proliferation of miss-forming microbial mats after the late neoproterozoic glaciations: evidence...
TRANSCRIPT

Pg
Za
b
c
a
ARRAA
KMsNTN
1
m2ttaeelzMGt
tT
0h
Precambrian Research 224 (2013) 529– 550
Contents lists available at SciVerse ScienceDirect
Precambrian Research
journa l h omepa g e: www.elsev ier .com/ locate /precamres
roliferation of MISS-forming microbial mats after the late Neoproterozoiclaciations: Evidence from the Kimberley region, NW Australia
hong-Wu Lana,c,∗, Zhong-Qiang Chenb
State Key Laboratory of Lithospheric Evolution, Institute of Geology and Geophysics, Chinese Academy of Sciences, Beijing 100029, ChinaState Key Laboratory of Biogeology and Environmental Geology, China University of Geosciences (Wuhan), Wuhan 430074, ChinaKey Lab of Petroleum Resources Research, Institute of Geology and Geophysics, Chinese Academy of Sciences, Beijing 100029, China
r t i c l e i n f o
rticle history:eceived 9 July 2012eceived in revised form 12 October 2012ccepted 4 November 2012vailable online 15 November 2012
eywords:ISS (microbially induced sedimentary
tructures)eoproterozoic glaciationshe Kimberley
a b s t r a c t
A total of 20 morphological types of microbially induced sedimentary structures (MISS) are recognizedfrom the late Neoproterozoic tillite-bearing successions of the Kimberley, northwestern Australia whichinclude alpha-petees, triradiate cracks, multidirectional linear ridges, millimeter ripples, erosional rem-nants and pockets and so on. Of these, triradiate cracks, multidirectional linear ridges, and millimeterripples are reported for the first time from the Neoproterozoic worldwide. Most Kimberley MISS aremorphologically comparable with their ancient and present-day counterparts whose biogenicity havebeen well demonstrated.
In the Kimberley, during the interglacial period MISS have colonization preference to relativelycoarse sandy substrates (Ranford Formation) and their abundance decreased towards the younger strata(Throssell Shale). In contrast, MISS abundance increased from the older strata (Yurabi Formation) towards
orthwestern Australia the younger strata (Flat Rock Formation) during the postglacial period. However, overall, both MISS abun-dance and bedding plane coverage percentage increased from the Marinoan-younger Marinoan/Gaskiersinterglacial successions to younger Marinoan/Gaskiers postglacial successions, indicating a proliferationof microbial mats after the younger Marinoan/Gaskiers glaciation. The increase of microbial mats couldhave ameliorated marine biogeochemical conditions and ecosystems, and thus laid the foundation forthe rise of Ediacaran biota in Australian basins.
. Introduction
The earth underwent repeated glaciations between 750 and 580illions years ago (Fairchild and Kennedy, 2007; Hoffman and Li,
009), the younger two of which are Marinoan and Gaskiers glacia-ions. Neoproterozoic glaciations were believed to have changedhe chemical and oxygenic conditions in oceans, and thus have setn agenda for the rise of the well-known Ediacaran biota (Knollt al., 2004; Kaufman et al., 2007; Shen et al., 2008). However, thexact driving forces for the emergence of the Ediacaran biota haveong been disputed (McCall, 2006), although the late Neoprotero-oic ocean oxygenation (Grey and Calver, 2007a; Fike et al., 2007;cFadden et al., 2008) or asteroid impact (e.g. Acraman impact,
rey et al., 2003; Gostin et al., 2010) are believed to have facilitatedhe diversification of the Ediacaran biota.
∗ Corresponding author at: State Key Laboratory of Lithospheric Evolution, Insti-ute of Geology and Geophysics, Chinese Academy of Sciences, Beijing 100029, China.el.: +86 10 82998445; fax: +86 10 62010846.
E-mail address: [email protected] (Z.-W. Lan).
301-9268/$ – see front matter © 2012 Elsevier B.V. All rights reserved.ttp://dx.doi.org/10.1016/j.precamres.2012.11.008
© 2012 Elsevier B.V. All rights reserved.
Within marine ecosystems microbes were responsible for pro-ducing energy and nutrition for other animals, and mediatinggeochemical conditions in oceans (Ehrlich, 1998; Madigan et al.,2009). Their abundance and ecologic activities directly affected theprimary productivity of marine ecosystems and compositions ofconsuming communities (Berglund et al., 2007; Solic et al., 2010).From this perspective, geomicrobiological study of microbial com-munities during and after glaciations is crucial for understandingthe evolution of the Ediacaran ecosystem and the possible linkbetween marine ecosystem changes and rise of Ediacaran biota.As one of the most important fossil records of microbes in Pre-cambrian successions, microbially induced sedimentary structures(MISS) are common in Proterozoic siliciclastic successions (Noffkeet al., 2002; Schieber, 2007), and thus would provide necessaryinformation relevant with shallow marine microbial communities.
In Australia, the Ediacaran MISS and biota have been docu-mented from the late Neoproterozoic successions not only incentral and South Australia (Bland, 1984; Logan et al., 1999;
McCall, 2006; Willman et al., 2006; Maloof et al., 2010), but alsoin the Kimberley region, northwestern Australia (Lan and Chen,2012a,b). In the Kimberley, the Neoproterozoic succession is upto 5300 m thick and dominated by the shallow marine siliciclastic
5 rian R
rers
oiidTpeueTabaica
2K
l∼icNs
F2M
30 Z.-W. Lan, Z.-Q. Chen / Precamb
ocks sandwiched with Marinoan and younger Marinoan glaciationquivalent tillites capped with carbonate rocks which are well cor-elated with the Marinoan and younger Marinoan/Gaskiers glacialuccessions in central and South Australia (Corkeron, 2007, 2008).
Aside from the Yurabi Formation (Lan and Chen, 2012a),ther inter- and postglacial successions yield also abundant MISSn the Kimberley. Preliminary investigation suggests that thesenterglacial and postglacial MISS have different associations, abun-ance, and bedding plane coverage density from one another.he MISS abundance changes from the interglacial to postglacialeriods therefore may indicate a change of the microbe-dominatedcosystems to some extent. The Kimberley MISS therefore aid innderstanding the evolutionary traits of the microbe-dominatedcosystems between and after the late Neoproterozoic glaciations.he aims of this paper are to describe a suit of MISS from the inter-nd postglacial successions in the Kimberley, and evaluate micro-ial changes in the ocean during this critical period using MISSbundance and bedding plane coverage percentage changes dur-ng the inter- and postglacial periods. We also discuss in a broadontext the possible relationship between microbial developmentnd rise of Ediacaran biota in Australia.
. Geological and MISS-bearing stratigraphic settings of theimberley region
Proterozoic successions are well exposed in the Kimber-ey region, northwestern Australia that occupies an area of300,000 km2 (Fig. 1a). The Proterozoic successions are divided
nto the folded rocks of the Kimberley Basin succession and adja-ent volcanics, granitoids and sedimentary rocks (Griffin, 1989).eoproterozoic strata unconformably overlie the Kimberley Basin
uccession and are unconformably overlain by the Cambrian
ig. 1. (a) Geological map showing the distribution of Neoproterozoic rocks in the Kimber000; Tyler, 2004). (b) Exposure of the Mount House Group in the Mount House area (afount Ramsay area (after Corkeron, 2007). (d) Exposure of the Duerdin and Albert Edwa
esearch 224 (2013) 529– 550
Antrim Plateau Volcanics (Hanley and Wingate, 2000). In the Kim-berley the Neoproterozoic rocks are mainly distributed in theMount House, Mount Ramsay and east Kimberley areas (Fig. 1a) andcomprise five major groups namely the Mount House, Kuniandi,Louisa Downs, Duerdin, and Albert Edward Groups. The first groupis confined to the Mount House area and characterized by the WalshTillite overlain by a sandstone-mudstone succession (Griffin et al.,1993; Fig. 1b). The Kuniandi and Louisa Downs Groups contain theLandrigan Tillite and Egan Formation, respectively and other sili-ciclastic units; they are mainly exposed along the O’Donnell andLouisa Synclines in the Mount Ramsay area (Fig. 1c). The Duerdinand Albert Edward Groups comprise the Neoproterozoic rocksexposed along the eastern margin of the Halls Creek Orogen inthe east Kimberley (Fig. 1d). The former group is typified by thepresence of the Fargoo and Moonlight Valley Tillites and other sili-ciclastic units, whereas the latter is characterized by the siliciclasticsuccessions interbedded with dolomite.
To date, two schemes have been proposed for the correlationsof the Neoproterozoic units in the Kimberley (Dow and Gemuts,1969; Coats and Preiss, 1980; Corkeron, 2007, 2008). Corkeron’s(2007, 2008) stratigraphic correlation scheme is followed hereinin that the Walsh, Landrigan, and Fargoo-Moonlight Valley tilliteunits are correlative with the Marinoan glaciation equivalents ofthe Olympic Formation in central Australia, and the Elatina Forma-tion in South Australia, whereas the tillite-bearing Egan Formationand the Boonall Dolomite are correlated with the younger Marinoanor possible Gaskiers glaciation equivalents of the Julie Formation ofcentral Australia and Wonoka Formation of South Australia (Greyand Corkeron, 1998; Table 1). The 17O record from the late Neo-
proterozoic tillites and cap carbonates of Kimberley reveals thatthe Moonlight Valley tillite represents the sedimentation of theMarinoan glaciations, supporting the “snowball earth” hypothesis,whereas the Egan tillite represents the deposition of a differentley region (modified from Griffin et al., 1993; Tyler et al., 1998; Hanley and Wingate,ter Corkeron, 2003). (c) Exposure of the Kuniandi and Louisa Downs Groups in therd Groups in the east Kimberley (after Corkeron, 2008).

Z.-W. Lan, Z.-Q. Chen / Precambrian Research 224 (2013) 529– 550 531
Table 1Neoproterozoic stratigraphic units and correlations with their counterparts in South and central Australia (modified from Grey and Corkeron,1998). The light grey units are glacial deposits capped by carbonates rocks. The dark grey units suggest correlation using stromatolitebiostratigraphy.
gpVa(5
lemBFGRSrpTaFoF(uatcS
Kws1r
laciation in the Kimberley, suggestive of the younger Marinoan orossible Gaskiers glaciations (Bao et al., 2012). The Antrim Plateauolcanics capping the Neoproterozoic rocks in the Mount Ramsaynd the east Kimberley areas gave a zircon U–Pb age of 513 ± 12 MaHanley and Wingate, 2000) and 40Ar/39Ar ages of 505 ± 2 Ma to08 ± 2 Ma (Glass and Phillips, 2006).
The interglacial successions between the Fargoo/Moonlight Val-ey or Landrigan or Walsh Tillites and Egan Formation and itsquivalents include the Traine, Throssell Shale, and Estaughs For-ations of the Mount House Group, the Stein, Wirara, and Mt
ertram Sandstone Formations of the Kuniandi Group, and therank River Sandstone, and Ranford Formation of the Duerdinroup (Gehling and Droser, 2009; Table 1). Among these, only theanford Formation and Throssell Shale yield MISS. The Throssellhale is equivalent to the upper Ranford Formation and thus rep-esents the interglacial successions (Gellatly et al., 1975). Theostglacial successions include the Yurabi Formation, McAlly Shale,ean Formation and Lubbock Formation of the Louisa Downs Group,nd the Boonall Dolomite, Timperley Shale, Nyuless Sandstone andlat Rock Formation of the Albert Edward Group (Table 1). Of these,nly the Yurabi and Flat Rock Formations yield MISS. The Yurabiormation is equivalent to the lower part of the Timperley ShaleGrey and Corkeron, 1998; Lan and Chen, 2012b; Table 1). The laternit is stratigraphically positioned below the Flat Rock Formationt the Palm Spring section (Corkeron, 2008). The Yurabi Forma-ion therefore is older than the Flat Rock Formation. The latter wasorrelated with the Ediacaran fossil-bearing Rawnsley Quartzite inouth Australia (Grey and Calver, 2007b).
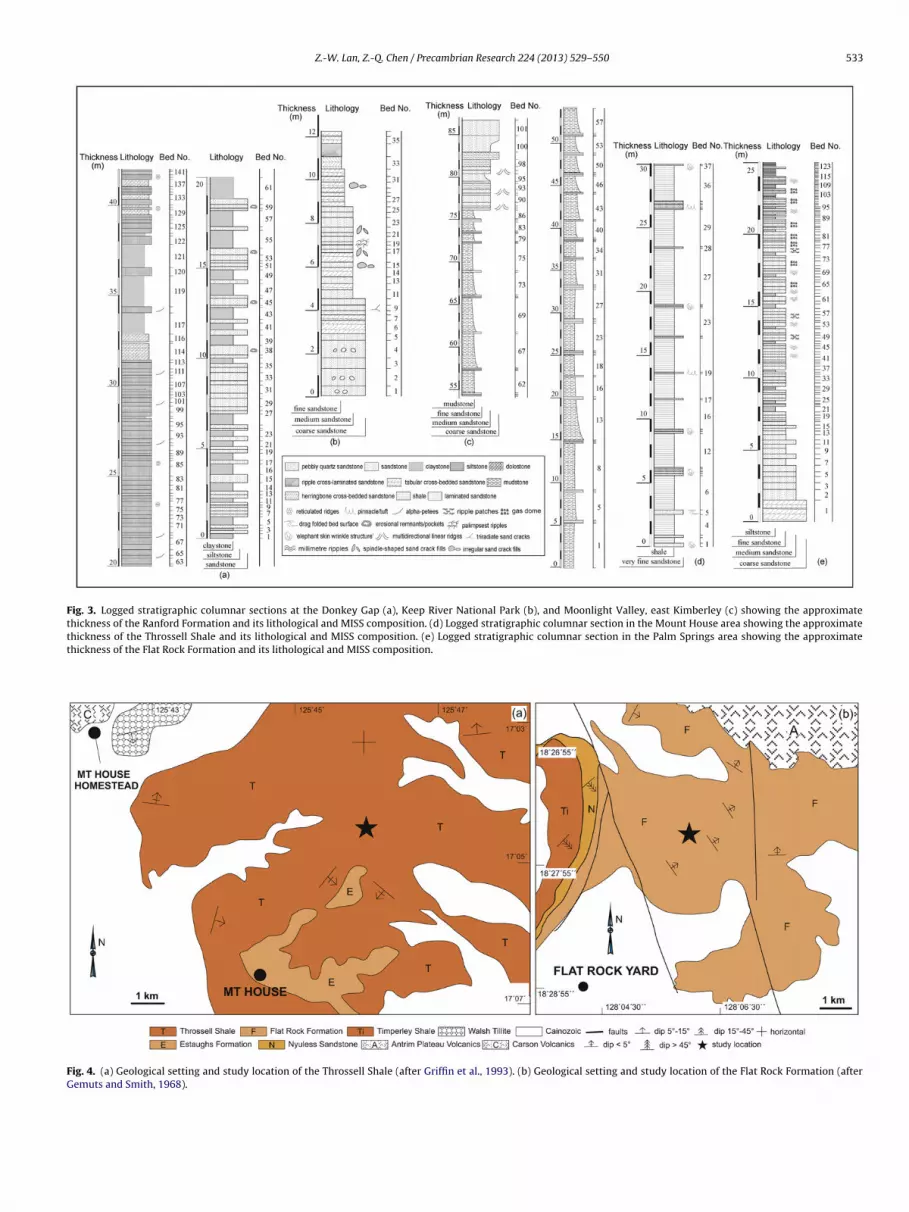
The Ranford Formation was studied at the Donkey Gap, theeep River National Park, and the Moonlight Valley sections,
hereas the Throssell Shale was studied only at the Mount Houseection (Fig. 2a). The Donkey Gap section (GPS: 16◦04′33.4′′S,28◦57′53.4′′E) is located near the Western Australia-Northern Ter-itory border. Therein, the Ranford Formation and Moonlight Valley
Tillite of the Duerdin Group are exposed (Fig. 2b). The upper Ran-ford Formation, assigned to the Jonny Cake Shale Member (Table 1;Grey and Calver, 2007b), is 42 m thick and dominated by finesandstone and clay at the lower part (Fig. 3a, Beds 1–61), andlaminated siltstone, clay and dolomite at the upper part (Fig. 3a,Beds 62–141). MISS are present mainly on fine sandstones andsiltstones, including erosional remnants, erosional pockets, alpha-petees, and reticulated ridges. The MISS-bearing strata are 32 mthick. Accumulative thickness of MISS-bearing single beds is up to3.4 m (Table 2).
At the studied location of the Keep River National Park (GPS:15◦53′20′′S, 129◦05′00′′E), the exposed Duerdin Group includes theBeasly Knob Member of the Ranford Formation and Moonlight Val-ley Formation (Fig. 2c). Of these, the Beasly Knob Member, 12.3 mthick, comprises massive sandstone. The Beasley Knob Memberis equivalent to the Jarrad Sandstone Member of the lower Ran-ford Formation (Dunster et al., 2000). MISS are mainly present onthe upper surface of Beasley Knob Member sandstone bed at a7-m-thick interval which include spindle-shaped sand crack fills,palimpsest ripples, irregular sand crack fills, and triradiate cracks.Accumulative thickness of single MISS-bearing beds is up to 0.85 m(Table 2).
At the Moonlight Light Valley section (GPS: 17◦05′46.2′′S,128◦20′54.2′′E; Fig. 2d), the Jarrad Sandstone Member of theRanford Formation conformably overlies the Moonlight Valley For-mation. The lower Jarrad Sandstone Member is dominated by asuit of cyclic mudstone and laminated fine sandstone, whereas theupper part of the member is characterized by thick-bedded quartzsandstone (Fig. 3c). MISS are mainly present on sandstone bed topswith an interval of 11.8 m and are dominated by multidirectional
linear ridges. Accumulative thickness of single MISS-bearing bedsis up to 1.3 m (Table 2).The Mount House section (GPS: 17◦04′29′′S, 125◦45′58′′E) islocated about 8 km southeast of the Mount House Homestead, and

532 Z.-W. Lan, Z.-Q. Chen / Precambrian Research 224 (2013) 529– 550
F in theG e after
aTirootMs
bsFM2(r
3
uiergotsowi
ig. 2. Distribution of the Neoproterozoic Duerdin Group and Albert Edward Groupap (b), Keep River National Park (c) and Moonlight Valley (d). (a), (b), (c), and (d) ar
espectively.
bout 6 km northeast of the Mount House (Fig. 4a). Therein, thehrossell Shale is dominated by shale with minor fine sandstonenterbeds (Fig. 3d) and is about 100 m thick. The Throssell Shale cor-elates with the Jonny Cake Shale Member of the Ranford Formationf the east Kimberley region (Table 1; Grey and Calver, 2007b). MISSccur mainly on a 30 m interval of fine sandstone beds. Accumula-ive thickness of single MISS-bearing bed is up to 2.5 m (Table 2).
ISS from this locality are represented by ‘elephant skin’ wrinkletructure, pinnacles/tufts and drag folded bed surface.
MISS from the Yurabi Formation have been described in detaily Lan and Chen (2012a) and thus will not be repeated herein. Thetudied location (GPS: 18◦27′40′′S, 128◦06′01.5′′E) of the Flat Rockormation is about 6 km northwest of the Flat Rock Yard (Fig. 4b).ISS are preserved on fine sandstone bed tops with an interval of
5 m (Fig. 3e). Of these, the MISS-bearing succession is 5 m thickTable 2). MISS include gas domes, ripple patches and millimetreipples.
. Methodology
Thin sections were prepared by cutting samples perpendic-lar to the MISS-bearing bedding plane surface so as to detect
nternal microstructures under the Nikon Eclipse E800 Microscopequipped with a Nikon DS-Fi 1 camera using transmitted andeflected lights. This is because microtextures such as cementedrains and diagenetic pyrite or carbonated laminae can only bebserved in cross sections. The Zeiss 1555 VP-FESEM was utilizedo detect microbial organics or diagenetic minerals of micron to
ubmicron meters. The SEM was specifically manipulated to get anptimal resolution at 50,000–200,000. It was tuned to an optimalorking distance of 6–12 mm and a voltage of 10–20 kV. Exam-nation of the biogenicity of MISS in the present paper followed
east Kimberley region (a) and study locations of the Ranford Formation at Donkeyr Corkeron (2003), Thorne et al. (1999), Dunster et al. (2000), and Tyler et al. (1998),
the six steps of the biogenicity described by Noffke (2009). Rela-tive abundance of MISS assemblages is evaluated based on theirbedding plane coverage percentage and accumulative thickness ofsingle MISS-bearing beds, as described by Lan and Chen (2012a).Statistics of inter- and postglacial MISS abundance was conductedto assess the effect of microbial ecosystem changes on the rise ofEdiacaran biota in Australia.
4. MISS description
Overall, twenty different kinds of MISS are described in thispaper. They are alpha-petees, reticulated ridges, ‘elephant skin’wrinkle structures, millimeter ripples, spindle-shaped sand crackfills, palimpsest ripples, irregular sand crack fills, triradiate cracks,multidirectional linear ridges, drag folded bed surface, pyrite con-centrated laminae, micrite laminae, gas domes, worm-like cracks,polygonal cracks, bamboo leaf-like cracks, erosional remnants andpockets, ripple patches, pinnacles/tufts, and oriented grains.
4.1. Alpha-petees
Smooth linear ridges, preserved as positive epirelief on siltstonebed tops, are 4–10 cm long and 0.5–1 cm wide (Fig. 5a and b). Indi-vidual ridges are straight to slightly curved, nearly parallel to oneanother, and relatively consistent in diameter. Inter-ridge distancesvary from 4 cm to 14 cm. All the ridges have a smooth surface andare sub-elliptical in cross section (Fig. 5c and d). The ridged area iscomposed mainly of quartz cement, with several dark clay laminae
(Fig. 5c and d). In contrast, the non-ridged area is dominated by clayminerals with subsidiary fine quartz grains.These materials presented here are morphologically similar tothe structures documented by Noffke et al. (2002) from Harughas
Z.-W. Lan, Z.-Q. Chen / Precambrian Research 224 (2013) 529– 550 533
Fig. 3. Logged stratigraphic columnar sections at the Donkey Gap (a), Keep River National Park (b), and Moonlight Valley, east Kimberley (c) showing the approximatethickness of the Ranford Formation and its lithological and MISS composition. (d) Logged stratigraphic columnar section in the Mount House area showing the approximatethickness of the Throssell Shale and its lithological and MISS composition. (e) Logged stratigraphic columnar section in the Palm Springs area showing the approximatethickness of the Flat Rock Formation and its lithological and MISS composition.
Fig. 4. (a) Geological setting and study location of the Throssell Shale (after Griffin et al., 1993). (b) Geological setting and study location of the Flat Rock Formation (afterGemuts and Smith, 1968).

534 Z.-W. Lan, Z.-Q. Chen / Precambrian Research 224 (2013) 529– 550
Table 2Statistics of the number of the MISS-bearing beds and their thickness of the studied Ranford Formation atthe Donkey Gap, Keep River National Park, and Moonlight Valley, the Throssell Shale near the Mount Housestation, the Yurabi Formation at the Egan Range and the Flat Rock Formation near the Flat Rock Yard.
Formation Bed No. Bed thickness (cm) Formation Bed No. Bed thickness (cm)
Ran
ford
For
mat
ion
at D
onke
y G
ap
38 62
Thr
osse
ll Sh
ale
Form
atio
n
1 4045 33 5 4054 30 7 1559 20 8 1064 10 9 868 8 10 876 30 11 785 47 13 1291 12 14 8100 18 15 7110 15 19 7118 38 24 8130 10 25 7139 7 26 7Total 340 30 7
Ran
ford
For
mat
ion
at K
eep
Riv
er
Nat
iona
l Par
k
9 15 31 1015 13 32 1517 10 33 819 5 34 1022 27 35 830 15 37 8Total 85 Total 250
Ran
ford
at
Moo
nlig
ht
Val
ley
88 23
Flat
Roc
k Fo
rmat
ion
42 892 55 45 2296 52 49 27
Total 130 53 28
Yur
abi F
orm
atio
n
13 26 56 2018 24 61 4529 20 63 2231 58 65 5033 42 69 1542 26 72 5346 12 76 4449 20 78 3361 17 81 3563 15 86 3879 17 93 2584 23 99 15Total 300 105 8
aatbpPvtp(tbocq
nd Bullsport, and thus can be assigned to alpha-petees or its vari-nts. Noffke et al. (2002) ascribed the formation of alpha-peteeso thick microbial mats. A similar microbial mat genesis has alsoeen proposed by Banerjee and Jeevankumar (2005) based on theresence of silicified bacterial filaments in alpha-petees from thealaeoproterozoic Koldaha Shale, central India. Actualistic obser-ations from supratidal pool at the Redfish Bay, Texas, USA suggesthat filamentous bacterial growth could induce a series of sub-arallel linear ridges on the top surface of siliciclastic sedimentHagadorn and Bottjer, 1997, 1999). As a result, the Ranford Forma-ion linear ridges could indicate the pre-existence of filamentous
acterial colonies in the unconsolidated sandy sediments. Therganic bacterial filaments were lithified and replaced with quartzement during subsequent diagenesis as evidenced by the higheruartz content in the ridged areas (Konhauser and Urrutia, 1999;114 12Total 500
Briggs, 2003; Callow and Brasier, 2009). In situ growth experimentsalso show that microbe can be interacted with silica during whichmicrobes play a significant role in precipitating and binding silica ontheir cell surface via extracellular polymeric substances (Handleyet al., 2008; Orange et al., 2009). As such, the quartz cement couldrepresent silicified bacteria. The smooth surface of the linear ridgescould be ascribed to the wave or current erosion after their forma-tion. The wind-induced ‘wrinkle marks’ described by Robb (1992)could have the same origin as the alpha-petees documented here.
4.2. Reticulated ridges
Wrinkle structures in the form of reticulated ridges are pre-served as positive relief on the upper surface of siltstone beds(Fig. 6a and b). Individual wrinkle structures are 0.4–1.5 cm wide

Z.-W. Lan, Z.-Q. Chen / Precambrian Research 224 (2013) 529– 550 535
Fig. 5. (a) Photograph of linear ridges preserved as positive epirelief on sandstone bed tops. (b) Close-up of a showing two near parallel linear ridges on sandstone bed tops.(c) Amalgamated thin section view of the cross section of linear ridges showing the elliptical area is dominated by quartz cement with minor presence of dark laminae. (d)S
aToa0e(
i(osmraCirbfiptr
chematic line drawing of (c).
nd surrounded by irregular, sharp to flat, subrounded margins.he weaving of individual ridges comprises reticulated morphol-gy. The wrinkled area has a sharp margin (Fig. 6a, indicated by therrow), which makes it possible to estimate a thickness of about.2 cm for the wrinkled surface. Thin section view shows the pres-nce of wavy dark lamina on the upper part of the wrinkled surfaceFig. 6c).
These Ranford wrinkle structures could not be formed in phys-cal dewatering processes due to absence of dewatering channelsBouougri and Porada, 2002). The absence of sandstone remnantsn the bed surface and sand-mud couplets in the host rock alsouggests that the reticulated ridges could not be purely physicaliniature load structures (Allen, 1985). As such, the reticulated
idges should have been formed through microbial mat growth,s interpreted elsewhere (e.g. Kumar and Pandey, 2007; Lan andhen, 2012b). Upon diagenesis microbial mats were degraded
n the anoxic zone with involvement of decay inducing bacte-ia (e.g. sulphate reducing bacteria) and were thus accompaniedy the simultaneous formation of fine grain carbonate and pore-
lling argillaceous material (Seilacher, 1999; Kühl et al., 2003). Theresence of thin dark laminae in the Ranford wrinkle structuresherefore should be associated with fossilized microbial mats aseported elsewhere (Kühl et al., 2003).4.3. ‘Elephant skin’ wrinkle structures
‘Elephant skin’ wrinkle structures comprises elongate to sub-rounded bulges (Figs. 7a and b and 8a and b) separated by narrowcontinuous or discontinuous grooves, or rectangular elongatesseparated by narrow grooves preserved on bed soles (Fig. 8c andd). The elongate to subrounded bulges are commonly 0.3–0.6 cmlong, 0.2–0.5 cm wide, preserved as convex epirelief on the uppersurface of fine sandstone beds. Sinuous or sigmoidal surface of thebulges is covered by a dark thin layer clay lamina in cross section(Fig. 7c and e). Beneath the dark clay lamina are sporadicallydistributed pyrite grains which preferentially concentrated at thelower relief areas (see circled areas in Fig. 7c). SEM images showthat pyrites are euhedral with a size range of 7–20 �m (Fig. 7f andg), and geochemically dominated by Fe and S. Individual rectangu-lar elongates are 1–6 cm long, 0.2–0.6 cm wide. Images of slab cutperpendicular to the bedding surface show that the ‘elephant skin’wrinkle structures-bearing bed (Fig. 8c) is composed of massive,fine sandstone (Fig. 8e).
Absence of typical sand-over-mud doublets suggests theThrossell Shale ‘elephant skin’ wrinkle structures could not havebeen produced through soft-sediment loading deformation (Allen,1985). Euhedral morphology of pyrite suggests that they could not

536 Z.-W. Lan, Z.-Q. Chen / Precambrian Research 224 (2013) 529– 550
F urfacet resen
bbtott1bamiMaaoslts2
4
utt
ig. 6. (a) Field photo showing reticulated ridges on the upper sandstone bedding shin section view of the cross section of reticulated ridges structures showing the p
e detrital grains transported from other places, but must haveeen formed in situ (Guy et al., 2010). As such, the concentra-ion of pyrite grains in the concave areas indicates the decay ofrganic layers during diagenesis. The ‘elephant skin’ wrinkle struc-ures therefore should represent microbial mat surface featureshat have been induced by microbial mats (Hagadorn and Bottjer,997, 1999; Gerdes et al., 2000). The microbial mat was then loadedy freshly deposited sediments which brought a loading pressurend caused the deformation of the underlying cohesive microbialat surface, thus forming the wrinkle structures on the overly-
ng bed soles (Noffke et al., 2002; Bottjer and Hagadorn, 2007).icrobial mat acted as a cohesive veneer between the overlying
nd underlying sandstone beds and prohibited them from beingggregated (Bottjer and Hagadorn, 2007). During diagenesis, therganic microbial mats were degraded with the involvement ofulphate reducing bacteria and replaced with pyrite grains. Fol-owing erosion of the overlying sandstone beds, microbial mat washen eroded away. Morphologically similar ‘elephant skin’ wrinkletructures have been described elsewhere (Bottjer and Hagadorn,007).
.4. Millimetre ripples
Millimetre ripples are preserved as positive epirelief on thepper surface of fine sandstone beds. Tiny ripple marks are millime-er sized, parallel to one another (Fig. 9a–c). The crests are sharpo round in cross section. These ripple marks have a wavelength
. (b) Close-up of a showing detail of reticulated ridges structures. (c) Amalgamatedce of dark lamina near the top of the fine sandstone bedding surface.
of 1–3 mm, ripple height of 0.5–1.5 mm, ripple index (RI) of 2–6,and ripple symmetrical index (RSI) of 1–1.1. These millimeter-sizedripple marks are unevenly distributed on the bed surfaces. Crosssection views show that the ripple crests share the same lithologicalcomposition with the host rock (Fig. 9g).
The morphological features and sizes of the Flat Rock For-mation ripple marks agree well with the concept of millimetreripples (Singh and Wunderlich, 1978; Pflüger, 1999). The wrinklepatterns suggest that these ripple marks can be categorized to thewrinkle structures documented by Hagadorn and Bottjer (1997)(Fig. 1a–g). The sharp to rounded crests of the millimetre ripplesare distinguished from the flat crests of Kinneyia wrinkle marks(Pflüger, 1999). These ripple marks also can not be confused withwind-driven foams because the ripple crests have the same litho-logical composition as the host rock, whereas the latter possesshollow crests (Allen, 1967). Both rain-impact ripples and adhesionripples have similar structures to the millimeter ripples describedhere, but differ from the latter in having asymmetrical morphologyand different lithological composition between ripple crests andthe host rock (Robb, 1992). Absence of sand-over-mud doubletsor other soft deformation related features also distinguishesthe Flat Rock Formation millimeter ripples from soft sedimentloading deformation structures (Allen, 1985; Parizot et al., 2005).
The formation of these millimetre ripples requires preexistingflexible, cohesive microbial mats on which wave or currentinduced shear stress would cause their deformation into variousmorphologies (Dalrymple, 1979; Pflüger, 1999). Millimetre ripples
Z.-W. Lan, Z.-Q. Chen / Precambrian Research 224 (2013) 529– 550 537
Fig. 7. (a) Field photo showing ‘elephant skin’ wrinkle structures preserved as positive epirelief on sandstone bed tops. (b) Close-up of a showing ‘elephant skin’ wrinklestructures are composed of elongate to elliptical bulges. (c) Amalgamated thin section view showing the semi-elliptical cross section of the bulges, the presence of dark claylamina on the surface, and pyrite aggregates in the low relief areas. (d) and (e) Close-up photos in (c) showing pyrite aggregates in the low relief areas. (f) and (g) SEM imagesshowing the presence of euhedral pyrite from the pyrite aggregates in (d) and (e). Coin in A is 2 cm in diameter.

538 Z.-W. Lan, Z.-Q. Chen / Precambrian Research 224 (2013) 529– 550
Fig. 8. (a) Field photo showing sub-rounded bulges composed reticulate wrinkle structures on sandstone bed tops. (b) Close-up of (a) showing reticulate wrinkle structureson sandstone bed tops. (c) Field photo showing rectangular bulges composed reticulate wrinkle structures on sandstone bed soles. Solid line indicates slab cutting position.(d) Close-up of (c) showing reticulate bulges composed ‘elephant skin’-like wrinkle structures on sandstone bed tops. (e) Photo of slab cut perpendicular to bedding surfaces
h(
4
eeomia(n
howing the wrinkle structure bearing bed is composed of massive fine sandstone.
ave been frequently documented from Precambrian strataPflüger, 1999).
.5. Spindle-shaped sand crack fills
A series of spindle-shaped sand crack fills preserved as positivepirelief on massive sandstone bed tops (Fig. 10). Spindles are gen-rally 0.8–2.2 cm long, 0.2–0.5 cm wide, and stand out 0.1–0.3 cmn bed surface. Some spindles are slightly deformed to curvedorphology (Fig. 10c). Most of them are randomly aligned with
ndividual spindles meeting at an acute or right angle (Fig. 10a, cnd d). Occasionally, a preferred parallel alignment is observedFig. 10b). Despite their multiple orientations, the sand crack fillsever form polygons. No superposition is observed among these
crack fills. The crack fills are composed of the same compositionand grain size as the host rock.
The spindle-shaped morphology of the Ranford sand crack fillssuggests that they should not be moulds of evaporitic gypsumcrystals (Astin and Rogers, 1991). Earthquake events should notbe responsible for the formation of these sand cracks becausetypical earthquake-related structures such as mud diapirs andball- and pillow structures are absent in the crack-bearing strata(e.g. Pratt, 1998). Desiccation of mudstone layer between two sandlayers may also generate similar structures to the spindle-shaped
sand crack fills. However, the laterally extending mudstone layersimmediately above the crack-fills bearing sandstone beds, charac-teristic of desiccation of mudstone layer (Eriksson et al., 2007), arenot seen in the field.
Z.-W. Lan, Z.-Q. Chen / Precambrian Research 224 (2013) 529– 550 539
Fig. 9. Millimetre ripple marks on sandstone bed tops. (a) Curved to sinuous round crested ripple marks. (b) Near parallel round crested ripple marks. (c) Straight to sinuous,parallel to near parallel sharp crested ripple marks. (d), (e) and (f) are close-ups of (a), (b) and (c), respectively. (g) Photo of polished slab cut perpendicular to bedding surfaceshows that the ripple mark bearing rock is composed of massive fine sandstone, and the crests of ripple marks have an indistinguishable lithological composition from thehost rock.

540 Z.-W. Lan, Z.-Q. Chen / Precambrian Research 224 (2013) 529– 550
Fig. 10. Field photos of spindle shaped sand cracks developed as positive epirelief. (a), (c) and (d) Randomly aligned spindle shaped sand cracks, individual cracks mostlycrossing at an acute angle. (b) Spindle shaped sand cracks aligned near parallel. Coin in all figures is 2 cm in diameter.
riamb1sMa2otssm
4
sres
The spindle-shaped morphology and clustered characteristicsecalls Rhysonetron from the Lower Proterozoic Bar River Formationn Elliot Lake, Ontario (Hofmann, 1967, 1971). Rhysonetron is gener-lly characterized by symmetrical paired corrugations separated byedial longitudinal furrows (Hofmann, 1967, 1971), and has also
een interpreted as microbial mat in origin (Pflüger, 1999; Gehling,999; Porada and Löffler, 2000). Spindle-shaped sand crack fillseparated by median furrows have also been reported from theesoproterozoic Chorhat Sandstone in India (Sarkar et al., 2006)
nd from the Ediacaran Sonia Sandstone in India (Sarkar et al.,008). These Indian structures were interpreted as deformationf crack fills resulted from disposition of microbial mat towardshe median areas of cracks fills. As a result, the Ranford spindle-haped sand crack fills share the same origin as Rhysonetron and thepindle-shaped sand crack fills and were produced due to involve-ent of microbial mats.
.6. Palimpsest ripples
Two suits of ripples that crossed at a nearly right angle on
andstone bed tops (Fig. 11a and b). The wavelength of theipples ranges from 3 cm to 5 cm. The primary ripples (formedarlier) generally have narrow, subrounded crests, whereas theecondary ripples (formed later) typically have wide, roundedripple crests (Fig. 11b). The overlapping of secondary ripplesonto the primary ripples formed many square-like structures.The secondary ripples are easily recognized as they preserved asisolated patches (outlined area in Fig. 11b).
Bidirectional crests of the palimpsest ripples indicate that theywere formed in two depositional events. Seilacher (1997) con-sidered that palimpsest ripples could be generated by depositionof a thin layer of microbial mat between the two depositionalevents. The microbial mat provided erosion-resisted cohesion andprotected the earlier formed ripples from being destroyed by thelater formed ones (Seilacher, 1997; Pflüger, 1999; Noffke et al.,2001). Actual field studies also show that palimpsest ripples formedby wave or currents are commonly overgrown by cyanobacterialmats which bound and stabilized the sediments in the rippledsurface (Noffke et al., 2001). Comparable palimpsest ripples havebeen documented from the Mesoproterozoic Chorhat Sandstonenear Chorhat, India (Seilacher, 1999), and the Palaeoprotero-zoic Magaliesberg Formation sandstones, South Africa (Mostert,2001).
4.7. Irregular sand crack fills
A set of irregular sand crack fills developed as convex epireliefon the rippled sandstone bed tops (Fig. 11c). They are densely

Z.-W. Lan, Z.-Q. Chen / Precambrian Research 224 (2013) 529– 550 541
Fig. 11. (a) Field photo showing palimpsest ripples on sandstone bedding surface. The squared area is figure B. The person is about 1.68 m in height. (b) Close-up of (a)showing later formed ripples overlapping on earlier formed ones. The preservation of these ripples attributed to the previous agglutinating microbial mat. (c) Field photoshowing mat growth induced irregular sand cracks on ripple crests and troughs. (d) Field photo showing triradiate sand cracks on ripple crests and troughs. Coin in all figuresi
ddaAis
ic(ssnansobhnpe
s 2 cm in diameter.
istributed along the ripple trough and ripple crest with a surfaceensity of 90%. Individual crack fill varies from 0.4–4 cm in length,nd 0.2–0.4 cm in width, projecting 0.1–0.3 cm from bed surface.ll of the crack fills are randomly aligned. Superposition among
ndividual crack fills is common. Crack fills are composed of theame composition and grain size as the host rock.
Absence of the laterally extending mudstone layers overly-ng the crack-bearing bed tops indicates that these irregular sandrack fills should not be sand-filled mudstone desiccation cracksEriksson et al., 2007). The irregular morphology of the Ranfordand cracks suggests they could not be moulds of evaporitic gyp-um crystals (Astin and Rogers, 1991). Earthquake events couldot produce these sand cracks because mud diapirs and ball-nd pillow structures, typical of earthquake-related structures, areot seen in the crack-bearing strata (e.g. Pratt, 1998). Unlike thepindle-shaped sand crack fills which show low density and lackf superposition, the irregular sand crack fills are characterizedy higher density and common superposition. Thus, they must
ave formed in alternating mat and sediment layers through alter-ating subaerial exposure and flooding, as shown in present-dayeritidal depositional environment (Gerdes et al., 2000; Noffket al., 2001).4.8. Triradiate cracks
Abundant triradiate cracks developed preferentially along theripple troughs of sandstone bed surfaces and preserved as nega-tive epirelief (Fig. 11d). The ripples commonly have broad rippletrough, 7–8 cm in width, and narrow ripple crest, 2–3 cm in width.Single triradiate cracks never connect each other, and thus nopolygonal cracks are observed. Each triradiate crack is composedof three spindle-shaped or linear cracks meeting at crack tails andforming a triradiate pattern. Single spindle-shaped or linear cracksare 0.4–2.5 cm long and 0.1–0.5 cm wide.
Neither mudstone remnants nor the laterally extending mud-stone layers immediately overlying the crack-bearing sandstonebeds occur in the examined strata. These cracks therefore are lesslikely to be mudstone desiccation cracks (c.f. Eriksson et al., 2007).Triradiate cracks represent the combination of multiple crackswhich were initially formed as spindle-shaped or linear tears at theearly stage of growth (Eriksson et al., 2007). Subsequent combina-
tion, organization and alignment of tear-like cracks facilitated thegrowth of triradiate cracks (Porada and Löffler, 2000; Eriksson et al.,2007). Crack patterns may have formed because of variation in thethickness of microbial mat originally covered on bed tops (Eriksson
5 rian R
eR2P
4
iwl3listtsbttogrFncoteIe(
wFsowdpacehtgnir
ioHt1rac
tmscdT
42 Z.-W. Lan, Z.-Q. Chen / Precamb
t al., 2007). Similar structures have been found in the middleiphean Bezymyannyi Formation of the Turukhansk Uplift (Petrov,002) and the Mesoproterozoic Makgabeng Formation, Limpoporovince, South Africa (Eriksson et al., 2007).
.9. Multidirectional linear ridges
These variously oriented linear ridges are all preserved as pos-tive epirelief on reddish medium bedded sandstone bed surfaces
hich are somewhat weathered. Ridges are sharp crested, paral-el to subparallel, slightly curved (Fig. 12a–c) or circular (Fig. 12d),–50 cm long, 0.8–2 cm wide, separated by wide groove of 5–50 cm
ong, 1.8–4 cm wide, projecting 1–3 cm from bed surface. Fieldnvestigation reveals that individual ridges have no longitudinaltriations on external surface. Nor do they have internal ornamen-ation (e.g. concentric cross section). Rather, all the ridges havehe same color and mineral composition as the host rock. Theharp crest of individual ridge makes it easy to differentiate theoundaries between neighboring individuals, and to trace theirerminations along extending directions. Some ridges are not con-inuous but intermittent (e.g. Fig. 12b and d), but no divergencef ridges are present. Ridges curving in the same direction tend toroup as clusters. There are five clusters (a–e) of different orientedidges in Fig. 12c, two in Fig. 12b, and one in Fig. 12a and d. Inigs. 12b and c, clusters of different orientations converge but doot intersect, and generally form an acute angle of 30–45◦. Somelusters of ridges are truncated by cracks (e.g. Fig. 12a–c), whereasthers are bordered by cracks (e.g. Fig. 12d). It should be noted thathe subparallel ridges (Fig. 12b and c) tend to constrict towardsnds, constituting radiating arrangement from an unknown point.n contrast, the curved ridges (Fig. 12a) do not converge to eithernds but show a subparallel alignment. The intermittent ridgesFig. 12d) aligned in a circular pattern.
Allen (1982) experimentally produced some radiating ridges,hich resemble the Ranford materials in many aspects (e.g.
ig. 12b, d, f and h). However, the modern analogues possesstraight ridges having pervasive pimple-like markings and beingccasionally bilobed. The Ranford structures have linear ridges,hich tend to radiate anteriorly and are curved to a commonirection, and lack external pimple-like markings or bilobed mor-hology. Furthermore, the experimentally produced ridges have
rectangular cross section, whereas a pyramidal cross sectionharacterizes the Ranford materials. Unidirectional currents shearrosion can not explain the circularly patterned ridges (Fig. 12d and). Physical scouring could not generate these structures as posi-ive epirelief on bed surface, but only a cluster of straight linearroove structures with longitudinal surface striations preserved asegative epirelief (Prothero and Schwab, 1996). As such, pure phys-
cal process could not produce the Ranford multidirectional linearidges.
Like multidirectional linear ridges, the late Neoproterozoicndex fossil Arumberia is characterized by radiating characteristicsf linear ridges (Glaessner and Walter, 1975; McIlroy et al., 2005).owever, Arumberia has subparallel, linear ridges that preferen-
ially developed on sandstone bed soles (e.g. Glaessner and Walter,975). In addition, Arumberia has the smoothly crested, subparallelidges (McIlroy et al., 2005) and the clustered ridges that are alwaysligned to one direction (McIlroy et al., 2005). Arumberia thereforean not accommodate the Ranford multidirectional linear ridges.
The clustered, multidirectional ridges of the Ranford mul-idirectional linear ridges are superficial to the present-day
ultidirectional ripple marks reported from the Mellum Island,
outhern North Sea (Noffke, 1998). However, the latter areharacterized by subrounded to rounded sinuous ripple crestsividing smooth, subrounded to rounded sinuous ripple troughs.he Ranford materials are composed of an array of multidirectionalesearch 224 (2013) 529– 550
sharp, straight or slightly curved linear crests separated by widegrooves of various reliefs. In addition, transition from ripple crestto ripple trough changed gradually in modern multidirectionalripple marks, whereas it changed sharply from ridge top to bottomin the Ranford materials. The multidirectional linear ridges shouldhave been formed during microbial mat growth by means ofmicrobial mat growth as confirmed by the actualistic observationsfrom the present-day supratidal pool at the Redfish Bay, Texas,USA (Hagadorn and Bottjer, 1997, 1999).
4.10. Drag folded bed surface
These structures are seen on the upper surface of sandstonebeds. They show different relief on different places, but thickentowards the fold hinges (Fig. 13a and b). Drag folds are present asmultidirectional clusters on the top of the crumpled bed surface.Folds in the same cluster usually curve towards the same direction.The axes of drag folds are nearly parallel to the bedding surface.Folds vary from 1 cm to 3 cm in width.
Similar structures documented from the Ediacaran Sonia Sand-stone of India have been interpreted as the involvement ofmicrobial mat in the flexible deformation of sandstone bed surface,which would have bound granular detrital sediments and providedcohesion so that a smooth, folded surface could have been produced(Sarkar et al., 2008). Drag folds due to deformation of microbialmats could have been influenced by shear stress triggered by wind,wave or current (Bouougri and Porada, 2002). As such, that the dragfolds show multidirections on top of the crumpled bed surface sug-gests they may have been influenced by varying wave or currentactions which brought various directions of shear stress. The taper-ing ends of the folded area indicate the ductile shear stress direction(c.f. Sarkar et al., 2008).
4.11. Gas domes
These domal structures are developed on the rippled sandstonebed tops and are preserved as positive epirelief (Fig. 14a and b).Domes generally have round to flat tops. Individual domes varyfrom 0.2 cm to 0.5 cm in diameter and project 0.05–0.2 cm frombed surface. Some domes are isolated and randomly distributed,whereas others are coalesced.
Bottjer and Hagadorn (2007) termed similar domal structuresas domal sand buildups, and suggested they could be resulted frombiofilm binding quartz sand grains in vertical direction (Bottjer andHagadorn, 2007). Periodic accretion due to the supply of detritalgrains eventually formed these structures. However, SEM imagesfrom the morphologically similar domal structures of the YurabiFormation suggest they lack internal laminae, but possess somepyrite framboids (Lan and Chen, 2012a). As such, they could haveformed in microbial metabolic process during which the domalstructures were formed due to organic decay with the involvementof sulphate reducing bacteria (Skyring et al., 1989). Gases, possiblyCO2, CH4 and H2S, were generated during this process. Accumu-lation of gas pressure to certain extent will eventually cause theupward movement of the sediments from below to form thesedomal structures. Similar domal structures with microbial mat ori-gin have been documented from the Late Cambrian Elk MoundGroup, Wisconsin, U.S.A. (Bottjer and Hagadorn, 2007).
4.12. Erosional remnants and pockets
Erosional remnants occur as concave, irregular mat remnant,
3–3.5 cm in diameter, on low relief area of the upper surface ofsandstone beds (Fig. 15a and b). The mat remnant has a sharpmargin compared to the host rock. This distinct margin makes itpossible to estimate a thickness of about 0.1–0.2 cm for the mat
Z.-W. Lan, Z.-Q. Chen / Precambrian Research 224 (2013) 529– 550 543
Fig. 12. Multidirected linear ridges. (a) Field photo showing parallel to sub-parallel linear ridges on the upper sandstone surface. (b) and (c) Field photos showing sub-parallelto curved linear ridges on the upper sandstone surface. (d) Field photo showing circular ridges on the upper sandstone surface. (e), (f), (g) and (h) are line drawings of (a),(b), (c) and (d), respectively. Visible length of hammer in (a) is about 28 cm. The pencil in (b) is about 11 cm in length. The hammer in (c) is about 33 cm in length. The bookin (d) is about 18 cm in length.

544 Z.-W. Lan, Z.-Q. Chen / Precambrian Research 224 (2013) 529– 550
Fig. 13. (a) Field photo showing the drag folded sandstone surface due to deformation of microbial bound sandstone bedding surface. Note the multidirections of the foldedsurface which may reflect multidirectional current induced shearing stress. (b) Close-up of (a) showing the single fold in each cluster curved in the same direction as responseto shearing stress. (c) Field photo showing pinnacle/tuft projecting through the sandstone bedding surface. (d) Close-up of (c) showing pinnacle/tufts projecting through thes
rtiiPsoidaBo(watr
pPcms
andstone bedding surface.
emnant. No mudstone interval or mud drapes are seen betweenhe host sandstone beds, but the host sandstone beds are dividednto several thin layers and are wrinkled and deformed withrregular surface and margins. The host sandstone beds are fragile.orosity or sponge fabrics are common on the upper surface ofandstone beds (Fig. 15a and b indicated by the arrow). Subpolyg-nal biofilms were detected (Fig. 15d) and they are embeddednto the quartz cement. Margins of biofilm polygon occasionallyropped off, otherwise are conspicuous (Fig. 15d indicated byrrows). Complete polygons range from 2 �m to 4 �m in size.iofilms are geochemically dominated by Si. The erosional pocketsccur as irregular low relief structures bound by microbial matindicated by the arrow), and preserved on the same bed surfaceith erosional remnants (Fig. 15c). The exposed erosional pockets
rea appears an irregular outline with a diameter of 5–7 cm and ahickness of 0.05–0.1 cm. Ragged margins can be seen on the hostock surface.
Sandstone chips become fragile upon lithification due to theresence of organic bacteria bodies and biofilms (e.g. Grey and
lanavsky, 2009). The silicified sub-polygonal biofilms and theirross-cutting relationships with host rock indicate that theyust have played an active role in trapping as well as bindingiliciclastic sediments. Because of different organic substances
distribution which will cause differential binding abilities, unevenerosion and destruction resulted from water current reworkingpeeled off the loosely microbial bounded sand surface formingerosional pockets, and left only the strongly microbial boundmat erosional remnants (Noffke, 1999; Friend et al., 2008). Theporous sandstone surface may be another layer of microbial boundsediments that indicates organic mat remnants were originallyexited in sandstone beds, and later experienced decay whichproduced gases. Escape of gases left these porous or spongysurfaces. Morphologically similar erosional remnants and pocketsof microbial mat origin have been documented from the lowersupratidal setting in Mellum Island (Noffke, 1999), and the LowerCretaceous Dakota Sandstone around Denver, Colorado, USA(MacKenzie, 1972). Such microbial mediated stabilization of sand-stone bed surface can also be formed in experiments (Friend et al.,2008).
4.13. Ripple patches
Ripple patches occupy an area of about 900 cm2, covering sand-stone bed tops. The ripple patches are characterized by indentedmargins (Fig. 14c and d). Ripples have a RI of 5, and a RSI of 3.5.Some crests are deformed and curve towards one direction.

Z.-W. Lan, Z.-Q. Chen / Precambrian Research 224 (2013) 529– 550 545
Fig. 14. (a) Field photo showing abundant gas domes preserved as positive epirelief on sandstone bed tops. Coin is 2 cm in diameter. (b) Close-up of c showing denselyclustered gas domes. (c) Field photo showing ripple patches on sandstone bed tops. The hammer is about 33 cm in length. (d) Close-up of (c) showing ripple patches withstraight to slightly curved parallel ripple crests.
tsotolaprtslmpstmitts
Sarkar et al. (2006) described similar ripple patches fromhe Mesoproterozoic Chorhat Sandstone in India. The jagged andtepped margins of the ripple patches together with shear erosionf bed surface between ripple patches prompted Sarkar et al. (2006)o propose a microbial mat coating origin for the ripple patches leftn bed tops. Sarkar et al. (2008) documented morphologically simi-ar ripple patches from the Ediacaran Sonia Sandstone in India. Thebsence of mudstone layers underlying and overlying the rippleatches beds led Sarkar et al. (2008) to surmise a microbial matelated genesis that the ripple patches were left by uni- or bidirec-ional wave shearing erosion on microbial mat covered sandstoneurface. The waves of currents are strong enough to peel off theoosely bound ripple surfaces, but not sufficient to plane off the
icrobial bound rippled surfaces (Sarkar et al., 2006, 2008). Mor-hologically similar ripple patches presented by Schieber (1998)how smooth margins to adjacent bed surface, contradictory tohose documented by Sarkar et al. (2006, 2008) that show indented
argins. The smooth margin was interpreted as indication of shear-
ng erosion of preexisting microbial mat (Schieber, 1998). As such,he unidirectional curving of ripple crests of the Flat Rock Forma-ion ripple patches could indicate a unidirectional wave or currenthearing erosion.4.14. Pinnacles/tufts
They have patterns of densely arranged pinnacles/tufts-shaped protrusions projecting through the sandstone bed surface,preserved as convex epirelief, 2–4 mm high (Fig. 13c and d). Pro-trusions range from 3 mm to 8 mm in width. Individual protrusioncommonly has a wide base and sharp, tapering top. Protrusionsare fragile and easily broken off.
Gerdes et al. (2000) termed such pinnacle-shaped protru-sions as pinnacles/tufts or ministromatolites that is formed asa result of clustered filamentous bacteria changing their grow-ing direction in response to gravity induced sediment fallout.The surface of pinnacles/tufts was initially coated with higheramount of cyanobacteria biomass that strengthens the flexi-bility of tufts by producing EPS (Gerdes et al., 2000; Gerdes,2007; Noffke et al., 2008). Abundant organic matters wouldhave caused the pinnacles/tufts to become rigid and fragileafter lithification. Similar pinnacles/tufts structures have been
documented from the present-day lower supratidal shallows inTunisian (Gerdes et al., 2000) and from the Late Archean Pon-gola Supergroup in South Africa (Gerdes, 2007; Noffke et al.,2008).
546 Z.-W. Lan, Z.-Q. Chen / Precambrian Research 224 (2013) 529– 550
Fig. 15. Erosional remnants and pockets. (a) and (b) Field photos showing erosional remnants on the upper sandstone bedding surface. (c) Indoor photo showing erosionalpocket on the upper sandstone bedding surface. (d) SEM image showing silicified sub-polygonal biofilm detected from the erosional remnants.
5
5
totMioB5tFfiteSFpF
. MISS abundance changes in the Kimberley
.1. MISS abundance in the interglacial stratigraphic units
MISS from the Ranford Formation have been examined athree localities in the Kimberley. MISS have occurring frequencyf 10.6% throughout the examined strata (Table 3; Fig. 16a) athe Donkey Gap section. Bedding plane coverage percentage of
ISS varies from 3% to 21% throughout the examined stratigraphicnterval (Table 3; Fig. 17). At the Keep River National Park section,ccurring frequency of MISS is up to 12.1% (Table 3; Fig. 16a).edding plane coverage percentage of MISS varies from% to 20% (Table 3; Fig. 17). At the Moonlight Valley sec-ion, MISS have occurring frequency up to 11% (Table 3;ig. 16a). Bedding plane coverage percentage of MISS variesrom 12% to 20% throughout the examined stratigraphicnterval (Table 3; Fig. 17). The MISS occurring frequency ofhe Ranford Formation is averagely 11.2%, with bedding plane cov-rage percentage varying from 3% to 21%. MISS from the Throssell
hale Formation have occurring frequency of 8.3% (Table 3;ig. 16a). Throughout the examined stratigraphic interval, beddinglane coverage percentage of MISS varies from 4% to 22% (Table 3;ig. 17).5.2. MISS abundance in the postglacial stratigraphic units
MISS from the Yurabi Formation have accumulative thickness ofsingle MISS-bearing beds of about 3 m. Their occurring frequency isup to 15% (Table 3; Fig. 16a). Bedding plane coverage percentage ofMISS varies from 30% to 68% throughout the examined stratigraphicinterval (Table 3; Fig. 17). The Flat Rock Formation MISS possessoccurring frequency up to 20% (Table 3; Fig. 16a). Bedding planecoverage percentage of MISS varies from 30% to 70% throughoutthe examined stratigraphic interval (Table 3; Fig. 17).
5.3. MISS abundance changes from the interglacial to postglacialperiods
MISS have average occurring frequency of 9.8% during theinterglacial period in the Kimberley (Fig. 16b). The average occur-ring frequency of MISS increased to 17.5% during the postglacialperiod (Fig. 16b). Clearly, MISS underwent a significant increasein average occurring frequency from interglacial to postglacial
period. Occurring frequency of MISS decreased from 12.1% (BeaslyKnob Member) or 11% (Jarrad Sandstone Member) immediatelyafter the Marinoan glaciation to 8.3% (Throssell Shale) or 10.6%(Jonny Cake Shale) in the late stage of the interglacial period. After
Z.-W. Lan, Z.-Q. Chen / Precambrian Research 224 (2013) 529– 550 547
Table 3Statistics of MISS abundance and bed surface density during each stratigraphic time interval.
Formation Total thickness of studiedstratigraphic unit (m)
Accumulative thickness ofsingle MISS-bearing beds (m)
MISS abundance MISS bed surface density
Flat Rock Formation 25 5 20% 30–70%Yurabi Formation 20 3 15% 30–68%Ranford Formation (Donkey Gap) 32 3.4 10.6% 3–21%Ranford Formation (Keep River Park) 7 0.85 12.1% 5–20%Ranford Formation (Moonlight Valley) 11.8 1.3 11% 12–20%Throssell Shale 30 2.5 8.3% 4–22%
0.0 0%
5.0 0%
10.0 0%
15.0 0%
20.0 0%
25.0 0%
Thro ssell S hale Ranford F orm ati on Yura bi Form ati on Flat Rock F orm ati on
MIS
S a
bu
nda
nce
(a)
MIS
S a
bu
nd
ance
0.0 %
2.0 %
4.0 %
6.0 %
8.0 %
10.0 %
12.0 %
14.0 %
16.0 %
18.0 %
20.0 %
interglacia l postgla cia l
(b)
Fig. 16. (a) MISS abundance changes during individual stratigraphic unit depositional period. (b) MISS abundance changes during interglacial and postglacial period.
0-10% 11-25% 26-50% 51-75% 76-100%
RanfordMoonlightValley
0.2
5 m
ThrossellShale
0.5
m
Flat RockFormation
0.5
m
YurabiFormation
0.5
m
RanfordKeep RiverNational Park
0.2
5 m
MISS bedsurfacedensity
MISS bedsurfacedensity
MISS bedsurfacedensity
MISS bedsurfacedensity
MISS bedsurfacedensity
RanfordDonkey Gap
0.5
m
5%
7%
3%
6%6%4%12%
20%
5%6%4%
6%
18%21%
MISS bedsurfacedensity
6%
12%
16%
5%
20%
15%
12%
15%
20%
5%
7%
12%
15%18%13%12%20%22%14%7%15%16%16%6%8%8%5%4%4%
16%
55%
52%
55%
30%
45%
35%
60%62%57%68%
65%
60%
60%56%30%65%
62%
70%
55%
45%
52%
32%
35%
30%30%46%68%48%49%64%
ng ind
t(
aR3rcf
Fig. 17. MISS bed surface density changes duri
he Gaskiers glaciation, MISS occurring frequency increased to 15%Yurabi Formation) and 20% (Flat Rock Formation), respectively.
During the interglacial period, bedding plane coverage percent-ges of MISS are 4–22% in the Throssell Shale and 3–21% in theanford Formation (Table 3; Fig. 17). The same proxy surges to
0–68% (Yurabi Formation) and 30–70% (Flat Rock Formation),espectively during the postglacial period. Clearly, bedding planeoverage percentages of MISS underwent a significant increaserom interglacial to postglacial period.ividual stratigraphic unit depositional period.
6. Discussions
All the MISS bearing sandstone and siltstone have experiencedmetamorphism of not more than prehnite-pumpellyite facies. MISSin the Kimberley mainly occur in fine sandstone and siltstone
beds, the siliciclastic grain is neither too coarse nor too fine andis favorable for the development and preservation of MISS. TheMISS bearing stratigraphic intervals were all deposited at tur-ning points of regression-transgression where peritidal hydraulics
5 rian R
pedMtbp
fhcdibqmeptRGce
wNt(sibatobisacAgametTnmo2
7
aucdpsgmahA
48 Z.-W. Lan, Z.-Q. Chen / Precamb
redominates. All MISS can be compared in morphology with mod-rn and ancient MISS analogues whose biogenicity has been wellemonstrated. At last, thin section observations on representativeISS also show they have microtextures left by microbial activi-
ies. Accordingly, all the prerequisites are in accordance with theiogenic criteria catalogued by Noffke (2009), and thus all MISSresented in this paper should be biogenic in origin.
MISS mainly occur on fine grained sandstone beds, only veryew on coarse and siltstone beds during the interglacial period. Aigher occurring frequency in sandstone successions (Fig. 17) indi-ates that MISS had colonization preference to quartz-rich sandyeposits as has been documented by Noffke et al. (2002), dur-
ng the interglacial period. This is because fine grained sandstoneeds generally reflect a moderate erosional environment with ade-uate light penetration which is favourable for photosyntheticicrobes to build microbial mats (Noffke et al., 2002). A prefer-
ntial occurrence on quartz sandstone beds is also witnessed inostglacial period during which MISS abundance increased towardshe younger stratigraphic unit (from the Yurabi Formation to Flatock Formation), indicating an increase in MISS abundance fromaskiers glaciation towards the terminal Ediacaran time. This isonsistent with the MISS abundance change trends through thentire postglacial period.
Although microbial mats could well develop in both cold andarmer climate conditions (Bertrand-Sarfati and Monty, 1994;offke, 2000), the Kimberley region was located in proximity
o equatorial regions during the inter- and postglacial periodsHoffman and Li, 2009), which means the Kimberley MISS exclu-ively occurred in warmer marine water settings during thenter- and postglacial periods. As such, temperature should note responsible for the MISS abundance changes between inter-nd postglacial periods. As MISS are formed from the interac-ions between microbes and siliciclastic sediments, their presencen bedding plane surface and vertical distributions therefore cane treated as an important proxy assessing microbial biomass
n the oceans. As a result, if sedimentation rate remains con-tant during the deposition of all MISS-bearing units in the studyrea, the increase in MISS abundance indicates an increasingontribution of microbial mats to sediments/sedimentary rocks.ccordingly, more MISS have been produced during the post-lacial period in comparison to the interglacial period. Low MISSbundance in the interglacial successions reflect low microbialass that could have created an ecosystem with low oxygen and
nergy that was responsible for the rare appearance of Ediacara-ype animals in the Kimberley region (Lan and Chen, 2012b).he increase of MISS abundance following the younger Mari-oan/Gaskiers glaciation could have reflected an amelioration ofarine ecosystem that contributed to the pervasive appearance
f Ediacara-type animals in Western and South Australia (McCall,006).
. Conclusions
Both occurring frequency and bedding plane coverage percent-ge of MISS increased from the interglacial units to postglacialnits in the Kimberley. During the interglacial period MISS haveolonization preference to quartz sandy substrates and their abun-ance decreased towards the younger strata. In contrast, during theostglacial period MISS abundance increased towards the youngertrata. Overall, MISS abundance increased from interglacial to post-lacial periods, implying an increase in microbial mats, which
ay indicate amelioration of the microbe-dominated ecosystemsfter the Ediacaran glaciation. The increase in microbial mats mayave contributed to pervasive emergence of Ediacaran biota inustralia.
esearch 224 (2013) 529– 550
Acknowledgments
Thanks the editor and two anonymous reviewers for their con-structive comments which have significantly improved the qualityof this paper. Janet Muhling and Steve Parry are thanked for assis-tance with obtaining SEM and BSE images at Centre for Microscopy,Characterisation, and Analysis (CMCA) at the University of WesternAustralia. ZWL’s PhD study is supported by a joint scholarship fromChina Scholarship Council and The University of Western Australia.This project is supported by an Australian Research Council discov-ery grant (DP0770398 to ZQC) and the National Natural ScienceFoundation of China (Grants 2012M510537 and 41273069 to ZWL,41272023 to ZQC). This study was also supported by a grant in aidfrom the State Key Laboratory of Geological Process and Resources,China University of Geosciences (Wuhan) (GPMR200901 to ZQC).
References
Allen, J.R.L., 1967. A beach structure due to wind-driven foam. Journal of SedimentaryPetrology 37, 691–692.
Allen, J.R.L., 1982. Sedimentary structures, their character and physical basis, Part II.In: Developments in Sedimentology Series. Elsevier, Amsterdam, pp. 1–676.
Allen, J.R.L., 1985. Wrinkle-marks: an intertidal sedimentary structure due to aseis-mic soft-sediment loading. Sedimentary Geology 41, 75–95.
Astin, T.R., Rogers, D.A., 1991. Subaqueous shrinkage cracks’ in the Devonian ofScotland reinterpreted. Journal of Sedimentary Petrology 61, 850–859.
Banerjee, S., Jeevankumar, S., 2005. Microbially originated wrinkle structures onsandstone and their stratigraphic context: Palaeoproterozoic Koldaha Shale,central India. Sedimentary Geology 176, 211–224.
Bao, H., Chen, Z.Q., Zhou, C., 2012. An 17O record of late Neoproterozoic glaciation inKimberley, Western Australia. Precambrian Research 216–219, 152–161.
Berglund, J., Müren, U., B ´amstedt, U., Andersson, A., 2007. Efficiency of aphytoplankton-based and a bacteria-based food web in a pelagic marine system.Limnology and Oceanography 52, 121–131.
Bertrand-Sarfati, J., Monty, C., 1994. Phanerozoic Stromatolites II. Kluwer AcademicPublishers, 471 p.
Bland, B.H., 1984. Arumberia Glaessner and Walter, a review of its potential forcorrelation in the region of the Precambrian Cambrian boundary. GeologicalMagazine 121, 625–633.
Bottjer, D., Hagadorn, J.W., 2007. Mat growth features. In: Schieber, J., Bose, P.K.,Eriksson, P.G., Banerjee, S., Sarkar, S., Altermann, W., Catuneau, O. (Eds.), Atlas ofMicrobial Mat Features Preserved Within the Siliciclastic Rock Record, Atlasesin Geosciences. Elsevier, Amsterdam, pp. 53–71.
Bouougri, E., Porada, H., 2002. Mat-related sedimentary structures in Neoprotero-zoic peritidal passive margin deposits of the West African Craton (Anti-Atlas,Morocco). Sedimentary Geology 153, 85–106.
Briggs, D.E.G., 2003. The role of decay and mineralization in the preservation ofsoft-bodied fossils. Annual Review of Earth and Planetary Sciences 31, 275–301.
Callow, R.H.T., Brasier, M.D., 2009. Remarkable preservation of microbial mats inNeoproterozoic siliciclastic settings: implications for Ediacaran taphonomicmodels. Earth-Science Reviews 96, 207–219.
Coats, R.P., Preiss, W.V., 1980. Stratigraphic and geochronological reinterpretationof Late Proterozoic glaciogenic sequences in the Kimberley Region, WesternAustralia. Precambrian Research 13, 181–208.
Corkeron, M., 2003. Neoproterozoic glacial events in the Kimberley region, West-ern Australia: their sedimentology and regional correlation in the context ofcontinental- and global-scale Neoproterozoic glaciation. PhD Thesis. Universityof Western Australia, Perth, Australia.
Corkeron, M., 2007. ‘Cap carbonates’ and Neoproterozoic glacigenic successionsfrom the Kimberley region, north-west Australia. Sedimentology 54, 871–903.
Corkeron, M., 2008. Deposition and palaeogeography of a glacigenic Neoproterozoicsuccession in the east Kimberley. Australia. Sedimentary Geology 204, 61–82.
Dalrymple, R.W., 1979. Wave-induced liquefaction: a modern example from the Bayof Fundy. Sedimentology 26, 825–844.
Dow, D.B., Gemuts, I., 1969. Geology of the Kimberley Region, Western Australia: theEast Kimberley. In: Western Australian Geological Survey Bulletin 1969/120.
Dunster, J.N., Beier, P.R., Burgess, J.M., Cutovinos, A., 2000. Auvergne, NorthernTerritory 1:250 000 Geological Series Explanatory Notes SD 52–15. NorthernTerritory Geological Survey.
Ehrlich, H.L., 1998. Geomicrobiology: its significance for geology. Earth-ScienceReview 45, 45–60.
Eriksson, P.G., Porada, H., Banerjee, S., Bouougri, E., Sarkar, S., Bumby, A.J., 2007.Mat-destruction features. In: Schieber, J., Bose, P.K., Eriksson, P.G., Banerjee, S.,Sarkar, S., Altermann, W., Catuneau, O. (Eds.), Atlas of Microbial Mat FeaturesPreserved Within the Siliciclastic Rock Record, Atlases in Geosciences. Elsevier,
Amsterdam, pp. 76–105.Fairchild, I.J., Kennedy, M.J., 2007. Neoproterozoic glaciation in the Earth System.Journal of the Geological Society 164, 895–921.
Fike, D.A., Grotzinger, J.P., Pratt, L.M., Summons, R.E., 2007. Reply to Grey K andCalver C R Arising from oxidation of the Ediacaran ocean. Nature 450, E18.

rian R
F
G
G
G
G
G
G
G
G
G
G
G
G
G
G
G
G
G
H
H
H
H
H
H
H
H
K
K
K
K
K
L
L
Z.-W. Lan, Z.-Q. Chen / Precamb
riend, P.L., Lucas, C.H., Holligan, P.M., Collins, M.B., 2008. Microalgal mediation ofripple mobility. Geobiology 6, 70–82.
ehling, J.G., 1999. Microbial mats in terminal Proterozoic siliciclastics: Ediacarandeath masks. Palaios 14, 40–57.
ehling, J.G., Droser, M.L., 2009. Textured organic surfaces associated with the Edi-acara biota in South Australia. Earth-Science Reviews 96, 196–206.
ellatly, D.C., Derrick, G.M., Plumb, K.A., 1975. The geology of the Lansdowne1:250,000 sheet area, Western Australian. Australia Bureau of Mineral andResources Geology and Geophysics Report 152.
emuts, I., Smith, J.W., 1968. Gordon Downs, Western Australian. Western Aus-tralian Geological Survey, 1:250,000 Geological Series Explanatory Notes.
erdes, G., Klenke, T., Noffke, N., 2000. Microbial signatures in peritidal siliciclasticsediments: a catalogue. Sedimentology 47, 279–308.
laessner, M.F., Walter, M.R., 1975. New Precambrian fossils from the ArumberaSandstone, Northern Territory, Australia. Alcheringa 1, 59–69.
lass, L.M., Phillips, D., 2006. Large igneous province in Australia with possible linksto faunal extinctions. Geology 34, 461–464.
erdes, G., 2007. Structures left by modern microbial mats in their host sediments.In: Schieber, J., Bose, P.K., Eriksson, P.G., Banerjee, S., Sarkar, S., Altermann,W., Catuneau, O. (Eds.), Atlas of Microbial Mat Features Preserved Withinthe Siliciclastic Rock Record, Atlases in Geosciences. Elsevier, Amsterdam,pp. 5–38.
ostin, V.A., McKirdy, D.M., Webster, L.J., Williams, G.E., 2010. Ediacaran ice-raftingand coeval asteroid impact, South Australia: insights into the terminal Protero-zoic. Australian Journal of Earth Sciences 57, 859–869.
rey, K., Corkeron, M., 1998. Late Neoproterozoic stromatolites in glacigenic suc-cessions of the Kimberley region, Western Australia: evidence for a youngerMarinoan glaciation. Precambrian Research 92, 65–87.
rey, K., Walter, M.R., Calver, C.R., 2003. Neoproterozoic biotic divesification: snow-ball earth or aftermath of the Acraman impact? Geology 31, 459–462.
rey, K., Calver, C.R., 2007a. Ediacaran oxidation and biotic evolution. Nature 450,E17.
rey, K., Calver, C.R., 2007b. Correlating the Ediacaran of Australia. In: Vickers-Rich,P., Komarower, P. (Eds.), The Rise and Fall of the Ediacaran Biota, 286. GeologicalSociety of London Special Publication, pp. 115–135.
rey, K., Planavsky, N.J., 2009. Microbialites of Lake Thetis, Cervantes, WesternAustralia—A field guide. Government of Western Australia, Department of Minesand Petroleum Record 2009/11.
riffin, T.J., 1989. A major thrust in the King Leopold Orogen, West Kimberley region.Geological Survey of Western Australia Report 26, 69–81.
riffin, T.J., Tyler, I.M., Playford, P.E., 1993. Lennard River, Western Australia, 3rd ed.Western Australia Geological Survey, 1:250,000 Geological Series ExplanatoryNotes.
uy, B.M., Beukes, N.J., Gutzmer, J., 2010. Paleoenvironmental controls on the textureand chemical composition of pyrite from non-conglomeratic sedimentary rocksof the Mesoarchean Witwatersrand Supergroup, South Africa. South AfricanJournal of Geology 113, 195–228.
agadorn, J.W., Bottjer, D.J., 1997. Wrinkle structures: microbially mediated sed-imentary structures in siliciclastic settings at the Proterozoic-Phanerozoictransition. Geology 25, 1047–1050.
agadorn, J.W., Bottjer, D.J., 1999. Restriction of a late Neoproterozoic biotope:suspect-microbial structures and trace fossils at the Vendian-Cambrian tran-sition. Palaios 14, 73–85.
aines, P.W., 2000. Problematic fossils in the late Neoproterozoic Wonoka Forma-tion, South Australia. Precambrian Research 100, 97–108.
andley, K.M., Turner, S.J., Campbell, K.A., Mountain, B.W., 2008. Silicifying biofilmexopolymers on a hot-spring microstromatolite: templating nanometer-thicklaminae. Astrobiology 8, 747–770.
anley, L.M., Wingate, M.T.D., 2000. SHRIMP zircon age for an Early Cambriandolerite dyke: an intrusive phase of the Antrim Plateau Volcanics of northernAustralia. Australia Journal of Earth Sciences 47, 1029–1040.
offman, P.F., Li, Z.X., 2009. A palaeogeographic context for Neoproterozoic glacia-tion. Palaeogeography, Palaeoclimatology, Palaeoecology 277, 158–172.
ofmann, H.J., 1967. Precambrian Fossils (?) near Elliot Lake. Ontario. Science 157,500–504.
ofmann, H.J., 1971. Precambrian fossils, pseudofossils and problematica in Canada.Geological Survey of Canada Bulletin 189.
aufman, A.J., Corsetti, F.A., Varni, M.A., 2007. The effect of rising atmospheric oxy-gen on carbon and sulphur isotope anomalies in the Neoproterozoic JohnnieFormation. Death Valley, USA. Chemical Geology 237, 47–63.
noll, A.H., Walter, M.R., Narbonne, G.M., Christie-Blick, N., 2004. A new period forthe geologic time scale. Science 305, 621–622.
onhauser, K.O., Urrutia, M.M., 1999. Bacterial clay authigenesis: a common biogeo-chemical process. Chemical Geology 161, 399–413.
ühl, M., Fenchel, T., Kazmierczak, J., 2003. Growth, structure and calcificationpotential of an artificial cyanobacterial mat. In: Krumbein, W.E., Paterson, D.M.,Zavarzin, G.A. (Eds.), Fossil and Recent Biofilms. Kluwer, Dordrecht, pp. 77–102.
umar, S., Pandey, S.K., 2007. Microbial mat-induced sedimentary structures in theNeoproterozoic Bundi Hill Sandstone, Indargarh area, Rajasthan. Current Science93, 1009–1012.
an, Z.W., Chen, Z.Q., 2012a. Exceptionally preserved microbially induced sedimen-
tary structures from the Ediacaran post-glacial successions in the Kimberleyregion, northwestern Australia. Precambrian Research 200–203, 1–25.an, Z.W., Chen, Z.Q., 2012b. Possible animal body fossils from the Late Neoprotero-zoic interglacial successions in the Kimberley region, northwestern Australia.Gondwana Research 21, 293–301.
esearch 224 (2013) 529– 550 549
Logan, G.A., Calver, C.R., Gorjan, P., Summons, R.E., Hayes, J.M., Walter, M.R., 1999.Terminal Proterozoic mid-shelf benthic microbial mats in the Centralian Super-basin and their environmental significance. Geochimica et Cosmochimica Acta63, 1345–1358.
MacKenzie, D.B., 1972. Tidal sand deposits in lower Cretaceous Dakota Group nearDenver, Colorado. Mountain Geologist 9, 269–277.
Madigan, M.T., Martinko, J.M., Dunlap, P.V., Clark, D.P., 2009. Brock Biology ofMicroorganisms, 12th ed. Pearson Benjamin-Cummings, San Francisco.
Maloof, A.C., Rose, C.V., Beach, R., Samuels, B.M., Calmet, C.C., Erwin, D.H., Poirier,G.R., Yao, N., Simons, F.J., 2010. Possible animal-body fossils in pre-Marinoanlimestones from South Australia. Nature Geoscience 3, 653–659.
McCall, G.J.H., 2006. The Vendian (Ediacaran) in the geological record: enigmasin geology’s prelude to the Cambrian explosion. Earth-Science Reviews 77, 1–229.
McFadden, K.A., Huang, J., Chu, X.L., Jiang, G.Q., Kaufman, A.J., Zhou, C.M., Yuan,X.L., Xiao, S.H., 2008. Pulsed oxidation and biological evolution in the Edi-acaran Doushantuo Formation. Proceedings of National Academy of Sciences105, 3179–3202.
McIlroy, D., Crimes, T.P., Pauley, J.C., 2005. Fossils and matgrounds from the Neopro-terozoic Longmyndian Supergroup, Shropshire, UK. Geological Magazine 142,441–455.
Mostert, P., 2001. Shoreline conditions in the Magaliesberg Formation, PretoriaGroup, Transvaal Supergroup. Honours Thesis. University of Pretoria, Pretoria,South Africa.
Noffke, N., 1998. Multidirected ripple marks rising from biological and sedimento-logical processes in modern lower supratidal deposits (Mellum Island, southernNorth Sea). Geology 26, 879–882.
Noffke, N., 1999. Erosional remnants and pockets evolving from biotic-physicalinteractions in a Recent lower supratidal environment. Sedimentary Geology123, 175–181.
Noffke, N., 2000. Extensive microbial mats and their influences on the erosional anddepositional dynamics of a siliciclastic cold water environment (Lower Areni-gian, Montagne Noire, France). Sedimentary Geology 136, 207–215.
Noffke, N., Gerdes, G., Klenke, T., Krumbein, W.E., 2001. Microbially induced sed-imentary structures – A new category within the classification of primarysedimentary structures. Journal of Sedimentary Research 71, 649–656.
Noffke, N., Knoll, A.H., Grotzinger, J.P., 2002. Sedimentary controls on the formationand preservation of microbial mats in siliciclastic deposits: a case study fromthe Upper Neoproterozoic Nama Group, Namibia. Palaios 17, 533–544.
Noffke, N., Beukes, N., Bower, D., Hazen, R.M., Swift, D.J.P., 2008. An actualis-tic perspective into Archean worlds—(cyano-)bacterially induced sedimentarystructures in the siliciclastic Nhlazatse Section, 2. 9 Ga Pongola Supergroup,South Africa. Geobiology 6, 5–20.
Noffke, N., 2009. The criteria for the biogenicity of microbially induced sedimen-tary structures (MISS) in Archean and younger, sandy deposits. Earth-ScienceReviews 96, 173–180.
Orange, F., Westall, F., Disnar, J.R., Prieur, D., Bienvenu, N., Le Romancer, M., Défarge,C.H., 2009. Experimental silicification of the extremophilic Archaea Pyrococ-cus abyssi and Methanocaldococcus jannaschii: applications in the search forevidence of life in early Earth and extraterrestrial rocks. Geobiology 7, 403–418.
Parizot, M., Eriksson, P.G., Aifa, T., Sarkar, S., Banerjee, S., Catuneanu, O., Alter-mann, W., Bumby, A.J., Bordy, E.M., van Rooy, J.L., Boshoff, A.J., 2005. Suspectedmicrobial mat-related crack-like sedimentary structures in the Palaeoprotero-zoic Magaliesberg Formation sandstones, South Africa. Precambrian Research138, 274–296.
Petrov, P.Yu., 2002. Microbial mats and terrigenous sedimentation settings in thePrecambrian: The middle Riphean Bezymyannyi Formation of the TurukhanskUplift. Lithology and Mineral Resources 37, 546–572.
Pflüger, F., 1999. Matground structures and redox facies. Palaios 14, 25–39.Porada, H., Löffler, T., 2000. Microbial shrinkage cracks in siliciclastic rocks
of the Neoproterozoic Nosib Group (Damara Supergroup) of central Namibia.Communications Geological Survey of Namibia 12, 63–72.
Pratt, B.R., 1998. Syneresis cracks: subaqueous shrinkage in argillaceous sedimentscaused by earthquake-induced dewatering. Sedimentary Geology 117, 1–10.
Prothero, D.R., Schwab, F., 1996. Sedimentary Geology: An Introduction to Sedimen-tary Rocks and Stratigraphy. Freeman, New York.
Robb III, A.J., 1992. Rain-impact microtopography (RIM): an experimental analoguefor fossil examples from the Maroon Formation, Colorodo. Journal of Sedimen-tary Petrology 62, 530–535.
Sarkar, S., Banerjee, S., Samanta, P., Jeevankumar, S., 2006. Microbial mat-inducedsedimentary structures in siliciclastic sediments examples from the 1.6 GaChorhat Sandstone, Vindhyan Supergroup, MP, India. Journal of Earth SystemScience 115, 49–60.
Sarkar, S., Bose, P.K., Samanta, P., Sengupta, P., Eriksson, P.G., 2008. Microbial matmediated structures in the Ediacaran Sonia Sandstone, Rajasthan, India, andtheir implications for Proterozoic sedimentation. Precambrian Research 162,248–263.
Schieber, J., 1998. Possible indicators of microbial mat deposits in shales and sand-stones: Examples from the Mid-Proterozoic Belt Supergroup, Montana, U.S.A.Sedimentary Geology 120, 105–124.
Schieber, J., 2007. Diagenetic features related to mat metabolism and decay.In: Schieber, J., Bose, P.K., Eriksson, P.G., Banerjee, S., Sarkar, S., Altermann,W., Catuneau, O. (Eds.), Atlas of Microbial Mat Features Preserved Withinthe Siliciclastic Rock Record, Atlases in Geosciences. Elsevier, Amsterdam,pp. 72–75.

5 rian R
S
SS
S
S
50 Z.-W. Lan, Z.-Q. Chen / Precamb
eilacher, A., 1997. Fossil Art. An exhibition of the Geologisches Institut, Tuebin-gen University, Germany. Royal Tyrell Museum of Paleontology, Drumheller,Alberta.
eilacher, A., 1999. Biomat-related lifestyles in the Precambrian. Palaios 14, 86–93.hen, B., Xiao, S.H., Kaufman, A.J., Bao, H.M., Zhou, C.M., Wang, H.F., 2008. Stratifica-
tion and mixing of a post-glacial Neoproterozoic ocean: evidence from carbonand sulphur isotopes in a cap dolostone from northwest China. Earth and Plan-etary Science Letters 265, 209–228.
ingh, I.B., Wunderlich, F., 1978. On the terms wrinkle marks (Runzelmarken), mil-limetre ripples, and mini ripples. Sencken-bergiana Maritime 10, 75–83.
kyring, G.W., Lynch, R.M., Smith, G.D., 1989. Quantitative relationships betweencarbon, hydrogen, and sulphur metabolism in cyanobacterial mats. In: Cohen,C., Rosenberg, E. (Eds.), Microbial Mats, Physiological Ecology of BenthicMicrobial Communities. American Society of Microbiology, Washington, D.C.,pp. 170–179.
esearch 224 (2013) 529– 550
Solic, M., Krstulovic, N., Kuspilic, G., Nincevic, G.Z., Bojanic, N., Sestanovic, S., Santic,D., Ordulj, M., 2010. Changes in microbial food web structure in response tochanged environmental trophic status: a case study of the Vranjic Basin (AdriaticSea). Marine Environmental Research 70, 239–249.
Thorne, A.M., Sheppard, S., Tyler, I.M., 1999. Lissadell, Western Australia (2nd ed.).Western Australia Geological Survey, 1:250,000 Geological Series ExplanatoryNotes.
Tyler, I.M., Griffin, T.J., Sheppard, S. 1998. Geology of the Dockrell 1:100,000 Sheet.Western Australia Geological Survey, 1:100,000 Geological Series ExplanatoryNotes.
Tyler, I.M., 2004. Geology of the Dixon 1:100,000 Sheet. Geological Survey of WesternAustralia, 1:100,000 Geological Series Explanatory Notes.
Willman, S., Moczydłowska, M., Grey, K., 2006. Neoproterozoic (Ediacaran) diversifi-cation of acritarchs: a new record from the Murnaroo 1 drillcore, eastern OfficerBasin, Australia. Review of Palaeobotany and Palynology 139, 17–39.