retraction: induction of apoptosis by hiv-1-infected monocytic cells
TRANSCRIPT

Induction of Apoptosis by HIV-1-Infected Monocytic Cells1
Kirk Sperber, 2*† Prarthana Beuria,* † Netai Singha,† Irwin Gelman, ‡ Patricia Cortes,†
Houchu Chen,*† and Thomas Kraus†
We have previously described a soluble 6000-Da peptide produced by an HIV-1-infected human macrophage cell line, clone 43HIV ,which induces apoptosis in T and B cells. We have identified this factor as the novel cDNA clone FL14676485 that encodes for thehuman hypothetical protein, FLJ21908. The FL14676485 cDNA clone was isolated from a 43HIV � ZAP Escherichia coli expressionlibrary and screened with a panel of rabbit and mouse anti-apoptotic Abs. We transfected the FL14676485 clone into Bosc cellsand non-HIV-1-infected 43 cells. Western blot analysis of lysates from the FL14676485-transfected 43 cells and Bosc cells usinganti-proapoptotic factor Abs revealed a protein with a molecular mass of 66 kDa corresponding to the size of the full-length geneproduct of the FL14676485 clone, while Western blot of the supernatant demonstrated a doublet of 46-kDa and 6000-Da peptidethat corresponds to our previously described proapoptotic factor. Primary HIV-1BaL-infected monocytes also produce theFLJ21908 protein. Supernatants from these transfected cells induced apoptosis in PBMC, CD4�, and CD8� T and B cells similarto the activity of our previously described proapoptotic factor. PCR analysis of 43 cells and 43HIV cells revealed a base pairfragment of 420 bp corresponding to the FL14676485 gene product in 43HIV cells, but not in 43 cells. The FLJ21908 proteininduces apoptosis through activation of caspase-9 and caspase-3. We have further demonstrated that the FLJ21908 protein hasapoptotic activity in the SH-SY5Y neuronal cell line and can be detected in brain and lymph tissue from HIV-1-infected patientswho have AIDS dementia. The FLJ21908 protein may contribute to the apoptosis and dementia observed in AIDS patients.TheJournal of Immunology, 2003, 170: 1566–1578.
Progressive depletion of CD4� T cells is a characteristicfeature of HIV-1 infection (1). Both virologic and immu-nologic mechanisms have been thought to play an impor-
tant role in the loss of CD4� T cells based on in vitro and in vivoobservations. Apoptosis or programmed cell death has been pro-posed as an alternative explanation for T cell loss in HIV-1-in-fected individuals (2). Spontaneous apoptosis of CD4� and CD8�
T cells and activation-induced apoptosis have been reported inPBL and lymph nodes during HIV-1 infection (3–9). The accel-erated apoptosis may relate to cross-linking of CD4 by gp120,leading to aberrant T cell signaling (10–12), cytokines (2), Fas andFas ligand (FasL)3 interactions (13–15), superantigen activity en-coded by HIV-1 products (16, 17), or the involvement of accessorycells. Several lines of evidence implicate accessory cells, includingmonocytes and dendritic cells, in the induction of apoptosis duringthe course of HIV-1 infection. Monocytes and dendritic cells serveas reservoirs for HIV-1, providing virions and the envelope proteingp120 to target CD4� T cells (18). APC dysfunction as a result ofHIV-1 infection may cause defective T cell activation resulting in
apoptosis instead of cellular activation (19–23). HIV-1 infectionor cross-linking of CD4 on monocytes results in the up-regulationof FasL expression that could induce apoptosis in uninfected by-stander CD4� T cells (24–26).
Soluble proapoptotic factors, especially those produced by HIV-1-infected macrophages, may also be playing a role. Macrophageshave been reported to produce proapoptotic chemokines and cy-tokines as well as apoptosis-promoting low molecular mass mol-ecules such as reactive oxygen species, PG, and NO (2). The che-mokine stromal-derived growth factor, which signals through theCXCR4 chemokine receptor, delivers a death signal to CD8� Tcells and to neuronal cell lines (27, 28). Stromal-derived growthfactor-1 blocks infection of T cells by T cell tropic viruses and mayplay an important role in the regulation of cell differentiation, pro-liferation, and migration of CD8� T cells in inflammatory re-sponses (29). After HIV-1 infection, there is increased productionof proinflammatory cytokines, including IL-6, IFN-�, TGF-�, andTNF-� (30). In HIV-1-infected individuals, this cytokine imbal-ance may contribute to apoptosis. TNF-�, TGF-�, as well as IFN-�promote apoptosis (31–38). Macrophages are crucial in the devel-opment of HIV-1-associated dementia (HAD)3 because they, alongwith microglia, are the only resident cells that can be productivelyinfected with HIV-1 in the CNS (39), although a nonproductive orlatent infection of astrocytes has been observed (40). HIV-1-in-fected macrophages migrate into the brain and constitute the prin-cipal route of viral entry into the CNS (41). HIV-infected or im-mune-stimulated macrophages/microglia produce neurotoxins, andmacrophages/microglia are required for HIV-1- or gp120-inducedneurotoxicity (42–44). Macrophage/Microglia damages neuronsby releasing excitotoxic substances that produce excessive activa-tion of glutamate receptors, primarily having theN-methyl-D-as-partate subtype (NMDAR) that induces apoptosis (39).
We have been interested in studying HIV-1 monocyte interac-tions using a series of human monocyte and macrophage hybrid-omas obtained by fusing monocytes and macrophages with a mu-tagenized U937 promonocytic cell line (45–50). We demonstrated
*Division of Clinical Immunology,†Immunobiology Institute, and‡Department ofMedicine, Mount Sinai School of Medicine, New York, NY 10029
Received for publication August 5, 2002. Accepted for publication November22, 2002.
The costs of publication of this article were defrayed in part by the payment of pagecharges. This article must therefore be hereby markedadvertisement in accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.1 This work was supported by National Institutes of Health Grants AI 45343, MH 59724,AI 45996, CA 94108, and AI 44236. Confocal laser-scanning microscopy was performedat the Mount Sinai Confocal Laser Scanning facility, supported with funding from theNational Institutes of Health Shared Instrumentation Grant 1S10RR09145 and the Na-tional Science Foundation Major Research Instrumentation Grant DB1-9724504.2 Address correspondence and reprint requests to Dr. Kirk Sperber, Box 1089, Divi-sion of Clinical Immunology, 1425 Madison Avenue, New York, NY 10029. E-mailaddress: [email protected] Abbreviations used in this paper: FasL, Fas ligand; CM, complete medium; GFP,green fluorescent protein; HAD, HIV-1-associated dementia; NMDAR,N-methyl-D-aspartate subtype; PARP, poly(ADP-ribose) polymerase; TPR, tetratricopeptide.
The Journal of Immunology
Copyright © 2003 by The American Association of Immunologists, Inc. 0022-1767/03/$02.00
RETRACTED
15 December 2005

that chronically HIV-1-infected human macrophage hybridomasinduce apoptosis in CD4� and CD8� T and B cells by multiplemechanisms, including gp120, FasL expression, and the produc-tion of a soluble 6000-Da proapoptotic peptide (49). In this study,we identify a novel cDNA clone isolated from the chronicallyHIV-1-infected human macrophage cell line, 43HIV, that inducesapoptosis in T and B cells, and in a neuronal cell line similar to ourpreviously described proapoptotic factor. Furthermore, using spe-cific Abs generated against the novel cDNA clone, we demonstratethat the proapoptotic factor is present in brain and lymphoid tissuefrom patients that are HIV-1 infected with HAD, but not in non-HIV-1-infected controls, Alzheimer’s patients, and non-HIV-1 en-cephalitis patients.
Materials and MethodsMonocyte isolation and HIV-1 infection
Mononuclear cells were separated from buffy coats obtained from normalhealthy volunteers by Ficoll-Hypaque (Pharmacia, Piscataway, NJ) densitygradient centrifugation. The cells were washed three times with sterile PBSand resuspended in RPMI 1640 (Life Technologies, Grand Island, NY)supplemented with 10% FCS (Life Technologies), 2 mM L-glutamine, and1% penicillin/streptomycin (Life Technologies), henceforth called com-plete medium (CM). Freshly isolated PBMC were incubated at 37oC in CMand allowed to adhere for 45 min. The nonadherent cells were removed,and adherent cells were washed with sterile PBS, harvested with a rubberpoliceman, and stained with monocyte-specific anti-CD14 mAbs to assessthe purity of the preparation. Ninety percent of the isolated cells expressedCD14 (45). Monocytes were infected with HIV-1BaL, as previously de-scribed (49). The HIV-1BaL virus was obtained from the AIDS Researchand Reference Reagent Program, Division of AIDS, National Institute ofAllergy and Infectious Diseases, National Institutes of Health (Bethesda,MD). Dilutions of HIV-1 supernatant standardized to contain reverse-tran-scriptase activity to 80,000 cpm/ml were incubated for 90 min, followed bythree washes with PBS.
Human macrophage hybridomas, Bosc cells, THB cells, andSH-SY5Y cells
Human macrophage hybridomas were obtained by fusing macrophages(obtained by allowing monocytes to mature into macrophages in Teflonbag cultures) with a hypoxanthine-guanine phosphoribosyltransferase-de-ficient promonocytic line (U937), as previously described (45). We haveuniformly infected and characterized one clone, 43, with different strains ofHIV-1 (43HIV) (47–50). Bosc cells are derived from L23T cells and are agift of K. Horvath (Immunobiology Institute, Mount Sinai School of Med-icine) (51). The SH-SY5Y and THB cells were purchased from the Amer-ican Type Culture Collection (Manassas, VA) (52).
PBMC, CD4�, and CD8� T and B cells
PBMC were separated from buffy coats obtained from normal healthy vol-unteers by Ficoll-Hypaque (Pharmacia) density gradient centrifugation.The cells were washed three times with sterile PBS and resuspended inRPMI 1640 (Life Technologies) supplemented with 10% FCS (Life Tech-nologies), 2 mM L-glutamine, and 1% penicillin/streptomycin (Life Tech-nologies), henceforth called CM (45). We isolated purified CD4� andCD8� T and B cell populations by RosetteSep (Stem Cell Technologies,Vancouver, British Columbia, Canada). RosetteSep is a rapid, easy cellseparation kit for the isolation of highly purified CD4� and CD8� T andB cells from whole blood. Whole blood is added to a RosetteSep cocktail,and cells are cross-linked with tetramer complexes. The cells are thenincubated at room temperature, layered over Ficoll-Hypaque, and centri-fuged for 20 min, and the enriched CD4� and CD8� T and B cells wereisolated (53).
Spleen cells
BALB/c mice used for the spleen cultures were purchased from CharlesRiver Laboratories (Wilmington, MA). Spleen cells were taken and treatedwith 0.17 M Tris-NH4Cl RBC lysis buffer to remove RBC by previouslydescribed methods (54). The isolated spleen cells were used in the apo-ptosis assay.
Acetone precipitation
Acetone precipitation was used to isolate the proapoptotic factor fromcrude supernatants from the 43 and 43HIV cell lines. Acetone (Sigma-Al-drich, St. Louis, MO) was chilled in an ice-salt bath to attain a temperaturebelow 0°C. Proteins were fractionated from 43 and 43HIV supernatants byprecipitation in 95% (v/v) acetone. The precipitated proteins were collectedby centrifugation, and the residual acetone in the precipitates was removedby vacuum centrifugation in a Speed-Vac (Savant, Piscataway, NJ) (49).
Reverse-phase HPLC analysis
Reverse-phase HPLC was performed on the acetone precipitate from the 43and 43HIV cells using a C18 (4.6 � 250-mm) column. Elution of boundproteins was developed using a linear gradient of 0.1% (v/v) trifluoroaceticacid. A gradient of 60 ml was developed at a flow rate of 1 ml/min. Elutionprofiles were monitored at an absorbance of 215 nm. Solvent in the protein-containing fractions was removed by vacuum centrifugation in a Speed-Vac (Savant) (49).
Anion exchange chromatography
Anion exchange chromatography was performed with a Mono-Q HR5/5(5 � 50-mm) column on an FLPC system (Pharmacia). The elution gra-dient was developed using 20 mM Tris-HCl, pH 7.5 (buffer A), and 1 MNaCl in buffer A (buffer B) at a flow rate of 1 ml/min. Samples wereprepared for anion exchange chromatography by exhaustive dialysis inbuffer A. The protein elution profile was monitored by absorbance at 280nm (49).
Generation of Abs
To clone the proapoptotic factor, we generated anti-proapoptotic factormAbs and polyclonal Abs by techniques previously established in our lab-oratory (55). mAbs were generated by boosting two BALB/c mice immu-nized with HPLC-fractionated supernatant from the 43HIV cell line, as pre-viously described, followed by fusion to the non-IgG-secreting mousemyeloma cell line SP2/0 (55). We screened the mAbs by immunoblot andby inhibition of apoptosis in the bioassay using PBMC as target cells. Forthe immunoblot screening, crude fraction 5 of the 43HIV supernatant wasapplied to nitrocellulose membranes using a Bio-dot apparatus (Bio-Rad,Richmond, CA) that permits application of uniform dots. Test supernatantsfrom the fusion were added to the fraction 5 containing nitrocellulose paperin 96-well microtiter plates for 2 h at 25°C, followed by five PBS washesand the addition of HRP-conjugated goat anti-mouse Ab (Life Sciences,Burlingame, CA) for 2 h at 25°C. Irrelevant murine mAbs of all isotypeswill be used as specificity controls in each assay. After extensive PBSwashing, substrate (3, 3�-diaminobenzidine tetrahydrochloride; Pierce,Rockford, IL) was added, and the plates were read for the appearance of ablue color at 30 min, indicating a positive result. Because many mAbs donot react in Western blots, we also screened by inhibition of apoptosis inthe bioassay. In addition, we generated rabbit anti-proapoptotic factor Absby i.m. injecting two rabbits (Mount Sinai Animal Facilities) with frac-tionated supernatant from the 43HIV cell line with CFA (Sigma-Aldrich)with two booster injections (55). Polyclonal antisera were prepared andpooled. IgG was purified by protein A-Sepharose (Pharmacia), as previ-ously described (55). Ouchterlony immunodiffusion in gels was used to testantiserum generated against the proapoptotic fractions. Immunoelectro-phoresis was performed in 2% sodium dodecylsulfate (Sigma-Aldrich) thatcontained 0.01 ml of ethylenediaminetetraacetic acid (Sigma-Aldrich).Rabbit serum against purified proapoptotic factor developed as a singleband in Ouchterlony immunodiffusion. Rabbit Abs were also assayed bytheir ability to block apoptosis in vitro using annexin V staining. ProteinG-purified anti-proapoptotic bodies were isolated from culture supernatantsand coupled to cyanogen bromide Sepharose 4B beads (Pharmacia) bymethods established in our laboratory (55). Coupling efficiency was deter-mined by measuring the protein concentration of the remaining Ab solu-tion. Greater than 90% of the Ab was bound to the beads. One milliliter ofthe Ab-coated beads was rotated overnight (18–20 h) at 4°C with super-natant from 43HIV and Escherichia coli transfected with the FL14676485gene. After washing, unabsorbed proapoptotic factor activity was com-pared with absorbed proapoptotic factor activity.
Annexin V and intracytoplasmic staining for activated caspase-3
FITC-labeled annexin V, a phospholipid-binding protein of the annexinfamily (56, 57), was used to measure apoptosis using a commercially avail-able kit (Coulter, Hialeah, FL). After incubating supernatants containingconcentrations of the FLJ21908 protein with PBMC, T cells, and B cells,the cell samples were washed with ice-cold PBS, followed by centrifuga-tion at 500 � g at 4°C. The cells were incubated with Annexin VFITC at
1567The Journal of Immunology
RETRACTED
15 December 2005

room temperature for 10 min in the dark. In other experiments, propidiumiodide (100 �g/ml) was added to assess cell necrosis. The cells were thenanalyzed by flow cytometry to measure the annexin V� population, gatingon the live cells (49). Cells were also stained intracytoplasmically for ac-tivated caspase-3 (BD Biosciences, San Diego, CA). Cells were perme-abilized with 70% ethanol, washed three times with PBS, and then stainedwith the FITC-labeled Abs directed against activated caspase-3. The cellswere then analyzed by flow cytometry gating on live cells (49)
� ZAP EcoRI/XhoI 43HIV cDNA Library
A 43HIV library was synthesized using the ZAP-cDNA synthesis method(Stratagene, La Jolla, CA) (58). The linker-primer was designed with aGAGA sequence to protect the XhoI restriction site and an 18-base ply (dT)sequence. The restriction site allows the finished cDNA to be inserted ontothe vector unidirectionally in the sense orientation with respect to the lacZpromoter. The linker-primer is a 50-base oligonucleotide with the followingsequence: 5�-GAGAGAGAGAGAGAGAGAAACTAGTCTCGAG(T)18-3�.
GAGA sequence XhoI
The adaptors are comprised of 9- and 13-mer oligonucleotides that are com-plementary to each other and have an EcoRI cohesive end. The adaptors havethe following sequence: 5�-AATTCGGCACGAG-3�; 3�-GCCGTGTC-5�.
The amplified library was grown in XL1-Blue MRF� strain. Two dif-ferent helper phages were used with the ZAP Express library, the ExAssist(Stratagene) interference-resistant helper phage and the 408 helper phage.To screen for the proapoptotic factor expressing clones, E. coli were grownon agar and the colonies were harvested onto 10 mM isopropyl-�-D-thio-galactopyranoside-soaked nitrocellulose filters for 16 h overnight. Themembranes were washed three times in PBS and Western blotted witheither rabbit or mouse anti-proapoptotic Abs, followed by either goat anti-rabbit- or anti-mouse-labeled horseradish-labeled secondary Abs (Pharma-cia), and developed using a commercially available ECL kit (DuPont, Wil-mington, DE) (50).
Western blot
The 43, 43HIV, Bosc, THB, and SH-SY5Y cells, and E. coli were lysedusing buffer containing PMSF (100 mM), aprotinin (10 mg/ml), leupeptin(10 mg/ml), iodoacteamide (1.8 mg/ml), and 0.1% Triton X-100 (Sigma-Aldrich). The lysates were resolved on a 10% SDS-PAGE gel, transferredonto a nitrocellulose membrane, blocked with 5% milk in PBS at roomtemperature for 60 min, then incubated with the anti-proapoptotic Abs(37o) for 43, 43HIV, Bosc cells, and E. coli, and with anti-caspase-3 andanti-poly(ADP-ribose) polymerase (PARP) Abs (BD Biosciences) for theSH-SY5Y cells, and with caspase-8 and caspase-9 Abs (BD Biosciences,La Jolla, CA) at 4°C overnight. A secondary horseradish-labeled goat anti-mouse Ig or goat anti-rabbit (Tago, Burlingame, CA) was then added at25°C for 2 h, and the blot was developed by chemiluminescence using acommercially available ECL kit (DuPont) (50).
Purification of the 6000d peptide
We purified the 6000-Da peptide from the supernatant of E. coli expressingthe FLJ21908 protein. To this end, we concentrated 2 L of bacterial su-pernatant by lyophilization, resuspended the concentrated material in 100mM NaCl and 20 mM Tris-HCl pH 7.5, and loaded it onto a DEAE Sepha-rose column (Pharmacia), increasing the salt concentration from 100 mMto 1 M NaCl eluted fractions. The isolated fractions were run on a 10% poly-acrylamide gel, transferred onto nitrocellulose paper, and Western blotted withthe rabbit anti-FLJ21908 Abs. The Western blot-positive fractions were thensilver stained to ascertain the purity of the protein separation.
Transfection with FL14676485
The 43 and Bosc cells were transiently transfected with a bacterial plasmidcontaining the FL14676485 cDNA clone using CaPO4, Superfect (Qiagen,Valencia, CA), or DEAE-dextran (50). The bacteria containing theFL14676485 cDNA clone were cultured, ethanol precipitated, extracted,and then centrifuged and resuspended in TBS. For CaPO4, a DNA/CaCl2/H2O mixture (500 �l) containing 20 �g of expressed FL14676485 plasmid,ssDNA carrier, 438 �l of H20, and 62 �l 2 M CaCl2 was added to HBSSand incubated with either Bosc cells or 43 cells for 5 h. The 43 and Bosccells were washed, and the medium was replaced. The cells were harvestedat 48–60 h posttransfection. For Superfect (Qiagen), 5 �g of FL14676485DNA was dissolved in TE buffer, pH 7.4, with medium that contains noserum proteins or antibiotics to a total volume of 150 �l. A total of 30 �lof Superfect (Qiagen) reagent was added to the DNA solution, mixed, andthen added to the Bosc and 43 cells for 10 min at 25°C to allow fortransfection complex formation. The cells were then washed once with 4
ml of PBS, and then 1 ml of CM was added. The cells were incubated withthe transfection complex for 3 h at 37°C. The medium was removed, andthe cells were washed in PBS. Fresh CM was added, and the cell super-natants were assayed at 48 and 60 h for apoptotic activity. For the DEAE-dextran method, the resuspended DNA was added to 10 mg/ml of DEAE-dextran (Sigma-Aldrich) and incubated with the 43 and Bosc cells for 4 hat 37°C. After aspirating the DEAE-dextran, the cells were shocked byadding 5 ml of 10% DMSO (Sigma-Aldrich) in PBS for 1 min at roomtemperature, washed with sterile PBS, and resuspended in CM for 48 h at37°C. In some experiments, the 43 and Bosc cells were treated with CaP04
alone, Superfect alone, or DEAE-dextran alone without the FL14767485cDNA clone, while in other experiments the cells were treated with CaP04,Superfect (Qiagen), DEAE-dextran, and jellyfish green fluorescent protein(GFP; Promega, Milwaukee, WI) to assess the efficacy of transfection.
Polymerase chain reaction
RNA was extracted from 43 and 43HIV using acid quanidium thiocynate/phenol/chloroform, as described previously (RNAzol, Linnai, TX) (33).Known quantities of RNA were mixed with 1 �g total cellular RNA andreverse transcribed at 37°C for 60 min in 20 �l of buffer containing 10 mMTris (Sigma-Aldrich), pH 8.3; 50 mM KCl (Sigma-Aldrich); 5 mM MgCl2(Sigma-Aldrich); 1 mM each of dATP, dCTP, dGTP, and dTTP (Sigma);20 U RNase inhibitor (Promega); 0.1 �g oligo(dT)15 (Boehringer Mann-heim, Indianapolis, IN); and 50 U murine leukemia virus reverse transcrip-tase (Bethesda Research Laboratories, Bethesda, MD). The PCR forFL14676485 was performed using the upstream primer 5�-TAGAAAACTGGGAAAAAGACATTA-3� and the downstream primer 5�-TTGGCAACACGGGATTA-3� for 40 cycles at 51.7°C for 1 min, at 50°C for 1 min,and at 68°C for 3 min, with a 20-s elongation step per cycle. Reactionswere stopped by heat inactivation for 10 min at 95°C, annealed for 2.5 min,and extended at 65°C. Negative controls were performed omitting RNAfrom the cDNA synthesis and specific amplification. PCR products wereseparated in a 2% NuSieve agarose (FMC, Rockland, ME) or a 5% poly-acrylamide gel (50).
Immunofluorescence
Frozen sections of brain tissue and lymph tissue were provided by theManhattan AIDS Brain Bank (New York, NY) through the assistance of S.Morgello (Mt. Sinai School of Medicine). The cells were fixed with 1%paraformaldehyde and then stained with murine anti-proapoptotic Abs asthe primary Abs and fluorescein-conjugated F(ab�)2 goat anti-mouse IgG(Tago) as a secondary Ab. mAb W6/32 (anti-class I) and IgG1 isotypecontrols were used as positive and negative controls, respectively. Thesections were mounted with Immun-mount (Shandon, Pittsburgh, PA) beforebeing viewed by a Leica fluorvent laser-scanning confocal microscope (Leica,Deerfield, IL) at a step position of 1 �m on the x-y- or x-z-axis (48).
ResultsFurther characterization of the proapoptotic factor
In our previous work, we demonstrated proapoptotic activity fromfractionated supernatant from the chronically HIV-1-infected cellline, 43HIV (49). Proapoptotic activity could not be precipitatedwith acetone at a concentration lower than 80% saturation, a char-acteristic observed with smaller peptides. Results from peptidebinding to anion exchange matrices at different pH indicated thatthe pI of the proapoptotic activity was between 6.5 and 7.0. Whenactive fractions were electrophoresed on a 10% SDS-PAGE gel, aband corresponding to a Mr of 6000 Da was detected. Furthermore,active fractions from 43HIV were electrophoresed on a nondena-turing SDS-PAGE gel, and proapoptotic activity was electroelutedfrom gel slices corresponding to a Mr of less than 10,000. Althoughwe identified two proapoptotic fractions from 43HIV supernatant inour previous studies, fractions 5 and 6, we investigated only frac-tion 5 in this study. We further characterized fraction 5 from the43HIV supernatant by reverse-phase HPLC analysis. We comparedHLPC elution profiles of fraction 5 and demonstrated that eightpeaks were present in fraction 5 from 43HIV supernatant, but not inthe uninfected 43 supernatant. We then wanted to determinewhether any of the unique fractions from the 43HIV supernatanthad proapoptotic activity in our bioassay. Unfortunately, none ofthe fractions from the HPLC had activity in the apoptosis assay
1568 APOPTOSIS IN HIV-1 INFECTION
RETRACTED
15 December 2005

measuring annexin V staining in bystander T cells. It is possiblethat during the isolation procedure, biological activity was lost. Wethen developed a panel of murine mAbs and rabbit polyclonal Absby immunizing mice and rabbits with the eight unique subfractionsof fraction 5. We first screened by dot-blot reaction and then at-tempted to block apoptosis induced by crude 43HIV fraction 5. Absdirected against subfractions 2, 5, and 8 of fraction 5 of 43HIV were
all capable of blocking apoptosis in unstimulated target T cells(Fig. 1A).
Identification of the proapoptotic factor
Using the panel of Abs directed against subfractions 2, 5, and 8 offraction 5, we have identified the proapoptotic factor by screeninga cDNA library (�ZAPII) generated from HIV-1-infected 43 cells.
FIGURE 1. FLJ21908 protein. A, Block-ing studies. Abs generated against subfrac-tions 2, 5, and 8 blocked apoptosis inducedby crude 43HIV supernatant. Abs were addedto T cell cultures along with 50% crude 43HIV
supernatant for 2 h at 37°C. The cells werethen simultaneously stained with annexin Vand propidium iodide to assess apoptosis andnecrosis. The percentage of positively stain-ing cells is indicated in the right upper cornerof each panel. This is representative of an ex-periment repeated three times. B, Proteinstructure of the proapoptotic factors of thehypothetical protein FLJ21908. C, Westernblot analysis of the FJ21908 protein. Lysateand supernatant from E. coli expressing theFLJ21908 protein were run on a 10% poly-acrylamide gel, transferred onto nitrocellu-lose paper, blotted with the rabbit polyclonalanti-proapoptotic factor Ab, and then devel-oped. This is representative of an experimentrepeated five times.
1569The Journal of Immunology
RETRACTED
15 December 2005

The 43HIV cDNA library was expressed in E. coli, and in thescreening process the expressed proteins were transferred ontonitrocellulose filters. The expressed proteins were lifted off ontonitrocellulose membranes and the membranes were blocked with milkproteins (Carnation milk 10% w/v) and subsequently incubated withthe rabbit and mouse anti-apoptotic factor Abs (10 �g/ml) or a pre-immune serum/mAb negative control. Ab bound to the expressed pro-teins was detected by incubation with either goat anti-rabbit IgG orgoat anti-mouse IgG conjugated with HRP. Initially, we establishedthe integrity of our library by screening with Abs directed againstproteins expressed in 43HIV cells (e.g., IL-10, IL-8, IL-6, and HIVviral products). We screened the library first by Western blot with therabbit polyclonal Abs and then screened the positive clones by West-ern blotting with the murine mAbs. The plaques that we identifiedwere picked, regrown, rescreened, and expanded, and eventually apure clone was obtained. The cDNA isolated from this clone wassequenced at the Mount Sinai DNA core sequencing facility andfound to be the recently described FL14767485 gene that encodes forthe hypothetical protein FLJ21980 (Fig. 1B).
Our initially described proapoptotic factor had a molecular massof 6000 Da, while the FLJ21908 protein has a molecular mass of66 kDa (49). To reconcile this discrepancy, we performed Westernanalysis on lysates and supernatants from E. coli expressing theFLJ21908 protein (Fig. 1C). In the lysate, a 66-kDa band wasidentified in accord with the molecular mass of the full-lengthFLJ21908 protein, while in the supernatant a doublet of 46-kDaand a 6000-Da band consistent with our previously described fac-tor was identified (Fig. 1c). In our initial characterization of pro-apoptotic activity from the 43HIV supernatant, active proapoptoticfractions were isolated with 95% acetone. Ninety-five percent ac-etone treatment eliminates proteins with molecular mass greater
than 10 kDa, including the 46-kDa doublet from the FLJ21908protein. We purified the 6000-Da peptide from the supernatant ofE. coli expressing the FLJ21908 protein by lyophilizing and re-suspending bacterial supernatant in 100 mM NaCl and 20 mMTris-HCl, pH 7.5, and loading it onto a DEAE Sepharose column.Fractions were eluted by increasing the salt concentration. Theisolated fractions were Western blotted with the rabbit anti-FLJ21908 Abs. The Western blot-positive fractions were then sil-ver stained to ascertain the purity of the protein separation. Frac-tions 39–49 that were eluted with 1 M NaCl demonstrated a bandat 6000 Da consistent with the previously described proapoptoticfactor (Fig. 2).
Biological activity of the FLJ21908 protein
Because we identified the FLJ21908 protein as the proapoptoticfactor, we then wanted to determine whether it had the same ap-optotic activity as our initially defined factor (49). We first usedannexin V staining of unstimulated PBMC as target cells to de-termine the biological activity of the FLJ21908 protein. Superna-tant from the FLJ21908-expressing E. coli induced apoptosis inunstimulated target PBMC (Fig. 3). To ensure that the FLJ21908protein was our initially defined proapoptotic factor, polyclonalrabbit anti-proapoptotic factor and mouse anti-apoptotic mAbswere used to block the apoptotic activity of the FLJ21908 protein(Fig. 3). The rabbit and murine anti-proapoptotic Abs blockedFLJ21908-inducted apoptosis. Similar to our previously describedproapoptotic factor (49), increased apoptosis was also observedwhen the FLJ21908 protein was added to PHA- and anti-CD3-stimulated PBMC (data not shown).
FIGURE 2. The FLJ21908 protein induces apoptosis in unstimulated PBMC. Supernatant from E. coli and supernatant from E. coli containing theFLJ21908 protein were incubated with PBMC for 2 h, and apoptosis was determined by annexin V staining. In blocking experiments, either the rabbitpolyclonal or murine anti-apoptotic factor Abs or preimmune rabbit serum or isotype-specific (IgG1) irrelevant murine mAbs were added to supernatantcontaining the FLJ21908 for 2 h, and apoptosis was evaluated by annexin V staining. The percentage of positively staining cells is indicated in the rightupper corner of each panel. This is representative of an experiment repeated five times using PBMC from different donors.
1570 APOPTOSIS IN HIV-1 INFECTION
RETRACTED
15 December 2005

Transfection of the FL14767485 gene into Bosc cells
To further confirm the apoptotic activity of the FLJ21908 protein, wetransfected the FL14676485-containing plasmid that encodes theFLJ21908 protein and GFP to assess the efficiency of transfection intoa human cell expression system, Bosc cells, and into non-HIV-1-infected 43 cells. Bosc cells are derived from the 243T human embryokidney cell line and are efficiently transfected with CaPO4 (51). Mocktransfection of the Bosc cells using GFP demonstrated that 90% of thecells were transfected (data not shown). We also used CaP04 to trans-
fect the 43 cells with the FL14767485 cDNA. In the 43 cells, theefficiency of the transfection using GFP was 30% (data nor shown).Transfection of the FL14767485 gene with DEAE dextran and Su-perfect produced similar rates of transfection in the 43 cells (data notshown). After transfection, apoptotic activity was determined in thesupernatant from Bosc cells and the 43 cells by annexin V stainingusing unstimulated PBMC as target cells. A dose-dependent increasein annexin V staining was observed in PBMC incubated with super-natants from the Bosc and 43 cells transfected with FL14767485gene, but not in Bosc and 43 cells transfected with GFP (Fig. 4A). Todemonstrate that the FLJ21908 protein is necessary and sufficient forapoptotic activity, we made an Ab affinity column using the three Absthat blocked apoptosis against subfractions 2, 5, and 8 (Fig. 1A). Wepassed supernatant from 43HIV and E. coli transfected with theFL14676485 gene over the Ab affinity column and assessed apoptoticactivity as determined by intracytoplasmic staining using FITC-la-beled Abs directed against activated caspase-3. There was no apopto-tic activity from supernatant passed over the Ab column in both 43HIV
and E. coli transfected with the FL14676485 gene using PBMC as thetarget cells (Fig. 4B).
Demonstration of FLJ21908 protein
We next preformed Western blot analysis using the supernatantand lysate of the untransfected and transfected 43 and Bosc cells
FIGURE 3. Purification of the 6000-Da peptide. Two liters of E. colisupernatant containing the FLJ21908 protein were lyophilized and thenpassed over a DEAE Sepharose column, increasing the concentration ofNaCl from 100 mM to 1 M eluted fractions. Only fractions that wereWestern blot positive were run on a 10% polyacrylamide gel and silverstained. This representative of an experiment was repeated three times.
FIGURE 4. Transfection with FL14676485. A,Supernatant from the FL14676485-transfected Boscand 43 cells induces apoptosis in target PBMC. Dif-ferent concentrations (50, 25, 10, and 0%) of super-natant from the FL14676485- and GFP-transfectedBosc and 43 cells were cocultured with freshly iso-lated PBMC for 2 h and then assessed for apoptosisby annexin V staining. The percentage of positivelystaining cells is indicated in the right upper cornerof each panel. This is representative of an experi-ment repeated five times. B, Supernatant from 43HIV
and E. coli transfected with FL14676485 43 werepassed over an anti-proapoptotic Ab column, cocul-tured with freshly isolated PBMC for 2 h, and thenassessed for apoptosis by staining with FITC-labeledAbs directed against active caspase-3. The percent-age of positively staining cells is indicated in theright upper corner. This is representative of an ex-periment repeated three times.
1571The Journal of Immunology
RETRACTED
15 December 2005

along with 43HIV cells using the polyclonal rabbit anti-proapop-totic factor Abs to determine whether the 6000-Da peptide wasbeing produced (Fig. 5). A protein with a molecular mass of 66kDa corresponding to the FLJ21908 protein was detected in thelysate of the FL14676485-transfected 43 and Bosc cells and 43HIV
cells, while a doublet of 46- and a 6-kDa band were found in thesupernatant. The 6000-Da molecular mass protein corresponds toour previously described proapoptotic factor. There was no detect-able protein in either the lysate or supernatant of the untransfected43 and Bosc cells (Fig. 5A). We next determined whether RNA forthe FLJ21908 protein is constitutively expressed in 43 cells orwhether it is induced after HIV-1 infection. We investigated byPCR whether we could demonstrate the presence of RNA for theFL14767485 gene in the uninfected 43 cells and in 43HIV cells.Forty-three cells were either left alone in culture or infected witheither HIV-1IIIB or HIV-1BaL for 35 days, and RNA was harvestedfor PCR analysis. Usage of HIV-1IIIB base pair fragments consis-tent with the predicted size of the FL14767485 (420 bp) was ob-served in the 43HIV cells, but not in the non-HIV-1-infected 43cells, demonstrating that HIV-1 infection induces the FLJ21908
protein (Fig. 5B). Actin (661 bp) was the positive control. Similarresults were observed with 43 cells infected with HIV-1BaL (datanot shown). In these experiments, we used uninfected 43 super-natant for the mock infection. We have tried to induce proapoptoticactivity from 43 cells and primary monocytes using UV-treatedHIV-1, but failed. Direct HIV-1 infection of 43 cells and primarymonocytes appears to be necessary for FLJ21908 production be-cause infection with HIV-2 and influenza virus did not induce theproduction of the FLJ21908 protein (data not shown). We havealso used different HIV-1 proteins, including pol, gag, env, rev, tat,and nef, to alter the expression of the FLJ21908 protein in bothHIV-1-infected and uninfected 43 cells and monocytes, and againfound that there was no effect (data not shown). We were alsounable to induce production of the FLJ21908 protein in 43 cellsand primary monocytes using different cytokines, includingTNF-�, IL-10, IL-12, IFN-�, and GM-CSF. These cytokines alsofailed to either up-regulate or down-regulate the production of theFLJ21908 protein in 43HIV cells and primary HIV-1BaL-infectedmonocytes (data not shown). In addition, HIV-1-infected primaryT cell and T cell lines did not produce the FLJ21908 protein (data
FIGURE 5. Production of the FLJ21908 protein. A,Western blot analysis of the lysate and supernatantfrom FL14767485 cDNA-transfected and untrans-fected 43 and Bosc cells and 43HIV cells. Forty-eighthours after transfection of the 43 and Bosc cells, thetransfected and untransfected cells along with 43HIV
were lysed, and the lysates and supernatants were runon a 10% polyacrylamide gel, analyzed by Westernblot analysis using the rabbit polyclonal anti-apoptoticfactor Abs, and then developed. This representative ofan experiment was repeated three times. B, PCR anal-ysis for FLJ14676485 mRNA in 43 and 43HIV cells.RNA was extracted from 43 and 43HIV cells, reversetranscribed, probed with an FLJ21908 or actin probe,then run on a 5% agarose gel. Omitting the RNA fromthe DNA amplification step performed negative con-trols. This is representative of an experiment repeatedfive times. c, Primary monocytes were isolated fromPBMC and then infected with HIV-1BaL for 14 days.The cells were lysed, run on a 10% polyacrylamidegel, transferred to nitrocellulose membranes, analyzedby Western blot using the polyclonal rabbit anti-proapoptotic factor Ab, and then developed.
1572 APOPTOSIS IN HIV-1 INFECTION
RETRACTED
15 December 2005

not shown). We infected 10 different monocyte preparations withHIV-1BaL to determine whether the FLJ21908 protein was producedin vivo to validate our findings in the 43 cells. Similar to our previousdescription of the 6000-Da proapoptotic factor, we found that 3 of 10monocyte preparations produced the FLJ21908 after HIV-1BaL infec-tion. Interestingly, Western blot analysis demonstrated the presence ofonly the 6000-Da peptide, but not the 46-kDa doublet (Fig. 5c).
Apoptotic signaling pathways used by the FLJ21908 protein
Although it is beyond the scope of this study to investigate in detailthe mechanisms involved in FLJ21908-induced apoptosis, it wouldbe important to identify some of the apoptotic pathways. The ma-jor effectors of the apoptotic process are the caspase enzymes.Caspases exist in cells in physical associations with adaptor pro-teins that serve as connections between the caspases and upstreamregulators of apoptosis (59). Caspase-3 is the major effectorcaspase in apoptosis and can be activated by other caspases, in-cluding caspase-8 and caspase-9 (59). Caspase-8 is a 55-kDa cy-tosolic protein with homology with CD95/Fas-associated signaltransducer, Fas-associated death domain protein/MORTI-1, aswell as other caspase cysteine proteases (59). Caspase-8 is pro-duced as a proenzyme (55 kDa) that is proteolytically cleaved intosmaller subunits of 40/36 and 23 kDa (59). Caspase-9 is anothermember of the cysteine protease family that can activate caspase-3(59). Procaspase-9 (44–48 kDa) when activated is processed intolarge (37-kDa) and small (10-kDa) subunits and is activated in thepresence of cytochrome c (59). To determine the apoptotic signal-ing pathways used by the FLJ21908 protein, we used the THB Tcell line. There was a marked increase in the expression of acti-vated caspase-3 following treatment with 50% supernatant fromFL14676485-transfected Bosc cells containing theFLJ21908 pro-tein (Fig. 6A). We next investigated whether the activation ofcaspase-3 was occurring through caspase-8 or caspase-9. To thisend, we performed Western blot analysis for caspase-8 (Fig. 6B)and caspase-9 (Fig. 6C) using lysates from the same THB cells thatdemonstrated activated caspase-3. There was no activation ofcaspase-8 in the THB cells, as a 55-kDa doublet consistent with themolecular mass of procaspase-8 was present (Fig. 6B). In thesame lysates, however, there was activation of caspase-9, as a37-kDa band representing an active subunit of caspase-9 wasdetected (Fig. 6C).
Induction of apoptosis in CD4� and CD8� cells and B cells bythe FJ21908 protein
In the initial studies describing the proapoptotic factor, apoptoticactivity was demonstrated in CD4� and CD8� T cells as well asB cells (49). To further prove that the FLJ21908 protein is ourpreviously described proapoptotic factor, we determined whetherthe supernatant containing the FLJ21908 protein had similar pro-apoptotic activity. We isolated purified CD4� and CD8� T and Bcell populations and used different concentrations of supernatantcontaining the FLJ21908 protein (50, 25, 10, and 0%) to demon-strate the induction of apoptosis. Similar to the 43HIV-derived pro-apoptotic factor, apoptotic activity from supernatants of theFL14676485-transfected Bosc cells containing the FLJ21908 pro-tein was demonstrated for CD4�, CD8� T cells, and B cells byannexin V staining (Fig. 7).
Demonstration of apoptotic activity in murine splenocytes
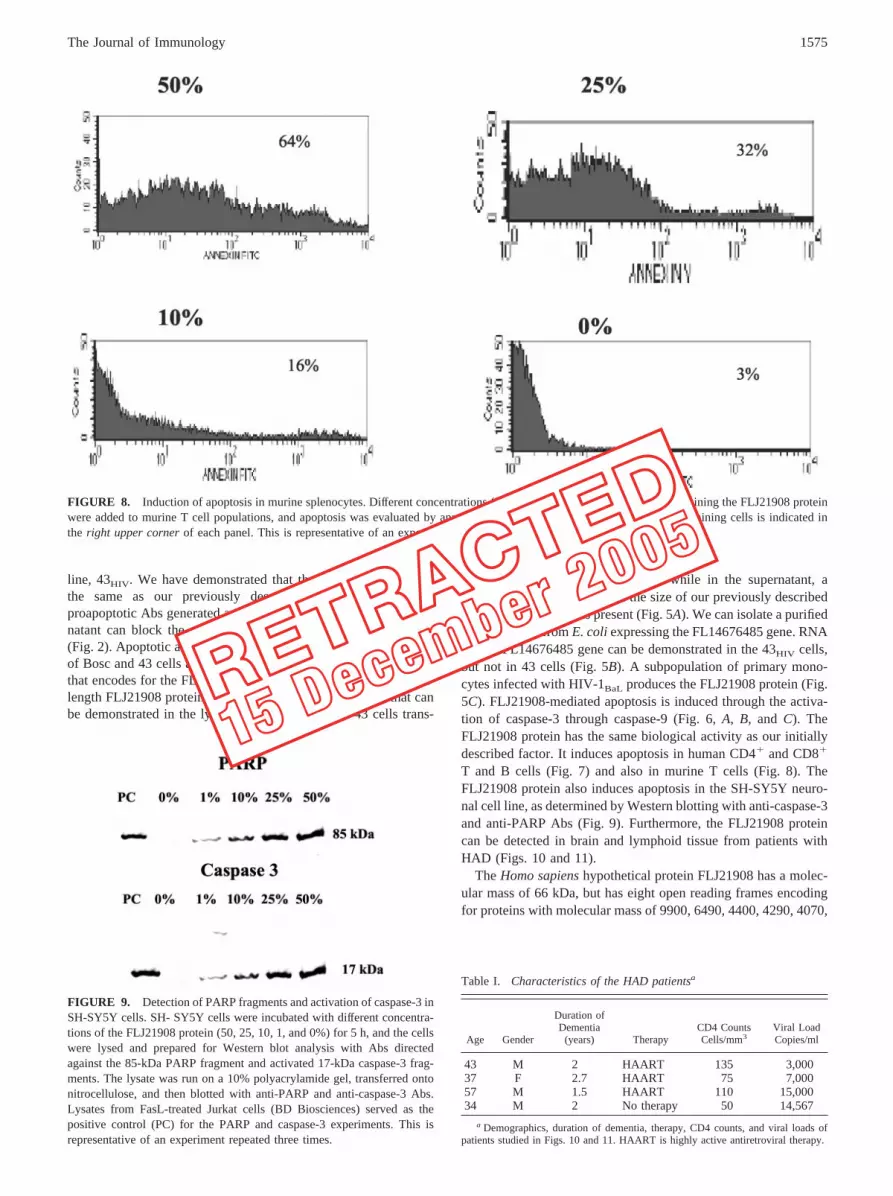
In our initial studies characterizing the proapoptotic factor, weused human PBMC to demonstrate apoptosis (49). It is possiblethat the proapoptotic factor might be conserved in other animalspecies. If the FLJ21908 protein has proapoptotic activity in otherspecies, it would be particularly useful in further studies to deter-mine its biologic significance. To test this hypothesis, we at-tempted to induce apoptosis in murine splenocytes generously pro-vided by T. Moran (Mount Sinai School of Medicine, Departmentof Microbiology). Different concentrations of the FLJ21908 pro-tein (50, 25, 10, and 0%) from the supernatants of Bosc cells wereadded to the murine splenocytes for 2 h, similar to the approachthat we used with the human T cells and apoptosis assessed byannexin V staining. In line with the results obtained with the hu-man T cells, supernatant containing the FLJ21908 protein inducedapoptosis in the murine splenocytes (Fig. 8).
Role of the FLJ21908 protein in neuronal apoptosis
Apoptosis of neurons is a prominent feature of AIDS dementia(41). Macrophages play an important role in this process. Macro-phages and microglia cells infected with HIV-1 produce neurotox-ins that damage neurons by releasing excitotoxins that produceexcessive activation of glutamate receptors, primarily of theNMDAR (41). To determine a role for the FLJ21908 protein inneuronal apoptosis, we investigated whether the FLJ21908 protein
FIGURE 6. Apoptotic signaling pathways. A,Treatment with the FLJ21908 protein activatescaspase-3 in the THB cell line. THB cells weretreated with 50% supernatant from the FL14676485-transfected Bosc cells for 16 h or left untreated andthen stained with FITC-labeled Abs directed againstactivated caspase-3. The percentage of positivelystaining cells is indicated in the right upper cornerof each panel. This is representative of an experi-ment repeated three times. B, The same THB cellsthat were used in A were lysed, run on a 10% poly-acrylamide gel, transferred onto nitrocellulose pa-per, incubated with anti-caspase-8 Abs, and then de-veloped. Fas-treated Jurkat cells (BD Bioscience)served as the positive control (PC). This is represen-tative of an experiment repeated three times. C, Thesame THB cells that were used in A were run on a10% polyacrylamide gel, transferred onto nitrocel-lulose paper, incubated with anti-caspase-9 Abs, andthen developed. Cytochrome c-treated U937 cells(BD Bioscience) served as the positive control (PC).This is representative of an experiment repeatedthree times.
1573The Journal of Immunology
RETRACTED
15 December 2005

induces apoptosis in the neuroblastoma cell line SH-SY5Y. Inthese experiments, different concentrations of the FLJ21908 pro-tein (50, 25, 10, and 0%) derived from the supernatants of Bosccells were added to cultures of the neuronal cell line SH-SY5Y,and apoptosis was assessed by the induction of caspase-3 andPARP. Active capase-3 consists of a heterodimer of 17- and 12-kDa subunits that are derived from the 32-kDa proenzyme. Activecaspase-3 proteolytically cleaves and activates other caspases aswell as relevant targets in the cytoplasm, e.g., D4-GDI and Bcl-2and PARP in the nucleus. PARP is a 116-kDa nuclear chromatin-associated enzyme that catalyzes the transfer of ADP-ribose unitsfrom NAD� to a variety of nuclear proteins, including topoisom-erases, histones, and PARP itself (59). During apoptosis, PARP iscleaved from its 166-kDa intact form into 85- and 25-kDa frag-ments. In the SH-SY5Y cultures incubated with supernatant fromBosc cells containing the FLJ21908 protein, the 17-kDa subunit ofcaspase-3 (60) was demonstrated along with the 85-kDa fragment(61) of PARP (Fig. 9).
Presence of the proapoptotic factor in macrophages in HAD
We next investigated whether we could detect the presence of theFLJ21908 protein in histological sections from patients with HAD.In these experiments, we used normal brain, Alzheimer’s disease,and non-HIV-1 encephalitis as controls. We were also able to ob-tain lymph tissue from the same patients. Mount Sinai MedicalCenter is a part of a national NeuroAIDS consortium that provideswell-characterized CNS and peripheral nervous system tissue sam-
ples and fluids from HIV-1-infected patients (62). All of the pa-tients that were studied met the criteria for HAD as established bythe NeuroAIDS consortium (62) (Table I). S. Morgello of the Pa-thology Department at the Mount Sinai School of Medicine gen-erously provided these tissue sections. Widespread reactive astro-cytosis, myelin pallor, and infiltration predominantly bymonocytoid cells, including blood-derived macrophages, residentmicroglia, and multinucleated giant cells, characterize the neuro-pathology associated with HIV infection of the brain (63). Neuro-logical apoptosis is not specific for HAD, but is a feature of manydifferent types of dementia caused by infectious agents and otherneurodegenerative diseases, including Alzheimer’s disease (41).Using the murine anti-FLJ21908 Ab, punctate green staining con-sistent with the presence of the FLJ21908 protein was detected inpatients with HAD, but not in normal, Alzheimer’s disease or non-HIV-1 encephalitis patients (Fig. 10). We also stained lymphnodes from the same patients for the presence of the FLJ21908protein that was used to study brain tissue. Similar to the resultsobtained with the brain tissue, the FLJ21908 protein was present inlymph nodes from the HIV-1-infected patients (Fig. 11).
DiscussionWe have extended our previous studies that first described apo-ptosis in T and B cells induced by chronically HIV-1-infectedhuman macrophage hybridomas. We have identified a proapoptoticfactor, the FLJ21908 protein (Fig. 1, B and C), produced by thechronically HIV-1-infected human macrophage hybridoma cell
FIGURE 7. Purified populations of CD4� and CD8� T and B cells were isolated by RosetteSep and incubated with different concentrations (50, 25, 10,and 0%) of the FLJ21908 protein for 2 h, and apoptosis was determined by annexin V staining. The percentage of positively staining cells is indicated inthe right upper corner of each panel. This is representative of an experiment repeated five times.
1574 APOPTOSIS IN HIV-1 INFECTION
RETRACTED
15 December 2005
line, 43HIV. We have demonstrated that the FLJ21908 protein isthe same as our previously described factor because anti-proapoptotic Abs generated against partially purified 43HIV super-natant can block the apoptotic activity of the FLJ21908 protein(Fig. 2). Apoptotic activity can be demonstrated in the supernatantof Bosc and 43 cells after transfection with the FL14767485 genethat encodes for the FLJ21908 protein (Fig. 4, A and B). The full-length FLJ21908 protein has a molecular mass of 66 kDa that canbe demonstrated in the lysate of 43HIV, Bosc, and 43 cells trans-
fected with the FL14767485 gene, while in the supernatant, a6000d protein consistent with the size of our previously describedproapoptotic factor was present (Fig. 5A). We can isolate a purified6000d peptide from E. coli expressing the FL14676485 gene. RNAfor the FL14676485 gene can be demonstrated in the 43HIV cells,but not in 43 cells (Fig. 5B). A subpopulation of primary mono-cytes infected with HIV-1BaL produces the FLJ21908 protein (Fig.5C). FLJ21908-mediated apoptosis is induced through the activa-tion of caspase-3 through caspase-9 (Fig. 6, A, B, and C). TheFLJ21908 protein has the same biological activity as our initiallydescribed factor. It induces apoptosis in human CD4� and CD8�
T and B cells (Fig. 7) and also in murine T cells (Fig. 8). TheFLJ21908 protein also induces apoptosis in the SH-SY5Y neuro-nal cell line, as determined by Western blotting with anti-caspase-3and anti-PARP Abs (Fig. 9). Furthermore, the FLJ21908 proteincan be detected in brain and lymphoid tissue from patients withHAD (Figs. 10 and 11).
The Homo sapiens hypothetical protein FLJ21908 has a molec-ular mass of 66 kDa, but has eight open reading frames encodingfor proteins with molecular mass of 9900, 6490, 4400, 4290, 4070,
FIGURE 8. Induction of apoptosis in murine splenocytes. Different concentrations (50, 25, 10, and 0%) of supernatant containing the FLJ21908 proteinwere added to murine T cell populations, and apoptosis was evaluated by annexin V staining. The percentage of positively staining cells is indicated inthe right upper corner of each panel. This is representative of an experiment repeated three times.
FIGURE 9. Detection of PARP fragments and activation of caspase-3 inSH-SY5Y cells. SH- SY5Y cells were incubated with different concentra-tions of the FLJ21908 protein (50, 25, 10, 1, and 0%) for 5 h, and the cellswere lysed and prepared for Western blot analysis with Abs directedagainst the 85-kDa PARP fragment and activated 17-kDa caspase-3 frag-ments. The lysate was run on a 10% polyacrylamide gel, transferred ontonitrocellulose, and then blotted with anti-PARP and anti-caspase-3 Abs.Lysates from FasL-treated Jurkat cells (BD Biosciences) served as thepositive control (PC) for the PARP and caspase-3 experiments. This isrepresentative of an experiment repeated three times.
Table I. Characteristics of the HAD patientsa
Age Gender
Duration ofDementia
(years) TherapyCD4 CountsCells/mm3
Viral LoadCopies/ml
43 M 2 HAART 135 3,00037 F 2.7 HAART 75 7,00057 M 1.5 HAART 110 15,00034 M 2 No therapy 50 14,567
a Demographics, duration of dementia, therapy, CD4 counts, and viral loads ofpatients studied in Figs. 10 and 11. HAART is highly active antiretroviral therapy.
1575The Journal of Immunology
RETRACTED
15 December 2005

4100, 3960, and 3740 Da (National Center for Biotechnology In-formation Open Reading Frame Finder). In addition, motif analy-sis of the FLJ21908 protein revealed two tetratricopeptide (TPR)repeats, which is a repeat structure of 34 aa first described in yeastand later found to occur in a large number of proteins (64) (Fig.1a). A common feature of TRP repeats is protein-protein interac-tions (65). It has also been proposed that TPR proteins preferen-tially interact with WD-40 repeat proteins, but in many instancesTPR aggregate to form multiprotein complexes (64). TPR repeatshave been implicated in apoptosis (66). In our initial characteriza-tion of the proapoptotic activity from the 43HIV supernatant, wedetermined by molecular mass fractionation that proapoptotic ac-tivity was present from the fractions that corresponded to a mo-lecular mass less than 10,000 Da (49). Furthermore, fractionationof supernatants from HIV-1BAL-infected monocytes that inducedapoptosis in target PBMC revealed that activity was also present inthose fractions with a molecular mass less than 10 Da similar to the43HIV cell line (49). The role of the full-length 66-kDa FLJ21908protein in apoptosis is unclear. However, the 66-kDa protein is notsecreted in 43HIV cells and in uninfected 43 and Bosc cells after
transfection with the FL14676485 gene (Fig. 5A). We are presentlyattempting to obtain sufficient quantities of the 6000d fragment ofthe FLJ21908 protein for protein sequence analysis.
The regulation of the FLJ21908 protein as it relates to apoptosisinvolves not only the induction of the FL14676485 gene, but alsoits cleavage into its 6000-Da proapoptotic form. Bosc cells trans-fected with the FL14676485 gene produce the 6000-Da peptide,demonstrating that normal host enzymes are capable of cleavingthe parent FLJ21908 protein (Fig. 4). Apoptotic activity from the43HIV cell line occurred only after prolonged infection (greaterthan 35 days) (49). As noted above, in the transfection experiments(Fig. 5) Bosc cells and uninfected 43 cells produced the 6000-Dapeptide, proving that the 66-kDa protein is cleaved by host pro-teases. It is possible that other factors may be involved in thegeneration of the 6000-Da peptide from the parent FLJ21908 pro-tein. One possibility is HIV-1 protease that is well known to cleavehost cell proteins and transfected HIV-1 protease kill host cells(67). HIV-1 protease inhibitor-based therapy to treat AIDS inhibitsapoptosis (68). HIV-1 protease inhibitors have had little impact onHAD because they have limited penetration across the blood-brain
FIGURE 11. Detection of the FJ21908 proteinfrom lymph nodes. Immunofluorescence was alsoperformed on sections of lymph nodes from thesame patients in Fig. 10. The sections were stainedwith FITC-labeled murine anti-proapoptotic factorAb and analyzed by confocal microscopy. Isotype-matched FITC-labeled IgG1 served as the negativecontrol. Two observers routinely observed 10 sepa-rate fields.
FIGURE 10. Detection of the proapoptoticfactor in patients with HAD. Immunofluorescencestaining was performed using tissue sections fromnormal brain, HAD, Alzheimer’s disease, andnon-HIV-1 encephalitis. The sections werestained with murine FITC-labeled anti-FLJ21908Abs and analyzed by confocal microscopy. Iso-type-matched FITC-labeled IgG1 served as thenegative control. Two observers routinely ob-served 10 separate fields.
1576 APOPTOSIS IN HIV-1 INFECTION
RETRACTED
15 December 2005

barrier (69, 70). The FLJ21908 protein may be induced by otherpathogens and not be specific to HIV-1. However, HIV-2 and in-fluenza infection of 43 cells did not result in the production of theFLJ21908.
The FLJ21908 protein induces apoptosis through the activationof caspase-9 (Fig. 6). Members of the Bcl-2 family control signal-ing of apoptosis through caspase-9. Studies are in progress to bet-ter characterize the apoptotic signaling of the FLJ21908 protein.The FLJ21908 protein induces apoptosis in CD4� and CD8� Tand B cells (Fig. 7) and can induce apoptosis in murine splenocytes(Fig. 8). The FLJ21908 protein may have a broader biological role.Although we identified the FLJ21908 protein as being produced byHIV-1-infected macrophages, it is possible that other cell typeshave the capacity to produce this protein, and in fact it may be anormal constituent of cell growth and regulation. This may notonly play a role in the T cell loss during HIV-1 infection, but maybe important in the regulation of inflammatory responses.
Although it has been proposed that HIV-1 disease progression isassociated with a shift in cytokine profile from type 1 to type 2profiles in vitro (71), the dominant in vivo shift is that of inducedTNF-�, IFN-�, IL-10, and IL-6 with reduced IL-2, as determinedby studying cytokine mRNA expression in lymph nodes (72).These cytokines have been implicated in the induction of apoptosisand may further contribute to HIV-1 disease progression by aug-mentation of apoptosis induced by the FLJ21908 protein. The keyquestion in our studies is whether the FLJ21908 protein contrib-utes to the cell loss in HIV-1-infected patients. One area of par-ticular interest is the neurological complications of HIV-1 infec-tions. As noted above, in HIV-1 infection of the CNS, the virus isselectively localized within the perivascular and infiltrating paren-chyma blood-borne macrophages and microglia (73). In particular,a subpopulation of macrophages, CD16highCD14low has been iden-tified as being important in the neurological manifestations ofAIDS (74). This subpopulation of macrophages binds to endothe-lial cells in the brain, migrates into the brain, and causes neuronaldamage. Interestingly, the human macrophage hybridomas includingclone 43 are CD16highCD14low (50). These activated macrophagesadhere to the normal endothelium of the brain microvasculature,transmigrate, and then trigger a number of deleterious processes.
The discrepancy between the number and localization of HIV-1-infected cells and the severity of the neurological abnormalitiesin AIDS has suggested that soluble factors produced by the in-fected brain macrophages may be responsible for the pathologyseen (41). Different cytokines, including IL-1 and TNF-�, chemo-kines (75), along with excitotoxic substances that cause excessiveactivation of NMDARs that damage neurons contribute to neuro-nal loss in HIV-1-infected patients (76). The FLJ21908 proteininduced apoptosis in the SH-SY5Y neuronal cell lines (Fig. 9), andby immunofluorescence it was present in brain and lymph tissuefrom patients with HAD (Figs. 10 and 11). The FLJ21908 proteinmay directly induce apoptosis, or alternatively, it might be an ex-citotoxin. Because we only used one neuronal cell line and per-formed immunofluorescence in only four patients in each of thestudy groups, HAD, Alzheimer’s disease, encephalitis, and nor-mal, it is difficult to make any generalizations regarding the role ofthe FLJ21908 protein in the neurological manifestations of HIV-1infections. We are extending these preliminary experiments to bet-ter characterize the mechanism by which the FLJ21908 proteininduces apoptosis in both lymphoid and neuronal cells, and tostudy more patients.
In conclusion, we have previously described a soluble 6000dpeptide produced by an HIV-1-infected human macrophage hy-bridoma, clone 43HIV, that induces apoptosis in bystander, Ag-,and mitogen-stimulated T cells, as well as B cells. We have iden-
tified this factor as the novel cDNA clone FL14676485 that en-codes for the human hypothetical factor FLJ21908. Apoptosis in-duced by the FLJ21908 protein may contribute to the apoptosisand dementia observed in AIDS patients.
References1. Pantaleo, G., C. Graziosi, and A .S. Fauci. 1993. The immunopathogenesis of
human immunodeficiency virus infection. N. Engl. J. Med. 328:327.2. Oyaizu, N., and S. Pahwa. 1995. Role of apoptosis in HIV pathogenesis. J. Clin.
Immunol. 15:217.3. Meyaard, L., S. A. Otto, R. R. Jonker, M. J. Mynster, R. P. M. Keet, and
F. Miedema. 1992. Programmed cell death of T cells in HIV-1 infection. Science257:217.
4. Groux, H., G. Tarpier, D. Monte, Y. Mounter, A. Capon, and J. C. Ameisen.1992. Activation-induced death by apoptosis in CD4� T cells from human im-munodeficiency virus-infected asymptomatic individuals. J. Exp. Med. 175:331.
5. Oyaizu, N., T. W. McCloskey, M. Coronesi, N. Chirmule, V. S. Kalayanarmann,and S. Pahwa. 1993. Accelerated apoptosis in peripheral blood mononuclear cells(PBMC) from human immunodeficiency virus type-1 infected patients and inCD4 cross-linked PBMC from normal individuals. Blood 82:3392.
6. Corbonari, M., M. Cibota, M. Cherchi, D. Sbarigia, A. M. Pesce, L. Dell’Anna,A. Modica, and M. Fiorelli. 1994. Detection and characterization of apoptoticperipheral blood lymphocytes in human immunodeficiency virus infection andcancer chemotherapy by a novel flow immunocytometric method. Blood 83:1268.
7. Sarin, A., M. Clerici, S. P. Blatt, C. W. Hendrix, G. M. Shearer, andP. A. Henkart. 1994. Inhibition of activation-induced programmed cell death andrestoration of defective immune response of HIV� donors by cysteine proteaseinhibitors. J. Immunol. 153:862.
8. Meyaard, L., S. A. Otto, J. D. M. Keet, M. T. C. Roos, and F. Miedema. 1994.Programmed death of T cells in human immunodeficiency virus infection. J. Clin.Invest. 93:982.
9. Lewis, D. E., D. S. Ng Tang, A. Adu-Oppong, W. Schober, and J. R. Rodgers.1994. Anergy and apoptosis in CD8� T cells from HIV-infected persons. J. Im-munol. 153:412.
10. Diamond, D. C., B. P. Sleckman, T. Gregory, L. A. Lasky, J. L. Greenstien, andS. J. Burakoff. 1988. Inhibition of CD4� T cell function by the HIV envelopeprotein gp120. J. Immunol. 141:3715.
11. Chirmule, N., V. S. Karayanarmann, N. Oyaizu, H. Slade, and S. Pahwa. 1990.Inhibition of functional properties of tetanus-antigen specific T cell clones byenvelope proteins of HIV-1. Blood 75:152.
12. Oyaizu, N., N. Chirmuli, V. S. Karayanarmann, W. W. Hall, R. A. Good, andS. Pahwa. 1990. Human immunodeficiency virus type 1 envelope protein gp120produces immune defects in CD4� T lymphocytes by inhibiting interleukin-2mRNA. Proc. Natl. Acad. Sci. USA 84:2379.
13. Debatin, K.-M., A. Fahrig-Faisser, S. Enenkel-Stoodt, W. Kreuz, A. Brenner, andP. H. Krammer. 1994. High expression of Apo-1 (CD95) on T lymphocytes fromhuman immunodeficiency virus-type-1 infected children. Blood 83:3101.
14. McClosky, T. W., N. Oyaizu, M. Kaplan, and S. Pahwa. 1995. Expression of theFas antigen in patients infected with human immunodeficiency virus. Cytometry22:111.
15. Kabayoshi, N., Y. Hamanato, N. Yamomato, A. Ishu, M. Yonehara, andS. Yonehara. 1990. Anti-Fas monoclonal antibody is cytocidal to human immu-nodeficiency virus-infected cells without augmenting viral replication. Proc.Natl. Acad. Sci. USA 90:7573.
16. Hugin, A. W., M. S. Vacchio, and H. C. Morse III. 1991. A virus-encoded “su-perantigen” in a retrovirus-induced immunodeficiency syndrome of mice. Science252:424.
17. Aziz, D. C., Z. Hammer, and P. Jolicoeur. 1989. Severe immunodeficiency dis-ease induced by a defective murine leukemia virus. Nature 338:505.
18. Pope, M., M. G. H. Betjes, N. Romani, H. Hermonil, P. U. Cameron,L. H. Hoffman, S. Gezelter, G. Schuler, and R.M. Steinman. 1994. Conjugate ofdendritic cells and memory T cells from the skin facilitates productive infectionof HIV-1. Cell 78:389.
19. Mosier, D. E., R. J. Gulizia, P. D. MacIsaac, B. E. Torbett, and J. A. Levy. 1993.Rapid loss of CD4� T cells in human PBL-SCID mice by monocytopathic HIVisolates. Science 260:689.
20. Watanabe, M., R. J. Ringler, P. N. Fultz, J. J. MacKey, J. E. Bolysen,C. G. Levine, and N. L. Levtvin. 1991. A chimpanzee passaged human immu-nodeficiency virus isolate is cytopathic for chimpanzee cells but does not inducedisease. J. Virol. 65:3853.
21. Johnson, B. K., G. A. Stow, M. S. Godec, D. M. Asher, D. C. Gajduseh, andC. J. Gibbs, Jr. 1993. Long-term observation of human immunodeficiency virus-infected chimpanzees. AIDS Res. Hum. Retroviruses 9:375.
22. Gendelman, H. E., G. D. Erlich, L.M. Boca, S. Conley, J. Ribas, D. C. Kotler,M. S. Meltzer, B .J. Poeiz, and P. Nara. 1991. The inability of human immuno-deficiency virus to infect chimpanzee monocytes can be overcome by serial pas-sage in vivo. J. Virol. 65:3865.
23. Schuitemaker, H., L. Mayard, N. A. Kootstra, R. Dubbs, S. A. Otto, M. Ternette,J .L. Heeney, and F. Miedema. 1993. Lack of T cell dysfunction and programmedcell death in human immunodeficiency virus type-1 infected chimpanzees corre-late with the absence of monocytopathic variants. J. Infect. Dis. 168:1140.
24. Badley, A. P., J. A. McElhiney, P. J. Leison, D. H. Lynch, M. R. Alderson, andC. V. Paige. 1996. Up-regulation of Fas ligand expression by human immuno-deficiency virus in human macrophages mediates apoptosis of uninfected T lym-phocytes. J. Virol. 70:199.
1577The Journal of Immunology
RETRACTED
15 December 2005

25. Oyaizu, N., A. Yasusha, F. Hashimoto, T. W. McCloskey, N. Hosaka,N. Kayagaki, H. Yagita, and S. Pahwa. 1997. Monocytes express Fas ligand uponCD4 cross-linking and induce CD4� T cell apoptosis. J. Immunol. 158:2456.
26. Wu, X., J. Daley, R. A. Rasmussen, and S. F. Schlossman. 1994. Monocytes arerequired to prime peripheral blood T cells to undergo apoptosis. Proc. Natl. Acad.Sci. USA 92:1525.
27. Herbein, G., U. Mahlkecht, F. Batliwalla, P. Gregersen, T. Pappas, J. Butler,W. A. O’Brien, and E. Verdin. 1998. Apoptosis of CD8� T cells is mediated bymacrophages through interaction of HIV gp120 with chemokine receptorCXCR4. Nature 395:189.
28. Hesselgesser, J., D. Taub, P. Baskar, M. Greenberg, J. Hoxie, D. L. Kolson, andR. Horuk. 1998. Neuronal apoptosis induced by gp120 and chemokine SDF-1�is mediated by the chemokine receptor CXCR4. Curr. Biol. 8:595.
29. Ameisen, J. C. 1998. Setting death in motion. Nature 395:117.30. Poli, G., and A. S. Fauci. 1992. Effect of cytokine and pharmacologic agents on
chronic HIV infection. AIDS Res. Hum. Retroviruses 8:191.31. Zauli, G., M. Vitale, D. Gibellini, and S. Capitani. 1996. Inhibition of purified
CD34� hematopoietic progenitor cells by HIV-1 or gp120 mediated by endog-enous TGF-�-1. J. Exp. Med. 183:99.
32. Clements, M. V., and I. Stamenkouri. 1994. Fas and tumor necrosis factor re-ceptor-mediated cell death: similarities and distinctions. J. Exp. Med. 180:557.
33. Wang, J., S. A. Stolman, and G. Dennert. 1994. TCR cross-linking induces CTLdeath via of TNF. J. Immunol. 152:3842.
34. Grell, M., G. Zimmermann, D. Hulsen, K. Pfizenmaier, and P. Scheuril. 1993.TNF receptor TR60 and TR80 can mediate apoptosis via induction of distinctsignal pathways. J. Immunol. 153:1963.
35. Liu, Y., and C. A. Janeway. 1990. Interferon-� plays a critical role in induced celldeath of effector T cells: a possible mechanism of self tolerance. J. Exp. Med.172:1735.
36. Groux, H. D., D. Monte, B. Plouvier, A. Capon, and J. C. Ameisen. 1993. CD3mediated apoptosis of medullary thymocytes and activated T cells: respectiveroles of interleukin-1, interleukin-2, interferon-�, and accessory cells. Eur. J. Im-munol. 23:1623.
37. Novelli, F., F. Pierro, P. F. di Celli, S. Bertini, P. Affaticati, G. Garotti, andG. Formi. 1994. Environmental signals influence expression of the IFN-� recep-tors on human T cells control whether IFN-� promotes proliferation or apoptosis.J. Immunol. 152:496.
38. Clerici, M., A. Sarin, R. L. Coffman, T. A. Wynn, S. Blatt, C. W. Hendrix,S. F. Wolf, G. M. Shearer, and P. A. Henkart. 1994. Type 1/type 2 cytokinemodulation of T cell programmed cell death as a model for human immunode-ficiency virus pathogenesis. Proc. Natl. Acad. Sci. USA 91:11811.
39. Lipson, S. A., and H. E. Gendelman. 1995. Dementia associated with acquiredimmunodeficiency syndrome. N. Engl. J. Med. 332:934.
40. Canki, M., J. N. Thai, W. Chao, A. Ghorpade, M. J. Potash, and D. J. Volsky.1997. Isolation and long-term culture of primary ocular human immunodefi-ciency virus type 1 isolates in primary astrocytes. J. Neurovirol. 3:10.
41. Kaul, M., G. A. Garden, and S. A. Lipton. 2001. Pathways to neuronal injury andapoptosis in HIV-asociated dementia. Nature 410:988.
42. Giulian, D., K. Vaca, and C. A. Noonan. 1991. Secretion of neurotoxins bymononuclear phagocytes infected with HIV-1. Science 250:1593.
43. Giulian, D., E. Wendt, K. Vaca, and C. A. Noonan. 1993. The envelope glyco-protein of human immunodeficiency virus type 1 stimulates release of neurotox-ins from monocytes. Proc. Natl. Acad. Sci. USA 90:2769.
44. Dreyer, E. B., P. K. Kaiser, J. T. Offerman, and S. A. Lipton. 1990. HIV-1 coatprotein neurotoxicity prevented by calcium channel antagonists. Science 248:364.
45. Sperber, K., A. Pizzimenti, V. Najfeld, and L. Mayer. 1990. Identification ofsubpopulations of human macrophages through the generation of human macro-phage hybridomas. J. Immunol. Methods 129:31.
46. Sperber, K, G. Hamrung, M. J. Louie, T. Kalb, R. Banerjee, H .S. Choi,F. Paronetto, and L. Mayer. 1993. Progressive impairment of monocytic functionin HIV-1 infected human macrophage hybridomas. AIDS Res. Hum. Retroviruses9:657.
47. Yoo, J., H. Chen, T. Kraus, D. Hirsch, S. Polyak, I. George, and K. Sperber. 1996.Altered cytokine production and accessory cell function after HIV-1 infection.J. Immunol. 157:1313.
48. Polyak, S., H. Chen, D. Hirsch, I. George, R. Herschberg, and K. Sperber. 1997.Impaired class II expression and antigen uptake in monocytic cells after HIV-1infection. J. Immunol. 159:2177.
49. Chen, H., Y. K. Yip, I. George, E. Salik, M. Tyorkin, and K. Sperber. 1998.Induction of apoptosis in co-cultured T cells by HIV-1 infected monocytic cell.J. Immunol. 161:4257.
50. Rakoff-Nahoum, S., H. Chen, T. Kraus, I. George, E. Oei, and K. Sperber. 2001.Regulation of class II expression in monocytic cells after HIV-1 infection. J. Im-munol. 167:2331.
51. Pear, W. S., G. P. Nolan, M. L. Scott, and D. Baltimore. 1993. Production ofhigh-titer helper-free retroviruses by transient transfection. Proc. Natl. Acad. Sci.USA 90:8392.
52. Ross, R. A., B. A. Spengler, and J. L. Biedler. 1983. Coordinate morphologicaland biochemical interconversion of human neuroblastoma cells. J. Natl. CancerInst. 71:741.
53. Bader, P., J. Beck, P. G. Schlegel, R. Handgretinger, D. Niethammer, andT. Klingebiel. 1999. Additional immunotherapy on the basis of increasing mixedhematopoietic chimerism after allogeneic BMT in children with acute leukemia:is there an option to prevent relapse? Transplant 20:79.
54. Lopez, C. B., A. Fernandez-Sesma, J. L. Schulman, and T. M. Moran. 2001.Myeloid dendritic cells stimulate both Th1 and Th2 immune responses dependingon the nature of the antigen. J. Interferon Cytokine Res. 21:763.
55. Sperber, K., E. Gollub, S. Goswami, T. Kalb, L. Mayer, and Z. Marom. 1992. Invivo detection of a novel macrophage-derived protein involved in the regulationof mucus-like glycoconjugate secretion. Am. Rev. Respir. Dis. 146:1589.
56. Fadak, V. A., D. R. Voelker, D. A. S. Campbell, J. J. Cohen, D. L. Bratton, andP. M. Henson. 1992. Exposure of phosphatidylserine on the surface of apoptoticlymphocytes triggers specific recognition and removal by macrophages. J. Im-munol. 148:2207.
57. Koopman, G., C. P .M. Reidelingsperger, G. A. M. Kuijtessin, R. M. J. Keelmen,S. T. Pals, and M. H. J. Van Oeis. 1994. Annexin V for flow cytometric detectionof phosphatidylserine expressed on B cells undergoing apoptosis. Blood 84:1415.
58. Short, J. M., J. M. Fernandez, J. A. Sorge, and W. P. Huse. 1989. Effect of �packaging extract MCR restriction activity on DNA cloning. Nucleic Acids Res.16:7583.
59. Strasser, A., L. O’Connor, and V. M. Dixit. 2000. Apoptosis signaling. Annu.Rev. Biochem. 69:217.
60. Patel, T., G. J. Gores, and S. H. Kaufmann. 1996. The role of proteases duringapoptosis. FASEB J. 10:587.
61. D’Amores, D., S. Desnoyers, I. D’Silva, and G. G. Poirier. 1999. Poly (ADP-ribose) in the regulation of nuclear functions. Biochem. J. 342:249.
62. Morgello, S., B. B. Gelman, P. B. Kozlowski, V. H. Winters, E. Masliah,M. Conford, W. Cavett, C. Marra, and E. J. Singer. 2001. The National Neu-roAIDS Tissue Consortium: a new paradigm in brain banking with an emphasison infectious disease. Neuropathol. Appl. Neurobiol. 27:326.
63. Lipson, S. A., and H. E. Gendelman. 1995. Dementia associated with acquiredimmunodeficiency syndrome. N. Engl. J. Med. 332:934.
64. Das, A. K., and G. T. W. Cohen. 1998. The structure of the tetratricopeptiderepeats of protein phophatases: implications for TRP-mediated protein-proteininteractions. EMBO J. 17:11192.
65. Lamb, J. R., S. Tugendreich, and P. Hieter. 1995. Tetratrico peptide repeatsinteractions: to repeat or not to TRP? Trends Biochem. Sci. 20:257.
66. Demonacos, C., M. K. Demonacos, and N. B. Thangue. 2001. A TPR motifcofactor contributes to p300 activity in the p53 response. Mol. Cell 8:71.
67. Memon, S. A., D. Petrak, M. B. Moreno, and C. M. Zacharchuk. 1995. A simplemethod for examining the effect of transiently expressed genes on programmedcell death. J. Immunol. Methods 180:15.
68. Bohler, T., J. Walcher, G. Holzl-Wenig, M. Geiss, B. Buchholz, R. Linde, andK. M. Debatin. 1999. Early effects of antiretroviral combination therapy on ac-tivation, apoptosis and regeneration of T cells in HIV-1-infected children andadolescents. AIDS 13:779.
69. Dore, G. J., P. K. Correll, Y. Li, J. M. Kaldor, D. A. Cooper, and B. J. Drew.1999. Changes to AIDS dementia complex in the era of highly active antiretro-viral therapy. AIDS 13:1249.
70. Enting, R. H., R. M. Hoetelmans, J. M. Lange, D. M. Burger, J. H. Beijnen, andP. Portegies. 1998. Antiretroviral drugs and the central nervous system. AIDS12:1941.
71. Shearer, G. M., and M. Clerici. 1998. Cytokine profiles in HIV type 1 disease andprotection. AIDS Res. Hum. Retroviruses 14 (Suppl. 2):S149.
72. Graziosi, C., G. Pantoleo, K. R. Gantt, J. P. Fortin, J. F. Demarrest, O. J. Cohen,R. P. Sekaly, and A. S. Fauci. 1994. Lack of evidence for the dichotomy of TH1and TH2 predominance in HIV-infected individuals. Science 265:248.
73. Koenig, S., H. Gendelman, and J. M. Orenstein. 1986. Detection of AIDS virusin macrophages in brain tissue from patients with encephalopathy. Science 233:1089.
74. Fisher-Smith, T., S. Croul, A. E. Sverstuik, C. Capini, D. L’Heureux,E. G. Reguilar, M. W. Richardson, S. Amini, S. Morgello, K. Khalili, andJ. Rapport. 2001. CNS invasion of CD14�/CD16� peripheral blood-derivedmonocytes in HIV dementia: perivascular accumulation and reservoir of HIVinfection. J. Neurovirol. 7:528.
75. Asensio, V., and I. L. Campbell. 1999. Chemokines in the CNS: plurifunctionalmediators in diverse states. Trends Neurosci. 22:504.
76. Gartner, S. 2000. HIV infection and dementia. Science 287:602.
1578 APOPTOSIS IN HIV-1 INFECTION
RETRACTED
15 December 2005