postnatal histomorphogenesis of the mandible in the house mouse
TRANSCRIPT

Postnatal histomorphogenesis of the mandible in thehouse mouseCayetana Martinez-Maza,1,2,3 Laetitia Montes,1,2 Hayat Lamrous,1,2 Jacint Ventura4 andJorge Cubo1,2
1UPMC Univ Paris 06, UMR 7193, ISTEP, Paris, France2CNRS, UMR 7193, ISTEP, Paris, France3Department of Paleobiology, Museo Nacional de Ciencias Naturales – CSIC, Madrid, Spain4Departament de Biologia Animal, de Biologia Vegetal i d’Ecologia, Facultat de Biociencies, Universitat Autonoma de
Barcelona, Barcelona, Spain
Abstract
The mandible of the house mouse, Mus musculus, is a model structure for the study of the development and
evolution of complex morphological systems. This research describes the histomorphogenesis of the house
mouse mandible and analyses its biological significance from the first to the eighth postnatal weeks. Histologi-
cal data allowed us to test a hypothesis concerning modularity in this structure. We measured the bone growth
rates by fluorescent labelling and identified the bone tissue types through microscopic analysis of histological
cross-sections of the mandible during its postnatal development. The results provide evidence for a modular
structure of the mouse mandible, as the alveolar region and the ascending ramus show histological differences
throughout ontogeny. The alveolar region increases in length during the first two postnatal weeks by bone
growth in the posterior region, while horizontally positioned incisors preclude bone growth in the anterior
region. In the fourth postnatal week, growth dynamics shows a critical change. The alveolar region drifts later-
ally and the ramus becomes more vertical due to the medial growth direction of the coronoid region and the
lateral growth of the ventral region of the ramus. Diet changes after weaning are probably involved in these
morphological changes. In this way, the development of the masticatory muscles that insert on the ascending
ramus may be particularly related to this shape modeling of the house mouse mandible.
Key words: bone histology; development; mandible; modularity; morphology; Mus musculus; postnatal growth.
Introduction
The house mouse, Mus musculus, represents a well estab-
lished model in mammalian biological research. In particu-
lar, the mandible has been the subject of genetic,
embryological, and functional anatomical studies providing
an extensive background on mechanisms explaining pheno-
typic variation (Atchley & Hall, 1991). Many works have
used the house mouse mandible as a model system to study
the development and evolution of complex morphological
structures (Atchley & Hall, 1991; Atchley, 1993; Klingenberg
& Leamy, 2001; Klingenberg, 2002; Klingenberg et al. 2004;
Monteiro et al. 2005). As well, functional approaches have
shown the influence of the masticatory and paramasticato-
ry activities on the morphology of the mammalian mandi-
ble (Kesner, 1980; Satoh, 1997; Ravosa et al. 2007; Enomoto
et al. 2010; Renaud et al. 2010; Ventura & Casado-Cruz,
2011). However, the histomorphogenesis and the bone
growth dynamics associated with the morphology of the
mouse mandible have never been analyzed.
The mandible originates from neural crest cells that
migrate to form mesenchymal condensations that differen-
tiate into the six major morphogenetic units (the ramus, the
molar and incisor alveolar components, and the coronoid,
condylar and angular processes) and Meckel’s cartilage
(Atchley & Hall, 1991; Ramaesh & Bard, 2003). The early
alveolar region forms by intramembranous ossification
surrounding Meckel’s cartilage and the processes are
formed by secondary cartilages located at the proximal end
that are replaced later on by bone through endochondral
Correspondence
Cayetana Martinez-Maza, Department of Paleobiology, Museo
Nacional de Ciencias Naturales – CSIC, Jose Gutierrez Abascal 2,
28006 Madrid, Spain. T: + 34 91 5668981; E: martinezmaza.
[email protected] and [email protected]
Jorge Cubo, Universite Pierre et Marie Curie, Equipe Biomineralisa-
tions et Environnements Sedimentaires – UMR7193. 4, Pl. Jussieu BC
19, 75005 Paris, France. T: + 33 14 4273124; E: jorge.cubo_garcia@
upmc.fr
Accepted for publication 1 February 2012
Article published online 28 February 2012
ªª 2012 The AuthorsJournal of Anatomy ªª 2012 Anatomical Society
J. Anat. (2012) 220, pp472–483 doi: 10.1111/j.1469-7580.2012.01488.x
Journal of Anatomy

ossification (Hall, 2003; Ramaesh & Bard, 2003). Later in the
intrauterine development, the mandible grows and two pri-
mary functional units or modules are identified: the alveo-
lar region, which is the anterior part bearing the teeth, and
the ascending ramus, which articulates with the cranium
and provides surfaces for muscle attachment (Atchley &
Hall, 1991). Geometric morphometric analyses have sup-
ported the division of the mandible into these two develop-
mental modules ‘that are separate from each other but
they are not completely independent’ (Klingenberg et al.
2003; see also Zelditch et al. 2008; Klingenberg, 2009).
After birth, the mouse mandible undergoes morphologi-
cal changes involving size, shape, and its relative position
into the craniofacial system to fulfill the functional require-
ments (Moss, 1960; Enlow & Hans, 1996). These morphologi-
cal changes are often mediated by the action of soft tissues
(e.g. muscular insertions) and the external (e.g. biomechani-
cal) and internal stimuli (e.g. hormonal factors; Moss, 1960;
Enlow & Hans, 1996). Bone growth occurs through the
modeling mechanism involving the coordinated activity of
osteoblasts (bone-producing cells) and osteoclasts (bone-
removing cells) (e.g. Bloom & Fawcett, 1994; Enlow & Hans,
1996). Osteoblasts or bone-forming cells secrete an organic
matrix mainly composed by collagen fibers and then bio-
mineralize it with calcium phosphate. After mineralization
of the matrix, osteoblasts lose their ability to undergo mito-
sis and become osteocytes, which communicate with each
other through a network of canaliculi. Osteoclasts are mul-
tinucleated cells that remove the bone mineral and degrade
the organic bone matrix. These cellular activities are
recorded in the bone tissue as structural features containing
information about morphogenetic changes (bone tissue
types reflecting bone growth rates; Amprino, 1947). There-
fore, histological organization of bone tissue provides infor-
mation about the growth directions and rates of the
functional regions and developmental modules underlying
the genesis of the mandibular shape. Previous works have
approached the analysis of the postnatal development from
a histological perspective in the mandible of the mink
(Buffrenil & Pascal, 1984), rabbit (Bang & Enlow, 1967), and
several species of primates (Enlow, 1963; Enlow & Hans,
1996). These studies have provided information on both the
bone tissue structure and the bone growth rates of differ-
ent mandible regions (Buffrenil & Pascal, 1984) as well as
the bone growth directions (Bang & Enlow, 1967; Enlow &
Hans, 1996). Histological data obtained for each species
have pointed out the particular growth dynamics explain-
ing the morphological changes of the mandible during the
postnatal development.
The aim of this study is to characterize the histomorpho-
genesis of the house mouse mandible from birth to the
eighth postnatal week analyzing the bone histology, the
growth directions and the bone growth rates. These results
are discussed in the light of similar studies to infer specific
bone growth dynamics and to establish a bone growth
model for the normal postnatal development of the
M. musculus mandible. In addition, we will use these histo-
logical data to test the division of the mouse mandible into
the alveolar and the ascending ramus modules as hypothe-
sized in prior studies (e.g. Atchley & Hall, 1991; Klingenberg
et al. 2003). According to this hypothesis and considering
the developmental and functional differences proposed for
these two modules (Atchley & Hall, 1991), we predict that
both modules will differ in their histological organization
of bone tissue and bone growth rates. Understanding how
these modules grow and interrelate during development
will provide useful information about the biological pro-
cesses underlying the development and evolution of mor-
phological structures in the house mouse mandible.
Materials and methods
Thirty-eight mice of the strain C57BL ⁄ 6J (Janvier Inc., France)
from the first to eight postnatal weeks were used in this study.
Mice were housed in standard cages in an animal room under
controlled environmental conditions and provided with food
and water ad libitum. Animals were handled in accordance with
the European Communities Council Directive (86 ⁄ 609 ⁄ EEC). In
this study, only female mice were employed. According to most
literature, no sexual dimorphism is observed in mice, although
Renaud et al. (2010) have suggested that strain C57BL ⁄ 10 pre-
sents some degree of sexual dimorphism that may affect model-
ing processes. However, a detailed analysis of strain C57BL ⁄ 6J
has yielded inconclusive results (Boell et al. 2011). Thus, we
assume that sexual dimorphism will be very limited, particularly
in young animals.
To study the periosteal growth rates, mice received an intra-
peritoneal injection of dicarboxymethyl aminomethyl fluores-
cein (DCAF) 25 mg kg)1 of body weight. These fluorescent dyes
specifically color the mineralizing zone of growing bone tissue.
A week after the intraperitoneal injection of DCAF, mice were
sacrificed by cervical dislocation. Mandibles were dissected and
cleaned by hand. They were dehydrated in graded ethanol, de-
fatted in acetone and trichloroethylene and finally dried at 38–
40 �C in a stove. Right mandibles were embedded in a polyester
resin. Histological cross-sections 100 ± 10 lm thick were made
using a diamond-tipped circular saw in four representative man-
dibular regions: diastema, first molar, second molar, and ascend-
ing ramus at the level of the coronoid process. Each thin section
was ground and polished before being mounted on a slide.
They were observed under ultraviolet and natural light micro-
scope (Zeiss Axiovert 35; Jena, Germany) and digitalized
through a camera (Olympus, Japan). To determine the perio-
steal growth rates (lm day)1), pictures taken under ultraviolet
light were analyzed through the image processing program IMA-
GEJ. The bone growth rate was obtained using the distance
between the DCAF-label (fluorescein) and the bone surface
divided by the time elapsed (7 days). Figure 1 shows the points
in the histological cross-sections used to calculate this rate.
Results
Histological cross-sections (diastema, first molar, second
molar, and ascending ramus) from the first to the eighth
ªª 2012 The AuthorsJournal of Anatomy ªª 2012 Anatomical Society
Postnatal growth of the mouse mandible, C. Martinez-Maza et al. 473

postnatal week are shown in Figs 2–5. To provide a detailed
description, we first describe the bone histology of the four
mandibular regions and then analyze the bone growth
dynamics (growth directions and rates). Finally, we use the
spatial and temporal distribution of bone tissue types and
modeling activities in the mandible to test the modular
structure proposed in previous studies (Atchley & Hall, 1991;
Cheverud et al. 1997; Mezey et al. 2000; Klingenberg et al.
2003).
Histological description
Diastema region
In the first and second postnatal weeks, this mandibular
region showed woven bone tissue (Fig. 6A) with low
ordered spatial arrangement of the collagen fiber bundles,
randomly distributed rounded osteocytic lacunae, and high
vascularity (Fig. 2: diastema, weeks 1 and 2; woven bone:
label w). The third week was characterized by a histological
change consistent with a more parallel-fibered arrange-
ment of collagen fibers including rounded osteocytic lacu-
nae in the dorsal half of the labial side (Fig. 2: diastema
week 3; parallel-fibered bone: label pf) and with flattened
osteocytic lacunae in an aligned distribution in the lingual
side (Fig. 2: diastema week 3, label pf). The diastema
showed woven bone tissue with high vascularity and ran-
domly distributed osteocytic lacunae (Fig. 2: diastema, week
3, label w) dorsally and in the ventral half of the labial side.
In the fourth week, the dorsal region and ventral half of
the labial side showed woven bone tissue (Fig. 2: diastema,
week 4, label w), while the dorsal half of the labial side dis-
played parallel-fibered bone tissue (Fig. 2: diastema, week
4, label pf; Fig. 6b). The lingual side displayed an internal
region (endosteal) of woven bone tissue (Fig. 2: diastema,
week 4, label w), whereas the outer region (periosteal)
showed parallel-fibered bone tissue with flattened osteocy-
tic lacunae in an ordered disposition (Fig. 2: diastema, week
4, label pf). In addition, the bone histology structure and
the DCAF label of the vascular cavity located in the dorsal
internal ridge suggest a centripetal bone growth associated
with the formation of the primary osteon (Fig. 2: diastema,
week 4; primary osteon: label po). The diastema region in
the fifth postnatal week showed an increase of the woven
bone tissue in the lingual and the labial sides (Fig. 2: dia-
stema week 5, w), whereas the parallel-fibered bone tissue
was located in the dorsal area of this region (Fig. 2: dia-
stema week 5, pf). The vascular cavity of the dorsal internal
ridge showed concentric bone lamellae related to the cen-
tripetal bone growth that could be involved in the forma-
tion of a primary osteon (Fig. 2: diastema, week 5, po). In
the sixth and seventh postnatal weeks, the diastema region
showed parallel-fibered bone tissue with flattened osteocy-
tic lacunae in the outer region (periosteal) of the labial and
lingual sides (Fig. 2: diastema, week 6 and diastema, week
7, pf), whereas woven bone tissue was observed in the
internal region (endosteal) of labial sides (Fig. 2: diastema,
week 6 and 7, label w) and in the ventral region (Fig. 2: dia-
stema, week 7, label w). The distribution of the bone tissues
and the concentric lamellae observed around some vascular
cavities or primary osteons (Fig. 2A: diastema, week 6 and
7, po) suggest a decrease of the bone growth in this postna-
tal week. In the eighth postnatal week, the labial and lin-
gual sides of the diastema region showed parallel-fibered
bone tissue with flattened osteocytic lacunae (Fig. 2: D8,
Fig. 1 Histological cross-sections from four regions of the Mus musculus mandible (from left to right): diastema, first molar region, second molar
region, and ascending ramus. Pictures taken under ultraviolet microscope show the green label (DCAF). Numbers on the DCAF line denote the
points used to measure the bone growth rates in this study. Scale bar: 1 mm.
ªª 2012 The AuthorsJournal of Anatomy ªª 2012 Anatomical Society
Postnatal growth of the mouse mandible, C. Martinez-Maza et al.474

label pf), whereas the ventral and dorsal regions displayed
woven bone tissue with rounded osteocytic lacunae
randomly distributed (Fig. 2: diastema, week 8, label w).
The first and second molar regions
Both regions showed, in the first and second postnatal
weeks, woven bone tissue with high vascularity (Fig. 2: first
molar, weeks 1 and 2 and Fig. 3: second molar, weeks 1
and 2, label w), which is related to high osteogenesis. In
the third week, mandibles showed a histological change in
the first molar region but not in the second one. The first
molar region displayed woven bone tissue in the ventral
half of the labial side (Fig. 2: first molar, week 3, label w),
whereas the dorsal half of this side and the lingual side
Fig. 2 Histological cross-sections under natural light of the diastema region (bottom) and the first molar region (top) obtained in the Mus
musculus mandible from the first (right) to the eighth postnatal week (left). Labels indicate the bone tissue type observed in different areas of
each mandibular region, w: woven bone tissue; pf: parallel-fibered bone tissue; po: primary osteon. Scale bar: 1 mm.
Fig. 3 Histological cross-sections under natural light of the second molar region (bottom) and the ascending ramus region (top) obtained in the
Mus musculus mandible from the first (right) to the eighth postnatal week (left). Labels indicate the bone tissue type observed in different areas of
each mandibular region, w: woven bone tissue; pf: parallel-fibered bone tissue; po: primary osteon. Scale bar: 1 mm.
ªª 2012 The AuthorsJournal of Anatomy ªª 2012 Anatomical Society
Postnatal growth of the mouse mandible, C. Martinez-Maza et al. 475

displayed parallel-fibered bone tissue with rounded osteo-
cytic lacunae in an ordered disposition (Fig. 2: first molar,
week 3, label pf). However, the second molar region was
characterized by woven bone tissue with a random distri-
bution of the osteocytic lacunae (Fig. 3: second molar,
week 3, label w). In the fourth and the fifth weeks, the
region of the first and second molars was characterized by
an increase of parallel-fibered bone tissue with rounded
osteocytic lacunae in an ordered disposition in a small area
of the dorsal half of the labial side and in the lingual side
(Fig. 2: first molar, week 4 and Fig. 3: second molar, week
4, pf), whereas the labial side displayed woven bone tissue
(Fig. 2: first molar, week 4 and Fig. 3: second molar week
4, w). In addition, both labial and lingual sides showed
concentric lamellae in some vascular cavities related to the
formation of primary osteons (Figs 2 and 3, po). The bone
histology of the molar region suggests a high bone growth
rate in the ventral half and slow bone growth in the lin-
gual and the dorsal half of the labial side. In the sixth and
seventh postnatal weeks, the first and second molar
regions showed parallel-fibered bone tissue with flattened
osteocytic lacunae in an ordered disposition on the lingual
side, the ventral and the outer regions (periosteal) of the
dorsal half of the labial side (Fig. 2: first molar, weeks 6
and 7, Fig. 3: second molar, weeks 6 and 7, pf). Woven
bone tissue was identified in two areas of the labial side,
the internal area (endosteal) of the dorsal half, around the
molar root, and close to the mandibular crest in the ven-
tral half (Fig. 2: first molar, weeks 6 and 7, Fig. 3: second
molar, weeks 6 and 7, label w). In the eighth postnatal
week, the molar region showed parallel-fibered bone tis-
sue with flattened osteocytic lacunae on the lingual side,
the ventral region, and in the dorsal half of the labial side
(Fig. 2: first molar, week 8, pf). The ventral half of the
labial side displayed woven bone tissue with randomly dis-
tributed rounded osteocytic lacunae (Fig. 2: first molar,
week 8, w). Also, the second molar region was character-
ized by parallel-fibered bone tissue with flattened osteocy-
tic lacunae in an ordered disposition (Fig. 3: second molar,
week 8, pf) and primary osteons close to the mandibular
crest and in the dorsal area of the lingual side (Fig. 3:
second molar, week 8, label po).
Ascending ramus region
From the first to the fifth postnatal week, the ascending
ramus was characterized by woven bone tissue with
rounded osteocytic lacunae indicating fast bone formation
(Fig. 3: ramus, weeks 1–5, label w). In the third and fifth
weeks, centripetal bone growth was observed in the vascu-
lar cavities, associated with the formation of a primary
osteon (Fig. 3: ramus, weeks 3 and 5, label po). In the sixth
week, the ascending ramus was characterized by woven
bone tissue, except in the labial side of the area surround-
ing the mandibular foramen, which showed parallel-
fibered bone tissue with flattened osteocytic lacunae
(Fig. 3: ramus, week 6, pf). There was a histological change
in the seventh week; the ascending ramus displayed paral-
lel-fibered bone tissue with flattened osteocytic lacunae on
the labial and lingual sides of the area surrounding the
mandibular foramen and in the labial side of the ventral
half of the ramus (Fig. 3: ramus, week 7, label pf), whereas
the coronoid and the lingual side of the ventral half of the
ramus displayed woven bone tissue (Fig. 3: ramus, week 7,
label w). The ascending ramus in the eighth week showed
parallel-fibered bone tissue with flattened osteocytic
lacunae on the labial and lingual side of the area
Fig. 4 Histological cross-sections under ultraviolet light of the diastema region (bottom) and the first molar region (top) obtained in the Mus
musculus mandible, from the first (right) to the eighth postnatal week (left). Labels indicate the bone modeling activity observed in different areas
of each mandibular region, bf: bone formation activity; BR: bone resorption activity. Scale bar: 1mm.
ªª 2012 The AuthorsJournal of Anatomy ªª 2012 Anatomical Society
Postnatal growth of the mouse mandible, C. Martinez-Maza et al.476

surrounding mandibular foramen and on the labial side of
the ventral half of the ramus (Fig. 3: ramus, week 8, pf),
whereas other areas were characterized by a transition
between woven bone and parallel-fibered bone tissue with
randomly distributed, rounded osteocytic lacunae.
Bone growth dynamics
Fluorescence-DCAF labeling provided information about
bone growth directions and rates of different mandibular
regions during postnatal development. On the one hand,
presence or absence of the DCAF-label was associated with
bone formation or resorption activities, respectively. As the
lack of fluorescence label could result from bone resorption
activity in the periosteal or endosteal surface, we analyzed
the profile of the bone surface to identify the region (perio-
steal or endosteal surface) where bone is resorbed (Enlow &
Hans, 1996). Bone growth rates were measured at different
points of each mandibular region (Fig. 1) to determine the
growth changes during development.
Diastema region
The histological data and the DCAF label observed in the
first 3 weeks showed periosteal bone formation surfaces
that indicated a general growth in all directions. The bone
resorption activity in the endosteal surface of the ventral
and lingual sides (Fig. 4: diastema, weeks 1–3, label br)
reveals bone growth in a ventral direction. From the fourth
to the eighth week, the labial side and the ventral area
were characterized by bone formation, but the lingual side
showed a field of bone resorption of variable size (Fig. 4:
diastema, weeks 4–8, label br). This bone modeling field dis-
tribution was associated with a lateral growth of the dia-
stema.
The bone growth rate data showed a high rate of osteo-
genesis during the first three postnatal weeks, followed
by a marked decrease of the rates in the fourth week and,
from the fifth week on, growth slowed gradually until the
eighth postnatal week. A thorough analysis of the bone
growth rates throughout development allows us to distin-
guish three areas in the diastema. The first area, which
includes the dorsal region and the dorsal half of the labial
side (from point 1 to 5, Fig. 1), was characterized by low
growth rate (4–5 lm day)1) in the first and second weeks, a
slight increase in the third and fourth weeks, mainly occur-
ring on the labial side (from point 4 to 5), and a gradual
decrease of the bone growth in the last weeks
(< 1 lm day)1). The second (the ventral half of the labial
side from point 6 to 11, labial side), showed a high bone
growth rate during the first four postnatal weeks and then
a gradual decrease of the rate with age (< 2 lm day)1). Part
of the ventral border of the diastema region (from point 9
to 11) displayed a high growth rate (> 20 lm day)1) during
the first 3 weeks. Growth of this area decreased abruptly in
the fourth week (� 9 lm day)1) and subsequently showed
a gradual decrease, with growth rates similar to those of
other points of this region. The third area corresponds to
the lingual side (from point 12 to 18) and is characterized
by lower ventral bone growth rates (< 3 lm day)1) than the
growth registered on the labial side during the first 3 weeks
of development. From the fourth week to the eighth, this
lingual side showed bone resorption.
First and second molar regions
These mandibular regions showed similar growth directions
throughout postnatal development. The DCAF label indi-
cated periosteal bone formation surfaces (see label bf in
Fig. 4: first molar and Fig. 5: second molar) but in the first
3 weeks, the lack of the DCAF label associated with bone
resorption activity in the ventro-lingual area of the endos-
teal surface indicated a dorso-ventral growth (Fig. 4: first
molar, weeks 1–3 and Fig. 5: second molar, weeks 1–3, label
br). From the fourth to the sixth weeks, the molar regions
showed bone resorption on the lingual side of their alveoli.
During the seventh and eighth weeks, mandibles showed
bone formation surfaces in both molar regions but bone
resorption activity occurred in the lingual side, as revealed
by the lack of the DCAF label and the histological data (see
label br in Fig. 4: first molar and Fig. 5: second molar, weeks
7–8).
The analysis of bone growth rates revealed differences
between the first and second molar regions. The first molar
region showed an increase of bone growth rates from the
first to the third postnatal weeks but subsequently the
growth rates decrease gradually until the eighth week. In
this mandibular region, four areas characterized by a partic-
ular variation of the bone growth rates during develop-
ment were identified. The first area corresponds with the
dorsal half of the labial side (from point 1 to 5) and showed
a gradual decrease of the bone growth (from 4–5 to
< 1 lm day)1). The second area (the mandibular crest of
the labial side from point 6 to 8) showed high growth rates
during the first 2 weeks (20–16 lm day)1), a decrease in
growth rates in the fourth week (6 lm day)1) and a marked
new decrease in the sixth week (2 lm day)1). The third area
included the ventral border (from point 9 to 11) and
showed high bone growth rates during the first two post-
natal weeks and in the fifth week (11–15 lm day)1). Both
in the third and fourth weeks and from the sixth postnatal
week onwards, the bone growth rates decreased in this
mandibular region (< 4 lm day)1). The fourth area was
associated with the lingual side (from point 12 to 17) and
showed lower ventral growth rates than in the labial side.
This side of the first molar displayed a gradual decrease in
bone growth rates during development (< 2 lm day)1).
Only the area associated with the incisor alveoli (points 12
and 13) showed a high growth rate (4–6 lm day)1) from
the first to the fifth week and then abruptly decreased
growth rates (� 1 lm day)1) until the eighth postnatal
week.
ªª 2012 The AuthorsJournal of Anatomy ªª 2012 Anatomical Society
Postnatal growth of the mouse mandible, C. Martinez-Maza et al. 477

In the second molar region, three areas with similar varia-
tions in bone growth rate during development were distin-
guished. The first area corresponds with the labial side
(from point 1 to 5) and showed low growth rates in the first
2 weeks (3–4 lm day)1), except in the area of the mandibu-
lar crest (points 4 and 5), which showed high growth rates
(� 16 lm day)1). In this region, all individuals showed a
constant rate (2–3 lm day)1) from the third to the eighth
weeks. The second area, which covers the area from the
labial crest to the ventral border (points 6–10), showed the
highest bone growth rates (10–30 lm day)1) during the first
three postnatal weeks; growth rates gradually decreased
from the fourth to the eighth weeks (� 5 lm day)1). The
third area (points 11–14) was associated with the lingual
side and was characterized by low growth rates
(� 3 lm day)1) that gradually decreased during the postna-
tal development (1 lm day)1).
Ascending ramus region
This region was characterized by the lack of the DCAF label
due to the periosteal and endosteal resorption activity. The
fluorescence label and the histomorphometric data indicate
complex growth dynamics in this region (Fig. 5: ramus
weeks, 1 and 2, label bf). The first and the second weeks
showed the DCAF label only in small areas of the labial side
of the coronoid tip and in the ventral area of the ramus,
which is associated with endosteal bone formation. The lack
of fluorescence was due to high endosteal resorption activ-
ity and suggests growth in all directions by periosteal bone
formation. In the third week, the ramus was entirely deposi-
tory, revealing general growth, but in the labial side of the
coronoid region there was a small area of bone resorption
activity (Fig. 5: ramus week 3, label br). From the fourth to
the eighth postnatal weeks, there was a lack of the DCAF-
label in the labial side of the coronoid tip and in the lingual
side of the ventral half of the ramus (Fig. 5: ramus, weeks
from 4 to 8, label br).
The analysis of bone growth rates was limited by the lack
of the DCAF label. Nevertheless, histomorphometric data
provided information indicating ranges of osteogenesis. In
the first and second postnatal weeks, the ascending ramus
showed regions without fluorescence label that revealed
high bone growth rates with endosteal resorption. These
data were associated with the increase in size of the ramus.
From the third postnatal week on, the ramus showed a
change in bone growth dynamics. In the third week, the
ramus showed bone formation surfaces except on the labial
side of the coronoid (points 1–3), as well as bone resorption
activity. Bone growth rates indicated a generally high
osteogenesis (3–8 lm day)1), particularly on the lingual
side, close to the mandibular foramen (points 15), which
showed the highest rate (> 10 lm day)1) in that week.
From the fourth to the eighth postnatal weeks, bone mod-
eling fields allowed us to distinguish two areas character-
Fig. 5 Cross-sections under ultraviolet light of the second molar region (bottom) and the mandibular ramus region (top) obtained in the Mus
musculus mandible from the first (right) to the eighth postnatal week (left). Labels indicate the bone modeling activity observed in different areas
of each mandibular region, bf: bone formation activity; BR: bone resorption activity. Scale bar: 1 mm.
ªª 2012 The AuthorsJournal of Anatomy ªª 2012 Anatomical Society
Postnatal growth of the mouse mandible, C. Martinez-Maza et al.478

ized by bone resorption fields: the labial side of the coro-
noid (points 1–3) and the lingual side of the ventral area of
the ramus (points 12–14). Likewise, two zones showed bone
formation surfaces: the lingual side of the coronoid (points
18–20) and the labial side of the ventral half of the ramus
(points 6–9). During the last four postnatal weeks, bone
growth rates revealed a general growth with a slight
decrease in rates in the areas with bone formation surfaces,
the lingual side of the coronoid, the mandibular foramen
area (labial and lingual sides), and the labial side of the ven-
tral-half of the ramus.
Modularity in the mouse mandible
The histological data provide new evidence to test the
hypothesis about modularity in the mouse mandible pro-
posed in previous works (e.g. Atchley & Hall, 1991; Klingen-
berg et al. 2003). As illustrated in Table 1 and Fig. 7A,
histological features observed in the diastema, the first and
second molar regions (from here on, we consider these
three regions the alveolar region) are similar to each other
but different from the ascending ramus at different times
of postnatal ontogeny. Although both modules (alveolar
region and ascending ramus) are characterized by woven
bone in the first 2 weeks after birth, the alveolar region
seems to mature earlier and presents fibrolamellar bone,
whereas the ascending ramus retains the woven bone until
the seventh week. Parallel ontogenetic changes are
observed in the modeling patterns (Fig. 7B). Both modules
show bone deposition in their external surfaces during the
first 3 weeks. Later on, the alveolar region shows a consis-
tent pattern characterized by resorption on the lingual side.
On the contrary, the ramus shows a complex pattern char-
acterized by the occurrence of resorption in the labial side
of the coronoid and the lingual side of the ventral area.
Together, bone histological data agree with the existence
of two natural modules showing different developmental
patterns. Differences between both modules become
apparent in the third postnatal week.
Discussion
The mouse mandible is a model structure for the study of
the development and evolution of complex morphological
systems (Atchley & Hall, 1991). In this study, our results on
the microstructure of bone tissue and the bone growth
rates allowed us to determine the histomorphogenesis of
the M. musculus mandible during postnatal development.
In addition, our histological data suggest that the house
mouse mandible shows a modular structure consisting of
the alveolar region and the ascending ramus. The growth
pattern obtained provides clues to the dynamics and the
biological significance of the histological changes that occur
after birth in the house mouse mandible. Patterns of bone
growth in the house mouse mandible were compared with
available data obtained from the growth models reported
for other species, such as the rabbit Oryctolagus cuniculus
(Bang & Enlow, 1967), the American mink Mustela vison
(Buffrenil & Pascal, 1984), the rhesus monkey Macaca mulat-
ta, and modern human Homo sapiens (e.g. Kurihara et al.
1980; Enlow & Hans, 1996). Of these studies, only Buffrenil
& Pascal (1984) and Kurihara et al. (1980) documented the
histological changes during the postnatal growth in
Mustela vison and Homo sapiens mandibles, respectively.
In the first two postnatal weeks, the M. musculus mandi-
ble (both the alveolar region and the ascending ramus)
grows in all directions, increasing its size, as suggested by
the presence of woven bone tissue and confirmed by mea-
sured growth rates. Furthermore, bone growth rate data
from labial and lingual sides of the diastema and molar
regions, and the resorption activity on the endosteal surface
of their ventral area indicate a main lateral and dorsoven-
tral growth while the mandible increases its size. In the
ascending ramus, the highest growth rates registered reveal
a dorsoventral and lateral growth. A similar dynamics has
A
B
Fig. 6 Bone tissue types observed in the Mus musculus mandible. (A)
Woven bone tissue (first postnatal week, buccal side of the first molar
section), and (B) parallel-fibered bone tissue (eighth postnatal week,
buccal side of the first molar section). Scale bar: 50 lm.
ªª 2012 The AuthorsJournal of Anatomy ªª 2012 Anatomical Society
Postnatal growth of the mouse mandible, C. Martinez-Maza et al. 479
been also described in minks at the age of 2 months and in
humans in the first postnatal year, both characterized by
bone formation surfaces (Buffrenil & Pascal, 1984; Enlow &
Hans, 1996). The inferred bone growth dynamics in mice
are in agreement with Buffrenil & Pascal’s (1984) interpreta-
tion of the increase in height of the alveolar region of the
mink, mainly in a ventral direction. In addition, and consid-
ering the interpretations of the growth in length in other
mammals (Bang & Enlow, 1967; Buffrenil & Pascal, 1984; En-
low & Hans, 1996), we hypothesize that the increase in
length of the mouse mandible occurs by bone formation in
the posterior region of the alveolar region, while it grows
mainly laterally and the ramus is relocated in a posterior
position. Nevertheless, Buffrenil & Pascal (1984) suggested
that the increase in length of the mink alveolar region was
associated with bone growth in both posterior and anterior
regions. This anterior and posterior growth is similar to that
observed in M. mulatta and H. sapiens (Enlow & Hans,
1996) but differ from the growth in length of the mouse
and rabbit mandibles, which show only posterior bone
growth (Bang & Enlow, 1967). Differences among these spe-
cies are related to the position of the incisors with respect
to the alveolar region. Mink and primate mandibles show a
vertical position of the incisors, whereas in the rabbit and
mouse mandible, the incisors show a horizontal position,
preventing bone growth in the anterior region of the man-
dible (Buffrenil & Pascal, 1984). Therefore, the posterior
growth of the alveolar region proposed in this study for the
mouse mandible may be associated with the horizontal
position of the incisor.
In the third postnatal week, the mouse mandible shows
growth dynamics similar to those in previous weeks,
although differences in the histological organization and in
bone growth rates were observed between the alveolar
region and the ascending ramus. In the alveolar module,
parallel-fibered bone tissue is related to low osteogenesis,
whereas bone resorption fields identified in the diastema
and the first molar regions suggest an increase in height of
the alveolar region in a mainly ventral direction. The
ascending ramus is characterized by woven bone tissue and
depository surfaces, indicating a general growth in all direc-
tions. Interestingly, the second molar region shows similari-
ties with the ascending ramus (both characterized by
woven bone). This result supports the statement of Klingen-
berg et al. (2003) arguing that ‘modularity in the mandible
appears to be a question of degrees’. Differential labial ⁄lingual growth rates indicate a lateral growth of the alveo-
lar region and ramus. Growth dynamics inferred for the
alveolar region in M. musculus are in agreement with the
increase in height of the alveolar region and the lateral
(labial) drift of the mandible suggested in other species pre-
viously analyzed (Bang & Enlow, 1967; Buffrenil & Pascal,
1984; Kurihara et al. 1980; Enlow & Hans, 1996).
In the fourth postnatal week, the growth dynamics of
the mouse mandible shows a noticeable change compared
with previous weeks. The particular bone modeling field
distribution observed in the fourth week in the alveolar
region and in the ascending ramus was also observed in the
next postnatal weeks, suggesting similar growth directions.
From the fourth to eighth postnatal weeks, the bone mod-
eling data indicate a lateral drift of the alveolar region,
whereas the ascending ramus has a vertical arrangement
due to medial growth of the coronoid region and lateral
growth of the ventral part of the ramus. Although these
growth dynamics are similar from the fourth to the eighth
postnatal weeks, bone tissue type distribution and bone
growth rates show slight differences that suggest similar
growth dynamics but different intensity of bone growth.
On the one hand, the alveolar region module shows a slight
growth in the ventral half of the labial side and in the
ventral area which is simultaneous with the lateral drift.
Such dynamics indicate a main dorsoventral direction of
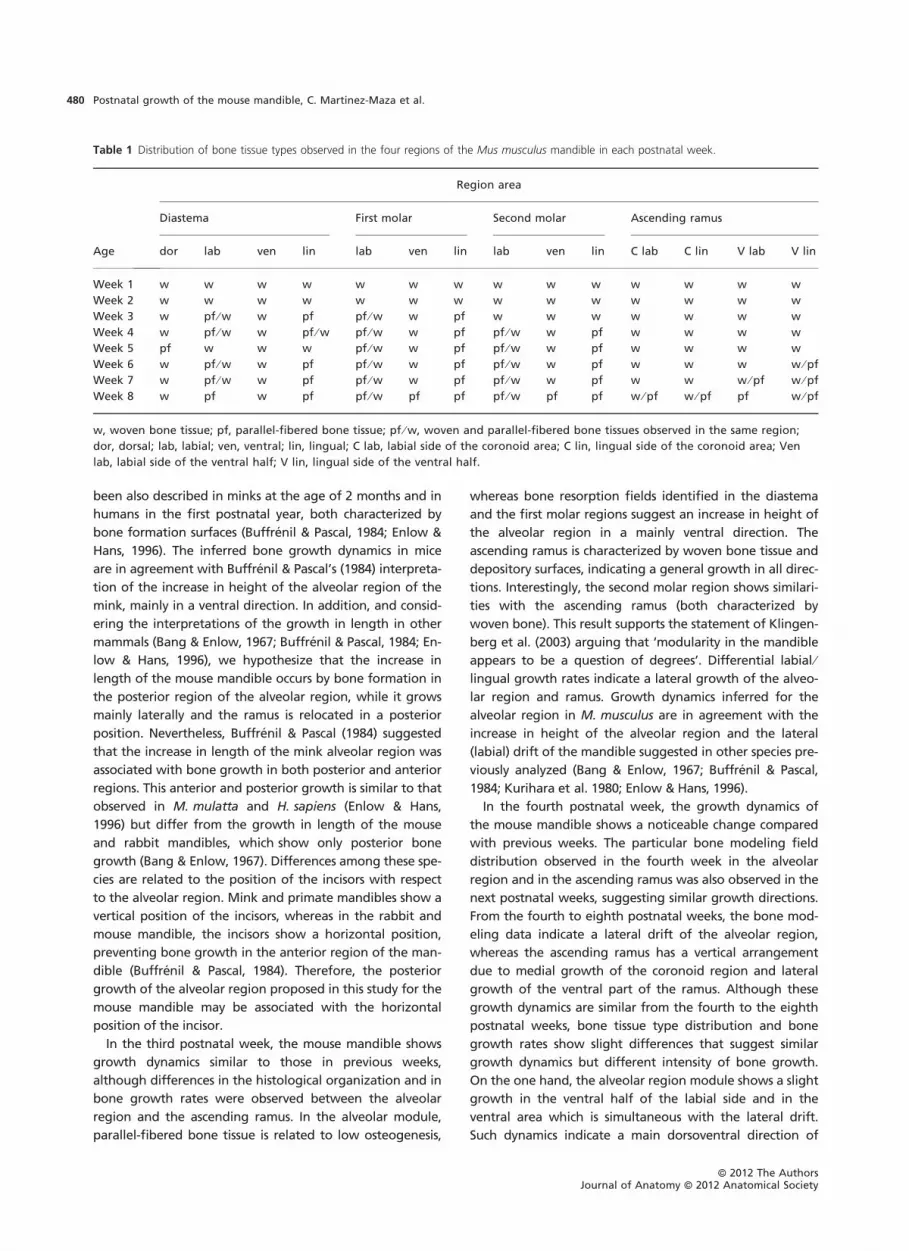
Table 1 Distribution of bone tissue types observed in the four regions of the Mus musculus mandible in each postnatal week.
Region area
Age
Diastema First molar Second molar Ascending ramus
dor lab ven lin lab ven lin lab ven lin C lab C lin V lab V lin
Week 1 w w w w w w w w w w w w w w
Week 2 w w w w w w w w w w w w w w
Week 3 w pf ⁄ w w pf pf ⁄ w w pf w w w w w w w
Week 4 w pf ⁄ w w pf ⁄ w pf ⁄ w w pf pf ⁄ w w pf w w w w
Week 5 pf w w w pf ⁄ w w pf pf ⁄ w w pf w w w w
Week 6 w pf ⁄ w w pf pf ⁄ w w pf pf ⁄ w w pf w w w w ⁄ pf
Week 7 w pf ⁄ w w pf pf ⁄ w w pf pf ⁄ w w pf w w w ⁄ pf w ⁄ pf
Week 8 w pf w pf pf ⁄ w pf pf pf ⁄ w pf pf w ⁄ pf w ⁄ pf pf w ⁄ pf
w, woven bone tissue; pf, parallel-fibered bone tissue; pf ⁄ w, woven and parallel-fibered bone tissues observed in the same region;
dor, dorsal; lab, labial; ven, ventral; lin, lingual; C lab, labial side of the coronoid area; C lin, lingual side of the coronoid area; Ven
lab, labial side of the ventral half; V lin, lingual side of the ventral half.
ªª 2012 The AuthorsJournal of Anatomy ªª 2012 Anatomical Society
Postnatal growth of the mouse mandible, C. Martinez-Maza et al.480

growth increasing the height of the alveolar region and a
lateral growth increasing the width of the ventral part of
the alveolar region. This is in agreement with the results
obtained for minks by Buffrenil & Pascal (1984) that sug-
gested (from the resorption of the alveolar area) an
increase in height of the mandible, particularly in the
molar region. The growth rate and the gradual variation
from woven bone (high osteogenesis) to fibrolamellar
bone tissue (low osteogenesis) indicate lateral and dorso-
ventral growths that gradually decrease from the fourth
to the sixth postnatal week. The growth model obtained
in this work for the mouse alveolar region is highly similar
to that reported for the rabbit (Bang & Enlow, 1967).
These growth dynamics suggest an increase in size of the
alveolar region in proportion to the overall size of the
entire growing mandible, whereas the lingual resorption
of the diastema region may indicate a downward develop-
ment of the genial tuberosity area (Bang & Enlow, 1967).
These similarities may be related to the presence of the
diastema in both species.
On the other hand, the ascending ramus shows specific
growth dynamics in the mouse mandible different from
those observed in the alveolar region. These bone growth
dynamics indicate a vertical arrangement of this region that
may be related to the proximity of the condyles to maintain
the contact with the cranial base through the temporoman-
dibular joint during development. The particular growth of
the ascending ramus is influenced by the large complex of
muscles that insert in this mandibular region (Atchley &
Hall, 1991). As reported in other rodent species, diet
changes after weaning are involved in the shape modeling
of the mandible during growth (Cardini & Tongiorgi, 2003;
Ventura & Casado-Cruz, 2011). In M. musculus the weaning
occurs around 21 days after birth and at the age of the
fourth week there is an increase of activities related to
drinking and feeding (Marques & Olson, 2007). The mandi-
ble responds to physiological loads resulting from the new
dietary requirements during the postweaning period
through the modeling mechanism. As a consequence, dif-
ferences in the bone mineral density have been reported
also in the mouse mandible (Ravosa et al. 2007). Therefore,
we hypothesize that changes here reported in the growth
dynamics in the fourth week are related to the diet change.
From the seventh to eighth postnatal weeks, the mouse
mandible shows similar growth dynamics to that in previous
weeks but bone growth rates decrease notably, and both
the alveolar region and the ascending ramus show bone tis-
sue types associated with low osteogenesis.
Considering the mandible as a whole, the growth pattern
of this structure in the house mouse shows a characteristic
growth based on the V principle established by Enlow
(1963), which has also been reported in the rabbit (Bang &
Enlow, 1967) and the macaque (Enlow & Hans, 1996).
During the postnatal development, the alveolar region is
relocated laterally and increases its length posteriorly, while
the ascending ramus is placed in a posterior and medial
position. Furthermore, histological data have shown a
change from immature bone tissue type (woven bone with
high vascularity) to mature bone tissue (fibrolamellar bone
tissue with flat osteocytic lacunae without vascularization
B
A
Fig. 7 Schematic maps show the main postnatal changes in structural organization of bone tissues (A) and in the distribution of bone modeling
fields (B) in the Mus musculus mandible. (A) A series of three mandibles from the first (left) to the eight postnatal weeks (right). On the left, the
mandible is characterized by woven bone (light gray) in the alveolar region and the ascending ramus (the first and second postnatal weeks). In the
middle, the mandible displays parallel-fibered bone tissue (dark gray) in the alveolar region and woven bone in the ascending ramus (from the
third to the sixth postnatal week). On the right, the mandible shows predominantly parallel-fibered bone tissue (the seventh and eighth postnatal
week). The broken line in the second mandible delimited the natural modules established in previous works (Atchley & Hall, 1991; Klingenberg
et al., 2003). (B) Two generalized bone modeling patterns observed throughout ontogeny. On the left, the mandible is characterized by bone
depository surfaces (light gray; from the first to the third postnatal week). On the right, mandible displays the distribution of the bone depository
and bone resorption (dark gray) fields (from the fourth to eighth postnatal week).
ªª 2012 The AuthorsJournal of Anatomy ªª 2012 Anatomical Society
Postnatal growth of the mouse mandible, C. Martinez-Maza et al. 481

or with primary osteons). This histological change occurred
anteroposteriorly, that is, changes were first observed in
the diastema and later in the molar and the ascending
ramus. Our findings provide new evidence supporting the
hypothesis that the mouse mandible is divided into two
developmental modules (alveolar region and the ascending
ramus), which agrees with results obtained in previous work
(Atchley et al. 1985; Leamy, 1993; Cheverud et al. 1997;
Mezey et al. 2000; Klingenberg et al. 2003). Results
obtained here reveal differences in the timing of bone tis-
sue development and the growth dynamics between the
alveolar region and the ascending ramus modules during
the postnatal development. Histological analyses allowed
us to establish the bone growth mechanism of the mouse
mandible, providing useful information to understand the
normal histomorphogenesis of the mandible. This growth
model can be useful for future studies focused on the deter-
mination of the growth changes associated with pheno-
typic variability due to genetic or epigenetic factors.
Acknowledgements
We thank two anonymous referees for their valuable comments
and their constructive suggestions that have contributed to
improve this article. We also thank Michel Laurin (Museum
National d’Histoire Naturelle, Paris, France) for checking the
English of this manuscript. Support for this study was provided
by a grant from the Spanish government, Ministerio de Ciencia
e Innovacion (CGL2010-15243). C.M.M. is funded by the
‘National Programme of Mobility and Humans Resources from
the I-D+I 2008-2011 National Plan’ of the Spanish Ministry of Sci-
ence and Innovation (MICINN). We thank the ‘Plateforme Ani-
malerie Rongeurs’ de l’IFR 83 de l’Universite Pierre et Marie
Curie.
References
Amprino R (1947) La structure du tissu osseux envisagee comme
expression de differences dans la vitesse de l’accroissement.
Arch Biol 58, 315–330.
Atchley WR (1993) Genetic and developmental aspects of
variability in the mammalian mandible. In The Skull, Vol. 1.
(eds Hanken J, Hall BK), pp. 207–247. Chicago: The University
of Chicago Press.
Atchley WR, Hall B (1991) A model for development and
evolution of complex morphological structures. Biol Rev 66,
101–157.
Atchley WR, Plummer AA, Riska B (1985) Genetics of mandible
form in the mouse. Genetics 111, 555–577.
Bang S, Enlow DH (1967) Postnatal growth of the rabbit
mandible. Arch Oral Biol 12, 993–998.
Bloom W, Fawcett DW (1994) A Textbook of Histology, 12th
edn. New York: Chapman & Hall.
Boell L, Gregorova S, Forejt J, et al. (2011) A comparative
assessment of mandible shape in a consomic strain panel of
the house mouse (Mus musculus) – implications for epistasis
and evolvability of quantitative traits. BMC Evol Biol 11, 309.
Buffrenil V, Pascal M (1984) Croissance et morphogenese
postnatales de la mandibule du vison (Mustela vison
Schreiber): donnees sur la dynamique et l’interpretation
fonctionnelle des depots osseux mandibulaires. Can J Zool 62,
2026–2037.
Cardini A, Tongiorgi P (2003) Yellow-bellied marmots ‘in the
shape space’: sexual dimorphism, growth and allometry of the
mandible. Zoomorphology 122, 11–23.
Cheverud JM, Routman EJ, Irschick DJ (1997) Pleiotropic effects
of individual gene loci on mandibular morphology. Evolution
51, 2006–2016.
Enlow DH (1963) Principles of Bone Remodeling. Springfield:
Charles C Thomas.
Enlow DH, Hans MG (1996) Essentials of Facial Growth.
Philadelphia: WB Saunders Co.
Enomoto A, Watahiki J, Yamaguchi T, et al. (2010) Effects of
mastication on mandibular growth evaluated by
microcomputed tomography. Eur J Orthod 32, 66–70.
Hall BK (2003) Unlocking the black box between genotype and
phenotype. Cell condensations as morphogenetic (modular
units). Biol Philos 18, 219–247.
Kesner MH (1980) Functional morphology of the masticatory
musculature of the Rodents subfamily microtine. J Morphol
165, 205–222.
Klingenberg CP (2002) Morphometrics and the role of the
phenotype in studies of the evolution of developmental
mechanisms. Gene 287, 3–10.
Klingenberg CP (2009) Morphometric integration and
modularity in configurations of landmarks: tools for
evaluating a priori hypotheses. Evol Dev 11, 405–421.
Klingenberg CP, Leamy LJ (2001) Quantitative genetics of
geometric shape in the mouse mandible. Evolution 55, 2342–
2352.
Klingenberg CP, Mebus K, Auffray JC (2003) Developmental
integration in a complex morphological structure: how
distinct are the modules in the mouse mandible? Evol Dev 5,
522–531.
Klingenberg CP, Leamy LJ, Cheverud JM (2004) Integration
and modularity of quantitative trait locus effects on
geometric shape in the mouse mandible. Genetics 166,
1909–1921.
Kurihara S, Enlow DH, Rangel RD (1980) Remodeling reversals in
anterior parts of the human mandible and maxilla. Angle
Orthod 50, 98–106.
Leamy L (1993) Morphological integration of fluctuating
asymmetry in the mouse mandible. Genetics 89, 139–153.
Marques JM, Olson IAS (2007) The effect of preweaning and
postweaning housing on the behaviour of the laboratory
mouse (Mus musculus). Lab Anim 41, 92–102.
Mezey JG, Cheverud JM, Wagner GP (2000) Is the genotype-
phenotype map modular? A statistical approach using mouse
quantitative trait loci data. Genetics 156, 305–311.
Monteiro LR, Bonato V, dos Reis SF (2005) Evolutionary
integration and morphological diversification in complex
morphological structures: mandible shape divergence in spiny
rats (Rodentia, Echimyidae). Evol Dev 7, 429–439.
Moss ML (1960) A functional approach to craniology. Am J Phys
Anthropol 18, 281–292.
Ramaesh T, Bard JBL (2003) The growth and morphogenesis of
the early mouse mandible: a quantitative analysis. J Anat 203,
213–222.
Ravosa MJ, Klopp EB, Pinchoff J, et al. (2007) Plasticity of
mandibular biomineralization in myostatin-deficient mice. J
Morphol 268, 275–282.
ªª 2012 The AuthorsJournal of Anatomy ªª 2012 Anatomical Society
Postnatal growth of the mouse mandible, C. Martinez-Maza et al.482

Renaud S, Auffray J-C, de la Porte S (2010) Epigenetic effects on
the mouse mandible: common features and discrepancies in
remodeling due to muscular dystrophy and response to food
consistency. BMC Evol Biol 10, 28.
Satoh K (1997) Comparative functional morphology of
mandibular forward movement during mastication of two
murid rodents, Apodemus speciosus (Murinae) and
Clethrionomys rufocanus (Arvicolinae). J Morphol 231, 131–142.
Ventura J, Casado-Cruz M (2011) Post-weaning ontogeny of the
mandible in fossorial water voles: ecological and evolutionary
implications. Acta Zool 92, 12–20.
Zelditch ML, Wood AR, Bonett RM, et al. (2008) Modularity of
the rodent mandible: integrating bones, muscles, and teeth.
Evol Dev 10, 756–768.
ªª 2012 The AuthorsJournal of Anatomy ªª 2012 Anatomical Society
Postnatal growth of the mouse mandible, C. Martinez-Maza et al. 483