polyploidy, hybridization and reticulate evolution - uenf
TRANSCRIPT

Polyploidy, hybridization and reticulate evolution:
lessons from the Brassicaceae
K. Marhold1,2 and J. Lihova
1
1Institute of Botany, Slovak Academy of Sciences, Bratislava, Slovak Republic2Department of Botany, Charles University, Praha, Czech Republic
Received November 26, 2005; accepted December 15, 2005Published online: June 19, 2006� Springer-Verlag 2006
Abstract. The family Brassicaceae is well knownfor its large variation in chromosome numbers,common occurrence of polyploids and manyreports of interspecific gene flow. The presentreview summarizes studies from the past decadeson polyploidization and hybridization events,recognizing them as important evolutionaryforces in the family. Attention is drawn to theissue of the reconstruction of reticulated patternof evolution resulting from allopolyploid andhomoploid hybrid speciation. The research ofvarious authors on several Brassicaceae genera ispresented and discussed in the context of ourcurrent understanding of polyploid and hybridevolution. Model species, Arabidopsis thalianaand Brassica taxa, are referred to only margin-ally, major focus is on a comprehensive survey ofstudies on about a dozen best explored non-model genera (e.g. Cardamine, Draba, Rorippa,Thlaspi). The increasing amount of genetic andgenomic resources available for Brassicaceaemodel species provides excellent opportunitiesfor comparative genetic and genomic studies.Future research directions and challenges arethus outlined, in order to obtain more detailedinsights into the evolution of polyploid andhybrid genomes.
Key words: Allopolyploidy, autopolyploidy,Cruciferae, chromosomes, hybrids, phylogeny.
Introduction
Polyploidization is undoubtedly a frequentmode of diversification and speciation inplants. Otto and Whitton (2000) suggestedthat polyploidization may be the most com-mon mechanism of sympatric speciation inplants. Moreover, recent data indicate thatmost plants have undergone one or moreepisodes of polyploidization during their evo-lutionary history (Soltis et al. 2004b). Formany polyploids, multiple origins in spaceand time have been proven, along withincreased genetic diversity and complexity,whereas in other cases a single origin isproposed (Soltis and Soltis 1999, Soltis et al.2004b). Hybrid speciation is another impor-tant evolutionary phenomenon. It occurs intwo possible ways, as allopolyploid or homop-loid hybrid speciation (Hegarty and Hiscock2005), thus hybridization and polyploidizationevents are usually tightly coupled. Theseprocesses often result in a reticulated patternof evolution. While in polyploids reticulation isalways suspected, homoploid hybrid speciationevents may be more difficult to identify andunravel. Linder and Rieseberg (2004), andVriesendorp and Bakker (2005) have pointed
Pl. Syst. Evol. 259: 143–174 (2006)DOI 10.1007/s00606-006-0417-x

to the fact that the evolutionary history ofmany plant groups does not follow divergentevolutionary patterns, and hardly can beunravelled in a tree-building procedure.Rather, it is like a network, which displays anumber of reticulate evolutionary events. Thefamily Brassicaceae is not an exception, andpolyploidy has certainly played one of the keyroles in its evolution. This is clearly illustratedby the recently discovered genome triplication,recognized as a major evolutionary event thatgave rise to the whole tribe Brassiceae. It hasbeen assumed that the ancestral Brassiceaegenome became triplicated via allohexaploidythat occurred 7.9–14.6 Mya (Lysak et al.2005).
The large variation in base chromosomenumbers (x = 4–13 ()17); Warwick and Al-Shehbaz 2006) in the family indicates complexkaryotype evolution. Species with higher basenumbers are suspected to represent palaeopo-lyploids that have undergone substantial gen-ome diploidization (e.g. Brassica; Osborn2004). Chromosome counts are currently avail-able for 232 out of 338 accepted Brassicaceaegenera (68.6%), and for 1558 out of the 3709(42.0%) recognized species (Warwick and Al-Shehbaz 2006). Approximately 37% of thespecies are assumed to be polyploid (Warwickand Al-Shehbaz 2006). This estimate, however,can be much higher, if we consider that severaldiploid species (e.g. from the genera Brassica,Physaria) are in fact palaeopolyploids. Thepercentage of polyploids in the family can insuch cases reach at least 50% (Koch et al.2003a). Diploidization processes hamper theidentification of ancient polyploidizationevents, which, however, can be suspected fromlarge-scale genome duplications (see Lysaket al. 2005). Polyploidy is widespread in manygenera, and some of them (e.g. Crambe,Moricandia) even seem to be exclusively poly-ploid (Koch et al. 2003a). In the genus Card-amine we identified 32% diploid, 10% bothdiploid and polyploid, and 58% entirely poly-ploid taxa (Kucera et al. 2005). For someother Brassicaceae genera, the correspondingfrequencies are as follows (based on the
database by Warwick and Al-Shehbaz 2006):Draba ) 25% diploid, 7% both diploid andpolyploid, 68% entirely polyploid taxa; Lepi-dium ) 34% diploid, 14% both diploid andpolyploid, 52% entirely polyploid taxa;and Rorippa ) 48% diploid, 39% both diploidand polyploid, and 13% entirely polyploidtaxa. These estimates clearly point to theevolutionary importance of polyploidy innumerous Brassicaceae genera. Besides poly-ploidization, hybridization and hybrid specia-tion are common evolutionary phenomena inthe Brassicaceae. Numerous examples ofhybrids are reported for this family (e.g. forthe British Isles summarized by Stace 1975),and some genera, such as Rorippa (Bleeker andHurka 2001), Cardamine (Lihova and Mar-hold 2006) or Boechera (Koch et al. 2003b),are characterized by frequent hybridizationand introgression.
With the advent of new molecular tools,especially in the past decade, our understand-ing of the above-mentioned evolutionary phe-nomena has been significantly improved.Advances in Brassicaceae research in thisrespect were recently summarized in Kochet al. (2003a). The vast majority of studies onpolyploid origins and subsequent genetic andgenomic evolution, have focused on modelplant species. The immensely increasingamount of genetic and genomic resourcesavailable for the model species Arabidopsisthaliana provides an excellent opportunity totransfer this knowledge to non-model relatives,as well as to use the methodological progress incomparative studies with related genera. Com-parative genetic and genomic analyses involv-ing this model species can reveal much aboutthe evolutionary patterns within this family(Hall et al. 2002, Lysak et al. 2003).
The aim of this review is to focus onpolyploidization and hybridization as twoimportant evolutionary forces in the familyBrassicaceae, as well as to explore the resultingpatterns of reticulate evolution. Case studiesare presented from the genus Cardamine,which has been extensively studied by ourresearch group and other authors as well, and
144 K. Marhold and J. Lihova: Polyploidy, hybridization and reticulate evolution

from about a dozen other genera, which havereceived particular attention in the past years.A better understanding of the evolutionarypatterns in Brassicaceae can be achieved onlyby a comprehensive survey of published stud-ies, as conducted here. New challenges andopen questions are then identified, stimulatingfurther studies.
Chromosome number and genome size diversity
A continuous series of base chromosomenumbers ranging from x = 4 to 13 (17) hasbeen reported for the Brassicaceae, with arelatively high percentage of taxa (37%) basedon x = 8 (Warwick and Al-Shehbaz 2006).The lowest chromosome number known forthis family (n = 4) is found in two unrelatedgenera, the Australian Stenopetalum and thewestern North American Physaria (Al-Sheh-baz 1984). The highest chromosome numbersknown in the family were reported in NorthAmerican Cardamine diphylla and C. concate-nata (= Dentaria laciniata), both up to 2n =256 or n = 128, corresponding to 32-ploidylevel (Easterly 1963, Al-Shehbaz 1988). How-ever, in the former species there is considerableintrapopulational and even intraindividualvariation in the chromosome numbers re-ported, and thus, it is difficult to assess whetherthe high numbers represent regular stabilizedchromosome sets. Arabidopsis thaliana hasx = 5 chromosomes, while many of its closerelatives have x = 8. Several reductions inbase chromosome number from x = 8 tox = 5–7 have been recorded within the tribeArabideae, suggesting that numbers lower thanx = 8 are phylogenetically derived. This alsoholds true for the genome of A. thaliana, whichis considered to be highly derived (Koch et al.1999). Chromosome number variation in thefamily is indeed enormous (see Warwick andAl-Shehbaz 2006). Several genera exhibit verylarge variation, as illustrated by Aethionema,Draba, Erysimum, Cochlearia (Warwickand Al-Shehbaz 2006 and references therein)and Cardamine (Kucera et al. 2005, Lihovaand Marhold 2006). This variation apparently
reflects the occurrence of several cytogeneticphenomena, such as polyploidy, aneuploidyand dysploidy. Complex karyotype evolutioninvolving extensive chromosomal structuralrearrangements can also be expected withinthe family, as already illustrated by compara-tive genetic mapping of Arabidopsis, Capsellaand Brassica species (Lagercrantz 1998, Lanet al. 2000, Boivin et al. 2004, Kuittinen et al.2004, Koch and Kiefer 2005). This approach,together with cytogenetic techniques such as insitu hybridization or comparative chromosomepainting, can elucidate significant patterns ofchromosome and karyotype evolution in theBrassicaceae (Schmidt et al. 2001, Hall et al.2002, Lysak et al. 2003, Lysak and Lexer2006).
Along with chromosome number determi-nation and karyotype studies, techniques thatmeasure DNA amount per nucleus (flowcytometry or microdensitometry) provide use-ful tools to study cytogenetic diversity (Dolezeland Bartos 2005, Bennett and Leitch 2005b).Genome size varies enormously among angio-sperms, and several plant families, such as theFabaceae, Poaceae or Liliaceae, exhibit largeranges in DNA content (Bennett and Leitch2005a). Despite the extensive structural gen-ome evolution observed in the Brassicaceae,the species analyzed so far have shown smallnuclear DNA content levels, with a narrowerrange in comparison to other families (up to1C = 1.95 pg, Johnston et al. 2005). Super-imposing genome size onto a robust phylogenyallows an assessment of ancestral DNA con-tent, as well as the ability to trace genome sizeevolution. Such studies have supported thetheory of bi-directional dynamic changes ingenome size, i.e. that both genome size expan-sion and contraction occur commonly (seeWendel et al. 2002). A first attempt to tracegenome size evolution in Brassicaceae has beenrecently published by Johnston et al. (2005).Monoploid genome sizes were mapped onto asingle-most parsimonious tree based on theinternal transcribed spacer of nuclear ribo-somal DNA (nrDNA ITS) sequence data from34 taxa, including both diploid and polyploid
K. Marhold and J. Lihova: Polyploidy, hybridization and reticulate evolution 145

taxa. Although the number of species analyzedwas not high, some general trends can be seen.The DNA content ranged over 8-fold (1C =0.16–1.31 pg) and 4.4 fold (1C = 0.16–0.71 pg) when allotetraploid Brassica specieswere excluded. The ancestral genome size wasestimated at ca. 1C = 0.2 pg, and thepresented data unequivocally supported theconcept of a dynamic nature of genome sizeevolution in this family, involving bothincreases and decreases. The smallest genomewas identified in Arabidopsis thaliana (1C =0.16 pg) that apparently underwent an evolu-tionary decrease. A decrease in genome sizewas also revealed in extant allopolyploids ofBrassica species and in Arabidopsis suecica(that originated from A. thaliana and A. aren-osa, for details see below). In all these cases,the genome size of allopolyploids was reportedto be less than the sum of their diploidancestors (Johnston et al. 2005).
In polyploid series that are of recent origin,increase in the DNA content in polyploids isexpected to be in direct proportion to theploidy level (Bennett et al. 2000). Indeed, sucha pattern was found in several Draba species,which included diploids, triploids, tetraploidsand hexaploids, whose recent origin was sup-ported by molecular markers (Grundt et al.2005). In some polyploids (as documented forArabidopsis suecica, Johnston et al. 2005; andA. lyrata subsp. petraea, Dart et al. 2004),however, the additivity has not been retained,but genome size contraction has apparentlyoccurred. Genome expansion following poly-ploidization has been observed as well,although it appears less common than genomecontraction (Leitch and Bennett 2004). In ourrecent study focused on the allopolyploidCardamine asarifolia (Lihova et al. 2006), wesuggested that this hexaploid had experiencedgenome expansion subsequent to its origin.This particular case, however, has been com-plicated by the assumed extinction of one ofthe parental species, and consequently, com-parisons were limited to other most closelyrelated species. Increase in genome size hasusually been associated with the accumulation
of retroelements which might be activated byallopolyploidization, while various mecha-nisms for the deletion of redundant genomicsequences leading to genome downsizing havealso been suggested (reviewed in Wendel et al.2002, Kellogg and Bennetzen 2004, Leitch andBennett 2004).
Other aspects of genome size variation inthe Brassicaceae than presented here, arediscussed by Lysak and Lexer (2006).
Spontaneous interspecific hybridizations
and their evolutionary consequences:
molecular evidence
The many examples of hybridization, intro-gression and hybrid speciation reported for theBrassicaceae indicate that this is a significantevolutionary force in the family. The compre-hensive work on hybridization of the taxaoccurring in the British Isles (Stace 1975)provided data on as many as 39 interspecifichybrids within this family. Yet, there are not somany recent studies, in which the hybrid originand the parentage of putative hybrid plantshave been proven using molecular or otherreliable evidence. Nevertheless, some generahave been subjected to thorough studiesexploring interspecific gene flow, and it isapparent that hybridization has significantlycontributed to their evolution and speciesdiversity. They certainly belong to the best-studied hybrid systems and provide interestingexamples of interspecific hybridization andhybrid speciation (e.g. hybrids in Rorippa,Bleeker and Hurka 2001, Bleeker 2003; Boe-chera, Schranz et al. 2005; Cardamine ·insuetaand C. schulzii, Urbanska et al. 1997). Inter-specific hybridization has often been recog-nized as a source of genetic variation andgenetic novelties, and in some cases successfulhybridization events have promoted rapidradiation (Seehausen 2004). This seems to bethe case for Australian/New Zealand Lepidiumspecies originating in the Pliocene/Pleistocene,for which hybridogenous genomic constitutionhas been recently revealed (Mummenhoff et al.2004). Ancient trans-oceanic dispersals from
146 K. Marhold and J. Lihova: Polyploidy, hybridization and reticulate evolution

California and South Africa, respectively,followed by hybridization have been assumedto occur. Low levels of sequence divergenceobserved among the species from Australiaand New Zealand suggested a rapid and recentradiation following the hybridization event, ascan be seen from the current species diversityin this area (Mummenhoff et al. 2004).
Hybrid establishment and persistence im-ply that the hybrid has become isolated fromboth of its parental taxa. This can be achievedeither by chromosomal rearrangements andsubsequent postzygotic genetic isolation,which have been observed in newly formedhybrids. Alternatively, ecological divergence,e.g. spread into new ecological niches notoccupied by the parents results in spatialisolation and gene flow restriction. Otherecological barriers, such as temporal or polli-nator divergence, can contribute to reproduc-tive isolation as well (Arnold 1997, Gross andRieseberg 2005, Hegarty and Hiscock 2005).The role of environmental disturbance (oftenman-induced) for creating new habitats avail-able for hybrids is widely recognized, andapparently was crucial also for the origin ofseveral Brassicaceae hybrids, such as Card-amine ·insueta (Urbanska 1987) or Rorippaaustriaca · R. sylvestris hybrids (R. ·armora-cioides, Bleeker 2003).
Molecular markers: detection of hybridiza-
tion events in the Brassicaceae. A variety ofmolecular techniques has been used to detectancient or more recent hybridization eventsand to identify parental taxa. Isozyme analysesand isoelectric focusing analysis of Rubisco(ribulose-1,5-bisphosphate carboxylase/oxy-genase) subunits have proven useful in manystudies of the Brassicaceae genera, e.g. inRorippa (Bleeker and Hurka 2001), Nasturtium(Bleeker et al. 1999), Arabidopsis (Mummen-hoff and Hurka 1995), Diplotaxis (Mummen-hoff et al. 1993, Eschmann-Grupe et al. 2003)and Cardamine (Urbanska et al. 1997), wherespecies-specific alleles identified in the parentshave been found in assumed hybrids. ThenrDNA ITS regions is among the most widelyused nuclear-encoded markers (Alvarez and
Wendel 2003). Its high evolutionary ratepermits discrimination of closely related puta-tive parents and identification of additivepatterns in hypothesized hybrids. Althoughconcerted evolution can erase nucleotide addi-tivity, bi-directional homogenization or inter-genomic recombination resulting in a mosaicstructure of ITS sequences would still retaintraces of past hybridization. This marker hassignificantly contributed to the identificationof hybrids or hybridogenous origins in Card-amine (Franzke and Mummenhoff 1999;Lihova et al. 2006), Boechera (Koch et al.2003b), Lepidium (Mummenhoff et al. 2004),Thlaspi (Mummenhoff et al. 1997), and Draba(Widmer and Baltisberger 1999). Fingerprint-ing methods, such as RAPD, AFLP or ISSR,have been frequently used due to their highresolution. Intermediate genotypes and theadditivity of individual fragments or allelesprovided convincing evidence for hybrid ori-gins (e.g. Rorippa austriaca · R. sylvestris,Bleeker and Matthies 2005; Cardamine ·insu-eta, Urbanska et al. 1997; Diplotaxis muralis,Warwick and Anderson 1997a, Martın andSanchez-Yelamo 2000; Erucastrum gallicum,Warwick and Anderson 1997b; Boechera ·di-varicarpa, Dobes et al. 2004a). Studies ofcpDNA haplotype diversity can identify thematernal parent, as well as demonstratebi-directional and multiple hybrid origins.Recently, we proved both bi-directional andrecurrent polytopic hybridizations between thehexaploid Cardamine asarifolia and diploidC. amara. The hybrids (C. ·ferrarii) displayedmultiple cpDNA haplotypes present in eitherof the parental species from the same or closelocality (Lihova et al. 2006). Similarly, bi-directional hybridization has been assumedfrom the presence of both parental species-specific trnL or trnL-trnF length variants inRorippa amphibia · R. sylvestris (R. ·anceps,Bleeker and Hurka 2001) and R. austriaca ·R. sylvestris (R. ·armoracioides, Bleeker andMatthies 2005).
As mentioned above, isozymes have thepotential to confirm the parentage of putativehybrids, provided that enough variation and
K. Marhold and J. Lihova: Polyploidy, hybridization and reticulate evolution 147

especially species-specific alleles can be de-tected. This is, however, not always the case.Bleeker et al. (1999) aimed to confirm theparentage of hexaploid Nasturtium ·sterile(tetraploid N. officinale · octoploid N. micro-phyllum), first suggested by Manton (1935).However, while N. microphyllum displayedspecies-specific fixed alleles, no such alleleswere found in N. officinale. Therefore,although N. ·sterile possessed the same band-ing patterns as N. microphyllum, providingevidence that this species is one of the parents,it was not possible to confirm N. officinale asthe second parent using available isozymedata. So far, no other marker systems havebeen applied that could provide more resolu-tion. There is, however, circumstantial evi-dence originating from classical data thatfavours this parentage (Howard and Manton1946, Bleeker et al. 1999): (1) intermediateploidy level of the hybrid, (2) morphologicalintermediacy, (3) experimental crosses result-ing in progeny morphologically stronglyresembling N. ·sterile, and (4) the occurrenceof N. ·sterile in areas where the putativeparents come into contact.
In the following survey we focus on a fewBrassicaceae studies, where often multiplemolecular markers have been used to provethe assumed hybridization events, and discussresulting patterns. As demonstrated by thesestudies, the use of additional data from othersources (e.g. chromosome numbers, morphol-ogy, artificially-produced hybrids) often helpsupport the evidence of hybridization.
Hybridization and introgression in Rorippa:hybrid fitness and intrapopulational variation.
Hybridization among European species ofRorippa has been frequently reported byseveral authors (e.g. Jonsell 1968, Tomsovic1969). In recent years, more detailed studiesemploying molecular data have been publishedby Bleeker and co-authors (Bleeker 2003, 2004;Bleeker and Hurka 2001; Bleeker and Matthies2005). Four closely related species belonging tosect. Rorippa, R. amphibia (2n = 16, 32), R.palustris (2n = 32), R. sylvestris (2n = 32, 48)and R. austriaca (2n = 16) have been
frequently involved in hybrid formation andintrogressive hybridization. An interestingmodel system to explore the dynamics ofinterspecific gene flow is represented by threehybrid zones observed between invasive R.austriaca and native R. sylvestris in Germany(Bleeker 2003, Bleeker and Matthies 2005). Inall three contact zones, hybridization wasinferred from the additivity of diagnosticparental AFLP markers of the local popula-tions, and from the overall genetic variationpatterns revealed by principal coordinate anal-ysis (intermediate position of hybrids). Inaddition, three individuals, morphologicallycorresponding to R. sylvestris, possessedAFLP profiles attributable to hybrids, provid-ing evidence of introgression. Different cyto-types of R. sylvestris (tetraploid and hexaploid)were found in the respective hybrid zones,resulting also in different ploidy levels in thehybrid progeny (triploid and pentaploid).While the origin of the triploid hybrid can beeasily explained by the fusion of reducedgametes, the prevalence of R. sylvestris mark-ers and the lower level of additivity in thepentaploids suggested that they may haveoriginated via first generation tetraploid plantsand further backcrosses with R. sylvestris(Bleeker and Matthies 2005). Hybrid fitnessdiffered between the triploids and pentaploids,the latter showing better seed set and germi-nation rate. Later generation hybrids areexpected to undergo genetic recombination,which, together with backcrossing and selec-tion, can result in a highly fertile and successfulhybrid species (Song et al. 1995, Rieseberget al. 2000). The higher fitness found in thepentaploid Rorippa hybrids can, thus, beexplained by these processes. The pentaploidchromosome number may cause meiotic irreg-ularities, however, viable gametes can still beproduced through polarized segregation ofchromosomes, as observed in Cardamine ·in-sueta (Urbanska et al. 1997). Another inter-esting issue that arose from these two Rorippastudies (Bleeker 2003, Bleeker andMatthies 2005) was the observed associationof intrapopulational variation and invasive
148 K. Marhold and J. Lihova: Polyploidy, hybridization and reticulate evolution

success. R. austriaca, an introduction toGermany, showed different levels of intrapop-ulational variation, with the highest oneobserved in areas where it behaves mostinvasively. This observation seems to be inaccordance with the assumption that accumu-lation of genetic variation either throughmultiple introductions or hybridization withcongeneric species may be a prerequisite forthe evolution of invasiveness (Lee 2002, Ell-strand and Schierenbeck 2000).
Another case, where interspecific gene flowhas shown a profound impact on intrapopula-tional and intraspecific variation, is reportedfor two self-incompatible species Rorippaamphibia and R. sylvestris (Bleeker and Hurka2001). Both species are native in the areastudied (Germany) and show different ecolog-ical preferences, but in periodically disturbedhabitats the isolation barriers can temporarilybreakdown. Hybridization and bi-directionalintrogression between these two species havebeen confirmed at the river Elbe in areascharacterized by natural dynamic changes(Bleeker and Hurka 2001, Bleeker 2004). Bothspecies exhibited increased intrapopulationalgenetic variation in that hybrid zone, whencompared with populations not influenced byhybridization. The same pattern was alsoobserved for seed set, being higher in thehybrid zone. Increased genetic variation dis-played at the level of isozyme variability, thus,might also affect variation at the S locus(through the exchange of alleles), which con-trols self-incompatibility. As a result, higherintrapopulational variation at the S locus maypositively affect successful fertilization andseed set (Bleeker 2004).
Hybridization and hybrid speciation in
Cardamine: the role of man-induced habitat
disturbances. A model of hybrid speciationassociated with man-induced habitat distur-bance is represented by triploid and hexaploidCardamine hybrids discovered in Switzerland.Hybridization between two diploids, C. prat-ensis (erroneously reported under the nameC. rivularis) and C. amara (subsp. amara) ledto successful establishment of triploid hybrids,
described as C. ·insueta (Urbanska-Wor-ytkiewicz and Landolt 1972). Autopolyploidi-zation of these hybrids resulted in the forma-tion of the highly fertile and viable hexaploidspecies C. schulzii (Urbanska-Worytkiewicz1977b). Since the discovery of this hybridcomplex, extensive biosystematic and molecu-lar investigations have been performed to gaininsights into its origin and ongoing microevo-lutionary processes. Earlier studies were fo-cused on cytogenetics and reproduction (chro-mosome numbers, karyotype, meiosis andchromosome segregation, pollen quality), aswell as population biology (size and spatialstructure of populations, population dynam-ics, flowering intensity), and the role of humaninterference (Urbanska-Worytkiewicz andLandolt 1972; Urbanska-Worytkiewicz1977a,b, 1980). It was concluded that thetriploid hybrid arose by fertilization of anunreduced C. pratensis gamete with a reducedC. amara gamete, and that it has only recentlyestablished and expanded into suitable man-made habitats. More recent molecular investi-gations included several markers, RFLP ofcpDNA, isozymes, RAPD fingerprinting, andITS sequences (Neuffer and Jahncke 1997,Urbanska et al. 1997, Franzke and Mummen-hoff 1999). cpDNA restriction site mutationsallowed the differentiation between the chlo-roplast genomes of the progenitor species C.pratensis and C. amara, and showed that allindividuals of C. ·insueta and C. schulziidisplayed patterns identical with C. pratensis,herewith providing the evidence for the mater-nal parent (Urbanska et al. 1997). Both RAPDmarkers and isozymes revealed additivity inthe triploid and hexaploid, supporting theirhybrid origin. The predominant vegetativereproduction observed in C. ·insueta, how-ever, somewhat contradicted its high level ofgenetic diversity, the latter being comparableto that in the outcrossing parental species.High genetic variation in the triploid can bemost likely attributed to backcrossing andrecurrent hybridizations between the parentalspecies. In accordance with its assumed recentorigin (dated not earlier than 1900), only a few
K. Marhold and J. Lihova: Polyploidy, hybridization and reticulate evolution 149

non-parental RAPD markers were detected inC. ·insueta, indicating that its genome has notevolved far from the parental ones (Neufferand Jahncke 1997). ITS sequences revealed arather pronounced sequence divergence be-tween the diploids C. pratensis and C. amaraand very rapid sequence homogenization inboth C. ·insueta and C. schulzii with a strongbias towards the maternal sequence type(Franzke and Mummenhoff 1999).
It seems that despite genetic differentiationbetween congeneric species, incompatibilitybarriers can occasionally break down, andplants with similar ecological requirements andsympatric occurrence hybridize. Similar to thecase of Cardamine ·insueta, we can report onanother example from this genus, where man-induced environmental disturbances openedthe possibility for the establishment of a newhybrid. Inferring from our preliminary mor-phological and molecular (AFLP markers)investigation, extensive hybridization, back-crossing and introgression have occurred onseveral sites between high polyploids of Card-amine pratensis s.str. and C. raphanifolia s.str.in the Cordillera Cantabrica Mts. (NW Spain;Lihova et al., unpubl.). Parental species to-gether with the scarce occurrence of intermin-gled hybrids or introgressed individuals havebeen observed to grow in relatively stable andless disturbed habitats along brooks on pas-tures and wet meadows, while dense popula-tions consisting exclusively of hybrids arespreading in ditches along the road.
Natural hybridization at the diploid levelwas documented between two eastern Pyre-nean endemics, Cardamine amara subsp. pyre-naea and C. crassifolia (Marhold et al. 2002b).Both species show very similar ecologicalpreferences and occupy almost the same dis-tribution area. Recent molecular phylogeneticanalyses showed that they belong to twodifferent lineages, and thus are geneticallydivergent (Lihova et al. 2004a). Nevertheless,two hybrid populations (named as C. ·enri-quei) were found. In both cases they formedsmall but rather dense populations that werepartially spatially separated from the parents,
and growing on somewhat man-influencedsites. The hybrid status was confirmed withmorphology, pollen sterility, and AFLP mark-ers. Similarly to C. ·insueta, the hybrid pop-ulations of C. ·enriquei exhibited considerablemorphological and genetic variation, suggest-ing recurrent origins and/or backcrossing withparents. Again, vegetative propagation hasmost likely made a major contribution to theestablishment of C. ·enriquei (Marhold et al.2002b).
Recurrent and polytopic hybridizations: the
case of Boechera ·divaricarpa. Hybridizationand polyploidization, together with apomixis,have played an important role in the evolutionof the North American genus Boechera. Thisgenus, originally classified within Arabis,includes about 62 species (Al-Shehbaz 2003)which presumably radiated before Pleistoceneglaciations. However, the frequent occurrenceof hybrids and allopolyploids indicates thatreproductive isolation of these taxa is incom-plete (Schranz et al. 2005 and references there-in). Most studies have focused on the wide-spread and largely sympatric species, Boecheraholboellii (= Arabis holboellii) and B. stricta(= Arabis drummondii). Recent studiesemploying several molecular markers haveindicated a complex evolutionary history with-in this group (Sharbel and Mitchell-Olds 2001;Koch et al. 2003b; Dobes et al. 2004a, b;Sharbel et al. 2005). As part of the thoroughstudies, assumed hybridization between thesetwo species, resulting in conspicuous patterns,has received much attention. The putativehybrid (B. ·divaricarpa) showed substantialpart of the genetic variation present in itsparental species. Almost all chloroplast paren-tal haplotypes were found in the hybridspecies. Their geographic distribution wascorrelated with the distribution of the respec-tive haplotypes found in the putative parents,and thus, suggested multiple hybridizations. Avery complex pattern was observed in ITSsequences, with the following variants identi-fied in different hybrid accessions: (1) a singlesequence variant typically found in either ofthe parents, (2) multiple ITS sequence variants
150 K. Marhold and J. Lihova: Polyploidy, hybridization and reticulate evolution

of which at least one was found in one of theparental species but not in the other, (3)multiple ITS sequence variants where bothparental ITS types were retained, and (4) arecombinant mosaic ITS sequence. Such apattern is consistent with different evolution-ary fates reported for divergent parental ITSsequences following their merger in the hybrids(see Alvarez and Wendel 2003). Nevertheless,the occurrence of such a broad spectrum in asingle hybrid species is intriguing. Definitely,hybridization must have been very commonand occurring multiple times and in multiplegeographical locations (as seen from thecpDNA data). The most recent studies, how-ever, have shown that the identification of theparentage of B. ·divaricarpamay be even morecomplicated, and B. holboellii might not be oneof its parents as currently believed. B. holboelliiitself is a highly polymorphic and polyphyleticspecies that may be of hybrid origin (Dobeset al. 2004a, b; Schranz et al. 2005). It has beenrecently suggested that B. stricta as the mostwidespread species of this genus hybridizeswith almost every sympatric diploid Boecheraspecies (Windham in Al-Shehbaz and Beilstein2006).
Gene flow between (transgenic) crop plants
and their wild relatives. An important aspect ofinterspecific hybridization that should also beaddressed here, is the possibility and the risk ofgene flow from crop plants to their wildrelatives. The family Brassicaceae includesmany economically important plants that arewidely cultivated. Spontaneous hybridizationbetween cultivated plants and their wild rela-tives can occasionally occur, and has beenindeed documented for several crops includingBrassica species (Ellstrand et al. 1999, Chevreet al. 2004). The probability of interspecifichybridization depends on various factors, suchas phylogenetic relatedness, mating systems,and density and spatial distribution of wildrelatives (Warwick et al. 2003). Darmencyet al. (1998) pointed to the fact that a self-incompatible wild species when growingisolated within or near a crop field can receivea considerable amount of pollen from the
cultivated plants, resulting in a rather highchance of cross-fertilization. Recently, therehas been a major concern about transgeniccrop plants that could transfer engineeredtraits to weedy relatives and become beneficialfor them. The escape of the gene(s) fromtransgenic plants into natural populations mayhave unpredictable evolutionary and ecologi-cal consequences (Warwick et al. 2003). Thismay be especially relevant for the oilseed rape(Brassica napus), a major crop commonlygrown in Europe, North America, Argentina,and China. This partially allogamous cropplant is of hybridogenous allopolyploid origin(originated from wild B. rapa and B. oleracea)and has numerous compatible wild relativeswhich often occur sympatrically with the crop.Several studies assessing the ability and fre-quency of interspecific gene flow and geneintrogression from B. napus to various wildrelatives have been performed both in exper-imental field trials and in commercial fields(reviewed by Chevre et al. 2004). While prob-abilities of gene flow from transgenic B. napusto several relatives (e.g. Raphanus raphani-strum, Sinapis arvensis, Erucastrum gallicum)have been estimated to be very low, a highpotential exists for hybridization with itsprogenitor B. rapa, as well as for the estab-lishment and spread of introgressed geneswithin its natural populations (Warwick et al.2003, Chevre et al. 2004). Weeds with noveltraits such as insect or disease resistance, orresistance to environmental stress, are likely todisplay increased overall fitness and competi-tiveness, and their spread may have seriousagronomic and environmental implications(Warwick et al. 2003).
Future tasks and challenges for hybridiza-
tion studies. Novel molecular and cytogeneticapproaches provide the opportunity not onlyto detect the origin and parentage of hybrids,but also to study hybrid speciation and theimpact of hybridization at the level of genomeorganization and gene expression (Hegartyand Hiscock 2005, Lysak and Lexer 2006).Comparative genetic and physical mapping,chromosome painting and other methods
K. Marhold and J. Lihova: Polyploidy, hybridization and reticulate evolution 151

allow examination of genome evolutionfollowing hybridization. There is increasingevidence that chromosomal rearrangementsmay take place rapidly in newly formedhybrids. Other consequences reported aretransposon activation, gene silencing orsequence elimination (see Hegarty and Hiscock2005). Within the Brassicaceae, extensivegenetic and genomic analyses in this contexthave been undertaken primarily in Arabidopsisand Brassica species (see Schmidt et al. 2001,Hall et al. 2002, Lysak and Lexer 2006).Expanding research activities to relatives ofArabidopsis thaliana is of great importance andopens future perspectives for studies within theBrassicaceae (Comai et al. 2003, Lysak et al.2003, Lysak and Lexer 2006).
Auto- and allopolyploidy – tracing the origin
of polyploids
The high percentage of polyploids in theBrassicaceae clearly shows that polyploidiza-tion processes must have been fundamental forspeciation in this family. The origin of polyp-loids, their successful establishment, geneticand genomic consequences, as well as evolu-tionary implications of polyploidy are chal-lenging topics. Especially with the advances ofmolecular biology techniques in recent dec-ades, such studies have received much atten-tion (reviewed e.g. by Wendel 2000, Osbornet al. 2003). Investigations of several modelsystems (e.g. Gossypium, Triticum, Aegilops,Nicotiana, Brassica) have demonstrated thehighly dynamic nature of polyploids, withincreasing evidence that each polyploid systemmay respond and evolve uniquely. Severalgenetic and genomic attributes of polyploidshave been suggested to account for theirevolutionary success, such as increased hetero-zygosity, increased genetic diversity throughmultiple formation, genome rearrangements,and changes in gene expression (Soltis andSoltis 2000). Major and rapid alterations togenome and chromosome organization havebeen observed in some allopolyploids (e.g.Levy and Feldman 2004, Lim et al. 2004),
while structural genomic stasis has beensuggested in others (Adams and Wendel2004, Ainouche et al. 2004). Numerous casesof changes in gene expression due to polyplo-idization have been reported as well (e.g.Kashkush et al. 2002, Levy and Feldman2004, Soltis et al. 2004a).
Allopolyploidy and long-distance dispersal,respectively, are regarded as prominent factorsin plant evolution and biogeography. Mum-menhoff and Franzke (2006) review the rarecases of prehistorical (not man-mediated)intercontinental long-distance dispersal ofplants combined with allopolyploidy. Allexamples given (Gossypium, Lepidium, Mi-croseris, Nicotiana) indicate a Late Tertiary/Quaternary evolution of the polyploid lineagesin the newly colonized continent. Late Ter-tiary, and especially Quaternary climatic fluc-tuations affected all parts of the world andthese changes might have created novel habi-tats providing new niches for rapid radiation(Mummenhoff and Franzke 2006).
Brassica, a model genus for the study of
polyploid evolution. Extensive genome rear-rangements, suggested to occur in a numberof polyploid crops, including tobacco, maize,soybeans and Brassica, are recognized as animportant source of genetic novelty in thepolyploids. Studies on synthetic experimentalallopolyploids produced from interspecificcrosses between Brassica rapa (AA genome,2n = 20) and B. nigra (BB genome, 2n = 16)and between B. rapa and B. oleracea (CCgenome, 2n = 18) (and subsequent self-pollinations up to the F5 generation) indicatedextensive genome changes in the early gener-ations after the polyploid formation (Songet al. 1995). Southern hybridization studiesusing several nuclear DNA probes haverevealed unexpected fragment profiles in thepolyploids, involving loss and/or gain ofparental restriction fragments and appearanceof novel fragments. Song et al. (1995) dis-cussed several potential causes of thesechanges in RFLP patterns, including chromo-some rearrangements, point mutations,gene conversion or DNA methylation. They
152 K. Marhold and J. Lihova: Polyploidy, hybridization and reticulate evolution

concluded that polyploids can generateextensive genetic diversity within a short timeperiod, which could significantly contribute totheir evolutionary success. Comparison ofgenetic linkage maps based on RFLP markersbetween natural allopolyploid B. juncea(AABB, 2n = 36) and its parental species(B. rapa and B. nigra) showed a high degree ofconservation of the A- and B-genomes in theallopolyploid and the diploids (Axelsson et al.2000). The collinearity observed in the geneticmaps indicated that the genome changesrevealed by Song et al. (1995) in the synthe-tized B. juncea are most probably not due toextensive intergenomic translocations, butoriginated from other genetic processes.
Studies on resynthesized Brassica polyp-loids also provided evidence of novel pheno-typic variation in several life-history traits andchanges in phenotypic plasticity, which havebeen associated with polyploidization andsubsequent genetic changes (Schranz and Os-born 2004). The research on Brassica speciesreviewed by Osborn (2004) provides evidencethat several mechanisms can contribute to denovo variation observed in newly formedpolyploids. The impact of polyploidy on dos-age-regulated genes was illustrated by studieson replicated FLC loci (flowering time regula-tor) in Brassica polyploids. Variation in flow-ering time observed within palaeopolyploidBrassica rapa may be due to the increasedallelic variation at multiple FLC loci. Newphenotypic variation can be generated inpolyploids also through intergenomic (homo-eologous) transpositions (suggested for B.napus), or epigenetic changes and alteredregulatory networks that affect gene expressionpatterns (Osborn et al. 2003, Osborn 2004).
Several other extensive genetic and molec-ular studies have been undertaken on Brassicaspecies but it is not in the scope of this reviewto deal with all of them in detail, instead wefocus on non-model examples of Brassicaceaepolyploids. They definitely represent new chal-lenges for future genetic and genomic studiescomparable to those in Brassica and Arabid-opsis, and have the potential to provide new
insights into the evolution of polyploidgenomes and to extend our current under-standing of this evolutionary phenomenon. Inthe following paragraphs, we will first presentstudies on several polyploid species illustratingtheir origins and evolutionary histories. Nextwe will point to some common or contrastingpatterns arising from those studies: the level ofgenetic variation found in polyploids in com-parison with their diploid progenitors, differ-ences in mating systems in diploids and theirpolyploid derivatives, the distinction betweenauto- and allopolyploidizations, and patternsassociated with single vs. polytopic and recur-rent polyploid origins.
Allopolyploid origins in Arabidopsis, Card-amine and Diplotaxis. To unravel polyploidorigin is undoubtedly a challenging task. It isnot always straightforward, and in some caseseven multiple molecular approaches do notbring unequivocal results. Most paleopolyp-loids have undergone intergenomic recombi-nation, genome rearrangement or diploidiza-tion, which often obscure their polyploidorigins and hamper identification of theirparentage. Genomes of both the originaldiploid progenitor and the polyploid havebecome substantially differentiated since thepolyploid formation and much effort is neededto elucidate their origin and evolutionaryhistory (Soltis et al. 2004b). Relatively recentallopolyploidization events, on the other hand,are usually much easier to reconstruct, as theaffinities to diploids and overall additivity aremostly still retained. The Brassicaceae familyoffers examples of both well-explored andrather simple allopolyploid origins, and com-plicated reticulate evolutionary histories ofhigh-polyploid complexes.
One of the best explored polyploids in thefamily, Arabidopsis suecica (2n = 26), hasattracted much interest, because one of itsparents is the model species A. thaliana. Withthe immense knowledge gathered on A. thali-ana, the tetraploid A. suecica offers uniqueopportunities for polyploid research, aspresented by Comai et al. (2000, 2003), Aliet al. (2004) and Madlung et al. (2005).
K. Marhold and J. Lihova: Polyploidy, hybridization and reticulate evolution 153

The parentage of A. suecica was first suggestedby Hylander (1957), based solely on morpho-logical and phytogeographical evidence.A. sue-cica was suggested to be an allopolyploid thathad arisen through hybridization betweenA. thaliana (2n = 2x = 10 ) and Cardamin-opsis arenosa (= A. arenosa, 2n = 2x,4x = 16, 32). Isoelectric focusing analysis ofpolypeptide composition of the large and smallsubunits of Rubisco provided molecular evi-dence for A. thaliana as the maternal parentand A. arenosa as the paternal one (Mummen-hoff and Hurka 1994). Further studies employ-ing restriction site analysis of cpDNA, isozymeanalyses and ITS sequence data corroboratedthis hypothesis (Mummenhoff and Hurka1995, O’Kane et al. 1996). As suggested forseveral other Brassicaceae polyploids (e.g. inCardamine, Draba, see below), its origin andspread to the current distribution area (south-ern Sweden and Finland) has most probablybeen associated with Pleistocene glaciation,more specifically, with glacier retreat providinglarge open areas. Interestingly, the parents ofA. suecica display different breeding systemsand extent of genetic variation. While A. tha-liana is a selfing species, characterized by highindividual homozygosity, low within- and highbetween-population differentiation, A. arenosais self-incompatible with high genetic diversity(Lind-Hallden et al. 2002, Sall et al. 2004). InA. suecica, low RAPD variation was found(especially within populations, 80% of thevariation was distributed among populations)indicating selfing (Lind-Hallden et al. 2002).Even the occurrence of apomixis wasdiscussed, which is extremely rare in theBrassicaceae, and otherwise observed only inBoechera species (Schranz et al. 2005). Micro-satellite genetic segregation patterns studied inA. suecica clearly showed that it reproducessexually with a high level of selfing (Sall et al.2004). This observation implies that the self-incompatibility of A. arenosa has broken downin the formation of the allopolyploid. The self-incompatibility in Brassicaceae is apparentlycontrolled by a multiallelic sporophytic sys-tem, and similar cases of shifts in mating
systems associated with polyploidization orhybridization have been found also in Capsellaand Diplotaxis species (see below). SyntheticA. suecica-like allopolyploids were producedby Comai et al. (2000), which together withnatural A. suecica were subjected to cytoge-netic (GISH and FISH) studies (Comai et al.2003, Ali et al. 2004). Both natural andsynthetic allopolyploids showed a patternconsistent with a genomic constitution of 10A. thaliana chromosomes and 16 A. arenosachromosomes. Homologous chromosomepairing observed in the synthetic polyploidsuggested that suppression of homoeologouspairing occurs already in early generationsafter polyploid formation, and was notachieved by adaptive mutations or by genomereshuffling as it might be expected (Comaiet al. 2003).
Recent molecular studies showed that thetraditional taxonomical circumscription ofArabidopsis and Arabis is largely artificial.Both were revealed to be polyphyletic, consist-ing of several independent lineages (Kochet al. 1999, Koch et al. 2003a, Al-Shehbazand Beilstein 2006). Al-Shehbaz et al. (1999)listed 59 binomials, previously assigned toArabidopsis, which are currently classified in14 genera. On the other hand, taxa formerlyplaced in Cardaminopsis are now included inArabidopsis, as well as several taxa, which wereoriginally described within Arabis. This holdstrue for the tetraploid taxa Arabis lyrata subsp.kamchatica and Arabis kawasakiana (Miyash-ita et al. 1998; Koch et al. 2000, 2001; Savo-lainen et al. 2000; Hoffmann 2005). There hasbeen much confusion about their taxonomicposition and circumscriptions, but the recentstudy by Shimizu et al. (2005) resolved thiscontroversy and also shed some light on theirpolyploid origin. Recently, O’Kane and Al-Shehbaz (1997) recognized three subspeciesunder Arabidopsis lyrata, the North Americansubsp. lyrata (2n = 2x = 16), the Europeansubsp. petraea (2n = 2x, 4x = 16, 32), andthe Far East/North American subsp. kam-chatica (2n = 4x = 32). This taxonomicconcept has also been supported by recent
154 K. Marhold and J. Lihova: Polyploidy, hybridization and reticulate evolution

ITS sequence data, where the three subspeciesformed a cohesive and well-supported clade inthe phylogenetic tree including other Arabid-opsis species (Hoffmann 2005). In their treat-ment, O’Kane and Al-Shehbaz (1997)considered Arabis kawasakiana as a synonymof Arabidopsis lyrata subsp. kamchatica. How-ever, as pointed by Shimizu et al. (2005), theformer is a winter annual species occurring onlow-altitudinal sandy shores of western Japan,while the latter is a mountain perennial morewidely distributed (Eastern Asia, northern-most North America). In addition, subsp.kamchatica appears to be distinct from A.lyrata subsp. lyrata and subsp. petraea, and thebest treatment reflecting its evolutionary his-tory is as a separate species A. kamchatica.Results from the nuclear single-copy gene ofchalcone synthase (CHS; see Koch et al. 2000)showed the presence of two homoeologous lociin the tetraploid A. kamchatica, which werehighly homologous to sequences retrievedfrom diploids A. halleri subsp. gemmifera andA. lyrata subsp. lyrata, respectively (Shimizuet al. 2005). This molecular data, together withthe morphological and cytological study byMulligan (1995) strongly suggest that A. kam-chatica is an allotetraploid species originatingfrom the above mentioned diploids. There aresome preliminary indications that A. kawasa-kiana is a later derivative of A. kamchatica,and their subspecific treatment as A. kamchat-ica subsp. kawasakiana and subsp. kamchaticawas suggested (Shimizu et al. 2005).
The use of low- or single-copy genes isstrongly encouraged for studies on polyploids,as they usually better reflect biparental lin-eages than commonly employed nrDNA andcpDNA markers (Mort and Crawford 2004,Small et al. 2004). Despite the technical dif-ficulties associated with polyploidy, theirapplication can provide valuable data ongenome composition of polyploids. The sin-gle-copy gene CHS has proven useful intracing the origin of the hexaploid Cardamineasarifolia (Lihova et al. 2006). Phylogeneticincongruence observed between nrDNA andcpDNA markers were resolved by the results
from CHS sequencing. Three CHS homoeo-logues were isolated from C. asarifolia, con-sistent with its hexaploid status and providingevidence for its allopolyploid origin. Phyloge-netic placements of the homoeologous se-quences pointed to the potential parentalspecies; C. amara s.l. was identified as thematernal parent, while one CHS homoeo-logue close to C. hirsuta most likely origi-nated from an extinct parent (Lihova et al.2006).
The genus Diplotaxis comprising around 30species distributed mainly in the EuropeanMediterranean has no economic importance,but it represents one of the nearest wildrelatives to Brassica. It may provide a valuablegenetic resource for plant breeding programsand crop improvement, and studies on thisgenus may be therefore of special relevance(Gomez-Campo 1999, Martın and Sanchez-Yelamo 2000). Phylogenetic relationshipswithin the genus as well as the polyploid originof the tetraploid Diplotaxis muralis have beenaddressed in several studies employing differ-ent markers. Based on morphological andcytological evidence, the allopolyploid originof D. muralis (2n = 42) from D. tenuifolia (2n= 22) and D. viminea (2n = 20) was firstsuggested by Harberd and McArthur (1972).Its genomic constitution has been subsequentlystudied by isoelectric focusing patterns ofRubisco, isozymes, restriction site variationof the cpDNA, RAPDs, and microsatellites(Warwick et al. 1992; Warwick and Anderson1997a; Mummenhoff et al. 1993; Martın andSanchez-Yelamo 2000; Eschmann-Grupe et al.2003, 2004). All the data confirmed the allo-tetraploid origin of D. muralis with the par-entage as originally proposed by Harberd andMcArthur (1972), and indicated that D. vim-inea was the maternal parent. Both the allote-traploid D. muralis and diploid D. tenuifoliaare successful colonizers; D. muralis indeeddisplays some common features of colonizingplants, such as polyploidy, annual to bienniallife form and selfing, while D. tenuifolia isperennial and strictly allogamous (Eschmann-Grupe et al. 2004). Strong differences in
K. Marhold and J. Lihova: Polyploidy, hybridization and reticulate evolution 155

genetic variation between these two specieswere found. Tetraploid D. muralis in contrastto D. tenuifolia displayed very low geneticvariation in RAPDs and isozymes. Thispattern may be explained by its young phylo-genetic origin, but also population history(colonization history involving e.g. geneticdrift, founder effects and local extinctions)might have caused the limited genetic variationin this tetraploid (Eschmann-Grupe et al.2003, 2004).
Biogeographic and evolutionary history
of polyploids in conjunction with Pleistocene
climatic changes: Cardamine and Microthlaspi.It has been widely recognized that the currentdistribution of genetic variation and geo-graphic patterns in the Northern Hemispherehave been significantly shaped by Pleistoceneclimatic changes. Plant migration duringglacial periods may have restricted or dis-rupted their original continuous range and ledto population diversification, but secondarycontact has often occurred during recoloniza-tion (Hewitt 2004, for Brassicaceae examplessee Koch and Kiefer 2006). Geographicpatterns and distribution of genetic diversityin a given polyploid, particularly whenassessed from multiple markers, can helpelucidate both its evolutionary and biogeo-graphic history, which are often tightly cou-pled. Two examples can be mentioned wherehybridization and subsequent polyploidizationmost likely predated Pleistocene glaciationprocesses, and where climatic changes havesignificantly impacted on their geographicdistribution and genetic variation patterns:tetraploid Cardamine amporitana (Lihovaet al. 2004a, b) and diploid to hexaploidMicrothlaspi perfoliatum (Koch and Bernhardt2004). There have also been numerous studiesdating the origin of polyploids to the time ofPleistocene population migrations and second-ary contacts, as also will be discussed for thehexaploid Cardamine silana (Perny et al.2005a, Lihova et al. 2004a).
Disjunct distribution has been observed inCardamine amporitana occurring in Catalonia(NE Spain) and central Italy (Lihova et al.
2004a, b). Although no substantial geneticdifferentiation has been found between thesetwo disjunct areas, Catalonian populationsdisplayed strong genetic depauperation incomparison with Italian ones. Alternativebiogeographic hypotheses to explain the abovepattern were discussed: (1) origin in Italy andlater colonization of Catalonia either by long-distance dispersal or migration through theregions connecting both peninsulas, associatedwith the reduction of genetic diversity throughfounder effects; (2) fragmentation of originallarger distribution area, disappearance of con-necting populations and genetic impoverish-ment in the western colonization route (Lihovaet al. 2004a, b). Despite the use of severalmarkers (AFLPs, cpDNA and ITS sequences),the polyploid origin of C. amporitana couldnot be unequivocally proved, and both ancientautopolyploid and allopolyploid origin withinthe C. amara group were considered as twoequally plausible hypotheses (Lihova et al.2004a, b).
On the other hand, the resolution of theorigin and parentage of polyploid Microthlaspiperfoliatum was more straightforward (Mum-menhoff et al. 1997, Koch et al. 1998b, Kochand Hurka 1999). Microthlaspi perfoliatum isan annual species widely distributed in Europeand represented by diploid, tetraploid andhexaploid cytotypes. Together with three otherdiploids, it has usually been treated as theM. perfoliatum aggregate. Although the differ-ent cytotypes can be hardly distinguishedmorphologically, based on molecular markersthey probably should be treated as two sepa-rate species (diploid and polyploid). Severalinvestigations using different markers (iso-zymes, cpDNA and nrDNA data) have docu-mented the allopolyploid origin of M.perfoliatum populations from the diploids M.perfoliatum and M. natolicum (Mummenhoffet al. 1997, Koch et al. 1998b, Koch andHurka 1999). Compared to the diploids, thepolyploid populations displayed significantlyhigher levels of genetic diversity as well as awider distribution area. Apparently, glaciationmust have had a greater influence on diploid
156 K. Marhold and J. Lihova: Polyploidy, hybridization and reticulate evolution

than on polyploid M. perfoliatum. Extinctionof most of the genetic variability in the diploidpopulations has been assumed, whereas polyp-loids maintained genetically differentiated pop-ulations (Koch and Hurka 1999). Differentbiogeographic histories and glacial refugiawere revealed for the diploid and polyploidcytotypes. The polyploid populations showedthe classical European pattern with three maindiversity centers in Iberia, Italy and the Bal-kans, while the diploids experienced substan-tial fragmentation and extinction, and wereforced into two refugia (SE France and Aus-tria/Croatia; Koch and Bernhardt 2004).
Molecular marker and morphological stud-ies have revealed the origin of the Calabrian(southern Italy) hexaploid endemic Cardaminesilana. The results provided strong supportthat two diploid species were involved in itspolyploid origin, the central Italian C. apenn-ina and Balkan C. acris (Perny et al. 2005a,Lihova et al. 2004a, see also Lihova andMarhold 2006). ITS sequences from C. silanaindicated that this multicopy DNA region,typically subjected to sequence homogeniza-tion, still retained (at least) two differentsequence variants. Intra-individual polymor-phisms detected in C. silana displayed additivepatterns, strongly suggesting that one ITSsequence variant comes from C. apennina,while the other is shared by several relatedspecies. The AFLP-fingerprinting profile, rep-resenting variation distributed throughout thewhole genome, on the other hand, revealedstrong affinity to the Balkan C. acris. Geo-graphic separation of both assumed parentalspecies from C. silana might be at first sightsurprising. There is, however, clear evidencefor past migration and gene flow between theApennine and Balkan Peninsulas seen in phy-logeographic studies (e.g. Fineschi et al. 2002),as well as Italian-Balkan distribution patternsof numerous taxa (for examples see Pernyet al. 2005a). In addition, Calabria is wellknown as an important refugium duringPleistocene climatic changes, where popula-tions of several species distributed in higherlatitudes (including C. apennina) could have
found favourable conditions during colderperiods (Taberlet et al. 1998, Fineschi et al.2002, Palme and Vendramin 2002). The pres-ent data allow us only to speculate on whereand when both presumable parental speciesmet and whether the few remaining popula-tions are remnants of a previously morewidespread species, or were always restrictedto the Sila Mts. Populations of C. silana do notappear genetically strongly depauperate (asshown by AFLP and cpDNA sequence data),but the number of individuals analyzed wastoo low to assess the level and distribution ofgenetic variation that could shed light on itsphylogeographic history (Perny et al. 2005a).
Complex polyploid speciation: evolutionary
scenaria in Draba, Cardamine and Cochlea-ria. When reviewing polyploidy as an impor-tant speciation mode in the Brassicaceae,Draba, the largest genus in the family atapprox. 350 species, cannot be omitted. Thefrequency of polyploid taxa is around 70%(based on data from Warwick and Al-Shehbaz2006), and both auto- and allopolyploidy haveapparently been common in its evolutionaryhistory. The ploidy level ranges from diploidsup to 18-ploids. The genus is well known for itscomplexity in terms of morphological varia-tion, reticulate evolution and consequentlytaxonomy (Grundt et al. 2004). It has beenhypothesized that the complexity may becompounded by several factors, such as recur-rent formation of the polyploids, gene flowacross ploidy levels, repeated migration andcolonization resulting in secondary contactsbetween previously isolated populations(Brochmann et al. 1992a, d; Koch and Al-Shehbaz 2002; Grundt et al. 2004). There havebeen numerous investigations, which aimed toresolve this evolutionary and taxonomic com-plexity in selected species groups (e.g. Broch-mann et al. 1992a, b, c, 1993; Koch and Al-Shehbaz 2002). While there is unequivocalevidence for the allopolyploid origin of theSwiss endemic D. ladina (Widmer and Baltis-berger 1999), much more complex pictureshave been found in arctic-alpine groupswhich involve high-polyploids. Additivity of
K. Marhold and J. Lihova: Polyploidy, hybridization and reticulate evolution 157

ITS sequences and the cpDNA haplotypefound in the tetraploid D. ladina supportedD. aizoides as the paternal and D. tomentosa asthe maternal parent. Despite the high tomoderate intraspecific genetic variation pres-ent in these widespread diploid parents, novariation has been observed in D. ladina,strongly supporting its single origin (Widmerand Baltisberger 1999). As for the arctic-alpinegroup, fixed heterozygosity observed in thecircumpolar tetraploid/hexaploid D. lacteaimplied genetic allopolyploidy, and diploidsD. fladnizensis, D. nivalis and/or D. subcapitatawere suggested as the putative parents (Broch-mann et al. 1992a). A recent study, however,based on several markers including the single-copy nuclear gene RPD2 showed that D. lacteaand other related polyploids (D. turczaninovii,D. porsildii) may have originated from singlediploid lineages. If allopolyploidization wasthe case, this must have involved very similargenomes (Grundt et al. 2004). It has beenhypothesized that hexaploid D. lactea aroserecurrently from the Beringian diploid D.palanderiana lineage via tetraploid D. lactea.This apparent contradiction (genetic allopoly-ploidy vs. taxonomic autopolyploidy) can beexplained by the existence of cryptic specieswithin diploid taxonomic species. Crossingexperiments have indicated that several diploidinbreeding Draba species comprise numerous‘strains’, which are cross-incompatible. Occa-sionally, the crossing barriers can break downand the cryptic species may produce a geneticallopolyploid (Brochmann et al. 2004, Grundtet al. 2004).
We may face a similar situation of crypticspecies also in diploid Cardamine pratensiss.str. This species includes several widely dis-tributed cytotypes from diploids up to heptap-loids. Molecular studies suggest that thepolyploids have evolved repeatedly from dip-loids, and can be considered as taxonomicautopolyploids (Franzke and Hurka 2000,Lihova et al. 2003). However, diploid popula-tions from the Alps and adjacent areas havebeen reported to be ecologically and evencytogenetically (based on karyotype character-
istics) mutually differentiated (Urbanska-Wor-ytkiewicz and Landolt 1974). The molecularmarkers applied, however, did not yieldenough resolution to find genetic divergenceamong those diploid populations.
Polyploidization has also been an impor-tant speciation driving force in Cochlearia. Thesection Cochlearia is a highly polymorphicspecies complex with considerable cytogeneticdiversity (with two base chromosome numbersand several polyploids), different ecologicaladaptations and geographic patterns. It isconsidered to be of recent Pleistocene origin,showing only little morphological differentia-tion and genetic divergence among the taxa(Koch et al. 1996). Several studies devoted tothis species group (Koch et al. 1996, 1998a;Koch 2002) indicated a complex evolutionaryhistory, including both auto- and allopolyploidorigins. The scenario displayed in Fig. 1 hasbeen proposed based on molecular data fromseveral markers, and is also supported bymorphological, cytological and ecological evi-dence. Apparently, the tetraploid C. officinalis,distributed along the northern coasts ofEurope and assumed to originate via auto- orallopolyploidization from ancestors of thediploid C. aestuaria, have played a central rolein speciation processes in this section. Thisspecies, showing extensive morphological andecological variation, contributed to the originof several higher polyploids, both via auto- (C.anglica) and allopolyploidizations (C. polonica,C. bavarica, C. tatrae). An interesting biogeo-graphic pattern has been observed within thehexaploid C. bavarica restricted to two disjunctregions in S Germany (Bavaria). Significantgenetic differentiation revealed between thesetwo regions raised the question whether itreflects: (1) polytopic origin, (2) migrationfrom the original range and subsequent geneticdifferentiation, or (3) disruption of a formerwider distribution area through populationextinctions. The third scenario was favouredbased on recent isozyme data (Koch 2002).
Autopolyploid origins reported in Card-amine, Biscutella and Capsella. Recent studiesfrom various angiosperm lineages indicate that
158 K. Marhold and J. Lihova: Polyploidy, hybridization and reticulate evolution

autopolyploidy is much more common thantraditionally considered and apparently hasplayed an important role in plant evolution(Levin 2002, Soltis et al. 2004b). As in Coch-learia (see above and Fig. 1), a few cases withevidence or indication for autopolyploidy havebeen documented in Cardamine and Biscutella.In Capsella, despite extensive studies, thepolyploid origin of C. bursa-pastoris has notbeen unequivocally resolved, but ancient auto-polyploidization has been favoured by Hurkaand Neuffer (1997).
Tetraploid populations of Cardamine amar-a occupying the area of the Eastern Alps andadjacent regions (C. amara subsp. austriaca)have been studied using several molecularmarkers, including isozymes, nuclear and chlo-roplast DNA sequences, as well as RAPD andAFLP fingerprinting (Marhold 1999; Lihovaet al. 2000, 2004b; Marhold et al. 2002a). The
results showed very low genetic differentiationbetween diploid C. amara subsp. amara andtetraploid subsp. austriaca, and only very fewalleles or fragments unique to either of thesubspecies (in contrast to other three subspe-cies). This, together with only slight morpho-logical differentiation and clear geographicalpatterns, strongly favoured autopolyploidorigin of the tetraploid subspecies. As thedistribution of subsp. austriaca coincides withthe area heavily affected by Pleistocene glaci-ation, we assume that the autopolyploidizationand establishment of this new tetraploid taxontook place in the last interglacial or postglacialperiod when new habitats became availableafter glacier retreat. Whether the autotetra-ploid has a single origin and colonized the Alpsspreading from a single refuge or whethermultiple events and different colonizationroutes can be traced, remains an open question.
diploid ancestral Cochlearia
C. aestuaria2n=2x=12
C. officinalis2n=4x=24
C. danica2n=6x=42
subsp. officinalis
subsp. norvegica
subsp. integrifolia
C. excelsa2n=2x=12
C. macrorrhiza2n=2x=12
C. pyrenaica2n=2x=12
C. polonica C. bavarica C. tatrae
C. anglica2n=8x,10x=48,60
C. groenlandica
2n=6x
2n=6x=36 2n=6x=42
2n=2x=14 2n=2x=12
2n=4x
Fig. 1. Evolutionary scenario in Cochlearia sect. Cochlearia according to Koch et al. (1998). Differentcolours indicate different ploidy levels in polyploids. Reproduced and modified with permission from thefirst author
K. Marhold and J. Lihova: Polyploidy, hybridization and reticulate evolution 159

Another suspected autopolyploid is Card-amine majovskyi, a recently described tetraploidspecies occurring in Central and southeasternEurope. C. majovskyi and its assumed progen-itor, diploid C. matthioli, were included in thecomplex molecular systematic and biogeo-graphic study of theC. pratensis group (Franzkeand Hurka 2000, Lihova et al. 2003). Allanalyses involving isozymes, nuclear andcpDNA sequences, RAPD and AFLP markersshowed a very close relationship between thesetwo species. Cardamine majovskyi is largelysympatric with C. matthioli. This fact, togetherwith slight morphological differences observedamong populations of C. majovskyi from dif-ferent parts of its distribution area (Lihova andMarhold 2003; and K. Marhold, unpubl. data)might indicate its recurrent and independentorigins. Thorough sampling and high-reso-lution molecular markers will be needed infuture studies to corroborate this assumption.
In the highly variable species Biscutellalaevigata, genetic autopolyploidy has beenfound in tetraploid populations (classified assubsp. laevigata) based on isozyme data (Tre-metsberger et al. 2002). These populationsexhibited tetrasomic inheritance and thusabsence of fixed heterozygosity, and showedonly moderate divergence from the diploids,implying moreover their recent origin. As thetetraploids showed genetic resemblance todifferent diploid subgroups, their multipleorigins have been hypothesized. Presumably,the resulting high genetic variation and greatergenomic plasticity have contributed to theevolutionary success of tetraploids over dip-loids in alpine habitats (Tremetsberger et al.2002).
An interesting case of polyploid speciationis also reported in Capsella. Only three specieshave been recognized in this genus, two geo-graphically restricted diploids (C. rubella, C.grandiflora) and a single tetraploid, the world-wide successful colonizer C. bursa-pastoris. Inspite of extensive studies (Hurka et al. 1989,Mummenhoff and Hurka 1990, Hurka andNeuffer 1997), the origin of the tetraploid hasnot been unequivocally resolved, most likely
due to its ancient origin. New alleles identifiedat isozyme loci not found in the diploids,presence of null alleles indicating gene silenc-ing, and considerable isozyme and RAPDvariability favour an ancient polyploidizationevent, supported also from fossil records(Hurka and Neuffer 1997). Both allopolyploidand autopolyploid origin from C. grandifloracan be considered. Disomic allozyme inheri-tance and fixed heterozygosity would suggestthe allotetraploid origin. Nevertheless, C.rubella seems to be a younger derivative of C.grandiflora, and no other extant species fromanother genus appears to be closely related tohybridize with Capsella. It can be hypothesizedthat the second parent is extinct, but based onavailable evidence, Hurka and Neuffer (1997)concluded that C. bursa-pastoris most likelyarose via autopolyploidization from C. gran-diflora a long time ago. Recently publishedreports on comparative chromosome paintingof C. rubella and C. bursa-pastoris are prom-ising and might in future studies bring moreresolution in this respect, provided that C.grandifora will display a different paintingpattern than C. rubella (Lysak et al. 2003).Similar to Arabidopsis suecica, speciation pro-cesses in Capsella were accompanied by aswitch in mating systems. While C. grandiflorais self-incompatible, both its assumed deriva-tives, C. rubella and C. bursa-pastoris, arepredominantly selfing species (Hurka andNeuffer 1997).
Contrasting patterns of genetic diversity in
polyploids and their possible determinants. Theabove survey of several recent studies focusingon the origin and evolution of polyploids in theBrassicaceae points to some interesting pat-terns that can be further discussed. There havebeen numerous studies and reviews relatinggenetic diversity in polyploids to their evolu-tionary success (see e.g. Soltis and Soltis 2000,Soltis et al. 2004b). Increased genetic diversityin polyploids, when compared to putativediploid parents, has been observed in poly-ploid Microthlaspi perfoliatum and it appar-ently correlates with increased morphologicalvariation and larger geographic distribution.
160 K. Marhold and J. Lihova: Polyploidy, hybridization and reticulate evolution

Preglacial polyploid origin and different bi-ogeographic history than in diploids have beenproposed (Koch and Bernhardt 2004). In-creased variation and successful colonizationof predominantly alpine areas have also beenrevealed in tetraploid Biscutella laevigatasubsp. laevigata that most likely reflect itsmultiple origins (Tremetsberger et al. 2002).The best example is probably Capsella bursa-pastoris, the tetraploid with high genetic var-iation and world-wide distribution, whichcontrasts with restricted areas of the relateddiploids (Hurka and Neuffer 1997). Despitethe limited sampling, several markers indicatedthat the assumed autotetraploid Cardamineamara subsp. austriaca harbours genetic vari-ation comparable to that of its putative parentsubsp. amara. This would favour multipleautopolyploidization events, supported alsofrom the presence of several cpDNA haplo-types in the tetraploid subspecies (Lihova et al.2000, 2004a, b; Marhold et al. 2002a). On theother hand, the allopolyploid Diplotaxis mu-ralis distributed across most of Europe dis-played surprisingly low genetic variation,attributable to its recent origin and/or loss ofgenetic variation through extinctions, bottle-neck and genetic drift effects (Eschmann-Grupe et al. 2004). Low genetic diversity dueto single origins has also been found in Drabaladina and Arabidopsis suecica, and both butespecially the former remained localized withina small geographic area (Widmer and Baltis-berger 1999, Lind-Hallden et al. 2002). InArabidopsis suecica, the low level of geneticvariation is explained also by its autogamousreproduction mode (Sall et al. 2004).
Shifts in mating systems associated with
polyploid speciation. An important determi-nant of the distribution of genetic variationand evolutionary success of a particular poly-ploid species is its reproduction mode. Matingsystem can significantly influence polyploidestablishment. It has been hypothesized thatself-fertilization in polyploids may facilitatetheir establishment because of limited back-crossing to their parents resulting in sterileprogeny. Selfing also ensures the persistence of
a low frequency or single hybridization events.In addition, theoretical models predict thatpolyploids relative to their diploid parent(s)should have reduced inbreeding depression,allowing for increased selfing rates. Fewempirical studies addressing this issue havebeen published (reviewed by Soltis and Soltis2000, see also Rausch and Morgan 2005). InDraba, inbreeding is common, and as indicatedby Brochmann (1993), diploid arctic Drabaspecies are genetically depauperate due tohigher levels of selfing, while polyploids dis-playing either predominantly autogamous ormixed mating maintain high genetic variation.Although the hypothesis that polyploids ex-hibit higher selfing rates than diploids has notbeen supported for the studied Draba species,the allopolyploidy probably serves here as anescape from genetic depauperation caused bydrift and inbreeding at the diploid level. Fixedheterozygosity in allopolyploids ensures thatgenetic diversity is maintained through gener-ations despite inbreeding (Brochmann 1993,Brochmann et al. 2004). In several Brassica-ceae polyploid species, shifts in mating systemtowards inbreeding have indeed been ob-served, in contrast to outcrossing or evenself-incompatibility in their diploid parent(s).We can hypothesize that the high level ofselfing found in the polyploids Arabidopsissuecica (Sall et al. 2004), Capsella bursa-pasto-ris (Hurka and Neuffer 1997) and Diplotaxismuralis (Eschmann-Grupe et al. 2004) hascontributed to their evolutionary success.
Distinction between auto- and allopolyploid
origins. Auto- vs. allopolyploid origins canusually be distinguished from genetic or geno-mic patterns, nevertheless, in some casesespecially in ancient polyploidization eventsthis distinction may not be conclusive. Geneticallopolyploids are characterized by disomicinheritance at each locus and fixed (nonsegre-gating) heterozygosity, while in autopolyploidsmultisomic inheritance typically occurs (Soltisand Soltis 2000). In Biscutella laevigata subsp.laevigata the absence of fixed heterozygosityand tetrasomic inheritance observed in iso-zyme data confirm its autopolyploid origin
K. Marhold and J. Lihova: Polyploidy, hybridization and reticulate evolution 161

(Tremetsberger et al. 2002). Close geneticresemblance and the absence of uniquealleles/fragments in a polyploid when com-pared with a putative diploid progenitor canalso be considered a reliable indicator ofautopolyploidy, as suggested for Cardamineamara subsp. amara (Lihova et al. 2000,2004b; Marhold et al. 2002a) and C. majovskyi(Franzke and Hurka 2000, Lihova et al. 2003).Very low genetic divergence between thediploids, however, may hamper the distinctionbetween auto- and allopolyploidy, unlessmarkers specific for either of the diploids arefound. Although fixed heterozygosity has beenobserved in Draba polyploids, results from thesingle-copy RPD2 gene implied that eitherautopolyploidy or allopolyploidy involvingvery similar genomes (from cryptic species)had occurred (Grundt et al. 2004). Similarly,the origins of tetraploids Capsella bursa-pasto-ris (Hurka and Neuffer 1997), Cardamineamporitana (Lihova et al. 2004a) and Cochlea-ria officinalis (Koch 2002) have not beenelucidated so far despite extensive studies.Future investigations employing other markersor newly available techniques may providemore resolution. Allopolyploid origins can beclearly proven from additive genetic patterns,as observed at isozyme loci (Arabidopsis sue-cica: Mummenhoff and Hurka 1995, Microthl-aspi perfoliatum: Koch and Hurka 1999), inmultilocus fingerprinting marker systems ofAFLPs or RAPDs (Cochlearia bavarica: Kochet al. 1996, Diplotaxis muralis: Eschmann-Grupe et al. 2004), and in nrDNA ITSsequences (Cardamine silana: Lihova et al.2004a, Microthlaspi perfoliatum: Mummenhoffet al. 1997, Draba ladina: Widmer and Baltis-berger 1999). Nuclear encoded single-copygenes have the potential to identify allopoly-ploid origins and involved parental speciesmore efficiently. The presence of two or morehomoeologous loci in a polyploid, correspond-ing to the sequences resolved from its diploidrelatives has been documented for Cardamineasarifolia (Lihova et al. 2006) and Ara-bidopsis kamchatica subsp. kamchatica (Shi-mizu et al. 2005). Genomic studies, such as
GISH can identify and separate parentalgenomes in the studied polyploids, as wasdemonstrated for Arabidopsis suecica (Aliet al. 2004).
Single vs. polytopic (multiple) origins of
polyploid taxa. Molecular studies have contin-ued to reveal that multiple origins of eitherauto- or allopolyploids from the same diploidprogenitors are common (Soltis and Soltis2000, Soltis et al. 2004b). It seems that poly-topic and recurrent polyploid origins are muchmore widespread than traditionally consid-ered, and may represent a significant source ofgenetic diversity in polyploids (Soltis et al.2004b). To prove the single vs. multiple originsunequivocally, however, it is crucial to sampleallelic (or haplotype) diversity in both thepolyploid and its diploid progenitor(s) (Doyleet al. 2004). Taxonomic and evolutionarycomplexity in the highly polymorphic genusDraba has been explained by multiple originsof several widespread high-polyploid species,as clearly proved in Draba lactea (Brochmannet al. 2004, Grundt et al. 2004). Multipleorigins have also been proposed for Arabidop-sis suecica based on isozyme data (Mummen-hoff and Hurka 1995), although extremely lowRAPD variation may be an indication for asingle origin (Lind-Hallden et al. 2002). Theformer scenario is favoured by the presence oftwo fixed heterozygous genotypes in A. suecicafor the Pgm loci, corresponding to the allelicpolymorphism at the same locus in its progen-itors. Although this intraspecific variation inA. suecicamight be explained by evolution of anew allele following a single polyploid origin,this hypothesis is unlikely (Mummenhoff andHurka 1995). Multiple autopolyploid originsfrom genetically differentiated progenitor pop-ulations have been proposed for Biscutellalaevigata subsp. laevigata (Tremetsberger et al.2002). Single origin, on the other hand, hasbeen strongly favoured for the local endemicDraba ladina with less than 12 known popu-lations. Lack of intraspecific variation inseveral individuals from two distinct locations,despite the significant variation present in bothparents, is a strong indication for the single
162 K. Marhold and J. Lihova: Polyploidy, hybridization and reticulate evolution

origin (Widmer and Baltisberger 1999).Although multiple cpDNA haplotypes with aclear geographic structure were found in boththe hexaploid Cardamine asarifolia and itsmaternal parent (C. amara), they shared onlyone of them. The remaining haplotypes wereunique to C. asarifolia and, based on statisticalparsimony analysis, they were apparentlyderived from the shared one. Thus, it has beensuggested that cpDNA diversification occurredafter the polyploidization event, but furtherstudies are still needed (Lihova et al. 2006).
The above studies illustrate that the Brass-icaceae harbour polyploids with very differentevolutionary and biogeographic histories, andprovide an immense resource for future stud-ies. As new molecular markers and techniquesfor genomic studies become available, thespectrum of powerful tools to study thegenomic composition and evolution of polyp-loids increases, and hopefully will soon find itswide application beyond the model plants.
Reticulate evolution
Polyploidization and hybridization are events,which often result in a reticulated pattern ofevolution. Reticulation complicates recon-struction of evolutionary relationships andmakes phylogeny inferences a challengingundertaking. Reticulate evolution can bedetected via incongruence between gene trees,although other processes can sometimes pro-duce similar discordant patterns. Disentan-gling reticulate from divergent relationshipsrequires use of multiple independent markerswith different modes of inheritance (Doyle1992, Linder and Rieseberg 2004, Vriesendorpand Bakker 2005). Investigation of the addi-tivity of molecular markers, as well as addi-tional data such as morphology, chromosomenumbers, GISH or FISH patterns, C-values,and geographic distribution, are often crucialto unravel ancient hybridization events and todistinguish them from population geneticprocesses that may confound the phylogeneticsignal (Linder and Rieseberg 2004, Vriesen-dorp and Bakker 2005). Numerous examples
of incongruent patterns attributed to reticu-lation can be found in recent Brassicaceaestudies, which are here shortly reviewed.
A highly reticulate evolution has beenidentified in Lepidium, a large worldwidedistributed genus with conspicuous floral var-iation and a predominantly autogamous mat-ing system. The incongruence of speciesrelationships based on maternally inheritedcpDNA versus nrDNA ITS markers togetherwith the common occurrence of polyploids hassuggested predominance of allopolyploid spe-ciation in this genus (Bowman et al. 1999,Mummenhoff et al. 2001, 2004). This indica-tion has been further supported by the resultsfrom the single-copy nuclear gene PI (PISTIL-LATA), since multiple phylogenetically dis-tinct sequences have been found in severalinvestigated species from Americas and Aus-tralia (Fig. 2, designated as group 1; Lee et al.2002). On the basis of previous cpDNA andITS sequence data, and ploidy levels of thosespecies, allopolyploidization (and thus homo-eologous origin of the multiple sequences) hasbeen strongly favoured to explain the observedpatterns. Four major groupings of allopolyp-loids composed of two genomes, and onegrouping of taxa composed of even threegenomes have been identified (Fig. 2). In someof them, multiple but very closely relatedsequence types have been found even withina clade, and these may have their origin eitherin allelic variation (proposed e.g. for L. monta-num) or in gene duplication (proposed for L.densiflorum). The origin of several Australian/New Zealand polyploid species (Fig. 2, desig-nated as group 2) still remains unresolved.Although only a single PI intron sequence hasbeen resolved from each species, their positionsin the ITS and cpDNA gene trees wereincongruent (Lee et al. 2002, Mummenhoffet al. 2004). In addition, there are indicationsfrom RFLP analyses that they possess twohomoeologous loci, but one locus apparentlyhas not been detected by PCR due to eitherpseudogenization or interlocus recombination(Lee et al. 2002). Inferring from these data, itis likely that they are allopolyploids as well.
K. Marhold and J. Lihova: Polyploidy, hybridization and reticulate evolution 163

Reticulation has been suggested to occuralso in the evolution of the Eurasian/NorthAmerican group of Braya and Neotorulariaspecies, with the assumption of both recentand more ancient hybridizations. Discordantphylogenetic placements of Braya rosea andNeotorularia brachycarpa in respect of thenrDNA ITS and trnL cpDNA gene trees,and an apparent ITS sequence homogenizationare in favour of an ancient reticulation. On theother hand, nucleotide additivity in ITSsequences observed in several accessions ofvarious Braya species probably reflects recentor even on-going hybridization (Warwick et al.2004).
Much controversy has surrounded thetaxonomic treatment of the Chinese endemics
recently treated within three genera Yinshania,Hilliella and Cochleariella. Molecular phylo-genetic analyses of ITS and trnL sequencedata have identified two main lineages, onecorresponding to the diploid Yinshania, thesecond lineage including the remaining twopolyploid genera (Koch and Al-Shehbaz2000). Comparisons of the two gene treesrevealed incongruences, which strongly sug-gested gene flow between these two mainlineages, and all taxa should probably betreated within a single widely conceived genus.Hybridizations within the polyploid Hilliella/Cochleariella clade and subsequent differentialITS sequence homogenization have beenproven as well. This has been illustrated alsoby the split decomposition analysis, which has
L. phlebopetalumL. leptopetalum (3x)L. heterophyllum (2x)L. hirtum ssp. calycotrichum (2x)L. campastre (2x)L. perfoliatum (2x)
L. ruderale (2x, 4x)L. cordatumL. latifolium (3x, 6x)L. pinnatifidum (2n=28)L. vesicarium (2x)
L. spinosum (2x, 3x)
L. africanum (2x)L. myriocarpumL. allaudii (2x)
L. graminifolium (6x)
L. lyratum (2x)L. persicum (2x)
L. sativum (3x, 4x)L. apetalum (4x)
B
C
L. oxytrichumL. flexicaule
L. densiflorumL. lasiocarpumL. virginicumL. montanum
L. bippinatifidumL. nitidum
L. meyeniiL. oblongum
L. pseudohyssopifolium
L. bonarienseL. chichicara
D
A
L. aschersoniiL. desvauxiiL. pseudotasmanicum (2n=28)L. hyssopifoliumL. oleraceumL. fremontii
L. quitenseL. dictyotum
L. monoplocoides
L. flavum (4x)
L. armoracia
(3x)
(4x)(4x)(4x)(4x)
(4x, 8x)(4x)
(4x)
(4x, 8x)
(4x)(4x)
(4x)
(4x, 8x)
(8x)
(4x)
group 1 group 2
Fig. 2. Polyploid speciation in Lepidium based on the PI intron gene tree. Group 1 is represented byallopolyploid species composed of genomes originating in two or three different clades (A-D). In taxa from thegroup 2, a single PI intron sequence has been resolved from each species, but at least for some taxa, theirallopolyploid origin has been favoured from other evidence. Australian/New Zealand taxa are set in bold.Reproduced and modified with permission from Lee et al. (2002); � 2002, National Academy of Sciences,U.S.A.
164 K. Marhold and J. Lihova: Polyploidy, hybridization and reticulate evolution

the potential to display conflicting phyloge-netic signal within a single marker. Accessionsof hybrid origin with unidirectional ITSsequence homogenization as well as with thepresence of both paternal sequence types werefound. One accession of H. sinuata showed amosaic recombinant sequence type (Koch andAl-Shehbaz 2000).
Complex evolutionary history includinghybridization, polyploidization and facultativeapomixis in the North American genus Boec-hera has been recently addressed in severalstudies. It has been assumed that extensivereticulation has played an important role in theevolution of the highly polymorphic taxonB. holboellii, which includes diploids, triploidsand aneuploids. As many as 70 different ITSsequence types have been found in this species,which did not form a single well-supportedclade, but were placed in several unresolvedbranches sister to another species, B. stricta(Dobes et al. 2004a). This, together with thefrequent occurrence of intra-individual ITSpolymorphism strongly suggests extensivereticulation. ITS, cpDNA as well as microsat-ellite data provided clear evidence that trip-loids have originated repeatedly from diploidlineages, and that hybridization between genet-ically distinct lineages most likely contributedto the high genetic diversity found in thisspecies (Sharbel and Mitchell-Olds 2001,Dobes et al. 2004a). In addition, occasionalintrogression of B. holboellii towards B. strictahas been detected from microsatellite variationpatterns (Dobes et al. 2004a). The most recentstudy further suggests that B. holboellii is ofpolyphyletic hybrid origin, as its cpDNAhaplotypes have been found interspersedamong the other investigated Boechera taxa(Schranz et al. 2005).
Reconstruction of evolutionary history ingenera strongly affected by reticulation andpolyploidization is definitely not an easy task.Forcing reticulation to be displayed in thebranching topology of a phylogenetic treemight lead to biased patterns, lack of supportfor the resolved clades, and collapse of hierar-chical structure (Bachmann 2000, Vriesendorp
and Bakker 2005). To decrease the risk ofproducing false phylogenies in such cases, onecould suggest the removal of all knownpolyploids from the phylogenetic analyses, asstrongly advocated by Bachmann (2000). Thisapproach was followed in the study on Card-amine species traditionally treated within threedifferent polyploid complexes of mainly Euro-pean distribution, by including only diploids inthe analyses (Marhold et al. 2004). The poly-ploid complexes of Cardamine amara, C. raph-anifolia and C. pratensis have always been atleast implicitly considered to represent coher-ent and monophyletic groups. To investigatephylogenetic relationships among them, twoindependent molecular data sets were used,nrDNA ITS sequences and AFLP markers.The phylogenetic trees obtained were largelycongruent, and revealed two main lineages.While the C. amara group was resolved as awell-defined monophyletic group, neither ofthe markers supported current taxonomicseparation of remaining diploids into twogroups (the C. pratensis and C. raphanifoliagroups). Instead, all these taxa formed a singleclade with poorly resolved relationships. It washypothesized that either hybridization andintrogression among the taxa obscured geneticdifferentiation between the two groups, or thattraditional taxonomic treatment is incorrectand the taxa form a single monophyletic groupwith a common ancestor. In addition, twomorphologically distinct Caucasian diploids(C. tenera and C. uliginosa) which are currentlyallopatric and ecologically differentiated andthus do not come into contact, displayed tracesof past hybridization events. They both pos-sessed intraindividual ITS polymorphisms,and in both the ITS and AFLP trees theywere found intermingled. It has been assumedthat glacial-induced migrations may havebrought them together, and lack of reproduc-tion barriers allowed them to hybridize (Mar-hold et al. 2004).
The lack of supported hierarchical struc-ture, together with the evidence for ancienthybridization and introgression amongCaucasian diploids in that study, indicated
K. Marhold and J. Lihova: Polyploidy, hybridization and reticulate evolution 165
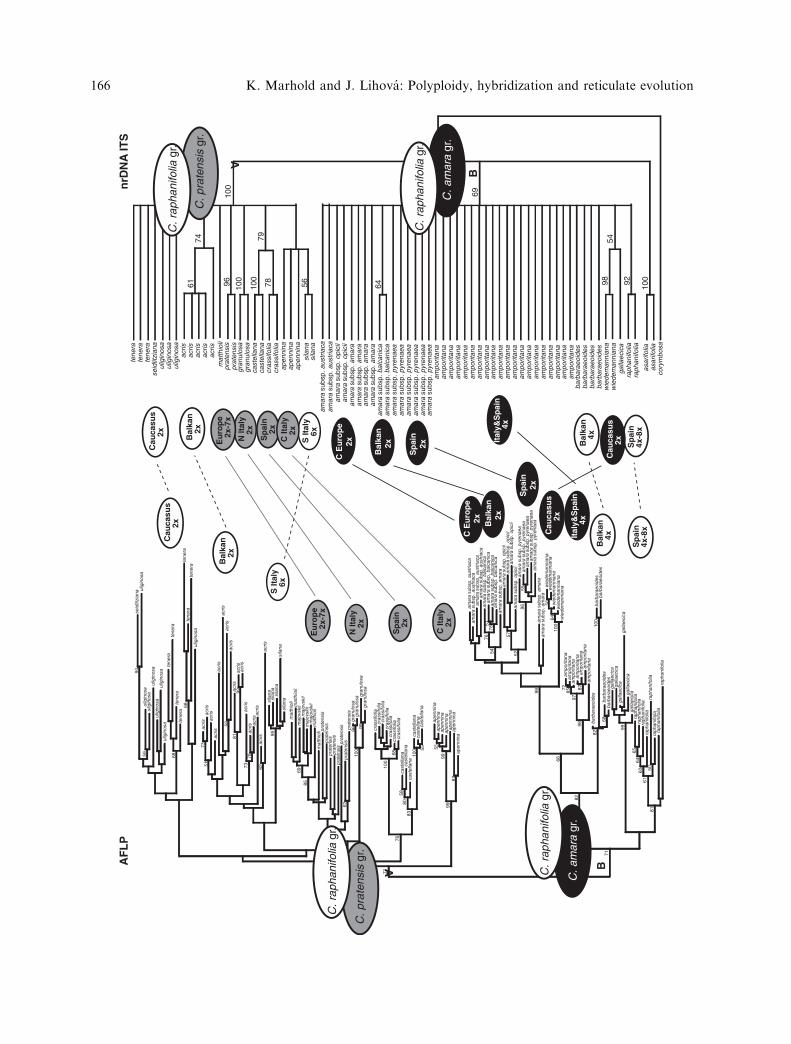
mat
thio
lim
atth
ioli
mat
thio
lim
atth
ioli
maj
ovsk
iim
ajov
skii
maj
ovsk
iim
ajov
skii
prat
ensi
spr
aten
sis
prat
ensi
spr
aten
sis
prat
ensi
spr
aten
sis
prat
ensi
spr
aten
sis
gran
ulos
agr
anul
osa gr
anul
osa
gran
ulos
a
cras
sifo
liacr
assi
folia
cras
sifo
liacr
assi
folia
cras
sifo
liacr
assi
folia
cras
sifo
liaca
stel
lana
cast
ella
naca
stel
lana
cast
ella
naca
stel
lana
cast
ella
na
apen
nina
apen
nina
apen
nina
apen
nina
apen
nina
apen
nina
apen
nina
amar
a su
bsp.
aus
tria
caam
ara
subs
p. a
ustr
iaca
amar
a su
bsp.
aus
tria
caam
ara
subs
p. a
ustr
iaca
amar
a su
bsp.
bal
cani
caam
ara
subs
p. b
alca
nica
amar
a su
bsp.
bal
cani
caam
ara
subs
p. b
alca
nica
amar
a su
bsp.
am
ara
amar
a su
bsp.
opi
cii
amar
a su
bsp.
opi
cii
amar
a su
bsp.
opi
cii
amar
a su
bsp.
opi
cii
amar
a su
bsp.
pyr
enae
aam
ara
subs
p. p
yren
aea
amar
a su
bsp.
pyr
enae
aam
ara
subs
p. p
yren
aea
amar
a su
bsp.
pyr
enae
aam
ara
subs
p. a
mar
aam
ara
subs
p. a
mar
aw
iede
man
nian
aw
iede
man
nian
aw
iede
man
nian
aw
iede
man
nian
aw
iede
man
nian
aam
porit
ana
ampo
ritan
aam
porit
ana
ampo
ritan
aam
porit
ana
ampo
ritan
a
barb
arae
oide
sba
rbar
aeoi
des
barb
arae
oide
sba
rbar
aeoi
des
barb
arae
oide
sga
llaec
ica
galla
ecic
aga
llaec
ica
galla
ecic
aga
llaec
ica
galla
ecic
ara
phan
ifolia
raph
anifo
liara
phan
ifolia
raph
anifo
liara
phan
ifolia
raph
anifo
liara
phan
ifolia
raph
anifo
lia
6985
62
96
9510
0
100
6876
8380
58
100 55
9999
87
55
71
57
5296
55
98
100
6456
7678
5154
98
ampo
ritan
a
9785
8772
82
66
82
100
6871
99
8150
6564
6061
acris
acris
acris
acris
acris
acris
acris
acris
acris
acris
acris
acris
acris ac
risac
risac
risac
rissi
lana
sila
nasi
lana
sila
nasi
lana
7351
62
8199
7378
59
99
seid
litzi
ana ul
igin
osa
ulig
inos
aul
igin
osa
ulig
inos
aul
igin
osa
ulig
inos
aul
igin
osa
tene
ra
ulig
inos
a
tene
rate
nera
tene
ra
tene
rate
nera
tene
ra
94
68
68
59
acris
ac
risac
risac
risac
ris
tene
rate
nera
tene
rase
idlit
zian
aul
igin
osa
ulig
inos
aul
igin
osa
amar
a su
bsp.
opi
cii
amar
a su
bsp.
opi
cii
amar
a su
bsp.
am
ara
amar
a su
bsp.
am
ara
amar
a su
bsp.
am
ara
amar
a su
bsp.
am
ara
amar
a su
bsp.
bal
cani
caam
ara
subs
p. b
alca
nica
amar
a su
bsp.
pyr
enae
aam
ara
subs
p. p
yren
aea
amar
a su
bsp.
pyr
enae
aam
ara
subs
p. p
yren
aea
amar
a su
bsp.
pyr
enae
aam
ara
subs
p. p
yren
aea
ampo
ritan
aam
porit
ana
ampo
ritan
aam
porit
ana
ampo
ritan
aam
porit
ana
ampo
ritan
aam
porit
ana
ampo
ritan
aam
porit
ana
ampo
ritan
aam
porit
ana
ampo
ritan
aam
porit
ana
ampo
ritan
aam
porit
ana
ampo
ritan
aam
porit
ana
ampo
ritan
aam
porit
ana
barb
arae
oide
sba
rbar
aeoi
des
barb
arae
oide
sba
rbar
aeoi
des
asar
ifolia
asar
ifolia
cory
mbo
sa
100
69B
64
100
A
7461
mat
thio
lipr
aten
sis
prat
ensi
s
apen
nina
apen
nina
apen
nina
sila
nasi
lana
96 56
gran
ulos
agr
anul
osa
100
cast
ella
naca
stel
lana
cras
sifo
liacr
assi
folia
100
7879
amar
a su
bsp.
aus
tria
caam
ara
subs
p. a
ustr
iaca
raph
anifo
liara
phan
ifolia
92
wie
dem
anni
ana
wie
dem
anni
ana
galla
ecic
a
5498
B
A
Sp
ain
2x
Ital
y&S
pai
n
4xBal
kan
2x
Bal
kan
4x
Sp
ain
4x-8
xS
pai
n4x
-8x
Bal
kan
4x
Cau
casu
s
2x
Cau
casu
s
2x
AF
LP
nrD
NA
ITS
Ital
y&S
pai
n
4x
Sp
ain
2x
Sp
ain
2x
Bal
kan
2x
C E
uro
pe
2x
C E
uro
pe
2xS It
aly
6x
S It
aly
6x
C It
aly
2x
Sp
ain
2x
C It
aly
2x
N It
aly
2x
N It
aly
2x
Eu
rop
e
2x-7
x
Eu
rop
e
2x-7
x
Cau
casu
s
2x
Cau
casu
s
2x
Bal
kan
2x
C. a
mar
a gr
.
C. a
mar
a gr
.
C. r
apha
nifo
lia g
r.
C. p
rate
nsis
gr.
C. r
apha
nifo
lia g
r.
C. p
rate
nsis
gr.
C. r
apha
nifo
lia g
r.
C. r
apha
nifo
lia g
r.
Bal
kan
2x
166 K. Marhold and J. Lihova: Polyploidy, hybridization and reticulate evolution

extensive past reticulate evolution even at thediploid level (Marhold et al. 2004). Neverthe-less, we were interested to investigate geneticaffinities of polyploid representatives of thecomplex to the respective diploids. Afterpolyploids were included (see Fig. 3), theresolved phylogenetic pattern was largelyretained. The distinction between the two mainlineages remained, but interestingly, polyploidsfrom the C. raphanifolia group were splitbetween the two clades. Iberian and Balkanpolyploids showed high affinity to the C. amararepresentatives, a pattern that would indicatetheir progenitor(s) in the C. amara group,whereas south Italian polyploid C. silana (asdiscussed in more detail above) apparentlyoriginated within the C. pratensis + C. raph-anifolia clade (Fig. 3). The traditionally recog-nized polyploid complexes evidently do notrepresent isolated lineages with their ownevolutionary history, as putative parents of atleast some polyploid taxa are likely to be foundin more than one of these groups. Althoughtwo very different markers were used (ITSsequences known to undergo concerted evolu-tion, and AFLP markers representing poly-morphisms across the major part of thegenome), results of cladistic and distance-based analyses of respective data sets weremostly congruent and revealed a similarpattern.
In the Brassicaceae, which comprises manygenera with polyploid and hybridogenousspecies, phylogenetic signatures of hybridiza-tion or hybrid speciation such as polytomiesand character conflict are expected to be foundrather frequently, but much caution is neededto distinguish them from population geneticprocesses (Linder and Rieseberg 2004). To
reconstruct reticulate relationships resultingfrom either homoploid or allopolyploid hybridspeciation more efficiently, network recon-struction methods and approaches have oftenproven useful. Numerous methods have beendiscussed by Vriesendorp and Bakker (2005)that are powerful in detecting character con-flict or in depicting alternative evolutionaryscenaria. Statistical parsimony (applied e.g. inthe Boechera study, Dobes et al. 2004b) andsplit decomposition (see the Yinshania study byKoch and Al-Shehbaz 2000) have been mostoften employed, but several other algorithmsand software are available as well. Phyloge-netic network reconstruction is, however, stillat an early stage of development and we mayexpect that future studies will significantlycontribute to this challenging issue (Linderand Rieseberg 2004).
This work was supported by the ScientificGrant Agency of the Ministry of Education of theSlovak Republic and the Slovak Academy ofSciences, VEGA, Bratislava (grant no. 3042 to K.M.), by the Alexander von Humboldt foundation(research fellowship to J. L.), by the GermanAcademic Exchange Service DAAD (project 323-PPP to K. M.), and by the Ministry of Education,Youth and Sports of the Czech Republic (grant no.0021620828, to K. M.). The authors are grateful toSuzanne Warwick and the editors of the presentissue for useful comments on the manuscript.
References
Adams K. L., Wendel J. F. (2004) Exploring thegenomic mysteries of polyploidy in cotton. Biol.J. Linn. Soc. 82: 573–581.
Ainouche M. L., Baumel A., Salmon A. (2004)Spartina anglica C. E. Hubbard: a natural modelsystem for analyzing early evolutionary changes
Fig. 3. Relationships among taxa from three traditionally recognized polyploid complexes of mainly Europeandistribution, the Cardamine amara, C. raphanifolia and C. pratensis groups. Neighbour-joining tree of AFLPdata (left) and strict-consensus tree of parsimony analysis of nrDNA ITS sequence data (right) are presented.White, grey and black colours show assignments to the complexes, with the indication of ploidy levels anddistribution areas. Bootstrap values are shown above the branches. The nrDNA ITS tree is based on data fromLihova et al. (2004a); the AFLP tree on re-analyses of separate data sets published in Lihova et al. (2003),Lihova et al. (2004b) and Perny et al. (2005b)
b
K. Marhold and J. Lihova: Polyploidy, hybridization and reticulate evolution 167

that affect allopolyploid genomes. Biol. J. Linn.Soc. 82: 475–484.
Ali H. B. M., Lysak M. A., Schubert I. (2004)Genomic in situ hybridization in plants withsmall genomes is feasible and elucidates thechromosomal parentage in interspecific Arabid-opsis hybrids. Genome 47: 954–960.
Al-Shehbaz I. A. (1984) The tribes of Cruciferae(Brassicaceae) in the southeastern United States.J. Arnold Arbor. 65: 343–373.
Al-Shehbaz I. A. (1988) The genera of Arabideae(Cruciferae; Brassicaceae) in the southeasternUnited States. J. Arnold Arbor. 69: 85–166.
Al-Shehbaz I. A. (2003) Transfer of most NorthAmerican species of Arabis to Boechera (Brass-icaceae). Novon 13: 381–391.
Al-Shehbaz I. A., Beilstein M. A. (2006) System-atics and phylogeny of the Brassicaceae (Cru-ciferae): an overview. Pl. Syst. Evol. 259: 89–120.
Al-Shehbaz I. A., O’Kane S. L., Price R. A. (1999)Generic placement of species excluded fromArabidopsis (Brassicaceae). Novon 9: 296–307.
Alvarez I., Wendel J. F. (2003) Ribosomal ITSsequences and plant phylogenetic inference.Molec. Phylogenet. Evol. 29: 417–434.
Arnold M. L. (1997) Natural hybridization andevolution. Oxford University Press, New York.
Axelsson T., Bowman C. M., Sharpe A. G.,Lydiate D. J., Lagercrantz U. (2000) Amphidip-loid Brassica juncea contains conserved progen-itor genomes. Genome 43: 679–688.
Bachmann K. (2000) Molecules, morphology andmaps: New directions in evolutionary genetics.Pl. Spec. Biol. 15: 197– 210.
Bennett M. D., Bhandol P., Leitch I. J. (2000)Nuclear DNA amounts in angiosperms and theirmodern uses – 807 new estimates. Ann. Bot.(Oxford) 86: 859–909.
Bennett M. D., Leitch I. J. (2005a) Plant DNA C-values database (release 4.0, October 2005).htttp://www.rbgkew.org.uk/cval/home-page.html.
Bennett M. D., Leitch I. J. (2005b) Plant genomesize research: a field in focus. Ann. Bot. (Oxford)95: 1–6.
Bleeker W. (2003) Hybridization and Rorippaaustriaca (Brassicaceae) invasion in Germany.Molec. Ecol. 12: 1831–1841.
Bleeker W. (2004) Genetic variation and self-incompatibility within and outside a Rorippa
hybrid zone (Brassicaceae). Pl. Syst. Evol. 246:35–44.
Bleeker W., Hurka H. (2001) Introgressive hybrid-ization in Rorippa (Brassicaceae): gene flow andits consequences in natural and anthropogenichabitats. Molec. Ecol. 10: 2013–2022.
Bleeker W., Huthmann M., Hurka H. (1999)Evolution of hybrid taxa in Nasturtium R. Br.(Brassicaceae). Folia Geobot. 34: 421–433.
Bleeker W., Matthies A. (2005) Hybrid zonesbetween invasive Rorippa austriaca and nativeR. sylvestris (Brassicaceae) in Germany: ploidylevels and patterns of fitness in the field. Heredity94: 664–670.
Boivin K., Acarkan A., Mbulu R.-S., Clarenz O.,Schmidt R. (2004) The Arabidopsis genomesequence as a tool for genome analysis inBrassicaceae. A comparison of the Arabidopsisand Capsella rubella genomes. Pl. Physiol. (Lan-caster) 135: 735–744.
Bowman J. L., Bruggemann H., Lee J.-Y., Mum-menhoff K. (1999) Evolutionary changes in floralstructure within Lepidium L. (Brassicaceae). Int.J. Pl. Sci. 160: 917–929.
Brochmann C. (1993) Reproductive strategies ofdiploid and polyploid populations of arcticDraba (Brassicaceae). Pl. Syst. Evol. 185: 55–83.
Brochmann C., Borgen L., Stedje B. (1993) Cross-ing relationships and chromosome numbers ofNordic populations of Draba (Brassicaceae),with emphasis on the D. alpina complex. Nord.J. Bot. 13: 121–147.
Brochmann C., Brysting A. K., Alsos I. G., BorgenL., Grundt H. H., Scheen A.-C., Elven R. (2004)Polyploidy in arctic plants. Biol. J. Linn. Soc. 82:521–536.
Brochmann C., Soltis P. S., Soltis D. E. (1992a)Recurrent formation and polyphyly of nordicpolyploids in Draba (Brassicaceae). Amer. J.Bot. 79: 673–688.
Brochmann C., Soltis P. S., Soltis D. E. (1992b)Multiple origins of the octoploid Scandinavianendemic Draba cacuminum: electrophoretic andmorphological evidence. Nord. J. Bot. 12: 257–272.
Brochmann C., Soltis D. E., Soltis P. S. (1992c)Electrophoretic relationships and phylogeny ofNordic polyploids in Draba (Brassicaceae). Pl.Syst. Evol. 182: 35–70.
168 K. Marhold and J. Lihova: Polyploidy, hybridization and reticulate evolution

Brochmann C., Stedje B., Borgen L. (1992d) Geneflow across ploidal levels in Draba (Brassica-ceae). Evol. Trends Pl. 6: 125–134.
Chevre A.-M., Ammitzbøll H., Breckling B., Dietz-Pfeilstetter A., Eber F., Fargue A., Gomez-Campo C., Jenczewski E., Jørgensen R., LavigneC., Meier M., den Nijs H. C. M., Pascher K.,Seguin-Swartz G., Sweet J., Stewart Jr. C. N.,Warwick S. (2004) A review on interspecific geneflow from oilseed rape to wild relatives. In: denNijs H. C. M., Bartsch D., Sweet J. (eds.)Introgression from genetically modified plantsinto wild relatives. CABI Publishing, Walling-ford, pp. 235–251.
Comai L., Tyagi A. P., Lysak M. A. (2003) FISHanalysis of meiosis in Arabidopsis allopolyploids.Chromosome Res. 11: 217–226.
Comai L., Tyagi A. P., Winter K., Holmes-DavisR., Reynolds S. H., Stevens Y., Byers, B. (2000)Phenotypic instability and rapid gene silencing innewly formed Arabidopsis tetraploids. Pl. Cell12: 1551–1567.
Darmency H., Lefol E., Fleury A. (1998) Sponta-neous hybridizations between oilseed rape andwild radish. Molec. Ecol. 7: 1467–1473.
Dart S., Kron P., Mable B. K. (2004) Character-izing polyploidy in Arabidopsis lyrata usingchromosome counts and flow cytometry. Canad.J. Bot. 82: 185–197.
Dobes C., Mitchell-Olds T., Koch M. A. (2004a)Intraspecific diversification in North AmericanBoechera stricta (= Arabis drummondii), Boech-era · divaricarpa, and Boechera holboellii (Brass-icaceae) inferred from nuclear and chloroplastmolecular markers – an integrative approach.Amer. J. Bot. 91: 2087–2101.
Dobes C., Mitchell-Olds T., Koch, M. A. (2004b)Extensive chloroplast haplotype variation indi-cates Pleistocene hybridization and radiation ofNorth American Arabis drummondii, A. · divari-carpa, and A. holboellii (Brassicaceae). Molec.Ecol. 13: 349–370.
Dolezel J., Bartos J. (2005) Plant DNA flowcytometry and estimation of nuclear genomesize. Ann. Bot. (Oxford) 95: 99–110.
Doyle J. J. (1992) Gene trees and species trees:molecular systematics as one-character taxon-omy. Syst. Bot. 17: 144–163.
Doyle J. J., Doyle J. L., Rauscher J. T., BrownA. H.D. (2004) Evolution of the perennial soybean
polyploid complex (Glycine subgenus Glycine): astudy of contrasts. Biol. J. Linn. Soc. 82: 583–597.
Easterly N. W. (1963) Chromosome numbers ofsome northwestern Ohio Cruciferae. Castanea28: 39–42.
Ellstrand N. C., Prentice H. C., Hancock J. F.(1999) Gene flow and introgression from domes-ticated plants into their wild relatives. AnnualRev. Ecol. Syst. 30: 539–563.
Ellstrand N. C., Schierenbeck K. A. (2000) Hybrid-ization as a stimulus for the evolution ofinvasiveness in plants? Proc. Natl. Acad. Sci.U.S.A. 97: 7043–7050.
Eschmann-Grupe G., Hurka H., Neuffer B. (2003)Species relationships within Diplotaxis (Brassica-ceae) and the phylogenetic origin of D. muralis.Pl. Syst. Evol. 243:13–29.
Eschmann-Grupe G., Hurka H., Neuffer B. (2004)Extent and structure of genetic variation in twoDiplotaxis specis (Brassicaceae) with contrastingbreeding systems. Pl. Syst. Evol. 244: 31–43.
Fineschi S., Taurchini D., Grossoni P., Petit R. J.,Vendramin G. G. (2002) Chloroplast DNAvariation of white oaks in Italy. Forest Ecol.Managem. 156: 103–114.
Franzke A., Hurka H. (2000) Molecular systemat-ics and biogeography of the Cardamine pratensiscomplex (Brassicaceae). Pl. Syst. Evol. 224: 213–234.
Franzke A., Mummenhoff K. (1999) Recent hybridspeciation in Cardamine (Brassicaceae) – con-version of nuclear ribosomal ITS sequences instatu nascendi. Theor. Appl. Genet. 98: 831–834.
Gomez-Campo C. (1999) Taxonomy. In: Gomez-Campo C. (ed.) Biology of Brassica coenospe-cies. Elsevier Science, Amsterdam, pp. 3–32.
Gross B. L., Rieseberg L. H. (2005) The ecologicalgenetics of homoploid hybrid speciation. J.Heredity 96: 241–252.
Grundt H. H., Obermayer R., Borgen L. (2005)Ploidal levels in the arctic-alpine polyploidDraba lactea (Brassicaceae) and its low-ploidrelatives. Bot. J. Linn. Soc. 147: 333–347.
GrundtH.H., PoppM., BrochmannC., OxelmanB.(2004) Polyploid origins in a circumpolar complexin Draba (Brassicaceae) inferred from clonednuclear sequences and fingerprints.Molec. Phylo-genet. Evol. 32: 695–710.
Hall A. E., Fiebig A., Preuss D. (2002) Beyond theArabidopsis genome: opportunities for compar-
K. Marhold and J. Lihova: Polyploidy, hybridization and reticulate evolution 169

ative genomics. Pl. Physiol. (Lancaster) 129:1439–1447.
Harberd D. J., McArthur E. E. (1972) The chro-mosome constitution of Diplotaxis muralis (L.)DC. Watsonia 9: 131–135.
Hegarty M. J., Hiscock S. J. (2005) Hybridspeciation in plants: new insights from molecularstudies. New Phytol. 165: 411–423.
Hewitt G. M. (2004) Genetic consequences ofclimatic oscillations in the Quaternary. Philos.Trans., Ser. B 359: 183–195.
Hoffmann M. H. (2005) Evolution of the realizedclimatic niche in the genus Arabidopsis (Brassic-aceae). Evolution 59: 1425–1436.
Howard H. W., Manton I. (1946) Autopolyploidand allopolyploid watercress with the descriptionof a new species. Ann. Bot. (London) 10: 1–13.
Hurka H., Freundner S., Brown A. H. D., Plan-tholt U. (1989) Aspartate aminotransferase iso-zymes in the genus Capsella (Brassicaceae):subcellular location, gene duplication, and poly-morphism. Biochem. Genet. 27: 77–90.
Hurka H., Neuffer B. (1997) Evolutionary pro-cesses in the genus Capsella (Brassicaceae). Pl.Syst. Evol. 206: 295–316.
Hylander N. (1957) Cardaminopsis suecica (Fr.)Hiit., a northern amphidiploid species. Bull.Jard. Bot. Etat 27: 591–604.
Johnston J. S., Pepper A. E., Hall A. E., Chen Z. J.,Hodnett G., Drabek J., Lopez R., Price J. (2005)Evolution of genome size in Brassicaceae. Ann.Bot. (Oxford) 95: 229–235.
Jonsell B. (1968) Studies in the north-west Euro-pean species of Rorippa s. str. Symb. Bot. Upsal.19: 1–121.
Kashkush K., Feldman M., Levy A. A. (2002)Gene loss, silencing and activation in a newlysynthesized wheat allotetraploid. Genetics 160:1651–1659.
Kellogg E. A., Bennetzen J. L. (2004) The evolutionof nuclear genome structure in seed plants.Amer. J. Bot. 91: 1709–1725.
Koch M. (2002) Genetic differentiation and speci-ation in prealpine Cochlearia: allohexaploidCochlearia bavarica (Brassicaceae) compared toits diploid ancestor Cochlearia pyrenaica inGermany and Austria. Pl. Syst. Evol. 232: 35–49.
Koch M., Al-Shehbaz I. A. (2000) Molecularsystematics of the Chinese Yinshania (Brassica-ceae): evidence from plastid and nuclear ITS
sequence data. Ann. Missouri Bot. Gard. 87:246–272.
Koch M., Al-Shehbaz I. A. (2002) Molecular dataindicate complex intra- and intercontinentaldifferentiation of American Draba (Brassica-ceae). Ann. Missouri Bot. Gard. 89: 88–109.
Koch M., Al-Shehbaz I. A., Mummenhoff K.(2003a) Molecular systematics, evolution andpopulation biology in the mustard family (Brass-icaceae). Ann. Missouri Bot. Gard. 90: 151–171.
Koch M., Bishop J., Mitchell-Olds T. (1999)Molecular systematics and evolution of Arabid-opsis and Arabis. Pl. Biol. 1: 529–537.
Koch M., Bernhardt K.-G. (2004) Comparativebiogeography of the cytotypes of annual Mic-rothlaspi perfoliatum (Brassicaceae) in Europeusing isozymes and cpDNA data: refugia, diver-sity centers, and postglacial colonization. Amer.J. Bot. 91: 115–124.
Koch M. A., Dobes C., Mitchell-Olds T. (2003b)Multiple hybrid formation in natural popula-tions: concerted evolution of the internal tran-scribed spacer of nuclear ribosomal DNA (ITS)in North American Arabis divaricarpa (Brassic-aceae). Molec. Biol. Evol. 20: 338–350.
Koch M., Haubold B., Mitchell-Olds T. (2000)Comparative evolutionary analysis of chalconesynthase and alcohol dehydrogenase loci inArabidopsis, Arabis and related genera (Brassi-caceae). Molec. Biol. Evol. 17: 1483–1498.
Koch M., Haubold B., Mitchell-Olds T. (2001)Molecular systematics of the Brassicaceae: evi-dence from coding plastidic matK and nuclearChs sequences. Amer. J. Bot. 88: 534–544.
Koch M., Hurka H. (1999) Isozyme analysis in thepolyploid complex Microthlaspi perfoliatum (L.)F. K. Meyer: morphology, biogeography andevolutionary history. Flora 194: 33–48.
Koch M., Hurka H., Mummenhoff K. (1996)Chloroplast DNA restriction site variation andRAPD-analyses in Cochlearia (Brassicaceae):Biosystematics and speciation. Nordic J. Bot.16: 585–603.
Koch M., Huthmann M., Hurka H. (1998a)Isozymes, speciation and evolution in the poly-ploid Cochlearia L. (Brassicaceae). Bot. Acta111: 411–425.
Koch M., Kiefer C. (2006) Molecules and migra-tions: biogeographical studies in cruciferousplants. Pl. Syst. Evol. 259: 121–142.
170 K. Marhold and J. Lihova: Polyploidy, hybridization and reticulate evolution

Koch M., Kiefer M. (2005) Genome evolutionamong cruciferous plants – a lecture from thecomparison of the genetic maps of three diploidspecies: Capsella rubella, Arabidopsis lyrata ssp.petraea and Arabidopsis thaliana. Amer. J. Bot.92: 761–767.
Koch M., Mummenhoff K., Hurka H. (1998b)Molecular biogeography and evolution of theMicrothlaspi perfoliatum s.l. polyploid complex(Brassicaceae): chloroplast DNA and nuclearribosomal DNA restriction site variation. Ca-nad. J. Bot. 76: 382–396.
Kucera J., Valko I., Marhold K. (2005) On-linedatabase of the chromosome numbers of thegenus Cardamine (Brassicaceae). Biologia (Bra-tislava) 60: 473–476 (database available on-lineat http://147.213.100.117/cardamine/index.php).
Kuittinen H., de Haan A. A., Vogl C., OikarinenS., Leppala J., Koch M., Mitchell-Olds T.,Langley C., Savolainen O. (2004) Comparingthe linkage maps of the close relatives Arabid-opsis lyrata and A. thaliana. Genetics 168: 1575–1584.
Lagercrantz U. (1998) Comparative mappingbetween Arabidopsis thaliana and Brassica nigraindicates that Brassica genomes have evolvedthrough extensive genome replication accompa-nied by chromosomal fusions and frequentrearrangements. Genetics 150: 1217–1228.
Lan T. H., DelMonte T. A., Reischmann K.,Hyman J., Kowalski S. P., McFerson J., Kres-ovich S., Paterson A. H. (2000) An EST-enrichedcomparative map of Brassica oleracea and Ara-bidopsis thaliana. Genome Res. 10: 776–788.
Lee C. E. (2002) Evolutionary genetics of invasivespecies. Trends Ecol. Evol. 17: 386–391.
Lee J.-Y., Mummenhoff K., Bowman J. L. (2002)Allopolyploidization and evolution of specieswith reduced floral structures in Lepidium L.(Brassicaceae). Proc. Natl. Acad. Sci. U.S.A. 99:16835–16840.
Leitch I. J., Bennett M. D. (2004) Genome down-sizing in polyploid plants. Biol. J. Linn. Soc. 82:651–663.
Levin D. A. (2002) The role of chromosomalchange in plant evolution. Oxford UniversityPress, New York.
Levy A. A., Feldman M. (2004) Genetic andepigenetic reprogramming of the wheat genomeupon allopolyploidization. Biol. J. Linn. Soc. 82:607–613.
Lihova J., Fuertes Aguilar J., Marhold K., NietoFeliner G. (2004a) Origin of the disjunct tetra-ploid Cardamine amporitana (Brassicaceae) as-sessed with nuclear and chloroplast DNAsequence data. Amer. J. Bot. 91: 1231–1242.
Lihova J., Marhold K. (2003) Taxonomy anddistribution of the Cardamine pratensis group(Brassicaceae) in Slovenia. Phyton (Horn,Austria) 43: 241–261.
Lihova J., Marhold K. (2006) Phylogenetic anddiversity patterns inCardamine (Brassicaceae) – agenus with conspicuous polyploid and reticulateevolution. In: Sharma A. K., Sharma A. (eds.)Plant genome: biodiversity and evolution, vol. 1C:Phanerogams (Angiosperms – Dicotyledons).Science Publishers, Inc., Enfield, pp. 149–186.
Lihova J., Marhold K., Neuffer B. (2000) Taxon-omy of Cardamine amara (Brassicaceae) in theIberian Peninsula. Taxon 49: 747–763.
Lihova J., Marhold K., Tribsch A., Stuessy T. F.(2004b) Morphological and AFLP re-evaluationof tetraploid Cardamine amara (Brassicaceae) inthe Mediterranean. Syst. Bot. 29: 134–146.
Lihova J., Shimizu K. K., Marhold K. (2006)Allopolyploid origin of Cardamine asarifolia(Brassicaceae): incongruence between plastidand nuclear ribosomal DNA sequences solvedby a single copygene. Molec. Phylogenet. Evol.(in press).
Lihova J., Tribsch A., Marhold K. (2003) TheCardamine pratensis (Brassicaceae) group in theIberian Peninsula: taxonomy, polyploidy anddistribution. Taxon 52: 783–802.
Lim K. Y., Matyasek R., Kovarik A., Leitch A. R.(2004) Genome evolution in allotetraploid Nico-tiana. Biol. J. Linn. Soc. 82: 599–606.
Lind-Hallden C., Hallden C., Sall T. (2002) Geneticvariation in Arabidopsis suecica and its parentalspecies A. arenosa and A. thaliana. Hereditas136: 45–50.
Linder C. R., Rieseberg L. H. (2004) Reconstruct-ing patterns of reticulate evolution in plants.Amer. J. Bot. 91: 1700–1708.
Lysak M. A., Koch M. A., Pecinka A., Schubert I.(2005) Chromosome triplication found acrossthe tribe Brassiceae. Genome Res. 15: 516–525.
Lysak M. A., Lexer C. (2006) Towards the era ofcomparative evolutionary genomics in Brassica-ceae. Pl. Syst. Evol. 259: 175–198.
Lysak M. A., Pecinka A., Schubert I. (2003) Recentprogress in chromosome painting of Arabidopsis
K. Marhold and J. Lihova: Polyploidy, hybridization and reticulate evolution 171

and related genera. Chromosome Res. 11: 195–204.
Madlung A., Tyagi A. P., Watson B., Jiang H.,Kagochi T., Doerge R. W., Martienssen R.,Comai L. (2005) Genomic changes in syntheticArabidopsis polyploids. Pl. J. 41: 221–230.
Manton I. (1935) The cytological history of water-cress (Nasturtium officinale R. Br.). Z. Indukt.Abstammungs-Vererbungsl. 69: 132–157.
Marhold K. (1999) Taxonomic evaluation of thetetraploid populations of Cardamine amara(Brassicaceae) from the Eastern Alps and adja-cent areas. Bot. Helv. 109: 67–84.
Marhold K., Huthmann M., Hurka H. (2002a)Evolutionary history of the polyploid complex ofCardamine amara (Brassicaceae): isozyme evi-dence. Pl. Syst. Evol. 233: 15–28.
Marhold K., Lihova J., Perny M., Bleeker W.(2004) Comparative ITS and AFLP analysis ofdiploid Cardamine (Brassicaceae) taxa fromclosely related polyploid complexes. Ann. Bot.(Oxford) 93: 507–520.
Marhold K., Lihova J., Perny M., Grupe R.,Neuffer B. (2002b) Natural hybridization inCardamine (Brassicaceae) in the Pyrenees: evi-dence from morphological and molecular data.Bot. J. Linn. Soc. 139: 275–294.
Martın J. P., Sanchez-Yelamo M. D. (2000)Genetic relationships among species of the genusDiplotaxis (Brassicaceae) using inter-simple se-quence repeat markers. Theor. Appl. Genet. 101:1234–1241.
Miyashita N. T., Kawabe A., Innan H., TerauchiR. (1998) Intra- and interspecific DNA variationand codon bias of the alcohol dehydrogenase(Adh) locus in Arabis and Arabidopsis species.Molec. Biol. Evol. 15: 1420–1429.
Mort M. E., Crawford D. J. (2004) The continuingsearch: low-copy nuclear sequences for lower-level plant molecular phylogenetic studies. Tax-on 53: 257–261.
Mulligan G. A. (1995) Synopsis of the genus Arabis(Brassicaceae) in Canada, Alaska and Green-land. Rhodora 97: 109–163.
Mummenhoff K., Bruggemann H., Bowman J. L.(2001) Chloroplast DNA phylogeny and bioge-ography of Lepidium (Brassicaceae). Amer. J.Bot. 88: 2051–2063.
Mummenhoff K., Eschmann-Grupe G., Zunk K.(1993) Subunit polypeptide composition of RU-BISCO indicates Diplotaxis viminea as maternal
parent species of amphidiploid Diplotaxis mu-ralis. Phytochemistry 34: 429–431.
Mummenhoff, K., Franzke A. (2006) Gone with thebird: Late Tertiary and Quaternary interconti-nental long-distance dispersal and allopolyploi-dization in plants. Syst. Biodivers. 4 (in press).
Mummenhoff K., Franzke A., Koch M. (1997)Molecular phylogenetics of Thlaspi s.l. (Brassi-caceae) based on chloroplast DNA restrictionsite variation and sequences of the internaltranscribed spacers of nuclear ribosomal DNA.Canad. J. Bot. 75: 469–482.
Mummenhoff K., Hurka H. (1990) Evolution ofthe tetraploid Capsella bursa-pastoris (Brassica-ceae): isloelectric focusing analysis of Rubisco.Pl. Syst. Evol. 172: 205–213.
Mummenhoff K., Hurka H. (1994) Subunit poly-peptide composition of Rubisco and the originof allopolyploid Arabidopsis suecica (Brassica-ceae). Biochem. Syst. Ecol. 22: 807–812.
Mummenhoff K., Hurka H. (1995) Allopolyploidorigin of Arabidopsis suecica (Fries) Norrlin:evidence from chloroplast and nuclear genomemarkers. Bot. Acta 108: 449–456.
Mummenhoff K., Linder P., Friesen N., BowmanJ. L., Lee J.-Y., Franzke A. (2004) Molecularevidence for bicontinental hybridogenous geno-mic constitution in Lepidium sensu stricto(Brassicaceae) species from Australia and NewZealand. Amer. J. Bot. 91: 254–261.
Neuffer B., Jahncke P. (1997) RAPD analyses ofhybridization events in Cardamine (Brassica-ceae). Folia Geobot. Phytotax. 32: 57–67.
O’Kane S. L., Al-Shehbaz I. A. (1997) A synopsisof Arabidopsis (Brassicaceae). Novon 7: 323–327.
O’Kane S. R. J., Schaal B. A., Al-Shehbaz I. A.(1996) The origins of Arabidopsis suecica (Brass-icaceae) as indicated by nuclear rDNA se-quences. Syst. Bot. 21: 559–566.
Osborn T. C. (2004) The contribution of poly-ploidy to variation in Brassica species. Physiol.Pl. (Copenhagen) 121: 531–536.
Osborn T. C., Pires J. C., Birchler J. A., Auger D.L., Chen Z. J., Lee H.-S., Comai L., MadlungA., Doerge R. W., Colot V., Martienssen R. A.(2003) Understanding mechanisms of novelexpression in polyploids. Trends Genet. 19:141–147.
Otto S. P., Whitton J. (2000) Polyploid incidenceand evolution. Annual Rev. Genet. 34: 401–437.
172 K. Marhold and J. Lihova: Polyploidy, hybridization and reticulate evolution

Palme A. E., Vendramin G. G. (2002) ChloroplastDNA variation, postglacial recolonization andhybridization in hazel, Corylus avellana. Molec.Ecol. 11: 1769–1779.
Perny M., Tribsch A., Stuessy T. F., Marhold K.(2005a) Allopolyploid origin of Cardamine silana(Brassicaceae) from Calabria (Southern Italy):karyological, morphological and molecular evi-dence. Bot. J. Linn. Soc. 148: 101–116.
Perny M., Tribsch A., Stuessy T., Marhold K.(2005b) Taxonomy and cytogeography of Card-amine raphanifoliaandC.gallaecica (Brassicaceae)in the IberianPeninsula. Pl. Syst. Evol. 254: 69–91.
Rausch J. H., Morgan M. T. (2005) The effect ofself-fertilization, inbreeding depression, andpopulation size on autopolyploid establishment.Evolution 59: 1867–1875.
Rieseberg L. H., Baird S. J. E., Gardner K. A.(2000) Hybridization, introgression, and linkageevolution. Pl. Molec. Biol. 42: 205–224.
Sall T., Lind-Hallden C., Jakobsson M., HalldenC. (2004) Mode of reproduction in Arabidopsissuecica. Hereditas 141: 313–317.
Savolainen O., Langley C. H., Lazzaro B. P.,Freville H. (2000) Contrasting patterns of nucle-otide polymorphism at the alcohol dehydroge-nase locus in the outcrossing Arabidopsis lyrataand the selfing Arabidopsis thaliana. Molec. Biol.Evol. 17: 645–655.
Schmidt R., Acarkan A., Boivin K. (2001) Com-parative structural genomics in the Brassicaceaefamily. Pl. Physiol. Biochem. (Montrouge) 39:253–262.
Schranz M. E., Dobes Ch., Koch M. A., Mitchell-Olds T. (2005) Sexual reproduction, hybridiza-tion, apomixis, and polyploidization in the genusBoechera (Brassicaceaea). Amer. J. Bot. 92:1797–1810.
Schranz M. E., Osborn T. C. (2004) De novovariation in life-history traits and responses togrowth conditions of resynthesized polyploidBrassica napus (Brassicaceae). Amer. J. Bot. 91:174–183.
Seehausen O. (2004) Hybridization and adaptiveradiation. Trends Ecol. Evol. 19: 198–207.
Sharbel T. F., Mitchell-Olds T. (2001) Recurrentpolyploid origins and chloroplast phylogeogra-phy in the Arabis holboellii complex (Brassica-ceae). Heredity 87: 59–68.
Sharbel T. F., Mitchell-Olds T., Dobes C., Kan-tama L., de Jong H. (2005) Biogeographic
distribution of polyploidy and B chromosomesin the apomictic Boechera holboellii complex.Cytogenet. Genome Res. 109: 283–292.
Shimizu K. K., Fujii S., Marhold K., Watanabe K.,Kudoh H. (2005) Arabidopsis kamchatica (Fisch.ex DC.) K. Shimizu & Kudoh and A. kamchaticasubsp. kawasakiana (Makino) K. Shimizu &Kudoh, new combinations. Acta Phytotax. Geo-bot. 56: 163–172.
Small R. L., Cronn R. C., Wendel J. F. (2004) Useof nuclear genes for phylogeny reconstruction inplants. Austral. Syst. Bot. 17: 145–170.
Soltis D. E., Soltis P. S. (1999) Polyploidy: recur-rent formation and genome evolution. TrendsEcol. Evol. 14: 348–352.
Soltis D. E., Soltis P., Pires J. Ch., Kovarik A.,Tate J. A., Mavrodiev E. (2004a) Recent andrecurrent polyploidy in Tragopogon (Astera-ceae): cytogenetic, genomic and genetic compar-isons. Biol. J. Linn. Soc. 82: 485–501.
Soltis D. E, Soltis P. S., Tate J. A. (2004b)Advances in the study of polyploidy since Plantspeciation. New Phytol. 161: 173–191.
Soltis P. S., Soltis D. E. (2000) The role of geneticand genomic attributes in the success of polyp-loids. Proc. Natl. Acad. Sci. U.S.A. 97: 7051–7057.
Song K., Lu P., Tang K., Osborn T. C. (1995)Rapid genome change in synthetic polyploids ofBrassica and its implication for polyploid evolu-tion. Proc. Natl. Acad. Sci. U.S.A. 92: 7719–7723.
Stace C. (ed.) (1975) Hybridization and the flora ofthe British Isles. Academic Press, London NewYork San Francisco.
Taberlet P., Fumagalli L., Wust-Saucy A. G.,Cosson J. F. (1998) Comparative phylogeogra-phy and postglacial colonization routes inEurope. Molec. Ecol. 7: 453–464.
Tomsovic P. (1969) Nejdulezitejsı vysledky revizeceskoslovenskych rukvı (Rorippa Scop. em. Rei-chenbach). Preslia 41: 21–38.
Tremetsberger K., Konig C., Samuel R., PinskerW., Stuessy T. F. (2002) Infraspecific geneticvariation in Biscutella laevigata (Brassicaceae):new focus on Irene Manton’s hypothesis. Pl.Syst. Evol. 233: 163–181.
Urbanska-Worytkiewicz K. (1977a) Reproductionin natural triploid hybrids (2n = 24) betweenCardamine rivularis Schur and C. amara L. Ber.Geobot. Inst. ETH Stiftung Rubel 44: 42–85.
K. Marhold and J. Lihova: Polyploidy, hybridization and reticulate evolution 173

Urbanska-Worytkiewicz K. (1977b) An autoallo-hexaploid in Cardamine L., new to the Swissflora. Ber. Geobot. Inst. ETH Stiftung Rubel 44:86–103.
Urbanska-Worytkiewicz K. (1980) Reproductivestrategies in a hybridogenous population ofCardamine L. Acta Oecol./Oecol. Pl. 1 (15):137–150.
Urbanska K. (1987) Disturbance, hybridizationand hybrid speciation. In: van Andel J. et al.(eds.) Disturbance in grasslands. Dr W. JunkPublishers, Dordrecht, pp. 285–301.
Urbanska K. M., Hurka H., Landolt E., NeufferB., Mummenhoff K. (1997) Hybridization andevolution in Cardamine (Brassicaceae) at Urn-erboden, Central Switzerland: biosystematic andmolecular evidence. Pl. Syst. Evol. 204: 233–256.
Urbanska-Worytkiewicz K., Landolt E. (1972)Naturliche Bastarde zwischen Cardamine amaraL. und C. rivularis Schur aus den SchweizerAlpen. Ber. Geobot. Inst. ETH. Stiftung Rubel41: 88–100.
Urbanska-Worytkiewicz K., Landolt E. (1974)Biosystematic investigations in Cardamine prat-ensis L. s.l. I. Diploid taxa from Central Europeand their fertility relationships. Ber. Geobot.Inst. ETH Stiftung Rubel 42: 42–139.
Vriesendorp B., Bakker F. T. (2005) Reconstruct-ing patterns of reticulate evolution in angio-sperms: what can we do? Taxon 54: 593–604.
Warwick S. I., Anderson J. K. (1997a) Isozymeanalysis of parentage in allopolyploid Diplotaxismuralis (L.) DC. (Brassicaceae). EucarpiaCruciferae Newslett. 19: 35–36.
Warwick S. I., Anderson J. K. (1997b) Isozymeanalysis of parentage in allopolyploid Eruca-strum gallicum (L.) DC. (Brassicaceae). EucarpiaCruciferae Newslett. 19: 37–38.
Warwick S. I., Al-Shehbaz I. A. (2006) Brassica-ceae: chromosome number index and databaseon CD-Rom. Pl. Syst. Evol. 259: 237–248.
Warwick S. I., Al-Shehbaz I. A., Suader C., HarrisJ. G., Koch M. (2004) Phylogeny of Braya andNeotorularia (Brassicaceae) based on nuclearribosomal internal transcribed spacer and chlo-roplast trnL intron sequences. Canad. J. Bot. 82:376–392.
Warwick S. I., Black L. D., Aguinagalde I. (1992)Molecular systematics of Brassica and alliedgenera (Subtribe Brassicinae, Brassiceae) – Chlo-roplast DNA variation in the genus Diplotaxis.Theor. Appl. Genet. 83: 839–850.
Warwick S. I., Simard M.-J., Legere A., Beckie H.J., Braun L., Zhu B., Mason P., Seguin-SwartzG., Stewart C. N. (2003) Hybridization betweentransgenic Brassica napus L. and its wild rela-tives: Brassica rapa L., Raphanus raphanistrumL., Sinapis arvensis L., and Erucastrum gallicum(Willd.) O. E. Schulz. Theor. Appl. Genet. 107:528–539.
Wendel J. F. (2000) Genome evolution in polyp-loids. Pl. Molec. Biol. 42: 225–249.
Wendel J. F., CronnR. C., Johnston J. S., Price H. J.(2002) Feast and famine in plant genomes. Gene-tica 115: 37–47.
Widmer A., Baltisberger M. (1999) Molecularevidence for allopolyploid speciation and asingle origin of the narrow endemic Draba ladina(Brassicaceae). Amer. J. Bot. 86: 1282–1289.
Addresses of the authors: Karol Marhold(e-mail: [email protected]) Institute of Bot-any, Slovak Academy of Sciences, Dubravska cesta14, 845 23 Bratislava, Slovak Republic; Departmentof Botany, Charles University, Benatska 2, 128 01Praha, Czech Republic; Judita Lihova (e-mail:[email protected]), Institute of Botany, SlovakAcademy of Sciences, Dubravska cesta 14, 845 23Bratislava, Slovak Republic.
174 K. Marhold and J. Lihova: Polyploidy, hybridization and reticulate evolution