promoter recognition and discrimination by esigmas rna polymerase
TRANSCRIPT

Promoter recognition and discrimination by EsS RNApolymerase
Tamas Gaal,1 Wilma Ross,1 Shawn T. Estrem,1 Lam H.
Nguyen,2 Richard R. Burgess2 and Richard L.
Gourse1*1Department of Bacteriology, University of Wisconsin,
Madison WI 53706, USA.2Department of Oncology, University of Wisconsin,
McArdle Lab, Madison WI 53706, USA.
Summary
Although more than 30 Escherichia coli promoters
utilize the RNA polymerase holoenzyme containing sS
(EsS), and it is known that there is some overlap
between the promoters recognized by EsS and by the
major E. coli holoenzyme (Es70), the sequence
elements responsible for promoter recognition by
EsS are not well understood. To define the DNA
sequences recognized best by EsS in vitro, we started
with random DNA and enriched for EsS promoter
sequences by multiple cycles of binding and selec-
tion. Surprisingly, the sequences selected by EsS
contained the known consensus elements (210 and
235 hexamers) for recognition by Es70. Using genetic
and biochemical approaches, we show that EsS and
Es70 do not achieve specificity through ‘best fit’ to
different consensus promoter hexamers, the way that
other forms of holoenzyme limit transcription to
discrete sets of promoters. Rather, we suggest that
EsS-specific promoters have sequences that differ
significantly from the consensus in at least one of the
recognition hexamers, and that promoter discrimi-
nation against Es70 is achieved, at least in part, by the
two enzymes tolerating different deviations from
consensus. DNA recognition by EsS versus Es70
thus presents an alternative solution to the problem of
promoter selectivity.
Introduction
Escherichia coli RNA polymerase (RNAP) consists of b,
b’, s, v and two a subunits. The core enzyme (a2bb’v) is
capable of transcription elongation, whereas s is required
for specific promoter recognition and transcription
initiation. Escherichia coli has seven different s subunits:
s70 (encoded by rpoD ), s54 (rpoN ), s32 (rpoH ), sS (rpoS ),
sE (rpoE ), s28 (rpoF ) and sFecI ( fecI ), each responsible
for directing RNAP to a different set of promoters, and, in
so doing, changing the pattern of gene expression (Gross
et al., 1998; Ishihama, 2000).
Es70, the holoenzyme responsible for transcription of
the majority of genes in E. coli, recognizes three DNA
sequence elements (all sequences provided below are for
the non-template strand, 50 to 30). The 210 element,
consensus TATAAT, is centred approximately 10 bp
preceding the transcription start site and is recognized
by regions 2.3 and 2.4 of s70. The 235 element,
consensus TTGACA, is 16–19 bp upstream from the
210 element and interacts with region 4.2 of s70 (Gross
et al., 1998). The UP element, an AT-rich region upstream
of the 235 hexamer, is recognized by the C-terminal
domains of the a subunits (Ross et al., 1993; Blatter et al.,
1994). In addition, some Es70 promoters contain an
extension of the 210 hexamer, TGTGn, immediately
upstream of the 210 motif, that is recognized by region 2.5
of s70 (Barne et al., 1997; Burr et al., 2000). Generally, the
more of these elements present in a particular promoter
and the more similar their sequences are to the consensus
sequences, the better the binding by RNAP (Gross et al.,
1998; Ross et al., 1998).
The synthesis, accumulation, and activity of sS (also
referred to as s38 or KatF) are controlled by a number of
positive and negative regulatory systems (Hengge-Aronis,
1996a; Ishihama, 2000). As a result, sS accumulates in the
cell at the beginning of stationary phase and directs RNAP
to transcribe stationary phase-specific genes (Mulvey and
Loewen, 1989; Nguyen et al., 1993). In addition, the
presence of specific transcription factors, high intracellular
osmolarity, and/or negative supercoiling can favour
transcription by EsS at certain promoters (Hengge-Aronis
et al., 1991; Kusano et al., 1996; Hengge-Aronis, 1996b;
Ishihama, 2000). As sS is involved in responses to osmotic
and oxidative stress, it is sometimes considered a master
regulator of stress responses (Hengge-Aronis, 1996b).
EsS is also required for virulence in a number of
pathogenic bacteria (Beltrametti et al., 1999; Suh et al.,
1999).
Although promoters recognized by EsS have been
compiled, they do not display a distinctive consensus
sequence (Espinosa-Urgel et al., 1996; Ishihama, 2000).
It has been suggested that EsS recognizes 210
elements similar, but not identical, to the consensusAccepted 6 September, 2000. *For correspondence. E-mail [email protected]; Tel. (11) 608 262 9813; Fax (11) 608 262 9865.
Molecular Microbiology (2001) 42(4), 939–954
Q 2001 Blackwell Science Ltd

recognized by Es70 (Espinosa-Urgel et al., 1996; Lee
and Gralla, 2001). It has also been suggested that a
235 consensus sequence for EsS, if it exists at all, is
different from the consensus element utilized by Es70,
for example, curved DNA with no specific 235 sequence
(Espinosa-Urgel et al., 1996), CC instead of TT at the
upstream end of the 235 element (Wise et al., 1996) or
CTGCAA as a 235 element (Bohannon et al., 1991;
Vicente et al., 1991; Ballesteros et al., 1998). Therefore,
the consensus DNA sequence for recognition by EsS is
still unclear.
We chose an in vitro approach to determine the
sequences recognized best by EsS, reasoning that
understanding the biochemical behaviour of the enzyme
in the absence of other factors, and then integrating these
data with the extensive information about EsS obtained in
vivo, might facilitate understanding of the behaviour of the
enzyme in the complex environment of the cell. We
generated a DNA library containing every possible
sequence variant in the s recognition regions and used
EsS to select those sequences that bound preferentially in
vitro. The 210 and 235 hexamers obtained were identical
to those recognized by Es70, raising an interesting
problem about promoter selectivity of the two holoen-
zymes. Based on genetic and biochemical approaches,
we show that the two forms of holoenzyme prefer the same
consensus hexamers for transcription, and that specificity
can be achieved by differential tolerance of the enzyme for
deviations from the consensus. This model is consistent
with the observation that many stationary phase-specific
genes are transcribed in vitro by both Es70 and EsS
(Tanaka et al., 1993; Tanaka et al., 1995), and that sS
shows considerable sequence homology to s70 in the
regions responsible for specific promoter recognition, 2.3,
2.4, 2.5 and 4.2 (Lonetto et al., 1992), but it differs
conceptually from the standard model for promoter
recognition in which each holoenzyme has its own discrete
DNA recognition elements responsible for transcribing
discrete sets of promoters.
Results
Rationale and experimental design
Footprinting experiments with an EsS-specific promoter,
bolA1, indicated previously that EsS protects about 70 bp
of promoter sequence (from about 257 to 117), similar in
length and position to that protected by Es70 (Nguyen and
Burgess, 1997). Based on this, and on the similarity in the
amino acid sequences of the two s factors in the regions
responsible for DNA binding (regions 2.3–2.5, and 4.2),
we decided to sample a population of DNA sequences
containing all possible sequences from 238 to 12.
The number of potential variants for 40 bp of DNA is
about 440 (¼1024) different DNA sequences, far too many
to test in vivo. Therefore, to test all possible recognition
sequences for optimal EsS binding, we used an in vitro
selection procedure with DNA fragments containing
randomized promoter sequences. Similar approaches
have been utilized previously to identify optimal binding
sequences for numerous nucleic acid binding proteins
(Blackwell and Weintraub, 1990; Pollock and Treisman,
1990; Tuerk and Gold, 1990; Wright et al., 1991; Estrem
et al., 1998; 1999).
One copy of each possible 40 bp sequence variant
would generate about 1 kg of DNA, far exceeding what can
be manipulated experimentally even in vitro. Therefore, we
performed the in vitro selection in two steps, randomizing a
20 bp segment to generate libraries containing at least one
copy of 420 (,1012) different sequences in each step. In
the first step, a selection was performed with promoter
fragments randomized from 218 to 12. This was followed
by a second selection with promoter fragments random-
ized from 238 to 219 in the context of the selected 218 to
12 sequence.
Highly purified EsS, containing no detectable s70 or
other s factors (Nguyen et al., 1993), was added to the
library of DNA fragments under limiting conditions so that
only 0.5–5% of the DNA fragments formed complexes
with RNAP. After separation of the complexes from free
DNA on a non-denaturing polyacrylamide gel, DNA from
the bound fraction was eluted, PCR-amplified and the
procedure was repeated for multiple cycles to enrich for
the best binding sequences (Fig. 1A). As EsS-DNA
complexes migrated to a location different from
Es70-DNA complexes (Fig. 1B), contamination with DNA
fragments bound by undetectable levels of Es70 (if
present) would be minimized at each cycle in the
enrichment. We note that our binding assay selects for
sequences resulting in the maximal rate of open complex
formation, but these sequences need not be optimal for
Fig. 1. A. Steps in the in vitro selection process. See text for details.B. Electrophoretic mobility-shift illustrating that EsS-promotercomplexes migrate to a different position than EsS-promotercomplexes containing the same 32P-labelled DNA fragment.
940 T. Gaal et al.
Q 2001 Blackwell Science Ltd, Molecular Microbiology, 42, 939–954

each step leading to individual intermediates in the
transcription initiation pathway.
Selection of the 210 promoter region by EsS
To identify EsS promoter determinants in the region
containing the 210 element, EsS was incubated with a
promoter fragment library containing randomized
sequences from 218 to 12 and bolA1 promoter
sequences from 254 to 219 and from 13 to 116 (210
rnd; Fig. 2A). After 17 cycles of selection by EsS, 16 DNA
fragments were cloned and sequenced (see Experimental
procedures ). A close match to the 11 bp sequence
TGTGCTATA(C/A)T was present on each of 12 promoter
fragments (Fig. 2B and C; the other four fragments
contained large deletions and probably were selected for
artifactual reasons described in Experimental pro-
cedures ). The 11-mer sequence (e.g. in 210 sel 1)
showed limited similarity to the bolA1 promoter but
contained the Es70-like 210 hexamers, TATAAT or
TATACT (Fig. 2). The 11-mer also contained the con-
sensus ‘extended 2100 motif TGTG, that is found
upstream of the 210 hexamer in a subset of E. coli
promoters (Burr et al., 2000), and a C between the
extended 210 motif and the 210 hexamer. The other 9 bp
of the randomized region differed among the selected
promoter fragments, indicating the promoters were of
independent origin.
The position of the selected 11-mer was relatively
constant within the randomized region (from 217 to 27 in
eight cases, from 216 to 26 in three cases, and from 218
to 28 in one case). These data suggest that sequences
outside the randomized region contribute to positioning
EsS on the DNA and, therefore, have an important role in
recognition by EsS.
Selection of the 235 promoter region by EsS
To identify promoter determinants for EsS binding
upstream of 218, a second selection was performed
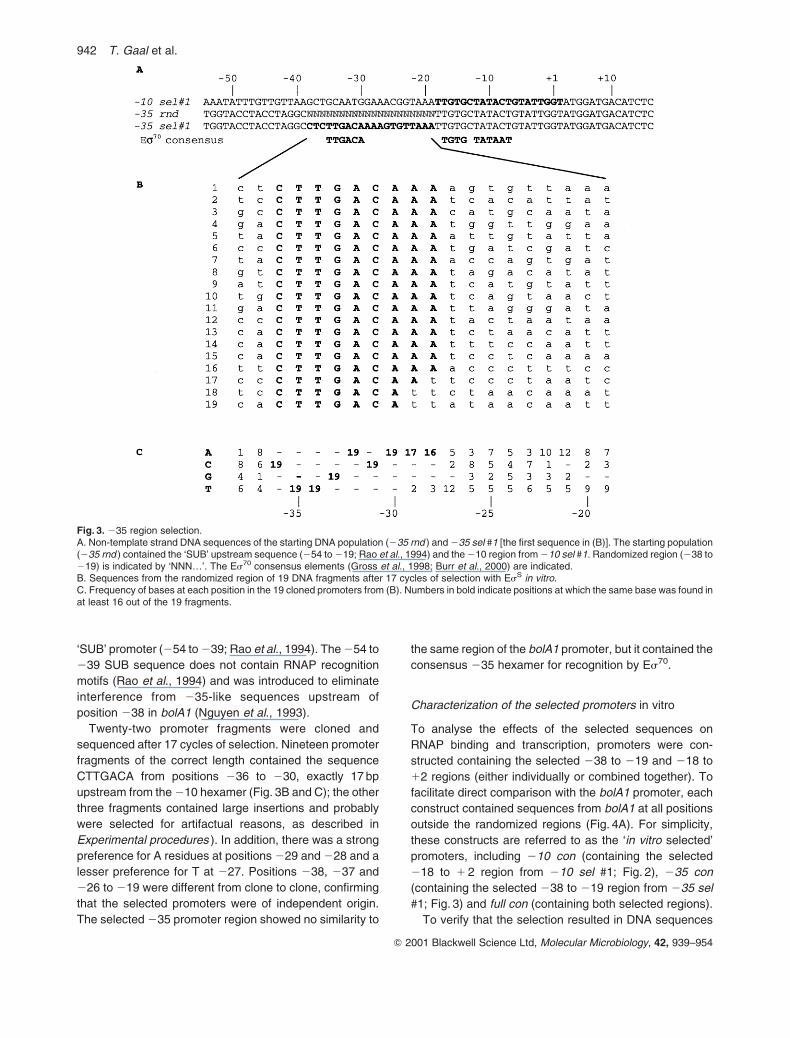
with a fragment library (235 rnd; Fig. 3A) containing
randomized basepairs from 238 to 219. Flanking
sequences in the starting population derived from the
210 sel #1 promoter (218 to 116; Fig. 2) and from the
Fig. 2. 210 region selection.A. Non-template strand DNA sequences of the bolA1 promoter, the starting DNA population (210 rnd ) and 210 sel #1 [the first sequence in (B)]. Therandomized region in 210 rnd (218 to 1 2) is indicated by ‘NNN…’. The Es70 consensus elements (Gross et al., 1998; Burr et al., 2000) areindicated.B. Sequences from the randomized region of 12 DNA fragments cloned after 17 cycles of selection with EsS in vitro. Sequences were aligned byidentities in the 210 region.C. Frequency of bases at each position in the 12 cloned promoters from (B). Numbers in bold indicate positions at which the same base was found inat least 10 out of the 12 promoters or at which either of two bases were found in all 12 promoters.
Promoter recognition and discrimination by Ess RNAP 941
Q 2001 Blackwell Science Ltd, Molecular Microbiology, 42, 939–954
‘SUB’ promoter (254 to 239; Rao et al., 1994). The 254 to
239 SUB sequence does not contain RNAP recognition
motifs (Rao et al., 1994) and was introduced to eliminate
interference from 235-like sequences upstream of
position 238 in bolA1 (Nguyen et al., 1993).
Twenty-two promoter fragments were cloned and
sequenced after 17 cycles of selection. Nineteen promoter
fragments of the correct length contained the sequence
CTTGACA from positions 236 to 230, exactly 17 bp
upstream from the 210 hexamer (Fig. 3B and C); the other
three fragments contained large insertions and probably
were selected for artifactual reasons, as described in
Experimental procedures ). In addition, there was a strong
preference for A residues at positions 229 and 228 and a
lesser preference for T at 227. Positions 238, 237 and
226 to 219 were different from clone to clone, confirming
that the selected promoters were of independent origin.
The selected 235 promoter region showed no similarity to
the same region of the bolA1 promoter, but it contained the
consensus 235 hexamer for recognition by Es70.
Characterization of the selected promoters in vitro
To analyse the effects of the selected sequences on
RNAP binding and transcription, promoters were con-
structed containing the selected 238 to 219 and 218 to
12 regions (either individually or combined together). To
facilitate direct comparison with the bolA1 promoter, each
construct contained sequences from bolA1 at all positions
outside the randomized regions (Fig. 4A). For simplicity,
these constructs are referred to as the ‘in vitro selected’
promoters, including 210 con (containing the selected
218 to 1 2 region from 210 sel #1; Fig. 2), 235 con
(containing the selected 238 to 219 region from 235 sel
#1; Fig. 3) and full con (containing both selected regions).
To verify that the selection resulted in DNA sequences
Fig. 3. 235 region selection.A. Non-template strand DNA sequences of the starting DNA population (235 rnd ) and 235 sel #1 [the first sequence in (B)]. The starting population(235 rnd ) contained the ‘SUB’ upstream sequence (254 to 219; Rao et al., 1994) and the 210 region from 210 sel #1. Randomized region (238 to219) is indicated by ‘NNN…’. The Es70 consensus elements (Gross et al., 1998; Burr et al., 2000) are indicated.B. Sequences from the randomized region of 19 DNA fragments after 17 cycles of selection with EsS in vitro.C. Frequency of bases at each position in the 19 cloned promoters from (B). Numbers in bold indicate positions at which the same base was found inat least 16 out of the 19 fragments.
942 T. Gaal et al.
Q 2001 Blackwell Science Ltd, Molecular Microbiology, 42, 939–954

that bound EsS better than bolA1, we compared the
affinities of the DNA fragments for EsS under conditions
identical to those used for the selection (see Experimental
procedures ). The three selected promoters bound EsS
substantially better than bolA1 (data not shown), confirming
that the selection worked as expected. The relative order of
binding was full con . 235 con . 210 con .. bolA1.
To assess whether the increase in EsS binding by the
selected promoters relative to bolA1 resulted in increased
transcription, the activities of the three selected promoters
were compared with the activity of the bolA1 promoter in vitro.
When transcribed by EsS, all three selected promoters were
much more active than bolA1, and the 235 con promoter was
the strongest, in buffers containing 100–800 mM K-gluta-
mate or 10–400 mM KCl (Fig. 4B and data not shown). The
difference in activity between bolA1 and the selected
promoters was less dramatic at high RNAP concentrations
(data not shown). As the selected promoters also contained
the known consensus sequences for Es70 recognition, we
tested their transcription by Es70 in vitro under the same
conditions used for EsS. The selected promoters were
transcribed very efficiently by Es70, whereas transcription
from the bolA1 promoter was hardly detectable (Fig. 4C).
The selected promoters were transcribed similarly by
both holoenzymes in buffers containing 10–400 mM KCl
(Fig. 4D and data not shown) or 100–800 mM K-glutamate
(Fig. 4E and data not shown). In no case were the
promoters transcribed more than about twofold better by
EsS holoenzyme than by Es70. Therefore, in contrast to
the situation for some osmotically regulated promoters that
are transcribed only by EsS at high concentrations of
K-glutamate (Ding et al., 1995; Kusano and Ishihama,
Fig. 4. In vitro transcription.A. DNA sequences (254 to 116) of the promoters analysed. Sequences outside of the in vitro selected regions are identical in all four promoters andderive from bolA1. The fragments were inserted into a plasmid that contains a transcription terminator ,170 bp downstream of the transcription startsite (11).B. Transcription from the four promoters with EsS. The transcripts from the four promoters are indicated as ‘test’. The RNA I transcript from thevector is also indicated. Reactions contained Transcription buffer (see Experimental procedures ), 100 mM KCl and approximately 1 nM RNAP.C. Transcription from the same templates as in (B) but with Es70 instead of EsS. EsS and Es70 were assembled by addition of fivefold excess of s tothe same amount of core enzyme (see Experimental procedures ). The faint transcript noticeable above the test transcripts in the Es70 reactionsoriginates from a weak promoter whose putative 235 hexamer derives from bolA1 sequences about 10–13 bp upstream of the 235 hexamerresponsible for the major transcript (Nguyen et al., 1993).D. Transcription of the selected promoters at different KCl concentrations in vitro under otherwise identical reaction conditions as in (B) and (C). They-axis is the ratio of specific product (from either the 210 con, 235 con or full con promoter) resulting from transcription by EsS vs. Es70.E. Ratio of transcription by EsS to Es70 at different K-glutamate concentrations.
Promoter recognition and discrimination by Ess RNAP 943
Q 2001 Blackwell Science Ltd, Molecular Microbiology, 42, 939–954
1997), the selected promoters did not exhibit a large
preference for EsS at high osmolarity.
Because EsS and Es70 were prepared by adding
saturating amounts of s factor to aliquots of the same
preparation of core enzyme, it is valid to compare the
relative activities of EsS and Es70 on each of the
promoters (see Experimental procedures ). For each
selected promoter, the level of transcription by EsS and
Es70 was very similar, although the three selected
promoters differed in activity from each other. In contrast,
the wild-type bolA1 promoter was transcribed substantially
better by EsS than by Es70, whereas the plasmid-derived
RNA I promoter was transcribed much better by Es70 than
by EsS. We conclude that the in vitro selected promoters
are transcribed much better than the bolA1 promoter in
vitro, but they do not exhibit specificity for EsS over Es70.
Transcription from the full consensus promoter ( full
con ) was always weaker than from the promoter contain-
ing only the consensus 235 region, 235 con (Fig. 4B and
C). The full con promoter’s relative inactivity results from a
defect in promoter escape (T. Gaal, R.L. Gourse and
N. Shimamoto, unpublished).
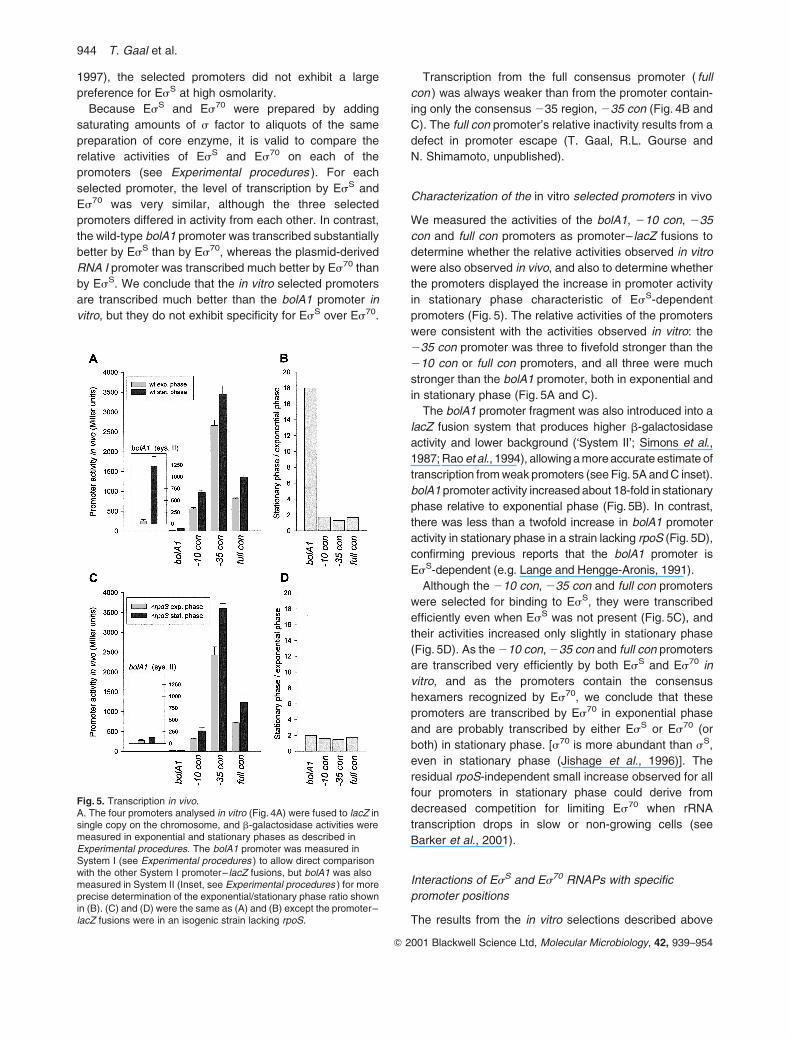
Characterization of the in vitro selected promoters in vivo
We measured the activities of the bolA1, 210 con, 235
con and full con promoters as promoter– lacZ fusions to
determine whether the relative activities observed in vitro
were also observed in vivo, and also to determine whether
the promoters displayed the increase in promoter activity
in stationary phase characteristic of EsS-dependent
promoters (Fig. 5). The relative activities of the promoters
were consistent with the activities observed in vitro: the
235 con promoter was three to fivefold stronger than the
210 con or full con promoters, and all three were much
stronger than the bolA1 promoter, both in exponential and
in stationary phase (Fig. 5A and C).
The bolA1 promoter fragment was also introduced into a
lacZ fusion system that produces higher b-galactosidase
activity and lower background (‘System II’; Simons et al.,
1987; Rao et al., 1994), allowing a more accurate estimate of
transcription from weak promoters (see Fig. 5A and C inset).
bolA1 promoter activity increased about 18-fold in stationary
phase relative to exponential phase (Fig. 5B). In contrast,
there was less than a twofold increase in bolA1 promoter
activity in stationary phase in a strain lacking rpoS (Fig. 5D),
confirming previous reports that the bolA1 promoter is
EsS-dependent (e.g. Lange and Hengge-Aronis, 1991).
Although the 210 con, 235 con and full con promoters
were selected for binding to EsS, they were transcribed
efficiently even when EsS was not present (Fig. 5C), and
their activities increased only slightly in stationary phase
(Fig. 5D). As the 210 con, 235 con and full con promoters
are transcribed very efficiently by both EsS and Es70 in
vitro, and as the promoters contain the consensus
hexamers recognized by Es70, we conclude that these
promoters are transcribed by Es70 in exponential phase
and are probably transcribed by either EsS or Es70 (or
both) in stationary phase. [s70 is more abundant than sS,
even in stationary phase (Jishage et al., 1996)]. The
residual rpoS-independent small increase observed for all
four promoters in stationary phase could derive from
decreased competition for limiting Es70 when rRNA
transcription drops in slow or non-growing cells (see
Barker et al., 2001).
Interactions of EsS and Es70 RNAPs with specific
promoter positions
The results from the in vitro selections described above
Fig. 5. Transcription in vivo.A. The four promoters analysed in vitro (Fig. 4A) were fused to lacZ insingle copy on the chromosome, and b-galactosidase activities weremeasured in exponential and stationary phases as described inExperimental procedures. The bolA1 promoter was measured inSystem I (see Experimental procedures ) to allow direct comparisonwith the other System I promoter– lacZ fusions, but bolA1 was alsomeasured in System II (Inset, see Experimental procedures ) for moreprecise determination of the exponential/stationary phase ratio shownin (B). (C) and (D) were the same as (A) and (B) except the promoter–lacZ fusions were in an isogenic strain lacking rpoS.
944 T. Gaal et al.
Q 2001 Blackwell Science Ltd, Molecular Microbiology, 42, 939–954
suggested that both EsS and Es70 prefer the same 210,
235 and extended 210 consensus sequences. However,
the identification of sequences in in vitro selection
experiments does not imply that each selected position
has the same relative importance to RNAP binding, as
even small increments to binding will result in selection,
nor do these experiments address whether EsS and Es70
interact with a particular promoter in the same manner. To
address whether the two enzymes recognize the same
promoter sequence differently, we used interference
footprinting, a technique in which DNA fragments with
modifications at different positions in the population can all
be assayed for protein binding at the same time.
Specifically, if a modification at a certain position interferes
with RNAP binding, the promoter fragment containing that
modification will be underrepresented in the bound
population. Interference footprinting thus allows estimation
of the relative contribution of individual positions in a
promoter to RNAP binding (although of course the effects
of every functional group or even every basepair are not
tested by any one interference probe).
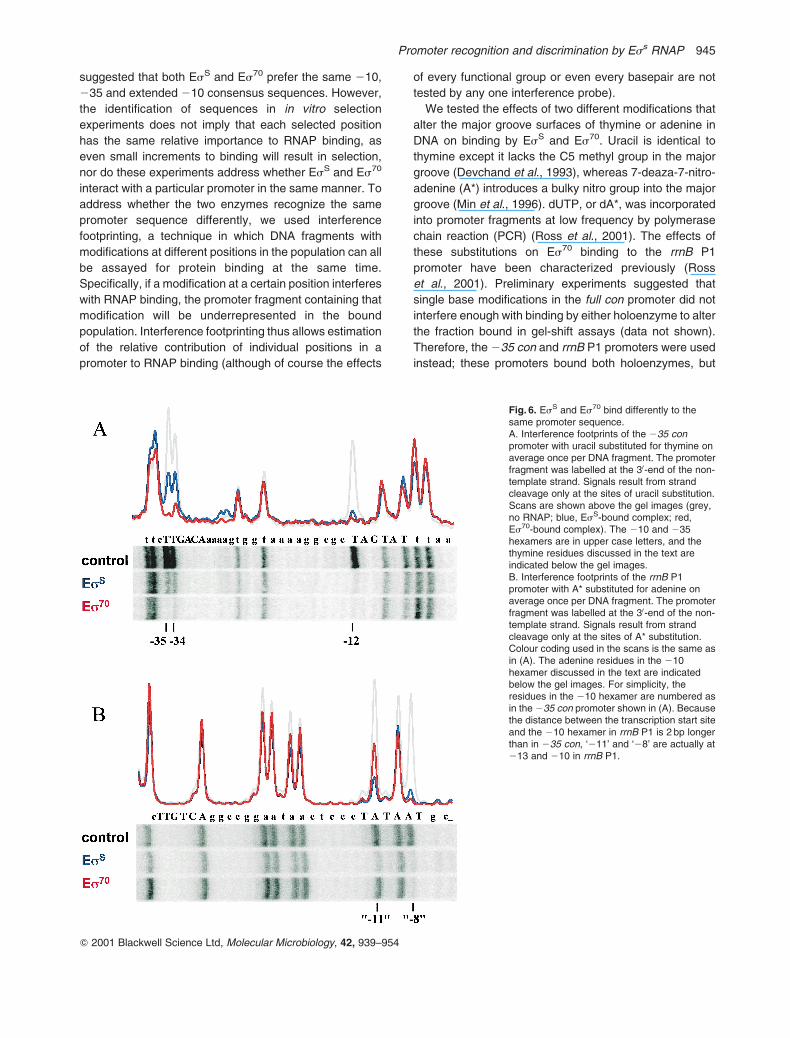
We tested the effects of two different modifications that
alter the major groove surfaces of thymine or adenine in
DNA on binding by EsS and Es70. Uracil is identical to
thymine except it lacks the C5 methyl group in the major
groove (Devchand et al., 1993), whereas 7-deaza-7-nitro-
adenine (A*) introduces a bulky nitro group into the major
groove (Min et al., 1996). dUTP, or dA*, was incorporated
into promoter fragments at low frequency by polymerase
chain reaction (PCR) (Ross et al., 2001). The effects of
these substitutions on Es70 binding to the rrnB P1
promoter have been characterized previously (Ross
et al., 2001). Preliminary experiments suggested that
single base modifications in the full con promoter did not
interfere enough with binding by either holoenzyme to alter
the fraction bound in gel-shift assays (data not shown).
Therefore, the 235 con and rrnB P1 promoters were used
instead; these promoters bound both holoenzymes, but
Fig. 6. EsS and Es70 bind differently to thesame promoter sequence.A. Interference footprints of the 235 conpromoter with uracil substituted for thymine onaverage once per DNA fragment. The promoterfragment was labelled at the 30-end of the non-template strand. Signals result from strandcleavage only at the sites of uracil substitution.Scans are shown above the gel images (grey,no RNAP; blue, EsS-bound complex; red,Es70-bound complex). The 210 and 235hexamers are in upper case letters, and thethymine residues discussed in the text areindicated below the gel images.B. Interference footprints of the rrnB P1promoter with A* substituted for adenine onaverage once per DNA fragment. The promoterfragment was labelled at the 30-end of the non-template strand. Signals result from strandcleavage only at the sites of A* substitution.Colour coding used in the scans is the same asin (A). The adenine residues in the 210hexamer discussed in the text are indicatedbelow the gel images. For simplicity, theresidues in the 210 hexamer are numbered asin the 235 con promoter shown in (A). Becausethe distance between the transcription start siteand the 210 hexamer in rrnB P1 is 2 bp longerthan in 235 con, ‘211’ and ‘28’ are actually at213 and 210 in rrnB P1.
Promoter recognition and discrimination by Ess RNAP 945
Q 2001 Blackwell Science Ltd, Molecular Microbiology, 42, 939–954
with lower affinity than the full con promoter, so that uracil
or A* substitutions resulted in changes in the fraction of the
DNA population bound by RNAP.
Uracil incorporation at three non-template strand
positions in the 235 con promoter (one in the 210 region
at 212, and two in the 235 region at 234 and 235)
reduced binding by both EsS and Es70 (Fig. 6A). Uracil at
212 virtually eliminated binding by both enzymes. In
contrast, uracil at either 234 or 235 greatly reduced Es70
binding, but only moderately reduced EsS binding. Similar
differential effects of uracil substitution at 234 and 235 on
Es70 versus EsS binding were also observed on the rrnB
P1 promoter (data not shown). These results suggested
that even though both holoenzymes prefer the same
promoter sequence, they interact with that promoter
differently.
The effects of A* substitutions for adenine in the non-
template strand on Es70 and EsS binding to the rrnB P1
promoter are illustrated in Fig. 6B; qualitatively similar
results were obtained for the 210 con and 235 con
promoters (data not shown). Modification of either of two
positions in the 210 hexamer reduced RNAP binding,
whereas no effects were observed in the 235 region.
Binding of the two holoenzymes was affected differentially
at one of the 210 region positions, the highly conserved
A211 in the 210 hexamer. At this position, A* decreased
EsS binding by about 60% but decreased Es70 binding by
only about 30%. As with the uracil interference exper-
iments, the A* footprints indicate that EsS and Es70 bind
differently to the same promoter sequence. The less than
total inhibition observed from introduction of A* for the
highly conserved A211 could be attributable to the fact
that the N1 position on the base is most crucial for A211
function (Matlock and Heyduk, 2000), not the 6 and 7
positions on the base altered in A*.
A* substitution at A29 had a smaller effect than at
A211, and A* substitution at A28 had a larger effect than
at A211, but in both cases the effects were similar for the
two holoenzymes. The strong inhibition of RNAP binding
by A* at 28 could be attributable to steric clash by
introduction of the bulky nitro group, as typically mutations
at 28 have less severe effects on RNAP interactions than
at 211.
Basis for promoter selectivity by EsS versus Es70
The similarity in the intrinsic promoter recognition proper-
ties of EsS and Es70 raises the issue of how some
promoters could be almost entirely dependent on EsS.
That is, what accounts for some promoters being
transcribed only by EsS when both EsS and Es70 are
present in stationary phase, and for these same promoters
not being transcribed by Es70 in exponential phase? In this
section, we propose a model to address these questions
and then we provide experimental support for this
proposal.
Our data suggest that the preferred hexamer sequences
might be identical for EsS and Es70. However, naturally
occurring promoter sequences rarely match the consen-
sus perfectly for a particular RNAP. Differences from the
Es70 consensus sequence, in conjunction with the effects
of specific activators and repressors, are responsible for
differences in levels of transcription initiation from
individual Es70 promoters. The interference footprints
shown above indicate that certain modifications of the 210
and 235 hexamers affect binding by EsS and Es70
differently. Therefore, to explain differences in transcrip-
tion by EsS versus Es70, we suggest that non-preferred
basepairs in a promoter sequence might have different
effects on recognition by the two enzymes.
To test this general concept, we chose two positions in
the 235 element for detailed examination. The choice of
these positions was dictated by the results of our
interference footprinting studies (Fig. 6A) indicating that
the loss of the methyl groups at positions 234 and 235
affect discrimination by Es70 versus EsS and by previous
genetic evidence suggesting that C substitutions at 234
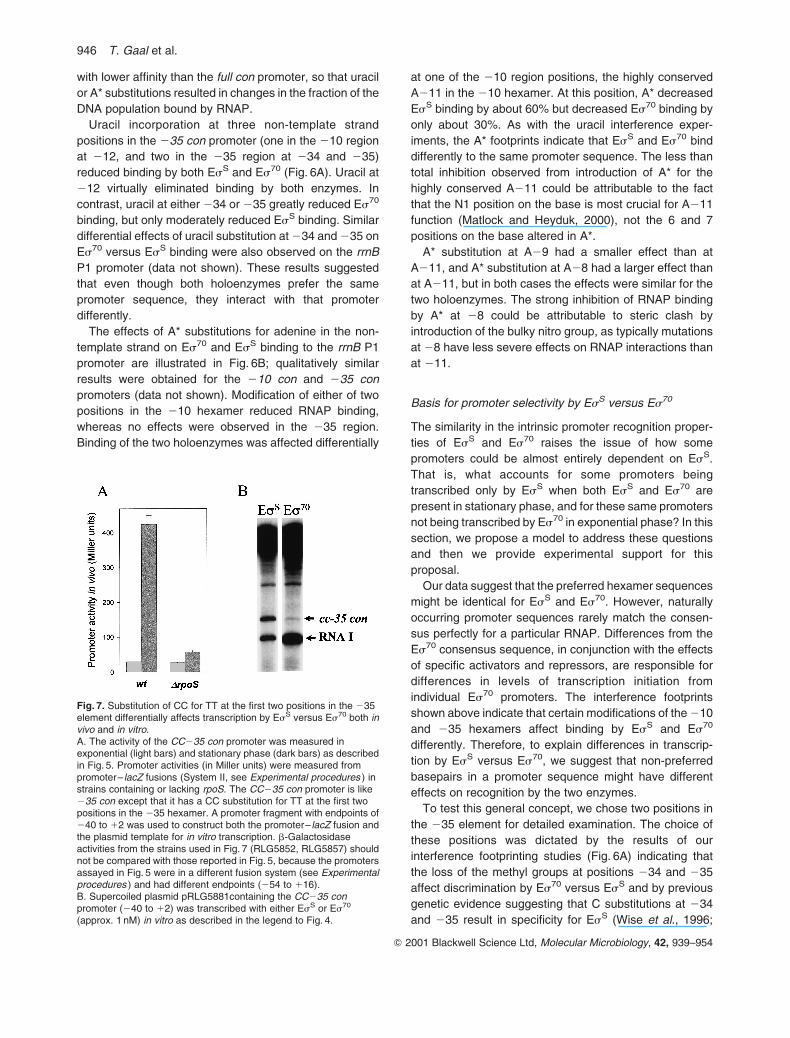
and 235 result in specificity for EsS (Wise et al., 1996;
Fig. 7. Substitution of CC for TT at the first two positions in the 235element differentially affects transcription by EsS versus Es70 both invivo and in vitro.A. The activity of the CC235 con promoter was measured inexponential (light bars) and stationary phase (dark bars) as describedin Fig. 5. Promoter activities (in Miller units) were measured frompromoter– lacZ fusions (System II, see Experimental procedures ) instrains containing or lacking rpoS. The CC235 con promoter is like235 con except that it has a CC substitution for TT at the first twopositions in the 235 hexamer. A promoter fragment with endpoints of240 to 12 was used to construct both the promoter– lacZ fusion andthe plasmid template for in vitro transcription. b-Galactosidaseactivities from the strains used in Fig. 7 (RLG5852, RLG5857) shouldnot be compared with those reported in Fig. 5, because the promotersassayed in Fig. 5 were in a different fusion system (see Experimentalprocedures ) and had different endpoints (254 to 116).B. Supercoiled plasmid pRLG5881containing the CC235 conpromoter (240 to 12) was transcribed with either EsS or Es70
(approx. 1 nM) in vitro as described in the legend to Fig. 4.
946 T. Gaal et al.
Q 2001 Blackwell Science Ltd, Molecular Microbiology, 42, 939–954
Bordes et al., 2000). To confirm that the identities of 234
and 235 contribute to discrimination by Es70 versus EsS,
we constructed a CC-35 con promoter (i.e. a 235 con
promoter containing T-34C and T-35C substitutions) fused
to lacZ. The promoter DNA fragment used in this fusion
extended only to 240 to eliminate any contribution to
activity in vivo from a weak Es70-dependent promoter
upstream of the 235 hexamer that derives from bolA1
sequences (see Fig. 4C).
CC235 con was considerably weaker than 235 con and
displayed the characteristics of an EsS-specific promoter
in vivo (Fig. 7A). Transcription from CC235 con greatly
increased in stationary relative to exponential phase, and
this increase was almost completely dependent on the
presence of rpoS. (The promoter endpoints and the lacZ
reporter system used in Fig. 7 differ from those used in
Fig. 5. Thus, the absolute b-galactosidase activities in the
two figures should not be compared directly (see also
Experimental procedures ). We also tested transcription
from CC235 con with EsS and Es70 in vitro (Fig. 7B).
Consistent with the results obtained in vivo, CC235 con
was preferentially utilized by EsS in vitro. EsS tolerated the
CC substitution much better than did Es70 under every
condition tested.
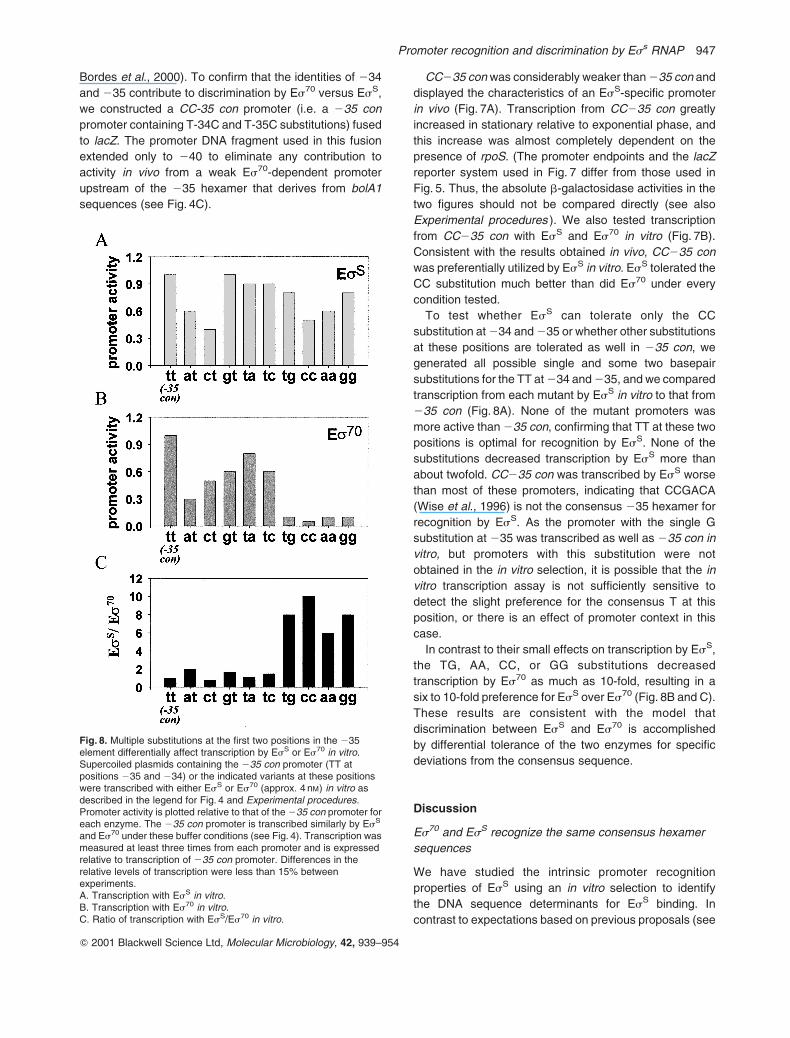
To test whether EsS can tolerate only the CC
substitution at 234 and 235 or whether other substitutions
at these positions are tolerated as well in 235 con, we
generated all possible single and some two basepair
substitutions for the TT at 234 and 235, and we compared
transcription from each mutant by EsS in vitro to that from
235 con (Fig. 8A). None of the mutant promoters was
more active than 235 con, confirming that TT at these two
positions is optimal for recognition by EsS. None of the
substitutions decreased transcription by EsS more than
about twofold. CC235 con was transcribed by EsS worse
than most of these promoters, indicating that CCGACA
(Wise et al., 1996) is not the consensus 235 hexamer for
recognition by EsS. As the promoter with the single G
substitution at 235 was transcribed as well as 235 con in
vitro, but promoters with this substitution were not
obtained in the in vitro selection, it is possible that the in
vitro transcription assay is not sufficiently sensitive to
detect the slight preference for the consensus T at this
position, or there is an effect of promoter context in this
case.
In contrast to their small effects on transcription by EsS,
the TG, AA, CC, or GG substitutions decreased
transcription by Es70 as much as 10-fold, resulting in a
six to 10-fold preference for EsS over Es70 (Fig. 8B and C).
These results are consistent with the model that
discrimination between EsS and Es70 is accomplished
by differential tolerance of the two enzymes for specific
deviations from the consensus sequence.
Discussion
Es70 and EsS recognize the same consensus hexamer
sequences
We have studied the intrinsic promoter recognition
properties of EsS using an in vitro selection to identify
the DNA sequence determinants for EsS binding. In
contrast to expectations based on previous proposals (see
Fig. 8. Multiple substitutions at the first two positions in the 235element differentially affect transcription by EsS or Es70 in vitro.Supercoiled plasmids containing the 235 con promoter (TT atpositions 235 and 234) or the indicated variants at these positionswere transcribed with either EsS or Es70 (approx. 4 nM) in vitro asdescribed in the legend for Fig. 4 and Experimental procedures.Promoter activity is plotted relative to that of the 235 con promoter foreach enzyme. The 235 con promoter is transcribed similarly by EsS
and Es70 under these buffer conditions (see Fig. 4). Transcription wasmeasured at least three times from each promoter and is expressedrelative to transcription of 235 con promoter. Differences in therelative levels of transcription were less than 15% betweenexperiments.A. Transcription with EsS in vitro.B. Transcription with Es70 in vitro.C. Ratio of transcription with EsS/Es70 in vitro.
Promoter recognition and discrimination by Ess RNAP 947
Q 2001 Blackwell Science Ltd, Molecular Microbiology, 42, 939–954

Introduction ), the binding sites selected by EsS contained
the consensus hexamer sequences for binding Es70, and
they initiated transcription at high levels by both
holoenzymes.
Many EsS-dependent promoters rely predominantly on
recognition of the 210 region (e.g. Tanaka et al., 1995;
Espinosa-Urgel et al., 1996; Colland et al., 1999; Lee and
Gralla, 2001). However, the results of the in vitro selection
and the extraordinary strength of the 235 con promoter,
relative to the bolA1 promoter, when transcribed by EsS in
vitro, suggest that the 235 element can also play a
prominent role in recognition by EsS in some promoter
contexts. In fact, some EsS-dependent promoters (e.g.
osmE; Bordes et al., 2000) have almost consensus 235
regions and appear to be strongly dependent on 235
hexamer interactions.
Differential tolerance of RNAP holoenzymes for deviations
from the same consensus hexamers
As both Es70 and EsS prefer the same consensus
hexamer sequences, there must be a mechanism for
preventing transcription from stationary phase-specific
promoters by Es70 in exponential phase. (Preventing
transcription from these promoters by EsS in exponential
phase is accomplished by limiting expression of sS.) We
suggest that holoenzyme specificity might rely, at least in
part (see below), on tolerance of the two enzymes for
different deviations from the same consensus sequences.
This hypothesis is supported by interference footprinting
studies showing that EsS and Es70 recognize the same
promoter sequence differently (Fig. 6) and by the effects of
certain promoter mutations constructed as a test of this
concept (Figs. 7 and 8). We found that, in the context of the
235 con promoter, substitutions of TT to TG, CC, AA, or
GG at the first two positions of the 235 hexamer resulted
in transcription in vitro that was strongly dependent on
EsS, in contrast to 235 con. These results are consistent
with genetic studies on the proU promoter, in which TT to
CC substitutions at the same positions strongly reduced
transcription and made it dependent on rpoS in vivo, and
on the osmY promoter, in which CC to TT substitutions
decreased discrimination between EsS and Es70 in vivo
(Wise et al., 1996).
We did not evaluate the relative preferences of EsS and
Es70 for each possible basepair at every position in the
consensus hexamers. However, several results strongly
suggest that, in addition to positions in the 235 hexamer,
there are positions in the 210 hexamer at which
interactions with the two RNAPs are differentially affected
by the same base modification or base substitution. These
include the results of our interference footprints (Fig. 6),
the fact that the promoter differing from bolA1 only in the
210 region (210 con ) is recognized by EsS but has lost its
EsS specificity (Fig. 4), and recent studies showing there is
differential binding of EsS and Es70 to fork-junction
templates with substitutions in the 210 region (Lee and
Gralla, 2001).
At position 28, A and C were about equally represented
in the sequences selected by EsS, whereas A is
considered the consensus for Es70-dependent promoters.
Although there are some differences between s70 and sS
in the region predicted to interact with position 28 (region
2.3; Lonetto et al., 1992; Malhotra et al., 1996; Fenton
et al., 2000), we suggest that the identity of 28 does not
play a major role in discrimination between the two
holoenzymes. We constructed promoters differing only by
A or C at this position and found they were transcribed
similarly by the two enzymes in vitro (data not shown), and
previous reports suggest that both Es70 and EsS tolerate A
or C at 28 (Oliphant and Struhl, 1988; Kolb et al., 1995;
Tanaka et al., 1995; Espinosa-Urgel et al., 1996).
Recognition of either A or C at 28 by both enzymes
could potentially be attributable to an interaction with the
same functional group on both bases, for example the
amino group on position 6 of A, or 4 of C.
Other potential contributions to differential promoter
recognition by EsS and Es70
Recent studies have proposed that a C at position 213
(i.e. the position just upstream from the 210 hexamer) is
preferred by EsS, and G213 is preferred by Es70 in certain
contexts, for example in the csiD and osmY promoters
(Becker and Hengge-Aronis, 2001) and in fork-junction
templates (Lee and Gralla, 2001). The results of our in vitro
selection strongly support the preference of EsS for C at
213. However, C213 is clearly not sufficient to
discriminate against recognition by Es70 in the context of
the in vitro selected promoters described here (Figs. 4 and
5). In addition, the rrn P1 promoters that are transcribed
much more efficiently by Es70 than by EsS (data not
shown) contain a C at this position.
Residues in addition to the consensus hexamers and
C-13 were selected by EsS in our in vitro binding
experiments, including a cytosine at the upstream flank
of the 235 hexamer (236), two adenines at the positions
just downstream of the 235 hexamer (229 and 228) and
the sequence TGTG at positions 214 to 217 (the
extended 210 region); 236, 229, and 228 are not
usually considered as being determinants of Es70 binding
and, therefore, it is formally possible that they might
contribute to specific EsS recognition. However, we
consider it improbable that these basepairs, by them-
selves, are responsible for discrimination between EsS
and Es70, as (i) C is also preferred by Es70 at 236 in the
lac and rrnB P1 promoters (Reznikoff, 1976; Josaitis et al.,
1990), and (ii) A to G substitutions for the A residues at
948 T. Gaal et al.
Q 2001 Blackwell Science Ltd, Molecular Microbiology, 42, 939–954

229 and 228, at least in the context of the 235 con
promoter, did not affect activity substantially in vitro with
either holoenzyme (data not shown). [We note, however,
that an A-tract downstream of the 235 hexamer was
identified as a requirement for efficient rRNA promoter
activity by Chlamydia trachomatis RNAP (Tan et al.,
1998)]. The interaction between the extended 210 motif
(TGTG) and region 2.5 of s has been well documented as
an important determinant of both EsS- and Es70-depen-
dent transcription (Voskuil et al., 1995; Barne et al., 1997;
Becker et al., 1999; Colland et al., 1999; Burr et al., 2000).
A spacer length of 17 bp between the 210 and 235
hexamers was strongly favoured for recognition by EsS in
our in vitro selection. We found that Es70 tolerated a 16 bp
spacer in the context of the rrnB P1 promoter much better
than EsS, but a 16 bp spacer reduced the activity of the full
con promoter approximately the same for both holoen-
zymes (data not shown). Therefore, spacer-length could
contribute to EsS versus Es70 discrimination at some
promoters, but our preliminary studies indicate that the
effect of spacer length on the two holoenzymes is context-
dependent and complex.
Finally, our hypothesis does not exclude a role for
additional DNA-binding proteins in promoter discrimination
by EsS and Es70. That is, expression of some promoters
(although not those investigated here) is influenced by
differential effects of transcription factors or nucleoid-
associated proteins on the two holoenzymes (Arnqvist
et al., 1994; Bouvier et al., 1998; Colland et al., 2000).
s determinants for differential recognition of the same
consensus hexamers
The similarity between the EsS and Es70 consensus
sequences is consistent with the similarity of the motifs in
the two s factors responsible for promoter recognition.
Sixteen out of 28 amino acids in region 4.2 (residues 572–
599 of s70) and 25 out of 40 amino acids in regions 2.3 and
2.4 (residues 417–456 of s70) are identical in the two s
factors (Lonetto et al., 1992), including several residues
implicated in direct interactions with specific basepairs in
the 210 and 235 hexamers (e.g. Q437 in region 2.4, and
R584 and R588 in region 4.2; s70 numbering, Gross et al.,
1998).
Superimposed on the overall conservation between the
two s factors in the regions responsible for DNA binding
are discrete regions of divergence that could contribute to
promoter discrimination. Two clusters of four amino acids
in region 4.2 (residues 578–581 and 591–594 of s70) are
virtually identical among s70 members from different
species and among different sS members from different
species, but these sequences differ between s70 and sS
(Lonetto et al., 1992) and could potentially play a role in
discrimination between deviations from the consensus
235 hexamer. K173 of sS has been implicated in
recognition of C213 (Becker and Hengge-Aronis, 2001).
The corresponding residue in s70 is different (E458),
consistent with their different DNA recognition prefer-
ences. As interactions with the 210 element are complex,
involving binding to both single-stranded and double-
stranded DNA in multiple steps during the process of
transcription initiation, it is more difficult with present
information to predict specific amino acid residues in s70
and sS that might play a role in differential 210 region
recognition.
Promoters recognized by both EsS and Es70
The observed overlap in the sequences recognized by
EsS and Es70 potentially allows some promoters to be
transcribed by both holoenzymes under some conditions,
and yet to be transcribed by only one or the other
holoenzyme under other conditions. Even though a
promoter might be transcribed by both enzymes under
some conditions, changes in the environment that alter
template geometry (e.g. superhelicity) or solute concen-
trations (e.g. anions or cations) might be sufficient to alter
the ratio of transcription by the two holoenzymes
(Ishihama, 2000).
It has been reported that the two holoenzymes possess
different tolerances for high concentrations of K-glutamate
or trehalose, and that this favours specific promoter
recognition by EsS (Ding et al., 1995; Kusano and
Ishihama, 1997; Nguyen and Burgess, 1997). We
compared the activities of EsS and Es70 on the bolA1,
210 con, 235 con, and full con promoters in buffers
containing 100–800 mM K-glutamate or 10–400 mM KCl
(data not shown). As predicted from previous studies
(Leirmo et al., 1987), the anion glutamate was less
disruptive to RNAP–promoter interactions than chloride,
so transcription by both holoenzymes was generally higher
at the same cation concentration in glutamate buffers.
Tolerance for high salt concentrations may contribute to
the ability of certain promoters to be transcribed only by
EsS (Ding et al., 1995; Kusano and Ishihama, 1997).
However, the in vitro selected promoters described here
are transcribed by both holoenzymes at high osmolarity;
high salt does not result in holoenzyme specificity.
The degree of negative supercoiling of the bacterial
chromosome, on average, is somewhat lower in stationary
phase than in exponential growth, and it has been
suggested that reduced supercoiling may enhance
transcription by EsS rather than by Es70 (Kusano et al.,
1996). Our promoter selections were performed on DNA
fragments (linking number¼ 0), but the promoters were
also transcribed efficiently by both holoenzymes on
supercoiled plasmid templates (linking
number¼<20.06). Therefore, it is probable that the
Promoter recognition and discrimination by Ess RNAP 949
Q 2001 Blackwell Science Ltd, Molecular Microbiology, 42, 939–954

same consensus hexamers are recognized best by both
EsS and Es70 on templates with a wide range of
superhelicities. Nevertheless, as with changes in osmo-
larity, conditions that alter template geometry could
potentially favour transcription by one holoenzyme or the
other in a specific promoter context.
Conclusions
We have shown for the first time that two RNAP
holoenzymes present in the same organism recognize
the same consensus hexamer sequences. This poses a
potential problem for promoter specificity, but we have
proposed a model that might resolve this dilemma. The
principle of tolerance for different deviations from the same
consensus sequence was recognized long ago as the
basis for differential binding of the Cro and cI repressor
proteins from bacteriophages l and 434 (e.g. Hochschild
et al., 1986; Harrison and Aggarwal, 1990; Albright and
Matthews, 1998). Cro and cI bind to the same consensus
operator sequences, but the two proteins prefer different
non-consensus bases at particular positions in the DNA
sequence.
Differential tolerance of two holoenzymes for certain
deviations from the same consensus is apparently not the
method used for discrimination between the other E. coli
RNAP holoenzymes, as the other E. coli s factors have
diverged from each other more than sS and s70 and
recognize qualitatively different DNA sequences. Even in
the case of promoters recognized by EsS versus Es70,
context is important to the effects of individual promoter
positions on RNAP recognition, and thus it is difficult to
predict which deviations from consensus account for
discrimination simply by inspection of a particular promoter
sequence. However, we suggest that the concept of
tolerance for different deviations from the same consensus
sequence as a mechanism for achieving specificity should
be considered in transcription studies of organisms with
multiple closely related RNAP holoenzymes, a common
feature in bacteria. For example, recent genome analysis
suggests that Streptomyces coelicolor might have as
many as 40 ECF s factors (http://www.sanger.ac.uk/
Projects/S_coelicolor/ ). We speculate that the principle
described here for discrimination between E. coli EsS and
Es70 might be utilized for discrimination between members
of s factor families in other bacteria.
Table 1. Bacterial strains and plasmids.
Name Genotypea Source
StrainsRLG3499 VH1000¼MG1655 lacZ, lacI, pyrE 1 Gaal et al. (1997)UM122 rpoS::Tn10 Xu and Johnson (1995)RLG3773 VH1000 lI bolA1– lacZ(254 to 116) This workRLG3774 VH1000 lI bolA1– lacZ rpoS::Tn10 This workRLG3760 VH1000 lII bolA1– lacZ (254 to 116) This workRLG3764 VH1000 lII bolA1– lacZ rpoS::Tn10 This workRLG3750 VH1000 lI 210 con– lacZ (254 to 116) This workRLG3769 VH1000 lI 210 con– lacZ rpoS::Tn10 This workRLG3752 VH1000 lI 235 con– lacZ (254 to 116) This workRLG3770 VH1000 lI 235 con– lacZ rpoS::Tn10 This workRLG3754 VH1000 lI full con– lacZ (254 to 116) This workRLG3771 VH1000 lI full con– lacZ rpoS::Tn10 This workRLG5852 VH1000 lII CC 235 con– lacZ (240 to 12) This workRLG5857 VH1000 lII CC 235 con– lacZ rpoS::Tn10 This workPlasmidspRLG770 pKM2 with rrnB T1T2 terminators Ross et al. (1990)pRLG3405 pRLG770 with bolA1 promoter (–54 to 116) This workpRLG3747 pRLG770 with 210 con promoter (–54 to 116) This workpRLG3748 pRLG770 with 235 con promoter (–54 to 116) This workpRLG3749 pRLG770 with full con promoter (–54 to 116) This workpRLG3790 pRLG770 with AT 235 con promoter (–54 to 116) This workpRLG3791 pRLG770 with CT 235 con promoter (–54 to 116) This workpRLG3792 pRLG770 with GT 235 con promoter (–54 to 116) This workpRLG3793 pRLG770 with TA 235 con promoter (–54 to 116) This workpRLG3794 pRLG770 with TC 235 con promoter (–54 to 116) This workpRLG3795 pRLG770 with TG 235 con promoter (–54 to 116) This workpRLG3798 pRLG770 with AA 235 con promoter (–54 to 116) This workpRLG3796 pRLG770 with CC 235 con promoter (–54 to 116) This workpRLG3799 pRLG770 with GG 235 con promoter (–54 to 116) This workpRLG5881 pRLG770 with CC 235 con promoter (–40 to 12) This work
a. lI refers to lacZ fusion system I, and lII refers to lacZ fusion system II (see Experimental procedures ).Numbers in parentheses refer to sequence endpoints of DNA fragments used to construct promoter– lacZ fusions and plasmids.
950 T. Gaal et al.
Q 2001 Blackwell Science Ltd, Molecular Microbiology, 42, 939–954

Experimental procedures
Promoter fragment library
In vitro selections with EsS were performed with libraries of86 bp promoter fragments containing 20 bp of random DNA
sequence. Non-template strand oligonucleotides (Integrated
DNA Technologies) for the 210 region selection contained(from 50 to 30) an Eco R1 site, bolA1 promoter sequences from
254 to 219, random sequences from 218 to 12 generatedfrom equimolar mixtures of the four nucleotides, bolA1
sequences from 13 to 116, and a HindIII site. Non-templatestrand oligonucleotides for the 235 region selection contained
an Eco R1 site, the ‘SUB’ sequence (Rao et al., 1994) from254 to 239, random sequences from 238 to 219, selected
210 region sequence #1 (Fig. 2B) from 218 to 12, bolA1sequences from 13 to 116 and a Hind III site. The template
strand for each of the two selections was generated byannealing 60 pmol of the non-template strand with 60 pmol of
a 21-mer complementary to the sequence from 13 to the HindIII site. The reactions were incubated in 25ml Sequenase
buffer (US Biochemical) for 5 min at 958C, slowly cooled toroom temperature, dNTPs were added to 1 mM, and the
annealed oligos were extended with 20 units of Sequenase for20 min at 378C. The DNAs were digested with Eco RI and
HindIII, and aliquots were cloned into M13mp18 for sequenceanalysis before selection with EsS to ensure that the
randomized regions contained approximately equal percen-tages of all four bases. The rest of the DNA was labelled at
both ends with [a232P]-dATP using Sequenase before use inthe selection.
RNA polymerases
EsS was prepared from core RNAP and highly purified sS
(Nguyen et al., 1993). The preparations contained no
detectable other s factors as judged on silver-stained gels(data not shown). EsS was reconstituted by mixing purified
core with a fivefold excess of sS (1mM) for 45 min at 308C.Es70 was reconstituted using the same core RNAP
preparation and a fivefold excess of purified s70. Increasingthe concentration of either s factor further did not increase
transcription (data not shown). Therefore, the two holoen-zyme preparations contain the same number of active RNAPs
(unless there are s molecules that bind to core but are notactive in transcription).
In vitro selection
The selection consisted of repeated cycles of RNAP binding,separation of the bound population from free DNA by
electrophoresis on 4% polyacrylamide gels in 0.5X Tris-Borate-EDTA buffer for 2–3 h at ,10 V cm21 and PCR
amplification (Fig. 1). For the initial 50ml binding reaction, 2mg(0.7mM) of the double-stranded DNA fragment library was
incubated with 20 nM EsS in 50 mM HEPES (pH 7.0), 100 mMKCl, 10 mM Mg-acetate, 0.1 mM dithiothreitol (DTT),
100mg ml21 BSA, 5% glycerol at room temperature for20 min. Then, 25mg ml21 heparin was added to prevent
further RNAP binding, and the samples were immediately
loaded on the gel. The RNAP–promoter complexes (0.5–5%
of the total DNA) were visualized by phosphorimaging,excised, eluted, extracted with phenol and precipitated with
ethanol. The recovered DNA was used as a template for PCRamplification using Taq DNA polymerase for 15 cycles (958C
for 1 min; 558C for 1 min; 728C for 1 min). For the 210selection, the PCR primers were complementary to 20 nt at
each end of the promoter DNA fragment, whereas in the 235selection, the primers were complementary to all except the
randomized positions (238 to 219). Aliquots were checkedby gel electrophoresis on 4% agarose gels and staining with
ethidium bromide for successful amplification. The amplifiedDNA was digested with HindIII, end-labelled and used for the
next cycle of RNAP binding. The binding time wasprogressively shortened, and the RNAP concentration was
gradually decreased, in subsequent selection cycles, until inthe last round the reaction time was 2 min and the RNAP
concentration was 4 nM. The progress of the selection wasfollowed after 10, 14 and 17 cycles by cloning samples into
M13 and sequencing a number of individual clones from the
selected population. The selection was stopped after 17cycles because inspection indicated that an obvious
consensus sequence had been reached.Heteroduplexes (which bind RNAP very efficiently) some-
times arose in the final cycles of the PCR from annealing of
single-stranded DNAs that contained different sequences inthe randomized region or from annealing of insertions and
deletions that arose from mistakes during oligonucleotidesynthesis or amplification. These appear on agarose gels as a
blurred region of stained material, migrating slower than theband of the expected size. If contamination from hetero-
duplexes was detected, 5ml of the reaction was used astemplate for an additional two cycles of PCR amplification in a
50ml reaction.
Strains and plasmids
Plasmids and strains are listed in Table 1. Cloning with M13and plasmids was carried out using standard techniques.
Plasmid templates for in vitro transcription were constructedby insertion of promoter fragments into pRLG770 (Ross et al.,
1990). Site-directed promoter mutations in 235 con weregenerated by PCR using plasmid pRLG3748 as template and
primers containing the desired sequence alterations. DNAsequencing was performed using a Sequenase kit supplied by
US Biochemicals.l lysogens were constructed containing promoter– lacZ
fusions as described (Rao et al., 1994). Two promoter– lacZ
fusion systems were used; ‘System II’ (Rao et al., 1994) wasused for measuring transcription from the bolA1 and CC235
con promoters as indicated. This fusion system has a very lowbackground, but it cannot tolerate very strong promoters.
‘System I’ (Rao et al., 1994), which has higher background butcan accommodate strong promoters, was used in all other
cases. The same promoter makes at least 6.7-fold more b-galactosidase in System II than in System I. The CC235 con
and bolA1 promoters in the lacZ fusions used in Fig. 7(Table 1; RLG5852, RLG5857, RLG5861 and RLG5862)
contain sequences only from 240 to 12 to eliminateinterference from the weak Es70 binding site upstream of
the 235 hexamer (see Fig. 4 and Nguyen et al., 1993). As the
reporter systems and the downstream endpoints in the
Promoter recognition and discrimination by Ess RNAP 951
Q 2001 Blackwell Science Ltd, Molecular Microbiology, 42, 939–954

promoters used for these fusions (12) are different from thoseused for the fusions in Fig. 5 (116), probably resulting in
different mRNA half-lives for the resulting transcripts, the b-galactosidase activities shown in Fig. 7 should not be
compared directly with those shown in Fig. 5. Strains lackingsS were constructed by transduction of rpoS::Tn10 from
RLG3237 (¼UM122; Xu and Johnson, 1995) with P1vir.
Measurement of promoter activity in vivo
Cells were grown in Luria–Bertani (LB) medium at 308C, and
b-galactosidase activity was measured at regular intervalsthroughout a growth cycle. To estimate promoter activity in
exponential phase, cells were grown to an A600 of < 0.25–0.5and then diluted 100-fold and grown again to ensure that b-
galactosidase, accumulated previously during stationaryphase, was minimized. The promoter activity at A600 0.25
and 0.5 was essentially identical. To estimate promoteractivity in stationary phase, b-galactosidase activity was
measured from cells grown to an A600 of 2.6 and 4.0 (in whichb-galactosidase activity was essentially identical).
Interference footprinting
Interference footprinting was performed as described in detail
elsewhere (Ross et al., 2001). The DNA contained, onaverage, one modified base (U instead of T; Devchand et al.,
1993; or A* instead of A; Min et al., 1996) per fragment,incorporated by PCR. In brief, RNAP (2 nM EsS or Es70) was
incubated with an excess of 32P end-labelled DNA fragment(10 nM) for 10 min at 228C in 50ml of 10 mM Tris-Cl (pH 7.9),
100 mM KCl, 10 mM MgCl2, and 1 mM DTT, so that about 5%of the DNA formed complexes. Heparin (final concentration
25mg ml21) was added, RNAP–promoter complexes wereseparated from unbound DNA and the DNA was eluted from
the gel. DNA fragments were cleaved at the position of A*incorporation by treatment with 1 M piperidine at 908C for
30 min. Fragments containing U were first treated with uracil
DNA glycosylase (UDG; NE Biolabs) and then cleaved withpiperidine. Fragments were analysed by phosphorimaging
(Molecular Dynamics). After electrophoresis on 10% denatur-ing polyacrylamide gels, lanes were normalized to correct for
loading differences and graphed using SigmaPlot (JandelScientific).
In vitro transcription
Reactions contained supercoiled plasmid DNA (20 ng), 10 mM
Tris-Cl (pH 7.9), 10 mM MgCl2, 1 mM DTT, 100mg ml21 ofBSA, 200mM ATP, GTP, and CTP, 10mM UTP, 4mCi [a-32P]-
UTP (NEN) and 10–800 mM KCl or K-glutamate. Transcrip-tion was initiated by addition of RNAP (1–4 nM EsS or Es70)
and terminated by addition of stop solution (Ross et al., 1993)after 15 min at 228C. Samples were electrophoresed on 5.5%
polyacrylamide 7 M urea gels and quantified byphosphorimaging.
Acknowledgements
We thank A. Ernst and G. Verdine (Harvard University) for
providing 7-deaza-7-nitro-adenine (A*), and M. Barker, J.Gralla, J. Helmann, A. Hochschild and R. Hengge-Aronis for
helpful discussions. This work was supported by NationalInstitutes of Health Grant GM37048 to R.L.G., by a Hatch
grant from the US Department of Agriculture to R.L.G. and byNIH grant GM28575 to R.R.B.
References
Albright, R.A., and Matthews, B.W. (1998) How Cro and l
repressor distinguish between operators: the structuralbasis underlying a genetic switch. Proc Natl Acad Sci USA
95: 3431–3436.Arnqvist, A., Olsen, A., and Normark, S. (1994) sS-dependent
growth-phase induction of the csgA promoter in Escherichiacoli can be achieved by s70 in the absence of the nucleoid
associated protein H-NS. Mol Microbiol 13: 1021–1032.Ballesteros, M., Kusano, S., Ishihama, A., and Vicente, M.
(1998) The ftsQ1p gearbox promoter of Escherichia coli is amajor sigma S-dependent promoter in the ddlB-ftsA region.
Mol Microbiol 30: 419–430.Barker, M.M., Gaal, T., and Gourse, R.L. (2001) Mechanism
of regulation of transcription initiation by ppGpp.II. Modelsfor positive control based on properties of RNAP mutants
and competition for RNAP. J Mol Biol 305: 689–702.Barne, K.A., Bown, J.A., Busby, S.J., and Minchin, S.D.
(1997) Region 2.5 of the Escherichia coli RNA polymerase
sigma70 subunit is responsible for the recognition of the‘extended-10’ motif at promoters. EMBO J 16: 4034–4040.
Becker, G., and Hengge-Aronis, R. (2001) What makes anEscherichia coli promoter sS-dependent? Role of the -13/-
14 nucleotide promoter positions and region 2.5 of sS. MolMicrobiol 39: pp.1153–1165.
Becker, G., Klauck, E., and Hengge-Aronis, R. (1999)Regulation of RpoS proteolysis in Escherichia coli: the
response regulator RssB is a recognition factor thatinteracts with the turnover element in RpoS. Proc Natl
Acad Sci USA 96: 6439–6444.Beltrametti, F., Kresse, A.U., and Guzman, C.A. (1999)
Transcriptional regulation of the esp genes of enterohemor-rhagic Escherichia coli. J Bacteriol 181: 3409–3418.
Blackwell, T.K., and Weintraub, H. (1990) Differences andsimilarities in DNA-binding preferences of MyoD and E2A
protein complexes revealed by binding site selection.Science 250: 1104–1110.
Blatter, E.E., Ross, W., Tang, H., Gourse, R.L., and Ebright,R.H. (1994) Domain organization of RNA polymerase alpha
subunit. C-terminal 85 amino acids constitute a domaincapable of dimerization and DNA binding. Cell 78:
889–896.Bohannon, D.E., Connell, N., Keener, J., Tormo, A.,
Espinosa-Urgel, M., Zambrano, M.M., and Kolter, R.(1991) Stationary-phase-inducible ‘gearbox’ promoters:
differential effects of katF mutations and role of sigma 70.J Bacteriol 173: 4482–4492.
Bordes, P., Repoila, F., Kolb, A., and Gutierrez, C. (2000)Involvement of differential efficiency of transcription by EsS
and Es70 RNA polymerase holoenzymes in growth phaseregulation of the Escherichia coli osmE promoter. Mol
Microbiol 35: 845–853.
Bouvier, J., Gordia, S., Kampmann, G., Lange, R., Hengge-
952 T. Gaal et al.
Q 2001 Blackwell Science Ltd, Molecular Microbiology, 42, 939–954

Aronis, R., and Gutierrez, C. (1998) Interplay between
global regulators of Escherichia coli: effect of RpoS, Lrp and
H-NS on transcription of the gene osmC. Mol Microbiol 28:
971–980.
Burr, T., Mitchell, J., Kolb, A., Minchin, S., and Busby, S.
(2000) DNA sequence elements located immediately
upstream of the -10 hexamer in Escherichia coli promoters:
a systematic study. Nucleic Acids Res 28: 1864–1870.
Colland, F., Fujita, N., Kotlarz, D., Bown, J.A., Meares, C.F.,
Ishihama, A., and Kolb, A. (1999) Positioning of sigma (S),
the stationary phase sigma factor, in Escherichia coli RNA
polymerase-promoter open complexes. EMBO J 18:
4049–4059.
Colland, F., Barth, M., Hengge-Aronis, R., and Kolb, A. (2000)
s factor selectivity of Escherichia coli RNA polymerase: role
for CRP, IHF and Lrp transcription factors. EMBO J 19:
3028–3037.
Devchand, P.R., McGhee, J.D., and van de Sande, J.H.
(1993) Uracil-DNA glycosylase as a probe for protein: DNA
interactions. Nucleic Acids Res 21: 3437–3443.
Ding, Q., Kusano, S., Villarejo, M., and Ishihama, A. (1995)
Promoter selectivity control of Escherichia coli RNA
polymerase by ionic strength: differential recognition of
osmoregulated promoters by E sigma D and E sigma S
holoenzymes. Mol Microbiol 16: 649–656.
Espinosa-Urgel, M., Chamizo, C., and Tormo, A. (1996) A
consensus structure for sigma S-dependent promoters
[letter]. Mol Microbiol 21: 657–659.
Estrem, S.T., Gaal, T., Ross, W., and Gourse, R.L. (1998)
Identification of an UP element consensus sequence for
bacterial promoters. Proc Natl Acad Sci USA 95:
9761–9766.
Estrem, S.T., Ross, W., Gaal, T., Chen, Z.W., Niu, W.,
Ebright, R.H., and Gourse, R.L. (1999) Bacterial promoter
architecture: subsite structure of UP elements and
interactions with the carboxy-terminal domain of the RNA
polymerase alpha subunit. Genes Dev 13: 2134–2147.
Fenton, M.S., Lee, S.J., and Gralla, J.D. (2000) Escherichia
coli promoter opening and 210 recognition: mutational
analysis of s70. EMBO J 19: 1130–1137.
Gaal, T., Bartlett, M.S., Ross, W., Turnbough, C.L. Jr and
Gourse, R.L. (1997) Transcription regulation by initiating
NTP concentration. rRNA synthesis in bacteria. Science
278: 2092–2097.
Gross, C.A., Chan, C., Dombroski, A., Gruber, T., Sharp, M.,
Tupy, J., and Young, B. (1998) The functional and
regulatory roles of sigma factors in transcription. Cold
Spring Harb Symp Quant Biol 63: 141–155.
Harrison, S.C., and Aggarwal, A.K. (1990) DNA recognition by
proteins with the helix-turn-helix motif. Annu Rev Biochem
59: 933–969.
Hengge-Aronis, R. (1996a) Back to log phase: sigma S as a
global regulator in the osmotic control of gene expression in
Escherichia coli. Mol Microbiol 21: 887–893.
Hengge-Aronis, R. (1996b) Regulation of gene expression
during entry into stationary phase. In Escherichia Coli and
Salmonella Cellular and Molecular Biology, Vol 1.
Neidhardt, F.C. (ed.). Washington, DC: American Society
for Microbiology Press, pp. 1497–1512.
Hengge-Aronis, R., Klein, W., Lange, R., Rimmele, M., and
Boos, W. (1991) Trehalose synthesis genes are controlled
by the putative sigma factor encoded by rpoS and are
involved in stationary-phase thermotolerance in Escher-
ichia coli. J Bacteriol 173: 7918–7924.
Hochschild, A., Douhan, J., and Ptashne, M. (1986) How l
Repressor and l Cro distinguish between OR1 and OR3.
Cell 47: 807–816.
Ishihama, A. (2000) Functional Modulation of Escherichia coli
RNA polymerase. Annu Rev Microbiol 54: 499–518.
Jishage, M., Iwata, A., Ueda, S., and Ishihama, A. (1996)
Regulation of RNA polymerase s subunit synthesis in
Escherichia coli. Intracellular levels of four species of sigma
subunit under various growth conditions. J Bacteriol 178:
5447–5451.
Josaitis, C.A., Gaal, T., Ross, W., and Gourse, R.L. (1990)
Sequences upstream of the-35 hexamer of rrnB P1 affect
promoter strength and upstream activation. Biochim
Biophys Acta 1050: 307–311.
Kolb, A., Kotlarz, D., Kusano, S., and Ishihama, A. (1995)
Selectivity of the Escherichia coli RNA polymerase Es38 for
overlapping promoters and ability to support CRP acti-
vation. Nucleic Acids Res 23: 819–826.
Kusano, S., and Ishihama, A. (1997) Stimulatory effect of
trehalose on formation and activity of Escherichia coli RNA
polymerase Es38 holoenzyme. J Bacteriol 179:
3649–3654.
Kusano, S., Ding, Q., Fujita, N., and Ishihama, A. (1996)
Promoter selectivity of Escherichia coli RNA polymerase
Es70 and Es38 holoenzymes. Effect of DNA supercoiling. J
Biol Chem 271: 1998–2004.
Lange, R., and Hengge-Aronis, R. (1991) Growth phase-
regulated expression of bolA and morphology of stationary-
phase Escherichia coli cells are controlled by the novel s
factor sS. J Bacteriol 173: 4474–4481.
Lee, S.J., and Gralla, J.D. (2001) Sigma 38 (rpoS ) RNA
polymerase promoter engagement via 210 region nucleo-
tides. J Biol Chem 276: 30064–30071.
Leirmo, S., Harrison, C., Cayley, D.S., Burgess, R.R., and
Record, M.T., Jr. (1987) Replacement of potassium
chloride by potassium glutamate dramatically enhances
protein–DNA interactions in vitro. Biochemistry 26:
2095–2101.
Lonetto, M., Gribskov, M., and Gross, C.A. (1992) The sigma
70 family. sequence conservation and evolutionary
relationships. J Bacteriol 174: 3843–3849.
Malhotra, A., Severinova, E., and Darst, S.A. (1996) Crystal
structure of the s70 subunit fragment from E. coli RNA
polymerase. Cell 87: 127–136.
Matlock, D.L., and Heyduk, T. (2000) Sequence determinants
for the recognition of the fork junction DNA containing the
210 region of promoter DNA by E. coli RNA polymerase.
Biochemistry 39: 12274–12283.
Min, C., Cushing, T.D., and Verdine, G.L. (1996) Template-
directed interference footprinting of protein-adenine con-
tacts. JACS 118: 6116–6120.
Mulvey, M.R., and Loewen, P.C. (1989) Nucleotide sequence
of katF of Escherichia coli suggest that KatF protein is a
novel sigma transcription factor. Nucleic Acids Res 17:
9979–9991.
Nguyen, L.H., and Burgess, R.R. (1997) Comparative
analysis of the interactions of Escherichia coli sigma S
and sigma 70 RNA polymerase holoenzyme with the
Promoter recognition and discrimination by Ess RNAP 953
Q 2001 Blackwell Science Ltd, Molecular Microbiology, 42, 939–954

stationary-phase-specific bolAp1 promoter. Biochemistry36: 1748–1754.
Nguyen, L.H., Jensen, D.B., Thompson, N.E., Gentry, D.R.,and Burgess, R.R. (1993) In vitro functional characteriz-
ation of overproduced Escherichia coli katF/rpoS geneproduct. Biochemistry 32: 11112–11117.
Oliphant, A.R., and Struhl, K. (1988) Defining the consensussequences of E. coli promoter elements by random
selection. Nucleic Acids Res 16: 7673–7683.Pollock, R., and Treisman, R. (1990) A. sensitive method for
the determination of protein-DNA binding specificities.Nucleic Acids Res 18: 6197–6204.
Rao, L., Ross, W., Appleman, J.A., Gaal, T., Leirmo, S.,Schlax, P.J., et al. (1994) Factor independent activation of
rrnB P1: An ‘extended’ promoter with an upstream elementthat dramatically increases promoter strength. J Mol Biol
235: 1421–1435.Reznikoff, W.S. (1976) Formation of the RNA polymerase-lac
promoter open complex. In RNA Polymerase. Losick, R.,
and Chamberlin, M. (eds). Cold Spring Harbor, NY: ColdSpring Harbor Laboratory Press, pp. 441–454.
Ross, W., Thompson, J.F., Newlands, J.T., and Gourse, R.L.(1990) E. coli Fis protein activates ribosomal RNA
transcription in vitro and in vivo. EMBO J 9: 3733–3742.Ross, W., Gosink, K.K., Salomon, J., Igarashi, K., Zou, C.,
Ishihama, A., et al. (1993) A third recognition element inbacterial promoters. DNA binding by the alpha subunit of
RNA polymerase. Science 262: 1407–1413.Ross, W., Aiyar, S.E., Salomon, J., and Gourse, R.L. (1998)
Escherichia coli promoters with UP elements of differentstrengths: modular structure of bacterial promoters. J
Bacteriol 180: 5375–5383.Ross, W., Ernst, A., and Gourse, R.L. (2001) Fine structure of
E. coli RNA polymerase–promoter interactions: a subunitbinding to the UP element minor groove. Genes & Dev 15:
491–506.Simons, R.W., Houman, F., and Kleckner, N. (1987) Improved
single and multicopy lac-based cloning vectors for proteinand operon fusions. Gene 53: 85–96.
Suh, S.J., Silo-Suh, L., Woods, D.E., Hassett, D.J., West,S.E., and Ohman, D.E. (1999) Effect of rpoS mutation on
the stress response and expression of virulence factors inPseudomonas aeruginosa. J Bacteriol 181: 3890–3897.
Tan, M., Gaal, T., Gourse, R.L., and Engel, J.N. (1998)Mutational analysis of the Chlamydia trachomatis rRNA P1
promoter defines four regions important for transcription invitro. J Bacteriol 180: 2359–2366.
Tanaka, K., Takayanagi, Y., Fujita, N., Ishihama, A., andTakahashi, H. (1993) Heterogeneity of the principal s factor
in Escherichia coli: the rpoS gene product, s38, is a secondprincipal s factor of RNA polymerase in stationary-phase
Escherichia coli. Proc Natl Acad Sci USA 90: 3511–3515.Tanaka, K., Kusano, S., Fujita, N., Ishihama, A., and
Takahashi, H. (1995) Promoter determinants for Escher-ichia coli RNA polymerase holoenzyme containing s38 (the
rpoS gene product). Nucleic Acids Res 23: 827–834.Tuerk, C., and Gold, L. (1990) Systematic evolution of ligands
by exponential enrichment. RNA ligands to bacteriophage
T4 DNA polymerase. Science 249: 505–510.Vicente, M., Kushner, S.R., Garrido, T., and Aldea, M. (1991)
The role of the ‘gearbox’ in the transcription of essentialgenes. Mol Microbiol 5: 2085–2091.
Voskuil, M.I., Voepel, K., and Chambliss, G.H. (1995) The216 region, a vital sequence for the utilization of a promoter
in Bacillus subtilis and Escherichia coli. Mol Microbiol 17:271–279.
Wise, A., Brems, R., Ramakrishnan, V., and Villarejo, M.(1996) Sequences in the 235 region of Escherichia coli
rpoS-dependent genes promote transcription by E sigma S.J Bacteriol 178: 2785–2793.
Wright, W.E., Binder, M., and Funk, W. (1991) Cyclicamplification and selection of targets (CASTing) for the
myogenin consensus binding site. Mol Cell Biol 11:4104–4110.
Xu, J., and Johnson, R.C. (1995) Identification of genesnegatively regulated by Fis: Fis and RpoS comodulate
growth-phase-dependent gene expression in Escherichiacoli. J Bacteriol 177: 938–947.
954 T. Gaal et al.
Q 2001 Blackwell Science Ltd, Molecular Microbiology, 42, 939–954