race and the odd history of human paleontology
TRANSCRIPT

ARTICLE
Race and the Odd History of Human PaleontologyJEFFREY H. SCHWARTZ*
Although the late 17th century witnessed the recognition of fossils as the remains of extinct organisms—becausethey could be incorporated into the creation story embodied in the Great Chain of Being—acceptance of humanantiquity through the indisputable demonstration of the contemporaneity of human bones, stone tools, and acceptedfossils was not forthcoming for nearly 2 centuries thereafter. When it did occur, however, ancient humans were notseen as presenting a pattern of diversity similar to that seen in the fossil records of nonhuman organisms. Instead,human evolution then, as now, has typically been interpreted as being unilinear. This belief can be traced to Huxley(1863), who argued that the Feldhofer Grotto Neanderthal skullcap was merely an extension into the past ofmorphology seen in the Australian Aborigine, whom he took to represent the primitive end of an extreme range ofvariation he thought characterized Homo sapiens. During the mid-20th century, Mayr and Dobzhansky (mis)used theirclout as founders of the evolutionary synthesis to cement in paleoanthropology the idea that human evolutionaryhistory was characterized by nonspeciation. As such, anything that could be interpreted as potentially representingtaxic diversity was relegated to the status of individual variation. Lack of understanding of the history of humanpaleontology, and the biases that constrained its perspective on human evolution, continue to affect the ways inwhich most paleoanthropologists pigeonhole human fossils. Anat Rec (Part B: New Anat) 289B:225–240, 2006.© 2006 Wiley-Liss, Inc.
KEY WORDS: Huxley; King; Darwin; Neanderthal; Engis; Mayr; Dobzhansky; taxic diversity; paleoanthropology; systematics
INTRODUCTIONTo most observers of the discoveryand interpretation of human fossils,the field of human paleontology—pa-leoanthropology—probably seemsexotic. And why not? What otherdiscipline can declare as its goal doc-
umenting the emergence of modernhumans from a trail of increasinglyhuman-like forms? Perhaps no otherarea of paleontology grabs the imagi-nation so completely, especially be-cause it deals with a basic question allhumans share: Where did I comefrom?
The practice of human fossil hunt-ing also captures our attention, andthat of the media, which plays on (per-haps more precisely, preys off) thepersonalities and aura of romanticismassociated with fieldwork. Sunburnedbut dedicated men and women whothrow caution to the wind as theyspend field season after grueling fieldseason coping with the parching sun,lack of water and food, dangerouspredators and disease, and even at-tack by bands of nomads. After yearsof crawling around on hands andknees, picking up every tiny fragmentof animal bone, stone, or root, theyeventually find “it”—the “it” often be-ing the specimen or specimens that itsdiscoverers then put forth as thatyear’s no longer “missing link” in thehuman evolutionary saga. And as of-
ten as not this now critical ancestor ofour own species is recognized as anew species if not also as a new genus.Even if “it” turns out to be only ahandful of teeth or a fragmentary jaw,the discovery is usually touted as thebasis for the most definitive declara-tions about the evolutionary relation-ships of this undoubted new species topreviously known human relatives (towhich, with humans, I will refer ashominids). The cachet that accompa-nies the successful discovery of a fos-sil hominid is more than worth itsweight in academic gold.
On a more serious note, however,the interpretation of a new hominidfossil is typically bounded by, and fil-tered through, a received wisdom thatis largely unspoken, and probably noteven understood, by the very paleoan-thropologists who make these pro-nouncements. Indeed, an increasingnumber of paleoanthropologists ap-pear to believe that only they have theright to publish on “their” specimens,and to decide whether or not theyshould be allocated to new taxa. Inaddition, especially over the last de-
A fellow of the World Academy of Artand Science, Dr. Schwartz is professorof physical anthropology and of historyand philosophy of science at the Univer-sity of Pittsburgh and a research asso-ciate at the American Museum of Natu-ral History. He recently published thefirst study of virtually the entire humanfossil record (The Human Fossil Recordwith I. Tattersall) and a revised edition ofThe Red Ape, which explores the as-sumptions underlying molecular andmorphological approaches to phyloge-netic reconstruction. Just published is arevised edition of his textbook on hu-man osteology, Skeleton Keys, is forth-coming.*Correspondence to: Jeffrey H.Schwartz, Departments of Anthropol-ogy and History and Philosophy of Sci-ence, University of Pittsburgh, Pitts-burgh, PA 15260. Fax: 412-648-7509;E-mail: [email protected]
DOI 10.1002/ar.b.20119Published online in Wiley InterScience(www.interscience.wiley.com).
THE ANATOMICAL RECORD (PART B: NEW ANAT.) 289B:225–240, 2006
© 2006 Wiley-Liss, Inc.

cade, the frequency with which notedpaleoanthropologists have kept theirprecious fossils off limits to other pa-leoanthropologists has increased at analarming rate. Ignoring the abuse ofthe rules of the International Code ofZoological Nomenclature through theclaim that newly named taxa havemerely been “announced” and notproperly described, such paleontolo-gists often justify denying access tospecimens by declaring that sincethey, the discoverers, had to endurethe hardships in the field that led tothe recovery of these fossils, they havethe right to determine access to them(Gibbons, 2002). Unfortunately, thismindset, well known for years as “dis-coverer’s bias,” is only as good as thediscoverer’s background in compara-tive morphology and, more impor-tantly, in systematic theory and prac-tice.
With regard to paleoanthropology,it is an odd and also tragic state ofaffairs that the rigor of systematicpractice typically prevailing in otherareas of comparative evolutionarymorphology is not now, and actuallyhas never been, central to the phylo-genetic and taxonomic interpretationof human fossils. Yet in almost re-verse correlation with this history,during the past decade there has beena marked increase in the number ofnewly proposed hominid taxa (generaand/or species, and even the elusivenotion of subspecies) and in a self-imposed pressure on the part of thediscoverers to situate their finds defin-itively in a schema of hominid evolu-tion (e.g., White et al., 1994; Leakey etal., 2001; Senut et al., 2001; Brunet,2002; Mallegni et al., 2003). I seriouslyquestion the claims about how thesespecimens (especially the recentlypublished very early specimens) arerelated to other hominids—if, indeed,they are even hominid (Schwartz,2004a, 2004b; Schwartz and Tatter-sall, 2005). Nevertheless, the rash ofdiscoveries, accompanied with themedia blitz that coincides with theirpublication in a prestigious journal,gives the general public and even mostphysical anthropologists the impres-sion that systematics is alive and wellin paleoanthropology. Unfortunately,once a specimen is named, its claimedtaxonomic status assumes a life of im-mortality, which, given the relative in-
accessibility of certain specimens,only the discoverers will later be ableto alter or reevaluate.
Is there, however, a necessary cor-relation between taxonomy, which isthe art of naming a new taxon, andsystematics, which involves trying todelineate groups of specimens on thebasis of shared derived features (i.e.,synapomorphies), and deciphering apattern of relationships between thesehypothesized groups? I think not(Schwartz, 2003). Nevertheless, thereis a tendency not only for the media-conscious general public, but also forthe professional consumers of taxon-omists’ pronouncements, to presumethat taxonomy and systematics aresynonymous. In turn, this leads to anunwarranted conflation of these his-torically, theoretically, and method-ologically distinct endeavors. Frommy perspective as a practicing system-atist and taxonomist, the results ofthis confusion are particularly easilyseen in paleoanthropology.
THE QUESTION OF HOMINIDTAXIC DIVERSITYThe general public, and even the in-terested but not necessarily well-in-formed academic community, are fre-quently led to believe that most of thedebates in paleoanthropology revolvearound whether hominids ever were ataxically diverse group. I do not meanto suggest that the question, “howmuch taxic diversity is there and hasthere been?” is not of general concernwith regard to metazoans (as opposedto bacteria, for instance, in which cel-lular lineages typically transformwithout diversification). Yet, unlikethe paleontological and systematicstudy of other organisms, the study ofhominids has developed a uniquelyodd tradition, which continues to beperpetuated. Namely, in contrast tothe typical paleontological experienceof discovering new taxa as new sitesare opened or as already-known sitescontinue to be excavated, it is not un-common to find paleoanthropologists(e.g., Wolpoff, 1996; Curnoe andThorne, 2003; White, 2003) arguingagainst the possibility that hominidscould have been as speciose in thepast as undoubtedly appears to have
been the case for other groups of or-ganisms, whether vertebrate or inver-tebrate.
Because of this bias against recog-nizing taxic diversity in the humanfossil record, the motivations underly-ing the presentation of a new hominidgenus and/or species are yet furtherdistinguished from the normal prac-tice of taxonomy and systematics.This is because, for many paleoan-thropologists, a newly designatedtaxon rarely embraces a biological en-tity that arose via branching specia-tion (i.e., cladogenesis). Were this tobe the case, the presentation of thisnew taxon would be in the form of atestable hypothesis. The typical paleo-anthropological notion of a “transi-tional form” is, however, untestable.In addition, a theory of relatednessintegrating this new hominid taxonwith other taxa that also emerged viacladogenesis would also be formu-lated as a testable hypothesis, at ahigher level within the monophyleticgroup or clade (i.e., a group of organ-isms descended from a common an-cestor). However, because paleoan-thropologists typically think in termsof nondiversity (i.e., nonbranchingspeciation), a new genus and/or spe-cies is often used as a place marker inwhat is seen from the beginning as anongoing process of unilinear (i.e.,anagenetic) transformation, which,by its very nature of continual trans-formation, precludes the recognitionof distinct taxa.
Such a perspective—in which unilin-earity and taxic nondiversity are heavilyemphasized—is reminiscent of a mind-set that was popular during the 1970samong a small number of vertebratepaleontologists. As Darwin (1859) haddone before them, these paleontologistsargued that the only reason one thinksone can delineate species as distinct en-tities is because of taphonomic effectson, and other inconvenient distortionsof, what had once been a seamlessrecord of organismal transformation(e.g., Gingerich, 1976). A complete fos-sil record would reveal a picture oftransforming lineages with fewer in-stances of branching.
Nevertheless, the emphasis onsearching for ever-elusive transitionalforms was steadily replaced by thetheoretically and methodologicallymore rigorous procedure of phyloge-
226 THE ANATOMICAL RECORD (PART B: NEW ANAT.) ARTICLE

netic reconstruction afforded bycladism. As any recent issue of thejournal Systematic Biology and theJournal of Vertebrate Paleontology willdemonstrate, the hypothetico-deduc-tive strengths of cladistic analysishave led to its becoming the method-ology of choice not only among pale-ontologists, but among systematists ingeneral. This is in large part becausecladism’s emphasis on extensivecross-taxic comparison and hypothe-sis testing precludes requiring a priorithat either diversity or nondiversitycharacterized the evolutionary historyof any group of organisms (Schwartz,2003). Curiously, paleoanthropology,while a discipline that must also con-front evolutionary history, and thattherefore should share the same theo-retical and methodological concernsas other systematic enterprises, doesnot (Tattersall, 1986; Tattersall andSchwartz, 2000).
Typically in paleoanthropology, taxaare defined not on the basis of unique-nesses or apomorphies that are delin-eated through a process of broad mor-phological comparison, but in thecontext of an a priori hypothesis of howone’s specimen(s) is related to previ-ously known hominid fossils. Increas-ingly, the determination of relatednessis through the temporal sequencing offossils (e.g., White, 2003; White et al.,2006). Thus, a newly described hominid“taxon” is likely to represent a phase ina transforming lineage [e.g., the middle-late Pleistocene specimens from Herto,Ethiopia, that were declared to repre-sent the immediate ancestor of livinghumans and placed in a new subspe-cies, H. sapiens indaltu (White et al.,2003)], or the antecedent of more thanone descendent [e.g., specimens fromthe c. 780 kya TD-6 level of Gran Do-lina, Atapuerca, that were claimed torepresent the ancestor of both H. nean-derthalensis and H. sapiens and conse-quently placed in the new species H.antecessor (Bermudez de Castro et al.,1997)].
White (2003: p. 1997) has criticizedother paleoanthropologists for prac-ticing “diversity systematics” which,he believes, derives from a zeal forrecognizing taxic diversity in the hu-man fossil record that overrides theassessment of diversity through “thecanons of modern biology.” As a resultof suggesting that many taxa, such as
Australopithecus anamensis (Leakeyet al., 1995) and Homo antecessor,were named prematurely and/or onthe basis of inadequate specimens(and may even represent previouslydiagnosed taxa), White was obliged toquestion the validity of two taxa hehad created, i.e., Ardipithecus ramidus(White et al., 1994, 1995) and the geo-logically younger Australopithecusgarhi (Asfaw et al., 1999). He also ar-gued that another cause of overesti-mating taxic diversity was “expandingmatrix distortion” (EMD). Because ofEMD, specimens become so reshapedthat they “fool” paleoanthropologistsinto thinking they represent new taxa.His primary example from the humanfossil record was the skull of Kenyan-thropus platyops (Leakey et al., 2001),which is, indeed, composed of a myr-iad fragments held together by matrix.
Although perhaps too dismissive ofthis specimen’s preservation of rele-vant morphology (many facial fea-tures are preserved bilaterally), Whiteultimately rests his case for rejectingclaims of hominid taxic diversity onan article by John Noble Wilford inthe 25 March 2001 issue of the NewYork Times. There, Wilford arguedthat because modern anthropologists“have been steeped in multicultural-ism and diversity, and see [newly dis-covered hominid fossils] in a favor-able light,” an appreciation of modernethnic diversity may have predisposedpaleoanthropologists to looking forand subsequently “discovering” diver-sity in the human fossil record. Inother words, as White apparently seesthe situation, the embracing of ethnicdiversity (“race”) by anthropologistsin general (and by cultural anthropol-ogists specifically) led to a strange ver-sion of politically correct paleoan-thropology, in which fossils that werereally only variants of the same spe-cies could be recognized as distinctspecies. In reaction to this purportedmotive, White strongly urges paleoan-thropologists to return to the taxo-nomic conformity (in reality, taxo-nomic minimalism) that the foundersof the evolutionary synthesis had im-posed decades earlier on the humanfossil record.
It is not my intention here to discusssystematic theory or how one mightdeal practically with the question oftaxic diversity, whether among homin-
ids or any other group of organisms. Ihave addressed many of these issueselsewhere (e.g., Schwartz, 1999a, 2003).Of relevance here is the cultural ele-ment of race by which White, via Wil-ford, accuses paleoanthropologists ofbeing unduly influenced in their at-tempts to decipher pattern in the hu-man fossil record. Also significant isWhite’s promotion of Mayr (1950)—one of the “fathers” of the evolutionarysynthesis—as an example of taxonomiclevel-headedness who tried to bring or-der to paleoanthropology, which Mayrcharacterized as a field of a “bewilder-ing diversity of names.”
Superficially, it might seem reason-able that White should invoke the au-thority of Mayr in resolving taxo-nomic issues in paleoanthropology.For, after all, Mayr trained as an orni-thologist and systematist and his ma-jor monographic contribution to theevolutionary synthesis, Systematicsand the Origin of Species (Mayr, 1942),did present his view of that subjectmatter. But in view of his acceptanceof taxic diversity among birds andother organisms, it is odd that Mayrargued so adamantly for whittlingdown the “bewildering diversity” ofpotential hominid taxa to only threespecies within a single genus, Homo(Mayr, 1950). Was the impetus purelyscientific, merely the neutral act ofone whose primary interest lay solelyin the science of evolutionary biology?Or did Mayr, as others before and af-ter him, fall victim to the mindset thatcomes from belonging to a species,Homo sapiens, that is so unique (espe-cially in being the only species of itskind alive today) that biological objec-tivity can easily be submerged by non-biological considerations?
The only way in which we can beginto understand why the study of humanfossils and the interpretation of humanevolution is and has been so differentfrom systematics and paleontology ingeneral is by understanding the dispar-ate histories of these disciplines.
FOSSILS AS REAL ENTITIES: THEEMERGENCE OFPALEONTOLOGY AND THERECOGNITION OF TAXICDIVERSITYThe fossilized remains of once-livingorganisms or the stone tools manufac-
ARTICLE THE ANATOMICAL RECORD (PART B: NEW ANAT.) 227

tured by earlier humans were not al-ways recognized for what they are. In1565, Konrad Gesner, creator of thegenus (Gesner, 1551–1558), classifiedobjects that came from the ground.He referred stone tools, which wereoften called “thunderbolts” or “thun-derstones” because of their presumedextraterrestrial origin, to “ceraunia,” aterm whose Greek and Latin etymo-logical underpinnings refer to thun-der. Some eight decades later, UlisseAldrovandi (1648) rejected Gesner’sceraunia and argued that like miner-als, natural stones and rocks, potter’sclay, and fossils, stone tools wereformed in the earth.
Aldrovandi’s and other 17th-centuryGreat Chain of Being scholars’ beliefthat all earth-borne objects had a sim-ilar origin is at odds with Michel Mer-cati’s widely circulated manuscript of1574 (published only in 1717), inwhich he argued, by analogy withstone implements that explorers saw“primitive” non-Westerners using,that these unnatural-looking stone ob-jects were actually human-made stonetools (Grayson, 1983).
The demise of the practice of lump-ing together all objects that camefrom the earth began in 1669, with thepublication of Steno’s studies on thereality of fossils (Winter, 1968). Com-bining chemistry with the nascentprinciples of geology and micropal-eontology, Steno demonstrated notonly that the crystalline structure ofearth-derived minerals was totally un-like that of the shells of fossilized in-vertebrates, but also that the crystal-line structures of the shells offossilized and extant invertebrateswere similar. Thus, rock-hard objectswith shapes reminiscent of biologicalentities—such as clamshells, shark’steeth, and mammal bones—were trulythe remains of once-living organisms.In turn, this understanding permittedconsidering life in a broader anddeeper historical context—at leastnonhuman life, and within the timeconstraints of the creation myth.
Once the notion that all earth-de-rived objects had a common originbegan to crumble, the reality of stone-tool production by earlier generationsof humans could also be embraced,although not without certain conse-quences. For Mercati’s argument,based as it was on analogy with the
stone implements of “primitive” soci-eties, led to the “scholarly” construc-tion of a technological hierarchy thatproceeded from the lowliest of stoneimplements to the metallurgic sophis-tication of western Europe (Schwartz,1999b). In turn, this bias made it pos-sible for natural historians such asWoodward (1728), to argue that,whenever one found stone tools, onewas dealing with savage and barbarichumans, whether they were of the un-civilized past and had no choice inavailable tool-making material, or in-habitants of the present who, for what-ever reason, had failed to advancetechnologically to metallurgy as “true”civilized human societies had done.
Steno’s work had a truly significantimpact on biological science. The re-ality of fossils meant that the searchfor “missing links” to fill in the gaps ofthe Great Chain of Being could be ex-panded into the past (Blumenbach,1969; Schwartz, 1999b). But this didnot apply to humans. Humans, beingthe last to be created in accordancewith the creation myth and the Bookof Genesis, could not be (appreciably)antediluvian (Grayson, 1983). Never-theless, the availability of hordes ofpreviously unacknowledged extinctlife forms opened up new avenues ofstudy. In particular, the nascent fieldsof vertebrate and invertebrate paleon-tology allowed the Great Chain of Be-ing-motivated taxonomist to revel inthe machinations of a divine creator.As the diversity of extant life had beenrecognized by earlier taxonomists, so,too, could diversity of past life now bedocumented (Schwartz, 1999b). Andbecause of this, the acts of describing,cataloguing, and organizing fossils ina systematic manner—albeit not froman evolutionary perspective but ac-cording to a classificatory representa-tion of the Great Chain of Being(Gregory, 1910; Lovejoy, 1942)—be-came the backbone of biology and sys-tematics, reaching an apogee by theearly 19th century, especially throughthe field, comparative, and interpre-tive work of Cuvier and other Euro-pean paleontologists (Gregory, 1910).
The legacy of such a long history ofallocating fossils to new or previouslynamed taxa and of systematically or-ganizing them is still with us today,e.g., in museums of natural historyand university departments that foster
paleontological research. As anyonevisiting these collections knows, fos-sils, like specimens of extant taxa, areextensively curated, with jaws, teeth,and other skeletal remains allocatedto genus and species and arranged indrawers, and drawers arranged withincabinets, according to a larger classi-ficatory scheme. This is not to say thatone might not find that the occasionalspecimen of living or extinct animalwas incorrectly identified to taxon orwas not recognized as a different ge-nus and/or species (Schwartz, 1995,1996). But, when one does use collec-tions to pursue a systematic study of agroup of animals, it is usually the casethat at least most of the initial identi-fication of genus and species (i.e., al-pha taxonomy) has already been quiteefficiently done in accordance with aprocedure that began centuries ago.
HUMANS ARE SOMEHOWDIFFERENTSuch regularized taxonomic and sys-tematic practice has not, however,carried over completely to human fos-sils (Schwartz and Tattersall, 2002b,2003, 2005). During my nearly de-cade-long study with I. Tattersall ofvirtually the entire human fossilrecord, and in contrast to my experi-ence with collections of nonhumanfossil primates and other mammals, Ihave found that the level of attentionto morphological detail in the cura-tion of human fossils has often beenminimal. In some cases, as with mid-dle to late Pleistocene specimens,fossils are often identified by site orprovenance (e.g., the Steinheim, Pe-tralona, or Kabwe skulls), by an infor-mal name corresponding to a per-ceived time period of that hominid’sexistence (e.g., the Krapina, La-Chapelle-aux-Saints, or Spy Neander-thals), or by a formal species namethat is associated with a time periodin which that taxon is supposed tohave existed (e.g., Homo erectus, toencompass all hominid fossils from c.1.8- to 0.5-million-year-old deposits ofthe Old World). The situation withearlier, Plio-Pleistocene, presumedhominid fossils is more tenuous and isfurther complicated by the fact thatthere has been no useful definition ofwhat “hominid” means (Schwartz,2001, 2007). Thus, it is not uncom-
228 THE ANATOMICAL RECORD (PART B: NEW ANAT.) ARTICLE

mon to find boxes of isolated teethwhose catalogue numbers identifythem simply as “hominid” or referthem all to a specific hominid taxon,such as “Australopithecus africanus.”Often, however, even cursory inspec-tion reveals that many or even none ofthese specimens can unequivocally beidentified as hominid (Grine andFranzen, 1994; Schwartz et al., 1995;Schwartz and Tattersall, 2005). Howcould the approach to human fossilsbe so dramatically superficial com-pared to the way in which other ex-tinct taxa are so routinely scrutinizedsystematically? Interestingly, one canactually point to two specific publica-tions that had this effect on paleoan-thropology.
In 1965, Simons and Pilbeam pub-lished a major taxonomic revision ofthe then-known Miocene hominoid(! the group consisting of hominidsand apes) fossils in which they notonly sank or synonymized a consider-able number of species and genera,but also further constrained hominoiddiversity by taking the three generathat survived their taxonomic lump-ing—the Indo-Pakistani Sivapithecus,the East African Proconsul, and theEuropean Dryopithecus—and demot-ing each to a subgenus of Dryopithe-cus. As they (also Pilbeam, 1970) sawit, Dryopithecus (Sivapithecus) was theancestor of the living Asian orangutanas well as of the large and extinct Gi-gantopithecus from the Plio-Pleisto-cene of Indo-Pakistan and southernChina; large and small species ofDryopithecus (Proconsul) gave rise, re-spectively, to the gorilla and chimpan-zee; and Dryopithecus (Dryopithecus)went extinct without issue. But whilethey collapsed most Miocene taxa intoone genus, Simons and Pilbeam alsorecognized Ramapithecus, which atthe time and for at least 15 yearsthereafter was interpreted as the an-cestor of “proper” hominids becauseof similarities in premolar and molarmorphology, thickness of molarenamel, and also, for a while, palatalshape (Simons, 1964, 1965).
As drastic as Simons and Pilbeam’srevision of Miocene hominoids was inreducing the number of accepted taxa,it had been preceded by any evenmore drastic revision. For Ernst Mayrhad already truncated the picture of
human evolution, first to three andthen to four time-successive species ofhominid.
It is clear from Mayr’s (1950) initialappraisal of paleoanthropology thathe was displeased with what he con-sidered the paleoanthropologists’ lackof standards when naming newly dis-covered human fossils. In large part,his disapproval is still warranted.From the beginning, paleoanthropol-ogy was essentially devoid of the pro-cedural rigor that characterized non-hominid paleontology, particularlywith regard to adhering to the rules ofthe International Code of Zoolog-ical Nomenclature (Tattersall andSchwartz, 2003).
But, aside from the fact that itspractitioners were schooled primarilyin biomedicine rather than compara-tive morphology and systematics,there are other reasons why paleoan-thropology differs so markedly inpractice from its sister disciplines.Much of this difference derives firstfrom the preevolutionary theory his-tory of human paleontology, and thenfrom Huxley’s (1863) assessment ofthe then-known human fossils. But wemust also return to Mayr. For, despitehis complaints about paleoanthro-pologists’ shoddy practice of system-atics, Mayr himself could not havedone more to deter scientificallyhealthy, alternative thinking in paleo-anthropology.
HUXLEY’S LEGACY, PART I: THEACCEPTANCE OF HUMANANTIQUITYIn 1863, Thomas Henry Huxley pub-lished Man’s Place in Nature, whichcontained three essays: “On the Natu-ral History of the Man-like Apes” (abrief review of historical referencesto, and conceptions about, the apes);“On the Relations of Man to the LowerAnimals” (the first developmental andcomparative morphological argumentfor why humans should be classifiednot only as primates, but in a sub-group with the apes); and “On SomeFossil Remains of Man” (a discussionof the Feldhofer Grotto Neanderthalskullcap and other real or presumedfossil crania).
With regard to the second essay,students of human evolution continueto claim incorrectly that Huxley dem-
onstrated a close evolutionary rela-tionship between humans and the Af-rican apes (e.g., Satta et al., 2000).Yet, as Huxley (1896) stated in thepreface to the second edition of thisvolume, and as is clear in the originalessay, he did not intend to provide anevolutionary argument for the related-ness of humans and any ape(Schwartz, 1987). Rather, as seen inthe quote below, Huxley’s conclusionwas taxonomic in nature, and consis-tent with his stated goal of validatingLinnaeus’ classifying humans withother animals:
“The structural differences betweenMan and the Man-like apes certainlyjustify our regarding him as constitut-ing a family apart from them; though,inasmuch as he differs less from themthan they do from other families ofthe same order, there can be no justi-fication for placing him in a distinctorder” (Huxley, 1863: p. 125).
J. Marks (personal communication)has suggested that we should read thispassage in light of Darwin’s (1859)view of the Linnaean taxonomic hier-archy as reflecting evolutionary pro-cesses that yield lines of descent. If wewere to do so, however, we would notconclude that humans are closely re-lated to the African apes. Rather, wewould interpret Huxley’s passage asindicating that humans are related toa group consisting of all large-bodied(“Man-like”) apes. Nevertheless, it be-came commonplace in the 20th cen-tury for paleoanthropologists to citethis essay as the first demonstration ofa close evolutionary relationship be-tween human and African apes(Schwartz, 2005), and this misconcep-tion continues to the present.
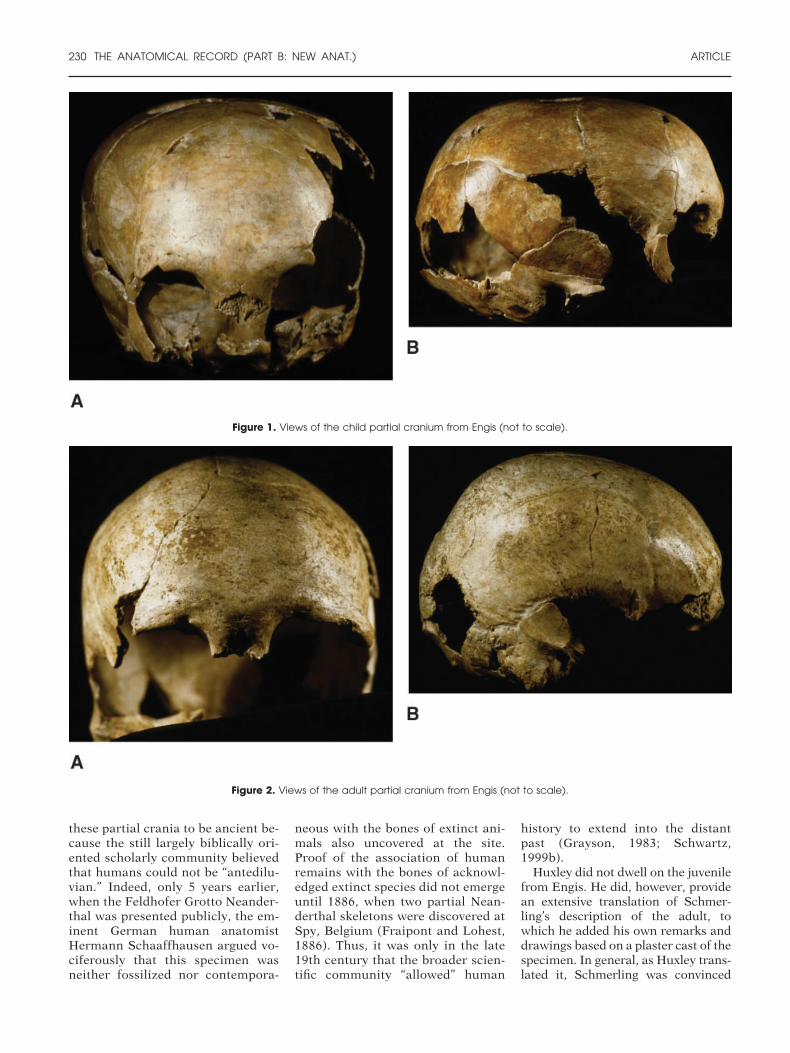
In the essay “On Some Fossil Re-mains of Man,” Huxley focused ontwo partial adult crania and a some-what disarticulated juvenile skull, allof which he embraced as being ofsome antiquity because of Lyell’s as-sessment of the geology and fauna ofthe sites from which they came. Sch-merling had excavated the juvenileand one adult specimen in the 1830sat Engis, near Liege, Belgium (Figs.1 and 2). The other adult craniumhad been unearthed during lime-stone quarrying at Feldhofer Grotto,near Dusseldorf, Germany, in theNeander Valley (Neander Thal). It issignificant that Huxley considered
ARTICLE THE ANATOMICAL RECORD (PART B: NEW ANAT.) 229
these partial crania to be ancient be-cause the still largely biblically ori-ented scholarly community believedthat humans could not be “antedilu-vian.” Indeed, only 5 years earlier,when the Feldhofer Grotto Neander-thal was presented publicly, the em-inent German human anatomistHermann Schaaffhausen argued vo-ciferously that this specimen wasneither fossilized nor contempora-
neous with the bones of extinct ani-mals also uncovered at the site.Proof of the association of humanremains with the bones of acknowl-edged extinct species did not emergeuntil 1886, when two partial Nean-derthal skeletons were discovered atSpy, Belgium (Fraipont and Lohest,1886). Thus, it was only in the late19th century that the broader scien-tific community “allowed” human
history to extend into the distantpast (Grayson, 1983; Schwartz,1999b).
Huxley did not dwell on the juvenilefrom Engis. He did, however, providean extensive translation of Schmer-ling’s description of the adult, towhich he added his own remarks anddrawings based on a plaster cast of thespecimen. In general, as Huxley trans-lated it, Schmerling was convinced
Figure 1. Views of the child partial cranium from Engis (not to scale).
Figure 2. Views of the adult partial cranium from Engis (not to scale).
230 THE ANATOMICAL RECORD (PART B: NEW ANAT.) ARTICLE
that these as well as human-like bonesfrom Engihoul, a site across the riverMeuse from Engis, were fossilizedand contemporaneous with the nu-merous bones of extinct animals alsoexcavated. In particular, Huxley(1863: p. 145) focused on Schmer-ling’s finding that “a broken fragmentof an ulna was soldered to a like frag-ment of a radius by stalagmite, a con-dition frequently observed among thebones of the [extinct] Cave Bear (Ur-sus spelaeus), found in the Belgiancaverns,” and also that “ProfessorSchmerling found, incrusted with sta-lagmite and joined to a stone, thepointed bone implement . . . andworked flints.”
Although, according to Huxley,Schmerling remarked that even if theface of the Engis adult had been pre-served it “would not have been possi-ble to pronounce, with certainty, upona single specimen” (Huxley, 1863: p.149), he proceeded to state that “theelongated and narrow form of theforehead,” together with “the slight el-evation of the frontal, its narrowness,and the form of the orbit, approxi-mate it more nearly to the cranium ofan Ethiopian than to that of an Euro-pean” (Huxley, 1863: p. 150). Giventhe apparent association of the bonesof a presumed primitive human(which “approximate” an Ethiopian’s)not only with the bones of acknowl-edged extinct animals, but also withbone and stone tools (hallmarks of“savages”), it is not surprising thatSchmerling then declared:
“At whatever conclusion we may ar-rive as to the origin of the man fromwhence this fossil skull proceeded, wemay express an opinion without ex-posing ourselves to a fruitless contro-versy. Each may adopt the hypothesiswhich seems to him most probable:for my own part, I hold it to be dem-onstrated that this cranium has be-longed to a person of limited intellec-tual faculties, and we conclude thencethat it belonged to a man of a lowdegree of civilization: a deductionwhich is borne out by contrasting thecapacity of the frontal with that of theoccipital region” (Huxley, 1863: p.150).
With regard to the overall morphol-ogy of the skull, I agree with Huxley’sdescription: it is relatively long, butnot very narrow (especially across thefront), with a high-domed frontal,well-arced sagittal profile, and littlemuscle scarring (Schwartz and Tat-tersall, 2002b). Of particular impor-tance for discussion here is that theonly region of the skull for whichHuxley (1863: p. 148–149) providesmuch morphological detail is the su-praorbital region:
“The supraciliary prominences orbrow-ridges . . . are well, but not ex-cessively, developed, and are sepa-rated by a median depression. Theirprincipal elevation is disposed so ob-liquely that I judge them to be due tolarge frontal sinuses.”
The illustration of the supraorbitalregion correctly portrays the “princi-pal” elevation as moderately swollen
bilaterally, with right and left portionsoriented somewhat superolaterally(Schwartz and Tattersall 1996, 2000,2002b, 2003). Although Huxley didnot describe other features of the su-praorbital region, they are faithfullyrepresented in the drawing. Specifi-cally, the bone lateral to each swellingis flatter and smoothly continuouswith the surrounding frontal bone.This particular configuration, whichis apparently unique to Homo sapiens,is identified in the paleoanthropologi-cal literature as a “bipartite brow”(Stringer et al., 1984; Schwartz andTattersall, 2002a). Given Huxley’sprowess as a comparative anatomist, Idoubt that these few morphologicaldetails escaped his notice inasmuch asthe supraorbital region assumes someimportance in his subsequent discus-sion.
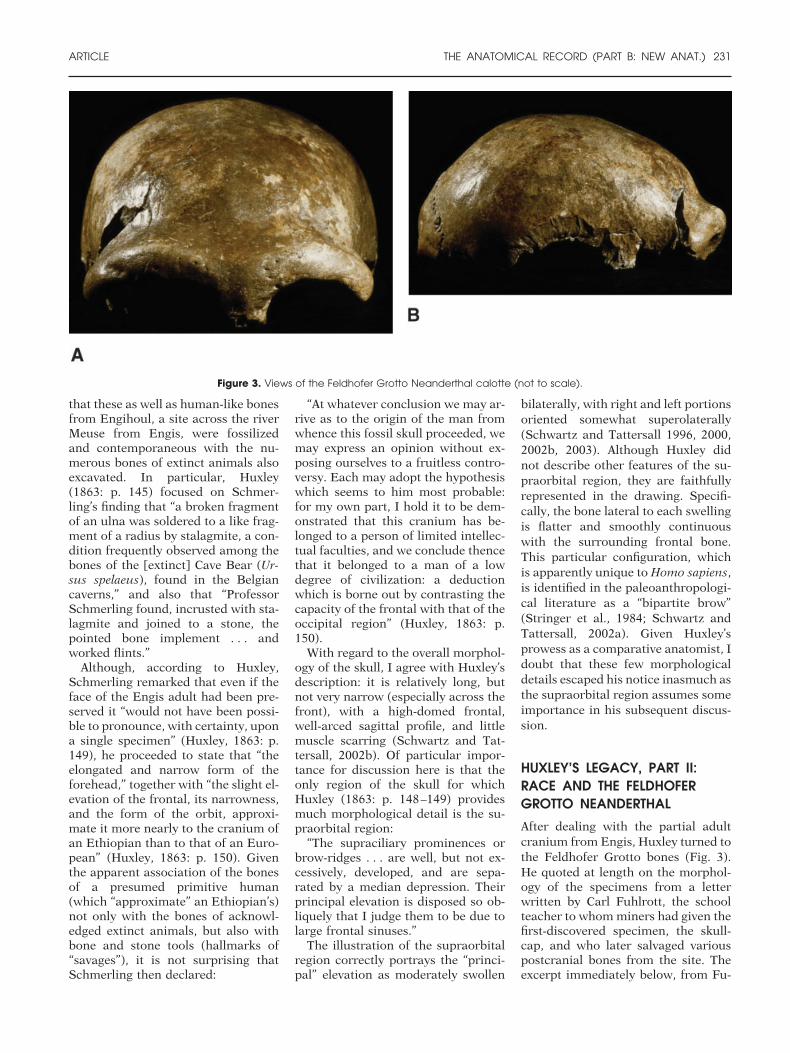
HUXLEY’S LEGACY, PART II:RACE AND THE FELDHOFERGROTTO NEANDERTHALAfter dealing with the partial adultcranium from Engis, Huxley turned tothe Feldhofer Grotto bones (Fig. 3).He quoted at length on the morphol-ogy of the specimens from a letterwritten by Carl Fuhlrott, the schoolteacher to whom miners had given thefirst-discovered specimen, the skull-cap, and who later salvaged variouspostcranial bones from the site. Theexcerpt immediately below, from Fu-
Figure 3. Views of the Feldhofer Grotto Neanderthal calotte (not to scale).
ARTICLE THE ANATOMICAL RECORD (PART B: NEW ANAT.) 231

hlrott, and others that follow are rele-vant for comparison with the adultEngis cranium:
“The cranium is of unusual size,and of a long-elliptical form. A mostremarkable peculiarity is at once ob-vious in the extraordinary develop-ment of the frontal sinuses, owing towhich the superciliary ridges, whichcoalesce completely in the middle, arerendered so prominent, that the fron-tal bone exhibits a considerable hol-low or depression above or rather be-hind them, whilst a deep depression isalso formed in the situation of theroot of the nose. The forehead is nar-row and low, though the middle andhinder portions of the cranial arch arewell developed. . . . [T]he superior oc-cipital ridges . . . are greatly devel-oped, and almost conjoined so as toform a horizontal eminence” (Huxley,1863: p. 151–152).
Huxley (1863: p. 152 et seq) thencites large sections from the anato-mist George Busk’s translation ofSchaaffhausen’s report, which wasprimarily a refutation of Fuhlrott’sclaim that the Feldhofer Grotto hu-man bones were fossilized and con-temporaneous with those of extinctnonhuman animals. After comparingthe Feldhofer Grotto skullcap withboth recent and what were thenconsidered ancient human crania,Schaaffhausen concluded:
“But the human bones and craniumfrom the [Feldhofer Grotto] Neander-thal exceed all the rest in those pecu-liarities of conformation which lead tothe conclusion of their belonging to abarbarous and savage race. Whetherthe cavern in which they were found,unaccompanied with any trace of hu-man art, were [sic] the place of theirinterment, or whether, like the bonesof extinct animals elsewhere, they hadbeen washed into it, they may still beregarded as the most ancient memo-rial of the early inhabitants of Eu-rope.”
Huxley (1863: p. 161–162) followsthe above quote with the comment:“Mr. Busk . . . has enabled us to forma very vivid conception of the de-graded character of the Neanderthalskull, by placing side by side with itsoutline, that of the skull of a Chim-panzee, drawn to the same absolutesize.” And, indeed, his discussion of
the Neanderthal skullcap is essentiallya justification of this impression of it.
Huxley tells his audience that hecame to study a plaster cast of theNeanderthal calotte for a second time,but then in detail, in order to provideLyell with a diagram that would illus-trate its oddities in comparison withother human skulls, which he repre-sented by the adult cranium from En-gis. For standardization, Huxleychose two cranial landmarks throughwhich he drew a line: i.e., glabella,which is the most anterior point onthe midline of the skull above the na-sal region, and inion, which is themost posterior point in the midline ofthe skull. He chose inion, which hesituated at the juncture of the occipi-tal protuberance and the superiorsemicircular curved line, for two rea-sons: it corresponds with the intracra-nial transverse sinuses and the line ofattachment of the tentorium, whichtethers the brain to the inside of thebraincase and also approximates thelower limits of the occipital lobes ofthe cerebrum (the “thinking” part ofthe brain).
Huxley aligned outlines of the Ne-anderthal and Engis partial craniaalong the glabella-inion axis. To hissurprise, he found that “the differencewas so vast and the flattening of theNeanderthal skull so prodigious . . .that I at first imagined I must havefallen into some error” (Huxley, 1863:p. 162). He asked: “Was it possible fora human being to have the brain thusflattened and depressed; or, on theother hand, had the muscular ridges[left by the nuchal muscles] shiftedtheir positions?” (Huxley, 1863: p.162). He then cited his correspon-dence with Fuhlrott, who had corrob-orated the configuration of the occip-ital region of the Neanderthal skullcapand had also sent photographs thatconfirmed Huxley’s (1863: p. 164) ear-lier finding (“the posterior lobe of thebrain of the Neanderthal man musthave been as much flattened as I sus-pected it to be”). Huxley (1863: p. 165)concluded this section of the essay onthe Neanderthal skullcap with thiscomment:
“[The Neanderthal skull cap] is par-ticularly interesting, as it suggeststhat, notwithstanding the flattenedcondition of the occiput, the posteriorcerebral lobes must have projected
considerably beyond the cerebellum,and as it constitutes one among sev-eral points of similarity between theNeanderthal cranium and certainAustralian skulls.”
The choice of “certain Australianskulls” for comment is not withouthistorical significance. For the GreatChain of Being-based hierarchical ar-rangement of life, which also rankedhumans from those closest to the“brutes” up to the most civilized, wasnot disavowed by 19th-century evolu-tionists. Among Great Chain of Beingtaxonomists and certain 19th-centuryevolutionists, contenders for the low-est of the humans included Hotten-tots, “bushmen,” and Australian Ab-origines, the latter of which wasHuxley’s choice as the most primitiveliving human (Schwartz, 1987).
But Huxley did not immediately of-fer an interpretation of the points ofsimilarity between the Neanderthalskullcap and Aboriginal skulls.Rather, he spent considerable timefirst emphasizing, as Blumenbach andothers had done before him, the enor-mous range of variability that a globalcomparison of human crania suppos-edly revealed. He then dismissed theways in which other scholars hadtried to equate human variation withracial distinctiveness, which, he be-lieved, failed to represent the funda-mental aspects that determine cranialshape because they were not “foundedupon the establishment of a relativelyfixed base line, to which the measure-ments, in all cases, must be referred”(Huxley, 1863: p. 171). By arguingthat skull growth in mammals fol-lowed a general pattern, he concluded“that the base of the skull may bedemonstrated developmentally to beits relatively fixed part, the roof andsides being relatively moveable . . .[which] . . . is exemplified by the studyof the modifications which the skullundergoes in ascending from thelower animals up to man” (Huxley,1863: p. 171). Essentially, Huxleyfound that from “lower” to “higher”crania, the basicranial axis becameshorter relative to skull length, the an-gle of the face to the neurocraniumbecame more acute, as the face be-came increasingly flexed downwardrelative to the braincase, and thebraincase became increasingly more
232 THE ANATOMICAL RECORD (PART B: NEW ANAT.) ARTICLE
vaulted or arched, with concomitantheightening and backward extensionof the cerebrum.
It is one thing to generalize from aseries of mammalian skulls that onelines up in a hypothetical series fromthe lowest (e.g., Huxley’s choice, thebeaver) to the highest (“man”). It is an-other, however, to continue this hierar-chical projection within the speciesHomo sapiens. Huxley (1863: p. 175)raises the question and answers it:
“Now comes the important ques-tion, can we discern, between the low-est and the highest forms of the hu-man cranium, anything answering, inhowever slight a degree, to this revo-lution of the side and roof bones of theskull upon the basicranial axis ob-served upon so great a scale in themammalian series? Numerous obser-vations lead me to believe that wemust answer this question in the affir-mative.”
Huxley’s “demonstration” is basedin part on comparing in one image thesagittal outlines of the skull of an Aus-tralian Aborigine and that of a Tartar
and, in another, the sagittal outlinesof the skull of a “Negro” with one froma cemetery in Constantinople. The Ab-origine and the “Negro” crania aremarkedly prognathic, with their ante-riorly projecting lower faces and up-per jaws, whereas the Tartar and es-pecially the other “modern-looking”skull are short-faced or orthognathic.Huxley likens these differences tothose between “lower animals” and“Man,” but admits that the prognathicindividuals are less ape-like becausetheir frontal cerebral lobes are situ-ated a bit more anteriorly than in theorthognathic individuals. He alsostates that a round skull (the skullfrom Constantinople) may exhibitgreater posterior projection than along one (the “Negro”). The accompa-nying drawing, however, representsthe “Negro” skull as the round oneand the other skull, which actuallydoes have a prominent posterior ex-tension, as the long one. Perhaps theartist confused the outlines of thesetwo skulls and the error escaped Hux-ley’s notice.
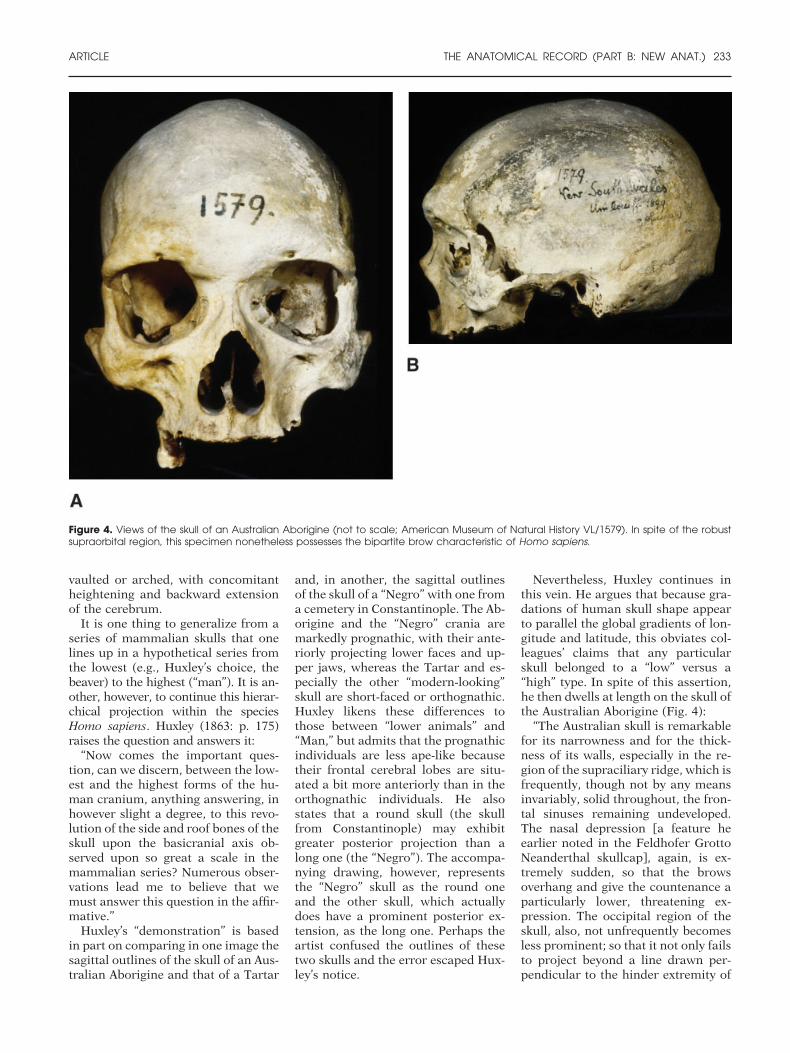
Nevertheless, Huxley continues inthis vein. He argues that because gra-dations of human skull shape appearto parallel the global gradients of lon-gitude and latitude, this obviates col-leagues’ claims that any particularskull belonged to a “low” versus a“high” type. In spite of this assertion,he then dwells at length on the skull ofthe Australian Aborigine (Fig. 4):
“The Australian skull is remarkablefor its narrowness and for the thick-ness of its walls, especially in the re-gion of the supraciliary ridge, which isfrequently, though not by any meansinvariably, solid throughout, the fron-tal sinuses remaining undeveloped.The nasal depression [a feature heearlier noted in the Feldhofer GrottoNeanderthal skullcap], again, is ex-tremely sudden, so that the browsoverhang and give the countenance aparticularly lower, threatening ex-pression. The occipital region of theskull, also, not unfrequently becomesless prominent; so that it not only failsto project beyond a line drawn per-pendicular to the hinder extremity of
Figure 4. Views of the skull of an Australian Aborigine (not to scale; American Museum of Natural History VL/1579). In spite of the robustsupraorbital region, this specimen nonetheless possesses the bipartite brow characteristic of Homo sapiens.
ARTICLE THE ANATOMICAL RECORD (PART B: NEW ANAT.) 233

the glabello-occipital line, but even, insome cases, begins to shelve awayfrom it, forwards, almost immedi-ately. In consequence of this circum-stance the parts of the occipital bonewhich lie above and below the tuber-osity make a much more acute anglewith one another than is usual,whereby the hinder part of the base ofthe skull appears obliquely truncated.Many Australian skulls have a consid-erable height, quite equal to that ofthe average of any other race, butthere are others in which the cranialroof becomes remarkably depressed”(Huxley, 1863: p. 179).
Also on this page is a profile linedrawing of an Aborigine’s skull andmandible with the outline of the an-teroposteriorly much longer Feld-hofer Grotto skullcap superimposedon it (Fig. 5). The cranial roof of thelatter is much lower than in the extanthuman. The brow (described and il-lustrated earlier by Huxley as contin-uous along the orbits and confluentacross glabella, and not bipartite) ismore massive, superoinferiorly thick-ened, and somewhat anteriorly pro-trusive. And the occipital region (alsopreviously described and illustratedby Huxley as quite different fromother humans, including the Engisadult cranium) is distinctive, even inprofile. Yet in spite of the number andmagnitude of differences that Huxleyas a critical comparative nonhumanvertebrate anatomist would have em-braced as worthy of distinction at thespecies if not genus level, he pro-claims: “A small additional amount offlattening and lengthening, with a cor-responding increase of the supracili-ary ridges, would convert the Austra-lian brain case into a form identicalwith that of the aberrant fossil” (Hux-ley, 1863: p. 179–180). A few pageslater, after commenting on how theNeanderthal cranium is rather pithe-coid or ape-like in some ways and yet,in others, compares well with “primi-tive” variants of human crania, Hux-ley (1863: p. 183) turns his hypotheti-cal continuum of morphologicaltransformation the other way aroundand states:
“[I]ndeed, though truly the mostpithecoid of known human skulls, theNeanderthal cranium is by no meansso isolated as it appears to be at first,but forms, in reality, the extreme term
of a series leading gradually from it tothe highest and best developed of hu-man crania.”
Clearly this does not represent thekind of morphological transformationHuxley had earlier argued was thenorm when he used the basicranialline as the standard on which to com-pare mammalian skulls. In addition,there is nothing in Huxley’s hypothet-ical sequence of altered cranial shapethat would lead to different details ofcranial morphology, such as bipartitesupraciliary arches (all recent andnear-recent human crania, includingthe adult Engis skull) versus continu-ous and smoothly rounded supraor-bital tori that merge at glabella (Nean-derthal skullcap). Or to a sulcusposterior to these supraorbital tori(Neanderthal) rather than a frontalthat rises laterally directly from thesupraorbital margin (bipartite-browedhuman crania, including the adultEngis skull). Or to a short but verywide occiput with a relatively straightoccipital “torus” and a pitted suprai-niac depression above (Neanderthal)versus a tall and triangular occiputwith an inferiorly oriented externaloccipital protuberance (human cra-nia, including the adult Engis skull)(Schwartz and Tattersall, 1996,2000, 2002b). These are the kinds ofdetails, rather than the general na-ture of craniofacial hafting, thatHuxley, following 2 centuries duringwhich taxonomists and systematistshad worked toward perfecting theirdiscipline, should have embraced asimportant in delineating taxa. In hisclassifications of mammals, he did.In the case of humans, he did not.
Why did Huxley, one of England’spremier comparative anatomists,abandon generally accepted and forthe time fairly rigorous systematicprocedure when dealing with the Ne-anderthal specimen? His emphasis onthe gradual transformation of what hesaw “as the most pithecoid of humancrania yet discovered” (Huxley, 1863:p. 181) into a more recognizablyHomo sapiens-like skull (albeit that ofa “primitive” Australian Aborigine)becomes even more perplexing inlight of the fact that by inclination hewas a saltationist (Huxley, 1860, 1863,1876). And for saltationists, differ-ences between organisms result fromchanges in development that lead tothe abrupt appearance of morpholog-ical novelty and, thus, to the abruptappearance of species (Mivart, 1871).As such, even though Huxley addedthat the great brain size of the Nean-derthal would have reflected an orga-nization not representative of the ape-like features imprinted in theskullcap, one would have expectedhim to see in the distinctive, non-Homo sapiens-like features of its browand occiput the hallmarks of a speciesin its own right. But, to reiterate, hedid not.
Insight into Huxley’s abandonmentof saltationism in favor of gradualmorphological transformation—inwhich the Feldhofer Grotto Neander-thal is appended as the most primitivevariant of a continuum of changeleading up to the most advanced cra-nial form—comes from his wonder-ing whether these bones could “be re-garded as the remains of a humanbeing intermediate between Men andApes” (Huxley, 1863: p. 181–182). Hadthese fossilized bones been the re-mains of any other mammal, doubt-less Huxley would not have hesitatedto interpret them as representing aspecies “intermediate” between more“primitive” and more “advanced” spe-cies. But in spite of admitting thatforms intermediate between humansand the ultimate “pithecoid” ancestorhad existed, he is unwilling to fill thisevolutionary void with the FeldhoferGrotto Neanderthal. He concludes hisessay:
“I may say, that the fossil remains ofMan hitherto discovered do not seemto me to take us appreciably nearer tothat lower pithecoid form, by the
Figure 5. Profile of Feldhofer Grotto skull capsuperimposed on drawing skull of AustralianAborigine (modified from Huxley, 1983).
234 THE ANATOMICAL RECORD (PART B: NEW ANAT.) ARTICLE

modification of which he has, proba-bly become what he is. And consider-ing what is now known of the mostancient Races of men; seeing that theyfashioned flint axes and flint knivesand bone-skewers, of much the samepattern as those fabricated by the low-est savages at the present day, andthat we have every reason to believethe habits and modes of living of suchpeople to have remained the samefrom the time of the Mammoth andthe tichorhine Rhinoceros till now, Ido not know that this result is otherthan might be expected.
“Where, then, must we look for pri-maeval Man? Was the oldest Homosapiens pliocene or miocene, or yetmore ancient? In still older strata dothe fossilized bones of an Ape moreanthropoid, or a Man more pithecoid,than any yet known await the re-searches of some unborn paleontolo-gist?
“Time will show. But, in the mean-while, if any form of the doctrine ofprogressive development is correct,we must extend by long epochs themost liberal estimate that has yet beenmade of the antiquity of Man” (Hux-ley, 1863: p. 183–184).
In spite of this challenging and evenfor the time heretical speculationabout humans having extinct relativesof considerable antiquity, Huxley sub-sumes the Feldhofer Grotto Neander-thal within the species Homo sapiensas an extension into the past of a con-tinuum of a racial hierarchy—por-trayed in morphology and in intelli-gence as inferred from associatedlevels of technological sophistica-tion—that was widely perceived to ex-ist among modern “races” of humans(Darwin, 1871). In downplaying, if notjust ignoring, the details of morpho-logical difference between the Nean-derthal specimen and Homo sapiens,Huxley conflated within-species dif-ferences due to individual variation(i.e., differences in the degree of ex-pression of a particular feature, as inrobustly versus weakly developedbrows) with between-species differ-ences that reflect taxic diversity (i.e.,differences of kind or configuration ofa feature, as in bipartite versus con-tinuous brows).
With the clarity that hindsight pro-vides, it is obvious that Huxley’s em-phasis on degrees of difference in
overall size and shape of features (e.g.,pronounced versus unpronouncedbrow regions, distended versus flatteroccipital regions, or more roundedversus more elongated skulls) laid thegroundwork for what became com-monplace in paleoanthropologicalthinking: that common themes in in-dividual variation (e.g., robustnessversus gracility) are as systematicallyrelevant, if not more so, than the mor-phological details of the regions beingscrutinized (e.g., bipartite versus con-tinuous brow, whether either is robustor gracile) (e.g., Hrdlicka, 1927;Broom, 1951; Smith and Ranyard,1980; Brauer, 1981; Thorne and Wol-poff, 1982; Wolpoff, 1989; Rightmire,1990; Rosas and Bermudez de Castro,1998; Asfaw et al., 2002). For, as morespecimens of Neanderthals and otherhominids became known during thelate 19th century and on into the 20th,Huxley’s suggested morphologicalcontinuum, although based on a ra-cial hierarchy that was seemingly ex-punged from the domain of paleoan-thropology, influenced how these andsubsequently discovered fossils wereincorporated into what was seen as analmost limitless realm of human vari-ability (e.g., Hrdlicka, 1927; McCownand Keith, 1939; Dobzhansky, 1944;Weidenreich, 1947; Mayr, 1950; How-ell, 1952, 1960; Thorne and Wolpoff,1982; Wolpoff, 1989). In turn, thisperception of unbounded human vari-ability was transformed into a self-fulfilling prophecy: the more differentobjects one lumps together and de-fines as variants of the same thing, thegreater becomes a presumed realm ofvariability, which then dilutes the sys-tematic significance of difference andmakes increasingly easy the justifica-tion of each subsequent addition tothis construct of variability.
AN ALTERNATIVE TO HUXLEY?WILLIAM KING ANDNEANDERTHALApproximately the same year as thepublication of Man’s Place in Nature, aprofessor of geology at Queen’s Col-lege, Galway, Ireland, William King,read a now-lost paper at a meeting ofthe British Association in which heconcluded that the differences be-tween the Feldhofer Grotto Neander-thal and humans were sufficient to
warrant recognition of the former inits own species, for which he coinedthe binomen Homo neanderthalensis.A year or so later, in an article pub-lished in 1864, King reviewed in detailthe morphology of the Neanderthalremains and resoundingly rejectedwhat he saw as Huxley’s superficialanalysis of them. As King (1864: p. 92)summarized:
“Huxley, while admitting that it isthe most ape-like and most brutal ofall human skulls yet discovered, statesthat it is ’closely approached’ by someAustralian forms, and ’even moreclosely affined to the skulls of certainancient people, who inhabited Den-mark during the Stone period.’ I haveno intention to deny that there aregeneral features of resemblance be-tween the Australian, Neanderthal,and ancient Danish crania; but it ap-pears to me, judging from the figures. . . in the deeply philosophical work,’Man’s Place in Nature,’ that a closerresemblance is assumed than reallyexists. No one would have any hesita-tion in admitting that the Borreby[Australian Aborigine] skull, repre-sented under one of the figures, isstrictly human,—nay, from what Ihave seen myself, I have no hesitationin saying that precisely the same cra-nial conformation is often repeated inthe present day; but it has yet to beshown that any skulls hitherto foundare more than approximately similarto the one under consideration.”
King (1864: p. 96) believed that theNeanderthal, while showing some fea-tures seen in humans during fetal de-velopment, was more chimpanzee-like and thus had its “closest affinitiesto the apes.” This alternative to Hux-ley’s conclusion should not be surpris-ing considering the fact that, asidefrom a smattering of not very ancienthuman skeletal remains, the primarycomparisons available to King and toHuxley for the Feldhofer Grotto Nean-derthal were with extant apes and hu-mans. While Huxley thought that thelarge Neanderthal brain was lesspithecoid than the shape of its skullmight suggest, King (1864: p. 96) in-terpreted the supposed pithecoid-likecranial features as reflecting a brainwhose “thoughts and desires . . . neversoared beyond those of the brute.”Like Huxley, however, King (1864: p.96) was not averse to ranking human
ARTICLE THE ANATOMICAL RECORD (PART B: NEW ANAT.) 235

races on the basis of a perceived“primitiveness” of skull and/or intel-lectual prowess (e.g., “the Andamaner. . . stand[s] next to brute beknighted-ness”). But whatever transpired inKing’s mind between his oral presen-tation, when he named Homo nean-derthalensis, and his composition ofthe 1864 article, he came to believethat the Feldhofer Grotto Neanderthalshould not even be included in thegenus Homo. Regardless of their dif-ferent taxonomic conclusions, it isclear that Huxley’s and King’s argu-ments came from the same mold: try-ing to decide where to draw the linebetween the lowest of human racesand the highest of the brutes.
TWENTIETH-CENTURYPALEOANTHROPOLOGY ANDTHE STATE OF HUMANEVOLUTIONWhat is interesting about 20th-cen-tury paleoanthropologists is thatmany of them appear to have losttouch with the social and historicalbeliefs that shaped the interpretationsof their heroes, Huxley and especiallyDarwin. Thus, as already mentionedabove, Huxley is said to have demon-strated a close evolutionary relation-ship between humans and the Africanapes when in fact he did not. Rather, if(rightly or wrongly) we imbue Hux-ley’s essay with a phylogenetic theme,we must conclude that he argued forthe relatedness of humans to a greatape group. A similar misunderstand-ing exists with regard to the claim thatHuxley demonstrated that Neander-thals were not a species distinct fromHomo sapiens. Nevertheless, althoughneither of these inferences is accurate,Huxley at least discussed morphology.Darwin did not.
Beginning with Raymond Dart’s(1925) discussion of the human-likeTaung specimen, paleoanthropolo-gists have continually praised Darwin(1871) on two points: his insightsabout humans being closely allied tothe African apes, and originating inAfrica. Yet Darwin did not provideone whit of morphology in The De-scent of Man or any other discourse insupport of either assertion (Schwartz,1987, 1999b). Nevertheless, his sup-posed demonstration of a close hu-man-African ape relationship is now
so entrenched in paleoanthropologi-cal lore that one can only concludethat those who cite this assertion havenot actually read The Descent. Buteven a casual reading of part 1 of thisopus reveals that Darwin’s statementis not based on morphology. Instead,it is based on a selectionist argumentfor why human evolution could nothave occurred in the “danger-free”setting of southeast Asia (the home ofthe red ape, the orangutan) as well ason sustaining a hierarchically race-based notion of the evolution of civi-lized from primitive “man” (Schwartz,1987, 1999b). Even though Darwindid not state this outright either in TheDescent or in his notebooks or letters,it seems reasonable, given his Victo-rian mindset, to infer that his empha-sis on Africa as the seat of humanorigins rested on the presence on thatcontinent of both primitive humans(“Negroes” as in many of his exam-ples) and black African apes.
To return to Mayr’s description ofhominid taxonomy as a “bewilderingdiversity of names,” it is certainly truethat the years following King’s recog-nition of Neanderthal as a species ofHomo witnessed the naming of aplethora of hominid genera and spe-cies. This should not, however, be sur-prising. The history of paleoanthro-pology did not begin in 1669, whenSteno infused fossils with biologicalreality. Rather, it was not until the1886 publication of the Spy Neander-thals that humans were broadly rec-ognized as having a fossil record. Con-sequently, when Mayr insertedhimself into paleoanthropology, thescience of seeking, discovering, inter-preting, and potentially naming ex-tinct human taxa was still quiteyoung. Furthermore, in contrast tothe centuries-older discipline of non-human paleontology, whose practitio-ners were comparative anatomistsand geologists, the study of humanfossils was dominated by human anat-omists (e.g., beginning with Schaaff-hausen), whose focus typically wasand still is on the minutiae of humanvariation. Thus, rather than being in-corporated into the broader arena ofpaleontology and systematics, fromits very beginnings paleoanthropologyremained a distinctly separate disci-pline, no doubt torn between Huxley’slegacy of the ancientness of human
variability (and the appeal of that tohuman anatomists), the tradition ofpaleontology, systematics, and taxon-omy, and a history of centuries of de-bate on the nature of “being human.”The issue of what it meant to be “hu-man” was clearly as relevant to Hux-ley’s and King’s discussions of theFeldhofer Grotto Neanderthal as itwas to earlier philosophers (e.g.,Rousseau and Monboddo) and natu-ralists (e.g., Linnaeus and Buffon),who endeavored to reflect qualities ofhumanness versus brutishness intheir classifications (Schwartz, 1987).
Another element of the history ofhuman paleontology appears to derivefrom the legitimization of human an-tiquity. First and foremost, this ac-creditation made possible explorationthat was motivated by the goal of find-ing human fossils. Indeed, in the1890s the Dutch paleontologist, geol-ogist, comparative anatomist, and hy-drologist Eugene Dubois was the firstto seek purposefully and discover hu-man fossils, which he initially referredto the taxon Anthropopithecus erectusbut subsequently changed to Pithe-canthropus erectus (Dubois, 1892,1894). By the third decade of the 20thcentury, additional Neanderthal spec-imens as well as the Mauer or Heidel-berg mandible (the holotype of Homoheidelbergensis) had been discoveredin Europe, a skull from what is nowZambia had had bestowed on it thespecies name Homo rhodesiensis, Si-nanthropus pekinensis had been basedon remains excavated from northernChina, and a handful of species, di-vided between three different genera(Australopithecus, Plesianthropus, andParanthropus), had been recognizedfrom a few sites in South Africa. Dur-ing the 1940s, as excavations for hu-man fossils proceeded apace in theOld World, the unearthing of new andoften unexpectedly different-lookingspecimens led to their being givennew species or even genus and speciesnames [e.g., a somewhat crushedmandible with some teeth from theSouth African site of Swartkrans wasnamed Telanthropus capensis becauseit was smaller and less robust thanspecimens from the same site allo-cated to Paranthropus crassidens (Tat-tersall and Schwartz, 2000)].
236 THE ANATOMICAL RECORD (PART B: NEW ANAT.) ARTICLE

ERNST MAYR, RACE, AND THEFATE OF PALEOANTHROPOLOGYWhen Mayr published his 1950 arti-cle, he was confronted both by a his-tory of seeking human fossils that hadnot yet spanned 60 years and by adiscipline that had barely had time toconfront a sampling of human fossilsthat was only then becoming largeenough for substantial critical com-parative study and reflection. As such,and in contrast to his proclaimed in-terest in the history of evolutionarybiology, Mayr’s antipathy toward thethen-current state of paleoanthropol-ogy seems insensitive to the disci-pline’s particular history. Never-theless, with characteristic self-assur-ance. Mayr assumed the role of thesystematist who, although unfamiliarwith the specimens themselves, wasgoing to clean up the mess.
Mayr was also clearly unapprecia-tive of perhaps the most significantconsequence of human evolution: al-most uniquely among organisms,Homo sapiens is the only survivingspecies of its immediate clade. Conse-quently, in contrast to the taxic worldin which Mayr typically operated, andfrom which systematists gainedknowledge that they applied to thestudy of the fossil record, one cannotachieve insight into human evolutionby studying living humans alone.Thus, if one adopts Huxley’s and sub-sequently Mayr’s notion of a pur-ported vastness of human variation inboth the present and the past, one willsee human fossils in this context andblur the distinction between morpho-logical variation within species anddifferences among taxa. If, however,one embraces the general pattern ofdiversity among extant nonhumantaxa as a model for interpreting anyfossil record, human or otherwise,one will recognize fossil taxa on thebasis of kinds (versus degrees) of dif-ference. With regard to either focus,the biggest mental and visual hurdleone must overcome when engaging inhominid systematics is that, no matterwhich ape is the closest living humanrelative, it does not look very muchlike Homo sapiens.
With choices available to him—di-versity versus variation—why didMayr (1950), the systematist of thesynthesis, opt for collapsing the bulk
of the then-known human fossilrecord into a mere three specieswithin the same genus, Homo? He be-gan with an example from the fruit flygenus Drosophila, in which at least600 species had then been recognizedand which, he claimed, “are probablymore different from each other thanare the various primates and certainlymore than the species of the suborderAnthropoidea” (Mayr, 1950: p. 109).This is clearly an overstatement inas-much as the morphological criteria onwhich Drosophila species were typi-cally based were often little more thandifferences in thoracic bristle numberor wing or antenna shape (Morgan etal., 1926). In fact, many fruit flyspecies, including one Dobzhansky(1935) named, D. miranda, were dis-tinguished not so much on the basis ofmorphology, but on the basis of dif-ferent aspects of their chromosomesor karyotypes (see also Spencer,1963).
Mayr’s (1950: p. 110) next argumentrests on the claim that “recent studiesindicate that the genus is not merely amorphological concept but that it hasa very distinct biological meaning.Species that are united in a given ge-nus occupy an ecological situationwhich is different from that occupiedby the species of another genus, or, touse the terminology of Sewall Wright,they occupy a different adaptive pla-teau. It is part of the task of the tax-onomist to determine the adaptivezones occupied by the various genera.The adaptive plateau of the genus isbased on a more fundamental differ-ence in ecology than that between theecological niches of species.”
Aside from the circularity of thisstatement, it is important to note thatMayr is focusing on ecological situa-tions—niches, not morphology—inmaking taxonomic judgments. Thismight seem logical, inasmuch as theconcept of the ecological niche is thecrux of the biological species conceptthat Mayr (1942) proposed 8 yearsearlier: in order for a new species toarise (i.e., for speciation to begin),there must first be a vacant econicheinto which a subspecies of an existentspecies can invade. Once there, asMayr envisioned the process, geo-graphic isolation in conjunction withthe different circumstances of varia-tion and selection will gradually alter
this incipient species until it also be-comes reproductively isolated fromthe parent species. If, sometime later,a member of this new species were ina position to mate with individuals ofits parental species, the offspringwould be infertile and/or not viable.
With these assumptions in place,Mayr (1950: p. 110) then declares: “Af-ter due consideration of the many dif-ferences between Modern man, Javaman, and the South African ape-man,I did not find any morphological char-acters that would necessitate separat-ing them into several genera. Not evenAustralopithecus has unequivocalclaims for separation. This form ap-pears to possess what might be con-sidered the principal generic charac-ter of Homo, namely, upright posturewith its shift to a terrestrial mode ofliving and the freeing of the anteriorextremity for new functions which, inturn, have stimulated brain evolu-tion.”
This statement alone should leadone to question Mayr’s knowledge ofhominid comparative morphology.Perhaps less comprehensible, how-ever, is why any paleoanthropologistshould have taken or should still takeMayr’s argument for taxonomic mini-malism seriously (e.g., see White,2003). Indeed, it is difficult to under-stand how and/or why Mayr wouldgloss over and diminish differencesbetween various hominid fossils andmodern humans that are so strikingthey make the criteria on which dif-ferent species of equid were at thetime distinguished pale by compari-son (e.g., see Simpson, 1944). I shallreturn to this point in a moment. Butto continue, having embraced anotion of presumed widespreadgeographical and “racial” variationamong living humans, Mayr (1950: p.112) makes the following unsubstan-tiated assertions:
“I interpret the available literatureto indicate that primitive man showedmore geographical as well as individ-ual variation than modern man.
“Why primitive man should havebeen more variable than modern manis not entirely clear. . . . Whatever thereasons, we should not use the vari-ability within populations of modernman as a yardstick by which to judgethe probable variability of extinct pop-ulations.
ARTICLE THE ANATOMICAL RECORD (PART B: NEW ANAT.) 237

“This point is important because itbears on the question whether or notmore than one species of hominid hasever existed on the earth at any onetime. Indeed, all the now available ev-idence can be interpreted as indicat-ing that, in spite of much geographi-cal variation, never more than onespecies of man existed on the earth atany one time.”
How can Mayr state that there hasnever been at one time more than onespecies of hominid? Because the de-gree of inferred variation (inferred aposteriori after first grouping speci-mens biochronologically) is used tosuggest that there are not now andnever were empty econiches thathominids could invade (Mayr’s re-quirement for speciation). To bolsterhis claim, Mayr cites an argument for-mulated by Dobzhansky (1944), thefruit fly population geneticist who au-thored the first monograph in the se-ries that provoked the evolutionarysynthesis, Genetics and the Originof Species (Dobzhansky, 1937). AsDobzhansky (1944) imagined it, asculture-bearing organisms hominidsexempted themselves from natural se-lection because they could produce,manipulate, and carry with them theirown “environmental” circumstances.Along with their widespread geo-graphic variation, hominids thus be-come even more unique and furtherseparated from nature. From this per-spective, Mayr then concludes thatthere was only one genus, Homo, and,in ascending order of succession, thespecies transvaalensis, erectus, and sa-piens, which, of course, because of thetransformational nature of this se-quence, are only arbitrary constructs.
But another—probably the ulti-mate—reason underlying Mayr’s(1950: p. 116) unsubstantiated asser-tion about humans being the onlyknown nonspeciating metazoan is re-vealed in this passage:
“If fossils of Congo pygmies and ofWatusi were to be found in the samedeposit by a paleontologist, a millionyears hence, he might well think thatthey belonged to two different species.As stated previously, the known diver-sity of fossil man can be interpreted asbeing the result of geographic varia-tion within a single species ofHomo. . . . What is the cause for thispuzzling trait of the human stock to
stop speciating in spite of its eminentevolutionary success? It seems to methat the reason is man’s great ecolog-ical diversity. Man has, so to speak,specialized in despecialization. Manoccupies more different ecologicalniches than any known animal. If thesingle species man occupies success-fully all the niches that are open for aHomo-like creature, it is obvious thathe cannot speciate.”
The real crux of Mayr’s argument isthus the following: if systematists ac-corded species status to Neanderthalsand other human-like fossils, or, moredangerously, recognized some ofthese fossil hominids as separate gen-era, this would leave the door open forthose who wished to do so to classifyhuman races at a taxonomic levelabove the subspecies, as some earliertaxonomists, such as Bory St. Vincent,had done (Schwartz, 1987) and evencontemporaneous taxonomists stilladvocated [e.g., Gates (1944), who, inthe same issue of the American Jour-nal of Physical Anthropology in whichDobzhansky argued that humanswere a wildly polytypic species, recog-nized five living species of human]. If,however, systematists minimized thesignificance of difference between fos-sil hominids at any point in time andbetween fossil and extant hominids—differences that, under other, non-human-focused systematic circum-stances, would be given taxonomic va-lence—then whatever differences(real or the result of prejudice) thatmight be perceived to exist among liv-ing humans were further diminished.
As a systematist who has spent acareer analyzing human skeletal mor-phology and also nearly a decade in-tensively studying virtually the entirehuman fossil record, I must rejectMayr’s dramatic oversimplification ofthe differences between “Congo pyg-mies” and Watusi. Theirs are differ-ences in degree of expression in, notin kind of, morphology (Schwartz,2003, 2006). But if we put morpholog-ical accuracy aside for the momentand try to appreciate the above quotein its historical context—the immedi-ately post-World War II years of NaziGermany—we may better understandthe raison d’etre, the underlying, un-spoken motivation, behind Mayr’sseemingly systematically neutral es-say: if supposedly primitive-looking
fossils such as Neanderthals and theZambian Kabwe skull and many othermore or less contemporaneous fossils(all of which differ markedly from anyliving human and among which areclearly defined morphological group-ings) are collapsed into the same spe-cies, it then becomes ludicrous even topretend that any perceived differencesbetween living humans are biologi-cally, and thus evolutionarily, signifi-cant (see discussion by Muller-Willie,2007). But the two points of Mayr’sessay are not as connected as onemight think. On the basis of his bio-logical species concept alone, one canargue that all living humans, as far asbiology is concerned, are members ofthe same species.
Nevertheless, the effect on paleoan-thropology of Mayr’s (1950) unin-formed sense both of human variabil-ity and of the human fossil record,together with Dobzhansky’s (1944,1955, 1962) notion that humans neverspeciated because the advent of cul-ture removed them from the pro-cesses of natural selection, was a sti-fling of intellectual curiosity that forover half a century prevented this dis-cipline from embracing the more rig-orous theoretical and methodologicalunderpinnings of paleontology andsystematics in general. Given the cloutMayr and Dobzhansky had as “fa-thers” of the evolutionary synthesis—and which Mayr maintained until hisrecent death—they should have exhib-ited greater restraint in imposing theirscenarios about human uniquenessonto the interpretation of the humanfossil record. But they did not, and theunfortunate consequence was that pa-leoanthropology became mired in asystematic “dark age” from which al-ternative thinking was largely ex-cluded. Regrettably there was someconsistency in Mayr and Dobzhan-sky’s actions, because this is preciselythe effect the evolutionary synthesishad on evolutionary biology in gen-eral: to squelch debate, and to brandas “antievolution” those who offeredalternative ideas (Mayr, 1949).
CONCLUSIONTo return to the beginning of this con-tribution, with the references to Whiteand Wilford, is it viable to claim that asurge in multiculturalism underlies
238 THE ANATOMICAL RECORD (PART B: NEW ANAT.) ARTICLE

those recent analyses of the humanfossil record that have recognizedtaxic diversity in it? If true, this wouldsuggest that cultures and species areof equal biological significance. Yet noone, including these individuals,would accept the assertion that differ-ent present-day cultures are the prod-ucts of different human species. Somaybe it is just coincidental that witha rise in multiculturalism, there hasbeen a growing appreciation amongpaleoanthropologists that the taxicblindfold that first Huxley, and thenMayr and Dobzhansky, imposed onhominid systematics has long outlivedwhatever sociological relevance itmay once have had. The history ofpaleoanthropology may have gottenoff to a rough start, but it is certainlyabout time that it took its scientificplace in the wider intellectual do-mains of paleontology and systemat-ics.
ACKNOWLEDGMENTSM. Sommer’s invitation to participatein a session on race at the 2003 meet-ing of the International Society for theHistory, Philosophy and Social Stud-ies of Biology was the impetus for thiscontribution. I thank her, as well as I.Tattersall and J. Marks for providinguseful criticism of earlier versions,and also S. Muller-Willie for sharinghis unpublished manuscript.
LITERATURE CITEDAldrovandi U. 1648. Musaeum metallicum.
Bonn: Ferronii.Asfaw B, White TD, Lovejoy O, Latimer B,
Simpson S, Suwa G. 1999. Australopithe-cus garhi: a new species of early hominidfrom Ethiopia. Science 284:629–635.
Asfaw B, Gilbert WH, Beyenne Y, HartWK, Rennel RR, WoldeGabriel G, VrbaES, White TD. 2002. Remains of Homoerectus from Bouri, Middle Awash, Ethi-opia. Nature 416:317–320.
Bermudez de Castro JM, Arsuaga J, Car-bonell E, Rosas A, Martinez I, MosqueraM. 1997. A hominid from the LowerPleistocene of Atapuerca, Spain: possibleancestor to Neandertals and modern hu-mans. Science 276:1392–1395.
Blumenbach JF. 1969. On the natural vari-eties of mankind. New York: BergmanPublishers.
Brauer G. 1981. New evidence on the tran-sitional period between Neanderthal andmodern man. J Hum Evol 10:467–474.
Broom R. 1951. Finding the missing link,2nd ed. London: Watts.
Brunet MEA. 2002. A new hominid fromthe Upper Miocene of Chad, Central Af-rica. Nature 418:145–151.
Curnoe D, Thorne A. 2003. Number of an-cestral human species: a molecular per-spective. Homo 53:201–224.
Dart R. 1925. Australopithecus africanus:the man-ape of South Africa. Nature 115:195–199.
Darwin C. 1859. On the origin of species bymeans of natural selection, or the pres-ervation of favored races in the strugglefor life. London: John Murray.
Darwin C. 1871. The descent of man andselection in relation to sex. London:John Murray.
Dobzhansky T. 1935. Drosophila miranda,a new species. Genetics 20:377–391.
Dobzhansky T. 1937. Genetics and the or-igin of species. New York: ColumbiaUniversity Press.
Dobzhansky T. 1944. On species and racesof living and fossil man. Am J Phys An-thropol 2:251–265.
Dobzhansky T. 1955. Evolution, genetics,and man. New York: John Wiley.
Dobzhansky T. 1962. Mankind evolving:the evolution of the human species. NewHaven, CT: Yale University Press.
Dubois E. 1892. Palaeontologische onder-zoekingen op Java (Paleontological in-vestigations on Java). Verslag van hetMijnwesen 3:10–14.
Dubois E. 1894. Pithecanthropus erectus,eine Mesnchenahnliche Ubergangsformaus Java. Batavia: Landesdruckerei.
Fraipont J, Lohest M. 1886. La race deNeanderthal ou de Cannstadt, en Bel-gique: recherches ethnographiques surdes ossements humains decourts dansdes depots quaternaires d’une grotte aSpy et determination de leur age ge-ologique—note Preliminaire. Bull AcadRoy Sci Lettres de Beaux-Arts 12:741–784.
Gates RR. 1944. Phylogeny and classifica-tion of hominids and anthropoids. Am JPhys Anthropol 2:279–292.
Gesner K. 1551–1558. Historia animalium.Zurich.
Gesner K. 1565–1566. De rerum fossilium,lapidium et gemmarum Maxime. Zurich:Tiguri.
Gibbons A. 2002. Glasnost for hominids:seeking access to fossils. Science 297:1464–1468.
Gingerich PD. 1976. Paleontology and phy-logeny: patterns of evolution at the spe-cies level in early Tertiary mammals.Am J Sci 276:1–28.
Grayson DK. 1983. The establishment ofhuman antiquity. New York: AcademicPress.
Gregory WK. 1910. The order of mammals.Bull Am Mus Nat Hist 27:1–524.
Grine F, Franzen J. 1994. Fossil hominidteeth from the Sangiran Dome (Java, In-donesia). Cour Forsch Senck 171:75–103.
Howell FC. 1952. Pleistocene glacial ecol-ogy and the evolution of “classic Nean-derthal” man. Southwestern J Arch 8:377–410.
Howell FC. 1960. European and northwestAfrican middle Pleistocene hominids.Curr Anthropol 1:195–232.
Hrdlicka A. 1927. The Neanderthal phaseof Man: the Huxley Memorial Lecturefor 1927. J Roy Anthropol Inst 57:249–273.
Huxley TH. 1860. Review of “the origin ofspecies.” Westminster Rev.
Huxley TH. 1863. Man’s place in nature.New York: D. Appleton.
Huxley TH. 1876. Lay sermons, addresses,and reviews. New York: D. Appleton.
Huxley TH. 1896. On the relation of man tothe lower animals. In: Man’s place innature. New York: D. Appleton. p 77–156.
King W. 1864. The reputed fossil man ofthe Neanderthal. Quart J Sci 1:88–97.
Leakey MD, Feibel CS, McDougall I,Walker AC. 1995. New four-million-year-old hominid species from Kanapoi andAllia Bay, Kenya. Nature 376:565–571.
Leakey MD, Spoor F, Brown FH, GathogoPN, Kiarie C, Leakey LN, McDougall I.2001. New hominid genus from easternAfrica shows diverse middle Pliocene lin-eages. Nature 410:433–443.
Lovejoy AO. 1942. The great chain of be-ing. Cambridge, MA: Harvard UniversityPress.
Mallegni F, Carnieri E, Bisconti M, Tar-tarelli G, Ricci S, Biddittu I, Segre A.2003. Homo cepranensis sp. nov. and theevolution of African-European MiddlePleistocene hominids. Comptes RendAcad Sci Paleo 2:153–159.
Mayr E. 1942. Systematics and the originof species. New York: Columbia Univer-sity Press.
Mayr E. 1949. Speciation and systematics.In: Jepsen GL, Simpson GG, Mayr E,editors. Genetics, paleontology, and evo-lution. New York: Atheneum. p 281–298.
Mayr E. 1950. Taxonomic categories infossil hominids. Cold Spring HarborSymposium on Quantitative Biology 15:109–118.
McCown TD, Keith A. 1939. The Stone Ageof Mount Carmel: the fossil remainsfrom the Levalloiso-Mousterian. Oxford:Clarendon Press.
Mercati M. 1717. Metallotheca vaticana.Rome.
Mivart SG. 1871. On the genesis of species.London: John Murray.
Morgan TH, Sturtevant AH, Muller HJ,Bridges CB. 1926. The mechanism ofMendelian heredity. New York: HenryHolt.
Muller-Willie S. 2007. Race and ethnicity:human diversity and the UNESCO state-ment on race (1950–1951). In: 60 yearsof UNESCO’s history. Paris: UNESCO(in press).
Pilbeam D. 1970. Gigantopithecus and theorigins of Hominidae. Nature 225:516.
Rightmire P. 1990. The evolution of Homoerectus. Cambridge: Cambridge Univer-sity Press.
Rosas A, Bermudez de Castro J.-M. 1998.On the taxonomic affinities of the Dma-nisi mandible (Georgia). Am J Phys An-thropol 107:145–162.
ARTICLE THE ANATOMICAL RECORD (PART B: NEW ANAT.) 239

Satta Y, Klein J, Takahata N. 2000. DNAarchives and our nearest relative: the tri-chotomy problem revisited. Mol PhylEvol 14:259–275.
Schwartz JH. 1987. The red ape. Boston:Houghton Mifflin.
Schwartz JH, Long VT, Cuong NL, Tatter-sall I. 1995. A review of the Pleistocenehominoid fauna of Vietnam (excludingHylobatidae). Anthropol Pap Am MusNat Hist 76:1–24.
Schwartz JH. 1995. Pseudopotto martini: anew genus and species of extant lorisi-form primate. Anthropol Pap Am MusNat Hist 78:1–14.
Schwartz JH, Tattersall I. 1996. Towarddistinguishing Homo neanderthalensisfrom Homo sapiens, and vice versa. An-thropologie 43:129–138.
Schwartz JH. 1999a. Can we really identifyspecies, fossil or living? Anthropologie37:221–230.
Schwartz JH. 1999b. Sudden origins: fos-sils, genes, and the emergence of species.New York: John Wiley and Sons.
Schwartz JH, Tattersall I. 2000. Variabilityin hominid evolution: putting the cartbefore the horse? In: Antunes M, editor.Last Neandertals in Portugal: odonto-logic and other evidence. Lisboa: Aca-demia das Ciencias de Lisboa. p 367–401.
Schwartz JH. 2001. A review of the system-atics and taxonomy of Hominoidea: his-tory, morphology, molecules, and fossils.Ludus Vitalis 9:15–45.
Schwartz JH, Tattersall I. 2002a. Bodo andthe concept of Homo heidelbergensis. In:Seidler H, Begashaw K, editors. Twenty-five years of Bodo: proceedings of the 4thPhillip V. Tobias Lecture on HumanEvolution. Addis Ababa: National Mu-seum. p 107–127.
Schwartz JH, Tattersall I. 2002b. The hu-man fossil record, vol. 1, terminologyand craniodental morphology of genusHomo (Europe). New York: Wiley-Liss.
Schwartz JH. 2003. Another perspective onhominid diversity. Science 301:763.
Schwartz JH, Tattersall I. 2003. The hu-man fossil record, vol. 2, craniodentalmorphology of genus Homo (Africa andAsia). New York: Wiley-Liss.
Schwartz JH. 2004a. Barking up the wrongape: australopiths and the quest for
chimpanzee characters in hominid fos-sils. Collegium Antropologicum 28:87–101.
Schwartz JH. 2004b. Issues in hominidsystematics. In: Baquedano E, RubioJara S, editors. Miscelanea en homenajea Emiliano Aguirre. Madrid: Museo Ar-queologico Regional de la Comunidad deMadrid. p 360–371.
Schwartz JH. 2005. The red ape: orangu-tans and human origins, 2nd rev ed.Boulder, CO: Westview Press.
Schwartz JH, Tattersall I. 2005. The hu-man fossil record, vol. 4, craniodentalmorphology of Australopithecus, Paran-thropus, and Orrorin. New York: Wiley-Liss.
Schwartz JH. 2006. Skeleton Keys: an in-troduction to human skeletal morphol-ogy, development, and analysis, 2nd reved. New York: Oxford University Press.
Schwartz JH. 2007. Defining Hominidae.In: Henke W, Rothe H, Tattersall I, edi-tors. Handbook of palaeoanthropology,volume 3, phylogeny of Hominines. Ber-lin: Axel Springer p 1–30.
Senut B, Pickford M, Gommery D, Mein P,Cheboi K, Coppens Y. 2001. First homi-nid from the Miocene (Lukeino forma-tion, Kenya). Comptes Rend Acad SciParis Ser IIa 332:137–144.
Simons EL. 1964. On the mandible of Ra-mapithecus. Proc Natl Acad Sci USA 51:528–535.
Simons EL, Pilbeam D. 1965. Preliminaryrevision of the Dryopithecinae (Pongi-dae, Anthropoidea). Folia Primatol 3:81–152.
Simpson GG. 1944. Tempo and mode inevolution. New York: Columbia Univer-sity Press.
Smith F, Ranyard G. 1980. Evolution of thesupraorbital region in Upper Pleistocenefossil hominids from south-central Eu-rope. Am J Phys Anthropol 53:589–610.
Spencer WP. 1963. Gene homologies andthe mutants of Drosophila hydei. In:Jepsen GL, Simpson GG, Mayr E, edi-tors. Genetics, paleontology and evolu-tion. New York: Atheneum. p 23–44.
Stringer CB, Hublin JJ, Vandermeersch B.1984. The origin of anatomically modernhumans in western Europe. In: SpencerF, Smith F, editors. The origins of mod-ern humans: a world survey from thefossil evidence. New York: Alan R. Liss. p51–135.
Tattersall I. 1986. Species recognition inhuman paleontology. J Hum Evol 15:165–175.
Tattersall I, Schwartz JH. 2000. Extinct hu-mans. Boulder, CO: Westview Press.
Tattersall I, Schwartz JH. 2003. Is paleoan-thropology science? naming new fossilsand control of access to them. Anat Rec(New Anat) 269B:239–241.
Thorne A, Wolpoff M. 1982. Regional con-tinuity in Australasian Pleistocene hom-inid evolution. Am J Phys Anthropol 55:337–349.
Weidenreich F. 1947. Fact and specula-tions concerning the origin of Homo sa-piens. Am Anthropol 49:39–48.
White TD, Suwa G, Asfaw B. 1994. Austra-lopithecus ramidus, a new species ofearly hominid from Aramis, Ethiopia.Nature 371:306–312.
White TD, Suwa G, Asfaw B. 1995. Austra-lopithecus ramidus, a new species ofearly hominid from Aramis, Ethiopia:corrigengum. Nature 375:88.
White TD. 2003. Early hominids: diversityor distortion? Science 299:1994–1997.
White TD, Asfaw B, DeGusta D, Gilbert H,Richards GD, Suwa G, Howell FC. 2003.Pleistocene Homo sapiens from MiddleAwash, Ethiopia. Nature 423:742–747.
White TD, WoldeGabriel G, Asfaw B, Am-brose S, Beyene Y, Bernor RL, BoisserieJ-R, Currie B, Gilbert H, Haile-SelassieY, Hart WK, Hlusko LJ, Howell FC,Kono RT, Lehmann T, Louchart A, Love-joy CO, Renne PR, Saegusa H, Vrba ES,Wesselman H, Suwa G. 2006. Asa Issie,Aramis and the origin of Australopithe-cus. Nature 440:883–889.
Winter JG. 1968. The prodromus of Nico-laus Steno’s dissertation concerning asolid body enclosed by process of naturewithin a solid. New York: Hafner.
Wolpoff MH. 1989. Multiregional evolu-tion: the fossil alternative to Eden. In:Mellars P, Stringer CB, editors. The hu-man revolution: behavioral and biologi-cal perspectives on the origins of modernhumans. Edinburgh: Edinburgh Univer-sity Press. p 62–108.
Wolpoff MH. 1996. Human evolution. NewYork: McGraw-Hill.
Woodward J. 1728. Fossils of all kinds, di-gested into a method, suitable to theirmutual relations and affinity. London:Wilkin.
240 THE ANATOMICAL RECORD (PART B: NEW ANAT.) ARTICLE