.
The discrepancies observed above suggest that mPFC circuit operation may vary
across different types of fear memory or species. Moreover, distinct cortical layers that have
evolved differently within mPFC might serve dramatically different functions in emotions.
These issues will need further investigation. In particular, it will be crucial to keep recordings
in precise register with mPFC subregion and layer position, using standardized definitions.
For instance greater spatial sampling and precision could be achieved with the use of high
density silicon probes (Csicsvari et al., 2003, Einevoll et al., 2012). Finally, PL and IL
boundaries which are best defined by their connectivity are determined quite arbitrarily in
most studies. It will be necessary in the near future to identify molecular markers that will
allow a precise separation between the IL and PL.

- 31 -
III/ Neuronal encoding of fear behaviour in the cortex
As mentioned earlier, AC and mPFC neuronal elements likely play a pivotal role in
fear behaviour. However, the fear related processes and the neuronal mechanisms involved
are still largely unknown. Most of the progress in the understanding of the cortical encoding
of fear behaviour comes from studies reporting correlations between firing rates and
behavioural outcome (Burgos-Robles et al., 2009; Quirk et al., 1997; Milad & Quirk, 2002).
This coding scheme is referred to as ‘‘rate coding’’ and can account for many aspects of
neuronal coding in the brain. It implies that precise firing patterns of neurons are less
important than their average firing rates, and that encoding results from changes in these
average firing rates. However, in addition, neurons with different and specific firing
sequences may cooperate and collectively provide information, which is referred to as
‘‘temporal coding’’ (Buzsaki & Chrobak, 1995; Macleod et al., 1998; Nicolelis et al., 1995;
Singer, 1999). In temporal coding, precise timing of firing is important, while average firing
rates can remain the same following learning. Furthermore, this temporal organization of the
firing sequences of different neurons leads to their transient and simultaneous activation in
order to form a neuronal assembly (Hebb, 1949). One obvious advantage of temporal coding
is the plasticity it affords. Thus, neurons might rapidly shift between multiple networks
according to sensory and internal inputs, a mechanism that strongly enhances coding power.
Importantly, temporal coding allows for the synchronization of spiking activity within and
among neuronal circuits for the accurate transmission of information during behavioural
responses. Indeed, synchronized inputs have a stronger impact on targeted neurons than
unsynchronized ones (Buzsaki & Draguhn, 2004; Sakata et al., 2005; Salinas & Sejnowski,
2001; Singer, 1999; Varela et al., 2001; Womelsdorf et al., 2007). A necessary condition of
temporal coding is a tight control of firing sequences. Brain oscillations are thought to be
instrumental in temporal coding through, binding cell assemblies, organizing individual firing
into meaningful collective activity, and coordinating remote areas. A prominent property of
cortical circuits is their capacity to sustain rhythmic activity, also referred to as cortical
oscillations. Cortical oscillations can be studied in vivo with local field potential (LFP)
recordings that reflect current (i.e. ion) flow through plasma membranes, including synaptic
currents, voltage-dependent membrane oscillations, and calcium spikes. Indeed, LFPs are
thought to reflect global ongoing activity in neuronal populations surrounding the recording
electrode (Buzsaki et al., 2012). LFPs are characterized by three principle properties:
amplitude, frequency and phase. LFP recordings provide a powerful tool to explore cortical

- 32 -
activity at the level of population dynamics within and between brain regions. Cortical
oscillations have been observed in different species, in multiple behavioural tasks, either
related to the behavioural state or evoked by external stimuli (Benchenane et al., 2010; Gray
& Singer, 1989; Peyrache et al., 2009; Sakata et al., 2005; Siapas et al., 2005; Sirota et al.,
2008).
Coordinated oscillations could for instance increase synaptic strength, through
coincident pre and post-synaptic activation and simultaneous convergence of multiple inputs
(Buzsaki & Draguhn, 2004; Fell & Axmacher, 2011; Markram et al., 1997). Coordinated
activity between the cortex and its synaptic partners represents a possible mechanism that
facilitates communication between remote structures to control fear behaviour. For instance,
in the mPFC, following fear conditioning, coherent theta (3-12 Hz) oscillations between the
mPFC and BLA during paradoxical sleep are positively correlated with fear memory
consolidation (Popa et al., 2010). Directional analysis indicated that the BLA led this
coordinated activity, in agreement with the potentiation observed in the BLA-mPFC pathway
(Popa et al., 2010; Vouimba & Maroun, 2011). Similar analysis performed during extinction
learning revealed decreased correlations between mPFC and hippocampal or BLA theta
oscillations (Lesting et al., 2011). Interestingly, genetic knockout of the serotonin transporter
led to both impaired extinction memory and persistent theta correlation between LA and IL
during and after extinction (Narayanan et al., 2011). Moreover, theta synchronization between
the vHPC and the mPFC was recently shown to be enhanced during anxious behaviour
(Adhikari et al., 2010, 2011). More recently, in an elegant study using a partial reinforcement
learning task that confers resistance to extinction in monkey, Livneh & Paz (2012) observed
that the synchronized firing of mPFC and amygdala neurons was correlated with resistance of
fear extinction (Livneh & Paz, 2012), possibly by a potentiation of the mPFC-BLA pathway
(Vouimba & Maroun, 2011). These results strongly suggest that neuronal synchrony between
mPFC, BLA, and hippocampus is associated with high fear states and anxious behaviour,
perhaps because it promotes synaptic plasticity. Conversely, neuronal loss of correlation
between these neuronal structures is associated with inhibition of fear behaviour. Decreased
correlation might promote depotentiation of fear-related synaptic links and allow a takeover
of extinction networks.
As for the AC, to our knowledge, there is no report on coordinated oscillatory activity
between AC and its synaptic partners, but some studies have identified oscillatory activity
within the AC linked to fear behaviour. For instance, these studies suggested that fast
oscillations in the gamma range (30-80 Hz) play a key role in local plastic changes observed

- 33 -
during fear acquisition (Headley & Weinberger, 2011, 2013; Weinberger et al., 2006, 2013).
Gamma oscillations are thought to be essential in numerous cortical functions such as sensory
processes, attentional selection and memory performance by allowing the transfer of
information between cortical areas and/or the synchronization of neuronal ensembles
(Buzsaki & Wang, 2012; Canolty et al., 2006; Cardin et al., 2009; Engel et al., 2001; Fries et
al., 2007). In particular, gamma oscillations are supposed to play a role in synaptic plasticity
by synchronizing neurons in the critical time window of spike-timing-dependent plasticity
(around 10-30 ms; Buzsaki & Wang, 2012; Harris et al., 2003). During fear acquisition, Au1
gamma oscillations increased in parallel with the strength of fear acquisition and its relative
plastic changes. Indeed, in two recent electrophysiological studies, Headley & Weinberger
(2011 and 2013) indicate that gamma oscillation power and gamma phase locking of Au1
neurons progressively increased during fear acquisition (Headley & Weinberger, 2011 and
2013). Furthermore, gamma oscillations in Au1 could be elicited by the release of
acetylcholine from the NBM (Metherate et al., 1992; Weinberger et al., 2006) which is
required for plastic changes induced by fear conditioning (Froemke et al., 2007). Taken
together, these data suggested that Au1 gamma oscillations participate to the establishment of
plastic changes occurring during fear conditioning.
Despite these recent progresses, additional work will be required to dissect the
principles that govern the encoding of fear behaviour in cortical areas. In particular, it would
be interesting to determine the contribution of both rate and temporal coding, but also to
precisely define the role of cortical oscillations and their link with the activity of the different
neuronal elements.
Parts of this introduction have been published as a review paper:
“Medial prefrontal cortex neuronal circuits in fear behavior”.
Julien Courtin, Thomas C. Bienvenu, Einar O. Einarsson & Cyril Herry.
Neuroscience 240, 219-242 (2013).

- 34 -
AIM OF THE THESIS
This thesis addresses the fundamental question of how behaviour is controlled at the
level of defined neuronal circuits. In the past decades, a number of seminal studies have
identified precisely the role of subcortical structures and their specific neuronal elements in
the acquisition, expression and extinction of conditioned fear behaviour. In contrast, the role
of cortical structures in fear behaviour is still poorly understood and only a few studies have
investigated in detail the role of specific cortical neuronal elements and mechanisms in the
regulation of fear responses. Previous reports have indicated that the AC might be involved in
the formation of conditioned fear memories, whereas the dorsal medial prefrontal cortex
(dmPFC) might be selectively involved in the expression of these memories. It is likely that
neuronal activity in the AC and dmPFC depends on specific interactions between INs and
PNs. In particular, PV INs are known to potently control cortical activity. Whereas the
anatomical and electrophysiological properties of PV INs are well described, their functional
role in the acquisition and expression of associative fear memory is largely unknown.
During my thesis, our objectives were twofold: firstly, in collaboration with the group
of Andreas Lüthi in Basel we investigated the role of AC inhibitory circuits during the
acquisition of auditory fear conditioning. Secondly, we evaluated the role of specific
populations of dmPFC PV INs and oscillatory mechanisms in the regulation of conditioned
fear expression. To achieve these goals we used a combination of single unit recordings and
optogenetic manipulations in behaving mice submitted to auditory fear conditioning.

- 35 -
MATERIAL AND METHODS
I/ Animals
Male C57BL6/J mice (3 months old, Janvier) and PV-IRES-Cre mice (3 months old,
Jackson Labs, B6;129P2-Pvalbtm1(cre)Arbr
/J) (Hippenmeyer et al., 2005) were individually
housed for at least 7 days prior to all experiments, under a 12 h light/dark cycle, and provided
with food and water ad libitum. All procedures were performed in accordance with standard
ethical guidelines (European Communities Directive 86/60-EEC) and were approved by the
committee on Animal Health and Care of Institut National de la Santé et de la Recherche
Médicale and French Ministry of Agriculture and Forestry (authorization A3312001).
II/ Behaviour
A/ Fear conditioning and extinction paradigm
Habituation and fear conditioning took place in context A consisting of a square
transparent plexiglas box (25cm side, 80 cm high) with a shock grid floor made of stainless
steel rods placed inside a sound attenuating and temperature regulated cubicle. The walls of
the cubicle were made of sound attenuating black foam. Inside the cubicle the floor and walls
of the plexiglas square were cleaned with a 70 % ethanol solution before and after each
session. Extinction learning and post-extinction fear retrieval were performed in context B
consisting of a cylinder transparent plexiglas (25 cm diameter, 40 cm high) with a grey plastic
floor placed inside a sound attenuating and temperature regulated cubicle. The walls of the
cubicle were made of sound attenuating white foam. The floor and walls of the plexiglas
cylinder were cleaned with a 1 % acetic acid solution before and after each session. To score
freezing behaviour an automated infrared beam detection system located on the bottom of the
experimental chambers was used (Coulbourn Instruments). The animals were considered to be
freezing if no movement was detected for 2 s. On day 1, C57BL6/J mice were submitted to an
habituation session in context A, in which they received 4 presentations of the CS+ and of the
CS- (total CS duration: 30 s, consisting of 27, 50 ms pips repeated at 0.9 Hz, 2 ms rise and
fall, pip frequency: 7.5 kHz or white-noise, 80 dB sound pressure level). Discriminative fear
conditioning was performed on the same day by pairing the CS+ with a US (1 s foot-shock,
0.6 mA, 5 CS+/US pairings; inter-trial intervals: 20-180 s). The onset of the US coincided
with the offset of the CS+. The CS
- was presented after each CS
+/US association but was

- 36 -
never reinforced (5 CS- presentations, inter-trial intervals: 20-180 s). The frequencies used for
CS+ and CS
- were counterbalanced across animals. On day 2 and day 3, conditioned mice
were submitted to extinction training (Post-FC and Ext. sessions) in context B during which
they received 4 and 12 presentations of the CS- and CS
+, respectively. Retrieval of fear was
tested 7 days later in context B, with 4 presentations of the CS- and the CS
+. For optogenetic
experiments using Archeorhodopsin (ArchT), PV-IRES-Cre mice were submitted on day 1 to
a pre-fear conditioning session (pre-FC) in context A during which they received yellow light
stimulations (250 ms pulses repeated at 0.9 Hz during 2 min). Fear conditioning was
performed on day 2 in context A, by pairing the CS+ with the US (1 s foot-shock, 0.6 mA, 5
CS+/US pairings; inter-trial interval: 20-180 s). On day 2 and day 3, conditioned mice were
submitted to extinction training (Post-FC and Ext. sessions) in context B during which they
received 12 presentations of the CS+. At the end of the last extinction session they received an
additional 4 presentations of the CS+ coupled to yellow light stimulations (each CS
+ pip was
paired with a 250 ms light pulse). For optogenetic experiments using Channelrhodopsin
(ChR2), PV-IRES-Cre mice were submitted on day 1 to the same fear conditioning protocol
as above. A post-conditioning test (Post-FC) was performed on day 2 in context B and
consisted of 4 presentations of the CS+ alone followed by 4 presentations of the CS
+ coupled
to blue light stimulations (each CS+ pip was paired with a 250 ms light pulse). On day 3, mice
were submitted to a second test in context B (Test) in which they received 4 CS+
presentations.
B/ Fear conditioning to complex tone paradigm
In collaborative with the Dr. Andreas Lüthi from Basel in Switzerland, we used
complex tones instead of pure tones during fear conditioning in order to engage the AC in the
fear acquisition process. For these experiments, we employed 30-s trains of frequency-
modulated sweeps as complex auditory conditioned stimuli (500 ms duration, logarithmically
modulated between 5 and 15 kHz, 50 ms rise and fall) delivered at 1 Hz at a sound pressure
level of 70 dB. The CS+ (up sweep, frequency-modulated from 5 to 15 kHz) was paired with a
periorbital shock (2 s, 2.5 mA, 15 CS+-shock pairings; inter-trial interval: 20-180 s) which
was used instead of foot shocks to minimize electrical artifacts in order to evaluate US-
evoked activity in the recorded neurons. The onset of the periorbital shock coincided with the
onset of the last sweep in the CS+. The CS
- (down sweep, frequency-modulated from 15 to 5
kHz) was presented after each CS+-footshock association, but was never reinforced (15 CS
-

- 37 -
presentations, inter-trial interval: 20-180 s). On the next day, conditioned mice were
submitted to fear retrieval in context B, during which they received 4 and 4 presentations of
the CS- and the CS
+, respectively. CS
+ and CS
- were counterbalanced across animals.
C/ Place avoidance paradigm
For the place avoidance experiment, we used an apparatus composed of two plexiglas
compartments (20 x 10 cm each) connected by an alleyway. The two compartments differed
tactilely (smooth plastic versus metal bars) and visually (grey plexiglas with red horizontal
stripes or grey plexiglas). The time spent in each compartment was automatically recorded by
an infrared beam detection system located on the bottom of the apparatus (Imetronic). On day
1, mice were allowed to freely explore the entire apparatus during a 15 min pre-exposure.
Following pre-exposure, the compartment in which the mice spent the most time was
designated as the preferred compartment. On day 2, mice were submitted to a 15 min test
session during which light pulses (250 ms repeated at 0.9 Hz) were delivered while the
animals occupied the preferred compartment, but not when they occupied the non-preferred
compartment.
III/ Surgery and recordings
Mice were anesthetized with isoflurane (induction 3%, maintenance 1.5%) in O2.
Body temperature was maintained at 37 °C with a temperature controller system (FHC). Mice
were secured in a stereotaxic frame and unilaterally implanted in the left dmPFC with a multi-
wire electrode array aimed at the following coordinates (Franklin et al., 1997): 2 mm anterior
to bregma; 0.3 mm lateral to the midline and 0.8 to 1.4 mm ventral to the cortical surface. A
subset of animals (n = 10) were also implanted in the dorsal hippocampus at the following
coordinates (Franklin et al., 1997): 2 mm posterior to bregma; 1.2 mm lateral to midline and
1.2 to 1.4 mm ventral to the cortical surface. For the collaborative work on the AC, another
subset of animals (n = 11) was implanted with a pair of insulated silver wires (170-mm
diameter) beneath the skin of each eyelid for delivery of periorbital shocks. In addition, these
mice were unilaterally implanted in the AC with a multi-wire electrode aimed at the following
coordinates (Franklin et al., 1997): 2.46 mm posterior of bregma, 4.5 mm lateral to midline,
and 0.6 mm to 0.85 mm below the cortical surface. The electrodes consisted of 16
individually insulated nichrome wires (13 μm inner diameter, impedance 30-100 KΩ;

- 38 -
Kanthal) contained in a 26 gauge stainless steel guide cannula. The wires were attached to an
18 pin connector (Omnetics). For mice that received dmPFC and dHip multi-wire implants,
two connectors were used. All implants were secured using Super-Bond cement (Sun
Medical). After surgery mice were allowed to recover for 7 days and were habituated to
handling. Analgesia was applied before, and 1 day after surgery (Metacam, Boehringer).
Electrodes were connected to a headstage (Plexon) containing sixteen unity-gain operational
amplifiers. The headstage was connected to a 16-channel preamplifier (gain 100x, bandpass
filter from 150 Hz to 9 kHz for unit activity and from 0.7 Hz to 170 Hz for field potentials,
Plexon). Spiking activity was digitized at 40 kHz and bandpass filtered from 250 Hz to 8 kHz,
and isolated by time-amplitude window discrimination and template matching using a
Multichannel Acquisition Processor system (Plexon). At the conclusion of the experiment,
recording sites were marked with electrolytic lesions before perfusion, and electrode tips
locations were reconstructed with standard histological techniques.
IV/ Single-unit analyses
Single-unit spike sorting was performed using Off-Line Spike Sorter (OFSS, Plexon)
for all behavioural sessions. Principal component (PC) scores were calculated for unsorted
waveforms and plotted in a 3D PC space; clusters containing similar valid waveforms were
manually defined. A group of waveforms were considered to be generated from a single
neuron if the waveforms formed a discrete, isolated, cluster in the PC space and did not
contain a refractory period less than 1 ms, as assessed by using auto-correlogram analyses
(Fig. 8).
Figure 8. Separation of putative pyramidal neurons and putative interneurons. a, Left: superimposed
waveforms recorded from 2 different units. Right: Spikes originating from individual units were sorted using
3D-principal component analysis. b, Corresponding auto-correlograms displaying clear refractory periods.

- 39 -
To avoid analysis of the same neuron recorded on different channels, we computed
cross-correlation histograms. If a target neuron presented a peak of activity at a time that the
reference neuron fired, only one of the two neurons was considered for further analysis. After
fear conditioning, if the same neuron was sequentially recorded during different behavioural
sessions, we considered only the first behavioural session in which it was recorded. To
separate putative INs from PNs we used an unsupervised cluster algorithm based on Ward’s
method. Briefly, the Euclidian distance was calculated between all neuron pairs based on the
3D space defined by each neuron’s average half spike width (measured from trough to peak),
the firing rate and the area under the hyperpolarization phase of the spike. An iterative
agglomerative procedure was then used to combine neurons into groups based on the matrix
of distances such that the total number of groups was reduced to give the smallest possible
increase within-group sum of square deviation. To assess the significance of cross-
correlogram analyses performed between pairs of recorded neurons, a mean firing rate with
95% confidence limits of the target neuron was calculated. Significant short-latency inhibitory
or excitatory interactions were retained if the number of action potentials of the target neuron
was inferior or superior to the 95% confidence limits, respectively. Moreover, to show that
cross-correlations were not simply occurring by chance or were due to CS presentations, we
performed two controls. First, the spike train of the neuron was shuffled 100 times and a
shuffled cross-correlogram was computed. Absence of short-latency interaction in the
shuffled cross-correlogram was indicative that the cross-correlations were not due to chance.
Second, to control that short-latency interactions were not artificially induced by stimulus
presentations, we computed a shift predictor and subtracted it from the original cross-
correlogram. Persistence of short-latency cross-correlations indicates that the neuronal
interactions were not due to CS presentations. CS or light-induced neural activity of recorded
neurons was calculated by comparing the firing rate after stimulus onset with the firing rate
recorded during the 500 ms before stimulus onset (bin size: 10 ms) using a z-score
transformation. Z-score values were calculated by subtracting the average baseline firing rate
established over the 500 ms preceding stimulus onset from individual raw values and by
dividing the difference by the baseline standard deviation. Only CS+ responsive neurons (at
least one significant positive or negative z-score bin within 100 ms following CS onset) were
considered for further analysis. For statistical analysis, z-score comparisons were performed
using the average z-score value calculated during the 150 ms after CS onset. In some analyses,
we compared PSTH established using different numbers of reference events. In these
conditions, the baseline neuronal activity displayed strong fluctuations that prevented an

- 40 -
appropriate z-score comparison. In these cases, we compared the firing activity of the neurons
normalized to the baseline firing, a measure less sensitive to fluctuation in baseline neuronal
activity. To identify the main firing patterns among INs, we used an unbiased principal
component analysis (PCA) based on the neuronal activity evoked by CS+ presentations (z-
score 500 ms before and after CS+ presentations, CS
+ presentations 1-4 in Post-FC, Extinction
and Retrieval sessions, each CS+ consisting of 27 individual sound pips; bin size: 10 ms).
Only the first PC was considered (PC1) because it explained most of the variance of our
dataset. Type1 and Type 2 INs were defined as correlated and inversely correlated with PC1
at the P < 0.001 significance level. Co-firing between recorded PNs pairs was established by
quantifying the number of non-overlapping 30 ms time windows following CS+ presentations
during which co-firing events occurred (each pip presentation, CS+ presentations 1-4, 108
pips, Post-FC, Extinction or Retrieval sessions). We then calculated a ratio of coincident
firing by dividing the number of co-firing occurrences during CS+ presentations by those
obtained during CS- presentations. This coincident firing ratio was normalized to the pre-CS
period (500 ms pre CS) using a z-score transformation. To control that the changes in
coincident firing between CS+ and CS
- conditions were not due to an increase in PNs firing
rate during CS+ presentations, the same analysis was performed but this time, the number of
co-firing events in each 30 ms time window was normalized by the total number of spikes of
the two neurons in this particular time window. Statistical analyses were performed using
paired Student’s t-tests post hoc comparisons at the P < 0.05 level of significance unless
indicated otherwise. Results are presented as mean ± s.e.m.
V/ Field potential analyses
Local field potential (LFP) signals were filtered in the theta range (8-12Hz) using a
zero-phase-delay filter implemented in the NeuroExplorer software (NeuroExplorer) and theta
peaks and troughs were identified. To analyze theta phase resetting evoked by CS or light
presentations, we first created a pure phase signal with a standardized amplitude by assigning
to theta peaks and troughs a zero and 180° value, respectively. The signal was then linearly
interpolated between these two points and cosined. Theta phase resetting precision was
quantified by calculating the variance of the time of first theta peak occurrence following
stimulus onset over all trials (CS or light). In this way, greater resetting precision is reflected
in smaller variance. To evaluate theta phase locking of individual neurons, we computed the
instantaneous theta phase of the 8-12 Hz filtered LFP using the Hilbert transform. For a given

- 41 -
neuron, each spike was assigned its corresponding LFP theta phase value from the LFP signal
recorded on the same wire. Theta phase locking was calculated using Rayleigh’s test for
circular uniformity and statistical significance was assessed using Rayleigh’s test Z value.
Logarithmic transformation was applied to tighten the Z distribution. For significantly phased-
locked PNs, we quantified phase locking strength using the Mean Resultant Length (MRL),
i.e. the length of the mean vector, and the concentration parameter of the Von Mises
distribution fit of the neuron’s phase histogram (κ). These two parameters estimate the
circular concentration of spikes. Low or high values for the two parameters are respectively
indicative of a spread or a concentrated circular distribution of spikes around the preferred
phase. All these analyses were performed on entire recordings for INs and on the 500 ms
recordings following stimulus onset for PNs (CS or light presentations, CS presentations 1-4,
Post-FC, extinction or retrieval sessions). Importantly, both MRL and κ estimate are highly
dependent on the sample size. We adopted two conditions to limit this bias in our analysis.
First, we computed MRL and κ only for significantly phase-locked neurons displaying at least
50 spikes during the 500 ms period following CS presentations. The minimum spike number
was reduced to 15 for optogenetic experiments in which a limited number of neurons were
recorded. Second, we controlled that the number of spikes did not significantly differ between
the conditions that we compared. Therefore, at least the relative comparisons of theta
modulation strength between conditions should not be affected by sample sizes (number of
spikes). To evaluate the dynamics changes of PNs neuronal synchronization with local theta
during phase resetting, we calculated the mean preferred phase of individual PNs for the theta
cycle before and the three theta cycles following CS or light stimulations. In these analyses, a
phase of 0° corresponds to the peak of prefrontal theta oscillations. For the CS+ condition, we
extracted the time periods of the first three theta cycles following CS+ presentations for each
trial (each CS+ pip), and calculated the phase of individual PNs during CS
+ pip presentations
relative to these time periods. Because prefrontal theta phase resetting occurs only during high
fear states (i.e. CS+ conditions), analyses during CS
- conditions were performed for each mice
by using the onset of the first theta cycle that was defined by the mean latency of the first
theta peak identified in the CS+ condition. For a given neuron, the mean preferred phase was
computed over 108 CS pip trials by using the mean circular direction. To quantify the
difference between two conditions (for instance between CS+ and CS
- conditions), we
calculated the deviation from circular uniformity of the preferred phases of PNs for each theta
cycle using the Rayleigh’s test. To control the underlying assumption of Von Mises
distribution in the Rayleigh test, we also performed the analysis using the non-parametric

- 42 -
Hodjes-Ajne test, which makes no such assumption. Because both tests gave similar results,
we only considered the statistical values obtained with the Rayleigh’s test.
VI/ Virus injections and optogenetics
For optical control of PV INs, conditional AAV encoding ArchT (AAV-FLEX-
ArchT-GFP, serotype 9,Vector Core, University of North Carolina) or ChR2 (AAV-EF1a-
DIO-hChR2(H134R)-EYFP, serotype 5, Vector Core, University of North Carolina) were
bilaterally injected into the dmPFC of PV-IRES-CRE mice from glass pipettes (tip diameter
10-20 µm) connected to a picospritzer (parker Hannifin Corporation; approximately 0.4 µL
per hemisphere) at the following coordinates (Franklin et al., 1997): 2 mm anterior to bregma;
0.4 mm lateral to midline and 0.9 to 1.2 mm ventral to the cortical surface. 1-2 weeks after the
injection mice were implanted bilaterally with optic fibres (diameter: 200 µm; numerical
aperture: 0.37; flat tip; Doric Lenses) at the same coordinates. Control experiments were
performed using an AAV containing the DNA construct for only GFP (AAV-FLEX-GFP,
Vector Core, University of North Carolina). All implants were secured using Super-Bond
cement (Sun Medical). For experiments using optogenetic stimulation coupled to single unit
recordings, one of the two optic fibres was combined to the array of 16 or 32 individually
insulated nichrome wires. Single unit recordings during the manipulation of PV INs were
performed as described in the “Surgery and recordings” section. Behavioural and recording
experiments were performed 3-5 weeks post-injection. The light (approximately 14mW per
implanted fibre) was bilaterally conducted from the laser (OptoDuet 473/593 nm, Ikecool) to
the mice via two fibre-optic patch cords (diameter: 200 µm, Doric Lenses), connected to a
rotary joint (1x2 fibre-optic rotary joint, Doric Lenses) that allowed mice to freely move in
the behavioural apparatus. For optogenetic manipulation of PV INs during behaviour, we used
250 ms or 500 ms light pulses delivered at 0.9 Hz. After behavioural and recording
experiments, mice were perfused and histological analysis was performed.
VII/ Anatomical analysis
Mice were euthanized with isoflurane and perfused through the left ventricle with 4%
w/v paraformaldehyde (PFA) in 0.1 M PBS. Brains were dissected out and post-fixed for 4 h
at 4˚C in the same solution. For localization of recording and electrical stimulation sites
(electrolytic lesions), mice were directly transcardially perfused with PFA 4%. 60μm-thick

- 43 -
sections were cut, mounted on gelatin-coated microscope slides, and dried. Sections were
stained with toluidine blue, dehydrated and mounted. Electrolytic lesions were identified with
conventional transmission light microscopy. Only recordings with confirmed lesions in
cingulate and prelimbic areas of dmPFC were included in our analyses. For verification of
muscimol (MUS) injections in medial septum (MS) and viral injections in dmPFC, serial 80
μm-thick slices containing the MS and dmPFC (respectively) were imaged using an
epifluorescence system (Leica DM 5000) fitted with a 10x dry objective. The location and the
extent of the injections/infections were visually controlled. All included MUS injections were
targeted and limited to the MS. Similarly, only infections accurately targeting the dmPFC
were considered for behavioural and electrophysiological analyses. The specificity and
sensitivity of PV INs infection was assessed as follows (Fig. 9). Immunofluorescence was
performed to visualize PV and to enhance fluorescence induced by viral GFP expression, and
colocalization of PV and ArchT was assessed. We used two mice that were not involved in
electrophysiological or optogenetics experiments to avoid confounding effects of brain
damage. Mice were first perfused with 10 mL of room-temperature PBS (pH 7.4), and fixed
with 75 mL of 4% PFA at 4˚C (TAAB, pH 7.3; 25 mL at ~10 mL/min and 50 mL at ~6
mL/min). After short post-fixation, sixty micrometer-thick coronal sections were cut and kept
in PB 0.1 until use. All reagents were diluted in PB 0.1 M containing triton X-100 0.3% v/v.
Free-floating sections were blocked in 20 % normal goat serum (NGS, Vector laboratories)
for 1 h at room temperature and incubated at 4˚C for 2 days in 1:2,000 anti-PV antibody made
in guinea pig (Synaptic Systems 195-004) with 2% NGS. Sections were washed and
incubated at 4˚C overnight in 1:500 Alexa 633-conjugated goat anti-guinea pig and 1:250
Alexa 488-conjugated anti-GFP rabbit antibody (Invitrogen) with 2% NGS. After extensive
washes, sections were mounted in Vectashield (Vector Laboratories). Three confocal images
stacks (1 μm steps, slice thickness 1 Airy unit) were acquired (Leica DM2500 TCS SPE 40x
oil immersion 1.3 NA objective) for each animal, from different sections of dmPFC, close to
the virus injection sites. Sequential acquisition was used to prevent cross-talk between Alexa
488 and Alexa 633 signals. Immunoreactivity of cell bodies for PV and GFP was assessed
independently for each stack (n = 200 PV+ cells). Cells were then marked as PV
+/GFP
-,
PV+/GFP
+ or PV
-/GFP
+ with Image J “Cell counter” plug-in. Specificity was calculated as
(number of PV+/GFP
+ neurons / total number of GFP
+ cells ). Sensitivity was calculated as
(number of PV+ / GFP
+ cells / total number of PV neurons). PV-expressing cells in PFC
comprise perisomatic-innervating basket cells as well as axon-initial segment innervating axo-
axonic cells (Massi et al., 2012). We also examined the dendritic arborizations of GFP-
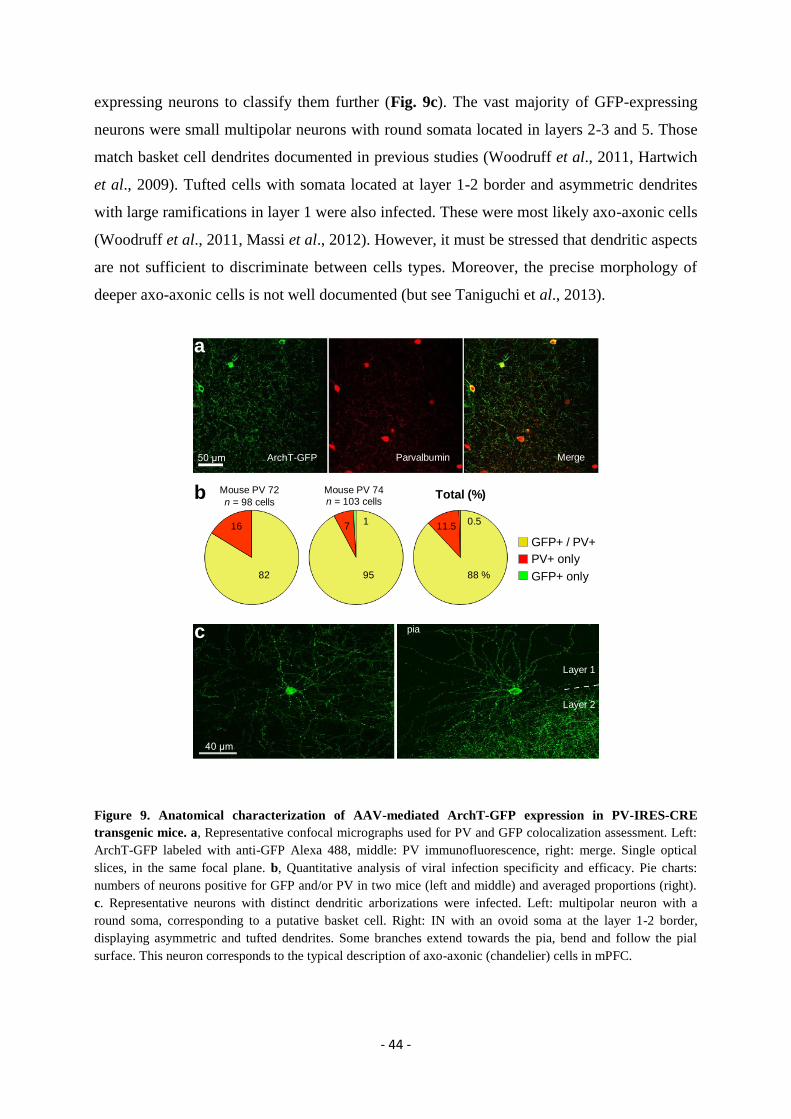
- 44 -
expressing neurons to classify them further (Fig. 9c). The vast majority of GFP-expressing
neurons were small multipolar neurons with round somata located in layers 2-3 and 5. Those
match basket cell dendrites documented in previous studies (Woodruff et al., 2011, Hartwich
et al., 2009). Tufted cells with somata located at layer 1-2 border and asymmetric dendrites
with large ramifications in layer 1 were also infected. These were most likely axo-axonic cells
(Woodruff et al., 2011, Massi et al., 2012). However, it must be stressed that dendritic aspects
are not sufficient to discriminate between cells types. Moreover, the precise morphology of
deeper axo-axonic cells is not well documented (but see Taniguchi et al., 2013).
Figure 9. Anatomical characterization of AAV-mediated ArchT-GFP expression in PV-IRES-CRE
transgenic mice. a, Representative confocal micrographs used for PV and GFP colocalization assessment. Left:
ArchT-GFP labeled with anti-GFP Alexa 488, middle: PV immunofluorescence, right: merge. Single optical
slices, in the same focal plane. b, Quantitative analysis of viral infection specificity and efficacy. Pie charts:
numbers of neurons positive for GFP and/or PV in two mice (left and middle) and averaged proportions (right).
c. Representative neurons with distinct dendritic arborizations were infected. Left: multipolar neuron with a
round soma, corresponding to a putative basket cell. Right: IN with an ovoid soma at the layer 1-2 border,
displaying asymmetric and tufted dendrites. Some branches extend towards the pia, bend and follow the pial
surface. This neuron corresponds to the typical description of axo-axonic (chandelier) cells in mPFC.
b Mouse PV 72
n = 98 cells
Mouse PV 74n = 103 cells
Total (%)
82 95 88 %
16 7 11.50.51
a
50 µm
GFP+ / PV+
PV+ only
GFP+ only
ArchT-GFP Parvalbumin Merge
c
40 µm
pia
Layer 1
Layer 2

- 45 -
VIII/ Muscimol inactivation
Mice were unilaterally implanted with stainless steel guide cannula (26 gauge; Plastics
One) aimed at the MS using an angle of 10° and recording electrodes were implanted in the
dmPFC and the dHip as described in the “surgery and recording” methodological section. To
target the MS, we used the following coordinates (Franklin et al., 1997): 1 mm anterior to
bregma; 0.7 mm lateral to midline and 3 to 3.3 mm ventral to the cortical surface with an
angle of 10° in the coronal plane. The cannula was secured using Super-Bond cement (Sun
Medical). On the injection day, muscimol (Muscimol-Bodipy-TMR-X conjugate, Invitrogen;
0.8 mM in PBS 0.1 M) was infused at a rate of 0.2 µL/min during 2 min (total volume of 0.4
µL). On the injection day, muscimol was infused 15 min before the behavioural test. After the
end of the experiment, muscimol was again infused with the same parameters to control for
the drug diffusion within the MS and mice were perfused. Brains were collected for
histological analyses as described above.
IX/ Extracellular stimulation
At the end of the behavioural procedure, mice were anesthetized with urethane (1.4 g
kg-1
) and concentric stimulating electrodes (FHC) were lowered in the BLA (1.7 mm posterior
to bregma; 3.1 mm lateral to midline and 3.8 to 4.6 mm ventral to the cortical surface) and the
PAG (4.8 mm posterior to bregma; 0.6 mm lateral to midline and 1 to 2 mm ventral to the
cortical surface) (Franklin et al., 1997). During the experiments, the stimulation electrodes
were advanced in steps of 5 µm by a motorized micromanipulator (FHC) and dmPFC-evoked
responses were recorded. Stimulation-induced and spontaneous spikes were sorted as
described in the section “Surgery and recordings”. Stimulation-induced spike waveforms were
quantitatively compared to the waveforms of units previously identified in the awake animal
and recorded on the same wire using correlation analysis. This ensured that the same neurons
were recorded across behaviour and anesthesia. To be classified as antidromic, evoked-
responses had to meet at least two out of three criteria: (1) stable latency (< 0.3 ms jitter), (2)
collision with spontaneously occurring spikes, and (3) ability to follow high-frequency
stimulation (250 Hz). At the end of the experiments, stimulating sites were marked with
electrolytic lesions before perfusion, and electrode locations were verified (see above).

- 46 -
RESULTS
I/ Results Part 1
A/ Introduction
The aim of my thesis was to identify the functional role of cortical PV INs in the
acquisition and expression of associative fear memory. During the first few months of my
PhD, in collaboration with the group of Dr. Andreas Lüthi, Basel, Switzerland, we evaluated
the role of the AC PV INs in the acquisition of auditory fear conditioning to complex tones. In
a first step, our collaborators demonstrated using reversible and targeted inactivation, that the
AC is necessary for auditory fear conditioning (Letzkus et al., 2011). They next observed
using calcium imaging in anesthetized animals that the vast majority of AC layer 1 INs (L1)
were strongly activated during footshock following tone presentations. These L1 INs receive
glutamatergic inputs from other cortical areas as well as cholinergic inputs from the basal
forebrain, the main source of acetylcholine in the brain. Using cholinergic antagonists in
anesthetized animals they were able to block the footshock-induced increase in neuronal
activity of cortical L1 INs. Moreover nicotinic puff applications or cholinergic inputs
stimulations strongly increase L1 INs activity. These data strongly suggest that neuronal
activity in cholinergic inputs is necessary and sufficient to drive L1 cortical INs during
auditory fear conditioning. In a second step, they evaluated how the excitatory neuronal
responses in L1 INs during footshock presentations modify local microcircuits. Notably, they
demonstrated that L1 INs activation during footshock presentations induces a strong
inhibition of the vast majority of PV INs in cortical layers 2/3 (L2/3) which disinhibit L2/3
PNs. Finally, using optogenetic approaches in behaving animals during auditory fear
conditioning they were able to show that optical stimulation of L2/3 PV INs during footshock
presentations blocks the acquisition of auditory fear conditioning.
My contribution to this work consisted in evaluating if the disinhibitory mechanism
mediated by PV INs was also observed in awake, freely moving animals. By using single unit
recordings in behaving mice, we showed that during acquisition of auditory fear conditioning,
presentation of the US was associated with a strong inhibition of putative PV INs leading to
the disinhibition of putative PNs located in AC superficial layers. My contribution to this
collaborative study is fully described the following section.
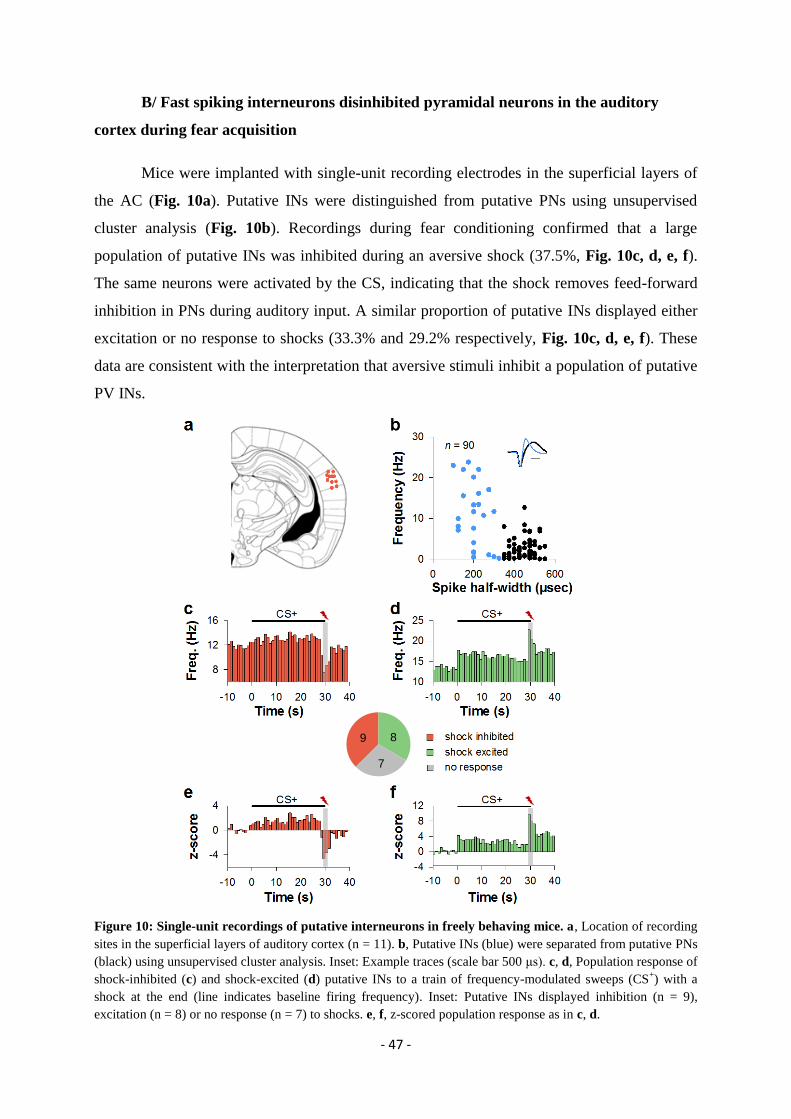
- 47 -
B/ Fast spiking interneurons disinhibited pyramidal neurons in the auditory
cortex during fear acquisition
Mice were implanted with single-unit recording electrodes in the superficial layers of
the AC (Fig. 10a). Putative INs were distinguished from putative PNs using unsupervised
cluster analysis (Fig. 10b). Recordings during fear conditioning confirmed that a large
population of putative INs was inhibited during an aversive shock (37.5%, Fig. 10c, d, e, f).
The same neurons were activated by the CS, indicating that the shock removes feed-forward
inhibition in PNs during auditory input. A similar proportion of putative INs displayed either
excitation or no response to shocks (33.3% and 29.2% respectively, Fig. 10c, d, e, f). These
data are consistent with the interpretation that aversive stimuli inhibit a population of putative
PV INs.
Figure 10: Single-unit recordings of putative interneurons in freely behaving mice. a, Location of recording
sites in the superficial layers of auditory cortex (n = 11). b, Putative INs (blue) were separated from putative PNs
(black) using unsupervised cluster analysis. Inset: Example traces (scale bar 500 μs). c, d, Population response of
shock-inhibited (c) and shock-excited (d) putative INs to a train of frequency-modulated sweeps (CS+) with a
shock at the end (line indicates baseline firing frequency). Inset: Putative INs displayed inhibition (n = 9),
excitation (n = 8) or no response (n = 7) to shocks. e, f, z-scored population response as in c, d.
9
7
8

- 48 -
PV INs provide strong, perisomatic inhibition to local PNs (Markram et al 2004,
Freund & Katona 2007, Kawaguchi & Kubota 1997, Kruglikov & Rudy 2008). We test
directly whether disinhibition was the main effect of aversive stimulation in AC L2/3 PNs.
Our collaborators showed that footshocks elicited the disinhibition of L2/3 PNs, in head-
fixed, anaesthetized mice. We confirmed this result by showing in freely moving mice, that
the coincidence of tone and eyelid shock excited putative PNs much more than the tone alone
(Fig. 11a). This effect was highly supralinear in both experiments because eyelid shocks
alone elicited almost no activity in these neurons (Fig. 11b). These data indicate that L2/3
PNs are disinhibited by aversive stimuli. In summary, these data are consistent with the
interpretation that aversive stimuli serve to remove both spontaneous and feed-forward
inhibition provided by PV INs to surrounding PNs during fear acquisition, in behaving mice.
Figure 11. Aversive shocks disinhibit L2/3 pyramidal neurons. a, Mean z-score transformation of CS-
and
CS+/US-evoked firing responses of putative PNs (n = 24) with significant response to the CS
+ in freely-behaving
mice during fear conditioning. Note strong activation by coincidence of sweep and shock. b, Comparison of
average z-scores in putative PNs during single-unit recordings in freely behaving mice in response to the shock
alone (gray, n = 16), the end of the CS- without shock and the end of the CS
+ with shock (n = 24). Note strong
boosting of the sweep response by the shock (p < 0.05, two-tailed paired Student’s t-test), exceeding the
arithmetic sum of the responses to shock and the end of the CS- (black). 6 of the neurons recorded during sweep
and compound were also recorded during shock alone. Values presented as mean ± s.e.m.
C/ Article 1: “A disinhibitory microcircuit for associative fear learning in the
auditory cortex”
Johannes J. Letzkus, Steffen B. E.Wolff, Elisabeth M. M. Meyer, Philip Tovot, Julien
Courtin, Cyril Herry & Andreas Lüthi.
Nature, 480, 331-335 (26 October 2011).

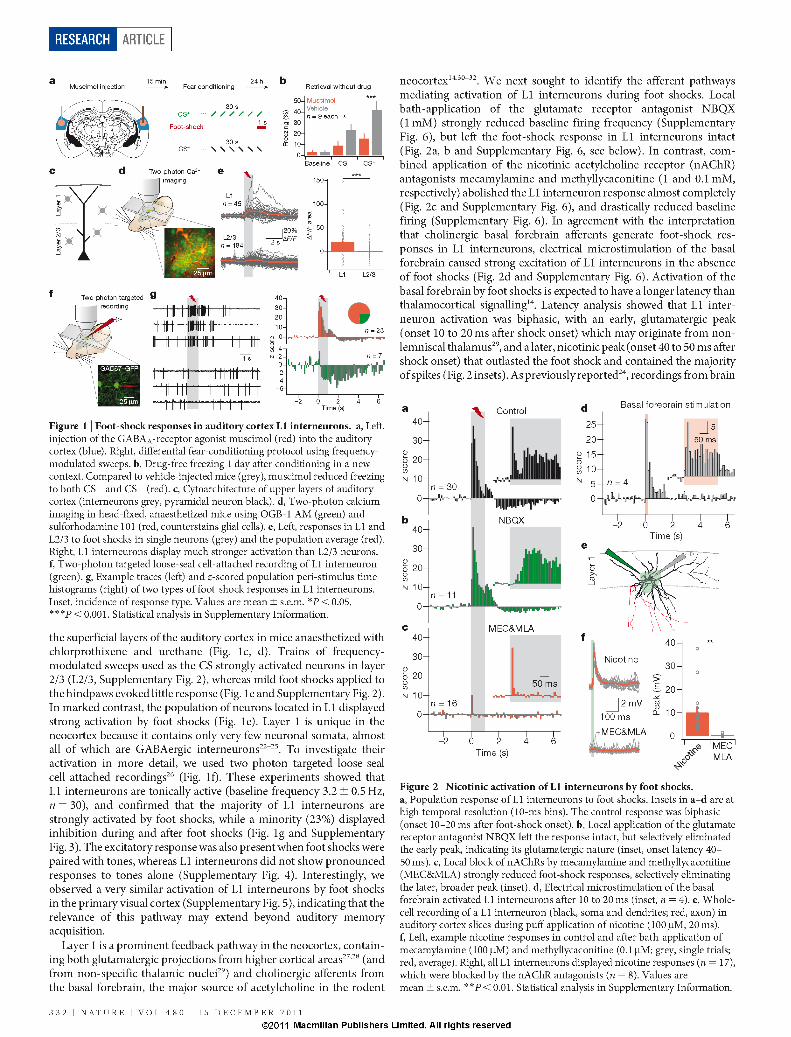
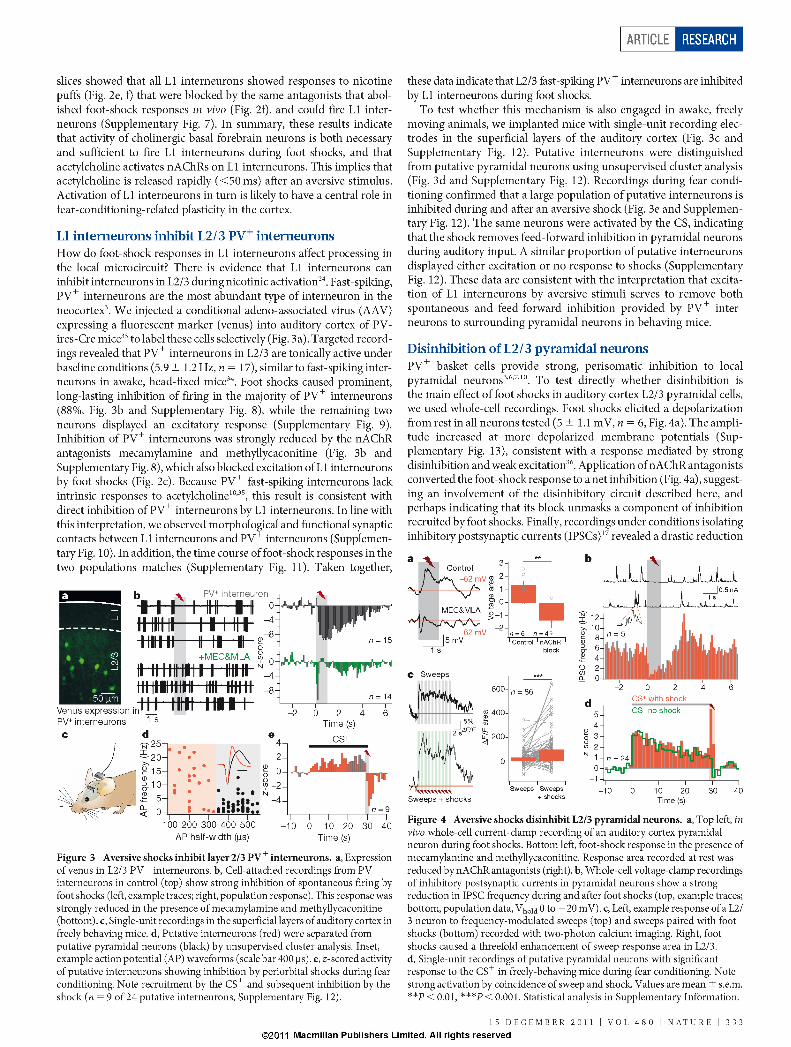





w w w. n a t u r e . c o m / n a t u r e | 1
SuPPLementarY InFormatIondoi:10.1038/nature10674
A disinhibitory microcircuit for associative fear learning in auditory cortex Johannes J. Letzkus1,3*, Steffen B.E. Wolff1*, Elisabeth M.M. Meyer1, Philip Tovote1, Julien Courtin2, Cyril Herry2, Andreas Lüthi1,3 1Friedrich Miescher Institute for Biomedical Research, Maulbeerstrasse 66, CH-4058 Basel, Switzerland. 2INSERM U862, Neurocentre Magendie, 146 Rue Léo-Saignat, 33077 Bordeaux, France. *These authors contributed equally to this work. 3Authors for correspondence: [email protected] and [email protected]
Statistical analysis Figure 1b: Effects of muscimol injection on acquisition of fear memory. Comparison of freezing during memory retrieval (24h after conditioning, drug-free) between a group of mice injected with muscimol during training (n=9) and a vehicle-injected control group (n=9). Two-way ANOVA indicated significant main effects between treatment groups: F(1,53)=12.536, p<0.001; as well as between test phases (baseline, CS-, CS+): F(2,53)=14.451, p<0.001. Significant interaction was detected between test phase and treatment: F(2,53)=3.823, p<0.05. Vehicle-injected control mice exhibited significantly higher freezing behaviour during CS+ presentation compared to baseline (p<0.001) and during CS- presentation compared to baseline (p<0.05), whereas there was no significant increase in freezing during CS- or CS+ presentation in muscimol treated mice. Post-hoc pair-wise Tukey’s tests revealed significantly reduced freezing during CS+ presentation (p<0.001) and CS- presentation (p<0.05) in muscimol injected animals compared to vehicle. Figure 1e right: Comparison of ∆F/F area in response to a foot-shock in L1 interneurons (n=45) and L2/3 neurons (n=184). Two-tailed, unpaired Student’s t-test revealed significant difference (p<0.001). Figure 2f right: Comparison of peak response to a nicotine puff in L1 interneurons in control (n=17) and after bath application of mecamylamine (100 µM) and methyllycaconitine (0.1 µM, n=8). Two-tailed, unpaired Student’s t-test revealed significant difference (p<0.01). Figure 4a right: Comparison of voltage response area elicited by a foot-shock in L2/3 pyramidal neurons in control (n=6) and in the presence of mecamylamine (1 mM) and methyllycaconitine (0.1 mM, n=4). Two-tailed, unpaired Student’s t-test revealed significant difference (p<0.01). Figure 4c right: Comparison of the response (∆F/F area) of L2/3 neurons (n=86) to a train of frequency-modulated sweeps, and to an identical train in
which each sweep was paired with a foot-shock. Two-tailed, paired Student’s t-test revealed significant difference (p<0.001). Figure 5b: Effects of mecamylamine and methyllycaconitine injection on acquisition of fear memory. Comparison of freezing during memory retrieval (24h after conditioning, drug-free) between a group of mice injected with drug during training (n=11) and a vehicle-injected control group (n=14). Two-way ANOVA indicated significant main effects between treatment groups: F(1,74)=20.060, p<0.001; as well as between test phases (baseline, CS-, CS+): F(2,74)=32.591, p<0.001. Significant interaction was detected between test phase and treatment: F(2,74)=3.497, p<0.05. Vehicle-injected control mice exhibited significantly higher freezing behaviour during CS+ presentation compared to baseline (p<0.001) and during CS- presentation compared to baseline (p<0.001). Drug-injected mice exhibited significantly higher freezing behaviour during CS+ presentation compared to baseline (p<0.01), but not during CS- presentation compared to baseline (p>0.05). Post-hoc pair-wise Tukey’s tests revealed significantly reduced freezing during CS+ presentation (p<0.001) and CS- presentation (p<0.01) in drug injected animals compared to vehicle. Figure 5f: Effects of optogenetic stimulation of PV+-interneurons during and after foot-shocks on acquisition of fear memory. Comparison of freezing during memory retrieval (24h after conditioning, no optogenetic manipulation) in three groups: optogenetics (n=8), reconditioning (n=8), sham (n=8). Two-way ANOVA indicated significant main effects between groups: F(2,71)=12.348, p<0.001; as well as between test phases (baseline, CS-, CS+): F(2,71)=73.043, p<0.001. Significant interaction was detected between test phase and treatment: F(4,71)=4.409, p<0.05. Post-hoc pairwise Tukey’s tests revealed significantly reduced freezing during CS+ presentation in the optogenetics group compared to both sham and reconditioning (p<0.001), whereas mice in sham and reconditioning groups exhibited similarly high CS+-induced freezing (p>0.05). During CS- presentation there were no significant differences between optogenetics, sham and reconditioning groups.

SUPPLEMENTARY INFORMATION
2 | w w w. n a t u r e . c o m / n a t u r e
RESEARCH
which each sweep was paired with a foot-shock. Two-tailed, paired Student’s t-test revealed significant difference (p<0.001). Figure 5b: Effects of mecamylamine and methyllycaconitine injection on acquisition of fear memory. Comparison of freezing during memory retrieval (24h after conditioning, drug-free) between a group of mice injected with drug during training (n=11) and a vehicle-injected control group (n=14). Two-way ANOVA indicated significant main effects between treatment groups: F(1,74)=20.060, p<0.001; as well as between test phases (baseline, CS-, CS+): F(2,74)=32.591, p<0.001. Significant interaction was detected between test phase and treatment: F(2,74)=3.497, p<0.05. Vehicle-injected control mice exhibited significantly higher freezing behaviour during CS+ presentation compared to baseline (p<0.001) and during CS- presentation compared to baseline (p<0.001). Drug-injected mice exhibited significantly higher freezing behaviour during CS+ presentation compared to baseline (p<0.01), but not during CS- presentation compared to baseline (p>0.05). Post-hoc pair-wise Tukey’s tests revealed significantly reduced freezing during CS+ presentation (p<0.001) and CS- presentation (p<0.01) in drug injected animals compared to vehicle. Figure 5f: Effects of optogenetic stimulation of PV+-interneurons during and after foot-shocks on acquisition of fear memory. Comparison of freezing during memory retrieval (24h after conditioning, no optogenetic manipulation) in three groups: optogenetics (n=8), reconditioning (n=8), sham (n=8). Two-way ANOVA indicated significant main effects between groups: F(2,71)=12.348, p<0.001; as well as between test phases (baseline, CS-, CS+): F(2,71)=73.043, p<0.001. Significant interaction was detected between test phase and treatment: F(4,71)=4.409, p<0.05. Post-hoc pairwise Tukey’s tests revealed significantly reduced freezing during CS+ presentation in the optogenetics group compared to both sham and reconditioning (p<0.001), whereas mice in sham and reconditioning groups exhibited similarly high CS+-induced freezing (p>0.05). During CS- presentation there were no significant differences between optogenetics, sham and reconditioning groups.
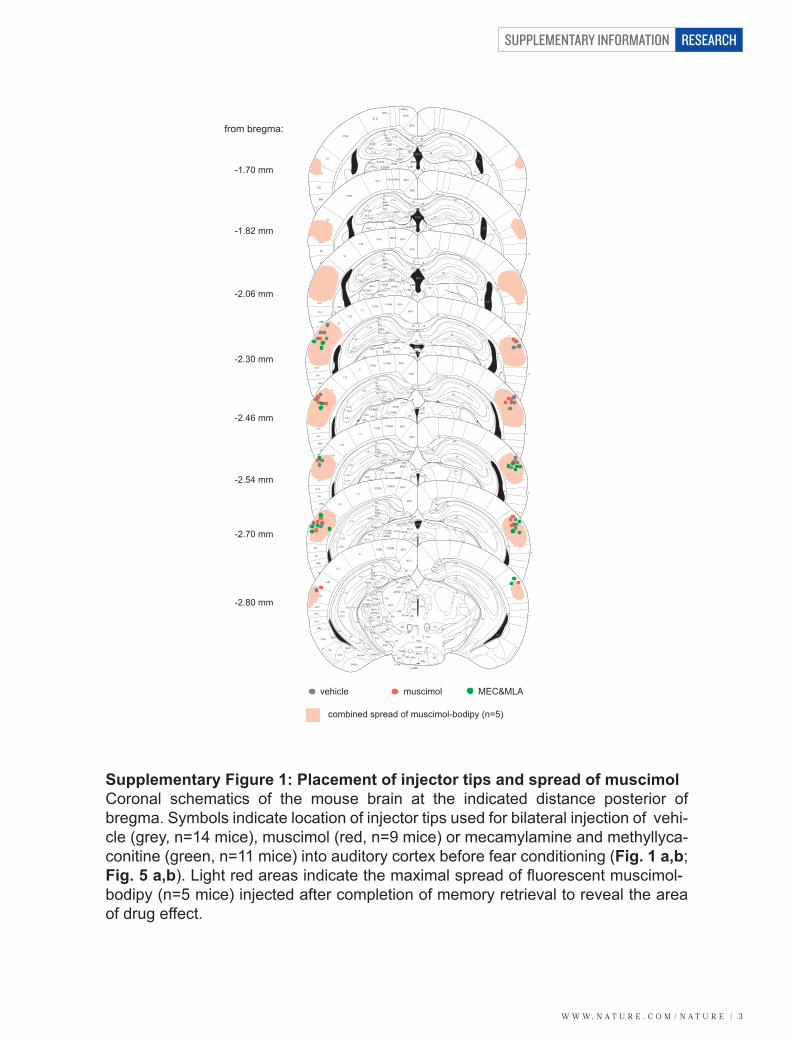
w w w. n a t u r e . c o m / n a t u r e | 3
SUPPLEMENTARY INFORMATION RESEARCH
from bregma:
-2.06 mm
-1.82 mm
-2.30 mm
-2.46 mm
-2.54 mm
-2.70 mm
-2.80 mm
S2
Ect CPuRt
LGPPRh
VLG
DEn LaVLCeL
CeC
Pir
BSTIA
BMPI
VEn
BLV
VPL VPMOPC
PCMDM
VMZID SubZIV
MCLH DM
VRe
SubI
STh
MePDPeF
LHVMHDM
VMHCVMHVL
ACo
MePV
PLCo
ME
ArcLArcD
fsox
optPe
Xi
Re
Rh
CM
mt
ns
ml
MDC
MDLPV
IMD
CLLHb
MHb
fr
VL
Po
DLG LDVL
LDDMLPMR
PoDG
DGSLu
CA2
CA3
Or
CA1
RSG
PyRadLMolMol
GrDG
RSA
S1BF
dhc
cc
cg
dfalv
ec
eml
fi
st
ic
st
I
rf
AStr
BMA
D3V
3V
FC
BLA
LV
hf
LaDL
S1Tr
AuV
MPtALPtA
IG
vehicle muscimol MEC&MLA
combined spread of muscimol-bodipy (n=5)
RSA
RSG
S1BF
CA2
Or
CA1RadPy
LMolMol
GrDG FC
DGCA3
DLGLPLR
LPMR
CLVLG
VPLVPM
OPC
PC
MDL PV
MDC
MDM
IMD
CM
SLu
PoDG
LHbM
LHbL
MHb
Po
S2
CPu
st
cc
dhc
df
cg
hbc
fr
Ect
PRh AStr
Rt
DEn
LaVM
CeL
CeCLaVL
Pir
VEnBLV
BMP
BSTIA
PLCo PMCo
I
ME
VMHVLVMHC
VMHDM
DMV
DMDDMC
MCLH
STh
MePD
MePV
LH
ZIV
VMSub
Rh
Re
mt
PH
ZIDVRe
Pe
PeFTe
MTu ArcDArcL
f
ns
ml
cpopt
Isox
st
eml st
ic
alv
ec
rf
fi
LV
hf
BLA
3V
D3V
LaDL
S1Tr
AuV
TeA
LPtAMPtA
IG
LPtA RSA
RSG
CA1
CA2
OrPyRadLMolMol
GrDG
PoDG
FC
AuDCA3
VLGMC CL LHb
MHb
DG
LPMRLPLRDLG
AuV
Ect
CPu
PRh LaDL AStr
DEn LaVMLaVL
BLA SThZIV ZID
Rt VPL VPM
Po
VLGPC IMA
PC
PFPV
OPC MD
IMD
PoMn
CM
Gus
SPFVM
PirVEn
BLP
BMP
BLV
PLCo
AHiAL MePV
BSTIA
MePD
PMCo
LH PeF
TeMTu
VMH
ArcL ME
ArcD
DMDMC
PH
f
nsmt
3V
scp
ml
sox
cp optCeL
cg
cc df
dhc
alv
hbc
freml
fi
ic st
ec
rf
D3V
LV
S1
hf
PSTh
SLu
A12
MPtA
V2L
TeA
IG
RSA
RSG
Or
S1
AuD
Au1
AuV
Ect
CPu
VLGPCVLGMC
CA3SLu
CA2
MolLMol
Py
CA1Rad
GrDG
PoDG DG
LPLRDLG
IGL
VPLPo
LPMRAPTD
SCOPrC
hbc
FC
cc df
dhc
cg
alv
emlPVPPF
SubG
PRh
LEntDEn
La
PirVEn
BLP STh
PSTh
Gem
ZIV ZID
VPM
GusSPF
fr
ml
opt
sox
cpnsmtPH
BMP
PLCoPMCo
DG LH
VTM
TePMV
ArcMP
ArcLPMe
LaDL
LaVM
SMTDTM
f
MePD
rf
ec
fi
D3V
3V
3V
LV
pv
pc
FF PR
hf
IMA
scp
AHiAL
TeA
V2L
V1V2ML
V2MM
IG
RSA
RSG
Or
Or
AuD
Au1
AuV
Ect VLGPCVLGMC
CA3
SLuCA2
MolLMol
Py
CA1
Rad
GrDG
PoDG DG
LPLRDLG
IGLIMA
LPMC
PrC
FC
ccdhc
cg
alv
PF
SubGPRh
LEnt DEn
Pir VEn BLP
Gem
ZIV
ZID
VPM
fr
mlopt
cpns
PH
BMPAHiAL AHiPM
PLCoPMCo
DG
VTMPMV
ArcMPArcLP
LaDL
LaVM
SMTDTM
f
rf
ec
fi
3V
LV
V2MMV2ML
V2L
S
SPFPCRI
PRF
SNR
PMD
SuMM
mt
D3V
pc
APir
Py
CA3
str
OPT
hf
PSTh
LH
scp
LPMR
APTD
PoTeA
V1
IG
mt
CA3
Or
VLGPCVLGMC
CA3
SLu
CA2
MolLMol
Py
CA1Rad
GrDG
PoDGDG
LPLR
DLG
IGLIMA
LPMR
APTD
FC
PFVPM
S
D3V
pc
OPT
RSG
RSAV2MMV2ML
V2L
AuD
Au1
AuV
Ect
PRh
LEnt
scc
cg
ec
alv
str
fiopt
cp
ns
ml
mtg
f
dhc
PAG
SCO
alvOr
DEn
PirBLP
AHiPMAPir
BMP
PLCo PMV
PMD
alv
ArcLP
ArcMP
SuMMSMT
SNR
SNC
ZID
ZIV
PR scp
fr
Eth
ScPo
PPT
RIF
PH
SPFPCSubG
DTMDG
LH
LT
bsc
Py
Py CA3
VTM
LV
rf
LPMC
3V
RPF
PSTh
hf
PMCo
AHiAL
TeA
V1
Or
VLGPC
VLGMC
CA3
SLu
CA2
MolLMol
Py
CA1
Rad
GrDG
PoDGDG
LPLRDLG
IGL
IMA LPMRAPTD
FCS
D3V
D3V
pc
OPT
RSG
RSAV2MMV2ML
V1
V2L
AuD
Au1
AuV
Ect
PRh
LEnt
cg
ec
alv
alv
str
opt
cp
ns
ml
mtg
dhc
SCO
Or
Or
DEn
PirBLP
APirBMP
PMCo PMD
ArcMP
SuMM
SNR
ZIDZIV
PR scpfr
Eth
REth
PPT
RIFPH
SPFPCSubG
DG
LT
bsc
Py
Py
VTM
SLu
LM ML MMMMn
MRe
SuML
sumxLH
PCom
OT
APTV
MPT
Po
PP
LPMC
CA1
PAG
SNC
SNC
fmj
CA3LV
RPF
MCPC
hf
pm
rf
AHiPM
f
TeA
pm
Or
VLGPCVLGMC
CA3
SLu
CA2
MolLMol
Py
CA1Rad
GrDG
hf
PoDG
DG
DLG
IGL
IMA
APTD
FCS
D3V
pc
OPT
RSG
RSAV2MMV2ML
V1
V2L
AuD
Au1
AuV
Ect
PRh
LEnt
cg
ec
alv
alvopt
cp
ns
ml
mtg
dhc
SCO
Or
Or
DEn
PirBLP
APir
PMCoArcMP
SuMMSNR
ZIDZIV
PR scpfr
MCPC
Eth
REth
PPT
RI
SPFPC
SubG
GrDG
LT
bsc
Py
VTM
AHiPM
SLu
LM ML MMMMn
MRe
SuML
sumx
PCom
OT
APTV
MPT
PoT
PP
LPMCLPLC
PAG
SNC
SNL
fmj
PLi
DkPILIntG
MGV
SG
MGM
MGD
EW
LH
f
CA3
rf
CA2
SComRPF
LV
TeA
-1.70 mm
Supplementary Figure 1: Placement of injector tips and spread of muscimolCoronal schematics of the mouse brain at the indicated distance posterior of bregma. Symbols indicate location of injector tips used for bilateral injection of vehi-cle (grey, n=14 mice), muscimol (red, n=9 mice) or mecamylamine and methyllyca-conitine (green, n=11 mice) into auditory cortex before fear conditioning (Fig. 1 a,b; Fig. 5 a,b). Light red areas indicate the maximal spread of fluorescent muscimol-bodipy (n=5 mice) injected after completion of memory retrieval to reveal the area of drug effect.
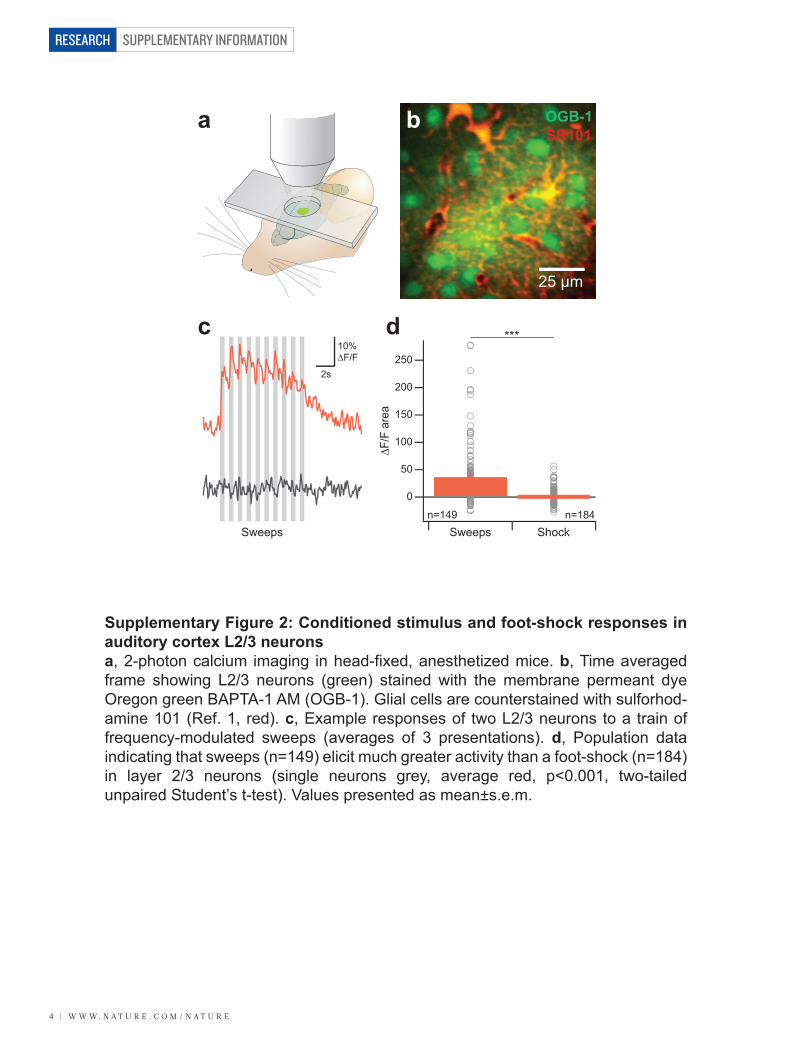
SUPPLEMENTARY INFORMATION
4 | w w w. n a t u r e . c o m / n a t u r e
RESEARCH
25 µm
10%∆F/F
2s
a b
c d
OGB-1SR101
Sweeps Shock
0
100
200
250
∆F/F
are
a
***
Sweepsn=149 n=184
150
50
Supplementary Figure 2: Conditioned stimulus and foot-shock responses in auditory cortex L2/3 neuronsa, 2-photon calcium imaging in head-fixed, anesthetized mice. b, Time averaged frame showing L2/3 neurons (green) stained with the membrane permeant dye Oregon green BAPTA-1 AM (OGB-1). Glial cells are counterstained with sulforhod-amine 101 (Ref. 1, red). c, Example responses of two L2/3 neurons to a train of frequency-modulated sweeps (averages of 3 presentations). d, Population data indicating that sweeps (n=149) elicit much greater activity than a foot-shock (n=184) in layer 2/3 neurons (single neurons grey, average red, p<0.001, two-tailed unpaired Student’s t-test). Values presented as mean±s.e.m.
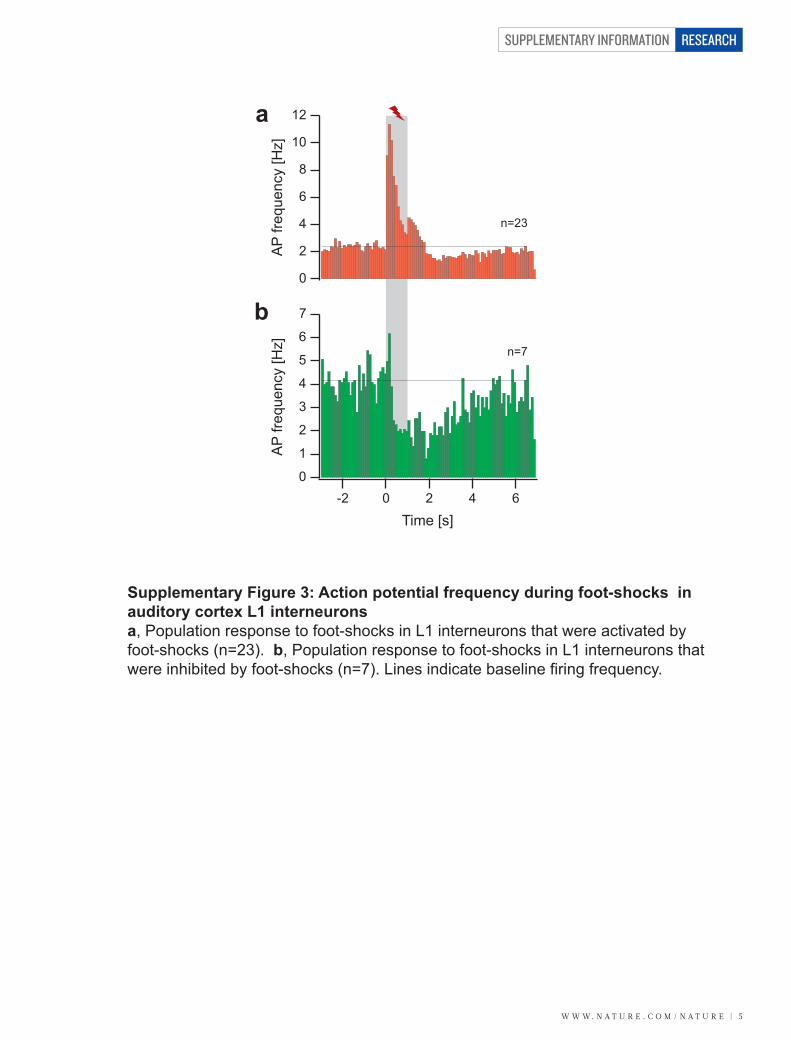
w w w. n a t u r e . c o m / n a t u r e | 5
SUPPLEMENTARY INFORMATION RESEARCH
a
bn=7
n=23
AP
frequ
ency
[Hz]
0
1
2
3
4
5
6
7
0
2
4
6
10
8
12
0 2 4 6-2
Time [s]
AP
frequ
ency
[Hz]
Supplementary Figure 3: Action potential frequency during foot-shocks in auditory cortex L1 interneuronsa, Population response to foot-shocks in L1 interneurons that were activated by foot-shocks (n=23). b, Population response to foot-shocks in L1 interneurons that were inhibited by foot-shocks (n=7). Lines indicate baseline firing frequency.
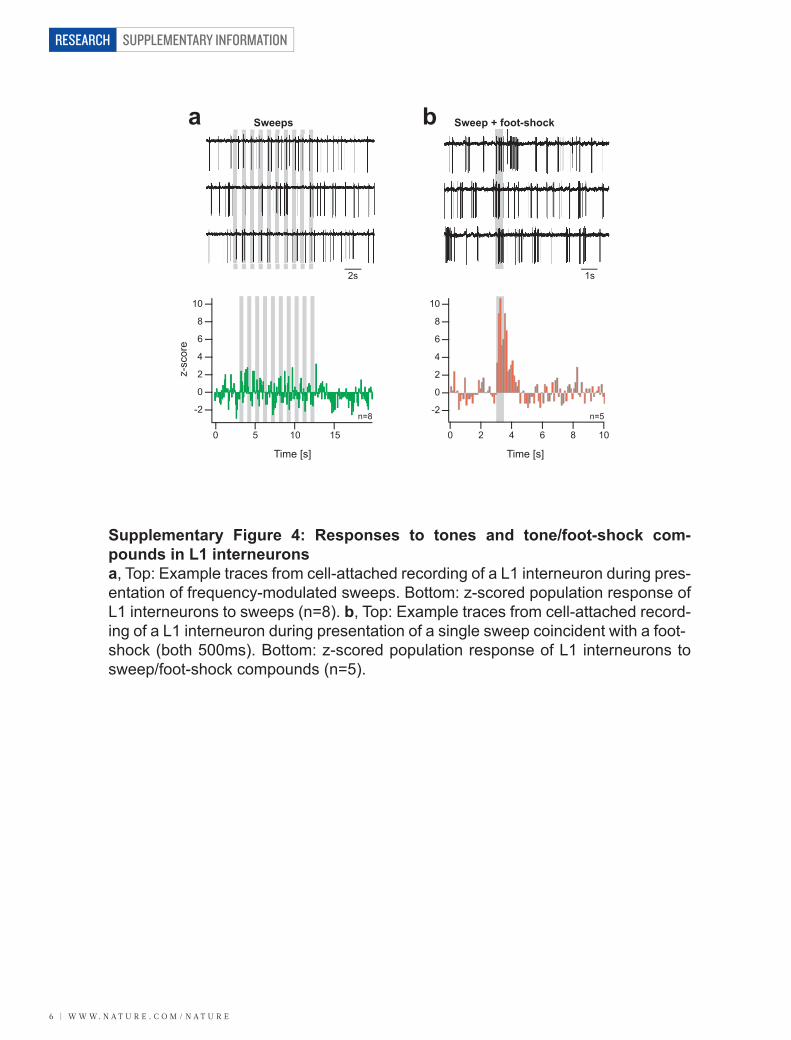
SUPPLEMENTARY INFORMATION
6 | w w w. n a t u r e . c o m / n a t u r e
RESEARCH
1s2s
a bSweeps Sweep + foot-shock
z-sc
ore
Time [s]
0 5 10 15
-2
0
2
4
6
8
10
0 2 4 6 8 10
Time [s]
n=8 n=5-2
0
2
4
6
8
10
Supplementary Figure 4: Responses to tones and tone/foot-shock com-pounds in L1 interneuronsa, Top: Example traces from cell-attached recording of a L1 interneuron during pres-entation of frequency-modulated sweeps. Bottom: z-scored population response of L1 interneurons to sweeps (n=8). b, Top: Example traces from cell-attached record-ing of a L1 interneuron during presentation of a single sweep coincident with a foot-shock (both 500ms). Bottom: z-scored population response of L1 interneurons to sweep/foot-shock compounds (n=5).
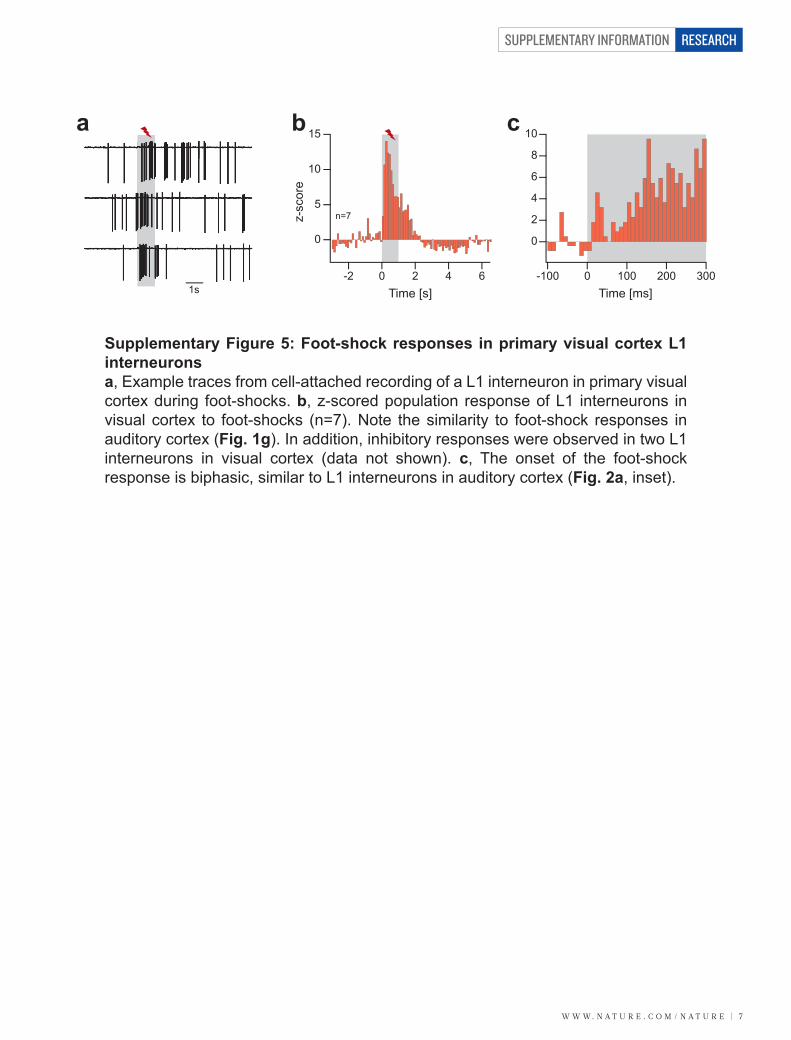
w w w. n a t u r e . c o m / n a t u r e | 7
SUPPLEMENTARY INFORMATION RESEARCH
1s
a b c
0
5
10
15
z-sc
ore
0-2 2 4 6Time [s]
-100 0 100 200 300Time [ms]
10
8
6
4
2
0
n=7
Supplementary Figure 5: Foot-shock responses in primary visual cortex L1 interneuronsa, Example traces from cell-attached recording of a L1 interneuron in primary visual cortex during foot-shocks. b, z-scored population response of L1 interneurons in visual cortex to foot-shocks (n=7). Note the similarity to foot-shock responses in auditory cortex (Fig. 1g). In addition, inhibitory responses were observed in two L1 interneurons in visual cortex (data not shown). c, The onset of the foot-shock response is biphasic, similar to L1 interneurons in auditory cortex (Fig. 2a, inset).
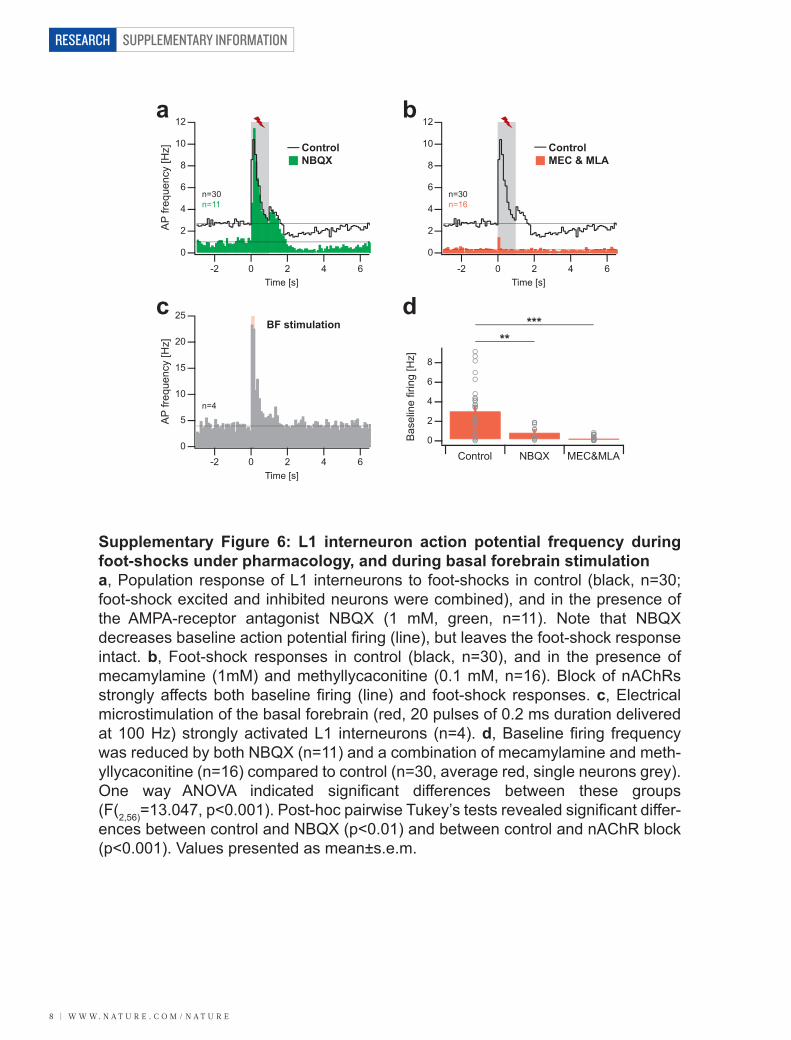
SUPPLEMENTARY INFORMATION
8 | w w w. n a t u r e . c o m / n a t u r e
RESEARCH
a b
*****
c d
ControlNBQX
ControlMEC & MLA
BF stimulation
n=30n=11
n=30n=16
n=4
0
2
4
6
8
10
12
AP
frequ
ency
[Hz]
0
2
4
6
8
10
12
0 2 4 6-2Time [s]
0 2 4 6-2Time [s]
0
5
10
15
20
25
AP
frequ
ency
[Hz]
0 2 4 6-2Time [s]
Bas
elin
e fir
ing
[Hz]
0
2
4
6
8
Control NBQX MEC&MLA
Supplementary Figure 6: L1 interneuron action potential frequency during foot-shocks under pharmacology, and during basal forebrain stimulationa, Population response of L1 interneurons to foot-shocks in control (black, n=30; foot-shock excited and inhibited neurons were combined), and in the presence of the AMPA-receptor antagonist NBQX (1 mM, green, n=11). Note that NBQX decreases baseline action potential firing (line), but leaves the foot-shock response intact. b, Foot-shock responses in control (black, n=30), and in the presence of mecamylamine (1mM) and methyllycaconitine (0.1 mM, n=16). Block of nAChRs strongly affects both baseline firing (line) and foot-shock responses. c, Electrical microstimulation of the basal forebrain (red, 20 pulses of 0.2 ms duration delivered at 100 Hz) strongly activated L1 interneurons (n=4). d, Baseline firing frequency was reduced by both NBQX (n=11) and a combination of mecamylamine and meth-yllycaconitine (n=16) compared to control (n=30, average red, single neurons grey). One way ANOVA indicated significant differences between these groups (F(2,56)=13.047, p<0.001). Post-hoc pairwise Tukey’s tests revealed significant differ-ences between control and NBQX (p<0.01) and between control and nAChR block (p<0.001). Values presented as mean±s.e.m.
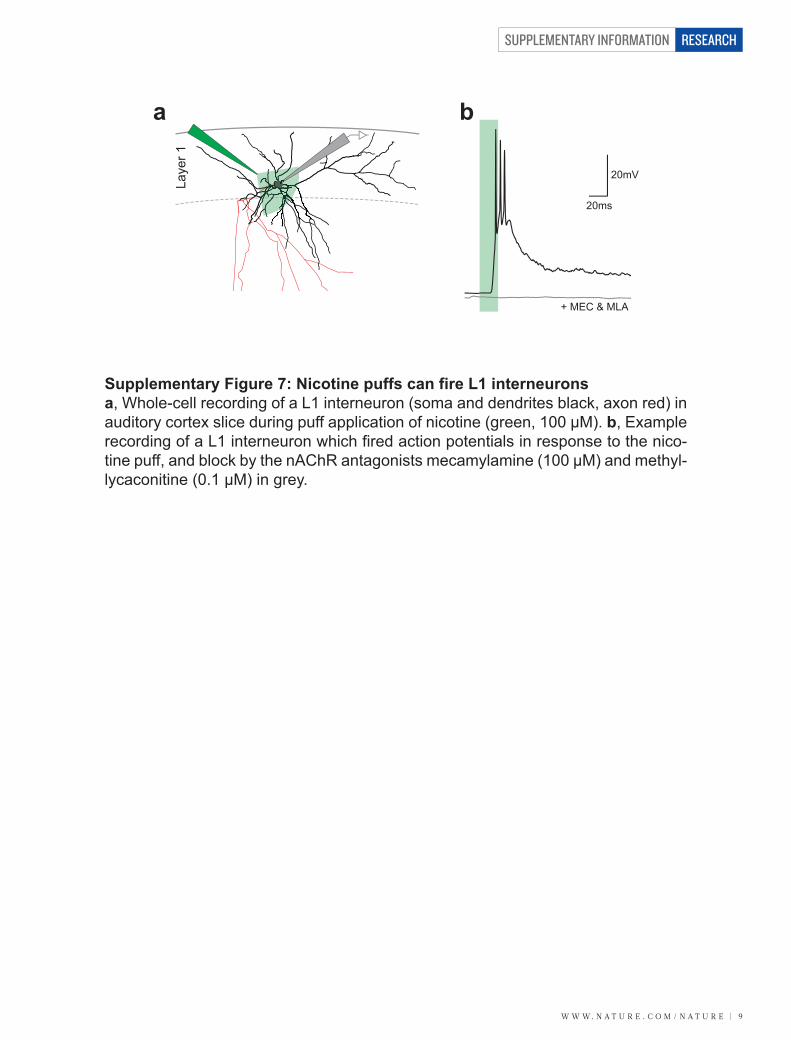
w w w. n a t u r e . c o m / n a t u r e | 9
SUPPLEMENTARY INFORMATION RESEARCH
20mV
20ms
a
Laye
r 1
b
+ MEC & MLA
Supplementary Figure 7: Nicotine puffs can fire L1 interneuronsa, Whole-cell recording of a L1 interneuron (soma and dendrites black, axon red) in auditory cortex slice during puff application of nicotine (green, 100 µM). b, Example recording of a L1 interneuron which fired action potentials in response to the nico-tine puff, and block by the nAChR antagonists mecamylamine (100 µM) and methyl-lycaconitine (0.1 µM) in grey.
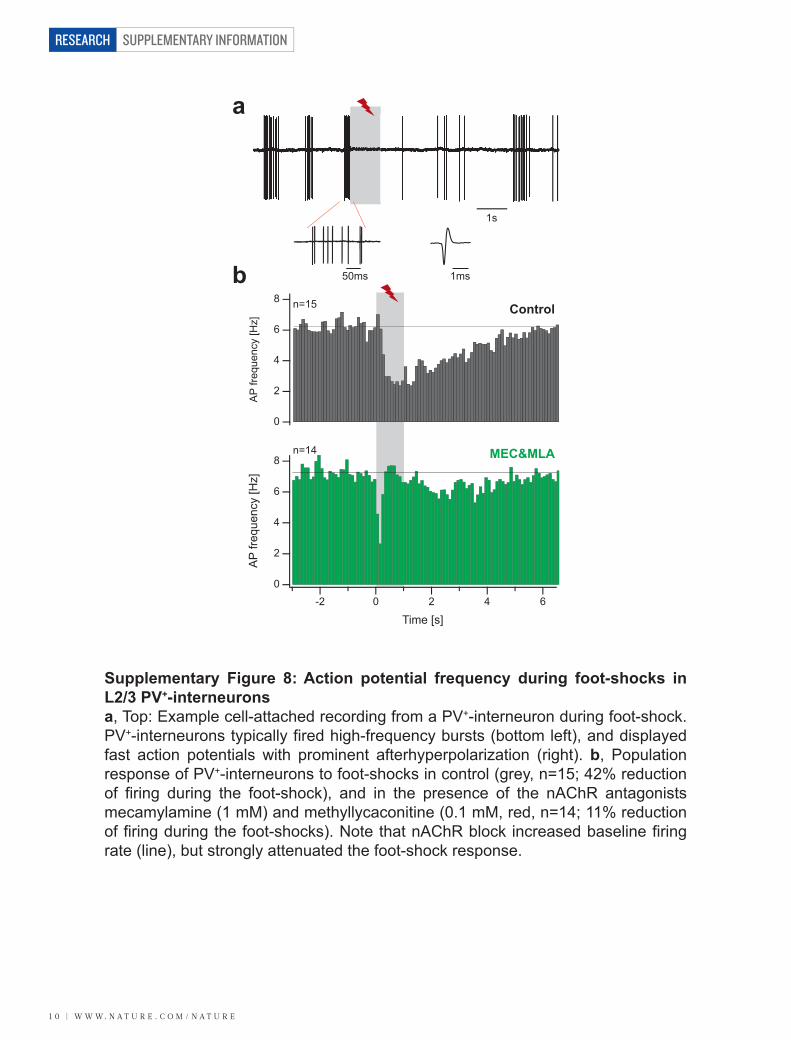
SUPPLEMENTARY INFORMATION
1 0 | w w w. n a t u r e . c o m / n a t u r e
RESEARCH
50ms 1ms
1s
a
bControl
MEC&MLA
0
2
4
6
8
AP
frequ
ency
[Hz]
0
2
4
6
8
AP
frequ
ency
[Hz]
0 2 4 6-2
Time [s]
n=14
n=15
Supplementary Figure 8: Action potential frequency during foot-shocks in L2/3 PV+-interneuronsa, Top: Example cell-attached recording from a PV+-interneuron during foot-shock. PV+-interneurons typically fired high-frequency bursts (bottom left), and displayed fast action potentials with prominent afterhyperpolarization (right). b, Population response of PV+-interneurons to foot-shocks in control (grey, n=15; 42% reduction of firing during the foot-shock), and in the presence of the nAChR antagonists mecamylamine (1 mM) and methyllycaconitine (0.1 mM, red, n=14; 11% reduction of firing during the foot-shocks). Note that nAChR block increased baseline firing rate (line), but strongly attenuated the foot-shock response.
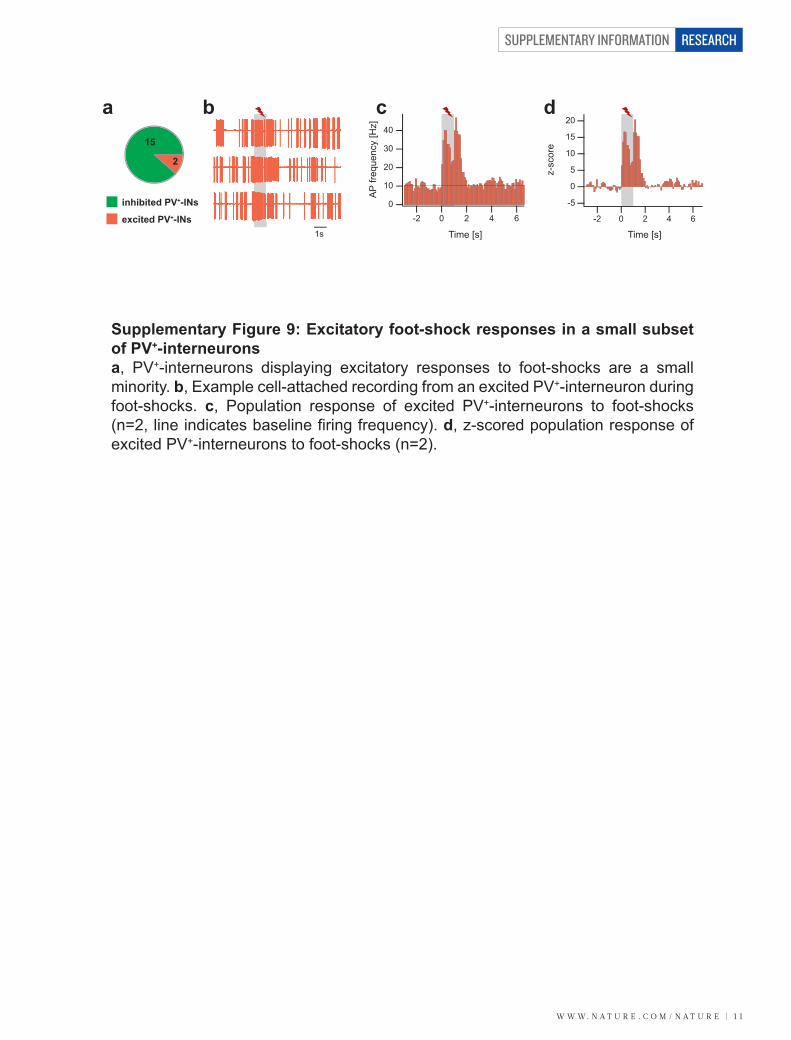
w w w. n a t u r e . c o m / n a t u r e | 1 1
SUPPLEMENTARY INFORMATION RESEARCH
15
2
1s
a b c d
inhibited PV+-INs
excited PV+-INs0
10
20
30
40
AP
frequ
ency
[Hz]
0 2 4 6-2
Time [s]
0 2 4 6-2
Time [s]
0
z-sc
ore
-5
5
10
15
20
Supplementary Figure 9: Excitatory foot-shock responses in a small subset of PV+-interneuronsa, PV+-interneurons displaying excitatory responses to foot-shocks are a small minority. b, Example cell-attached recording from an excited PV+-interneuron during foot-shocks. c, Population response of excited PV+-interneurons to foot-shocks (n=2, line indicates baseline firing frequency). d, z-scored population response of excited PV+-interneurons to foot-shocks (n=2).
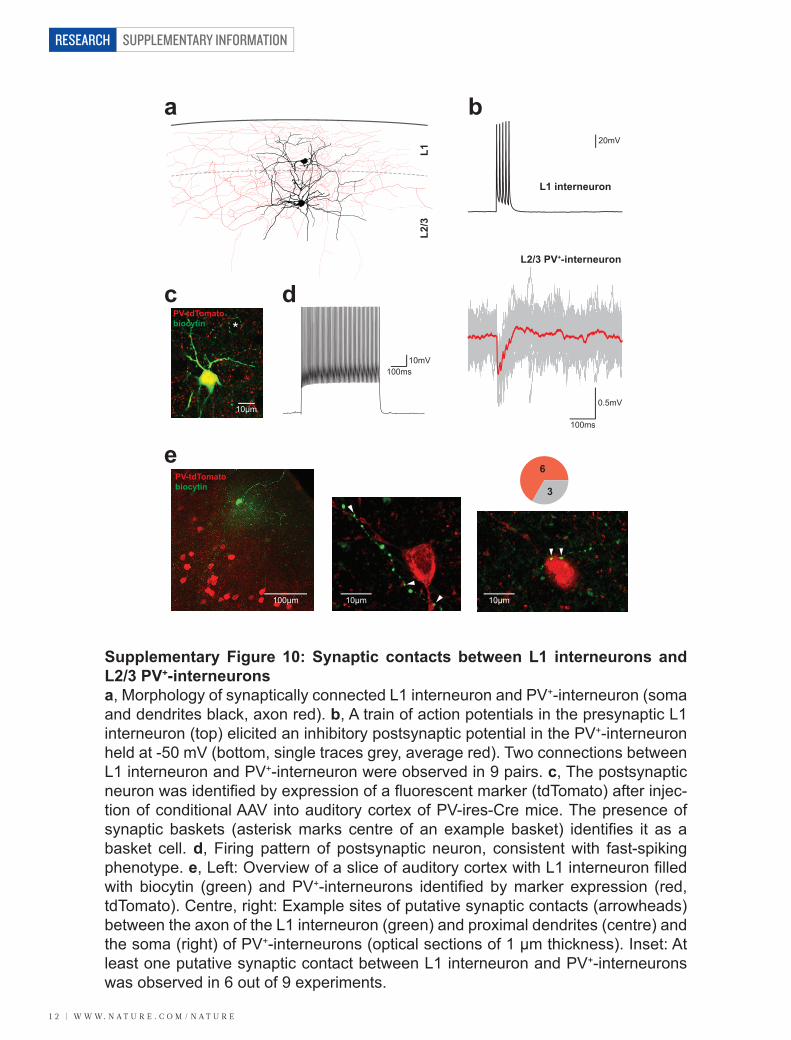
SUPPLEMENTARY INFORMATION
1 2 | w w w. n a t u r e . c o m / n a t u r e
RESEARCH
*
10µm
PV-tdTomatobiocytin
a
100ms10mV
100ms
0.5mV
20mV
L2/3 PV+-interneuron
L1 interneuron
b
c d
L1L2
/3
ePV-tdTomatobiocytin
100µm 10µm 10µm
6
3
Supplementary Figure 10: Synaptic contacts between L1 interneurons and L2/3 PV+-interneuronsa, Morphology of synaptically connected L1 interneuron and PV+-interneuron (soma and dendrites black, axon red). b, A train of action potentials in the presynaptic L1 interneuron (top) elicited an inhibitory postsynaptic potential in the PV+-interneuron held at -50 mV (bottom, single traces grey, average red). Two connections between L1 interneuron and PV+-interneuron were observed in 9 pairs. c, The postsynaptic neuron was identified by expression of a fluorescent marker (tdTomato) after injec-tion of conditional AAV into auditory cortex of PV-ires-Cre mice. The presence of synaptic baskets (asterisk marks centre of an example basket) identifies it as a basket cell. d, Firing pattern of postsynaptic neuron, consistent with fast-spiking phenotype. e, Left: Overview of a slice of auditory cortex with L1 interneuron filled with biocytin (green) and PV+-interneurons identified by marker expression (red, tdTomato). Centre, right: Example sites of putative synaptic contacts (arrowheads) between the axon of the L1 interneuron (green) and proximal dendrites (centre) and the soma (right) of PV+-interneurons (optical sections of 1 µm thickness). Inset: At least one putative synaptic contact between L1 interneuron and PV+-interneurons was observed in 6 out of 9 experiments.
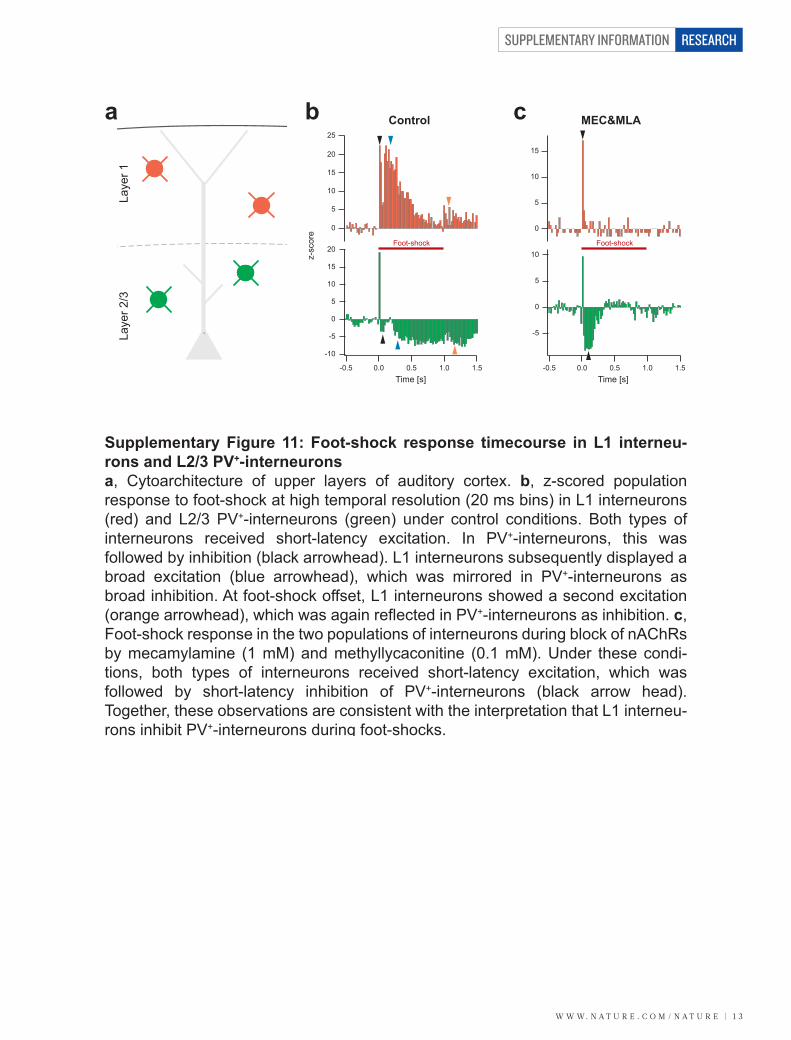
w w w. n a t u r e . c o m / n a t u r e | 1 3
SUPPLEMENTARY INFORMATION RESEARCH
Laye
r 1La
yer 2
/3a b cControl MEC&MLA
0
5
10
15
20
25
z-sc
ore
0
5
10
15
20
-5
-10
0.0-0.5 0.5 1.51.0Time [s]
0.0-0.5 0.5 1.51.0Time [s]
0
5
10
-5
0
5
10
15
Foot-shock Foot-shock
Supplementary Figure 11: Foot-shock response timecourse in L1 interneu-rons and L2/3 PV+-interneuronsa, Cytoarchitecture of upper layers of auditory cortex. b, z-scored population response to foot-shock at high temporal resolution (20 ms bins) in L1 interneurons (red) and L2/3 PV+-interneurons (green) under control conditions. Both types of interneurons received short-latency excitation. In PV+-interneurons, this was followed by inhibition (black arrowhead). L1 interneurons subsequently displayed a broad excitation (blue arrowhead), which was mirrored in PV+-interneurons as broad inhibition. At foot-shock offset, L1 interneurons showed a second excitation (orange arrowhead), which was again reflected in PV+-interneurons as inhibition. c, Foot-shock response in the two populations of interneurons during block of nAChRs by mecamylamine (1 mM) and methyllycaconitine (0.1 mM). Under these condi-tions, both types of interneurons received short-latency excitation, which was followed by short-latency inhibition of PV+-interneurons (black arrow head). Together, these observations are consistent with the interpretation that L1 interneu-rons inhibit PV+-interneurons during foot-shocks.
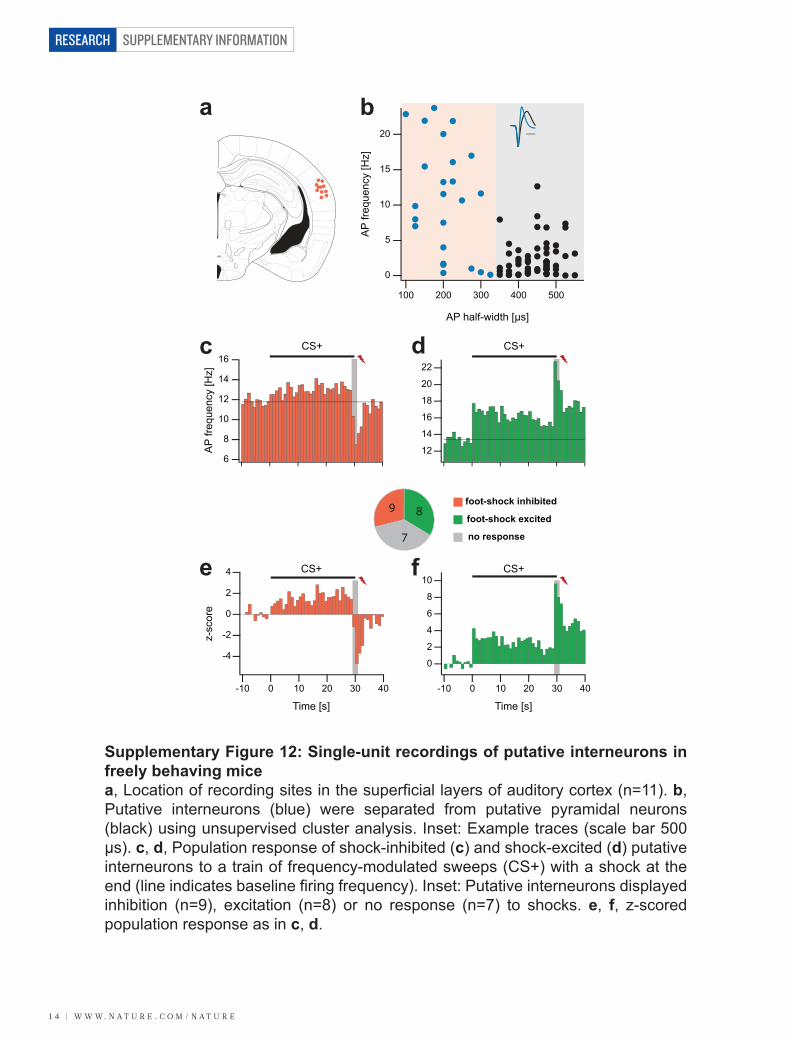
SUPPLEMENTARY INFORMATION
1 4 | w w w. n a t u r e . c o m / n a t u r e
RESEARCH
9
7
8
c d
e f
AP
frequ
ency
[Hz]
6
8
10
12
14
16
12
16
20
-10 0 10 20 30 40
Time [s]
-10 0 10 20 30 40
Time [s]
0
-2
-4
2
4
z-sc
ore
0
4
8
CS+ CS+
CS+ CS+
foot-shock inhibited
foot-shock excited
no response
RSA
RSG
Or
Or
AuD
Au1
AuV
Ect VLGPCVLGMC
CA3
SLuCA2
MolLMol
Py
CA1
Rad
GrDG
PoDG DG
LPLRDLG
IGLIMA
LPMC
PrC
FC
PF
SubGPRh
LEnt DEn
Pir VEn BLP
Gem
ZIV
ZID
VPM
BMPAHiAL AHiPM
PLCoPMCo
DG
VTMPMV
ArcLP
SMT
V2MMV2ML
V2L
S
SPFPCRI
PRF
SNR
PMD
SuMM
APir
Py
CA3
OPT
PSTh
LH
LPMR
APTD
PoTeA
V1
IG
a b20
15
10
5
0
AP
frequ
ency
[Hz]
100 200 300 400 500
AP half-width [µs]
Supplementary Figure 12: Single-unit recordings of putative interneurons in freely behaving micea, Location of recording sites in the superficial layers of auditory cortex (n=11). b, Putative interneurons (blue) were separated from putative pyramidal neurons (black) using unsupervised cluster analysis. Inset: Example traces (scale bar 500 µs). c, d, Population response of shock-inhibited (c) and shock-excited (d) putative interneurons to a train of frequency-modulated sweeps (CS+) with a shock at the end (line indicates baseline firing frequency). Inset: Putative interneurons displayed inhibition (n=9), excitation (n=8) or no response (n=7) to shocks. e, f, z-scored population response as in c, d.
14
18
22
2
6
10

w w w. n a t u r e . c o m / n a t u r e | 1 5
SUPPLEMENTARY INFORMATION RESEARCH
Pea
k (n
orm
aliz
ed)
Vm [mV]
1.0
0.8
0.6
0.4
0.2
-100 -90 -80 -70 -60 -50
n=6
-62mV
-80mV
-97mV
5mV
1s
Supplementary Figure 13: Positive relationship between foot-shock response amplitude and membrane potential in pyramidal neuronsLeft: Example whole-cell current-clamp recording of foot-shock responses in an auditory cortex pyramidal neuron at different membrane potentials. Right: Positive relationship between normalized response amplitude and membrane potential (n=6). This is consistent with disinhibition, because the effect of disinhibition increases with increasing distance from the GABAA reversal potential. However, in most of the pyramidal neurons recorded the reversal potential of the response is more hyperpolarized than expected for pure disinhibition (calculated ECl for the solu-tions used is -84 mV). This likely suggests that the foot-shock response is mediated by weak excitation along with strong disinhibition, since such compound responses can reverse at more hyperpolarized potentials than pure disinhibition2.
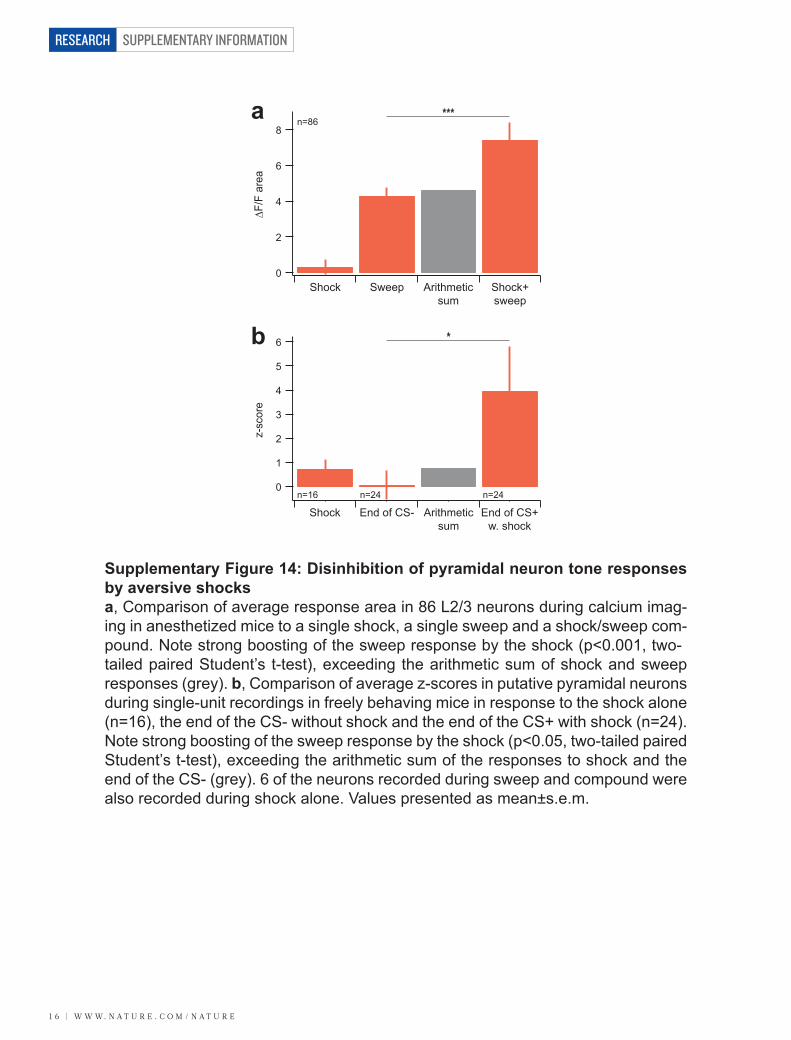
SUPPLEMENTARY INFORMATION
1 6 | w w w. n a t u r e . c o m / n a t u r e
RESEARCH
a
0∆F
/F a
rea
2
4
6
8
Shock Sweep Shock+sweep
b
z-sc
ore
0
1
2
3
4
5
6
Shock End of CS- End of CS+w. shock
***n=86
n=16 n=24 n=24
*
Arithmeticsum
Arithmeticsum
Supplementary Figure 14: Disinhibition of pyramidal neuron tone responses by aversive shocksa, Comparison of average response area in 86 L2/3 neurons during calcium imag-ing in anesthetized mice to a single shock, a single sweep and a shock/sweep com-pound. Note strong boosting of the sweep response by the shock (p<0.001, two-tailed paired Student’s t-test), exceeding the arithmetic sum of shock and sweep responses (grey). b, Comparison of average z-scores in putative pyramidal neurons during single-unit recordings in freely behaving mice in response to the shock alone (n=16), the end of the CS- without shock and the end of the CS+ with shock (n=24). Note strong boosting of the sweep response by the shock (p<0.05, two-tailed paired Student’s t-test), exceeding the arithmetic sum of the responses to shock and the end of the CS- (grey). 6 of the neurons recorded during sweep and compound were also recorded during shock alone. Values presented as mean±s.e.m.
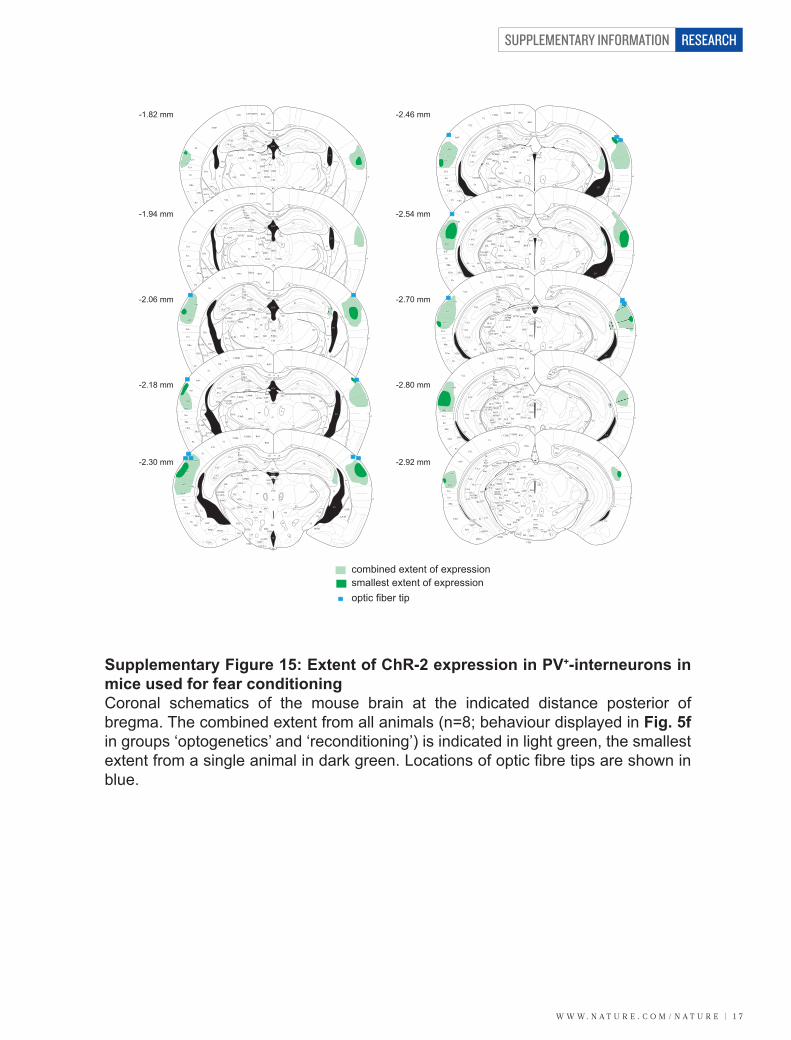
w w w. n a t u r e . c o m / n a t u r e | 1 7
SUPPLEMENTARY INFORMATION RESEARCH
combined extent of expressionsmallest extent of expressionoptic fiber tip
RSA
RSG
S1BF
CA2
Or
CA1RadPy
LMolMol
GrDG FC
DGCA3
DLGLPLR
LPMR
CLVLG
VPLVPM
OPC
PC
MDL PV
MDC
MDM
IMD
CM
SLu
PoDG
LHbM
LHbL
MHb
Po
S2
CPu
st
cc
dhc
df
cg
hbc
fr
Ect
PRh AStr
Rt
DEn
LaVM
CeL
CeCLaVL
Pir
VEnBLV
BMP
BSTIA
PLCo PMCo
I
ME
VMHVLVMHC
VMHDM
DMV
DMDDMC
MCLH
STh
MePD
MePV
LH
ZIV
VMSub
Rh
Re
mt
PH
ZIDVRe
Pe
PeFTe
MTu ArcDArcL
f
ns
ml
cpopt
Isox
st
eml st
ic
alv
ec
rf
fi
LV
hf
BLA
3V
D3V
LaDL
S1Tr
AuV
TeA
LPtAMPtA
IG
AuV
-2.30 mm
LPtAMPtA RSA
cg
alv
cc df
dhcFC
hbcMHb
LHbMLHbLCL
LPMRLPLR
SLu
CA2
S1BF
CA3
Or
CA1RadPy
LMolMol
GrDG
DGPoDG
VLG
Po
MDL
PVIMD
fr
MDC
MDM PoMn
CM
VM
VPL
VPMRtPC
OPC
ZIV ZIDA11
PePHDMC
DMD
mt
scp
ml
soxnsMCLH
STh
AStr
LaVM
CeL
BSTIABLA
BMP
BLP
LaDL
CPu
DEn
LaVL
VEnPir
PRh
Ect
AuV
emlfi
ic
st
ic
opt
I
cp
ec
f
ArcDArcL
ME
VMHDMVMHCVMHVL
DMVLH
PeF
MTu
MePVTe
PMCo
AHiAL
PLCo
BLV
D3V
3V
Gus
rf
LV
hf
RSG
DLG
MePD
A12
V2L
TeA
IG
LPtA RSA
RSG
CA1
CA2
OrPyRadLMolMol
GrDG
PoDG
FC
AuDCA3
VLGMC CL LHb
MHb
DG
LPMRLPLRDLG
AuV
Ect
CPu
PRh LaDL AStr
DEn LaVMLaVL
BLA SThZIV ZID
Rt VPL VPM
Po
VLGPC IMA
PC
PFPV
OPC MD
IMD
PoMn
CM
Gus
SPFVM
PirVEn
BLP
BMP
BLV
PLCo
AHiAL MePV
BSTIA
MePD
PMCo
LH PeF
TeMTu
VMH
ArcL ME
ArcD
DMDMC
PH
f
nsmt
3V
scp
ml
sox
cp optCeL
cg
cc df
dhc
alv
hbc
freml
fi
ic st
ec
rf
D3V
LV
S1
hf
PSTh
SLu
A12
MPtA
V2L
TeA
IG
AuD
AuV
Rt
AuD
Au1
AuV
Ect
PRh
LEntDEn
Pir
VEn
BLV
BMP
BLP
LaVLLaVM
LaDL
AStr
CPu
CA3SLu
CA2
IGL VLGMCVLGPC
DLG LPLRLPMR
PoDG
GrDGMolLMol
PyOr
S1Rad
FCdhc
cc df
DG
MHbLHb
PVP
hbc
frPF
Po
VPLVPM
ZIVZID
OPC
Gus
SPFVM
STh
PSTh
MePVBSTIA
AHiAL
Gem PH
DTM
mt ns
scp
ml
alv
eml
opt
ec
rf
fi
st
ic
opt
sox
cp
MePDst
cg
RSA
CA1
RSG
PLCoPMCo
DG LHSMT
DMf
ArcMPArcLP
Te
ME
VTM
IMA
LV
PrC
3V
D3V
hf
pv
APTD
V2MMV2MM
V1V2L
TeA
IG
AuV
TeATeAT
RSA
RSG
Or
S1
AuD
Au1
AuV
Ect
CPu
VLGPCVLGMC
CA3SLu
CA2
MolLMol
Py
CA1Rad
GrDG
PoDG DG
LPLRDLG
IGL
VPLPo
LPMRAPTD
SCOPrC
hbc
FC
cc df
dhc
cg
alv
emlPVPPF
SubG
PRh
LEntDEn
La
PirVEn
BLP STh
PSTh
Gem
ZIV ZID
VPM
GusSPF
fr
ml
opt
sox
cpnsmtPH
BMP
PLCoPMCo
DG LH
VTM
TePMV
ArcMP
ArcLPMe
LaDL
LaVM
SMTDTM
f
MePD
rf
ec
fi
D3V
3V
3V
LV
pv
pc
FF PR
hf
IMA
scp
AHiAL
TeA
V2L
V1V2ML
V2MM
IG
AuD
Au1
AuVCPu
TeATeAT
-1.82 mm
-1.94 mm
-2.06 mm
-2.18 mm
-2.46 mm
-2.54 mm
-2.70 mm
-2.80 mm
RSA
RSG
Or
Or
AuD
Au1
AuV
Ect VLGPCVLGMC
CA3
SLuCA2
MolLMol
Py
CA1
Rad
GrDG
PoDG DG
LPLRDLG
IGLIMA
LPMC
PrC
FC
ccdhc
cg
alv
PF
SubGPRh
LEnt DEn
Pir VEn BLP
Gem
ZIV
ZID
VPM
fr
mlopt
cpns
PH
BMPAHiAL AHiPM
PLCoPMCo
DG
VTMPMV
ArcMPArcLP
LaDL
LaVM
SMTDTM
f
rf
ec
fi
3V
LV
V2MMV2ML
V2L
S
SPFPCRI
PRF
SNR
PMD
SuMM
mt
D3V
pc
APir
Py
CA3
str
OPT
hf
PSTh
LH
scp
LPMR
APTD
PoTeA
V1
IG
Au1
AuV
mt
CA3
Or
VLGPCVLGMC
CA3
SLu
CA2
MolLMol
Py
CA1Rad
GrDG
PoDGDG
LPLR
DLG
IGLIMA
LPMR
APTD
FC
PFVPM
S
D3V
pc
OPT
RSG
RSAV2MMV2ML
V2L
AuD
Au1
AuV
Ect
PRh
LEnt
scc
cg
ec
alv
str
fiopt
cp
ns
ml
mtg
f
dhc
PAG
SCO
alvOr
DEn
PirBLP
AHiPMAPir
BMP
PLCo PMV
PMD
alv
ArcLP
ArcMP
SuMMSMT
SNR
SNC
ZID
ZIV
PR scp
fr
Eth
ScPo
PPT
RIF
PH
SPFPCSubG
DTMDG
LH
LT
bsc
Py
Py CA3
VTM
LV
rf
LPMC
3V
RPF
PSTh
hf
PMCo
AHiAL
TeA
V1
AuD
Au1
AuV
Or
VLGPC
VLGMC
CA3
SLu
CA2
MolLMol
Py
CA1
Rad
GrDG
PoDGDG
LPLRDLG
IGL
IMA LPMRAPTD
FCS
D3V
D3V
pc
OPT
RSG
RSAV2MMV2ML
V1
V2L
AuD
Au1
AuV
Ect
PRh
LEnt
cg
ec
alv
alv
str
opt
cp
ns
ml
mtg
dhc
SCO
Or
Or
DEn
PirBLP
APirBMP
PMCo PMD
ArcMP
SuMM
SNR
ZIDZIV
PR scpfr
Eth
REth
PPT
RIFPH
SPFPCSubG
DG
LT
bsc
Py
Py
VTM
SLu
LM ML MMMMn
MRe
SuML
sumxLH
PCom
OT
APTV
MPT
Po
PP
LPMC
CA1
PAG
SNC
SNC
fmj
CA3LV
RPF
MCPC
hf
pm
rf
AHiPM
f
TeA
AuD
Au1
AuV
pm
Or
VLGPCVLGMC
CA3
SLu
CA2
MolLMol
Py
CA1Rad
GrDG
hf
PoDG
DG
DLG
IGL
IMA
APTD
FCS
D3V
pc
OPT
RSG
RSAV2MMV2ML
V1
V2L
AuD
Au1
AuV
Ect
PRh
LEnt
cg
ec
alv
alvopt
cp
ns
ml
mtg
dhc
SCO
Or
Or
DEn
PirBLP
APir
PMCoArcMP
SuMMSNR
ZIDZIV
PR scpfr
MCPC
Eth
REth
PPT
RI
SPFPC
SubG
GrDG
LT
bsc
Py
VTM
AHiPM
SLu
LM ML MMMMn
MRe
SuML
sumx
PCom
OT
APTV
MPT
PoT
PP
LPMCLPLC
PAG
SNC
SNL
fmj
PLi
DkPILIntG
MGV
SG
MGM
MGD
EW
LH
f
CA3
rf
CA2
SComRPF
LV
TeA
AuD
Au1
AuV
Aq
pm
Or
VLGPCVLGMC
CA3SLu
SLu
CA2
Mol
LMol
Py
CA1
Rad
Rad
GrDGPoDG
DG
DLGIGL
APTD
S
pc
RSG
RSA
Ect
PRh
cg
ec
alv
alvopt
cp
ml
mp
Or
Or
BLP
APir
PMCo
SuMSNR
fr
MCPC
REth
PPT
SPFPCSubG
LT
bsc
Py
VTM
VTMAHiPMLM ML MM
MMn
sumx
PCom
OT
APTV
PoT
LPMC
LPLC
PAG
SNC
SNL
fmj
PLi
DkPIL
MGV SGMGMMZMG
MGD
EW
VTA
f
CA3
LV
SC
RPC
ZI
CA1
PoDGGrDG
PBP
DpMe
PP
rf
dhc
CA2
LEnt
hf
RPF
InC
InCG
mtg
scp
AuD
Au1
AuV
V2L
V1
V2MMV2ML
TeA
Au1
AuV
-2.92 mm
Supplementary Figure 15: Extent of ChR-2 expression in PV+-interneurons in mice used for fear conditioningCoronal schematics of the mouse brain at the indicated distance posterior of bregma. The combined extent from all animals (n=8; behaviour displayed in Fig. 5f in groups ‘optogenetics’ and ‘reconditioning’) is indicated in light green, the smallest extent from a single animal in dark green. Locations of optic fibre tips are shown in blue.
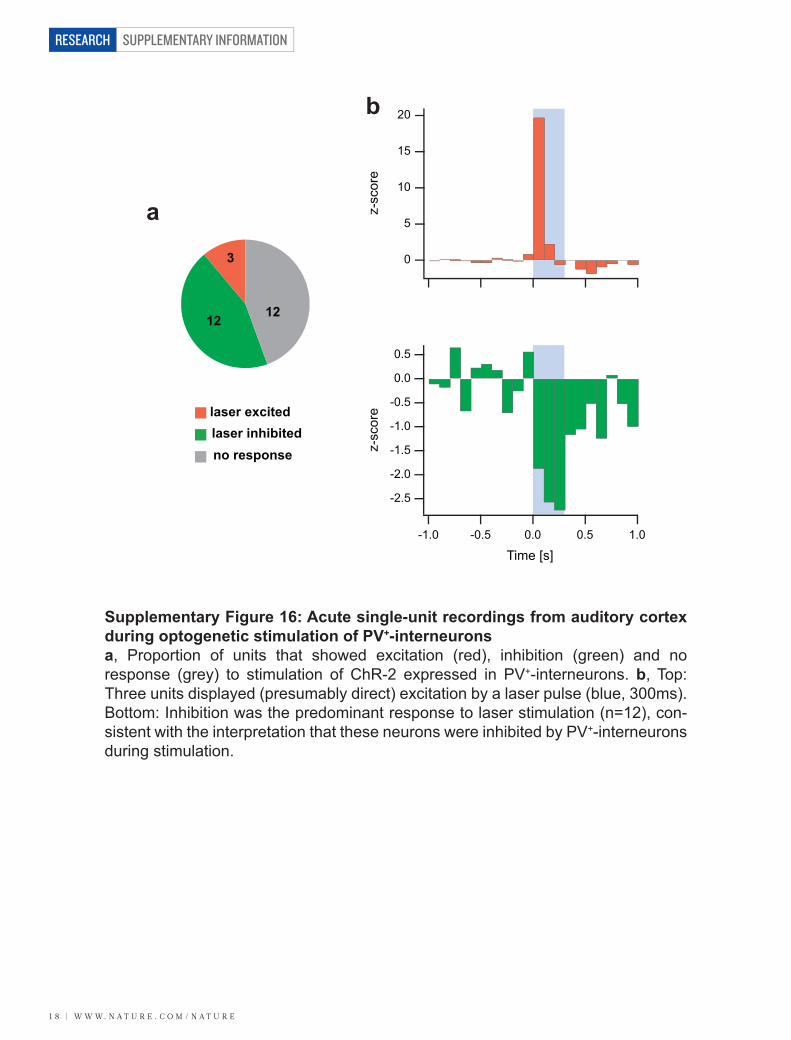
SUPPLEMENTARY INFORMATION
1 8 | w w w. n a t u r e . c o m / n a t u r e
RESEARCH
a
b
laser excitedlaser inhibitedno response
3
1212
0
5
10
15
20
z-sc
ore
0.0
0.5
-0.5
-1.0
-1.5
-2.0
-2.5
z-sc
ore
0.0 0.5-0.5-1.0 1.0
Time [s]
Supplementary Figure 16: Acute single-unit recordings from auditory cortex during optogenetic stimulation of PV+-interneuronsa, Proportion of units that showed excitation (red), inhibition (green) and no response (grey) to stimulation of ChR-2 expressed in PV+-interneurons. b, Top: Three units displayed (presumably direct) excitation by a laser pulse (blue, 300ms). Bottom: Inhibition was the predominant response to laser stimulation (n=12), con-sistent with the interpretation that these neurons were inhibited by PV+-interneurons during stimulation.
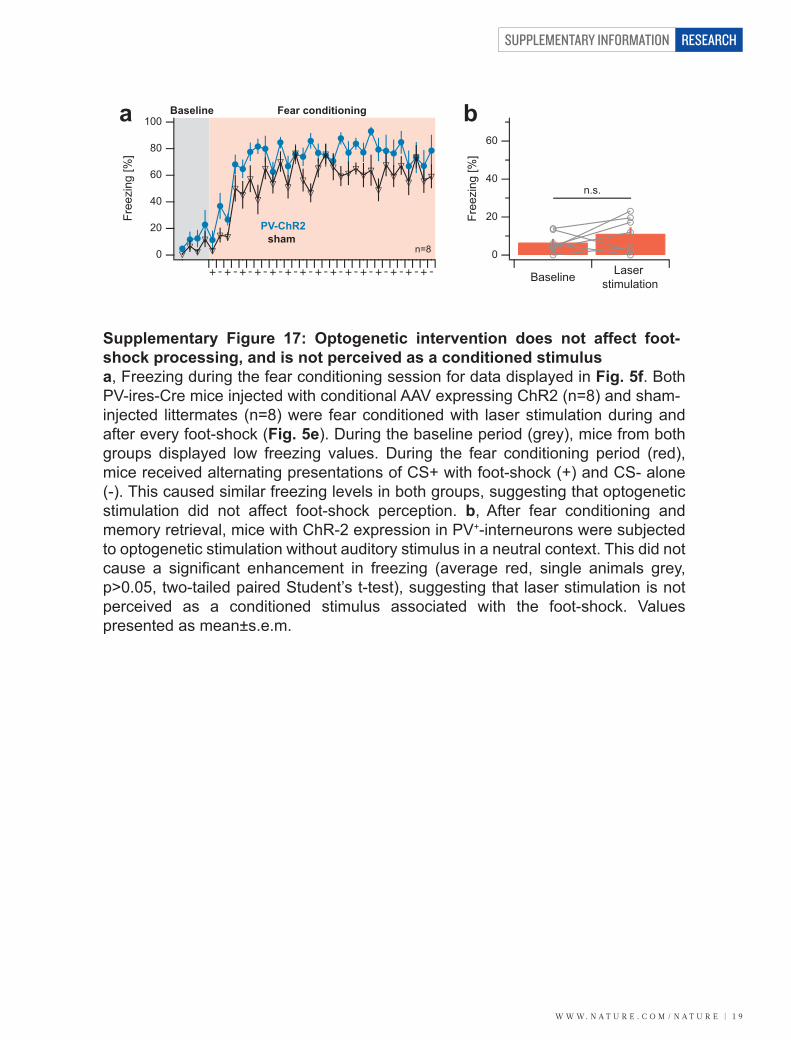
w w w. n a t u r e . c o m / n a t u r e | 1 9
SUPPLEMENTARY INFORMATION RESEARCH
a b
n.s.
PV-ChR2sham
0
20
40
60
80
100Fr
eezi
ng [%
]Baseline Fear conditioning
n=8
Baseline Laser stimulation
0
20
40
60
Free
zing
[%]
Supplementary Figure 17: Optogenetic intervention does not affect foot-shock processing, and is not perceived as a conditioned stimulusa, Freezing during the fear conditioning session for data displayed in Fig. 5f. Both PV-ires-Cre mice injected with conditional AAV expressing ChR2 (n=8) and sham-injected littermates (n=8) were fear conditioned with laser stimulation during and after every foot-shock (Fig. 5e). During the baseline period (grey), mice from both groups displayed low freezing values. During the fear conditioning period (red), mice received alternating presentations of CS+ with foot-shock (+) and CS- alone (-). This caused similar freezing levels in both groups, suggesting that optogenetic stimulation did not affect foot-shock perception. b, After fear conditioning and memory retrieval, mice with ChR-2 expression in PV+-interneurons were subjected to optogenetic stimulation without auditory stimulus in a neutral context. This did not cause a significant enhancement in freezing (average red, single animals grey, p>0.05, two-tailed paired Student’s t-test), suggesting that laser stimulation is not perceived as a conditioned stimulus associated with the foot-shock. Values presented as mean±s.e.m.

SUPPLEMENTARY INFORMATION
2 0 | w w w. n a t u r e . c o m / n a t u r e
RESEARCH
References 1 Nimmerjahn, A., Kirchhoff, F., Kerr, J.N., & Helmchen, F., Sulforhodamine 101 as a specific marker of astroglia in the neocortex in vivo. Nat Methods 1 (1), 31-37 (2004). 2 Manookin, M.B., Beaudoin, D.L., Ernst, Z.R., Flagel, L.J., & Demb, J.B., Disinhibition combines with excitation to extend the operating range of the OFF visual pathway in daylight. J Neurosci 28 (16), 4136-4150 (2008).

- 49 -
II/ Results Part 2
A/ Introduction
The second objective of my thesis was to identify the functional role of cortical PV
INs in the expression of associative fear memory. Because the mPFC is known to be a key
structure involved in fear memory expression, we concentrated our analysis on this neuronal
structure. By using single unit recordings, we characterized the changes in neuronal activity of
prefrontal neurons evoked by the conditioned stimulus (CS+, associated with the US during
fear conditioning) during fear expression in behaving mice. Our results revealed that
following fear conditioning, CS+ presentations induced a powerful inhibition of prefrontal fast
spiking putative INs, a population of neurons displaying characteristic of PV INs. By using a
combination of single unit recordings and optogenetic approaches in behaving mice, we next
demonstrated that these CS+-inhibited INs were indeed PV INs. Furthermore, we showed that
the neuronal inhibition of PV INs was causally related to conditioned fear expression via two
main mechanisms: the disinhibition of prefrontal PNs and the resetting of local theta
oscillations that synchronizes prefrontal PNs. In a last experiment, we demonstrated using
extracellular stimulations that the PNs displaying disinhibition, preferentially target the BLA.
Our results identify two complementary neuronal mechanisms both mediated by prefrontal
PV INs that precisely coordinate and enhance the neuronal efficiency of prefrontal PNs to
drive fear expression.
B/ Article 2: “Prefrontal parvalbumin interneurons shape neuronal activity to
drive fear expression”
Julien Courtin, Fabrice Chaudun, Robert R. Rozeske, Nikolas Karalis, Cecilia Gonzalez-
Campo, Hélène Wurtz, Azzedine Abdi, Jerome Baufreton, Thomas C.M. Bienvenu and Cyril
Herry.
In press at Nature.
ABSTRACT
Accurate transmission of information in the brain requires precise temporal pattern of
activity and synchronous firing of pyramidal neurons to efficiently drive targeted neuronal
regions. The medial prefrontal cortex regulates fear behaviour via projections to the
amygdala, a neuronal structure encoding associative fear memories. However, the prefrontal

- 50 -
neuronal mechanisms allowing for the precise temporal control of fear behaviour are largely
unknown. Using single unit recordings and optogenetic manipulations in behaving mice, we
show that expression of fear behaviour is causally related to the phasic inhibition of prefrontal
parvalbumin-expressing interneurons. Inhibition of parvalbumin interneuron activity
disinhibits prefrontal pyramidal neurons and synchronizes their firing by resetting local theta
oscillations, leading to fear expression. Our results identify two complementary neuronal
mechanisms both mediated by prefrontal parvalbumin-expressing interneurons that precisely
coordinate and enhance the neuronal efficiency of prefrontal pyramidal neurons to drive fear
expression.
INTRODUCTION
Synchronization of spiking activity within and among neuronal networks is a
fundamental process that allows for the precise and accurate transmission of information to
drive behavioural responses (Buzsaki & Draguhn 2004, Salinas & Sejnowski 2001, Singer
1999, Varela et al 2001, Womelsdorf et al 2007). In cortical areas, synchronization of
pyramidal neuron spiking activity is an effective mechanism for information coding that is
precisely regulated by GABAergic interneurons through the generation of neuronal
oscillations (Cardin et al 2009, Cobb et al 1995, Markram, H. et al 2004, Royer et al 2012).
Although neuronal synchrony has been demonstrated to be crucial for sensory, motor, and
cognitive processing (Benchenane et al 2010, Friedrich et al 2004, Ishikane et al 2005, Riehle
et al 1997, Stopfer et al 1997), it has not been investigated at the level of defined neuronal
circuits involved in the control of emotional behaviour.
In mammals, emotional behaviour is often studied using auditory fear conditioning, a
robust learning paradigm in which animals learn to associate an initially neutral stimulus (the
conditioned stimulus, CS) with a coincident aversive foot-shock (the unconditioned stimulus,
US). Re-exposure to the CS induces the expression of a broad range of conditioned fear
responses, including an immobilization reaction called freezing. Converging evidence
indicates that following learning, the behavioural expression of conditioned fear memories is
precisely regulated by the dorsal medial prefrontal cortex (dmPFC), including the cingulate
and prelimbic subregions (Burgos-Robles et al 2009, Corcoran et al 2007, Sierra-Mercado et
al 2011, Tang et al 2005, Vidal-Gonzalez et al 2006). This control over fear behaviour is
thought to rely on the activation of specific prefrontal projections to the basolateral complex
of the amygdala (BLA), a structure known to encode associative fear memories (Knapska et
al 2012, LeDoux 2000, Pape & Pare 2010). However, it remains to be established how the

- 51 -
precise temporal control of fear behaviour is achieved at the level of specific prefrontal
neuronal circuits.
Here we investigate the contribution of dmPFC interneurons and the associated
neuronal oscillatory mechanisms involved in the temporal organization of spiking activity
underlying conditioned fear behaviour. Our results demonstrate that the inhibition of neuronal
activity in a subpopulation of prefrontal parvalbumin-expressing (PV) interneurons disinhibits
and temporally synchronizes firing activity of pyramidal neurons during precise time
windows, via a phase resetting of local theta oscillations, to accurately drive fear expression.
RESULTS
Prefrontal inhibitory neurons involved in fear behaviour
To identify the prefrontal circuitry involved in conditioned fear behaviour, C57BL6/J
mice were chronically implanted with recording electrodes aimed at the dmPFC, and were
submitted to a discriminative fear conditioning protocol consisting of two different auditory
conditioned stimuli (CS) (Fig. 12a). The first CS (CS+) was repeatedly associated with an
aversive foot-shock (US) whereas the second CS (CS-) was not, and served as an internal
control. To increase the likelihood of recording from both pyramidal neurons and interneurons
during fear expression, mice were submitted to several behavioural sessions following fear
conditioning. These included two extinction sessions and one retrieval session, all performed
in a different context from the conditioning context (Fig. 12a). Twenty-four hours following
fear conditioning, mice displayed a selective increase in freezing during CS+
presentations
which returned to baseline levels by the end of the second extinction session (Fig. 12b). One
week later, CS+ presentations in the extinction context induced a selective recovery of fear
responses (Fig. 12b).
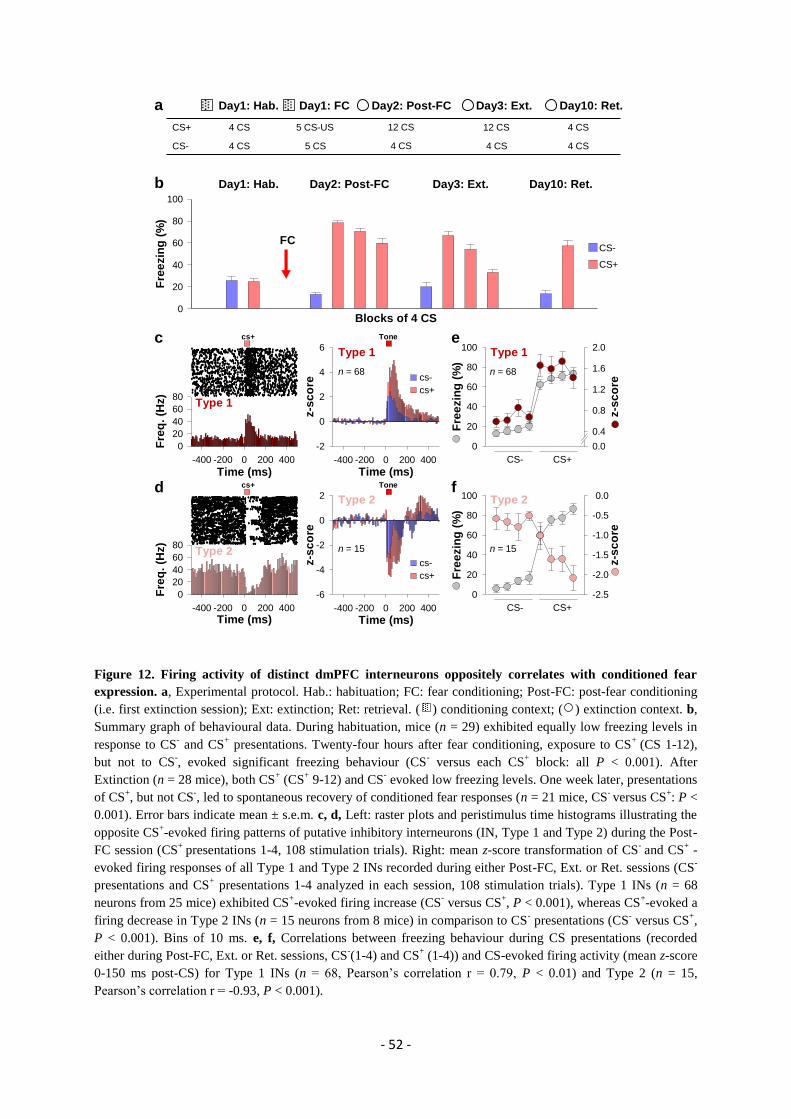
- 52 -
Figure 12. Firing activity of distinct dmPFC interneurons oppositely correlates with conditioned fear
expression. a, Experimental protocol. Hab.: habituation; FC: fear conditioning; Post-FC: post-fear conditioning
(i.e. first extinction session); Ext: extinction; Ret: retrieval. ( ) conditioning context; ( ) extinction context. b,
Summary graph of behavioural data. During habituation, mice (n = 29) exhibited equally low freezing levels in
response to CS- and CS
+ presentations. Twenty-four hours after fear conditioning, exposure to CS
+ (CS 1-12),
but not to CS-, evoked significant freezing behaviour (CS
- versus each CS
+ block: all P < 0.001). After
Extinction (n = 28 mice), both CS+ (CS
+ 9-12) and CS
- evoked low freezing levels. One week later, presentations
of CS+, but not CS
-, led to spontaneous recovery of conditioned fear responses (n = 21 mice, CS
- versus CS
+: P <
0.001). Error bars indicate mean ± s.e.m. c, d, Left: raster plots and peristimulus time histograms illustrating the
opposite CS+-evoked firing patterns of putative inhibitory interneurons (IN, Type 1 and Type 2) during the Post-
FC session (CS+
presentations 1-4, 108 stimulation trials). Right: mean z-score transformation of CS- and CS
+ -
evoked firing responses of all Type 1 and Type 2 INs recorded during either Post-FC, Ext. or Ret. sessions (CS-
presentations and CS+ presentations 1-4 analyzed in each session, 108 stimulation trials). Type 1 INs (n = 68
neurons from 25 mice) exhibited CS+-evoked firing increase (CS
- versus CS
+, P < 0.001), whereas CS
+-evoked a
firing decrease in Type 2 INs (n = 15 neurons from 8 mice) in comparison to CS- presentations (CS
- versus CS
+,
P < 0.001). Bins of 10 ms. e, f, Correlations between freezing behaviour during CS presentations (recorded
either during Post-FC, Ext. or Ret. sessions, CS-(1-4) and CS
+ (1-4)) and CS-evoked firing activity (mean z-score
0-150 ms post-CS) for Type 1 INs (n = 68, Pearson’s correlation r = 0.79, P < 0.01) and Type 2 (n = 15,
Pearson’s correlation r = -0.93, P < 0.001).
FC
b
c
Time (ms)-400 -200 0 200 400
Fre
q.
(Hz)
0
20
40
60
80
Time (ms)-400 -200 0 200 400
Fre
q.
(Hz)
0
20
40
60
80
Tone
Tone
Blocks of 4 CS
cs+
cs+
Fre
ezin
g (
%)
0
20
40
60
80
100
a
Day1: Hab. Day2: Post-FC Day3: Ext. Day10: Ret.
Time (ms)-400 -200 0 200 400
z-s
co
re
-6
-4
-2
0
2
Time (ms)-400 -200 0 200 400
z-s
co
re
-2
0
2
4
6
Fre
ezin
g (
%)
0
20
40
60
80
100
z-s
co
re
-2.5
-2.0
-1.5
-1.0
-0.5
0.0
Fre
ezin
g (
%)
0
20
40
60
80
100
z-s
co
re
0.0
0.4
0.8
1.2
1.6
2.0
Type 1
Type 2
e
d f
Day1: Hab. Day1: FC Day3: Ext.
4 CS 5 CS-US
4 CS5 CS4 CS
12 CS
Day2: Post-FC
4 CS
12 CSCS+
CS-
Day10: Ret.
4 CS
4 CS
CS-
CS+
cs-
cs+
cs-
cs+
CS- CS+
CS- CS+
n = 15
Type 2
n = 68
Type 1
n = 68
Type 1
n = 15
Type 2
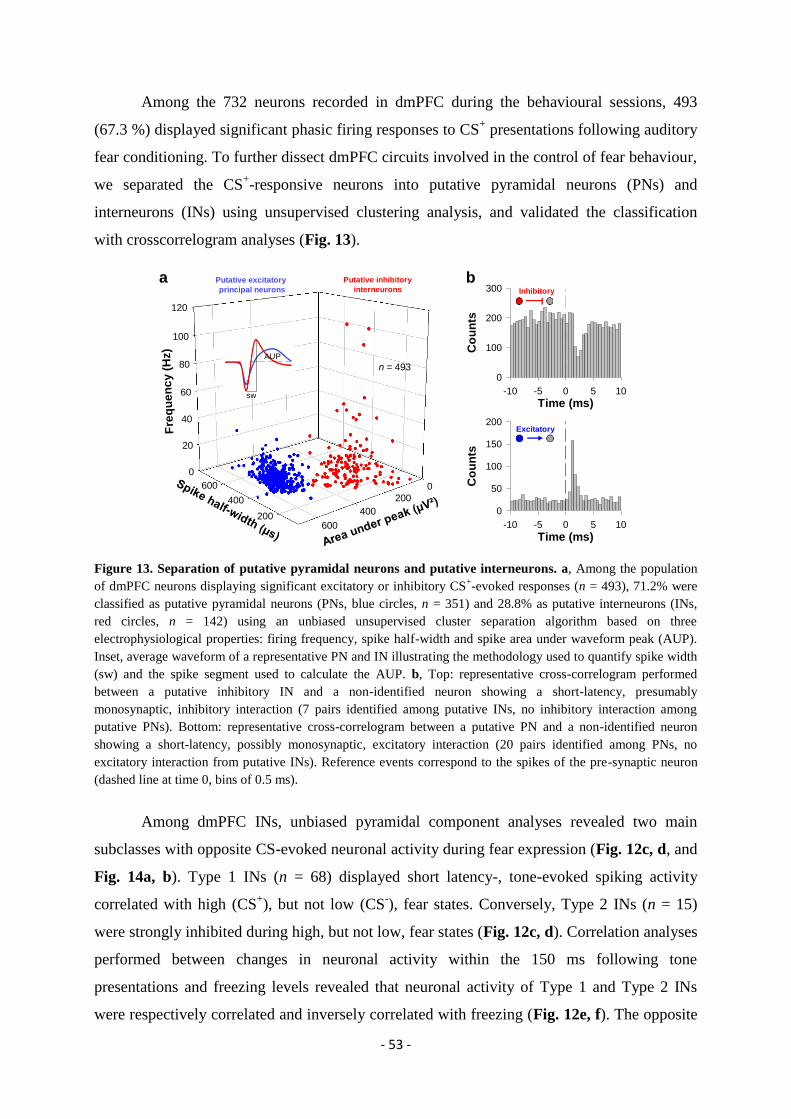
- 53 -
Among the 732 neurons recorded in dmPFC during the behavioural sessions, 493
(67.3 %) displayed significant phasic firing responses to CS+ presentations following auditory
fear conditioning. To further dissect dmPFC circuits involved in the control of fear behaviour,
we separated the CS+-responsive neurons into putative pyramidal neurons (PNs) and
interneurons (INs) using unsupervised clustering analysis, and validated the classification
with crosscorrelogram analyses (Fig. 13).
Figure 13. Separation of putative pyramidal neurons and putative interneurons. a, Among the population
of dmPFC neurons displaying significant excitatory or inhibitory CS+-evoked responses (n = 493), 71.2% were
classified as putative pyramidal neurons (PNs, blue circles, n = 351) and 28.8% as putative interneurons (INs,
red circles, n = 142) using an unbiased unsupervised cluster separation algorithm based on three
electrophysiological properties: firing frequency, spike half-width and spike area under waveform peak (AUP).
Inset, average waveform of a representative PN and IN illustrating the methodology used to quantify spike width
(sw) and the spike segment used to calculate the AUP. b, Top: representative cross-correlogram performed
between a putative inhibitory IN and a non-identified neuron showing a short-latency, presumably
monosynaptic, inhibitory interaction (7 pairs identified among putative INs, no inhibitory interaction among
putative PNs). Bottom: representative cross-correlogram between a putative PN and a non-identified neuron
showing a short-latency, possibly monosynaptic, excitatory interaction (20 pairs identified among PNs, no
excitatory interaction from putative INs). Reference events correspond to the spikes of the pre-synaptic neuron
(dashed line at time 0, bins of 0.5 ms).
Among dmPFC INs, unbiased pyramidal component analyses revealed two main
subclasses with opposite CS-evoked neuronal activity during fear expression (Fig. 12c, d, and
Fig. 14a, b). Type 1 INs (n = 68) displayed short latency-, tone-evoked spiking activity
correlated with high (CS+), but not low (CS
-), fear states. Conversely, Type 2 INs (n = 15)
were strongly inhibited during high, but not low, fear states (Fig. 12c, d). Correlation analyses
performed between changes in neuronal activity within the 150 ms following tone
presentations and freezing levels revealed that neuronal activity of Type 1 and Type 2 INs
were respectively correlated and inversely correlated with freezing (Fig. 12e, f). The opposite
0
20
40
60
80
100
120
200
400
600 0
200
400
600
Fre
qu
en
cy (
Hz)
Spike half-width (µs)Area under peak (µV²)
a
Time (ms)-10 -5 0 5 10
Co
un
ts
0
50
100
150
200
Time (ms)-10 -5 0 5 10
Co
un
ts
0
100
200
300b
Inhibitory
Excitatory
n = 493
Putative excitatory
principal neurons
Putative inhibitory
interneurons
sw
AUP
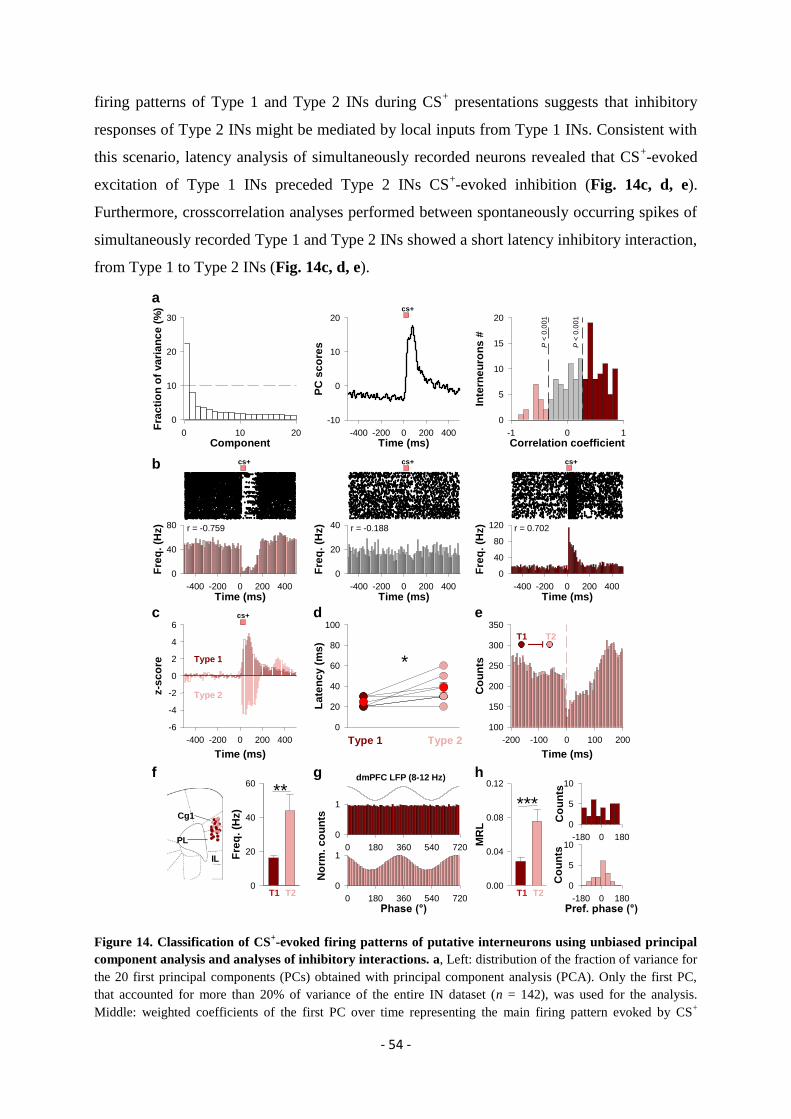
- 54 -
firing patterns of Type 1 and Type 2 INs during CS+ presentations suggests that inhibitory
responses of Type 2 INs might be mediated by local inputs from Type 1 INs. Consistent with
this scenario, latency analysis of simultaneously recorded neurons revealed that CS+-evoked
excitation of Type 1 INs preceded Type 2 INs CS+-evoked inhibition (Fig. 14c, d, e).
Furthermore, crosscorrelation analyses performed between spontaneously occurring spikes of
simultaneously recorded Type 1 and Type 2 INs showed a short latency inhibitory interaction,
from Type 1 to Type 2 INs (Fig. 14c, d, e).
Figure 14. Classification of CS+-evoked firing patterns of putative interneurons using unbiased principal
component analysis and analyses of inhibitory interactions. a, Left: distribution of the fraction of variance for
the 20 first principal components (PCs) obtained with principal component analysis (PCA). Only the first PC,
that accounted for more than 20% of variance of the entire IN dataset (n = 142), was used for the analysis.
Middle: weighted coefficients of the first PC over time representing the main firing pattern evoked by CS+
Time (ms)-400 -200 0 200 400
PC
sc
ore
s
-10
0
10
20
a
b
Component0 10 20
Fra
cti
on
of
va
ria
nc
e (
%)
0
10
20
30
Time (ms)-400 -200 0 200 400
Fre
q.
(Hz)
0
40
80
Time (ms)-400 -200 0 200 400
Fre
q.
(Hz)
0
20
40 r = -0.188r = -0.759
cs+cs+
cs+
Time (ms)
-400 -200 0 200 400
z-s
co
re
-6
-4
-2
0
2
4
6
*Type 1
cs+
La
ten
cy (
ms
)
0
20
40
60
80
100
Type 2Type 1
Type 2
c d e
Correlation coefficient-1 0 1
Inte
rne
uro
ns
#
0
5
10
15
20
Time (ms)-400 -200 0 200 400
Fre
q.
(Hz)
0
40
80
120 r = 0.702
cs+
Time (ms)
-200 -100 0 100 200
Co
un
ts
100
150
200
250
300
350
T2T1
P <
0.0
01
P <
0.0
01
0 180 360 540 720
0
1
No
rm.
co
un
ts
dmPFC LFP (8-12 Hz)
Fre
q.
(Hz)
0
20
40
60
T1 T2
**gf h
Cg1
PL
IL
Pref. phase (°)-180 0 180
Co
un
ts
0
5
10
***
T1 T2
Phase (°)0 180 360 540 720
0
1
MR
L
0.00
0.04
0.08
0.12
-180 0 180
Co
un
ts
0
5
10

- 55 -
presentations (CS+ onset at time 0) of the entire IN dataset. Right: distribution of prefrontal INs relative to their
correlation coefficients with the first PC. The dashed lines indicate the levels of significance for positive and
negative correlations (P < 0.001). Among the 142 INs, 83 (58.5 %) displayed a significant positive (n = 68, 48
%, dark red bars) or negative (n = 15, 10.6%, light red bars) correlation with the first PC, whereas 41.5 % INs (n
= 59, grey bars) were not correlated with the first PC. b, Raster plots and PSTH examples of individual INs
whose firing negatively correlated (left, Type 2 IN), not correlated (middle) or positively correlated (right, Type
1 IN) with the first PC. Type 1 INs were excited, whereas Type 2 INs were inhibited by CS+ presentations. Bins
of 10 ms. c, Peristimulus time histogram of all Type 1 (n = 68) and Type 2 (n = 15) INs illustrating the selective
CS+-evoked responses (either Post-FC, Ext. or Ret. Sessions, CS
+ presentations 1-4). CS
+-evoked neuronal
activation in Type 1 INs temporally matched CS+-evoked inhibition of Type 2 INs. Bins of 10 ms. d, Individual
(Type 1 INs, dark red dots; Type 2 INs, light red dots) and averaged (red dots) latencies of the first significant
time bin (z-score < -1.65 or > +1.65) following CS+ presentations for Type 1 and Type 2 INs recorded
simultaneously (n = 7 pairs recorded in 5 mice). CS+-evoked excitation in Type 1 INs preceded CS
+-evoked
inhibition in Type 2 INs (mean latency: Type 1: 24.3 ± 2 ms; Type 2: 38.6 ± 4.6 ms; paired t test, * P < 0.05).
Error bars indicate mean ± s.e.m. e, Cross-correlation analysis performed between a Type 1 and a Type 2 INs
recorded simultaneously outside CS presentations. The cross-correlogram shows a short latency, potentially
monosynaptic, inhibitory interaction. Reference event corresponds to the spikes of the Type 1 IN (dashed line at
time 0). Bins of 5 ms.
Interestingly, whereas Type 1 INs displayed moderate firing rates (mean firing rate:
16.2 ± 1.5 Hz) and were not modulated with local theta oscillations, Type 2 INs showed fast
firing activity (mean firing rate: 43.9 ± 9.7 Hz) and were strongly modulated with local theta
oscillations, suggesting that Type 2 INs are PV-expressing interneurons (PV INs, Fig. 15a, b,
c). To further address this possibility, we selectively infected PV INs with targeted injections
of a conditional adeno-associated virus (AAV) encoding for archeorhodopsin (ArchT) in the
dmPFC of PV-IRES-Cre mice (Fig. 15d). Using this strategy, we were able to optically
silence the neuronal firing of Type 2 (n = 5/5 identified Type 2 INs) but not Type 1 INs (n =
0/9 identified Type 1 INs), indicating that Type 2 INs indeed belonged to the PV INs subclass
(Fig. 15e). Remarkably, among light reactive PV INs (n = 9), only Type 2 PV INs (n = 5)
displayed significant decreases in tone-evoked activity following fear conditioning,
suggesting a functional role of this subpopulation of PV INs during fear behaviour (Fig. 16e,
f). In summary, we identified two functional subclasses of INs in the dmPFC whose activities
oppositely correlate with fear behaviour. Furthermore, we demonstrated that Type 2 INs are
PV INs.
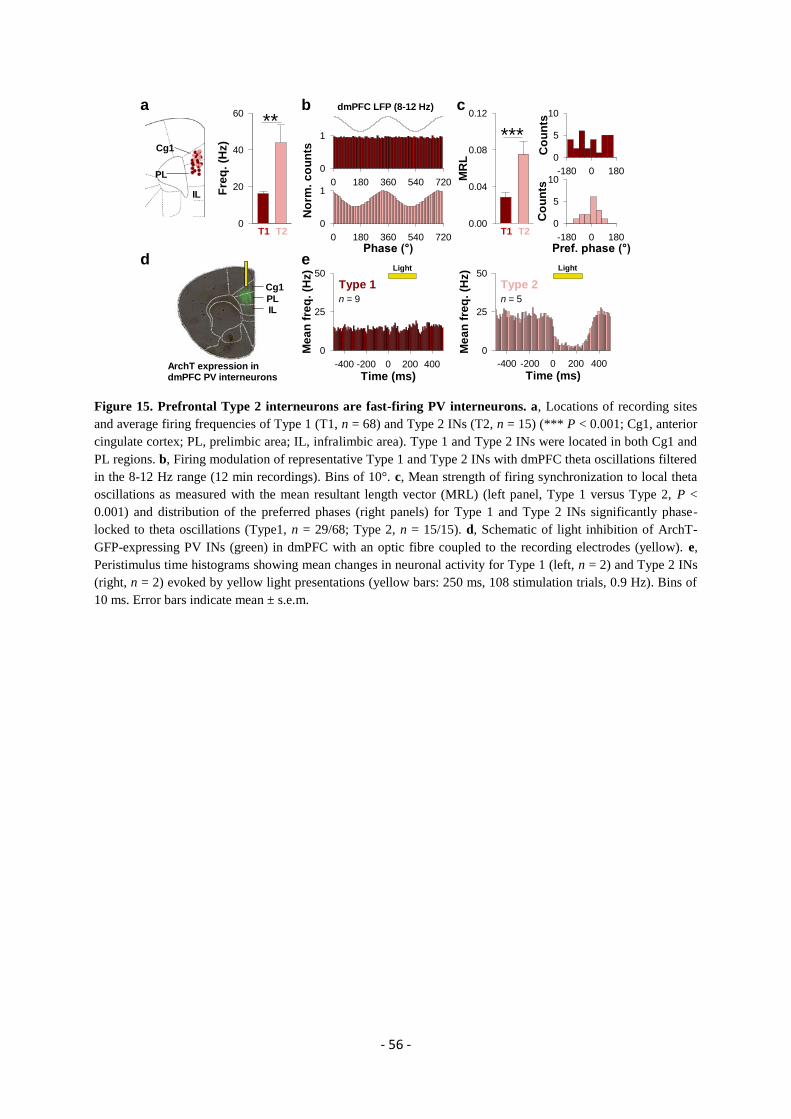
- 56 -
Figure 15. Prefrontal Type 2 interneurons are fast-firing PV interneurons. a, Locations of recording sites
and average firing frequencies of Type 1 (T1, n = 68) and Type 2 INs (T2, n = 15) (*** P < 0.001; Cg1, anterior
cingulate cortex; PL, prelimbic area; IL, infralimbic area). Type 1 and Type 2 INs were located in both Cg1 and
PL regions. b, Firing modulation of representative Type 1 and Type 2 INs with dmPFC theta oscillations filtered
in the 8-12 Hz range (12 min recordings). Bins of 10°. c, Mean strength of firing synchronization to local theta
oscillations as measured with the mean resultant length vector (MRL) (left panel, Type 1 versus Type 2, P <
0.001) and distribution of the preferred phases (right panels) for Type 1 and Type 2 INs significantly phase-
locked to theta oscillations (Type1, n = 29/68; Type 2, n = 15/15). d, Schematic of light inhibition of ArchT-
GFP-expressing PV INs (green) in dmPFC with an optic fibre coupled to the recording electrodes (yellow). e,
Peristimulus time histograms showing mean changes in neuronal activity for Type 1 (left, n = 2) and Type 2 INs
(right, n = 2) evoked by yellow light presentations (yellow bars: 250 ms, 108 stimulation trials, 0.9 Hz). Bins of
10 ms. Error bars indicate mean ± s.e.m.
***
b
0 180 360 540 720
0
1
Phase (°)0 180 360 540 720
0
1
MR
L
0.00
0.04
0.08
0.12a
T1 T2
-180 0 180
Co
un
ts
0
5
10
Pref. phase (°)-180 0 180
Co
un
ts
0
5
10
No
rm.
co
un
ts
dmPFC LFP (8-12 Hz)
Time (ms)-400 -200 0 200 400
Me
an
fre
q.
(Hz)
0
25
50
Time (ms)-400 -200 0 200 400
Me
an
fre
q.
(Hz)
0
25
50
Type 1 Type 2
n = 9 n = 5
c
d e
ArchT expression indmPFC PV interneurons
Cg1
PLIL
Light Light
Fre
q.
(Hz)
0
20
40
60
T1 T2
Cg1
PL
IL
**
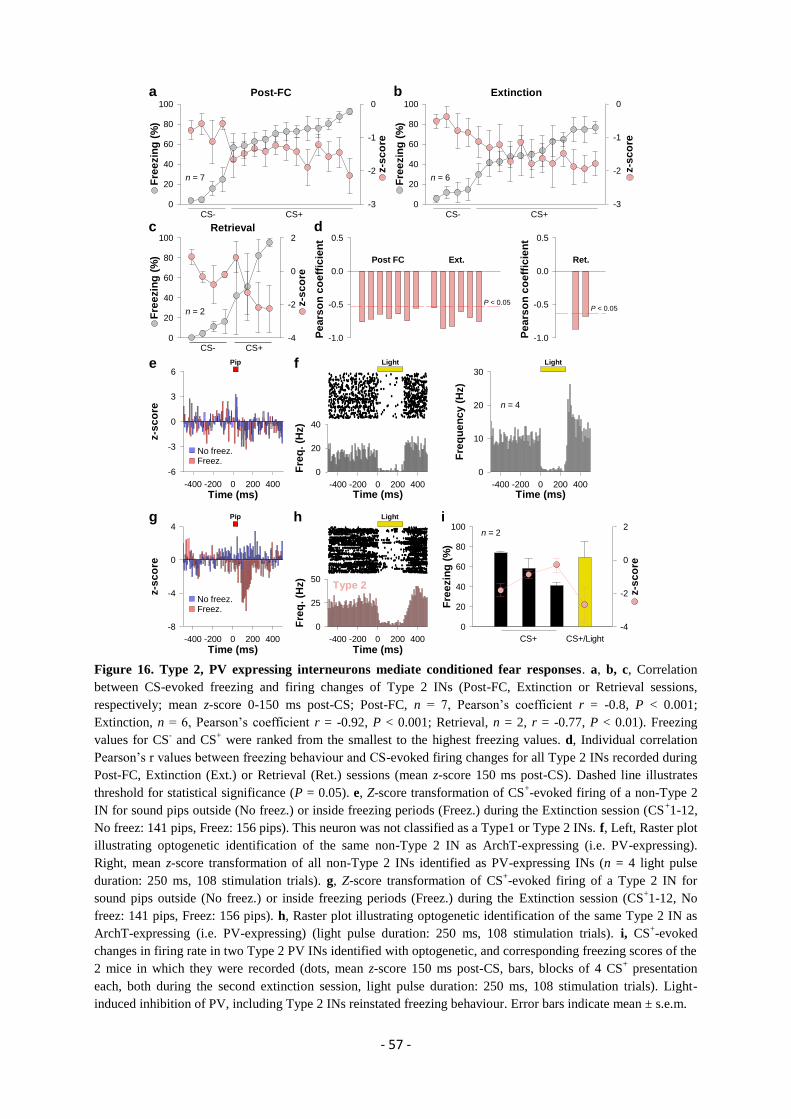
- 57 -
Figure 16. Type 2, PV expressing interneurons mediate conditioned fear responses. a, b, c, Correlation
between CS-evoked freezing and firing changes of Type 2 INs (Post-FC, Extinction or Retrieval sessions,
respectively; mean z-score 0-150 ms post-CS; Post-FC, n = 7, Pearson’s coefficient r = -0.8, P < 0.001;
Extinction, n = 6, Pearson’s coefficient r = -0.92, P < 0.001; Retrieval, n = 2, r = -0.77, P < 0.01). Freezing
values for CS- and CS
+ were ranked from the smallest to the highest freezing values. d, Individual correlation
Pearson’s r values between freezing behaviour and CS-evoked firing changes for all Type 2 INs recorded during
Post-FC, Extinction (Ext.) or Retrieval (Ret.) sessions (mean z-score 150 ms post-CS). Dashed line illustrates
threshold for statistical significance (P = 0.05). e, Z-score transformation of CS+-evoked firing of a non-Type 2
IN for sound pips outside (No freez.) or inside freezing periods (Freez.) during the Extinction session (CS+1-12,
No freez: 141 pips, Freez: 156 pips). This neuron was not classified as a Type1 or Type 2 INs. f, Left, Raster plot
illustrating optogenetic identification of the same non-Type 2 IN as ArchT-expressing (i.e. PV-expressing).
Right, mean z-score transformation of all non-Type 2 INs identified as PV-expressing INs (n = 4 light pulse
duration: 250 ms, 108 stimulation trials). g, Z-score transformation of CS+-evoked firing of a Type 2 IN for
sound pips outside (No freez.) or inside freezing periods (Freez.) during the Extinction session (CS+1-12, No
freez: 141 pips, Freez: 156 pips). h, Raster plot illustrating optogenetic identification of the same Type 2 IN as
ArchT-expressing (i.e. PV-expressing) (light pulse duration: 250 ms, 108 stimulation trials). i, CS+-evoked
changes in firing rate in two Type 2 PV INs identified with optogenetic, and corresponding freezing scores of the
2 mice in which they were recorded (dots, mean z-score 150 ms post-CS, bars, blocks of 4 CS+ presentation
each, both during the second extinction session, light pulse duration: 250 ms, 108 stimulation trials). Light-
induced inhibition of PV, including Type 2 INs reinstated freezing behaviour. Error bars indicate mean ± s.e.m.
z-s
co
re
-3
-2
-1
0
Fre
ezin
g (
%)
0
20
40
60
80
100
Fre
ezin
g (
%)
0
20
40
60
80
100
z-s
co
re
-3
-2
-1
0
Fre
ezin
g (
%)
0
20
40
60
80
100
CS- CS-CS+ CS+
Post-FC Extinction
P < 0.05z-s
co
re
-4
-2
0
2
Pe
ars
on
co
eff
icie
nt
-1.0
-0.5
0.0
0.5
Pe
ars
on
co
eff
icie
nt
-1.0
-0.5
0.0
0.5
Post FC Ext. Ret.
P < 0.05
CS- CS+
Retrieval
a
c d
n = 7 n = 6
n = 2
b
e f
Time (ms)-400 -200 0 200 400
Fre
q. (H
z)
0
20
40
Light
Time (ms)-400 -200 0 200 400
z-s
co
re
-6
-3
0
3
6
Time (ms)-400 -200 0 200 400
Fre
qu
en
cy (
Hz)
0
10
20
30
n = 4
No freez.Freez.
Pip Light
Time (ms)-400 -200 0 200 400
z-s
co
re
-8
-4
0
4
Fre
ezin
g (
%)
0
20
40
60
80
100
z-s
co
re-4
-2
0
2
Type 2
Time (ms)-400 -200 0 200 400
Fre
q. (H
z)
0
25
50
n = 2
CS+ CS+/Light
Lighth ig Pip
No freez.Freez.
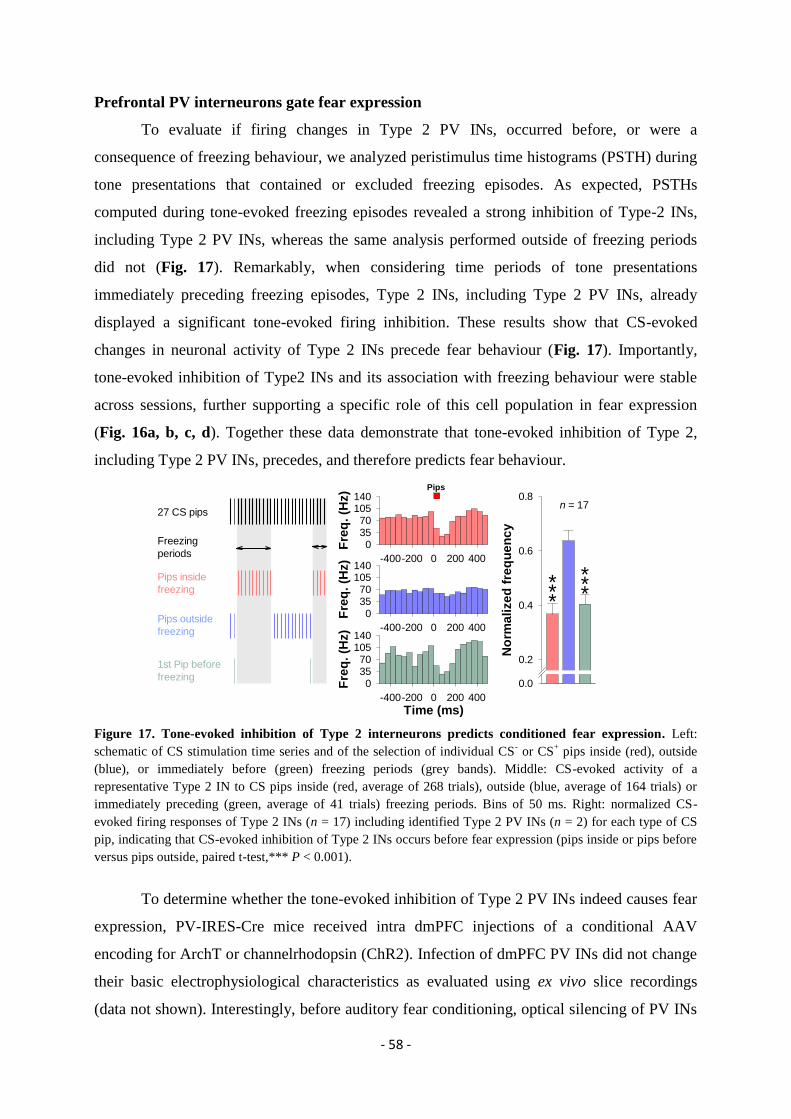
- 58 -
Prefrontal PV interneurons gate fear expression
To evaluate if firing changes in Type 2 PV INs, occurred before, or were a
consequence of freezing behaviour, we analyzed peristimulus time histograms (PSTH) during
tone presentations that contained or excluded freezing episodes. As expected, PSTHs
computed during tone-evoked freezing episodes revealed a strong inhibition of Type-2 INs,
including Type 2 PV INs, whereas the same analysis performed outside of freezing periods
did not (Fig. 17). Remarkably, when considering time periods of tone presentations
immediately preceding freezing episodes, Type 2 INs, including Type 2 PV INs, already
displayed a significant tone-evoked firing inhibition. These results show that CS-evoked
changes in neuronal activity of Type 2 INs precede fear behaviour (Fig. 17). Importantly,
tone-evoked inhibition of Type2 INs and its association with freezing behaviour were stable
across sessions, further supporting a specific role of this cell population in fear expression
(Fig. 16a, b, c, d). Together these data demonstrate that tone-evoked inhibition of Type 2,
including Type 2 PV INs, precedes, and therefore predicts fear behaviour.
Figure 17. Tone-evoked inhibition of Type 2 interneurons predicts conditioned fear expression. Left:
schematic of CS stimulation time series and of the selection of individual CS- or CS
+ pips inside (red), outside
(blue), or immediately before (green) freezing periods (grey bands). Middle: CS-evoked activity of a
representative Type 2 IN to CS pips inside (red, average of 268 trials), outside (blue, average of 164 trials) or
immediately preceding (green, average of 41 trials) freezing periods. Bins of 50 ms. Right: normalized CS-
evoked firing responses of Type 2 INs (n = 17) including identified Type 2 PV INs (n = 2) for each type of CS
pip, indicating that CS-evoked inhibition of Type 2 INs occurs before fear expression (pips inside or pips before
versus pips outside, paired t-test,*** P < 0.001).
To determine whether the tone-evoked inhibition of Type 2 PV INs indeed causes fear
expression, PV-IRES-Cre mice received intra dmPFC injections of a conditional AAV
encoding for ArchT or channelrhodopsin (ChR2). Infection of dmPFC PV INs did not change
their basic electrophysiological characteristics as evaluated using ex vivo slice recordings
(data not shown). Interestingly, before auditory fear conditioning, optical silencing of PV INs
27 CS pips
Freezing
periods
Pips inside
freezing
Pips outside
freezing
1st Pip before
freezing
No
rma
lize
d f
req
ue
nc
y
0.0
0.2
0.4
0.6
0.8n = 17
-400-200 0 200 400
Fre
q.
(Hz)
03570
105140
Time (ms)-400-200 0 200 400
Fre
q.
(Hz)
03570
105140
Pips
-400-200 0 200 400
Fre
q.
(Hz)
03570
105140
***
***
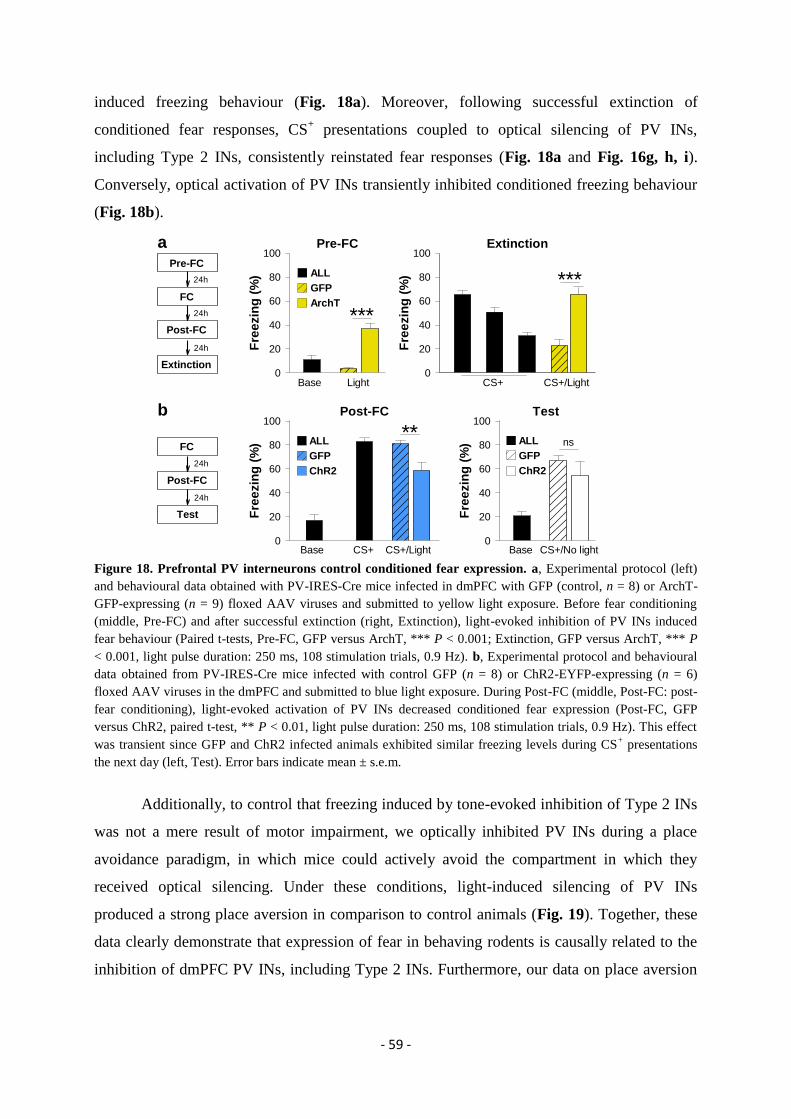
- 59 -
induced freezing behaviour (Fig. 18a). Moreover, following successful extinction of
conditioned fear responses, CS+ presentations coupled to optical silencing of PV INs,
including Type 2 INs, consistently reinstated fear responses (Fig. 18a and Fig. 16g, h, i).
Conversely, optical activation of PV INs transiently inhibited conditioned freezing behaviour
(Fig. 18b).
Figure 18. Prefrontal PV interneurons control conditioned fear expression. a, Experimental protocol (left)
and behavioural data obtained with PV-IRES-Cre mice infected in dmPFC with GFP (control, n = 8) or ArchT-
GFP-expressing (n = 9) floxed AAV viruses and submitted to yellow light exposure. Before fear conditioning
(middle, Pre-FC) and after successful extinction (right, Extinction), light-evoked inhibition of PV INs induced
fear behaviour (Paired t-tests, Pre-FC, GFP versus ArchT, *** P < 0.001; Extinction, GFP versus ArchT, *** P
< 0.001, light pulse duration: 250 ms, 108 stimulation trials, 0.9 Hz). b, Experimental protocol and behavioural
data obtained from PV-IRES-Cre mice infected with control GFP (n = 8) or ChR2-EYFP-expressing (n = 6)
floxed AAV viruses in the dmPFC and submitted to blue light exposure. During Post-FC (middle, Post-FC: post-
fear conditioning), light-evoked activation of PV INs decreased conditioned fear expression (Post-FC, GFP
versus ChR2, paired t-test, ** P < 0.01, light pulse duration: 250 ms, 108 stimulation trials, 0.9 Hz). This effect
was transient since GFP and ChR2 infected animals exhibited similar freezing levels during CS+ presentations
the next day (left, Test). Error bars indicate mean ± s.e.m.
Additionally, to control that freezing induced by tone-evoked inhibition of Type 2 INs
was not a mere result of motor impairment, we optically inhibited PV INs during a place
avoidance paradigm, in which mice could actively avoid the compartment in which they
received optical silencing. Under these conditions, light-induced silencing of PV INs
produced a strong place aversion in comparison to control animals (Fig. 19). Together, these
data clearly demonstrate that expression of fear in behaving rodents is causally related to the
inhibition of dmPFC PV INs, including Type 2 INs. Furthermore, our data on place aversion
***
***
a
b
Fre
ezin
g (
%)
0
20
40
60
80
100
Base Light CS+
Pre-FC
Fre
ezin
g (
%)
0
20
40
60
80
100Extinction
CS+/Light
GFP
ArchT
ALL
**
Fre
ezin
g (
%)
0
20
40
60
80
100
Fre
ezin
g (
%)
0
20
40
60
80
100
Base CS+
ns
Post-FC Test
Base CS+/No lightCS+/Light
24h
24h
24h
Pre-FC
FC
Post-FC
Extinction
24h
24h
FC
Post-FC
Test
GFP
ChR2
ALL
GFP
ChR2
ALL

- 60 -
also indicate that the functional inhibition of dmPFC PV INs might also control the formation
and not only the expression of aversive behaviours.
Figure 19. Optogenetic inhibition of prefrontal PV interneurons induces place aversion. a, On day 1, GFP
(n = 11) and ArchT (n = 13) infected mice were exposed to a two-compartment place aversion apparatus during
15 min. Following pre-exposure, the most visited compartment was selected for each animal. On day 2,
systematic yellow light-induced inhibition of PV INs was triggered in the most visited compartment (and no
light stimulation in the less visited compartment) during a 15 min exposure session. On day 3, GFP (n = 6) and
ArchT (n = 6) infected mice were re-exposed to the place aversion apparatus during 15 min to evaluate the long-
term effect of yellow light stimulation on day 2. b, Percentage of time spent in the most and less visited
compartments on day 1 for individual infected mice (GFP (n = 11) and ArchT (n = 13)). c, Average percentage
of time spent in the most visited compartment on day 1,2 and 3 for GFP and ArchT infected mice. On day 2,
light inhibition of PV INs induced an avoidance of the most visited compartment for ArchT infected animals in
comparison to day 1 (ArchT mice: day 1 versus day 2, paired t-test, ** P < 0.01) and to GFP controls on day 2
(day 2: ArchT versus GFP, unpaired t-test, * P < 0.05; 250 ms pulses delivered at 0.9 Hz). On day 3, ArchT mice
did not avoid any more the most visited compartment (ArchT mice: day 2 versus day 3, unpaired t-test, * P <
0.05). Error bars indicate mean ± s.e.m.
What could be the functional consequence of CS+-evoked inhibition of Type 2 PV
INs, at the level of prefrontal pyramidal neurons? PV INs, target the perisomatic region of
PNs thereby providing powerful inhibition of prefrontal output neuronal activity (Freund &
Katona, 2007). Therefore, CS+-evoked inhibition of PV INs during fear behaviour might
disinhibit PNs, a permissive mechanism that would gate conditioned fear responses.
Consistent with this possibility, the vast majority of tone-reactive PNs (n = 308/351, 87.7%)
displayed a significant increase in neuronal activity upon CS+ presentations. This effect was
greater during CS+ in comparison with CS
- presentations (Fig. 20a). To further test for a
disinhibitory mechanism, we optogenetically manipulated PV INs while simultaneously
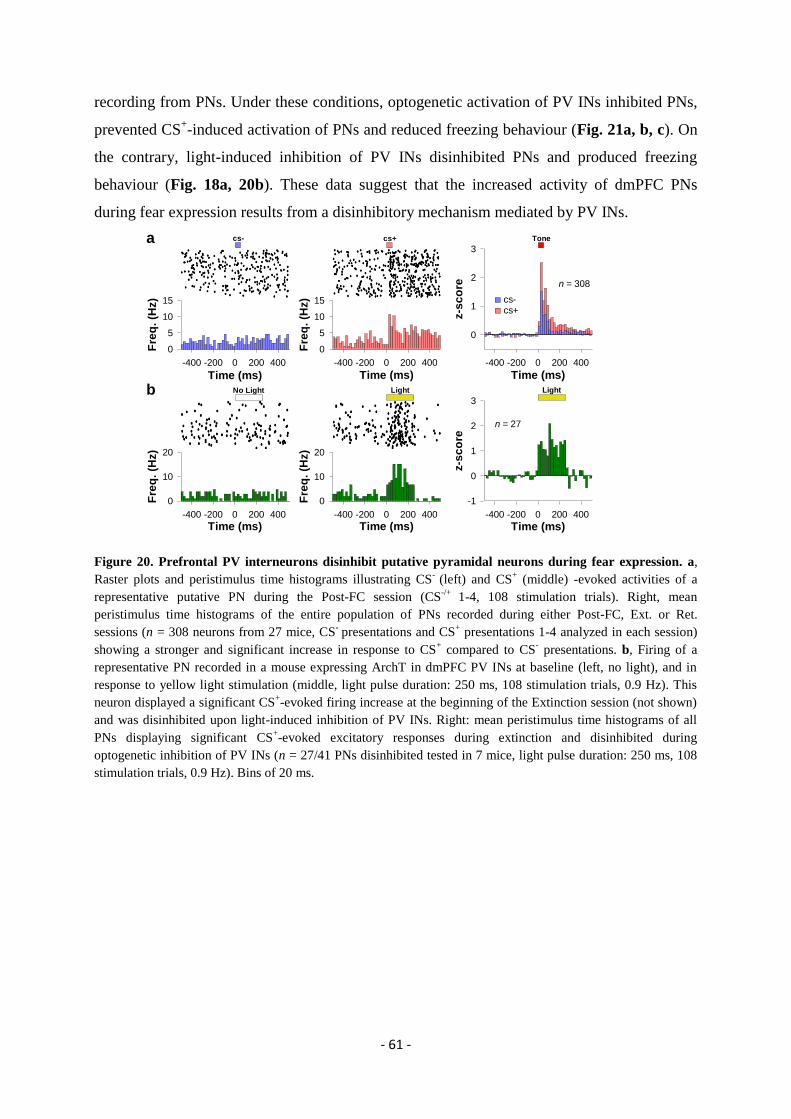
- 61 -
recording from PNs. Under these conditions, optogenetic activation of PV INs inhibited PNs,
prevented CS+-induced activation of PNs and reduced freezing behaviour (Fig. 21a, b, c). On
the contrary, light-induced inhibition of PV INs disinhibited PNs and produced freezing
behaviour (Fig. 18a, 20b). These data suggest that the increased activity of dmPFC PNs
during fear expression results from a disinhibitory mechanism mediated by PV INs.
Figure 20. Prefrontal PV interneurons disinhibit putative pyramidal neurons during fear expression. a,
Raster plots and peristimulus time histograms illustrating CS-
(left) and CS+ (middle) -evoked activities of a
representative putative PN during the Post-FC session (CS-/+
1-4, 108 stimulation trials). Right, mean
peristimulus time histograms of the entire population of PNs recorded during either Post-FC, Ext. or Ret.
sessions (n = 308 neurons from 27 mice, CS- presentations and CS
+ presentations 1-4 analyzed in each session)
showing a stronger and significant increase in response to CS+ compared to CS
- presentations. b, Firing of a
representative PN recorded in a mouse expressing ArchT in dmPFC PV INs at baseline (left, no light), and in
response to yellow light stimulation (middle, light pulse duration: 250 ms, 108 stimulation trials, 0.9 Hz). This
neuron displayed a significant CS+-evoked firing increase at the beginning of the Extinction session (not shown)
and was disinhibited upon light-induced inhibition of PV INs. Right: mean peristimulus time histograms of all
PNs displaying significant CS+-evoked excitatory responses during extinction and disinhibited during
optogenetic inhibition of PV INs (n = 27/41 PNs disinhibited tested in 7 mice, light pulse duration: 250 ms, 108
stimulation trials, 0.9 Hz). Bins of 20 ms.
Time (ms)-400 -200 0 200 400
Fre
q.
(Hz)
0
5
10
15
Time (ms)-400 -200 0 200 400
Fre
q.
(Hz)
0
5
10
15
Time (ms)-400 -200 0 200 400
z-s
co
re
0
1
2
3
Courtin et al., Figure 4
a
b
cs-cs+
n = 308
cs+cs- Tone
Time (ms)-400 -200 0 200 400
z-s
co
re
-1
0
1
2
3
Time (ms)-400 -200 0 200 400
Fre
q.
(Hz)
0
10
20
n = 27
Time (ms)-400 -200 0 200 400
Fre
q.
(Hz)
0
10
20
No Light Light Light
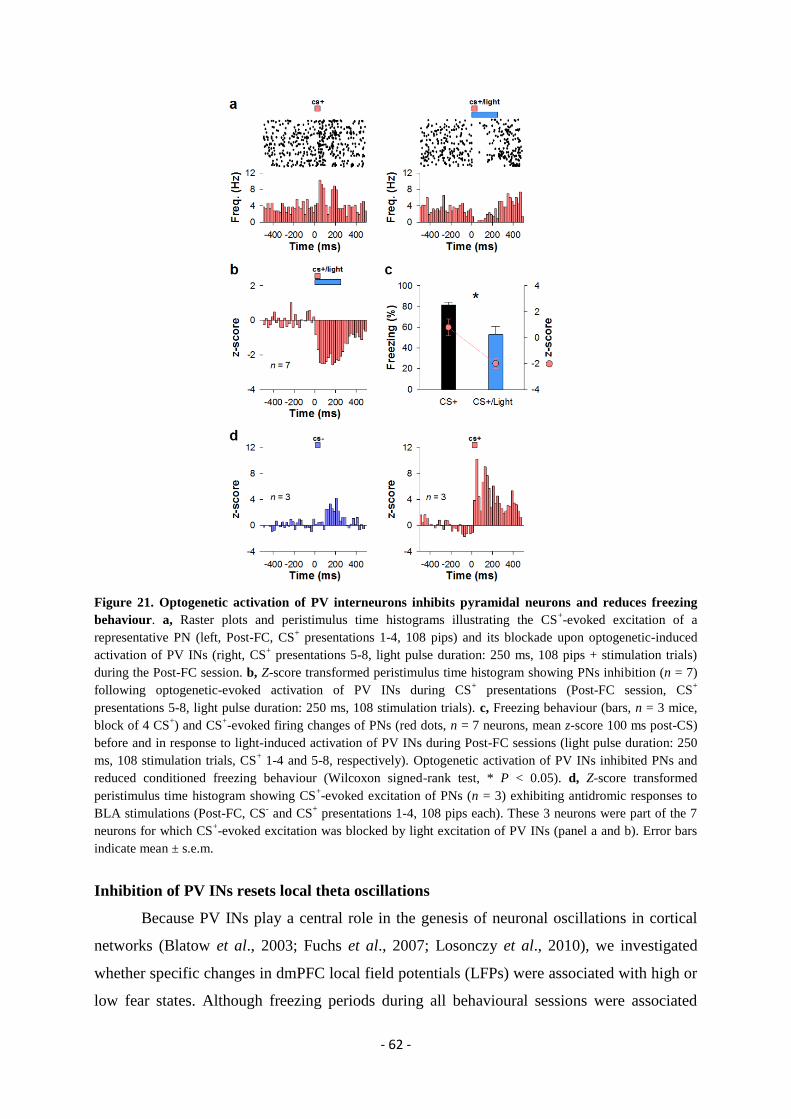
- 62 -
Figure 21. Optogenetic activation of PV interneurons inhibits pyramidal neurons and reduces freezing
behaviour. a, Raster plots and peristimulus time histograms illustrating the CS+-evoked excitation of a
representative PN (left, Post-FC, CS+ presentations 1-4, 108 pips) and its blockade upon optogenetic-induced
activation of PV INs (right, CS+ presentations 5-8, light pulse duration: 250 ms, 108 pips + stimulation trials)
during the Post-FC session. b, Z-score transformed peristimulus time histogram showing PNs inhibition (n = 7)
following optogenetic-evoked activation of PV INs during CS+ presentations (Post-FC session, CS
+
presentations 5-8, light pulse duration: 250 ms, 108 stimulation trials). c, Freezing behaviour (bars, n = 3 mice,
block of 4 CS+) and CS
+-evoked firing changes of PNs (red dots, n = 7 neurons, mean z-score 100 ms post-CS)
before and in response to light-induced activation of PV INs during Post-FC sessions (light pulse duration: 250
ms, 108 stimulation trials, CS+ 1-4 and 5-8, respectively). Optogenetic activation of PV INs inhibited PNs and
reduced conditioned freezing behaviour (Wilcoxon signed-rank test, * P < 0.05). d, Z-score transformed
peristimulus time histogram showing CS+-evoked excitation of PNs (n = 3) exhibiting antidromic responses to
BLA stimulations (Post-FC, CS- and CS
+ presentations 1-4, 108 pips each). These 3 neurons were part of the 7
neurons for which CS+-evoked excitation was blocked by light excitation of PV INs (panel a and b). Error bars
indicate mean ± s.e.m.
Inhibition of PV INs resets local theta oscillations
Because PV INs play a central role in the genesis of neuronal oscillations in cortical
networks (Blatow et al., 2003; Fuchs et al., 2007; Losonczy et al., 2010), we investigated
whether specific changes in dmPFC local field potentials (LFPs) were associated with high or
low fear states. Although freezing periods during all behavioural sessions were associated
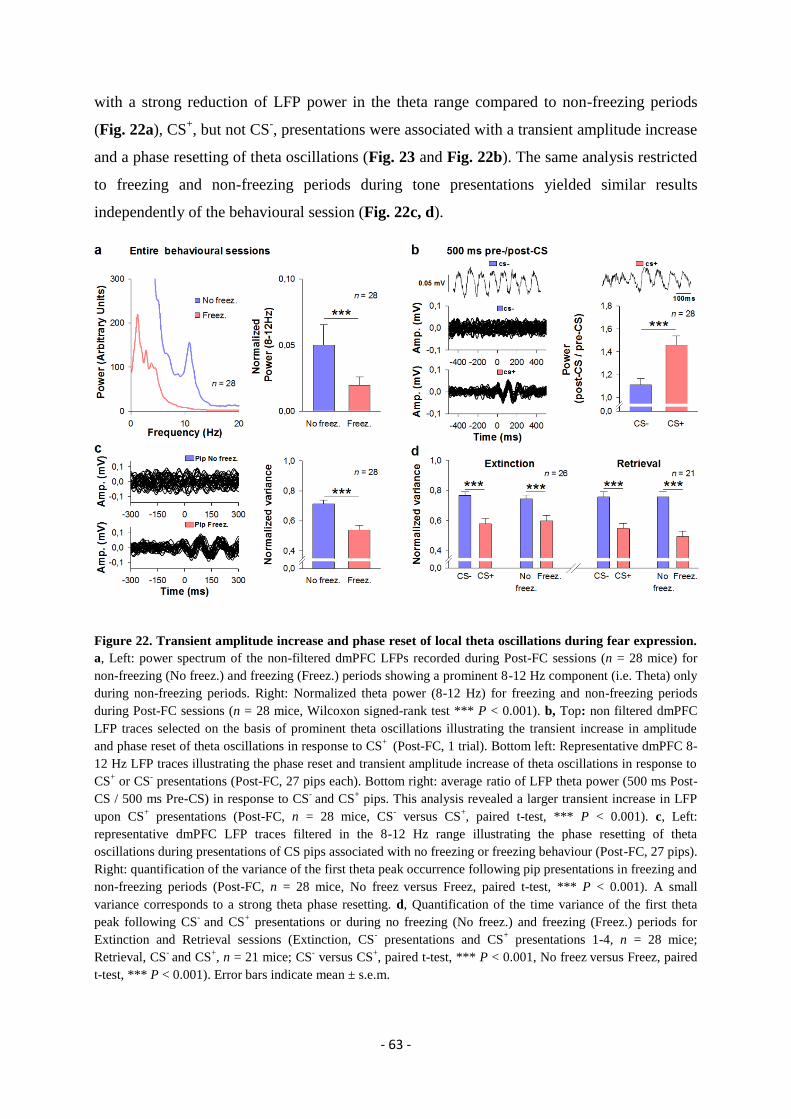
- 63 -
with a strong reduction of LFP power in the theta range compared to non-freezing periods
(Fig. 22a), CS+, but not CS
-, presentations were associated with a transient amplitude increase
and a phase resetting of theta oscillations (Fig. 23 and Fig. 22b). The same analysis restricted
to freezing and non-freezing periods during tone presentations yielded similar results
independently of the behavioural session (Fig. 22c, d).
Figure 22. Transient amplitude increase and phase reset of local theta oscillations during fear expression.
a, Left: power spectrum of the non-filtered dmPFC LFPs recorded during Post-FC sessions (n = 28 mice) for
non-freezing (No freez.) and freezing (Freez.) periods showing a prominent 8-12 Hz component (i.e. Theta) only
during non-freezing periods. Right: Normalized theta power (8-12 Hz) for freezing and non-freezing periods
during Post-FC sessions (n = 28 mice, Wilcoxon signed-rank test *** P < 0.001). b, Top: non filtered dmPFC
LFP traces selected on the basis of prominent theta oscillations illustrating the transient increase in amplitude
and phase reset of theta oscillations in response to CS+
(Post-FC, 1 trial). Bottom left: Representative dmPFC 8-
12 Hz LFP traces illustrating the phase reset and transient amplitude increase of theta oscillations in response to
CS+ or CS
- presentations (Post-FC, 27 pips each). Bottom right: average ratio of LFP theta power (500 ms Post-
CS / 500 ms Pre-CS) in response to CS- and CS
+ pips. This analysis revealed a larger transient increase in LFP
upon CS+ presentations (Post-FC, n = 28 mice, CS
- versus CS
+, paired t-test, *** P < 0.001). c, Left:
representative dmPFC LFP traces filtered in the 8-12 Hz range illustrating the phase resetting of theta
oscillations during presentations of CS pips associated with no freezing or freezing behaviour (Post-FC, 27 pips).
Right: quantification of the variance of the first theta peak occurrence following pip presentations in freezing and
non-freezing periods (Post-FC, n = 28 mice, No freez versus Freez, paired t-test, *** P < 0.001). A small
variance corresponds to a strong theta phase resetting. d, Quantification of the time variance of the first theta
peak following CS-
and CS+ presentations or during no freezing (No freez.) and freezing (Freez.) periods for
Extinction and Retrieval sessions (Extinction, CS- presentations and CS
+ presentations 1-4, n = 28 mice;
Retrieval, CS- and CS
+, n = 21 mice; CS
- versus CS
+, paired t-test, *** P < 0.001, No freez
versus Freez, paired
t-test, *** P < 0.001). Error bars indicate mean ± s.e.m.
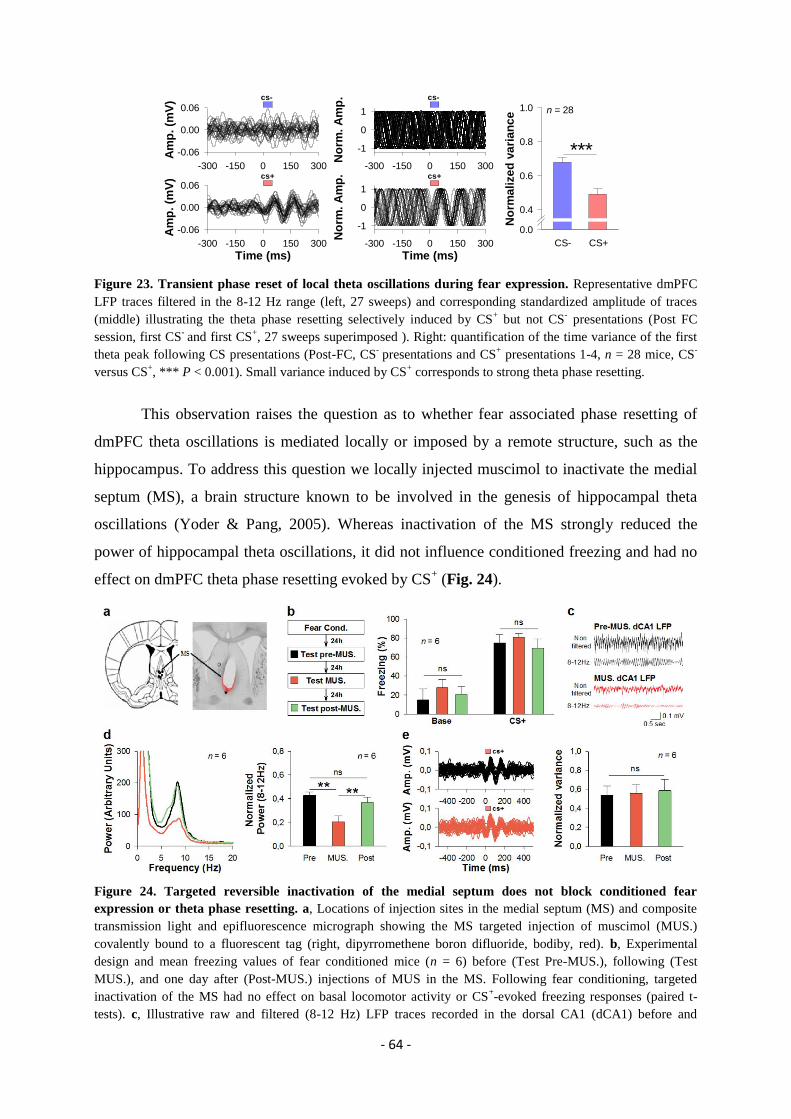
- 64 -
Figure 23. Transient phase reset of local theta oscillations during fear expression. Representative dmPFC
LFP traces filtered in the 8-12 Hz range (left, 27 sweeps) and corresponding standardized amplitude of traces
(middle) illustrating the theta phase resetting selectively induced by CS+ but not CS
- presentations (Post FC
session, first CS- and first CS
+, 27 sweeps superimposed ). Right: quantification of the time variance of the first
theta peak following CS presentations (Post-FC, CS- presentations and CS
+ presentations 1-4, n = 28 mice, CS
-
versus CS+, *** P < 0.001). Small variance induced by CS
+ corresponds to strong theta phase resetting.
This observation raises the question as to whether fear associated phase resetting of
dmPFC theta oscillations is mediated locally or imposed by a remote structure, such as the
hippocampus. To address this question we locally injected muscimol to inactivate the medial
septum (MS), a brain structure known to be involved in the genesis of hippocampal theta
oscillations (Yoder & Pang, 2005). Whereas inactivation of the MS strongly reduced the
power of hippocampal theta oscillations, it did not influence conditioned freezing and had no
effect on dmPFC theta phase resetting evoked by CS+ (Fig. 24).
Figure 24. Targeted reversible inactivation of the medial septum does not block conditioned fear
expression or theta phase resetting. a, Locations of injection sites in the medial septum (MS) and composite
transmission light and epifluorescence micrograph showing the MS targeted injection of muscimol (MUS.)
covalently bound to a fluorescent tag (right, dipyrromethene boron difluoride, bodiby, red). b, Experimental
design and mean freezing values of fear conditioned mice (n = 6) before (Test Pre-MUS.), following (Test
MUS.), and one day after (Post-MUS.) injections of MUS in the MS. Following fear conditioning, targeted
inactivation of the MS had no effect on basal locomotor activity or CS+-evoked freezing responses (paired t-
tests). c, Illustrative raw and filtered (8-12 Hz) LFP traces recorded in the dorsal CA1 (dCA1) before and
Time (ms)-300 -150 0 150 300
Am
p. (m
V)
-0.06
0.00
0.06
Time (ms)-300 -150 0 150 300
No
rm. A
mp
.
-1
0
1
-300 -150 0 150 300A
mp
. (m
V)
-0.06
0.00
0.06
-300 -150 0 150 300
No
rm. A
mp
.
-1
0
1
cs-
n = 28
cs+
cs-
cs+
No
rma
lize
d v
ari
an
ce
0.0
0.4
0.6
0.8
1.0
CS- CS+
***
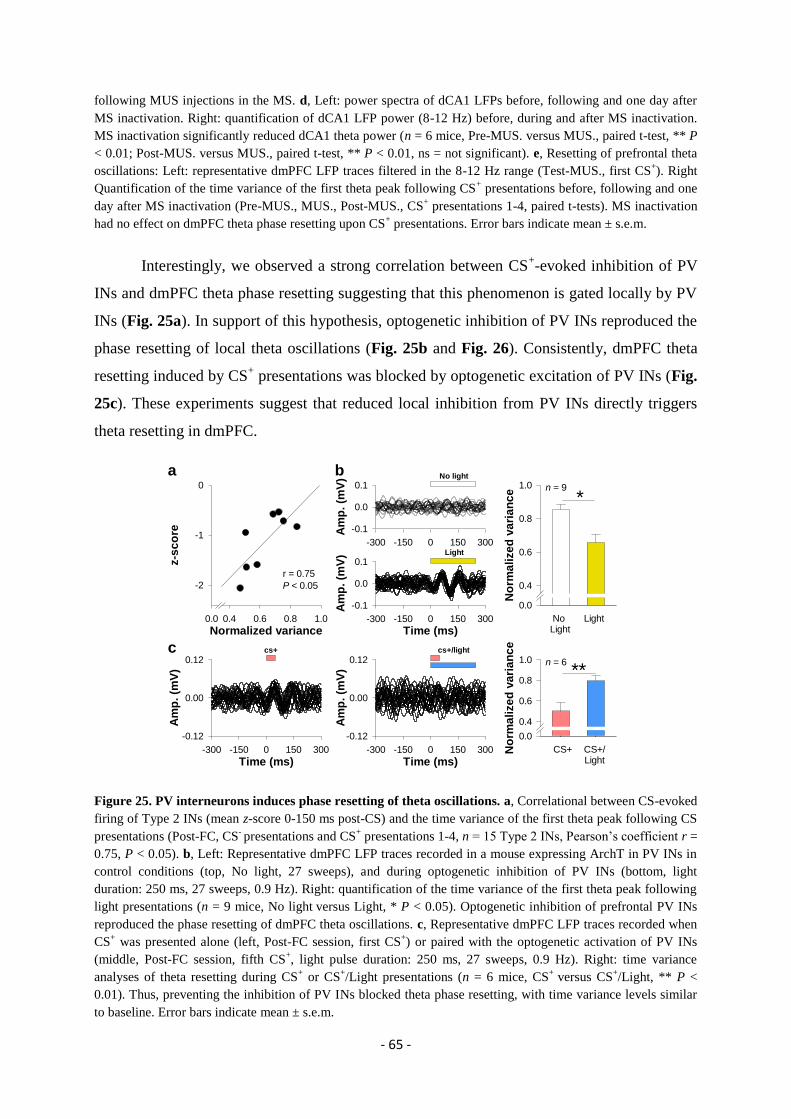
- 65 -
following MUS injections in the MS. d, Left: power spectra of dCA1 LFPs before, following and one day after
MS inactivation. Right: quantification of dCA1 LFP power (8-12 Hz) before, during and after MS inactivation.
MS inactivation significantly reduced dCA1 theta power (n = 6 mice, Pre-MUS. versus MUS., paired t-test, ** P
< 0.01; Post-MUS. versus MUS., paired t-test, ** P < 0.01, ns = not significant). e, Resetting of prefrontal theta
oscillations: Left: representative dmPFC LFP traces filtered in the 8-12 Hz range (Test-MUS., first CS+). Right
Quantification of the time variance of the first theta peak following CS+ presentations before, following and one
day after MS inactivation (Pre-MUS., MUS., Post-MUS., CS+ presentations 1-4, paired t-tests). MS inactivation
had no effect on dmPFC theta phase resetting upon CS+ presentations. Error bars indicate mean ± s.e.m.
Interestingly, we observed a strong correlation between CS+-evoked inhibition of PV
INs and dmPFC theta phase resetting suggesting that this phenomenon is gated locally by PV
INs (Fig. 25a). In support of this hypothesis, optogenetic inhibition of PV INs reproduced the
phase resetting of local theta oscillations (Fig. 25b and Fig. 26). Consistently, dmPFC theta
resetting induced by CS+ presentations was blocked by optogenetic excitation of PV INs (Fig.
25c). These experiments suggest that reduced local inhibition from PV INs directly triggers
theta resetting in dmPFC.
Figure 25. PV interneurons induces phase resetting of theta oscillations. a, Correlational between CS-evoked
firing of Type 2 INs (mean z-score 0-150 ms post-CS) and the time variance of the first theta peak following CS
presentations (Post-FC, CS- presentations and CS
+ presentations 1-4, n = 15 Type 2 INs, Pearson’s coefficient r =
0.75, P < 0.05). b, Left: Representative dmPFC LFP traces recorded in a mouse expressing ArchT in PV INs in
control conditions (top, No light, 27 sweeps), and during optogenetic inhibition of PV INs (bottom, light
duration: 250 ms, 27 sweeps, 0.9 Hz). Right: quantification of the time variance of the first theta peak following
light presentations (n = 9 mice, No light versus Light, * P < 0.05). Optogenetic inhibition of prefrontal PV INs
reproduced the phase resetting of dmPFC theta oscillations. c, Representative dmPFC LFP traces recorded when
CS+ was presented alone (left, Post-FC session, first CS
+) or paired with the optogenetic activation of PV INs
(middle, Post-FC session, fifth CS+, light pulse duration: 250 ms, 27 sweeps, 0.9 Hz). Right: time variance
analyses of theta resetting during CS+ or CS
+/Light presentations (n = 6 mice, CS
+ versus CS
+/Light, ** P <
0.01). Thus, preventing the inhibition of PV INs blocked theta phase resetting, with time variance levels similar
to baseline. Error bars indicate mean ± s.e.m.
a
Time (ms)-300 -150 0 150 300
Am
p. (m
V)
-0.1
0.0
0.1
Time (ms)-300 -150 0 150 300
Am
p. (m
V)
-0.12
0.00
0.12
Time (ms)-300 -150 0 150 300
Am
p. (m
V)
-0.12
0.00
0.12
Normalized variance0.0 0.4 0.6 0.8 1.0
z-s
co
re
-2
-1
0
r = 0.75
P < 0.05
n = 6
n = 9
c
-300 -150 0 150 300
Am
p. (m
V)
-0.1
0.0
0.1
cs+ cs+/light
No
rma
lize
d v
ari
an
ce
0.0
0.4
0.6
0.8
1.0
No
rma
lize
d v
ari
an
ce
0.0
0.4
0.6
0.8
1.0
CS+/Light
CS+
Light
No light
No Light
Light
b
**
*
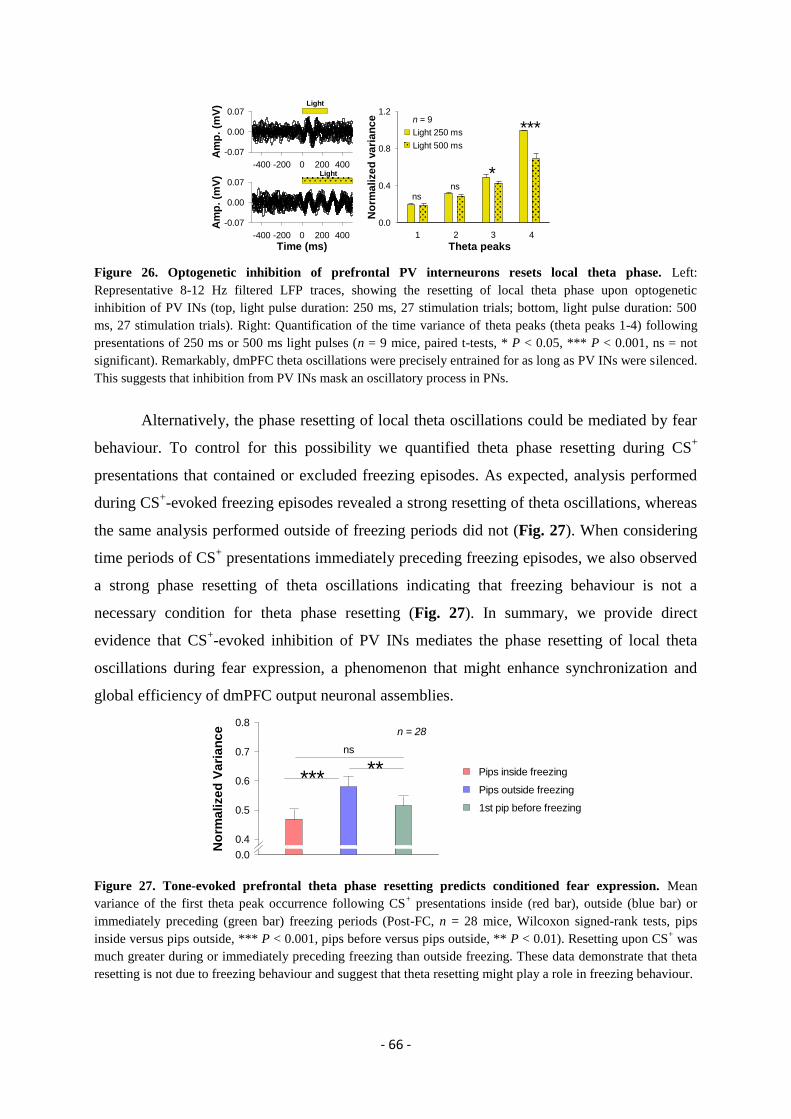
- 66 -
Figure 26. Optogenetic inhibition of prefrontal PV interneurons resets local theta phase. Left:
Representative 8-12 Hz filtered LFP traces, showing the resetting of local theta phase upon optogenetic
inhibition of PV INs (top, light pulse duration: 250 ms, 27 stimulation trials; bottom, light pulse duration: 500
ms, 27 stimulation trials). Right: Quantification of the time variance of theta peaks (theta peaks 1-4) following
presentations of 250 ms or 500 ms light pulses (n = 9 mice, paired t-tests, * P < 0.05, *** P < 0.001, ns = not
significant). Remarkably, dmPFC theta oscillations were precisely entrained for as long as PV INs were silenced.
This suggests that inhibition from PV INs mask an oscillatory process in PNs.
Alternatively, the phase resetting of local theta oscillations could be mediated by fear
behaviour. To control for this possibility we quantified theta phase resetting during CS+
presentations that contained or excluded freezing episodes. As expected, analysis performed
during CS+-evoked freezing episodes revealed a strong resetting of theta oscillations, whereas
the same analysis performed outside of freezing periods did not (Fig. 27). When considering
time periods of CS+ presentations immediately preceding freezing episodes, we also observed
a strong phase resetting of theta oscillations indicating that freezing behaviour is not a
necessary condition for theta phase resetting (Fig. 27). In summary, we provide direct
evidence that CS+-evoked inhibition of PV INs mediates the phase resetting of local theta
oscillations during fear expression, a phenomenon that might enhance synchronization and
global efficiency of dmPFC output neuronal assemblies.
Figure 27. Tone-evoked prefrontal theta phase resetting predicts conditioned fear expression. Mean
variance of the first theta peak occurrence following CS+ presentations inside (red bar), outside (blue bar) or
immediately preceding (green bar) freezing periods (Post-FC, n = 28 mice, Wilcoxon signed-rank tests, pips
inside versus pips outside, *** P < 0.001, pips before versus pips outside, ** P < 0.01). Resetting upon CS+ was
much greater during or immediately preceding freezing than outside freezing. These data demonstrate that theta
resetting is not due to freezing behaviour and suggest that theta resetting might play a role in freezing behaviour.
Time (ms)-400 -200 0 200 400
Am
p. (m
V)
-0.07
0.00
0.07
Theta peaks1 2 3 4
nsns
*
***n = 9
Light
Light
Light 250 ms
Light 500 ms
No
rma
lize
d v
ari
an
ce
0.0
0.4
0.8
1.2
-400 -200 0 200 400
Am
p. (m
V)
-0.07
0.00
0.07
No
rma
lize
d V
ari
an
ce
0.0
0.4
0.5
0.6
0.7
0.8n = 28
ns
***** Pips inside freezing
Pips outside freezing
1st pip before freezing
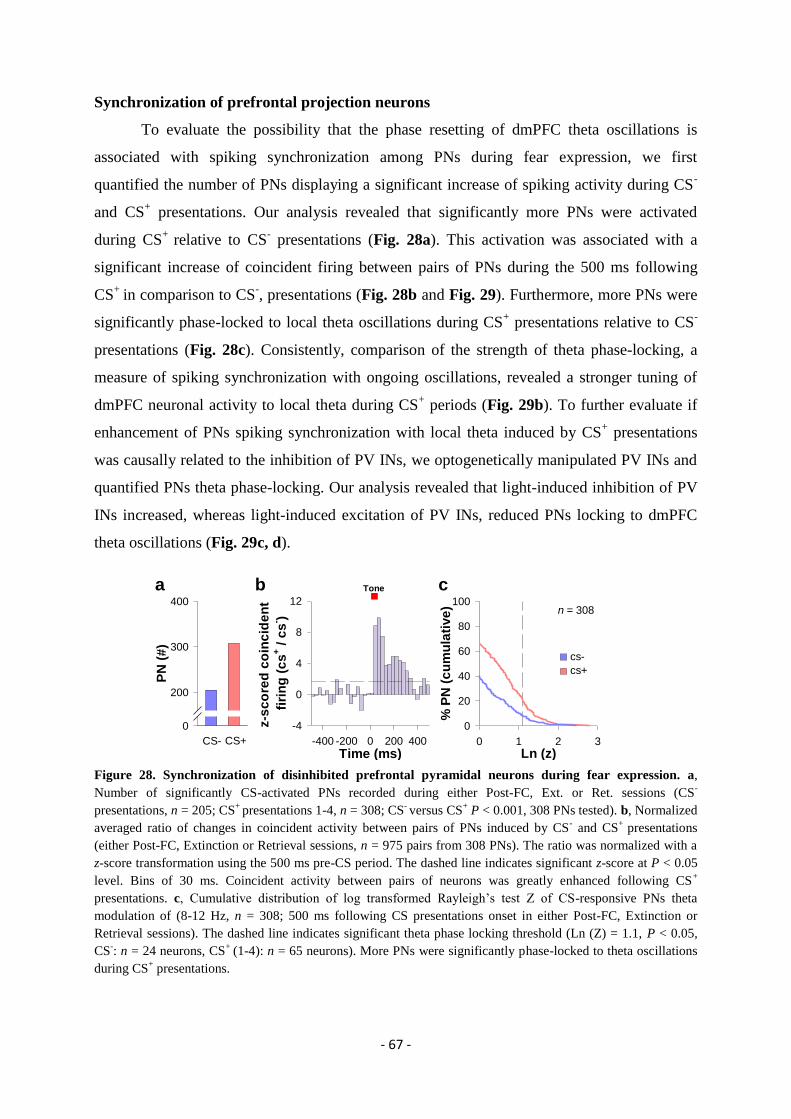
- 67 -
Synchronization of prefrontal projection neurons
To evaluate the possibility that the phase resetting of dmPFC theta oscillations is
associated with spiking synchronization among PNs during fear expression, we first
quantified the number of PNs displaying a significant increase of spiking activity during CS-
and CS+ presentations. Our analysis revealed that significantly more PNs were activated
during CS+
relative to CS- presentations (Fig. 28a). This activation was associated with a
significant increase of coincident firing between pairs of PNs during the 500 ms following
CS+
in comparison to CS-, presentations (Fig. 28b and Fig. 29). Furthermore, more PNs were
significantly phase-locked to local theta oscillations during CS+ presentations relative to CS
-
presentations (Fig. 28c). Consistently, comparison of the strength of theta phase-locking, a
measure of spiking synchronization with ongoing oscillations, revealed a stronger tuning of
dmPFC neuronal activity to local theta during CS+ periods (Fig. 29b). To further evaluate if
enhancement of PNs spiking synchronization with local theta induced by CS+ presentations
was causally related to the inhibition of PV INs, we optogenetically manipulated PV INs and
quantified PNs theta phase-locking. Our analysis revealed that light-induced inhibition of PV
INs increased, whereas light-induced excitation of PV INs, reduced PNs locking to dmPFC
theta oscillations (Fig. 29c, d).
Figure 28. Synchronization of disinhibited prefrontal pyramidal neurons during fear expression. a,
Number of significantly CS-activated PNs recorded during either Post-FC, Ext. or Ret. sessions (CS-
presentations, n = 205; CS+
presentations 1-4, n = 308; CS- versus CS
+ P < 0.001, 308 PNs tested). b, Normalized
averaged ratio of changes in coincident activity between pairs of PNs induced by CS- and CS
+ presentations
(either Post-FC, Extinction or Retrieval sessions, n = 975 pairs from 308 PNs). The ratio was normalized with a
z-score transformation using the 500 ms pre-CS period. The dashed line indicates significant z-score at P < 0.05
level. Bins of 30 ms. Coincident activity between pairs of neurons was greatly enhanced following CS+
presentations. c, Cumulative distribution of log transformed Rayleigh’s test Z of CS-responsive PNs theta
modulation of (8-12 Hz, n = 308; 500 ms following CS presentations onset in either Post-FC, Extinction or
Retrieval sessions). The dashed line indicates significant theta phase locking threshold (Ln (Z) = 1.1, P < 0.05,
CS-: n = 24 neurons, CS
+ (1-4): n = 65 neurons). More PNs were significantly phase-locked to theta oscillations
during CS+ presentations.
Ln (z)0 1 2 3
% P
N (
cu
mu
lati
ve
)
0
20
40
60
80
100
b ca
n = 308
PN
(#
)
0
200
300
400
CS+CS-
Time (ms)-400 -200 0 200 400
z-s
co
red
co
inc
ide
nt
firi
ng
(c
s+ /
cs
- )
-4
0
4
8
12
cs-
cs+
Tone
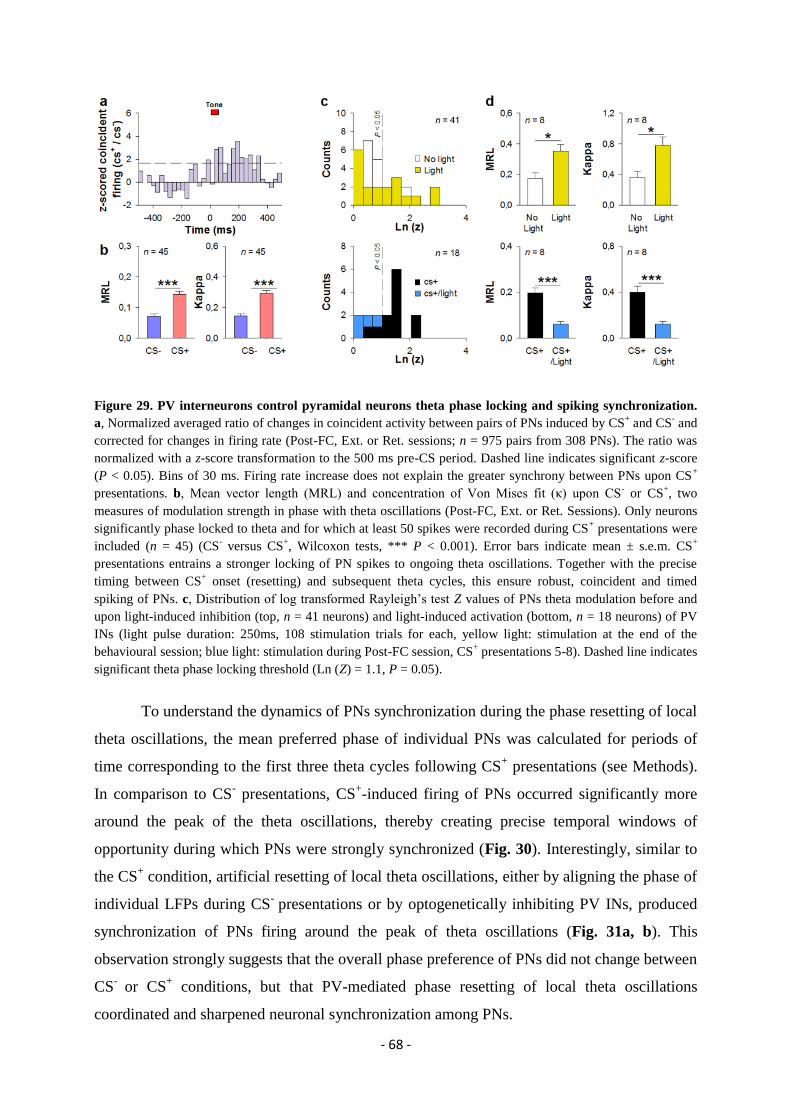
- 68 -
Figure 29. PV interneurons control pyramidal neurons theta phase locking and spiking synchronization.
a, Normalized averaged ratio of changes in coincident activity between pairs of PNs induced by CS+ and CS
- and
corrected for changes in firing rate (Post-FC, Ext. or Ret. sessions; n = 975 pairs from 308 PNs). The ratio was
normalized with a z-score transformation to the 500 ms pre-CS period. Dashed line indicates significant z-score
(P < 0.05). Bins of 30 ms. Firing rate increase does not explain the greater synchrony between PNs upon CS+
presentations. b, Mean vector length (MRL) and concentration of Von Mises fit (κ) upon CS- or CS
+, two
measures of modulation strength in phase with theta oscillations (Post-FC, Ext. or Ret. Sessions). Only neurons
significantly phase locked to theta and for which at least 50 spikes were recorded during CS+ presentations were
included (n = 45) (CS- versus CS
+, Wilcoxon tests, *** P < 0.001). Error bars indicate mean ± s.e.m. CS
+
presentations entrains a stronger locking of PN spikes to ongoing theta oscillations. Together with the precise
timing between CS+ onset (resetting) and subsequent theta cycles, this ensure robust, coincident and timed
spiking of PNs. c, Distribution of log transformed Rayleigh’s test Z values of PNs theta modulation before and
upon light-induced inhibition (top, n = 41 neurons) and light-induced activation (bottom, n = 18 neurons) of PV
INs (light pulse duration: 250ms, 108 stimulation trials for each, yellow light: stimulation at the end of the
behavioural session; blue light: stimulation during Post-FC session, CS+ presentations 5-8). Dashed line indicates
significant theta phase locking threshold (Ln (Z) = 1.1, P = 0.05).
To understand the dynamics of PNs synchronization during the phase resetting of local
theta oscillations, the mean preferred phase of individual PNs was calculated for periods of
time corresponding to the first three theta cycles following CS+ presentations (see Methods).
In comparison to CS- presentations, CS
+-induced firing of PNs occurred significantly more
around the peak of the theta oscillations, thereby creating precise temporal windows of
opportunity during which PNs were strongly synchronized (Fig. 30). Interestingly, similar to
the CS+ condition, artificial resetting of local theta oscillations, either by aligning the phase of
individual LFPs during CS- presentations or by optogenetically inhibiting PV INs, produced
synchronization of PNs firing around the peak of theta oscillations (Fig. 31a, b). This
observation strongly suggests that the overall phase preference of PNs did not change between
CS-
or CS+ conditions, but that PV-mediated phase resetting of local theta oscillations
coordinated and sharpened neuronal synchronization among PNs.

- 69 -
Figure 30. Dynamics of pyramidal neurons synchronization during fear expression. Theta modulation of
PNs significantly phase locked to theta and displaying at least 15 spikes during No light and Light conditions.
Modulation with local theta was measured with the MRL (top left, n= 8 neurons, yellow light stimulation, paired
t-tests, No light versus Light: * P < 0.05; bottom left, n= 8 neurons, blue light stimulation, No light versus Light:
* P < 0.05) and κ (top right, n = 8 neurons, yellow light stimulation, paired t-tests, No light versus Light: *** P <
0.001; bottom left, n = 8 neurons, blue light stimulation, No light versus Light: *** P < 0.001). Error bars
indicate mean ± s.e.m. These results show that inhibiting PV INs is both sufficient to increase PNs’ modulation
with local theta, and necessary for theta entrainment of PNs evoked by CS+.
Figure 31. Artificially-induced phase resetting of prefrontal theta synchronizes pyramidal neurons. a, Top:
distribution of PNs preferred theta phase (n = 308) during cycles around CS-. The phase of individual LFPs were
aligned to the first theta peak following CS- onset to mimic phase resetting of local theta (one theta cycle before,
and 3 theta cycles following CS were included, bins of 45°). Bottom: distribution of individual PNs preferred
theta phases during theta cycles around CS-
showing a synchronization of PNs around the peak of the LFP
(Rayleigh’s test for circular uniformity: first theta cycle post CS, P < 0.001 indicating that the circular
distribution is not uniform). b, Top: distribution of PNs preferred theta phase (n = 41) during theta cycles outside
light stimulation (left, 15.8% freezing) and upon light-induced resetting of theta oscillations (right, 36.8%
freezing; one theta cycle before, and 3 theta cycles following CS were included, bins of 45°). Bottom:
corresponding distributions of individual PNs preferred theta phase outside and upon light stimulation. Despite a
low number of neurons and a moderate freezing induced by light inhibition of dmPFC PV INs (36.8 % freezing),
this analysis revealed that light-induced reset of local theta oscillations promotes neuronal synchronization of
PNs (Rayleigh’s test for circular uniformity: first theta cycle post CS, Light P < 0.001, No light P = ns).
How do PV INs inhibition-mediated increase in PNs synchronization promote fear
behaviour? Converging evidence indicates that dmPFC PNs target both the BLA and the
periaqueductal grey (PAG), two brain structures strongly involved in the encoding and
expression of fear behaviour (DiScala et al., 1987; Gabbott et al., 2005; Herry et al., 2008;
PN
#
0
40
80
PN
#
0
40
80
Preferred phase (°)
300
200
100
0
PN
#
Preferred phase (°)
300
200
100
0
PN
#
0-360 360 720 108010807203600-360
CS- (14.7% freez.) CS+ (69.8% freez.)
PN
#
0
40
80
300
200
100
0
CS-
Preferred phase (Deg.)
PN
#
0-360 360 720 1080
a b
PN
#
0
5
10
No light (15.8% freezing) Light (36.8% freezing)
Preferred phase (Deg.)
0-360 360 720 1080
Preferred phase (Deg.)
PN
#
0-360 360 720 10800
20
40
0
PN
#
0
5
10
PN
#
20
40
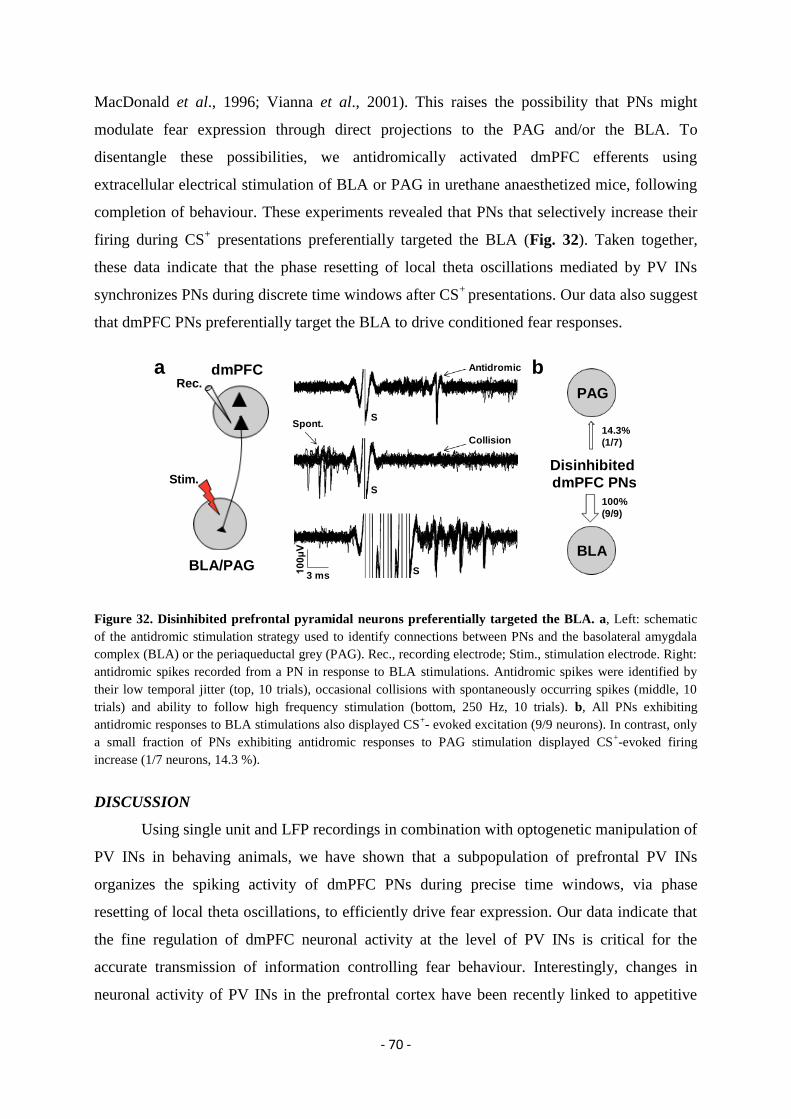
- 70 -
MacDonald et al., 1996; Vianna et al., 2001). This raises the possibility that PNs might
modulate fear expression through direct projections to the PAG and/or the BLA. To
disentangle these possibilities, we antidromically activated dmPFC efferents using
extracellular electrical stimulation of BLA or PAG in urethane anaesthetized mice, following
completion of behaviour. These experiments revealed that PNs that selectively increase their
firing during CS+ presentations preferentially targeted the BLA (Fig. 32). Taken together,
these data indicate that the phase resetting of local theta oscillations mediated by PV INs
synchronizes PNs during discrete time windows after CS+
presentations. Our data also suggest
that dmPFC PNs preferentially target the BLA to drive conditioned fear responses.
Figure 32. Disinhibited prefrontal pyramidal neurons preferentially targeted the BLA. a, Left: schematic
of the antidromic stimulation strategy used to identify connections between PNs and the basolateral amygdala
complex (BLA) or the periaqueductal grey (PAG). Rec., recording electrode; Stim., stimulation electrode. Right:
antidromic spikes recorded from a PN in response to BLA stimulations. Antidromic spikes were identified by
their low temporal jitter (top, 10 trials), occasional collisions with spontaneously occurring spikes (middle, 10
trials) and ability to follow high frequency stimulation (bottom, 250 Hz, 10 trials). b, All PNs exhibiting
antidromic responses to BLA stimulations also displayed CS+- evoked excitation (9/9 neurons). In contrast, only
a small fraction of PNs exhibiting antidromic responses to PAG stimulation displayed CS+-evoked firing
increase (1/7 neurons, 14.3 %).
DISCUSSION
Using single unit and LFP recordings in combination with optogenetic manipulation of
PV INs in behaving animals, we have shown that a subpopulation of prefrontal PV INs
organizes the spiking activity of dmPFC PNs during precise time windows, via phase
resetting of local theta oscillations, to efficiently drive fear expression. Our data indicate that
the fine regulation of dmPFC neuronal activity at the level of PV INs is critical for the
accurate transmission of information controlling fear behaviour. Interestingly, changes in
neuronal activity of PV INs in the prefrontal cortex have been recently linked to appetitive
a b
S
Collision
Antidromic
S
S
Spont.
dmPFC
Stim.
BLA/PAG
Rec.PAG
10
0µ
V
3 ms
Disinhibited
dmPFC PNs
100%
(9/9)
14.3%
(1/7)
BLA

- 71 -
associative learning suggesting that PV IN might be involved in both aversive and appetitive
situations (Kvitsiani et al., 2013). Our data indicated that inhibition of Type 2 PV INs during
CS+ presentations was causally related to the expression of conditioned fear responses and
suggest that Type 1 INs might directly inhibit Type 2 INs, although it is possible that
inhibition of Type 2 INs may arise from a third population of unidentified dmPFC INs.
Whereas the origin of CS-mediated Type 1 INs excitatory neuronal responses remains to be
determined, it is likely that they receive inputs from neuronal structures involved in the
encoding or modulation of conditioned fear such as BLA or hippocampus (Sotres-Bayon et
al., 2012; Tierney et al., 2004). Moreover, future work will be required to investigate the
precise cellular identity of Type 1 INs.
What neuronal mechanisms can account for our observation that inhibition of PV INs
is both necessary and sufficient for the expression of fear responses? Cortical PV INs are
known to inhibit PNs through powerful perisomatic inhibition (Freund & Katona, 2007). As a
consequence, CS+-evoked neuronal inhibition in PV INs induced a strong disinhibition of
PNs, a permissive mechanism that gated neuronal responses during fear expression. These
results indicate that tone-evoked activity in dmPFC PNs during conditioned fear expression
result in part from a disinhibitory mechanism. Notably, freezing behaviour was not entirely
prevented by PV INs activation, indicating that some dmPFC PNs may escape inhibitory
control, or that other brain regions promote fear responses in concert with dmPFC.
Importantly, it is possible that the increase neuronal activity observed in dmPFC PNs upon
CS+ presentations arises from direct BLA or hippocampal inputs in addition to the direct
disinhibition of PNs exerted by PV INs. Indeed, disinhibition of PNs is a powerful permissive
mechanism that opens precise temporal windows during which excitatory inputs onto PNs
could be enhanced. However the observation that light-induced activation of PV INs blocked
tone-evoked activity in PNs (Fig. 21a, b, c) strongly suggests that the main regulatory
neuronal mechanism of PNs neuronal activity is provided by direct PV INs inputs. Although
most of the dmPFC PNs neurons recorded displayed excitatory responses during CS+
presentations (87.7 %, n = 308/351), a small fraction of dmPFC PNs displayed CS-evoked
inhibitory responses (12.3 %, n = 43/351), that could be mediated by amygdala inputs-
mediated feed-forward inhibition onto dmPFC PNs as recently observed (Dilgen et al., 2013).
Although fear behaviour was globally associated with a reduction in dmPFC theta
oscillations magnitude, CS+-evoked inhibition of PV INs induced a robust and transient theta
phase resetting spanning two to three theta cycles. Transient phase resetting of theta
oscillations has been previously observed in cortical regions following electrical or sensory

- 72 -
stimulations, including appetitive conditioned stimuli (Buzsaki et al., 1979; Makeig et al.,
2002; McCartney et al., 2004; Rizzuto et al., 2003; Takehara-Nishiuchi et al., 2011; Williams
et al., 2003; Zugaro et al., 2005). Our findings provide the first mechanistic explanation of
phase resetting at the cellular level and extend this phenomenon to the control of emotional
behaviour. Functionally, we observed that phase resetting of local theta oscillations strongly
synchronized PNs around theta peaks without changing the preferred phases of individual
PNs. This observation has important consequences because it suggests that resetting of local
theta oscillations, but not the preferred phases of individual PNs to the local LFP, is critically
involved in the behavioural expression of fear responses. Thus, theta phase resetting
represents a powerful mechanism for reliable fear expression because it creates an optimal
temporal relationship that precisely binds spiking activity with sensory information provided
by CS. Ultimately, phase resetting of oscillations is an efficient mechanism that enhances the
impact of input signals and enables precise transmission of information to downstream
targets. Our data also show that reduction of rhythmic inhibition from PV INs paradoxically
increases synchrony. Suppression of interference between two oscillators may account for this
effect. Future work will be needed to identify the origin of dmPFC theta oscillations that are
unmasked by PV INs inhibition.
How could synchronized PNs accurately control fear expression? Previous findings
suggest that putative dmPFC PNs displaying sustained or transient changes in their spiking
activity promote fear expression through activation and synchronization of BLA neurons
(Chang et al., 2010; Lesting et al., 2011; Livneh & Paz, 2012; Pape & Pare, 2010; Sotres-
Bayon et al., 2012). In line with these studies, our results demonstrate that PNs exhibiting
CS+-evoked synchronized firing during fear expression preferentially project to the BLA
where they may target specific populations of neurons that are activated during fear behaviour
(Bienvenu et al., 2012; Goosens et al., 2003; Herry et al., 2008; Quirk et al., 1995). Although
our data demonstrate that specific inhibitory neuronal circuits in the dmPFC control freezing
behaviour by disinhibiting and synchronizing dmPFC neurons projecting to the BLA, the
most parsimonious interpretation of our results would be that the behavioural changes
observed are directly mediated by BLA or CeA output neurons. Indeed, most of dmPFC PNs
projecting to the PAG were not activated during CS+ presentations. This observation suggests
that this dmPFC-PAG projecting pathway does not play an active role during freezing
expression, although it might be required for other behavioural manifestation of fear
responses. Further work will be required to precisely identify the role of dmPFC-PAG
projection pathway during fear behaviour.

- 73 -
Finally our findings suggest that persistent fear behaviour, which is at the core of
psychiatric conditions such as anxiety disorders, might be finely regulated at the level of
dedicated prefrontal inhibitory circuits.

- 74 -
GENERAL DISCUSSION
The identification of factors and circuit elements controlling neuronal activity during
learning and memory represent one of the most important challenges for neurosciences
research. By using a combination of single unit recordings, pharmacological and optogenetic
manipulations in behaving mice submitted to classical auditory fear conditioning, we
identified the specific cortical neuronal elements and mechanisms involved in the acquisition
and expression of conditioned fear behaviour. Numerous recent reports highlighted the
important role played by cortical circuits in the regulation of fear behaviour. However, the
anatomical and functional characteristics of the individual circuit elements remain largely
unknown. During my thesis, we described that both AC and mPFC inhibitory neuronal
elements form a disinhibitory microcircuit required for the acquisition and the expression of
auditory conditioned fear behaviour, respectively. In both cases, PV INs constitute the central
element of the circuit and were phasically inhibited during presentations of conditioned tones.
Our data suggest that this phasic inhibition is provided by another population of INs and
causes the disinhibition of PNs. From a functional point of view, disinhibition is an attractive
mechanism that opens precise temporal windows during which tonic perisomatic inhibition
mediated by PV INs onto PNs is removed (Fig. 33a). Thereby, this disinhibitory microcircuit
can precisely control the temporal excitability of PNs. In particular, in the AC, we
demonstrated that layer 1 INs-mediated disinhibition of PNs is required to ensure stimulus
convergence, which is essential for the acquisition of associative fear memory. Secondly, in
the mPFC, we demonstrated that the optogenetic inhibition of prefrontal PV INs activity,
disinhibits PNs and synchronizes their firing by resetting local theta oscillations, leading to
fear expression (Fig. 33). Several questions that remain unanswered or that emerged during
my thesis are discussed below.
First of all, it would be particularly interesting, in both cortices, to identify precisely
the identity and the characteristics of the presynaptic inhibitory neurons that provide the
inhibition to PV INs. Concerning our experiments in the AC, we have already identified that
these presynaptic inhibitory neurons are located in layer 1, that they receive a massive
cholinergic input from the NBM and that they expressed nicotinic receptors. However, their
precise cellular identity is still unknown. Concerning our experiments in the mPFC, our data
strongly suggest that the population of prefrontal Type 1 INs directly inhibits PV INs during
conditioned tone presentations. However, the direct connectivity between Type 1 and 2 IN
could not be demonstrated, just suggested by our experiments. To address this point further, it

- 75 -
will be necessary to identify specific markers of Type 1 INs in order to manipulate or labeled
them. Nevertheless, a careful review of the literature on the connectivity between INs
revealed that PV INs are often mutually connected and received inputs originating mainly
from SOM, VIP and CR INs (Burkhalter, 1999; Caputi et al., 2009; Gentet, 2012; Gonchar
and Meskenaite, 1997; Gabbott et al., 1997; Hioki et al., 2013; Pfeffer et al., 2013; Xu et al.,
2013). Because we know that Type 1 INs are not PV INs, this finding open the possibility that
Type 1 INs belongs to the SOM, VIP or CR subclass, an hypothesis that will need to be
demonstrated using genetic mouse models and optogenetic strategies. Ultimately, the
identification of specific markers for Type 1 INs could help us to firmly establish that they
mediate Type 2 PV INs inhibition, to investigate their properties in vivo and in vitro and to
define their anatomical and functional inputs.
During my thesis, our two studies, along with other recent publications, identified
neuronal disinhibition as a key neuronal mechanism involved in cortical processes in general,
and fear behaviour in particular (Gentet, 2012; Kuhlman et al., 2013; Lovett-Barron et al.,
2012; Pfeffer et al., 2013; Pi et al., 2013; Xu et al., 2013). Disinhibition is a recent concept
that emerged mainly from anatomical observations of IN/IN connections. Almost all of the
studies reporting a serial circuit linking two INs emphasized a potential disinhibitory function
(Deniau & Chevalier, 1985; Freund & Antal, 1988; Gabbott et al., 1997; Meskenaite, 1997;
Tamas et al., 1998). A simple disinhibitory circuit is often composed by a first IN (the
inhibitor IN) contacting a second IN (the disinhibitor IN) and the target neurons of the latter
one, in our case PNs (Fig. 33a). Functionally, disinhibition is as a powerful mechanism
allowing the accurate control of downstream PNs excitability. Specifically, the activation of
the inhibitor INs disinhibits PNs by reducing their tonic inhibition on disinhibitor INs (Fig.
33b). An important aspect concerning this disinhibitory mechanism is the relation between the
duration of the disinhibitor INs inhibition and PNs activation. Indeed, in the mPFC we
showed that the length of the optogenetic-mediated PV INs inhibition determined the duration
of the PNs disinhibition. Thereby, disinhibitory circuits are able to open precise temporal
widows during which PNs can integrate neuronal inputs. In the AC, disinhibition has proven
to be an attractive mechanism for fear learning because it is permissive for strong activation
and concomitant plasticity induction. Indeed, the available sensory input at the time of the
aversive stimulus can select the circuit elements for plasticity induction. Interestingly, some
studies indicated that disinhibition can be provided by non PV disinhibitor INs such as SOM
INs (Gentet, 2012; Lovett-Barron et al., 2012; Pfeffer et al., 2013; Xu et al., 2013). Given the
fact that PV and SOM INs target differentially the somatic or dendritic regions of PNs, a

- 76 -
disinhibitory circuit engaging PV or SOM INs would respectively regulate the PNs
excitability at the level of their output or input compartments. Furthermore, according to their
presynaptic inputs, PV or SOM INs would control PNs either via an inhibition or a
disinhibition. Together, these observations highlight the importance of precisely
understanding the wiring layout of distinct INs to understand their specific functions.
Figure 33. Prefrontal disinhibitory circuits controlling fear expression. a, Schematic of the dmPFC
disinhibitory circuit composed by Type 1 INs (brown) , Type 2 PV INs contacted by Type 1 (pink) and the
targets pyramidal neurons of Type 2 INs projecting to the BLA (grey). b Scheme of the disinhibition of
prefrontal PNs during high fear state. c, Disinhibition and synchronization of BLA projecting PNs via the
inhibition of type 2 PV INs and the resetting of local theta oscillations.
Furthermore, our study conducted in the mPFC revealed that disinhibition is a critical
mechanism to efficiently control the spiking activity of prefrontal PNs to drive fear
expression. Specifically, we showed that prefrontal PNs are not only activated during the time
window of disinhibition, but their spiking activity is accurately organized via phase resetting
of local theta oscillations during fear expression. Indeed, CS+-evoked inhibition of PV INs
induced a robust and transient theta phase resetting spanning two to three theta cycles.
Functionally, we observed that phase resetting of local theta oscillations synchronized PNs
around theta peaks without changing their individual preferred phases and drove fear
expression (Fig. 33c). This observation has important consequences because it suggests that
the synchronization of PNs is due to a resetting of local theta oscillations, but not to a shift of
the preferred phases of individual PNs to the local LFP, as has been previously described
(Benchenane et al., 2010; Cobb et al., 1995). Moreover, we showed that PV INs inhibition is
both necessary and sufficient to generate the resetting of dmPFC theta oscillations.
-400 0 400-400 0 400 -400 0 400Time (ms)
BLA projectingPN
Type 1putative IN
Type 2PV IN
Type 1 IN
Type 2 PV IN
BLA projecting PN
100 ms
CS+
BLA
IN
PN
PV
IN
IN
PV
PNPN
PN
a b
c CS+
dmPFC LFP(8 – 12 Hz)

- 77 -
Resetting of theta oscillations could be mediated by at least two different mechanisms.
First of all, the fact that inhibition of Type 2 INs induced this oscillation strongly suggests
that under normal conditions, this oscillation is actively blocked by the tonic inhibition
emanating from Type 2 INs. This could be mediated for instance by a tonic inhibition of
glutamatergic afferences originating from remote structures, such as the hippocampus. At odd
with this possibility, local targeted inactivation of the MS, a neuronal structure involved in the
genesis of theta oscillations (Yoder and Pang, 2005) did not influence theta resetting. While
these data suggests that resetting of theta oscillation is a local prefrontal phenomenon, we
cannot rule out the involvement of different neuronal structures in the genesis of this
oscillation. A second possibility relies on the existence within the dmPFC of two neuronal
oscillators that will oscillate in the theta range but with an opposite phase, one of which would
comprise Type 2 INs. Under normal condition the oscillations will cancel each other because
of their opposite phase. However, during high fear states, the strong inhibition of one of
oscillator (comprised of Type 2 INs) will reveal the second oscillation mediated by the second
oscillator. Although this mechanism is a very interesting one, our experiments did not allow
us to identify the second oscillator in the dmPFC. Further work will be required to precisely
identify this second oscillator and validate the dual oscillator model we proposed here.
Finally, we provided evidence that prefrontal PNs involved in the disinhibition process
preferentially target the BLA but not the PAG. Additional analyses indicated that most of the
PAG projecting neurons were not activated during CS+ presentations but were rather
inhibited. Thus, in accordance with previous findings, these observations suggest that dmPFC
PNs promote fear expression by activating BLA neurons (Fig. 33, for a detailed review see
Pape and Pare, 2010). In the future, it would be interesting to causally demonstrate the exact
contribution of dmPFC-BLA and dmPFC-PAG pathways in the regulation of fear behaviour.
The data cumulated during my thesis confirmed the general principle that the anatomical and
functional specialization of cortical INs confers them their functional specificities (Buzsaki et
al., 2004; Gentet, 2012; Isaacson & Scanziani, 2011; Merchant et al., 2012; Moore et al.,
2010). Indeed, by revealing the neuronal elements and the function of PV dependent
disinhibitory microcircuit, we contributed to the understanding of the cortical processes
governing the learning and expression of fear memories. Gaining an exhaustive knowledge of
the anatomical and functional characteristics of cortical INs represents an exciting and
challenging research avenue that could potentially provide new therapeutic strategies for
pathological fear behaviour.

- 78 -
PUBLICATIONS AND COMMUNICATIONS
PUBLICATIONS
Jalabert, M., Bourdy, R., Courtin, J., Veinante, P., Manzoni, O., Barrot, M. & Georges, F.
Neuronal circuits underlying acute morphine action on dopamine neurons. Proceedings of the
National Academy of Sciences of the United States of America 108, 16446-16450 (2011).
Letzkus, J., Wolff, S., Meyer, E., Tovote, P., Courtin, J., Herry, C. & Lüthi, A. A
disinhibitory microcircuit for associative fear learning in the auditory cortex. Nature 480,
331-335 (2011).
Courtin, J., Gonzalez-Campo, C. & Herry, C. Neural mechanisms of extinction learning and
retrieval. In Encyclopedia of the Sciences of Learning, Seel. NM. (Ed). Springer Publishers,
New York (2012).
Courtin, J., Bienvenu, T. C., Einarsson, E. O. & Herry, C. Medial prefrontal cortex neuronal
circuits in fear behavior. Neuroscience 240, 219-242 (2013).
Courtin, J., Karalis, N., Gonzalez-Campo, C., Wurtz, H. & Herry, C. Persistence of
amygdala gamma oscillations during extinction learning predicts spontaneous fear recovery.
Neurobiology of learning and memory (2013).
Courtin, J., Chaudun, F., Rozeske, R. R., Karalis, N., Gonzalez-Campo, C., Wurtz, H., Abdi,
A., Baufreton, J., Bienvenu, T. C. M. & Herry, C. Prefrontal parvalbumin interneurons shape
neuronal activity to drive in fear expression. In press at Nature.
POSTERS
Courtin, J. and Herry, C. Lesions of the ventral medial prefrontal cortex enhanced contextual
fear generalization in C57BL6/J mice. ECNP meeting, Nice, France, March 2011.
Courtin, J. and Herry, C. A subpopulation of prefrontal putative inhibitory interneurons
controls conditioned fear expression. 10th
meeting of the French Society for Neurosciences,
Marseille, France, May 2011.
Courtin, J., Gonzalez-Campo, C., Chaudun, F. & Herry, C. Prefrontal neuronal circuits
controlling fear behavior. FENS meeting, Barcelona, Spain, July 2012.
Courtin, J., Chaudun, F., Rozeske, R. R., Karalis, N., Gonzalez-Campo, C., Wurtz, H.,
Bienvenu, T. C. M. & Herry, C. Prefrontal parvalbumin-expressing interneurons control fear
expression. 11th
meeting of the French Society for Neurosciences, Lyon, France, May 2013.

- 79 -
REFERENCES
Adesnik, H. & Scanziani, M. Lateral competition for cortical space by layer-specific
horizontal circuits. Nature 464, 1155-1160 (2010).
Adhikari, A., Topiwala, M. A. & Gordon, J. A. Single units in the medial prefrontal cortex
with anxiety-related firing patterns are preferentially influenced by ventral hippocampal
activity. Neuron 71, 898-910 (2011).
Adhikari, A., Topiwala, M. A. & Gordon, J. A. Synchronized activity between the ventral
hippocampus and the medial prefrontal cortex during anxiety. Neuron 65, 257-269 (2010).
Akirav, I., Raizel, H. & Maroun, M. Enhancement of conditioned fear extinction by infusion
of the GABA(A) agonist muscimol into the rat prefrontal cortex and amygdala. The European
journal of neuroscience 23, 758-764 (2006).
Alexander, G. E. Basal ganglia-thalamocortical circuits: their role in control of movements. J
Clin Neurophysiol 11, 420-431 (1994).
Alexander, G. E. & Crutcher, M. D. Functional architecture of basal ganglia circuits: neural
substrates of parallel processing. Trends Neurosci 13, 266-271 (1990a).
Alexander, G. E., Crutcher, M. D. & DeLong, M. R. Basal ganglia-thalamocortical circuits:
parallel substrates for motor, oculomotor, "prefrontal" and "limbic" functions. Progress in
brain research 85, 119-146 (1990b).
Alexander, G. E., DeLong, M. R. & Strick, P. L. Parallel organization of functionally
segregated circuits linking basal ganglia and cortex. Annual review of neuroscience 9, 357-
381 (1986).
Amano, T., Duvarci, S., Popa, D. & Pare, D. The fear circuit revisited: contributions of the
basal amygdala nuclei to conditioned fear. The Journal of neuroscience: the official journal of
the Society for Neuroscience 31, 15481-15489 (2011).
Anglada-Figueroa, D. & Quirk, G. J. Lesions of the basal amygdala block expression of
conditioned fear but not extinction. The Journal of neuroscience: the official journal of the
Society for Neuroscience 25, 9680-9685 (2005).
Antunes, R. & Moita, M. A. Discriminative auditory fear learning requires both tuned and
nontuned auditory pathways to the amygdala. The Journal of neuroscience: the official
journal of the Society for Neuroscience 30, 9782-9787 (2010).
Ascoli, G. A. et al. Petilla terminology: nomenclature of features of GABAergic interneurons
of the cerebral cortex. Nature reviews. Neuroscience 9, 557-568 (2008).

- 80 -
Atallah, B. V., Bruns, W., Carandini, M. & Scanziani, M. Parvalbumin-expressing
interneurons linearly transform cortical responses to visual stimuli. Neuron 73, 159-170
(2012).
Audinat, E., Conde, F. & Crepel, F. Cortico-cortical connections of the limbic cortex of the
rat. Experimental brain research. Experimentelle Hirnforschung. Experimentation cerebrale
69, 439-443 (1988).
Bacon, S. J., Headlam, A. J., Gabbott, P. L. & Smith, A. D. Amygdala input to medial
prefrontal cortex (mPFC) in the rat: a light and electron microscope study. Brain research
720, 211-219 (1996).
Baeg, E. H. et al. Fast spiking and regular spiking neural correlates of fear conditioning in the
medial prefrontal cortex of the rat. Cereb Cortex 11, 441-451 (2001).
Bakin, J. S. & Weinberger, N. M. Induction of a physiological memory in the cerebral cortex
by stimulation of the nucleus basalis. Proceedings of the National Academy of Sciences of the
United States of America 93, 11219-11224 (1996).
Bannister, A. P. Inter- and intra-laminar connections of pyramidal cells in the neocortex.
Neuroscience research 53, 95-103 (2005).
Baran, S. E., Armstrong, C. E., Niren, D. C. & Conrad, C. D. Prefrontal cortex lesions and sex
differences in fear extinction and perseveration. Learn Mem 17, 267-278 (2010).
Bartos, M., Vida, I. & Jonas, P. Synaptic mechanisms of synchronized gamma oscillations in
inhibitory interneuron networks. Nature reviews. Neuroscience 8, 45-56 (2007).
Beckstead, R. M. An autoradiographic examination of corticocortical and subcortical
projections of the mediodorsal-projection (prefrontal) cortex in the rat. The Journal of
comparative neurology 184, 43-62 (1979).
Benchenane, K. et al. Coherent theta oscillations and reorganization of spike timing in the
hippocampal- prefrontal network upon learning. Neuron 66, 921-936 (2010).
Benchenane, K., Tiesinga, P. H. & Battaglia, F. P. Oscillations in the prefrontal cortex: a
gateway to memory and attention. Current opinion in neurobiology 21, 475-485 (2011).
Berendse, H. W., Galis-de Graaf, Y. & Groenewegen, H. J. Topographical organization and
relationship with ventral striatal compartments of prefrontal corticostriatal projections in the
rat. The Journal of comparative neurology 316, 314-347 (1992).
Berretta, S., Pantazopoulos, H., Caldera, M., Pantazopoulos, P. & Pare, D. Infralimbic cortex
activation increases c-Fos expression in intercalated neurons of the amygdala. Neuroscience
132, 943-953 (2005).

- 81 -
Bienvenu, T.C. et al. Cell-type-specific recruitment of amygdala interneurons to hippocampal
theta rhythm and noxious stimuli in vivo. Neuron 74, 1059-1074 (2012).
Bissiere, S. et al. The rostral anterior cingulate cortex modulates the efficiency of amygdala-
dependent fear learning. Biol Psychiatry 63, 821-831 (2008).
Blair, H. T., Schafe, G. E., Bauer, E. P., Rodrigues, S. M. & LeDoux, J. E. Synaptic plasticity
in the lateral amygdala: a cellular hypothesis of fear conditioning. Learn Mem 8, 229-242
(2001).
Blatow, M. et al. A novel network of multipolar bursting interneurons generates theta
frequency oscillations in neocortex. Neuron 38, 805-817 (2003).
Bliss, T. V. & Collingridge, G. L. A synaptic model of memory: long-term potentiation in the
hippocampus. Nature 361, 31-39 (1993).
Boatman, J. A. & Kim, J. J. A thalamo-cortico-amygdala pathway mediates auditory fear
conditioning in the intact brain. The European journal of neuroscience 24, 894-900 (2006).
Bouton, M. E. & King, D. A. Contextual control of the extinction of conditioned fear: tests for
the associative value of the context. Journal of experimental psychology. Animal behaviour
processes 9, 248-265 (1983).
Bouton, M. E., Garcia-Gutierrez, A., Zilski, J. & Moody, E. W. Extinction in multiple
contexts does not necessarily make extinction less vulnerable to relapse. Behaviour research
and therapy 44, 983-994 (2006).
Brown, S. P. & Hestrin, S. Intracortical circuits of pyramidal neurons reflect their long-range
axonal targets. Nature 457, 1133-1136 (2009).
Burgos-Robles, A., Vidal-Gonzalez, I. & Quirk, G. J. Sustained conditioned responses in
prelimbic prefrontal neurons are correlated with fear expression and extinction failure. The
Journal of neuroscience: the official journal of the Society for Neuroscience 29, 8474-8482
(2009).
Burgos-Robles, A., Vidal-Gonzalez, I., Santini, E. & Quirk, G. J. Consolidation of fear
extinction requires NMDA receptor-dependent bursting in the ventromedial prefrontal cortex.
Neuron 53, 871-880 (2007).
Burkhalter, A. Many specialists for suppressing cortical excitation. Front Neurosci 2, 155-167
(2008).
Busti, D. et al. Different fear states engage distinct networks within the intercalated cell
clusters of the amygdala. The Journal of neuroscience: the official journal of the Society for
Neuroscience 31, 5131-5144 (2011).

- 82 -
Buzsaki, G. & Chrobak, J. J. Temporal structure in spatially organized neuronal ensembles: a
role for interneuronal networks. Current opinion in neurobiology 5, 504-510 (1995).
Buzsaki, G. & Draguhn, A. Neuronal oscillations in cortical networks. Science 304, 1926-
1929 (2004).
Buzsaki, G. & Wang, X. J. Mechanisms of gamma oscillations. Annual review of
neuroscience 35, 203-225 (2012).
Buzsaki, G., Anastassiou, C. A. & Koch, C. The origin of extracellular fields and currents--
EEG, ECoG, LFP and spikes. Nature reviews. Neuroscience 13 (2012).
Buzsaki, G., Geisler, C., Henze, D. A. & Wang, X. J. Interneuron Diversity series: Circuit
complexity and axon wiring economy of cortical interneurons. Trends in neurosciences 27,
186-193 (2004).
Buzsaki, G., Grastyan, E., Tveritskaya, I. N. & Czopf, J. Hippocampal evoked potentials and
EEG changes during classical conditioning in the rat. Electroencephalography and clinical
neurophysiology 47, 64-74 (1979).
Campeau, S. & Davis, M. Involvement of subcortical and cortical afferents to the lateral
nucleus of the amygdala in fear conditioning measured with fear-potentiated startle in rats
trained concurrently with auditory and visual conditioned stimuli. The Journal of
neuroscience: the official journal of the Society for Neuroscience 15, 2312-2327 (1995).
Cannich, A. et al. CB1 cannabinoid receptors modulate kinase and phosphatase activity
during extinction of conditioned fear in mice. Learn Mem 11, 625-632 (2004).
Canolty, R. T. et al. High gamma power is phase-locked to theta oscillations in human
neocortex. Science 313, 1626-1628 (2006).
Cardin, J. A. et al. Driving fast-spiking cells induces gamma rhythm and controls sensory
responses. Nature 459, 663-667 (2009).
Carlen, M. et al. A critical role for NMDA receptors in parvalbumin interneurons for gamma
rhythm induction and behaviour. Molecular psychiatry 17, 537-548 (2012).
Carlyon, R. P. How the brain separates sounds. Trends in cognitive sciences 8, 465-471
(2004).
Carr, D. B. & Sesack, S. R. Hippocampal afferents to the rat prefrontal cortex: synaptic
targets and relation to dopamine terminals. The Journal of comparative neurology 369, 1-15
(1996).
Cassell, M. D. & Wright, D. J. Topography of projections from the medial prefrontal cortex to
the amygdala in the rat. Brain research bulletin 17, 321-333 (1986).

- 83 -
Cauli, B. et al. Molecular and physiological diversity of cortical nonpyramidal cells. The
Journal of neuroscience: the official journal of the Society for Neuroscience 17, 3894-3906
(1997).
Caviness, V. S., Jr. Architectonic map of neocortex of the normal mouse. The Journal of
comparative neurology 164, 247-263 (1975).
Chandler, D. J., Lamperski, C. S. & Waterhouse, B. D. Identification and distribution of
projections from monoaminergic and cholinergic nuclei to functionally differentiated
subregions of prefrontal cortex. Brain research 1522, 38-58 (2013).
Chang, C. H. & Maren, S. Strain difference in the effect of infralimbic cortex lesions on fear
extinction in rats. Behavioural neuroscience 124, 391-397 (2010).
Chang, C. H., Berke, J. D. & Maren, S. Single-unit activity in the medial prefrontal cortex
during immediate and delayed extinction of fear in rats. PloS one 5, e11971 (2010).
Chang, Y. M. & Luebke, J. I. Electrophysiological diversity of layer 5 pyramidal cells in the
prefrontal cortex of the rhesus monkey: in vitro slice studies. Journal of neurophysiology 98,
2622-2632 (2007).
Chiu, C. Q. et al. Compartmentalization of GABAergic inhibition by dendritic spines. Science
340, 759-762 (2013).
Choi, D. C. et al. Prelimbic cortical BDNF is required for memory of learned fear but not
extinction or innate fear. Proceedings of the National Academy of Sciences of the United
States of America 107, 2675-2680 (2010).
Ciocchi, S. et al. Encoding of conditioned fear in central amygdala inhibitory circuits. Nature
468, 277-282 (2010).
Cobb, S. R., Buhl, E. H., Halasy, K., Paulsen, O. & Somogyi, P. Synchronization of neuronal
activity in hippocampus by individual GABAergic interneurons. Nature 378, 75-78 (1995).
Corcoran, K. A. & Quirk, G. J. Activity in prelimbic cortex is necessary for the expression of
learned, but not innate, fears. The Journal of neuroscience: the official journal of the Society
for Neuroscience 27, 840-844 (2007).
Corcoran, K. A. & Quirk, G. J. Activity in prelimbic cortex is necessary for the expression of
learned, but not innate, fears. The Journal of neuroscience: the official journal of the Society
for Neuroscience 27, 840-844 (2007).
Courtin, J., Bienvenu, T. C., Einarsson, E. O. & Herry, C. Medial prefrontal cortex neuronal
circuits in fear behavior. Neuroscience 240, 219-242 (2013).
Csicsvari, J. et al. Massively parallel recording of unit and local field potentials with silicon-
based electrodes. Journal of neurophysiology 90, 1314-1323 (2003).

- 84 -
DeFelipe, J. & Farinas, I. The pyramidal neuron of the cerebral cortex: morphological and
chemical characteristics of the synaptic inputs. Prog Neurobiol 39, 563-607 (1992).
DeFelipe, J. et al. New insights into the classification and nomenclature of cortical
GABAergic interneurons. Nature reviews. Neuroscience 14, 202-216 (2013).
DeFelipe, J. Types of neurons, synaptic connections and chemical characteristics of cells
immunoreactive for calbindin-D28K, parvalbumin and calretinin in the neocortex. J Chem
Neuroanat 14, 1-19 (1997).
DeFelipe, J., Hendry, S. H. & Jones, E. G. Visualization of chandelier cell axons by
parvalbumin immunoreactivity in monkey cerebral cortex. Proceedings of the National
Academy of Sciences of the United States of America 86, 2093-2097 (1989).
Degenetais, E., Thierry, A. M., Glowinski, J. & Gioanni, Y. Electrophysiological properties of
pyramidal neurons in the rat prefrontal cortex: an in vivo intracellular recording study. Cereb
Cortex 12, 1-16 (2002).
Di Scala, G., Mana, M. J., Jacobs, W. J. & Phillips, A. G. Evidence of Pavlovian conditioned
fear following electrical stimulation of the periaqueductal grey in the rat. Physiology &
behaviour 40, 55-63 (1987).
Dilgen, J., Tejeda, H.A., & O’Donnell, P. Amygdala inputs drive feedforward inhibition in
the medial prefrontal cortex. Journal of Neurophysiology 110, 221-229 (2013).
Donoghue, J. P. & Wise, S. P. The motor cortex of the rat: cytoarchitecture and
microstimulation mapping. The Journal of comparative neurology 212, 76-88 (1982).
Doucet, J. R., Molavi, D. L. & Ryugo, D. K. The source of corticocollicular and corticobulbar
projections in area Te1 of the rat. Experimental brain research. Experimentelle
Hirnforschung. Experimentation cerebrale 153, 461-466 (2003).
Douglas, R. J. & Martin, K. A. Neuronal circuits of the neocortex. Annual review of
neuroscience 27, 419-451 (2004).
Doyere, V., Schafe, G. E., Sigurdsson, T. & LeDoux, J. E. Long-term potentiation in freely
moving rats reveals asymmetries in thalamic and cortical inputs to the lateral amygdala. The
European journal of neuroscience 17, 2703-2715 (2003).
Druga, R. Neocortical inhibitory system. Folia biologica 55, 201-217 (2009).
Edeline, J. M., Hars, B., Maho, C. & Hennevin, E. Transient and prolonged facilitation of
tone-evoked responses induced by basal forebrain stimulations in the rat auditory cortex.
Experimental brain research. Experimentelle Hirnforschung. Experimentation cerebrale 97,
373-386 (1994).

- 85 -
Ehret, G. The auditory cortex. Journal of comparative physiology. A, Sensory, neural, and
behavioural physiology 181, 547-557 (1997).
Einevoll, G. T., Franke, F., Hagen, E., Pouzat, C. & Harris, K. D. Towards reliable spike-train
recordings from thousands of neurons with multielectrodes. Current opinion in neurobiology
22, 11-17 (2012).
Engel, A. K., Fries, P. & Singer, W. Dynamic predictions: oscillations and synchrony in top-
down processing. Nature reviews. Neuroscience 2, 704-716 (2001).
Fanselow, M. S. & LeDoux, J. E. Why we think plasticity underlying Pavlovian fear
conditioning occurs in the basolateral amygdala. Neuron 23, 229-232 (1999).
Farinelli, M., Deschaux, O., Hugues, S., Thevenet, A. & Garcia, R. Hippocampal train
stimulation modulates recall of fear extinction independently of prefrontal cortex synaptic
plasticity and lesions. Learn Mem 13, 329-334 (2006).
Fell, J. & Axmacher, N. The role of phase synchronization in memory processes. Nature
reviews. Neuroscience 12, 105-118 (2011).
Fernandez Espejo, E. Prefrontocortical dopamine loss in rats delays long-term extinction of
contextual conditioned fear, and reduces social interaction without affecting short-term social
interaction memory. Neuropsychopharmacology: official publication of the American College
of Neuropsychopharmacology 28, 490-498 (2003).
Floresco, S. B. & Tse, M. T. Dopaminergic regulation of inhibitory and excitatory
transmission in the basolateral amygdala-prefrontal cortical pathway. The Journal of
neuroscience: the official journal of the Society for Neuroscience 27, 2045-2057 (2007).
Floyd, N. S., Price, J. L., Ferry, A. T., Keay, K. A. & Bandler, R. Orbitomedial prefrontal
cortical projections to distinct longitudinal columns of the periaqueductal gray in the rat. The
Journal of comparative neurology 422, 556-578 (2000).
Floyd, N. S., Price, J. L., Ferry, A. T., Keay, K. A. & Bandler, R. Orbitomedial prefrontal
cortical projections to hypothalamus in the rat. The Journal of comparative neurology 432,
307-328 (2001).
Fontanez-Nuin, D. E., Santini, E., Quirk, G. J. & Porter, J. T. Memory for fear extinction
requires mGluR5-mediated activation of infralimbic neurons. Cereb Cortex 21, 727-735
(2011).
Franklin, K. J. B. P., G. The Mouse Brain in Stereotaxic Coordinates (Academic, 1997).
Freund, T. F. & Buzsaki, G. Interneurons of the hippocampus. Hippocampus 6, 347-470
(1996).

- 86 -
Freund, T. F. & Katona, I. Perisomatic inhibition. Neuron 56, 33-42 (2007).
Freund, T. F. Interneuron Diversity series: Rhythm and mood in perisomatic inhibition.
Trends in neurosciences 26, 489-495 (2003).
Freund, T. F., Magloczky, Z., Soltesz, I. & Somogyi, P. Synaptic connections, axonal and
dendritic patterns of neurons immunoreactive for cholecystokinin in the visual cortex of the
cat. Neuroscience 19, 1133-1159 (1986).
Friedrich, R. W., Habermann, C. J. & Laurent, G. Multiplexing using synchrony in the
zebrafish olfactory bulb. Nature neuroscience 7, 862-871 (2004).
Fries, P., Nikolic, D. & Singer, W. The gamma cycle. Trends in neurosciences 30, 309-316
(2007).
Frisina, R. D., & Walton, J. P. Neuroanatomy of the mouse auditory system. In: Handbook of
Mouse Auditory Res: From Behaviour to Molecular Biology (ed. Willott JP), pp 243-77. CRC
Press, New York (2001).
Froemke, R. C., Merzenich, M. M. & Schreiner, C. E. A synaptic memory trace for cortical
receptive field plasticity. Nature 450, 425-429 (2007).
Frysztak, R. J. & Neafsey, E. J. The effect of medial frontal cortex lesions on respiration,
"freezing," and ultrasonic vocalizations during conditioned emotional responses in rats. Cereb
Cortex 1, 418-425 (1991).
Fuchs, E. C. et al. Recruitment of parvalbumin-positive interneurons determines hippocampal
function and associated behaviour. Neuron 53, 591-604 (2007).
Fukuda, T. Structural organization of the gap junction network in the cerebral cortex. The
Neuroscientist : a review journal bringing neurobiology, neurology and psychiatry 13, 199-
207 (2007).
Fuster, J.M. The Prefrontal Cortex (Fourth Edition). Academic Press, London (2008).
Gabbott, P. et al. Amygdala afferents monosynaptically innervate corticospinal neurons in
rat medial prefrontal cortex. The Journal of comparative neurology 520, 2440-2458 (2012).
Gabbott, P. L. & Bacon, S. J. Calcineurin immunoreactivity in prelimbic cortex (area 32) of
the rat. Brain research 747, 352-356 (1997).
Gabbott, P. L., Warner, T. A., Jays, P. R., Salway, P. & Busby, S. J. Prefrontal cortex in the
rat: projections to subcortical autonomic, motor, and limbic centers. The Journal of
comparative neurology 492, 145-177 (2005).

- 87 -
Galarreta, M. & Hestrin, S. Electrical and chemical synapses among parvalbumin fast-spiking
GABAergic interneurons in adult mouse neocortex. Proceedings of the National Academy of
Sciences of the United States of America 99, 12438-12443 (2002).
Gale, G. D. et al. Role of the basolateral amygdala in the storage of fear memories across the
adult lifetime of rats. The Journal of neuroscience: the official journal of the Society for
Neuroscience 24, 3810-3815 (2004).
Garcia, R., Chang, C. H. & Maren, S. Electrolytic lesions of the medial prefrontal cortex do
not interfere with long-term memory of extinction of conditioned fear. Learn Mem 13, 14-17
(2006).
Gentet, L. J. Functional diversity of supragranular GABAergic neurons in the barrel cortex.
Frontiers in neural circuits 6, 52 (2012).
Geracitano, R., Kaufmann, W. A., Szabo, G., Ferraguti, F. & Capogna, M. Synaptic
heterogeneity between mouse paracapsular intercalated neurons of the amygdala. The Journal
of physiology 585, 117-134 (2007).
Gewirtz, J. C., Falls, W. A. & Davis, M. Normal conditioned inhibition and extinction of
freezing and fear-potentiated startle following electrolytic lesions of medical prefrontal cortex
in rats. Behavioural neuroscience 111, 712-726 (1997).
Gibson, J. R., Beierlein, M. & Connors, B. W. Two networks of electrically coupled
inhibitory neurons in neocortex. Nature 402, 75-79 (1999).
Gigg, J., Tan, A. M. & Finch, D. M. Glutamatergic hippocampal formation projections to
prefrontal cortex in the rat are regulated by GABAergic inhibition and show convergence with
glutamatergic projections from the limbic thalamus. Hippocampus 4, 189-198 (1994).
Gonchar, Y. A., Johnson, P. B. & Weinberg, R. J. GABA-immunopositive neurons in rat
neocortex with contralateral projections to S-I. Brain research 697, 27-34 (1995).
Goosens, K. A., Hobin, J. A. & Maren, S. Auditory-evoked spike firing in the lateral
amygdala and Pavlovian fear conditioning: mnemonic code or fear bias? Neuron 40, 1013-
1022 (2003).
Gray, C. M. & Singer, W. Stimulus-specific neuronal oscillations in orientation columns of
cat visual cortex. Proceedings of the National Academy of Sciences of the United States of
America 86, 1698-1702 (1989).
Gray, E. G. Axo-somatic and axo-dendritic synapses of the cerebral cortex: an electron
microscope study. Journal of anatomy 93, 420-433 (1959).
Groenewegen, H. J. Organization of the afferent connections of the mediodorsal thalamic
nucleus in the rat, related to the mediodorsal-prefrontal topography. Neuroscience 24, 379-
431 (1988).

- 88 -
Groenewegen, H. J., Berendse, H. W., Wolters, J. G. & Lohman, A. H. The anatomical
relationship of the prefrontal cortex with the striatopallidal system, the thalamus and the
amygdala: evidence for a parallel organization. Progress in brain research 85, 95-116;
discussion 116-118 (1990).
Groenewegen, H. J., Wright, C. I. & Uylings, H. B. The anatomical relationships of the
prefrontal cortex with limbic structures and the basal ganglia. J Psychopharmacol 11, 99-106
(1997).
Guldin, W. O., Pritzel, M. & Markowitsch, H. J. Prefrontal cortex of the mouse defined as
cortical projection area of the thalamic mediodorsal nucleus. Brain, behaviour and evolution
19, 93-107 (1981).
Gupta, A., Wang, Y. & Markram, H. Organizing principles for a diversity of GABAergic
interneurons and synapses in the neocortex. Science 287, 273-278 (2000).
Harris, K. D., Csicsvari, J., Hirase, H., Dragoi, G. & Buzsaki, G. Organization of cell
assemblies in the hippocampus. Nature 424, 552-556 (2003).
Hars, B., Maho, C., Edeline, J. M. & Hennevin, E. Basal forebrain stimulation facilitates tone-
evoked responses in the auditory cortex of awake rat. Neuroscience 56, 61-74 (1993).
Hartwich, K., Pollak, T. & Klausberger, T. Distinct firing patterns of identified basket and
dendrite-targeting interneurons in the prefrontal cortex during hippocampal theta and local
spindle oscillations. The Journal of neuroscience: the official journal of the Society for
Neuroscience 29, 9563-9574 (2009).
Hazama, M., Kimura, A., Donishi, T., Sakoda, T. & Tamai, Y. Topography of corticothalamic
projections from the auditory cortex of the rat. Neuroscience 124, 655-667 (2004).
Headley, D. B. & Weinberger, N. M. Gamma-band activation predicts both associative
memory and cortical plasticity. The Journal of neuroscience: the official journal of the
Society for Neuroscience 31, 12748-12758 (2011).
Headley, D. B. & Weinberger, N. M. Fear conditioning enhances gamma oscillations and
their entrainment of neurons representing the conditioned stimulus. The Journal of
neuroscience: the official journal of the Society for Neuroscience 33, 5705-5717 (2013).
Hebb D.O., The organization of behaviour: a neuropsychological theory, Wiley, New York,
(1949).
Hefner, K. et al. Impaired fear extinction learning and cortico-amygdala circuit abnormalities
in a common genetic mouse strain. The Journal of neuroscience: the official journal of the
Society for Neuroscience 28, 8074-8085 (2008).

- 89 -
Heidbreder, C. A. & Groenewegen, H. J. The medial prefrontal cortex in the rat: evidence for
a dorso-ventral distinction based upon functional and anatomical characteristics.
Neuroscience and biobehavioural reviews 27, 555-579 (2003).
Herry, C. & Mons, N. Resistance to extinction is associated with impaired immediate early
gene induction in medial prefrontal cortex and amygdala. The European journal of
neuroscience 20, 781-790 (2004).
Herry, C. et al. Neuronal circuits of fear extinction. Eur J Neurosci 31, 599-612 (2010).
Herry, C. et al. Switching on and off fear by distinct neuronal circuits. Nature 454, 600-606
(2008).
Herry, C., Trifilieff, P., Micheau, J., Luthi, A. & Mons, N. Extinction of auditory fear
conditioning requires MAPK/ERK activation in the basolateral amygdala. Eur J Neurosci 24,
261-269 (2006).
Hestrin, S. & Galarreta, M. Electrical synapses define networks of neocortical GABAergic
neurons. Trends in neurosciences 28, 304-309 (2005).
Hevner, R. F. et al. Beyond laminar fate: toward a molecular classification of cortical
projection/pyramidal neurons. Developmental neuroscience 25, 139-151 (2003).
Hikind, N. & Maroun, M. Microinfusion of the D1 receptor antagonist, SCH23390 into the IL
but not the BLA impairs consolidation of extinction of auditory fear conditioning.
Neurobiology of learning and memory 90, 217-222 (2008).
Hippenmeyer, S. et al. A developmental switch in the response of DRG neurons to ETS
transcription factor signaling. PLoS biology 3, e159 (2005).
Hirel, J. et al. The hippocampo-cortical loop: spatio-temporal learning and goal-oriented
planning in navigation. Neural networks: the official journal of the International Neural
Network Society 43, 8-21 (2013).
Hobin, J. A., Goosens, K. A. & Maren, S. Context-dependent neuronal activity in the lateral
amygdala represents fear memories after extinction. The Journal of neuroscience: the official
journal of the Society for Neuroscience 23, 8410-8416 (2003).
Hok, V., Chah, E., Save, E. & Poucet, B. Prefrontal cortex focally modulates hippocampal
place cell firing patterns. The Journal of neuroscience: the official journal of the Society for
Neuroscience 33, 3443-3451 (2013).
Holmes, A. et al. Chronic alcohol remodels prefrontal neurons and disrupts NMDAR-
mediated fear extinction encoding. Nature neuroscience 15, 1359-1361 (2012).
Holson, R. R. Mesial prefrontal cortical lesions and timidity in rats. I. Reactivity to aversive
stimuli. Physiology & behaviour 37, 221-230 (1986).

- 90 -
Hopkins, D. A. & Holstege, G. Amygdaloid projections to the mesencephalon, pons and
medulla oblongata in the cat. Exp Brain Res 32, 529-547 (1978).
Howard, A., Tamas, G. & Soltesz, I. Lighting the chandelier: new vistas for axo-axonic cells.
Trends in neurosciences 28, 310-316 (2005).
Hu, B., Senatorov, V. & Mooney, D. Lemniscal and non-lemniscal synaptic transmission in
rat auditory thalamus. The Journal of physiology 479 ( Pt 2), 217-231 (1994).
Hubel, D. H. & Wiesel, T. N. Receptive fields, binocular interaction and functional
architecture in the cat's visual cortex. The Journal of physiology 160, 106-154 (1962).
Huggenberger, S., Vater, M. & Deisz, R. A. Interlaminar differences of intrinsic properties of
pyramidal neurons in the auditory cortex of mice. Cereb Cortex 19, 1008-1018 (2009).
Hugues, S., Chessel, A., Lena, I., Marsault, R. & Garcia, R. Prefrontal infusion of PD098059
immediately after fear extinction training blocks extinction-associated prefrontal synaptic
plasticity and decreases prefrontal ERK2 phosphorylation. Synapse 60, 280-287 (2006).
Hugues, S., Deschaux, O. & Garcia, R. Postextinction infusion of a mitogen-activated protein
kinase inhibitor into the medial prefrontal cortex impairs memory of the extinction of
conditioned fear. Learn Mem 11, 540-543 (2004).
Isaacson, J. S. & Scanziani, M. How inhibition shapes cortical activity. Neuron 72, 231-243
(2011).
Ishikane, H., Gangi, M., Honda, S. & Tachibana, M. Synchronized retinal oscillations encode
essential information for escape behaviour in frogs. Nature neuroscience 8, 1087-1095
(2005).
Jarrell, T. W., Gentile, C. G., Romanski, L. M., McCabe, P. M. & Schneiderman, N.
Involvement of cortical and thalamic auditory regions in retention of differential bradycardiac
conditioning to acoustic conditioned stimuli in rabbits. Brain research 412, 285-294 (1987).
Jay, T. M., Glowinski, J. & Thierry, A. M. Selectivity of the hippocampal projection to the
prelimbic area of the prefrontal cortex in the rat. Brain research 505, 337-340 (1989).
Ji, W. & Suga, N. Development of reorganization of the auditory cortex caused by fear
conditioning: effect of atropine. Journal of neurophysiology 90, 1904-1909 (2003).
Ji, W., Gao, E. & Suga, N. Effects of acetylcholine and atropine on plasticity of central
auditory neurons caused by conditioning in bats. Journal of neurophysiology 86, 211-225
(2001).
Johansen, J. P. et al. Optical activation of lateral amygdala pyramidal cells instructs
associative fear learning. Proceedings of the National Academy of Sciences of the United
States of America 107, 12692-12697 (2010).

- 91 -
Johansen, J. P., Cain, C. K., Ostroff, L. E. & LeDoux, J. E. Molecular mechanisms of fear
learning and memory. Cell 147, 509-524 (2011).
Karube, F., Kubota, Y. & Kawaguchi, Y. Axon branching and synaptic bouton phenotypes
in GABAergic nonpyramidal cell subtypes. The Journal of neuroscience: the official journal
of the Society for Neuroscience 24, 2853-2865 (2004).
Kawaguchi, Y. & Kubota Y. GABAergic cell subtypes and their synaptic connections in rat
frontal cortex. Cereb Cortex. 7, 476-86. (1997)
Kawaguchi, Y. & Kubota, Y. Correlation of physiological subgroupings of nonpyramidal
cells with parvalbumin- and calbindinD28k-immunoreactive neurons in layer V of rat frontal
cortex. Journal of neurophysiology 70, 387-396 (1993).
Kawaguchi, Y. & Kubota, Y. Physiological and morphological identification of somatostatin-
or vasoactive intestinal polypeptide-containing cells among GABAergic cell subtypes in rat
frontal cortex. The Journal of neuroscience: the official journal of the Society for
Neuroscience 16, 2701-2715 (1996).
Kawaguchi, Y. Physiological subgroups of nonpyramidal cells with specific morphological
characteristics in layer II/III of rat frontal cortex. The Journal of neuroscience: the official
journal of the Society for Neuroscience 15, 2638-2655 (1995).
Kim, S. C., Jo, Y. S., Kim, I. H., Kim, H. & Choi, J. S. Lack of medial prefrontal cortex
activation underlies the immediate extinction deficit. The Journal of neuroscience: the official
journal of the Society for Neuroscience 30, 832-837 (2010).
Kimura, A., Donishi, T., Okamoto, K. & Tamai, Y. Efferent connections of "posterodorsal"
auditory area in the rat cortex: implications for auditory spatial processing. Neuroscience 128,
399-419 (2004).
Kimura, A., Donishi, T., Okamoto, K., Imbe, H. & Tamai, Y. Efferent connections of the
ventral auditory area in the rat cortex: implications for auditory processing related to emotion.
The European journal of neuroscience 25, 2819-2834 (2007).
Klavir, O., Genud-Gabai, R. & Paz, R. Low-frequency stimulation depresses the primate
anterior-cingulate-cortex and prevents spontaneous recovery of aversive memories. The
Journal of neuroscience: the official journal of the Society for Neuroscience 32, 8589-8597
(2012).
Knapska, E. & Maren, S. Reciprocal patterns of c-Fos expression in the medial prefrontal
cortex and amygdala after extinction and renewal of conditioned fear. Learn Mem 16, 486-
493 (2009).

- 92 -
Knapska, E. et al. Functional anatomy of neural circuits regulating fear and extinction.
Proceedings of the National Academy of Sciences of the United States of America 109, 17093-
17098 (2012).
Krettek, J. E. & Price, J. L. The cortical projections of the mediodorsal nucleus and adjacent
thalamic nuclei in the rat. The Journal of comparative neurology 171, 157-191 (1977).
Krimer, L. S. et al. Cluster analysis-based physiological classification and morphological
properties of inhibitory neurons in layers 2-3 of monkey dorsolateral prefrontal cortex.
Journal of neurophysiology 94, 3009-3022 (2005).
Krook-Magnuson, E., Varga, C., Lee, S. H. & Soltesz, I. New dimensions of interneuronal
specialization unmasked by principal cell heterogeneity. Trends in neurosciences 35, 175-184
(2012).
Kruglikov, I. & Rudy, B. Perisomatic GABA release and thalamocortical integration onto
neocortical excitatory cells are regulated by neuromodulators. Neuron 58, 911-924 (2008).
Kubota, Y. & Kawaguchi, Y. Two distinct subgroups of cholecystokinin-immunoreactive
cortical interneurons. Brain research 752, 175-183 (1997).
Kubota, Y., Hattori, R. & Yui, Y. Three distinct subpopulations of GABAergic neurons in rat
frontal agranular cortex. Brain research 649, 159-173 (1994).
Kuhlman, S. J. et al. A disinhibitory microcircuit initiates critical-period plasticity in the
visual cortex. Nature 501, 543-546 (2013).
Kuroda, M., Murakami, K., Kishi, K. & Price, J. L. Thalamocortical synapses between axons
from the mediodorsal thalamic nucleus and pyramidal cells in the prelimbic cortex of the rat.
The Journal of comparative neurology 356, 143-151 (1995a).
Kuroda, M., Murakami, K., Oda, S., Shinkai, M. & Kishi, K. Direct synaptic connections
between thalamocortical axon terminals from the mediodorsal thalamic nucleus (MD) and
corticothalamic neurons to MD in the prefrontal cortex. Brain research 612, 339-344 (1993).
Kuroda, M., Murakami, K., Shinkai, M., Ojima, H. & Kishi, K. Electron microscopic
evidence that axon terminals from the mediodorsal thalamic nucleus make direct synaptic
contacts with callosal cells in the prelimbic cortex of the rat. Brain research 677, 348-353
(1995b).
Kvitsiani, D. et al. Distinct behavioural and network correlates of two interneuron types in
prefrontal cortex. Nature 498, 363-366 (2013).
Lacroix, L., Spinelli, S., Heidbreder, C. A. & Feldon, J. Differential role of the medial and
lateral prefrontal cortices in fear and anxiety. Behavioural neuroscience 114, 1119-1130
(2000).

- 93 -
Laurent, V. & Westbrook, R. F. Inactivation of the infralimbic but not the prelimbic cortex
impairs consolidation and retrieval of fear extinction. Learn Mem 16, 520-529 (2009).
Lauzon, N. M., Bishop, S. F. & Laviolette, S. R. Dopamine D1 versus D4 receptors
differentially modulate the encoding of salient versus nonsalient emotional information in the
medial prefrontal cortex. The Journal of neuroscience: the official journal of the Society for
Neuroscience 29, 4836-4845 (2009).
Laviolette, S. R. & Grace, A. A. Cannabinoids Potentiate Emotional Learning Plasticity in
Neurons of the Medial Prefrontal Cortex through Basolateral Amygdala Inputs. The Journal
of neuroscience: the official journal of the Society for Neuroscience 26, 6458-6468 (2006).
Laviolette, S. R., Lipski, W. J. & Grace, A. A. A subpopulation of neurons in the medial
prefrontal cortex encodes emotional learning with burst and frequency codes through a
dopamine D4 receptor-dependent basolateral amygdala input. The Journal of neuroscience:
the official journal of the Society for Neuroscience 25, 6066-6075 (2005).
Lebron, K., Milad, M. R. & Quirk, G. J. Delayed recall of fear extinction in rats with lesions
of ventral medial prefrontal cortex. Learn Mem 11, 544-548 (2004).
LeDoux, J. E. Emotion circuits in the brain. Annual review of neuroscience 23, 155-184
(2000).
LeDoux, J. E., Farb, C. R. & Romanski, L. M. Overlapping projections to the amygdala and
striatum from auditory processing areas of the thalamus and cortex. Neuroscience letters 134,
139-144 (1991).
LeDoux, J. E., Sakaguchi, A. & Reis, D. J. Subcortical efferent projections of the medial
geniculate nucleus mediate emotional responses conditioned to acoustic stimuli. The Journal
of neuroscience: the official journal of the Society for Neuroscience 4, 683-698 (1984).
Lee, C. C. & Sherman, S. M. On the classification of pathways in the auditory midbrain,
thalamus, and cortex. Hearing research 276, 79-87 (2011).
Lee, Y. K. & Choi, J. S. Inactivation of the medial prefrontal cortex interferes with the
expression but not the acquisition of differential fear conditioning in rats. Exp Neurobiol 21,
23-29 (2012).
Leonard, C. M. The prefrontal cortex of the rat. I. Cortical projection of the mediodorsal
nucleus. II. Efferent connections. Brain research 12, 321-343 (1969).
Lesting, J. et al. Patterns of coupled theta activity in amygdala-hippocampal-prefrontal
cortical circuits during fear extinction. PloS one 6, e21714 (2011).
Letinic, K., Zoncu, R. & Rakic, P. Origin of GABAergic neurons in the human neocortex.
Nature 417, 645-649 (2002).

- 94 -
Letzkus, J. J. et al. A disinhibitory microcircuit for associative fear learning in the auditory
cortex. Nature 480, 331-335 (2011).
Likhtik, E., Pelletier, J. G., Paz, R. & Pare, D. Prefrontal control of the amygdala. The Journal
of neuroscience: the official journal of the Society for Neuroscience 25, 7429-7437 (2005).
Lin, H. C., Mao, S. C., Su, C. L. & Gean, P. W. The role of prefrontal cortex CB1 receptors in
the modulation of fear memory. Cereb Cortex 19, 165-175 (2009).
Little, J. P. & Carter, A. G. Subcellular synaptic connectivity of layer 2 pyramidal neurons in
the medial prefrontal cortex. The Journal of neuroscience: the official journal of the Society
for Neuroscience 32, 12808-12819 (2012).
Livneh, U. & Paz, R. Amygdala-prefrontal synchronization underlies resistance to extinction
of aversive memories. Neuron 75, 133-142 (2012).
Llano, D. A. & Sherman, S. M. Evidence for nonreciprocal organization of the mouse
auditory thalamocortical-corticothalamic projection systems. The Journal of comparative
neurology 507, 1209-1227 (2008).
Losonczy, A., Zemelman, B. V., Vaziri, A. & Magee, J. C. Network mechanisms of theta
related neuronal activity in hippocampal CA1 pyramidal neurons. Nature neuroscience 13,
967-972 (2010).
Lovett-Barron, M. et al. Regulation of neuronal input transformations by tunable dendritic
inhibition. Nature neuroscience 15, 423-430, S421-423 (2012).
MacLeod, K., Backer, A. & Laurent, G. Who reads temporal information contained across
synchronized and oscillatory spike trains? Nature 395, 693-698 (1998).
Madison, D. V., Malenka, R. C. & Nicoll, R. A. Mechanisms underlying long-term
potentiation of synaptic transmission. Annual review of neuroscience 14, 379-397 (1991).
Maher, B. A. & McIntire, R. W. The extinction of the CER following frontal ablation.
Journal of comparative and physiological psychology 53, 549-552 (1960).
Makeig, S. et al. Dynamic brain sources of visual evoked responses. Science 295, 690-694
(2002).
Malmierca, M. S. The structure and physiology of the rat auditory system: an overview.
International review of neurobiology 56, 147-211 (2003).
Mamiya, N. et al. Brain region-specific gene expression activation required for
reconsolidation and extinction of contextual fear memory. The Journal of neuroscience: the
official journal of the Society for Neuroscience 29, 402-413 (2009).

- 95 -
Maren, S. & Hobin, J. A. Hippocampal regulation of context-dependent neuronal activity in
the lateral amygdala. Learn Mem 14, 318-324 (2007).
Maren, S. & Quirk, G. J. Neuronal signalling of fear memory. Nature reviews. Neuroscience
5, 844-852 (2004).
Maren, S. Neurobiology of Pavlovian fear conditioning. Annual review of neuroscience 24,
897-931 (2001).
Markram, H. et al. Interneurons of the neocortical inhibitory system. Nature reviews.
Neuroscience 5, 793-807 (2004).
Maroun, M., Kavushansky, A., Holmes, A., Wellman, C. & Motanis, H. Enhanced extinction
of aversive memories by high-frequency stimulation of the rat infralimbic cortex. PLoS One
7, e35853 (2012).
Marsicano, G. et al. The endogenous cannabinoid system controls extinction of aversive
memories. Nature 418, 530-534 (2002).
Mascagni, F., McDonald, A. J. & Coleman, J. R. Corticoamygdaloid and corticocortical
projections of the rat temporal cortex: a Phaseolus vulgaris leucoagglutinin study.
Neuroscience 57, 697-715 (1993).
Massi, L. et al. Temporal Dynamics of Parvalbumin-Expressing Axo-axonic and Basket Cells
in the Rat Medial Prefrontal Cortex In Vivo. J Neurosci 32, 16496-16502 (2012).
McCartney, H., Johnson, A. D., Weil, Z. M. & Givens, B. Theta reset produces optimal
conditions for long-term potentiation. Hippocampus 14, 684-687 (2004).
McDonald, A. J. Cortical pathways to the mammalian amygdala. Prog Neurobiol 55, 257-332
(1998).
McDonald, A. J. Organization of amygdaloid projections to the mediodorsal thalamus and
prefrontal cortex: a fluorescence retrograde transport study in the rat. The Journal of
comparative neurology 262, 46-58 (1987).
McDonald, A. J. Organization of amygdaloid projections to the prefrontal cortex and
associated striatum in the rat. Neuroscience 44, 1-14 (1991).
McDonald, A. J., Mascagni, F. & Guo, L. Projections of the medial and lateral prefrontal
cortices to the amygdala: a Phaseolus vulgaris leucoagglutinin study in the rat. Neuroscience
71, 55-75 (1996).
McKernan, M. G. & Shinnick-Gallagher, P. Fear conditioning induces a lasting potentiation
of synaptic currents in vitro. Nature 390, 607-611 (1997).

- 96 -
Merchant, H., de Lafuente, V., Pena-Ortega, F. & Larriva-Sahd, J. Functional impact of
interneuronal inhibition in the cerebral cortex of behaving animals. Progress in neurobiology
99, 163-178 (2012).
Meskenaite, V. Calretinin-immunoreactive local circuit neurons in area 17 of the cynomolgus
monkey, Macaca fascicularis. The Journal of comparative neurology 379, 113-132 (1997).
Metherate, R. & Ashe, J. H. Basal forebrain stimulation modifies auditory cortex
responsiveness by an action at muscarinic receptors. Brain research 559, 163-167 (1991).
Metherate, R., Cox, C. L. & Ashe, J. H. Cellular bases of neocortical activation: modulation
of neural oscillations by the nucleus basalis and endogenous acetylcholine. The Journal of
neuroscience: the official journal of the Society for Neuroscience 12, 4701-4711 (1992).
Micheyl, C. et al. The role of auditory cortex in the formation of auditory streams. Hearing
research 229, 116-131 (2007).
Milad, M. R. & Quirk, G. J. Neurons in medial prefrontal cortex signal memory for fear
extinction. Nature 420, 70-74 (2002).
Milad, M. R., Vidal-Gonzalez, I. & Quirk, G. J. Electrical stimulation of medial prefrontal
cortex reduces conditioned fear in a temporally specific manner. Behavioural neuroscience
118, 389-394 (2004).
Miles, R., Toth, K., Gulyas, A. I., Hajos, N. & Freund, T. F. Differences between somatic and
dendritic inhibition in the hippocampus. Neuron 16, 815-823 (1996).
Miller, E. K. & Cohen, J. D. An integrative theory of prefrontal cortex function. Annual
review of neuroscience 24, 167-202 (2001).
Millhouse, O. E. The intercalated cells of the amygdala. The Journal of comparative
neurology 247, 246-271 (1986).
Minciacchi, D. & Granato, A. Development of the thalamocortical system: transient-crossed
projections to the frontal cortex in neonatal rats. The Journal of comparative neurology 281,
1-12 (1989).
Mizuseki, K., Diba, K., Pastalkova, E. & Buzsaki, G. Hippocampal CA1 pyramidal cells form
functionally distinct sublayers. Nature neuroscience 14, 1174-1181 (2011).
Molnar, Z. & Cheung, A. F. Towards the classification of subpopulations of layer V
pyramidal projection neurons. Neuroscience research 55, 105-115 (2006).
Moore, C. I., Carlen, M., Knoblich, U. & Cardin, J. A. Neocortical interneurons: from
diversity, strength. Cell 142, 189-193 (2010).

- 97 -
Morawska, M. M. & Fendt, M. The effects of muscimol and AMN082 injections into the
medial prefrontal cortex on the expression and extinction of conditioned fear in mice. J Exp
Biol 215, 1394-1398 (2012).
Morgan, M. A. & LeDoux, J. E. Differential contribution of dorsal and ventral medial
prefrontal cortex to the acquisition and extinction of conditioned fear in rats. Behavioural
neuroscience 109, 681-688 (1995).
Morgan, M. A., Romanski, L. M. & LeDoux, J. E. Extinction of emotional learning:
contribution of medial prefrontal cortex. Neuroscience letters 163, 109-113 (1993).
Morgan, M. A., Schulkin, J. & LeDoux, J. E. Ventral medial prefrontal cortex and emotional
perseveration: the memory for prior extinction training. Behavioural brain research 146, 121-
130 (2003).
Morishima, M. & Kawaguchi, Y. Recurrent connection patterns of corticostriatal pyramidal
cells in frontal cortex. The Journal of neuroscience: the official journal of the Society for
Neuroscience 26, 4394-4405 (2006).
Morishima, M., Morita, K., Kubota, Y. & Kawaguchi, Y. Highly differentiated projection-
specific cortical subnetworks. The Journal of neuroscience: the official journal of the Society
for Neuroscience 31, 10380-10391 (2011).
Morrow, B. A., Elsworth, J. D., Inglis, F. M. & Roth, R. H. An antisense oligonucleotide
reverses the footshock-induced expression of fos in the rat medial prefrontal cortex and the
subsequent expression of conditioned fear-induced immobility. The Journal of neuroscience:
the official journal of the Society for Neuroscience 19, 5666-5673 (1999a).
Morrow, B. A., Elsworth, J. D., Rasmusson, A. M. & Roth, R. H. The role of mesoprefrontal
dopamine neurons in the acquisition and expression of conditioned fear in the rat.
Neuroscience 92, 553-564 (1999b).
Mountcastle, V. B., Davies, P. W. & Berman, A. L. Response properties of neurons of cat's
somatic sensory cortex to peripheral stimuli. Journal of neurophysiology 20, 374-407 (1957).
Mueller, D., Olivera-Figueroa, L. A., Pine, D. S. & Quirk, G. J. The effects of yohimbine and
amphetamine on fear expression and extinction in rats. Psychopharmacology (Berl) 204, 599-
606 (2009).
Mueller, D., Porter, J. T. & Quirk, G. J. Noradrenergic signaling in infralimbic cortex
increases cell excitability and strengthens memory for fear extinction. The Journal of
neuroscience: the official journal of the Society for Neuroscience 28, 369-375 (2008).
Muigg, P. et al. Impaired extinction of learned fear in rats selectively bred for high anxiety--
evidence of altered neuronal processing in prefrontal-amygdala pathways. The European
journal of neuroscience 28, 2299-2309 (2008).

- 98 -
Myers, K. M. & Davis, M. Mechanisms of fear extinction. Molecular psychiatry 12, 120-150
(2007).
Nicolelis, M. A., Baccala, L. A., Lin, R. C. & Chapin, J. K. Sensorimotor encoding by
synchronous neural ensemble activity at multiple levels of the somatosensory system. Science
268, 1353-1358 (1995).
Nieuwenhuys, R. Comparative neuroanatomy: place, principles, practice and programme.
European journal of morphology 32, 142-155 (1994).
Olah, S. et al. Regulation of cortical microcircuits by unitary GABA-mediated volume
transmission. Nature 461, 1278-1281 (2009).
Ongur, D. & Price, J. L. The organization of networks within the orbital and medial prefrontal
cortex of rats, monkeys and humans. Cereb Cortex 10, 206-219 (2000).
Orozco-Cabal, L. et al. A novel rat medial prefrontal cortical slice preparation to investigate
synaptic transmission from amygdala to layer V prelimbic pyramidal neurons. J Neurosci
Methods 151, 148-158 (2006).
Orsini, C. A., Kim, J. H., Knapska, E. & Maren, S. Hippocampal and prefrontal projections to
the basal amygdala mediate contextual regulation of fear after extinction. The Journal of
neuroscience: the official journal of the Society for Neuroscience 31, 17269-17277 (2011).
Padilla-Coreano, N., Do-Monte, F. H. & Quirk, G. J. A time-dependent role of midline
thalamic nuclei in the retrieval of fear memory. Neuropharmacology 62, 457-463 (2012).
Pape, H. C. & Pare, D. Plastic synaptic networks of the amygdala for the acquisition,
expression, and extinction of conditioned fear. Physiological reviews 90, 419-463 (2010).
Pare, D., Quirk, G. J. & Ledoux, J. E. New vistas on amygdala networks in conditioned fear.
Journal of neurophysiology 92, 1-9 (2004).
Parent, M. A., Wang, L., Su, J., Netoff, T. & Yuan, L. L. Identification of the hippocampal
input to medial prefrontal cortex in vitro. Cereb Cortex 20, 393-403 (2010).
Pavlov, I. P. Conditioned reflexes. Oxford: Oxford University Press (1927).
Peter, M. et al. Induction of immediate early genes in the mouse auditory cortex after auditory
cued fear conditioning to complex sounds. Genes, brain, and behaviour 11, 314-324 (2012).
Peyrache, A., Khamassi, M., Benchenane, K., Wiener, S. I. & Battaglia, F. P. Replay of rule-
learning related neural patterns in the prefrontal cortex during sleep. Nature neuroscience 12,
919-926 (2009).

- 99 -
Pezze, M. A., Bast, T. & Feldon, J. Significance of dopamine transmission in the rat medial
prefrontal cortex for conditioned fear. Cereb Cortex 13, 371-380 (2003).
Pfeffer, C. K., Xue, M., He, M., Huang, Z. J. & Scanziani, M. Inhibition of inhibition in
visual cortex: the logic of connections between molecularly distinct interneurons. Nature
neuroscience 16, 1068-1076 (2013).
Pfeiffer, U. J. & Fendt, M. Prefrontal dopamine D4 receptors are involved in encoding fear
extinction. Neuroreport 17, 847-850 (2006).
Pi, H. J. et al. Cortical interneurons that specialize in disinhibitory control. Nature (2013).
Pitman, R. K. et al. Biological studies of post-traumatic stress disorder. Nature reviews.
Neuroscience 13, 769-787 (2012).
Plendl, W. & Wotjak, C. T. Dissociation of within- and between-session extinction of
conditioned fear. The Journal of neuroscience: the official journal of the Society for
Neuroscience 30, 4990-4998 (2010).
Popa, D., Duvarci, S., Popescu, A. T., Lena, C. & Pare, D. Coherent amygdalocortical theta
promotes fear memory consolidation during paradoxical sleep. Proceedings of the National
Academy of Sciences of the United States of America 107, 6516-6519 (2010).
Pouille, F. & Scanziani, M. Enforcement of temporal fidelity in pyramidal cells by somatic
feed-forward inhibition. Science 293, 1159-1163 (2001).
Quirk, G. J. & Mueller, D. Neural mechanisms of extinction learning and retrieval.
Neuropsychopharmacology: official publication of the American College of
Neuropsychopharmacology 33, 56-72 (2008).
Quirk, G. J. Memory for extinction of conditioned fear is long-lasting and persists following
spontaneous recovery. Learn Mem 9, 402-407 (2002).
Quirk, G. J., Armony, J. L. & LeDoux, J. E. Fear conditioning enhances different temporal
components of tone-evoked spike trains in auditory cortex and lateral amygdala. Neuron 19,
613-624 (1997).
Quirk, G. J., Repa, C. & LeDoux, J. E. Fear conditioning enhances short-latency auditory
responses of lateral amygdala neurons: parallel recordings in the freely behaving rat. Neuron
15, 1029-1039 (1995).
Quirk, G. J., Russo, G. K., Barron, J. L. & Lebron, K. The role of ventromedial prefrontal
cortex in the recovery of extinguished fear. The Journal of neuroscience: the official journal
of the Society for Neuroscience 20, 6225-6231 (2000).

- 100 -
Ramon y Cajal, S. Comparative study of the sensory areas of the human cortex. Harvard College
Library (1989).
Ray, J. P. & Price, J. L. The organization of the thalamocortical connections of the
mediodorsal thalamic nucleus in the rat, related to the ventral forebrain-prefrontal cortex
topography. The Journal of comparative neurology 323, 167-197 (1992).
Repa, J. C. et al. Two different lateral amygdala cell populations contribute to the initiation
and storage of memory. Nature neuroscience 4, 724-731 (2001).
Rescorla, R. A. & Heth, C. D. Reinstatement of fear to an extinguished conditioned stimulus.
Journal of experimental psychology. Animal behaviour processes 1, 88-96 (1975).
Rescorla, R. A. Pavlovian conditioning. It's not what you think it is. The American
psychologist 43, 151-160 (1988).
Resstel, L. B., Joca, S. R., Guimaraes, F. G. & Correa, F. M. Involvement of medial prefrontal
cortex neurons in behavioural and cardiovascular responses to contextual fear conditioning.
Neuroscience 143, 377-385 (2006).
Riehle, A., Grun, S., Diesmann, M. & Aertsen, A. Spike synchronization and rate modulation
differentially involved in motor cortical function. Science 278, 1950-1953 (1997).
Rizzuto, D. S. et al. Reset of human neocortical oscillations during a working memory task.
Proceedings of the National Academy of Sciences of the United States of America 100, 7931-
7936 (2003).
Rogan, M. T., Staubli, U. V. & LeDoux, J. E. Fear conditioning induces associative long-term
potentiation in the amygdala. Nature 390, 604-607 (1997).
Romanski, L. M. & LeDoux, J. E. Equipotentiality of thalamo-amygdala and thalamo-cortico-
amygdala circuits in auditory fear conditioning. The Journal of neuroscience: the official
journal of the Society for Neuroscience 12, 4501-4509 (1992).
Romanski, L. M. & LeDoux, J. E. Organization of rodent auditory cortex: anterograde
transport of PHA-L from MGv to temporal neocortex. Cereb Cortex 3, 499-514 (1993a).
Romanski, L. M. & LeDoux, J. E. Information cascade from primary auditory cortex to the
amygdala: corticocortical and corticoamygdaloid projections of temporal cortex in the rat.
Cereb Cortex 3, 515-532 (1993b).
Rosen, J. B. et al. Lesions of the perirhinal cortex but not of the frontal, medial prefrontal,
visual, or insular cortex block fear-potentiated startle using a visual conditioned stimulus. The
Journal of neuroscience: the official journal of the Society for Neuroscience 12, 4624-4633
(1992).

- 101 -
Rosenkranz, J. A. & Grace, A. A. Dopamine-mediated modulation of odour-evoked amygdala
potentials during pavlovian conditioning. Nature 417, 282-287 (2002).
Royer, S. et al. Control of timing, rate and bursts of hippocampal place cells by dendritic and
somatic inhibition. Nature neuroscience 15, 769-775 (2012).
Rutkowski, R. G. & Weinberger, N. M. Encoding of learned importance of sound by
magnitude of representational area in primary auditory cortex. Proceedings of the National
Academy of Sciences of the United States of America 102, 13664-13669 (2005).
Sacchetti, B., Baldi, E., Lorenzini, C. A. & Bucherelli, C. Differential contribution of some
cortical sites to the formation of memory traces supporting fear conditioning. Experimental
brain research. Experimentelle Hirnforschung. Experimentation cerebrale 146, 223-232
(2002).
Sakata, S., Yamamori, T. & Sakurai, Y. 7-12 Hz cortical oscillations: behavioural context and
dynamics of prefrontal neuronal ensembles. Neuroscience 134, 1099-1111 (2005).
Salinas, E. & Sejnowski, T. J. Correlated neuronal activity and the flow of neural information.
Nature reviews. Neuroscience 2, 539-550 (2001).
Santini, E. & Porter, J. T. M-type potassium channels modulate the intrinsic excitability of
infralimbic neurons and regulate fear expression and extinction. The Journal of neuroscience:
the official journal of the Society for Neuroscience 30, 12379-12386 (2010).
Santini, E., Ge, H., Ren, K., Pena de Ortiz, S. & Quirk, G. J. Consolidation of fear extinction
requires protein synthesis in the medial prefrontal cortex. The Journal of neuroscience: the
official journal of the Society for Neuroscience 24, 5704-5710 (2004).
Santini, E., Quirk, G. J. & Porter, J. T. Fear conditioning and extinction differentially modify
the intrinsic excitability of infralimbic neurons. The Journal of neuroscience: the official
journal of the Society for Neuroscience 28, 4028-4036 (2008).
Santini, E., Sepulveda-Orengo, M. & Porter, J. T. Muscarinic receptors modulate the intrinsic
excitability of infralimbic neurons and consolidation of fear extinction.
Neuropsychopharmacology: official publication of the American College of
Neuropsychopharmacology 37, 2047-2056 (2012).
Schafe, G. E. & LeDoux, J. E. Memory consolidation of auditory pavlovian fear conditioning
requires protein synthesis and protein kinase A in the amygdala. The Journal of neuroscience:
the official journal of the Society for Neuroscience 20, RC96 (2000).
Schreiner, C. E. & Winer, J. A. Auditory cortex mapmaking: principles, projections, and
plasticity. Neuron 56, 356-365 (2007).

- 102 -
Sesack, S. R., Deutch, A. Y., Roth, R. H. & Bunney, B. S. Topographical organization of the
efferent projections of the medial prefrontal cortex in the rat: an anterograde tract-tracing
study with Phaseolus vulgaris leucoagglutinin. The Journal of comparative neurology 290,
213-242 (1989).
Shehadi, K. & Maroun, M. Different effects of low frequency stimulation to infralimbic
prefrontal cortex on extinction of aversive memories. Brain research (2012).
Shi, C. J. & Cassell, M. D. Cortical, thalamic, and amygdaloid projections of rat temporal
cortex. The Journal of comparative neurology 382, 153-175 (1997).
Shin, L. M. & Liberzon, I. The neurocircuitry of fear, stress, and anxiety disorders.
Neuropsychopharmacology: official publication of the American College of
Neuropsychopharmacology 35, 169-191 (2010).
Shin, R. M., Tsvetkov, E. & Bolshakov, V. Y. Spatiotemporal asymmetry of associative
synaptic plasticity in fear conditioning pathways. Neuron 52, 883-896 (2006).
Shinonaga, Y., Takada, M. & Mizuno, N. Topographic organization of collateral projections
from the basolateral amygdaloid nucleus to both the prefrontal cortex and nucleus accumbens
in the rat. Neuroscience 58, 389-397 (1994).
Siapas, A. G., Lubenov, E. V. & Wilson, M. A. Prefrontal phase locking to hippocampal theta
oscillations. Neuron 46, 141-151 (2005).
Sierra-Mercado, D., Jr., Corcoran, K. A., Lebron-Milad, K. & Quirk, G. J. Inactivation of the
ventromedial prefrontal cortex reduces expression of conditioned fear and impairs subsequent
recall of extinction. The European journal of neuroscience 24, 1751-1758 (2006).
Sierra-Mercado, D., Padilla-Coreano, N. & Quirk, G. J. Dissociable roles of prelimbic and
infralimbic cortices, ventral hippocampus, and basolateral amygdala in the expression and
extinction of conditioned fear. Neuropsychopharmacology: official publication of the
American College of Neuropsychopharmacology 36, 529-538 (2011).
Silberberg, G., Gupta, A. & Markram, H. Stereotypy in neocortical microcircuits. Trends in
neurosciences 25, 227-230 (2002).
Singer, W. Neuronal synchrony: a versatile code for the definition of relations? Neuron 24,
49-65, 111-125 (1999).
Sirota, A. et al. Entrainment of neocortical neurons and gamma oscillations by the
hippocampal theta rhythm. Neuron 60, 683-697 (2008).
Smith, M. A., Banerjee, S., Gold, P. W. & Glowa, J. Induction of c-fos mRNA in rat brain by
conditioned and unconditioned stressors. Brain research 578, 135-141 (1992).

- 103 -
Sohal, V. S., Zhang, F., Yizhar, O. & Deisseroth, K. Parvalbumin neurons and gamma
rhythms enhance cortical circuit performance. Nature 459, 698-702 (2009).
Somogyi, J. et al. GABAergic basket cells expressing cholecystokinin contain vesicular
glutamate transporter type 3 (VGLUT3) in their synaptic terminals in hippocampus and
isocortex of the rat. The European journal of neuroscience 19, 552-569 (2004).
Somogyi, P. A specific 'axo-axonal' interneuron in the visual cortex of the rat. Brain research
136, 345-350 (1977).
Somogyi, P. et al. Identified axo-axonic cells are immunoreactive for GABA in the
hippocampus and visual cortex of the cat. Brain research 332, 143-149 (1985).
Somogyi, P., Priestley, J. V., Cuello, A. C., Smith, A. D. & Bolam, J. P. Synaptic connections
of substance P-immunoreactive nerve terminals in the substantia nigra of the rat. A correlated
light- and electron-microscopic study. Cell and tissue research 223, 469-486 (1982).
Somogyi, P., Tamas, G., Lujan, R. & Buhl, E. H. Salient features of synaptic organisation in
the cerebral cortex. Brain research. Brain research reviews 26, 113-135 (1998).
Sotres-Bayon, F. & Quirk, G. J. Prefrontal control of fear: more than just extinction. Curr
Opin Neurobiol 20, 231-235 (2010).
Sotres-Bayon, F., Bush, D. E. & LeDoux, J. E. Acquisition of fear extinction requires
activation of NR2B-containing NMDA receptors in the lateral amygdala.
Neuropsychopharmacology: official publication of the American College of
Neuropsychopharmacology 32, 1929-1940 (2007).
Sotres-Bayon, F., Diaz-Mataix, L., Bush, D. E. & LeDoux, J. E. Dissociable roles for the
ventromedial prefrontal cortex and amygdala in fear extinction: NR2B contribution. Cereb
Cortex 19, 474-482 (2009).
Sotres-Bayon, F., Sierra-Mercado, D., Pardilla-Delgado, E. & Quirk, G. J. Gating of fear in
prelimbic cortex by hippocampal and amygdala inputs. Neuron 76, 804-812 (2012).
Spruston, N. Pyramidal neurons: dendritic structure and synaptic integration. Nature reviews.
Neuroscience 9, 206-221 (2008).
Staiger, J. F., Freund, T. F. & Zilles, K. Interneurons immunoreactive for vasoactive intestinal
polypeptide (VIP) are extensively innervated by parvalbumin-containing boutons in rat
primary somatosensory cortex. The European journal of neuroscience 9, 2259-2268 (1997).
Stevenson, C. W. Role of amygdala-prefrontal cortex circuitry in regulating the expression of
contextual fear memory. Neurobiology of learning and memory 96, 315-323 (2011).
Stopfer, M., Bhagavan, S., Smith, B. H. & Laurent, G. Impaired odour discrimination on
desynchronization of odour-encoding neural assemblies. Nature 390, 70-74 (1997).

- 104 -
Streb, J. M. & Smith, K. Frontal lobotomy and the elimination of conditioned anxiety in the
rat. Journal of comparative and physiological psychology 48, 126-129 (1955).
Suga, N. & Ma, X. Multiparametric corticofugal modulation and plasticity in the auditory
system. Nature reviews. Neuroscience 4, 783-794 (2003).
Sugino, K. et al. Molecular taxonomy of major neuronal classes in the adult mouse forebrain.
Nature neuroscience 9, 99-107 (2006).
Sun, Y. J., Kim, Y. J., Ibrahim, L. A., Tao, H. W. & Zhang, L. I. Synaptic mechanisms
underlying functional dichotomy between intrinsic-bursting and regular-spiking neurons in
auditory cortical layer 5. The Journal of neuroscience: the official journal of the Society for
Neuroscience 33, 5326-5339 (2013).
Swanson, L. W. A direct projection from Ammon's horn to prefrontal cortex in the rat. Brain
research 217, 150-154 (1981).
Takagishi, M. & Chiba, T. Efferent projections of the infralimbic (area 25) region of the
medial prefrontal cortex in the rat: an anterograde tracer PHA-L study. Brain research 566,
26-39 (1991).
Takehara-Nishiuchi, K., Maal-Bared, G. & Morrissey, M. D. Increased Entorhinal-Prefrontal
Theta Synchronization Parallels Decreased Entorhinal-Hippocampal Theta Synchronization
during Learning and Consolidation of Associative Memory. Frontiers in behavioural
neuroscience 5, 90 (2011).
Tamas, G., Somogyi, P. & Buhl, E. H. Differentially interconnected networks of GABAergic
interneurons in the visual cortex of the cat. The Journal of neuroscience: the official journal
of the Society for Neuroscience 18, 4255-4270 (1998).
Tang, J. et al. Pavlovian fear memory induced by activation in the anterior cingulate cortex.
Molecular pain 1, 6 (2005).
Taniguchi, H., Lu, J. & Huang, Z. J. The spatial and temporal origin of chandelier cells in
mouse neocortex. Science 339, 70-74 (2013).
Teich, A. H. et al. Role of auditory cortex in the acquisition of differential heart rate
conditioning. Physiology & behaviour 44, 405-412 (1988).
Terreberry, R. R. & Neafsey, E. J. The rat medial frontal cortex projects directly to autonomic
regions of the brainstem. Brain research bulletin 19, 639-649 (1987).
Thierry, A. M., Blanc, G., Sobel, A., Stinus, L. & Golwinski, J. Dopaminergic terminals in
the rat cortex. Science 182, 499-501 (1973).

- 105 -
Thompson, B. M. et al. Activation of the infralimbic cortex in a fear context enhances
extinction learning. Learn Mem 17, 591-599 (2010).
Tian, S. et al. Ventrolateral prefrontal cortex is required for fear extinction in a modified
delay conditioning paradigm in rats. Neuroscience 189, 258-268 (2011).
Tierney, P. L., Degenetais, E., Thierry, A. M., Glowinski, J. & Gioanni, Y. Influence of the
hippocampus on interneurons of the rat prefrontal cortex. The European journal of
neuroscience 20, 514-524 (2004).
Toledo-Rodriguez, M. et al. Correlation maps allow neuronal electrical properties to be
predicted from single-cell gene expression profiles in rat neocortex. Cereb Cortex 14, 1310-
1327 (2004).
Tsvetkov, E., Carlezon, W. A., Benes, F. M., Kandel, E. R. & Bolshakov, V. Y. Fear
conditioning occludes LTP-induced presynaptic enhancement of synaptic transmission in the
cortical pathway to the lateral amygdala. Neuron 34, 289-300 (2002).
Uematsu, M. et al. Quantitative chemical composition of cortical GABAergic neurons
revealed in transgenic venus-expressing rats. Cereb Cortex 18, 315-330 (2008).
Uylings, H. B. & van Eden, C. G. Qualitative and quantitative comparison of the prefrontal
cortex in rat and in primates, including humans. Progress in brain research 85, 31-62 (1990).
Uylings, H. B., Groenewegen, H. J. & Kolb, B. Do rats have a prefrontal cortex? Behavioural
brain research 146, 3-17 (2003).
Van Eden, C. G. & Uylings, H. B. Cytoarchitectonic development of the prefrontal cortex in
the rat. The Journal of comparative neurology 241, 253-267 (1985).
Varela, F., Lachaux, J. P., Rodriguez, E. & Martinerie, J. The brainweb: phase
synchronization and large-scale integration. Nature reviews. Neuroscience 2, 229-239 (2001).
Veening, J. G., Swanson, L. W. & Sawchenko, P. E. The organization of projections from the
central nucleus of the amygdala to brainstem sites involved in central autonomic regulation: a
combined retrograde transport-immunohistochemical study. Brain Res 303, 337-357 (1984).
Vertes, R. P. Differential projections of the infralimbic and prelimbic cortex in the rat.
Synapse 51, 32-58 (2004).
Vertes, R. P. Interactions among the medial prefrontal cortex, hippocampus and midline
thalamus in emotional and cognitive processing in the rat. Neuroscience 142, 1-20 (2006).
Vianna, D. M., Graeff, F. G., Brandao, M. L. & Landeira-Fernandez, J. Defensive freezing
evoked by electrical stimulation of the periaqueductal gray: comparison between dorsolateral
and ventrolateral regions. Neuroreport 12, 4109-4112 (2001).

- 106 -
Vianna, M. R., Szapiro, G., McGaugh, J. L., Medina, J. H. & Izquierdo, I. Retrieval of
memory for fear-motivated training initiates extinction requiring protein synthesis in the rat
hippocampus. Proceedings of the National Academy of Sciences of the United States of
America 98, 12251-12254 (2001).
Vidal-Gonzalez, I., Vidal-Gonzalez, B., Rauch, S. L. & Quirk, G. J. Microstimulation reveals
opposing influences of prelimbic and infralimbic cortex on the expression of conditioned fear.
Learn Mem 13, 728-733 (2006).
Vouimba, R. M. & Maroun, M. Learning-induced changes in mPFC-BLA connections after
fear conditioning, extinction, and reinstatement of fear. Neuropsychopharmacology: official
publication of the American College of Neuropsychopharmacology 36, 2276-2285 (2011).
Vouimba, R. M., Garcia, R., Baudry, M. & Thompson, R. F. Potentiation of conditioned
freezing following dorsomedial prefrontal cortex lesions does not interfere with fear reduction
in mice. Behavioural neuroscience 114, 720-724 (2000).
Wang, Y., Gupta, A., Toledo-Rodriguez, M., Wu, C. Z. & Markram, H. Anatomical,
physiological, molecular and circuit properties of nest basket cells in the developing
somatosensory cortex. Cereb Cortex 12, 395-410 (2002).
Watakabe, A. Comparative molecular neuroanatomy of mammalian neocortex: what can gene
expression tell us about areas and layers? Development, growth & differentiation 51, 343-354
(2009).
Watakabe, A. et al. Comparative analysis of layer-specific genes in Mammalian neocortex.
Cereb Cortex 17, 1918-1933 (2007).
Waterhouse, I. K. Effects of prefrontal lobotomy on conditioned fear and food responses in
monkeys. Journal of comparative and physiological psychology 50, 81-88 (1957).
Weinberger, N. M. Associative representational plasticity in the auditory cortex: a synthesis
of two disciplines. Learn Mem 14, 1-16 (2007).
Weinberger, N. M. Physiological memory in primary auditory cortex: characteristics and
mechanisms. Neurobiology of learning and memory 70, 226-251 (1998).
Weinberger, N. M. Specific long-term memory traces in primary auditory cortex. Nature
reviews. Neuroscience 5, 279-290 (2004).
Weinberger, N. M. The medial geniculate, not the amygdala, as the root of auditory fear
conditioning. Hearing research 274, 61-74 (2011).
Weinberger, N. M., Miasnikov, A. A. & Chen, J. C. The level of cholinergic nucleus basalis
activation controls the specificity of auditory associative memory. Neurobiology of learning
and memory 86, 270-285 (2006).

- 107 -
Weinberger, N. M., Miasnikov, A. A., Bieszczad, K. M. & Chen, J. C. Gamma band plasticity
in sensory cortex is a signature of the strongest memory rather than memory of the training
stimulus. Neurobiology of learning and memory 104, 49-63 (2013).
Weisskopf, M. G. & LeDoux, J. E. Distinct populations of NMDA receptors at subcortical
and cortical inputs to principal cells of the lateral amygdala. Journal of neurophysiology 81,
930-934 (1999).
Wilensky, A. E., Schafe, G. E., Kristensen, M. P. & LeDoux, J. E. Rethinking the fear circuit:
the central nucleus of the amygdala is required for the acquisition, consolidation, and
expression of Pavlovian fear conditioning. The Journal of neuroscience: the official journal of
the Society for Neuroscience 26, 12387-12396 (2006).
Willard, F. J. & Ryugo, D. K. Anatomy of the central auditory system. In: The auditory
psychobiology of the mouse, (ed. Willott JF), 201–304 Thomas, Springfield, IL (1983).
Williams, J. M. & Givens, B. Stimulation-induced reset of hippocampal theta in the freely
performing rat. Hippocampus 13, 109-116 (2003).
Wilson, N. R., Runyan, C. A., Wang, F. L. & Sur, M. Division and subtraction by distinct
cortical inhibitory networks in vivo. Nature 488, 343-348 (2012).
Womelsdorf, T. et al. Modulation of neuronal interactions through neuronal synchronization.
Science 316, 1609-1612 (2007).
Woodruff, A. R. et al. State-dependent function of neocortical chandelier cells. The Journal of
neuroscience: the official journal of the Society for Neuroscience 31, 17872-17886 (2011).
Xu, X., Roby, K. D. & Callaway, E. M. Immunochemical characterization of inhibitory
mouse cortical neurons: three chemically distinct classes of inhibitory cells. The Journal of
comparative neurology 518, 389-404 (2010).
Yang, C. R., Seamans, J. K. & Gorelova, N. Electrophysiological and morphological
properties of layers V-VI principal pyramidal cells in rat prefrontal cortex in vitro. The
Journal of neuroscience: the official journal of the Society for Neuroscience 16, 1904-1921
(1996).
Yizhar, O. et al. Neocortical excitation/inhibition balance in information processing and
social dysfunction. Nature 477, 171-178 (2011).
Yoder, R. M. & Pang, K. C. Involvement of GABAergic and cholinergic medial septal
neurons in hippocampal theta rhythm. Hippocampus 15, 381-392 (2005).

- 108 -
Zaitsev, A. V., Povysheva, N. V., Gonzalez-Burgos, G. & Lewis, D. A. Electrophysiological
classes of layer 2/3 pyramidal cells in monkey prefrontal cortex. Journal of neurophysiology
108, 595-609 (2012).
Zugaro, M. B., Monconduit, L. & Buzsaki, G. Spike phase precession persists after transient
intrahippocampal perturbation. Nature neuroscience 8, 67-71 (2005).