rad5a ubiquitin ligase is involved in ubiquitination of arabidopsis thaliana proliferating cell...
TRANSCRIPT

Journal of Experimental Botany, Vol. 64, No. 4, pp. 859–869, 2013doi:10.1093/jxb/ers368 Advance Access publication 10 January, 2013
<journal-title>doi:<DOI>
© The Author [2013]. Published by Oxford University Press [on behalf of the Society for Experimental Biology]. All rights reserved. For permissions, please email: [email protected]
ReseaRch papeR
RAD5a ubiquitin ligase is involved in ubiquitination of Arabidopsis thaliana proliferating cell nuclear antigen
Wojciech Strzalka1,†, Filip Bartnicki1,*, Katarzyna Pels1,*, Agata Jakubowska1, Toshiki Tsurimoto2 and Katsunori Tanaka3
1 Department of Plant Biotechnology, Faculty of Biochemistry, Biophysics and Biotechnology, Jagiellonian University, Gronostajowa 7, 30–387 Krakow, Poland2 Department of Biology, School of Sciences, Kyushu University, 6-10-1 Hakozaki, Higashi-ku, Fukuoka 812–8581, Japan3 Department of Bioscience and Research Centre for Environmental Bioscience, School of Science and Technology Kwansei Gakuin University, Sanda, Hyogo 669–1337, Japan
* These authors contributed equally to this work.† To whom correspondence should be addressed. E-mail: [email protected]
Received 29 June 2012; Revised 3 December 2012; Accepted 3 December 2012
Abstract
The proliferating cell nuclear antigen (PCNA) is post-translationally modified by ubiquitin in yeast and mammalian cells. It is widely accepted that in yeast mono- and polyubiquitinated PCNA is involved in distinct pathways of DNA postrep-lication repair. This study showed an interaction between plant ubiquitin and PCNA in the plant cell. Using different approaches, it was demonstrated that Arabidopsis RAD5a ubiquitin ligase is involved in the post-translational modi-fication of plant PCNA. A detailed analysis of the properties of selected Arabidopsis ubiquitin-conjugating enzymes (AtUBC) has shown that a plant homologue of yeast RAD6 (AtUBC2) is sufficient to monoubiquitinate AtPCNA in the absence of ubiquitin ligase. Using different combinations of selected AtUBC proteins together with AtRAD5a, it was demonstrated that plants have potential to use different pathways to ubiquitinate PCNA. The analysis of Arabidopsis PCNA1 and PCNA2 did not demonstrate substantial differences in the ubiquitination pattern between these two pro-teins. The major ubiquitination target of Arabidopsis PCNA, conserved in eukaryotes, is lysine 164. Taken together, the presented results clearly demonstrate the involvement of Arabidopsis UBC and RAD5a proteins in the ubiquitination of plant PCNA at lysine 164. The data show the complexity of the plant ubiquitination system and open new questions about its regulation in the plant cell.
Key words: Arabidopsis thaliana, PCNA, ubiquitin.
Introduction
The proliferating cell nuclear antigen (PCNA) was discovered in human cells over three decades ago. This protein is a key component of the DNA replication machinery present in the nuclei of all dividing cells in eukaryotes. In the first years of studies, PCNA was shown to play the role of an accessory factor for DNA polymerase δ, required for the synthesis of a new DNA strand (Tan et al., 1986; Bravo et al., 1987; Prelich et al., 1987). The comparative analysis of yeast (Krishna et al., 1994), human (Gulbis et al., 1996), and Arabidopsis (Strzalka
et al., 2009) PCNA crystal structures indicates high conser-vation of this protein within eukaryotes. In general, PCNA forms a trimeric ring structure with pseudo-6-fold symmetry, encircling the DNA helix (Krishna et al., 1994; Gulbis et al., 1996; Strzalka et al., 2009). The other evidence supporting the conserved function of PCNA originates from unrelated organisms (plants and mammals) and was shown in studies where recombinant rice and runner bean PCNA were able to stimulate the enzymatic activity of human DNA polymerase
by guest on May 7, 2016
http://jxb.oxfordjournals.org/D
ownloaded from

860 | Strzalka et al.
δ (Matsumoto et al., 1994; Strzalka et al., 2010). Conversely, studies on mammalian PCNA have shown its ability to stimu-late the activity and processivity of two wheat δ-like polymer-ases (Laquel et al., 1993).
PCNA, as a sliding clamp, also plays the role of a docking station for other proteins required in DNA metabolism (DNA replication and repair) and cell cycle control (Strzalka and Ziemienowicz, 2011). The best-characterized PCNA-binding motif present in PCNA-interacting proteins is called PIP box (Vivona and Kelman, 2003). Until now, the majority of stud-ies have concentrated on yeast and human PCNA. As a con-sequence of this tendency, knowledge about plant PCNA is very poor. The number of identified plant PCNA-interacting proteins is very limited in comparison with human and yeast (Vivona and Kelman, 2003). For a long time, studies on plant PCNA involved mainly the identification of PCNA genes in, for example, rice (Suzuka et al., 1991) and carrot (Hata et al., 1992) and, more recently, common bean (Strzalka and Ziemienowicz, 2007) and runner bean (Strzalka et al., 2010).
In the last decade, the importance of PCNA post-trans-lational modifications in genome stability and DNA repair has been reported (Strzalka and Ziemienowicz, 2011). Among several types of PCNA post-translational modifica-tions, including acetylation, methylation, and phosphoryla-tion and two others, sumoylation and ubiquitination, seem to play a major role in the maintenance of genomic stability (Stoimenov and Helleday, 2009). PCNA sumoylation was shown for yeast (Hoege et al., 2002), Xenopus (Leach and Michael, 2005), chicken (Arakawa et al., 2006) and recently Arabidopsis (Strzalka et al., 2012). Ubiquitin, similarly to SUMO, may post-translationally modify a great number of cellular proteins. Ubiquitin is synthesized as an inactive pre-cursor and must be cleaved by a specific protease to form a mature polypeptide with a glycine residue at the C-terminus (Kerscher et al., 2006). In the next step, ubiquitin is activated by the ubiquitin-activating enzyme (E1). From E1, ubiquitin is passed on to the ubiquitin-conjugating enzyme (E2 or UBC) and finally it is conjugated to the substrate, either directly or with the aid of E3 ubiquitin ligase, resulting in the cova-lent modification of the target lysine residue in the substrate (Kerscher et al., 2006). Ubiquitin mediates a wide spectrum of processes, for example protein degradation, kinase activation, DNA repair, and vesicle trafficking (Sun and Chen, 2004). There is a multitude of different E2 and E3 in eukaryotes (Sun and Chen, 2004; Kraft et al., 2005; Vierstra, 2009). The spec-ificity of target protein ubiquitination, the site of ubiquitin attachment to the substrate protein, and the chain length and type of linkage between ubiquitin molecules is dependent on the E2/E3 combination (Hershko and Ciechanover, 1998).
The ubiquitination of PCNA was reported in budding yeast (Hoege et al., 2002), fission yeast (Frampton et al., 2006), chicken (Arakawa et al., 2006), and Xenopus (Leach and Michael, 2005) cells. The modification of PCNA occurs in response to the stalling DNA replication fork, which acti-vates a specific mechanism called DNA postreplication repair (PRR, Bergink and Jentch, 2009; Strzalka and Ziemienowicz, 2011). Monoubiquitinated PCNA promotes the recruit-ment of translesion (TLS) DNA polymerases that facilitate
DNA damage bypass (Lee and Myung, 2008). On the other hand, polyubiquitinated PCNA is thought to promote TLS-independent damage avoidance through template switching, although the molecular mechanism is not clearly understood (Lee and Myung, 2008).
Following previous studies of plant PCNA sumoylation, this study demonstrates, as far as is known for the first time, that plant PCNA can also be post-translationaly ubiquit-inated. Using a plant ubiquitination system reconstituted in bacteria, the ubiquitination of Arabidopsis PCNA is charac-terized in detail and a putative model of plant PCNA ubiqui-tination pathways is proposed.
Materials and methods
Plasmid constructionThe open reading frames of mature ubiquitin AtUBQ (At2g36170), AtUBA1 (At2g30110), AtUBC2 (AT2G02760), AtUBC30 (AT5G56150), AtUBC35 (AT1G78870), and AtMMZ1 (At1g23260) were purchased from the Arabidopsis Biological Stock Centre. AtRAD5a and AtUBQ K63/R were purchased from GeneScript USA. The source of AtPCNA1 (AT1G07370) and AtPCNA2 (AT2G29570) was previously described (Strzalka et al., 2009). The open reading frames were amplified using PCR. The PCR reactions were performed in a 100 µL volume, containing 1 × PCR buffer with 2 mM MgCl2, 0.2 µM dNTPs, 5 units of Easy-A DNA polymerase (Agilent Technologies), 2 mM of each primer (Supplementary Table S1, available at JXB online) and 1 ng plasmid template. The prelimi-nary denaturation step was performed at 94 °C for 3 min. Next, there were 30 cycles of denaturation at 94 °C for 40 s, annealing at 50 °C for 30 s, and DNA synthesis at 72 °C for 4 min, followed by incubation at 72 °C for 5 min, in a thermocycler (Eppendorf). For Escherichia coli expression vectors (Supplementary Table S1), PCR products were purified, digested with restriction enzymes, ligated using T4 DNA ligase (Fermentas), and sequenced. To prepare agrobacterial vec-tors used in the bimolecular fluorescence complementation (BiFC) assay (Supplementary Table S1), the purified PCR products were first introduced into the pDONR221 vector using a gateway BP reaction, followed by sequencing, and then transferred to the final agrobacte-rial vector using an LR reaction. AtPCNA1/2 K164/R mutants were prepared using the QuikChange II Site Directed Mutagenesis Kit (Stratagene) according to the protocol supplied by the producer.
Bimolecular fluorescence complementation analysisThe BiFC assay was performed according to the protocol described by Yang et al. (2000) with some modifications. Wild-type Nicotiana benthamiana plants were grown in soil under long-day conditions (16/8 light/dark cycles) at 23 °C. Binary vectors carrying AtPCNA1_GFP, AtPCNA1_CtermGFP, AtPCNA2_GFP, AtPCNA2_CtermGFP, GFP_AtUBQ (GG), NtermGFP_AtUBQ (GG), AtCYCA1_NtermGFP, NtermGFP_AtPCNA1, Nterm_AtPCNA2, CtermGFP_AtRAD5a, and GFP_AtRAD5a were introduced into Agrobacterium tumefaciens C58. Next, Agrobacterium strains were grown for 2 days at 28 °C in YEB medium supplemented with appro-priate antibiotics (l–1: 20 mg gentamycin, 100 mg spectinomycin, and 100 mg rifampicin). After incubation, Agrobacterium cells were cen-trifuged and resuspended in infiltration buffer (10 mM MES, 10 mM MgCl2, 100 µM acetosyringone; pH 5.6). The final OD600 of the cul-ture was adjusted to 0.5. For coexpression studies, 500 µl of each bacterial culture were mixed and incubated at room temperature for 2–5 h. The final bacterial mixture was injected to the lower epider-mis of tobacco leaves. After 3–5 days, the infiltrated area was tested using a Zeiss LSM 510 META laser scanning microscope. Three independent infiltrations per combination were analysed.
by guest on May 7, 2016
http://jxb.oxfordjournals.org/D
ownloaded from

Arabidopsis PCNA ubiquitination | 861
Preparation of columns with bound AtPCNA2 and BSAAtPCNA2 was purified as described previously (Strzalka et al., 2009). BSA was purchased from BioShop Canada. Recombinant AtPCNA2 or BSA (20 mg) in binding buffer (25 mM MES, 100 mM NaCl, 0.01% Nonidet P-40; pH 6.0) were mixed with 2 ml Affi-Gel 15 (Bio-Rad) for 6 h at 4 °C. After removal of the supernatant, the beads were incubated with 0.1 M ethanolamine (pH 8.2), at 4 °C overnight and washed with 10 ml plant protein extraction buffer (30 mM Tris, 100 mM NaCl, 1 mM EDTA, 1 mM DTT, 1 mM PMSF, 10%, v/v, glycerol, 0.01%, v/v, Nonidet P-40; pH7.4).
Preparation of plant protein extract: AtPCNA2 affinity chromatographyArabidopsis thaliana T87 suspension cell culture was grown as described in a protocol published by Ogawa et al. (2008). An ali-quot (20 g) of a 10-day-old culture was centrifuged (5000 g, 15 min, 4 °C) washed with 30 mM Tris (pH 7.4) and centrifuged again. The pellet was frozen in liquid nitrogen and ground in a mortar. Next, the disrupted cells were resuspended in the plant protein extraction buffer supplemented with plant protease inhibitor cocktail (Sigma) and centrifuged (30,000 g, 60 min, 4 °C). The supernatant was first loaded on the 2 ml BSA column and the flow-through was loaded on the 2 ml AtPCNA2 column. Next the AtPCNA2 column was washed with 50 volumes of extraction buffer and the bound proteins were eluted in the same buffer containing 0.3 M NaCl.
Mass spectrometry analysisThe protein sample eluted from the AtPCNA2 column was sepa-rated in a 5–20% gradient SDS-polyacrylamide gel and stained with Coomassie Brilliant Blue. The gel lane containing the separated sam-ple was divided into 24 sectors. The substrate proteins in each sector were reduced, alkylated, and digested with trypsin as described previ-ously (Ohta et al., 2002). After digestion, the peptides were extracted and analysed by LC/MS/MS mass spectrometry at the Medical Insitute of Bioregulation in Kyushu University. Molecular masses of the detected peptides were checked against the non-redundant NCBI database for high fits with the ion spectrum data generated by LC/MS/MS, using the MASCOT program (Matrix Science).
In bacteria reconstitution of the PCNA ubiquitination systemE. coli BL21 (DE3) cells were transformed with the appropriate set of plasmids (Supplementary Tables S1 and S2). The expression of all proteins was tested by Western blotting (data not shown). The transformed cells were grown at 37 °C in 20 ml LB medium con-taining the appropriate antibiotic (l–1: 25 mg chloramphenicol, 50 mg kanamycin, 100 mg spectinomycin or 100 mg ampicillin). When the culture reached OD595 0.6, the temperature was decreased to 20 °C and protein overexpression was induced using 0.2 mM isopropyl-β-d-thiogalactopyranoside. After 20 h induction, 10 ml cells were har-vested by centrifugation (5000 g for 15 min at 4 °C), resuspended in 200 µl of PBS containing polyhistidine-tagged protein inhibitor (Sigma), and lysozyme (1 mg ml–1) and kept on ice for 30 min, fol-lowed by sonication for 1 min (5 s pulses, 10 s break, 4 °C). The sonicated cells were centrifuged at 35,000 g for 20 min at 4 °C. After centrifugation, protein concentration was measured using the BIO-RAD protein assay (Bio-Rad). Total protein extract (40 µg) were mixed with SDS-PAGE loading buffer, denatured at 100 °C for 5 min, and kept at –20 °C until use.
Western blotting analysisTotal protein extract (40 µg) from E. coli cells overexpressing the reconstituted plant ubiquitination system and AtPCNA were sepa-rated in 12% SDS-PAGE and electrotransferred onto a PVDF mem-brane (0.2 µm, Millipore), as described previously (Towbin et al., 1979). After the transfer, the membrane was washed in 1 × PBS
containing 0.5% Tween 20 (PBS-T) and blocked with PBST supple-mented with 5% fat-free milk (PBS-TB) for 30 min. The membrane was incubated overnight with a monoclonal anti-c-myc antibody at 4 °C (Sigma, dilution 1:20,000). After washing in PBS-TB, the mem-brane was incubated for 1 h with a goat anti-mouse IgG-horseradish peroxidase conjugate (Pierce, dilution 1:20,000). After extensive washes in PBS-T, proteins were detected using Super Signal West Pico Chemiluminescent substrate (Pierce) with the help of a Multi Spectral Imaging System (UVP).
Results
Analysis of the interaction between AtPCNA, ubiquitin, and AtRAD5a in the plant cell
Leaves of N. benthamiana were used to study the interac-tion of Arabidopsis PCNA and ubiquitin in vivo using BiFC analysis. Epidermal cells of N. benthamiana were transformed with selected combinations of A. tumefaciens strains carry-ing different plasmids: one containing an N-terminal GFP fragment fused to ubiquitin (AtUBQ), with two glycine (NtermGFP_AtUBQ GG) residues at the C-terminus, and the other with AtPCNA fused to a C-terminal GFP frag-ment (AtPCNA1_CtermGFP or AtPCNA2_CtermGFP). During confocal microscopy analysis of transiently trans-formed leaves, GFP fluorescence could be seen in the samples where AtPCNA_CtermGFP and NtermGFP_AtUBQ GG (which can covalently modify proteins) were overexpressed, as well as in the samples expressing AtPCNA1, 2 and AtUBQ fused to full-length GFP (Fig. 1). In addition, epidermal cells of N. benthamiana were transformed using a mixture of A. tumefaciens strains, one containing an C-terminal GFP fragment fused to AtRAD5a (Cterm_AtRAD5a) and the other with the N-terminal GFP fragment fused to AtPCNA (Nterm_AtPCNA1 or Nterm_AtPCNA2). During confocal microscopy analysis of transiently transformed leaves, GFP fluorescence could be observed in the samples overexpress-ing Nterm_AtPCNA and CtermGFP_AtRAD5a as well as in the sample with full-length GFP fused to AtRAD5a (Fig. 1). In the control samples (NtermGFP_AtUbq GG, AtPCNA1_CtermGFP, AtPCNA2_CtermGFP, AtPCNA1_CtermGFP and AtCYCA1_NtermGFP, NtermGFP_AtPCNA, Cterm GFP_AtRAD5a), no fluorescence signal was observed. The results of this analysis show that interactions between plant PCNA and ubiquitin occur both in the cytoplasm and in the nucleus. Moreover, the experiment revealed that AtRAD5a is localized the nucleus and that the interaction between AtPCNA and AtRAD5a occurs in the nucleus and not in the cytoplasm.
Investigating the potential of AtUBC2, AtUBC30, and AtUBC35 to directly ubiquitinate AtPCNA
To elucidate whether ubiquitin can be directly transferred from the AtUBC-ubiquitin complex onto AtPCNA, an Arabidopsis ubiquitination system reconstituted in bacteria was used. AtUBA1 (E1), selected AtUBC (E2), AtUBC2 (ScRAD6 homologue), AtUBC30 (ScUBC4 homologue), or AtUBC35 (ScUBC13 homologue), and AtUBQ (GG or
by guest on May 7, 2016
http://jxb.oxfordjournals.org/D
ownloaded from

862 | Strzalka et al.
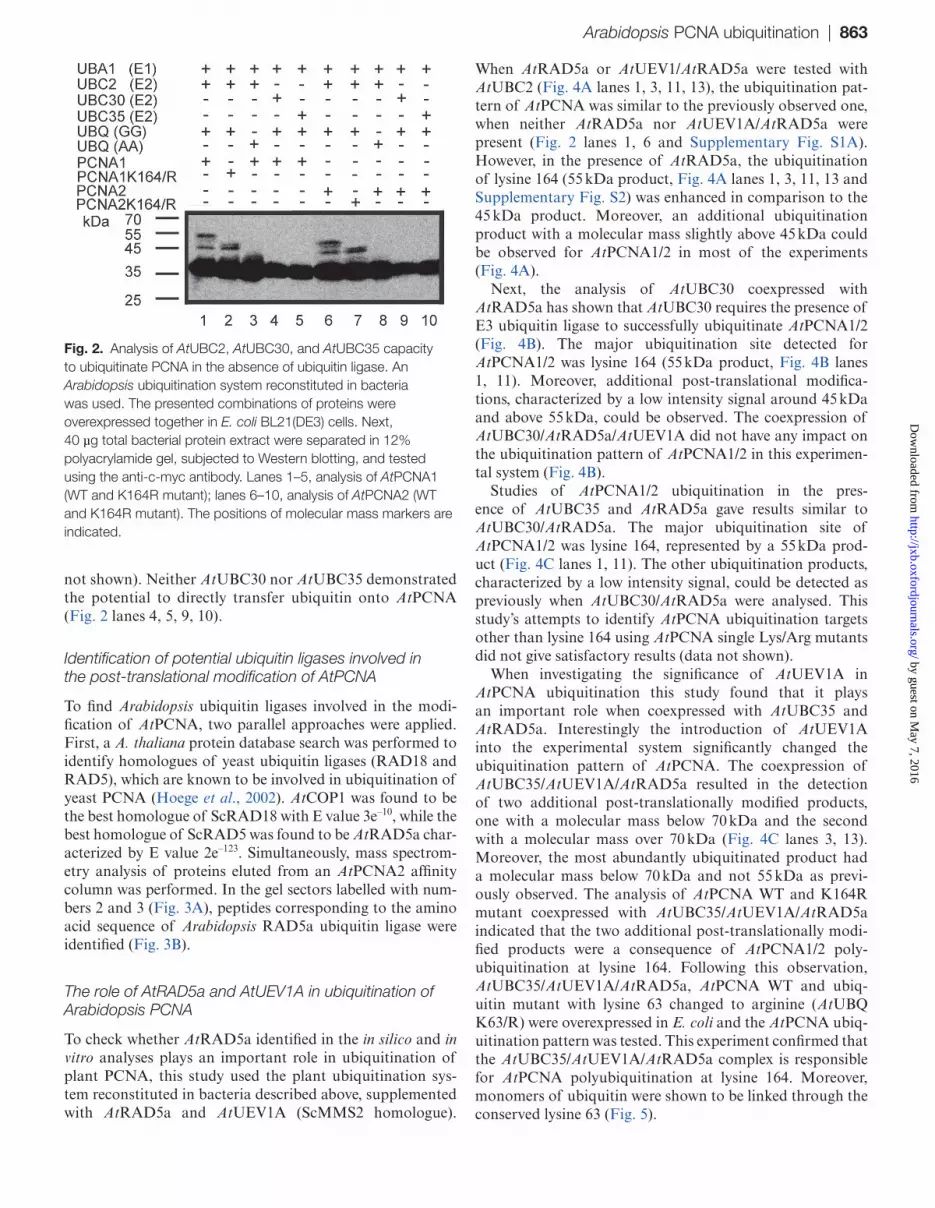
AA C-terminal motif variant) were simultaneously overex-pressed with either His-c-myc-AtPCNA1/2 wild type (WT) or His-c-myc-AtPCNA1/2 K164/R mutant (with lysine 164 changed to arginine) in E. coli host cells (Supplementary Table S2). The post-translational modification pattern of AtPCNA proteins was analysed by Western blotting using anti-c-myc antibodies. The ubiquitination of AtPCNA was observed only when AtUBQ GG variant was overexpressed with AtUBC2 (Fig. 2 lanes 1, 2, 6, 7). In this case, two
different products of ubiquitination, characterized by dif-ferent molecular masses, were observed for both AtPCNA1 and AtPCNA2. The first product had a molecular mass around 45 kDa, the second around 55 kDa (Fig. 2). When AtPCNA1/2 K164R mutants were tested, the modification product of about 55 kDa was not observed (Fig. 2 lanes 2, 7). The analysis of other lysine/arginine AtPCNA mutants failed to identify a second target residues, the modification of which would result in a product of around 45 kDa (data
Fig. 1. Images of Nicotiana benthamiana leaf epidermal cells showing transient expression of: (A) split GFP complex formed between AtPCNA1_CtermGFP and NtermGFP_AtUBQ (GG), (B) AtPCNA1_CtermGFP, (C) AtPCNA1_GFP, (D) split GFP complex formed between AtPCNA2_CtermGFP and NtermGFP_AtUBQ (GG), (E) AtPCNA2_CtermGFP, (F) AtPCNA2_GFP, (G) split AtPCNA1_CtermGFP and AtCYCA1_NtermGFP, (H) NtermGFP_AtUBQ (GG), (I) GFP_AtUBQ (GG), (J) split CtermGFP_AtRAD5a and NtermGFP_PCNA1, (K) Nterm_AtPCNA1, (L) CtermGFP_AtRAD5a and NtermGFP_AtPCNA2, (M) NtermGFP_AtPCNA2, (N) CtermGFP_AtRAD5a, and (O) GFP_AtRAD5a. All the images are overlays of bright field and green fluorescence channel. AtUBQ (GG) represent ubiquitin which can covalently modify proteins.
by guest on May 7, 2016
http://jxb.oxfordjournals.org/D
ownloaded from
Arabidopsis PCNA ubiquitination | 863
not shown). Neither AtUBC30 nor AtUBC35 demonstrated the potential to directly transfer ubiquitin onto AtPCNA (Fig. 2 lanes 4, 5, 9, 10).
Identification of potential ubiquitin ligases involved in the post-translational modification of AtPCNA
To find Arabidopsis ubiquitin ligases involved in the modi-fication of AtPCNA, two parallel approaches were applied. First, a A. thaliana protein database search was performed to identify homologues of yeast ubiquitin ligases (RAD18 and RAD5), which are known to be involved in ubiquitination of yeast PCNA (Hoege et al., 2002). AtCOP1 was found to be the best homologue of ScRAD18 with E value 3e–10, while the best homologue of ScRAD5 was found to be AtRAD5a char-acterized by E value 2e–123. Simultaneously, mass spectrom-etry analysis of proteins eluted from an AtPCNA2 affinity column was performed. In the gel sectors labelled with num-bers 2 and 3 (Fig. 3A), peptides corresponding to the amino acid sequence of Arabidopsis RAD5a ubiquitin ligase were identified (Fig. 3B).
The role of AtRAD5a and AtUEV1A in ubiquitination of Arabidopsis PCNA
To check whether AtRAD5a identified in the in silico and in vitro analyses plays an important role in ubiquitination of plant PCNA, this study used the plant ubiquitination sys-tem reconstituted in bacteria described above, supplemented with AtRAD5a and AtUEV1A (ScMMS2 homologue).
When AtRAD5a or AtUEV1/AtRAD5a were tested with AtUBC2 (Fig. 4A lanes 1, 3, 11, 13), the ubiquitination pat-tern of AtPCNA was similar to the previously observed one, when neither AtRAD5a nor AtUEV1A/AtRAD5a were present (Fig. 2 lanes 1, 6 and Supplementary Fig. S1A). However, in the presence of AtRAD5a, the ubiquitination of lysine 164 (55 kDa product, Fig. 4A lanes 1, 3, 11, 13 and Supplementary Fig. S2) was enhanced in comparison to the 45 kDa product. Moreover, an additional ubiquitination product with a molecular mass slightly above 45 kDa could be observed for AtPCNA1/2 in most of the experiments (Fig. 4A).
Next, the analysis of AtUBC30 coexpressed with AtRAD5a has shown that AtUBC30 requires the presence of E3 ubiquitin ligase to successfully ubiquitinate AtPCNA1/2 (Fig. 4B). The major ubiquitination site detected for AtPCNA1/2 was lysine 164 (55 kDa product, Fig. 4B lanes 1, 11). Moreover, additional post-translational modifica-tions, characterized by a low intensity signal around 45 kDa and above 55 kDa, could be observed. The coexpression of AtUBC30/AtRAD5a/AtUEV1A did not have any impact on the ubiquitination pattern of AtPCNA1/2 in this experimen-tal system (Fig. 4B).
Studies of AtPCNA1/2 ubiquitination in the pres-ence of AtUBC35 and AtRAD5a gave results similar to AtUBC30/AtRAD5a. The major ubiquitination site of AtPCNA1/2 was lysine 164, represented by a 55 kDa prod-uct (Fig. 4C lanes 1, 11). The other ubiquitination products, characterized by a low intensity signal, could be detected as previously when AtUBC30/AtRAD5a were analysed. This study’s attempts to identify AtPCNA ubiquitination targets other than lysine 164 using AtPCNA single Lys/Arg mutants did not give satisfactory results (data not shown).
When investigating the significance of AtUEV1A in AtPCNA ubiquitination this study found that it plays an important role when coexpressed with AtUBC35 and AtRAD5a. Interestingly the introduction of AtUEV1A into the experimental system significantly changed the ubiquitination pattern of AtPCNA. The coexpression of AtUBC35/AtUEV1A/AtRAD5a resulted in the detection of two additional post-translationally modified products, one with a molecular mass below 70 kDa and the second with a molecular mass over 70 kDa (Fig. 4C lanes 3, 13). Moreover, the most abundantly ubiquitinated product had a molecular mass below 70 kDa and not 55 kDa as previ-ously observed. The analysis of AtPCNA WT and K164R mutant coexpressed with AtUBC35/AtUEV1A/AtRAD5a indicated that the two additional post-translationally modi-fied products were a consequence of AtPCNA1/2 poly-ubiquitination at lysine 164. Following this observation, AtUBC35/AtUEV1A/AtRAD5a, AtPCNA WT and ubiq-uitin mutant with lysine 63 changed to arginine (AtUBQ K63/R) were overexpressed in E. coli and the AtPCNA ubiq-uitination pattern was tested. This experiment confirmed that the AtUBC35/AtUEV1A/AtRAD5a complex is responsible for AtPCNA polyubiquitination at lysine 164. Moreover, monomers of ubiquitin were shown to be linked through the conserved lysine 63 (Fig. 5).
Fig. 2. Analysis of AtUBC2, AtUBC30, and AtUBC35 capacity to ubiquitinate PCNA in the absence of ubiquitin ligase. An Arabidopsis ubiquitination system reconstituted in bacteria was used. The presented combinations of proteins were overexpressed together in E. coli BL21(DE3) cells. Next, 40 µg total bacterial protein extract were separated in 12% polyacrylamide gel, subjected to Western blotting, and tested using the anti-c-myc antibody. Lanes 1–5, analysis of AtPCNA1 (WT and K164R mutant); lanes 6–10, analysis of AtPCNA2 (WT and K164R mutant). The positions of molecular mass markers are indicated.
by guest on May 7, 2016
http://jxb.oxfordjournals.org/D
ownloaded from

864 | Strzalka et al.
Discussion
In eukaryotes, nuclear genetic material is replicated by DNA Polδ/ε (Burgers, 2008). However, when damage occurs in the template strand, replicative polymerases, which synthesize DNA with a high degree of accuracy, are blocked by lesions and cannot continue the synthesis of new DNA (Prakash et al., 2005). Studies on yeast have shown that eukaryotic cells can utilize the DNA PRR pathway to complete the rep-lication process. Two branches of PRR are known (Prakash et al., 2005; Strzalka and Ziemienowicz, 2011). The first branch involves monoubiquitinated PCNA and TLS poly-merases which are able to synthesize DNA across lesions. Studies of Arabidopsis TLS polymerase mutants have sug-gested that in plant Polζ, Rev1 are involved in error-prone and Polη in error-free TLS repair of UV-induced DNA dam-age (Nakagawa et al., 2011). The second PRR branch, called error-free, involves PCNA polyubiquitination mediated by RAD5. Its molecular mechanism is not clearly under-stood (Strzalka and Ziemienowicz, 2011). The Arabidopsis RAD5a protein is suggested to be a functional homologue
of the yeast RAD5 (Chen et al., 2008, Wang et al., 2011). ScRAD5 protein contains an SWI/SNF family domain, a conserved ATPase sequence (Johnson et al., 1992), and a RING finger motif required to interact with ScUBC13 (Ulrich, 2003). Moreover, ScRAD5 possesses ATPase (Gangavarapu et al., 2006), helicase (Blastyak et al., 2007), and E3 ligase activity (Parker and Ulrich, 2009). Analysis of rad5a mutant plants has shown their hypersensitivity to DNA-damaging agents such as mitomycin C, cis-platin and methyl methane sulphonate (Chen et al., 2008). This mutant also demonstrated deficiency in homologous recombination after treatment with a double-strand break-inducing agent (Chen et al., 2008).
The nature of AtPCNA – ubiquitin interaction
This study successfully demonstrated an interaction between AtPCNA and ubiquitin in the plant cell (Fig. 1). The observed interaction between AtPCNA and AtUBQ in the nucleus is in agreement with other in vitro studies which have shown that PCNA ubiquitination is dependent on the loading of
Fig. 3. Identification of AtRAD5a as an AtPCNA2-interacting protein by mass spectrometry analysis. (A) AtPCNA2-interacting proteins extracted from an Arabidopsis T87 suspension cell culture were isolated using an AtPCNA2 affinity column. The isolated proteins were separated in 12% polyacrylamide gel, stained in Coomassie brilliant blue and the gel fragment containing proteins was divided into 24 sectors. The positions of molecular mass markers are indicated. (B) Sequences of the AtRAD5a peptides identified in the gel sectors number 2 and 3 by mass spectrometry analysis.
by guest on May 7, 2016
http://jxb.oxfordjournals.org/D
ownloaded from

Arabidopsis PCNA ubiquitination | 865
Fig. 4. Analysis of AtRAD5a and AtUEV1A impact on AtPCNA ubiquitination in the presence of AtUBC2 (A), AtUBC30 (B), and AtUBC35 (C). An Arabidopsis ubiquitination system reconstituted in bacteria was used. The presented combinations of proteins were overexpressed together in E. coli BL21(DE3) cells. Next 40 µg total bacterial protein extract were separated in 12% polyacrylamide gel, subjected to Western blotting, and tested using the anti-c-myc antibody. Lanes 1–10, analysis of AtPCNA1 (WT and K164R mutant); lanes 11–20, analysis of AtPCNA2 (WT and K164R mutant). The positions of molecular mass markers are indicated.
by guest on May 7, 2016
http://jxb.oxfordjournals.org/D
ownloaded from

866 | Strzalka et al.
the PCNA homotrimer onto DNA by replication factor C (Haracska et al., 2006). However the interaction between AtPCNA and AtUBQ observed in the cytoplasm was unex-pected. Testing the total protein extract prepared from the N. benthamiana leaf cells overexpressing AtPCNA and AtUBQ by Western blotting failed to detect the covalently ubiquitinated AtPCNA (data not shown). This result sug-gested that this study observed the non-covalent interaction between ubiquitin and PCNA. Examples of proteins which can non-covalently interact with ubiquitin have been already reported (Hicke et al., 2005). The unsuccessful detection of covalently ubiquitinated PCNA in the plant cells may result from the fact that most of the leaf cells of N. benthamiana used for BiFC analysis were non-dividing cells. The present in vivo studies of AtRAD5a and AtPCNA1 interaction showed that these two proteins interact exclusively in the nucleus. This result supports the in vitro and in bacteria studies sug-gesting the involvement of AtRAD5a in the ubiquitination of AtPCNA in the plant cell.
In bacteria analysis of plant PCNA ubiquitination pathways
In yeast cells, two types of PCNA ubiquitination (monoubiq-uitination and polyubiquitination), which are dependent on different E2/E3 complexes, have been reported (Hoege et al.,
2002). In A. thaliana, 41 genes are predicted to encode func-tional proteins with a ubiquitin-conjugating domain and a cysteine active site, which includes four that are shown or predicted to function with ubiquitin-like proteins (Kraft et al., 2005). This study focused on three different E2 pro-teins (AtUBC2, AtUBC30, and AtUBC35) that correspond to yeast homologues involved in ScPCNA ubiquitination. The data from the present in bacteria studies have undoubt-edly shown that, among the tested E2, only AtUBC2 was able to directly monoubiquitinate AtPCNA. These observa-tions suggest that AtUBC2-mediated AtPCNA ubiquitina-tion (E3-independent) may play a relevant role in plant DNA repair signalling. Other data supporting this hypothesis were presented for the yeast rad6 mutant in which the UV sensi-tivity phenotype could be complemented by AtUBC2 (Zwirn et al., 1997).
In contrast to a relatively small number of proteins with predicted UBC domains, the Arabidopsis genome has more than 1400 genes that are predicted to encode for E3 enzymes (Vierstra, 2009). This study tested the role of two different ubiquitin ligases, AtCOP1 and AtRAD5a in AtPCNA ubiqui-tination. In contrast to AtCOP1 (data not shown), AtRAD5a played an important role in plant PCNA ubiquitination (Fig. 4B, C). In normal yeast cells, PCNA can be monoubiq-uitinated by the RAD6/RAD18 complex (Hoege et al., 2002). Moreover, recent studies on the yeast cdc9 mutant (deficient in DNA ligase I) have shown that ScPCNA can be monoubiq-uitinated by ScUBC4/ScMMS2/ScRAD5 at lysine 107 (Das-Bradoo et al., 2010). The present data are partly in agreement with the widely accepted model describing ubiquitination of yeast PCNA. First, this study found that AtRAD5a might be involved in AtPCNA monoubiquitination dependent on AtUBC2. Second, this study did not find AtUEV1A neces-sary for AtUBC30/AtRAD5a-dependent AtPCNA mon-oubiquitination (Fig. 4B).
The analysis of AtPCNA ubiquitination using either AtUBC30/AtRAD5a or AtUBC35/AtRAD5a gave simi-lar results. This raised the question whether AtUBC30 and AtUBC35 might have redundant functions in the plant cell. The introduction of an additional protein (AtUEV1A) into the experimental system has clearly shown the different properties of AtUBC30 and AtUBC35 (Fig. 4B, C). Only the AtUBC35/AtUEV1A/AtRAD5a complex showed simi-lar activity as yeast UBC13/MMS2/RAD5 responsible for PCNA polyubiquitination (Hoege et al., 2002). AtUEV1A is one of the four homologues (AtUEV1A, B, C and D) of the yeast MMS2 protein in the Arabidopsis genome (Wen et al., 2008). Yeast MMS2 and Arabidopsis UEV1A and D were shown to promote the assembly of a lysine-63-linked polyubiquitin chain (Hoffman and Pickart, 1999; Wen et al., 2008). Moreover, it was demonstrated that UEV1D is involved in the DNA damage response (Wen et al., 2008). The present experimental results are in agreement with previous reports. AtUEV1A together with AtUBC35/AtRAD5a was able to form a polyubiquitin chain on AtPCNA (Fig. 4C). Moreover, this study also confirmed the linkage of ubiquitin units through the conserved lysine 63 as described for yeast PCNA (Hoege et al., 2002, Fig. 5).
Fig. 5. Identification of the AtUBQ lysine 63 involved in the formation of a polyubiquitin chain on AtPCNA. An Arabidopsis ubiquitination system reconstituted in bacteria was used. The presented combinations of proteins were overexpressed together in E. coli BL21 (DE3) cells. Next, 40 µg total bacterial protein extract were separated in 12% polyacrylamide gel, subjected to Western blotting, and tested using the anti-c-myc antibody. Lanes 1–3, analysis of AtPCNA1 (WT); lanes 4–6, analysis of AtPCNA2 (WT). The positions of molecular mass markers are indicated. UBQ (GG) K63R represents the ubiquitin mutant with lysine 63 changed to arginine.
by guest on May 7, 2016
http://jxb.oxfordjournals.org/D
ownloaded from

Arabidopsis PCNA ubiquitination | 867
The results of this analysis are coherent with data presented previously for rad5a and rev3 mutants. The results of rad5a mutant plant analysis suggested that the RAD5-dependent error-free branch of PRR might be conserved between yeast and plants (Chen et al., 2008). The quantitative analysis of rad5a and rev3 mutant plants indicated the synergistic effect of these mutations (Wang et al., 2011). Based on the present in bacteria results, it is proposed that rad5a mutant plants might still have active AtUBC2-dependent PRR which may involve the monoubiquitinated PCNA-dependent TLS branch. Moreover, the data demonstrate that, in Arabidopsis, RAD5a might be involved in two branches of PPR, one dependent on monoubiquitinated (TLS-dependent) and the second on poly-ubiquitinated (TLS-independent) PCNA. This would be in agreement with other studies where the Saccharomyces cerevi-siae UBC13/MMS2 complex and RAD5 were each suggested to independently stimulate the TLS-dependent PRR branch in PCNA sumoylation-deficient yeast cells (Halas et al., 2011).
In this study a new tool for the analysis of AtPCNA ubiq-uitination pathways involved in plant PPR was constructed. Using this system it has been shown that AtPCNA is post-translationally ubiquitinated at the conserved lysine 164. The results suggest that, in plants, ubiquitination of PCNA may occur either in an AtRAD5a-dependent or -independent man-ner. Moreover, this study has demonstrated that plant PCNA polyubiquitination requires AtUEV1. The results, together with data published by others, suggest a similar function of ubiquitinated plant PCNA as proposed for yeast and mam-malian cells. The presented results did not demonstrate any difference between ubiquitination patterns of AtPCNA1/2 suggesting the equivalent role of these two proteins in PRR. Based on these results, a putative model of a plant PCNA ubiquitination scenario is proposed (Fig. 6).
Supplementary material
Supplementary data are available at JXB online.Supplementary Fig. S1. Control experiment for AtRAD5a
role in polyubiquitination of AtPCNA.Supplementary Fig. S2. Analysis of AtRAD5a impact on
AtPCNA ubiquitination.Supplementary Table S1. List of the vectors and primers
used in the study.
Supplementary Table S2. Analysis of ubiquitination-over-expression scheme.
Acknowledgements
This project was supported by the Polish Ministry of Science and Higher Education (project no. N N301 474438). W.S., F.B., K.P., and A.J. were supported by the Polish National Centre for Research and Development (project no. LIDER/28/54/L-/10/NCBiR/2011). WS was supported by the Polish Ministry of Science and Higher Education (project Iuventus Plus no. IP2011 052571). The Faculty of Biochemistry, Biophysics and Biotechnology of Jagiellonian University is a beneficiary of structural funds from the European Union (grant no. POIG.02.01.00-12-064/08).
References
Arakawa H, Moldovan GL, Saribasak H, Saribasak NN, Jentsch S, Buerstedde JM. 2006. A role for PCNA ubiquitination in immunoglobulin hypermutation. PLoS Biology 4, e366.
Bergink S, Jentch S. 2009. Principles of ubiquitin and SUMO modifications in DNA repair. Nature 458, 461–467.
Blastyak A, Pinter L, Unk I, Prakash L, Prakash S, Haracska L. 2007. Yeast Rad5 protein required for postreplication repair has a DNA helicase activity specific for replication fork regression. Molecular Cell 28, 167–175.
Das-Bradoo S, Nguyen HD, Wood JL, Ricke RM, Haworth JC, AK Bielinsky. 2010. Defects in DNA ligase I trigger PCNA ubiquitination at lysine 107. Nature Cell Biology 12, 74–20.
Bravo R, Frank R, Blundell PA, Macdonald-Bravo H. 1987. Cyclin/PCNA is the auxiliary protein of DNA polymerase-delta. Nature 326, 515–517.
Burgers PM. 2009. Polymerase dynamics at the eukaryotic DNA replication fork. Journal of Biological Chemistry 284, 4041–4045.
Chen I, Mannuss A, Orel N, Heitzeberg F, Puchta H. 2008. A homolog of ScRAD5 is involved in DNA repair and homologous recombination in Arabidopsis. Plant Physiology 146, 1786–1796.
Frampton J, Irmisch A, Green CM, Neiss A, Trickey M, Ulrich HD, Furuya K, Watts FZ, Carr AM, Lehmann. 2006. Postreplication repair and PCNA modification in Schizosaccharomyces pombe. Molecular Biology of the Cell 17, 2976–2985.
Gangavarapu V, Haracska L, Unk I, Johnson RE, Prakash S, Prakash L. 2006. Mms2-Ubc13-dependent and-independent roles of Rad5 ubiquitin ligase in postreplication repair and translesion DNA synthesis in Saccharomyces cerevisiae. Molecular Cell Biology 26, 7783–7790.
Gulbis JM, Kelman Z, Hurwitz J, O’Donnell M, Kuriyan J. 1996. Structure of the C-terminal region of p21(WAF1/CIP1) complexed with human PCNA. Cell 87, 297–306.
Halas A, Podlaska A, Derkacz J, McIntyre J, Skoneczna A, Sledziewska-Gojska E. 2011. The roles of PCNA SUMOylation, Mms2-Ubc13 and RAD5 in translesion DNA synthesis in Saccharomyces cerevisiae. Molecular Microbiology 80, 786–797.
Fig. 6. Putative model of proliferating cell nuclear antigen ubiquitination pathways in plants.
by guest on May 7, 2016
http://jxb.oxfordjournals.org/D
ownloaded from

868 | Strzalka et al.
Haracska L, Unk I, Prakash L, Prakash S. 2006. Ubiquitylation of yeast proliferating cell nuclear antigen and its implications for translesion DNA synthesis. Proceedings of the National Academy of Sciences, USA 103, 6477–6482.
Hata S, Kouchi H, Tanaka Y, Minami E, Matsumoto T, Suzuka I, Hashimoto J. 1992. Identification of carrot cDNA clones encoding a second putative proliferating cell nuclear antigen, DNA polymerase delta auxiliary protein. European Journal of Biochemistry 203, 367–371.
Hershko A, Ciechanover A. 1998. The ubiquitin system. Annual Review of Biochemistry 67, 425–479.
Hicke L, Schubert HL, Hill CP. 2005. Ubiquitin binding domains. Nature Reviews 6, 610–621.
Hoege C, Pfander B, Moldovan GL, Pyrowolakis G, Jentsch S. 2002. RAD6-dependent DNA repair is linked to modification of PCNA by ubiquitin and SUMO. Nature 419, 135–141.
Hofmann RM, Pickart CM. 1999. Noncanonical MMS2-encoded ubiquitin-conjugating enzyme functions in assembly of novel polyubiquitin chains for DNA repair. Cell 96, 645–653.
Johnson RE, Henderson ST, Petes TD, Prakash S, Bankmann M, Prakash L. 1992. Saccharomyces cerevisiae RAD5-encoded DNA repair protein contains DNA helicase and zinc-binding sequence motifs and affects the stability of simple repetitive sequences in the genome. Molecular Cell Biology 12, 3807–3818.
Kerscher O, Felberbaum R, Hochstrasser M. 2006. Modification of proteins by ubiquitin and ubiquitin-like proteins. Annual Review of Cell and Developmental Biology 22, 159–158.
Kraft E, Stone SL, Ma L, Su N, Gao Y, Lau O, Deng X, Callis J. 2005. Genome analysis and functional characterization of the E2 and RING-type E3 ligase ubiquitination enzymes of Arabidopsis. Plant Physiology 139, 1597–1611.
Krishna TS, Kong XP, Gary S, Burgers PM, Kuriyan J. 1994. Crystal structure of the eukaryotic DNA polymerase processivity factor PCNA. Cell 79, 1233–1243.
Laquel P, Litvak S, Castroviejo M. 1993. Mammalian proliferating cell nuclear antigen stimulates the processivity of two wheat embryo DNA polymerases. Plant Physiology 102, 107–114.
Leach CA, Michael WM. 2005, Ubiquitin/SUMO modification of PCNA promotes replication fork progression in Xenopus laevis egg extracts. Journal of Cell Biology 171, 947–954.
Lee KY, Myung K. 2008. PCNA modifications for regulation of postreplication repair pathways. Molecules and Cells 26, 5–11.
Matsumoto T, Hata S, Suzuka I, Hashimoto J. 1994. Expression of functional proliferating cell nuclear antigen from rice (Oryza sativa) in Escherichia coli. Activity in association with human DNA polymerase delta. European Journal of Biochemistry 223, 179–187.
Nakagawa M, Takahashi S, Tanaka A, Narumi I, Sakamoto AN. 2011. Role of AtPolζ, AtRev1, and AtPolη in UV light-Induced mutagenesis in Arabidopsis. Plant Physiology 155, 414–420.
Ogawa Y, Dansako T, Yano K, Sakurai N, Suzuki H, Aoki K, Noji M, Saito K, Shibata D. 2008. Efficient and high-throughput vector construction and Agrobacterium-mediated transformation of Arabidopsis thaliana suspension-cultured cells for functional genomics. Plant Cell Physiology 49, 242–250.
Ohta S, Shiomi Y, Sugimoto K, Obuse C, Tsurimoto T. 2002. A proteomics approach to identify proliferating cell nuclear antigen (PCNA)-binding proteins in human cell lysates. Journal of Biological Chemistry 277, 40362–40367.
Parker JL, Ulrich HD. 2009. Mechanistic analysis of PCNA poly-ubiquitylation by the ubiquitin protein ligases Rad18 and Rad5. EMBO Journal 28, 3657–3666.
Prakash S, Johnson RE, Prakash L. 2005. Eukaryotic translesion synthesis DNA polymerases: specificity of structure and function. Annual Review of Biochemistry 74, 317–353.
Prelich G, Tan CK, Kostura M, Mathews MB, So AG, Downey KM, Stillman B. 1987. Functional identity of proliferating cell nuclear antigen and a DNA polymerase-delta auxiliary protein. Nature 326, 517–520.
Stoimenov I, Helleday T. 2009. PCNA on the crossroad of cancer. Biochemical Society Transaction 37, 605–613.
Strzalka W, Kaczmarek A, Naganowska B, Ziemienowicz A. 2010. Identification and functional analysis of PCNA1 and PCNA-like1 genes of Phaseolus coccineus. Journal of Experimental Botany 61, 873–888.
Strzalka W, Labecki P, Bartnicki F, Aggarwal C, Rapala-Kozik M, Tani C, Tanaka K, Gabrys H. 2012. Arabidopsis thaliana proliferating cell nuclear antigen has several potential sumoylation sites. Journal of Experimental Botany 63, 2971–2983.
Strzalka W, Oyama T, Tori K, Morikawa K. 2009. Crystal structure of the Arabidopsis thaliana proliferation cell nucear antigen 1 and 2 proteins complexed with the human p21 C-terminal segment. Protein Science 18, 1072–1080.
Strzalka W, Ziemienowicz A. 2007. Molecular cloning of Phaseolus vulgaris cDNA encoding proliferating cell nuclear antigen. Journal of Plant Physiology 164, 209–213.
Strzalka W, Ziemienowicz A. 2011. Proliferating cell nuclear antigen (PCNA): a key factor in DNA replication and cell cycle regulation. Annals of Botany 107, 127–1140.
Sun L, Chen ZJ. 2004. The novel functions of ubiquitination in signaling. Current Opinion in Cell Biology 16, 119–126.
Suzuka I, Hata S, Matsuoka M, Kosugi S, Hashimoto J. 1991. Highly conserved structure of proliferating cell nuclear antigen (DNA polymerase delta auxiliary protein) gene in plants. European Journal of Biochemistry 195, 571–575.
Tan CK, Castillo C, So AG, Downey KM. 1986. An auxiliary protein for DNA polymerase delta from fetal calf thymus. Journal of Biological Chemistry 261, 12310–12316.
Towbin H, Staehelin T, Gordon J. 1979. Electrophoretic transfer of proteins from polyacrylamide gels to nitrocellulose sheets: procedure and some applications. Proceedings of the National Academy of Sciences, USA 76, 4350–4354.
Ulrich HD. 2003. Protein-protein interactions within an E2-RING finger complex. Implications for ubiquitin-dependent DNA damage repair. Journal of Biological Chemistry 278, 7051–7058.
Wang S, Wen R, Shi X, Lambrecht A, Wang H, Xiao W. 2011. RAD5a and REV3 function in two alternative pathways of DNA-damage tolerance in Arabidopsis. DNA Repair 10, 620–628
Wen R, Torres-Acosta JA, Pastushok L, Lai X, Pelzer L, Wang H, Xiao W. 2008. Arabidopsis UEV1D promotes lysine-63–linked
by guest on May 7, 2016
http://jxb.oxfordjournals.org/D
ownloaded from

Arabidopsis PCNA ubiquitination | 869
polyubiquitination and is involved in DNA damage response. The Plant Cell 20, 213–227.
Vierstra RD. 2009. The ubiquitin–26S proteasome system at the nexus of plant biology. Nature Reviews Molecular Cell Biology 10, 385–397.
Vivona JB, Kelman Z. 2003. The diverse spectrum of sliding clamp interacting proteins. FEBS Letters 546, 167–172.
Yang Y, Li R, Qi M. 2000. In vivo analysis of plant promoters and transcription factors by agroinfiltration of tobacco leaves. The Plant Journal 22, 543–551.
Zwirn P, Stary S, Luschnig C, Bachmair A. 1997. Arabidopsis thaliana RAD6 homolog AtUBC2 complements UV sensitivity, but not N-end rule degradation deficiency, of Saccharomyces cerevisiae rad6 mutants. Current Genetics 32, 309–314.
by guest on May 7, 2016
http://jxb.oxfordjournals.org/D
ownloaded from