protective effects of alcoholic beverages and their constituent
TRANSCRIPT

1
Chemoprevention of Cancer and DNA Damage by Dietary Factors
35a. Protective Components of Alcoholic Beverages - Wine
Philipp Saiko* & Thomas Szekeres
Clinical Institute of Medical and Chemical Laboratory Diagnostics
Medical University of Vienna, General Hospital of Vienna
Waehringer Guertel 18-20
A-1090 Vienna, Austria
*Corresponding author:
DDr. Philipp Saiko
Phone: (43) 1 40400 ext. 54960
Fax: (43) 1 292 77 08 17
E-mail: [email protected]
1. INTRODUCTION
1.1. General information & historical background
Wine is an alcoholic beverage made from fermentation of fruit or grape juice. Concerning grapes, a
wine may consist of a single type of grape, or may contain a blend of different grapes. The natural
chemical balance of grapes is such that they can ferment without the addition of sugars, acids,
enzymes or other nutrients. Although other fruits such as apples and berries can also be fermented,
the resultant "wines" are normally named after the fruit from which they are produced (e.g., apple
wine) and are generically known as fruit or country wine. Others, such as barley wine and rice wine
(e.g., sake) are made from starch-based materials and resemble beer more than wine, while ginger
wine is fortified with brandy. In these cases, the use of the term "wine" is a reference to the higher
alcohol content rather than to the production process. Law in many jurisdictions protects the

2
commercial use of the English word „wine“ and its equivalent in other languages. Wine is produced
by fermenting crushed grapes using various types of yeast that consume the sugars being included
in the grapes and convert them into alcohol. Various varieties of grapes and strains of yeasts are
used depending on the types of wine produced.
The scientific history of wine stretches back much longer than its first written account in the Bible,
with the earliest evidence dating around 5000 BC. A pottery jar recovered in present-day Iran
provides the earliest chemical evidence so far discovered [1]. The wine was identified by the
presence of calcium salts of tartaric acid, only present in large amounts in grapes and in the resins
of terebinth trees [2]. This resin was widely used in ancient times as an additive to wine to inhibit
bacterial growth. Though wild grape trunks have been found to originate as far back as the eighth
millennium, this archaeological discovery marks the earliest scientific record of fermented wine as
part of human culture. From the fifth millennium BC wine spread from its postulated origins in the
Southern Caucasus to Palestine, Syria, Egypt and Mesopotamia and subsequently to the
Mediterranean [2], but after the fall of the Roman Empire wine making declined. During the Dark
Ages, wine making was kept alive mainly through the efforts of Christian monasteries. As the
Church extended their monasteries, they began to develop some of the finest vineyards in Europe.
In this Medieval period, wine was still considered a staple of everyday diet, because most of Europe
lacked reliable sources of drinking water. However, in the following centuries wine had to face the
rival of a clean and readily available supply of drinking water, and was no longer needed as a major
part of the daily diet.
1.2. The health effects of wine
In 1992, Renaud and de Lorgeril observed a lower mortality rate of coronary heart disease in France
in comparison to other Northern European countries and the USA, despite a similar intake of high
levels of saturated fat, identical smoking habits and lack of exercise [3]. This observation was
brought to the attention of the medical society, lay public, and became known as the “French

3
Paradox”. The authors explained the paradox by the consumption of the so-called „Mediterranean
diet”, with an abundance of vegetables, fruits, olive oil, and – especially – red wine. Concerning
France, they suggested that this difference is attributed to the high consumption of (red) wines by
the French population. In the USA, a boom in red wine consumption was initiated in the 1990s by
“60 Minutes“, and other news reports on the “French paradox”. Population studies have observed a
J curve association between wine consumption and the risk of heart disease. This means that
abstainers and heavy drinkers have an elevated risk, whilst moderate drinkers have a lower risk.
They also found that moderate consumption of other alcoholic beverages might be cardioprotective,
though the association is considerably stronger for wine.
1.3. Ingredients of wine
Wine is a rich source of biologically active phytochemicals, chemicals found in plants.
Phytochemicals are divided into distinct subgroups according to their structure and function in the
plant. The polyphenols are one of the most prominent groups in disease prevention. Currently, more
than 8000 of them have been identified, which are ubiquitous in plant-borne foods. Polyphenols are
associated not only with color and with sensory properties, but also linked to the health benefits
ascribed to fruits, vegetables, and wine.
The grape polyphenols may be classified into the following three groups:
1. Nonflavonoids, derived from hydroxycinnamic acids (paracoumaric acid, caffeic acid
chlorogenic acid, and ferulic acid) and hydroxybenzoic acids (gallic acid, protecatechouric
acid, and vanillic acid),
2. Flavonoids, comprising the largest class (several thousand) of phenolic compounds
including flavonols (e.g., querecetin and myricetin), isoflavonols, flavanones, flavanals (e.g.,
catechin, epicatechin, and procyanidin), and anthocyanins (e.g., delphinidin, cyanidin, and
malvidin),

4
3. Stilbenes, constituting a relatively small group of phenolic compounds that are usually
synthesized in plants in response to stress conditions. Their structures contain two benzene
rings connected by a methylene bridge. Resveratrol (3,4’,5-trihydroxy-trans-stilbene; RV) is
the most extensively studied stilbene derivative.
Most flavonoids occurring in plants are conjugated to sugar-moieties, pectins, and organic acids, or
are polymerized with other flavonoids to polymers. Products of many fruits and vegetables, such as
strawberries, blueberries, green and black tea, tomatoes, yellow onions, soy, and chocolate, contain
considerable amounts of polyphenols. Red grapes and wine of the Vitis vinifera varieties are
especially rich in these polyphenols, and more than 500 phenolic compounds have been recognized
in wine thus far [2]. Evidence from laboratory studies suggests that red wine may possess superior
health benefits including the prevention of cancer because it contains more polyphenols than white
wine, which is due to the production process (see below). Red wines obtained by traditional
maceration can have a polyphenol content of more than 3 g/L. RV is thought to be at least partly
responsible for the health benefits of red wine, since it has been shown to exert a range of both
cardioprotective as well as chemoprotective mechanisms, thus being the most prominent of these
polyphenols. Red wine contains much greater amounts of RV than white wine does, since RV is
concentrated in the grape skins and seeds, and the manufacturing procedure of red wine includes
prolonged contact of grape juice with these parts.
Plant polyphenols are recognized for their antioxidative activities, thereby protecting cells from
oxidative damage caused by free radicals. Electron acceptors such as molecular oxygen react easily
with them to become reactive oxygen species (ROS). Polyphenols scavenge free radicals, thus
breaking the free radical chain reaction of lipid peroxidation, which has been implicated in the
development of cancer. However, it is inherently difficult to evaluate the beneficial effects of
specific polyphenolic antioxidants, since a large number of individual compounds may occur in a

5
single food. For example, over sixty different chemically distinct flavonoids are known to occur in a
given red wine. Numerous scientific studies have been conducted to attempt to arrive at one
consistent index for food antioxidant power. Since it has been proven that the dietary intake of
compounds exerting antioxidant activity is of great medical value, a number of chemical, biological,
and electrochemical methods have been proposed to evaluate the antioxidant potential of naturally
occurring agents such as RV.
2. PHYSICO-CHEMICAL PROPERTIES OF ACTIVE COMPOUNDS, OCCURRENCES,
AND CHEMICAL STRUCTURES
2.1. Resveratrol
2.1.1. History & sources
Resveratrol (3,4’,5,-trihydroxy-trans-stilbene; RV; Fig. 1) was first isolated in 1940 as an
ingredient of the roots of white hellebore (Veratrum grandiflorum O. Loes) and has since been
found in a wide variety of about 70 plant species, including grapes, mulberries, and peanuts [4]. RV
is a polyphenol and has been classified as a phytoalexin for being synthesized in spermatophytes in
response to injury, UV irradiation, and fungal attack. RV was identified in 1963 as the active
constituent of the dried roots of Polygonum cuspidatum, also called Ko-jo-kon in Japanese, and
used in traditional Asian medicine against suppurative dermatitis, gonorrhea, favus, and
hyperlipemia [5]. RV was first detected in grapevines (Vinis vitifera) in 1976, and then in wine in
1992 [6]. In grapes, especially when infected with Botrytis cinerea, RV is exclusively synthesized
in the leaf epidermis and in the grape skins, but not in the flesh. Fresh grape skins contain 50-
100mg RV per g, corresponding to 5-10% of their biomass [7]. Since the grape skins are not
fermented in the production process of white wines, only red wines contain considerable amounts of
RV. Its concentrations measured in a sampling of red wine varieties ranged from 2 to 40µM [7].
Figure 1. Chemical structure of resveratrol (RV).

6
2.1.2. French paradox
Epidemiological studies have revealed an inverse correlation between red wine consumption and
the incidence of cardiovascular disease, a phenomenon commonly known as the “French Paradox”,
i.e. the fact that the incidence of heart infarction in France is about 40% lower than in the rest of
Europe, despite a diet being traditionally rich in saturated fat [3]. This led to the suggestion that RV
might be the active principle of red wine. Indeed, RV protects the cardiovascular system by a large
number of mechanisms including defense against ischemic-reperfusion injury, promotion of
vasorelaxation, protection and maintenance of intact endothelium, anti-atherosclerotic properties,
inhibition of low-density lipoprotein oxidation, suppression of platelet aggregation, and estrogen-
like actions [8, 9].
2.1.3. Effects of resveratrol
Besides its effects on the cardiovascular system, RV exhibits a remarkable inhibitory potential in
various stages of tumor development [9]. The antitumor activity of RV was first revealed by its
ability to reduce the incidence of carcinogen-induced development of cancers in experimental
animals [10]. Subsequently, RV has been shown to exert numerous effects that may block tumor
development at several discrete stages during the multigenic process of carcinogenesis, involving
interactions between RV and manifold targets [11]. These targets include kinases [12], steroid
hormone receptors [13], reactive oxygen species [14], ribonucleotide reductase [15], and DNA
polymerases [16]. RV causes an arrest at the S/G2 phase transition of the cell cycle [17] and is
capable of inducing differentiation and apoptosis in a multitude of human tumor cell lines. RV was
also identified as an effective inhibitor of ribonucleotide reductase (RR) [15, 18]. RR catalyzes the
rate-limiting step of de novo DNA synthesis, namely the reduction of ribonucleotides into the
corresponding deoxyribonucleoside triphosphates (dNTPs). The importance of all these targets for

7
cancer development is well known and therefore RV can beneficially contribute to cancer
prevention.
As tissue inflammation provokes tumor promotion, anti-inflammatory agents are viewed as a
valuable chemopreventive modality against this mechanism of carcinogenesis [14]. RV has been
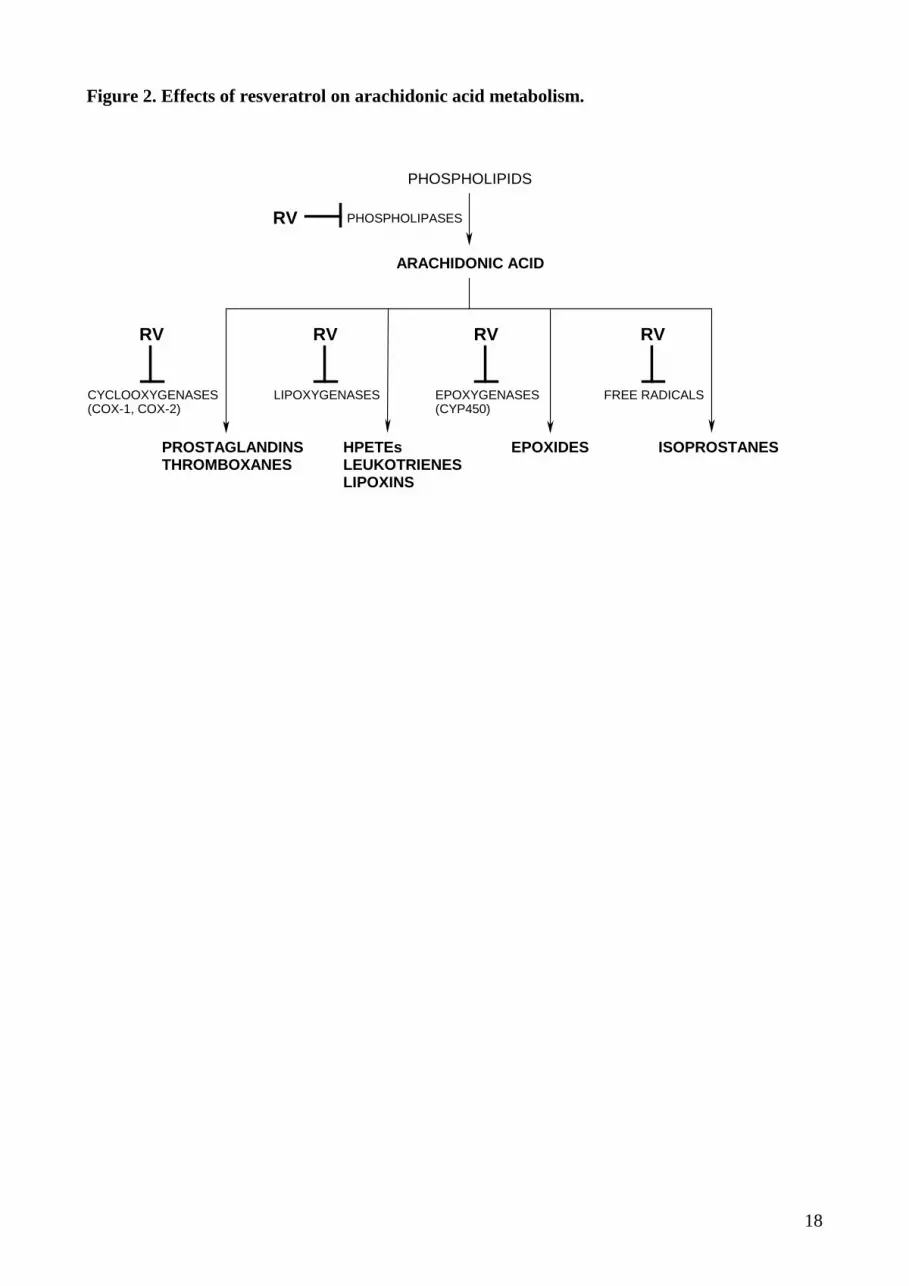
shown to exert substantial antiphlogistic activity in an in vivo rat model [19]. The key molecular
targets implicated herein are cyclooxygenases (COX-1 and COX-2). COX-1 and COX-2 are
respectively constitutive and inducible enzymes that catalyze the production of pro-inflammatory
prostaglandins from arachidonic acid [14]. Prostaglandins stimulate tumor growth by acting on cell
proliferation, angiogenesis and immunosupression. RV effects against cellular COX activity involve
its direct inhibitory action against COX-1 and COX-2 and its suppression of transcriptional COX-2
upregulation [20]. As prostaglandins not only stimulate tumor cell growth but also suppress
immune surveillance, COX enzymes are likely important targets to the cancer preventive activity of
RV [20]. Arachidonic acid is also metabolized via lipoxygenase (LOX) to produce
hydroperoxyeicosatetraenoic acids (HPETEs) or leukotrienes. Arachidonic acid metabolites derived
from LOX pathways play an important role in growth-related signal transduction, implying that
intervention through these pathways could be useful for attenuating cancer progression. RV inhibits
LOX and COX in K562 myelogenous leukemia cells [21]. LOX-derived metabolites have an
(indirect) influence on development as well as progression of human cancers [22].
Figure 2. Effects of resveratrol on arachidonic acid metabolism.
Furthermore, RV induces a multitude of effects that depend on the cell type (e.g. NF-κB modulation
in cancer cells vs. neural cells), cellular condition (normal, stressed or malignant) and concentration
(proliferative vs. growth arrest) and can have opposing activities. The final read-out depends on the
balance of these partially opposing effects. Single alterations in cell physiology, signaling and

8
metabolism result often in a cascade of changes that cannot always be restored by reversion of the
single original change. RV targets whole pathways and sets of intracellular events rather than a
single enzyme and therefore offers a less specific but more gentle (fewer side effects) and possibly
more effective strategy for therapy to restore homoestasis.
Therefore, keen interest has emerged in RV due to its evident value as a cancer preventive and
cardioprotective dietary substance. RV may provide an alternative (and early) intervention approach
that could prevent/delay disease onset, emend the course of disease and/or prevent further damage.
Since the identification of RVs health benefits are largely owed to its high abundance in certain
plants and foods, the discovery of further naturally occurring stilbenes, as well as, chemically
modified analogs that are superior to RV in their cancer chemopreventive properties may be
expected.
This overview of molecular targets implicated in the antagonism of cancer by RV underscores the
complexity underlying biological responses to this drug, which is probably common to many other
molecules generated for self-defense. The efficacy of RV against distinct mechanisms of disease
development is an indicator of its potential value for the prevention of various human diseases.
Subsequently, the search and identification of more effective preventive agents among stilbene
natural products is warranted.
Table 1. Cancer-related targets, effects, and benefits of resveratrol.
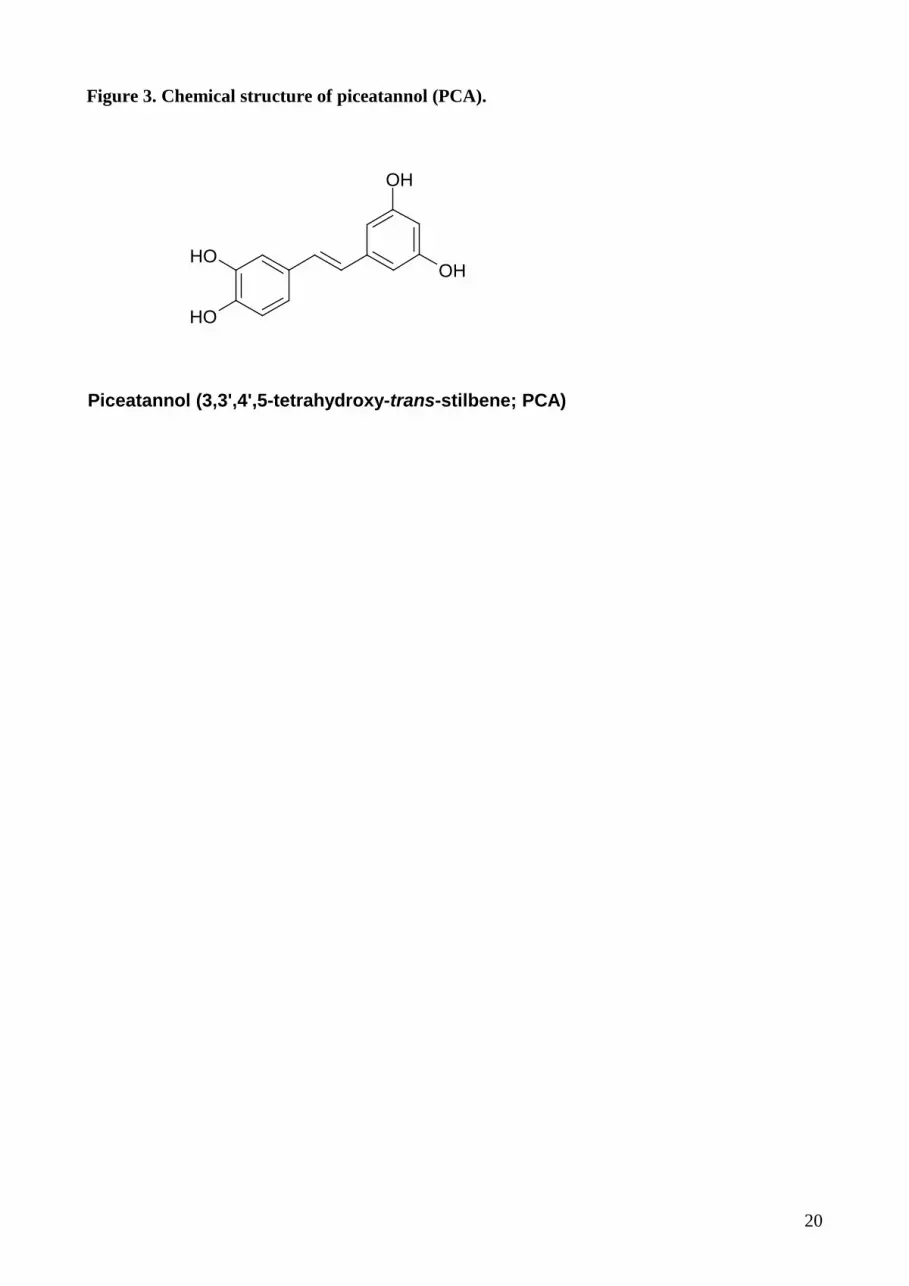
2.2. Piceatannol – a naturally occurring resveratrol metabolite
In contrast to the detailed knowledge of RV activities in biological systems, little is known about
the effects of other polyhydroxylated stilbens. RV undergoes cytochrome P450 catalyzed
hydroxylation to piceatannol (3,3’,4’,5-tetrahydroxy-trans-stilbene; PCA; Fig. 3) and two other

9
unidentified mono- and dihydroxy-RV analogs. This demonstrates that a natural dietary cancer
preventative agent can be converted to a compound with known chemopreventive and anticancer
activity by the enzyme CYP1B1, which is overexpressed in a wide variety of human tumors.
Importantly, these findings give insight into the functional role of the cytochrome P450 enzyme
CYP1B1 and provide evidence for the concept that CYP1B1 in tumors might be serving as a
growth suppressor enzyme [23]. Comparable with RV, PCA displays cytotoxic activity in acute
leukemia and lymphoma cells and exerts anti-proliferative activity in colon cancer cells [24].
Figure 3. Chemical structure of piceatannol (PCA).
2.3. Gallic acid
Gallic acid (3,4,5-trihydroxybenzoic acid; GA; Fig. 4) is found in gallnuts, sumac, tea leaves, oak
bark, grapes, various herbs, and in red and white wines. In particular, red wine has a high content of
this phenolic acid. GA can be present as free molecule or as part of the tannin molecule
(gallotannin). It was recently shown that GA antagonizes P-selectin mediated platelet leucocyte
interactions [25] and could be jointly responsible for the beneficial effects of red wine and the
“French paradox”. Other beneficial effects might be the antidiabetic and antiangiogenic effects of
GA containing fruit extracts [26] and the induction of Ca2+ dependent apoptosis in leukemia cells
[27]. Altogether, GA was described as an excellent free radical scavenger and as inducer of
differentiation as well as programmed cell death in numerous tumor cell lines and might play an
important role in the prevention of malignant transformation and cancer development.
Figure 4. Chemical structure of gallic acid (GA).
3. BIOAVAILABILITY AND METABOLISM OF ACTIVE COMPOUNDS
3.1. Bioavailability of resveratrol

10
Several in vivo studies in animals and humans demonstrated a very low intestinal uptake of RV
leading to trace amounts in the bloodstream based on extensive metabolism in the gut and liver.
Rapid metabolism is also the main reason for the short initial half-life of the primary molecule (~8-
14 min) [28]. The bulk of an intravenous dose of RV is converted to sulfate conjugates within ~30
min in humans. A detailed analysis of plasma metabolites after oral dosing was not possible;
however, both sulfate and glucuronide conjugates were detected [29].
3.2. Metabolites of resveratrol – glucuronide and sulfate conjugates
Although modifications such as glucuronidation and sulfation typically reduce the cell permeability
of drugs and aid in their excretion, the undeniable in vivo efficacy of administered RV, despite its
low bioavailability, has led to the suggestion that its metabolites are likely to be the active principle.
However, resveratrol-3-sulfate fails to inhibit CYPs [30] and there is currently no evidence that any
metabolite is able to cross the plasma membrane.
Recently, the absorptive efficiency of three polyphenolic constituents (trans-resveratrol, +-catechin
and quercetin) was investigated after oral application to healthy human subjects in three different
media (white wine, grape juice, and vegetable juice/homogenate) [31]. All compounds were present
in serum and urine predominantly as glucuronide and sulfate conjugates, reaching peak
concentrations in the former around 30 min after consumption. The absorption of these three
polyphenols was broadly equivalent in aqueous and alcoholic matrices; however, their peak plasma
concentrations reached only 10-40nM, whereas in vitro biologic activities have been studied at 5-
100µM [31].
3.3. Bioavailability of resveratrol in grape juice compared to its pure aglycone
In grape juice, the level of free RV is rather low, and cis- and trans-Piceid (RV-3-O-β-D-glucoside;
Polydatin) are the major RV derivatives. This suggests a lower bioavailability of RV glycosides in
grape juice in comparison to its pure aglycone in wine [32]. These findings were confirmed by

11
another study reporting that RV concentrations in Italian red wine ranged from 8.6 to 24.8µM,
whereas the amount in grape juice was only 1.6µM, respectively [33].
Given that in vivo concentrations of individual metabolites can be much higher than those of the
native compound, further studies are needed to determine (1) whether the metabolites represent
inactivated forms of the drug, (2) act as a pool from which free RV can be released in various
tissues, or (3) are themselves active in promoting many of the health benefits attributed to RV.
4. MECHANISMS OF PROTECTION
4.1. Results of in vitro studies
A large number of RV’s beneficial health effects, such as anticancer, antiviral, neuroprotective,
anti-aging, and anti-inflammatory effects, have been reported in vitro. Table 2 summarizes a variety
of RV’s anticancer activities.
Table 2. Results of in vitro studies.
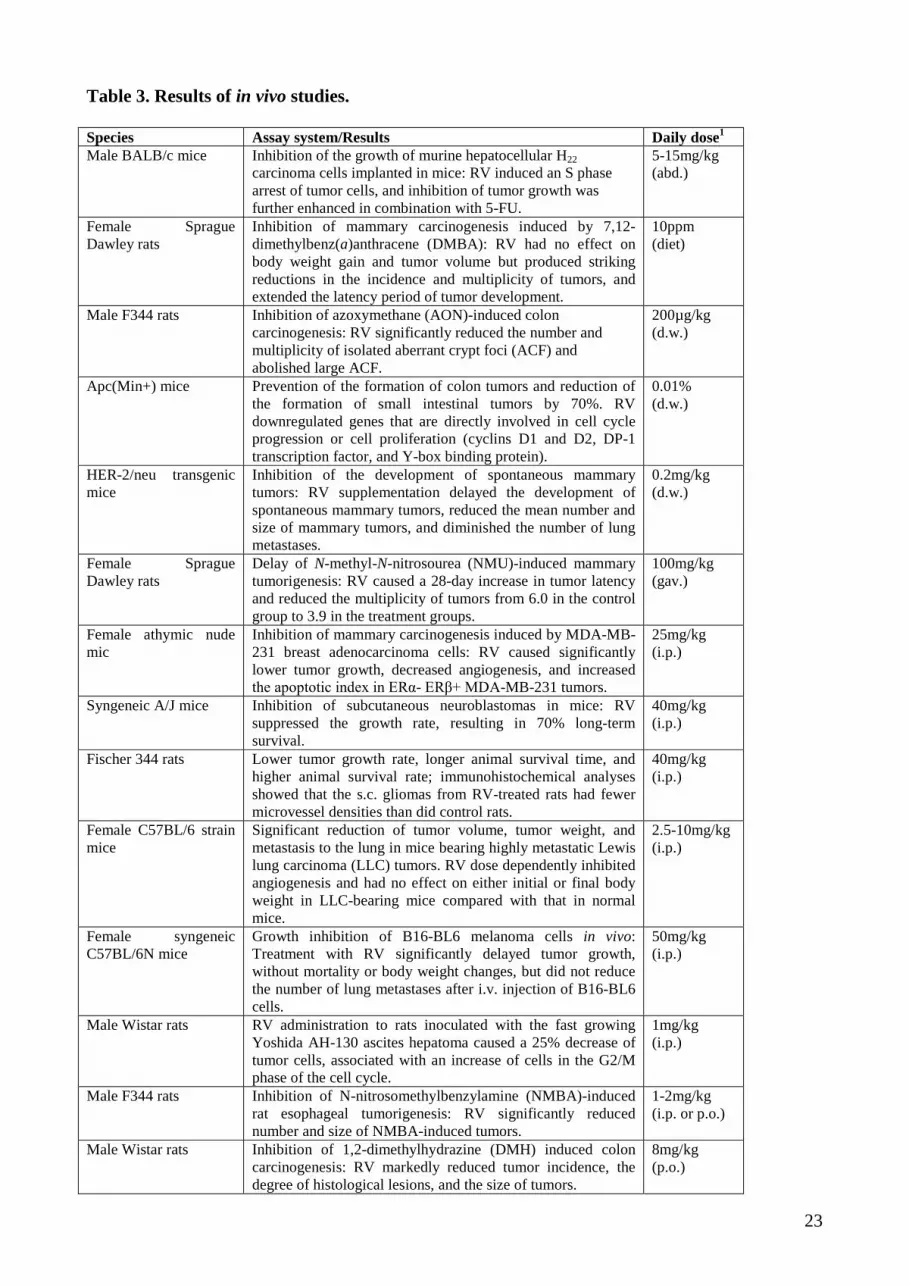
4.2. Results of in vivo studies
As the mechanisms of RV's broad cancer chemopreventive effects are not completely understood,
continued efforts are needed, especially well designed pre-clinical studies in animal models that
closely mimic/represent human disease, to establish the usefulness of RV as cancer
chemopreventive agent. Table 3 gives an overview of such pre-clinical studies including the
observed effects.
Table 3. Results of in vivo studies.
5. RESULTS OF HUMAN STUDIES

12
Overall, in vivo studies in rodents clearly show great promise for RV in the prevention and
treatment of cancers. However, only a few Phase I clinical trials are currently underway for oral RV
in humans at doses up to 7.5 g per day. A National Cancer Institute-sponsored study has been
completed recently [34], suggesting that consumption of RV does not cause serious adverse events.
RV and six metabolites were recovered from plasma and urine, among them two monoglucuronides
and RV-3-sulfate. The area under the plasma concentration curve (AUC) values for these
metabolites were up to 23 times greater than those of RV [34]. Cancer chemopreventive effects of
RV in cells in vitro require levels of at least 5 mM per litre, intimating that consumption of high-
dose RV might be insufficient to elicit systemic levels synonymous with cancer chemopreventive
efficacy. On the other hand, the high systemic levels of RV metabolites clearly suggest that they
might be the active principle of the parent drug.
6. CONCLUSIONS
Chemopreventive agents such as RV might be used not only to prevent, but also to treat cancer
since the molecular targets are similar. Due of their pharmacological safety, RV and its
metabolites/analogs may be applied in combination with other chemotherapeutic agents in order to
enhance their efficacy thus minimizing chemotherapy-induced toxicity. By inhibition of COX and
cytochrome P450 enzymes and by induction of quinone reductase, RV can simultaneously inhibit
promutagen bioactivation, stimulate carcinogen detoxification and protect the organism against the
adverse effects of diverse environmental toxins. Furthermore, RV inhibits prostaglandins, NO
formation and the generation of ROS, therefore preventing their stimulative effect on tumor
development. Besides acting as a chemopreventive agent, several in vitro, ex vivo, and in vivo
experiments have shown that RV suppresses the growth of various cancer cell lines by inhibition of
DNA polymerases, RR, and by inducing cell cycle arrest or apoptosis. Although the most exciting
in vivo data relate to its cancer chemopreventive and chemotherapeutic activity, some studies also
demonstrate beneficial effects on cardiovascular, neurological, and hepatic systems.

13
RV is commonly referred to as a “dirty” molecule, meaning that it seems to interact with many
different proteins, including cyclooxygenases, ribonucleotide reductase and DNA polymerases.
Thus, its activity cannot be resumed in a unique mechanism of action but likely results from various
complementary actions of different biochemical pathways.
However, the question of whether RV itself can accumulate to bioactive levels in target organs
remains to be addressed. Opposing results and controversies involving the available data are leading
to the suggestion that RV metabolites might be the active principle, and call for additional
experiments. Long-term in vivo epidemiological studies are highly warranted to determine the
preventive and therapeutic efficacy of dietary or supplemented RV on tumor development and
progression.

14
References
1. McGovern, P. E., Glusker, D. L., Exner, L. J., and Voigt, M. M. (1996) Neolithic resinated wine, Nature 381, 480-481.
2. Soleas, G. J., Diamandis, E. P., and Goldberg, D. M. (1997) Wine as a biological fluid: history, production, and role in disease prevention, Journal of clinical laboratory analysis 11, 287-313.
3. Renaud, S., and de Lorgeril, M. (1992) Wine, alcohol, platelets, and the French paradox for coronary heart disease, Lancet 339, 1523-1526.
4. Baur, J. A., and Sinclair, D. A. (2006) Therapeutic potential of resveratrol: the in vivo evidence, Nat Rev Drug Discov 5, 493-506.
5. Nonomura, S., Kanagawa, H., and Makimoto, A. (1963) [Chemical Constituents Of Polygonaceous Plants. I. Studies On The Components Of Ko-J O-Kon. (Polygonum Cuspidatum Sieb. Et Zucc.)], Yakugaku Zasshi 83, 988-990.
6. Siemann, E. H., L.L. Creasy. (1992) Concentration of phytoalexin resveratrol in wine, Am. J. Enol. Vitic. 43, 49-52.
7. Gusman, J., Malonne, H., and Atassi, G. (2001) A reappraisal of the potential chemopreventive and chemotherapeutic properties of resveratrol, Carcinogenesis 22, 1111-1117.
8. Hao, H. D., and He, L. R. (2004) Mechanisms of cardiovascular protection by resveratrol, J Med Food 7, 290-298.
9. Saiko, P., Szakmary, A., Jaeger, W., and Szekeres, T. (2008) Resveratrol and its analogs: defense against cancer, coronary disease and neurodegenerative maladies or just a fad?, Mutation research 658, 68-94.
10. Dong, Z. (2003) Molecular mechanism of the chemopreventive effect of resveratrol, Mutation research 523-524, 145-150.
11. Aggarwal, B. B., and Shishodia, S. (2006) Molecular targets of dietary agents for prevention and therapy of cancer, Biochemical pharmacology 71, 1397-1421.
12. Stewart, J. R., Christman, K. L., and O'Brian, C. A. (2000) Effects of resveratrol on the autophosphorylation of phorbol ester-responsive protein kinases: inhibition of protein kinase D but not protein kinase C isozyme autophosphorylation, Biochemical pharmacology 60, 1355-1359.
13. Bhat, K. P., Lantvit, D., Christov, K., Mehta, R. G., Moon, R. C., and Pezzuto, J. M. (2001) Estrogenic and antiestrogenic properties of resveratrol in mammary tumor models, Cancer research 61, 7456-7463.
14. Jang, M., Cai, L., Udeani, G. O., Slowing, K. V., Thomas, C. F., Beecher, C. W., Fong, H. H., Farnsworth, N. R., Kinghorn, A. D., Mehta, R. G., Moon, R. C., and Pezzuto, J. M. (1997) Cancer chemopreventive activity of resveratrol, a natural product derived from grapes, Science (New York, N.Y 275, 218-220.

15
15. Fontecave, M., Lepoivre, M., Elleingand, E., Gerez, C., and Guittet, O. (1998) Resveratrol, a remarkable inhibitor of ribonucleotide reductase, FEBS letters 421, 277-279.
16. Locatelli, G. A., Savio, M., Forti, L., Shevelev, I., Ramadan, K., Stivala, L. A., Vannini, V., Hubscher, U., Spadari, S., and Maga, G. (2005) Inhibition of mammalian DNA polymerases by resveratrol: mechanism and structural determinants, Biochem J 389, 259-268.
17. Ragione, F. D., Cucciolla, V., Borriello, A., Pietra, V. D., Racioppi, L., Soldati, G., Manna, C., Galletti, P., and Zappia, V. (1998) Resveratrol arrests the cell division cycle at S/G2 phase transition, Biochem Biophys Res Commun 250, 53-58.
18. Horvath, Z., Saiko, P., Illmer, C., Madlener, S., Hoechtl, T., Bauer, W., Erker, T., Jaeger, W., Fritzer-Szekeres, M., and Szekeres, T. (2005) Synergistic action of resveratrol, an ingredient of wine, with Ara-C and tiazofurin in HL-60 human promyelocytic leukemia cells, Experimental hematology 33, 329-335.
19. Stewart, J. R., Ward, N. E., Ioannides, C. G., and O'Brian, C. A. (1999) Resveratrol preferentially inhibits protein kinase C-catalyzed phosphorylation of a cofactor-independent, arginine-rich protein substrate by a novel mechanism, Biochemistry 38, 13244-13251.
20. Subbaramaiah, K., Chung, W. J., Michaluart, P., Telang, N., Tanabe, T., Inoue, H., Jang, M., Pezzuto, J. M., and Dannenberg, A. J. (1998) Resveratrol inhibits cyclooxygenase-2 transcription and activity in phorbol ester-treated human mammary epithelial cells, The Journal of biological chemistry 273, 21875-21882.
21. MacCarrone, M., Lorenzon, T., Guerrieri, P., and Agro, A. F. (1999) Resveratrol prevents apoptosis in K562 cells by inhibiting lipoxygenase and cyclooxygenase activity, European journal of biochemistry / FEBS 265, 27-34.
22. Steele, V. E., Holmes, C. A., Hawk, E. T., Kopelovich, L., Lubet, R. A., Crowell, J. A., Sigman, C. C., and Kelloff, G. J. (2000) Potential use of lipoxygenase inhibitors for cancer chemoprevention, Expert opinion on investigational drugs 9, 2121-2138.
23. Potter, G. A., Patterson, L. H., Wanogho, E., Perry, P. J., Butler, P. C., Ijaz, T., Ruparelia, K. C., Lamb, J. H., Farmer, P. B., Stanley, L. A., and Burke, M. D. (2002) The cancer preventative agent resveratrol is converted to the anticancer agent piceatannol by the cytochrome P450 enzyme CYP1B1, Br J Cancer 86, 774-778.
24. Wolter, F., Clausnitzer, A., Akoglu, B., and Stein, J. (2002) Piceatannol, a natural analog of resveratrol, inhibits progression through the S phase of the cell cycle in colorectal cancer cell lines, J Nutr 132, 298-302.
25. Appeldoorn, C. C., Bonnefoy, A., Lutters, B. C., Daenens, K., van Berkel, T. J., Hoylaerts, M. F., and Biessen, E. A. (2005) Gallic acid antagonizes P-selectin-mediated platelet-leukocyte interactions: implications for the French paradox, Circulation 111, 106-112.
26. Liu, Z., Schwimer, J., Liu, D., Greenway, F. L., Anthony, C. T., and Woltering, E. A. (2005) Black raspberry extract and fractions contain angiogenesis inhibitors, Journal of agricultural and food chemistry 53, 3909-3915.
27. Isuzugawa, K., Inoue, M., and Ogihara, Y. (2001) Ca2+-Dependent caspase activation by gallic acid derivatives, Biological & pharmaceutical bulletin 24, 844-847.

16
28. Marier, J. F., Vachon, P., Gritsas, A., Zhang, J., Moreau, J. P., and Ducharme, M. P. (2002) Metabolism and disposition of resveratrol in rats: extent of absorption, glucuronidation, and enterohepatic recirculation evidenced by a linked-rat model, The Journal of pharmacology and experimental therapeutics 302, 369-373.
29. Walle, T., Hsieh, F., DeLegge, M. H., Oatis, J. E., Jr., and Walle, U. K. (2004) High absorption but very low bioavailability of oral resveratrol in humans, Drug Metab Dispos 32, 1377-1382.
30. Yu, C., Shin, Y. G., Kosmeder, J. W., Pezzuto, J. M., and van Breemen, R. B. (2003) Liquid chromatography/tandem mass spectrometric determination of inhibition of human cytochrome P450 isozymes by resveratrol and resveratrol-3-sulfate, Rapid Commun Mass Spectrom 17, 307-313.
31. Goldberg, D. M., Yan, J., and Soleas, G. J. (2003) Absorption of three wine-related polyphenols in three different matrices by healthy subjects, Clinical biochemistry 36, 79-87.
32. Meng, X., Maliakal, P., Lu, H., Lee, M. J., and Yang, C. S. (2004) Urinary and plasma levels of resveratrol and quercetin in humans, mice, and rats after ingestion of pure compounds and grape juice, J Agric Food Chem 52, 935-942.
33. Wang, Y., Catana, F., Yang, Y., Roderick, R., and van Breemen, R. B. (2002) An LC-MS method for analyzing total resveratrol in grape juice, cranberry juice, and in wine, J Agric Food Chem 50, 431-435.
34. Boocock, D. J., Faust, G. E., Patel, K. R., Schinas, A. M., Brown, V. A., Ducharme, M. P., Booth, T. D., Crowell, J. A., Perloff, M., Gescher, A. J., Steward, W. P., and Brenner, D. E. (2007) Phase I dose escalation pharmacokinetic study in healthy volunteers of resveratrol, a potential cancer chemopreventive agent, Cancer Epidemiol Biomarkers Prev 16, 1246-1252.

17
Figure 1. Chemical structure of resveratrol (RV).
OH
OH
OH
Resveratrol (3,4',5-trihydroxy-trans-stilbene; RV)
18
Figure 2. Effects of resveratrol on arachidonic acid metabolism.
RV RVRV
RV
RV
ARACHIDONIC ACID
PHOSPHOLIPIDS
CYCLOOXYGENASES(COX-1, COX-2)
LIPOXYGENASES EPOXYGENASES(CYP450)
FREE RADICALS
ISOPROSTANESEPOXIDESHPETEsLEUKOTRIENESLIPOXINS
PROSTAGLANDINSTHROMBOXANES
PHOSPHOLIPASES

19
Table 1. Cancer-related targets and biological effects of resveratrol.
CLASS OF TARGETS MOLECULAR TARGETS BIOLOGICAL EFFECTS
Direct radical scavenging ROS DNA stability Lipidoxidation Apoptosis/cell survival
Antioxidant/phase II enzymes
SOD, catalase, GR DNA stability Lipidoxidation Apoptosis/cell survival
Heme oxidase 1 (HO-1) Radical scavenging Anti-inflammatory Anti-apoptotic
GST, NQO1, UDP-glucuronyl transferase
ARE/EpRE activation ERβ-activated
Arachidonic acid related COX 1+2, (COX 2 via NF-κB) Anti-inflammatory Anti-tumor promoting
Estrogen-related
Selective estrogen receptor modulation (SERM)
Superagonist Agonist Antagonist
CYP 1A1, CYP 1B1 Inhibition of estrogen-metabolizing phase I enzymes
Modulation of signalling kinases Raf, Src, MAPK, PKD, PKCδ
Cell growth arrest Cell death Differentiation
Modulation of global gene expression through chromatin remodelling
p300 (Acetylase), SIRT1 (Deacetylase) Cell survival Apoptosis delay Inflammatory response
Transcription factors p53/p21 Cell cycle arrest
Apoptosis IκB kinase/ NF-κB Cell survival AP1 Tumor growth promoter
Other cell cycle related
Ribonucleotide reductase, replicative DNA polymerases
DNA synthesis Cell cycle arrest
Survivin Cell survival Apoptosis
Cell death related
TRAIL/DR4+5 Apoptosis
Fas/CD95 Apoptosis Mitochondria-dependent, cytochrome c, Apaf-1 Apoptosis
PI3K/Akt, ERα-dependent Cell survival
Ceramide Apoptosis
20
Figure 3. Chemical structure of piceatannol (PCA).
OH
OHOH
OH
Piceatannol (3,3',4',5-tetrahydroxy-trans-stilbene; PCA)

21
Figure 4. Chemical structure of gallic acid (GA).
OH
OHOH
OH
O
Gallic acid (3,4,5-trihydroxybenzoic acid; GA)

22
Table 2. Results of in vitro studies. Mechanism Assay system µM1 Estrogenic/antiestrogenic and scavenging properties
MCF-7 and MVLN breast cancer cells 0.1-25
Inhibition of aryl hydrocarbon-induced cytochrome P-450 1A1 enzyme activity and CYP1A1 expression
HepG2 hepatoma cells and MCF-7 breast cancer cells
0.5-5
Modulation of the catalytic activity and mRNA expression of the procarcinogen-activating human cytochrome P450 1B1
Cultured MCF-7 human breast carcinoma cells
1-20
Inhibition of cell growth, G1-phase arrest, and induction of apoptosis
Human A431 epidermoid carcinoma cells
1-50
Inhibition of cyclooxygenase-2 transcription and activity in phorbol ester-treated human mammary epithelial cells
Human mammary 184B5/HER epithelial cells (and premalignant MSK Leuk1 oral epithelial cells)
2.5-40
Suppression of TNF-induced activation of nuclear transcription factor NF-κB, activator protein-1, and apoptosis
U937 myeloid leukemia cells, Jurkat lymphoid and HeLa and H4 epithelial cells
5
Inhibition of the binding of labeled estradiol to the estrogen receptor
MCF-7 breast cancer cells 10
Stimulation of the proliferation of estrogen-dependent breast cancer cells
T47D breast cancer cells 10
Growth inhibition, induction of apoptosis by down-regulation of Bcl-2 and up-regulation of bax
EC-9706 esophageal cancer cells 10
Modulation of ERE-Luciferase Activity and Estrogen-inducible Protein Expression
MCF-7, T47D, LY2, and S30 breast cancer cells
10-15
Inhibition of the clonal growth of tumor cells 32Dp210, HL-60, U937, and L1210 leukemia cells
10-80
Depletion of intracellular dCTP, dTTP, dATP, and dGTP pools (inhibition of ribonucleotide reductase)
HL-60 leukemia cells 12.5
Inhibition of 14C-cytidine incorporation into DNA (inhibition of ribonucleotide reductase)
HL-60 leukemia cells 12.5
Growth inhibition, accumulation of cells at the S/G2 phase transition of the cell cycle, and significant decrease of ornithine decarboxylase (ODC) activity
CaCo-2 colon cancer cells 25
Cell division cycle arrest at S/G2 phase transition
HL-60 leukemia cells 30
Increase in the content of cyclins A and B1 as well as cyclin-dependent kinases Cdk1 and Cdk2, and promotion of Cdk1 phosphorylation
SW480 colon cancer cells 30
Increase in the expression and kinase activities of positive G1/S and G2/M regulators, resulting in cell cycle blockade at the S-phase and apoptosis induction
MCF-7 breast cancer cells < 50
Inhibition of the expression and function of the androgen receptor
LNCaP prostate cancer cells 50-100
Decrease of the expression and kinase activities of positive G1/S and G2/M cell cycle regulators and inhibition of ribonucleotide reductase activity
MDA-MB-231 breast cancer cells < 200
1Lowest concentration at which reproducible changes have been observed, or IC50 or EC50, if provided.
23
Table 3. Results of in vivo studies. Species Assay system/Results Daily dose1 Male BALB/c mice Inhibition of the growth of murine hepatocellular H22
carcinoma cells implanted in mice: RV induced an S phase arrest of tumor cells, and inhibition of tumor growth was further enhanced in combination with 5-FU.
5-15mg/kg (abd.)
Female Sprague Dawley rats
Inhibition of mammary carcinogenesis induced by 7,12-dimethylbenz(a)anthracene (DMBA): RV had no effect on body weight gain and tumor volume but produced striking reductions in the incidence and multiplicity of tumors, and extended the latency period of tumor development.
10ppm (diet)
Male F344 rats Inhibition of azoxymethane (AON)-induced colon carcinogenesis: RV significantly reduced the number and multiplicity of isolated aberrant crypt foci (ACF) and abolished large ACF.
200µg/kg (d.w.)
Apc(Min+) mice Prevention of the formation of colon tumors and reduction of the formation of small intestinal tumors by 70%. RV downregulated genes that are directly involved in cell cycle progression or cell proliferation (cyclins D1 and D2, DP-1 transcription factor, and Y-box binding protein).
0.01% (d.w.)
HER-2/neu transgenic mice
Inhibition of the development of spontaneous mammary tumors: RV supplementation delayed the development of spontaneous mammary tumors, reduced the mean number and size of mammary tumors, and diminished the number of lung metastases.
0.2mg/kg (d.w.)
Female Sprague Dawley rats
Delay of N-methyl-N-nitrosourea (NMU)-induced mammary tumorigenesis: RV caused a 28-day increase in tumor latency and reduced the multiplicity of tumors from 6.0 in the control group to 3.9 in the treatment groups.
100mg/kg (gav.)
Female athymic nude mic
Inhibition of mammary carcinogenesis induced by MDA-MB-231 breast adenocarcinoma cells: RV caused significantly lower tumor growth, decreased angiogenesis, and increased the apoptotic index in ERα- ERβ+ MDA-MB-231 tumors.
25mg/kg (i.p.)
Syngeneic A/J mice Inhibition of subcutaneous neuroblastomas in mice: RV suppressed the growth rate, resulting in 70% long-term survival.
40mg/kg (i.p.)
Fischer 344 rats Lower tumor growth rate, longer animal survival time, and higher animal survival rate; immunohistochemical analyses showed that the s.c. gliomas from RV-treated rats had fewer microvessel densities than did control rats.
40mg/kg (i.p.)
Female C57BL/6 strain mice
Significant reduction of tumor volume, tumor weight, and metastasis to the lung in mice bearing highly metastatic Lewis lung carcinoma (LLC) tumors. RV dose dependently inhibited angiogenesis and had no effect on either initial or final body weight in LLC-bearing mice compared with that in normal mice.
2.5-10mg/kg (i.p.)
Female syngeneic C57BL/6N mice
Growth inhibition of B16-BL6 melanoma cells in vivo: Treatment with RV significantly delayed tumor growth, without mortality or body weight changes, but did not reduce the number of lung metastases after i.v. injection of B16-BL6 cells.
50mg/kg (i.p.)
Male Wistar rats RV administration to rats inoculated with the fast growing Yoshida AH-130 ascites hepatoma caused a 25% decrease of tumor cells, associated with an increase of cells in the G2/M phase of the cell cycle.
1mg/kg (i.p.)
Male F344 rats Inhibition of N-nitrosomethylbenzylamine (NMBA)-induced rat esophageal tumorigenesis: RV significantly reduced number and size of NMBA-induced tumors.
1-2mg/kg (i.p. or p.o.)
Male Wistar rats Inhibition of 1,2-dimethylhydrazine (DMH) induced colon carcinogenesis: RV markedly reduced tumor incidence, the degree of histological lesions, and the size of tumors.
8mg/kg (p.o.)

24
C57Bl6/J mice Treatment with RV significantly inhibited the growth of murine T241 fibrosarcoma in mice and suppressed FGF-2- and VEGF-stimulated angiogenesis.
1mg/kg (p.o.)
Nude mice Inhibition of implanted human primary gastric carcinoma cells: RV could significantly inhibit carcinoma growth when it was injected near the carcinoma and induced implanted tumor cells to undergo apoptosis.
500-1500mg/kg (s.c.)
Female SKH-1 hairless mice
Topical application of skin with RV (both pre- and post- treatment) resulted in a highly significant inhibition in tumor incidence and in a delay in the onset of tumorigenesis.
25-50µM (top.)
1Lowest concentration at which reproducible changes have been observed, or IC50 or EC50, if provided. abd. – abdominal injection, d.w. – drinking water, gav. – gavage, i.p. – intraperitoneal, p.o. – per os, s.c. – subcutaneous injection, top. – topical application.