quantitative analysis of finger motion coordination in hand manipulative and gestic acts
TRANSCRIPT

Human Movement Science 22 (2004) 661–678
www.elsevier.com/locate/humov
Quantitative analysis of fingermotion coordination in hand manipulative
and gestic acts
Peter Braido, Xudong Zhang *
Department of Mechanical and Industrial Engineering, University of Illinois at Urbana-Champaign,
140 Mechanical Engineering Building, MC 244, 1206 West Green Street, Urbana, IL 61801, USA
Abstract
This article reports an experimental study that aimed to quantitatively analyze motion
coordination patterns across digits 2–5 (index to little finger), and examine the kinematic syn-
ergies during manipulative and gestic acts. Twenty-eight subjects (14 males and 14 females)
performed two types of tasks, both right-handed: (1) cylinder-grasping that involved concur-
rent voluntary flexion of digits 2–5, and (2) voluntary flexion of individual fingers from digit 2
to 5 (i.e., one at a time). A five-camera opto-electronic motion capture system measured tra-
jectories of 21 miniature reflective markers strategically placed on the dorsal surface land-
marks of the hand. Joint angular profiles for 12 involved flexion–extension degrees of
freedom (DOF’s) were derived from the measured coordinates of surface markers. Principal
components analysis (PCA) was used to examine the temporal covariation between joint an-
gles. A mathematical modeling procedure, based on hyperbolic tangent functions, character-
ized the sigmoidal shaped angular profiles with four kinematically meaningful parameters. The
PCA results showed that for all the movement trials (n ¼ 280), two principal components ac-
counted for at least 98% of the variance. The angular profiles (n ¼ 2464) were accurately char-
acterized, with the mean (±SD) coefficient of determination (R2) and root-mean-square-error
(RMSE) being 0.95 (±0.12) and 1.03� (±0.82�), respectively. The resulting parameters which
quantified both the spatial and temporal aspects of angular profiles revealed stereotypical pat-
terns including a predominant (87% of all trials) proximal-to-distal flexion sequence and char-
acteristic interdependence – involuntary joint flexion induced by the voluntarily flexed joint.
The principal components’ weights and the kinematic parameters also exhibited qualitatively
similar variation patterns. Motor control interpretations and new insights regarding the
* Corresponding author. Tel.: +1-217-265-8031; fax: +1-217-244-6534.
E-mail address: [email protected] (X. Zhang).
0167-9457/$ - see front matter � 2003 Elsevier B.V. All rights reserved.
doi:10.1016/j.humov.2003.10.001

662 P. Braido, X. Zhang / Human Movement Science 22 (2004) 661–678
underlying synergistic mechanisms, particularly in relation to previous findings on force syn-
ergies, are discussed.
� 2003 Elsevier B.V. All rights reserved.
PsycINFO classification: 2240; 2260; 2330
Keywords: Finger motion; Coordination; Kinematic synergy; Principal components analysis; Mathemat-
ical modeling
1. Introduction
Dexterity of the human hand is a fundamental attribute that enables one tomanipulate objects and convey gestic information. It is afforded by coordinated
movements of a complex and hyper-redundant biomechanical system consisting of
27 bones, 39 muscles, and over 30 degrees of freedom (DOF’s), mostly by those of
the multiple finger joints (Hepp-Reymond, Huesler, & Maier, 1996; Pagowski &
Piekarski, 1977). In fact, the fingers (or digits) of the hand are so vital in human per-
formance that amputation at the metacarpophalangeal (MCP) joints is considered to
be 54% impairment of an entire person (Engelberg, 1988). The number of possible
motion combinations that can be created by the hand biomechanical system is over-whelming. For example, merely considering the sign of the motion (e.g., + flexion/
) extension) at each of the DOF’s possessed by the hand would yield millions of
possibilities (Soechting & Flanders, 1997). Amazingly, human beings are able to
organize these DOF’s in production of a vast array of deft, purposeful, yet concerted
hand movement acts.
The process of controlling a complex and redundant system via coordinative
structures or functional synergies is evident in the performance of a variety of human
motions (Flash, 1990; Gelfand, Gurfinkel, Tsetlin, & Shik, 1971; Kelso, Southard, &Goodman, 1979; Kelso, Tuller, Bateson, & Fowler, 1984; Saltzman, 1979; Valero-
Cuevas, 2000). Such functional synergies have been identified in multi-finger iso-
metric force production as the enslaving effects (Zatsiorsky, Li, & Latash, 2000) or
lack of complete independence (Kilbreath & Gandevia, 1992, 1994; Schieber,
1995). They are believed to be due to one or a combination of three mechanisms:
(1) peripheral mechanical coupling, (2) multi-digit motor units in extrinsic and intrin-
sic flexor muscles, and (3) diverging central commands (Zatsiorsky et al., 2000). A
natural subsequent hypothesis would be that these control mechanisms may manifestthemselves in coordination of finger movement acts as kinematic synergies – classes
of movement patterns having associated kinematic characteristics. Identification of
such synergies or coordinative patterns can help elucidate or consolidate understand-
ing of the exact mechanism(s) through which the excessive DOF’s of the hand are
organized into controllable units according to a specific motor command (Fish &
zSoechting, 1992; Hepp-Reymond et al., 1996; Leijnse et al., 1993; Santello, Flan-
ders, & Soechting, 2002). Quantitative assessment of the synergies may also facilitate
mathematical modeling of complex human hand motions, which entails resolving theredundancy problem (Bernstein, 1967). The models in turn can aid in developing

P. Braido, X. Zhang / Human Movement Science 22 (2004) 661–678 663
products and systems such as hand prosthetics, hand-held tools, robotic hands, vir-
tual reality, and tele-operation. Furthermore, quantitative descriptions of normative
synergistic behavior can serve as the base-line references for diagnosis of functional
impairment as well as evaluation of rehabilitative progress or surgical success (Chiu,
1995; Chiu, Lin, Su, Wang, & Hsu, 2000).Despite considerable attention devoted to investigating the synergistic control
mechanisms underlying finger motor acts, direct in vivo kinematic studies with a
multi-finger multi-joint task paradigm have yet to be systematically explored. Elec-
tromyography (EMG) and force data during static finger actions have been rather
common measures for investigating muscle coordination or synergy (Kinoshita,
Kawai, & Ikuta, 1995; Li, Latash, & Zatsiorsky, 1998a, 1998b; Maier & Hepp-
Reymond, 1995a, 1995b; Ohtsuki, 1981; Reilly & Hammond, 2000; Schieber,
1995; Valero-Cuevas, 2000; Zatsiorsky et al., 2000). These measures do not containspatial–temporal information but usually only offer indirect inferences about motion
coordination strategies. Data gloves have also been employed to measure the joint
kinematics and identify temporal synergies of finger motions (H€ager-Ross &
Schieber, 2000; Santello et al., 2002; Soechting & Flanders, 1997). The resulting data
have permitted the use of kinematic analysis methods developed previously for gross
body movements, providing more temporal information pertinent to the motion syn-
ergies. Nevertheless, the reliability and accuracy of data glove measurement remain
questionable: for instance, the measurement error reportedly can be as large as 6�(Kessler, Hodges, & Walker, 1995). Encouragingly, recent years have witnessed an
increasing number of studies demonstrating the efficacy of surface-based opto-elec-
tronic motion capture systems in measuring finger kinematics (Chiu, Su, Wang, &
Hsu, 1998; Chiu et al., 2000; Darling, Cole, & Miller, 1994; H€ager-Ross & Schieber,
2000; Rash, Belliappa,Wachowiak, Somia, &Gupta, 1999; Somia, Rash,Wachowiak,
& Gupta, 1998). A newly proposed computational method for identifying the finger
joint centers can further enhance the accuracy in quantifying the ‘‘true’’ underlying
rigid-bodykinematics (Zhang,Lee,&Braido, 2003). Todate, however, studies employ-ing the opto-electronic measurement systems have been limited to analyses of single
finger postures or motions (Chiu et al., 1998; Darling et al., 1994; Rash et al., 1999),
fingertip traces (H€ager-Ross & Schieber, 2000), or joint flexion–extension initiation
and sequence (Somia et al., 1998).
The purpose of this study was to quantitatively examine the coordinative motion
patterns and synergies within and across digits 2–5 (index to little finger) during hand
manipulative and gestic acts, based on more direct measurement and in-depth anal-
ysis. The study focused on two types of movements: cylinder-grasping that entailsconcurrent voluntary flexion of digits 2–5, and voluntary flexion of individual fingers
from digit 2 to 5 (one at a time). We chose these movements because they are the
most elemental multi-finger manipulative and gestic acts, and had been previously
studied using different measurement or analysis methods (H€ager-Ross & Schieber,
2000; Santello et al., 2002) – a comparative look was intended. Two complimentary
analysis techniques, principal components analysis (PCA) and parametric movement
profile modeling, were integrated to quantify the spatial, temporal, and coordina-
tive characteristics. The associations between results generated by the two analysis

664 P. Braido, X. Zhang / Human Movement Science 22 (2004) 661–678
methods were inspected in an attempt to gain additional insights into finger motion
coordination and control.
2. Methods
2.1. Experimental protocol
Asymptomatic subjects were selected based on the stature through a rigorous
screening process, using a seven-stratum scheme (1st, 5th, 25th, 50th, 75th, 95th,
and 99th percentile) in reference to a normative database in the literature (Eastman
Kodak Company, 1986). Four subjects (two males and two females) with the appro-
priate heights were designated to each stratum with a tolerance of ±1 cm. The mean(±SD) of weight and age for the male subjects were 72.3 (±9.25) kg and 23.6 (±3.3)
years. The corresponding values for the female subjects were 57.8 (±8.53) kg and
24.4 (±6.3) years.
The subjects performed two types of movement tasks during the experiment: (1)
cylinder-grasping, and (2) voluntary flexion of individual fingers, both right-handed.
All movements began in a consistent seated posture with the torso upright, the right
upper arm approximately vertical and forearm horizontal on an armrest, the fingers
in natural full extension (abduction/adduction not specified), and the palm facingmedially. The seat height and the armrest could be adjusted separately. In the first
task, subjects reached forward over a distance of approximately 25 cm to grasp a ver-
tical cylindrical handle (45 mm in diameter) by concurrent voluntary flexion of digits
2–5, returned to the initial posture, paused, and repeated the grasping motion once.
Note that arm transport motion, without significant forearm pronation/supination
and torso assistance, was involved. In the second task, subjects maintained the same
initial posture as in the first task, but the right forearm was comfortably strapped to
the armrest. Each subject performed two consecutive repetitions of individuated flex-ion – voluntary flexion of individual fingers from digit 2 to 5, one digit at a time. For
the latter task, the subjects were instructed not to consciously control involuntary
joint flexion of the non-intended fingers; only the distal pulp of the voluntarily flexed
finger was allowed to contact the palmer surface (which signified the end of the flex-
ion), and movement of the distal interphalangeal (DIP) joint was minimized to better
homogenize the task conditions. The subjects were allowed to practice and rest be-
fore each trial until they felt ready to proceed with the movement that would be actu-
ally recorded. They each completed 10 trials (five different movements, tworeplicates). The experimental protocol was approved by the Institutional Review
Board of the University of Illinois at Urbana-Champaign, and informed consent
was obtained from all the participants.
2.2. Data acquisition and processing
Miniature (5-mm diameter) reflective markers were secured to the dorsum of the
subjects’ right hand at 21 palpable surface landmarks (Fig. 1a). A five-camera Vicon

Fig. 1. (a) Twenty-one miniature reflective markers were placed on the dorsal aspect of the right hand at
landmarks corresponding to various joints, and were measured during the movement tasks by a five-cam-
era opto-electronic motion capture system (Vicon 250); (b) a local coordinate system X 0–Y 0–Z 0 was con-
structed using measured coordinates of the MCP2; 3; 5 and CMC3 markers. Note that the movements of
the thumb markers were not analyzed in the current study.
P. Braido, X. Zhang / Human Movement Science 22 (2004) 661–678 665
250 opto-electronic motion capture system (Oxford Metrics Ltd., UK) recorded thereflective marker movements at a sampling frequency of 120 Hz, and then output
the time-varying marker coordinates in a three-dimensional laboratory coordinate
system (X–Y –Z) established through prior calibration. The motion data were pro-
cessed through a second order Butterworth low-pass filter with a cutoff frequency
of 5 Hz.
A local coordinate system X 0–Y 0–Z 0 was established to facilitate kinematic descrip-
tions and definitions (Fig. 1b). The origin of this local coordinate system was the

666 P. Braido, X. Zhang / Human Movement Science 22 (2004) 661–678
marker adhered to the dorsal landmark of the carpometacarpal joint of digit 3
(CMC3). The CMC3–MCP2 and CMC3–MCP5 vectors (vectors connecting the
respective CMC and MCP markers) formed a plane of the coordinate system. The
X 0-axis was the projection of the CMC3–MCP3 vector onto this plane. The Y 0-axis
lay in the plane, pointing radially while being perpendicular to the X 0-axis. The Z 0-axis was therefore normal to the plane, pointing dorsally. Coordinates of the
markers measured in the global (laboratory) coordinate system (X–Y –Z) were trans-formed and expressed in the local coordinate system (X 0–Y 0–Z 0). From the local coor-
dinates, the time-varying joint angles measuring all the involved flexion–extension
DOF’s were derived through a computational procedure that determined the finger
segmental centers of rotation (Zhang et al., 2003).
Only the flexion portions of angular profiles for the MCP, proximal interphalan-
geal (PIP), and DIP joints of digits 2–5, a total of 12 DOF’s, were analyzed inthe current study. A data truncating algorithm was established to identify the initi-
ation and termination times of the flexion motions. The initiation time was defined
as the starting frame of a 10-frame (1/12 s) time window during which the first
10% increase in the moving average of joint angles (across the 10 frames) occur-
red, and the termination time was defined as the ending frame of a 10-frame time
window during which the last 10% decrease in the moving average of joint angles
(across the 10 frames) occurred. The total movement time period (T ) was deter-
mined as the difference between the earliest initiation time and latest terminationtime among all the joint angles, and was later utilized to normalize the angular pro-
files.
2.3. Data analyses
Principal components analysis (PCA) was employed to investigate the synergistic
behavior among finger joints by reducing the dimensionality of the original set of
angular data into a more manageable and understandable format (Dunteman,1989). In other words, when more than one variables are correlated and reflecting
the same behavioral principle governing the system, the PCA could reveal to what
extent that occurs (Mah, Hulliger, Lee, & O’Callaghan, 1994). Specifically, PCA
examined the temporal covariation between the joint angles, providing a set of
ortho-normal basis functions, the dimensionality of which corresponded to the num-
ber of joint angles involved in a movement (Soechting & Lacquaniti, 1989). The
movement thus was characterized by a compilation of principal components
(PC’s), where each PC represented simultaneous changes in the joint angles by aset of fixed ratios (Santello & Soechting, 1997; Soechting & Flanders, 1997). The first
PC was the function or eigenvector that on average gave the best fit to all the data;
the second PC was the next best fit and so on in descending order of the eigenvalue
magnitude. The relative contribution of each element of a PC is referred to as the
weight (signed normalized ratio) that corresponds to a particular variable or angular
profile (Mah et al., 1994). The contribution of a component to the movement, or per-
cent variance accounted for (PVAF) due to a single component PCi (i ¼ 1; . . . ; k,where k is the number of DOF’s) was defined by
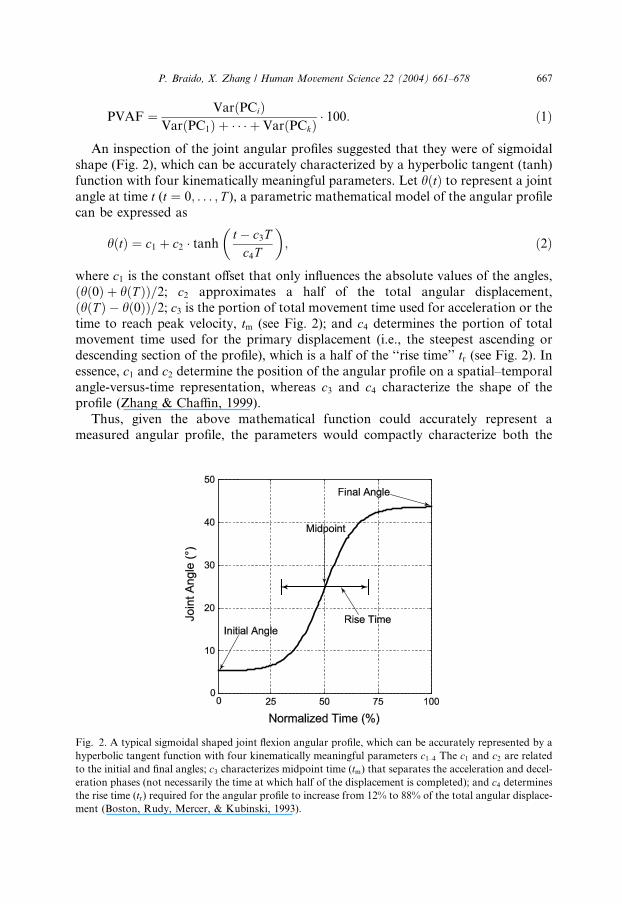
Fig. 2
hyperb
to the
eration
the rise
ment (
P. Braido, X. Zhang / Human Movement Science 22 (2004) 661–678 667
PVAF ¼ VarðPCiÞVarðPC1Þ þ � � � þ VarðPCkÞ
� 100: ð1Þ
An inspection of the joint angular profiles suggested that they were of sigmoidalshape (Fig. 2), which can be accurately characterized by a hyperbolic tangent (tanh)
function with four kinematically meaningful parameters. Let hðtÞ to represent a joint
angle at time t (t ¼ 0; . . . ; T ), a parametric mathematical model of the angular profile
can be expressed as
hðtÞ ¼ c1 þ c2 � tanht � c3Tc4T
� �; ð2Þ
where c1 is the constant offset that only influences the absolute values of the angles,
ðhð0Þ þ hðT ÞÞ=2; c2 approximates a half of the total angular displacement,
ðhðT Þ � hð0ÞÞ=2; c3 is the portion of total movement time used for acceleration or the
time to reach peak velocity, tm (see Fig. 2); and c4 determines the portion of totalmovement time used for the primary displacement (i.e., the steepest ascending or
descending section of the profile), which is a half of the ‘‘rise time’’ tr (see Fig. 2). Inessence, c1 and c2 determine the position of the angular profile on a spatial–temporal
angle-versus-time representation, whereas c3 and c4 characterize the shape of the
profile (Zhang & Chaffin, 1999).
Thus, given the above mathematical function could accurately represent a
measured angular profile, the parameters would compactly characterize both the
. A typical sigmoidal shaped joint flexion angular profile, which can be accurately represented by a
olic tangent function with four kinematically meaningful parameters c1–4 The c1 and c2 are relatedinitial and final angles; c3 characterizes midpoint time (tm) that separates the acceleration and decel-
phases (not necessarily the time at which half of the displacement is completed); and c4 determines
time (tr) required for the angular profile to increase from 12% to 88% of the total angular displace-
Boston, Rudy, Mercer, & Kubinski, 1993).
668 P. Braido, X. Zhang / Human Movement Science 22 (2004) 661–678
amplitude and timing aspects of a spatial–temporal pattern. A non-linear least-
squares curve fitting procedure was created to determine the set of parameter values
that best characterized each joint angular profile, as indicated by the coefficient of
determination (R2) and root-mean-square-error (RMSE). The search was facilitated
by setting good initial estimates of the parameters: c1 ¼ ðhð0Þ þ hðT ÞÞ=2,c2 ¼ ðhðT Þ � hð0ÞÞ=2, c3 ¼ 0:5, and c4 ¼ 0:25 (Zhang & Chaffin, 1999). Once the
angular profiles were fitted, the PC weights and c1–c4 parameters corresponding to
particular profiles were examined to identify their possible association.
All the data processing and analysis procedures were implemented using MAT-
LAB� (The MathWorks, Inc., Natick, MA). For each measured movement, these
procedures generated (1) one set of PC’s along with the PVAF values, (2) 12 sets
of PC weights, one for each DOF, and (3) 12 sets of c1–c4 parameters and corre-
sponding R2 and RMSE values.
3. Results
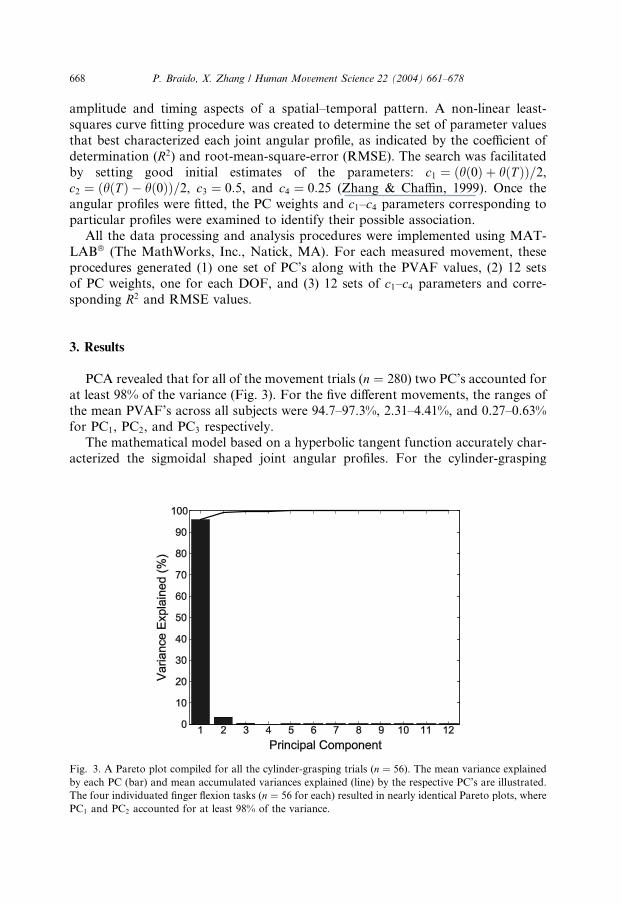
PCA revealed that for all of the movement trials (n ¼ 280) two PC’s accounted for
at least 98% of the variance (Fig. 3). For the five different movements, the ranges of
the mean PVAF’s across all subjects were 94.7–97.3%, 2.31–4.41%, and 0.27–0.63%
for PC1, PC2, and PC3 respectively.The mathematical model based on a hyperbolic tangent function accurately char-
acterized the sigmoidal shaped joint angular profiles. For the cylinder-grasping
Fig. 3. A Pareto plot compiled for all the cylinder-grasping trials (n ¼ 56). The mean variance explained
by each PC (bar) and mean accumulated variances explained (line) by the respective PC’s are illustrated.
The four individuated finger flexion tasks (n ¼ 56 for each) resulted in nearly identical Pareto plots, where
PC1 and PC2 accounted for at least 98% of the variance.
P. Braido, X. Zhang / Human Movement Science 22 (2004) 661–678 669
movements, the mean (±SD) R2 and RMSE values in fitting all 672 joint angular pro-
files (28 subjects, 12 joint angles, and 2 replicates) were 0.995 (±0.009) and 1.08�(±0.65�), respectively. For the movements involving voluntary flexion of different
individual fingers, the mean (±SD) R2 and RMSE values in fitting 448 angular pro-
files of the voluntarily flexed joints (28 subjects, 4 voluntary fingers, 2 joint angles perfinger, and 2 replicates) were 0.986 (±0.071) and 1.53� (±1.13�), respectively. Theangular profiles of the involuntarily flexed joints (i.e., joint flexion of the non-
intended fingers) were fitted less accurately than those of the voluntarily flexed joints:
the mean (±SD) R2 and RMSE values in fitting 1344 angular profiles (28 subjects, 3
sets of four involuntary fingers, 2 joint angles per finger, and 2 replicates) were 0.892
(±0.206) and 0.836� (±0.796�), respectively.Within a digit, a proximal-to-distal flexion sequence, as indicated by c3 (normal-
ized duration of the acceleration phase), was evident for all the tasks. The MCP–PIPsequence occurred during 93%, 86%, 80%, and 82% of the trials, respectively, in indi-
viduated flexion of digits 2, 3, 4, and 5 (n ¼ 56 for each). For cylinder-grasping, the
MCP–PIP–DIP and PIP–MCP–DIP flexion sequences occurred during 89% and
11% of the finger flexions (n ¼ 224), respectively. These sequences are reflected by
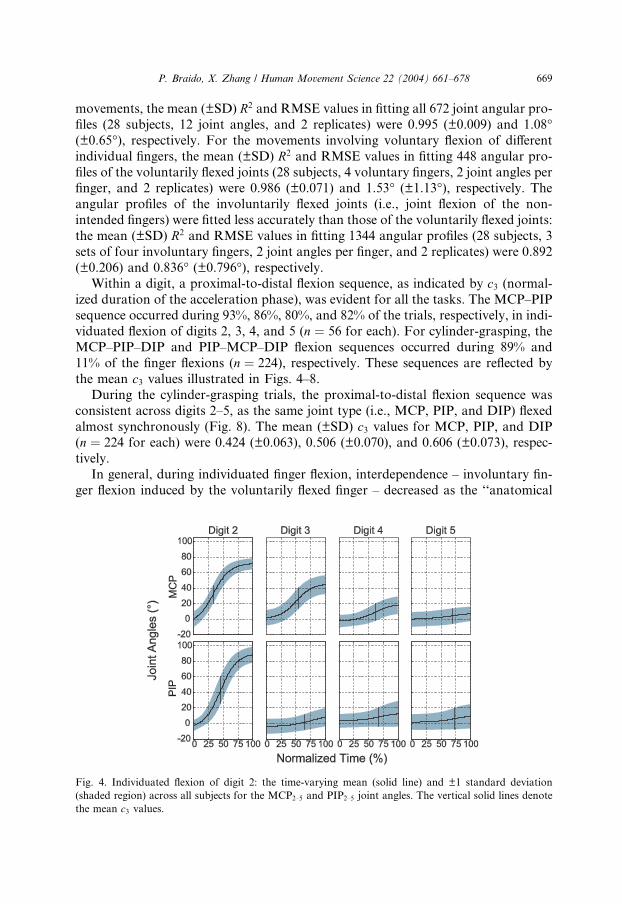
the mean c3 values illustrated in Figs. 4–8.
During the cylinder-grasping trials, the proximal-to-distal flexion sequence was
consistent across digits 2–5, as the same joint type (i.e., MCP, PIP, and DIP) flexed
almost synchronously (Fig. 8). The mean (±SD) c3 values for MCP, PIP, and DIP(n ¼ 224 for each) were 0.424 (±0.063), 0.506 (±0.070), and 0.606 (±0.073), respec-
tively.
In general, during individuated finger flexion, interdependence – involuntary fin-
ger flexion induced by the voluntarily flexed finger – decreased as the ‘‘anatomical
Fig. 4. Individuated flexion of digit 2: the time-varying mean (solid line) and ±1 standard deviation
(shaded region) across all subjects for the MCP2–5 and PIP2–5 joint angles. The vertical solid lines denote
the mean c3 values.
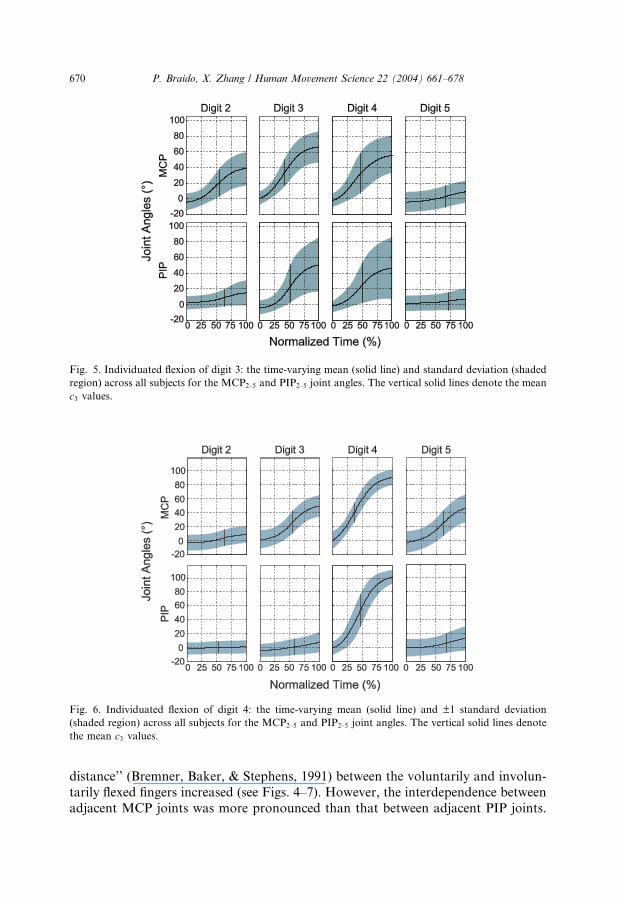
Fig. 5. Individuated flexion of digit 3: the time-varying mean (solid line) and standard deviation (shaded
region) across all subjects for the MCP2–5 and PIP2–5 joint angles. The vertical solid lines denote the mean
c3 values.
Fig. 6. Individuated flexion of digit 4: the time-varying mean (solid line) and ±1 standard deviation
(shaded region) across all subjects for the MCP2–5 and PIP2–5 joint angles. The vertical solid lines denote
the mean c3 values.
670 P. Braido, X. Zhang / Human Movement Science 22 (2004) 661–678
distance’’ (Bremner, Baker, & Stephens, 1991) between the voluntarily and involun-
tarily flexed fingers increased (see Figs. 4–7). However, the interdependence between
adjacent MCP joints was more pronounced than that between adjacent PIP joints.
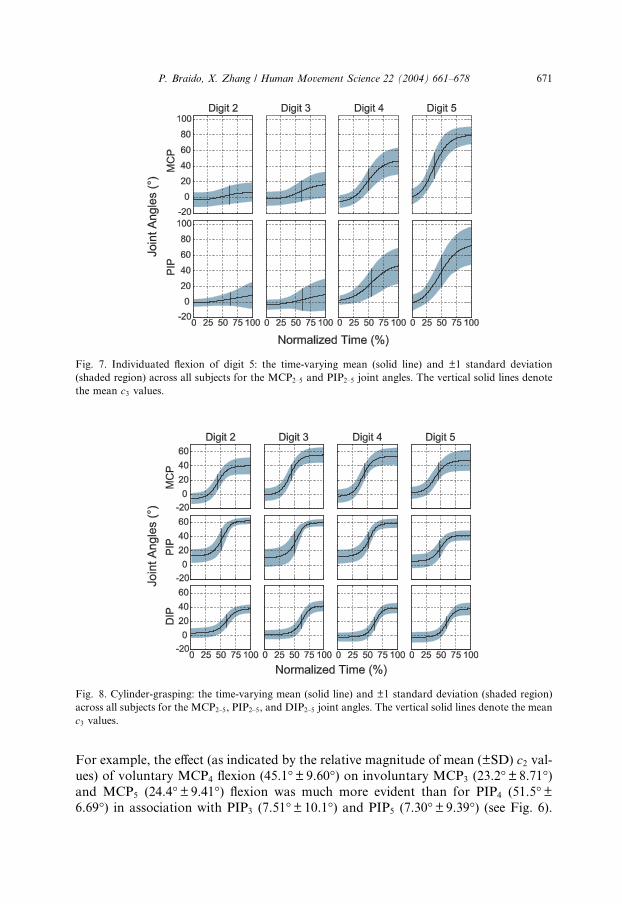
Fig. 7. Individuated flexion of digit 5: the time-varying mean (solid line) and ±1 standard deviation
(shaded region) across all subjects for the MCP2–5 and PIP2–5 joint angles. The vertical solid lines denote
the mean c3 values.
Fig. 8. Cylinder-grasping: the time-varying mean (solid line) and ±1 standard deviation (shaded region)
across all subjects for the MCP2–5, PIP2–5, and DIP2–5 joint angles. The vertical solid lines denote the mean
c3 values.
P. Braido, X. Zhang / Human Movement Science 22 (2004) 661–678 671
For example, the effect (as indicated by the relative magnitude of mean (±SD) c2 val-ues) of voluntary MCP4 flexion (45.1�±9.60�) on involuntary MCP3 (23.2�±8.71�)and MCP5 (24.4�±9.41�) flexion was much more evident than for PIP4 (51.5�±6.69�) in association with PIP3 (7.51�±10.1�) and PIP5 (7.30�±9.39�) (see Fig. 6).
672 P. Braido, X. Zhang / Human Movement Science 22 (2004) 661–678
The interdependence effects on involuntarily flexed fingers with equal ‘‘anatomical
distance’’ were nevertheless not the same in many cases. For instance, the effects
of voluntary MCP3 flexion (34.7�±14.1�) on involuntary MCP2 (22.6�±11.9�)and MCP4 (31.7�±14.8�) flexion were dissimilar, and the disparity was even more
apparent for PIP3 (28.7�±24.2�) in relation to PIP2 (7.92�±10.5�) and PIP4
(25.7�±23.9�) (see Fig. 5).
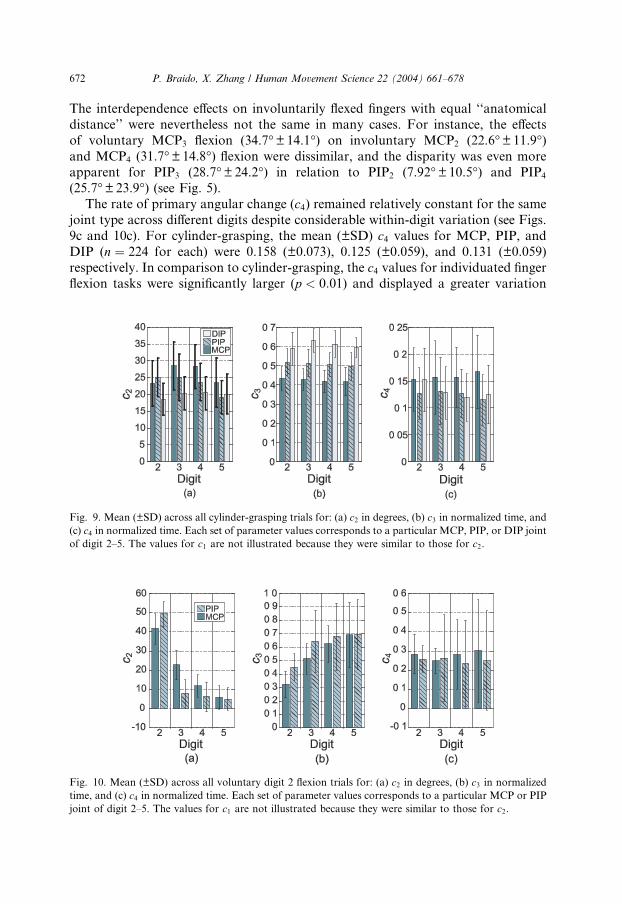
The rate of primary angular change (c4) remained relatively constant for the same
joint type across different digits despite considerable within-digit variation (see Figs.
9c and 10c). For cylinder-grasping, the mean (±SD) c4 values for MCP, PIP, and
DIP (n ¼ 224 for each) were 0.158 (±0.073), 0.125 (±0.059), and 0.131 (±0.059)
respectively. In comparison to cylinder-grasping, the c4 values for individuated finger
flexion tasks were significantly larger (p < 0:01) and displayed a greater variation
Fig. 9. Mean (±SD) across all cylinder-grasping trials for: (a) c2 in degrees, (b) c3 in normalized time, and
(c) c4 in normalized time. Each set of parameter values corresponds to a particular MCP, PIP, or DIP joint
of digit 2–5. The values for c1 are not illustrated because they were similar to those for c2.
Fig. 10. Mean (±SD) across all voluntary digit 2 flexion trials for: (a) c2 in degrees, (b) c3 in normalized
time, and (c) c4 in normalized time. Each set of parameter values corresponds to a particular MCP or PIP
joint of digit 2–5. The values for c1 are not illustrated because they were similar to those for c2.
P. Braido, X. Zhang / Human Movement Science 22 (2004) 661–678 673
(see Fig. 10c). For all individuated finger flexion tasks, the mean (±SD) c4 values forMCP and PIP (n ¼ 896 for each) were 0.276 (±0.163) and 0.228 (±0.200), respec-
tively.
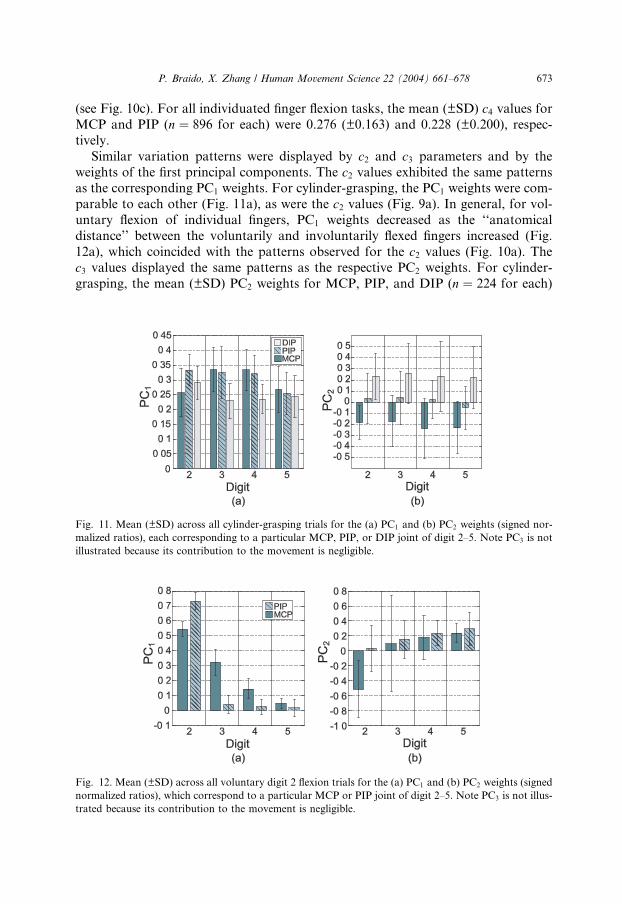
Similar variation patterns were displayed by c2 and c3 parameters and by the
weights of the first principal components. The c2 values exhibited the same patternsas the corresponding PC1 weights. For cylinder-grasping, the PC1 weights were com-
parable to each other (Fig. 11a), as were the c2 values (Fig. 9a). In general, for vol-
untary flexion of individual fingers, PC1 weights decreased as the ‘‘anatomical
distance’’ between the voluntarily and involuntarily flexed fingers increased (Fig.
12a), which coincided with the patterns observed for the c2 values (Fig. 10a). The
c3 values displayed the same patterns as the respective PC2 weights. For cylinder-
grasping, the mean (±SD) PC2 weights for MCP, PIP, and DIP (n ¼ 224 for each)
Fig. 11. Mean (±SD) across all cylinder-grasping trials for the (a) PC1 and (b) PC2 weights (signed nor-
malized ratios), each corresponding to a particular MCP, PIP, or DIP joint of digit 2–5. Note PC3 is not
illustrated because its contribution to the movement is negligible.
Fig. 12. Mean (±SD) across all voluntary digit 2 flexion trials for the (a) PC1 and (b) PC2 weights (signed
normalized ratios), which correspond to a particular MCP or PIP joint of digit 2–5. Note PC3 is not illus-
trated because its contribution to the movement is negligible.

674 P. Braido, X. Zhang / Human Movement Science 22 (2004) 661–678
were )0.206 (±0.222), 0.011 (±0.207), and 0.236 (±0.269) respectively (see also Fig.
11b). This increasing trend was in agreement with the previously described proximal-
to-distal flexion sequence indicated by the c3 values (see Fig. 9b). Furthermore, the
same patterns were noted for both the PC2 weights and c3 values for the voluntary
finger flexion tasks (see also Figs. 10b and 12b). However, the PC2 weights usuallyexhibited greater variation than the corresponding c3 values (see Figs. 9b, 10b,
11b and 12b). Overall, as the ‘‘anatomical distance’’ between the voluntarily and
involuntarily flexed fingers increased, interdependence decreased (c2 and PC1), and
phase lag increased (c3 and PC2), and the respective variation increased.
4. Discussion
Previous studies have employed various data acquisition techniques and analysis
methods to examine functional synergies underlying coordinated finger movements.
However, a comprehensive quantitative description of the synergies from a kine-
matic perspective under a multi-finger multi-joint task paradigm has been lacking.
The current study was an attempt to fulfill this void by utilizing the latest measure-
ment technology to attain in vivo kinematic data, and by combining both PCA and
mathematical modeling to achieve a thorough analysis of the data. The PCA dis-
closed the presence of a high degree of synergy in organizing the multi-finger mul-ti-joint movements. The mathematical modeling of stereotypical movement
profiles succinctly characterized the coordinative patterns and provided more direct
and physically meaningful representations of the synergistic behavior.
The advantages of more accurate measurement coupled with more in-depth anal-
ysis can be appreciated by comparing the results from the current study with those
obtained in the past. Santello et al. (2002) applied PCA to grasping motion data
measured using data gloves, and concluded that two principal components ac-
counted for about 75% of the variance. In contrast, we discovered the PVAF bythe first two PC’s were greater than 98%, suggesting a much higher level of synergy.
Some of the grasping tasks studied in that work (e.g., grasping bottle and pan han-
dle) should be similar to the cylinder-grasping studied in the current work. We spec-
ulate that this significant discrepancy is attributable to the motion artifacts and
errors introduced by the use of data gloves – these artifacts and errors inflated the
total variability of the measured data as well as the portion unexplainable by the first
two PC’s. H€ager-Ross and Schieber (2000) utilized a video camera and a data glove
simultaneously to record the fingertip positions and the finger joint angles duringindividuated finger flexion tasks (similar to the second task devised in this work).
From the recordings, four indexes were derived to quantify motion independence.
None of those indexes however characterized any temporal aspect (e.g., sequence,
phase difference) of the motion independence or synergy. Several past studies have
investigated the coordination of joint motions within a digit (as opposed to between
or across digits) in terms of characteristic ordering, temporal covariation, or muscle
activation pattern (Bendz, 1974; Darling et al., 1994; Landsmeer, 1963; Landsmeer &
Long, 1965; Somia et al., 1998). These investigations have largely been qualitative,

P. Braido, X. Zhang / Human Movement Science 22 (2004) 661–678 675
and yielded results not in concurrence with each other. For instance, Bendz (1974)
observed that the MCP3 and PIP3 joints moved synchronously during cylinder-
grasping. Darling et al. (1994) concluded that in index finger flexion–extension, by
and large, the PIP and DIP joints moved simultaneously, and there was more vari-
ation in start position between MCP and IP joints. But they also discovered substan-tial phase difference between DIP and PIP motions by some subjects. Somia et al.
(1998) reported that, during less controlled finger flexion motions, the most frequent
initiation sequence was DIP–PIP–MCP, which occurred 28% of the time. While the
ordering as well as the correlation could be task-dependent, it appears that a lack of
quantitative and discriminating measure(s) has plagued those previous investigations
and also amplified the inconsistency in the findings. In that regard, the c3 value
resulting from parametric modeling of joint angle profiles as in the current study
makes a potent candidate measure: it can not only describe the flexion sequence(which differs from the initiation sequence) but also discriminate whether two joint
motions are truly synchronous or perfectly correlated (a perfectly linear correlation
is evidenced in identical c3 values).
Meanwhile, there are noteworthy convergences between new findings on kine-
matic synergy yielded from this study and some previous findings on force synergy
or motor equivalence. The current study discovered a general trend of motion inter-
dependence decreasing with ‘‘anatomical distance’’ and also a clear lack of parity in
the interdependence for ‘‘anatomically equidistant’’ digits. These findings resemblewhat was reported by Bremner et al. (1991) based on recordings of discharges of mo-
tor units: motor units acting on fingers farther from each other were less synchro-
nized than motor units acting on the fingers closer to each other, but when the
‘‘anatomical distance’’ from the flexed digit was the same, the respective motor units
did not show the same amount of synchronization. The force synergistic ‘‘enslaving’’
effects during single finger exertions, as examined by Zatsiorsky et al. (2000), also
exhibited the similar characteristic patterns. Furthermore, Zatsiorsky et al. (2000)
demonstrated that force enslaving effects were invariant with respect to involvementof different muscle groups, thereby arguing for a more dominant role played by the
neural mechanism in finger force control than peripheral mechanical coupling or
functional organization of multitendoned extrinsic muscles. This argument is sup-
ported, from an alternative motion synergy point of view, by our observation that
a greater amount of motion interdependence occurred between adjacent MCP joints
than between adjacent PIP joints.
A comparative look within the study at the connection between outcomes of two
analysis methods and the difference between two movement tasks can also lead tosome new insights. First, the congruency in variation pattern between the kinematic
parameters and principal components (c2 versus PC1, and c3 versus PC2) suggests
that the variability is mostly due to the variation of c2 and c3. Since c2 represents
the total angular displacement (i.e., the total area underneath the velocity profile),
and c3 specifies when the peak velocity occurs, this finding may lend credence to
a velocity control strategy as an effective way of solving motor redundancy or sim-
plifying motion coordination (Zhang, Kuo, & Chaffin, 1998). Such an interpreta-
tion also echoes a prior discovery of a robust relationship between the dynamic

676 P. Braido, X. Zhang / Human Movement Science 22 (2004) 661–678
sensorimotor cortical activity and movement velocity (Kelso et al., 1998). Second,
the two movement tasks of distinctively different kinematics were shown to have
comparable levels of synergy or control simplification, as revealed by the PCA (first
two PC’s explained more than 98% of the variance). On the other hand, the individ-
uated flexions were carried out more ‘‘cautiously’’ – as they required a relativelylonger rise time (measured by c4), than the grasping. These insights provoke interest-
ing questions such as whether two kinematically different movements can be consid-
ered as equivalent at a certain rudimentary level and how movement complexity shall
be defined.
In conclusion, the present study quantified the coordinated finger motion behav-
ior and its synergistic mechanisms during several common manipulative and gestic
acts. Although such multi-finger multi-joint acts could be carried out in seemingly
an infinite number of ways, stereotypical motion patterns controlled via simplifyingkinematic synergies do emerge as evidenced in this study. Direct marker-based mea-
surement and in-depth analysis permitted a more accurate specification of the extent
to which the simplification of control (or mathematically, the condensation of
dimensionality) can be achieved. The study also rendered additional evidence to sup-
port the conjecture that while both peripheral and neural mechanisms are responsi-
ble for the synergistic finger motion behavior, the latter mechanism plays a more
dominant role in the coordination of multi-finger motions.
Acknowledgements
The authors acknowledge the financial support from Caterpillar Inc. and the
National Science Foundation (DMI 0200143), and the technical assistance by Ms.
Angela Bishop and Mr. Sang-Wook Lee.
References
Bendz, P. (1974). Systematization of the grip of the hand in relation to finger motor systems. A
kinesiologic study using a new method for recording finger joint motions. Scandinavian Journal of
Rehabilitation Medicine, 6, 158–165.
Bernstein, N. (1967). The coordination and regulation of movements. Oxford: Pergamon Press.
Boston, J. R., Rudy, T. E., Mercer, S. R., & Kubinski, J. A. (1993). A measure of body movement
coordination during repetitive dynamic lifting. IEEE Rehabilitation Engineering, 1, 137–144.
Bremner, F. D., Baker, J. R., & Stephens, J. A. (1991). Variation in the degree of synchronization
exhibited by motor units lying in different finger muscles in man. Journal of Physiology, 432, 381–399.
Chiu, H. Y. (1995). A method of two-dimensional measurement for evaluating finger motion impairment.
A description of the method and comparison with angular measurement. Journal of Hand Surgery,
20B, 691–695.
Chiu, H. Y., Lin, S. C., Su, F. C., Wang, S. T., & Hsu, H. Y. (2000). The use of the motion analysis system
for evaluation of loss of movement in the finger. Journal of Hand Surgery, 25, 195–199.
Chiu, H. Y., Su, F. C., Wang, S. T., & Hsu, H. Y. (1998). The motion analysis system and goniometry of
the finger joints. Journal of Hand Surgery, 23B, 788–791.
Darling, W. G., Cole, K. J., & Miller, G. F. (1994). Coordination of index finger movements. Journal of
Biomechanics, 27, 479–491.

P. Braido, X. Zhang / Human Movement Science 22 (2004) 661–678 677
Dunteman, G. H. (1989). Principal components analysis. Newbury Park: Sage Publications.
Eastman Kodak Company (1986). Ergonomic design for people at work (Vol. 1). New York: Van Nostrand
Reinhold.
Engelberg, A. L. (Ed.). (1988). Guides to the evaluation of permanent impairment (3rd ed.). Chicago, IL:
American Medical Association.
Fish, J., & Soechting, J. F. (1992). Synergistic finger movements in a skilled motor task. Experimental
Brain Research, 91, 327–334.
Flash, T. (1990). The organization of human arm trajectory control. In J. M. Winters & S.-Y. Woo (Eds.),
Multiple muscle systems: Biomechanics and movement organization (pp. 282–301). New York: Springer.
Gelfand, L. M., Gurfinkel, V. S., Tsetlin, M. L., & Shik, M. L. (1971). Some problems in the analysis of
movements. In L. M. Gurfinkel, V. S. Gurfinkel, S. V. Fomin, & M. L. Tsetlin (Eds.), Models of the
structural-functional organization of certain biological systems (pp. 329–345). Cambridge, MA: MIT
Press.
H€ager-Ross, C., & Schieber, M. H. (2000). Quantifying the independence of human finger movements:
Comparisons of digits, hands, and movement frequencies. The Journal of Neuroscience, 20, 8542–8550.
Hepp-Reymond, M.-C., Huesler, E. J., & Maier, M. A. (1996). Precision grip in humans: Temporal and
spatial synergies. In A. M. Wing, P. Haggard, & J. R. Flanagan (Eds.), Hand and brain (pp. 37–68).
San Diego, CA: Academic Press.
Kelso, J. A. S., Fuchs, A., Lancaster, R., Holroyd, T., Cheyne, D., & Weinberg, H. (1998). Dynamic
cortical activity in the human brain reveals motor equivalence. Nature, 392, 814–818.
Kelso, J. A. S., Southard, D. L., & Goodman, D. (1979). On the nature of human interlimb coordination.
Science, 203, 1029–1031.
Kelso, J. A. S., Tuller, B., Bateson, E.-V., & Fowler, C. A. (1984). Functionally specific articulatory
cooperation following jaw perturbations during speech: evidence of coordinative structure. Journal of
Experimental Psychology: Human Perception and Performance, 10, 812–832.
Kessler, G. D., Hodges, L. F., & Walker, N. (1995). Evaluation of the Cyberglove as a whole hand input
device. ACM Transactions on Computer–Human Interaction, 2(4), 263–283.
Kilbreath, S. L., & Gandevia, S. C. (1992). Independent control of the digits: Changes in perceived
heaviness over a wide range of force. Experimental Brain Research, 91, 539–542.
Kilbreath, S. L., & Gandevia, S. C. (1994). Limited independent flexion of the thumb and fingers in human
subjects. Journal of Physiology, 479, 487–497.
Kinoshita, H., Kawai, S., & Ikuta, K. (1995). Contributions and coordination of individual fingers in
multiple finger prehension. Ergonomics, 38, 1212–1230.
Landsmeer, J. M. F. (1963). The coordination of finger joint motions. Journal of Bone and Joint Surgery,
45A, 1654–1662.
Landsmeer, J. M. F., & Long, C. (1965). The mechanism of finger control based on electromyograms and
location analysis. Acta Anatomica, 60, 330–347.
Leijnse, J. N. A. L., Snijders, C. J., Bonte, J. E., Landsmeer, J. M. F., Kalker, J. J., Van Der Meulen, J. C.,
Sonneveld, G. J., & Hovius, S. E. R. (1993). The hand of the musician: The kinematics of the bidigital
finger system with anatomical restrictions. Journal of Biomechanics, 26, 1169–1179.
Li, Z. M., Latash, M. L., & Zatsiorsky, V. M. (1998a). Force sharing among fingers as a model of the
redundancy problem. Experimental Brain Research, 119, 276–286.
Li, Z. M., Latash, M. L., & Zatsiorsky, V. M. (1998b). Motor redundancy during maximum voluntary
contraction in four-finger tasks. Experimental Brain Research, 122, 71–78.
Mah, C. D., Hulliger, M., Lee, R. G., & O’Callaghan, I. S. (1994). Quantitative analysis of human
movement synergies: Constructive pattern analysis for gait. Journal of Motor Behavior, 26, 83–102.
Maier, M. A., & Hepp-Reymond, M. C. (1995a). EMG activation patterns during force production in
precision grip. I. Contribution of 15 finger muscles to isometric force. Experimental Brain Research,
103, 108–122.
Maier, M. A., & Hepp-Reymond, M. C. (1995b). EMG activation patterns during force production in
precision grip. II. Muscular synergies in the spatial and temporal domain. Experimental Brain
Research, 103, 123–136.
Ohtsuki, T. (1981). Inhibition of individual fingers during grip strength exertion. Ergonomics, 24, 21–
36.

678 P. Braido, X. Zhang / Human Movement Science 22 (2004) 661–678
Pagowski, S., & Piekarski, K. (1977). Biomechanics of the metacarpophalangeal joint. Journal of
Biomechanics, 10, 205–209.
Rash, G. S., Belliappa, P. P., Wachowiak, M. P., Somia, N. N., & Gupta, A. (1999). A demonstration of
the validity of a 3-D video motion analysis method for measuring finger flexion and extension. Journal
of Biomechanics, 32, 1337–1341.
Reilly, K. T., & Hammond, G. R. (2000). Independence of force production by digits of the human hand.
Neuroscience Letters, 290, 53–56.
Saltzman, E. (1979). Levels of sensorimotor representation. Journal of Mathematical Psychology, 20, 91–
163.
Santello, M., & Soechting, J. F. (1997). Matching object size by controlling finger span and hand shape.
Somatosensory & Motor Research, 14, 203–212.
Santello, M., Flanders, M., & Soechting, J. F. (2002). Patterns of hand motion during grasping and the
influence of sensory guidance. The Journal of Neuroscience, 22, 1426–1435.
Schieber, M. H. (1995). Muscular production of individual finger movements: The role of extrinsic finger
muscles. The Journal of Neuroscience, 15, 284–297.
Soechting, J. F., & Flanders, M. (1997). Flexibility and repeatability of finger movements during typing:
Analysis of multiple degrees of freedom. Journal of Computational Neuroscience, 4, 29–46.
Soechting, J. F., & Lacquaniti, F. (1989). An assessment of the existence of muscle synergies during load
perturbation and intentional movements of the human arm. Experimental Brain Research, 3, 535–548.
Somia, N., Rash, G. S., Wachowiak, M., & Gupta, A. (1998). The initiation sequence of digital joint
motion – a three dimensional motion analysis. Journal of Hand Surgery, 23B, 792–795.
Valero-Cuevas, F. J. (2000). Predictive modulation of muscle coordination pattern magnitude scales
fingertip force magnitude over the voluntary range. Journal of Neurophysiology, 83, 1469–1479.
Zatsiorsky, V. M., Li, Z. M., & Latash, M. L. (2000). Enslaving effects in multi-finger force production.
Experimental Brain Research, 131, 187–195.
Zhang, X., & Chaffin, D. B. (1999). The effects of speed variation on joint kinematics during multisegment
reaching movements. Human Movement Science, 18, 741–757.
Zhang, X., Kuo, A. D., & Chaffin, D. B. (1998). Optimization-based differential inverse kinematic
modeling exhibits a velocity-control strategy for dynamic posture determination in seated reaching
movements. Journal of Biomechanics, 31, 1035–1042.
Zhang, X., Lee, S.-W., & Braido, P. (2003). Determining finger segmental centers of rotation in flexion–
extension based on surface marker measurement. Journal of Biomechanics, 36, 1097–1102.