retinol binding protein in rainbow trout: molecular properties and mrna expression in tissues
TRANSCRIPT

A
rhd
hs
General and Comparative Endocrinology 123, 51–61 (2001)doi:10.1006/gcen.2001.7636, available online at http://www.idealibrary.com on
Retinol Binding Protein in Rainbow Trout: MolecularProperties and mRNA Expression in Tissues1
Marei Sammar,* Patrick J. Babin,† Michele Durliat,‡ Iris Meiri,§ Itai Zchori,¶bigail Elizur,§,2 and Esther Lubzens*,3
*Israel Oceanographic and Limnological Research, Haifa 31080, Israel; †Genomique et Physiologie des Poissons, USCINRA, Universite Bordeaux I, 33405 Talence Cedex, France; ‡UPRESA 8080 CNRS, Universite Paris-Sud,91405 Orsay Cedex, France; §Israel Oceanographic and Limnological Research, Eilat, Israel;and ¶Department of Zoology, Tel-Aviv University, Tel Aviv, Israel
Accepted February 2, 2001
Retinoids are important regulatory signaling molecules rainbow trout, 17b-estradiol treatment led to a decrease
A
during embryonic development. The molecular proper-ties of rainbow trout (Oncorhynchus mykiss) retinol-bind-ing protein (rtRBP), the specific retinol carrier in verte-brate plasma, were studied to elucidate its role intransporting retinols to developing fish oocytes. A 954-nucleotide rtRBP cDNA was cloned from the liver codingfor a 176-amino-acid (aa) mature protein, with an esti-mated molecular mass of 20,267 Da. The nucleotide se-quence suggests a putative 16-aa signal peptide andshows all the aa residues that were previously identifiedas critical for the retinol binding pocket. Five of the eightamino acid residues that are associated with the interac-tion of RBP and transthyretin in mammalian and non-mammalian species are conserved. The deduced aa se-quence of rtRBP shows 60–66% identity with zebrafish,chicken, mouse, rat, horse, bovine, and human RBPs and56% identity with Xenopus RBP. Northern blot analysisevealed a ;1.1-kb hepatic mRNA transcript. RBP isighly expressed in the liver, but low levels were alsoetected in the spleen, kidney, ovary, and brain. In the
1 The rainbow trout RBP cDNA sequence reported in this paperas been deposited in GenBank/EMBL Data Library under Acces-ion No. AF257326.
2 Current address: Bribie Island Aquaculture Research Centre,Queensland Department of Primary Industries, Queensland 4507,Australia.
3 To whom correspondence should be addressed.
0016-6480/01 $35.00Copyright © 2001 by Academic PressAll rights of reproduction in any form reserved.
51
in the RBP mRNA signal relative to that of the controls.The efficacy of the 17b-estradiol treatment was verifiedby an induction of vitellogenin (VTG) mRNA expressionin the liver and occurrence of VTG in the plasma. © 2001
cademic Press
Key Words: retinol-binding protein; cDNA; trout; vitel-logenin; liver; 17b-estradiol.
INTRODUCTION
Retinoids are a family of low-molecular-weight hy-drophobic molecules that are derived from, or arestructurally related to, retinol, known also as vitaminA, and function as important regulatory signalingmolecules (mainly as retinoic acid; RA). In the adultanimal, retinoids are essential for growth, vision(mainly as retinal), reproduction, hematopoiesis, im-mune function, and maintenance of epithelial cells.During embryogenesis, RA regulates embryonic bodypatterning involving the action of homeobox tran-scription factors in diverse animals ranging from nem-atodes to insects to mammals. Both deficiency in vita-min A and administration of teratogenic doses of RAresult in congenital malformations of many structures(Gudas, 1994; Conlon, 1995; Means and Gudas, 1995;Herman, 1995; Curley and Robage, 1997; Morris-Kay,

1997; Redfern, 1997; Takeuchi et al., 1998). The mode of bound to serum retinol-binding protein (RBP), a ;21-kapToasc1c(K1oo
R
y(a
(omec
52 Sammar et al.
retinoid transport to developing oocytes in oviparousvertebrates has therefore gained significant attentionin studies performed in Xenopus and chicken (Mc-Kearin et al., 1987; McKearin and Shapiro, 1988; Vieiraand Schneider, 1993; Vieira et al., 1995a,b), but similarstudies are not available for teleosts.
The retinoids and their carotenoid precursors arenot synthesized de novo by vertebrates. Vitamin A is apolyisoprenoid compound containing a cyclohexenylring and includes retinol, retinoic acid, and retinal, butonly retinol has full vitamin A activity. The mainsources of vitamin A are dietary and derived fromvegetable provitamin A carotenoids, retinyl estersfrom animal sources, or dietary supplements. Carote-noids such as b-carotene are cleaved symmetrically toyield two molecules of retinal, whereas other carote-noids may cleave to generate retinol. Retinal is re-duced to retinol in the intestinal mucosa by a specificretinaldehyde reductase, and a small fraction of theretinal is oxidized to retinoic acid. Retinyl esters arehydrolyzed by lipases or esterases in the intestinallumen prior to absorption. Most of the retinol is ester-ified with saturated fatty acids and incorporated intolymph chylomicrons, which enter the blood stream(Curley and Robage, 1997). These are converted tochylomicron remnants, which are taken up by theliver with their content of retinol. In the liver, retinol isstored as an ester in the lipocytes, and for transport itis hydrolyzed and retinol is bound to apo-retinol-binding protein. The resulting holo-retinol-bindingprotein is processed in the Golgi apparatus and se-creted into the plasma. Retinol and retinal are inter-converted by dehydrogenases or reductases, presentin many tissues. However, retinoic acid that is formedfrom retinol or retinal cannot be converted back intothese compounds and, therefore, retinoic acid cannotreplace retinal in its role in vision or retinol in itssupport of the reproductive system. Carotenoids arealso present in the serum bound to low-density li-poproteins (LDL) and presumably enter cells by LDLreceptors, where consequently they can be metabo-lized, via retinal, directly to retinoic acid. Unlike se-rum retinol levels, the level of serum carotenoidschanges with the dietary intake of carotenoids (Curleyand Robage, 1997; Redfern, 1997).
Whereas retinoic acid is transported in the plasmabound to albumin, retinol is mobilized from the liver
Copyright © 2001 by Academic PressAll rights of reproduction in any form reserved.
Da protein that is synthesized by hepatocytes and ismember of the lipocalin family of proteins that trans-ort lipophilic molecules (Pervaiz and Brew, 1987).he liver is one of the major sites for gene expressionf RBP and protein synthesis, but transcripts for RBPnd protein synthesis were also detected in other tis-ues, including the retinal pigment epithelium, kidneyells, and the seminal vesicle epithelium (see Blaner,989; Redfern, 1997). In birds and mammals, RBP cir-ulates in the plasma complexed with transthyretinTTR), a protein carrier of thyroxine (Heller, 1976;opelman et al., 1976a; Blaner, 1989; Blomhoff et al.,990). Three-dimensional studies of human RBP dem-nstrate that one tetramer of TTR binds two moleculesf RBP (Monaco et al., 1995). The association of RBP
with TTR increases the stability of the retinol–RBPcomplex and, it was suggested to prevent filtrationin the renal glomerulus of both the RBP and thebound retinol (Blaner, 1989). Whereas RBP may bethe major carrier of retinol in the blood, retinol mayalso be transported by serum albumin. Althoughretinol has a lower affinity for serum albumin, therelatively larger concentration of albumin can po-tentially transport significant amounts of retinol. Inthis case, the transfer of retinol will lack the speci-ficity afforded by cell surface receptors for retinolbound to RBP (Noy and Xu, 1990).
RBP has been purified from the plasma of severalnonmammalian vertebrates, including chicken, snap-ping turtle, bullfrogs, and fish [eel (Anguilla japonica),carp (Cyprinus carpio), blue shark (Pronace glauca), yel-lowtail (Seriola quinqueradia), and trout (Oncorhynchusmykiss)] (Mokady and Tal, 1974; Kopelman et al.,1976a,b; Shidoji and Muto, 1977; Hayashi et al., 1990;Berni et al., 1992; Vieira and Schneider, 1993]. Trout
BP was purified as a ;21-kDa protein and its aminoacid sequence was determined by Edman degradation(Zapponi et al., 1992). RBP from carp, blue shark, and
ellowtail showed a lower apparent molecular mass;16 kDa) and low abundance in the plasma (Shidojind Muto, 1977).Retinol is the major retinoid in chicken oocytic yolk
Joshi et al., 1973) but retinal is the predominant formf vitamin A in eggs of reptiles, amphibians, andarine teleosts (Plack and Pritchard, 1968; Branchaud
t al., 1995). The concentration of retinal in ovaries ofod increases during the period of yolk deposition,

corresponding to a sixfold increase in blood plasma,
cvr
liver, and due to the association of TTR with RBP in
rR
(
Trout Retinol-Binding Protein cDNA 53
and falls to low or zero levels in the blood and ovaries,respectively, after spawning. Injection of cod males orimmature females with 17b-estradiol increased theplasma levels of retinal to concentrations found nor-mally in maturing females, but had no effect on con-centrations of retinyl esters or retinol in the plasma(Plack and Pritchard, 1968). Unfertilized zebrafisheggs contain more retinal than retinol (2 mM t-retinalompared with 100 nM t-retinol) and retinal is con-erted to retinoic acid in embryos, but in the adults,etinol is the prevalent retinoid (Costaridis et al., 1996;
Maden and Pizzey, 1997). Large variations in theastaxanthin content were found in farmed salmoneggs, but no correlations between its quantities in eggsand the percentage of fertilization or the survival ofembryos or posthatch larvae were found (Torrissen,1984; Torrissen and Christiansen, 1995; Christiansenand Torrissen, 1997).
Studies on the transport and uptake of retinoids infish eggs have been initiated. The present studies fo-cus on the molecular structure and characterization ofa piscine RBP cDNA and follow the expression ofrtRBP mRNA in trout tissues and its possible regula-tion by 17b-estradiol.
MATERIAL AND METHODS
Animals
Rainbow trout (O. mykiss) were cultured at DanFarms (Kibbutz Dan, Israel). Treatments of fish with17b-estradiol were performed at the culture facilities.
Extraction of Cellular RNA
Total RNA was extracted from rainbow trout liver,spleen, kidney, ovaries, brain, and blood cells withTrizol reagent (GibcoBRL, Life Technologies) accord-ing to the manufacturer’s protocol. Several tissueswere investigated to determine whether RBP mRNA isexpressed in these tissues in addition to its expectedexpression in the liver. Expression of RBP mRNA wasreported in the rat kidney (Whitman et al., 1990). Inseveral vertebrate species, TTR mRNA is expressed inthe choroid plexus in addition to its expression in the
higher vertebrates, it seemed appropriate to investi-gate whether RBP mRNA is expressed in the rainbowtrout brain. Blood cells were chosen as putative neg-ative controls. Extraction of mRNA from rainbowtrout liver was performed with the Quick Prep MicromRNA purification kit according to the manufac-turer’s instructions (Amersham-Pharmacia-Biotech).Total RNA and mRNA concentrations were deter-mined by optical density at 260 nm and the purity wasvalidated from the ratio 260/280 nm. The integrity ofthe RNA was verified by ethidium bromide staining of18S and 28S ribosomal RNA bands in agarose gels.
First-Strand cDNA Synthesis and RT-PCR
Total RNA (5 mg) from rainbow trout liver wasreversed transcribed into a single-stranded cDNAwith Oligo(dt)12-18 and 200 units of transcript II (Gib-coBRL, Life Technologies) reverse transcriptase, ac-cording to the manufacturer’s instructions. The reac-tion mixture was incubated at 42° for 50 min and thenheated at 70° for 15 min. PCR amplification was car-ried out with 5 ml of the first-strand cDNA in 50 ml ofeaction mixture with two degenerate primers, (1)BP-5: 59-AAY ATH CAR GTN ATG CAR AAY-39
and (2) RBP-6: 59-CAY AAR AAR AAR GAR ATHTGY TTY-39, where Y 5 C/T, H 5 T/C/A, R 5 G/A,and N 5 T/C/A/G. The design of the primers wasbased on the trout RBP amino acid sequence derivedby Edman degradation (Berni et al., 1992; Zapponi etal., 1992). The thermal profile for RT-PCR consisted ofinitial denaturation at 94° for 2 min followed by 40cycles of denaturation (94° for 30 s), annealing (48° for1 min), and extension (68° for 2 min). Amplified prod-ucts were separated by 1.2% agarose gel electrophore-sis, and the resulting single-band product of 462 bp(RBP-a) was purified by QIAquick PCR purificationkit (Qiagen, USA) and subcloned into pGEM T-easy(Promega, USA) according to Sambrook et al. (1989).Several clones were isolated and analyzed.
3*- and 5*-Rapid Amplification of cDNA Ends (3*-and 5*-RACE) of Trout RBP
The full coding sequence of rainbow trout cDNAwas cloned with both 39- and 59-RACE proceduresFrohman et al., 1988) in combination with the stan-
Copyright © 2001 by Academic PressAll rights of reproduction in any form reserved.

dard RT-PCR procedure. For 39-RACE (RBP-b), first-
staGtp(Te
USA). RNA was prehybridized for 3 h in 50% form-a
54 Sammar et al.
strand cDNA was primed with an Oligo(dT) adaptorprimer, 59-GAC TCG AGT CGA CAT CGATTTTTTTTTTTTTTTTT-39, with AMV reverse tran-criptase (Promega, USA) followed by amplification ofhe first-strand cDNA with RBP-5 primer (forward) asspecific gene primer and a universal adaptor primerAC TCG AGT CGA CATCGA. After an initial dena-
uration cycle of 94° for 3 min, 50 PCR cycles wereerformed, each consisting of 1 min denaturation
94°), 1 min annealing (58°), and 2 min extension (68°).o ensure complete extension in the last cycle, thextension was increased to 7 min. For 59-RACE (RBP-
c), first-strand cDNA was tailed with terminal deoxy-nucleotidyl transferase (gibco/BRL) with 1 mM dCTPfollowing the specification of the manufacturer(gibco/BRL). Five microliters of the tailing reactionwas used as a template for 59-RACE PCR (RBP-c) withRBP-6 as a gene-specific primer in combination withthe Oligo(dG) adaptor primer: 59-AAG CTT GAA TTCGGA TCC C (GGGGGGGGGGGGGGGGG)-39. ThePCR thermal conditions were identical to the 39-RACEconditions except for the annealing temperature (48°).The resulting PCR products were separated on 1.2%agarose gel, purified, and subcloned into pGEM-TEasy vector (Promega), and clones containing insertswere isolated and sequenced with the BigDye Termi-nator Cycle Sequencing Kit from ABI in Applied Bio-systems DNA Sequencers at the Weizmann InstituteDNA Sequencing Unit (Rehovot, Israel).
Sequence Analysis
Sequence alignment and comparisons were per-formed with programs of GeneRunner (Version 3.0;Hastings Software Inc., 1994) software. Data basesearches and multiple local alignments were per-formed with the BLASTP suite of programs (http://www.ncbi.nlm.nih.gov/BLAST). Sequence signal pre-diction was performed according to Nielsen et al.(1997) (http://genome.cbs.dtu.dk/services/SignalP).
Northern Blot Analysis
Total RNA (15 mg) was extracted from rainbowtrout tissues, separated on 1% formaldehyde agarosegel, transferred to Hybond membrane (Amersham),and cross-linked with UV Stratalinker (Stratagene,
Copyright © 2001 by Academic PressAll rights of reproduction in any form reserved.
mide, 53 SSPE, 53 Denhard’s, 0.1% SDS, and 100mg/ml of calf thymus DNA. For detection of RBPtranscripts, a 462-bp RBP-a cDNA fragment was la-beled with [32P]adCTP by use of a random priming kit(New England Biolabs). Hybridization was performedovernight at 42° and the membranes were washedtwice with 23 SSPE, 2% SDS, followed by a final washat 55° in 0.1% SDS, 13 SSPE for 1 h, and exposed toX-ray film (Biomax, USA). After autoradiography andstripping of the probe, the membrane was reprobedunder the same conditions with [32P]adCTP-labeled18S rDNA (by random primers with a NEBlot kit;New England Biolabs) to standardize for loading. Sim-ilarly, Northern blot analysis was performed withmRNAs extracted from the liver of 17b-estradiol-treated and control animals. Detection of vitellogenin(VTG) transcripts was performed with a rainbow trout[32P]adCTP-labeled VTG probe [exons 5–10; Dr. J.Wolff, Universite de Bretagne Sud, Vannes, France(Mouchel et al., 1996)].
Treatment of Rainbow Trout with 17b-Estradiol
Five juvenile trout males weighing between 80 and120 g were injected twice (on day 0 and day 7 of theexperiment) intramuscularly with 4–5 mg of 17b-es-tradiol (Sigma, USA) dissolved in corn oil in eachinjection. Controls were injected with the carrier oil.On day 14, fish were anesthetized and blood wascollected in heparinized tubes containing 0.1 mMPefa-Block (Boehringer, Germany) to prevent proteol-ysis. The blood was centrifuged (1500g for 15 min at4°) and the plasma was collected. Livers, spleen, kid-ney, ovaries, and brain were collected and frozen im-mediately in liquid nitrogen, and plasma and tissueswere stored at 270°.
SDS–PAGE and Western Blot Analyses
Samples were subjected to polyacrylamide gel elec-trophoresis (PAGE) according to the procedure de-scribed by Laemmli (1970) with a mini PROTEAN IIapparatus (Bio-Rad, USA). Trout plasma (1-ml) sam-ples were diluted 310 in sample buffer and boiled for3 min before being loaded on gels. The protein profilewas visualized with Coomassie blue staining. ForWestern blot analyses, proteins were transferred to

nitrocellulose membranes (Bio-Rad). Nonspecific
waciat
o(bwa(tt
(8.2%), tyrosine (8.1%), and lysine (7.2%) and a lower
Trout Retinol-Binding Protein cDNA 55
binding sites were blocked with 5% nonfat milk in TBS(20 mM Tris–HCl, 150 mM NaCl, pH 7.5). For detec-tion of VTG, the membrane was incubated with VTGantiserum (obtained from Professor T. Braunbeck,University of Heidelberg, Germany) (Bierberstein etal., 1999), diluted 1:50,000 for 2 h at room temperature,
ashed three times with TBS containing 0.05% Tween,nd then incubated with horse-raddish peroxidaseonjugated to goat anti-rabbit IgG in TBS. After 1 h ofncubation, the membrane was washed as describedbove and the signal was visualized by an ECL detec-ion system (Amersham-Pharmacia-Biotech).
RESULTS
RBP cDNA Cloning and Sequence Analysis
A cDNA fragment of 462 bp (RBP-a) was amplified inthe first RT-PCR experiments from trout liver cDNAwith two degenerate primers, RBP-5 and RBP-6, as for-ward and reverse primers, respectively. This fragmentwas cloned and its deduced amino acid sequence re-vealed that it is homologous to an internal region of theRBP cDNA of other animal species. A second PCR prod-uct was generated by 59-RACE with the combination ofthe RBP-6 specific primer and an oligo(dG) adapter. Thisproduct of 620 bp (RBP-b) spans from the 59 end to
verlap with the known sequence of the internal regionRBP-a). To complete the full coding sequence of rain-ow trout RBP (rtRBP) cDNA, a 39-RACE PCR productas amplified by the RBP-5 primer and an oligo(dT)
daptor primer, yielding a cDNA fragment of 830 bpRBP-c). A 954-nucleotide rtRBP cDNA was then ob-ained by the aligning of the PCR product sequences ofhe internal and the 59- and 39-RACE clones (Fig. 1). The
59 untranslated region was 93 bp long based on thededuced first methionine residue. A 16-amino-acid (aa)-long signal peptide could be predicted by use of a neuralnetwork method resulting in a rtRBP mature protein of176 aa with an estimated molecular mass of 20,267 Da.The 282-bp 39 untranslated end region contained onepolyadenylation consensus signal (AATAAA) found 20nucleotides upstream from the poly(A) tail (Fig. 1). Thededuced aa composition of rtRBP revealed a high levelof aspartic acid (11.2%), arginine (9.5%), phenylalanine
abundance of proline (2.8%) and histidine (3.8%).
Sequence Comparison
Comparison of the rtRBP deduced amino acid se-quence to those of RBP I and RBP II determined byEdman degradation after purification of the proteinfrom the plasma (Berni et al., 1992; Zapponi et al., 1992)revealed that rtRBP differed by two aa residues fromRBP I and by eight aa residues from RBP II (Fig. 2).Comparisons with RBP sequences of zebrafish, Xeno-pus, and chicken revealed 66, 56, and 64% identity,respectively, whereas 60% identity was found with thesequences of mouse, rat, and bovine and 62% identitywas found with human RBP sequences (Fig. 2).
Tissue Distribution of rtRBP mRNA
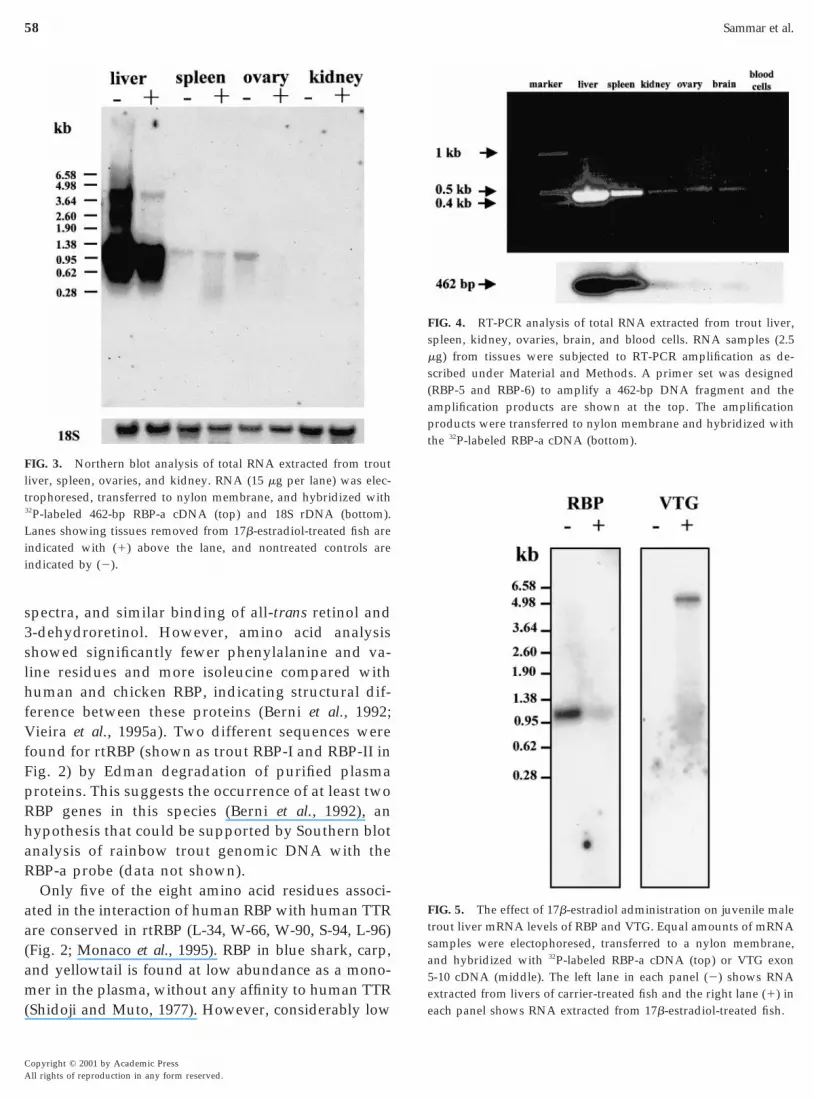
To characterize the tissue-specific expression of thertRBP mRNA, the RBP-a (462 bp) cDNA clone wasused as a probe in Northern blot analysis. As shown inFig. 3, a ;1.1-kb transcript of RBP mRNA was pre-dominantly expressed in the liver. A weak mRNAsignal was also found in the spleen, ovary, and kid-ney. These results were further extended and con-firmed by RT-PCR, showing that a 462-bp product wasamplified from the total RNA of all examined tissues,including liver, spleen, kidney, ovary, and brain, andwas absent from blood cells (Fig. 4). The identities ofthese bands were verified by probing with RBP-a inSouthern blot analysis (Fig. 4 bottom).
Regulation by Estrogen
The effect of 17b-estradiol on the expression ofrtRBP was examined by Northern blot analysis of totalRNA extracted from different tissues. As depicted inFig. 3, rtRBP transcript of ;1.1 kb was detected in thelivers of both the controls and the 17b-estradiol-treated individuals. No clear effect of 17b-estradioltreatment was observed in spleen, ovary, or kidney(Fig. 3), due to the low expression of rtRBP in thesetissues. The effect of 17b-estradiol treatment on theexpression of the rtRBP gene in the liver was betterresolved in another experiment, in which the 17b-estradiol treatment led to a decrease of the RBPmRNA signal relative to that of the control (Fig. 5).
Copyright © 2001 by Academic PressAll rights of reproduction in any form reserved.

56 Sammar et al.
The efficacy of the 17b-estradiol administration wasverified by the dramatic increase in VTG gene expres-sion in the liver and the appearance of VTG in theplasma, as revealed by Western blot analysis (Fig. 6).
DISCUSSION
Retinoids function as important regulatory signal-ing molecules for cell growth and differentiation dur-
FIG. 1. Nucleotide and deduced amino acid sequence of rainboware numbered on the right. The signal sequence as determined accoris indicated by boldface letters. The nucleotide sequence of the two rin PCR experiments are underlined. The potential polyadenylation
Copyright © 2001 by Academic PressAll rights of reproduction in any form reserved.
ing embryonic development and therefore must beavailable in the yolk stored in oviparous eggs for thefuture development of embryos. Their mode of trans-port and storage in piscine eggs has not been studiedin detail so far.
The molecular properties of rtRBP have been exam-ined to elucidate its role as a transporting protein intopiscine eggs. The present study revealed that the de-duced amino acid sequence of rtRBP (Fig. 1) showshigh homology with RBPs of zebrafish and other ver-tebrates (Fig. 2), including Xenopus, chicken, mouse,
etinol-binding protein cDNA. Nucleotide and amino acid residuesNielsen et al. (1997) (http://genome.cbs.dtu.dk/services/SignalP)
corresponding to primers RBP-5 (sense) and RBP-6 (antisense) usedsequence is indicated in italics and underlined.
trout rding toegionssignal

mei
RaRFQRsops
Trout Retinol-Binding Protein cDNA 57
rat, horse, bovine, and human. Moreover, except forthree amino acids, the rtRBP deduced amino acidsequence is identical with the trout RBP I amino acidsequence (Fig. 2), determined by Edman degradationof the protein purified from the blood (Berni et al.,1992). The estimated molecular mass of trout RBP is20,267 Da and is in accordance with the ;21-kDa
olecular mass determined for plasma protein (Bernit al., 1992). The positions of the six cysteines, involvedn the formation of the disulfide linkages in human
FIG. 2. Comparison of the deduced amino acid sequence of rainboXenopus, chicken, mouse, rat, horse, bovine, and human. The plasmthe following Accession Nos.: AJ236884 for zebrafish, sp (Swiss-PrP04916 for rat, sp P02758 for horse, sp P18902 for bovine, and sp P0amino acid residue is the same as the rtRBP deduced from its cdegradation of purified plasma proteins (Berni et al., 1992). Aminohuman RBP (Newcomb et al., 1984) are indicated by an asterisk on
BP, are conserved in rtRBP. Moreover, all aminocids identified by X-ray crystallography of humanBP as critical to the retinol binding pocket (L-34,-35, A-42, F-44, A-56, V-60, M-72, G-74, M-87, L-96,-97, H-103, Y-132, and F-134) are conserved in troutBP (Fig. 2; Newcomer et al., 1984). Piscine RBP isimilar to human and chicken RBPs in several ways:ne binding site for retinol, presence of a high trypto-han content, almost identical ultraviolet absorptionpectra and fluorescence excitation and emission
t retinol-binding protein (rtRBP) with RBP sequences of zebrafish,sequences were obtained from http://www.ncbi.nlm.nih.gov with172 for Xenopus, sp P41263 for chicken, sp Q00724 for mouse, sp
or human RBP sequences. A dash indicates that the correspondingTrout RBP-I and RBP-II sequences were determined by Edman
orresponding to those known to form the retinol-binding pocket inp of the relevant amino acid.
w troua RBPot) P062753 fDNA.
acids cthe to
Copyright © 2001 by Academic PressAll rights of reproduction in any form reserved.
V
haR
aa(am(
5ee
l
L
58 Sammar et al.
spectra, and similar binding of all-trans retinol and3-dehydroretinol. However, amino acid analysisshowed significantly fewer phenylalanine and va-line residues and more isoleucine compared withhuman and chicken RBP, indicating structural dif-ference between these proteins (Berni et al., 1992;
ieira et al., 1995a). Two different sequences werefound for rtRBP (shown as trout RBP-I and RBP-II inFig. 2) by Edman degradation of purified plasmaproteins. This suggests the occurrence of at least twoRBP genes in this species (Berni et al., 1992), an
ypothesis that could be supported by Southern blotnalysis of rainbow trout genomic DNA with theBP-a probe (data not shown).Only five of the eight amino acid residues associ-
ted in the interaction of human RBP with human TTRre conserved in rtRBP (L-34, W-66, W-90, S-94, L-96)Fig. 2; Monaco et al., 1995). RBP in blue shark, carp,nd yellowtail is found at low abundance as a mono-er in the plasma, without any affinity to human TTR
Shidoji and Muto, 1977). However, considerably low
FIG. 3. Northern blot analysis of total RNA extracted from troutiver, spleen, ovaries, and kidney. RNA (15 mg per lane) was elec-
trophoresed, transferred to nylon membrane, and hybridized with32P-labeled 462-bp RBP-a cDNA (top) and 18S rDNA (bottom).
anes showing tissues removed from 17b-estradiol-treated fish areindicated with (1) above the lane, and nontreated controls areindicated by (2).
Copyright © 2001 by Academic PressAll rights of reproduction in any form reserved.
FIG. 5. The effect of 17b-estradiol administration on juvenile maletrout liver mRNA levels of RBP and VTG. Equal amounts of mRNAsamples were electophoresed, transferred to a nylon membrane,and hybridized with 32P-labeled RBP-a cDNA (top) or VTG exon-10 cDNA (middle). The left lane in each panel (2) shows RNAxtracted from livers of carrier-treated fish and the right lane (1) inach panel shows RNA extracted from 17b-estradiol-treated fish.
FIG. 4. RT-PCR analysis of total RNA extracted from trout liver,spleen, kidney, ovaries, brain, and blood cells. RNA samples (2.5mg) from tissues were subjected to RT-PCR amplification as de-scribed under Material and Methods. A primer set was designed(RBP-5 and RBP-6) to amplify a 462-bp DNA fragment and theamplification products are shown at the top. The amplificationproducts were transferred to nylon membrane and hybridized withthe 32P-labeled RBP-a cDNA (bottom).

mdbatldlantS
akteom
1997). Results here show that there is little, if any,e
nryttSttSyz1rat
omEsctiiccRtaitgwca
1asp
mt
Trout Retinol-Binding Protein cDNA 59
binding affinities were revealed between rainbowtrout RBP-I and RBP-II and human TTR and the twotrout proteins seem not to produce plasma protein–protein complexes (Berni et al., 1992). Although RBP in
ammals and chicken circulates with TTR, there is noirect evidence for this association in teleosts. It haseen suggested that the binding of RBP to TTR is notn absolute requirement for the transport of retinol toissues (Schreiber and Richardson, 1997). Moreover,ow levels of plasma RBP and plasma retinol in TTR-eficient rat mutants did not affect their retinol tissue
evels. The amount of urine RBP in mutants was lownd similar to wild-type rats, indicating that there is aeed to reexamine the assumption that binding of RBP
o TTR prevents excretion of RBP into the urine (seechreiber and Richardson, 1997).RBP transcripts were abundant in trout liver (Figs. 3
nd 4) and were detected at low levels in the spleen,idney, ovaries, and brain but not in blood cells. RBPranscripts were reported in the rat kidney (Whitmant al., 1990). It has been suggested that the expressionf TTR first evolved in the brain choroid plexus anduch later in the liver (Schreiber and Richardson,
FIG. 6. The effect of 17b-estradiol administration on VTG plasmalevels in juvenile male trout. Plasma samples were removed fromrainbow trout before (Time 0, lane 1) and after 17b-estradiol treat-
ent (lane 3). Fish treated with the carrier oil used for administra-ion of 17b-estradiol served as controls (lane 2). Plasma proteins
were subjected to SDS-PAGE (7.5%) and either stained with Coo-massie blue (Panel A) or electro-transferred to a nitrocellulose mem-brane (Panel B) and subjected to Western blot analysis using VTGantiserum. The VTG signal was visualised with ECL detection sys-tem and corresponding VTG position in the Coomassie blue stainedgel is indicated by an arrow (for more details see Material andMethods).
xpression of RBP in the adult trout brain.From its role in embryogenesis, it is clear that retinol
eeds to be available in the egg yolk. In the chicken,elatively large quantities of RBP were detected in eggolk, where RBP was considered to be the major, if nothe only, source of retinol in the oocyte, and retinol ishe major retinoid in yolk (Vieira and Schneider, 1993).erum albumin–retinoic acid complexes or lipopro-ein-bound carotenoids do not contribute significantlyo the yolk retinoid pool (Joshi et al., 1973; Vieira andchneider, 1993). Similar studies are not available, aset, for fish in which retinal has been reported inebrafish as the main egg retinoid (Costaridis et al.,996). It remains, however, to be determined howetinoids reach the oocytes during oocyte enlargementnd whether it involves RBP, serum albumin, or caro-enoids transported by LDL or VTG.
To investigate the relationship between RBP andocyte vitellogenesis in fish, we compared the hor-onal regulation of RBP and VTG gene expression.
strogen treatment caused a reduction in the expres-ion of RBP mRNA in rainbow trout liver relative toontrol (Fig. 5). In parallel, there was an induction inhe expression of VTG mRNA (Fig. 5) and a dramaticncrease in the plasma levels of VTG (Fig. 6), indicat-ng that the administered estrogen was physiologi-ally effective. Similar results were reported for thehicken, in which an estrogen-dependent decrease inBP gene transcription was also observed in estrogen-
reated roosters (Vieira et al., 1995a). However, thepparent lower level of RBP mRNA may result fromts lower relative abundance in the cellular mRNA dueo the massive increase in mRNA of estrogen-inducedenes such as VTG. Some support for this propositionas found in Northern blot analysis showing a de-
rease in the relative content of apolipoprotein A-1nd elongation factor 1a mRNAs in estrogen-treated
fish compared to controls (data not shown).Estrogen increased hepatic mRNA and serum pro-
tein levels of yolk precursor lipoproteins such as VTGand very-low-density lipoproteins in fish, amphibians,and chicken (Baker and Shapiro, 1977; Jost et al., 1978;Chan et al., 1980; Wallace, 1985; Mommsen and Walsh,988; Wallaert and Babin, 1992; Vieira et al., 1995a). Inmphibians, estrogen induced hepatic RBP gene tran-cription (McKearin et al., 1987; McKearin and Sha-iro, 1988). In the rat, estrogen treatment induces RBP
Copyright © 2001 by Academic PressAll rights of reproduction in any form reserved.

gene transcription in the kidney, but not in the liver
rtcshaTi
B
B
B
B
B
near Montreal and a reference site. Can. J. Fish Aquat. 52, 1703–
C
C
C
C
C
F
G
H
H
H
J
J
M
60 Sammar et al.
(Whitman et al., 1990).Vieira et al. (1995a) proposed two modes of estrogen
egulation of hepatically derived yolk precursor pro-eins. On the one hand, estrogen induces higher cir-ulating levels of lipoproteins, such as VTG, that con-titute a prominent mass of the oocyte. On the otherand, minor yolk proteins such as TTR, RBP, andpolipoprotein A-I respond by lower plasma levels.he explanation for this dichotomy in chicken and fish
s awaited.
ACKNOWLEDGMENTS
We dedicate this publication to Professor R. A. Wallace, in appre-ciation of his lifelong contribution to the study of vitellogenesis.Professor Wallace suggested the study of transport of vitamins toeggs during the sabbatical stay of one of us (E.L.) at the WhitneyLaboratory, University of Florida. We thank Dan Fish Farms (Kib-butz Dan, Israel) and Mr. A. Hurowitz for their support and supplyof trout. Our sincere thanks go to Professor T. Braunbeck (Univer-sity of Heidelberg, Germany) for trout VTG antiserum and to Dr. J.Wolff (Universite de Bretagne Sud, Vannes, France) for trout VTGinserts. This research was supported by the Ministry of Agriculture(Grant 894-0083-97 to E.L. and A.E.). Dr. M. Sammar was a recipientof a grant from the Israeli Ministry for Absorption, Center forAbsorption in Science.
REFERENCES
Baker, H. J., and Shapiro, D. J. (1977). Kinetics of estrogen inductionin Xenopus laevis vitellogenin mRNA as measured by hybridiza-tion to cDNA. J. Biol. Chem. 252, 8428–8434.
erni, R., Stoppini, M., and Zapponi, M. C. (1992). The piscineplasma retinol-binding protein: Purification, partial amino acidsequence and interaction with mammalian transthyretin. Eur.J. Biochem. 204, 99–106.
ierberstein, U., Berbner, T., Islinger, M., and Braunbeck, T. (1999).Immunohistochemical localization of vitellogenin in rainbowtrout (Oncorhynchus mykiss) hepatocytes using immunofluores-cence. Sci. Total Environ. 233, 67–75.
laner, W. S. (1989). Retinol-binding protein: The serum transportprotein for vitamin A. Endocr. Rev. 26, 1241–1251.
lomhoff, R., Green, M. H., Berg, T., and Norum, K. R. (1990).Transport and storage of vitamin A. Science 250, 399–404.
ranchaud, A., Gendron, A., Fortin, R., Anderson, P. D., and Spear,P. A. (1995). Vitamin A stores, teratogenesis, and EROD activity inwhite sucker, Catostomus commersoni, from Riviere des Prairies
Copyright © 2001 by Academic PressAll rights of reproduction in any form reserved.
1713.han, L., Bradley, W. A., Jackson., R. L., and Means, A. R. (1980).Lipoprotein synthesis in the cockerel liver: Effects of estrogen onhepatic polysomal messenger ribonucleic acid activities for themajor apoproteins in very low and high density lipoproteins andfor albumin and evidence for precursors to these secretory pro-teins. Endocrinology 106, 275–283.
hristiansen, R., and Torrissen, O. J. (1997). Effect of dietary astax-anthin supplementation on fertilization and egg survival in At-lantic salmon (Salmo salar L.). Aquaculture 153, 51–62.
onlon, R. A. (1995). Retinoic acid and pattern formation in verte-brates. Trends Genet. 11, 314–319.
ostaridis, P., Horton, C., Zeitlinger, J., Holder, N., and Maden, M.(1996). Endogeneous retinoids in zebrafish embryo and adult.Dev. Dyn. 205, 41–51.
urley, R. W., and Robage, M. J. (1997). Retinoid structure, chem-istry, and biological active derivatives. In “Advances in OrganBiology; Retinoids: Their Physiological Function and TherapeuticPotential” (E. E. Bittar, Series Ed. and G. V. Sherbet, Guest Ed.),Vol. 3, pp. 35–77. JAI Press, London.
rohman, M. A., Dush, M. K., and Martin, G. R. (1988). Rapidproduction of full-length cDNAs from rare transcripts: Amplifi-cation using single gene-specific oligonucleotide primers. Proc.Natl. Acad. Sci. USA 85, 8998–9002.udas, L. J. (1994). Retinoids and vertebrate development. J. Biol.Chem. 269, 15399–15402.ayashi, S., Ooshiro, Z., Tanaka, Y., and Okamoto, H. (1990). Reti-nol binding protein isolated from eel serum. Nippon Suisan Gak-kaishi 56, 641–649.eller, J. (1976). Purification and evidence for the identity of chickenplasma and egg yolk retinol-binding protein–prealbumin com-plex. Dev. Biol. 51, 1–9.ermann, K. (1995). Teratogenic effects of retinoic acid and relatedsubstances on the early development of the zebrafish (Brachydaniorerio) as assessed by a novel scoring system. Toxic. Vitro 9, 267–283.
oshi, P. S., Mathur, S. N., Murthy, S. K., and Ganguly, J. (1973).Vitamin A economy of the developing chick embryo and of thefreshly hatched chick. Biochem. J. 136, 757–761.
ost, J.-P., Ohno, T., Panyim, S., and Scheurch, A. R. (1978). Appear-ance of mRNA sequence and rate of vitellogenin synthesis inchicken liver following primary and secondary stimulation by17b-estradiol. Eur. J. Biochem. 84, 355–361.
Kopelman, M., Cogan, U., Mokady, S., and Shinitzky, M. (1976a).The interaction between retinol binding proteins and prealbu-mins studied by fluorescence polarization. Biochim. Biophys. Acta439, 449–460.
Kopelman, M., Mokady, S., and Cogan, U. (1976b). Comparativestudies of human and chicken retinol binding proteins and pre-albumins. Biochim. Biophys. Acta 439, 442–448.
Laemmli, U. K. (1970). Cleavage of structural protein during theassembly of the bacteriophage T4. Nature 227, 680–685.aden, M., and Pizzey, J. (1997). The role of retinoids in patterningfish, amphibian, and chick embryos. In “Advances in Organ Bi-ology; Retinoids: Their Physiological Function and Therapeutic

Potential” (E. E. Bittar, Series Ed. and G. V. Sherbet, Guest Ed.),
M
M
M
M
M
M
M
N
N
N
P
P
R
Function and Therapeutic Potential” (E. E. Bittar, Series Ed. and
S
S
S
T
T
T
V
V
V
W
W
Trout Retinol-Binding Protein cDNA 61
Vol. 3, pp. 93–139. JAI Press, London.McKearin, D. M., Barton, M. C., Keller, M. J., and Shapiro, D. J.
(1987). Estrogen induced transcription of the Xenopus laevis serumretinol-binding protein gene. J. Biol. Chem. 262, 4939–4942.cKearin, D. M., and Shapiro, D. J. (1988). Persistent estrogeninduction of hepatic Xenopus laevis serum retinol-binding proteinmRNA. J. Biol. Chem. 263, 3261–3265.eans, A. L., and Gudas, L. J. (1995). The roles of retinoids invertebrate development. Annu. Rev. Biochem. 64, 201–233.okady, S., and Tal, M. (1974). Isolation and partial characterizationof retinol binding protein from chicken plasma. Biochim. Biophys.Acta 336, 361–366.ommsen, T. P., and Walsh, P. J. (1988). Vitellogenesis and oocyteassembly. In “Fish Physiology” (W. S. Hoar and D. J. Randall,Eds.), Vol. XIA, pp. 347–406. Academic Press, New York.onaco, H. L., Rizzi, M., and Coda, A. (1995). Structure of a com-plex of two plasma proteins: Transthyretin and retinol-bindingprotein. Science 268, 1039–1041.oriss-Kay, G. (1997). Retinoids in mammalian embryonic develop-ment. In “Advances In Organ Biology; Retinoids: Their PhysiologicalFunction and Therapeutic Potential” (E. E. Bittar, Series Ed. and G. V.Sherbet, Guest Ed.), Vol. 3, pp. 79–92. JAI Press, London.ouchel, N., Trichet, V., Betz, A., Pennec, J.-P., and Wolff, J. (1996).Characterization of vitellogenin from rainbow trout (Oncorhyn-chus mykiss). Gene 174, 59–64.ewcomer, M. E., Jone, T. A., Aqvist, J., Sundelin, J., Eriksson, U.,Rask, L., and Peterson, P. A. (1984). The three dimensional struc-ture of retinol-binding protein. EMBO J. 3, 1451–1454.ielsen, H., Engelbrecht, J., Brunak, S., and von Heijne, G. (1997).Identification of prokaryotic and eukaryotic signal peptides andprediction of their cleavage sites. Protein Eng. 10, 1–6.oy, N., and Xu, Z.-J. (1990). Interactions of retinol with bindingproteins: Implications for the mechanism of uptake by cells. Bio-chemistry 29, 3878–3883.
ervais, S., and Brew, K. (1987). Homology and structure functioncorrelations between a 1-acid glycoprotein and serum retinol-binding protein and its relatives. FASEB J. 1, 209–214.
lack, P. A., and Pritchard, D. J. (1968). Effect of oestradiol 3-ben-zoate on the concentration of retinal and lipids in cod plasma.Biochem. J. 106, 257–262.
edfern, C. P. F. (1997). Molecular mechanisms of retinoid function.In “Advances In Organ Biology; Retinoids: Their Physiological
G. V. Sherbet, Guest Ed.), Vol. 3, pp. 35–77. JAI Press, London.ambrook, J., Fritsch, E. F., and Maniatis, T. (1989). “MolecularCloning: A Laboratory Manual.” Cold Spring Harbor LaboratoryPress, Cold Spring Harbor, NY.
chreiber, G., and Richardson, S. J. (1997). The evolution of geneexpression, structure and function of transthyretin. Comp. Bio-chem. Physiol. 116B, 137–160.
hidoji, Y., and Muto, Y. (1977). Vitamin A transport in plasma ofthe non-mammalian vertebrates: Isolation and partial character-ization of piscine retinol-binding protein. J. Lipid Res. 18, 679–691.
akeuchi, T., Dedi, J., Haga, Y., Seikal, T., and Watanabe, T. (1998).Effect of vitamin A compounds on bone deformity in larvalJapanese flounder (Paralichthys olivaceus). Aquaculture 169, 155–165.
orrissen, O. J. (1984). Pigmentation of salmonids—Effect of caro-tenoids in eggs and start-feeding diet on survival and growthrate. Aquaculture 43, 185–193.
orrissen, O. J., and Christiansen, R. (1995). Requirements for caro-tenoids in fish diets. J. Appl. Ichthyol. 11, 225–230.
ieira, A. V., and Schneider, W. J. (1993). Transport and uptake ofretinol during chicken oocyte growth. Biochim. Biophys. Acta 1169,250–256.
ieira, A. V., Kuchler, K., and Schneider, W. J. (1995a). Retinol inavian oogenesis: Molecular properties of the carrier protein. DNACell Biol. 14, 403–410.
ieira, A. V., Sanders, E. J., and Schneider, W. J. (1995b). Receptor-mediated endocytosis of serum transthyretin by chicken oocytes.J. Biol Chem. 270, 2952–2956.allace, R. A. (1985). Vitellogenesis and oocyte growth in non-mammalian vertebrates. In “Developmental Biology” (L. Brow-der, Ed.), pp. 127–177. Plenum, New York.allaert, C., and Babin, P. J. (1992). Effects of 17b-estradiol andstarvation on trout plasma lipoproteins. Lipids 27, 1032–1041.
Whitman, M. M., Harnish, D. G., Sprano, K. J., and Soprano, D. R.(1990). Retinol-binding protein mRNA induced by estrogen in thekidney but not in the liver. J. Lipid Res. 31, 1483–1490.
Zapponi, M. C., Zanotti, G., Stoppini, M., and Berni, R. (1992). Theprimary structure of piscine (Oncorhynchus mykiss) retinol-bind-ing protein and a comparison with the three-dimensional struc-ture of mammalian retinol-binding protein. Eur. J. Biochem. 210,937–943.
Copyright © 2001 by Academic PressAll rights of reproduction in any form reserved.