relative growth and morphological sexual maturity of uca cumulanta (crustacea: decapoda: ocypodidae)...
TRANSCRIPT

This article was downloaded by: [UNESP]On: 05 February 2015, At: 03:24Publisher: Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House,37-41 Mortimer Street, London W1T 3JH, UK
Click for updates
Invertebrate Reproduction & DevelopmentPublication details, including instructions for authors and subscription information:http://www.tandfonline.com/loi/tinv20
Relative growth and morphological sexual maturity ofthe mangrove crab Aratus pisonii (H. Milne Edwards,1837) (Decapoda, Brachyura, Sesarmidae) on thesouthern coast of the state of São Paulo, BrazilRégis Augusto Pescinellia, Thiago Maia Davansoa & Rogerio Caetano da Costaa
a LABCAM (Laboratório de Biologia de Camarões Marinhos e de Água Doce), Faculdade deCiências, Departamento de Ciências Biológicas, UNESP, Bauru, SP, BrazilPublished online: 03 Feb 2015.
To cite this article: Régis Augusto Pescinelli, Thiago Maia Davanso & Rogerio Caetano da Costa (2015): Relative growthand morphological sexual maturity of the mangrove crab Aratus pisonii (H. Milne Edwards, 1837) (Decapoda, Brachyura,Sesarmidae) on the southern coast of the state of São Paulo, Brazil, Invertebrate Reproduction & Development, DOI:10.1080/07924259.2015.1006339
To link to this article: http://dx.doi.org/10.1080/07924259.2015.1006339
PLEASE SCROLL DOWN FOR ARTICLE
Taylor & Francis makes every effort to ensure the accuracy of all the information (the “Content”) containedin the publications on our platform. However, Taylor & Francis, our agents, and our licensors make norepresentations or warranties whatsoever as to the accuracy, completeness, or suitability for any purpose of theContent. Any opinions and views expressed in this publication are the opinions and views of the authors, andare not the views of or endorsed by Taylor & Francis. The accuracy of the Content should not be relied upon andshould be independently verified with primary sources of information. Taylor and Francis shall not be liable forany losses, actions, claims, proceedings, demands, costs, expenses, damages, and other liabilities whatsoeveror howsoever caused arising directly or indirectly in connection with, in relation to or arising out of the use ofthe Content.
This article may be used for research, teaching, and private study purposes. Any substantial or systematicreproduction, redistribution, reselling, loan, sub-licensing, systematic supply, or distribution in anyform to anyone is expressly forbidden. Terms & Conditions of access and use can be found at http://www.tandfonline.com/page/terms-and-conditions

Relative growth and morphological sexual maturity of the mangrove crab Aratus pisonii(H. Milne Edwards, 1837) (Decapoda, Brachyura, Sesarmidae) on the southern coast of thestate of São Paulo, Brazil
Régis Augusto Pescinelli, Thiago Maia Davanso and Rogerio Caetano da Costa*
LABCAM (Laboratório de Biologia de Camarões Marinhos e de Água Doce), Faculdade de Ciências, Departamento de CiênciasBiológicas, UNESP, Bauru, SP, Brazil
(Received 28 July 2014; accepted 6 January 2015)
Growth in crustaceans is characterized by ontogenetic differentiation during the development of their body structures, sostudies on relative growth are widely applied in this group. In this study, the growth pattern of the body structures wasverified through the analysis of relative growth, and then, morphological sexual maturity of the mangrove crab Aratuspisonii (H. Milne Edwards, 1837) was estimated. The carapace width (CW), cheliped propodus length (PL), chelipedpropodus height (PH), propodus width (PW), abdomen width (AW), and first pleopod length (FPL) of the crabs weremeasured. The relationships that best showed changes in the allometric coefficient among demographic categories wereFPL vs. CW for males and AW vs. CW for females. This study verified the increased size of the cheliped in terms oflength, width, and height, which occurred mainly in adult males. This increase reflects the importance of this structure inthe reproductive processes of A. pisonii. For females, the increase in abdomen growth reflects their reproductivepotential, since it is a structure that provides protection for eggs.
Keywords: allometry; development; reproduction
Introduction
Mangrove forests provide a high diversity of ecologicalniches for many brachyuran species (Kathiresan &Bingham 2001). One of these species, the mangrove crabAratus pisonii (H. Milne Edwards, 1837) from the familySesarmidae, inhabits the area above these coastalenvironments and is usually associated with aerial struc-tures (stems and roots) of plant species such asRhizophora mangle (Warner 1967; Diaz & Conde 1989).A. pisonii is considered one of the main species responsi-ble for energy transfer between pelagic and benthic foodwebs due to its production of thousands of larvae thatare eaten by many species that inhabit or spend part oftheir life cycle in mangroves (Schwamborn et al. 1999,Schwamborn et al. 2002).
Many studies focusing on this species have beenconducted to examine various aspects of its biology,such as life history (Warner 1967), behavior (Warner1970), population dynamics (Diaz & Conde 1989),fecundity (Leme & Negreiros-Fransozo 1998), foodselection (Erickson et al. 2008), population biology(Leme 2002), early development (Diaz & Bevilacqua1987), output (Leme 2006), and larval morphology(Cuesta et al. 2006). However, when it comes to popula-tions along the Brazilian coast, there seems to be noinformation in the literature regarding the size at which
this species reaches morphological sexual maturity, or onthe growth pattern of their body structures.
The growth of crustaceans is characterized by ontoge-netic differences that occur during the development oftheir body structures (Hartnoll 1978). As a result, the studyof relative growth is widely applied to this group, since itis an effective tool in the morphological observation of thetransition between juvenile and adult stages (Hartnoll1978; Vaninni & Gherardi 1988; Dalabona et al. 2005). Incrabs, structures such as chelipeds, abdomen, and gonopodmay be used to establish this feature (Castiglioni &Negreiros-Fransozo 2004). Estimating a size at whichspecies of the infraorder Brachyura reach morphologicalsexual maturity is critical in the understanding of the lifecycle of these organisms (Hartnoll & Gould 1988).
The goal of this study was to estimate the morpho-logical sexual maturity of the mangrove crab A. pisoniiand to verify the growth patterns of its body structuresthrough the analysis of relative growth.
Materials and methods
Study area
Sampling was performed in the mangrove of the estua-rine-lagoon complex of Cananéia-Iguape, located alongthe southernmost coast of São Paulo state (25°04′25.7′′ S
*Corresponding author. Email: [email protected]
© 2015 Taylor & Francis
Invertebrate Reproduction & Development, 2015http://dx.doi.org/10.1080/07924259.2015.1006339
Dow
nloa
ded
by [
UN
ESP
] at
03:
24 0
5 Fe
brua
ry 2
015

– 48°03′29.6′′ W). The northernmost limit of the com-plex is the city of of Iguape, while the eastern limit isthe island known as Ilha Comprida, and the westernmostlimit includes the Serra do Mar escarpment, the city ofCananeia, and Cardoso Island. This region is known forits high degree of conservation interest, and it is consid-ered one of the most productive estuarine ecosystems inthe world (UNESCO 2005).
Sampling
Sampling was performed bimonthly from February 2013to January 2014. Crabs were collected manually in aerialstructures of the trees (stems and roots) and were storedin plastic bags with ice and taken to the Laboratory ofBiology of Marine and Freshwater Shrimp (LABCAM).
Relative growth and determination of morphologicalsexual maturity
The sex of each crab was identified according to themorphology of the abdomen and the presence of pairedgonopods in males. The following measurements weretaken using a 0.01-mm caliper: carapace width (CW),cheliped propodus length (PL), cheliped propodus height(PH), cheliped propodus width (PW), abdomen width(AW), and first pleopod length (FPL) (Figure 1).
Changes in the growth pattern of the body structuresin relation to the independent variable (CW) were veri-fied by means of relative growth analysis. The allometricequation y = axb in the linearized version (log y = log
a + b log x) was used, where y is the dependent variableor the dimension studied, x is the independent variable,b is the allometric coefficient of the studied structure,and a is the point at which the straight line intersects theordinate axis. The allometric condition b for each struc-ture was analyzed (b = 1: isometry; b < 1: negativeallometry; b > 1: positive allometry) using Student’st-test (Ho: b = 1, α = 5%) (Zar 1996).
A non-hierarchical “K-means clustering” analysiswas performed on the transformed data (log10). Thismethod distributes data into a previously determinednumber of groups defined by an iterative process thatminimizes the variance within groups and maximizes thevariance among them. The result of the classification(K-means) was refined by applying a discriminant analy-sis (DA). The size of the smallest individual classified asan adult was used as the value at which the speciesreaches the morphological sexual maturity. This statisti-cal methodology was based on the work of Sampedroet al. (1999).
After the log-transformed data was divided intodemographic categories, an analysis of covariance(ANCOVA) was applied to test the angular and linearcoefficients between juveniles and adults for each rela-tionship and to check whether the groups found shouldbe represented by different linear equations.
To verify the difference between the average sizesreached by males and females, Student’s t-test and theMann–Whitney test were used for normal data and fornon-normal data, respectively (α = 0.05).
A DA was performed to check for possible sexualdimorphism, searching for evidence of morphometric dif-ferences between sexes, and determining which variables(body structures) were more important in this dimor-phism. The discriminatory power of each variable wasobserved by means of Wilks’lambda, combined with sta-tistical values of F and p (F values < 1 were included inthe DA) (Anastasiadou & Leonardos 2008).
Results
Sampling
A total of 438 crabs were captured. From these, 229were males and 209 were females, 58 of which were car-rying eggs. CW ranged from 5.2 to 24.39 mm (15.00± 3.96) in males and from 5.98 to 21.41 mm (13.51± 2.77) in females. When only the females with eggswere considered, CW ranged from 11.97 to 17.50 mm(13.61 ± 1.74).
Relative growth and determination of morphologicalsexual maturity
A detailed description of each of the relationshipsanalyzed can be seen in Table 1. The PL vs. CW
Figure 1. A. pisonii (H. Milne Edwards, 1837). Body dimen-sions used in the morphometric analyses (not in scale): (CW)carapace width, (PL) cheliped propodus length, (PH) chelipedpropodus height, (PW) cheliped propodus width, (FPL) firstpleopod length, and (AW) abdomen width. a = Female andb = Male.
2 R.A. Pescinelli et al.
Dow
nloa
ded
by [
UN
ESP
] at
03:
24 0
5 Fe
brua
ry 2
015
relationship showed positive allometric growth for bothjuvenile males and adults. For females, growth was neg-atively allometric for juveniles and isometric for adults.The AW vs. CW relationship showed positive allometricgrowth for juveniles and negative allometric growth inadults, for both males and females. When the PH vs.CW relationship was calculated, positively allometricgrowth was observed for males in both demographic cat-egories analyzed, while in females, growth was isometricfor juveniles and positive allometric for adults. The PWvs. CW relationship showed isometric growth for juve-nile males and a positive allometry for the adults. How-ever, a negative allometry for both female demographiccategories was observed. When the FPL vs. CW relation-ship was determined, the allometric growth was positivefor juveniles and allometry was negative for adults.
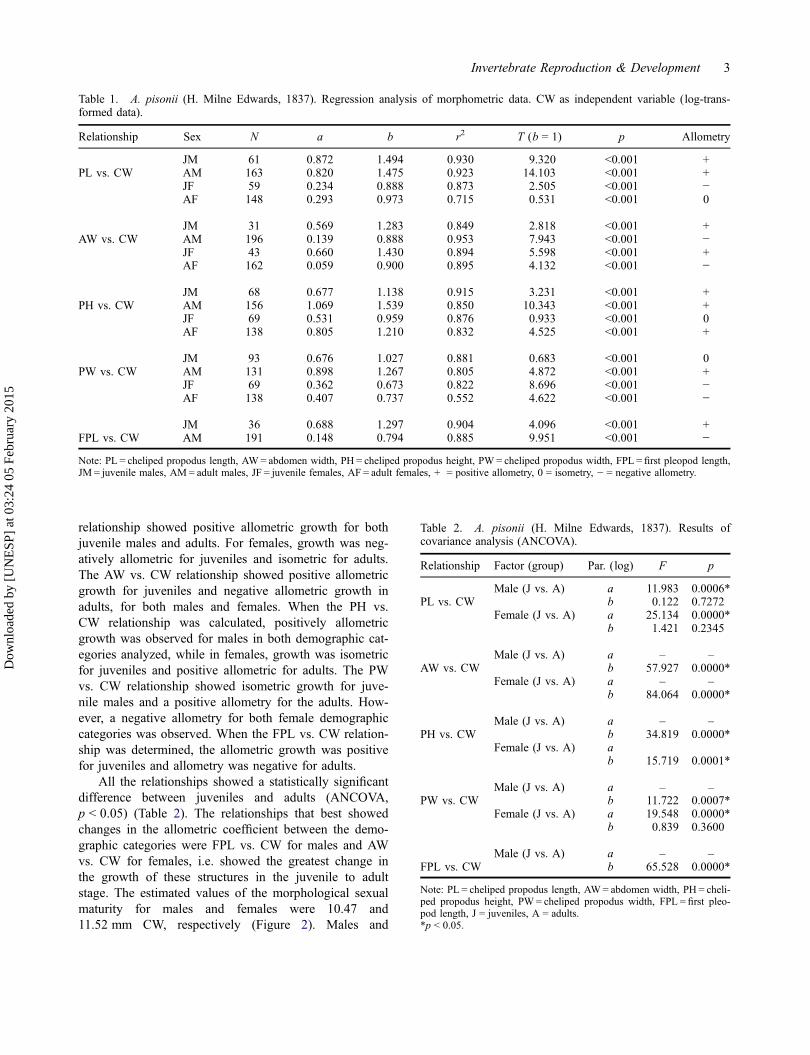
All the relationships showed a statistically significantdifference between juveniles and adults (ANCOVA,p < 0.05) (Table 2). The relationships that best showedchanges in the allometric coefficient between the demo-graphic categories were FPL vs. CW for males and AWvs. CW for females, i.e. showed the greatest change inthe growth of these structures in the juvenile to adultstage. The estimated values of the morphological sexualmaturity for males and females were 10.47 and11.52 mm CW, respectively (Figure 2). Males and
Table 1. A. pisonii (H. Milne Edwards, 1837). Regression analysis of morphometric data. CW as independent variable (log-trans-formed data).
Relationship Sex N a b r2 T (b = 1) p Allometry
JM 61 0.872 1.494 0.930 9.320 <0.001 +PL vs. CW AM 163 0.820 1.475 0.923 14.103 <0.001 +
JF 59 0.234 0.888 0.873 2.505 <0.001 −AF 148 0.293 0.973 0.715 0.531 <0.001 0
JM 31 0.569 1.283 0.849 2.818 <0.001 +AW vs. CW AM 196 0.139 0.888 0.953 7.943 <0.001 −
JF 43 0.660 1.430 0.894 5.598 <0.001 +AF 162 0.059 0.900 0.895 4.132 <0.001 −
JM 68 0.677 1.138 0.915 3.231 <0.001 +PH vs. CW AM 156 1.069 1.539 0.850 10.343 <0.001 +
JF 69 0.531 0.959 0.876 0.933 <0.001 0AF 138 0.805 1.210 0.832 4.525 <0.001 +
JM 93 0.676 1.027 0.881 0.683 <0.001 0PW vs. CW AM 131 0.898 1.267 0.805 4.872 <0.001 +
JF 69 0.362 0.673 0.822 8.696 <0.001 −AF 138 0.407 0.737 0.552 4.622 <0.001 −
JM 36 0.688 1.297 0.904 4.096 <0.001 +FPL vs. CW AM 191 0.148 0.794 0.885 9.951 <0.001 −
Note: PL = cheliped propodus length, AW= abdomen width, PH = cheliped propodus height, PW = cheliped propodus width, FPL = first pleopod length,JM = juvenile males, AM= adult males, JF = juvenile females, AF = adult females, + = positive allometry, 0 = isometry, − = negative allometry.
Table 2. A. pisonii (H. Milne Edwards, 1837). Results ofcovariance analysis (ANCOVA).
Relationship Factor (group) Par. (log) F p
Male (J vs. A) a 11.983 0.0006*PL vs. CW b 0.122 0.7272
Female (J vs. A) a 25.134 0.0000*b 1.421 0.2345
Male (J vs. A) a – –AW vs. CW b 57.927 0.0000*
Female (J vs. A) a – –b 84.064 0.0000*
Male (J vs. A) a – –PH vs. CW b 34.819 0.0000*
Female (J vs. A) ab 15.719 0.0001*
Male (J vs. A) a – –PW vs. CW b 11.722 0.0007*
Female (J vs. A) a 19.548 0.0000*b 0.839 0.3600
Male (J vs. A) a – –FPL vs. CW b 65.528 0.0000*
Note: PL = cheliped propodus length, AW= abdomen width, PH = cheli-ped propodus height, PW = cheliped propodus width, FPL = first pleo-pod length, J = juveniles, A = adults.*p < 0.05.
Invertebrate Reproduction & Development 3
Dow
nloa
ded
by [
UN
ESP
] at
03:
24 0
5 Fe
brua
ry 2
015
females were found to have differences in their averagesize (Mann–Whitney, p < 0.05). When the males andfemales were analyzed separately, a sexual dimorphismin relation to size was observed in both juveniles (Stu-dent’s t-test, p < 0.05) and adults (Mann–Whitney,p < 0.05). Additionally sexual dimorphism was observedin the cheliped PL and AW (DA, p < 0.05) (Table 3).
Discussion
For most organisms, a decrease in the absolute growthof adults in relation to juveniles becomes clear. Thischaracteristic can be observed in crustaceans, fish, “rep-tiles,” amphibians, and plants, organisms which continuegrowing after reaching sexual maturity, but slower due to
the energy cost directed toward reproduction (Taylor &Gabriel 1992) and also can affect in the relative growthwith a decrease in the growth of some body structures.This change in relative growth was noted in someA. pisonii relationships in the present study.
In crabs (mainly for males), the growth of the cheli-ped is differentiated: It is highlighted by a positiveallometry, since it is a fundamental structure for intra-and interspecific interactions such as territorial disputes,defense against predators, and female courting (Hartnoll1974; Castiglioni & Negreiros-Fransozo 2004). Theincrease in the size of the cheliped (particularly in adultmales and in terms of length, width, and height) verifiedin this study highlights the importance of this structurein the processes related to reproduction. In the naturalenvironment, A. pisonii males have been observed fight-ing for females (Warner 1970). Crabs with robust cheli-peds have advantages when competing with other malesand thus in terms of sexual selection by females(Mariappan et al. 2000).
In addition to territorial behavior and fights forfemales through physical combat, A. pisonii malesengage in a “display” performance, which starts with astrong vibration of the cheliped, followed by lifting andlateral positioning to intimidate and scare the opponent(Warner 1970). According to the author, the size of thecheliped in males reflects its dominance over othermales. After the fight, however, the losing individualreceives aggression from the dominant male. This factcould explain the cases of cannibalism reported by War-ner (1970), in which this behavior was recorded asoccurring among individuals whose larger chelipeds thanthose of the dead individuals.
The positive allometric growth in males and theprevalence of negative allometric and isometric growthin females in almost all relationships in which the cheli-ped was measured show the distinct strategies that eachsex uses in relation to the direction of energy. Inbrachyuran females, the lower increment of chelipedgrowth compared to that of males is due to the directionof energy toward egg production and toward abdomengrowth (Hartnoll 1974).
A. pisonii shows a sexual dimorphism of chelipedsand abdomens (Hartnoll 1965), where in the averagefemale cheliped is smaller and the agonistic behavior
Figure 2. A. pisonii (H. Milne Edwards, 1837). Estimated sizeat morphological sexual maturity for males and females. Theestimated size refers to the smallest individual after the inflec-tion point of the equations for juveniles and adults.
Table 3. A. pisonii (H. Milne Edwards, 1837). Results of the DA of morphometric data.
Body structures Factor (Group) Wilks’lambda F p
PL Male vs. Female 0.333157 0.1975 0.656930PW Male vs. Female 0.333470 0.6037 0.437589PH Male vs. Female 0.357012 31.1445 0.000000*AW Male vs. Female 0.444088 144.1062 0.000000*
Note: PL = cheliped propodus length, PW = cheliped propodus width, PH = cheliped propodus height, AW = abdomen width.*p < 0.05.
4 R.A. Pescinelli et al.
Dow
nloa
ded
by [
UN
ESP
] at
03:
24 0
5 Fe
brua
ry 2
015

rituals, such as the aforementioned display, are limited tomales (Warner 1970). The use of this structure in ritualsalso reflects in a more complex social organization ofthe species, in which the ritualized behavior of malesreduces physical damage that could occur during dis-putes. In species with little sexual dimorphism in thechelipeds, such as Goniopsis cruentata (Latreille, 1803),both males and females use this structure in interspecificfights and disputes. However, the high degree of sexualdimorphism of the chelipeds of A. pisonii is reflected inthe less offensive behavior of females compared to thatof males (Hartnoll 1974).
Abdomen growth in females is essential, because theabdomen is a structure that provides protection for eggsand thus creates better conditions for incubation of futuregenerations, in addition to protecting gonopores(Castiglioni & Negreiros-Fransozo 2004; Cobo & Alves2009). In this study, A. pisonii females showed positiveallometric growth in the juvenile stage, followed by anegative allometry in the adult stage. Therefore, energydirection toward abdomen growth in the periodpreceding morphological sexual maturity in femalesseems to be an evolutionarily selected characteristic thatprovides greater efficiency during the reproductive pro-cesses. This type of abdomen growth was also observedin the freshwater crab species Trichodactylus fluviatilisLatreille, 1828 (Pescinelli et al. 2014).
The abdomen in males protects the paired gonopod,structures formed by the first- and second-modified pleo-pods, with the function of transferring spermatophoresduring copulation (Castiglioni & Negreiros-Fransozo2004). The increase in abdomen size occurred in juvenilemales, followed by a negative allometric growth inadults. This also occurred in relation to the first pleopod,which shows a synchronism and a relationship betweenboth structures during development. This kind of growthin the adult stage of males prevents an overgrowth of thegonopod; this overgrowth could result in functional prob-lems since the size of this structure is proportional to thesize of the female genital opening (Hartnoll 1974). Thenegative allometry also reduces variation in the size ofthe gonopod between large and small males, allowing ahigher number of males to mate with a higher proportionof females from the population (Hartnoll 1974); in otherwords, males that recently reached morphological sexualmaturity and those that are already morphologicallymature can copulate with most of the female population.
In the present study, the cheliped and the gonopodshowed changes in growth patterns between the demo-graphic categories in the same size class for males. Thisresult shows that both structures may be used in theestimating of the morphological sexual maturity in thisspecie. However, considering the fact that the gonopodsare involved in the transfer of spermatophores to thefemale in the act of copulation, this structure becomes
indispensable for the reproductive success of crabs andwas used to estimate the morphological sexual maturityin males.
Sexual dimorphism related to size was also observedin A. pisonii in both demographic categories: Males werefound to be larger in size. Nevertheless, males reachmorphological sexual maturity at smaller sizes whencompared to females. This fact ensures that smallermales are able to participate in the reproductive process,since competition for females only occurs between malesof similar size (Warner 1970). Thus, when it comes tobody size, the size of cheliped will have a greaterinfluence in this dispute (Warner 1970).
Many previous studies have highlighted the dilemmaof energy allocation during all life phases of animals andplants, including studies by Cohen (1971), Mirmiraniand Oster (1978), Vincent and Pulliam (1980), King andRoughgarden (1981), and Schaffer et al. (1982). All ofthese authors pointed out that reproductive success isfavored when the direction of energy toward somaticgrowth and toward reproduction occur separately ontimescale, and this can also occur in the relative growth.In this study, A. pisonii showed greater growth in thejuvenile stage in that structures directly involved inreproduction, such as the first pleopod in males and theabdomen in females, followed by a decrease in the ofgrowth after morphological sexual maturity was reached.
AcknowledgmentsThe authors are grateful to “Fundação de Amparo à Pesquisado Estado de São Paulo” (FAPESP) Scholarship (#2013/12136-4 To RAP), Thematic Biota (#2010/50188-8 to RCC), “Conse-lho Nacional de Desenvolvimento Científico e Tecnológico”(CNPq) for Research Scholarships (PQ 304784/2011-7) to(RCC) and (CAPES) Scholarship (TMD). We also thank theLABCAM coworkers for help during field work, Msc. JoãoAlberto Farinelli Pantaleão for illustration of body dimensions,and Dr Gustavo Luis Hirose for assisting in the analysis. Allexperiments conducted during this study complied with currentapplicable state and federal laws in Brazil.
ReferencesAnastasiadou CA, Leonardos ID. 2008. Morphological varia-
tion among populations of Atyaephyra desmarestii (Millet,1831) (Decapoda: Caridea: Atyidae) from freshwater habi-tats of northwestern Greece. Journal of Crustacean Biology.28:240–247.
Castiglioni DS, Negreiros-Fransozo ML. 2004. Comparativeanalysis of the relative growth of Uca rapax (Smith) (Crus-tacea, Ocypodidae) from two mangroves in São Paulo. Bra-zil. Revista Brasileira de Zoologia. 21:137–144.
Cobo V, Alves DFR. 2009. Relative growth and sexual matu-rity of the spider crab Mithrax tortugae Rathbun, 1920(Brachyura, Mithracidae) on a continental island off thesoutheastern Brazilian coast. Crustaceana. 82:1265–1273.
Cohen D. 1971. Maximizing final yield when growth is limitedby time or by limiting resources. Journal of TheoreticalBiology. 33:299–307.
Invertebrate Reproduction & Development 5
Dow
nloa
ded
by [
UN
ESP
] at
03:
24 0
5 Fe
brua
ry 2
015

Cuesta JA, García-Guerreiro MU, Rodríguez A, Hendrickx M.2006. Larval morphology of the sesarmid crab, Aratuspisonii (H. Milne Edwards, 1837) (Decapoda, Brachyura,Grapsoidea) from laboratory-reared material. Crustaceana.79:175–196.
Dalabona G, Silva JL, Pinheiro MAA. 2005. Size at morpholog-ical maturity of Ucides cordatus (Linnaeus, 1763) (Brachy-ura, Ocypodidae) in the Laranjeiras Bay, Southern Brazil.Brazilian Archives of Biology and Technology. 48:139–145.
Diaz H, Bevilacqua M. 1987. Early developmental sequencesof Aratus pisonii (H. Milne-Edwards) (Brachyura, Grapsi-dae) under laboratory conditions. Journal of CoastalResearch. 3:63–70.
Diaz H, Conde JE. 1989. Population dynamics and life history ofthe mangrove crab Aratus pisonii (Brachyura: Grapsidae) in amarine environment. Bulletin of Marine Science. 45:148–163.
Erickson AA, Feller IC, Paul VJ, Kwiatkowski LM, Lee W.2008. Selection of an omnivorous diet by the mangrovetree crab Aratus pisonii in laboratory experiments. Journalof Sea Research. 59:59–69.
Hartnoll RG. 1965. Notes on the marine grapsid crabs ofJamaica. Proceedings of the Linnean Society of London.176:113–147.
Hartnoll RG. 1974. Variation in growth pattern betweensome secondary sexual characters in crabs (Decapoda,Brachyura). Crustaceana. 27:131–136.
Hartnoll RG. 1978. The determination of relative growth inCrustacea. Crustaceana. 34:281–293.
Hartnoll RG, Gould P. 1988. Brachyuran life history strategiesand the optimization of egg production. Symposia of theZoological Society of London. 59:1–9.
Kathiresan K, Bingham BL. 2001. Biology of mangroves andmangroves ecosystems. Advances in Marine Biology.40:84–251.
King D, Roughgarden J. 1981. Multiple switches betweenvegetative and reproductive growth in annual plants.Theoretical Population Biology. 21:194–204.
Leme MH. 2006. Investimento reprodutivo e produção de ovosem desovas consecutivas do caranguejo Aratus pisonii(H. Milne Edwards) (Crustacea, Brachyura, Grapsoidea).Revista Brasileira de Zoologia. 23:727–732.
Leme MHA. 2002. A comparative analysis of the populationbiology of the mangrove crabs Aratus pisonii and Sesarmarectum (Brachyura, Grapsidae) from the north coast of SãoPaulo state, Brazil. Journal of Crustacean Biology. 22:553–557.
Leme MHA, Negreiros-Fransozo ML. 1998. Fecundity ofAratus pisonii (Decapoda, Grapsidae) in Ubatuba region,
state of São Paulo, Brazil. Iheringia, Série Zoológica, PortoAlegre. 84:73–77.
Mariappan P, Balasundaram C, Schmitz B. 2000. Decapodcrustacean chelipeds: an overview. Journal of Biosciences.25:301–313.
Mirmirani M, Oster GF. 1978. Competition, kin selection, andevolutionary stable strategies. Theoretical PopulationBiology. 13:304–339.
Pescinelli RA, Pantaleão JAF, Davanso TM, Costa RC. 2014.Relative growth and morphological sexual maturity of thefreshwater crab Trichodactylus fluviatilis Latreille 1828(Decapoda, Trichodactylidae) from west central São PauloState, Brazil. Invertebrate Reproduction and Development.58:108–114.
Sampedro MP, Gonzáles-Gurriarán E, Freire J, Muiño R. 1999.Morphometry and sexual maturity in the spider crab Majasquinado (Decapoda: Majidae) in Galicia, Spain. Journal ofCrustacean Biology. 19:578–592.
Schaffer WM, Inouye RS, Whittam TS. 1982. Energyallocation by an annual plant when the effects of seasonali-ty on growth and reproduction are decoupled. TheAmerican Naturalist. 120:787–815.
Schwamborn R, Ekau W, Pinto AS, Silva TA, Saint-Paul U.1999. The contribution of estuarine decapod larvae to mar-ine macrozooplankton communities in northeast Brazil.Archive of Fishery Marine Research. 47:167–182.
Schwamborn R, Voss M, Ekau W, Saint-Paul U. 2002. Howimportant are mangroves as a carbon source for decapodcrustacean larvae in a tropical estuary? Marine EcologyProgress Series. 229:195–205.
Taylor BE, Gabriel W. 1992. To grow or not to grow: optimalresource allocation for Daphnia. The American Naturalist.139:248–266.
UNESCO. 2005. World network of biosphere reserves – SC/EES – June 2005. Paris: The MAB Program; p. 19.
Vaninni M, Gherardi F. 1988. Studies on the pebble crab,Eriphia smithi MacLeay 1838 (Xanthoidea Menippidae):patterns of relative growth and population structure. Tropi-cal Zoology. 1:203–216.
Vincent TL, Pulliam HR. 1980. Evolution of life historystrategies for an asexual annual plant model. TheoreticalPopulation Biology. 17:215–231.
Warner GF. 1967. The life history of the mangrove tree crabAratus pisonii. Journal of Zoology. 153:321–335.
Warner GF. 1970. Behaviour of two species of grapsid crabduring intraspecific encounters. Behaviour. 36:9–19.
Zar JH. 1996. Biostatistical analysis. Upper Saddle River (NJ):Prentice Hall; p. 662.
6 R.A. Pescinelli et al.
Dow
nloa
ded
by [
UN
ESP
] at
03:
24 0
5 Fe
brua
ry 2
015