regulation of human insulin gene transcription by the immunosuppressive drugs cyclosporin a and...
TRANSCRIPT

Abstract Cyclosporin A and tacrolimus are importantimmunosuppressive drugs. They share a diabetogenic ac-tion as one of their most serious adverse effects. In a singlestudy, tacrolimus (100 nM) inhibited human insulin genetranscription in the β-cell line HIT. Using transfections ofa human insulin-reporter gene into HIT cells, the presentstudy shows that this inhibition is seen only at high con-centrations of tacrolimus and is not caused by cyclospor-in A. However, after stimulation by the major second mes-sengers in the regulation of the insulin gene, cAMP anddepolarization-induced calcium influx, both tacrolimusand cyclosporin A inhibited human insulin gene transcrip-tion in a concentration-dependent manner with IC50 valuesof 1 nM and 30 nM, respectively. A further analysis offersa mechanism for this effect by revealing that the activa-tion by cAMP and calcium of human insulin gene tran-scription is mediated by the transcription factor cAMP-re-sponsive element binding protein (CREB) whose activityis inhibited by the immunosuppressants. These data demon-strate for the first time that cAMP- and calcium-inducedactivity of the human insulin gene is mediated by CREBand blocked by both tacrolimus and cyclosporin A at con-centrations that inhibit calcineurin phosphatase activity.Since also the immunosuppressive effects of cyclosporin Aand tacrolimus are thought to be secondary to inhibitionof calcineurin, the present study suggests that inhibitionof human insulin gene transcription by the immunosup-pressants is clinically important and may contribute to theirdiabetogenic effect.
Keywords Cyclosporin A · Tacrolimus · Calcineurin ·Human insulin gene transcription · CREB
Abbreviations CAT chloramphenicol acetyl transferase ·CRE cAMP-responsive element · CREBcAMP-responsive element binding protein · GLP-1glucagon-like peptide 1 · NFAT nuclear factor ofactivated T-cells · PKA protein kinase A · RSV Roussarcoma virus
Introduction
Cyclosporin A and tacrolimus are clinically important im-munosuppressive drugs which are widely used in order toprevent organ rejection after transplantation. The intro-duction of cyclosporin A in the early 1980s resulted in anextraordinary improvement of graft survival. In addition,both drugs are used in the therapy of an increasing num-ber of autoimmune diseases. The structurally distinctdrugs bind to their respective intracellular receptors, theimmunophilins. The drug/immunophilin complexes inter-act directly with the calcium/calmodulin-dependent phos-phatase calcineurin, and inhibit its activity (Ho et al. 1996).Inhibition of calcineurin blocks the dephosphorylationand nuclear translocation of the nuclear factor of activatedT-cells (NFAT), thereby preventing the activation of NFAT-regulated genes, including those necessary for T-cell pro-liferation, such as interleukin 2 (Ho et al. 1996; Rao et al.1997; Rühlmann and Nordheim 1997). Thus, inhibition ofNFAT-directed transcription and repression of an early stepin T-cell activation may be one mechanism to explain theimmunosuppressive action of cyclosporin A and tacrolimus(Ho et al. 1996; Rao et al. 1997; Rühlmann and Nordheim1997). However, other targets of calcineurin are likely tobe involved in this process, including the cAMP responseelement (CRE)-binding transcription factor CREB. Previ-ous studies have shown that cyclosporin A and tacrolimusinhibit CRE-directed transcription and CREB transcrip-tional activity by blocking calcineurin in the T cell lineJurkat (Krüger et al. 1997; Schwaninger et al. 1993b,1993c, 1995). Since CREB is required for T-cell activa-tion (Barton et al. 1996; Rudolph et al. 1998), these find-ings suggest that inhibition of CREB activity represents
Elke Oetjen · Daniela Grapentin · Roland Blume · Michael Seeger · Doris Krause · Anke Eggers · Willhart Knepel
Regulation of human insulin gene transcription by the immunosuppressive drugs cyclosporin A and tacrolimus at concentrations that inhibit calcineurin activity and involving the transcription factor CREB
Naunyn-Schmiedeberg’s Arch Pharmacol (2003) 367 : 227–236DOI 10.1007/s00210-003-0694-7
Received: 12 November 2002 / Accepted: 9 January 2003 / Published online: 20 February 2003
ORIGINAL ARTICLE
E. Oetjen · D. Grapentin · R. Blume · M. Seeger · D. Krause ·A. Eggers · W. Knepel (✉)Department of Molecular Pharmacology, University of Göttingen, Robert-Koch-Strasse 40, 37075 Göttingen, GermanyTel.: +49-551-395787, Fax: +49-551-399652,e-mail: [email protected]
© Springer-Verlag 2003

an additional mechanism underlying the immunosuppres-sive effects of cyclosporin A and tacrolimus.
The therapeutic application of cyclosporin A and tacro-limus is, however, limited by shared untoward effects(Kahan 1989; European FK506 Multicenter Study LiverGroup 1994; U.S. Multicenter FK506 Liver Study Group1994). Among the most serious adverse effects of im-munosuppressive therapy using cyclosporin A and tacro-limus is impaired glucose tolerance leading to hyper-glycemia and diabetes mellitus (Kahan 1989; EuropeanFK506 Multicenter Study Liver Group 1994; U.S. Multi-center FK506 Liver Study Group 1994; Docherty andClark 1994; Saltiel 2001; Kahn 1994). The incidence ofdiabetes mellitus associated with the use of cyclosporin Aand tacrolimus has been reported to be 10–30% (Kahan1989; European FK506 Multicenter Study Liver Group1994; U.S. Multicenter FK506 Liver Study Group 1994).It is manifested by insulin deficiency, insulin resistance,and the appearance of islet cell antibodies (Krentz et al.1993; Lohmann et al. 2000; Jindal et al. 1997). This ad-verse effect may be particularly important for islet trans-plantation, where immunosuppressive regimens also in-clude tacrolimus and cyclosporin A (Shapiro et al. 2000).The mechanism of the diabetogenic action of cyclospor-in A and tacrolimus is unclear. Since it is caused by bothdrugs, the diabetogenic effect of cyclosporin A and tacro-limus is generally assumed to be secondary to inhibitionof calcineurin (Ho et al. 1996). Furthermore, a direct effectof cyclosporin A and tacrolimus on pancreatic islets maybe involved. Cyclosporin A and tacrolimus decrease in-sulin mRNA levels, insulin content and ultimately insulinsecretion in isolated islets (Herold et al. 1993; Redmon etal. 1996; Teuscher et al. 1994). Since the immunosuppres-sants do not acutely alter insulin secretion or even en-hance insulin secretion acutely (Ebihara et al. 1996), theseeffects may be explained by a reduction in insulin genetranscription. Some studies have examined this assump-tion. Whereas cyclosporin A and tacrolimus did not in-hibit cAMP-induced rat insulin I gene transcription in HITcells (Siemann et al. 1999), tacrolimus decreased rat in-sulin I gene transcription after stimulation by high potas-sium-induced membrane depolarization or glucose in theβ-cell line INS-1 (Lawrence et al. 2001, 2002). Evidencewas presented to suggest that tacrolimus may interferewith calcium-induced activation of the rat insulin I genethat is mediated by NFAT binding to three sites within therat insulin I gene promoter (Lawrence et al. 2001). How-ever, very high concentrations of tacrolimus were re-quired to produce this effect (1–10 µM) (Lawrence et al.2001), raising doubts about both the involvement of cal-cineurin and the clinical importance of this effect. Fur-thermore, the effect of the immunosuppressants on the hu-man insulin gene remained unclear.
The human insulin gene promoter shows a 75% ho-mology with the rat insulin I gene promoter up to basepair 240 and then diverges more markedly (Walker et al.1983). Within the first 300 bp, the structural organizationof the human insulin gene promoter, as far as it is under-stood, is related to but distinct from the one of the rat in-
sulin I gene promoter (Walker et al. 1983). For example,the human insulin gene promoter lacks the consensus E2element and harbors four, rather than one, CREs (Inagakiet al. 1992). Thus, the regulation of the human insulingene promoter has to be studied in its own right. In a sin-gle report, the effect of tacrolimus on human insulin genetranscription was investigated. It was found that tacro-limus inhibited human insulin gene transcription in HITcells incubated in a low (0.4 mM) or high (20 mM) glucoseconcentration (Redmon et al. 1996). However, the con-centration of tacrolimus (100 nM) was very high, and cy-closporin A was not used (Redmon et al. 1996), raisingagain doubts about both the involvement of calcineurinand the clinical importance of this effect.
We, therefore, examined the effect of both drugs, cy-closporin A and tacrolimus, on the transcriptional activityof the human insulin gene. We confirmed the data ofRedmon et al. (1996) and found in addition that, when hu-man insulin gene transcription was stimulated by the twomajor intracellular second messengers, cAMP and cal-cium, much lower concentrations of tacrolimus and alsocyclosporin A were sufficient to inhibit human insulingene transcription (IC50 values of 1 and 30 nM, respec-tively). Further data support a mechanism according towhich cyclosporin A and tacrolimus reduce human insulingene transcription by inhibiting the calcineurin-dependenttranscriptional activity of the cAMP- and calcium-respon-sive transcription factor CREB that was found to bind tothe human insulin gene CREs. Since the concentrations ofcyclosporin A and tacrolimus reported here to inhibit hu-man insulin gene transcription are indistinguishable fromthose concentrations that inhibit calcineurin activity, ourdata strongly suggest that inhibition of human insulingene transcription by cyclosporin A and tacrolimus is clin-ically important and represents one mechanism of the dia-betogenic effect of these immunosuppressants.
Materials and methods
Plasmid construction. The plasmids 4xSomCRELuc and5xGal4E1BLuc, and the expression vectors RSV-PKA, RSV-CREB-327, and RSV-KCREB have been described before (Krüger et al.1997; Oetjen et al. 1994; Day et al. 1989; Walton et al. 1992). Forthe plasmid –339hInsLuc, the HindIII fragment of pINCAT1-Wt(Inagaki et al. 1992) was subcloned in the forward orientation intothe HindIII site of pXP2 (Nordeen 1988). For 4xhInsCRE2Luc and4xhInsCRE4Luc, the synthetic oligonucleotides hInsCRE2 andhInsCRE4 with 5’-GATC overhangs (shown in Table 1) weretetramerized and subcloned in the forward orientation into theBamHI site of pT81Luc (Nordeen 1988). After transfection of theblank luciferase vector pXP2 the transcriptional activity was1,000-fold less when compared to the transcriptional activity of the–339hInsLuc plasmid and was not influenced by any treatment.Subcloning and plasmid isolation were performed by standardmethods. The identity and the orientation of all constructs wereconfirmed by sequencing, using the enzymatic method.
Cell culture and transfection of DNA. F9 mouse teratocarcinomacells (DSM ACC 112) were grown in DMEM medium supple-mented with 10% fetal calf serum, 100 U of penicillin/ml, and 100 µg streptomycin/ml. F9 cells were transfected by the calciumphosphate method followed by glycerol shock using 5 µg of the in-dicator plasmid, 5 µg of an expression vector for the catalytic sub-
228

unit of PKA, and 0.8 ng to 6 µg of an expression vector for CREB-327 per 6-cm dish (gelatin coated). HIT-T15 syrian hamster cells(Schwaninger et al. 1993a) were grown in RPMI 1640 mediumsupplemented with 10% fetal calf serum, 5% horse serum, 100 Uof penicillin/ml, and 100 µg of streptomycin/ml. HIT cells weretransfected by the DEAE-dextran method (Schwaninger et al. 1993a)with 2 µg of indicator plasmid, and 2 µg of the expression vectorfor KCREB, when indicated. RSV-CAT (2.5 µg for F9 cells, 0.5 µg for HIT cells) was added as a second reporter to control forpotential differences in transfection efficiency. Cells were treatedwith nifedipine, cyclosporin A or tacrolimus (at concentrations asindicated) 7 h before harvest. Cells were stimulated with forskolin (10 µM) and with KCl (final concentration 45 mM) 6 h before har-vest. Cell extracts were prepared 48 h after transfection. The lu-ciferase assay was performed as described previously (Schwanin-ger et al. 1993a). A chromatographic CAT assay was performed asdescribed (Schwaninger et al. 1993a). Thin-layer chromatographyplates were scanned by a Fuji PhosphorImager (Fujisawa GmbH,Munich, Germany).
Nuclear extracts. Nuclear extracts were prepared according toDignam et al. (1983) with the modification that after the chromatinextraction step an ammonium sulfate (0.3 g/ml) precipitation wasincluded to concentrate proteins before dialysis.
Bacterial expression of recombinant CREB. The Escherichia colistrain BL21(DE3)pLysS was transformed with the plasmid for therecombinant CREB-327 protein (Oetjen et al. 1994). Bacteria weregrown and recombinant CREB was extracted as described (Vallejoet al. 1992).
Electrophoretic mobility shift assay. Synthetic complementaryoligonucleotides with 5’-GATC overhangs (Table 1) were annealedand labeled by a fill-in reaction using α32P[dCTP] and Klenow en-zyme. Using 15 µg of protein of nuclear extracts or 1 µg of recom-binant CREB, the electrophoretic mobility shift was performed asdescribed (Oetjen et al. 1994). Briefly, nuclear extracts or recom-binant CREB were incubated with 2 µg of poly(dI-dC) as nonspe-cific competitors and with specific competitors when indicated, in20 µl (total volume) of 20 mM Hepes, pH 7.9, 5 mM MgCl2, 1 mMEDTA, 0.5 mM dithiothreitol, 70 mM KCl, and 10% glycerol. Fol-lowing a 10-min incubation on ice, the labeled oligonucleotides(10,000 to 20,000 cpm, corresponding to about 4–8 fmol) wereadded, and the incubation was continued on ice for another 15 min.The reaction mixtures were resolved on a 5% nondenaturatingpolyacrylamide gel. The gel was dried down and autoradiographedat –70°C with an intensifying screen.
Materials. Luciferin, Tween 80, and forskolin were purchasedfrom Sigma Chemical Co. (St. Louis, MO). Tacrolimus was a giftfrom Fujisawa, cyclosporin A a gift from Sandoz (Basel, Switzer-land). Forskolin was dissolved in DMSO, tacrolimus and nifedip-ine were dissolved in ethanol. A stock solution of cyclosporin Awas prepared in ethanol with 20% Tween 80 and further diluted inRPMI 1640. Controls received the solvent only.
Results
Effect of tacrolimus and cyclosporin A on human insulin gene transcription
It has been shown previously that tacrolimus (100 nM) in-hibits transcription of the human insulin gene in the β-cellline HIT incubated in a low or high glucose concentration(Redmon et al. 1996). This effect is confirmed in Fig. 1Awhich shows that, when HIT cells were incubated in a highglucose concentration (11 mM), transcription of a luciferasereporter gene under the control of the human insulin genepromoter from –339 to +112 was inhibited by 100 nMtacrolimus. Maximum inhibition (63%) was achieved af-ter 24 h treatment with the immunosuppressant (Fig. 1A).The inhibition by tacrolimus of human insulin gene tran-scription seems to be promoter-specific, since tacrolimus(100 nM) only slightly inhibited transcriptional activity ofthe Rous sarcoma virus (RSV) promoter and the cy-tomegalovirus promoter (Fig. 1A). Tacrolimus inhibitedtranscription of the human insulin gene in a concentration-dependent manner (Fig. 1B). The IC50 value was about 20 nM (Fig. 1B). In contrast, cyclosporin A in concentra-tions up to 1 µM did not inhibit transcriptional activity ofthe human insulin gene (Fig. 1B). The tacrolimus analoguesirolimus (1.1 µM) did not inhibit human insulin genetranscription (data not shown). The high concentration oftacrolimus and the lack of effect of cyclosporin A call intoquestion the clinical importance of this effect.
Effect of cyclosporin A and tacrolimus on human insulin gene transcription stimulated by cAMP and calcium
It was investigated next whether cyclosporin A and tacro-limus affect human insulin gene transcription when stim-ulated by signaling pathways. Cyclic AMP and calciumare well known major second messengers of insulin genetranscription (Docherty and Clark 1994; Fehmann andHabener 1992; German et al. 1990). Therefore, cAMP andmembrane depolarization-induced calcium influx were usedas stimuli for human insulin gene transcription in HITcells.
229
Table 1 Sequences of theoligonucleotides used. Givenare the coding strands. Mutatedbases are underlined, deletedbases are marked by “-”
Sequence of oligonucleotides Name of oligonucleotides
5′-GATCCTCCTTGGCTGACGTCAGAGAGAGAGTA-3′ SomCRE (-58/-31)GATCCAGAGTTGTTGACGTCCAATGAGCGCTA rInsCRE (-193/-168)GATCCGACTCTAATGACCCGCTGGTCCTGAGA hInsCRE1 (-219/-194)GATCCGACTCTAATGACTTGCTGGTCCTGAGA hInsCRE1mGATCCAGAGGTGCTGACGACCAAGGAGATCTA hInsCRE2 (-191/-166)GATCCAGAGGTGCTGA-GACCAAGGAGATCTA hInsCRE2mGATCCGGACAGGCTGCATCAGAAGAGGCCATA hInsCRE3 (+10/+35)GATCCGGACAGGCTAAATCAGAAGAGGCCATA hInsCRE3mGATCCAGGGCCTTTGCGTCAGGTGGGCTCAGA hInsCRE4 (+53/+78)GATCCAGGGCCTTTAAATCAGGTGGGCTCAGA hInsCRE4m

Membrane depolarization alone did not increase humaninsulin gene transcription (data not shown). Forskolin, anactivator of adenylate cyclase, stimulated the transcrip-tional activity of the human insulin gene which was not
further enhanced by the combined stimulus membrane de-polarization plus forskolin (Fig. 2). To investigate whetherthe combined stimulus depended on calcium influx, theblocker of voltage-dependent L-type calcium channels,nifedipine, was used. Nifedipine slightly increased basalactivity (Fig. 2) (P<0.01). Relative to the respective con-trol (with or without nifedipine) the increase in the tran-scriptional activity of the human insulin-reporter gene in-duced by forskolin and forskolin plus membrane depolar-ization was reduced by about 30% and 88%, respectively,in the presence of nifedipine when compared to the respec-tive increases in the absence of nifedipine (Fig. 2). Thus,although by stimulation with forskolin plus membrane de-polarization human insulin gene transcription in HIT cellsis elevated to levels similar to those reached by stimula-tion with forskolin alone (Figs. 2, 3, and 6D), it now de-pends to a larger extent (about 88%) on calcium influx. Asshown in Fig. 3, cyclosporin A (5 µM) did not alter basal
230
Fig. 1A, B Effect of tacrolimus and cyclosporin A on transcrip-tional activity of the human insulin gene. A reporter genes underthe control of the promoter of the human insulin gene promoter(from –339 to +112) (-339Ins), the Rous sarcoma virus promoter(RSV) or the promoter of the cytomegalovirus (CMV) were tran-siently transfected into HIT cells. Cells were treated with 100 nMtacrolimus as indicated. Reporter gene activity is expressed rela-tive to the mean value in each experiment of the activity measuredin the respective controls. Values are means ± SEM of three in-dependent experiments each done in duplicate. B the plasmid–339hInsLuc was transiently transfected into HIT cells. Cells weretreated for 24 h with increasing concentrations of tacrolimus(FK506) or cyclosporin A (CsA). Luciferase activity is expressedrelative to the mean value in each experiment of the activity mea-sured in the respective controls. Values are means ± SEM of fourindependent experiments, each done in duplicate
Fig. 2 Effect of nifedipine on human insulin gene transcription.The plasmid –339hInsLuc was transiently transfected into HIT cells.Cells were treated with 10 µM forskolin (cAMP), forskolin plusmembrane depolarization (45 mM KCl), and nifedipine (10 µM) asindicated. Control, no cAMP or KCl treatment. Luciferase activityis expressed relative to the mean value in each experiment of theactivity measured in the respective controls without nifedipine.Values are means ± SEM of three independent experiments, eachdone in duplicate
Fig. 3 Cyclosporin A inhibits transcriptional activation of the hu-man insulin gene promoter. The plasmid –339hInsLuc was tran-siently transfected into HIT cells. The cells were stimulated by 10 µMforskolin (cAMP) or forskolin plus KCl (45 mM). Cyclosporin A (5 µM) (CsA) was added 7 h before harvest. Luciferase activity isexpressed relative to the mean value in each experiment of the ac-tivity measured in the respective controls. Values are means±SEMof three independent experiments, each done in duplicate
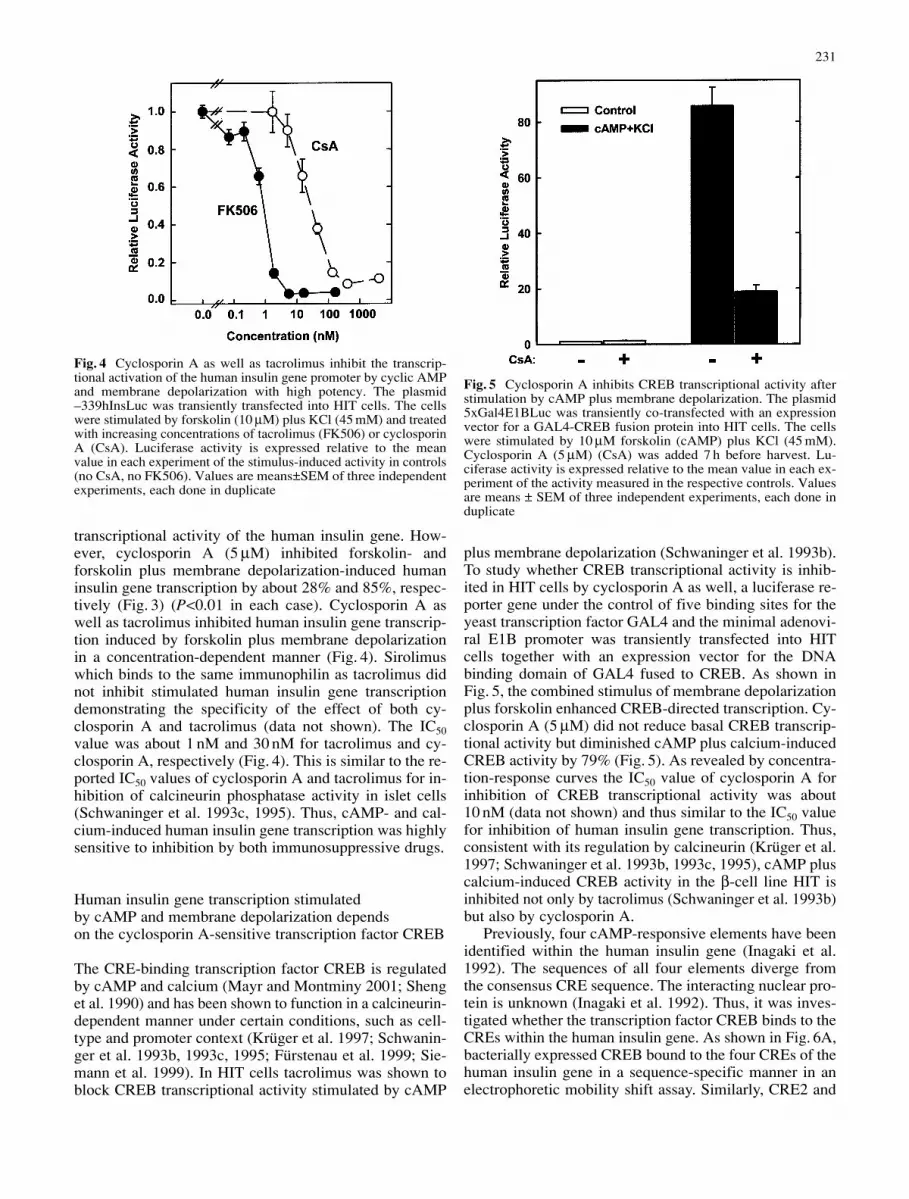
transcriptional activity of the human insulin gene. How-ever, cyclosporin A (5 µM) inhibited forskolin- andforskolin plus membrane depolarization-induced humaninsulin gene transcription by about 28% and 85%, respec-tively (Fig. 3) (P<0.01 in each case). Cyclosporin A aswell as tacrolimus inhibited human insulin gene transcrip-tion induced by forskolin plus membrane depolarizationin a concentration-dependent manner (Fig. 4). Sirolimuswhich binds to the same immunophilin as tacrolimus didnot inhibit stimulated human insulin gene transcriptiondemonstrating the specificity of the effect of both cy-closporin A and tacrolimus (data not shown). The IC50value was about 1 nM and 30 nM for tacrolimus and cy-closporin A, respectively (Fig. 4). This is similar to the re-ported IC50 values of cyclosporin A and tacrolimus for in-hibition of calcineurin phosphatase activity in islet cells(Schwaninger et al. 1993c, 1995). Thus, cAMP- and cal-cium-induced human insulin gene transcription was highlysensitive to inhibition by both immunosuppressive drugs.
Human insulin gene transcription stimulated by cAMP and membrane depolarization depends on the cyclosporin A-sensitive transcription factor CREB
The CRE-binding transcription factor CREB is regulatedby cAMP and calcium (Mayr and Montminy 2001; Shenget al. 1990) and has been shown to function in a calcineurin-dependent manner under certain conditions, such as cell-type and promoter context (Krüger et al. 1997; Schwanin-ger et al. 1993b, 1993c, 1995; Fürstenau et al. 1999; Sie-mann et al. 1999). In HIT cells tacrolimus was shown toblock CREB transcriptional activity stimulated by cAMP
plus membrane depolarization (Schwaninger et al. 1993b).To study whether CREB transcriptional activity is inhib-ited in HIT cells by cyclosporin A as well, a luciferase re-porter gene under the control of five binding sites for theyeast transcription factor GAL4 and the minimal adenovi-ral E1B promoter was transiently transfected into HITcells together with an expression vector for the DNAbinding domain of GAL4 fused to CREB. As shown inFig. 5, the combined stimulus of membrane depolarizationplus forskolin enhanced CREB-directed transcription. Cy-closporin A (5 µM) did not reduce basal CREB transcrip-tional activity but diminished cAMP plus calcium-inducedCREB activity by 79% (Fig. 5). As revealed by concentra-tion-response curves the IC50 value of cyclosporin A forinhibition of CREB transcriptional activity was about 10 nM (data not shown) and thus similar to the IC50 valuefor inhibition of human insulin gene transcription. Thus,consistent with its regulation by calcineurin (Krüger et al.1997; Schwaninger et al. 1993b, 1993c, 1995), cAMP pluscalcium-induced CREB activity in the β-cell line HIT isinhibited not only by tacrolimus (Schwaninger et al. 1993b)but also by cyclosporin A.
Previously, four cAMP-responsive elements have beenidentified within the human insulin gene (Inagaki et al.1992). The sequences of all four elements diverge fromthe consensus CRE sequence. The interacting nuclear pro-tein is unknown (Inagaki et al. 1992). Thus, it was inves-tigated whether the transcription factor CREB binds to theCREs within the human insulin gene. As shown in Fig. 6A,bacterially expressed CREB bound to the four CREs of thehuman insulin gene in a sequence-specific manner in anelectrophoretic mobility shift assay. Similarly, CRE2 and
231
Fig. 4 Cyclosporin A as well as tacrolimus inhibit the transcrip-tional activation of the human insulin gene promoter by cyclic AMPand membrane depolarization with high potency. The plasmid–339hInsLuc was transiently transfected into HIT cells. The cellswere stimulated by forskolin (10 µM) plus KCl (45 mM) and treatedwith increasing concentrations of tacrolimus (FK506) or cyclosporinA (CsA). Luciferase activity is expressed relative to the meanvalue in each experiment of the stimulus-induced activity in controls(no CsA, no FK506). Values are means±SEM of three independentexperiments, each done in duplicate
Fig. 5 Cyclosporin A inhibits CREB transcriptional activity afterstimulation by cAMP plus membrane depolarization. The plasmid5xGal4E1BLuc was transiently co-transfected with an expressionvector for a GAL4-CREB fusion protein into HIT cells. The cellswere stimulated by 10 µM forskolin (cAMP) plus KCl (45 mM).Cyclosporin A (5 µM) (CsA) was added 7 h before harvest. Lu-ciferase activity is expressed relative to the mean value in each ex-periment of the activity measured in the respective controls. Valuesare means ± SEM of three independent experiments, each done induplicate

232
Fig. 6A–D CREB binds to the human insulin gene promoter andmediates activation by cAMP and membrane depolarization. A Re-combinant CREB binding to the four CREs of the human insulingene as revealed in the electrophoretic mobility shift assay. CREBwas incubated with labeled wild-type or mutant oligonucleotidesas indicated. The sequences of the oligonucleotides used are shownin Table 1. Note that CREB binding to the four human insulin geneCREs was sequence-specific, since in each case mutations withinthe CRE sequences decreased binding. B CRE2 and CRE4 of thehuman insulin gene promoter compete for the binding of CREB innuclear extracts to labeled SomCRE in an electrophoretic mobilityshift assay. Nuclear extracts were incubated with labeled SomCREand competitors in 500-fold (lanes 1, 3, 5, 7, 10, 12, 14, 16, 19, and21) or 1,500-fold molar excess (lanes 2, 4, 6, 8, 11, 13, 15, 17, 20,and 22). Lanes 9 and 18, no competitor. Note that hInsCRE2 andhInsCRE4 of the human insulin gene compete for the binding ofCREB specifically since the competition by their respective mu-tant oligonucleotides was much less pronounced (compare lane 11with lane 13, and lane 20 with 22, respectively). A and B indicatecomplexes that have been shown to represent nuclear CREB bind-ing to the SomCRE (Oetjen et al. 1994). C CREB activates transcrip-
tion at CRE2 and CRE4 of the human insulin gene promoter. In-creasing amounts of an expression vector for CREB (RSV-CREB)were transiently transfected into F9 cells together with the plas-mids 4xSomCRELuc (SomCRE), 4xhInsCRE2Luc (hInsCRE2),and 4xhInsCRE4Luc (hInsCRE4) and a constant amount of an ex-pression vector for the catalytic subunit of PKA. Luciferase activ-ity is expressed relative to the mean value in each experiment ofthe activity measured for 4xSomCRELuc in the absence of CREB.Values are means ± SEM of three independent experiments, eachdone in duplicate. D CREB mediates the activation of the humaninsulin gene promoter by cAMP and cAMP plus membrane depo-larization as revealed by overexpression of the dominant-negativemutant KCREB. The plasmid –339hInsLuc was transiently trans-fected into HIT cells. An expression vector encoding a dominantnegative CREB mutant, KCREB, was cotransfected as indicated.Cells were treated with 10 µM forskolin (cAMP), or forskolin plusKCl (45 mM). Control, no treatment. Luciferase activity is ex-pressed relative to the mean value in each experiment of the activ-ity measured in the respective control (no KCREB). Values aremeans ± SEM of three independent experiments each done in du-plicate

CRE4 of the human insulin gene competed for the bindingof CREB in nuclear extracts from HIT cells to a highaffinity CRE (SomCRE) (Fig. 6B), suggesting that CREBis among the nuclear proteins that bind to CRE2 and CRE4of the human insulin gene. Because of overshadowingprotein binding, we were unable to clearly demonstrate nu-clear CREB binding to labeled hInsCRE2 and hInsCRE4in the electrophoretic mobility shift assay (data not shown).To test whether CREB is able to activate transcriptionthrough CRE2 or CRE4 of the human insulin gene, tran-sient transfections into the mouse teratoma cell line F9were performed. Luciferase reporter fusion genes underthe control of four copies of the SomCRE, the hInsCRE2or the hInsCRE4 were transfected together with a constantamount of an expression vector, encoding the catalyticsubunit of protein kinase A, and increasing amounts of anexpression vector for CREB-327. As shown in Fig. 6C,the expression of CREB enhanced transcription by allthree CREs. CREB enhanced transcription at the SomCRE8-fold, at the hInsCRE2 10-fold, and at the hInsCRE438-fold (Fig. 6C) Thus, CREB is able to confer transcrip-tional activity to CRE2 and to CRE4 of the human insulingene. Mutations of all four CRE-like elements within thehuman insulin gene promoter did not decrease the pro-moter’s basal transcriptional activity. However, after stim-ulation by the adenylate cyclase activator forskolin or bymembrane depolarization plus forskolin transcriptionalactivity was markedly reduced (data not shown). Thesefindings indicate that the cAMP plus calcium-inducedtranscriptional activity of the human insulin gene dependson intact CRE-like elements. To specifically address thequestion whether cellular CREB mediates activation ofthe human insulin gene, a dominant negative CREB mutant,KCREB, was employed (Walton et al. 1992). As shown inFig. 6D, the overexpression of KCREB did not change basaltranscriptional activity but inhibited almost completely thetranscriptional response to forskolin as well as the tran-scriptional response to the combined stimuli forskolinplus membrane depolarization (Fig. 6D). This effect wasspecific, since the overexpression of KCREB did not de-crease membrane depolarization-induced transcription ofthe G2 element within the rat glucagon gene promoter,binding NFAT and hepatocyte nuclear factor-3β (Fürste-nau et al. 1999) (data not shown). These data suggest thatthe transcriptional activity of the human insulin gene pro-moter stimulated by forskolin or by forskolin plus mem-brane depolarization is conferred by the tacrolimus- andcyclosporin A- sensitive transcription factor CREB orclosely related proteins.
Discussion
The present study demonstrates that human insulin genetranscription is highly sensitive to inhibition by cy-closporin A and tacrolimus when stimulated by cAMPand depolarization-induced calcium influx. This studyalso offers an explanation for this effect by establishingthat the calcineurin-dependent transcription factor CREB
binds to human insulin gene CREs and mediates its acti-vation in response to cAMP and calcium. Together thesedata suggest that inhibition of insulin gene transcription isone of the mechanisms underlying the diabetogenic actionof these clinically important immunosuppressant drugs.
A specific effect of tacrolimus on insulin gene tran-scription has been suggested only recently (Redmon et al.1996; Lawrence et al. 2001). Redmon et al. (1996) reportedthat tacrolimus (100 nM) inhibited the transcriptional ac-tivity of a human insulin promoter-reporter gene in the β-cell line HIT. Although this finding was confirmed inthe present study, further analysis revealed that tacrolimusinhibits human insulin gene transcription only at concen-trations considerably higher than those that are required to inhibit calcineurin phosphatase activity in islet cells(Schwaninger et al. 1995). Furthermore, cyclosporin Ahad no effect on human insulin gene transcription at con-centrations (up to 1 µM) that have been shown to practi-cally completely inhibit calcineurin activity in this β-cellline (Schwaninger et al. 1995). Thus, the results of thepresent study suggest that this effect of tacrolimus, due tothe high concentrations required, is unlikely to be clini-cally meaningful. However, the present study now demon-strates that much lower concentrations of tacrolimus andalso low concentrations of cyclosporin A are sufficient toinhibit the transcriptional activity of the human insulingene when stimulated by the major intracellular secondmessengers, cAMP and calcium.
Like neurons, pancreatic islet cells are electrically ex-citable. They express L-type voltage-gated calcium chan-nels, and it is well documented that membrane depolar-ization leads to an influx of calcium and an increase in cy-tosolic calcium concentration (Cook et al. 1991; Wollheimand Pralong 1990). Insulin release and gene transcriptionis stimulated by blood glucose, which has the ability todepolarize β cells and to increase the concentration of in-tracellular free calcium. A synergistic interaction betweenthe hormonally regulated cAMP-dependent signaling sys-tem and the glucose-regulated calcium-dependent signal-ing system gives β cells the ability to match the ambientconcentration of glucose to an appropriate insulin response(Holz and Habener 1992). Although the signaling path-ways activated by incretin hormones like GLP-1 and byglucose are by far more complex (Fehmann and Habener1992; German et al. 1990; Ohneda et al. 2000; Vaulont etal. 2000; Skoglund et al. 2000), the interaction betweencAMP and calcium is crucial for the regulation of insulingene transcription (Docherty and Clark 1994; Ohneda etal. 2000; Vaulont et al. 2000). These two major intracellu-lar messengers were therefore used in the present study.As has been reported before for the rat insulin I gene pro-moter (Oetjen et al. 1994), membrane depolarization didnot potentiate cAMP-induced activation of the human in-sulin gene promoter in the β-cell line HIT. However, sig-naling to the insulin promoter was different under thecombined stimulation, since the activation of the humaninsulin promoter now depended to a large extent (88%) oncalcium influx. The fact that, nevertheless, membrane de-polarization did not potentiate cAMP-induced insulin gene
233

transcription suggests that membrane depolarization influ-ences cAMP-induced insulin gene transcription throughmultiple pathways including at least one as yet unknownnegative signal as well as one positive signal resultingfrom the influx of calcium through voltage-dependent L-type calcium channels. Under this condition (cAMPplus calcium influx), the present study demonstrates thathuman insulin gene transcription is highly sensitive to in-hibition by both tacrolimus and cyclosporin A with IC50values of about 1 nM and 30 nM, respectively. The potencyof tacrolimus to inhibit human insulin gene transcriptionafter stimulation by cAMP plus calcium, as shown here, isthus about 100-fold and 1,000-fold higher than has beenreported previously for the inhibition by tacrolimus of the human (Redmon et al. 1996) or rat insulin I gene(Lawrence et al. 2001), respectively. The β-cell line HITshares many specific characteristics with fully differen-tiated normal β cells (Schwaninger et al.1993a, 1993b,Oetjen et al. 1994; Eggers et al. 1998) suggesting that theresults obtained in the present study hold true also for nor-mal β cells. This view is supported by recent data thatdemonstrate a potent inhibition of insulin gene transcrip-tion by cyclosporin A and tacrolimus in primary islet cellsusing islets from transgenic mice that carry a human in-sulin gene promoter-luciferase transgene (Oetjen, Baun,Beimesche, Krause, Cierny, Knepel; manuscript in prepa-ration).
Both tacrolimus and cyclosporin A have been associ-ated with decreased glucose tolerance following solid or-gan transplantation as a major side effect (see Introduc-tion). The mechanism is unclear. The present study showsthat, after stimulation by cAMP and calcium, cyclosporinA and tacrolimus inhibit human insulin gene transcriptionat concentrations that are known to inhibit calcineurinphosphatase activity (Schwaninger et al. 1995; Clipstoneand Crabtree 1992; O’Keefe et al. 1992). It has beenshown previously that in pancreatic islet cells, includingHIT cells, the IC50 value of inhibition of calcineurin ac-tivity is about 1 nM and 30 nM for tacrolimus and cy-closporin A, respectively (Schwaninger et al. 1993c; 1995).This is consistent with the drugs’ IC50 values for inhibi-tion of calcineurin activity in other tissues (Clipstone andCrabtree 1992; O’Keefe et al. 1992). Since the immuno-suppressive effects of cyclosporin A and tacrolimus arethought to be secondary to inhibition of calcineurin in T-cells, our data suggest that the concentrations of tacro-limus and cyclosporin A that cause immunosuppressionare indistinguishable from those that cause inhibition ofhuman insulin gene transcription. Our data thereby sug-gest that inhibition of insulin gene transcription is clini-cally important and a mechanism of the diabetogenic ef-fects associated with the clinical use of these drugs.
The results of the present study also suggest a mecha-nism through which tacrolimus and cyclosporin A inhibithuman insulin gene transcription. Both compounds wereused in the present study over a wide concentration range.Since cyclosporin A and tacrolimus are chemically distinctand bind to distinct intracellular receptors (immunophilins),they do not share other activities than inhibition of cal-
cineurin phosphatase activity (Ho et al. 1996). Thus, thecombined use of these potent inhibitors of calcineurin is apowerful tool to evaluate the role of calcineurin. The pre-sent study demonstrates for the first time an inhibition ofinsulin gene transcription by both drugs with IC50 valuessimilar to those for inhibition of calcineurin. These datastrongly suggest that calcineurin phosphatase activity isactivated by depolarization-induced calcium influx and isthen required for stimulation of human insulin gene tran-scription by cAMP and calcium. The best characterizedcAMP- and calcium-activated transcription factor is theCRE-binding protein CREB (Mayr and Montminy 2001;Sheng et al. 1990). It has been shown before that cal-cineurin phosphatase activity is required for CREB activ-ity in islet cells when stimulated by cAMP and depolar-ization-induced calcium influx (Schwaninger et al. 1993b,1993c, 1995). Consistent with this calcineurin dependency,cAMP and calcium-induced CREB activity in HIT cells isblocked by tacrolimus (Schwaninger et al. 1993b) and cy-closporin A (this study). The human insulin gene containsfour CREs that have been shown before to confer cAMPresponsiveness (Inagaki et al. 1992). The interacting tran-scription factor was unknown. The sequences of the hu-man insulin gene CREs are different from each other andfrom the consensus CRE motif (Inagaki et al. 1992).Therefore, the examination for the binding of CREB tothese CRE-like sequences was required. The present studydemonstrates that recombinant CREB binds to all fourand that cellular CREB binds to at least two of the fourCRE-like sequences in the human insulin gene promoter.The present results also strongly suggest that cellularCREB mediates the activation of the human insulin genepromoter by cAMP and calcium, since the overexpressionof a dominant-negative mutant of CREB, KCREB (Waltonet al. 1992), was found to abolish this response. Note-worthy, regulation of insulin gene transcription by CREBis highly conserved among species, although the rat in-sulin I gene carries one rather than four CREB bindingCREs which by position and sequence is closest to theCRE2 of the human insulin gene (Inagaki et al. 1992;Oetjen et al. 1994; Eggers et al. 1998; Ohneda et al. 2000).When taken together, the results suggest a model accord-ing to which tacrolimus and cyclosporin A inhibit humaninsulin gene transcription by inhibiting calcineurin and,thus, CREB activity, which is essential for the activationby cAMP and calcium of the human insulin gene.
Other CRE-binding transcription factors have beenproposed to function at the CREs of the human insulingene. Recombinant ATF-2 has been shown to bind to thehuman insulin CREs in vitro and, when overexpressed, toactivate the human insulin promoter (Inagaki et al. 1992;Ban et al. 2000). However, ATF-2 is unlikely to be involvedin the regulation of the human insulin gene by cAMP andmembrane depolarization studied here. First, ATF-2 activ-ity is not stimulated by cAMP (Ban et al. 2000). Second,the dominant-negative mutant KCREB abolished the acti-vation of the human insulin gene by cAMP and calcium(this study) but does not block ATF-2 activity (Walton etal. 1992). CREM proteins like CREMτ are closely related
234

to CREB and can heterodimerize with CREB as well asKCREB (Walton et al. 1992; Inada et al. 1999). AlthoughCREM proteins like CREMτ could thus be involved, theregulation of their transcriptional activity by calcineurinand the immunosuppressants has not been studied yet and,thus, remains unclear.
Recently, another transcription factor, NFAT, has beensuggested to mediate the inhibition of the rat insulin I geneby high concentrations of tacrolimus (1–10 µM) in INS-1cells (Lawrence et al. 2001). Originally discovered in T-cells, NFAT is now known to be expressed and regu-lated by calcineurin also in islet cells (Fürstenau et al. 1999).The rat glucagon gene has been shown to be activated inislet α-cells in response to membrane depolarizationthrough both NFAT, binding to the G2 element, and CREB,binding to a CRE (Fürstenau et al. 1999; Schwaninger etal. 1993a). By sequence inspection, two of the three NFATbinding sites in the rat insulin I gene may be conserved inthe human insulin gene (Lawrence et al. 2001). However,their functional significance is unknown. While the re-sults of the present study do not exclude the involvementof additional transcription factors like NFAT, they demon-strate an essential role of CREB or closely related proteinsin the stimulation by cAMP and calcium of the human in-sulin gene and its inhibition by the immunosuppressivedrugs tacrolimus and cyclosporin A.
Acknowledgements This work was supported by a grant fromthe Deutsche Forschungsgemeinschaft, SFB 402/A3. We appreci-ate the generous gift of pINCAT1-Wt from Dr. H. Imura, Kyoto,Japan. We thank Cordula Spinhoff for typing the manuscript.
References
Ban N, Yamada Y, Someya Y, Ihara Y, Adachi T, Kubota A,Watanabe R, Kuroe A, Inada A, Miyawaki K, Sunaga Y, ShenZP, Iwakura T, Tsukiyama K, Toyokumi S, Tsuda K, Seino Y(2000) Activating transcription factor-2 is a positive regulatorin CaM kinase IV-induced human insulin gene expression. Di-abetes 49:1142–1148
Barton K, Muthasamy N, Chanyangam M, Fischer C, ClendeninC, Leiden JM (1996) Defective thymocyte proliferation and IL-2production in transgenic mice expressing a dominant negativeform of CREB. Nature 379:81–85
Clipstone N, Crabtree GR (1992) Identification of calcineurin akey signalling enzyme in T lymphocyte activation. Nature357:695–697
Cook DL, Satin LS, Hopkins WF (1991) Pancreatic B cells arebursting, but how? Trends Neurosci 14:411–414
Day RN, Walder JA, Maurer RA (1989) A protein kinase inhibitorgene reduces both basal and multihormone-stimulated prolactingene expression. J Biol Chem 264:431–436
Dignam JD, Lebovitz RM, Roeder RG (1983) Accurate transcrip-tion initiation by RNA polymerase II in a soluble extract fromisolated mammalian nuclei. Nucleic Acids Res 11:1475–1489
Docherty K, Clark AR (1994) Nutrient regulation of insulin geneexpression. FASEB J 8:20–27
Ebihara K, Fukunaga K, Matsumoto K, Shichiri M, Miyamoto E(1996) Cyclosporin A stimulation of glucose-induced insulinsecretion in MIN6 cells. Endocrinology 137:5255–5263
Eggers A, Siemann G, Blume R, Knepel W (1998) Gene-specifictranscriptional activity of the insulin cAMP-responsive elementis conferred by NF-Y in combination with cAMP response ele-ment-binding protein. J Biol Chem 273:18499–18508
European FK506 Multicenter Study Liver Group (1994) Ran-domised trial comparing tacrolimus (FK506) and cyclosporinin prevention of liver allograft rejection. Lancet 344:423–428
Fehmann H-C, Habener JF (1992) Insulinotropic hormone glucagon-like peptide-I(7–37) stimulation of proinsulin gene expressionand proinsulin biosynthesis in insulinoma beta TC-1 cells. En-docrinology 130:159–166
Fürstenau U, Schwaninger M, Blume R, Jendrusch E-M, KnepelW (1999) Characterization of a novel calcium response ele-ment in the glucagon gene. J Biol Chem 274:5851–5860
German MS, Moss, LG, Rutter WJ (1990) Regulation of insulingene expression by glucose and calcium in transfected primaryislet cultures. J Biol Chem 265:22063–22066
Herold KC, Nagamatsu S, Buse JB, Kulsakdinum P, Steiner DF(1993) Inhibition of glucose stimulated insulin release fromβTC3 cells and rodent islets by an analog of FK506. Trans-plantation 55:186–192
Ho S, Clipstone N, Timmermann L, Northrop J, Graef I, FiorentinoD, Nourse J, Crabtree GR (1996) The mechanism of action ofcyclosporin A and FK506. Clin Immunol Immunopathol 80:S40–S45
Holz GG, Habener JF (1992) Signal transduction crosstalk in theendocrine system: pancreatic β-cells and the glucose compe-tence concept. Trends Biol Sci 17:388–393
Inada A, Someya Y, Yamda Y, Ihara Y, Kubota A, Ban N,Watanabe R, Tsuda K, Seino Y (1999) The cyclic AMP re-sponse element modulator family regulates the insulin genetranscription by interacting with the transcription factor IID. J Biol Chem 274:21095–21103
Inagaki N, Maekawa T, Sudo T, Ishii S, Seino Y, Imura H (1992)c-Jun represses the human insulin promoter activity that de-pends on multiple cAMP response elements. Proc Natl AcadSci USA 89:1045–1049
Jindal RM, Sidner RA, Milgrom ML (1997) Post-transplant dia-betes mellitus. Drug Safety 16:242–257
Kahan BD (1989) Cyclosporine. N Engl J Med 321:1725–1738Kahn CR (1994) Insulin action, diabetogenes, and the cause of
type II diabetes. Diabetes 43:1066–1084Krentz AJ, Dousset B, Mayer D, McMaster P, Buckels J, Cramb
R, Smith JM, Nattrass M (1993) Metabolic effects of cy-closporin A and FK 506 in liver transplant recipients. Diabetes42:1753–1759
Krüger M, Schwaninger M, Blume R, Oetjen E, Knepel W (1997)Inhibition of CREB- and cAMP response element-mediatedgene transcription by the immunosuppressive drugs cyclosporinA and FK506 in T cells. Naunyn-Schmiedebergs Arch Pharma-col 356:433–440
Lawrence MC, Bhatt HS, Watterson JM, Easom RA (2001) Regu-lation of insulin gene transcription by a Ca2+-responsive path-way involving calcineurin and nuclear factor of activated Tcells. Mol Endocrinol 15:1758–1767
Lawrence MC, Bhatt HS, Easom RA (2002) NFAT regulates in-sulin gene promoter activity in response to synergistic path-ways induced by glucose and glucagon-like peptide-1. Diabetes51:691–698
Lohmann T, List C, Lamesch P, Kohlhaw K, Wenzke M, SchwarzC, Richter O, Hauss J, Seissler J (2000) Diabetes mellitus andislet cell specific autoimmunity as adverse effects of immuno-suppressive therapy with FK506/tacrolimus. Exp Clin En-docrinol Diabetes 108:347–352
Mayr B, Montminy M (2001) Transcriptional regulation by thephosphorylation-dependent factor CREB. Nat Rev Mol CellBiol 2:599–609
Nordeen SK (1988) Luciferase reporter gene vectors for analysisof promoters and enhancers. Biotechnique 6:454–458
Oetjen E, Diedrich T, Eggers A, Eckert B, Knepel W (1994) Dis-tinct properties of the cAMP responsive element of the rat in-sulin I gene. J Biol Chem 269:27036–27044
Ohneda K, Ee H, German M (2000) Regulation of insulin genetranscription. Cell Dev Biol 11:227–233
235

O’Keefe S, Tamura J, Kincaid RL, Tocci MJ, O’Neill EA (1992)FK-506- and CsA-sensitive activation of the interleukin-2 pro-moter by calcineurin. Nature 357:692–694
Rao A, Luo C, Hogan PG (1997) Transcription factors of theNFAT family: regulation and function. Annu Rev Immunol 15:707–747
Redmon JB, Olson LK, Armstrong MB, Greene MJ, Robertson RP(1996) Effects of tacrolimus (FK506) on human insulin geneexpression, insulin mRNA levels, and insulin secretion in HITT15 cells. J Clin Invest 98:2786–2793
Rudolph D, Tafuri A, Gass P, Hammerling GJ, Arnold B, SchützG (1998) Impaired fetal T cell development and perinatallethality in mice lacking the cAMP response element bindingprotein. Proc Natl Acad Sci USA 95:4481–4486
Rühlmann A, Nordheim A (1997) Effects of the immunosuppres-sive drugs CsA and FK506 on intracellular signalling and generegulation. Immunobiology 198:192–206
Saltiel AR (2001) New perspectives into the molecular pathogene-sis and treatment of type 2 diabetes. Cell 104:517–529
Schwaninger M, Lux G, Blume R, Oetjen E, Hidaka H, Knepel W(1993a) Membrane depolarization and calcium influx induceglucagon gene transcription in pancreatic islet cells through thecyclic AMP-responsive element. J Biol Chem 268:5168–5177
Schwaninger M, Blume R, Oetjen E, Lux G, Knepel W (1993b)Inhibition of cAMP responsive element-mediated gene tran-scription by cyclosporin A and FK506 after membrane depo-larization. J Biol Chem 268:23111–23115
Schwaninger M, Blume R, Oetjen E, Knepel W (1993c) The im-munosuppressive drugs cyclosporin A and FK506 inhibit cal-cineurin phosphatase activity and gene transcription mediatedthrough the cAMP-responsive element in a non-immune cellline. Naunyn-Schmiedebergs Arch Pharmacol 348:541–545
Schwaninger M, Blume R, Krüger M, Lux G, Oetjen E, Knepel W(1995) Involvement of the Ca(2+) dependent phosphatase cal-cineurin in gene transcription that is stimulated by cAMPthrough cAMP response elements. J Biol Chem 270:8860–8866
Siemann G, Blume R, Grapentin D, Oetjen E, Schwaninger M,Knepel W (1999) Inhibition of cyclic AMP response element-binding protein/cyclic AMP response element-mediated tran-scription by the immunosuppressive drugs cyclosporin A andFK506 depends on the promoter context. Mol Pharmacol 55:1094–1100
Shapiro AM, Lakey JR, Ryan EA, Korbutt GS, Toth E, WarnockGL, Kneteman NM, Rajotte RV (2000) Islet transplantation inseven patients with type 1 diabetes mellitus using a glucocorti-coid-free immunosuppressive regimen. N Engl J Med 343:289–290
Sheng M, McFadden G, Greenberg ME (1990) Membrane depo-larization and calcium induce c-fos transcription via phosphor-ylation of transcription factor CREB. Neuron 4:571–582
Skoglund G, Hussain MA, Holz GG (2000) Glucagon-like peptide1 stimulates insulin gene promoter activity by protein kinaseA-independent activation of the rat insulin I gene cAMP re-sponse element. Diabetes 49:1156–1164
Teuscher AU, Seaquist ER, Robertson RP (1994) Diminished in-sulin secretory reserve in diabetic pancreas transplant and non-diabetic kidney transplant recipients. Diabetes 43:593–598
U.S. Multicenter FK506 Liver Study Group (1994) A comparisonof tacrolimus (FK 506) and cyclosporine for immunosuppres-sion in liver transplantation. N Engl J Med 331:1110–1115
Vallejo M, Penchuk L, Habener JF (1992) Somatostatin gene up-stream enhancer element activated by a protein complex con-sisting of CREB, Isl-1-like, and alpha-CBF-like transcriptionfactors. J Biol Chem 267:12876–12884
Vaulont S, Vasseur-Cognet M, Kahn A (2000) Glucose regulationof gene transcription. J Biol Chem 275:31555–31558
Walker MD, Edlund T, Boulet AM, Rutter WJ (1983) Cell-spe-cific expression controlled by the 5’ flanking regions of the in-sulin and chymotrypsin genes. Nature 306:557–581
Walton KM, Rehfuss RP, Chrivia JC, Lochner JE, Goodman RH(1992) A dominant repressor of cyclic adenosine 3’,5’-mono-phosphate (cAMP)-regulated enhancer binding protein activityinhibits the cAMP-mediated induction of the somatostatin pro-moter in vivo. Mol Endocrinol 6:647–655
Wollheim CB, Pralong WF (1990) Cytoplasmic calcium ions andother signalling events in insulin secretion. Biochem Soc Trans18:111–114
236