prior infection with an h1n1 swine influenza virus partially protects pigs against a low pathogenic...
TRANSCRIPT

Pa
KBa
b
c
a
ARA
KICHHPSP
1
bHobatlnamhl
0d
Vaccine 27 (2009) 6330–6339
Contents lists available at ScienceDirect
Vaccine
journa l homepage: www.e lsev ier .com/ locate /vacc ine
rior infection with an H1N1 swine influenza virus partially protects pigsgainst a low pathogenic H5N1 avian influenza virus
ristien Van Reeth a,∗, Debby Braeckmans a, Eric Cox b, Steven Van Borm c, Thierry van den Berg c,runo Goddeeris b, Annebel De Vleeschauwer a
Laboratory of Virology, Faculty of Veterinary Medicine, Ghent University, Salisburylaan 133, 9820 Merelbeke, BelgiumLaboratory of Immunology, Faculty of Veterinary Medicine, Ghent University, Salisburylaan 133, 9820 Merelbeke, BelgiumAvian Virology and Immunology, Veterinary and Agrochemical Research Centre, Groeselenberg 99, 1180 Brussels, Belgium
r t i c l e i n f o
rticle history:eceived 3 February 2009ccepted 5 March 2009
eywords:nfluenza virusross-subtype protection1N15N1igserum NI antibodyandemic
a b s t r a c t
Most humans lack virus neutralizing (VN) and haemagglutination inhibition (HI) antibodies to H5N1 avianinfluenza viruses (AIVs), but cross-reactive neuraminidase inhibition (NI) antibodies and cell-mediatedimmune (CMI) responses are common. These immune responses result largely from infections with sea-sonal human H1N1 influenza viruses, but the protective effect of H1N1 infection-immunity against H5N1infection has never been examined. To this purpose, we have used the pig model of influenza and alow pathogenic (LP) H5N1 AIV. Pigs were inoculated intranasally with sw/Belgium/1/98 (H1N1) 4 weeksbefore challenge with duck/Minnesota/1525/81 (H5N1). While the viruses failed to cross-react in HI andVN tests, the H1N1 infection induced high levels of H5N1 cross-reactive NI antibodies. Cross-reactive CMIwas demonstrated by measurements of lymphoproliferation and IFN-� secretion after in vitro restimula-tion of peripheral blood mononuclear cells. All control pigs showed clinical signs and H5N1 virus isolationfrom the respiratory tract post-challenge. The H1N1-immune pigs, in contrast, showed a complete clinical
protection and only 3 pigs out of 10 were H5N1 virus-positive. In a second and smaller experiment, H1N1virus infection also conferred cross-protection against a LP H5N2 AIV, while cross-reactive immunity wassolely detected in tests for CMI. Our data further support the notion that immunity induced by seasonalhuman H1N1 influenza virus infection may provide some protection against H5N1 or other H5 AIVs inthe absence of neutralizing H5 antibodies. Further studies should reveal whether cross-protection holdsagainst H5N1 viruses that are better adapted to replicate in mammals or with a more distantly related N1.. Introduction
Between December 2003 and the time of this writing, there haveeen approximately 400 cases of human infection with the Asian5N1 highly pathogenic avian influenza virus (HPAIV) and over 50%f these were fatal [1]. Because most humans lack neutralizing anti-odies to the H5 haemagglutinin (HA), the H5N1 virus could causehuman influenza pandemic if it would adapt to humans and gain
he ability to spread efficiently between humans, which it currentlyacks. At the same time, humans may have cross-reactive immu-ity to other viral proteins of the H5N1 virus. The internal proteins
re highly conserved between influenza A viruses and they are theajor targets for the cellular immune response [2]. In this context,umans with no known exposure to AIVs were found to have highevels of memory cytotoxic T lymphocytes that recognize multiple
∗ Corresponding author. Tel.: +32 9 2647369; fax: +32 9 2647495.E-mail address: [email protected] (K. Van Reeth).
264-410X/$ – see front matter © 2009 Elsevier Ltd. All rights reserved.oi:10.1016/j.vaccine.2009.03.021
© 2009 Elsevier Ltd. All rights reserved.
epitopes on AIVs, including HP H5N1 viruses isolated from patientsin Hong Kong in 1997 and in Vietnam in 2004 [3,4]. In addition, theH5N1 AIVs contain a neuraminidase (NA) of the same subtype asthe human H1N1 viruses that have circulated for around 90 yearsand anti-NA antibodies prevent the release of viruses from infectedcells in a similar way as NA inhibitors [5,6]. In a small serologicalsurvey, neuraminidase inhibition (NI) antibodies to the contempo-rary human A/New Caledonia/20/99 H1N1 virus were detected in31 of 38 humans tested and cross-reaction with the human H5N1isolates A/Hong Kong/213/03 and A/Vietnam/1203/04 was seen ineight and nine individuals respectively [7].
Though cross-reactive T lymphocytes or NA antibodies cannotprevent influenza virus infection, they did protect against severedisease and death caused by H5N1 HPAIVs in experiments in mice
and chickens. Mice infected with an H9N2 virus 4 weeks beforechallenge with an H5N1 virus still showed a temporary weight lossafter the challenge, but none of them died [8]. Similarly, chickensprimed with an H9N2 virus 3–70 d earlier were partially pro-tected against H5N1 virus shedding from the trachea and less so
ccine 2
fdlabigUv[albo[ccOmw[tftaHpei
oA–sdcciaHlutaiHf(t
2
2
avi(sUflvca
K. Van Reeth et al. / Va
rom the cloaca. Most H9N2-immune chickens showed only mildisease compared to the controls and survived the lethal chal-
enge [9,10]. In both instances, cross-reactive cellular immunityppeared to be vital for the observed cross-protection, as showny the absence of serologic cross-reaction in haemagglutination
nhibition (HI) and virus neutralization (VN) tests, as well as strate-ies to abrogate either the cellular or humoral immune response.nlike live influenza viruses, parenteral administration of inacti-ated influenza vaccines does not induce cross-subtype immunity2,11]. Consequently, conventional vaccines based on human H1N1nd/or H3N2 viruses largely failed to protect against H5N1 chal-enge in the mouse model [12–14]. Better results though haveeen obtained with novel vaccine formulations, novel adjuvantsr intranasal vaccination, or combinations of these approaches12–15]. The superior cross-protection to the traditional vaccinesould be explained by an enhanced stimulation of cross-reactiveytotoxic T cells, mucosal immunity or non-HI serum antibodies.ne highly interesting study used DNA vaccination to immunizeice specifically against the N1 of A/New Caledonia/20/99 (H1N1),hich has been a human influenza vaccine strain for several years
7]. Five out of 10 vaccinated mice survived a lethal challenge withhe A/Vietnam/1203/04 H5N1 HPAIV and passive transfer of serarom immunized to naive mice conferred an almost similar protec-ion. This was the first documentation of a role for N1 antibodiesgainst seasonal human H1N1 viruses in cross-protection against5N1. All these animal studies suggest that a portion of the humanopulation could have some degree of resistance to H5N1 influenza,specially if they have high N1 antibody titres, but it is almostmpossible to test this hypothesis in humans.
Pigs are natural influenza virus hosts with anatomical, physi-logical and immunological similarities to humans. The influenzavirus subtypes that occur in humans – H1N1, H3N2 and H1N2are enzootic in swine populations worldwide [16]. Pigs are also
usceptible to H5N1 HPAIVs under natural and experimental con-itions, but there is a clear species barrier as appears to be thease for humans [17–19]. We have previously shown that intratra-heal inoculation of pigs with low pathogenic (LP) H5 AIVs resultsn relatively high virus titres in the lungs and a mild to moder-te influenza-like illness [20]. These LP H5 viruses resemble theirP counterparts antigenically and genetically, but they require
ess stringent biosafety precautions. We have therefore started tose these viruses in porcine challenge experiments to test poten-ial immunization strategies against H5 HPAIVs. In this study, wesked three questions: (1) Can prior infection with an H1N1 swinenfluenza virus (SIV) partially protect pigs from challenge with an5N1 LPAIV? (2) Can H1N1 infection-immunity also protect pigs
rom challenge with an H5 AIV with a disparate NA, notably H5N2?3) Is protective immunity associated with cross-reactive antibodyo the N1 and/or cross-reactive cell-mediated immunity?
. Materials and methods
.1. Viruses
Swine/Belgium/1/98 (H1N1) was isolated during an outbreak ofcute respiratory disease in fattening pigs in Belgium in 1998. Theirus is representative of the avian-like H1N1 SIVs that are enzooticn swine populations of Western Europe. Duck/Minnesota/1525/81H5N1) was obtained from the virus repository of Dr. Robert Web-ter (St. Jude Children’s Research Hospital, Memphis, Tennessee,
S). Chicken/Belgium/150/99 (H5N2) was isolated from a chickenock in Belgium in 1999. Both H5 AIVs are low pathogenic. Theiruses were grown in the allantoic cavity of 10-d old embryonatedhicken eggs. The stocks used for inoculation of pigs, for geneticnalysis and for in vitro restimulation of peripheral blood mononu-7 (2009) 6330–6339 6331
clear cells represented the third or fourth egg passage. The stocksfor serological tests had undergone one more passage in eggs (HIand NI tests) or MDCK cells (VN tests).
2.2. Genetic characterisation
The complete coding sequence of the haemagglutinin (HA),neuraminidase (NA), matrix (M) and nucleoprotein (NP) genesegments was determined for the three viruses by directsequencing of overlapping RT-PCR products. The sequences ofduck/Minnesota/1525/81 HA, NP, and M, which were already avail-able in GenBank, were resequenced to avoid missing mutations thatmay arise during the passage history of the strain. Viral RNA wasextracted from allantoic fluid representing the virus stocks usedin the animal experiments using the High Pure Virus Nucleic AcidKit (Roche). The RNA was reverse transcribed using TranscriptorRT (Roche) and primer Uni12 [21]. The primer sequences used togenerate overlapping PCR amplicons are available on request. PCRamplicons were controlled on agarose gel and purified using theHigh Pure PCR product purification kit (Roche). Direct sequencingwas done using the BigdyeTerminator V3.1 kit (Applied Biosystems)starting from approximately 10 ng of template DNA. Sequencingreactions were purified using the BigdyeXterminator kit (AppliedBiosystems) and separated on an Applied Biosystems 310 geneticanalyzer. Assembly of the sequences was done using DNAsysMaxv2.09 (Hitachi Software Engineering) and manual editing.
The generated sequences were compared to selected sequencesfrom GenBank. Sequence alignment was done using the clustalWalgorithm within Bioedit V.09 [22]. Phylogenetic (Neighbour Join-ing, bootstrap with 1000 replicates) analysis was done using MEGAv3.1 [23].
2.3. Pigs and experimental design
The pigs were derived from a conventional herd that is negativefor influenza A viruses as shown by an anti-influenza A nucleocap-sid ELISA (ID Screen® Influenza A Antibody Competition, ID-VET).Each experimental group was housed in a separate isolation facility.The pigs were transferred to the experimental facilities at the ageof 4 weeks and they were 5-week-old at the time of the H1N1 virusinoculations.
Twenty-five pigs were used to investigate whether prior infec-tion with H1N1 confers cross-protection against an H5N1 AIV. Theywere inoculated first with swine/Belgium/1/98 (H1N1) and 4 weekslater with duck/Minnesota/1525/81 (H5N1) (H1N1-immune group,n = 13), or with the latter virus only at the age of 9 weeks (H5N1challenge control group, n = 12). The primary virus inoculation wasperformed intranasally, using 7.0 log10 EID50. The H5N1 challengewas performed both intranasally (7.0 log10 EID50) and intratra-cheally (7.5 log10 EID50). Two pigs per group were euthanatizeddaily on days 1–5 post-challenge (d PC), but only one control pig waseuthanatized 1 d PC. Samples from the entire respiratory tract werecollected for virus titrations. Whole blood for analyses of the cell-mediated immune (CMI) response was collected from six pigs pergroup at 1 week before the H5N1 challenge, and from the remainingthree pigs at 3 weeks PC. Blood samples for serology were collectedat 28, 14 and 0 d before challenge in all pigs, and at 14 and 28 d PCin the remaining pigs.
Fourteen pigs were used to investigate the extent of cross-protection between H1N1 and an H5 AIV with a distinct HA and NA.They were inoculated first with swine/Belgium/1/98 (H1N1) and 4
weeks later with chicken/Belgium/150/99 (H5N2) (H1N1-immunegroup, n = 7), or with the H5N2 virus only (H5N2 challenge controlgroup, n = 7). One pig per group was euthanatized at 1 and 2 d PCwith H5N2 for virus titrations, and two pigs per group at 3 d PC. Theremaining three pigs per group were used for analyses of the CMI
6 ccine
rlfi
2
achpd2t1nMutc
2
diittAcwdamwdtHwavrt
epctt
2
bb1sp
toe(tf
332 K. Van Reeth et al. / Va
esponse 1 week before and 3 weeks PC with H5N2. Virus inocu-ations and blood sampling for serology were performed as in therst experiment.
.4. Virus titrations
Samples from the nasal mucosa, nasopharynx, tonsil, tracheand lungs were collected at euthanasia. The cranial (apical andardiac) and caudal (diaphragmatic) lobes of the right and leftalves of the lung were kept separately to obtain four lung sam-les, because previous experiments had shown that H5 LPAIVso not replicate uniformly throughout all lung lobes. Ten or0% tissue homogenates were prepared and clarified by cen-rifugation. Virus titrations were performed by inoculation of0-fold dilutions of the supernatants in Madin Darby canine kid-ey (MDCK) cell cultures. After 72 h of incubation, virus-positiveDCK cells were demonstrated by immunoperoxidase staining
sing monoclonal antibodies against influenza virus nucleopro-ein, biotinylated sheep anti-mouse IgG, and a streptavidin–biotinomplex coupled to peroxidase.
.5. Serological assays
The serum samples were examined in HI, VN and NI tests toetermine antibody titres against the viruses used for inoculation
n the respective experiments. The HI test was performed accord-ng to standard methods with four haemagglutinating units ofhe respective viruses. Chicken erythrocytes (0.5%) were used inests with the H1N1 SIV and horse erythrocytes (1%) in tests withIVs. The sera were first adsorbed onto the respective erythro-ytes and treated with receptor-destroying enzyme. The VN testas performed as described previously [24]. Briefly, 2-fold serumilutions were incubated (1 h, 37 ◦C) with 100 TCID50 of MDCK cell-dapted virus in microtitre plates. MDCK cells (600,000 cells perl) were inoculated with the virus–serum mixture for 24 h, afterhich virus-positive cells were demonstrated by immunoperoxi-
ase staining as described higher. The NI test was performed in testubes of approximately 16 mm × 100 mm as described by Aymard-enry et al. [25]. Briefly, standard virus doses for use in the testere selected by an assay of NA activity based on the colorimetric
nalysis of sialic acid release from fetuin substrate. An appropriateirus dilution was incubated with 2-fold serum dilutions and theeduction of NA activity in each serum dilution was compared tohat in controls without serum.
All sera were tested in duplicate and antibody titres werexpressed as the reciprocal of the highest serum dilution that com-letely inhibited haemagglutination or virus replication in MDCKells, or that gave a 50% inhibition of NA activity. The starting dilu-ions were 1:2 in the VN test, 1:10 in the HI test and 1:20 in the NIest.
.6. Cell-mediated immune responses
Peripheral blood mononuclear cells (PBMCs) were obtainedy Ficoll-Hypaque density gradient centrifugation of heparinizedlood. The cells were resuspended in RPMI 1640 medium withmM sodium pyruvate, 1 mM l-glutamine, antibiotics, 5% fetal calf
erum and 50 �M �-mercaptoethanol at a concentration of 107 cellser ml.
For examination of lymphoproliferation after in vitro restimula-ion, PBMC were placed in triplicate wells (3 × 105 cells per well)
f 96-well microtitre plates. Each triplicate was incubated withach of the following stimuli: medium alone, allantoic fluid proteinnegative controls), concanavalin A (positive control; final concen-ration 10 �g/ml), H1N1, H5N1 and H5N2 influenza viruses usedor inoculation of the pigs. The H5N1 virus was left out in the H5N227 (2009) 6330–6339
challenge experiment. Sucrose gradient purified influenza viruseswere used at an MOI of 1. Allantoic fluid protein was harvestedfrom 10-d old noninfected chicken embryos, purified by sucrosegradient centrifugation and used at a similar dilution as the egg-grown influenza viruses. The PBMC were incubated at 37 ◦C for4 d and pulse-labeled with [3H]thymidine (1 �Ci per well, Amer-sham ICN) during the final 18 h. Thymidine uptake was measured incounts per minute (cpm) using a �-scintillation counter and lumi-nescence counter system (PerkinElmer). Lymphoproliferation wasexpressed as stimulation index, SI = mean cpm of cells stimulatedwith virus/mean cpm of cells stimulated with noninfected allan-toic fluid. The group mean cpm in response to allantoic fluid fromuninfected eggs was consistently <15% of the response to influenzaviruses in infection-immune pigs.
To examine IFN-� secretion after in vitro restimulation, PBMCwere placed in duplicate wells (107 cells per well) of 24-wellmicrotitre plates and incubated with the stimuli used in thelymphoproliferation assay. Concanavalin A was used at a final con-centration of 5 �g/ml. Supernatants were collected at 0, 18, 48 and72 h and stored at −70 ◦C. A capture ELISA was used to determinethe concentration of IFN-� by use of antibody pairs purchased fromInvitrogen (Swine IFN-� CytosetTM), following the manufacturer’sprotocol. Group mean IFN-� titres in response to noninfected allan-toic fluid were between 0.7 and 1.4 log10 pg/ml.
2.7. Statistical analysis
Standard two-sample t-tests were used to compare virus titres,antibody titres and CMI responses between groups. t-Tests werealso used to compare antibody and CMI responses to H1N1, H5N1and H5N2 viruses. Samples that tested negative for virus weregiven a numeric value of 1.6 log10 TCID50/g (trachea and lung sam-ples; detection limit 1.7 log10) or 1.8 log10 TCID50/g (nasal mucosa,nasopharynx and tonsil; detection limit 1.9 log10). Samples thattested negative in serological assays were assigned a value cor-responding with half of the minimum detectable antibody titre.p < 0.05 was taken as the level of significance.
3. Results
3.1. Genetic characterisation
The sequences generated in this study are availablethrough GenBank accession numbers FJ750564–FJ750571 andFJ805962–FJ805965. No changes could be observed in theduck/Minnesota/1525/81 (H5N1) challenge stock compared to theoriginal sequence information already available in GenBank for HA,NP and M (CY014726–CY014728). Phylogenetic analysis confirmsthe different origin of the viruses. All four analyzed genes (HA, NA,M and NP) for swine/Belgium/1/98 (H1N1) cluster within a group ofswine isolates, whereas the avian viruses duck/Minnesota/1525/81(H5N1) and chicken/Belgium/150/99 (H5N2) cluster within aNorth American and Eurasian group of avian viruses respectively(Fig. 1 for NA, data similar but not shown for other genes). Theamino acid identity (Table 1) shows a relatively high homol-ogy (90.7%) between the duck/Minnesota/1525/81 (H5N1) andswine/Belgium/1/98 (H1N1) NA and confirms the conserved natureof NP and M genes.
3.2. Protection against H5 AIVs in H1N1 infection-immune pigs
Within 1 d after the challenge with H5N1 all 12 control pigs were
depressed and had no appetite, and some pigs showed fever (5/12)and tachypnoea (6/12) (Table 2). Coughing was seen occasionally at4 and 5 d PC. The symptoms peaked 1 and 2 d PC and had completelyresolved by 6 d PC. Small areas of lung consolidation, involving anaverage 5.4% of the total lung, were seen in six out of nine control
K. Van Reeth et al. / Vaccine 27 (2009) 6330–6339 6333
Fig. 1. Genetic characterization of the N1 neuraminidase of duck/Minnesota/1525/81 (H5N1) and swine/Belgium/1/98 (H1N1) based on the complete neuraminidase codingsequence.
Table 1Comparison of genes of the H1N1 swine influenza virus used for primary infection and the H5 LPAI challenge viruses.
Virus % identitya against swine/Belgium/1/98 (H1N1) genes
HA NA NP M
pnt(ltsoliwfi
Duck/Minnesota/1525/81 (H5N1) 64.3Chicken/Belgium/150/99 (H5N2) 65.3
a % identity at the amino acid level.
igs euthanatized. The H5N1 challenge virus was isolated from alline pigs (Table 3a), but the virus did not replicate homogeneouslyhroughout the respiratory tract and only 42 of the total 72 samples58%) were virus-positive. The apical and cardiac lobes of the rightung were consistently positive and they tended to contain higheritres than the other lung lobes, trachea or upper respiratory tractamples. The H1N1-immune pigs, in contrast, did not show any sign
f disease after the H5N1 challenge. Only 2 out of 10 pigs showedung lesions, with an average 0.75% of the lung tissue involved. Virussolation rates and virus titres were strongly reduced in comparison
ith those in the control group (p < 0.05): only 3 out of 10 pigs orve of the total 80 samples (6%) tested virus-positive.
Table 2Clinical signs after challenge with an H5N1 LPAIV in control and H1N1-immune pigs.
d PCb Number of pigs with clinical signs/total number of pigs(mean clinical score)a
Challenge control group H1N1-immune group
0 0/12 0/131 12/12 (3.4) 0/132 11/11 (2.4) 0/113 8/9 (1.1) 0/94 3/7 (1.0) 0/75 2/5 (1.0) 0/56 0/3 0/37 0/3 0/38 0/3 0/3
a Mean of the individual scores of all pigs with symptoms. Individualscores were obtained by assigning a value of 1 to each of the followingsymptoms: fever (body temperature ≥40 ◦C), depression, loss of appetite,tachypnoea (breathing rate >45/min), abdominal respiration, coughing.
b d PC: days post-challenge.
90.7 95.5 96.442.2 95.3 96.0
The H5N2 challenge produced a transient depression, fever andtachypnoea in two out of seven control pigs at 1 and 2 d PC only(data not shown). Gross lung lesions, involving an average 6% ofthe total lung, were present in all four pigs euthanatized. The H5N2challenge virus was isolated from all control pigs or from 18 of thetotal 32 samples (56%). The right lung lobes tend to show the highestvirus isolation rates (Table 3b). The H1N1-immune pigs remainedasymptomatic and without lung lesions. The virus was recoveredfrom two out of four pigs and from four of the 32 samples (12.5%).Owing to the small group size, the difference with the control groupwas not statistically significant (p > 0.05).
3.3. Serological findings
At the start of the experiments all pigs were influenza-naiveas measured by an ELISA for the presence of antibodies specificfor the nucleoprotein. All pigs were monitored serologically untilthe time of challenge with H5 AIVs; only three pigs per groupwere not euthanatized and were also examined PC. In the H5N1challenge experiment, however, one pig of both groups died acci-dentally between 14 and 28 d PC and only two instead of three pigswere available for serologic testing at 28 d PC.
The H5N1 challenge control pigs were still negative in all anti-body assays at the time of the challenge and they had developedHI, VN and NI antibodies to the challenge virus at 14 and 28 d PC(Table 4a). There was no serological cross-reaction between H5N1
and H1N1 in HI and VN tests, but the pig with the highest NI anti-body titre to H5N1 (160 and 80 at 14 and 28 d PC respectively)showed cross-reaction with H1N1. The H1N1-immune pigs onlyhad HI and VN antibodies to the homologous H1N1 virus withwhich they had been inoculated before challenge. Twelve out of 13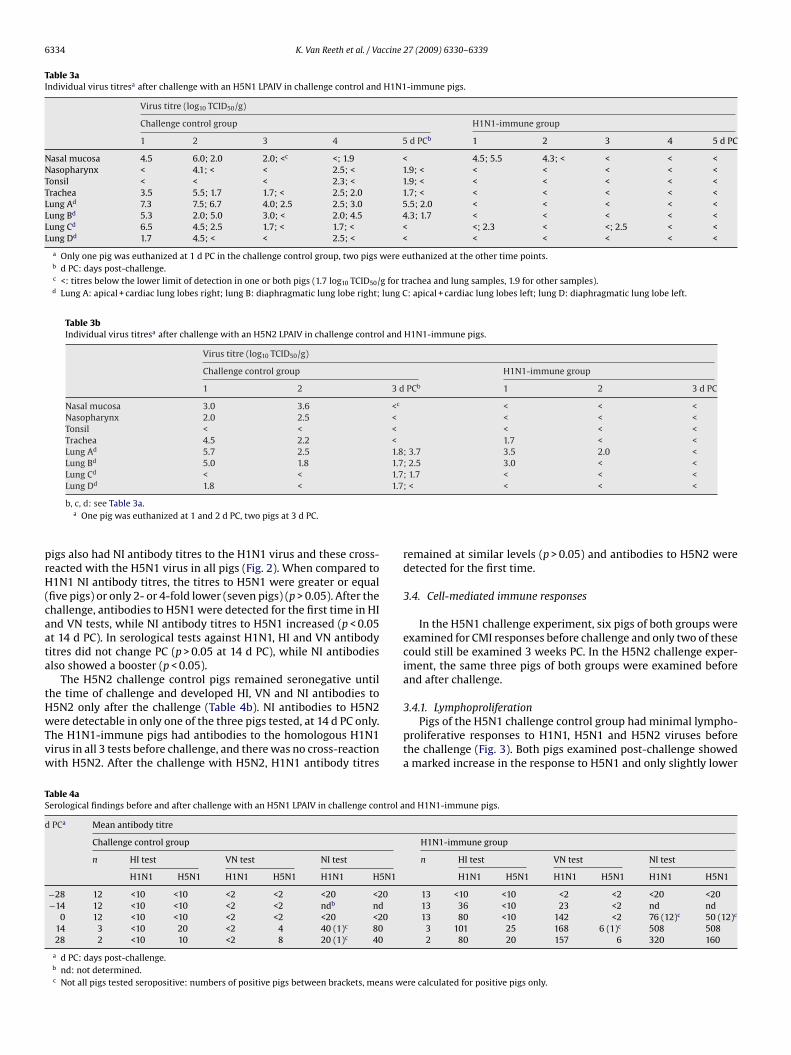
6334 K. Van Reeth et al. / Vaccine 27 (2009) 6330–6339
Table 3aIndividual virus titresa after challenge with an H5N1 LPAIV in challenge control and H1N1-immune pigs.
Virus titre (log10 TCID50/g)
Challenge control group H1N1-immune group
1 2 3 4 5 d PCb 1 2 3 4 5 d PC
Nasal mucosa 4.5 6.0; 2.0 2.0; <c <; 1.9 < 4.5; 5.5 4.3; < < < <Nasopharynx < 4.1; < < 2.5; < 1.9; < < < < < <Tonsil < < < 2.3; < 1.9; < < < < < <Trachea 3.5 5.5; 1.7 1.7; < 2.5; 2.0 1.7; < < < < < <Lung Ad 7.3 7.5; 6.7 4.0; 2.5 2.5; 3.0 5.5; 2.0 < < < < <Lung Bd 5.3 2.0; 5.0 3.0; < 2.0; 4.5 4.3; 1.7 < < < < <Lung Cd 6.5 4.5; 2.5 1.7; < 1.7; < < <; 2.3 < <; 2.5 < <Lung Dd 1.7 4.5; < < 2.5; < < < < < < <
a Only one pig was euthanized at 1 d PC in the challenge control group, two pigs were euthanized at the other time points.b d PC: days post-challenge.c <: titres below the lower limit of detection in one or both pigs (1.7 log10 TCID50/g for trachea and lung samples, 1.9 for other samples).d Lung A: apical + cardiac lung lobes right; lung B: diaphragmatic lung lobe right; lung C: apical + cardiac lung lobes left; lung D: diaphragmatic lung lobe left.
Table 3bIndividual virus titresa after challenge with an H5N2 LPAIV in challenge control and H1N1-immune pigs.
Virus titre (log10 TCID50/g)
Challenge control group H1N1-immune group
1 2 3 d PCb 1 2 3 d PC
Nasal mucosa 3.0 3.6 <c < < <Nasopharynx 2.0 2.5 < < < <Tonsil < < < < < <Trachea 4.5 2.2 < 1.7 < <Lung Ad 5.7 2.5 1.8; 3.7 3.5 2.0 <Lung Bd 5.0 1.8 1.7; 2.5 3.0 < <Lung Cd < < 1.7; 1.7 < < <
d 1.7
prH(caata
tHwTvw
TS
d
Lung D 1.8 <
b, c, d: see Table 3a.a One pig was euthanized at 1 and 2 d PC, two pigs at 3 d PC.
igs also had NI antibody titres to the H1N1 virus and these cross-eacted with the H5N1 virus in all pigs (Fig. 2). When compared to1N1 NI antibody titres, the titres to H5N1 were greater or equal
five pigs) or only 2- or 4-fold lower (seven pigs) (p > 0.05). After thehallenge, antibodies to H5N1 were detected for the first time in HInd VN tests, while NI antibody titres to H5N1 increased (p < 0.05t 14 d PC). In serological tests against H1N1, HI and VN antibodyitres did not change PC (p > 0.05 at 14 d PC), while NI antibodieslso showed a booster (p < 0.05).
The H5N2 challenge control pigs remained seronegative untilhe time of challenge and developed HI, VN and NI antibodies to
5N2 only after the challenge (Table 4b). NI antibodies to H5N2ere detectable in only one of the three pigs tested, at 14 d PC only.he H1N1-immune pigs had antibodies to the homologous H1N1irus in all 3 tests before challenge, and there was no cross-reactionith H5N2. After the challenge with H5N2, H1N1 antibody titres
able 4aerological findings before and after challenge with an H5N1 LPAIV in challenge control a
PCa Mean antibody titre
Challenge control group
n HI test VN test NI test
H1N1 H5N1 H1N1 H5N1 H1N1 H5N1
−28 12 <10 <10 <2 <2 <20 <20−14 12 <10 <10 <2 <2 ndb nd
0 12 <10 <10 <2 <2 <20 <2014 3 <10 20 <2 4 40 (1)c 8028 2 <10 10 <2 8 20 (1)c 40
a d PC: days post-challenge.b nd: not determined.c Not all pigs tested seropositive: numbers of positive pigs between brackets, means we
; < < < <
remained at similar levels (p > 0.05) and antibodies to H5N2 weredetected for the first time.
3.4. Cell-mediated immune responses
In the H5N1 challenge experiment, six pigs of both groups wereexamined for CMI responses before challenge and only two of thesecould still be examined 3 weeks PC. In the H5N2 challenge exper-iment, the same three pigs of both groups were examined beforeand after challenge.
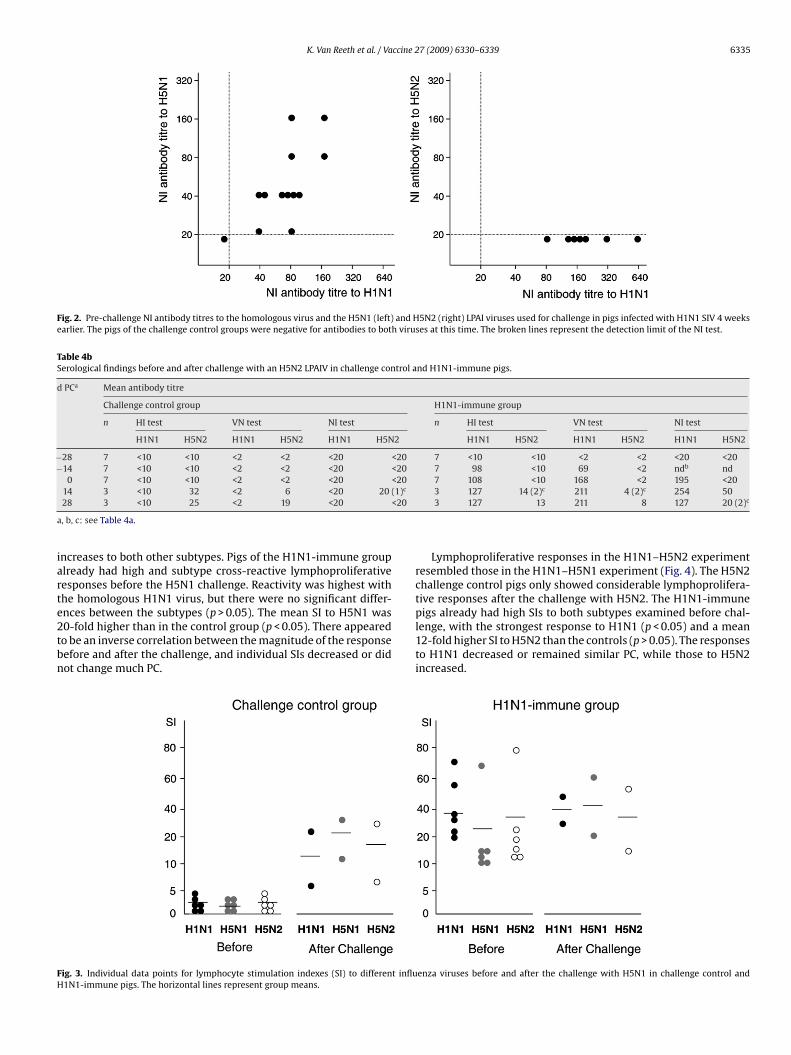
3.4.1. LymphoproliferationPigs of the H5N1 challenge control group had minimal lympho-
proliferative responses to H1N1, H5N1 and H5N2 viruses beforethe challenge (Fig. 3). Both pigs examined post-challenge showeda marked increase in the response to H5N1 and only slightly lower
nd H1N1-immune pigs.
H1N1-immune group
n HI test VN test NI test
H1N1 H5N1 H1N1 H5N1 H1N1 H5N1
13 <10 <10 <2 <2 <20 <2013 36 <10 23 <2 nd nd13 80 <10 142 <2 76 (12)c 50 (12)c
3 101 25 168 6 (1)c 508 5082 80 20 157 6 320 160
re calculated for positive pigs only.
K. Van Reeth et al. / Vaccine 27 (2009) 6330–6339 6335
Fig. 2. Pre-challenge NI antibody titres to the homologous virus and the H5N1 (left) and H5N2 (right) LPAI viruses used for challenge in pigs infected with H1N1 SIV 4 weeksearlier. The pigs of the challenge control groups were negative for antibodies to both viruses at this time. The broken lines represent the detection limit of the NI test.
Table 4bSerological findings before and after challenge with an H5N2 LPAIV in challenge control and H1N1-immune pigs.
d PCa Mean antibody titre
Challenge control group H1N1-immune group
n HI test VN test NI test n HI test VN test NI test
H1N1 H5N2 H1N1 H5N2 H1N1 H5N2 H1N1 H5N2 H1N1 H5N2 H1N1 H5N2
−28 7 <10 <10 <2 <2 <20 <20 7 <10 <10 <2 <2 <20 <20−14 7 <10 <10 <2 <2 <20 <20 7 98 <10 69 <2 ndb nd
0 7 <10 <10 <2 <2 <20 <20 7 108 <10 168 <2 195 <20(1)c
<20
a
iarte2tbn
FH
14 3 <10 32 <2 6 <20 2028 3 <10 25 <2 19 <20
, b, c: see Table 4a.
ncreases to both other subtypes. Pigs of the H1N1-immune grouplready had high and subtype cross-reactive lymphoproliferativeesponses before the H5N1 challenge. Reactivity was highest withhe homologous H1N1 virus, but there were no significant differ-
nces between the subtypes (p > 0.05). The mean SI to H5N1 was0-fold higher than in the control group (p < 0.05). There appearedo be an inverse correlation between the magnitude of the responseefore and after the challenge, and individual SIs decreased or didot change much PC.ig. 3. Individual data points for lymphocyte stimulation indexes (SI) to different influ1N1-immune pigs. The horizontal lines represent group means.
3 127 14 (2)c 211 4 (2)c 254 503 127 13 211 8 127 20 (2)c
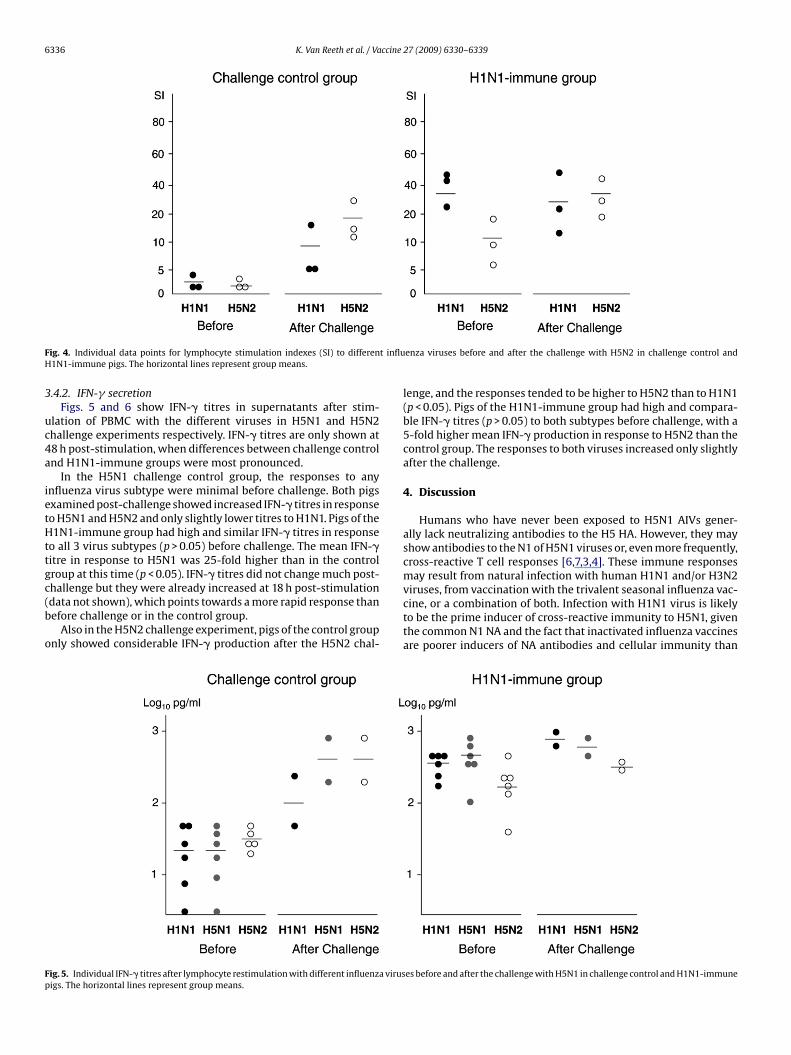
Lymphoproliferative responses in the H1N1–H5N2 experimentresembled those in the H1N1–H5N1 experiment (Fig. 4). The H5N2challenge control pigs only showed considerable lymphoprolifera-tive responses after the challenge with H5N2. The H1N1-immune
pigs already had high SIs to both subtypes examined before chal-lenge, with the strongest response to H1N1 (p < 0.05) and a mean12-fold higher SI to H5N2 than the controls (p > 0.05). The responsesto H1N1 decreased or remained similar PC, while those to H5N2increased.enza viruses before and after the challenge with H5N1 in challenge control and
6336 K. Van Reeth et al. / Vaccine 27 (2009) 6330–6339
F t influH
3
uc4a
ietHttgc(b
o
Fp
ig. 4. Individual data points for lymphocyte stimulation indexes (SI) to differen1N1-immune pigs. The horizontal lines represent group means.
.4.2. IFN-� secretionFigs. 5 and 6 show IFN-� titres in supernatants after stim-
lation of PBMC with the different viruses in H5N1 and H5N2hallenge experiments respectively. IFN-� titres are only shown at8 h post-stimulation, when differences between challenge controlnd H1N1-immune groups were most pronounced.
In the H5N1 challenge control group, the responses to anynfluenza virus subtype were minimal before challenge. Both pigsxamined post-challenge showed increased IFN-� titres in responseo H5N1 and H5N2 and only slightly lower titres to H1N1. Pigs of the1N1-immune group had high and similar IFN-� titres in response
o all 3 virus subtypes (p > 0.05) before challenge. The mean IFN-�itre in response to H5N1 was 25-fold higher than in the controlroup at this time (p < 0.05). IFN-� titres did not change much post-
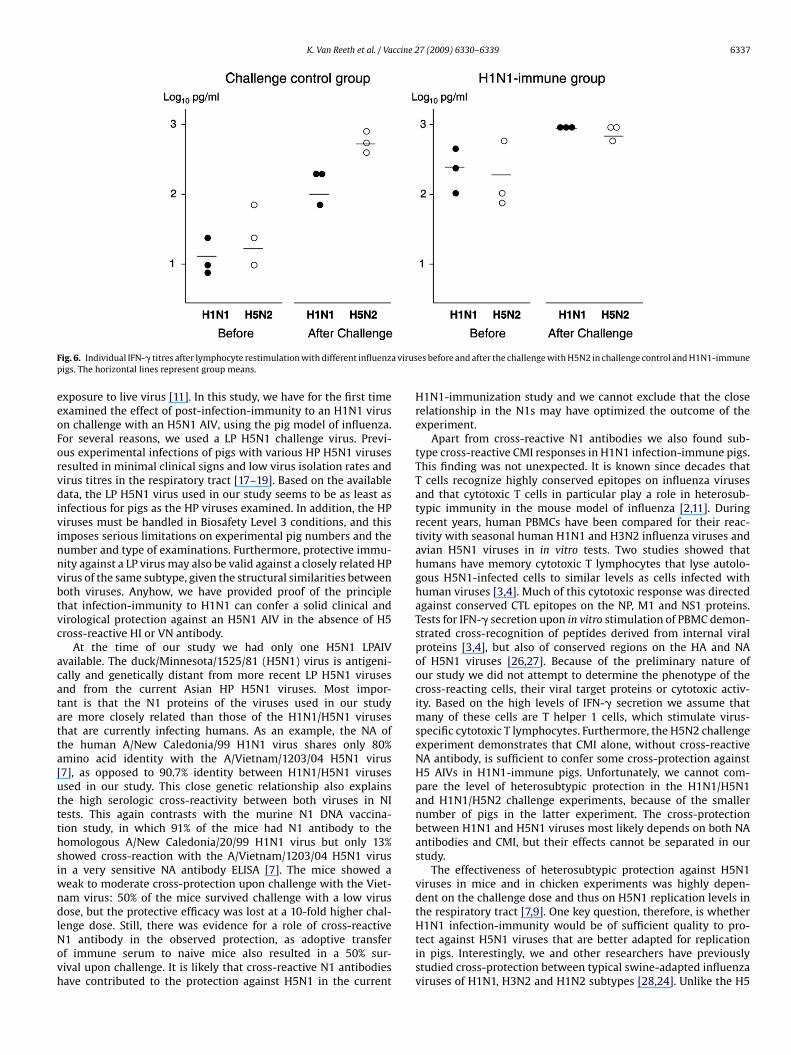
hallenge but they were already increased at 18 h post-stimulationdata not shown), which points towards a more rapid response thanefore challenge or in the control group.Also in the H5N2 challenge experiment, pigs of the control groupnly showed considerable IFN-� production after the H5N2 chal-
ig. 5. Individual IFN-� titres after lymphocyte restimulation with different influenza virusigs. The horizontal lines represent group means.
enza viruses before and after the challenge with H5N2 in challenge control and
lenge, and the responses tended to be higher to H5N2 than to H1N1(p < 0.05). Pigs of the H1N1-immune group had high and compara-ble IFN-� titres (p > 0.05) to both subtypes before challenge, with a5-fold higher mean IFN-� production in response to H5N2 than thecontrol group. The responses to both viruses increased only slightlyafter the challenge.
4. Discussion
Humans who have never been exposed to H5N1 AIVs gener-ally lack neutralizing antibodies to the H5 HA. However, they mayshow antibodies to the N1 of H5N1 viruses or, even more frequently,cross-reactive T cell responses [6,7,3,4]. These immune responsesmay result from natural infection with human H1N1 and/or H3N2
viruses, from vaccination with the trivalent seasonal influenza vac-cine, or a combination of both. Infection with H1N1 virus is likelyto be the prime inducer of cross-reactive immunity to H5N1, giventhe common N1 NA and the fact that inactivated influenza vaccinesare poorer inducers of NA antibodies and cellular immunity thanes before and after the challenge with H5N1 in challenge control and H1N1-immune
K. Van Reeth et al. / Vaccine 27 (2009) 6330–6339 6337
F a virusp
eeoForvdivinnvbtvc
acatatta[uttthsiwndlNovh
ig. 6. Individual IFN-� titres after lymphocyte restimulation with different influenzigs. The horizontal lines represent group means.
xposure to live virus [11]. In this study, we have for the first timexamined the effect of post-infection-immunity to an H1N1 virusn challenge with an H5N1 AIV, using the pig model of influenza.or several reasons, we used a LP H5N1 challenge virus. Previ-us experimental infections of pigs with various HP H5N1 virusesesulted in minimal clinical signs and low virus isolation rates andirus titres in the respiratory tract [17–19]. Based on the availableata, the LP H5N1 virus used in our study seems to be as least as
nfectious for pigs as the HP viruses examined. In addition, the HPiruses must be handled in Biosafety Level 3 conditions, and thismposes serious limitations on experimental pig numbers and theumber and type of examinations. Furthermore, protective immu-ity against a LP virus may also be valid against a closely related HPirus of the same subtype, given the structural similarities betweenoth viruses. Anyhow, we have provided proof of the principlehat infection-immunity to H1N1 can confer a solid clinical andirological protection against an H5N1 AIV in the absence of H5ross-reactive HI or VN antibody.
At the time of our study we had only one H5N1 LPAIVvailable. The duck/Minnesota/1525/81 (H5N1) virus is antigeni-ally and genetically distant from more recent LP H5N1 virusesnd from the current Asian HP H5N1 viruses. Most impor-ant is that the N1 proteins of the viruses used in our studyre more closely related than those of the H1N1/H5N1 viruseshat are currently infecting humans. As an example, the NA ofhe human A/New Caledonia/99 H1N1 virus shares only 80%mino acid identity with the A/Vietnam/1203/04 H5N1 virus7], as opposed to 90.7% identity between H1N1/H5N1 virusessed in our study. This close genetic relationship also explainshe high serologic cross-reactivity between both viruses in NIests. This again contrasts with the murine N1 DNA vaccina-ion study, in which 91% of the mice had N1 antibody to theomologous A/New Caledonia/20/99 H1N1 virus but only 13%howed cross-reaction with the A/Vietnam/1203/04 H5N1 virusn a very sensitive NA antibody ELISA [7]. The mice showed a
eak to moderate cross-protection upon challenge with the Viet-am virus: 50% of the mice survived challenge with a low virusose, but the protective efficacy was lost at a 10-fold higher chal-
enge dose. Still, there was evidence for a role of cross-reactive1 antibody in the observed protection, as adoptive transferf immune serum to naive mice also resulted in a 50% sur-ival upon challenge. It is likely that cross-reactive N1 antibodiesave contributed to the protection against H5N1 in the current
es before and after the challenge with H5N2 in challenge control and H1N1-immune
H1N1-immunization study and we cannot exclude that the closerelationship in the N1s may have optimized the outcome of theexperiment.
Apart from cross-reactive N1 antibodies we also found sub-type cross-reactive CMI responses in H1N1 infection-immune pigs.This finding was not unexpected. It is known since decades thatT cells recognize highly conserved epitopes on influenza virusesand that cytotoxic T cells in particular play a role in heterosub-typic immunity in the mouse model of influenza [2,11]. Duringrecent years, human PBMCs have been compared for their reac-tivity with seasonal human H1N1 and H3N2 influenza viruses andavian H5N1 viruses in in vitro tests. Two studies showed thathumans have memory cytotoxic T lymphocytes that lyse autolo-gous H5N1-infected cells to similar levels as cells infected withhuman viruses [3,4]. Much of this cytotoxic response was directedagainst conserved CTL epitopes on the NP, M1 and NS1 proteins.Tests for IFN-� secretion upon in vitro stimulation of PBMC demon-strated cross-recognition of peptides derived from internal viralproteins [3,4], but also of conserved regions on the HA and NAof H5N1 viruses [26,27]. Because of the preliminary nature ofour study we did not attempt to determine the phenotype of thecross-reacting cells, their viral target proteins or cytotoxic activ-ity. Based on the high levels of IFN-� secretion we assume thatmany of these cells are T helper 1 cells, which stimulate virus-specific cytotoxic T lymphocytes. Furthermore, the H5N2 challengeexperiment demonstrates that CMI alone, without cross-reactiveNA antibody, is sufficient to confer some cross-protection againstH5 AIVs in H1N1-immune pigs. Unfortunately, we cannot com-pare the level of heterosubtypic protection in the H1N1/H5N1and H1N1/H5N2 challenge experiments, because of the smallernumber of pigs in the latter experiment. The cross-protectionbetween H1N1 and H5N1 viruses most likely depends on both NAantibodies and CMI, but their effects cannot be separated in ourstudy.
The effectiveness of heterosubtypic protection against H5N1viruses in mice and in chicken experiments was highly depen-dent on the challenge dose and thus on H5N1 replication levels inthe respiratory tract [7,9]. One key question, therefore, is whether
H1N1 infection-immunity would be of sufficient quality to pro-tect against H5N1 viruses that are better adapted for replicationin pigs. Interestingly, we and other researchers have previouslystudied cross-protection between typical swine-adapted influenzaviruses of H1N1, H3N2 and H1N2 subtypes [28,24]. Unlike the H5
6 ccine
AtpvtHaiwsHfcph1awmiaatnn
tiplsiilsrapvit
awtahwr
A
aa1
R
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
338 K. Van Reeth et al. / Va
IVs used in our and other pig infection experiments [19,17,20],hese SIVs replicate consistently in all parts of the porcine res-iratory tract, at titres of 5.0 up to 8.0 ID50/g and with minimalariation between pigs. In one experiment we examined the pro-ective effect of a prior infection with H3N2 SIV on challenge with1N2 SIV, using virus isolates with a similar genetic relationships in the current H1N1/H5N1 experiment and a similar exper-mental design [24]. Protection against the H1N2 SIV, however,
as much weaker than that against the H5N1 AIV and the acutetage of H1N2 virus replication in the lungs was not affected in3N2-immune pigs. Still, there was a more rapid virus clearance
rom the lungs and a 2-d shorter nasal virus excretion than inhallenge control pigs and part of the pigs also showed clinicalrotection. Similarly, the population immunity against the H2N2uman influenza virus could not prevent an H3N2 pandemic in968, though some data point towards an impact on morbiditynd mortality [29,30]. Logic thus dictates that only immunizationith H5 viruses can induce near to complete protection against aammalian-adapted H5N1 virus. At the same time, heterosubtypic
mmunity could be sufficient to reduce virus replication and thessociated disease, and this may blunt the severity of a pandemic. Inddition, some types of inactivated influenza vaccine may enhancehe cross-reactivity of post-infection immune responses, thoughatural infection induces clearly the broadest immune response inaive subjects.
Though hardly used, pigs offer a valuable influenza immunopro-ection model. They are natural influenza virus hosts and, unliken mouse models, the pathogenesis and symptoms of influenza inigs resemble those in humans. Pigs are anatomically and physio-
ogically similar to man, also in the pulmonary system, and bothpecies reach a similar body weight in their adult life. Pigs aremmunologically closer to humans than mice [31] and many porcinemmunological reagents have become available, which is still aimitation in the ferret model of influenza [32,33]. Finally, hetero-ubtypic protection against influenza viruses appears to occur moreeadily in mice than in humans or in pigs and some researchersrgue that mice are not the model of choice to study cross-subtyperotection [33]. While mice continue to be essential for influenzairus research, potential immunization strategies against pandemicnfluenza viruses should also be tested in other animal species likehe pig.
In conclusion, we have demonstrated protection against poorlydapted H5N1 and H5N2 AIVs in pigs infected 4 weeks earlierith an H1N1 SIV. Our data further support the notion that part of
he human population may have some degree of cross-protectiongainst H5N1 because of exposure to H1N1 viruses. On the otherand, it remains to be proven whether significant cross-protectionould also occur against H5N1 viruses that are better adapted for
eplication in mammals or with a more distant N1.
cknowledgements
We thank Fernand De Backer and Geert Opsomer for help withnimal experiments, Lieve Sys and Melanie Bauwens for technicalssistance, and Dr. R. Webster for supplying the duck/Minnesota/525/81 (H5N1) virus.
eferences
[1] WHO website. Cumulative number of confirmed human cases of avianinfluenza A/(H5N1) reported to WHO, <http://www.who.int/csr/disease/avianinfluenza/country/cases table 2009 01 14/en/index.html>; 2009 [accessed on
14.01.09].[2] Grebe KM, Yewdell J, Bennink JR. Heterosubtypic immunity to influenza A virus:where do we stand? Microbes Infect 2008;10:1024–9.
[3] Jameson J, Cruz J, Terajima M, Ennis FA. Human CD8+ and CD4+ T lympho-cyte memory to influenza A viruses of swine and avian species. J Immunol1999;162:7578–83.
[
[
27 (2009) 6330–6339
[4] Kreijtz JHCM, de Mutsert G, van Baalen A, Fouchier RAM, Osterhaus ADME,Rimmelzwaan GF. Cross-recognition of avian H5N1 influenza virus by humancytotoxic T-lymphocyte populations directed to human influenza A virus. J Virol2008;82:5161–6.
[5] Johansson BE, Brett IC. Changing perspective on immunization againstinfluenza. Vaccine 2007;25:3062.
[6] Webby RJ, Sandbulte MR. A role for neuraminidase antibodies in protectionfrom pandemic influenza? In: Tripp RA, editor. Immunobiology of influenzainfection: approaches for an emerging zoonotic disease. Kerala, India: ResearchSignpost; 2008. p. 17–24.
[7] Sandbulte MR, Jimenez GS, Boon AC, Smith LR, Treanor JJ, Webby RJ. Cross-reactive neuraminidase antibodies afford partial protection against H5N1in mice and are present in unexposed humans. PLoS Med 2007;4(2):e59,doi:10.1371/journal.pmed.00400959.
[8] O’Neill E, Krauss SL, Riberdy JM, Webster RG, Woodland DL. Heterologous pro-tection against lethal A/HongKong/156/97 (H5N1) influenza virus infection inC57BL/6 mice. J Gen Virol 2000;81:2689–96.
[9] Seo SH, Webster RG. Cross-reactive, cell-mediated immunity and protectionof chickens from lethal H5N1 influenza virus infection in Hong Kong poultrymarkets. J Virol 2001;75:2516–25.
10] Seo SH, Peiris M, Webster RG. Protective cross-reactive cellular immunity tolethal A/Goose/Guandong/1/96-like H5N1 influenza virus is correlated with theproportion of pulmonary CD8(+) T cells expressing gamma interferon. J Virol2002;76:4886–90.
[11] Tamura S, Tanimoto T, Kurata T. Mechanisms of broad cross-protection providedby influenza virus infection and their application to vaccines. Jpn J Infect Dis2005;58:195–207.
12] Sambhara S, Kurichh A, Miranda R, Tumpey T, Rowe T, Renshaw M, et al. Het-erosubtypic immunity against human influenza A viruses, including recentlyemerged avian H5 and H9 viruses, induced by FLU-ISCOM vaccine in micerequires both cytotoxic T-lymphocyte and macrophage function. Cell Immunol2001;211:143–53.
13] Tumpey TM, Renshaw M, Clements JD, Katz JM. Mucosal delivery of inactivatedinfluenza vaccine induces B-cell-dependent heterosubtypic cross-protectionagainst lethal influenza A H5N1 virus infection. J Virol 2001;75:5141–50.
14] Ichinohe T, Tamura S, Kawaguchi A, Ninomiya A, Imai M, Itamura S, etal. Cross-protection against H5N1 influenza virus infection is afforded byintranasal inoculation with seasonal trivalent inactivated influenza vaccine. JID2007;196:1313–20.
15] Takada A, Matsushita S, Ninomiya A, Kawaoka Y, Kida H. Intranasal immu-nization with formalin-inactivated virus vaccine induces a broad spectrum ofheterosubtypic immunity against influenza A virus infection in mice. Vaccine2003;21:3212–8.
16] Olsen CW, Brown IH, Easterday BC, Van Reeth K. Swine influenza. In: Straw BE,Zimmerman JJ, d’Allaire S, Taylor SJ, editors. Disease of swine. Oxford: BlackwellPublishing; 2006. p. 469–82.
17] Choi YK, Nguyen TD, Ozaki H, Webby RJ, Puthavathana P, Buranathal C, et al.Studies of H5N1 influenza virus infection of pigs by using viruses isolated inVietnam and Thailand in 2004. J Virol 2005;79:10821–5.
18] Isoda N, Sakoda Y, Kishida N, Bai GR, Matsuda K, Umemura T, et al. Pathogenic-ity of a highly pathogenic avian influenza virus. A/chicken/Yamaguchi/7/04(H5N1) in different species of birds and mammals. Arch Virol 2006;151:1267–79.
19] Lipatov AS, Kwon YK, Sarmento LV, Lager KM, Spackman E, Suarez DL, etal. Domestic pigs have low susceptibility to H5N1 highly pathogenic avianinfluenza viruses. PloS Pathog 2008;4(7):e1000102.
20] De Vleeschauwer A, Barbé F, Van Reeth K. Pathogenesis of low pathogenicH5N2 avian influenza virus in pigs. In: Proceedings of the 5th InternationalSymposium on Emerging and Re-emerging Pig Diseases. 2007. p. 241.
21] Hoffmann E, Stech J, Guan Y, Webster RG, Perez DR. Universal primerset for the full-length amplification of all influenza A viruses. Arch Virol2001;146:2275–89.
22] Hall TA. BioEdit: a user-friendly biological sequence alignment editor and anal-ysis program for Windows 95/98/NT. Nucleic Acids Symp Ser 1999;41:95–8.
23] Kumar S, Tamura K, Nei M. MEGA3: integrated software for molecu-lar evolutionary genetic analysis and sequence alignment. Brief Bioinform2004;5:150–63.
24] Van Reeth K, Gregory V, Hay A, Pensaert M. Protection against a EuropeanH1N2 swine influenza virus in pigs previously infected with H1N1 and/or H3N2subtypes. Vaccine 2003;21:1375–81.
25] Aymard-Henry M, Coleman MT, Dowdle WR, Laver WG, Schild GC, Webster RG.Influenzavirus neuraminidase and neuraminidase-inhibition test procedures.Bull World Health Organ 1973;48:199–202.
26] Goy K, Von Bibra S, Lewis J, Laurie K, Barr I, Anderson D, et al. Heterosub-typic T-cell responses against avian influenza H5 haemagglutinin are frequentlydetected in individuals vaccinated against or previously infected with humansubtypes of influenza. IORV 2008;2:115–25.
27] Gioia C, Castilletti C, Tempestilli M, Piacentini P, Bordi L, Chiappini R, et al. Cross-subtype immunity against avian influenza in persons recently vaccinated forinfluenza. Emerg Infect Dis 2008;14:121–8.
28] Heinen PP, de Boer-Luijtze E, Bianchi ATJ. Respiratory and systemic humoraland cellular immune responses of pigs to a heterosubtypic influenza A virusinfection. J Gen Virol 2001;82:2697–707.
29] Murphy BR, Kasel JA, Chanock RM. Association of serum anti-neuraminidaseantibody with resistance to influenza in man. N Engl J Med 1972;286:1329–32.

ccine 2
[
[
K. Van Reeth et al. / Va
30] Gillim-Ross L, Subbarao K. Can immunity induced by the human influenza virusN1 neuraminidase provide some protection from avian influenza H5N1 viruses?PloS Med 2007;4(2):e91, doi:10.1371/journal.pmed.0040091.
31] Mestas J, Hughes CCW. Of mice and not men: differences between mouse andhuman immunology. J Immunol 2004;172:2731–8.
[
[
7 (2009) 6330–6339 6339
32] Renegar KB. Influenza virus infections and immunity: a review of human andanimal models. Lab Anim Sci 1992;42:222–32.
33] van der Laan JW, Herberts C, Lambkin-Williams R, Boyers A, Mann AJ,Oxford J. Animal models in influenza vaccine testing. Expert Rev Vaccines2008;7:783–93.