report - fortescue metals group
TRANSCRIPT

Report
Fortescue Metals Night Parrot Project Final Report Against the Approved Research Plan
Perth December 2018 CB-RP-EN-1087

Fortescue Metals Night Parrot Project
Final report against the approved research plan
12 December 2018

Page 2 of 27
Table of contents
1. Background ..................................................................................................................................... 3
2. Developing Detection Strategies for Night Parrots ......................................................................... 5
2.1. Introduction ............................................................................................................................. 5
2.2. Acoustics ................................................................................................................................. 5
2.2.1. Passive call detection ...................................................................................................... 5
2.2.2. Call detection using playback ......................................................................................... 9
2.3. Acoustic detection – conclusions .......................................................................................... 10
2.4. Camera trap detection ........................................................................................................... 11
3. Understanding Night Parrot habitat preferences and use .............................................................. 15
4. Increasing knowledge of Night Parrot distribution ....................................................................... 16
5. Understanding the threats to Night Parrots ................................................................................... 19
6. Human and social aspects ............................................................................................................. 20
7. Diet and drinking .......................................................................................................................... 21
8. Breeding biology ........................................................................................................................... 24
9. References ..................................................................................................................................... 25
10. Appendix 1 – Approved research plan ...................................................................................... 26
11. Appendix 2 – Approval letter .................................................................................................... 27
List of figures
Figure 1. Night Parrot calling patterns during the night, stratified by wet and dry periods .................... 7
Figure 2. The effect of rainfall on Night Parrot call rates ....................................................................... 8
Figure 3. The effect of distance on the detection on the Night Parrot dink-dink call ............................. 9
Figure 4. Cameras fail to detect at site with >3000 calls. ..................................................................... 13
Figure 5. Ad hoc Night Parrot detection on camera trap ...................................................................... 13
Figure 6. Location of the historical Night Parrot records and those surveyed during the research program
.............................................................................................................................................................. 18
List of tables
Table 1. Topics addressed during the research program ......................................................................... 4
Table 2. The nine Night Parrot calls recorded by the end of the research program (December 2016) ... 6
Table 3. Summary of strategies used to test the efficacy of camera traps for detecting Night Parrots . 12
Table 4. Summary of non-target species detected during camera trapping tests .................................. 12
Table 5. The ten historical Night Parrot record surveyed using acoustics ............................................ 17
Table 6. Night Parrot diet based on DNA sequencing results ............................................................... 22

Page 3 of 27
1. Background
The Night Parrot (Pezoporus occidentalis) is a small sized parrot (~100g; (Murphy et al. 2017b))
endemic to Australia’s arid zone. Once widespread and apparently common at least at some places,
the species seemed to decline sharply around the start of the 20th century (Olsen 2018) at a time
when many other arid zone native mammals were also disappearing. Night Parrots are now listed
as Endangered at the federal level (Environment Protection and Biodiversity Conservation Act
1999 (EPBC Act) declaration under s178, s181, and s183). The species are also listed on state
legislation:
NSW: Listed as Extinct (Biodiversity Conservation Act 2016)
NT: Listed as Critically Endangered (Territory Parks and Wildlife Conservation Act 2000)
QLD: Listed as Endangered (Nature Conservation Act 1992)
SA: Listed as Endangered (National Parks and Wildlife Act 1972)
WA: Listed as Critically Endangered (Wildlife Conservation Act 1950)
VIC: Listed as Regionally Extinct (Advisory List of Threatened Vertebrate Fauna in
Victoria)
This document presents the results of a multi-year intensive research program on Night Parrots,
mostly in Queensland. In November 2012, Fortescue Metals Group entered into an agreement with
the Australian Government to invest in Night Parrot research, associated with expansion of the
Cloudbreak iron ore mine, under EPBC Referral 2010/5696. Condition 10 states:
To compensate for any unavoidable impacts on habitat for the Night Parrot, a Night Parrot Research Plan must be prepared and submitted to the Minister for approval within 12 months of the date of this approval. The research activities much be funded to a value of no less than $1 million (GST Exclusive). The Night Parrot Research Plan must include, but not be limited to:
a) funding of a suitably qualified expert/s to (undertake) research which contributes to understanding the field ecology/biology of the Night Parrot. The research must investigate and develop a repeatable survey methodology that provides for the detection of the species
b) upon completion of the research identified by (condition 10 a) and no more than 4 years from the date of this approval, conduct or fund comprehensive targeted surveys for the Night Parrot in at least 3 sites in areas where confirmed sightings or specimens of Night Parrot have been recorded since 1 January 2000. The surveys must be informed by the outcomes of the research identified at (condition 10 a) and undertaken by a suitably qualified expert; and
c) indicative timeframes for funding, and completion of all research activities required under the Night Parrot Research Plan. The Night Parrot Research Plan must include timeframes for reporting the outcomes of the research.
The results of any research funded under the Night Parrot Research Plan must be provided to the department within 12 months of completion. During the implementation timeframe for the Night Parrot Research Plan, research results and annual progress of research activities must be reported annually in the Environmental Performance Report required at (condition 13). The approved Night Parrot Research Plan must be funded and implemented.
In line with Condition 10, a Night Parrot Research Plan (Appendix 1) was prepared and approved
by the Australian Government in January 2014 (Appendix 2). The research plan presented a
prioritised list of topics deemed to be potentially important for the long-term conservation and
understanding of Night Parrots. They were classified as “Critical”, “High” and “Nice to know”.
All critical research topics were included in the approved research plan, whereas the high and nice-
to-know topics were to be included only if the opportunity to investigate them arose. The topics
that were ultimately investigated during implementation of the research plan, and their priorities,
Page 4 of 27
are listed in Table 1. The remainder of this report outlines the results of each of these research
topics.
TABLE 1. TOPICS ADDRESSED DURING THE RESEARCH PROGRAM
Part Research topic Priority
1 Developing Detection Strategies for Night Parrots Critical
2 Understanding Night Parrot habitat preferences and use Critical
3 Increasing Knowledge of Night Parrot Distribution Critical
4 Understanding the threats to Night Parrots Critical
5 Human and social aspects High
6 Diet and drinking High
7 Breeding biology Nice to know
The approved research plan was implemented from January 2014 to December 2016, when
Fortescue’s Conditioned commitments were met. This report summarises all the findings of the
research completed under the plan.
Parts 2, 4, 5, 6 (in part) and 7 of the plan (see Table 1) have been published in peer-reviewed
scientific journals, and this report presents copies of these papers.
Results of three parts of the approved research plan about detection, diet and distribution (Parts 2,
3 and part of 6 – Table 1) are detailed in this report. These components have not yet been published
because they have been incorporated into an on-going research program, being led by Mr Nicholas
Leseberg as part of his PhD studies at the University of Queensland. This ongoing work is not part
of Fortescue funded research program required to meet Condition 10.
At the time the research plan was written, very little was known about the biology of Night Parrots.
Basic information about critical aspects such as preferred habitats and use, breeding biology,
movements, diet, social interactions and threatening processes were either entirely absent or based
on supposition and conjecture. Only a small amount of information was based on direct
observations of living birds that took place about 100 years ago (Andrews 1883; Wilson 1937).
Importantly, prior to 2013 there were no recordings of Night Parrot calls, which made detection
and verification of sightings extremely difficult. There were only written descriptions and
inferences based on Indigenous names (Olsen 2018). More detailed background about Night Parrot
ecology is presented in each of the sections that follow.
This report represents the final research report to meet the requirements of Condition 10 of
Fortescue’s EPBC Referral 2010/5696.

Page 5 of 27
2. Developing Detection Strategies for Night Parrots
2.1. Introduction
Developing detection strategies for Night Parrots was specified by the EPBC Condition as a topic
that must be investigated (the other parts refer only to “field ecology/biology”). As such, this topic
received a large amount of investment during development and implementation of the research
plan. As outlined in the plan, the focus of the detection work was to test readily available
technologies that could be used by anyone with a modicum of training. It was decided to focus on
testing acoustics and camera traps.
Camera trap testing has been completed and is reported here in full. In contrast, acoustic testing is
ongoing, reflecting an ever-increasing understanding of the repertoire of Night Parrots and ever-
increasing advancements in the automated detection of acoustic signals and associated hardware.
The on-going work is being led by Nick Leseberg as part of his PhD studies, in collaboration with
Steve Murphy.
2.2. Acoustics
At the time the Condition was developed by the Australian Government Environment Department
(then the Department of Sustainability, Environment, Water, Populations and Communities), there
were no recordings of Night Parrot calls but rather only written descriptions of calls and inferences
based on Aboriginal names for the species, which were assumed to be onomatopoeic. The great
breakthrough came sometime around 2013 when naturalist John Young discovered the first reliable
site for Night Parrots in about one hundred years and made the first recordings of their calls. Prior
to this, the only other somewhat reliably occupied sites were the Gawler Ranges (SA) in the late
1800s and the East Murchison (WA) in the 1920s/30s. Having a reliable site and some calls meant
that for the first time it was possible to conduct comprehensive and systematic research into
acoustic detection. The approved research plan divided the acoustic work into passive call
detection and call detection using playback.
2.2.1. Passive call detection
Passive call detection involves using automatic recording devices to record ambient sounds during
user-specified duty cycles. Broadly speaking, the settings used for the entire duration of the study
were to record all sounds all night using a high sampling rate (44.1 or 48 kHz) because we didn’t
know the full suite of calls, or when they might occur. The objectives of the study were to answer
whether or not automatic sound recorders and automatic computer processing software could
detect Night Parrots and at what level of reliability (i.e. what omission and commission error rates).
Song Meter models 2+ and 3 (Wildlife Acoustics Inc., MA, USA) were used exclusively
throughout the study as recording devices. During the official research project (January 2014 to
December 2016) we tested two computer programs for their ability to detect calls automatically:
Song Scope (Wildlife Acoustics) and Sound ID (Boucher et al. 2008). During preliminary testing
of Sound ID, it became clear that the software was under-developed in terms of its user interface
and ability to handle large quantities of data, and so that approach was dropped and is not
considered further in this report. Since December 2017, in collaboration with US-based firm
Conservation Metrics Inc. and UQ PhD candidate Nicholas Leseberg, we have also tested machine
learning approaches (Conservation Metrics) and packages in the computer coding environment
“R” (R Core Team 2016). These systems are still in development although preliminary results
indicate that both are outperforming Song Scope.
In this report we use results generated by Song Scope to answer the questions posed on the research
plan. In addition to the broad questions about the utility of hardware and software to simply detect
Night Parrots, the plan asked:

Page 6 of 27
What is the repertoire of Night Parrot calls?
What time of night do Night Parrots call?
Does calling change according to environmental factors, such as rainfall?
Over what distance can calls be detected?
Our results show convincingly that Night Parrot calls can be recorded on automated sound
recording devices. The equipment we used (Song Meter models SM2+ and SM3) was deployed
for extended periods (4-8 months, depending on settings) and withstood harsh-field conditions,
including extreme temperature fluctuations and rain deluges. Microphones needed annual (or
thereabouts) replacement and a small number of solar batteries also needed replacement. Otherwise
we did not experience any equipment failures during multi-year deployments of approximately 50
units.
Automated recognition is necessary to analyse the very large datasets that need to be collected to
detect Night Parrots. There are several programmes available but all suffer to varying degrees from
missing bona fide Night Parrot calls (false negatives or omission error) and incorrectly assigning
other sounds in the samples as Night Parrots (false positives or commission error). As noted above,
during the life of the research project we used Song Scope (Wildlife Acoustics) for detecting three
of the four main calls heard at the project area (dink-dink, ding-de-ding and dee-de-dee-de). The
other main call type that Song Scope was not capable of detecting without extreme levels of
commission error was the croak call.
The development of more effective call recognition (using machine learning and R packages) has
continued since the research project officially finished (as noted above). Preliminary results show
that the R detection models are outperforming both the machine learning approach and Song Scope
results: for three call types (dink-dink, ding-de-ding and dee-de-dee-de), R has roughly half the
omission error rate as the other two, and detects 86, 90, 69 and 41% of all four call types, noting
that 41% for the croak call compares to 6% for machine learning and 0% for Song Scope.
2.2.1.1. Night Parrot call repertoire
At the end of the official research period (December 2016) there were nine known Night Parrot
calls (Table 2). Since then, several other calls have been recorded, both at the Queensland site and
in Western Australia (Jackett et al. 2017).
TABLE 2. THE NINE NIGHT PARROT CALLS RECORDED BY THE END OF THE RESEARCH
PROGRAM (DECEMBER 2016)
Call Location Context Date first detected First recordist
dink-dink QLD contact/advertising/territorial pre-2013 John Young
croak QLD/WA contact/alarm pre-2013 John Young
dee-de-dee-de QLD contact/advertising/territorial pre-2013 John Young
distress QLD distress April 2014 Steve Murphy
ding-de-ding QLD contact/advertising/territorial April 2014 Steve Murphy
dee-de QLD contact/advertising/territorial April 2014 Steve Murphy
peel QLD breeding April 2015 Steve Murphy
chirrup QLD breeding April 2015 Steve Murphy
prat QLD breeding April 2015 Steve Murphy
Page 7 of 27
2.2.1.2. Nightly calling patterns
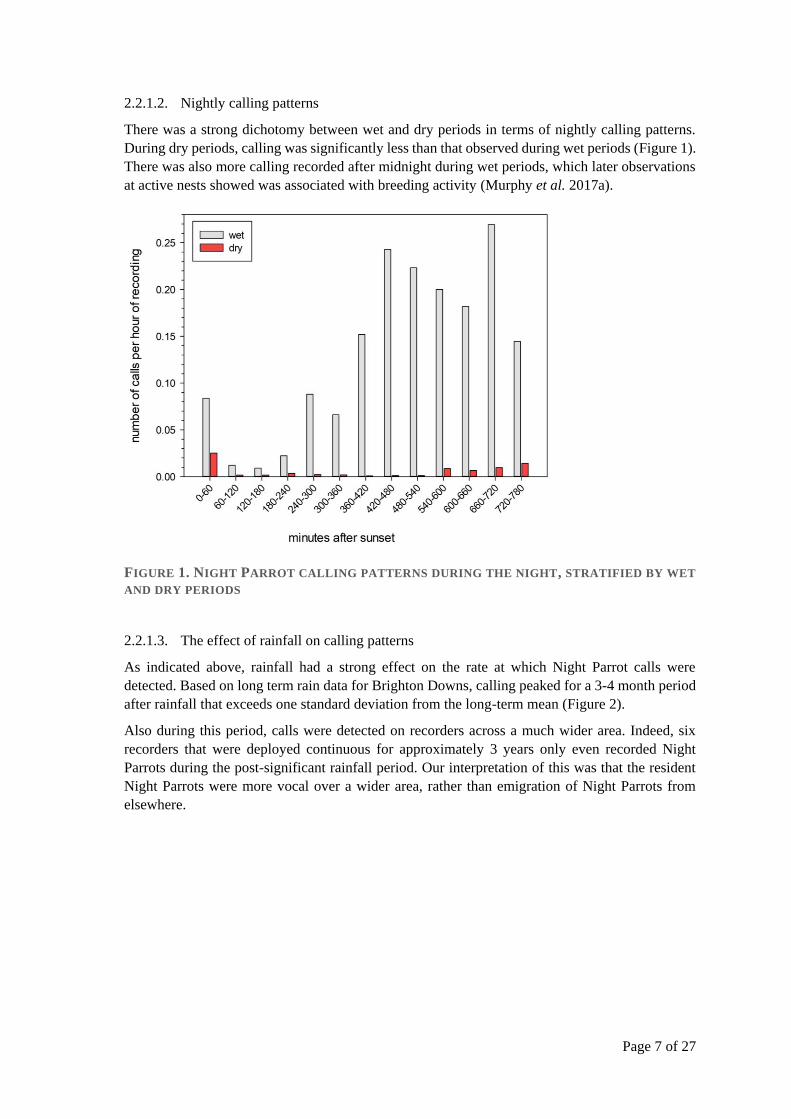
There was a strong dichotomy between wet and dry periods in terms of nightly calling patterns.
During dry periods, calling was significantly less than that observed during wet periods (Figure 1).
There was also more calling recorded after midnight during wet periods, which later observations
at active nests showed was associated with breeding activity (Murphy et al. 2017a).
FIGURE 1. NIGHT PARROT CALLING PATTERNS DURING THE NIGHT, STRATIFIED BY WET
AND DRY PERIODS
2.2.1.3. The effect of rainfall on calling patterns
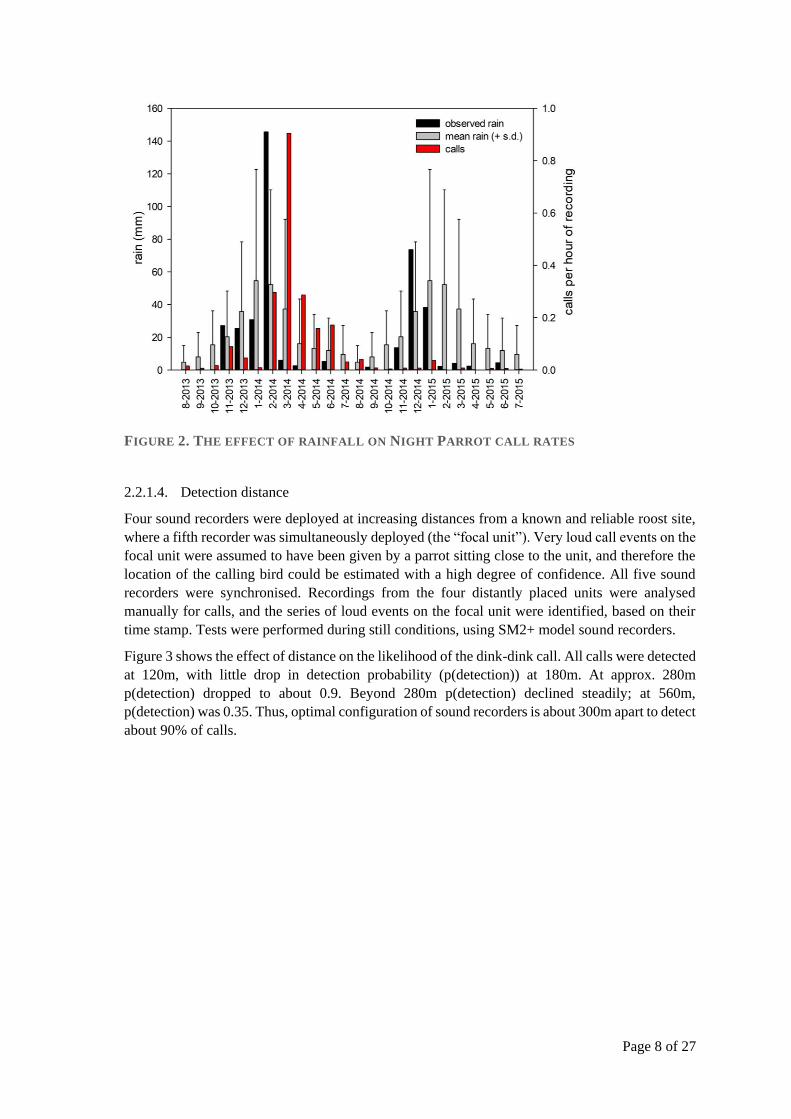
As indicated above, rainfall had a strong effect on the rate at which Night Parrot calls were
detected. Based on long term rain data for Brighton Downs, calling peaked for a 3-4 month period
after rainfall that exceeds one standard deviation from the long-term mean (Figure 2).
Also during this period, calls were detected on recorders across a much wider area. Indeed, six
recorders that were deployed continuous for approximately 3 years only even recorded Night
Parrots during the post-significant rainfall period. Our interpretation of this was that the resident
Night Parrots were more vocal over a wider area, rather than emigration of Night Parrots from
elsewhere.
Page 8 of 27
FIGURE 2. THE EFFECT OF RAINFALL ON NIGHT PARROT CALL RATES
2.2.1.4. Detection distance
Four sound recorders were deployed at increasing distances from a known and reliable roost site,
where a fifth recorder was simultaneously deployed (the “focal unit”). Very loud call events on the
focal unit were assumed to have been given by a parrot sitting close to the unit, and therefore the
location of the calling bird could be estimated with a high degree of confidence. All five sound
recorders were synchronised. Recordings from the four distantly placed units were analysed
manually for calls, and the series of loud events on the focal unit were identified, based on their
time stamp. Tests were performed during still conditions, using SM2+ model sound recorders.
Figure 3 shows the effect of distance on the likelihood of the dink-dink call. All calls were detected
at 120m, with little drop in detection probability (p(detection)) at 180m. At approx. 280m
p(detection) dropped to about 0.9. Beyond 280m p(detection) declined steadily; at 560m,
p(detection) was 0.35. Thus, optimal configuration of sound recorders is about 300m apart to detect
about 90% of calls.

Page 9 of 27
FIGURE 3. THE EFFECT OF DISTANCE ON THE DETECTION ON THE NIGHT PARROT DINK-
DINK CALL
2.2.2. Call detection using playback
To understand how Night Parrots respond to call-playback we collected information about the
response of our GPS-tagged bird upon broadcasting calls. While this is a small dataset, it
nonetheless provides unique information because during the tests we knew independently and with
certainty about the bird’s location and response even if it didn’t respond vocally and/or was not
seen.
This information was collected while we were attempting to re-capture the tagged bird for GPS
data download, which was our main priority. Consequently, we did not keep detailed records that
enable responses to be stratified by call type. However, we did keep a list of calls used, which
were: dink-dink, croak and dee-de-dee-de calls. We used them in an ‘interactive’ way with the
tagged bird, during 9 bouts of play-back during seven re-capture attempts (3 dawn, 4 dusk). The
following responses were recorded:
The tagged bird responded by both moving closer and calling on 6/9 occasions (67%)
It approached without calling once (11%); this was later in a netting attempt when
previously it had called in response.
It called without approaching once (11%).
Only on one occasion did it neither call nor approach after hearing playback. This was
one of the later capture attempts when it had already been exposed to many calls.
The most common response (by it and other non-tagged bird/s that came in on two
occasions) was the croak call and we had one or two episodes of many exchanges between
it and us lasting for up to 30 mins.
In conclusion, generally speaking, playback was very effective at getting the bird moving around
the mist-nets and can increase the likelihood of eliciting a response.
We also conducted playback tests in places where we did not have tagged birds. These were done
using the automated call broadcasting and recording function on SM3 units. Once our error

Page 10 of 27
detection rates for the automated call recognises improve (see Section 2.2.1) we will be in a
position to report on these tests.
2.3. Acoustic detection – conclusions
Our results show that Night Parrots can be detected and monitored using automatic sound
recorders, and that their calls can be detected automatically using software. Preliminary analyses
based on acoustic data from Queensland show that Night Parrot calling behaviour varies
significantly in relation to precedent rainfall. Specifically, when rainfall exceeds one standard
deviation from the long-term mean, calling rates increase, more calling occurs through the night
and calls are detected over a wider area away from the roosting area. During dry times, calling is
mostly limited to the core roosting areas and occurs mostly in the first hour after sunset and before
sunrise. It is clear, therefore, that rainfall affects the detectability of Night Parrots. During dry
times, when birds call less over a much smaller area, detection rates are low. Birds can typically
only be detected at the roost site, and sample sites must be selected that provide more or less
comprehensive spatial coverage for the survey area. In contrast, sampling during post-wet periods
can afford to be spatially separated because birds call over a wider area.

Page 11 of 27
2.4. Camera trap detection
The detailed knowledge of site occupancy by Night Parrots presented an opportunity to perform
robust tests of whether they could be detected using camera traps. To do so, we used HC600 units
(Reconyx, WI, USA) set to ‘rapid fire’ mode without delay, capturing five photographs per trigger.
At sites known to be used by Night Parrots for roosting (based on acoustics and field observation),
we positioned camera traps in three scenarios: (1) along trackways between hummocks, (2)
pointing skywards and (3) along drift-fences running through the Triodia patch. We also set a
camera pointing directly at a roost that we discovered during VHF-tracking in 2014. Deployments
also occurred at locations we suspected Night Parrots were using for feeding, which later GPS-
tracking confirmed were floristically similar to those that are actually used by parrots (“Gilgai on
ironstone” and “Stony Rises” in the habitat and movements paper – Section 3). Finally, we
deployed cameras at water points near known roosting areas that we suspected might be visited by
parrots and at one location where GPS-tracking confirmed was being visited for drinking. All
images, even non-target species, were catalogued.
No Night Parrots were photographed on any of the camera traps that were set specifically and
systematically for detection.
Table 3 shows camera trap deployment scenarios and effort. Of particular interest is the group of
cameras that were deployed (4158 trap nights) within a known roosting area at the same time that
over 3000 calls were detected on automatic recorders (Figure 4). Direct observation at this site
confirmed that Night Parrots began calling after dusk from within the Triodia patch being
surveyed, and so we were confident that the birds were roosting there rather than flying in from
elsewhere and calling without landing. In other words, we knew they were there but simply were
not detected on camera traps.
We did capture one Night Parrot on a camera trap that was deployed to inform capture as part of
the GPS-tracking work (Figure 5) although this is considered an ad hoc rather than systematic
deployment.
The non-target species detected during tests is shown in Table 4. Of particular interest are those
species that have similar habits to Night Parrots, especially in relation to being relatively small,
cryptic and ground dwelling, such as small mammals, quail, bushlarks and pipits. Also of interest
is that little-button quail were not detected on camera traps even though we observed them
occasionally in the patches where the cameras were deployed. Not shown in Table 4 is that some
detections involved relatively fast moving events (e.g. bee-eater and wedge-tailed eagle captured
on the wing) suggesting that the camera traps were capable of detecting Night Parrots flying.

Page 12 of 27
TABLE 3. SUMMARY OF STRATEGIES USED TO TEST THE EFFICACY OF CAMERA TRAPS
FOR DETECTING NIGHT PARROTS
TABLE 4. SUMMARY OF NON-TARGET SPECIES DETECTED DURING CAMERA TRAPPING
TESTS
Habitat Detection StrategyNo.
deploymentsTotal camera trap nights
Triodia roosting Set at gaps in 300mm high plastic drift fence
running through known roosting habitat
10 743
Triodia roosting Pointed upwards at edges of known roosting habitat
to detect flying Night Parrots
15 1450
Triodia roosting Set low along pathways running through known
roosting habitat
40 4566
Actual roost discovered during 2015
tracking
Pointed at opening of roost tunnel 1 108
Small waterhole in creekline near known
roosts
Pointed at edge of water 29 4284
Artificial stock "tank" where GPS-tagged
bird was recorded
Pointed at edge of water 1 162
Potential feeding habitat (Stony rises and
gilgai on ironstone)
Set low 39 1905
GRAND TOTALS 135 13218
MAMMALS n photos BIRDS n photos
euro 8230 spinifex pigeon 375
red kangaroo 1768 spinifexbird 163
cow 550 diamond dove 87
desert mouse 413 willie wagtail 82
macropod (unident.) 390 crested pigeon 68
cat 258 emu 62
dingo 254 wedgetailed eagle 26
rodent (unident.) 179 Australian magpie 20
small mammal (unident.) 138 spotted bowerbird 16
echidna 76 Australian raven 15
camel 59 masked woodswallow 15
dunnart (unident.) 27 Australian kestrel 11
hopping mouse (unident.) 17 Hall's Babbler 10
striped-faced dunnart 15 zebra finch 10
dasyurid 6 corvid 9
crested bellbird 9
REPTILE/AMPHIBIAN n photos hooded robin 9
Varanus panoptes 581 common bronzewing 8
Ctenotus sp. 81 budgerygah 6
Varanus giganteus 64 grey butcherbird 5
Tiliqua multifasciata 35 little eagle 5
skink (unident.) 29 Richard's pipit 5
Ctenotus pantherinus 27 singing bushlark 5
Varanus acanthurus 15 quail (unidentified) 2
Varanus sp. 7 unidentified bird 1
Cyclorana (unident.) 5 rainbow bee-eater 1
bearded dragon 5 spotted nightjar 1
Pogonona sp. 5
Varanus tristis 2 GRAND TOTAL 14262

Page 13 of 27
FIGURE 4. CAMERAS FAIL TO DETECT AT SITE WITH >3000 CALLS.
Yellow squares indicate camera traps that despite 4158 trap nights failed to detect Night Parrots,
even though more than 3000 Night Parrot calls were detected on Song Meters (red dots) at the
same time.
FIGURE 5. AD HOC NIGHT PARROT DETECTION ON CAMERA TRAP

Page 14 of 27
In conclusion in relation to camera traps, given the intensive camera trap effort at sites known to
be occupied and used by Night Parrots and our failure in detecting them using a variety of
strategies, it is clear that this survey technique is not a reliable way to detect the species. That we
were able to detect other cryptic species and fast moving events suggests that our lack of detection
was not related to how we used, configured or installed the cameras. It is noteworthy that other
cryptic species present at the sites that we surveyed were also not recorded.
Our conclusions that the survey technique is unreliable is not to say that Night Parrots cannot be
captured on camera traps, as our own ad hoc result shows. Further evidence of this came in 2017
from Paraku IPA in WA (Mills and Collins 2017), although we note that the image was only
confirmed as a Night Parrot once supplementary acoustic evidence was collected.

Page 15 of 27
3. Understanding Night Parrot habitat preferences and use
Murphy, S.A., Silcock, J., Murphy, R., Reid, J., and Austin, J.J. (2017). Movements and habitat
use of the night parrot Pezoporus occidentalis in south-western Queensland. Austral Ecology 42,
858-868.

Movements and habitat use of the night parrot Pezoporusoccidentalis in south-western Queensland
STEPHEN A. MURPHY,1* JENNIFER SILCOCK,2 RACHEL MURPHY,1
JULIAN REID3 AND JEREMY J. AUSTIN4
1Research Institute for the Environment and Livelihoods, Charles Darwin University, Casuarina,Northern Territory 0800 (Email: [email protected]), 2Threatened Species Recovery Hub, NationalEnvironmental Science Program, University of Queensland, St Lucia, Queensland, 3The Fenner Schoolof Environment and Society, Australian National University, Acton, Australian Capital Territory, and4Australian Centre for Ancient DNA, School of Biological Sciences, The University of Adelaide,Adelaide, South Australia, Australia
Abstract The nocturnal, cryptic and geographically remote nature of night parrots, combined with their appar-ent rapid decline, means that very little is known of their biology or ecology. The discovery of a resident popula-tion in south-western Queensland in 2013 provides the first opportunity to undertake detailed studies on thismost enigmatic of birds. We attached a radio tag to a bird for 20 days in April 2015 and a GPS tag to anotherbird for 5 days in May 2016 to study movement patterns and habitat use. Both birds displayed similar behaviourbut the GPS-tagging provided a much finer resolution of spatial data. They called at dusk from their diurnalroosts amongst spinifex hummocks and then flew to more floristically diverse habitats dominated by large-seededspecies to feed. We conducted floristic surveys to describe the feeding grounds of the GPS-tagged bird and makedietary inferences. This individual spent most of its time in highly diverse but ephemeral habitats, including sea-sonally inundated plains and depressions associated with the outer Diamantina floodplain and gilgais on iron-stone plains. Prolifically seeding ephemeral species, most notably the annual grass Uranthoecium truncatum,dominate these feeding grounds. This work suggests that the habitat mosaic containing roost sites in close prox-imity to feeding grounds with key seed-producing species is an important factor, rather than an association withspinifex or samphire alone. Further work is needed to examine movement patterns and habitat use in more typi-cal dry seasons and the impact of cattle grazing on night parrot feeding areas, particularly with regard to seedproduction. The information presented here is vital for both in situ conservation of the Pullen-Pullen-MtWindsor-Diamantina population and for setting future research and survey priorities.
Key words: arid zone, diet, endangered bird, granivory, roosting, telemetry.
INTRODUCTION
Night parrots (Pezoporus occidentalis) are cryptic, noc-turnal and endemic to Australia’s arid interior. Untilthe late 19th century they were widespread and rela-tively easily found at least at some locations. Forinstance, 14 of the 25 museum skins in existencecame from the Gawler Ranges in South Australiabetween 1871 and 1881 (Andrews 1883; Black2012). The last night parrot collected intentionallywas in Western Australia in 1912 (Wilson 1937).Then followed 78 years of unconfirmed reports span-ning all mainland states and the Northern Territory,until in 1990 a desiccated body was found by a road-side in western Queensland (Boles et al. 1994). In2006, another body was discovered by a national
parks ranger 200 km to the south-east of the 1990specimen (McDougall et al. 2009). In 2013, the firstphotographs of a living night parrot were capturedclose to the site of the 2006 specimen (Dooley 2013).Due to its apparent decline the species is listed asEndangered under the federal Environment Protectionand Biodiversity Conservation Act 1999, and includedon all mainland state and territory threatened speciesschedules (Australian Government 2016).Their cryptic nature, remote distribution and appar-
ently rapid decline mean that there is scant ecologicalinformation about night parrots. This includes infor-mation about habitat use and movements, which, froma conservation management perspective presents anespecially acute problem. Information which does existis based on incidental, anecdotal and/or unconfirmedencounters, or observations by early naturalists whowere both focused on obtaining specimens rather thancollecting ecological information and hampered by thelimited technologies available to them (Andrews 1883;
*Corresponding author.Present address: Adaptive NRM, Malanda, Queensland4885, Australia.Accepted for publication May 2017.
© 2017 Ecological Society of Australia doi:10.1111/aec.12508
Austral Ecology (2017) ��, ��–��
Howe & Tregellas 1914). Night parrots have beenassociated with habitats containing one or more spe-cies of the widespread grass genus Triodia, used forroosting, breeding and feeding (Andrews 1883; Whit-lock 1924; McGilp 1931; Wilson 1937). There havealso been numerous sightings and at least five speci-mens collected in or near habitats featuring species ofChenopodiaceae (Austin 1855; Andrews 1883;McGilp 1931; Powell 1970; Forshaw et al. 1976; Ellis1982; Boles et al. 1994; McDougall et al. 2009).There is even less information about the movements
of night parrots. Andrews (1883, p. 30) stated thatthey ‘come and go according to the nature of the sea-son’ and that if Triodia does not set seed due to dryconditions ‘. . . no birds are to be seen.’ This suggeststhat night parrots undertake large-scale, rainfall-drivenmovements, which is also reflected in comments fromAboriginal people who reported that night parrots dis-appear during dry spells (Cleland 1930; Wilson 1937).Parker (1980) suggested that night parrots may occupyhabitats dominated by Triodia spp. during wet periodsand move into chenopod habitats during dry periods(Parker 1980) but there is no evidence for this. Atsmaller scales or over shorter periods, there is also verylittle known about night parrot movements. Andrews(1883, p. 29) stated that he had ‘known them to fly adistance of four or five miles’ to visit water.The site in western Queensland where the first
photographs were taken in 2013 presented the firstopportunity to conduct systematic research on keyaspects of night parrot biology and ecology. Here wereport on the results from radio and GPS tracking oftwo individuals – the first night parrots trapped andreleased for scientific study – that were followed for20 and 5 days, respectively, to gain insights intohabitat use and movements. Sample sizes are neces-sarily limited due to sensitivities about invasiveresearch techniques on a population that is suspectedto be small. Our objectives were to (i) describe shortterm movements of night parrots; (ii) describe thehabitats used by night parrots in terms of landscapeposition, vegetation structure and composition; (iii)attempt to interpret the activities of the individual ineach habitat; (iv) describe the roosts made by nightparrots; (v) infer diet based on field measurementsand plant biology, and; (vi) suggest directions for fur-ther research. The findings will inform conservationmanagement of the known population, and assist inlocating additional populations.
METHODS
Study site
This study was undertaken on Pullen-Pullen Reserve andneighbouring Mount Windsor Station in south-western
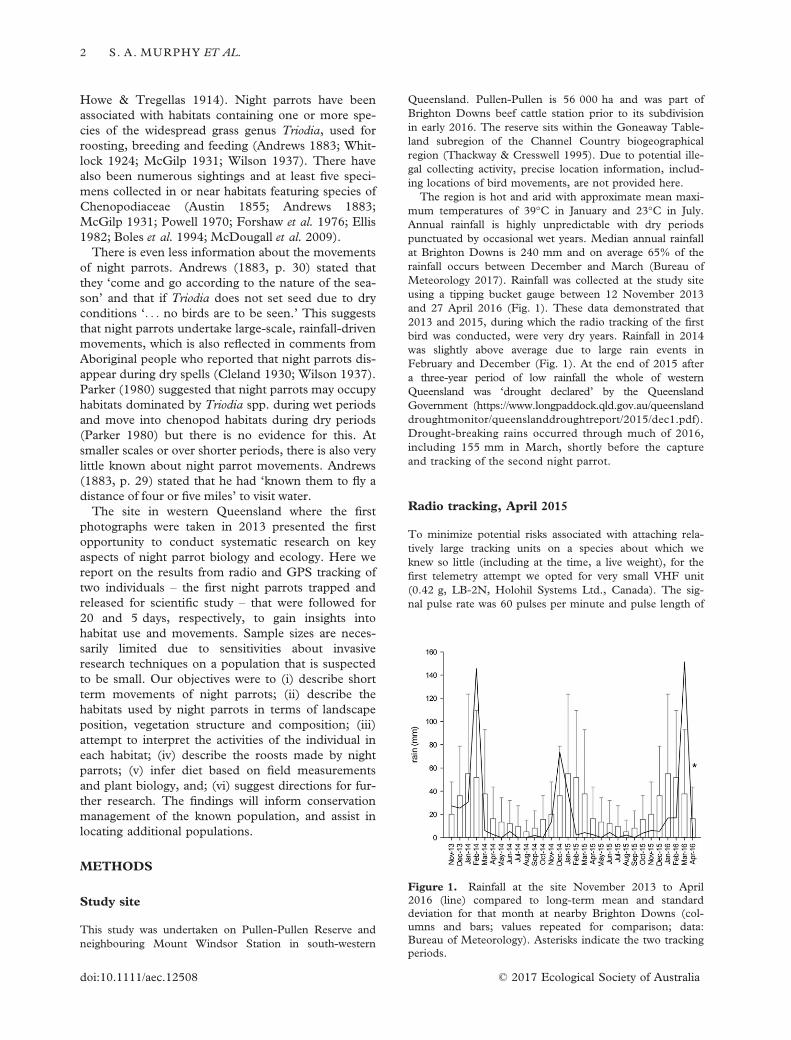
Queensland. Pullen-Pullen is 56 000 ha and was part ofBrighton Downs beef cattle station prior to its subdivisionin early 2016. The reserve sits within the Goneaway Table-land subregion of the Channel Country biogeographicalregion (Thackway & Cresswell 1995). Due to potential ille-gal collecting activity, precise location information, includ-ing locations of bird movements, are not provided here.
The region is hot and arid with approximate mean maxi-mum temperatures of 39°C in January and 23°C in July.Annual rainfall is highly unpredictable with dry periodspunctuated by occasional wet years. Median annual rainfallat Brighton Downs is 240 mm and on average 65% of therainfall occurs between December and March (Bureau ofMeteorology 2017). Rainfall was collected at the study siteusing a tipping bucket gauge between 12 November 2013and 27 April 2016 (Fig. 1). These data demonstrated that2013 and 2015, during which the radio tracking of the firstbird was conducted, were very dry years. Rainfall in 2014was slightly above average due to large rain events inFebruary and December (Fig. 1). At the end of 2015 aftera three-year period of low rainfall the whole of westernQueensland was ‘drought declared’ by the QueenslandGovernment (https://www.longpaddock.qld.gov.au/queenslanddroughtmonitor/queenslanddroughtreport/2015/dec1.pdf).Drought-breaking rains occurred through much of 2016,including 155 mm in March, shortly before the captureand tracking of the second night parrot.
Radio tracking, April 2015
To minimize potential risks associated with attaching rela-tively large tracking units on a species about which weknew so little (including at the time, a live weight), for thefirst telemetry attempt we opted for very small VHF unit(0.42 g, LB-2N, Holohil Systems Ltd., Canada). The sig-nal pulse rate was 60 pulses per minute and pulse length of
Figure 1. Rainfall at the site November 2013 to April2016 (line) compared to long-term mean and standarddeviation for that month at nearby Brighton Downs (col-umns and bars; values repeated for comparison; data:Bureau of Meteorology). Asterisks indicate the two trackingperiods.
doi:10.1111/aec.12508 © 2017 Ecological Society of Australia
2 S. A. MURPHY ET AL.

20 ms, giving a nominal operating life of 21 days. It wasattached to trimmed feathers on the back just above thehips using a cyanoacrylate glue so that it would be shedwithout harm to the bird when it next moulted, if notbefore. A single individual, weighing 104 g, was capturedwithin an 11 ha area of Triodia at 19.00 hours on 4 April2015, in a 38 mm 18 m polyester mist net, without the useof call-playback. In addition to fitting the transmitter, whilein the hand we recorded the bird’s weight using a 300 gspring balance and sampled a growing contour feather formolecular sex determination. The time from mist-netextraction to release was less than 20 min.
Tracking of the bird, using an Australis 26K receivingunit (Titley, Australia) and hand-held three-element anten-nae was achieved using a combination of ground (by foot,4 9 4 vehicle and 4 9 4 motorbike) and aerial searches(Robinson R-22 helicopter). Aerial searches were done atvarying heights from 100 to 800 m above ground, whichtesting using the same model transmitter showed gave us arange of approximately 2 km by holding the antennae out-side of the fuselage. The aerial search paths were a combi-nation of a systematic transects spaced approximately 1 kmapart covering all habitats and targeted examination of allTriodia patches. Locations where signals were detected werelogged using a handheld GPS unit and the signal bearingwas estimated using a compass. Precise locations of thebird could not be obtained. Sunset/sunrise during the radiotracking period ranged from 1831/0642 on 4 April to 1813/0650 on 24 April.
GPS tracking, May 2016
With lessons learned through the 2015 radio-tracking ses-sion, a larger GPS tracking device was used for a secondround of telemetry. A second night parrot weighing 102 gwas captured (6 km from the 2015 location) at 18.30 hourson 6 May, again without call-playback but this time in a60 mm 18 m nylon mist-nest. The tag incorporated a GPSunit (PinPoint 10) and VHF radio transmitter (PicoPipAg379) with 20 ms pulse length and 33 ppm pulse rate, fora nominal life of 34 days (Biotrack, U.K.). A small gauzepatch was also fitted to the underside of the tag to aidadherence to the bird, using the same attachment methodas described above. Total tag weight was 1.8 g. Handlingtime was approximately 15 min.
The tag was programmed to acquire GPS fixes in two‘series’ per night for five nights using the schedule outlinedin Appendix S1. The first GPS fixes were programmed tooccur the night after capture to allow the bird to settle. Forall or part of the series on three evenings, we increased thefix interval in the first hour to every 5 min in an attempt todetect drinking behaviour, because it was hypothesized thatnight parrots would be more likely to drink during thistime. Sunset/sunrise during the GPS-tracking period rangedfrom 1804/0656 on 7 May to 1802/0658 on 11 May.
At the time of sampling, there were no field-tested GPStags of suitable weight with remote data download function.To retrieve the tag the bird was recaptured using call-play-back in a mist-net on 23 May, 5.6 km from its original cap-ture point, where it was located using a helicopter to detectthe VHF signal in the manner described above.
Tag positional error was estimated by deploying the tag(prior to attachment) at three points over two nights usinga similar schedule to that described above. The mean andstandard deviation of distances between the position of thetag (as determined by handheld GPS) and the tag’s fixeswere then calculated, and these statistics helped wheninspecting and interpreting the locations where the individ-ual was later recorded, including in the design and place-ment of subsequent vegetation survey plots.
Several of the bird’s locations were represented by a sin-gle fix, and it was possible these may have been loggedwhile the bird was flying. We examined this using the pro-cedure outlined in Appendix S2. To calculate the timespent by the bird at each location, it was important to con-sider where the fixes at each location occurred in the pro-grammed series. If a location included the first and/or lastfixes in a series (‘end points’), we could only know the min-imum time the bird spent at that location based on thenumber of fixes and fix interval. If fixes were recorded at alocation with fixes from other locations either side in thesame series (‘nested points’), then it was possible to calcu-late the maximum time spent at that location. For locationswith single fixes, we applied the same logic; if it was anested point, the maximum time spent there was less thantwice the fix interval, whereas if it was an end point, theminimum time spent there was the time it took to get a fix,which we nominally annotate at 1+ min.
Descriptive statistics were calculated: (i) mean, maxi-mum and minimum distances moved within a series andbetween fixes; (ii) maximum, minimum and mean cumula-tive distances moved in a night, and; (iii) nightly and totalminimum convex polygons (MCPs). Cumulative measure-ments included movements to and from the roost (whichVHF tracking showed was stable during the GPS trackingperiod), and movements between series within a night.Given there were long periods each night when the GPStag did not acquire fixes, cumulative distances are mini-mum values. All spatial calculations were done using Arc-Map 10.0 (Environmental System Research Institute, USA).
Molecular sex determination
Full details of sexing methods are given in Appendix S3.
Vegetation surveys, June 2016
Between 12 and 17 June, we visited all fixes where theGPS-tagged night parrot had been recorded 1 month ear-lier. As discussed below, some sites were apparent feedinggrounds where the bird was recorded two or more times insuccession, whereas some sites involved repeated visits overextended periods. Others were apparently fly-overs or verybrief stops, with only single GPS fixes taken at some dis-tance from the preceding and following fixes. All sites werevisited, but more detailed surveys were done at sites wherethe bird had spent longer periods of time. Sites wereassigned to various ‘broad land types’, comprised of a land-form category combined with common vegetation structuralcharacteristics, and ‘habitats’, which we described floristi-cally.
© 2017 Ecological Society of Australia doi:10.1111/aec.12508
NIGHT PARROT MOVEMENTS AND HABITATS 3

Each site comprised between one and 35 GPS fixes.Where there was a single fix in a homogenous habitat, fourfloristic quadrats were placed within 30 m of the fix; thisdistance was based on estimates of GPS fix accuracy (seeResults). Where there were multiple fixes, a site encompass-ing all of them was marked using steel droppers and quadratselection was stratified by habitat type, with each type repre-sented by at least three quadrats. Quadrat selection within asite was biased towards GPS fixes. Where vegetation waspatchy (i.e. small vegetated areas on stony pavements),quadrats were placed within vegetated areas. Most sites wereseparated from each other by more than one kilometre, butfour clusters of fixes located within a square kilometre weretreated as discrete sites (labelled Nardoo, Lumpy, SucculentFlat and Mitchell Grass Flat – see Appendix S6) due tohabitat differences. The total number of quadrats at eachsite increased with the time spent there by the parrot andthe heterogeneity of the habitat.
Each 2 9 7 m quadrat was split into four sub-plots ofincreasing size. Plant species present in the first 0.3 9 2 msub-plot were assigned an abundance score of 4, new speciespresent in the next 0.7 9 2 m sub-plot an abundance of 3,new species in the next 2 9 2 m sub-plot an abundance of 2,and the final 2 9 4 m sub-plot an abundance of 1. Thismethod, involving unrepeated scoring of species presence,has been demonstrated to provide the best return (robustmeasure of rank abundance) for effort (no more time thanpresence-absence scores), thereby allowing for a relativelylarge quadrat size (Morrison et al. 1995). Voucher specimensof species have been lodged at the Queensland Herbarium.Nomenclature follows Bostock and Holland (2007). Thedata were analyzed using the R package (R Core Develop-ment Team 2016), and ordinated using non-metric multidi-mensional scaling package ‘vegan’ (Oksanen et al. 2017).
Lifeforms were assigned based on the above-ground partsof the plant, thus species with perennial rootstock butannual stems were classified as annual for this study. Short-lived species which can be biennial in higher-rainfall areas,or in consecutive high-rainfall years, were classified asannuals, as field observations, expert knowledge and exami-nation of herbarium specimen labels suggested they areshort-lived in the study area. Rain (10.6 mm) was recordedin the month between the last GPS fix and our flora sur-veys. Where field observations suggested that plants weremore or less abundant at the time of the parrot’s visit, thiswas noted. In particular, short-lived summer-growinggrasses were probably more common, but they were stilldetected in quadrats as seed heads and litter lying on theground.
RESULTS
Radio-tracking
We had difficulty locating the 2015 radio-taggedbird, which molecular sexing showed was a female.Apart from detecting it within Triodia 200 m fromthe capture point 5 h after release, ground-basedsearches failed to locate the bird for 3 days (until 7
April) when it was located 7 km to the north-east.We inspected this location on foot shortly after dis-covery, but the bird was inadvertently flushed at18.35 hours from a roost in an isolated Triodia hum-mock measuring 9.8 9 5.3 m across by 0.5 m high.The roost was a 25 cm long, horizontal tunnel, 8 cmin diameter (at entrance), constructed 8 cm aboveground, with the entrance facing inwards towards thebare centre of the ring-shaped hummock. The tunnelwas evidently constructed by a combination of chew-ing some leaves to length, and pushing and shapingothers (the majority). There was no chamber at theend. A camera-trap trained on the roost for 108 daysafter discovery showed that the bird did not returnafter being disturbed.Triodia in the vicinity of the new roost was sparse,
covering only 6.8% of a 25 ha sample area aroundthe roost (based on high resolution satellite imagery),the rest being an ironstone pavement dominated bysparse Sclerolaena longicuspsis (a habitat similar to thebroad land type ‘Stony Rise’, described below). Fol-lowing flushing, we did not detect a signal for12 days, despite approximately 1200 km of aerial and250 km of ground searching over 450 km2 of suitableroosting habitat, up to 50 km from the 7 April roost.The bird was relocated by air on 19 April only1.1 km away. Triodia cover within 25 ha around the19 April roost area was 1.9% and again, hummockswere embedded in a matrix of ironstone and sparseS. longicuspsis.The tag stayed attached to the bird for the life of
the battery. Between 19 and 24 April (when the bat-tery failed), we detected the tagged bird 16 timesfrom six locations, always in the early evening or justbefore sunrise. After a bout of calling involving one ortwo conspecifics, the tagged bird left the roost areaeach evening about 20–30 min after sunset andreturned to the roost area about 40–60 min beforesunrise. On most mornings, one or two calls wereheard as the bird settled. We failed to detect the birdoutside of these twilight periods despite approximately380 km of vehicle-based searching. The times, detec-tion positions (in relation to 19 April roost), bearings,signal strength (which is a proxy for distance betweenreceiver and tagged bird) and our interpretation ofthe bird’s activity are shown in Appendix S4. Despitethe small number of detections, we were able to con-clude that the female was moving considerable dis-tances (at least 5 km and possibly greater than10 km) away from her roost into habitats thatincluded floodplains and non-Triodia grasslands.
GPS-tracking
The mean positional error of the GPS tag was12.8 m (SD = 11.2 m; min. = 0.18 m; max. = 36.9 m;
doi:10.1111/aec.12508 © 2017 Ecological Society of Australia
4 S. A. MURPHY ET AL.

n = 48 fixes at three locations). Despite expectations,we were unable to determine from metadata if a fixwas taken on the wing or not (see Appendix S2).Based on molecular sexing, the GPS-tagged bird
was a male. The tag remained attached to the birduntil recapture on 23 May, when it fell off while thebird was being extracted from the mist net. The taglogged 127 fixes clustered into 18 sites. Excludingseries one on night one (the night after capture, whenthe bird was still settling from capture) and seriesnine and ten (which were less than 2 h), the meandistance moved within the remaining seven-two-hourseries was 8.9 km (SD = 5.3 km; max. = 17.62 km;min. = 1.3 km). The maximum distance betweenfixes within a series was 6.3 km between two loca-tions that were logged 10 min apart, which indicatesa minimum flight speed capability of ca. 38 km perhour.The mean minimum cumulative distance moved
per night was 29.9 km (SD = 9.6 km; max. =41.18 km; min. = 17.82 km; n = 4). The maximumstraight-line distance the bird was recorded away fromthe roost was 9.4 km at 04.10 hours on 12 May. Thenightly mean MCP area was 783 ha (SD = 605 ha;max. = 1821 ha; min. = 305 ha). The total MCParea for all points was 3344 ha.Four of the sites – Uranth Flat, Despair Flat,
Osteo and Stony Flat – appeared to be the favouredfeeding grounds during the tracking period, with aminimum of one hour spent at each (Appendix S6).The bird stayed for at least 20 min at a furtherthree sites (Mitchell Edge, Succulent Flat and Nar-doo, the latter being visited four times over threenights, including two possible fly-overs). The birdspent an extended period at the roost area early inthe night after capture and, given movement pat-terns over the subsequent four nights, we considerthis anomalous behaviour related to capture stress.Nine sites were single points and possible fly-overs.A single point was logged at an artificial water pointfor stock.
Broad land types and site-based descriptions
The roost site of the GPS-tagged night parrot waslocated in Triodia longiceps at the base of a low sand-stone range. Based on high resolution sub-1 m Digi-talGlobe (www.digitalglobe.com) satellite imagery,Triodia cover within a 25 ha area (centred on theroost) was approximately 10%, but this was inflateddue to the 25 ha area extending towards the plateaumargin where Triodia cover was more extensive. TheGPS fix at water was at the corner of an artificialstock ‘tank’ (earthen reservoir) that was approxi-mately 60 m wide. The remaining 16 sites encom-passed six broad land types, (with some sites
comprising more than one land type; Figure 2).Three sites were situated on a floodplain which isperiodically inundated by waters from the Dia-mantina River: an extensive plain dominated byephemeral species (Uranth Flat, in the broad landtype ‘floodplain’) and two small patches of Astreblagrassland (‘Mitchell grassland’) on the edge of thefloodplain, one of which graded into ephemeral herb-field (Mitchell Edge). One site was located within abroad depression fed by local run-off from nearbyhills (Despair Flat). One was on stony rises close to asandstone mesa dotted with gilgais, one was in anephemeral herbfield, four contained herbfields andgilgais on ironstone, while the remaining six siteswere situated on undulating ironstone plains dottedwith vegetated patches (gilgais and small drainagechannels) of varying sizes.Floristic diversity was concentrated in the ephem-
eral floodplain and depression land types, which hadthe highest average species richness (22.5 and 23 spe-cies per 14 m2 respectively), comprised primarily ofannual forbs (Fig. 3). On ironstone plains, diversityand biomass were mostly restricted to gilgais andsmall drainage channels, with average species rich-ness per quadrat of 13.8, compared to 9.3 on ephem-eral herbfields which sometimes occur adjacent tothem. No quadrats were located on ironstone plainsthemselves, which generally support no plants exceptoccasional small annuals Eriachne pulchella and/orTrianthema triquetra. The gilgai land type as a wholesupported a slightly higher richness and abundanceof annual grasses than other land types, including thefloodplain and depression (Fig. 3).The Mitchell grassland quadrats unsurprisingly had
the highest perennial grass frequency and rank abun-dance, but were otherwise quite similar to ironstonegilgais in floristic diversity and lifeform composition.Herbfields tended to occur adjacent to gilgais orclose to small drainage lines, and were dominated byephemeral flora with isolated perennial chenopodsand/or Astrebla tussocks and had the lowest overallspecies richness. The stony rise quadrats were alsofloristically similar to gilgais, but with lower annualgrass frequency and abundance (Fig. 3).Annuals (including geophytic species with annual
stems) comprised 70% of the flora, including 59herb, 22 grass and three sedge species. Annualherbs were the most frequent and abundant lifeformin all land types, with the exception of gilgais wherethey were slightly eclipsed by both perennial herbsand annual grasses. Twenty-six perennial herbs,seven perennial grasses, three shrubs and two treeswere recorded. Apart from sedges (three speciesrecorded at three sites), perennial grasses had by farthe lowest richness and abundance across all landtypes, with richness only greater than one speciesper 14 m2 in three land types (Depression,
© 2017 Ecological Society of Australia doi:10.1111/aec.12508
NIGHT PARROT MOVEMENTS AND HABITATS 5
(a)
(c) (d)
(f)
(e)
(b)
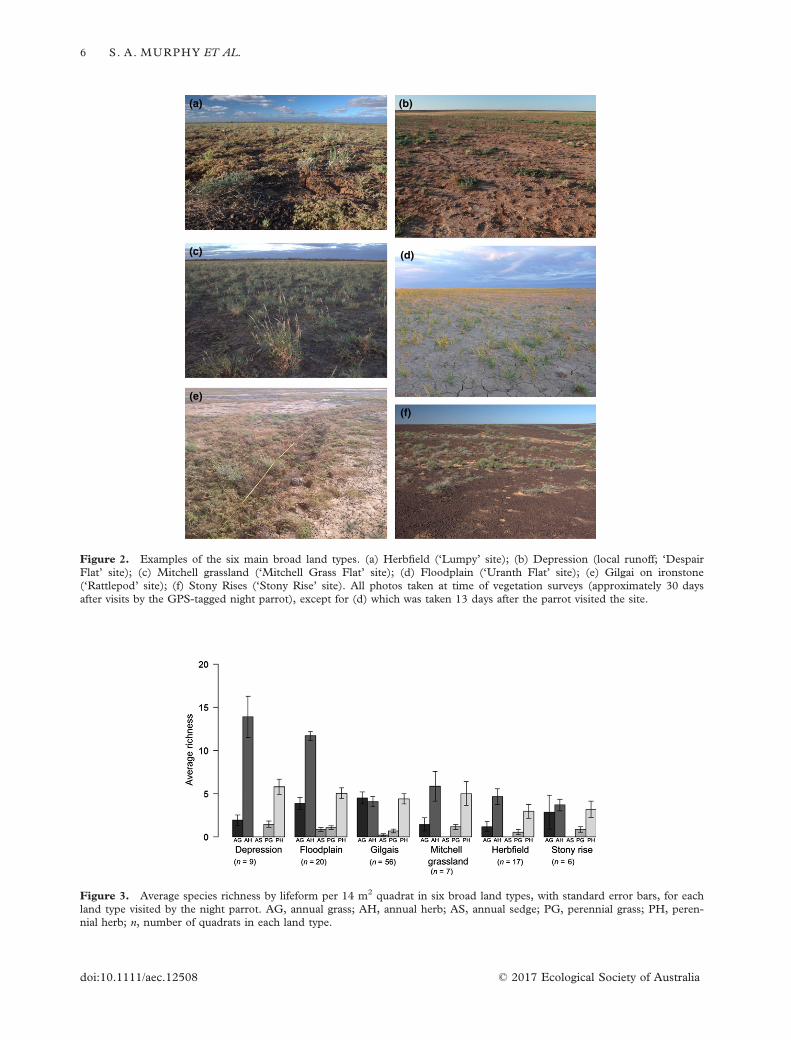
Figure 2. Examples of the six main broad land types. (a) Herbfield (‘Lumpy’ site); (b) Depression (local runoff; ‘DespairFlat’ site); (c) Mitchell grassland (‘Mitchell Grass Flat’ site); (d) Floodplain (‘Uranth Flat’ site); (e) Gilgai on ironstone(‘Rattlepod’ site); (f) Stony Rises (‘Stony Rise’ site). All photos taken at time of vegetation surveys (approximately 30 daysafter visits by the GPS-tagged night parrot), except for (d) which was taken 13 days after the parrot visited the site.
Figure 3. Average species richness by lifeform per 14 m2 quadrat in six broad land types, with standard error bars, for eachland type visited by the night parrot. AG, annual grass; AH, annual herb; AS, annual sedge; PG, perennial grass; PH, peren-nial herb; n, number of quadrats in each land type.
doi:10.1111/aec.12508 © 2017 Ecological Society of Australia
6 S. A. MURPHY ET AL.

Floodplain and Mitchell Grassland). With theexception of Mitchell grasslands, perennial grasseswere sparse and patchy, and only occurred in gilgaisand small drainage channels. Appendix S5 showsthe frequency and abundance of species that wererecorded in all quadrats.Four sites were apparently favoured feeding
grounds during the tracking period (Appendix S6).Two are low-lying, periodically inundated plains: anouter floodplain (Uranth Flat), where the bird spentat least 4 h over four nights, and a broad, shallowdepression fed from local run-off (Despair Flat; a sin-gle visit for at least one hour). These sites were thesole representative of the floodplain and depressionland types respectively (Appendix S6). Soils at bothsites are strongly self-mulching clays, and both areintersected by small drainage channels less than0.5 m deep. Despite their different hydrology, bothsites occur on fertile cracking clay soils and are char-acterized by a diverse ephemeral flora, having thehighest species richness across all sites and comprisedprimarily of annual herbs (Appendix S7). Perennialforbs were the next most frequent and abundant life-form at both sites, while annual grasses made a sub-stantial contribution at Uranth Flat (Appendix S7).There were occasional shrubs (Acacia farnesiana andEremophila polyclada) at Despair Flat, but no trees orshrubs at Uranth Flat.The two other main sites visited by the bird, Osteo
and Stony Flat, were located on ironstone plains dot-ted with vegetated gilgais of varying size and floristiccomposition (gilgais on ironstone land type). Sam-phire (Tecticornia spp.) was dominant in the wetterareas of Stony Flat, while deeper gilgais supportedscattered perennial grasses, mostly Astrebla pectinataand Eragrostis xerophila. Apart from the Nardoo site,all other ironstone gilgai sites appeared to be shortvisits or fly-overs. Although overall species richnesswas lower than at inundated sites, these sites had thehighest annual grass richness and abundance of allsites (Appendix S7).Analysis of dispersion (ANDOSIP) using species
abundance scores across all sites revealed thatquadrats within Nardoo, Hoofprint and Stony Risesites were significantly more variable than othersites because they spanned multiple land types.Uranth and Despair Flats were clear outliers in thefloristic ordination, although the quadrats from dee-per gilgais at Nardoo had some overlap with theformer (Fig. 4). Stony Flat and Osteo overlappedwith each other, but were mostly separate from theother sites in the ordination space, having someoverlap with Stony Rise. Although Analysis of Sim-ilarity (ANOSIM) revealed that differences in plantcomposition between sites were not significant,numerous species occurred only at the main feed-ing sites.
Diet inferences
Although it is impossible to determine with certaintywhat the bird was eating without direct observation,or scat or crop analyses, the floristic data providesome insights into what drew the bird repeatedlyand/or for lengthier periods to the four main sites,and a further three where it spent at least 20 minduring a single visit (here grouped as feeding sites).Fifty-five species were frequent (occurred in >50% ofquadrats) at one or more of these seven feeding sites(Appendix S8). Twenty of these were widespreadand abundant across the majority of the 16 sites.These include prolifically seeding short-lived herbs(notably Portulaca oleracea, T. triquetra and Boerhaviaschomburgkiana) and grasses (including A. pectinata,Dactyloctenium radulans, Enneapogon polyphyllus, Isei-lema vaginiflorum and Tripogon lolliiformis), which arepotential food sources but could be found through-out the landscape. A further nine species are mostlyrestricted to one or two land types, but locally abun-dant within them at both feeding and non-feedingsites (Appendix S8).
Figure 4. Two dimensional non-metric multidimensionalscaling (NMDS) ordination diagram of untransformedfloristic frequency data from 115 quadrats where the GPS-tagged night parrot spent time. Stress = 0.205. Key feedingsites identified as follows: black circles, Uranth Flat; hollowcircles, Despair Flat; stars, Osteo; crossed circles, Stonyflat. Other sites as follows: hollow squares, Nardoo; trian-gles inside squares, Succulent Flat; upside-down triangles,Mitchell Edge.; triangles, Flowering Lignum flat; crosses,Hoofprint; Grove, plus sign; black triangles, Lumpy;crossed squares, Mitchell grass flat; asterisks, Stony rise;circles with + sign, Rattlepod Gully; crossed squares, SandyBlight; black squares, Unlikely Plains. Ninety-five percentconfidence ellipses are placed around the favoured feedingsite clusters of Uranth Flat, Despair Flat, Stony Flat andOsteo quadrats.
© 2017 Ecological Society of Australia doi:10.1111/aec.12508
NIGHT PARROT MOVEMENTS AND HABITATS 7

Eleven species occurred across numerous sites butwere only frequent and abundant at one or more ofthe feeding sites, while fifteen occurred only at thefeeding sites. Of particular note are four prolificallyseeding, large-seeded annual grasses (U. truncatum,Xerochloa barbata, Brachyachne prostrata and Tragusaustraliense; the three former being locally restrictedto the mid-reaches of the Diamantina; QueenslandHerbarium records). Similarly, numerous large-seeded and prolific annual forbs were also foundcommonly at one or more feedings sites, includingPtilotus murrayi, two Portulaca species, two Curcur-bitaceae vines and Alternanthera denticulata.
DISCUSSION
Understanding species’ movements, habitat use andkey resources within habitats is vital for conservationmanagement. The discovery of a night parrot popula-tion in south-western Queensland in 2013 (Dooley2013), coupled with advances in the miniaturizationof tracking devices, has allowed us to provide the firstdetailed information about night parrot habitat useand movements. Our small sample size necessitatescaution in interpreting the data, but knowledge gath-ered in this study is unique and will underpin boththe management of the known populations andsearches for additional ones. It will also provide thebasis for further research, which is necessary givenonly two individuals were studied at a single location.
Activity patterns and movements
Both tagged birds displayed similar behaviour interms of a brief period of vocal socialization for about30 min after sunset and upon returning to theirroosts within an hour of sunrise. Both birds alsomoved similar (known maximum) distances awayfrom the roost (ca. 8 and 9.4 km for the female andmale respectively). The value of GPS tag technologywas demonstrated by the increased resolution it pro-vided, showing greater than expected movementincluding a total of at least 41 km travelled in a sin-gle night. This scale of nightly movement contrastsstarkly with that known for the congeneric easternground parrot (Pezoporus wallicus). For example, themean home range for 12 radio-tagged eastern groundparrots was 9.2 ha, compared to over 3000 ha forour GPS-tagged night parrot (McFarland 1991). Wesuggest the differences may be related to the morewidely dispersed resources used by night parrots intheir arid environment compared to mesic, coastalhabitats of eastern ground parrots. Night parrotroosting and foraging habitats were widely separatedin our study, whereas the heathlands and sedgelands
occupied by eastern ground parrots fulfil both roost-ing and feeding needs (McFarland 1991).At a broader scale we found no evidence of nomadic
behaviour; both tracked birds were captured approxi-mately 7 km apart in consecutive years (one dry, onewet) and moved within the same relatively small area(~70 km2). Furthermore, the species has continuouslyoccupied the same area for 4 years based on acousticmonitoring (S. Murphy, unpublished data), and weassume that at least some individuals have been resi-dent through the entire period. Indeed the populationmay have been resident for a much longer time giventhe close proximity of the 2006 specimen location(McDougall et al. 2009). This contrasts with previousassumptions of nomadism (Reid & Fleming 1992;Higgins 1999) that may have arisen because ofAndrews’ (1883) observations cited earlier. An impor-tant implication of the night parrot being more seden-tary than previously assumed is that all localities wherethe species was recorded historically should becomethe focus of renewed, intensive survey.On 9 May 2016 (day two of the tracking period),
38.2 mm of rain fell, and this may explain why thebird was only detected visiting a watering point once(Appendix S6). While it is possible that the bird vis-ited this site (or other permanent water points) onother nights given the intermittent tracking schedule,ephemeral water was abundant during the trackingperiod. Although we cannot conclude with certaintythat the bird actually drank while at water on nighttwo, it seems reasonable to assume that it did, andso this corroborates earlier assertions that night par-rots fly to water, sometimes over considerable dis-tances (Andrews 1883; McGilp 1931).Use of regular roost sites to which our tagged night
parrots returned before dawn each night confirms ear-lier accounts (Andrews 1883; McGilp 1931). However,both birds were absent at times from their respectiveroosting areas. The male relocated 5.6 km and thefemale apparently relocated to an unknown location for12 days. Both movements may have been in responseto capture stress (for the female), or disturbance whilerecapture attempts were being made (in the case of themale). Further studies are required to determine ifnight parrots preferentially use a single roost or can usemultiple roosts within a large home range, perhapsdepending on the proximity to feeding areas which wesuspect will shift through time and space.
Habitats
Both tagged birds roosted in long-unburnt Triodiahummocks, which is consistent with the establishedassociation between Triodia and night parrots(Andrews 1883; Whitlock 1924; McGilp 1931; Wil-son 1937). However, the total extent of Triodia
doi:10.1111/aec.12508 © 2017 Ecological Society of Australia
8 S. A. MURPHY ET AL.

patches at these sites was not large; one roosting areahad only 1.9% Triodia cover on a sparsely vegetatedironstone plain.From Triodia roosts, both birds travelled relatively
large distances into non-Triodia habitats. The GPSdata clearly show the bird making use of fertile,diverse but ephemeral parts of the landscape: flood-plains, run-on areas and gilgais. The two sites withthe highest plant species richness appeared to befavoured feeding grounds, as were the sites with thehighest diversity and abundance of prolifically seed-ing annual grasses, most notably U. truncatum, andherbs (Fig. 3; Appendix S7).It is important to note that, despite floristic similar-
ity, variation exists within broad land types and thismay explain why some sites were visited more fre-quently or for longer. Some ironstone sites, notablyOsteo, Stony Flat and Nardoo, had more extensivegilgai development and therefore more total seedproduction than others. The geographical context offavoured sites also appeared important, with StonyFlat and Osteo being closest to the roost site. Thespatial configuration of habitat types is likely to beimportant in determining optimal landscapes for sus-taining night parrot populations.The GPS tracking was done on one bird in an
exceptionally wet season. Many of the plant speciesrecorded only persist for a brief period (weeks).However, we suggest that seeds from these ephem-eral species might be available to night parrots wellinto dry spells, based on movements of the radio-tagged bird, which we detected in these same areasduring a dry period. Further anecdotal support isprovided by the long-term occupancy of the generalarea by night parrots through the 2013–2015 drought(S. Murphy, unpublished data). Despite the higheffort, cost and risk involved, we strongly recommendmore tracking data be obtained during dry periods.This information will be critical to informing conser-vation management.Apart from feeding, another reason for the GPS-
tagged night parrot to visit ironstone plains frequentlycould be for ingesting gastroliths – small stones swal-lowed to aid food comminution (Wings 2007).Autopsies revealed similar grit in the gizzards of twonight parrot specimens (Murie 1868; McDougallet al. 2009).
Habitat comparisons with other night parrotlocalities
There are broad similarities in habitat and floristicsbetween most published sightings of night parrotsand the study area. Most had areas of Triodia spp. onstony/hilly terrain juxtaposed with more fertile plainssupporting vegetation dominated by chenopod and/or
grass species (e.g. Black 2012; J. Reid & S. Murphy,unpubl. data, 2015; Paton 1970; Forshaw et al.1976; Higgins 1999; McDougall et al. 2009). Thisincludes the 1990 Boulia specimen locality, whichwas characterized by gilgais on an ironstone plainsand patches of Triodia nearby (contra Boles et al.1994). A similar range of landforms (alluvial plainsand hillslopes) and vegetation types (Triodia andmixed herb-grasslands) were recorded in the SelwynRanges (north-western Queensland) around theseven locations where night parrots were reported in1992–1993 (Garnett et al. 1993). The sighting of twonight parrots at water at Fortescue Marsh in WesternAustralia in 2005 occurred close to extensive plainsof samphire (a chenopod) alongside expanses of Trio-dia (Davis & Metcalf 2008), while the March 2017confirmed sighting in Western Australia was from aspinifex plain adjacent to an open chenopod shrub-land dotted with a chain of ephemeral pools fringedby samphire (Tecticornia spp.; N. Jackett andB. Greatwich, in litt. 8 April 2017).There are regions where night parrots occurred his-
torically which have no Triodia or limited amounts inthe general area of the observations. In these areas,shrubby samphire or lignum may have providedroosting sites (Andrews 1883). We concur with Gar-nett et al. (1993) that night parrots can be found in awide range of habitats, but highlight here for the firsttime the importance of feeding areas in proximity todense ground layer vegetation for roosting and nest-ing (Murphy et al. 2017). It is likely that the speciesneeds access to free-standing water, at least duringhot conditions (Kearney et al. 2016).
Conservation implications and future researchdirections
Available evidence strongly suggests that this popula-tion is resident rather than nomadic, but rangesacross a far greater area (>3000 ha over five nights)than previously known. In particular, we haverevealed extensive use by night parrots of habitatsthat are distant, and structurally and floristically dis-tinct, from their roost sites. These are the focus ofthe grazing industry in the region (Phelps et al.2007), but the impact of cattle grazing on the habi-tats used by the night parrot, particularly on seedproduction of key species, is unknown. Previousstudies have shown minimal impacts of grazing onfloristic composition in the Channel Country flood-plains and dunefields (Phelps et al. 2007; Silcock &Fensham 2012) and in Mitchell grasslands (Fenshamet al. 2014). However, floodplain grazing exclosureshave demonstrated significant reductions in vegeta-tion biomass and groundcover at typical levels ofcommercial cattle grazing (Phelps et al. 2007).
© 2017 Ecological Society of Australia doi:10.1111/aec.12508
NIGHT PARROT MOVEMENTS AND HABITATS 9

Studies in other areas have shown negative impactsof livestock on seed production of grasses and herbs(Friedel & James 1995; Crowley & Garnett 2001).The impacts of reduced biomass on seed productionand availability in the study area require furtherinvestigation, using grazing exclosures and repeatedmeasurements across different seasons.Further night parrot surveys in similar habitat are
also required, and our initial tracking results andhabitat descriptions provide a good basis for identify-ing potentially suitable habitat: that is, T. longicepsranges abutting areas containing an abundance oflarge-seeded, prolific species, particularly floodplainsand ironstone plains with drainage lines and/or gil-gais. Mapping of spinifex and flooded areas is cur-rently being undertaken using remote sensing andground validation. This habitat model will identifyhigh-priority areas to survey, using acoustic monitor-ing and targeted searching for nest/roost sites. Weanticipate that additional night parrot populationswill be located in the region broadly circumscribedby Cloncurry, Boulia, Winton and Jundah, due tothe similarities in landscapes and vegetation with ourstudy area, and also because of the number of nightparrot sightings in the region over the past 25 years(Boles et al. 1994; S. Murphy unpublished data,2017; Garnett et al. 1993).
ACKNOWLEDGEMENTS
We acknowledge the significant contribution JohnYoung made through his initial discovery of this site.We also acknowledge the support of Fortescue MetalsGroup (especially Cath Bozanich, Martin Heller andShaun Grein), Peter and Carol Britton, Whatty Camp-bell, Greg and Justine Clark, Bob McDonald, BushHeritage Australia and members of the Night ParrotRecovery Team. We thank Rod Fensham for helpfuldiscussions about habitat surveys and Boris Laffineurfor producing Figures 2 and 3. John Neldner, DaleRichter and Chris Pennay also provided valuable assis-tance with pre-release Regional Ecosystem informa-tion. We appreciate the Western Australian nightparrot habitat insights provided by Nigel Jacket andBruce Greatwich. JS thanks the National Environmen-tal Science Programme for supporting her contribu-tion to this work. This research was conducted underScientific Purposes Permit Number WISP14627114and QLD Department of Agriculture and FisheriesAnimal Ethics Approval number SA 2015/09/521.
REFERENCES
***: These references cited in Appendices S2 andS3.
Andrews F. W. (1883) Notes on the Night Parrot. Trans. Proc.Roy. Soc. S. Aust. 6, 29–30.
Austin R. (1855) Journal of Assistant-Surveyor R. Austin. . . E.Stirling. Government Printer, Perth.
Australian Government (2016) Species Profile and ThreatsDatabase: Pezoporus occidentalis – Night Parrot. Departmentof the Environment, Canberra.
Black A. B. (2012) Collection localities of the Night ParrotPezoporus (Geopsittacus) occidentalis (Gould, 1861). Bull. Br.Orn. Club 132, 277–82.
Boles W. E., Longmore N. W. & Thompson M. C. (1994) Arecent specimen of the Night Parrot Geopsittacusoccidentalis. Emu 94, 37–40.
Bostock P. D. & Holland A. (2007) Census of the QueenslandFlora 2007. Queensland Herbarium, Brisbane.
Bureau of Meteorology (2017) Climate Data Online; BrightonDowns, Station Number 37007.
Cleland J. B. (1930) Notes on the birds of Central Australia. S.Austr. Ornithol. 10, 160–6.
Crowley G. & Garnett S. (2001) Growth, seed production andeffect of defoliation in an early flowering perennial grass,Alloteropsis semialata (Poaceae), on Cape York Peninsula,Australia. Aust. J. Bot. 49, 735–43.
Davis R. & Metcalf B. (2008) The Night Parrot (Pezoporusoccidentalis) in northern Western Australia a recent sightingfrom the Pilbara region. Emu 108, 233–6.
Dooley S. (2013) Out of the shadows. Aust. Birdlife 2, 26–30.Ellis R. (1982) Bush Safari. Rigby Publishers, Adelaide.Fensham R. J., Silcock J. L. & Firn J. (2014) Managed livestock
grazing is compatible with the maintenance of plantdiversity in semi-desert grasslands. Ecol. Appl. 24, 503–17.
Forshaw J. M., Fullagar P. J. & Harris J. I. (1976) Specimensof the Night Parrot in museums throughout the world.Emu 76, 120–6.
***Fridolfsson A.-K. & Ellegren H. (1999) A simple anduniversal method for molecular sexing of non-ratite birds.J. Avian Biol. 30, 116–21.
Friedel M. & James C. D. (1995) How does grazing of nativepastures affect their biodiversity? In: ConservingBiodiversity: Threats and Solutions (eds A. Bradstock, T.Auld, D. Keith, R. Kingsford, D. Lunney & D. Sivertsen)pp. 249–59. Surrey Beatty & Sons, Chipping Norton, NewSouth Wales.
Garnett S., Crowley G., Duncan R., Baker N. & Doherty P.(1993) Notes on live night parrot sightings in North-Western Queensland. Emu 93, 292–6.
***Griffiths R., Double M. C., Orr K. & Dawson R. J. (1998)A DNA test to sex most birds. Mol. Ecol. 7, 1071–5.
Higgins P. J. (1999) Handbook of Australian, New Zealand andAntarctic Birds. Vol. 4. Parrots to Dollarbird. OxfordUniversity Press, Melbourne.
Howe F. E. & Tregellas T. H. (1914) Rarer birds of themallee. Emu 14, 71–84.
Kearney M. R., Porter W. P. & Murphy S. A. (2016) Anestimate of the water budget for the endangered nightparrot of Australia under recent and future climates. Clim.Change Responses 3, 14–31.
***Lotek (2016) PinPoint Host Software for PinPoint GPSTags. User Manual Revision 05. Ontario, Canada.
McDougall A., Porter G., Mostert M. et al. (2009) Anotherpiece in an Australian ornithological puzzle – a second NightParrot is found dead in Queensland. Emu 109, 198–203.
McFarland D. (1991) The biology of the Ground Parrot,Pezoporus wallicus, in Queensland. I. Microhabitat use,activity cycle and diet. Wildl. Res. 18, 169–84.
doi:10.1111/aec.12508 © 2017 Ecological Society of Australia
10 S. A. MURPHY ET AL.

McGilp J. N. (1931) Geopsittacus occidentalis, Night-Parrot. S.Austr. Ornithol. 11, 68–70.
Morrison D. A., Le Broque A. F. & Clarke P. J. (1995) Anassessment of some improved techniques for estimating theabundance (frequency) of sedentary organisms. Vegetatio120, 131–45.
Murie J. (1868) On the nocturnal Ground-Parrakeet (Geopsittacusoccidentalis, Gould). Proc. Zool. Soc. Lond. 1868, 158–65.
Murphy S. A., Austin J. J., Murphy R. K. et al. (2017)Observations on breeding Night Parrots (Pezoporus occidentalis)in western Queensland. Emu Aust. Ornithol. 117, 107–13.
Oksanen J., Blanchet F. G., Friendly M. et al. (2017) vegan:Community Ecology Package. R package version 2.4-2.[Cited 16 February 2017] Available for URL: http://CRAN.R-project.org/package=vegan
Parker S. (1980) Birds and conservation parks in the north-eastof South Australia. South Aust. Parks Conserv. 3, 11–8.
Paton J. B. (1970) Additional notes. S. Austr. Ornithol. 25,208–9.
Phelps D., Lynes B. C., Connelly P. T., Horrocks D. J., FraserG. W. & Jeffery M. R. (2007) Sustainable Grazing in theChannel Country Floodplains (Phase 2). Meat & LivestockAustralia, North Sydney.
Powell B. (1970) The Night Parrot. S. Aust. Ornithol. 25, 208.R Core Team. (2016) R: A Language and Environment for
Satistical Computing. R Foundation for StatisticalComputing, Vienna, Austria. [cited 16 February 2017]Available for URL: https://www.R-project.org/.
Reid J. & Fleming M. (1992) The conservation status of birdsin arid Australia. Rangeland J. 14, 65–91.
Silcock J. L. & Fensham R. J. (2012) Arid vegetation indisequilibrium with livestock grazing: evidence from long-term exclosures. Austral Ecol. 38, 57–65.
Thackway R. & Cresswell I. D. (1995) An interim biogeographicregionalisation for Australia: a framework for establishing anational system of reserves, version 4.0. Australian NatureConservation Agency, Canberra. Current version 7. [Cited19 September 2016.] Available from URL: http://www.environment.gov.au/parks/nrs/science/bioregion-framework/ibra/index.html
Whitlock F. L. (1924) Journey to Central Australia in search ofthe Night Parrot. Emu 23, 248–81.
Wilson H. (1937) Notes on the Night Parrot, with referencesto recent occurrences. Emu 37, 79–87.
Wings O. (2007) A review of gastrolith function withimplications for fossil vertebrates and a revisedclassification. Acta Palaeontol. Pol. 52, 1–16.
SUPPORTING INFORMATION
Additional Supporting Information may be found inthe online version of this article at the publisher’sweb-site:
Appendix S1. Acquisition schedule for GPS tag.Appendix S2. Interpreting night parrot behaviour atsingle GPS fixes.Appendix S3. Molecular sex determination methods.Appendix S4. Summary of signal detections for the2015 radio tagged night parrot.Appendix S5. Species frequency (% of quadratsoccurring in) and abundance (average abundancescore per quadrat) for each broad land type wherethe GPS-tagged night parrot was recorded.Appendix S6. Details of the locations visited by theGPS-tagged night parrot, and inferred type of activityat each.Appendix S7. Average species and lifeform richness(per 14 m2 quadrat) and abundance at night parrothabitat sites.Appendix S8. Frequent species (defined as occur-ring in >50% of quadrats at any one site) at sevennight parrot feeding sites.
© 2017 Ecological Society of Australia doi:10.1111/aec.12508
NIGHT PARROT MOVEMENTS AND HABITATS 11

Page 16 of 27
4. Increasing knowledge of Night Parrot distribution
This research topic relates to Part B of the Condition, which states:
b) upon completion of the research identified by (condition 10 a) and no more than 4 years from the date of this approval, conduct or fund comprehensive targeted surveys for the Night Parrot in at least 3 sites in areas where confirmed sightings or specimens of Night Parrot have been recorded since 1 January 2000. The surveys must be informed by the outcomes of the research identified at (condition 10 a) and undertaken by a suitably qualified expert;
It is worth noting that, at the time the Condition was written, there was only one (Diamantina
National Park (Cupitt and Cupitt 2008; McDougall et al. 2009), or arguably two including the
Cloudbreak record (Davis and Metcalf 2008), confirmed locations of Night Parrots since 2000, and
therefore the Condition could not be met. Nonetheless, we conducted Night Parrot surveys using
proven acoustic (discussed in Section 2.2) techniques at ten locations that had recent high veracity
reports as well as older (pre-2000) confirmed records. Figure 6 and Table 5 provides details of the
Song Meters deployed during the research program in relation to confirmed and unconfirmed Night
Parrot records. For some confirmed or high veracity records that we wished to visit, the exact
location was uncertain and in these cases we selected habitat that matched our understanding of
Night Parrot habitat that was close to the nominal location of the record. An example of this is the
1990 roadkilled specimen north of Boulia, which was collected at a location where there was no
Triodia, and indeed it may have been transported from a considerable distance to its eventual
resting place (Boles et al. 1994). As such, we deployed recorders west and north-west of that
location in extensive Triodia areas.
In addition to the ten historical Night Parrot locations surveyed in the eastern part of the
distribution, we also retrospectively analysed 2397 hours of acoustic data collected between 2013
and 2018 from Fortescue Marsh where the 2005 sighting was made that led to the EPBC Condition
(Davis and Metcalf 2008).
For analysis all acoustic data, we used the Song Scope recogniser that is known to be effective at
detecting three of the main call types. It is important to note that this recogniser has a high
commission error rate, which can be an advantage because it is likely to be able to detect calls that
are different to those used as training/reference calls; with the differences relating to geographic
variation. All putative Night Parrot detections were screened visually (using spectrograms) and
aurally.
No Night Parrot calls were detected at any of the locations away from Pullen-Pullen/Diamantina
National Park, using the Song Scope recogniser.
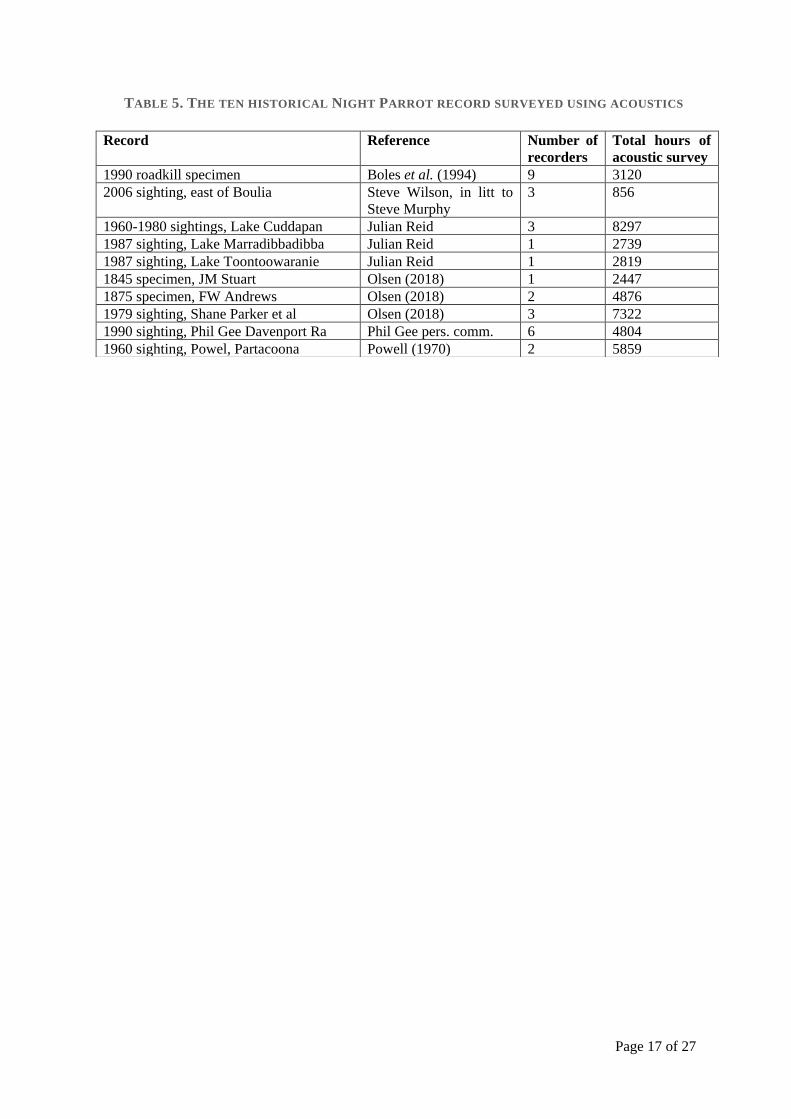
Page 17 of 27
TABLE 5. THE TEN HISTORICAL NIGHT PARROT RECORD SURVEYED USING ACOUSTICS
Record Reference Number of
recorders
Total hours of
acoustic survey
1990 roadkill specimen Boles et al. (1994) 9 3120
2006 sighting, east of Boulia Steve Wilson, in litt to
Steve Murphy
3 856
1960-1980 sightings, Lake Cuddapan Julian Reid 3 8297
1987 sighting, Lake Marradibbadibba Julian Reid 1 2739
1987 sighting, Lake Toontoowaranie Julian Reid 1 2819
1845 specimen, JM Stuart Olsen (2018) 1 2447
1875 specimen, FW Andrews Olsen (2018) 2 4876
1979 sighting, Shane Parker et al Olsen (2018) 3 7322
1990 sighting, Phil Gee Davenport Ra Phil Gee pers. comm. 6 4804
1960 sighting, Powel, Partacoona Powell (1970) 2 5859
Page 18 of 27
FIGURE 6. LOCATION OF THE HISTORICAL NIGHT PARROT RECORDS AND THOSE
SURVEYED DURING THE RESEARCH PROGRAM

Page 19 of 27
5. Understanding the threats to Night Parrots
Murphy, S.A., Paltridge, R., Silcock, J., Murphy, R., Kutt, A.S., and Read, J. (2017).
Understanding and managing the threats to Night Parrots in south-western Queensland. The Emu
- Austral Ornithology 117, 135-145.

Full Terms & Conditions of access and use can be found athttp://www.tandfonline.com/action/journalInformation?journalCode=temu20
Download by: [116.240.73.116] Date: 29 October 2017, At: 23:01
Emu - Austral Ornithology
ISSN: 0158-4197 (Print) 1448-5540 (Online) Journal homepage: http://www.tandfonline.com/loi/temu20
Understanding and managing the threats to NightParrots in south-western Queensland
Stephen A. Murphy, Rachel Paltridge, Jennifer Silcock, Rachel Murphy, Alex S.Kutt & John Read
To cite this article: Stephen A. Murphy, Rachel Paltridge, Jennifer Silcock, Rachel Murphy, AlexS. Kutt & John Read (2017): Understanding and managing the threats to Night Parrots in south-western Queensland, Emu - Austral Ornithology
To link to this article: http://dx.doi.org/10.1080/01584197.2017.1388744
Published online: 29 Oct 2017.
Submit your article to this journal
View related articles
View Crossmark data

Understanding and managing the threats to Night Parrots in south-westernQueenslandStephen A. Murphya,b, Rachel Paltridgeb,c, Jennifer Silcockd, Rachel Murphya,b, Alex S. Kutte,f and John Readg,h
aAdaptive NRM, Malanda, Australia; bResearch Institute for the Environment and Livelihoods, Charles Darwin University, Casuarina, Australia;cDesert Wildlife Services, Alice Springs, Australia; dThreatened Species Recovery Hub, National Environmental Science Program, University ofQueensland, St Lucia, Australia; eSchool of BioSciences, University of Melbourne, Parkville, Australia; fBush Heritage Australia, Melbourne,Australia; gSchool of Earth and Environmental Sciences, University of Adelaide, Adelaide, Australia; hEcological Horizons, Kimba, Australia
ABSTRACTSouth-western Queensland supports a suite of threatened native species, including Night Parrots.We investigated why this species has persisted in the region and discovered low prevalence ofthe typical factors that are thought to explain fauna attrition elsewhere in central Australia. Foxesappear to be completely absent. Feral cats were recorded relatively infrequently and showed asignificant preference for habitats less commonly used by Night Parrots, a partition that may bedriven by the presence of dogs that were detected twice as frequently as cats. Our study area hashad a long history of moderate grazing pressure, which is concentrated mostly in productivealluvial habitats. We detected very few herbivores, and dog scat analyses suggest that macropodpopulations are regulated by predation. Archival imagery shows that large fires are not a featureof this landscape, resulting in the long-term, stable availability of patchy Triodia habitats sepa-rated by natural no-fuel areas. Based on these empirical data, we postulate that low non-nativepredator pressure, long-term stable availability of Triodia cover and a productive landscape thathas had only moderate grazing pressure are the interacting factors that may explain why NightParrots have persisted in south-western Queensland. We present practical management actionsthat could enhance the suitability of this landscape for Night Parrots.
ARTICLE HISTORYReceived 3 May 2017Accepted 21 September 2017
KEYWORDSParrots; conservation;endangered species;nocturnal birds; threatenedspecies; wildlifemanagement
Introduction
Night Parrots (Pezoporus occidentalis) are small (~100g; Murphy et al. 2017b), nocturnal parrots that wereonce found across Australia’s vast arid zone. Specimenshave been collected from an area spanning more than1.2 million km2 and there is some evidence to suggestthat the species was relatively common at least at someplaces throughout this range. For example, 21 of the 28museum skins known to exist were collected between1871 and 1881 at a single location (the Gawler Ranges)in South Australia (Black 2012). At Alice SpringsTelegraph Station in 1892 pet cats killed enoughNight Parrots for their owner to make several mountedwall features from their wings and tails (Ashby 1924).
Despite their previously vast range and apparentabundance at some times and places, irrefutable NightParrot records were absent for most of the 20th cen-tury. The last intentionally collected specimen wastaken in 1912 in the Gascoyne region of WesternAustralia (Wilson 1937). It was 78 years before thenext indisputable record surfaced when museumornithologists found a desiccated body by a roadside
in south-western Queensland (Boles et al. 1994).Sixteen years after that record, a Queensland ParksService ranger found another dead bird in south-wes-tern Queensland (Cupitt and Cupitt 2008; McDougallet al. 2009) and in 2013 the first photographs of a livebird were obtained in roughly the same area (Dooley2013). Since then, field research in south-westernQueensland has started to build a better understandingof this enigmatic species, including aspects of breeding(Murphy et al. 2017a), home range and habitat use(Murphy et al. 2017b), vocalisations and diet.
The apparent precipitous decline of Night Parrots incentral Australia was not an isolated phenomenon.Bush Stone-Curlews (Burhinus grallarius), for example,went from being ‘seldom absent from the many local-ities visited during the expedition [to CentralAustralia]’ (Whitlock 1924, p. 256) to being absent orextremely rarely reported from central Australia incontemporary times (Reid and Fleming 1992; Baxterand Paton 1998). Although there are similar examplesinvolving declines in other bird species or groups (e.g.Grasswrens (Amytornis spp.) (Skroblin and Murphy
CONTACT Stephen A. Murphy [email protected]
EMU - AUSTRAL ORNITHOLOGY, 2018https://doi.org/10.1080/01584197.2017.1388744
© 2017 BirdLife Australia
Dow
nloa
ded
by [
116.
240.
73.1
16]
at 2
3:01
29
Oct
ober
201
7

2013) and Plains Wanderers (Pedionomus torquatus);Bennett 1983) it is the rapid declines of native mam-mals that have most dramatically underscored theseverity of ecosystem dysfunction in this region. Tenarid zone mammal species are wholly extinct, and afurther 21 taxa that are either regionally extinct or haveexperienced severe range contractions (EnvironmentProtection and Biodiversity Conservation Act 1999, ter-restrial mammal list, February 2017).
While the primary cause of this profound ecologicalperturbation is still debated, most ecologists agree thatit was likely to involve interacting factors rather than asole threatening process (Morton 1990; Dickman 1996;Johnson and Isaac 2009; Morton et al. 2011). In arecent review McKenzie et al. (2007) found that sixfactors interacted to cause fauna attrition acrossAustralia’s mainland bioregions. Four were intrinsiccharacteristics: phylogenetic similarity, body weight,distribution, area, and tendency for burrowing. Theother two were environmental variables, namely rain-fall (as a surrogate for regional productivity) and theextent of ecological change since European settlement,which included vegetation clearing, grazing pressure,predation by introduced mammals and changed fireregimes. Of the six interacting factors identified byMcKenzie et al. (2007) it is the suite related to ecolo-gical change since European settlement that can bemanipulated by conservation managers in centralAustralia and is therefore the focus of this paper.
Apart from some relatively small areas associatedwith mining and horticulture, large-scale vegetationclearing has not occurred in central Australia and istherefore not considered further here. However,Australia has more land area under managed grazingthan any other country, with much of central Australiaused for cattle and sheep grazing; the latter mostlyconfined to the eastern and southern portions (Asneret al. 2004). Adding to total grazing pressure, largemacropods (Macropus spp.) occur across the area,most abundantly in semi-arid regions (Pople andGrigg 1999), often alongside high densities of FeralGoats (Capra hircus; Pople and Froese 2012).European Rabbits (Oryctolagus cuniculus) no longerirrupt at the plague densities they once did thanks tothe introduction of myxomatosis and calici viruses inthe 1950s and 1990s, and this has led to dramaticincreases in the range and abundance of formerly raredesert mammals in some areas (Pedler et al. 2016).Nonetheless, they do remain at high densities in places(Scanlan et al. 2006). One-humped Camels (Camelusdromedaries) roam the central deserts (Saalfeld andEdwards 2010) while Feral Horses (Equus callabus),Donkeys (E. asinus) and Pigs (Sus scrofa) occur
patchily (Edwards et al. 2004). These domestic, feraland native herbivores contribute to elevated total graz-ing pressure throughout inland Australia, which his-torically supported relatively low densities of nativemacropods since the extinction of the Pleistocenemegafauna ~45 000 years ago (Johnson 2006;Fensham and Fairfax 2008). While the mechanismsand magnitude of ecological change wrought by live-stock production remain debated (Gill 2005; Silcocket al. 2013), it is clear that the past 150 years of alteredland management practices have modified manyregions, including places that once supported NightParrot populations such as the Gawler Ranges(Andrews 1883; Department for Environment andHeritage 2006).
Feral Cats (Felis catus) inhabit the entireAustralian continent in virtually all habitat types(Legge et al. 2017). Cat predation has contributed to28 mammal extinctions (Woinarski et al. 2014) andcontinues to threaten a further 70 mammals, 40birds, 21 reptiles and 4 amphibians (Commonwealthof Australia 2015). European Foxes (Vulpes vulpes)occur mostly across the southern two-thirds ofAustralia (Atlas of Living Australia (ALA), accessed9 January 2017) and are listed as a threat to 14species of bird, 48 mammals, 12 reptiles and 2amphibians (DEWHA 2008). Although mammalsare the most frequently eaten prey, 20–30% of foxscats from arid areas have been found to containbirds (Palmer 1995; Paltridge 2002; Cupples et al.2011), and ground-nesting birds are thought to beparticularly vulnerable to fox predation (DEWHA2008).
Dogs (treated here as including Dingoes (Canislupus dingo), Feral Dogs (Canis familiaris) and theirhybrids) are a third potential mammalian predator thatmay have contributed to fauna attrition in centralAustralia, being known predators of threatened speciessuch as Greater Bilbies (Macrotis lagotis) (Newsomeet al. 2014), Centralian Brush-tail Possums(Trichosurus vulpecula) (Kerle et al. 1992), somerodents (Allen and Leung 2012) and possibly evenNight Parrots (G. A. Keartland 1894 in Ashby 1924).Notwithstanding the potential for dogs to prey onfauna, several studies have shown that dogs can alsobenefit native species via their regulatory effects on catsand foxes (Newsome et al. 2001; Kennedy et al. 2012;Moseby et al. 2012). Evidence supporting this ‘meso-predator regulation’ effect is variable, which hasrecently fuelled a lively debate in the literature aboutthe scale at which it occurs and whether sufficientevidence exists for managers to maintain dogs in land-scapes to achieve conservation objectives (Allen et al.
2 S. A. MURPHY ET AL.
Dow
nloa
ded
by [
116.
240.
73.1
16]
at 2
3:01
29
Oct
ober
201
7

2013, 2014; Hayward and Marlow 2014; Nimmo et al.2015).
There has been dramatic spatial and temporalchange in contemporary fire patterns in centralAustralia, with a shift from the continuous small-scaleuse of fire under traditional Aboriginal managementtowards less frequent and much larger single-fireevents that typically follow years of above-average rain-fall (Burrows et al. 2006; Edwards et al. 2008).Contemporary fire patterns homogenise the landscapeand periodically remove cover over large areas(Edwards et al. 2008). Such homogenisation can altervegetation structure and floristics, promote the spreadof weeds, increase grazing pressure and make condi-tions favourable for hunting by introduced predators(Kutt and Woinarski 2007; Edwards et al. 2008; Milleret al. 2010; McGregor et al. 2015). As such, changes tofire patterns have been mooted as a potentially signifi-cant mechanism explaining faunal collapse in centralAustralia (Garnett et al. 2011; Woinarski et al. 2014). Itis important to note, however, that there is little evi-dence of changed fire regimes in the eastern parts ofcentral Australia (Silcock et al. 2013; Kimber andFriedel 2015).
It is within this context that we considered thepersistence and management of Night Parrots insouth-western Queensland. Tracking studies (Murphyet al. 2017b) and long-term acoustic monitoring (S.Murphy unpub. data) suggest that at least some of thepopulation is sedentary and this provides a clearimperative to maintain the population by understand-ing and managing the threats to it. In this paper, wefocus on the potential impacts of mammalian preda-tors, herbivores and fire on Night Parrots in south-western Queensland and in doing so present a concep-tual model that may explain why Night Parrots mayhave persisted in the region. We also discuss the keyconservation management options being implementedto enhance the inherent resilience that exists in thissystem.
Methods
Study area
The study was undertaken on Pullen-Pullen Reserveand Mt Windsor Station in south-westernQueensland. Pullen-Pullen is 56 000 ha and was partof Brighton Downs beef cattle station prior to its sub-division and acquisition by the conservation charityBush Heritage Australia in 2016. The reserve sits withinthe Goneaway Tableland subregion of the ChannelCountry biogeographic region (Thackway and
Cresswell 1995). Habitats include sandstone plateausvegetated with Spinifex (Triodia longiceps) andLancewood (Acacia shirleyi), extensive open ironstoneplains dotted with vegetated run-on areas, relativelywooded watercourses dominated by A. aneura and A.cambagei, and productive open floodplains of the outerDiamantina River. Detailed descriptions of habitats andtheir use by Night Parrots appear in Murphy et al.(2017b).
The region is hot and arid with mean maximumtemperatures of 39°C in January and 23°C in July.Annual rainfall is highly unpredictable, with dry peri-ods punctuated by occasional wet years. Median annualrainfall at Brighton Downs is 240 mm and on average65% of the rainfall occurs between December andMarch (Bureau of Meteorology station number37,007). Rainfall was collected at the study site usinga tipping bucket gauge between 12 November 2013 and27 April 2016. The years 2013 and 2015 were very dry,while rainfall in 2014 was slightly above average due tolarge events in February and December, and 2016 wasan extremely wet year, with regular falls including155 mm in March.
Mammalian predators
Photographs from 205 camera trap (Reconyx HC600)locations were examined to determine the presenceof mammalian predators including feral cats, dogsand foxes. Most locations were in the Mayne Rangewhere Night Parrot activity was concentrated (deter-mined by long-term acoustic data; S. Murphy unpub.data), with a smaller number of locations sampled inthe Nisbet Range. Both lured (dried meat) and lure-less cameras were positioned on trees, fence-postsand on star-pickets in all the major habitats in thestudy area. Cameras were set for several reasons aspart of the broader study (e.g. testing as a surveymethod for Night Parrots) and so some were notpositioned optimally to detect predators (Read et al.2015a). However, this did not affect the simpleassessment of predator presence or absence (althoughit was taken into account when calculating detectionrates and for inferential analyses – see below).Cameras were set to ‘rapid fire’ mode withoutdelay, capturing five photographs per trigger. Forinferential analyses and calculations of relative detec-tion rates (for herbivores – see below), individualevents were defined as those that occurred morethan 1 h after the last event (of the same type) oneach camera. Counts of individuals were notincluded in analyses. The average deployment periodwas 120 nights at each location.
EMU - AUSTRAL ORNITHOLOGY 3
Dow
nloa
ded
by [
116.
240.
73.1
16]
at 2
3:01
29
Oct
ober
201
7

No foxes were detected on cameras or during hun-dreds of spotlighting hours, nor were any fox tracks orscats found in the area, and so further presentation ofmethods excludes them. We were interested in whetherthe variation in where cats were detected could beexplained by habitat type and/or the presence of dogs.Using a subset of cameras that were positioned opti-mally (Read et al. 2015a) to detect predators withoutmeat lures (hereafter termed the ‘focal dataset’), weexpressed cat and dog detections in two ways. First,we calculated a continuous response variable – detec-tions per 100 trap nights – that accounted for variablesample effort. Second, we converted detections into acategorical variable (yes/no). This was valid becausethere was no significant relationship between thesecategorical expressions and sample effort (cats:F1,44 = 0.38, p = 0.541; dogs: F1,44 = 1.06, p = 0.31)and therefore variable sample effort could be ignored.To examine the relationship between cat and dogdetections, we used linear regression for continuousdata and contingency tables with chi-square tests forcategorical data. To examine the effect of habitat on catand dog detections, the focal dataset was attributedaccording to whether they occurred in open or woodedhabitats, based on habitat descriptions of the mostsuitable Regional Ecosystem (RE; Neldner et al. 2012)polygon at or nearby each point (there are one or moredescriptions, each assigned a percentage, for every REpolygon). Regional Ecosystems 5.3.21, 5.3.21c, 5.7.15,5.7.10x1, 5.9.3x3 and 5.9.5 were classed ‘open’, whileREs 5.3.10, 5.7.2, 5.7.3, 5.7.5, 5.7.6 and 5.9.2x1 wereclassed ‘wooded’ (Queensland Herbarium 2016). Weexamined the relationship between habitat type andcat detections using analysis of variance for continuousdetection data, and chi-square tests for categoricaldetection data.
Scat analysis
Predator scats were examined for evidence of mesopre-dator regulation and predation on Night Parrots. Nofox scats were found and only one cat scat collected, soanalyses were limited to dogs. Sixty dog scats wereanalysed, mostly collected along vehicle tracksthroughout the 56 000 ha study area between April2015 and October 2016. Dog scats were identifiedbased on size, smell, shape and colour (Triggs 1996).A single scat sample was defined as one or more faecalpellets that appeared to have been deposited together inone defecation event. Scat samples were placed intoindividual nylon bags, soaked in water for 24 h beforebeing washed through a domestic washing machine seton regular cycle. Samples were air-dried in the sun
prior to microscopic analysis. Undigested fragmentsof prey (hair, teeth, claws, feathers, scales, exoskele-tons) were initially examined under a dissecting micro-scope (20–40 × magnification) and compared withreference material from species known or expected tobe in the area. Prey were identified to the most specifictaxonomic level possible. Hair from each sample wasfurther examined through whole-mount and cross-sec-tion under compound microscope (100–400 × magni-fication) following the methods of Brunner and Triggs(2002). Scat analyses were conducted by R.P. but arepresentative hair sample of all mammal species iden-tified was confirmed by a second hair analyst (B. Triggsunpub. data).
Herbivores
Images from all cameras (irrespective of their position)were used to detect medium-large herbivores in thestudy area (Cattle (Bos taurus × indicus), horses, don-keys, goats, camels, pigs (included here as herbivoresalthough we acknowledge also their omnivoroushabits) and rabbits). The macropods CommonWallaroo (Macropus robustus) and Red Kangaroo (M.rufus) were collectively termed ‘macropod’. To obtain ameasure of their abundance, the relative detection ratesof large herbivores was calculated using the focal data-set as described above, expressed as detections per 100camera-trap nights.
Fire history
Given the role that altered fire regimes have played inchanging ecosystems in central Australia (Edwards et al.2008) and its recurring role in the decline of a diverserange of birds (Garnett et al. 2011), we examined the firehistory of the Pullen-Pullen–Mt Windsor site at a scalethat is relevant to Night Parrot biology (i.e. their use ofhabitat patches that can be less than 100 m wide;Murphy et al. 2017b). We used the automaticallymapped Landsat-based time series for Queensland for1988–2015 that has 30 m resolution (Goodwin andCollett 2014). This dataset is prone to both commissionand omission errors, which can vary in time and space(Goodwin and Collett 2014). To assess its reliability wemanually compared putative automatically derived firescars to Landsat TM, ETM+ and OLI images for1987–2016 for the scene that covers the Pullen-Pullen–Mt Windsor area (scene 097–076). These were processedas false colour images, where fire scars were highlightedat 30 m resolution using a combination of bands RGB:7/5, 4, 3 (TM and ETM+) or 7/6, 5, 4 (OLI) with pixelvalue stretching using ERDAS ER Mapper 2011
4 S. A. MURPHY ET AL.
Dow
nloa
ded
by [
116.
240.
73.1
16]
at 2
3:01
29
Oct
ober
201
7

(Hexagon Geospatial, GA, USA). False colour imageswere inspected manually for all putative fire scars withinthe study area. In addition, we selected 108 randompoints within putative fire scars for the entire Landsatscene and manually inspected our false colour imagelibrary for evidence of fires, scoring each point asfire = yes/no. We ignored commission errors that wereobviously related to Landsat 7 Scan Line Correctorinstrument failure, which were easily identified by theirstriped appearance. We extended the Landsat-based firehistory with archival air photographs from 1951 and1971 over the Mayne Range and that part of theNisbet Range that occurs on Pullen-Pullen. Digitisedimages were georeferenced using DigitalGlobe imagery(www.digitalglobe.com) and manually inspected for evi-dence of past fires.
Results
Mammalian predators
No foxes were detected in 19 108 camera-trap nights at205 locations. There were 51 and 111 cat and dogdetections, respectively. It was difficult to determineexactly how many individual cats were involved; sixcoat colours and patterns were recorded (light tabby,dark tabby, spotted tabby, medium tabby, ginger andpied). In one area of about 200 ha, five of the sixcolours were recorded. Given this diversity, and thedistances between some detections of cats with thesame coat colours (e.g. 11 km, 54 km) we suggest thatthe 51 encounters involved at least nine individuals,and probably more. Given the observed uniformity indog coat colours over a wide area (i.e. sandy with whitepoints) it was even more difficult to determine exactlyhow many dogs were involved in the 111 detections,although both singletons and packs of up to four dogswere recorded.
A total of 6206 camera-trap nights from 46 locationswere included in the focal dataset for detection ratecalculations and inferential analyses. Detection ratesfor cats and dogs are shown in Table 1. Variation incat detection was not explained by the presence of dogsfor either continuous or categorical expressions of thedata (F1,44 = 0.92, p = 0.34; X2
1 = 0.18, p = 0.67; n(cats) = 26; n(dogs) = 52). Similarly, variation in ourcontinuous expression of cat detection was notexplained by habitat type (open vs. wooded;F1,44 = 1.6, p = 0.2). In contrast, using the categoricalexpression of cat detections, cats were significantlymore likely to be detected in wooded habitats thanopen habitats (X2
1 = 4.7, p = 0.03).
Scat analysis
Analysis of 60 dog scats revealed a diet dominated bymacropods (82% of scats) including both RedKangaroos and Common Wallaroos. Size of claws orteeth in 16 samples indicated juveniles and adults wereeaten in similar proportions. Cat remains were foundin one dog scat (<2%), cattle hair in 15% of scats, butno sign of foxes was recorded. Rodents were infre-quently recorded (5% of samples). Evidence of birdsoccurred in three scats. Two could not be identified(but were not parrots), whereas one sample containedparrot feathers but the blue, green and yellow featherswere identified as an Australian Ringneck (Barnadiuszonarius). The presence of fly larvae in this sampleindicated the bird was consumed as carrion.
Herbivore detections
The medium-large herbivore assemblage in the studyarea, based on 19 108 camera-trap nights at 205 loca-tions, included macropods (Common Wallaroos andRed Kangaroos), cattle (managed as opposed to wild),camels (wild), horses (wild) and rabbits. No pigs, goatsor donkeys were detected, despite infrequent incidentalsightings of the former two. Detection rates are shownin Table 1. Only two rabbits were detected at locationsthat were not included in the focal dataset (hence donot appear in Table 1).
Fire history at Pullen-Pullen
We could find no evidence of any fire activity withinthe Pullen-Pullen–Mt Windsor study area in either theLandsat archive or air photograph sequence datingback to 1951. Beyond this area we did detect firescars in the remainder of the Landsat scene, but incor-rect assignment remained common; 78% of 108 pointswithin putative scars appeared to be commission errorsmainly associated with watercourses and ephemeralalluvial habitats that experience large change in ground
Table 1. Relative detection rates of mammalian predators andlarge herbivores (all based on 6206 camera-trap nights at 46locations, except for macropods that involved 4259 camera-trap nights at 26 locations).
SpeciesDetection rate
(events/100 nights)
Cats 0.42Dogs 0.84Macropods 4.60Cattle 0.73One-humped Camels 0.05Horses 0.05
EMU - AUSTRAL ORNITHOLOGY 5
Dow
nloa
ded
by [
116.
240.
73.1
16]
at 2
3:01
29
Oct
ober
201
7

cover over short periods. All correctly assigned scarsoccurred in the north-east of the scene, which is sup-ported by incidental field observations of fire scars inthis region (S. Murphy pers. obs.). We were unsureabout fire activity at three points, which we conserva-tively scored as being correct.
Discussion
With only one confirmed record in the late 20th cen-tury amidst a handful of unconfirmed sightings (Boleset al. 1994; S. Murphy unpub. data), many peoplethought Night Parrots were following the fate of somany Australian arid-zone mammals into extinction.Dramatically, and in keeping with the species’ myster-ious aura, Night Parrots were recently found alive insouth-western Queensland and at a second, widelyseparated, undisclosed location in Western Australia.An empirical assessment of actual or potential threatsoperating at each of these two populations, and theirsimilarities or differences, is the critical first step inimplementing practical conservation managementactions for Night Parrots. At the time of writing theWestern Australian population had only recently beendiscovered and research into this new population wasyet to commence. By contrast, Night Parrots in south-western Queensland have been studied intensively forabout 4 years and a reasonable understanding of thethreatening processes in that region is beginning toemerge. The analyses presented above suggest thatseveral of the main threats typically associated withhistorical and ongoing fauna attrition elsewhere incentral Australia are absent or occur only at low levelsat the south-western Queensland site.
The absence of large, single fires
Our analyses of both air photographs and Landsatimagery show that the large single-fire events thatcharacterise contemporary fire patterns elsewhere incentral Australia do not occur in south-westernQueensland (a pattern also presented by Kimber andFriedel 2015). Field observations show that ignitions dooccur (mostly by lightning; S. Murphy pers. obs.) butfire spread is limited by bare surfaces that are naturallydevoid of fuel. Given a run of wet years it is possiblethat some of these bare interstitial areas could supportcombustible vegetation (most likely annuals or short-lived perennials, especially among Triodia hummocksnear plateau margins), thus potentially fuelling largerfires (Nano et al. 2012). It is likely that that this sce-nario happens on much longer time scales than can becaptured by the 60+ year imagery archive that we
analysed. In some areas around the plateau margins,short burnt stumps of small trees (presumably Acaciashirleyi or A. aneura) provide evidence of previous fire.However, aggregations of these charcoaled stumpsoccur infrequently through the study area and do notprovide evidence of the very large fires that occur inTriodia-dominated systems elsewhere. From the par-rot’s perspective, this pattern of relatively small andvery infrequent fires has meant that Triodia cover hasbeen constantly available within a relatively small area,providing critical breeding (Murphy et al. 2017a),roosting (Murphy et al. 2017b) and feeding (S.Murphy unpub. data) habitat.
From a management perspective there is no justifi-cation for the application of an extensive prescribedburning programme of the kind implemented in manyother parts of central Australia (Edwards et al. 2008).Instead, we recommend that fuels loads be monitoredto ensure that currently isolated areas of Triodia do notbecome linked by vegetation following stock removaland/or significantly wet periods. Similarly, Buffel Grass(Cenchrus ciliaris) incursions should be removedbefore they establish in bare interstitial areas whichwould increase fire sizes and potentially act as wicksinto critical Night Parrot habitat. Buffel Grass currentlyoccurs as small, isolated outbreaks mostly along drai-nage lines but has the potential to invade large areasand significantly affect fire behaviour (Miller et al.2010). Observations on neighbouring DiamantinaNational Park suggest it has spread over the past dec-ade, and continues to do so (C. Mitchell pers. comm.).We recommend baseline mapping of Buffel Grass inthe study area so that spread can be monitored overtime, and encourage targeted control of Buffel in adja-cent regions and corridors leading to the Night Parrothabitat.
The absence of foxes
Despite our large survey effort, we failed to detect foxesat the site – a premise supported by the absence ofrecords within the ALA (accessed 9 January 2017) andfrom hundreds of hours of unpublished spotlightingdata amassed by bilby researchers in the same regionover the past two decades (P. McRae pers. comm.).Although there are no empirical data suggesting thatfoxes prey upon Night Parrots, in southern Australia,they are known to impact other ground-dwelling threa-tened birds such as Plains Wanderers (Llewellyn 1975)and Bush Stone-Curlews (Carter 2010). It seems likelythat the absence of foxes has contributed to the persis-tence of these species, Night Parrots and other
6 S. A. MURPHY ET AL.
Dow
nloa
ded
by [
116.
240.
73.1
16]
at 2
3:01
29
Oct
ober
201
7

threatened species such as Greater Bilbies and Kowaris(Dasyuroides byrnei) in this region.
The absence of foxes from this part of Queensland ispoorly understood. Distribution records (ALA) showthat foxes occur north, south, east and west of thestudy area thereby indicating that physiological con-straints related to climate alone do not explain theirabsence. While dogs are known to kill foxes (Mosebyet al. 2012), given that both species coexist elsewhere incentral Australia (R. Paltridge unpub. data) it seemsimprobable that foxes are being excluded from theregion solely because of pressure exerted by dogs,although we assume that predation efficacy of dingoeson foxes would be higher in this region than in habitatswith many subterranean fox refuges such as rabbitwarrens. Lack of preferred prey may also be a contri-buting factor to fox absence with very few rabbits,possibly due to extensive rocky substrates that preventburrowing (this study; ALA, accessed 17 April 2017)and normally low populations of small mammals(between irruptions), as demonstrated by the very lowfrequency of small mammals in the dog scats that weexamined. However, their catholic diet (Paltridge 2002)suggests that it is unlikely that foxes are absent solelydue to lack of food. Another untested hypothesis toexplain their absence is poisoning by naturally occur-ring toxic compounds that become systemic in the foodweb (Peacock et al. 2011; Read et al. 2015b). Given thatthe fox-free status of this region could be a majordriver explaining the persistence of Night Parrots andother threatened species, and is potentially useful toreplicate elsewhere for conservation management, weargue that it is an issue that warrants further research.
Cats in the presence of dogs
Low levels of cats and dogs were detected on camerasthroughout the study area. Camera trapping remains arelatively recent technique for surveying predators inAustralia and so there are few published and relevantdatasets that permit comparison of our detection rates.Two that are somewhat relevant both reported consid-erably higher detection rates of both predators (6.1 catsand 2.3 dogs per 100 camera-trap nights across 25cameras deployed continuously for 2 years in theSimpson Desert (Greenville et al. 2014), and 4 catsand 7.5 dogs per 100 camera-trap nights on 18 pastoralproperties across northern and central Australia; Brooket al. 2012). Confounding effects in these studies thatmake comparisons with our data questionable are thatGreenville et al. (2014) was conducted during a rodentirruption while Brook et al. (2012) included a moreproductive biome and used lures to attract predators,
rather than the passive technique we used to calculatedetection rates. A third unpublished study generatedmore comparative data (R. Paltridge unpub. data): 3years of continuous monitoring in Triodia grasslandsin the Tanami Desert revealed detection rates for catsthat were similar to our study area (0.7 detections per100 camera-trap nights). This work was conductedduring a period of average climatic conditions andincluded equal numbers of baited cameras set alongroads (1.1 cats/100 camera-trap nights), and unbaitedcameras set off roads at burrows of key prey species(0.3 cats/100 camera-trap nights). This suggests catdensities in our study area in south-westernQueensland may not be substantially different fromother parts of central Australia during typical seasonalconditions.
The presence of cat remains in one dog scat pro-vided some evidence of intraguild predation althoughthe extent to which this regulates cats is uncertain. Inmost parts of Australia cats form only a minor part ofdog diet; 0.63% reported by Allen et al. (2014).However, higher levels of cat remains in dog scatshave been found in other Triodia habitats (R.Paltridge unpub. data) and dogs are also known tokill cats and foxes without eating them (Moseby et al.2012). We found no relationship between dog and catpresence, but cats were more likely to be detected inwooded habitats possibly because trees provide effec-tive refuges from dog predation. Given that NightParrots preferentially use open habitats with a verylow density of trees and shrubs (Murphy et al.2017b), this spatial separation suggests there is reducedprobability that cats will encounter Night Parrots andthat this might be driven by the presence of dogs. Ourresults suggest that birds are not a common part of dogdiets in this area. Elsewhere, birds are eaten opportu-nistically by dogs, generally occurring in 2–20% of scats(Corbett 2001).
More research is required to examine predator–preyand predator–predator interactions, and habitat prefer-ences of cats during small mammal irruptions. Thismay help to determine trigger points for instigatingcat control (for example, as small mammals declineso as to mitigate prey switching), to inform the mostefficient placement of baits, find refuge areas to targetduring dry periods, and improve our understanding ofhow dogs might regulate cats. While cats are notor-iously difficult to control with traps, baits and shooting,dogs can be manipulated more easily through manage-ment decisions about control efforts, opening or clos-ing watering points and potentially the provision ofcarrion to help channel their activity to critical habitatareas. However, management tools that manipulate
EMU - AUSTRAL ORNITHOLOGY 7
Dow
nloa
ded
by [
116.
240.
73.1
16]
at 2
3:01
29
Oct
ober
201
7

dog activity by carrion provisioning must first examinethe likelihood of perverse outcomes (for example,increased dog predation, unintended increases in feralcats or increases in varanid populations near NightParrot breeding habitat).
Low levels of herbivory in a resilient system
Given the close proximity of the 2006 Night Parrotspecimen to our study area (McDougall et al. 2009), itseems likely that Night Parrots and cattle have coex-isted on Brighton Downs for at least 11 years andprobably much longer. Prior to its purchase by theBritton family in 2013, Brighton Downs had beenowned by the Australian Agricultural Company forover 60 years, during which time grazing pressurewas moderated by conservative stocking rates (P.Britton pers. comm.). Furthermore, herbivory bymacropods is likely to have been regulated to someextent by dogs, which have escaped intensive controlmeasures partly because of the rugged terrain andpartly due to a long history of targeted, as opposed towholesale, dog control measures in the study area (W.Campbell pers. comm.).
GPS tracking data presented in Murphy et al.(2017b) identify spatial overlap between productivecattle grazing areas and key Night Parrot feedingareas. Furthermore, ongoing diet studies (S. Murphyunpub. data) show that Night Parrots rely on a suite ofplants including some grasses that are preferred bycattle such as Astrebla spp. and Uranthoecium trunca-tum. Previous studies have failed to demonstrate anegative effect of grazing on floristic diversity in suchhabitats (Phelps et al. 2007; Silcock and Fensham 2012)and this may explain why Night Parrots have been ableto coexist with cattle grazing. In addition, these com-mon feeding areas are somewhat buffered from theimpacts of localised droughts because they occuralong a mid-section of a hydrological catchment thatextends northwards into much higher rainfall zones(Geoscience Australia 2011; Bureau of Meteorology2015). Hence, reliable monsoon-driven distant rainfallevents trigger localised flooding and pulses of theresources that both cattle and parrots require.
Despite the evidence that cattle and Night Parrotscan coexist and that cattle grazing does not lead tofloristic change in these systems, it remains a possibilitythat grazing could impact phenology resulting in lessfood for Night Parrots. A similar situation was demon-strated in Golden-shouldered Parrots (Psephotus chry-sopterygius) where even moderate levels of grazing leadto reduced seed availability in the key food plantAlloteropsis semialata (Crowley and Garnett 2001).
Given this possibility, a stock fence was completedalong the eastern boundary of Pullen-Pullen in 2016.Recently collected data from ongoing research showthere is already a significant difference in total seedabundance in some habitats either side of the fence (J.Silcock, R. Fairfax, and N. Leseberg unpub. data). Anindependent review of the risks (to Night Parrots col-liding with the fence) and benefits of destockingPullen-Pullen concluded that, with appropriate modi-fications (i.e. plain-top wire and high visibility broadtape, cf. three-strand barbed wire), the benefits fromfencing outweighed risks (Smales 2016). Other internal,and now redundant, fences have been removed and thenew fence is being monitored for its effectiveness instock exclusion and bird collision (not limited to NightParrots).
Watch or act? Enhancing ecological resiliencethrough adaptive management
Our conceptual model that explains the persistence ofNight Parrots at the study area includes that:
● The naturally patchy distribution of vegetation(predominately Triodia) limits the size of wild-fires, which has resulted in the long-term stableavailability of Triodia that is critical for breeding,roosting, feeding and escaping predators.
● The absence of foxes reduces total predation pres-sure, which is potentially also suppressed by theregulation of cats by dogs. There is evidence thatNight Parrots and cats are spatially separatedbecause of their different habitats preferences,and it is possible that the presence of dogs main-tains this separation.
● The impacts of grazing by introduced and nativeherbivores on Night Parrot habitats have beenlimited historically through relatively low stockingrates, few artificial water points, long-term stabledog populations potentially regulating macropodnumbers and very low densities of feral herbi-vores. Grazing also occurs mainly in a subset ofNight Parrot habitats (alluvial plains and run-onareas) that are inherently resistant to herbivory-mediated floristic change and are somewhat buf-fered from the effects of localised drought due toephemeral inflows from distant, higher rainfallareas.
Opportunities exist for management to enhance theinherent ecological resilience that occurs in the studyarea, rather than simply to rely upon it for maintainingNight Parrot populations. We acknowledge that some
8 S. A. MURPHY ET AL.
Dow
nloa
ded
by [
116.
240.
73.1
16]
at 2
3:01
29
Oct
ober
201
7

management actions may involve risk (such as main-taining or manipulating the ranging behaviour of dogsand removing livestock) but that this risk can beunderstood and mitigated by following the principlesof adaptive management (Salafsky et al. 2002).Similarly, grooming traps that target feral cats withoutaffecting dogs or their ecological roles are also beingtrialled at Pullen-Pullen (Read et al. 2014). Meanwhile,the prevalence of more idiosyncratic threats, such asnest predation by native predators (Murphy et al.2017a), is currently being investigated and the threatof disturbance by uncontrolled (and potentially illegal)human activity is being managed by legal instruments,patrolling and education. As new ecological informa-tion about Night Parrots emerges, so too do opportu-nities that can increase the efficacy of management. Forexample, recent water budget modelling by Kearneyet al. (2016) suggests that Night Parrots need accessto water during summer (as opposed to relying ondietary water), which may expose them to increasedrisk of predation by cats at artificial water points. Thisrisk could be lowered by targeted cat control at waterpoints, especially leading into summer. Such ecologicalinsights like this, coupled with strategic managementactions, will form the basis of ongoing conservation ofNight Parrots in south-western Queensland andelsewhere.
Acknowledgements
We acknowledge the significant contribution that JohnYoung made through his initial discovery of NightParrots on Brighton Downs. We also acknowledge thesupport of Fortescue Metals Group (especially CathBozanich, Shaun Grein and Martin Heller), Peter andCarol Britton, Whatty Campbell, Greg and JustineClark, Bob McDonald, Bush Heritage Australia (especiallyRob Murphy) and members of the Night Parrot RecoveryTeam. We thank Pete McRae and Katherine Moseby forfruitful discussions about the ecology of threatening pro-cesses, and Barbara Triggs for analysing some of the hairsamples. Chris Pennay,Dale Richter and John Neldnerfrom the Queensland Herbarium generously provideddraft Regional Ecosystem mapping for the study area.J.S. thanks the National Environmental ScienceProgramme for supporting her contribution to thiswork. This research was conducted under ScientificPurposes Permit Number WISP14627114 and QLDDepartment of Agriculture and Fisheries Animal EthicsApproval number SA 2015/09/521.
Funding
This work was supported by Fortescue Metals Group.
References
Allen, B. L., Allen, L. R., and Leung, L. K. P. (2014).Interactions between two naturalised invasive predatorsin Australia: Are feral cats suppressed by dingoes?Biological Invasions 17:761–776. doi:10.1007/s10530-014-0767-1
Allen, B. L., Fleming, P. J., Allen, L. R., Engeman, R. M.,Ballard, G., and Leung, L. K.-P. (2013). As clear as mud: Acritical review of evidence for the ecological roles ofAustralian dingoes. Biological Conservation 159:158–174.doi:10.1016/j.biocon.2012.12.004
Allen, B. L., and Leung, L. K. (2012). Assessing predation riskto threatened fauna from their prevalence in predatorscats: Dingoes and rodents in arid Australia. PLoS One 7:e36426. doi:10.1371/journal.pone.0036426
Andrews, F. W. (1883). Notes on the Night Parrot.Transactions and Proceedings of the Royal Society ofSouth Australia 6:29–30.
Ashby, E. (1924). Notes on extinct or rare Australian birds,with suggestions as to some of the causes of their disap-pearance. Emu 23:178–183.
Asner, G. P., Elmore, A. J., Olander, L. P., Martin, R. E., andHarris, A. T. (2004). Grazing systems, ecosystemresponses, and global change. Annual Review ofEnvironment and Resources 29:261–299.
Baxter, C., and Paton, P. A. (1998). Further notes on thebirds of the Gawler Ranges. South Australian Ornithologist33:1–15.
Bennett, S. (1983). A review of the distribution, status andbiology of the Plains-wanderer Pedionomus torquatus,Gould. Emu 83:1–11.
Black, A. B. (2012). Collection localities of the Night ParrotPezoporus (Geopsittacus) occidentalis (Gould, 1861).Bulletin of the British Ornithologists Club 132:277–282.
Boles, W. E., Longmore, N. W., and Thompson, M. C.(1994). A recent specimen of the Night ParrotGeopsittacus occidentalis. Emu 94:37–40.
Brook, L. A., Johnson, C. N., Ritchie, E. G., andDickman, C. (2012). Effects of predator control onbehaviour of an apex predator and indirect conse-quences for mesopredator suppression. Journal ofApplied Ecology 49:1278–1286.
Brunner, H., and Triggs, B. (2002). Hair ID [electronicresource]: An interactive tool for identifying Australianmammalian hair
Bureau of Meteorology. (2015). ‘Mean Monthly, Seasonaland Annual Rainfall Data (Base Climatological DataSets).’ (Commonwealth of Australia: Canberra).
Burrows, N. D., Burbidge, A. A., Fuller, P. J., and Behn, G.(2006). Evidence of altered fire regimes in the WesternDesert region of Australia. Conservation Science WesternAustralia 5:14–26.
Carter, A. (2010). ‘Improving Red Fox (Vulpes Vulpes)Management for Bush Stone-Curlew (BurhinusGrallarius) Conservation in South-Eastern Australia.’(Charles Sturt University: Albury-Wodonga).
Commonwealth of Australia. (2015). ‘Threat Abatement Planfor Predation by Feral Cats.’ (Department of theEnvironment: Canberra).
Corbett, L. K. (2001). ‘The Dingo Is Australian and Asia.,’2nd edn. (J.B. Books: Marleston, South Australia).
EMU - AUSTRAL ORNITHOLOGY 9
Dow
nloa
ded
by [
116.
240.
73.1
16]
at 2
3:01
29
Oct
ober
201
7

Crowley, G., and Garnett, S. (2001). Growth, seed productionand effect of defoliation in an early flowering perennialgrass, Alloteropsis semialata (Poaceae), on Cape YorkPeninsula, Australia. Australian Journal of Botany49:735–743.
Cupitt, R., and Cupitt, S. (2008). Another recent specimen ofthe Night Parrot Pezoporus occidentalis from westernQueensland. Australian Field Ornithology 25:69.
Cupples, J. B., Crowther, M. S., Story, G., and Letnic, M.(2011). Dietary overlap and prey selectivity among sympa-tric carnivores: Could dingoes suppress foxes throughcompetition for prey? Journal of Mammalogy 92:590–600.
Department for Environment and Heritage. (2006). ‘GawlerRanges National Park Management Plan.’ (Adelaide:South Australia).
DEWHA. (2008). ‘Background Document for the ThreatAbatement Plan for Predation by the European Red Fox.Department of the Environment, Water.’ (Heritage andthe Arts: Canberra).
Dickman, C. R. (1996). Impact of exotic generalist predatorson the native fauna of Australia. Wildlife Biology 2:185–195.
Dooley, S. (2013). Out of the shadows. Australian Birdlife2:26–30.
Edwards, G. P., Allan, G. E., Brock, C., Duguid, A., Gabrys,K., and Vaarzon-Morel, P. (2008). Fire and its manage-ment in central Australia. The Rangeland Journal 30:109–121.
Edwards, G. P., Pople, A. R., Saalfeld, K., and Caley, P.(2004). Introduced mammals in Australian rangelands:Future threats and the role of monitoring programmesin management strategies. Austral Ecology 29:40–50.
Fensham, R., and Fairfax, R. (2008). Water-remoteness forgrazing relief in Australian arid-lands. BiologicalConservation 141:1447–1460.
Garnett, S. T., Szabo, J. K., and Dutson, G. (2011). ‘TheAction Plan for Australian Birds 2010.’ (CSIRO:Collingwood, VC).
Geoscience Australia. (2011). ‘National CatchmentBoundaries V.1.1.4.’ (Commonwealth of Australia:Canberra).
Gill, N. (2005). Life and death in Australian ‘heartlands’:Pastoralism, ecology and rethinking the outback. Journalof Rural Studies 21:39–53.
Goodwin, N. R., and Collett, L. J. (2014). Development of anautomated method for mapping fire history captured inLandsat TM and ETM+ time series across Queensland,Australia. Remote Sensing of Environment 148:206–221.
Greenville, A. C., Wardle, G. M., Tamayo, B., and Dickman,C. R. (2014). Bottom-up and top-down processes interactto modify intraguild interactions in resource-pulse envir-onments. Oecologia 175:1349–1358.
Hayward, M. W., and Marlow, N. (2014). Will dingoes reallyconserve wildlife and can our methods tell? Journal ofApplied Ecology 51:835–838.
Johnson, C. N. (2006). ‘Australia’s Mammal Extinctions: A50,000 Year History.’ (Cambridge University Press:Melbourne).
Johnson, C. N., and Isaac, J. L. (2009). Body mass andextinction risk in Australian marsupials: The ‘CriticalWeight Range’ revisited. Austral Ecology 34:35–40.
Kearney, M. R., Porter, W. P., and Murphy, S. A. (2016). Anestimate of the water budget for the endangered nightparrot of Australia under recent and future climates.Climate Change Responses 3:14–31.
Kennedy, M., Phillips, B. L., Legge, S., Murphy, S. A., andFaulkner, R. A. (2012). Do dingoes suppress the activity offeral cats in northern Australia? Austral Ecology 37:134–139.
Kerle, J., Foulkes, J., Kimber, R., and Papenfus, D. (1992).The decline of the brushtail possum, Trichosurus vulpecula(Kerr 1798), in arid Australia. The Rangeland Journal14:107–127.
Kimber, R. G., and Friedel, M. H. (2015). Challenging theconcept of Aboriginal mosaic fire practices in the LakeEyre Basin. The Rangeland Journal 37:623–630.
Kutt, A. S., and Woinarski, J. C. Z. (2007). The effects ofgrazing and fire on vegetation and the vertebrate assem-blage in a tropical savanna woodland in north-easternAustralia. Journal of Tropical Ecology 23:95.
Legge, S., Murphy, B., McGregor, H., Woinarski, J.,Augusteyn, J., Ballard, G., Baseler, M., Buckmaster, T.,Dickman, C., and Doherty, T. (2017). Enumerating acontinental-scale threat: How many feral cats are inAustralia? Biological Conservation 206:293–303.
Llewellyn, L. C. (1975). Recent observations of the Plains-wanderer with a review of its past and present status. Emu75:137–142.
McDougall, A., Porter, G., Mostert, M., Cupitt, R., Cupitt, S.,Joseph, L., Murphy, S., Janetzki, H., Gallagher, A., andBurbidge, A. (2009). Another piece in an Australianornithological puzzle – a second Night Parrot is founddead in Queensland. Emu 109:198–203.
McGregor, H., Legge, S., Jones, M. E., and Johnson, C. N.(2015). Feral cats are better killers in open habitats,revealed by animal-borne video. PLoS One 10:e0133915.
McKenzie, N. L., Burbidge, A. A., Baynes, A., Brereton, R. N.,Dickman, C. R., Gordon, G., Gibson, L. A., Menkhorst, P.W., Robinson, A. C., Williams, M. R., and Woinarski, J. C.Z. (2007). Analysis of factors implicated in the recentdecline of Australia’s mammal fauna. Journal ofBiogeography 34:597–611.
Miller, G., Friedel, M., Adam, P., and Chewings, V. (2010).Ecological impacts of buffel grass (Cenchrus ciliaris L.) inva-sion in central Australia – does field evidence support a fire-invasion feedback? The Rangeland Journal 32:353–365.
Morton, S. R. (1990). The impact of European settlement onthe vertebrate animals of arid Australia: A conceptualmodel. Proceedings of the Ecological Society of Australia16:201–213.
Morton, S. R., Stafford Smith, D. M., Dickman, C. R.,Dunkerley, D. L., Friedel, M. H., McAllister, R. R. J.,Reid, J. R. W., Roshier, D. A., Smith, M. A., Walsh, F. J.,Wardle, G. M., Watson, I. W., and Westoby, M. (2011). Afresh framework for the ecology of arid Australia. Journalof Arid Environments 75:313–329.
Moseby, K. E., Neilly, H., Read, J. L., and Crisp, H. A. (2012).Interactions between a top order predator and exoticmesopredators in the Australian Rangelands.International Journal of Ecology 2012:1–15.
Murphy, S. A., Austin, J. J., Murphy, R. K., Silcock, J., Joseph,L., Garnett, S. T., Leseberg, N. P., Watson, J. E. M., andBurbidge, A. H. (2017a). Observations on breeding Night
10 S. A. MURPHY ET AL.
Dow
nloa
ded
by [
116.
240.
73.1
16]
at 2
3:01
29
Oct
ober
201
7

Parrots (Pezoporus occidentalis) in western Queensland.Emu – Austral Ornithology 32:858–868.
Murphy, S. A., Silcock, J., Murphy, R., Reid, J., and Austin, J.J. (2017b). Movements and habitat use of the night parrotPezoporus occidentalis in south-western Queensland.Austral Ecology. doi:10.1111/aec.12508
Nano, C. E. M., Clarke, P. J., and Pavey, C. R. (2012). Fireregimes in arid hummock grasslands and Acacia shrub-lands. In ‘Flammabel Australia. Fire Regimes, Biodiversityand Ecosystems in a Changing World.’ (Eds. R. A.Bradstock, A. M. Gill, and R. J. Williams). (CSIROPublishing: Collingwood, VC).
Neldner, V. J., Wilson, B. A., Thompson, E. J., andDillewaard, H. A. (2012). Methodology for survey andmapping of regional ecosystems and vegetation commu-nities in Queensland. Version 3.2. Updated August 2012.,Brisbane.
Newsome, A. E., Catling, P. C., Cooke, B. D., and Smyth, R.(2001). Two ecological universes separated by the dingobarrier fence in semi-arid Australia: Interactions betweenlandscapes, herbivory and carnivory, with and withoutdingoes. Rangeland Journal 23:71–98.
Newsome, T. M., Ballard, G.-A., Crowther, M. S., Fleming, P.J. S., and Dickman, C. R. (2014). Dietary niche overlap offree-roaming dingoes and domestic dogs: The role ofhuman-provided food. Journal of Mammalogy 95:392–403.
Nimmo, D. G., Watson, S. J., Forsyth, D. M., Bradshaw, C. J.A., and Frair, J. (2015). FORUM: Dingoes can help con-serve wildlife and our methods can tell. Journal of AppliedEcology 52:281–285.
Palmer, R. A. (1995). Diet of the red fox (Vulpes vulpes) insouth-western Queensland. The Rangeland Journal 17:99.
Paltridge, R. (2002). The diets of cats, foxes and dingoes inrelation to prey availability in the Tanami Desert,Northern Territory. Wildlife Research 29:389–403.
Peacock, D., Christensen, P., and Williams, B. (2011).Historical accounts of toxicity to introduced carnivoresconsuming bronzewing pigeons (Phaps chalcoptera andP. elegans) and other vertebrate fauna in south-westWestern Australia. Australian Zoologist 35:826–842.
Pedler, R. D., Brandle, R., Read, J. L., Southgate, R., Bird, P.,and Moseby, K. E. (2016). Rabbit biocontrol and land-scape-scale recovery of threatened desert mammals.Conservation Biology 30:774–782.
Phelps, D., Lynes, B. C., Connelly, P. T., Horrocks, D. J.,Fraser, G. W., and Jeffery, M. R. (2007). ‘SustainableGrazing in the Channel Country Floodplains (Phase 2).’(Meat & Livestock Australia: North Sydney).
Pople, A. R., and Grigg, G. C. (1999). ‘CommercialHarvesting of Kangaroos in Australia.’ (EnvironmentAustralia: Canberra).
Pople, T., and Froese, J. (2012). Distribution, abundance andharvesting of feral goats in the Australian rangelands1984–2011. Final Report to ACRIS Management Committee.Queensland Department of Employment, EconomicDevelopment and Innovation, Brisbane, Australia.
Queensland Herbarium. (2016). Regional ecosystem descrip-tion database (REDD). Version 10.0 (December 2016).
Queensland Department of Science, InformationTechnology and Innovation. Available at https://www.qld.gov.au/environment/plants-animals/plants/ecosystems/descriptions Verified February 2017.
Read, J., Gigliotti, F., Darby, S., and Lapidge, S. (2014). Dyingto be clean: Pen trials of novel cat and fox control devices.International Journal of Pest Management 60:166–172.
Read, J. L., Bengsen, A. J., Meek, P. D., and Moseby, K. E.(2015a). How to snap your cat: Optimum lures and theirplacement for attracting mammalian predators in aridAustralia. Wildlife Research 42:1.
Read, J. L., Peacock, D., Wayne, A. F., and Moseby, K. E.(2015b). Toxic Trojans: Can feral cat predation be mitigatedby making their prey poisonous? Wildlife Research 42:689.
Reid, J., and Fleming, M. (1992). The conservation status ofbirds in arid Australia. Rangeland Journal 14:65–91.
Saalfeld, W. K., and Edwards, G. P. (2010). Distribution andabundance of the feral camel (Camelus dromedarius) inAustralia. Rangeland Journal 32:1–9.
Salafsky, N., Margoluis, R., Redford, K. H., and Robinson, J.G. (2002). Improving the practice of conservation: A con-ceptual framework and research agenda for conservationscience. Conservation Biology 16:1469–1479.
Scanlan, J. C., Berman, D. M., and Grant, W. E. (2006).Population dynamics of the European rabbit(Oryctolagus cuniculus) in north eastern Australia:Simulated responses to control. Ecological Modelling196:221–236.
Silcock, J. L., and Fensham, R. J. (2012). Arid vegetation indisequilibrium with livestock grazing: Evidence from long-term exclosures. Austral Ecology 38:57–65.
Silcock, J. L., Piddocke, T. P., and Fensham, R. J. (2013).Illuminating the dawn of pastoralism: Evaluating therecord of European explorers to inform landscape change.Biological Conservation 159:321–331.
Skroblin, A., and Murphy, S. A. (2013). The conservationstatus of Australian malurids and their value as modelsin understanding land-management issues. Emu113:309.
Smales, I. (2016). A consideration of the risks to Night Parrotsof collisions with stock fences. Report for Bush HeritageAustralia. Project no. 22373. Melbourne, Australia.
Thackway, R., and Cresswell, I. D. (Eds.) (1995). An interimbiogeographic regionalisation for Australia: A frameworkfor establishing a national system of reserves, version 4.0.Australian Nature Conservation Agency: Canberra;Current version 6.1. Available at http://www.environment.gov.au/parks/nrs/science/bioregion-framework/ibra/index.html
Triggs, B. (1996). ‘Tracks, Scats, and Other Traces.’ (OxfordUniversity Press: Melbourne).
Whitlock, F. L. (1924). Journey to Central Australia in searchof the Night Parrot. Emu 23:248–281.
Wilson, H. (1937). Notes on the Night Parrot, with refer-ences to recent occurrences. Emu 37:79–87.
Woinarski, J., Burbidge, A., and Harrison, P. (2014). ‘TheAction Plan for Australian Mammals 2012.’ (CSIROPublishing: Melbourne).
EMU - AUSTRAL ORNITHOLOGY 11
Dow
nloa
ded
by [
116.
240.
73.1
16]
at 2
3:01
29
Oct
ober
201
7

Page 20 of 27
6. Human and social aspects
Garnett, S.T., Kleinschmidt, M., Jackson, M.V., Zander, K.K., and Murphy, S.A. (2016). Social
landscape of the night parrot in western Queensland, Australia. Pacific Conservation Biology 22,
360-366.

Social landscape of the night parrot inwestern Queensland, Australia
Stephen T. GarnettA,D, Mark KleinschmidtB, Micha V. JacksonA,Kerstin K. ZanderC and Stephen A. MurphyA
AResearch Institute for the Environment and Livelihoods, Charles Darwin University,
Casuarina, NT 0909, Australia.BAstrebla Services Pty Ltd, Suite One, 78 Smythe Drive, Highfields, Qld 4352, Australia.CThe Northern Institute, Charles Darwin University, Casuarina, NT 0909, Australia.DCorresponding author. Email: [email protected]
Abstract. The attitudes of the owners or managers of properties potentially supporting populations of night parrot(Pezoporus occidentalis) in western Queensland, Australia, were explored using interviews to understand whether they
would be sympathetic to the species’ conservation. Eighteen interviews were carried out by a former member of the localgrazing community and found a high level of support for conservation, especially if it did not unduly disrupt existinggrazing management practices and there was compensation in the event property management needed to change. This
included trying to limit burning and not overgrazing habitat in which the parrot might occur. It also included the cessationof wild dog baiting, which is conducted to reduce calf losses, although concern about wild dogs was deeply entrenched.While some graziers were indifferent, none were openly antagonistic to parrot conservation that might involve their
property. The results suggest that collaborative management with local graziers can contribute substantially toconservation of the night parrot in the region and any fears that graziers might be antagonistic to night parrot conservationare ill-founded.
Additional keywords: graziers, habitat management, threatened species.
Received 31 March 2016, accepted 30 May 2016, published online 1 August 2016
Introduction
The May 2013 discovery of a population of the night parrot(Pezoporus occidentalis) (Koch 2013), an arid-adapted parrot of
which no extant population had been located for over a century(Murphy 2013), suddenly made it both possible and urgent toidentify and mitigate threats to the new population (Pyke and
Ehrlich 2014). Formal investment from government prioritisedidentification of proximate biophysical threats such as cat pre-dation, fire or cattle grazing (Murphy 2014). However, while
biophysical threats to biodiversity are often analysed, almost allextinctions are a function of human agency (Szabo et al. 2012)and most threats are emergent properties of the human social
environment within which the biodiversity occurs (Ban et al.
2013). Thus effective management of threats requires not just anunderstanding of the biophysical drivers of population changebut also the social attitudes and behaviours of those who can
influence how those drivers operate (Manfredo 1989). Effectiveconservation management also needs to be cognisant of theinstitutional regimes, formal and informal, within which atti-
tudes and behaviours are expressed.The night parrot was discovered on land leased from the
Queensland Government for grazing by cattle or sheep. Parrots
were found in large long-unburnt patches of spinifex (Triodia
longiceps), a member of a distinctive Australian grass genus thatforms large spiny shrub-like hummocks that grow among lowstony hills. Concern was expressed privately to members of the
research team that some graziers with a potential for nightparrots to occur on their properties might not be sympatheticto their conservation and, indeed, might reduce the likelihood of
occurrence by burning potential habitat should they themselvesfeel threatened by efforts to conserve the species. There weresome grounds for nervousness from both graziers and those
wishing to conserve parrots. Within the memory of many lease-holders in the region, leases had been compulsorily resumed bythe State for the purposes of conservation (Fitzpatrick and others
vsCrown1992).On the other hand, in theUSA threatened speciesor their habitat have been destroyed soon after their discovery onprivate land to avoid intervention by the Federal Governmentusing the power of the Endangered Species Act to protect
threatened species (Adler 2008).There has also long been a recognition in Australia that
acquisition of habitat by the State is not necessarily the best
model for threatened species conservation and that off-parkconservation can play an important role (Cary andWebb 2001).Funding for protected area management is almost always less
than the amount required and tends to be spent on infrastructure
CSIRO PUBLISHING
Pacific Conservation Biology, 2016, 22, 360–366
http://dx.doi.org/10.1071/PC16014
Journal compilation � CSIRO 2016 www.publish.csiro.au/journals/pcb

for visitors ahead of biodiversity conservation (AustralianSenate 2007). Alternativemodels for off-park conservation vary
from acquisition and management by private individuals ororganisations for the express purpose of conservation (Kamalet al. 2015) through to agreements with existing land owners to
change their management if necessary to ensure conservation ofkey biodiversity assets.
This last option requires that landholders be sympathetic to
conservation and willing to change management if necessary,which usually (but not always) requires compensation to coverreduced production or opportunity costs (Moon and Cocklin2011). A recent study across Australia north of the area consi-
dered in this paper found that graziers would bewilling to acceptabout AU$11 per hectare per year to exclude stock fromsensitive habitat on a long-term basis, though this could be
reduced considerably if there was flexibility in the contract or ifexclusion was for only part of any year (Greiner 2014). Thewillingness of graziers to participate in such schemes was
strongly associated with underlying values, and could beenhanced with appropriate incentives (Greiner 2015).
However, changes in behaviour rarely happen in isolation.While theories of behavioural change such as the Reasoned
Action Approach (Fishbein and Ajzen 2011) suggest that inten-tion to act is influenced by beliefs of the individual and theircapacity to exercise control over a decision, these intentions are
also moderated by the normative opinions of others. Thereforeconservation actions of individual graziers need to be placed in aregional context to understand the types of pressure they might
be under to conform with wider community norms and whichmight constrain the actions of individuals.
To this end this paper describes a survey of the attitudes of
most of the owners and managers of pastoral and grazingproperties with suitable habitat in the region in which the nightparrot was discovered. The primary purpose of the survey was todevelop support mechanisms for the graziers to become involved
in threatened speciesmanagement.At the same timewewished toascertain which properties would entertain hosting additionalsurveys or research relating to night parrots as well as to deter-
mine who would consider changing their management to suit theparrot and, if so, in what ways. The sample size is relatively smallbut the area of land is very large so the attitudes of a small number
of people can affect a substantial proportion of Queensland, andmost of the potential habitat of the parrot in the region.
Methods
The owners and/or managers of the 44 properties that supportpotential habitat in the region where the night parrot was found
were contacted and invited to participate in interviews abouttheir attitude to the night parrot and approaches to landmanagement (see Table S1 available as supplementary material
to this paper). All were cattle properties covered in a mixof grassland, shrubland and low woodland, with most cattlegrazing occurring on plains dominated by annual grasses and
forbs, interspersed with perennials such as Mitchell grass(Astrebla spp.), away from areas dominated by spinifex. Theonly perennial water in the region was in waterholes along thelargest rivers, with grazing supported by provision of borewater.
The climate was cold at night in winter but otherwise warm to
hot, with temperatures commonly exceeding 408C in summer.Rainfall averaged below 250 mm a year, falling primarily from
December to March, but was highly variable with multiyeardroughts frequent.
In aiming to ascertain ways in which graziers could become
involved in threatened species conservation we sought theirviews on: conservation and means of effecting it (NationalParks, private conservation, stewardship); knowledge about
and interest in the night parrot; and, attitudes towards potentialmeans of managing the three most obvious short-term threats tothe night parrot’s persistence – overgrazing of the habitat bycattle, predation by feral cats and burning old spinifex. We also
explored the level of interest in receiving stewardship payments,andwhat for, and the potential of using the parrots themselves asan attraction for paying tourists.
While the original intention was to visit each target propertyin person, after introductory telephone calls it was obvious thatthis would be logistically impossible to do in a cost-effective
manner. Therefore it was decided to conduct the interviews bytelephone with answers recorded, via an electronic form, in aspreadsheet for subsequent analysis. Interviews were all carriedout by author MK, who was familiar to most interviewees. The
questionnaire was delivered in a conversational style withadditional elicited details recorded against each question.Answers to some questions were contingent on the answers to
other questions. Transcripts of the interview were then madeavailable for additional comment or correction by the graziers –16 emailed, one mailed. One responded with some refinements
to their answers and another responded with confirmation thatthey were happy with the accuracy of the record; from thebalance, there was no response.
Two forms of analysis were conducted. First, descriptivestatistics were tabulated to summarise knowledge and attitudes.Inferential statistical analysis was not undertaken because thoseinterviewed had responsibility for almost all properties in the
region that could support the parrots. While the total number ofproperties is relatively small, and the number of people whocould be invited even smaller since there was often one owner
for several properties, the area encompassed by these propertieswas very large (a combined area of 45 000 km2), stretchingacross western Queensland from near Winton to the Northern
Territory border (Fig. 1). These data therefore represent theknowledge and attitudes of those responsible for a large area ofpotential night parrot habitat and no inference is made aboutthe owners or managers of those who did not participate in the
survey.Second, the textual data was examined for passages that
encapsulated the principal messages emerging from the inter-
views so that these messages could be conveyed in thelanguage of those who delivered them, providing context forthe descriptive statistics.
Results
Twenty-six people were invited to participate in the survey.Collectively, they had responsibility for the 44 properties thatcontained habitat that could have supported populations of nightparrot at some stage in their recent history based on knowledge
of their local ecology at the time (S. Murphy, pers. obs.).
Social landscape of the night parrot Pacific Conservation Biology 361

Of these, 23 agreed. The 181 who were ultimately interviewed
had responsibility for 41 properties (5 managers, 14 owners2).All the owners expressed awish to pass the property on to familyin the future. The individual graziers had amedian of 35 years of
experience in the pastoral industry (range: 0–65 years) with amedian of 19 years for each property (range: 1–65 years).
To give respondent answers a general context, all grazierswere asked to rank the following policy issues in order
of importance: Economic growth, Conservation, Healthcare,Education, Employment, and Immigration. Several graziersexpressed discomfort with the question, with one expressing
that it could ‘be taken out of context easily’. A numbercommented that the variables were linked and in reality hadequal importance. Nonetheless, the majority of respondents
ranked economic growth as the most important (Table 1). Educa-tion came through as the second most important whereas conser-vation, employment and healthcare were all ranked similarly. All
but one respondent ranked immigration as the least important.All but four interviewees felt that the responsibility for mana-
ging threatened species on private land is jointly the responsibilityof government and landowners, butmost also felt that government
should bear the financial costs (Table 2). One suggested that thecost should not be great: ‘if you don’t interfere with the threatenedspecies, shouldn’t cost you too much to look after them’ but none
knew of government management of threatened species outsideNational Parks andmost had little knowledge of threatened speciesmanagement on National Parks. Those who did expressed largely
negative sentiments, which was consistent with a widespreadview that management of National Parks was inadequate. Theywere commonly viewed as ‘havens for feral animals with poor fire
management’, particularly those rarely visited by tourists. Thegraziers saw the lack of management of National Parks as defeat-ing their purpose,withone feeling theywere simplyan approach to
‘appease Green voters’. One expressed the view that NationalParks have ‘good people, but not enough of them’ because theylack funds and suggested that National Parks ‘could be run in
partnership with landholders’. Private conservation groups wereseen as more acceptable, though some graziers thought that theprivate conservation groups needed to own the land theywished toconserve. Others were ‘happy with the collaborative stewardship
concept’. One was of the view that ‘we only lease country’ and‘don’t have the right to interfere with nature’.
1This figure includes a couple who answered questions jointly and are therefore counted as one entity.2One respondent owned one property but managed a second; so is counted as both an owner and a manager.
Table 1. Ranking of social issues by landholders
Ranked from 1 (highest) to 6 (lowest); n = 18
Policy issue Average rank No. ranking highest No. ranking fifth
Economic growth 1.9 11 1
Education 2.4 5 1
Conservation 3.3 1 5
Healthcare 3.4 0 6
Employment 3.9 0 5
Immigration 5.7 1 0
138�E 139�E 140�E 141�E 142�E 143�E
21�S
22�S
23�S
24�S
25�S0 100 Km
QueenslandNor
ther
n Te
rrito
ry
BOULIA
MIDDLETON WINTON
Fig. 1. Map of properties whose owners/managers agreed to be interviewed about their attitudes to night
parrot conservation.
362 Pacific Conservation Biology S. T. Garnett et al.

Some graziers (7) had known about night parrots for a longtime (1990 or earlier), mostly from hearing that people werelooking for them.Most (11) heard about the rediscovery of night
parrots by John Young (Koch 2013), sometimes through localmedia, particularly radio (5 people) when it was announced. Fora few (3) the survey itself was the first they knew about the
species. Nine people thought theywould recognise a night parrotif they encountered one, mostly because they were familiar withother species on their property. One woman thought she had
seen night parrots – two green parrots that had flown off theground out of spinifex. Everyone who knew of the bird was alsoaware of its association with spinifex.
While few people knew of the night parrot project, whichwas
just starting at the time of the interviews, all but three land-holders were completely accepting, in principle, of researchoccurring on their property (Table 3), though details of any
potential research were not discussed. One felt research orsurveys were ‘one of those sort of things come back to hauntthe landholder, when you cooperate you end up being themeat in
the sandwich’. Another was cautious at this stage because he felthe knew too little. Few expected anything in return exceptcommon sense (‘leave gates as you find them’) and ‘general
courtesy’. Just one wanted ‘compensation depending on what’sinvolved and the impact on our grazing enterprise’, but anotherasked ‘do you have to expect something?’. Most graziers wouldwant to keep secret any encounters with night parrot on their
property. Some wished to remain anonymous because theyfeared disturbance of their cattle, that they might be compulso-rily acquired by the State, or that there might be public liability
implications; others were concerned the birds themselves mightbe disturbed. Just one was keen to share the location so that itcould complement other conservation or research activities.
Most people knew at least a little bit about night parrothabitat, and all respondents who commented on night parrothabitat named spinifex as an important habitat. Most people
were at least willing to consider changing their cattle grazingpractices for the sake of the night parrot. A number commentedthat the spinifex country where the night parrots are thought
most likely to occur is rarely grazed by stock (Table 4). The tworespondents most reluctant to change practices were morewilling to do so with the potential of compensation. Fencing
to exclude cattle from night parrot habitat was also generallyacceptable, though one interviewee was concerned about stockperishing against the new fence (as stock can do when a newfence is erected) and two stated explicitly that they would like
such fences to be paid for and erected by others. People werereluctant to estimate an area they would be prepared to fence offfor conservation but onemanager of a property associated with a
Table 2. Landholder attitudes to threatened species management
Issue Attitude No. of responses
Responsibility for managing
threatened species on private
land
Government only 2
Landholder only 1
Joint 14
Other 1
No response 0
Responsibility for cost of
managing threatened species
on private land
Government only 12
Landholder only 0
Joint 5
Other 1
No response 0
Government management of
threatened species on
National Parks
Positive 1
Negative 7
Unknown 10
No response 0
Government management of
National Parks (overall)
Somewhat positive 4
Negative 12
Unknown 2
No response 0
Government management of
threatened species off-park
conservation programs
Positive 0
Negative 2
Unknown 16
No response 0
Role for private conservation
organisations
Yes 10
No 2
Maybe 2
No response 4
Table 3. Landholder attitudes to night parrot research
Issue Attitude No. of responses
Knowledge of night parrot project None 12
Very little 3
Some 2
No response 1
Willing to allow access for night
parrot research/surveys
Yes 15
No 1
Maybe 2
No response 0
Desire to remain anonymous and keep
location secret if night parrots found
Yes 13
No 1
Maybe 4
No response 0
Table 4. Landholder attitudes to Night Parrot habitat management
Issue Attitude No. of
responses
Knowledge of night parrot habitat Nothing 5
A little 4
Some 8
No response 1
Willingness to change stock
management to improve habitat of
an area of spinifex country if night
parrots were found
Yes 12
Maybe 4
No 2
No response 0
Willingness to change stock
management of spinifex country if
paid for projected lost production
Yes 7
Maybe 6
Not applicable/
no payment required
1
Not asked 4
Willingness to consider fencing to
exclude cattle from spinifex country
for night parrot conservation if paid
for projected lost production
Yes 5
Maybe 3
Not applicable/
no payment required
1
Not asked 9
Social landscape of the night parrot Pacific Conservation Biology 363

mine, so not used intensively for pastoralism, volunteered that
‘50 or 60% of land area on this property not practical from apastoral perspective,’ andwould ‘consider any option that mightcomplement conservation generally, subject to mine operation
expectations’. Another was happy to have fencing ‘as long as itdidn’t affect the productive Mitchell Grass country’.
There is potential for dingo management to contribute tonight parrot conservation through the regulation of feral cats
(Ritchie and Johnson 2009). Most graziers had a negative viewof dingoes (Table 5). While one person viewed them as a nativeanimal, for most they were unmitigated pests, killing calves and
causing losses they estimated at up to $75 000 a year in actual oropportunity costs. Thoughmost (10) did not know the cost, threeventured losses of ,AU$10 000 and four ventured losses of
.$10 000. Most graziers killed dingoes on sight or throughvarious control programs and were critical of neighbours if theydid not contribute to dingo control. Indeed, one was prepared to
have a low density of dingoes on his property because ‘theyhelped keep ‘roo numbers down a bit’ and only participated inthe annual shire wild dog baiting program to be a goodneighbour and support the wider effort. Despite these attitudes,
most of the graziers interviewed would be willing to reconsidertheir approach if night parrots were found on their property,especially if they were offered compensation. However, one
respondent described the idea as ‘fairyland – I don’t think it’srelevant as not many dogs will kill a cat’ while another was
worried about his popularity with neighbours.Firemanagement, like dingomanagement, is also likely to be
important for night parrot conservation.Within the study region,
large single fire events aremost common in the north, whereas inthe south, expansive areas of naturally low or no-fuel habitatsrestrict the spatial impact of fires (S. Murphy, unpubl. data).
Most of the graziers in the northern part of the study area use firein their management, commonly putting in fire breaks to preventthe spread of fire or using it to induce ‘green pick’ for cattle
(Table 6). Of those using fire, most would be sympathetic tochanging the way they manage it if it would benefit the parrots,even without compensation (Table 7). The few who said theywould not change their fire management did so only because
they experienced it so rarely on their properties.One potential source of income for propertieswith populations
of night parrots is tourism, particularly birdwatchers hoping to see
the birds. However, while most people were cautious abouttourism for fear of stock disturbance and public liability, otherswould at least tolerate birdwatchers if they ‘only came to see the
bird and leave’ (Table 8).Of the twowho had had experiencewithtourism, only one might see it as a potential income sourcedepending on its impact on liability. Neither of the two who
could envisage tourism as a potential revenue source for theproperty had had previous experience in the industry.
In addition to the formal survey, several of the graziersoffered additional information and opinions. One owner of three
properties had already declared substantial nature reserves ontwo, another had fenced off rock-wallaby habitat and created anartificial swamp especially for waterbirds and a third set of three
Table 5. Landholder attitudes to dingoes and their control
Issue Attitude or behaviour No. of responses
General attitude to dingoes Shoot on sight 13
Tolerate 3
Positive 2
No response 0
Dingo control methods usedA Baiting 11
Shooting 12
Trapping 3
Non-lethal 1
No response 0
Willingness to modify dog
baiting if night parrots were
found
Yes 6
Maybe 4
No 3
No response 5
Willingness to modify dog
baiting if paid for projected
stock losses or damage
Yes 6
Maybe 6
No response 6
AMore than one answer possible.
Table 6. Use of fire by landholders
Reason for burning No. who burn
for this reasonA
Improved grazing 6
Fire breaks 7
Fire fighting 14
Not applicable – do not burn 1
No response 0
AMore than one answer possible.
Table 7. Landholder attitudes to fire management
Issue Attitude No. of responses
Willingness to change fire
management if night parrots found
and require a different regime
Yes 9
No 4
No response 5
Willingness to change fire
management if paid for projected
losses in productivity
Yes 6
No 5
No response 7
Table 8. Landholder attitudes to tourism/birdwatching related to
night parrots
Issue Attitude No. of responses
Tolerant of birdwatchers on
property if night parrots found
Yes 6
Maybe 6
No 6
No response 0
Previous involvement in tourism Yes 2
No 16
No response 0
View tourism as a potential
revenue source
Yes 2
Maybe 3
No 13
No response 0
364 Pacific Conservation Biology S. T. Garnett et al.

properties also had nature refuges in less productive countrysuch as that containing spinifex. Eight of the graziers, represent-
ing 18 of the properties, expressed positive comments about thebirds such as ‘I am interested in the night parrot and conservationmore generally, please keepme informed’, ‘happy to help in any
way we can’ and ‘hope they find them’.
Discussion
The survey revealed a high level of support for conservation ofnight parrots in the region. There was genuine pleasure amongmany of those interviewed at the thought that the birds might
occur on their property, and certainly that they still existed. Eventhose owners who were not interested were willing to contem-plate changing management should they be compensated ade-
quately. While this group of landholders does not include all thesites where night parrots might occur in the region, and some ofthose who were unable to be contacted or unwilling to be
interviewed may be less favourably disposed, the overallimpression is that local support for conservation is high over alarge part of western Queensland. This is in line with surveysfurther north onwillingness to accept conservation payments for
conservation (Moon and Cocklin 2011; Greiner 2014) and thelong-standing attachment to place among graziers in ruralQueensland (Everingham et al. 2014).
The graziers had comments pertinent tomanagement of knownmajor threats to the night parrot’s persistence – overgrazing of thehabitat by cattle, predation by feral cats, burning old spinifex – as
well as on whether the parrots could contribute directly to theirincome through tourism.
Raising of beef cattle is the main form of income for all the
properties interviewed. Cattle, however, are mostly grazed onflat lands supporting Mitchell grass. Spinifex, the principalroosting and breeding habitat of the night parrots, supportsfew cattle and many of the properties made little use of country
supporting spinifex. Fencing to exclude cattle from spinifextherefore posed little problem for most graziers in the region,although several suggested that it may not be necessary because
cattle rarely use the spinifex anyway. The few who use spinifexas a source of food during times of drought were still prepared tofence out cattle if they were paid appropriate compensation.
Thus, while grazing has been a major source of biodiversity lossacross northern Australia (Garnett et al. 2010), there is potentialto reduce its impact on the night parrot on working cattleproperties in western Queensland because the birds are thought
to mainly occupy the least productive part of the landscape. It isimportant to note that should the parrots also be found to usenon-spinifex habitat in this region, as suggested by Garnett et al.
(1993), the attitudes to managing habitat for parrots may be lesssympathetic.
During the day the night parrots shelter in large hummocks of
spinifex that have not been burnt for many years. Analyses ofsatellite and aerial imagery for the areas currently occupied bythe parrot could not find evidence of fire since 1956 (S. Murphy,
unpubl. data.). Fires smaller than the pixel size detectable byimagery have certainly occurred (based on field observation),but rarely, and most of the area shows no fire scars. While fires inarid areas can cover very large areas after heavy rainfall (Allan
and Southgate 2002), empirical evidence from satellite mapping
suggests that fires in western Queensland do not follow thispattern (S. Murphy, unpubl. data), mostly because of the expan-
sive low- or no-fuel habitats that have already been mentioned.Fire is used sparingly in the study area as a management tool,
unlike in the savannas to the north. Even burning of fire breaks is
not practised universally, presumably because fires, particularlylarge fires, are infrequent. As with grazing, therefore, there wasgeneral agreement that fire management could be changed for
night parrots if thought necessary and that this would have littleeconomic impact on the graziers.
In comparison to grazing and fire, dingoes and wild dogselicited a very strong reaction. There are good data to suggest
that the presence of dingoes benefits many threatened species(Wallach and O’Neill 2008). In the tropical savannas, thepresence of dingoes in the landscape reduces the activity of
cats, and potentially their absolute numbers (Kennedy et al.
2012), though there has been vigorous debate about the statisticsused to support this research (Allen et al. 2015). There is also
research showing that killing of stable pairs of adult dingoesallows country to be occupied by larger numbers of non-territorial dogs (Thomson et al. 1992). The belief among mostgraziers, however, was that dingoes kill many cattle, particularly
calves, and cause substantial financial losses. Actual costsappear to vary greatly between properties, and do not closelycorrespond with their control strategies, but the issue generated
the most animated answers of the issues discussed. While manyproperties may be willing to accept compensation for cattlelosses from dingoes if that was thought the best way of protect-
ing night parrots, there is evidentlymuch greater need to providestrong evidence of benefit and the more cost-effective strategymay be ongoing control of feral cats by other means.
Tourism was initially discussed as a potential source ofincome rather than a threat. Bird tourism is a growing sector intheAustralian tourismmarket (Steven et al. 2015) and tourism forbiodiversity is commonly touted as a potential ecosystem service
for which payments can be received (Balmford et al. 2009).However,many graziers saw tourism as a potential threat to eitherthemselves or the parrots and there was little interest in using the
parrots as away of generating income from tourism.Birdwatchersmay be tolerated on some stations but only if they do not disruptthe existing cattle grazing enterprise. As it is, most birdwatchers
will want to see the target bird. However, even when researchershave known the exact location of roosts or nests, individual nightparrots were difficult to observe (S. Murphy, pers. comm.).Furthermore, there is some evidence to suggest that disturbance
at one roost area (associated with the recapture of a GPS taggedbird) caused the temporary relocation of three night parrots to analternative roost site approximately 7 km away for about four
weeks (S. Murphy, unpubl. data). Given that night parrots aredifficult to observe and are known to move away from areasfollowing disturbance, it seems that the species does not lend
itself readily to eco-tourism.
Conclusions
Currently the social environment of the night parrot in westernQueensland is largely supportive. The harsh environment inwhich the night parrot is currently known to persist, which has
low productivity for cattle, and the limited use of fire by graziers,
Social landscape of the night parrot Pacific Conservation Biology 365

suggests that conservation of the parrot will impose little eco-nomic cost to graziers. The potential for reducing dingo control
so that they can control cats is more contentious, but notunsurmountable. Nevertheless both social and economic costs ofchanging dingo control strategies, including effects on reputation
with neighbours if dingo control is stopped in night parrot habitatonly, would need to be carefully weighed against the benefits ofcat control, particularly if other means of cat control are likely to
be effective. Tourism is unlikely to be welcomed by graziers,which may not matter as the species’ secretive nature means itwill not readily be seen by visiting birdwatchers.
This research provides the basis for discussions with graziers
about a range of potential conservation arrangements for thespecies. Low confidence in State-funded National Parks toprovide sufficient resources to manage the habitat makes this
the least favoured option among those interviewed. Privatelyfunded conservation seemed more acceptable, with conditions,while there seemed to be many opportunities for working with
existing leaseholders to modify land management withoutnecessarily changing tenure. It is unlikely, however, that nightparrots will contribute greatly to the local economy throughtourism.
Acknowledgements
We thank the team at Desert Channels Queensland for their support for this
project, John Young for finding this night parrot population and the parti-
cipating landholders and managers for making themselves available for
interview. The research was conducted under Human Ethics permit from
Charles Darwin University. Funding for the social research was provided by
Desert Channels Foundation. Funding for the wider research project is
provided by Fortescue Metals Group.
References
Adler, J. H. (2008). Money or nothing: the adverse environmental con-
sequences of uncompensated land use control. Boston College Law
Review. Boston College. Law School 49, 301–366.
Allan, G. E., and Southgate, R. I. (2002). Fire regimes in the spinifex
landscapes of Australia. In ‘Flammable Australia: the Fire Regimes and
Biodiversity of a Continent’. (Eds R. A. Bradstock, J. E. Williams, and
A. M. Gill.) pp. 145–176. (Cambridge University Press: Cambridge.)
Allen, B. L., Allen, L. R., and Leung, L. K. P. (2015). Interactions between
two naturalised invasive predators in Australia: are feral cats suppressed
by dingoes? Biological Invasions 17, 761–776. doi:10.1007/S10530-
014-0767-1
Australian Senate (2007). Conserving Australia: Australia’s national parks,
conservation reserves and marine protected areas. Environment, Commu-
nications, Information Technology and the Arts Reference Committee of
the Australian Senate, Commonwealth of Australia, Canberra.
Balmford, A., Beresford, J., Green, J., Naidoo, R.,Walpole,M., andManica,
A. (2009). A global perspective on trends in nature-based tourism. PLoS
Biology 7(6), e1000144. doi:10.1371/JOURNAL.PBIO.1000144
Ban, N. C., Mills, M., Tam, J., Hicks, C. C., Klain, S., Stoeckl, N., Bottrill,
M. C., Levine, J., Pressey, R. L., Satterfield, T., and Chan, K. M. (2013).
A social–ecological approach to conservation planning: embedding
social considerations. Frontiers in Ecology and the Environment 11,
194–202. doi:10.1890/110205
Cary, J., and Webb, T. (2001). Landcare in Australia: community participa-
tion and land management. Journal of Soil and Water Conservation 56,
274–278.
Everingham, J. A., Collins, N., Rifkin, W., Rodriguez, D., Baumgartl, T.,
Cavaye, J., and Vink, S. (2014). How farmers, graziers, miners, and gas-
industry personnel see their potential for coexistence in rural Queensland.
SPE Economics & Management 6, 122–130. doi:10.2118/167016-PA
Fishbein, M., and Ajzen, I. (2011). ‘Predicting and Changing Behavior: the
Reasoned Action Approach.’ (Taylor & Francis: New York.)
Fitzpatrick and others vsCrown (1992).Claim for compensation – resumption
for national park purposes. Land Court A94-44. http://www.courts.qld.
gov.au/__data/assets/pdf_file/0008/273374/A92-44.pdf
Garnett, S. T., Crowley, G. M., Duncan, R., Baker, N., and Doherty, P.
(1993). Notes on live night parrot sightings in north-western Queens-
land. Emu 93, 292–297. doi:10.1071/MU9930292
Garnett, S. T., Woinarski, J. C. Z., Crowley, G. M., and Kutt, A. S. (2010).
Biodiversity conservation in Australian tropical rangelands. In ‘Can
Rangelands be Wildlands?: Wildlife and Livestock in Semi-arid Eco-
systems’. (Eds J. du Toit, R. Kock, and J. Deutsch.) pp. 191–234.
(Blackwell Publishing: Oxford.)
Greiner, R. (2014). Environmental duty of care: from ethical principle
towards a code of practice for the grazing industry in Queensland
(Australia). Journal of Agricultural & Environmental Ethics 27,
527–547. doi:10.1007/S10806-013-9475-6
Greiner, R. (2015). Motivations and attitudes influence farmers’ willingness
to participate in biodiversity conservation contracts. Agricultural Systems
137, 154–165. doi:10.1016/J.AGSY.2015.04.005
Kamal, S., Grodzinska-Jurczak, M., and Brown, G. (2015). Conservation on
private land: a review of global strategies with a proposed classification
system. Journal of Environmental Planning and Management 58,
576–597. doi:10.1080/09640568.2013.875463
Kennedy, M., Phillips, B. L., Legge, S., Murphy, S. A., and Faulkner, R. A.
(2012). Do dingoes suppress the activity of feral cats in northern
Australia? Austral Ecology 37, 134–139. doi:10.1111/J.1442-9993.
2011.02256.X
Koch, T. (2013). Tweets in the night, a flash of green: is this ourmost elusive
bird? The Australian, 29 June.
Manfredo, M. J. (1989). Human dimensions of wildlife management.
Wildlife Society Bulletin 17, 447–449.
Moon, K., and Cocklin, C. (2011). Participation in biodiversity conserva-
tion: motivations and barriers of Australian landholders. Journal of
Rural Studies 27, 331–342. doi:10.1016/J.JRURSTUD.2011.04.001
Murphy, S. (2013). Night manoeuvres. Australian Birdlife 2, 20–22.
Murphy, S. (2014). Night parrot (Pezoporus occidentalis) research plan.
Map IT and Fortescue Metals Group, Yungaburra, Queensland, and
Perth, Western Australia.
Pyke, G. H., and Ehrlich, P. R. (2014). Conservation and the Holy Grail: the
story of the night parrot. Pacific Conservation Biology 20, 221–226.
doi:10.1071/PC140221
Ritchie, E. G., and Johnson, C. N. (2009). Predator interactions, meso-
predator release and biodiversity conservation. Ecology Letters 12,
982–998. doi:10.1111/J.1461-0248.2009.01347.X
Steven, R.,Morrison, C., Arthur, J.M., and Castley, J. G. (2015). Avitourism
and Australian important bird and biodiversity areas. PLoS One 10,
e0144445. doi:10.1371/JOURNAL.PONE.0144445
Szabo, J. K., Khwaja, N., Garnett, S. T., and Butchart, S. H. M. (2012).
Global patterns and drivers of avian extinctions at the species and
subspecies level. PLoS One 7, e47080. doi:10.1371/JOURNAL.PONE.
0047080
Thomson, P. C., Rose,K., andKok,N. E. (1992). The behavioural ecology of
dingoes in north-western Australia. III. Hunting and feeding behaviour,
and diet. Wildlife Research 19, 565–584. doi:10.1071/WR9920565
Wallach, A. D., andO’Neill, A. J. (2008). Persistence of endangered species:
is the dingo the key? South Australia Department for Environment and
Heritage, Adelaide.
www.publish.csiro.au/journals/pcb
366 Pacific Conservation Biology S. T. Garnett et al.

Page 21 of 27
7. Diet and drinking
Several types of samples that potentially contained fragments of Night Parrot plant food were
collected during the course of the study. They were:
scats (droppings) collected from nests and roosts
scats collected from birds in the hand
crop contents collected from dead nestlings (see Section 9).
These were sent to Dr. Jeremy Austin as the Australian Centre for Ancient DNA at the University
of Adelaide. DNA was extracted from samples using ancient DNA techniques which include
specialised protocols for dealing with low quality, highly degraded and small amounts of DNA
that also has a high potential for being contaminated with other sources of DNA.
Two loci were amplified and sequenced: ITS (plastid) and matK (nuclear). For ITS, two primer
pairs were used; one that amplified a long section of the locus and the other a shorter section which
was expected to more successfully amplify shorter sections of template that were likely to be
present due to degradation. Sequences were first compared to published sequences using a BLAST
search on GenBank (see https://blast.ncbi.nlm.nih.gov/Blast.cgi?PAGE_TYPE=BlastSearch) and
this often produced a genus or a few genera that were the most likely matches. To help further
define the foods to species level, plant reference material was collected from the site or sampled
from the Queensland Herbarium and extracted and sequenced. This included Uranthoecium
truncatum, Trianthema triquetra, Brachyachne ciliaris and B. prostrata. Results are presented in
Table 6.
In addition to the work on diet, we also completed an analysis of water requirements by Night
Parrots. A copy of that paper is included below.
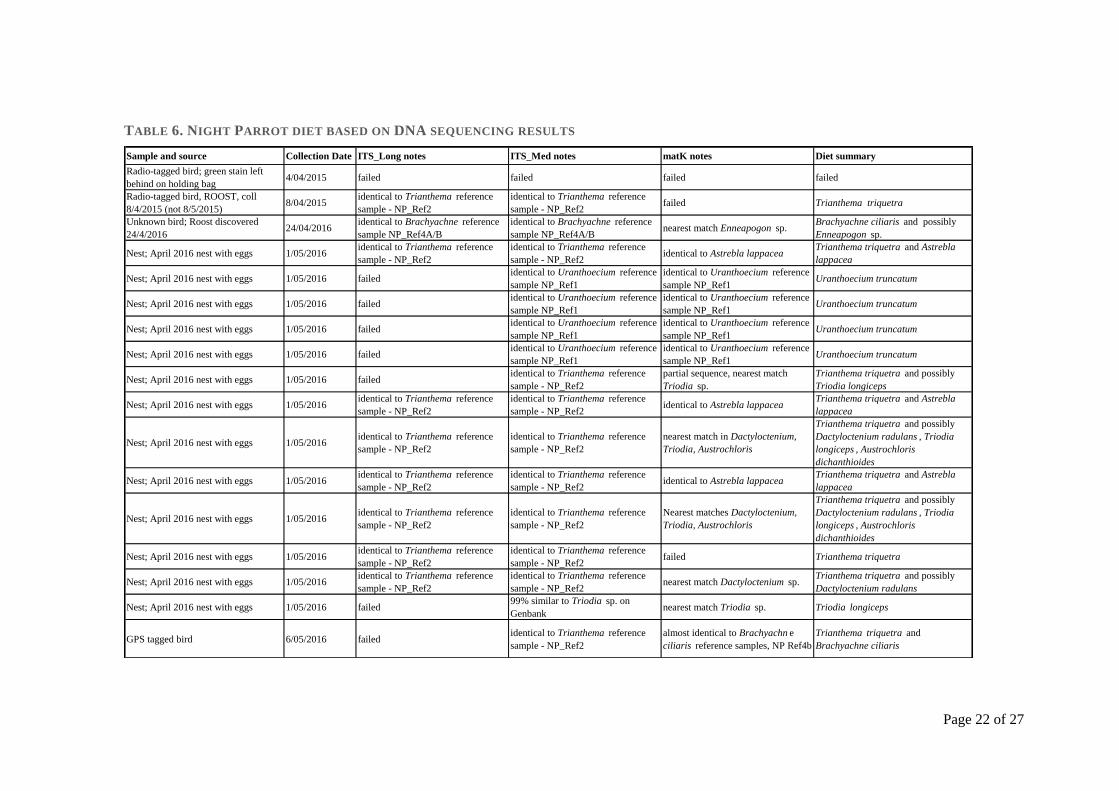
Page 22 of 27
TABLE 6. NIGHT PARROT DIET BASED ON DNA SEQUENCING RESULTS
Sample and source Collection Date ITS_Long notes ITS_Med notes matK notes Diet summary
Radio-tagged bird; green stain left
behind on holding bag 4/04/2015 failed failed failed failed
Radio-tagged bird, ROOST, coll
8/4/2015 (not 8/5/2015)8/04/2015
identical to Trianthema reference
sample - NP_Ref2
identical to Trianthema reference
sample - NP_Ref2failed Trianthema triquetra
Unknown bird; Roost discovered
24/4/201624/04/2016
identical to Brachyachne reference
sample NP_Ref4A/B
identical to Brachyachne reference
sample NP_Ref4A/Bnearest match Enneapogon sp.
Brachyachne ciliaris and possibly
Enneapogon sp.
Nest; April 2016 nest with eggs 1/05/2016identical to Trianthema reference
sample - NP_Ref2
identical to Trianthema reference
sample - NP_Ref2identical to Astrebla lappacea
Trianthema triquetra and Astrebla
lappacea
Nest; April 2016 nest with eggs 1/05/2016 failedidentical to Uranthoecium reference
sample NP_Ref1
identical to Uranthoecium reference
sample NP_Ref1Uranthoecium truncatum
Nest; April 2016 nest with eggs 1/05/2016 failedidentical to Uranthoecium reference
sample NP_Ref1
identical to Uranthoecium reference
sample NP_Ref1Uranthoecium truncatum
Nest; April 2016 nest with eggs 1/05/2016 failedidentical to Uranthoecium reference
sample NP_Ref1
identical to Uranthoecium reference
sample NP_Ref1Uranthoecium truncatum
Nest; April 2016 nest with eggs 1/05/2016 failedidentical to Uranthoecium reference
sample NP_Ref1
identical to Uranthoecium reference
sample NP_Ref1Uranthoecium truncatum
Nest; April 2016 nest with eggs 1/05/2016 failedidentical to Trianthema reference
sample - NP_Ref2
partial sequence, nearest match
Triodia sp.
Trianthema triquetra and possibly
Triodia longiceps
Nest; April 2016 nest with eggs 1/05/2016identical to Trianthema reference
sample - NP_Ref2
identical to Trianthema reference
sample - NP_Ref2identical to Astrebla lappacea
Trianthema triquetra and Astrebla
lappacea
Nest; April 2016 nest with eggs 1/05/2016identical to Trianthema reference
sample - NP_Ref2
identical to Trianthema reference
sample - NP_Ref2
nearest match in Dactyloctenium,
Triodia, Austrochloris
Trianthema triquetra and possibly
Dactyloctenium radulans , Triodia
longiceps , Austrochloris
dichanthioides
Nest; April 2016 nest with eggs 1/05/2016identical to Trianthema reference
sample - NP_Ref2
identical to Trianthema reference
sample - NP_Ref2identical to Astrebla lappacea
Trianthema triquetra and Astrebla
lappacea
Nest; April 2016 nest with eggs 1/05/2016identical to Trianthema reference
sample - NP_Ref2
identical to Trianthema reference
sample - NP_Ref2
Nearest matches Dactyloctenium,
Triodia, Austrochloris
Trianthema triquetra and possibly
Dactyloctenium radulans , Triodia
longiceps , Austrochloris
dichanthioides
Nest; April 2016 nest with eggs 1/05/2016identical to Trianthema reference
sample - NP_Ref2
identical to Trianthema reference
sample - NP_Ref2failed Trianthema triquetra
Nest; April 2016 nest with eggs 1/05/2016identical to Trianthema reference
sample - NP_Ref2
identical to Trianthema reference
sample - NP_Ref2nearest match Dactyloctenium sp.
Trianthema triquetra and possibly
Dactyloctenium radulans
Nest; April 2016 nest with eggs 1/05/2016 failed99% similar to Triodia sp. on
Genbanknearest match Triodia sp. Triodia longiceps
GPS tagged bird 6/05/2016 failedidentical to Trianthema reference
sample - NP_Ref2
almost identical to Brachyachn e
ciliaris reference samples, NP Ref4b
Trianthema triquetra and
Brachyachne ciliaris

Page 23 of 27
Kearney, M.R., Porter, W.P., and Murphy, S.A. (2016). An estimate of the water budget for the
endangered night parrot of Australia under recent and future climates. Climate Change Responses
3, 14-31.

RESEARCH Open Access
An estimate of the water budget for theendangered night parrot of Australia underrecent and future climatesMichael R. Kearney1*, Warren P. Porter2 and Stephen A. Murphy3,4
Abstract
Background: Endangered species management must now incorporate the potential effects of climate change, butthis is often in the context of limited data. The endangered night parrot was recently rediscovered in the Australianarid zone and a major effort is underway to ensure its survival. A key question is to what extent it is dependent onstanding water under current and future climates, as this has major implications for understanding and managingits habitat requirements. However, very little is known about its ecology and physiology, and its conservation statusprecludes invasive ecophysiological studies.
Method: Here we show how the methods of biophysical ecology permit strong inferences about this problemwith minimal data. We developed a biophysical model of both the parrot and its habitat at the site of itsrediscovery. We used allometrically-adjusted observations of the known physiology of a closely related desert-adapted Australian parrot, the budgerigar, to infer unknown aspects of the night parrot’s physiological responses,together with plumage measurements from museum specimens. We tested the microclimate model againstempirical data on microhabitat temperatures and compared the endotherm model predictions against an infra-redthermograph of the bird itself. We then used the model to predict the frequency with which the parrot wouldneed to find standing water under current and future climates depending on the water content of its food.
Results: Our field data show that air temperature in night parrot roosts during high summer typically exceeds theinferred resting core temperature (38 °C) and can exceed 45 °C. Our calculations imply that night parrots can persiston dry seed during winter conditions without exceeding dangerous levels of dehydration, but would need accessto water or succulent (55% water) food during summer. Air temperature at the site is projected to increase 3 °C by2070, which would lead to likely lethal (22% of body mass) levels of daily dehydration in some years even onsucculent food, and would dramatically increase its dependence on standing water.
Conclusion: Our findings have significant implications for the conservation management of the night parrot andprovide guidance for future research priorities.
Keywords: Biophysical model, Endangered species, Water budget, Heat stress, Arid zone, Endotherm, Australia
* Correspondence: [email protected] of BioSciences, The University of Melbourne, Parkville, VIC 3010,AustraliaFull list of author information is available at the end of the article
© The Author(s). 2016 Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, andreproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link tothe Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver(http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
Kearney et al. Climate Change Responses (2016) 3:14 DOI 10.1186/s40665-016-0027-y

BackgroundThe management of endangered species in the wild mustnow consider the potential influence of future climatechange on population persistence [1, 2]. This shouldideally be based on a mechanistic understanding of theconnections between species and climate, including basicecophysiological responses [3]. However, obtaining the re-quired data for developing such an understanding can beextremely difficult with rare and sensitive species. A casein point is the night parrot Pezoporus occidentalis, a spe-cies once widespread in the Australian arid zone but forwhich no confirmed live sightings had been made foraround 100 years until 2013, when a single populationwas discovered in western Queensland. A major effort isnow underway to conserve this population and to discoverand manage other populations. However, only about 8–10birds have been observed thus far at the site and researchactivities are necessarily highly limited.Birds and mammals have a range of adaptations to arid
environments, such as the production of highly concen-trated urine and dry faeces, but the physiological challengesof such places for endotherms may be unappreciated [4].Nocturnal birds such as the night parrot face especially pro-nounced challenges for two reasons. First, most birds donot use highly thermally buffered retreats such as burrowsand caves. Second, nocturnal birds must endure the entireday without water intake from food or drinking and arethus more susceptible to dehydration. The additional heatstress imposed by climate change is thus a serious issue fordesert birds such as the night parrot [5]. This is particularlyso in Australia where the magnitude of climate warming ispredicted to be greatest in the arid zone [6, 7] and wheremajor die-off events due to heat waves have been observedpreviously [5, 8]. McKechnie and Wolf [8] reviewed theissue of climate change impacts on Australian land birdsand outlined a conceptual framework for understandingand predicting the effects of climate change on birds in hotenvironments. They distinguished between the acute andchronic impacts of heat stress on bird survival and repro-ductive success. In the short term, intense heat waves maykill birds due to excessive dehydration from evaporativecooling responses, and hyperthermia when evaporativecooling responses fail. In the longer term, as climate warm-ing increases the frequency and duration of warm spells,birds may find it harder to balance their water budgets,which may reduce reproductive success or even cause deathfrom chronic dehydration. The ability of birds to deal withstressful climate warming is intimately linked to the avail-ability of buffering microclimates and water in their habitat.The ideal approach for understanding how species mightrespond to such changes, advocated by McKechnie andWolf [8], is to develop a mechanistic model of the interac-tions between the physiological tolerances, behaviouralresponses and available microclimates. This would allow
strong inferences to be made about how their survival,reproduction and habitat requirements will be altered byfuture climate change.Our aim in this paper is to develop such a model for the
night parrot, with two purposes in mind: 1) to makejudgements about the plight of this particular speciesunder recent and future climate change and how this mayinfluence its management, and 2) to illustrate a generalapproach that can be applied to tackle this kind of prob-lem for other species and habitats. To achieve this we em-ploy the methods of biophysical ecology, which involvethe application of physical principles of energy and masstransfer to organisms and their habitats to computefundamental constraints on a species’ behaviour, bodytemperature and food and water requirements [9–11].The approach involves solving heat budget equations fororganisms subject to the constraints of their particularmorphology, physiology and behaviour, as well as habitatconstraints on available microclimates. We apply themicroclimate and endotherm models of the Niche Mapperbiophysical modelling package (now available as an Rpackage, NicheMapR [12]) in conjunction with historicaldaily, 5 km resolution weather grids for Australia [13, 14].We use this system to compute the heat, energy and waterbudget of the night parrot at its current known location toask the following questions:
1) How well can we simulate microclimates available tothe night parrot in its habitat?
2) How much water must night parrots lose byevaporation to avoid overheating, and how has thischanged in historical times?
3) To what extent can night parrots survive at the sitewithout drinking water, and how is this influencedby dietary water?
4) How will these water requirements change underfuture climate change scenarios predicted for the site?
MethodsStudy species and siteThe night parrot is one of only two species of parrotworldwide that is fully nocturnal, with partial nocturnal-ity reported in only a handful of other species [15].Night parrots have long been associated with Triodiaspp. [16], an Australian endemic, perennial genus ofgrasses comprising more than 60 species colloquiallyknown as ‘spinifex’, most of which form very long lived,dense, spikey hummocks. There are also several crediblerecords of night parrots from chenopod (a salt-tolerant,arid-adapted shrub) dominated systems without Triodia,including the first specimen collected by explorer JohnMcDouall Stuart in 1845 [17]. Early observations, includ-ing those made by Aboriginal people [18, 19], museumcollectors [16] and graziers [20] suggest that night parrots
Kearney et al. Climate Change Responses (2016) 3:14 Page 2 of 17

use Triodia hummocks for daytime roosting and nesting.These behaviours have recently been confirmed during anintensive field study on the species in western Queensland(S. Murphy et al., unpubl. data) that is being implementedas part of an environmental offsets agreement [21]. Radio-and GPS-tag work on two individual night parrots (afemale in April 2015 and a male in April 2016, respect-ively) revealed that Night Parrots at least sometimes roostduring the day in Triodia hummocks and venture out intonon-Triodia habitats for most of the night to forage.These habitats include periodically inundated plains withstrongly self-mulching clay soils that support a high diver-sity of mostly ephemeral herbs and annual grasses, andironstone plains dotted with similarly floristically diverseephemeral plant communities growing in gilgai forma-tions (Murphy et al. in prep).Based on limited observations from specimens and a
captive live bird in the 1860s, we assume that the nightparrot is primarily a granivore [16, 19, 22]. Preliminarymolecular analyses based on scats (Murphy et al. in prep.)suggest that Night Parrots feed on a taxonomically diverserange of plants, including grasses (e.g. Triodia longiceps,Brachyachne spp.) and herbs (e.g. Trianthema triquetra).The closely related ground parrot feeds opportunisticallyon seeds of a wide range of species [23, 24], and prelimin-ary investigations of the night parrot at the Queenslandsite (S. Murphy in prep.) are consistent with this. How-ever, the night parrot has been observed to prefer greenfood in captivity [22] and there is circumstantial evidenceassociating it with a highly succulent desert plant genus,Sclerolaena spp. [25, 26].Due to concerns about potentially illegal collecting
activity, the exact location of the study area in westernQueensland is not provided here. However, the populationoccurs in the Goneaway Tableland subregion of the InterimBiogeographic Regionalisation of Australia (Version 7 [27]);the geographic centroid of this subregion is S 24.1096, E142.172. Key habitats include (1) Triodia longiceps on veryshallow rocky, loamy soils on the slopes and margins of dis-sected plateaus (2) Sclerolaena spp. dominant over otherchenopods on deep clay soils with a surface pavement ofironstone and silcrete, and (3) braided drainages dominatedby Acacia cambagei on deep alluvia.
Field observationsA single night parrot was captured using a mist-net atapproximately 1843 h on 6 May 2016, primarily for fittinga GPS tag to study movements and habitat use, and theseresults will be reported elsewhere. While in the hand werecorded the bird’s mass using a 300 g Pesola springbalance (Schindellegi, Switzerland) and measured surfaceskin temperature on the birds’ flank using a type-Kthermocouple (Digitech QM1601), just below the wing.We also took an infra-red thermograph with a FLIR T420
(FLIR Systems, Inc., Oregon, USA). During handling, wemeasured air temperature and relative humidity using ahygrochron iButton® (Maxim Integrated, San California,USA) suspended approximately 1.5 m above ground, andwind speed using a WindMate® WM-300 handheldanemometer (WeatherHawk, Utah USA) positioned ap-proximately 2 m above ground. Between December 2015and January 2016, six iButtons recorded temperature andrelative humidity in deep shade (n = 2), deep within largeTriodia hummocks (n = 2), in the open, adjacent to largehummocks (n = 1) and in a disused night parrot roost in arelatively smaller hummock that was discovered in April2015 (n = 1). The roost (Fig. 3e) was in an isolated Triodiahummock (size: 9.8 m x 5.3 m x 0.5 m H). The roost itselfwas a 25 cm long, horizontal tunnel, 8 cm in diameter (atentrance), constructed 8 cm above ground, with theentrance facing inwards towards the bare centre of thering-shaped hummock.
Museum measurementsWe measured plumage dimensions of three specimens ofthe night parrot (36,256, HLW54 and HLW55, collectedin the late 1800s) and three specimens of the budgerigar(B25905, B759 and B17320). Specifically, we measuredplumage depth at 20 locations from the top of the shoul-der to the base of the tail on the dorsal and ventral side(10 per side), as well as the length of three feathers fromthe dorsal side and three feathers from the ventral side(the latter was only done for two of the three night parrotspecimens). We also measured the body dimensions(length from beak to the point at which legs emerged fromthe feathers, width and depth at the shoulder) from whichto obtain a shape estimate.We measured plumage reflectance at three replicate
locations dorsally and ventrally of two of the night parrots(36,256, HLW54). Measurements were made with anOcean Optics dual-spectrometer system [Ocean Optics,Inc., USA; (Smith et al. 2016)] comprising two spectrome-ters (USB2000, 300–1000 nm and NIRQuest, 1000–2150 nm) with two light sources (PX-2 pulsed Xenon lightfor the UV-Visrange and HL-2000 tungsten halogen lightsfor the Vis-NIR range) connected with a quadrifurcatedfiber optic ending in a single probe (measurement area of5 mm x 3 mm oval). The probe was held in an OceanOptics RPH-1 probe holder (Ocean Optics, Inc., USA) ata constant angle (45°) and distance (approx. 1 cm) fromthe plumage skin and each measurement was expressedrelative to a Spectralon 99% white reflectance standard(Labsphere Inc., North Sutton, NH, USA). From reflectancespectra, we calculated solar reflectivity, which is a functionof reflectance and solar irradiance, from 300 – 2100 nmusing the ASTM G-173 standard irradiance spectrum fordry air derived from SMARTs v. 2.9.2 [28].
Kearney et al. Climate Change Responses (2016) 3:14 Page 3 of 17
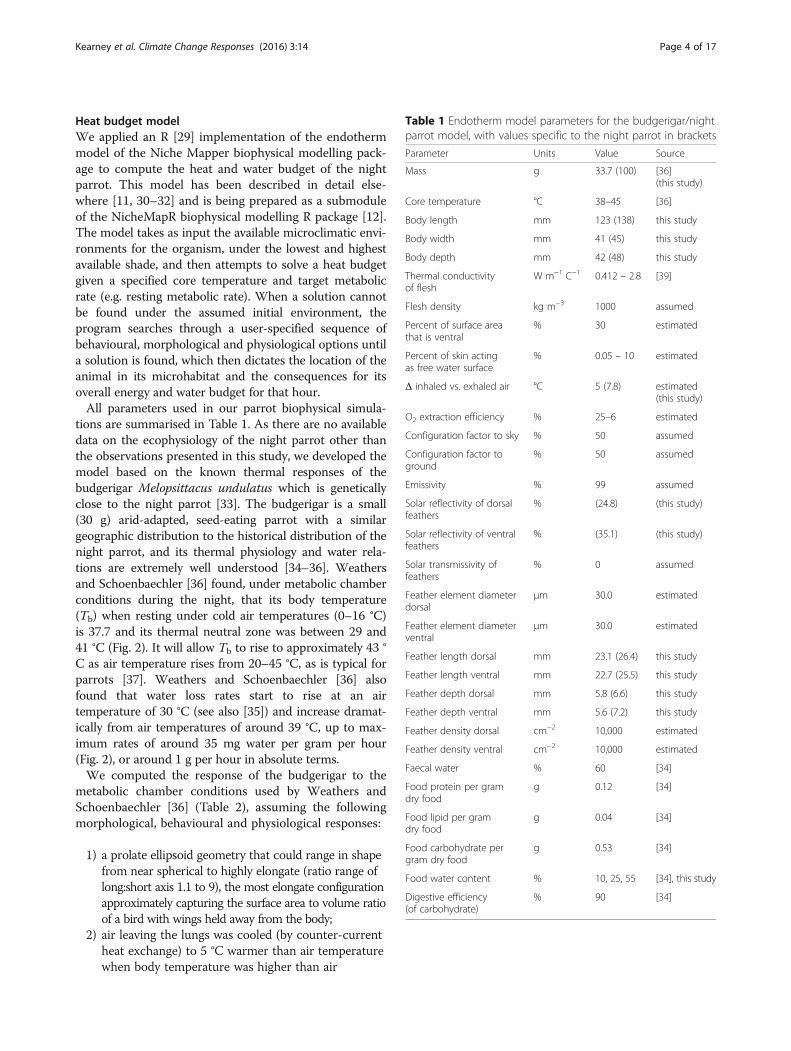
Heat budget modelWe applied an R [29] implementation of the endothermmodel of the Niche Mapper biophysical modelling pack-age to compute the heat and water budget of the nightparrot. This model has been described in detail else-where [11, 30–32] and is being prepared as a submoduleof the NicheMapR biophysical modelling R package [12].The model takes as input the available microclimatic envi-ronments for the organism, under the lowest and highestavailable shade, and then attempts to solve a heat budgetgiven a specified core temperature and target metabolicrate (e.g. resting metabolic rate). When a solution cannotbe found under the assumed initial environment, theprogram searches through a user-specified sequence ofbehavioural, morphological and physiological options untila solution is found, which then dictates the location of theanimal in its microhabitat and the consequences for itsoverall energy and water budget for that hour.All parameters used in our parrot biophysical simula-
tions are summarised in Table 1. As there are no availabledata on the ecophysiology of the night parrot other thanthe observations presented in this study, we developed themodel based on the known thermal responses of thebudgerigar Melopsittacus undulatus which is geneticallyclose to the night parrot [33]. The budgerigar is a small(30 g) arid-adapted, seed-eating parrot with a similargeographic distribution to the historical distribution of thenight parrot, and its thermal physiology and water rela-tions are extremely well understood [34–36]. Weathersand Schoenbaechler [36] found, under metabolic chamberconditions during the night, that its body temperature(Tb) when resting under cold air temperatures (0–16 °C)is 37.7 and its thermal neutral zone was between 29 and41 °C (Fig. 2). It will allow Tb to rise to approximately 43 °C as air temperature rises from 20–45 °C, as is typical forparrots [37]. Weathers and Schoenbaechler [36] alsofound that water loss rates start to rise at an airtemperature of 30 °C (see also [35]) and increase dramat-ically from air temperatures of around 39 °C, up to max-imum rates of around 35 mg water per gram per hour(Fig. 2), or around 1 g per hour in absolute terms.We computed the response of the budgerigar to the
metabolic chamber conditions used by Weathers andSchoenbaechler [36] (Table 2), assuming the followingmorphological, behavioural and physiological responses:
1) a prolate ellipsoid geometry that could range in shapefrom near spherical to highly elongate (ratio range oflong:short axis 1.1 to 9), the most elongate configurationapproximately capturing the surface area to volume ratioof a bird with wings held away from the body;
2) air leaving the lungs was cooled (by counter-currentheat exchange) to 5 °C warmer than air temperaturewhen body temperature was higher than air
Table 1 Endotherm model parameters for the budgerigar/nightparrot model, with values specific to the night parrot in brackets
Parameter Units Value Source
Mass g 33.7 (100) [36](this study)
Core temperature °C 38–45 [36]
Body length mm 123 (138) this study
Body width mm 41 (45) this study
Body depth mm 42 (48) this study
Thermal conductivityof flesh
W m−1 C−1 0.412 – 2.8 [39]
Flesh density kg m−3 1000 assumed
Percent of surface areathat is ventral
% 30 estimated
Percent of skin actingas free water surface
% 0.05 – 10 estimated
Δ inhaled vs. exhaled air °C 5 (7.8) estimated(this study)
O2 extraction efficiency % 25–6 estimated
Configuration factor to sky % 50 assumed
Configuration factor toground
% 50 assumed
Emissivity % 99 assumed
Solar reflectivity of dorsalfeathers
% (24.8) (this study)
Solar reflectivity of ventralfeathers
% (35.1) (this study)
Solar transmissivity offeathers
% 0 assumed
Feather element diameterdorsal
μm 30.0 estimated
Feather element diameterventral
μm 30.0 estimated
Feather length dorsal mm 23.1 (26.4) this study
Feather length ventral mm 22.7 (25.5) this study
Feather depth dorsal mm 5.8 (6.6) this study
Feather depth ventral mm 5.6 (7.2) this study
Feather density dorsal cm−2 10,000 estimated
Feather density ventral cm−2 10,000 estimated
Faecal water % 60 [34]
Food protein per gramdry food
g 0.12 [34]
Food lipid per gramdry food
g 0.04 [34]
Food carbohydrate pergram dry food
g 0.53 [34]
Food water content % 10, 25, 55 [34], this study
Digestive efficiency(of carbohydrate)
% 90 [34]
Kearney et al. Climate Change Responses (2016) 3:14 Page 4 of 17

temperature (and otherwise was at bodytemperature) [see 38];
3) skin thermal conductivity could vary from 0.412 to2.8 W m−1 °C−1 (typical range for animal tissue [39]);
4) core temperature could range from 38 °C to 43 °C;5) basal metabolic rate varies with mass according to
McKechnie and Wolf ’s [40] allometric equation,with a Q10 response of 2.5 to increased bodytemperature above 38 °C.
6) oxygen extraction efficiency could vary from 25%down to 6% under heat load to simulate observedmaximum level of pulmonary water loss;
7) cutaneous water loss could rise under heat load,with the percentage area of the skin area acting as afree water surface ranging from 0.05% to amaximum of 10% such that non-pulmonary waterloss rose to approximately 70% of total evaporativewater loss [see 41];
To scale this model to the night parrot, we allowed basalmetabolic rate to change as expected according to theabove-cited allometric equation from a 33 g bird to a 100 gbird, based on masses of two individual night parrots cap-tured in April 2015 and May 2016 (S. Murphy, unpubl.data), and changed the plumage characteristics based onmuseum specimen measurements, above. We also assumedthe difference between air temperature and exhaled air was7.8 °C to match the observed difference between air andcere temperature in Fig. 1. All other parameters were as-sumed to be identical to those of the budgerigar.The simulation started under the assumption of cold
conditions, with a spherical posture, low flesh thermalconductivity, base cutaneous water loss and high oxygenextraction efficiency. In field simulations, the parrots weresimulated to first seek shelter in Triodia grass hummocks(90% shade, 0.1 m s−1 wind speed, as described furtherbelow). In the field and metabolic chamber simulations,parrots under hot conditions were simulated to firstchange posture and flesh thermal conductivity to the ex-treme allowable values, then to allow Tb to rise to 41 °C,and finally to increase respiratory and cutaneous waterloss rates and core temperature up to the allowable limits;i.e. evaporative heat loss and extreme hyperthermia weresimulated as last resorts. Similarly, under cold conditions,elevated metabolic rates were only attempted when pos-tural and flesh conductivity options were exhausted.
Water budget modelAs with the heat budget model, we based the waterbudget model of the night parrot on that of the budgeri-gar. Water inputs include drinking water, preformed(dietary) water and metabolic water. The latter varieswith diet such that 0.4, 1.07 and 0.56 g of metabolicwater is produced per gram of protein, lipid and carbo-hydrate metabolised, respectively [42]. Following Cadeand Dybas [34] we assumed a diet with a dry mass com-position of 12% protein, 4% fats and 53% digestiblecarbohydrate. Water outputs include urinary, faecal andevaporative losses. Budgerigars fed on dry (10% water)seed and without drinking water produce faeces with60% water or less and no urinary water [34]. Dependingon the water vapour pressure deficit, budgerigars cansurvive indefinitely on dry seed without drinking waterat air temperatures between 14 and 20 °C and can go forlong periods with little mass loss at 30 °C [34, 35].We assumed maximum gross daily energy intake of
dry (10% water) seed scales with basal metabolic rate
Table 2 Environmental conditions for metabolic chambersimulations
Variable Units Value Source
Air temperature °C 0 – 45 [36]
Wind speed mm s−1 0.64 – 1.8 derived from [36]
Relative humidity % 15 – 20.8 [39]
Fig. 1 Long wavelength thermal infra-red (a) and associated visible (b)image of a 102 g night parrot 5 mins after capture at 18:48 on 6th May2016. Under the observed microclimatic conditions, assuming a corebody temperature of 42.0 °C, the biophysical model predicted a skintemperature of 40.2 °C and an outer feather temperature of 29.1 °C
Kearney et al. Climate Change Responses (2016) 3:14 Page 5 of 17

(BMR), and that digestible energy intake is roughly 4.6times BMR [43]. This produced an estimate of ≈ 10 gwet food per day for a 33 g budgerigar, which is consist-ent with a detailed study of their daily food intake incaptivity [44] (mean 6.92, range 1.24–15.47) but lowerthan that found by Cade and Dybas [34] (≈4 g). Thesame calculation produced an estimate of 21.8 g for a100 g night parrot. We explored different food watercontents, keeping the total mass of food constant, suchthat a parrot feeding on food with greater than 10%water content would obtain more preformed water, butless energy, per gram eaten.In simulations of parrots under natural conditions we
considered diets of seed with 10% water, but also succu-lent plant leaves with 25 and 55% water. The latter twofigures are based on pre- and post-oven dried masses oftwo samples of the chenopod Sclerolaena longicuspis.One sample (55%) was collected from a shallow run-onzone that supported succulent plants, while the otherwas from an area immediately adjacent to this. In April2015, a night parrot was observed in close proximity tothe run-on zone, leading us to speculate that night par-rots may derive substantial dietary water from this (andpotentially other) succulent plant species. Based on thesediets, we assessed how often parrots would exceed po-tentially lethal dehydration levels of 11% and 22% ofbody mass, following McKechnie and Wolf [5].
Microclimate model and climate change scenariosWe modelled microclimates using NicheMapR microcli-mate model [12]. This model, when applied in Australiausing interpolated daily weather data (air temperature,vapour pressure and solar radiation) and a continent-widesoil database as input, was recently shown to reliably predictactual soil temperatures (within 10% of observed values)across a wide range of environments [45]. We implementedthe model as reported in Kearney et al. [45] with theadditional incorporation of a daily gridded mean wind speedproduct [14], assuming maximum wind speed was double,and minimum wind speed was half, the mean value [45used long-term monthly mean values]. We estimatedair temperature, wind speed and relative humidity at4 cm above the ground either in zero or 90% shade, tocapture environments available to foraging and shelteringparrots, respectively, but assumed that parrots experiencedlow wind (0.1 m/s) when sheltering in Triodia tussocks.We simulated future climate by imposing projected
monthly changes in temperature, humidity, solar radiationand wind speed for 2070 onto interpolated daily weatherdata following the approach described in Briscoe et al. [32].Projections were obtained from six Global CirculationModels (GCMs), ACCESS 1.3, ACCESS 1.0, CanESM2,GDFL-CM3, HadGem2-CC and HadGem2-ES, which per-form well in capturing past climate in Australia [46].
ResultsMuseum specimen measurementsPlumage measurements are reported in Table 1, withdepth averaging 4.9 mm for the budgerigar and 6.9 mmfor the night parrot, and 20.1 and 26.0 mm, respectively,for feather lengths. The mean solar reflectivity of thenight parrot was 30%.
Heat and water budget modelThe endotherm model captured the response of the budg-erigar to increasing air temperature, showing close agree-ment with observed metabolic rate, body temperature andevaporative water loss (Fig. 2). Initially the maximumallowed ratio between the long and short axis of the birdwas set to 3, matching observed dimensions of budgerigarsat rest (Table 1), but this resulted in premature increase inevaporative water loss. It was found that a much closermatch to the observed data was possible if the surface areato volume ratio was allowed to increase further by allowingthe long to short axis ratio to extend to 9, which we pre-sume accounts for the birds holding their wings away fromtheir body. The model also predicted that budgerigarswould remain in positive water balance on a diet of dryseed only up to an air temperature of 14.5 °C, consistentwith Weathers and Schoenbaechler [36]. This threshold airtemperature would rise to 19.5 °C under a more typical in-door humidity level of 50% (a vapour pressure deficit basedon a humidity of 15% was assumed at this temperature inthe original simulation, Table 2).Converting the budgerigar model to the night parrot
(from 33 g to 100 g, increasing the plumage depth by theobserved amount, allometrically adjusting the basal meta-bolic rate) resulted in a reduction of the lower criticaltemperature from 27.5 to 24.5 °C, a shift in the onset ofthe evaporative cooling response from an air temperatureof 32.5 to 30.5 °C, and a reduction of the maximum mass-specific water loss rate by approximately 30% (Fig. 2). Themodel predictions were consistent with observed skin(40.2 °C) and outer feather temperature (mean ≈ 28 °C) ofa night parrot photographed with a long wavelength ther-mal infrared camera under known environmental condi-tions (Fig. 1) under the assumption of a body temperatureof 42 °C, rather than the assumed minimum value of 38 °C.The photo was taken 5 min after capture and the bird wasflying at the time of capture, all of which is consistent withthe inferred higher body temperature.
Microclimate observations and predictionsShaded air temperatures extracted for the study site fromthe continental 5 km weather grids were consistently 1.5 °Clower than both the shaded air temperatures measured withthe iButtons in trees (Fig. 2a,b) and the air temperatures re-corded by the on-site weather station, consistent with errorestimates of the weather grids for this region [13]. Thus, all
Kearney et al. Climate Change Responses (2016) 3:14 Page 6 of 17
predictions presented in this study are based on +1.5 °Cadjusted air temperatures. With this adjustment, predictedhourly air temperatures at 1.2 m showed good generalagreement with the observed values (Fig. 3a,b & Additionalfile 1: Figure S2). The main discrepancy with the 1.2 m airtemperature predictions was for daily minima to be
underestimated (Additional file 1: Figure S2), possiblyreflecting cold air drainage from the surrounding hills(mesas) [47]. Predictions of the two iButton shaded airtemperature measurements in trees (Fig. 3a,b) showedcorrelation coefficients (r) of 0.82 and 0.83, and root meansquared deviations (rmsd) of 3.7 and 4.1 °C, respectively.
Fig. 2 Observed a metabolic rate, b body temperature and c evaporative water loss rate of budgerigars in response to increasing temperature(adapted from Weathers and Schoenbaechler [36]) (red dots) compared to model predictions for the budgerigar (solid black line) and the nightparrot (dashed black line)
Kearney et al. Climate Change Responses (2016) 3:14 Page 7 of 17

Temperatures in the two large Triodia clumps (Fig. 2c,d)ranged from between 20 and 47 °C (Fig. 3c,d). This vari-ation was significantly underestimated when the microcli-mate model was run under the assumption of 90% shadeand 4 cm above the ground, but showed close agreementwhen assuming 70% shade at this same height (Fig. 3c,d),both showing correlations of 0.81, and rmsd of 4.1 and4.3 °C, respectively. In the night parrot simulations, how-ever, we conservatively assumed 90% shade as some dis-turbance of the Triodia necessarily occurs when insertingthe data loggers, and the parrots may be able to chooseparticularly dense locations and/or modify the leaves toform roosting structures that would increase shade.Temperatures in the autumn roost site (Fig. 2e) weremuch higher than those in the larger Triodia clumps(up to 55 °C) and were closely approximated by themicroclimate model predictions for air temperature at4 cm above the ground in full sun (r = 0.83, rmsd = 4.6 °C).Finally, the amplitude of fluctuations in the temperature
of a data logger in the open at 4 cm above the ground(Fig. 2f) was underestimated by the microclimate modelpredictions of air temperature in the open at the sameheight. However this logger was directly exposed to solarradiation and, while painted white to minimise solar influ-ences, its temperature was well approximated (Fig. 3f) bysimulating a 5 g cylindrical object absorbing 70% of thesolar radiation (r = 0.89, rmsd = 5.7 °C).Relative humidity (Additional file 2: Figure S3) and wind
speed (Additional file 1: Figure S2) were also adequatelycaptured by the microclimate model, although the modeltended to underestimate the maximum relative humidityand to overestimate the minimum wind speed.
Historical simulations under field conditionsSimulations of the year 2015 imply that air temperatures inlarge Triodia clumps providing 90% shade would regularlyexceed 38 °C through the warmer months of Novemberthrough to March, often approaching or slightly exceeding
Fig. 3 Locations of temperature data loggers in the habitat of the night parrot, measuring: a-b shaded air temperature at approximately 1.2 m intrees, c-d temperature 4 cm above the ground in centre of large Triodia tussocks potentially available to night parrots as roosts, e temperature intunnel of Autumn Triodia roost, f temperature near the ground representing the experienced temperature of a foraging night parrot
Kearney et al. Climate Change Responses (2016) 3:14 Page 8 of 17

45 °C (Fig. 5a). Over the same period, predicted ground sur-face temperature and air temperature in the open exceededthese conditions by 5 to 20 °C (Fig. 5a). Throughout thesame warm months, the biophysical model of the nightparrot predicted birds to be elevating core temperatureregularly, and that in the cool months the feathers wouldprovide strong insulation against cold conditions (gradientsfrom feather to skin temperature of around 30 °C, Fig. 5b).Deep (90%) shade would be required during the day toavoid heat stress on all but a few winter days, when shadelevels down to 60% were adequate (results not shown).Assuming a maximum intake of 21.8 g food per day,
night parrots would always be in positive energy balancewith over 200 kJ excess (discretionary) energy per daywhen feeding on seeds with 10% water, down to around50 kJ per day when feeding on food of the same energyvalue per gram of dry mass but with 55% water (Fig. 5c).Evaporative cooling was required at rates of over 1–2 g
per hour through the warmer months from Octoberthrough to early April (Fig. 5d). This put the birds intonegative daily water balance throughout this period if a dietof seed (10% water) was assumed, and to a lesser extent ifthe food was assumed to be 25% water, with a diet of 55%water causing negative water balances only occasionally(Fig. 5e). Parrots would regularly be in danger of dehydrat-ing by 11% of their original body mass each day on a diet ofseed only, but could substantially reduce this by feeding onfood containing 25% water, and eliminate it with foods con-taining 55% water (Fig 5e). They would be at close to neu-tral daily water balance from mid-April through to the endof August on a diet of seed, with substantial discretionarywater available (e.g. for reproduction) during these monthswhen feeding on moister food (Fig. 5e).Calculations of cumulative water balance, permitting
rehydration with rainfall events greater than 1 mm, sub-stantially exceeded 22% dehydration in the summer monthson a diet of dry seed (Fig. 5f). However, dehydration stayedalmost entirely below 22% on a diet of 25% water, and neverexceeded 11% on a diet of 55% water (Fig. 5f).Over the summers of 1996/7 to 2014/15, parrots eating
only dry seed would have regularly experienced daily de-hydration exceeding 11% for periods of 10 or more daysper year, with some exposure to this level of dehydrationon a diet of 25% water (Fig. 6a). However, daily dehydra-tion only exceeded 22% on a dry seed diet in five of the19 years. In contrast, cumulative dehydration exceeded11% and 22% in all years on a diet of dry seed (Fig. 6c,d).However, 22% dehydration was never exceeded on a dietof 55% water (Fig. 6b).
Climate change projectionsBoth maximum and minimum air temperatures are ex-pected to rise approximately 2.9 °C (range 2.4 to 3.3 °C)at the study site by 2070, with minor or inconsistent
change predicted in relative humidity, wind speed andsolar radiation (Table 3, Additional file 3: Figure S8,Additional file 4: FigureS9, Additional file 5: Figure S10,Additional file 6: Figure S11). While all six GCMs con-sistently predicted a small decline (mean 7%) in annualrainfall, four predicted an increase in summer rainfallwith an overall mean projected increase in summer rain-fall of 12% (Table 3, Additional file 7: Figure S12).Calculations of the water budget for 2015 under the
six climate change scenarios for 2070 produced similarestimates of water stress indices (Table 4). When werecalculated the water budget after imposing the ACCESS1.3 2070 scenario changes on the historical climate(Additional file 8: Figure S5), daily dehydration levelsexceeded 11% in almost all years irrespective of diet(Fig. 6e), and exceeded 22% on a diet of seed, and evenoccasionally on a diet of 55% water (Fig. 6f ). Similarly,cumulative dehydration exceeded the 11% thresholdevery year irrespective of diet (Fig. 6g) and exceeded22% every year on dry seed and 25% water diets(Fig. 6h). The 22% threshold was exceeded on 13 of the19 years even on the 55% water diet (Fig. 6h).
DiscussionInferring the ecophysiology of the night parrotThe amount of data required to develop a detailed mech-anistic model of a species’ climatic niche is not trivial [48,49]. Some endangered species are sufficiently well studiedin this regard, through captive populations and detailedfield studies (e.g. [50]). However, in the case of a data-poor, cryptic species like the night parrot, one must makeinferences for many parameters based on phylogeneticallyclosely-related and/or ecologically similar species.In this case we chose to develop a model to compute
the thermal and hydric responses of a related, arid-adapted species for which extensive data were available –the budgerigar - and then adjusted it to the specific knowndetails of the night parrot. In fact, the key parameters
Table 3 Climate change scenarios for the study site in 2070 forminimum (ΔTmin, °C) and maximum (ΔTmax °C) air temperature,relative humidity (ΔRH, %), wind speed (% Wind), solar radiation(% Solar), annual rainfall (% ARain) and summer rainfall (% SRain)
Scenario ΔTmax ΔTmin ΔRH % Wind % Solar % ARain % SRain
Access 1.3 3.2 2.9 −3.5 −1.4 0.2 −4.0 35.2
Access 1.0 2.8 3.0 −1.5 1.4 0.0 −0.4 18.7
CanESM2 3.3 2.9 −1.3 −2.5 0.5 −9.6 4.3
GDFLCM3 3.0 3.0 −4.5 3.5 1.7 −11.4 −4.6
HadGEM2-CC 2.6 2.7 −1.6 1.7 −0.7 −12.7 21.8
HadGEM2-ES 2.6 2.4 −1.3 −6.1 0.6 −3.2 −4.0
mean 2.9 2.8 −2.3 −0.6 0.4 −6.9 11.9
Air temperature and relative humidity are expressed as absolute change whilewind speed, solar radiation and rainfall are expressed as percent change
Kearney et al. Climate Change Responses (2016) 3:14 Page 9 of 17
needed to develop the budgerigar heat balance modelwere relatively few - the most important being its mass,shape, plumage depth, core body temperature range andbasal metabolic rate. The first three morphological traitsare easily measured, body temperature is highly phylogen-etically conserved in birds [37], and metabolic rate scalespredictably with body size [51] and body temperature[52]. The role of feathers in the heat transfer of birds iscomplex, especially with respect to the role of solar radi-ation [5]. However, the nocturnal habit of the night parrotrenders our predictions less sensitive to these complex-ities. Where having detailed data on thermal responses ismost important is in determining the particular responsesto heat and cold stress that the animal exhibits, and inwhat order. The detailed data on the physiological re-sponse of the budgerigar allowed us to infer this sequenceof responses (as described in the Methods section), whichis typical of desert-adapted birds [53] in the delayed onsetof evaporative cooling. Our final model showed closeagreement with the observed metabolic, body temperature
and evaporative water loss responses of the budgerigar be-tween air temperatures of 0 to 45 °C (Fig. 2).As expected, scaling the budgerigar up to the size
of a night parrot shifted the thermoneutral to lowertemperatures and resulted in reduced mass-specific evap-orative water loss rates (Fig. 2). The reduced mass-specificwater loss rate would make the night parrot more robustthan the budgerigar to desiccation under short-term expo-sures to high temperature [see 5]. Our calculations of thewater budget of the night parrot again relied on inferencesabout its water economy from the budgerigar, specificallyits faecal water content. Although the budgerigar exhibitsrelatively high levels of water conservation among birds[53], it is possible that the night parrot exhibits more ex-treme adaptations. For example, it has an unusually largecere [22] relative to other parrots, which may relate to itsability to save water by pre-cooling exhaled air [54].Conversely, they also have an unusually large eye (Fig. 1),presumably related to their nocturnal habit, which mayexacerbate water loss.
Table 4 Water stress indices under six different climate change scenarios for the year 2015
11% dehydration 22% dehydration
Scenario 10% H2O 25% H2O 55% H2O 10% H2O 25% H2O 55% H2O
a) Days exceeding 11 or 22% daily dehydration
Current 75 9 0 0 0 0
Access 1.3 141 71 13 20 3 0
Access 1.0 144 72 9 16 1 0
CanESM2 150 75 8 20 0 0
GDFLCM3 140 85 17 24 4 0
HadGEM2-CC 140 61 5 14 0 0
HadGEM2-ES 147 66 10 20 0 0
b) Number of events exceeding 11 or 22% cumulative dehydration
Current 15 14 0 14 3 0
Access 1.3 20 20 14 20 14 5
Access 1.0 16 16 9 15 15 3
CanESM2 16 12 11 14 16 3
GDFLCM3 14 15 14 15 16 7
HadGEM2-CC 13 16 10 15 13 1
HadGEM2-ES 15 15 10 13 17 4
c) Maximum run of days exceeding 11 or 22% cumulative dehydration
Current 38 12 0 16 2 0
Access 1.3 38 18 7 18 15 5
Access 1.0 39 18 7 20 10 3
CanESM2 39 25 10 25 12 2
GDFLCM3 45 24 10 24 17 5
HadGEM2-CC 39 18 9 18 11 1
HadGEM2-ES 39 28 10 28 12 5
Kearney et al. Climate Change Responses (2016) 3:14 Page 10 of 17

Microclimates available to the night parrotHalf the challenge of biophysical modelling is the accurateestimation of microclimates available to organisms. How-ever, recent developments in the availability of gridded cli-mate products for Australia [13, 14], and developments inmicroclimate modelling software [12], now make it pos-sible to infer microclimates from continent-scale datasetswith sufficient accuracy for biophysical calculations [45]. Inthe present case, the downscaling of the gridded daily wea-ther data to hourly microclimatic conditions was largelysuccessful, with good agreement between observed andpredicted hourly variation in air temperature, wind speedand relative humidity (Fig. 4, Additional file 9: Figure S1 &Additional file 1: Figure S2).Most importantly, the modelling system captured hourly
variation in the thermal environment inside potentialroosts of the night parrot to within ± 4 °C, or within 15%of the measured range (Fig. 4c,d). The main discrepanciesincluded a downward bias of 1.5 °C in the raw gridded airtemperatures (for which we corrected), and a tendency tounderestimate maximum humidity and minimum air tem-peratures at night time, potentially due to meso-scale cli-matic factors such as cold air drainage. These inaccuracieswould not influence our main conclusions since the mostimportant aspect of our calculations relate to heat stressduring the day when air temperature and relative humidityestimates were more accurate.
The water budget of the night parrotOur calculations of the water and energy budget of thenight parrot are likely to be conservative for two main rea-sons. First, we have assumed no additional heat or watercosts for activity, instead assuming metabolic rate will onlyvary from basal due to the effects of body temperatureand the need for metabolic heating in the cold. In relationto cold, the higher metabolic rates due to activity would,to some extent, cover the heating requirements of the par-rot (metabolic heat substitution [55]). Moreover, duringthe heat of the day in warm weather, many arid-adaptedbirds such as the budgerigar conserve water by remaininginactive [53], and thus would be near basal metabolic rateas we assumed. Nonetheless, tracking studies of the nightparrot (S. Murphy et al., in prep.) indicate that it can movetens of kilometres a night to feed and drink, and futurework will need to incorporate the associated costs into amore complete energy and water budget.Second, we assumed that night parrots were able to ob-
tain the allometrically-inferred maximum daily food intakerate of 21.8 g of food per 100 g bird. Food availability iscurrently unknown at the site and is likely to vary with thehighly erratic inter-annual rainfall that occurs here andthroughout the known range of the species.Despite our conservative assumptions about activity costs
and food availability, and our assumption that parrots can
shelter in 90% shade (measured temperatures in large Trio-dia clumps better matched model predictions for 70%shade, Fig. 3), simulated water costs for the night parrot atthe study site under historical conditions were very high(Fig. 5d-f & 6a-d). In our simulation of the year 2015, waterloss rates often exceeded 2 g per hour (Fig. 5d) and parrotswould be expected to regularly lose in excess of 11% oftheir body mass as water while inactive during the day insummer if feeding solely on seed (Fig. 5e). Such a scenariowould necessitate nightly drinking, because otherwise cu-mulative dehydration would exceed the likely lethal thresh-old of 22% multiple times through the year (Fig. 5f). Incontrast, with access to food of high water content (55%),our calculations imply that night parrots could have per-sisted without drinking. Extension of these calculations20 years into the past, however, shows that 2015 was a rela-tively mild year (Additional file 10: Figure S4); in someyears (summers of 2006/7 and 2013/14) daily dehydrationcould have exceeded 22% on a seed only diet (Fig. 6b) andcumulative dehydration could have exceeded 11% in someyears, even on a diet of 55% water content (Fig. 6c).One of the most effective behavioural options for ther-
mal buffering is to shelter below ground [56], an optionnot available to most desert birds [53]. There are anec-dotes of night parrots sheltering in rabbit burrows [18],caves [57] and even tunnels dug into sand [58]. To as-sess the potential significance of burrowing, we ran sim-ulations assuming that inactive night parrots shelter10 cm below ground in the shade (Additional file 11:Figure S6 & Additional file 12: Figure S7). This dramat-ically reduced water requirements, to the extent that theparrots could survive on seed-only diets without exceeding22% dehydration, and rarely exceeding 11% dehydration,considering both daily and cumulative water requirements(Fig. 6S). This is an aspect worthy of attention in futurefield studies of the species.
Climate change projections for the night parrot’s waterbudgetThe projected climate warming of approximately 3 °C by2070 at the study site had striking impacts on the ex-pected water budget (Fig. 6 and Additional file 10: FigureS4, Table 4). For example, this led to an increase in the fre-quency of daily dehydration events exceeding 11% from75 days per year to around 144 on a diet of seed, irrespect-ive of the climate change scenario (Table 4a). The propor-tional increase was even greater when food water contentwas increased to 25%, increasing from 9 to around 72 daysper year, and changed from no events to around 10 eventsper year under the 55% water diet. The mean airtemperature at the site in 2015 was 25.2 °C, thus an 11.5%increase in temperature imposed a 200-800% increase inthe frequency of stress events. The 11 and 22% dehydra-tion thresholds would even be exceeded under the
Kearney et al. Climate Change Responses (2016) 3:14 Page 11 of 17
assumption of a burrowing habit (Additional file 12:Figure S7). The reason for such non-linear and thresholdresponses to warming is easily seen in the sudden andsteep rise in evaporative water loss rate as environmentaltemperature approaches body temperature (Fig. 2c).There was an overall decline in projected rainfall across
all climate models which would act to reduce availablestanding water and food water content (Table 3). There
was a minor tendency towards increased summer rainfallat the site (Table 3, Additional file 7: Figure S12) whichmay partially offset the negative impacts of increased airtemperatures, but overall the greater evaporative power ofthe environment under a 3 °C warming will likely lead tolower water availability for the parrots.Our predictions indicate that climate change will impose
a substantial increase in the risk of heat wave induced die-
Fig. 4 Observed temperature in night parrot habitat (red lines) compared to microclimate model predictions driven by historical gridded dailyweather data (black lines). Logger locations are depicted in Fig. 3: a-b shaded air temperature at approximately 1.2 m in trees, c-d temperature 4cm above the ground in centre of large Triodia tussocks potentially available to night parrots as roosts, e temperature in tunnel of AutumnTriodia roost, f temperature near the ground representing the experienced temperature of a foraging night parrot
Kearney et al. Climate Change Responses (2016) 3:14 Page 12 of 17
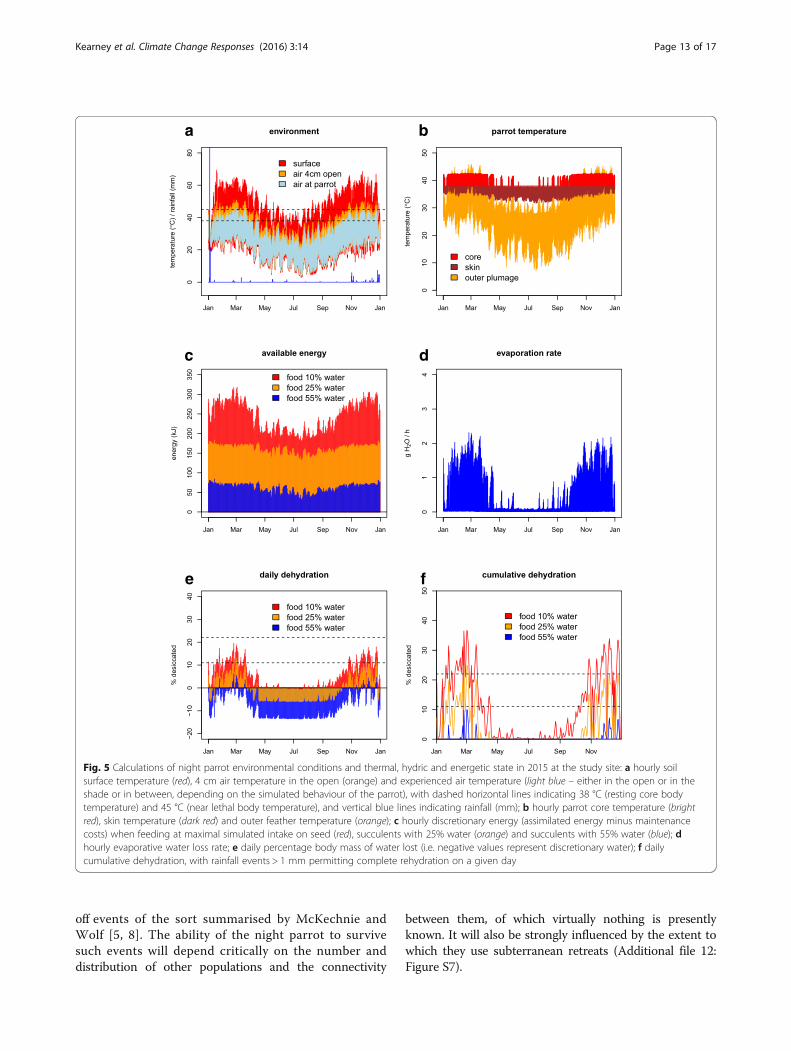
off events of the sort summarised by McKechnie andWolf [5, 8]. The ability of the night parrot to survivesuch events will depend critically on the number anddistribution of other populations and the connectivity
between them, of which virtually nothing is presentlyknown. It will also be strongly influenced by the extent towhich they use subterranean retreats (Additional file 12:Figure S7).
Fig. 5 Calculations of night parrot environmental conditions and thermal, hydric and energetic state in 2015 at the study site: a hourly soilsurface temperature (red), 4 cm air temperature in the open (orange) and experienced air temperature (light blue – either in the open or in theshade or in between, depending on the simulated behaviour of the parrot), with dashed horizontal lines indicating 38 °C (resting core bodytemperature) and 45 °C (near lethal body temperature), and vertical blue lines indicating rainfall (mm); b hourly parrot core temperature (brightred), skin temperature (dark red) and outer feather temperature (orange); c hourly discretionary energy (assimilated energy minus maintenancecosts) when feeding at maximal simulated intake on seed (red), succulents with 25% water (orange) and succulents with 55% water (blue); dhourly evaporative water loss rate; e daily percentage body mass of water lost (i.e. negative values represent discretionary water); f dailycumulative dehydration, with rainfall events > 1 mm permitting complete rehydration on a given day
Kearney et al. Climate Change Responses (2016) 3:14 Page 13 of 17
Extending the modelThe calculations presented here could be extended tocapture life history and phenological aspects of the nightparrot as influenced by climate. For example, repro-ductive potential could be computed [32, 59] based onestimates of egg energy and water content from mea-surements of egg dimensions and literature on typicalvalues for related birds [60]. General models of growthand reproduction such as the Dynamic Energy Budget
model [e.g. 49, 61] could also be developed based on lifehistory observations of the closely related ground parrotPezoporus wallacus. Such calculations could then be in-tegrated with estimates of water availability on thebasis of soil moisture and standing water availability[12, 50] to derive a more complete, whole life-cycleperspective on the impact of climate change and variabil-ity on this species of the sort envisioned by McKechnieand Wolf [8].
Fig. 6 The frequency that daily (a, b, e, f) or cumulative (c, d, g, h) dehydration of night parrots was predicted to exceed 11% (a, c, e, g) or 22%(b, d, f, h) over the summers 1996/7 to 2014/15 at the site under historical (a-d) and projected (e-h, 2070 ACCESS 1.3) climate
Kearney et al. Climate Change Responses (2016) 3:14 Page 14 of 17

ConclusionsConservation management of endotherms requires anunderstanding of how climate change may affect threat-ened and endangered species, especially in the contextof other pressures like habitat modification and feralpredators. Here we have illustrated how the methods ofbiophysical ecology can provide insight into the waterbudget of an endangered species under different climatescenarios. Our analysis predicts strong associations be-tween the night parrot and succulent vegetation in theabsence of standing water, points to a large advantage ofsubterranean retreats, and highlights heat-induced waterstress as a major issue for the future of this species.Effective predictions of heat stress in birds can be
made directly from empirical approaches of estimatingthermal responses to heat [5], but such predictions as-sume conditions similar to metabolic chambers and aretherefore less robust. Mechanistic approaches are alsomore likely to suggest new avenues for research, by allow-ing hypothetical scenarios to be explored as exemplifiedby our calculations of the consequences of burrowing forthe night parrot’s water budget. The generality of themodelling approach means it can be easily transferredto other taxa, the microclimate and endotherm modelsused here having been applied to a wide range of othertaxa [e.g. 32, 59].The ability to approximate microclimate conditions
relevant to night parrots (and other animals) fromcontinent-wide grids provides the potential to make thekinds of calculations illustrated here over much broaderscales of space and time. This will allow predictions ofthe relative suitability of different parts of Australia fornight parrots in recent history, under different scenariosof habitat quality, and thereby inform future survey workfor new populations.
Additional files
Additional file 1: Figure S2. Predicted (black) hourly wind speed (a)and air temperature (b) at the site compared to observations from theweather station (red) and, in late 2015 (green) the shaded 1.2 m airtemperature measurements from data loggers in trees as depicted inFig. 3a & b. (PDF 1117 kb)
Additional file 2: Figure S3. Observed relative humidity in night parrothabitat (red lines) compared to microclimate model predictions driven byhistorical gridded daily weather data (black lines). Logger locations aredepicted in Fig. 3. (PDF 289 kb)
Additional file 3: Figure S8. Predicted absolute monthly changes inminimum (blue) and maximum (red) air temperature at the study siteunder six different Global Circulation Models. (PDF 142 kb)
Additional file 4: Figure S9. Predicted absolute monthly changes inrelative humidity at the study site under six different Global CirculationModels. (PDF 125 kb)
Additional file 5: Figure S10. Predicted proportional monthly changesin wind speed at the study site under six different Global CirculationModels. (PDF 129 kb)
Additional file 6: Figure S11. Predicted proportional monthly changesin solar radiation at the study site under six different Global CirculationModels. (PDF 133 kb)
Additional file 7: Figure S12. Predicted proportional monthly changesin rainfall at the study site under six different Global Circulation Models.(PDF 137 kb)
Additional file 8: Figure S5. Predicted thermal conditions and hydricstate of the night parrot at the study site from 1996–2015 under theACCESS 1.3 global circulation model 2070 climate change scenario: a)maximum daily soil surface temperature (red), 4 cm air temperature inthe open (orange) and experienced air temperature (light blue – either inthe open or in the shade or in between, depending on the simulatedbehaviour of the parrot), with dashed horizontal lines indicating 38 °C(resting core body temperature) and 45 °C (near lethal body temperature),and vertical blue lines indicating rainfall (mm); b) daily maximum percentdehydration; c) daily cumulative dehydration, with rainfall events > 1 mmpermitting complete rehydration on a given day. (PDF 357 kb)
Additional file 9: Figure S1. Solar reflectivity across the wavelengths300–2100 nm for two specimens of the night parrot. (PDF 63 kb)
Additional file 10: Figure S4. Predicted thermal conditions and hydricstate of the night parrot at the study site from 1996–2015 under currentclimate: a) maximum daily soil surface temperature (red), 4 cm airtemperature in the open (orange) and experienced air temperature (lightblue – either in the open or in the shade or in between, depending onthe simulated behaviour of the parrot), with dashed horizontal linesindicating 38 °C (resting core body temperature) and 45 °C (near lethalbody temperature), and vertical blue lines indicating rainfall (mm); b)daily maximum percent dehydration; c) daily cumulative dehydration,with rainfall events > 1 mm permitting complete rehydration on a givenday. (PDF 294 kb)
Additional file 11: Figure S6. Calculations of night parrot environmentalconditions and thermal, hydric and energetic state in 2015 at the study siteas per Fig. 5, but assuming access to burrows 10 cm below shaded ground:a) hourly soil surface temperature (red), 4 cm air temperature in the open(orange) and experienced air temperature (light blue – either in the open orin the shade or in between, depending on the simulated behaviour of theparrot), with dashed horizontal lines indicating 38 °C (resting core bodytemperature) and 45 °C (near lethal body temperature), and vertical bluelines indicating rainfall (mm); b) hourly parrot core temperature (bright red),skin temperature (dark red) and outer feather temperature (orange); c)hourly discretionary energy (assimilated energy minus maintenance costs)when feeding at maximal simulated intake on seed (red), succulents with25% water (orange) and succulents with 55% water (blue); d) hourlyevaporative water loss rate; e) daily percentage body mass of water lost(i.e. negative values represent discretionary water); f) daily cumulativedehydration, with rainfall events > 1 mm permitting complete rehydrationon a given day. (PDF 253 kb)
Additional file 12: Figure S7. The frequency daily or cumulativedehydration of night parrots was predicted to exceed 11% or 22% overthe summers 1996/7 to 2014/15 at the site under historical and projected(2070 ACCESS 1.3) climate, as per Fig. 6, but assuming access to retreats10 cm below shaded ground. (PDF 78 kb)
AcknowledgementsMK thanks Katie Date and Monique Winterhoff from Museum Victoriaand Devi Stuart-Fox from The University of Melbourne for assistance withmeasurement of the museum specimens. SM thanks Rachel Murphyfor her assistance in the field, and also acknowledges the support ofCatherine Bozanich (Fortescue Metals Group) and Bush Heritage Australia.MK and WP acknowledge Natalie Briscoe and Paul Mathewson for theircontributions to the model development and testing. All authorsacknowledge the contribution that John Young made by his initialdiscovery of night parrots in western Queensland. We also thank JosephForshaw for his insights into night parrot burrowing.
FundingFunding for field work was provided by Fortescue Metals Group Limitedunder an agreement between them and the Australian Government
Kearney et al. Climate Change Responses (2016) 3:14 Page 15 of 17

associated with Environment Protection and Biodiversity Conservation Act(1999) approval EPBC No 2010/5696, condition 10. SM works under contractto Fortescue to develop and implement the required research investment, asoutlined in the condition. The modelling work was supported by theNational Environmental Research Program’s Threatened Species RecoveryHub, under funding from the Australian Government.
Availability of data and materialsAll data generated or analysed during this study are included in thispublished article [and its additional files].
Authors’ contributionsMK, SM and WP designed the study, SM collected the field data, MKcollected the museum data, MK and WP developed the models, MK ran thesimulations and analysed the data, MK, SM and WP wrote the manuscript. Allauthors read and approved the final manuscript.
Authors’ informationMK has had a long-term interest in arid zone biota and has spent the last 12 yearsdeveloping a system to apply biophysical models in the Australian context. WPwas a founder of the field of biophysical ecology and was the original and currentco-developer of the models used in this project. SM is a field ecologist with afascination for the night parrot since childhood and was contracted to undertakethe first scientific research project on the species between 2014 and 2016.
Competing interestsThe MRK and SAM declare that they have no competing interests. WPP isco-founder of Niche Mapper, LLC.
Consent for publicationNot applicable.
Ethics approvalThe field research in this project was done under scientific research permitsWISP14627114/WITK14627214, and Animal Ethics Committee permit SA 2015/09/521.
Author details1School of BioSciences, The University of Melbourne, Parkville, VIC 3010,Australia. 2Department of Zoology, The University of Wisconsin, Madison53706, USA. 3Research Institute for the Environment and Livelihoods, CharlesDarwin University, Casuarina, NT 0800, Australia. 4Bush Heritage Australia,Level 1, 395 Collins St, Melbourne, VIC 3000, Australia.
Received: 9 September 2016 Accepted: 28 November 2016
References1. Ruhl J. Climate change and the endangered species Act: building bridges to
the no-analog future. Boston Univ Law Rev. 2008;88(1):07–18.2. Hoegh-Guldberg O, Huges L, McIntyre S, Lindenmayer DB, Parmesan C,
Possingham HP, Thomas CD. Assisted colonization and rapid climatechange. Science. 2008;321:345–6.
3. Cooke SJ, Sack L, Franklin CE, Farrell AP, Beardall J, Wikelski M, Chown SL:What is conservation physiology? Perspectives on an increasingly integratedand essential science. Conserv Physiol 2013;1(1):1–23.
4. Walsberg GE. Small mammals in Hot deserts: some generalizations revisited.Bioscience. 2000;50(2):109–20.
5. McKechnie AE, Wolf BO. Climate change increases the likelihood ofcatastrophic avian mortality events during extreme heat waves. Biol Lett.2010;6(2):253–6.
6. Suppiah R, Hennessy KJ, Whetton PH, McInnes K, Macadam I, Bathols J,Ricketts J, Page CM. Australian climate change projections derived fromsimulations performed for the IPCC 4th assessment report. Aust MeteorolMag. 2007;56(3):131–52.
7. Reside A, VanDerWal J, Phillips B, Shoo L, Rosauer D, Anderson B,Welbergen J, Moritz C, Ferrier S, Harwood T, et al. Climate change refugiafor terrestrial biodiversity: defining areas that promote species persistenceand ecosystem resilience in the face of global climate change. In. GoldCoast: National Climate Change Adaptation Research Facility; 2013.
8. McKechnie AE, Hockey PAR, Wolf BO: Feeling the heat: Aust landbirds andclim chang. Emu 2012, 112(2):i-vii.
9. Gates DM. Biophysical ecology. New York: Springer Verlag; 1980.10. Porter WP, Gates DM. Thermodynamic equilibria of animals with
environment. Ecol Monogr. 1969;39(3):227–44.11. Porter WP, Budaraju S, Stewart WE, Ramankutty N. Calculating climate effects
on birds and mammals: impacts on biodiversity, conservation, populationparameters, and global community structure. Am Zool. 2000;40(4):597–630.
12. Kearney MR, Porter WP. NicheMapR - an R package for biophysicalmodelling: the microclimate model. Ecography. 2016.
13. Jones DA, Wang W, Fawcett R. High-quality spatial climate data-sets forAustralia. Aust Meteorol Oceanogr J. 2009;58:233–48.
14. McVicar TR, Van Niel TG, Li LT, Roderick ML, Rayner DP, Ricciardulli L,Donohue RJ. Wind speed climatology and trends for Australia, 1975–2006:capturing the stilling phenomenon and comparison with near-surfacereanalysis output. Geophys Res Lett. 2008;35(20):L20403.
15. Collar NJ. Psittacidae (parrots). In: del Hoyo J, Elliott A, Sargatal J, editors.Handbook of the birds of the world, Sandgrouse to cuckoos, vol. 4. Barcelona:Lynx Edicions; 1997.
16. Andrews FW. Notes on the night parrot. Trans Proc R Soc S Aust. 1883;6:29–30.17. Cleland JB. The history of ornithology in south Australia. Emu. 1936;36:197–221.18. Menkhorst P, Ryan E: CH. McLennan (‘mallee Bird’) and his aboriginal
informant jowley: the source of early records of the night parrot pezoporusoccidentalis in Victoria? Mem Mus Victoria. 2015;73:107–15.
19. Wilson H. Notes on the night parrot, with references to recent occurrences.Emu. 1937;37:79–87.
20. McGilp JN. Geopsittacus occidentalis, night-parrot. S Aust Ornithol. 1931;11:68–70.21. Murphy S. Night parrot (pezoporus occidentalis) research plan. In.
Yungaburra, QLD and Perth: Map IT and Fortescue Metals Group; 2014.22. Murie J. On the nocturnal ground-parrakeet (geopsittacus occidentalis,
Gould). Proc Zool Soc London. 1868;1868:158–65.23. McFarland D. The biology of the ground parrot, pezoporus wallicus, in
Queensland. I. Microhabitat use, activity cycle and diet. Wildl Res. 1991;18(2):169–84.24. McNab A, Sanders MG, 4/5/6. Consumption of exotic grass seeds (Poaceae:
Cynodon dactylon) by the eastern ground parrot (pezoporous wallicus).Queensland Nat. 2014;52:82–4.
25. Boles W, Longmore N, Thompson M. A recent specimen of the night parrotgeopsittacus occidentalis. Emu. 1994;94(1):37–40.
26. Ellis R. Bush safari. Adelaide: Rigby Publishers; 1982.27. Australia’s bioregions (IBRA) [https://www.environment.gov.au/land/nrs/
science/ibra]. Accessed 1 Dec 2016.28. Gueymard C. SMARTs, a simple model of the atmospheric radiative transfer
of sunshine: algorithms and performance assessment. In. Cocoa: FloridaSolar Energy Center; 1995.
29. R Development Core Team. R: A language and environment for statisticalcomputing. Vienna: R Foundation for Statistical Computing; 2012. URLhttp://www.R-project.org/. ISBN 3-900051-07-0.
30. Porter WP, Munger JC, Stewart WE, Budaraju S, Jaeger J. Endothermenergetics: from a scalable individual-based model to ecologicalapplications. Aust J Zool. 1994;42:125–62.
31. Porter WP, Vakharia NP, Klousie WD, Duffy D. Po’ouli landscapebioinformatics models predict energetics, behavior, diets and distributionon Maui. Integr Comp Biol. 2006;46:1143–58.
32. Briscoe NJ, Kearney MR, Taylor CA, Wintle BA. Unpacking the mechanismscaptured by a correlative species distribution model to improve predictionsof climate refugia. Glob Chang Biol. 2016;22(7):2425–39.
33. Leeton PRJ, Christidis L, Westerman M, Boles WE. Molecular phylogeneticaffinities of the night parrot (geopsittacus occidentalis) and the groundparrot (pezoporus wallicus). Auk. 1994;111(4):833–43.
34. Cade TJ, John A, Dybas Jr. Water economy of the budgerygah. Auk.1962;79(3):345–64.
35. Greenwald L, Stone WB, Cade TJ. Physiological adjustments of thebudgerygah (Melopsittacus undulatus) to dehydrating conditions. CompBiochem Physiol. 1967;22(1):91–100.
36. Weathers WW, Schoenbaechler DC. Regulation of body temperature in thebudgerygah. Melopsittacus undulatus Aust J Zool. 1976;24(1):39–47.
37. Prinzinger R, Preßmar A, Schleucher E. Body temperature in birds. CompBiochem Physiol A Physiol. 1991;99(4):499–506.
38. Schmidt-Nielsen K, Hainsworth FR, Murrish DE. Counter-current heatexchange in the respiratory passages: effect on water and heat balance.Respir Physiol. 1970;9(2):263–76.
39. Chato JC. In: Chao BT, editor. Advanced heat transfer. Urbana: U. of Ill. Press;1969. p. 395–412. Heat transfer in bioengineering. In.
Kearney et al. Climate Change Responses (2016) 3:14 Page 16 of 17

40. McKechnie AE, Wolf BO. The allometry of avian basal metabolic rate: goodpredictions need good data. Physiol Biochem zool: Ecol Evol Approaches.2004;77(3):502–21.
41. Hoffman TCM, Walsberg GE, DeNardo DF. Cloacal evaporation: an importantand previously undescribed mechanism for avian thermoregulation. J ExpBiol. 2007;210(5):741–9.
42. Hainsworth FR. Animal physiology. Reading: Addison-Wesley PublishingCompany; 1981.
43. Lindstrom A, Kvist A. Maximum energy intake rate is proportional to basalmetabolic rate in passerine birds. Proc R Soc Lond Ser B Biol Sci. 1995;261(1362):337–43.
44. Waterhouse CE, Hutcheson LM, Booker KM. Food consumption andpalatability studies in budgerigars. J Small Anim Pract. 1961;2(1–4):175–84.
45. Kearney MR, Shamakhy A, Tingley R, Karoly DJ, Hoffmann AA, Briggs PR,Porter WP. Microclimate modelling at macro scales: a test of a generalmicroclimate model integrated with gridded continental-scale soil andweather data. Methods Ecol Evol. 2014;5:273–86.
46. Watterson IG, Hirst AC, Rotstayn LD. A skill score based evalutation ofsimulated Australian climate. Aust Meteorol Oceanogr J. 2013;63:181–90.
47. Ashcroft MB, Gollan JR. Fine-resolution (25 m) topoclimatic grids of near-surface (5 cm) extreme temperatures and humidities across various habitats ina large (200 × 300 km) and diverse region. Int J Climatol. 2012;32(14):2134–48.
48. Kearney M, Porter WP. Mechanistic niche modelling: combiningphysiological and spatial data to predict species’ ranges. Ecol Lett.2009;12:334–50.
49. Kearney MR, Simpson SJ, Raubenheimer D, Kooijman SALM. Balancing heat,water and nutrients under environmental change: a thermodynamic nicheframework. Funct Ecol. 2013;27(4):950–66.
50. Mitchell N, Hipsey MR, Arnall S, McGrath G, Bin Tareque H, Kuchling G,Vogwill R, Sivapalan M, Porter WP, Kearney MR. Linking eco-energetics andeco-hydrology to select sites for the assisted colonisation of Australia’srarest reptile. Biology. 2013;2(1):1–25.
51. Burton S, Perrin MR, Downs CT. Thermal biology of African lovebirds andAustralian grass parakeets. J Therm Biol. 2008;33(6):355–62.
52. Angilletta MJ, Cooper BS, Schuler M, Boyles JG: The evolution of thermalphysiology in endotherms. Frontiers in Bioscience 2009, in press.
53. Maclean GL: Ecophysiology of Desert Birds. Berlin: Springer-Verlag54. Schmid WD. Temperature gradients in the nasal passage of some small
mammals. Comp Biochem Physiol A Physiol. 1976;54(3):305–8.55. Humphries MM, Careau V. Heat for nothing or activity for free? evidence
and implications of activity-thermoregulatory heat substitution. Integr CompBiol. 2011;51(3):419–31.
56. Sunday JM, Bates AE, Kearney MR, Colwell RK, Dulvy NK, Longino JT, HueyRB. Thermal-safety margins and the necessity of thermoregulatory behavioracross latitude and elevation. Proc Natl Acad Sci. 2014;11(15):5610–5.
57. Schodde R, Mason IJ. Nocturnal birds of Australia. Melbourne: Lansdowne; 1980.58. Forshaw JM, Cooper WT. Australian parrots, second edition edn. Lansdowne:
Melbourne; 1981.59. Kearney MR, Wintle BA, Porter WP. Correlative and mechanistic models of
species distribution provide congruent forecasts under climate change.Conserv Lett. 2010;3(3):203–13.
60. Sotherland PR, Rahn H. On the composition of bird eggs. Condor.1987;89(1):48–65.
61. Teixeira CMGL, Sousa T, Marques GM, Domingos T, Kooijman SALM. A newperspective on the growth pattern of the wandering albatross (Diomedeaexulans) through DEB theory. J Sea Res. 2014;94:117–27.
• We accept pre-submission inquiries
• Our selector tool helps you to find the most relevant journal
• We provide round the clock customer support
• Convenient online submission
• Thorough peer review
• Inclusion in PubMed and all major indexing services
• Maximum visibility for your research
Submit your manuscript atwww.biomedcentral.com/submit
Submit your next manuscript to BioMed Central and we will help you at every step:
Kearney et al. Climate Change Responses (2016) 3:14 Page 17 of 17

Page 24 of 27
8. Breeding biology
Murphy, S.A., Austin, J.J., Murphy, R.K., Silcock, J., Joseph, L., Garnett, S.T., Leseberg, N.P.,
Watson, J.E.M., and Burbidge, A.H. (2017). Observations on breeding Night Parrots (Pezoporus
occidentalis) in western Queensland. Emu - Austral Ornithology 117, 107-113.

Full Terms & Conditions of access and use can be found athttp://www.tandfonline.com/action/journalInformation?journalCode=temu20
Download by: [CSIRO Library Services] Date: 28 February 2017, At: 21:43
Emu - Austral Ornithology
ISSN: 0158-4197 (Print) 1448-5540 (Online) Journal homepage: http://www.tandfonline.com/loi/temu20
Observations on breeding Night Parrots(Pezoporus occidentalis) in western Queensland
Stephen A. Murphy, Jeremy J. Austin, Rachel K. Murphy, Jennifer Silcock, LeoJoseph, Stephen T. Garnett, Nicholas P. Leseberg, James E. M. Watson & AllanH. Burbidge
To cite this article: Stephen A. Murphy, Jeremy J. Austin, Rachel K. Murphy, Jennifer Silcock,Leo Joseph, Stephen T. Garnett, Nicholas P. Leseberg, James E. M. Watson & Allan H. Burbidge(2017): Observations on breeding Night Parrots (Pezoporus occidentalis) in western Queensland,Emu - Austral Ornithology
To link to this article: http://dx.doi.org/10.1080/01584197.2017.1292404
View supplementary material
Published online: 28 Feb 2017.
Submit your article to this journal
View related articles
View Crossmark data

Observations on breeding Night Parrots (Pezoporus occidentalis) in westernQueenslandStephen A. Murphya,b, Jeremy J. Austinc, Rachel K. Murphya, Jennifer Silcockd, Leo Josephe, Stephen T. Garnetta,Nicholas P. Lesebergf, James E. M. Watsonf,g and Allan H. Burbidgeh
aResearch Institute for the Environment and Livelihoods, Charles Darwin University, Casuarina, Australia; bBush Heritage Australia,Melbourne, Australia; cAustralian Centre for Ancient DNA, School of Biological Sciences, University of Adelaide, Adelaide, Australia; dSchoolof Biological Sciences, University of Queensland, St Lucia, Australia; eAustralian National Wildlife Collection, National Research CollectionsAustralia CSIRO, Canberra, Australia; fSchool of Geography, Planning and Environmental Management, University of Queensland, St Lucia,Australia; gWildlife Conservation Society, Global Conservation Program, New York, USA; hDepartment of Parks and Wildlife, Bentley, Australia
ABSTRACTA population of Night Parrots (Pezoporus occidentalis) was discovered in 2013 in westernQueensland and has become the primary focus of efforts aimed at conserving habitat andprotecting the species from extinction. Critical information on nesting habitat and location,breeding season and behaviour, clutch size and breeding success is currently limited to anecdotal19th-century observations and accounts by early natural historians. Here we describe severalbreeding attempts at Pullen Pullen Reserve. Our observations include nest and fledgling descrip-tions, habitat and clutch characteristics, breeding seasonality, adult breeding behaviour andvocalisations. We also identify a King Brown Snake (Pseudechis australis) as the predator respon-sible for one nesting failure. Our observations confirm historical reports of nesting habitat, eggand clutch size and breeding seasonality and provide important new information on vocalisationsand adult behaviour around an active nest. These new data provide a basis for future studies andconservation management of this enigmatic threatened species.
ARTICLE HISTORYReceived 8 November 2016Accepted 24 January 2017
KEYWORDSBreeding; conservation;Endangered species; nestbuilding; nocturnal birds;parrots; predation;threatened species
Introduction
The Night Parrot (Pezoporus occidentalis) is a small(~100 g; Murphy et al. in press) nocturnal parrot ende-mic to Australia’s arid zone. It is listed as Endangeredunder the Commonwealth Environment Protection andBiodiversity Conservation Act 1999. Although there areonly 25 specimens known to exist, judging by geogra-phically widespread specimen locations, including twoplaces (Gawler Ranges and Alice Springs) where multi-ple individuals were recorded over a relatively shortperiod, the species appears to have been widespreadand apparently relatively common until the late 19thcentury (Andrews 1883; Ashby 1924; Black 2012).However, after 1912 – when a specimen was shot inthe Gascoyne region of Western Australia (Wilson1937) – confirmed records of living birds ceased foralmost 100 years despite numerous searches (see Pykeand Ehrlich 2014 for examples). In 1990 and 2006,desiccated bodies were discovered in south-westernQueensland (Boles et al. 1994; McDougall et al. 2009).Finally, the first photographs of a living Night Parrot
were obtained in 2013 by naturalist John Young (Dooley2013). The location discovered by Young turned out tobe occupied over an extended period, presenting a rareand important opportunity to conduct research into thislittle-known species (Murphy 2014).
Before 2013 most information about NightParrots, including their breeding biology, was eitherbased on anecdotes or inference, or was non-existent.Several sources suggested that Night Parrots bred inhummocks of the (mostly) arid-adapted grass genusTriodia (Andrews 1883; Whitlock 1924; McGilp1931; Wilson 1937; Kershaw 1943; Menkhorst andRyan 2015). In the late 1800s, a South Australiancattle grazier, John McDonald, reportedly found‘three or four nests’ north-east of Coober Pedy, oneof which was under a low, dense samphire bush(McGilp 1931, p. 69). Aboriginal people suggestedthat breeding occurs after significant rain (Wilson1937) and Andrews (1883) ventured that it is trig-gered when rain induces prolific Triodia seedproduction.
CONTACT Stephen A. Murphy [email protected] data for this article can be accessed here.
EMU - AUSTRAL ORNITHOLOGY, 2017http://dx.doi.org/10.1080/01584197.2017.1292404
© 2017 BirdLife Australia
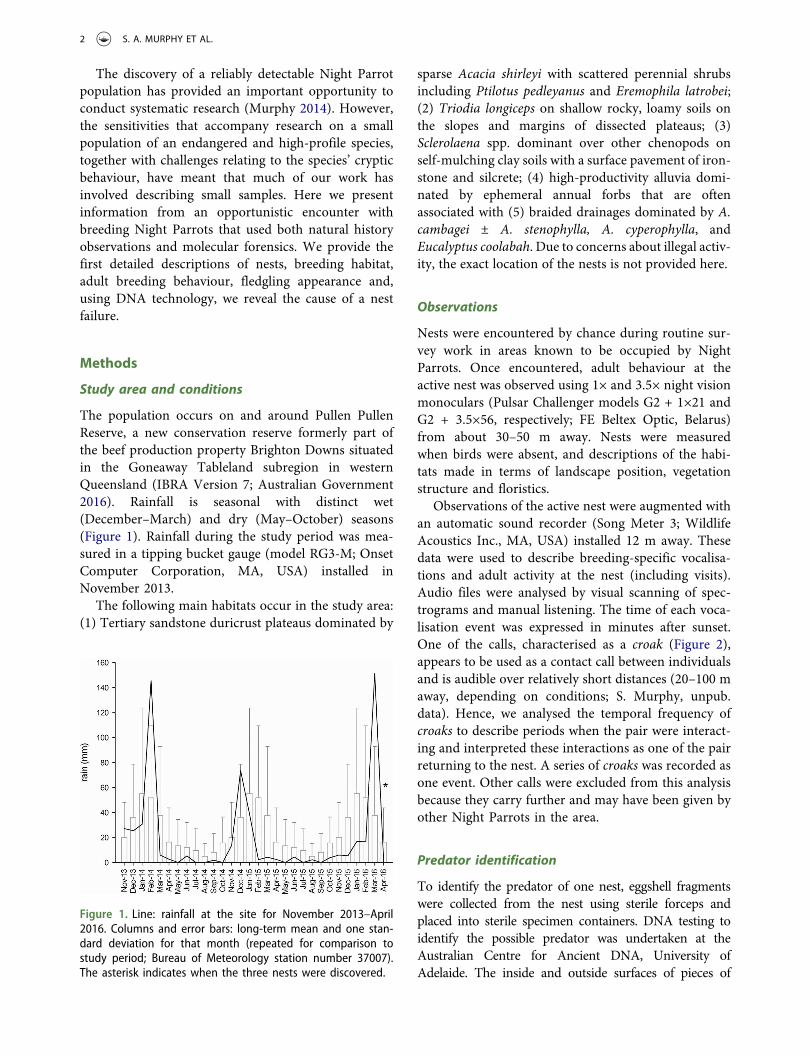
The discovery of a reliably detectable Night Parrotpopulation has provided an important opportunity toconduct systematic research (Murphy 2014). However,the sensitivities that accompany research on a smallpopulation of an endangered and high-profile species,together with challenges relating to the species’ crypticbehaviour, have meant that much of our work hasinvolved describing small samples. Here we presentinformation from an opportunistic encounter withbreeding Night Parrots that used both natural historyobservations and molecular forensics. We provide thefirst detailed descriptions of nests, breeding habitat,adult breeding behaviour, fledgling appearance and,using DNA technology, we reveal the cause of a nestfailure.
Methods
Study area and conditions
The population occurs on and around Pullen PullenReserve, a new conservation reserve formerly part ofthe beef production property Brighton Downs situatedin the Goneaway Tableland subregion in westernQueensland (IBRA Version 7; Australian Government2016). Rainfall is seasonal with distinct wet(December–March) and dry (May–October) seasons(Figure 1). Rainfall during the study period was mea-sured in a tipping bucket gauge (model RG3-M; OnsetComputer Corporation, MA, USA) installed inNovember 2013.
The following main habitats occur in the study area:(1) Tertiary sandstone duricrust plateaus dominated by
sparse Acacia shirleyi with scattered perennial shrubsincluding Ptilotus pedleyanus and Eremophila latrobei;(2) Triodia longiceps on shallow rocky, loamy soils onthe slopes and margins of dissected plateaus; (3)Sclerolaena spp. dominant over other chenopods onself-mulching clay soils with a surface pavement of iron-stone and silcrete; (4) high-productivity alluvia domi-nated by ephemeral annual forbs that are oftenassociated with (5) braided drainages dominated by A.cambagei ± A. stenophylla, A. cyperophylla, andEucalyptus coolabah. Due to concerns about illegal activ-ity, the exact location of the nests is not provided here.
Observations
Nests were encountered by chance during routine sur-vey work in areas known to be occupied by NightParrots. Once encountered, adult behaviour at theactive nest was observed using 1× and 3.5× night visionmonoculars (Pulsar Challenger models G2 + 1×21 andG2 + 3.5×56, respectively; FE Beltex Optic, Belarus)from about 30–50 m away. Nests were measuredwhen birds were absent, and descriptions of the habi-tats made in terms of landscape position, vegetationstructure and floristics.
Observations of the active nest were augmented withan automatic sound recorder (Song Meter 3; WildlifeAcoustics Inc., MA, USA) installed 12 m away. Thesedata were used to describe breeding-specific vocalisa-tions and adult activity at the nest (including visits).Audio files were analysed by visual scanning of spec-trograms and manual listening. The time of each voca-lisation event was expressed in minutes after sunset.One of the calls, characterised as a croak (Figure 2),appears to be used as a contact call between individualsand is audible over relatively short distances (20–100 maway, depending on conditions; S. Murphy, unpub.data). Hence, we analysed the temporal frequency ofcroaks to describe periods when the pair were interact-ing and interpreted these interactions as one of the pairreturning to the nest. A series of croaks was recorded asone event. Other calls were excluded from this analysisbecause they carry further and may have been given byother Night Parrots in the area.
Predator identification
To identify the predator of one nest, eggshell fragmentswere collected from the nest using sterile forceps andplaced into sterile specimen containers. DNA testing toidentify the possible predator was undertaken at theAustralian Centre for Ancient DNA, University ofAdelaide. The inside and outside surfaces of pieces of
Figure 1. Line: rainfall at the site for November 2013–April2016. Columns and error bars: long-term mean and one stan-dard deviation for that month (repeated for comparison tostudy period; Bureau of Meteorology station number 37007).The asterisk indicates when the three nests were discovered.
2 S. A. MURPHY ET AL.

eggshell were swabbed with sterile, DNA-free swabs(Copan 4N6FLOQSwabs; Copan, Murrieta, CA, USA).DNA was extracted from four swabs (shell1_inside, shel-l1_outside, shell2_inside, shell2_outside) using protocolsdescribed elsewhere (Austin et al. 2013) and with a nega-tive extraction control. A short (<140 base-pair, includingprimers) hypervariable segment of the mitochondrialDNA (mtDNA) 16S rRNA gene was PCR amplified andSanger sequenced as described byAustin et al. (2013) usingprimers 16Smam1 and 16Smam2 (Taylor 1996). Sequencechromatograms were manually edited using Geneiousv9.0.5 (Biomatters, Auckland, New Zealand) and consen-sus sequence (trimmed of primer sequence) searchedagainst the NCBI nucleotide database using BLASTsearches (‘nr’ database, blastn program) within Geneious.
Retrospective analysis of previously collectedacoustic data
Since August 2013, acoustic surveys using Song Metershave been conducted more or less continually at multi-ple sites on Pullen Pullen, which have resulted in alarge audio archive. We searched this archive for pat-terns of occurrence vocalisations that were similar towhat we observed at the active nest, which may suggestprevious breeding activity. The archive was searchedusing the automated acoustic detection software SongScope 4.1.3.A (Wildlife Acoustics Inc., Concord, MA,USA). In total we analysed 62 recording nights at onelocation for the 2013–2014 summer wet period, 191recording nights at five locations for the 2014–2015summer wet period and 325 recording nights at threelocations for the April–July 2015 dry period.
Results
Rainfall
A total of 185.2 mm fell between 26 January and 1 May2016, most of which occurred in March (151.4 mm).Compared to nearby historical rainfall data (Bureau ofMeteorology 2016), this was within the upper 5% per-centile for totals for the month of March.
Nest discovery and measurements
Breeding activity was first suspected in April 2016 whenadult behaviour after dusk changed from behavioursobserved during the same time period over the previous2 years. Crepuscular movement by the birds increased,including adults flying close to observers on severaloccasions, and three novel vocalisations (all archived atthe Australian National Wildlife Collection, CSIRO,Canberra) were recorded (referred to as peel, chirrupand prat calls – the latter resembling the call of theAustralian Pratincole Stiltia isabella). These calls wereadditional to the croak call (referred to by Andrews(1883); Figure 2), and dink-dink call (Murphy and TheNight Parrot Recovery Team 2015), and ding-de-dingand dee-de-dee-de calls described here for the firsttime. The latter three calls have a more or less bell-like, ringing quality and are detected in all seasons (S.Murphy, unpub. data). The first nest was discoveredwhen a bird was unintentionally flushed at approxi-mately 1900 h on 24 April 2016. The bird flushedsilently from a Triodia hummock and landed a fewmetres away from the hummock and the observer (S.M.). It was observed through 1× night vision for about 5s, during which time it ran upslope a little, before flyingoff, giving the croak call once or twice. S.M. and R.M.then discovered a nest containing two eggs (see supple-mentary material Figure S1). The area was vacatedimmediately to avoid causing any further disturbance.
We did not handle the eggs to determine age ordimensions. However, by comparing the eggs to theknown dimension of the bowl structure in photographs,the eggs were estimated to be 30 × 25mm. The eggs weresub-elliptical, immaculate white and had a bright, lus-trous appearance, suggesting they were recently laid;eggs typically go darker and duller as incubation pro-gresses (Ian Mason, pers. comm., 6 June 2016).
Nest descriptions, habitat and clutch characteristics
All nests were in large, more or less ring-shaped hum-mocks of T. longiceps (the only Triodia species in thearea). The first (active) nest comprised three parts: atunnel constructed through an apron of dead Triodia
Figure 2. Spectrogram of the Night Parrot croak call. This seriesof seven croaks was recorded at the nest at approximately0020h on 27 April 2016. This recording is stored at theAustralian National Wildlife Collection (ANWC) Sound Library,registration number ANWCX49087.
EMU – AUSTRAL ORNITHOLOGY 3

leaves that had accumulated inside the ring, leading toa chamber constructed low in the living part of thehummock, at the bottom of which was a depressionexcavated into the gravelly/sandy soil. The tunnel andthe chamber were made by a combination of chewingsome Triodia leaves, and pushing and shaping others(the majority). The depression was lined deeply andsolely with short sections of Triodia leaves that werealso chewed.
A search of many nearby hummocks revealed twoadditional nests. One (Nest 2) was discovered 16 mupslope of the active nest. One Night Parrot featherwas collected from its tunnel and although a bowl wasconstructed, it was not lined and nor were any scatsevident. In all respects Nest 2 was similar to the activenest, although some of the measurements differedslightly (Table 1). We assume this nest was neveractive. The third nest was situated 330 m west of Nest1. It was discovered at 1900 h on 8 May and rapidinspection at the time revealed that it contained onlyfeathers. Due to flooding rain that occurred later thatnight, this nest could not be revisited until 13 May,when nestling remains were collected. Evidently, thisnest was active but had failed before its discovery on 8May. Like the second nest, this third nest was similar inmost respects to the active nest (Table 1).
The three nests were in an area of gently undulatingironstone pavement, 700–1000 m from a Tertiary sand-stone range. Triodia occurs in dense stands around themargins of the range and along nearby shallow drai-nages, but exists only as isolated, small (~20 m) agglom-erations of large hummocks on the ironstone pavementwhere the nests were built (see supplementary materialFigure S2). Here, other vegetation cover is sparse andconfined mostly to small depressions or within hum-mocks, with common species including the annualgrasses Eriachne pulchella and Enneapogon polyphyllus,annual herbs Trianthema triquetra and Ptilotus nobilis,and perennial herbs Sclerolaena longicuspis, Maireanaspongiocarpa and Hemichroa mesembryanthema.
Observations at the active nest
While active, Nest 1 was observed on two evenings (25and 27 April). Our impression was that the non-incu-bating bird (sex unknown) roosted during the day inhummocks that were approximately 100 m downslopeof the nest. At approximately 1840h on both observa-tion days the non-incubating bird called for a briefperiod away from the nest before landing on theopen, stony surface within a few metres of the nesthummock. There it gave several loud peel and chirrupcalls. These were answered by the incubating bird withone or two faint dink-dink calls before both birds flewoff together, presumably to feed. On both evenings weceased observations after this initial period of activityto avoid disturbance when one or both adults returnedto the nest.
The automatic sound recorder captured 57.8 h ofaudio between 25 April and 1 May, within which 631Night Parrot vocalisations were detected (Table 2). Therecordings showed similar patterns of calling to ourdirect observations at the nest; namely, peel and chirrupcalls answered by dink-dink calls, mostly faint and fewin number. Analyses of 80 croak calls suggested therewere peaks of calling at dusk and dawn, interspersedwith periodic lulls in detection rate throughout thenight (Figure 3).
Nest fates
We visited Nest 1 a second time at 1915h on 30 April andfound it to contain only eggshell fragments. Judging bytypical levels of calling behaviour just before dawn that day,followed by unusually few distant calls that evening, weassume the active nest failed some time during the day on30 April. Only one of four swabs (shell2_inside) amplifiedfor themtDNA16S rRNA target. Sequencing of this ampli-con yielded 91 base-pairs of sequence (AAAAAAAAGAACTTCCAAAAACTTATTACATAATAATACT-AACCCGGCTAACAAGCCTATACAAGACCCGGTA-AAGCTGACAATTGAACCA). A BLAST search identified five sequences from Pseudechis australis (KingBrown Snake) as the most similar (97.8%, with two transi-tion substitutions) to the egg swab sequence; a further fiveTable 1. Dimensions of the three Night Parrot nests
Dimension Nest 1a Nest 2b Nest 3c
Tunnel entrance aspect (degrees) 164 240 46Tunnel length (cm) 20 30 33Tunnel angle (to the horizontalplane) (degrees)
0 30 25
Chamber entrance width (cm) 12 11 10Chamber entrance height (cm) 9 11 12Chamber diameter (cm) 28 20 25Bowl depth (cm) 3 4 4Bowl diameter (cm) 15 14 14
aDiscovered active with eggs.bUncompleted nest, never active.cDiscovered containing remains of nestling/s.
Table 2. Vocalisations detected at the active nestCall type Number detected
Peel 259Ding-de-ding 120Dink-dink 81Chirrup 81Croak 80Dee-de-dee-de 8Prat 2
4 S. A. MURPHY ET AL.

P. australis sequences being the next most similar (84.8–96.7%). The egg swab sequence is >20% divergent from allother Australian elapid snakes available on Genbank (>30genera, accessed 25 October 2016).
We do not know why the nestling(s) in Nest 3 died.Inspection of bones and feathers did not suggest thatmore than one nestling died in the nest. Most of theskeleton was missing, as were most of the feathers.Judging by the development of remiges, and by com-parison to descriptions of feathers in relation to age forEastern Ground Parrots (P. wallicus; Higgins 1999) weestimate the age at death to be 14–20 days. All partsthat we collected, plus the lining of the nest, are storedat the Queensland Museum (registration numberQMO.33208).
Breeding seasonality and retrospective acousticanalyses
The breeding we observed was during a period thatfollowed significant rain (Figure 1). In March,151.4 mm fell, which was well outside one standarddeviation from the mean (37.6 ± 55.2 mm) based onnearby long-term data (Bureau of Meteorology2016). Retrospective analyses of audio files fromother post-wet periods during the previous 2 yearsdetected one peel call in the post-February 2014 wetperiod (24 March 2014). In this period the pattern ofdink-dink and croak calls was similar to thatobserved at the active nest, in that calling was com-mon throughout the night, in addition to the normalpeaks observed at sunset and sunrise (Murphy andNPRT 2015). The pattern of calling during the dryperiod we sampled showed only dusk and dawn
peaks, with very few calls detected at other times. Afull treatment of Night Parrot calling behaviour isforthcoming.
Discussion
This paper describes our observations of an active NightParrot nest, the first recorded for over 100 years. Like ourobservations, most historical records of Night Parrotnests were in Triodia hummocks (Andrews 1883;Whitlock 1924; McGilp 1931; Wilson 1937; Kershaw1943). Although no previous nest records contain mea-sured dimensions, most descriptions are similar to thestructures that we observed with variations in the termsand phrases used. For instance, Andrews, in SouthAustralia, describes ‘. . . the inside [of the Triodia hum-mock] being pulled out and a snug retreat formed for itsprotection’ (Andrews 1883, p. 29), whereas a WesternAustralian Aboriginal informant, Masters, stated thatthey made ‘tunnels under the spinifex’ (Wilson 1937, p.84). The description of a nest found by Kelly in 1897includes a reference to a ‘slight depression’ (Kershaw1943, p. 196), while McDonald described ‘an enlargedcavity’ lined with ‘some small sticks’ (McGilp 1931, p.69). This is similar to the description given to Keartlandfrom Aboriginal people that mentions a ‘loosely-madecup nest under the shade of the spinifex’ (North 1897,p. 171). Our descriptions of the nests also closely accordwith those of Eastern Ground Parrots (Forshaw 2002).
Our estimate of egg size (30 × 25 mm) is similar tothose of the crested pigeon (30 × 22 mm; Beruldsen1980), a species that grazier John McDonald reportedas having eggs of similar size to the Night Parrot eggshe observed in the late 19th century (McGilp 1931).Similarly, the white colour and rounded, oval shape weobserved tallies with previous reports (McGilp 1931;Wilson 1937; Menkhorst and Ryan 2015) and ofrecords of the slightly smaller Eastern Ground Parroteggs (Forshaw 2002). Historical records suggest aclutch size range of between two and four (North1901–1914; McGilp 1931; Wilson 1937; Kershaw1943; Menkhorst and Ryan 2015) and our observationsfit with this, although we suspect that more eggs werelaid between discovery and failure.
Our analysis indicating that a King Brown Snake (P.australis) preyed on the eggs is the first documentedoccurrence of predation on Night Parrots. We have nodata to suggest that there are unusually or unnaturallyhigh densities of any snakes on Pullen Pullen Reserve.
All nests were discovered after a significant rainfallevent at Pullen Pullen Reserve. This accords with state-ments made by Andrews (1883) and Masters (inWilson 1937, p. 84). However, one significant
Figure 3. Mean number (and standard error) of croak contactcalls detected over 6 nights at the active Night Parrot nest.
EMU – AUSTRAL ORNITHOLOGY 5

departure from the observations made by Andrews(1883) is that the Triodia at Pullen Pullen during theearly 2016 breeding event did not coincide with largeamounts of Triodia seed. In fact, all florets in severaldozen T. longiceps flower spikes we examined aroundthis time were barren. Many species of Triodia areknown to produce barren flowers en masse (Wrightet al. 2014). Andrews’s (1883) account is so brief that itdoes not eliminate the possibility that he may havebeen deceived into thinking that seed was beingproduced.
It is intriguing that our retrospective analysis of voca-lisations revealed a within-night pattern of dink-dinkand croak calls in the 2013–2014 summer wet periodthat was similar to that we observed at the active nest,but that we recorded just one peel call. The explanationmay lie simply in the poor positioning of recordersrelative to other possible nests. Alternatively, birdsmay not have bred, or may not have committed to afull breeding attempt during other post-wet periods.
Although the three nests described here were allunsuccessful, we are buoyed by one positive footnote weare able to add: in late October 2016, N.L. and J.W. wereconducting preliminary fieldwork in the same area as partof a new phase of research. At 1820h on 29 October, aNight Parrot fledgling was flushed from a hummock ofspinifex, approximately 80 m west of the hummock thatcontained the nest with eggs found inApril. N.L. and J.W.watched the fledgling from 30–40 m away; it sat briefly atthe edge of the hummock with its back to the observersbefore it moved out of sight. It had a very short tail, andwas greyish-green in colour, with heavy black patterningon the head and back (see supplementary materialFigure S3). To avoid further disturbance no search for anest was undertaken. That evening an hour was spentobserving the hummock from a distance of 50 m usingnight vision equipment. At 1940h an adult bird landedclose to the hummock and over 15min gave several croakcalls. In each instance it was answered by several quietwhinnying calls, which we assume were fledglingresponses. These emanated from two locations, indicat-ing at least two fledglings were present. This most recentbreeding attempt also followed significant rain in thearea, and has shown that, at least in this year, breedingoccurred between April and October. This suggests thatbreeding might occur any time of the year followingsignificant rainfall, rather than being strictly seasonal.
These observations have contributed to a morerobust understanding of the natural history and biol-ogy of Night Parrots. Along with continuing studies onmovements, habitat use and diet, this information willunderpin conservation efforts at all sites where thisspecies is known to occur.
Acknowledgements
We acknowledge the support of Fortescue Metals Group, Peterand Carol Britton, Bush Heritage Australia and other membersof the Night Parrot Recovery Team. We also acknowledge thesignificant contribution that John Young made through hisinitial discovery of this site. Jen Silcock thanks the NationalEnvironmental Science Programme for supporting her contri-bution to this work. We thank the editor, associate editor andanonymous reviewers for helpful comments on the manuscript.This research was conducted under Scientific Purposes PermitNumber WISP14627114 and QLD Department of Agricultureand Fisheries Animal Ethics Approval number SA 2015/09/521.
References
Andrews, F. W. (1883). Notes on the Night Parrot.Transactions and Proceedings of the Royal Society ofSouth Australia 6, 29–30.
Ashby, E. (1924). Notes on extinct or rare Australian birds,with suggestions as to some of the causes of their disap-pearance. Emu 23, 178–183. doi:10.1071/MU923178
Austin, J. J., Joseph, L., Pedler, L., and Black, A. (2013).Uncovering cryptic evolutionary diversity in extant andextinct populations of the southern australian arid zonewestern and thick-billed grasswrens (passeriformes: mal-uridae: amytornis). Conservation Genetics 14, 1173–1184.doi:10.1007/s10592-013-0504-9
Australian Government. (2016). Australia’s bioregions(IBRA). (Department of the Environment and Energy.)Available at https://www.environment.gov.au/land/nrs/science/ibra [Verified 29 July 2016].
Beruldsen, G. R. (1980). ‘A Field Guide to Australian Birds’Nests and Eggs.’ (Rigby Publishers Limited: Adelaide.)
Black, A. B. (2012). Collection localities of the Night ParrotPezoporus (Geopsittacus) occidentalis (Gould, 1861).Bulletin of the British Ornithologists Club 132, 277–282.
Boles, W. E., Longmore, N. W., and Thompson, M. C.(1994). A recent specimen of the Night ParrotGeopsittacus occidentalis. Emu 94, 37–40. doi:10.1071/MU9940037
Bureau of Meteorology (2016). Climate Data Online;Brighton Downs, Station Number 37007. Accessed 14September 2016.
Dooley, S. (2013). Out of the shadows. Australian Birdlife 2,26–30.
Forshaw, J. M. (2002). ‘Australian Parrots. Illustrated by W.T. Cooper,’ 3rd edn. (Alexander Editions: Robina,Queensland).
Higgins, P. J. (Ed.) (1999). ‘Handbook of Australian, NewZealand and Antarctic Birds. Vol. 4. Parrots to Dollarbird’.(Oxford University Press: Melbourne.)
Kershaw, J. (1943). Concerning a rare parrot. VictorianNaturalist 59, 196.
McDougall, A., Porter, G., Mostert, M., Cupitt, R., Cupitt, S.,Joseph, L., Murphy, S., Janetzki, H., Gallagher, A., andBurbidge, A. (2009). Another piece in an Australian ornitho-logical puzzle – a second Night Parrot is found dead inQueensland. Emu 109, 198–203. doi:10.1071/MU08018
McGilp, J. N. (1931). Geopsittacus occidentalis, Night-Parrot.South Australian Ornithologist 11, 68–70.
6 S. A. MURPHY ET AL.

Menkhorst, P., and Ryan, E. (2015). CH McLennan (‘MalleeBird’) and his Aboriginal informant Jowley: The source ofearly records of the Night Parrot Pezoporus occidentalis inVictoria? Memoirs of Museum Victoria 73, 107–115.
Murphy, S. (2014). ‘Night Parrot (Pezoporus Occidentalis)Research Plan.’ (Map IT and Fortescue Metals Group:Yungaburra and Perth.)
Murphy, S. A., Silcock, J. L.,Murphy, R. K., Reid, J., andAustin, J.(submitted). ‘Movements and Habitat Use of the Night ParrotPezoporus Occidentalis in South-Western Queensland.’
Murphy, S., and The Night Parrot Recovery Team. (2015).Shining a light: The research unlocking the secrets of themysterious Night Parrot. Australian Birdlife 4, 30–35.
North, A. (1901–1914). ‘Nests and Eggs of Birds FoundBreeding in Australia and Tasmania.’ (F.W. White: Sydney.)
North, A. J. (1897). List of birds collected by the CalvertExploring Expedition in Western Australia. Transactionsof the Royal Society of South Australia 22, 125–192.
Pyke, G. H., and Ehrlich, P. R. (2014). Conservation and theHoly Grail: The Story of the Night Parrot. PacificConservation Biology 20, 221–226. doi:10.1071/PC140221
Taylor, P. G. (1996). Reproducibility of ancient dnasequences from extinct pleistocene fauna. MolecularBiology And Evolution 13, 283-285. doi:10.1093/oxford-journals.molbev.a025566
Whitlock, F. L. (1924). Journey to Central Australia in searchof the Night Parrot. Emu 23, 248–281. doi:10.1071/MU923248
Wilson, H. (1937). Notes on the Night Parrot, with refer-ences to recent occurrences. Emu 37, 79–87. doi:10.1071/MU937079
Wright, B. R., Zuur, A. F., and Chan, G. C. K. (2014).Proximate causes and possible adaptive functions of mastseeding and barren flower shows in spinifex grasses(Triodia spp.) in arid regions of Australia. TheRangeland Journal 36, 297–308. doi:10.1071/RJ13104
EMU – AUSTRAL ORNITHOLOGY 7

Page 25 of 27
9. References
Andrews, F.W. (1883). Notes on the Night Parrot. Transactions and Proceedings of the Royal
Society of South Australia 6, 29-30.
Boles, W.E., Longmore, N.W., and Thompson, M.C. (1994). A recent specimen of the Night
Parrot Geopsittacus occidentalis. Emu 94, 37-40.
Boucher, N.J., Burbidge, A.H., and Jinnai, M. (2008). Computer recognition of sounds that
have never been heard before. Presented at 18th National Australian Institute of Physics
Conference, Adelaide.
Cupitt, R., and Cupitt, S. (2008). Another recent specimen of the Night Parrot Pezoporus
occidentalis from western Queensland. Australian Field Ornithology 25, 69.
Davis, R., and Metcalf, B. (2008). The Night Parrot (Pezoporus occidentalis) in northern
Western Australia a recent sighting from the Pilbara region. Emu 108, 233-236.
Jackett, N., Greatwich, B., Swann, G., and Boyle, A. (2017). A nesting record and vocalisations
of the Night Parrot Pezoporus occidentalis from the East Murchison, Western Australia.
Australian Field Ornithology 34, 144-150.
McDougall, A., Porter, G., Mostert, M., Cupitt, R., Cupitt, S., Joseph, L., Murphy, S., Janetzki,
H., Gallagher, A., and Burbidge, A. (2009). Another piece in an Australian
ornithological puzzle - a second Night Parrot is found dead in Queensland. Emu 109,
198-203.
Mills, V., and Collins, B. (2017). Night parrot captured in blurry image in the Great Sandy
Desert. Australian Broadcasting Corporation http://www.abc.net.au/news/2017-10-
14/night-parrot-found-sandy-desert/9047462 Accessed 19 October 2017.
Murphy, S.A., Austin, J.J., Murphy, R.K., Silcock, J., Joseph, L., Garnett, S.T., Leseberg, N.P.,
Watson, J.E.M., and Burbidge, A.H. (2017a). Observations on breeding Night Parrots
(Pezoporus occidentalis) in western Queensland. Emu - Austral Ornithology 117, 107-
113.
Murphy, S.A., Silcock, J., Murphy, R., Reid, J., and Austin, J.J. (2017b). Movements and
habitat use of the night parrot Pezoporus occidentalis in south-western Queensland.
Austral Ecology 42, 858-868.
Olsen, P. (2018). Night Parrot: Australia's most elusive bird. (CSIRO Publishing; Clayton
South, Victoria).
Powell, B. (1970). The Night Parrot. South Australian Ornithologist 25, 208.
R Core Team (2016). R: A language and environment for statistical computing.
Wilson, H. (1937). Notes on the Night Parrot, with references to recent occurrences. Emu 37,
79-87.

Page 26 of 27
10. Appendix 1 – Approved research plan

Report
Night Parrot (Pezoporus
occidentalis) Research Plan
10 January 2014 CB-RP-EN-1048

Night Parrot Pezoporus occidentalis Research Plan Page 2 of 40
TABLE OF CONTENTS
1. INTRODUCTION ............................................................................................................ 6
1.1 The Night Parrot ................................................................................................ 6
1.2 Scope ................................................................................................................. 7
1.3 Development of the Research Plan .................................................................. 8
1.4 Support during the life of the research program ............................................. 9
1.5 General Approach in Developing the Plan ....................................................... 9
1.6 Program Framework ........................................................................................ 11
2. PROPOSED RESEARCH ............................................................................................. 13
2.1 Developing Detection Strategies for Night Parrots ....................................... 13
2.1.1 Background and relevance .................................................................. 13
2.1.2 How this relates to other parts of the research plan ............................. 13
2.1.3 Aims ..................................................................................................... 14
2.1.4 Detection using sound .......................................................................... 14
2.1.5 Detection using camera traps ............................................................... 16
2.1.6 Timeline and anticipated outcomes ...................................................... 17
2.2 Understanding Night Parrot habitat preferences and use ........................ 18
2.2.1 Background and relevance .................................................................. 18
2.2.2 How this relates to other parts of the research plan ............................. 18
2.2.3 Aims ..................................................................................................... 19
2.2.4 Identifying key habitats ......................................................................... 19
2.2.5 Habitat descriptions and preferences ................................................... 21
2.2.6 Timeline and anticipated outcomes ...................................................... 21
2.3 Increasing Knowledge of Night Parrot Distribution ...................................... 22
2.3.1 Background and relevance .................................................................. 22
2.3.2 How this relates to other parts of the research plan ............................. 22
2.3.3 Aims ..................................................................................................... 23
2.3.4 Finding additional Night Parrot populations .......................................... 23
2.3.5 Following-up third party sightings ......................................................... 23
2.3.6 Timeline and anticipated outcomes ...................................................... 24
2.4 Understanding the threats to Night Parrots ................................................... 24
2.4.1 Background and relevance .................................................................. 24

Night Parrot Pezoporus occidentalis Research Plan Page 3 of 40
2.4.2 How this relates to other parts of the research plan ............................. 25
2.4.3 Aims ..................................................................................................... 25
2.4.4 Fire history of Night Parrot habitats ...................................................... 26
2.4.5 Potential impacts of mammalian predators on Night Parrots ................ 26
2.4.6 Potential impacts of grazing on Night Parrot habitat ............................. 27
2.4.7 Timeline and anticipated outcomes ...................................................... 27
2.5 Budget Estimate .............................................................................................. 28
3. TIMELINE SUMMARY AND MILESTONES ................................................................. 29
4. REPORTING REQUIREMENTS ................................................................................... 33
5. REFERENCES ............................................................................................................. 34

Night Parrot Pezoporus occidentalis Research Plan Page 4 of 40
List of Tables
Table 1: How this plan meets the requirements of the Condition 10 ....................... 8
Table 2: Potential areas of research, priority, description and rationale ............. 10
Table 3: Budget Estimate .......................................................................................... 28

Night Parrot Pezoporus occidentalis Research Plan Page 5 of 40
LIST OF FIGURES
Figure 1: Research program framework .................................................................... 12
Figure 2: Proposed timeline. ...................................................................................... 31
Figure 3: Projected proportion of time analysing various research components .. 32

Night Parrot Pezoporus occidentalis Research Plan Page 6 of 40
1. INTRODUCTION
1.1 The Night Parrot
Night Parrots (Pezoporus occidentalis) are small to medium-sized nocturnal parrots that are
endemic to Australia’s arid zone. Along with a suite of medium-sized desert-dwelling mammals,
the distribution and density of Night Parrots has reduced dramatically over the 20th century, and
indeed they may be locally extinct over much of their former range (Murphy 2012; Murphy et al.
2009). Under various State and Commonwealth legislation they are listed as:
Endangered (and Migratory) under the Environment Protection and Biodiversity
Conservation Act 1999 (Commonwealth)
Critically Endangered under the Territory Parks and Wildlife Conservation Act 2000
(Northern Territory)
Endangered under the Nature Conservation Act 1992 (Queensland)
Endangered under the National Parks and Wildlife Act 1972 (South Australia)
Rare Or Likely To Become Extinct under the Wildlife Conservation Act 1950 (Western
Australia)
Presumed Extinct under the Threatened Species Conservation Act 1995 (NSW)
Regionally Extinct under the Advisory List of Threatened Vertebrate Fauna in Victoria
2003 (Victoria)
Little is known about the biology of the Night Parrot. There is no reliable information about basic
attributes such as preferred habitats and use, breeding biology, movements, diet, social
interactions and threatening processes. Gaining this knowledge is being hindered by the lack of
robust, repeatable detection methods. A comprehensive review of Night Parrot biology appears
in Murphy (2012). Knowledge gaps that are relevant to this proposal appear in the backgrounds
to the sections below.
In May 2013, at the Queensland Museum, Queensland naturalist John Young presented the
findings of his search for the Night Parrot, complete with the first photographs of living birds.
Importantly, John revealed that he had been repeatedly visiting a location for a year or more,
during which time Night Parrots were present and he made sound recordings of their calls. Prior
to this, the only other time a site seemed to be reliably occupied by Night Parrots was in the
1870-80s at the Gawler Ranges in South Australia, where the majority of museum specimens
were collected. John’s discovery presents a critical, possibly unique, opportunity to collect
systematic data about Night Parrots, and provides the basis for much of the proposed research
presented here.

Night Parrot Pezoporus occidentalis Research Plan Page 7 of 40
1.2 Scope
This research plan has been developed by Fortescue Metals Group (Fortescue) to satisfy
Condition 10 of EPBC 2010/5696, which states that:
To compensate for any unavoidable impacts on habitat for the Night Parrot, a Night
Parrot Research Plan must be prepared and submitted to the Minister for approval within
12 months of the date of this approval. The research activities much be funded to a
value of no less than $1 million (GST Exclusive). The Night Parrot Research Plan must
include, but not be limited to:
a) funding of a suitably qualified expert/s to (undertake) research which contributes to
understanding the field ecology/biology of the Night Parrot. The research must
investigate and develop a repeatable survey methodology that provides for the detection
of the species
b) upon completion of the research identified by (condition 10 a) and no more than 4
years from the date of this approval, conduct or fund comprehensive targeted surveys
for the Night Parrot in at least 3 sites in areas where confirmed sightings or specimens of
Night Parrot have been recorded since 1 January 2000. The surveys must be informed
by the outcomes of the research identified at (condition 10 a) and undertaken by a
suitably qualified expert; and
c) indicative timeframes for funding, and completion of all research activities required
under the Night Parrot Research Plan. The Night Parrot Research Plan must include
timeframes for reporting the outcomes of the research.
The results of any research funded under the Night Parrot Research Plan must be
provided to the department within 12 months of completion. During the implementation
timeframe for the Night Parrot Research Plan, research results and annual progress of
research activities must be reported annually in the Environmental Performance Report
required at (condition 13). The approved Night Parrot Research Plan must be funded
and implemented.

Night Parrot Pezoporus occidentalis Research Plan Page 8 of 40
Table 1: How this plan meets the requirements of the Condition 10
Conditional requirement Related proposed activity Section (Page)
10 a) research which contributes to understanding the field ecology/biology of the Night Parrot, focusing on developing repeatable survey methods
1. Refine detection methods at known sites
2. Build preferred habitat model by describing landscape and habitat variables that are potentially important determinants of Night Parrot occupancy, including threatening processes
3. Combine detection methods and habitat model to survey new likely locations
2.1 (13)
2.2 (18) and 2.4 (25)
2.3 (22)
10 b) targeted surveys for the Night Parrot in at least 3 sites in areas where confirmed sightings or specimens of Night Parrot have been recorded since 1 January 2000
1. In addition to surveys described above, conduct systematic surveys using refined detection methods at 3 or more locations where recent, high veracity observations have been made.
2.3 (22)
10 c) indicative timeframes for funding, and completion of all research activities required under the Night Parrot Research Plan, including timeframes for reporting
1. See Budget Estimate
2. See Timeline Summary and Milestones
3. See Reporting Requirements
2.5 (28)
3.0 (29)
4.0 (33)
1.3 Development of the Research Plan
Steve Murphy has been engaged by Fortescue as the “suitably qualified expert” to prepare and
implement the Night Parrot Research Plan as defined under the condition above. Prior to this
engagement, Steve assisted John Young with implementing systematic research at the SW
QLD site so as not to miss the opportunities presented by that situation. Once approved, the
proposal presented here will form the basis of all research that occurs at the SW QLD site, and
John will be involved in its implementation.
In preparing this research plan, a Night Parrot Research Advisory Panel (RAP) was established
to provide advice and feedback. Members were selected based on their knowledge of the Night
Parrot, arid zone ecology (especially threatening processes), and the recovery planning
process. They were:
Allan Burbidge (RAP Chair; Principal Research Scientist, WA Department of Parks and
Wildlife)
Julian Reid (Fenner School of Environment and Society, Australian National University)
Leo Joseph (Director, Australian National Wildlife Collection, CSIRO)
Rachel Paltridge (Director, Desert Wildlife Services, Alice Springs)

Night Parrot Pezoporus occidentalis Research Plan Page 9 of 40
Stephen Garnett (Professor of Conservation and Sustainability, Research Institute for
the Environment and Livelihoods, Charles Darwin University)
Wolf Sievers (Director Threatened Species Unit, Queensland Department of
Environment and Heritage Protection).
The RAP met face to face in Perth on 21st October 2013 to discuss research priorities.
1.4 Support during the life of the research program
Given the uncharted nature of this work, and the high likelihood of unforseen changes in the
direction the work will take, it is suggested that an advisory group be developed that allows
experts to provide advice and feedback as the research plan is implemented. The RAP was
designed only to provide advice about the development of the plan and will cease to exist once
the plan is approved by the Commonwealth. Sometime after this, it may be optimal to form a
new advisory group (Night Parrot Recovery Team) that can provide the support for the research
program. Membership would include some or all RAP members along with other key
stakeholders (NRM groups, Indigenous groups, pastoralists etc.). The responsibility of
convening this may be best placed with the Commonwealth as it will be independent of
Fortescue’s Research Plan obligations.
1.5 General Approach in Developing the Plan
To better understand the biology and field ecology of the Night Parrot a three-year time frame is
proposed for completing this research program. Potential areas of research were drawn from
the Night Parrot Interim Recovery Plan (Blyth 1996), the Approved Conservation Advice for the
Night Parrot (Australian Government 2008), the Night Parrot Workshop held in Perth on
22/08/2013, and a long history of discussions among colleagues. These were prioritised
according to their importance in contributing to the species’ long-term survival, given the current
state of knowledge. The priority scale extends from “critical → high → nice to know → not
important”. The potential research areas and their priorities are shown in Table 2.
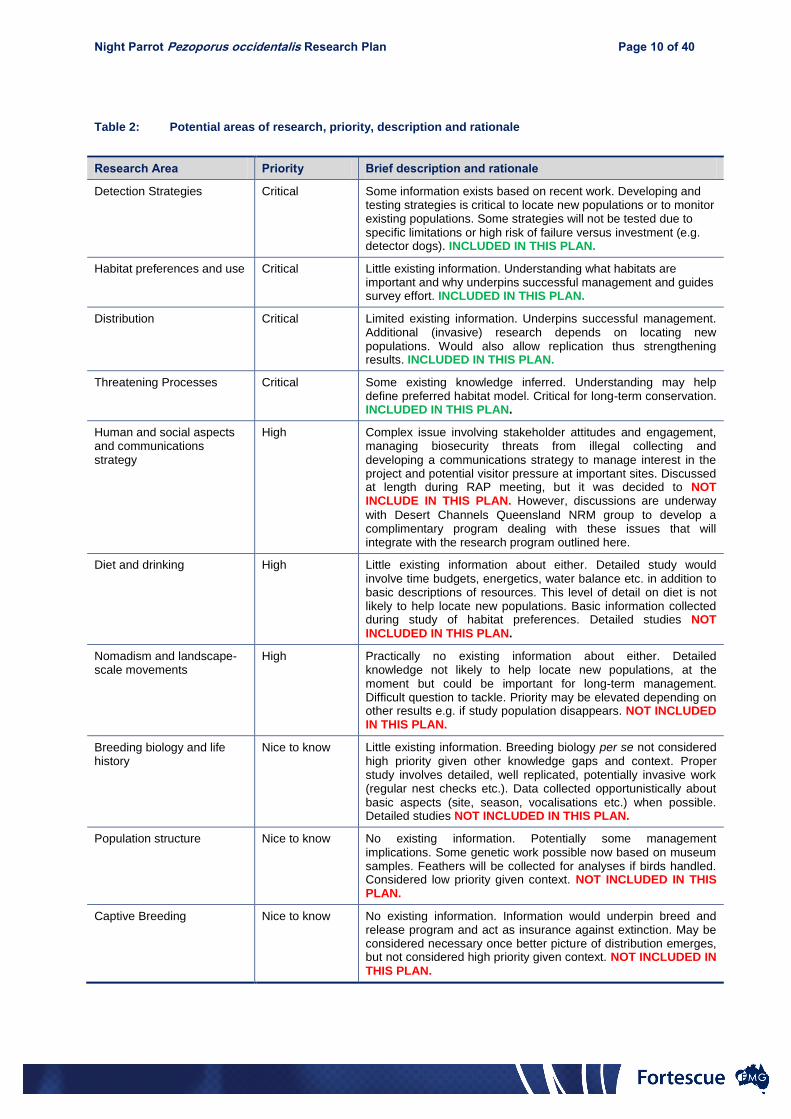
Night Parrot Pezoporus occidentalis Research Plan Page 10 of 40
Table 2: Potential areas of research, priority, description and rationale
Research Area Priority Brief description and rationale
Detection Strategies Critical Some information exists based on recent work. Developing and testing strategies is critical to locate new populations or to monitor existing populations. Some strategies will not be tested due to specific limitations or high risk of failure versus investment (e.g. detector dogs). INCLUDED IN THIS PLAN.
Habitat preferences and use Critical Little existing information. Understanding what habitats are important and why underpins successful management and guides survey effort. INCLUDED IN THIS PLAN.
Distribution Critical Limited existing information. Underpins successful management. Additional (invasive) research depends on locating new populations. Would also allow replication thus strengthening results. INCLUDED IN THIS PLAN.
Threatening Processes Critical Some existing knowledge inferred. Understanding may help define preferred habitat model. Critical for long-term conservation. INCLUDED IN THIS PLAN.
Human and social aspects and communications strategy
High Complex issue involving stakeholder attitudes and engagement, managing biosecurity threats from illegal collecting and developing a communications strategy to manage interest in the project and potential visitor pressure at important sites. Discussed at length during RAP meeting, but it was decided to NOT INCLUDE IN THIS PLAN. However, discussions are underway
with Desert Channels Queensland NRM group to develop a complimentary program dealing with these issues that will integrate with the research program outlined here.
Diet and drinking High Little existing information about either. Detailed study would involve time budgets, energetics, water balance etc. in addition to basic descriptions of resources. This level of detail on diet is not likely to help locate new populations. Basic information collected during study of habitat preferences. Detailed studies NOT INCLUDED IN THIS PLAN.
Nomadism and landscape-scale movements
High Practically no existing information about either. Detailed knowledge not likely to help locate new populations, at the moment but could be important for long-term management. Difficult question to tackle. Priority may be elevated depending on other results e.g. if study population disappears. NOT INCLUDED IN THIS PLAN.
Breeding biology and life history
Nice to know Little existing information. Breeding biology per se not considered high priority given other knowledge gaps and context. Proper study involves detailed, well replicated, potentially invasive work (regular nest checks etc.). Data collected opportunistically about basic aspects (site, season, vocalisations etc.) when possible. Detailed studies NOT INCLUDED IN THIS PLAN.
Population structure Nice to know No existing information. Potentially some management implications. Some genetic work possible now based on museum samples. Feathers will be collected for analyses if birds handled. Considered low priority given context. NOT INCLUDED IN THIS PLAN.
Captive Breeding Nice to know No existing information. Information would underpin breed and release program and act as insurance against extinction. May be considered necessary once better picture of distribution emerges, but not considered high priority given context. NOT INCLUDED IN THIS PLAN.

Night Parrot Pezoporus occidentalis Research Plan Page 11 of 40
When prioritising the nature and timing of research activities, the degree to which proposed
activities caused disturbance to individual Night Parrots or habitat was considered. At the time
of writing this plan, there is only one reliable location known that supports an unknown number
of birds. Invasive research methods that might cause birds to abandon the site cannot be
applied at this stage. Accordingly, an overarching principle of the plan is to first apply passive
methods before applying progressively more active and potentially invasive ones. At the
extreme end of this spectrum is handling individual Night Parrots and it is proposed to attempt
this only when additional occupied locations and populations are identified.
As discussed above, there exists a paucity of reliable information about nearly all aspects of
Night Parrot biology and ecology. Even though there is the opportunity to conduct some
systematic work at a site where Night Parrots can be reliably detected, the low sample size
means that a heuristic approach for much of this work is appropriate, especially initially. As
sample sizes increase, it may be possible to work with more robust statistical frameworks.
Finally, given that so little is known about Night Parrots and that this will be the first
systematically recorded information about their biology, it is not possible to fully or accurately
anticipate which direction various aspects of the research will head. Subsequently, what is
presented here is an initial program that is intended to answer research questions deemed to be
critical at this time. The program will remain flexible depending on findings and context, and the
yet-to-be-formed advisory group will play an important role in supporting this adaptability.
1.6 Program Framework
A schematic of how the research will be developed is provided in Figure 1. It illustrates how the
data collected (using non-invasive methods) at the single known site will be used to find
additional sites that will allow more invasive research to be conducted. It also shows how results
from the invasive research at additional sites will be used to refine knowledge of habitat
preferences and detection methods to ultimately locate more populations. Elements in the
research program presented here are inter-dependent and by applying this framework in an
iterative way, an ever-increasing level of refinement and confidence in the knowledge about
Night Parrots can be assured. The surveys that will be conducted to locate additional
populations will partly satisfy part 10(b) of the EPBC Condition (see ‘Background and
Relevance’ section of ‘Increasing Knowledge of Night Parrot Distribution’).
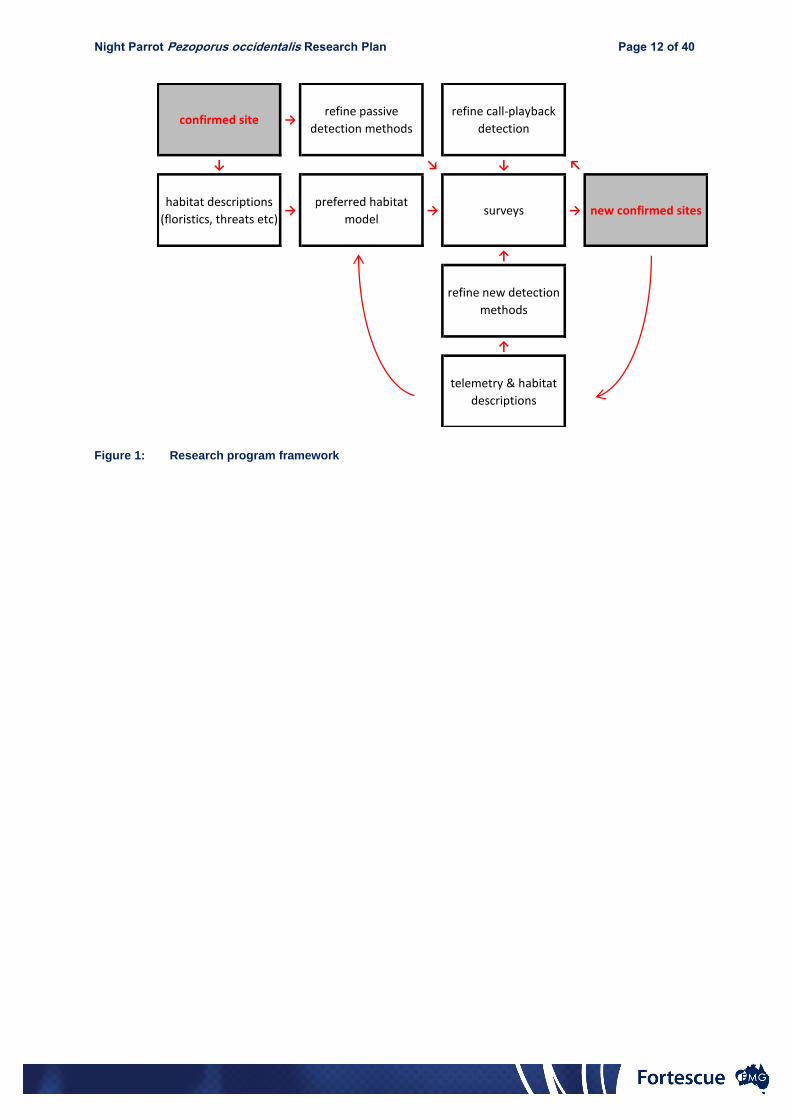
Night Parrot Pezoporus occidentalis Research Plan Page 12 of 40
Figure 1: Research program framework
confirmed site →refine passive
detection methods
refine call-playback
detection
↓ ↘ ↓ ↖
habitat descriptions
(floristics, threats etc)→
preferred habitat
model→ surveys → new confirmed sites
↑
refine new detection
methods
↑
telemetry & habitat
descriptions

Night Parrot Pezoporus occidentalis Research Plan Page 13 of 40
2. PROPOSED RESEARCH
2.1 Developing Detection Strategies for Night Parrots
2.1.1 Background and relevance
For cryptic species, such as the Night Parrot, developing robust, repeatable survey
methodology is critical for locating populations for active management and research. This is
also the primary intent of the Commonwealth’s Condition 10(a) that states the “research must
investigate and develop a repeatable survey methodology that provides for the detection of the
species.”
Night Parrots are difficult to detect, even when they are known, or suspected to be in an area
(Murphy 2012). Many attempts have been made to find Night Parrots using a variety of
techniques without success. However it is unknown whether the lack of detection is a failure of
the techniques used or whether Night Parrots were not present at the time surveys were
conducted. The SW QLD location provides a critically important opportunity to develop and test
detection strategies because there is a degree of certainty surrounding occupancy.
Until recently, there have been no recordings of Night Parrot calls and this has prevented the
development of useful survey techniques. Sound data recorded initially by John Young, and
then subsequently during preliminary systematic work at the SW QLD site, provides a promising
new detection tool. The research program proposes to invest heavily in testing and refining
sound-based detection strategies. This aligns with recommendations outlined in the Interim
Recovery Plan (Blyth 1996) and the Species Profile and Threat statement for Night Parrots
(Australian Government 2013).
Historically, the most common strategy employed for the Night Parrot was to survey at isolated
water during dry spells (Blyth and Boles 1999; Blyth et al. 1997a; Blyth et al. 1997b; Butler
1977; Maher 1995). This is an intuitive strategy, especially in the context of information
recorded by authorities who observed Night Parrots drinking (Andrews 1883; Bourgoin in Wilson
1937). However, given its limited success, the strategy needs to be tested systematically and it
is proposed to do this using camera traps. Additional detection methods may be developed and
tested once more information about Night Parrot habitat use and associated behaviours
emerges, such as active searches for feeding signs or tunnels through Triodia, camera traps set
is particular ways in specific feeding habitats etc.
2.1.2 How this relates to other parts of the research plan
Developing survey methods underpins most other aspects of this research proposal.
It will be combined with the ‘preferred habitat model’ (see below) to locate new
populations and thus refine knowledge of the species’ distribution

Night Parrot Pezoporus occidentalis Research Plan Page 14 of 40
Finding new populations will allow more invasive research methods (e.g. telemetry) to
be adopted
An understanding of detection probability is essential in the formulation of a credible
monitoring protocol.
2.1.3 Aims
To develop and measure quantitatively detection techniques on reliable populations, starting
with:
Sound-based detection, both passive and using call-playback
Camera traps in drinking, roosting and feeding habitats.
2.1.4 Detection using sound
Listening for bird calls is a widespread survey method (e.g. Allen et al. 2004; Burbidge et al.
2007; Kubel and Yahner 2007). For aural surveys to be conducted and interpreted properly, an
understanding of factors that influence detectability is required. These factors include
understanding the frequency of calling, whether detectability changes seasonally or due to other
factors (e.g. wind), the distance over which calls can be detected, and whether or not
detectability is increased in response to call-playback, and the context in which playback is
done (e.g. breeding season, distance of respondent to speaker etc.). Ultimately, some form of a
detection function is calculated using these and other variables that allows robust statistical
interpretation of survey results using distance sampling or occupancy modelling (Buckland et al.
2008; MacKenzie et al. 2006) – although see Welsh et al. (2013) for a discussion about the
vagaries of trying to adjust occupancy models to account for non-detection.
As mentioned elsewhere in this research plan, because of limited sample sizes it is necessary
to adopt a heuristic approach when developing and refining survey methods for Night Parrots.
At least initially, this will be an exercise in data collection and description, rather than being
strictly inferential. However, this will not preclude collecting quantitative data about factors that
affect detectability using Night Parrot calls.
Passive call detection using automated recording units
Recent advances in electronics and digital memory storage has allowed for the development of
commercially available products capable of autonomous operation for extended periods. In mid-
August 2013, 8 of these units were deployed at the SW QLD site to collect data while the
opportunity existed. ‘SM2+’ units were deployed (Wildlife Acoustics Inc., Massachusetts) with a
configuration that is power unlimited and capable of recording every night for 42 nights (i.e. they
are not triggered by target frequencies) before memory reaches capacity. Preliminary analyses
shows that these units are very capable of detecting Night Parrots. It is proposed to continue to
use multiple SM2+ units at sites known to be occupied to investigate the following:

Night Parrot Pezoporus occidentalis Research Plan Page 15 of 40
What is the full repertoire of Night Parrot calls?
What time of night do Night Parrots call?
How frequently do they call?
Over what distance can calls be detected, and under what conditions?
Does calling change according to environmental factors, such as rainfall?
Answers to these questions will directly inform the design of Night Parrot surveys. To answer
them adequately, it is proposed to maintain the 8 existing SM2+ units, with several more to be
permanently deployed in a set array at all sites where Night Parrots are confirmed. Rain and
wind logging instruments will be installed so these variables can be considered in calculating
detection probabilities. Recording will continue even if it appears that Night Parrots have
abandoned the site because seasonal occupancy of the site cannot be ruled out. Each unit will
be placed at pre-determined distances so the effect of distance on detectability can be
considered.
This high level of data collection requires a site visit approximately every 40 days to retrieve
data and replace memory cards. Such an extensive array of equipment recording more or less
continuously will generate a vast amount of data (130 gigabytes per SM2+ every 42 days).
‘Song Scope’ (Wildlife Acoustics Inc.) and ‘Sound ID’ (Boucher et al. 2008) software will be
trialled, which both employ an automatic sound-file processing function, based on sample of
reference calls. Preliminary use of Song Scope indicates that it is possible to detect Night Parrot
from sound files automatically, with about 80% success. As the library of reference calls
increases and the sound ‘recogniser’ algorithms are refined, it is expected that the performance
of this software will improve markedly.
Maintaining SM2+s at confirmed Night Parrot locations serves an additional benefit of
monitoring occupancy of sites (as distinct from collecting data about calling and/or testing this
as a detection method).
Call detection using call-playback
Call-playback is used commonly in bird surveys to elicit a vocal response thereby improving
detection probability. Gaining a proper understanding of the effect of playback on detection
probability requires knowledge of variables such as season, time of day/night, the distance of
the target bird to the speaker at the time the call is broadcast etc. (Allen et al. 2004; Buckland et
al. 2006; Kubel and Yahner 2007). With respect to speaker-respondent distance, investigation
of birds wearing radio transmitters will be considered, although only if there is certainty that it
will not unduly disrupt the main purpose of telemetry (i.e. investigating habitat preferences and
use).
Even if the distance between respondent and speaker in unknown, call-playback experiments
will still yield important information about detection probability. It is proposed that an automated
system be developed using a combination of SM2+s and custom-built automatic playback
devices. Broadly, this will involve broadcasting a Night Parrot call at realistic volume, time of

Night Parrot Pezoporus occidentalis Research Plan Page 16 of 40
night and frequency (based on preliminary data) over a 2 week period, with an SM2+ unit set up
to record responses. As with testing other detection methods, it will be important to conduct this
work at sites known to be occupied by Night Parrots. This system can also be used in an
opportunistic way for surveying new areas in the region from early-on in the life of the project
(see also section on Night Parrot distribution below).
To avoid excessive disturbance at the only known site, it is proposed to undertake call-playback
work only at sites that are additional to the currently known occupied site.
2.1.5 Detection using camera traps
Similar to sound equipment, the capabilities of infra-red triggered camera traps have improved
dramatically in recent years, with a concomitant reduction in price. It is now possible to
undertake extensive fauna surveys using this technology, and there are many examples of
programs that have successfully detected rare and/or cryptic bird species (reviewed in O'Brien
and Kinnaird 2008). There has also been an increasing amount of attention paid to developing
robust methods of analysing camera trap data (O'Brien and Kinnaird 2008; Rowcliffe et al. 2008
- see also the discussion below about estimating predator densities). While it may be a long-
term objective to apply such analytical frameworks to the Night Parrot data collected as part of
this research plan, initially (and in-line with the recurring heuristic theme), the focus will be on
developing methods for detection using camera traps on known birds.
As described previously in this section, surveying for Night Parrots at water points is an intuitive
and commonly adopted strategy that is intended to be investigated using the currently known
location in SW QLD. A combination of local knowledge, spatial data and ground searches will
be used to identify all surface water points within 10 km of the site. This approach aligns with
Andrews (1883) who described birds as flying “four or five miles” to water. Camera traps and
SM2 sound units will then be installed at each point in an attempt to detect Night Parrots. This
work will be repeated under dry and wet conditions to test whether there is a seasonal effect of
surface water dependency. It is possible that Night Parrots switch to a seed-based diet in wet
periods during which time they are dependent on surface water (inferred from comments in
Andrews 1883). During wet conditions there may be an issue of too many water points to
survey, in which case there will be a need to survey the transition period between wet and dry
when surveys become more feasible but while Night Parrots may still be feeding on seeds.
Another possibility is to attempt to detect Night Parrots at artificial water that is provided during
dry spells. However, caution will need to be applied if this is attempted due to the risk of
attracting mammalian predators to occupied sites. Further caution will be needed so that
important details about how Night Parrots obtain their water requirements naturally are not
obscured by providing artificial water supplies.
In addition to detection at water, we also propose to deploy camera traps among the day-time
roosting habitat at the known location to test this as a detection method. This would integrate
with the proposed study on assessing the potential impacts of mammalian predators at the site
(see below).

Night Parrot Pezoporus occidentalis Research Plan Page 17 of 40
Additional strategies using camera traps may become more apparent once Night Parrot habitat
use is better understood. For example, if birds feed in a predictable way in specific habitats,
then it may be possible to use camera traps to detect them.
2.1.6 Timeline and anticipated outcomes
Passive call detection
It is proposed to deploy SM2+ units for continual recording at all confirmed locations.
This is necessary to understand whether there are seasonal effects of call detection.
This work can begin immediately and, in fact, will be a continuation of the preliminary
work already being done. This will be repeated at all sites where Night Parrots are
confirmed to build adequate sample sizes for analyses. Consequently, this work will
continue throughout the life of the research project.
Call detection using play-back
The efficacy of call-playback will be tested using the systematic approach described
above once Night Parrots have been discovered at additional sites. The automated
call-playback system will be used opportunistically beforehand to assist in finding new
sites (see section on distribution).
Detection using camera traps
It is proposed to identify surface water points near the current SW QLD site and install
camera traps immediately, to take advantage of the present unseasonably dry
conditions in the region. Water points will be surveyed using camera traps when wet
conditions return, but at a time when surveying an adequate and appropriate sample of
water points is feasible (see above).
Camera traps in roosting habitat will be installed immediately at the SW QLD site. This
will integrate with work on assessing the potential impacts of mammalian predation.
This will be repeated once Night Parrots are detected at other sites.
New detection methods will be tested using camera traps once there is a clearer picture of
habitat use.
Upon completion of this component of the research project, survey methods for the Night Parrot
will have been designed, refined and tested. This will include the use of passive call detection
and call-playback, and various strategies using camera traps. Refinement and testing of
detection methods will occur at locations of known Night Parrot occupation. Therefore, there will
be opportunity to assess quantitatively the performance of various detection strategies. This
knowledge will be used to inform further surveys, both as part of this research program and also
by others conducting dedicated Night Parrot surveys as part of environmental impact
assessments. The knowledge will also be used to develop robust monitoring protocols that
could be used to assess the impact of management actions.

Night Parrot Pezoporus occidentalis Research Plan Page 18 of 40
2.2 Understanding Night Parrot habitat preferences and use
2.2.1 Background and relevance
Understanding species’ habitat preferences is central to understanding the biology and field
ecology of any species, and is critical for managing threatening processes. It also underpins the
selection of sites for surveys that aim to determine distribution. Both of these objectives are
highly relevant for the conservation of Night Parrots. It has particular relevance for habitat-
based assessments for the presence of Matters of National Environmental Significance during
development proposals, such as mining.
Effective habitat management also requires some understanding about what, specifically, Night
Parrots are doing in various habitat types and if there is a seasonal (temporal) component. This
includes the identification of:
Feeding habitat
Sites used for drinking (if indeed they do drink)
Breeding habitat
Roosting habitat
Temporal changes in each of these, according to seasonality or other factors
2.2.2 How this relates to other parts of the research plan
Understanding habitat preference and use relates to other parts of the proposed research
program. With respect to the components to be investigated, understanding habitat use relates
directly to:
Developing detection methods for Night Parrots, because some behaviours in some
habitats may offer new methods of detection (e.g. feeding in particular parts of the
landscape)
Increasing knowledge of Night Parrot distribution, because it will allow the identification
of areas with similar habitat for surveys
Understanding the threats to Night Parrots, because it enables better understanding of
the processes that may be specific to particular habitat types (e.g. grazing may impact
some habitats more than others). Particular threats may be important variables in
defining preferred Night Parrot habitat.
Though not a priority of this research plan, there may be an opportunity to collect basic
information about Night Parrot breeding. This would include nest site, season, recording
breeding-related vocalisations etc.

Night Parrot Pezoporus occidentalis Research Plan Page 19 of 40
2.2.3 Aims
To identify key habitats used by Night Parrots for feeding/drinking, roosting and
breeding
To describe these habitats in terms of vegetation structure and floristics, and position in
the landscape, in order to define Night Parrot habitat preferences
2.2.4 Identifying key habitats
To ensure the full range of habitats used by Night Parrots is understood, it is proposed to track
individuals using telemetry. This involves fixing small devices to individual birds so their location
can be recorded. While a lot can be gained from such studies, this is an invasive procedure that
carries potentially significant risks relating to individuals abandoning sites or sustaining injuries.
Therefore this component of the research will only be conducted once additional occupied sites
are located.
Capture
Standard mist-netting is a proven safe way of capturing birds (Spotswood et al. 2012). The
nominated lead researcher holds a current A-Class mist netting endorsement from the
Australian Bird and Bat Banding Scheme (Bander ID 2289, issued 1997) and has extensive
experience capturing small-medium sized parrots (e.g. Cyanoramphus spp., Psephotus spp.,
Neophema spp., Melopsittacus undulatus, Barnardius zonarius, Trichoglossus spp.). Handling
protocols will be streamlined to ensure restraint time is minimised. Handling will include
recording limited morphometric data (weight), collecting a small number of feathers for DNA
analyses, photography for aging and transmitter attachment.
Telemetry devices and attachment
There are two main types of telemetry devices potentially relevant for the study: (1) radio
telemetry units that transmit a signal that is followed in the field using hand-held receivers, and
(2) GPS units that store satellite-derived location data that is retrieved at a later date.
GPS tracking data is superior in that location information is accurate, regularly obtained and
collected without interference to the birds by field workers. The main downside is weight (the
smallest unit currently available weighs 2.5g). Most workers advocate that telemetry devices
should be no more than 2-5% of body weight, with recommendations tending towards 2% for
studies involvement individual movements (Barron et al. 2010; Murray and Fuller 2000). No
data exist for Night Parrot weight, although judging by their general size, they are not likely to be
more than 90-100g. This means that current GPS telemetry devices are probably not suitable to
use with Night Parrots (applying a 2% rule of thumb).
Given uncertainties about Night Parrot weight, how they move through their habitat (which
probably has implications for transmitter bulk), how they will respond behaviourally to wearing a
relatively large device and sensitivities surrounding their capture and handling, we propose to at

Night Parrot Pezoporus occidentalis Research Plan Page 20 of 40
least start the telemetry work using very small traditional VHF transmitting units. GPS units will
be considered for deployment at a later date once accurate weight data is collected and an
assessment is made of how individuals respond to wearing VHF transmitters. Key specifications
of the unit intended to be used are: 0.31g, 13-22 day lifespan (21 day nominal), and 8mm long x
5.3 mm wide x 2.8 mm high (http://www.holohil.com/lb2x.htm).
It is proposed to glue transmitters to trimmed feathers just above the synsacrum (on the back,
between the hips), which is the location least likely to affect flight performance or behaviour
because it is nearer the centre of gravity and does not interfere with the movement of the wings
(Raim 1978; Rappole and Tipton 1991). In a recent review comparing attachment methods,
dorsal gluing was the attachment method least likely to affect behaviour and breeding (Barron et
al. 2010).
Tracking
Studies on other nocturnal birds of similar size have attempted to locate individuals wearing
transmitters using triangulation from multiple fixed-point receiving stations (e.g. plains wanderer
Pedionomus torquatus (Baker-Gabb et al. 1990)). However, the error ellipses (up to several
hundred metres) produced by triangulation are likely to be too large for the purpose of this
(Murphy, unpubl. data). Additionally, as one of the research objectives is to determine why birds
are visiting particular parts of the landscape, it will be necessary to make direct observations.
Considering all this, it is proposed to track individuals on foot as much as possible while
transmitters remain on individuals. To avoid causing excess disturbance birds will be observed
from a distance using night vision equipment. Behaviours and associated details (e.g. feeding
and food items, drinking, social interactions, breeding etc.) will be recorded. Once birds have
moved away from their location, accurate location data will be recorded using handheld GPS
units, and associated details will be confirmed with close-up inspections.
There is the potential that locating birds wearing transmitters on foot will not be possible all of
the time. If birds cannot be located after two nights, it is proposed to locate them from the air
using a helicopter using suitably qualified, instrument-rated pilots .
Analyses of telemetry data
In the first instance, the tracking data will simply be used to identify those habitats used by Night
Parrots. As the number of recorded point locations increases, it will be possible to answer
questions relating to home-range size and the relative importance of areas using kernel density
estimation. Assuming individuals will carry transmitters for the nominal 21-day period at least
four fixes each night (separated by at least one hour) and two each day will be attempted to
reach the minimum number of fixes required for analysis (a minimum of 30 per individual but
preferably >50, Seaman et al. 1999). Refining home range estimates by considering movement
behaviour, and barriers and/or areas that are never traversed will also be attempted (Benhamou
and Cornélis 2010).

Night Parrot Pezoporus occidentalis Research Plan Page 21 of 40
2.2.5 Habitat descriptions and preferences
To describe habitats, National Vegetation Information System Level V protocols will be used,
because this is the national standard and will ensure habitat data are consistent and
comparable across state boundaries. Level V includes collecting information about dominant
growth form, height, cover and species (3 species) for the three traditional strata i.e. Upper, Mid
and Ground (Executive Steering Committee for Australian Vegetation Information 2003) .
Additionally, information will be collected that is specific to Night Parrots that may not be part of
the standard NVIS Level V protocols, such as size and spaces between Triodia clumps and a
measure of greenness in key species (e.g. chenopods).
During habitat surveys, information about potential threatening processes will also be collected
(e.g. herbivore and predator scat recording and collection). Habitat descriptions at the same or
similar times that Night Parrots are detected will be conducted to assess potential resource
availability i.e. habitat assessments will not be undertaken during wet periods if birds were
detected during dry spells.
Defining Night Parrot habitat preferences (i.e. building a preferred habitat model) will
incorporate both qualitative and quantitative variables. Some variables that are likely to be
important will be investigated in other parts of the plan – namely, threatening processes (fire,
grazing history etc.). Broadly, the variables that are likely to be used in describing preferred
habitat are:
Floristics and structure of roosting, feeding and breeding habitats
Spatial configuration of these habitat types
Fire history of the site and the availability of long-term fire refugia
The effect of season on habitat use
Grazing impacts in particular habitats
2.2.6 Timeline and anticipated outcomes
The capture and tracking of individual Night Parrots using telemetry will only be
attempted once new confirmed sites are identified.
Habitat assessments will be conducted at known sites soon after discovery. For the
currently occupied site this will be undertaken as soon as possible. These data will be
incorporated into understanding preferred Night Parrot habitat in an iterative way (i.e.
will be continually refined as new data are recorded).
This section will define where Night Parrots spend their time and why, in terms of vegetation
floristics and structure, and position in the landscape. It will form the basis of defining preferred
Night Parrot habitat, which will underpin site selection for subsequent surveys and increase
understanding of the potential threatening processes.

Night Parrot Pezoporus occidentalis Research Plan Page 22 of 40
2.3 Increasing Knowledge of Night Parrot Distribution
2.3.1 Background and relevance
At present only one site occupied by Night Parrots is known, which is in SW QLD. Together with
the discoveries of dead birds in 1990 and 2006 (Boles et al. 1994; McDougall et al. 2009), this
suggests an extant population in this region, perhaps extending into north-eastern South
Australia. There is a consistent record of Night Parrot sightings from the Pilbara region, Western
Australia, suggesting that there may also be an extant population there (Murphy 2012). In
contrast, Night Parrots seem to have retracted from the southern portion of their former range
(Murphy 2012).
It is critical that the knowledge about the distribution of Night Parrots is refined because this
underpins practically all aspects of conservation management and research. This importance is
reflected in Condition 10(b) of EPBC 2010/5696, which states:
“upon completion of the research identified by (condition 10 a) and no more than 4 years
from the date of this approval, conduct or fund comprehensive targeted surveys for the
Night Parrot in at least 3 sites in areas where confirmed sightings or specimens of Night
Parrot have been recorded since 1 January 2000. The surveys must be informed by the
outcomes of the research identified at (condition 10 a) and undertaken by a suitably
qualified expert.”
Exactly how these surveys integrate into the research plan was discussed at the Night Parrot
Research Advisory Panel meeting in October 2013, where it was unanimously agreed that the
surveys should be integrated into the research plan rather than delaying until the research has
been completed. However, it is important to note that it is not proposed to conduct surveys only
at locations where Night Parrot sightings have been made by third parties. Areas will also be
selected on the basis of current knowledge of preferred habitats. The benefits of this approach
are that:
Conducting surveys will allow us to test learnings about preferred Night Parrot habitat.
Similarly, surveys in new areas may be a good test of the detection strategies
Following up on high veracity sightings quickly, rather than after the research is completed,
increases the chances of detecting Night Parrots
2.3.2 How this relates to other parts of the research plan
As mentioned above, conducting surveys to determine distribution will test detection strategies
and understanding of preferred habitat. Also, identifying additional locations will allow some of
the more invasive aspects of this proposal to be undertaken, and will increase the opportunity to
increase samples sizes, generally. This will allow transition from a heuristic research approach
to one based on more rigorous, inferential analyses.

Night Parrot Pezoporus occidentalis Research Plan Page 23 of 40
2.3.3 Aims
To combine knowledge of detection methods and habitat preferences to locate
additional Night Parrot populations both within SW QLD and interstate.
To follow-up high veracity, third-party sightings during the research program using
refined detection methods, thus satisfying Condition 10(b) of EPBC 2010/5696.
2.3.4 Finding additional Night Parrot populations
As mentioned elsewhere, there will be no attempt to conduct invasive research at the confirmed
occupied site. Additional sites will need to be found before some critical information can be
collected that will improve understanding about the best ways to search for Night Parrots. The
circularity of this situation may not be as constraining as it first appears because two important
tools are available to locate additional sites immediately. Firstly Night Parrots are known to roost
in long unburnt Triodia sp. Secondly, there is evidence that Night Parrots call relatively
frequently in the first two hours after sunset from these roosting areas, and that this call can be
detected using automated recording devices. Related to this, methods have already been
developed to screen these sound data automatically for Night Parrot calls.
To identify long unburnt Triodia (and indeed other habitats as they become known) a
combination of on-ground site inspections and spatial analyses will be used. The latter will allow
systematic identification of likely areas to survey using supervised classification of multispectral
LANDSAT imagery. Generally, this involves mapping sample areas of known habitat, and then
classifying an entire image based on these ‘training areas’. Subsequently, field data are
normally collected to verify and refine the classification. This approach has been applied
commonly in similar situations (Debinski et al. 1999; Gottschalk et al. 2005; Grace 2003;
Leyequien et al. 2007; Saxon 1983; Wallin et al. 1992).
One potential problem is that it may not be possible to identify the small habitat patches that are
important to Night Parrots using LANDSAT (30m resolution). In this case, suitable habitats will
be mapped from other sources, such as Google Earth imagery or archived aerial photographs.
This has also been used successfully in other situations (Franklin and Steadman 1991; Harvey
and Hill 2001; Lauver et al. 2002; Robertson et al. 1990).
Using these tools, surveys of new likely areas can commence immediately and will be
maintained throughout the life of the project with an ever-increasing level of refinement about
when, where and how to look, as information becomes available.
2.3.5 Following-up third party sightings
Areas with previous Night Parrot sightings of high veracity may also be surveyed. Preliminary
analyses of a relatively large database of Night Parrot sightings suggests that there is a high
probability of misidentifying a Night Parrot (Murphy et al. 2009). As part of that analysis, a
protocol was developed to give individual sightings a score related to how likely the observation

Night Parrot Pezoporus occidentalis Research Plan Page 24 of 40
was of a Night Parrot. The protocol used elements of the observation, such as the physical
description of the bird and the habitat, and details about the observer and the conditions under
which the observation was made. It is proposed to adapt this scoring protocol to assess whether
or not effort should be invested in following up on new third-party sightings. It is also proposed
to assess the habitat based on knowledge assembled during this research. Once a decision to
follow up a sighting has been made, new detection strategies that have been developed will be
applied.
An example of a location where high veracity Night Parrot sightings have been made recently is
Lorna Glen – a conservation reserve in Western Australia managed by the Department of Parks
and Wildlife (Burbidge and Hamilton 2013). A Night Parrot sighting was reported by DPaW (then
DEC) staff in 2009, and despite follow-up surveys, no further evidence has been discovered. As
part of this proposal, it is intended to collaborate with DPaW staff to apply some of the detection
and analytical methods being developed as part of this project at Lorna Glen in 2014.
2.3.6 Timeline and anticipated outcomes
As soon as possible, opportunistic surveys will be conducted using the tools already
developed in likely areas identified by on-ground inspection and spatial analyses.
As knowledge of detection strategies and habitat preferences improves, a more refined and
systematic approach in the selection of survey sites will be applied.
The main outcome of this component of the research plan will be completed surveys in areas
that (a) have been identified systematically using spatial analyses and (b) have been identified
from Night Parrot sightings from third parties. This will satisfy point 10(b) of the EPBC Condition.
If surveys are successful in detecting Night Parrots, it will allow other, more invasive parts of the
research plan to proceed. It will also potentially increase the sample size of some aspects of the
research, therefore allowing a shift toward more rigorous inferential analyses.
2.4 Understanding the threats to Night Parrots
2.4.1 Background and relevance
Based on historical accounts suggesting high densities of Night Parrots at some places and
times (Andrews 1883; Ashby 1924) and the lack of confirmed sightings from about 1912
(Murphy 2012), it seems clear that the species has suffered a major range contraction and/or
reduction in population density in most places. Over the same period and in the same general
area, a suite of medium-sized mammals became either wholly or functionally extinct, with
changed fire patterns and introduced predators and herbivores identified as the main drivers
(Burbidge et al. 1988).

Night Parrot Pezoporus occidentalis Research Plan Page 25 of 40
It is intuitive that these same threatening processes, acting either alone or synergistically,
probably affected Night Parrots, and there is some anecdotal or circumstantial evidence that
supports this. Recent work in SW QLD and high veracity historical records demonstrate clearly
that Night Parrots rely on long-unburnt Triodia spp. Analyses of contemporary fire patterns in
central Australia show that Triodia is prone to large-scale disturbance by mismanaged fire
(Edwards et al. 2008) and this is most likely impacting on Night Parrots. This is not to advocate
fire exclusion, rather that there will be optimal fire patterns that maintain Night Parrot habitat.
Night Parrots appear to rely on chenopod systems (although it is not known exactly why), and
other birds dependent on these systems have declined due to grazing pressure (e.g. some
Grasswrens – reviewed in Skroblin and Murphy (2013)). Anecdotal evidence also suggests that
feral and domestic cats prey upon Night Parrots (Ashby 1924; McGilp 1931; Whitlock 1924). In
one case, a single domestic cat is thought to have been responsible for killing several Night
Parrots (“several picture frames were covered by the wings and tails of the ‘Porcupine Parrot’
which had been caught by a cat last summer” (Ashby 1924, p. 179). Also, Keartland claims to
have frequently found “the remains of those (Night Parrots) recently killed by dingoes…”
(quoted in Ashby 1924, p. 179). Although there are no reports of foxes (Vulpes vulpes) preying
upon Night Parrots, it is a reasonable proposition that they do so.
With regards to dingoes, irrespective of whether they are classed as native or not, they occupy
an important ecological position that is potentially relevant to Night Parrots. It is possible that
dingoes prey upon Night Parrots, although there is considerable evidence to suggest that
dingoes regulate cat and fox populations (Glen et al. 2007; Kennedy et al. 2012; Letnic et al.
2011), and could reduce the potential impact of these predators.
Clarifying the potential for these threats to impact Night Parrots and understanding their
dynamics (e.g. optimal fire patterns) is important for long-term management.
2.4.2 How this relates to other parts of the research plan
Understanding processes that may be threatening Night Parrots relies on identifying what parts
of the landscape Night Parrots are using. The presence of threatening processes and history of
fire patterns are likely to be important variables in defining preferred habitat.
2.4.3 Aims
To investigate the fire history at areas occupied by Night Parrots
To assess the potential impacts of predation by invasive species (cats/foxes) on Night
Parrots
To assess the potential impacts of grazing on Night Parrot habitat

Night Parrot Pezoporus occidentalis Research Plan Page 26 of 40
2.4.4 Fire history of Night Parrot habitats
It is proposed to create a fire history for areas occupied by Night Parrots, using the free
LANDSAT image archive (www.glovis.usgs.gov ). Images extending back to the mid-1970s will
be processed to highlight and map fire scars using supervised classification techniques in
remote sensing and GIS software (e.g. ER Mapper/Imagine and ArcMap). It is anticipated that
fire frequency will be quite low, and this process may not be as arduous as it first appears.
Vectorised fire scars will be attributed with date and image ID.
Importantly, the fire history will be extended into the area surrounding occupied sites in order to
better understand fire patterns at a landscape scale. This is necessary to investigate the
importance of fire refugia when core habitat areas are disturbed. In a sense, it is testing a
hypothesis that Night Parrots now only occur where landscape features ensure reliable long-
term fire refugia. This information will be incorporated into understanding Night Parrot habitat
preferences developed during other parts of the research plan.
2.4.5 Potential impacts of mammalian predators on Night Parrots
It is unlikely that the level of cat or fox predation on Night Parrots can be assessed directly.
Instead, it is proposed to assess the density of these predators (and dingoes) in the habitats
that are being used by Night Parrots, as defined by the section on habitat preferences and use.
In doing so, it will enable the potential impacts of these species on Night Parrots to be
estimated.
Assessing mammalian predator density is fraught with issues such as low densities (and thus
low statistical signals) and violations of non-independent sampling for some commonly used
methods such as counting foot prints along roadways (Wilson and Delahay 2001). Additionally,
the rocky surfaces in and around the SW QLD study area preclude the use of track-based
methods, and to import sand etc. for plots is not recommended due to the risk of inadvertently
attracting predators to the area. Instead, it is proposed to use arrays of passive infra-red
triggered cameras. This approach has been applied successful in estimating feral cat densities
using a capture-mark-recapture analytical framework where individuals were identifiable from
coat patterns (Bengsen et al. 2012). Camera traps have also been used to estimate densities
without relying on individual identification, and it is proposed to use one such method developed
by Rowcliffe et al. (2008). They combined information about individual daily movements and
group size, with camera-trap specifications relating to detection distance and angle to provide
reliable density estimates. Reliable information exists about daily movements for cats, foxes and
dingoes in arid environments (Edwards et al. 2001; Marsack and Campbell 1990; Molsher et al.
2005; Moseby et al. 2009b; Thomson 1992). Assumptions of the approach are that (i) the
movements of individuals with respect to conspecifics and landscape features is random and
independent (although the model was robust to some violation of this) (ii) animal movements
are largely independent of the cameras and (iii) the population is closed (i.e. migration,
recruitment and mortality are negligible).

Night Parrot Pezoporus occidentalis Research Plan Page 27 of 40
Most studies of predator activity using camera traps employ attractant lures to increase the
probability of detection. In the context of this research proposal, the use of lures risks attracting
predators to occupied sites and therefore it is proposed to use camera traps without lures. If the
statistical signal is too low without lures, an attempt will be made to determine predator
density/activity and the potential impacts on Night Parrots by systematic scat surveys within 2
ha area (Moseby et al. 2009a). Scats will be analysed using standard protocols (Paltridge
2002). Although more labour-intensive, such surveys have been used previously to investigate
predator-prey dynamics successfully (Paltridge 2002).
2.4.6 Potential impacts of grazing on Night Parrot habitat
Observations in the only habitat known to be used by Night Parrots suggest that grazing
impacts are low and are likely to remain low given the nature of the habitat (long-unburnt
Triodia). However, once a clearer picture of the full range of habitats used by Night Parrots
emerges from other components of this research plan, the potential impacts of grazing can be
better assessed. This is proposed by generating an index of grazing pressure based on
herbivore (cattle, horse, camel, rabbit and macropod) scat densities recorded during habitat
descriptions (see above). Alternative quantitative measures of grazing impacts using remote
sensing (LANDSAT) data will be considered for some areas where grazing pressure seem to be
particularly important. (Bastin et al. 1993). Given that the LANDSAT archive extends back to the
mid-1970s, this opens the possibility of examining historical grazing pressure also.
2.4.7 Timeline and anticipated outcomes
A fire history using archived LANDSAT imagery for all sites known to be used by Night
Parrots will be created. This will begin immediately for the currently occupied site in SW
QLD.
It is proposed to install camera traps at the SW QLD site as soon as possible, and at other
sites as they become known.
Data on potential herbivore impacts will be collected during the habitat descriptions of
confirmed sites. This will be done as soon as possible for the currently known SW QLD site.
The combined elements of this part of the research project will provide estimates of the level of
potential threat in various habitat types posed by feral predators and grazing animals. It will also
describe the fire history in and around sites occupied by Night Parrots, which will inform both
the optimal habitat model and practical fire management guidelines.

This page, which includes budget details, has been removed from this version of the Research Plan

Night Parrot Pezoporus occidentalis Research Plan Page 29 of 40
3. TIMELINE SUMMARY AND MILESTONES
As discussed, several key parts of this research plan rely on finding additional populations for
study. There is some risk associated with not being able to do this, although the probability of
finding additional occupied sites is considered very high. However, if after 12 months of
comprehensive surveys in likely habitats in the region no additional birds have been found,
contingency plans will be discussed with the yet-to-be-formed advisory group. A likely
contingency plan will be attempting to do some of the invasive research (e.g. call-playback
work) at the known site. If birds vacate the known site, investment in identifying areas of
suitable habitat and conducting surveys will be increased.
Given the uncertainty around when additional populations will be located that will enable several
key parts of the proposed research to proceed, it is difficult to be specific about the timing of
much of the proposal. Figure 2 shows a predicted timeline considering this uncertainty.
Figure 3 shows proportion of time analysing various parts of the proposed research plan. The
sound data is anticipated to take up 40% of analysis time, possibly more in the first year. It is
also anticipated that the ratio of field work to analyses and project management is 30:70.
Additionally, as a result of uncertainty it is not possible to provide precise dates for milestones
for the full duration of the research program. Having said this, the following indicative milestones
are proposed (assuming the program can commence in January 2014):
June 30, 2014:
1. All permits (including ethics) for invasive research granted (or at least submitted)
2. Methods for automated sound recording refined or at least at an advanced stage of
development
3. Preliminary analyses of passive call detection data collected to date completed
4. Preliminary analyses of camera trap data collected to date completed (both for threatening
processes and Night Parrot detection)
5. Habitat assessment of known site completed
6. Fire history of known site completed
7. At least 5 new areas identified from aerial photos and surveyed using SM2 units, and data
analysed
December 15, 2014:
8. Preliminary analyses of passive call detection data collected during subsequent sampling
period completed
9. Preliminary analyses of camera trap data completed (both for threatening processes and
Night Parrot detection)

Night Parrot Pezoporus occidentalis Research Plan Page 30 of 40
10. At least 5 new areas identified from aerial photos and surveyed using SM2 units, and data
analysed
11. Annual report completed presenting progress to date.
Night Parrot Pezoporus occidentalis Research Plan Page 31 of 40
CB-RP-EN-1048
Rport
Figure 2: Proposed timeline.
NB: Green blocks indicate estimated timing of those activities that are not dependent on finding Night Parrots at additional locations.
jan feb mar apr may jun jul aug sep oct nov dec jan feb mar apr may jun jul aug sep oct nov dec jan feb mar apr may jun jul aug sep oct nov dec
passive call detection permanent monitoring at all confirmed locations
detection using call-playback as soon as Night Parrots detected at additional locations, then ongoing until adequately sampled
detection using camera traps at currently known site until adequately sampledthen, as additional information about habitat use becomes known until adequately sampled
identifying key habitats using telemetry as soon as Night Parrots detected at additional locations, then ongoing until adequately sampled
habitat descriptions then, as additional sites become known
identify & survey likely areas ongoing, once we begin to define preferred habitat model
follow-up third party sightings as high veracity sightings become known
Fire history of habitats at known site then, as additional sites become known
Potential impacts of mammalian predators at known site, until adequately sampled then, as additional sites become known
Potential impacts of grazing then, as additional sites become known
2015 2016
Part Research activity
"
2014
Detection
Habitat preferences
Distribution
Threatening processes
Night Parrot Pezoporus occidentalis Research Plan Page 32 of 40
CB-RP-EN-1048
Rport
Figure 3: Projected proportion of time analysing various research components
sound habitat mapping fire scar mapping
camera trap telemetry habitat descriptions

Night Parrot Pezoporus occidentalis Research Plan Page 33 of 40
4. REPORTING REQUIREMENTS
As described in Condition 10 c), annual reporting will be provided to the Department of the Environment in December of each year of project implementation. Reporting will refer specifically to the research components outlined in this plan conducted in that year and include:
Number of survey nights using passive and call-playback sound detection, and camera traps, and results of these surveys
Number of areas for which detailed habitat surveys have been conducted, and the results of these surveys. This will include details about threatening processes potentially operating at each site.
Number of individual Night Parrots followed using radio telemetry, the number of tracking days and the results of these investigations
Preliminary discussion and conclusions drawn from Night Parrot ecology and survey/monitoring procedures
Preliminary maps of potential suitable habitat in key regions, based on knowledge of preferred habitat.
Also required under Condition 10 c) is a final report detailing results of the full and completed research plan, within 12 months of project completion. According, full analyses and discussion of results of all parts of the research plan will be submitted to the Department of the Environment no later than December 2017. It is anticipated that these results will also form the basis of a series of peer-reviewed scientific publications.

Night Parrot Pezoporus occidentalis Research Plan Page 34 of 40
CB-RP-EN-1048
Rport
5. REFERENCES
Allen, T., Finkbeiner, S.L., and Johnson, D.H. (2004) Comparison of detection rates of breeding
marsh birds in passive and playback surveys at Lacreek National Wildlife Refuge, South
Dakota. Waterbirds 27, 277-281.
Andrews, F.W. (1883) Notes on the Night Parrot. Transactions and Proceedings of the Royal
Society of South Australia 6, 29-30.
Ashby, E. (1924) Notes on extinct or rare birds, with suggestions as to some of the causes of
their disappearance. Emu 23, 178-183.
Australian Government (2008) Approved Conservation Advice for Pezoporus occidentalis (Night
Parrot).
Australian Government (2013) Species Profile and Threats Database. Department of the
Environment http://www.environment.gov.au/cgi-
bin/sprat/public/publicspecies.pl?taxon_id=59350#survey_guidelines accessed 30 October
2013
Baker-Gabb, D.J., Benshemesh, J.S., and Maher, P.N. (1990) A revision of the distribution,
status and management of the Plains-wanderer Pedionomus torquatus. Emu 90, 161-168.
Barron, D.G., Brawn, J.D., and Weatherhead, P.J. (2010) Meta‐analysis of transmitter effects on
avian behaviour and ecology. Methods in Ecology and Evolution 1, 180-187.
Bastin, G.N., Pickup, G., Chewings, V.H., and Pearce, G. (1993) Land degradation assessment
in central Australia using a grazing gradient method. The Rangeland Journal 15, 190-216.
Bengsen, A., Butler, J., and Masters, P. (2012) Estimating and indexing feral cat population
abundances using camera traps. Wildlife Research 38(8), 732-739.
Benhamou, S., and Cornélis, D. (2010) Incorporating Movement Behaviour and Barriers to
Improve Kernel Home Range Space Use Estimates. Journal of Wildlife Management 74(6),
1353-1360.
Blyth, J. (1996) Night parrot (Pezoporus occidentalis) interim recovery plan for Western
Australia 1996-1998. No. Interim Recovery Plan No. 4, Wanneroo, W.A.
Blyth, J., and Boles, W. (1999) An expedition to the Murchison, Gascoyne and East Pilbara
areas to search for Night Parrots, November 1997. Eclectus 6, 12-16.
Blyth, J., Burbidge, A., and Boles, W. (1997a) Another Night Parrot Search. Western Australian
Bird Notes 81, 13.

Night Parrot Pezoporus occidentalis Research Plan Page 35 of 40
Blyth, J., Burbidge, A., and Boles, W. (1997b) Report on an expedition to the Western Desert
and East Pilbara areas in search of the Night Parrot Pezoporus occidentalis. Eclectus 2, 25-30.
Boles, W.E., Longmore, N.W., and Thompson, M.C. (1994) A recent specimen of the Night
Parrot Geopsittacus occidentalis. Emu 94, 37-40.
Boucher, N.J., Burbidge, A.H., and Jinnai, M. (2008) Computer recognition of sounds that have
never been heard before. In '18th National Australian Institute of Physics Conference.' Adelaide)
Buckland, S.T., Marsden, S.J., and Green, R.E. (2008) Estimating bird abundance: making
methods work. Bird Conservation International 18(S1).
Buckland, S.T., Summers, R.W., Borchers, D.L., and Thomas, L.E.N. (2006) Point transect
sampling with traps or lures. Journal of Applied Ecology 43, 377-384.
Burbidge, A. (2013) Night Parrot sightings at Lorna Glen, WA. In 'Night Parrot Workshop,
Department of Parks and Wildlife.' Kensington, WA)
Burbidge, A.A., Johnson, K.A., Fuller, P.J., and Southgate, R.I. (1988) Aboriginal knowledge of
the mammals of the central deserts of Australia. Wildlife Research 15, 9-39.
Burbidge, A.H., Rolfe, J., McNee, S., Newbey, B., and Williams, M. (2007) Monitoring population
change in the cryptic and threatened Western Ground Parrot in relation to fire. 107, 79-88.
Butler, H. (1977) 'In The Wild.' (Australian Broadcasting Commission: Sydney)
Debinski, D., Kindscher, K., and Jakubauskas, M. (1999) A remote sensing and GIS-based
model of habitats and biodiversity in the Greater Yellowstone Ecosystem. International Journal
of Remote Sensing 20, 3281-3291.
Edwards, G.P., Allan, G.E., Brock, C., Duguid, A., Gabrys, K., and Vaarzon-Morel, P. (2008)
Fire and its management in central Australia. The Rangeland Journal 30, 109-121.
Edwards, G.P., De Preu, N., Shakeshaft, B.J., Crealy, I.V., and Paltridge, R.M. (2001) Home
range and movements of male feral cats (Felis catus) in a semiarid woodland environment in
central Australia. Austral Ecology 26, 93-101.
Franklin, J., and Steadman, D.W. (1991) The potential for conservation of Polynesian birds
through habitat mapping and species translocation. Conservation Biology 5(4), 506-521.
Glen, A.S., Dickman, C.R., SoulÉ, M.E., and Mackey, B.G. (2007) Evaluating the role of the
dingo as a trophic regulator in Australian ecosystems. Austral Ecology 32, 492-501.
Gottschalk, T., Huettmann, F., and Ehlers, M. (2005) Review article: Thirty years of analysing
and modelling avian habitat relationships using satellite imagery data: a review. International
Journal of Remote Sensing 26, 2631-2656.

Night Parrot Pezoporus occidentalis Research Plan Page 36 of 40
Grace, L.P. (2003) Mapping Bald Eagle Communal Night Roost Habitat in Northwest
Washington Using Satellite Imagery. Western Washington University,
Harvey, K., and Hill, G. (2001) Vegetation mapping of a tropical freshwater swamp in the
Northern Territory, Australia: a comparison of aerial photography, Landsat TM and SPOT
satellite imagery. International Journal of Remote Sensing 22, 2911-2925.
Kennedy, M., Phillips, B.L., Legge, S., Murphy, S.A., and Faulkner, R.A. (2012) Do dingoes
suppress the activity of feral cats in northern Australia? Austral Ecology 37, 134-139.
Kubel, J.E., and Yahner, R.H. (2007) Detection probability of Golden-winged Warblers during
point counts with and without playback recordings. Journal of Field Ornithology 78, 195-205.
Lauver, C.L., Busby, W.H., and Whistler, J.L. (2002) Testing a GIS model of habitat suitability
for a declining grassland bird. Environmental Management 30, 88-97.
Letnic, M., Greenville, A., Denny, E., Dickman, C.R., Tischler, M., Gordon, C., and Koch, F.
(2011) Does a top predator suppress the abundance of an invasive mesopredator at a
continental scale? Global Ecology and Biogeography 20, 343-353.
Leyequien, E., Verrelst, J., Slot, M., Schaepman-Strub, G., Heitkönig, I.M.A., and Skidmore, A.
(2007) Capturing the fugitive: Applying remote sensing to terrestrial animal distribution and
diversity. International Journal of Applied Earth Observation and Geoinformation 9, 1-20.
MacKenzie, D.I., Nichols, J.D., Royle, J.A., Pollock, K.H., Bailey, L.L., and Hines, J.E. (2006)
'Occupancy Estimation and Modeling: Inferring Patterns and Dynamics of Species Occurrence.'
(Elsevier: San Diego, California)
Maher, P. (1995) Night Parrot Survey, Techniques and Habitat Requirements with Notes on the
Fauna of the Winton and Cloncurry Districts. A Report to the Queensland Department of
Environment and Heritage. Deniliquin.
Marsack, P., and Campbell, G. (1990) Feeding-Behavior and Diet of Dingoes in the Nullarbor
Region, Western-Australia. Wildlife Research 17, 349-357.
Mayfield, H. (1975) Suggestions for calculating nest success. The Wilson Bulletin 87, 456-466.
McDougall, A., Porter, G., Mostert, M., Cupitt, R., Cupitt, S., Joseph, L., Murphy, S., Janetzki,
H., Gallagher, A., and Burbidge, A. (2009) Another piece in an Australian ornithological puzzle _
a second Night Parrot is found dead in Queensland. Emu 109, 198-203.
McGilp, J.N. (1931) Geopsittacus occidentalis, Night-Parrot. South Australian Ornithologist 11,
68-70.
Molsher, R., Dickman, C., Newsome, A., and Müller, W. (2005) Home ranges of feral cats (Felis
catus) in central-western New South Wales, Australia. Wildlife Research 32, 587-595.

Night Parrot Pezoporus occidentalis Research Plan Page 37 of 40
Moseby, K.E., Stott, J., and Crisp, H. (2009) Movement patterns of feral predators in an arid
environment-implications for control through poison baiting. Wildlife Research 36, 422-435.
Murphy, S. (2012) Independent expert advice on the likely impacts of the proposed Cloudbreak
Iron Ore Expansion Project (EPBC 2010/5696) on the EPBC Act listed endangered and
migratory Night Parrot (Pezoporus occidentalis). Map IT, Malanda.
Murphy, S., Burbidge, A.H., Joseph, L., McAllan, I.A.W., Venables, W., and King, E. (2009)
Wanted: a bigger needle or a smaller haystack. Explaining and predicting the occurrence of
Night Parrots in a vast landscape. In 'Fifth Biennial Australasian Ornithological Conference.'
(Birds Australia: Armidale, NSW)
Murray, D.L., and Fuller, M.R. (2000) A critical review of the effects of marking on the biology of
vertebrates. In 'Research techniques in animal ecology: controversies and consequences.'. pp.
15-64. (Columbia University Press: New York)
O'Brien, T.G., and Kinnaird, M.F. (2008) A picture is worth a thousand words: the application of
camera trapping to the study of birds. Bird Conservation International 18(S1).
Paltridge, R. (2002) The diets of cats, foxes and dingoes in relation to prey availability in the
Tanami Desert, Northern Territory. Wildlife Research 29, 389-403.
Raim, A. (1978) A radio transmitter attachment for small passerine birds. Bird Banding 49, 326-
332.
Rappole, J.H., and Tipton, A.R. (1991) New Harness Design for Attachment of Radio
Transmitters to Small Passerines. Journal of Field Ornithology 62, 335-337.
Robertson, J.G., Eknert, B., and Ihse, M. (1990) Habitat analysis from infra-red aerial
photographs and the conservation of birds in Swedish agricultural landscapes. Ambio, 195-203.
Rowcliffe, J.M., Field, J., Turvey, S.T., and Carbone, C. (2008) Estimating animal density using
camera traps without the need for individual recognition. Journal of Applied Ecology 45, 1228-
1236.
Saxon, E. (1983) Mapping the habitats of rare animals in the Tanami Wildlife Sanctuary (central
Australia): An application of satellite imagery. Biological conservation 27, 243-257.
Seaman, D.E., Millspaugh, J.J., Kernohan, B.J., Brundige, G.C., Raedeke, K.J., and Gitzen,
R.A. (1999) Effects of sample size on kernel home range estimates. The Journal of Wildlife
Management 63, 739-747.
Skroblin, A., and Murphy, S. (2013) The conservation status of Australian malurids and their
value as models to understand land management issues. Emu 113, 309-318.
Spotswood, E.N., Goodman, K.R., Carlisle, J., Cormier, R.L., Humple, D.L., Rousseau, J.,
Guers, S.L., and Barton, G.G. (2012) How safe is mist netting? evaluating the risk of injury and
mortality to birds. Methods in Ecology and Evolution 3, 29-38.

Night Parrot Pezoporus occidentalis Research Plan Page 38 of 40
Thomson, P.C. (1992) The behavioural ecology of dingoes in north-western Australia. IV. Social
and spatial organistaion, and movements. Wildlife Research 19, 543-563.
Wallin, D.O., Elliott, C.C., Shugart, H.H., Tucker, C.J., and Wilhelmi, F. (1992) Satellite remote
sensing of breeding habitat for an African weaver-bird. Landscape Ecology 7, 87-99.
Welsh, A.H., Lindenmayer, D.B., and Donnelly, C.F. (2013) Fitting and Interpreting Occupancy
Models. PloS one 8, e52015.
Whitlock, F.L. (1924) Journey to Central Australia in search of the Night Parrot. Emu 23, 248-
281.
Wilson, G.J., and Delahay, R.J. (2001) A review of methods to estimate the abundance of
terrestrial carnivores using field signs and observation. Wildlife Research 28, 151-164.
Wilson, H. (1937) Notes on the Night Parrot, with references to recent occurrences. Emu 37,
79-87.

Night Parrot Pezoporus occidentalis Research Plan Page 39 of 40
This page has been left blank intentionally

Page 27 of 27
11. Appendix 2 – Approval letter

