plant stem cell niches
TRANSCRIPT

PP63CH25-Laux ARI 27 March 2012 11:48
Plant Stem Cell NichesErnst Aichinger,1,2 Noortje Kornet,1
Thomas Friedrich,1 and Thomas Laux1,2
1BIOSS Centre for Biological Signalling Studies, Faculty of Biology, University of Freiburg,79104 Freiburg, Germany; email: [email protected] Institute of Advanced Studies (FRIAS), D-79104 Freiburg, Germany
Annu. Rev. Plant Biol. 2012. 63:615–36
First published online as a Review in Advance onFebruary 9, 2012
The Annual Review of Plant Biology is online atplant.annualreviews.org
This article’s doi:10.1146/annurev-arplant-042811-105555
Copyright c© 2012 by Annual Reviews.All rights reserved
1543-5008/12/0602-0615$20.00
Keywords
cambium, organizing center, quiescent center, boundary formation,signaling
Abstract
Multicellular organisms possess pluripotent stem cells to form new or-gans, replenish the daily loss of cells, or regenerate organs after in-jury. Stem cells are maintained in specific environments, the stem cellniches, that provide signals to block differentiation. In plants, stem cellniches are situated in the shoot, root, and vascular meristems—self-perpetuating units of organ formation. Plants’ lifelong activity—which,as in the case of trees, can extend over more than a thousand years—requires that a robust regulatory network keep the balance betweenpluripotent stem cells and differentiating descendants. In this review,we focus on current models in plant stem cell research elaborated duringthe past two decades, mainly in the model plant Arabidopsis thaliana. Weaddress the roles of mobile signals on transcriptional modules involvedin balancing cell fates. In addition, we discuss shared features of anddifferences between the distinct stem cell niches of Arabidopsis.
615
Ann
u. R
ev. P
lant
Bio
l. 20
12.6
3:61
5-63
6. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by U
nive
rsid
ad A
uton
oma
de Q
uere
taro
on
10/2
4/13
. For
per
sona
l use
onl
y.

PP63CH25-Laux ARI 27 March 2012 11:48
Contents
INTRODUCTION . . . . . . . . . . . . . . . . . . 616STRUCTURE AND FUNCTION
OF THE SHOOT APICALMERISTEM . . . . . . . . . . . . . . . . . . . . . . 617Regulation of the Cytokinin/
Gibberellic Acid Balance byKNOX Genes . . . . . . . . . . . . . . . . . . . 617
Local Control of Stem Cells byWUS . . . . . . . . . . . . . . . . . . . . . . . . . . 619
ZLL/AGO1 Balance DeterminesStem Cell Inhibitory MicroRNALevels . . . . . . . . . . . . . . . . . . . . . . . . . . 619
Control of the Stem Cell Number . . 620What Keeps the Stem Cell Niche at
the Tip of the Shoot? . . . . . . . . . . . 622THE ROOT STEM CELL NICHE . . 622
The Quiescent Center Functions asOrganizer of the Root Stem CellNiche . . . . . . . . . . . . . . . . . . . . . . . . . . 624
WOX5 Expression in the QuiescentCenter Is Required to MaintainStem Cells . . . . . . . . . . . . . . . . . . . . . . 624
The Stele-Borne SHR Signal IsRequired for Quiescent CenterFunction . . . . . . . . . . . . . . . . . . . . . . . 625
Long-Range Control of the StemCell Niche via Auxin and PLTGenes . . . . . . . . . . . . . . . . . . . . . . . . . . 625
Quiescence of the QuiescentCenter . . . . . . . . . . . . . . . . . . . . . . . . . 626
THE VASCULAR STEM CELLNICHE . . . . . . . . . . . . . . . . . . . . . . . . . . . 626Stem Cell Maintenance in the
Procambium. . . . . . . . . . . . . . . . . . . . 627More Boundaries Within the
Vascular Stem Cell Niche . . . . . . . 628Coordination of Longitudinal and
Lateral Growth . . . . . . . . . . . . . . . . . 629MOLECULAR SIGNATURE OF
PLANT STEM CELLS . . . . . . . . . . . 629
INTRODUCTION
Plants that are several hundred years old andyet generate whole new organs year after
year offer one of the most impressive examplesof plant developmental capacity. The origin ofeach new organ lies on the tip at each side ofthe plant body—the shoot apical meristem andthe root apical meristem. In addition, there aretwo meristems involved in the thickening of thestem, which are most prominent in trees: thering-shaped cambium of the vasculature, whichprovides transport and support, and the phel-logen/cork cambium, which replenishes theouter layer (bark) that is regularly shed. In allcases, meristems have two functions: to producenew cells and to initiate organ formation.
How new cells are produced has beendemonstrated by elegant cell-tracking experi-ments in the shoot meristem. In 1940, Satinaand coworkers (113) induced polyploidy in sin-gle cells of Datura through colchicine treatmentand found that the shoot meristem consistsof clonally separate layers of cells: an outerL1 layer from which the epidermis is derived,a subepidermal L2 layer, and an internal L3layer. This three-layer organization is typicalof the shoot meristems of dicotyledonous seedplants, but varies in monocotyledons (two lay-ers), gymnosperms (one layer), and more basalspecies (mosses, ferns) where all cells originatefrom a single apical cell. Thirty years later,Stewart & Dermen (125) provided elegantevidence that the cells in approximately one-third of the shoot circumference originatedfrom a single stem cell in the shoot meristem,indicating the presence of three stem cells inthis layer. Because each clone derived from asingle cell comprised all cell types in this layer,the shoot meristem stem cells can be viewedas pluripotent. Importantly, marked sectorsoccasionally broadened from one-third to two-thirds and even to the entire circumference atthe expense of the progeny of nonmarked stemcells and vice versa, indicating that stem cellscan be replaced and act as stem cells only aslong as they are in a specialized environment.Finally, when an L1 stem cell was displacedinto the subepidermal layer by an occasionalpericlinal division, it gave rise to L2 cell types,suggesting that the fate of its progeny is notdetermined by the stem cell’s history but rather
616 Aichinger et al.
Ann
u. R
ev. P
lant
Bio
l. 20
12.6
3:61
5-63
6. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by U
nive
rsid
ad A
uton
oma
de Q
uere
taro
on
10/2
4/13
. For
per
sona
l use
onl
y.

PP63CH25-Laux ARI 27 March 2012 11:48
by the position of the differentiating offspring.In 1965, Newman (98) had already describedshoot meristem stem cells as “temporary occu-pants of a permanent office”—a good metaphorfor what we call stem cell niches today.
How can plant stem cells exist for hun-dreds of years without accumulating mutations?That stem cells divide relatively infrequentlyhas been recognized by histological studies andhas been associated with the advantage of re-ducing the mutational load of DNA replica-tion (84). Immediate daughter cells of the stemcells might perform a finite number of divisionrounds before they are replaced, thus increas-ing the stem cell output as transit amplifyingcells (125). Fulcher & Sablowski (41) observedthat shoot and root meristem cells in Arabidopsisare hypersensitive to DNA double strand breaksand respond with cell death to DNA damage,allowing a quick clearing of compromised DNAfrom the stem cell system.
On the basis of these studies, together withstudies in animals that rely largely on test-ing stem cell potential in transplantation as-says (114), a general stem cell niche conceptfor multicellular organisms is apparent: (a) Cellsthat are protected from differentiation by sig-nals from neighboring cells in specific nichescan divide and thus function as stem cells, and(b) the cells that leave a niche are bound to dif-ferentiate. Differentiation of the daughter cellsleaving a niche appears to be dictated by the en-vironment rather than by cell origin (128). Invitro, however, the potential of stem cells canbe expanded experimentally (101).
The excitement that the progress in animalstem cell research has caused is mainly due toits potential in regenerative medicine. Is therea similar potential for plant stem cells in agri-culture and breeding? Indeed, despite multipleexamples of nonsexual propagation, regenera-tion in many crop species has remained elusiveand inefficient. Triggering an excess of stemcells can overcome this problem, as exemplifiedby the enhanced induction of somatic embryosin Arabidopsis stem-cell-surplus mutants (94).This review discusses current models and openquestions in plant stem cell research that have
been identified during the past two decadesby genetic studies, mainly in the model plantArabidopsis thaliana. For aspects not addressedhere, we refer readers to several excellent re-views covering the initiation of meristems in theembryo (9, 15, 72) and organ formation (18, 79).
STRUCTURE AND FUNCTION OFTHE SHOOT APICAL MERISTEM
The Arabidopsis shoot meristem is organizedinto a central zone (CZ) of slowly dividingcells that contains the stem cells, a surroundingperipheral zone (PZ) of more rapidly dividingcells from which lateral organs are derived,and an underlying rib zone where cells assumea flattened shape as the first indication ofdifferentiation toward central stem tissue(Figure 1a). Shoot meristem stem cells func-tion in a population mode—that is, individualdivisions are not strictly asymmetric, generat-ing one new stem cell and one differentiatingcell (73, 123). Rather, whether any givencell becomes a new stem cell or undergoesdifferentiation depends on its position. Geneticstudies of the past decade provided substantialinsight into two questions: How is stem cellidentity determined? And how are boundariesmaintained in the dividing cellular context ofthe shoot meristem?
Regulation of the Cytokinin/Gibberellic Acid Balanceby KNOX Genes
The first plant stem cell regulator gene wasisolated from the maize leaf mutant knotted(kn) (50). KN encodes a homeodomain proteinthat in the dominant active kn-1 mutant isectopically expressed in leaf veins, resultingin proliferating “knots.” KN is the foundingmember of the KNOTTED1-like homeobox(KNOX) genes. Its normal expression is inundifferentiated cells of the shoot meristemdome, but notably, it is absent from the cells inleaf anlagen, consistent with a model in whichKN promotes the undifferentiated cell state inthe shoot meristem (120).
www.annualreviews.org • Plant Stem Cell Niches 617
Ann
u. R
ev. P
lant
Bio
l. 20
12.6
3:61
5-63
6. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by U
nive
rsid
ad A
uton
oma
de Q
uere
taro
on
10/2
4/13
. For
per
sona
l use
onl
y.
PP63CH25-Laux ARI 27 March 2012 11:48
LP
CZPZ
LP
RZ
a
Expression ofkey shoot apical
meristem regulators
CLV3
WUS/AHK4
CLV1
ZLL
miR165/166
HD-ZIPIII
AGO1AGO1AGO1
d
KNOX
CKAS1
Auxin
GASTM/KNOX
AS1CK
b
WUS
ARR7/15
CK
CLV3
CLV1
CKCKRp
IPT7
LOG
?
STM
GAAS1
Auxin
c
SC
OC
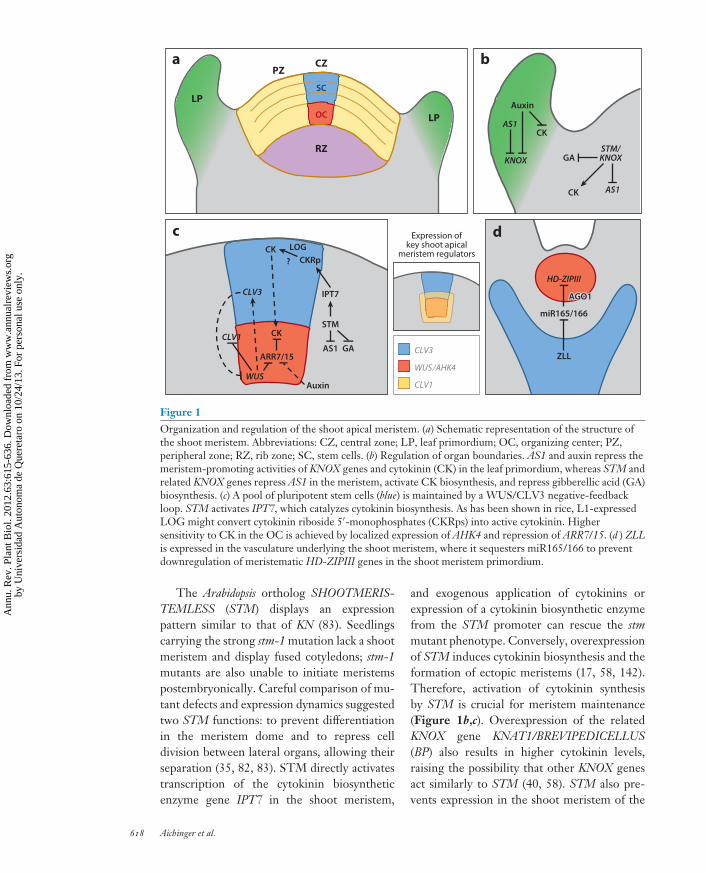
Figure 1Organization and regulation of the shoot apical meristem. (a) Schematic representation of the structure ofthe shoot meristem. Abbreviations: CZ, central zone; LP, leaf primordium; OC, organizing center; PZ,peripheral zone; RZ, rib zone; SC, stem cells. (b) Regulation of organ boundaries. AS1 and auxin repress themeristem-promoting activities of KNOX genes and cytokinin (CK) in the leaf primordium, whereas STM andrelated KNOX genes repress AS1 in the meristem, activate CK biosynthesis, and repress gibberellic acid (GA)biosynthesis. (c) A pool of pluripotent stem cells (blue) is maintained by a WUS/CLV3 negative-feedbackloop. STM activates IPT7, which catalyzes cytokinin biosynthesis. As has been shown in rice, L1-expressedLOG might convert cytokinin riboside 5′-monophosphates (CKRps) into active cytokinin. Highersensitivity to CK in the OC is achieved by localized expression of AHK4 and repression of ARR7/15. (d ) ZLLis expressed in the vasculature underlying the shoot meristem, where it sequesters miR165/166 to preventdownregulation of meristematic HD-ZIPIII genes in the shoot meristem primordium.
The Arabidopsis ortholog SHOOTMERIS-TEMLESS (STM) displays an expressionpattern similar to that of KN (83). Seedlingscarrying the strong stm-1 mutation lack a shootmeristem and display fused cotyledons; stm-1mutants are also unable to initiate meristemspostembryonically. Careful comparison of mu-tant defects and expression dynamics suggestedtwo STM functions: to prevent differentiationin the meristem dome and to repress celldivision between lateral organs, allowing theirseparation (35, 82, 83). STM directly activatestranscription of the cytokinin biosyntheticenzyme gene IPT7 in the shoot meristem,
and exogenous application of cytokinins orexpression of a cytokinin biosynthetic enzymefrom the STM promoter can rescue the stmmutant phenotype. Conversely, overexpressionof STM induces cytokinin biosynthesis and theformation of ectopic meristems (17, 58, 142).Therefore, activation of cytokinin synthesisby STM is crucial for meristem maintenance(Figure 1b,c). Overexpression of the relatedKNOX gene KNAT1/BREVIPEDICELLUS(BP) also results in higher cytokinin levels,raising the possibility that other KNOX genesact similarly to STM (40, 58). STM also pre-vents expression in the shoot meristem of the
618 Aichinger et al.
Ann
u. R
ev. P
lant
Bio
l. 20
12.6
3:61
5-63
6. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by U
nive
rsid
ad A
uton
oma
de Q
uere
taro
on
10/2
4/13
. For
per
sona
l use
onl
y.

PP63CH25-Laux ARI 27 March 2012 11:48
ASYMMETRIC LEAVES1 (AS1) gene, whichrepresses the meristem-promoting factorsKNAT1/BP and KNAT2 in lateral organ pri-mordia (20). Interestingly, the shoot meristemis recovered in as1 stm-1 double mutants,showing that meristem cells can be maintainedwithout STM activity. It is plausible that, inthis situation, KNAT genes can replace STMfunction.
STM (and related KNOX genes in otherspecies) furthermore downregulates gibberellicacid (GA) levels in the shoot meristem throughdirect repression of the GA-biosynthesis geneGA 20-oxidase and upregulation of the GA-degrading gene GA 2-oxidase (23, 58, 111).GAs promote differentiation and thus can beviewed as antagonists of meristem identity. GA2-oxidase is expressed at the base of organ pri-mordia. One attractive model is that GA move-ment from the leaves into the shoot meristemis thereby excluded (reviewed in 119).
Local Control of Stem Cells by WUS
Seedlings of the wuschel (wus) mutant lack ashoot meristem and display partially differenti-ated cells at the position of the stem cells, sug-gesting that WUS is required to prevent dif-ferentiation of stem cells (74, 91). In contrastto stm mutants, wus mutants can initiate ad-ventitious shoot meristems postembryonically,although these terminate after the generationof a few organs. Conversely, overexpression ofWUS leads to enlarged meristems, suggestingthat WUS is also sufficient to promote stem cellidentity (17, 76, 115, 141). Notably, this abilityappears to be limited to immature tissues, im-plying that other factors are also required. WUSencodes a plant-specific homeodomain proteinand is the founding member of the WUSCHEL-RELATED HOMEOBOX (WOX) gene family,which regulates diverse aspects of development(130). WUS expression in the shoot meristemdefines the organizing center (OC), which inseedlings is located in the fourth- and fifth-outermost cell layers, underneath the threestem cell layers (Figure 1a,c); in the inflores-cence meristem and floral meristem, however,
the OC variably overlaps with stem cells in thethird outermost cell layer. In addition to main-taining the undifferentiated nature of the stemcells, WUS is required for expression of theCLAVATA3 (CLV3; see below) gene in the stemcell region of shoot and floral meristems (17, 76,115). Because all three layers of stem cells areaffected by WUS expression in the OC, a stem-cell-promoting signal from the OC has beenpostulated (91). Yadav and colleagues (140)demonstrated that the WUS protein movesfrom the OC into the CZ, where it binds di-rectly to the CLV3 promoter, providing anotherexample of moving plant transcription factorsin cell-cell communication (97, 120). This in-tercellular movement appears to be critical forstem cell maintenance, as decreasing WUS mo-bility results in loss of the shoot meristem.
In OC cells, WUS directly downregulatesthe transcription of the type A ARABIDOPSISRESPONSE REGULATOR genes ARR7 andARR15 (Figure 1c), which encode inhibitorsof intracellular cytokinin signal transduction(75). Overexpression of a constitutively activeform of ARR7 caused shoot meristem defectssimilar to those in wus mutants, suggestingthat promoting cytokinin signal transductionin OC cells is an important part of how WUSacts in stem cell maintenance. Chromatinimmunoprecipitation experiments combinedwith transcriptional profiling revealed thatWUS acts on a larger set of genes involved inmeristem regulation, hormone pathways, andcell division control (19). Notably, this studypoints to additional roles of WUS in regulatingtwo phytohormones: jasmonate ( JA) throughrepression of the JA response factor JAZ5, andauxin (see below) through modulation of auxintransport and response genes. In summary, theOC-derived WUS protein must be present inboth the OC and the stem cells to maintain thestem cells in an undifferentiated state.
ZLL/AGO1 Balance Determines StemCell Inhibitory MicroRNA Levels
The ZWILLE/PINHEAD/AGO10 (ZLL) genehas been identified in mutant screens for
www.annualreviews.org • Plant Stem Cell Niches 619
Ann
u. R
ev. P
lant
Bio
l. 20
12.6
3:61
5-63
6. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by U
nive
rsid
ad A
uton
oma
de Q
uere
taro
on
10/2
4/13
. For
per
sona
l use
onl
y.

PP63CH25-Laux ARI 27 March 2012 11:48
meristem regulators and encodes an ARG-ONAUTE (AGO) protein, implying a functionin RNA silencing. Mutant zll seedlings displaya range of phenotypes, from flat apices of differ-entiated cells to a terminal organ in place of theshoot meristem (92, 95). However, WUS is stillexpressed in the OC of zll embryos, whereasCLV3 expression is initiated in the presumptivestem cells but is not maintained. In addition,accumulation of stem cells by overexpressionof WUS is suppressed in zll mutants (127).These findings suggest that ZLL is required topotentiate WUS-dependent stem cell signal-ing. ZLL expression in the underlying vascularprimordium is sufficient for shoot meristemdevelopment, indicating that the vasculatureplays a role in controlling the shoot meristem.
The primary cause for the loss of the shootmeristem in zll seedlings appears to be the ac-cumulation of microRNA165/166 (miR165/166)and the consequent downregulation of theirtarget HOMEODOMAIN-LEUCINE ZIPPERIII (HD-ZIPIII) messenger RNAs (mRNAs)in the shoot meristem (Figure 1d ) (80). HD-ZIPIII genes encode homeodomain proteinsthat act as key meristem regulators, amongother functions (9, 105). Multiple mutantsof HD-ZIPIII genes such as PHABULOSA(PHB), PHAVOLUTA (PHV), and REVO-LUTA (REV ) or overexpression of miR166lead to shoot meristem termination (34, 136),whereas upregulation of HD-ZIPIII genesleads to enlarged or ectopic meristems (93).How HD-ZIPIII proteins promote stem cellmaintenance is unclear.
A puzzling question is how ZLL protein inthe vasculature can prevent miR165/166 accu-mulation in the shoot meristem primordium.Surprisingly, molecular and genetic dataindicate that ZLL and its close homolog AGO1have not only overlapping but also antagonisticeffects on gene silencing and development(88). One proposed model is that ZLL mightsequester miR165/166 and thus block itsaccumulation in the shoot meristem (18, 24).Biochemical evidence demonstrated that ZLLbinds miR165/166 more efficiently than AGO1but does not seem to degrade HD-ZIPIII
mRNA as efficiently as AGO1, suggesting thatZLL sequesters miR165/166 away from the cat-alytically more active AGO1, and consequentlyHD-ZIPIII expression is increased (151).HD-ZIPIII activity also promotes adaxial leaffate, which had previously been shown topositively affect shoot meristem maintenance(132), whereas abaxial identity genes, suchas KANADI, antagonize it (reviewed in 9).However, it has not yet been distinguished howHD-ZIPIII expression in the shoot meristemand in the leaf differ in their effects on shootmeristem development.
Control of the Stem Cell Number
Although all shoot meristem cells divide, theshoot meristem keeps its shape and size (withseasonal changes occurring) and its internal or-ganization. How are boundaries in the shootmeristem controlled? Mutants with an ex-panded stem cell pool have been the start-ing point for some answers. Analysis of clavata(clv) mutants, which display gradually enlarg-ing shoot meristems and floral meristems thatproduce more organs than those of wild-typeplants (26, 27), led to the identification of asignal cascade central to stem cell regulation.
CLV3 (39) belongs to a family of 32small proteins called CLV3/EMBRYO SUR-ROUNDING REGION (CLE), which areposttranslationally processed into the activesignal peptides CLEp (29, 57). The expandedmeristem of clv3 mutants is caused by anenlargement of the WUS expression domain(115). Conversely, overexpression of CLV3 re-sulted in repression of WUS transcription anda phenocopy of the wus mutant (16, 77). CLV3is expressed in a wedge-shaped domain thatroughly coincides with the position of the stemcells. Notably, the number of cells in the L3layer that express CLV3 appears smaller thanthe number of cells in the L1 layer, indicatingthat CLV3 expression is not strictly linked tostem cells, if there are the same number ofstem cells in each layer. CLV3 transcript levelsare positively regulated by WUS, as CLV3expression is lost in wus mutants and expanded
620 Aichinger et al.
Ann
u. R
ev. P
lant
Bio
l. 20
12.6
3:61
5-63
6. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by U
nive
rsid
ad A
uton
oma
de Q
uere
taro
on
10/2
4/13
. For
per
sona
l use
onl
y.

PP63CH25-Laux ARI 27 March 2012 11:48
in WUS overexpressors (17, 115). This estab-lishes a negative-feedback loop in which WUSexpression in the OC is required for CLV3 ex-pression in the stem cells, which in turn delimitsthe WUS expression domain (Figure 1c). Thisfeedback loop provides a conceptual frame-work for how the size of the stem cell pool isdynamically assessed and kept stable. Notably,however, the meristem appears able to toleratesignificant alterations of CLV3 levels withoutphenotypic consequences, and initial down-regulation of WUS expression can be reversedat later time points, suggesting additional reg-ulatory mechanisms that buffer stem cell main-tenance against incidental fluctuations (96).
CLV3p is perceived by a multitude of recep-tor complexes (reviewed in 61). CLV1 encodesa leucine-rich repeat (LRR) receptor kinaseexpressed in a central domain of the shootmeristem, including the OC and L3 layer of thestem cells (25). CLV2 also codes for an LRR-receptor-like transmembrane protein that lacksan intracellular kinase domain. CLV2 interactswith CORYNE (CRN)/SUPPRESSOR OFLLP1 2 (SOL2) (which shares similaritywith serine/threonine kinases but lacks anextracellular LRR domain) to form a CLV3-receptor complex independently of CLV1 (48).Nimchuk et al. (99) recently provided evidencethat CRN is unable to autophosphorylateand can mediate its function without a func-tional kinase domain; the authors speculatedthat CRN could have scaffolding functionsanalogous to those of animal pseudokinases.Finally, a third important player in CLV3pperception is the recently identified receptorRECEPTOR-LIKE PROTEIN KINASE2 (RPK2)/TOADSTOOL2 (TOAD2) (64).The clv1 clv2 rpk2 triple mutant phenocopiesthe clv3 mutant, suggesting that these threereceptors constitute the three main pathwaysof CLV3p perception in the shoot meristem.
Where in the shoot meristem does CLV3activity repress WUS? In clv3 mutants, WUSexpression is both shifted one layer up andexpanded laterally, suggesting that CLV3normally represses WUS at the distal andlateral boundaries of the OC. Notably, CLV3
overexpression in the L1 layer can completelyrepress WUS expression in the OC, causinga wus phenocopy (77). How is this preventedunder normal conditions? An active CLV3–green fluorescent protein (GFP) fusion proteinspread away from the stem cells predominantlyin the lateral direction rather than into theunderlying OC. Both effects were blocked bythe presence of a functional CLV1 receptor,indicating that the CLV3-GFP fusion proteinremained uncleaved and that the observedspread was not due to clipped off GFP. Thesedata suggest that binding of CLV3p to its re-ceptors in the overlying cells contributes to OCstability (77). However, recent findings are atodds with such a mechanism: CLV3-dependentinternalization of a functional CLV1-2xGFP asa proxy for CLV3p binding was also detectedin deeper cell layers of the shoot meristem,irrespective of CLV1 levels, suggesting thatligand sequestering is not important (100).Curiously, most of the CLV1 receptors appearalready internalized in wild-type plants, sug-gesting a largely saturated response to CLV3,which raises the question of how increasedCLV3 levels can terminate the meristem andWUS expression as previously shown (16, 77).Notably, WUS was found to directly repressCLV1 expression, and thus might reduce theeffect of CLV3p in the OC cells, therebysustaining its own expression (19). Therefore,a decrease in sensitivity to CLV3p in OC cellsmay also contribute to stability of the OC.
Several decades ago, mechanical destruc-tion of the meristem center in Impatiens royleicaused induction of new meristems from thePZ, suggesting that this developmental optionis normally repressed in the PZ by lateralinhibition (81). By elegant laser ablation exper-iments in tomato apices, Reinhardt et al. (108)demonstrated that only ablation of the entireCZ, and not merely of the stem cell layers,causes rapid activation of WUS in adjacentPZ cells. Together, these results suggest thatthe OC promotes stem cell fate in its apicalneighbors but represses it in PZ cells, whichin turn implies the requirement of additionalfactors that modify cell responsiveness. In
www.annualreviews.org • Plant Stem Cell Niches 621
Ann
u. R
ev. P
lant
Bio
l. 20
12.6
3:61
5-63
6. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by U
nive
rsid
ad A
uton
oma
de Q
uere
taro
on
10/2
4/13
. For
per
sona
l use
onl
y.

PP63CH25-Laux ARI 27 March 2012 11:48
Arabidopsis, Reddy & Meyerowitz (107) alsoreported lateral inhibition when manipulatingthe stem cells. Using an inducible downregu-lation of CLV3 expression in the CZ by RNAinterference (RNAi), they observed that cellsclose to the CZ reverted back to stem cellfate (which potentially could be mediated byderepressed WUS expression), whereas cellsmore distant from the CZ displayed increasedcell division, separating independent functionsof CLV3. In a different set of experiments byYadav et al. (141), downregulation of WUS inthe OC resulted in enlargement and a gradualshift of the auxin maxima marking lateral organanlagen toward the CZ, implying that WUSalso has a non-cell-autonomous function incontrolling auxin responsiveness in the PZ.
In summary, control of WUS gene ex-pression and multiple levels of lateral inhi-bition contribute to robustly maintaining theboundaries between pluripotent stem cells anddifferentiating descendants.
What Keeps the Stem Cell Nicheat the Tip of the Shoot?
If cells above the OC undergo periclinaldivisions, why is the OC not gradually dis-placed away from the apex? Schoof et al. (115)proposed that in addition to the repressiveCLV3 signal, stem cells also emanate a gradedsignal that promotes WUS expression and thusanchors the stem cell niche to the tip of theplant. Several recent observations are consis-tent with a model in which cytokinin might beinvolved in this process. As said above, STMpromotes cytokinin synthesis throughout themeristem with the exception of incipient organprimordia. In rice, the LONELY GUY (LOG)gene, which encodes the enzyme catalyzingthe final step of cytokinin biosynthesis, isspecifically expressed in the L1 layer, includingthe stem cell domain (69). Functional evidencein Arabidopsis is still missing, but Yadav et al.(139) reported expression of LOG homologsin the shoot meristem. It is thus tempting tospeculate that by producing active cytokinin,the L1 layer provides apical-basal information
to the underlying cells. Expression of the geneencoding the cytokinin receptor ARABIDOP-SIS HISTIDINE KINASE 4 (AHK4) overlapswith the OC and is required for induction ofWUS expression by cytokinin (44). Togetherwith the finding that WUS promotes intracel-lular cytokinin signals (75), this suggests thatOC cells are dedicated within the shoot meris-tem to perceiving and transducing cytokininsignaling, and thus reinforce WUS expression.By contrast, in the stem cells, the cytokininsignaling inhibitors ARR7 and ARR15 arerequired for high levels of CLV3 expression(149).
Along the same lines, Yoshida et al. (144) as-sociated downregulation of cytokinin responsewith strongly increased CLV3 expression in thetomato shoot meristem. The surprising aspectof this work is that light is required for nor-mal cytokinin response in the shoot apex. Im-portantly, this requirement is independent ofphotosynthesis and therefore suggests that lightacts as a signal to increase cytokinin response,which in turn keeps the CLV signal pathway incheck and thus promotes proliferation. At thesame time, light is also required for the forma-tion of auxin maxima, involving PINFORMED1 (PIN1) localization. Notably, in this study,light was able to trigger lateral primordia initi-ation in the absence of leaves. Intuitively, onemight not assume that the shoot apex is a placefor light perception, because in many cases it iscovered by developing leaves. Thus, either sen-sitive photoreceptors might enable the shootmeristem to perceive light in a natural situa-tion as well, or the leaves might take over thisfunction when the apex is covered.
THE ROOT STEM CELL NICHE
Longitudinal root growth originates at the tipsof the roots where the root stem cells reside. Inthe center of the root tip is the quiescent center(QC), which is mitotically relatively inactive inArabidopsis. The stem cells directly surround theQC and give rise to the different cell files of theroot, the stele, the ground tissue (consisting ofendodermis and cortex), the epidermis, and the
622 Aichinger et al.
Ann
u. R
ev. P
lant
Bio
l. 20
12.6
3:61
5-63
6. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by U
nive
rsid
ad A
uton
oma
de Q
uere
taro
on
10/2
4/13
. For
per
sona
l use
onl
y.
PP63CH25-Laux ARI 27 March 2012 11:48
c
a b
TPSTexpression
PLTprotein
RGF1/2/3expression
RGFprotein
SHR
Quiescent centerColumellaLateral root capEpidermisCortexEndodermisStele
SCR
WOX5
ACR4
CLE40
CLE40
CLE40
SCR
SCR
ACR4ACR4
ACR4
SCR
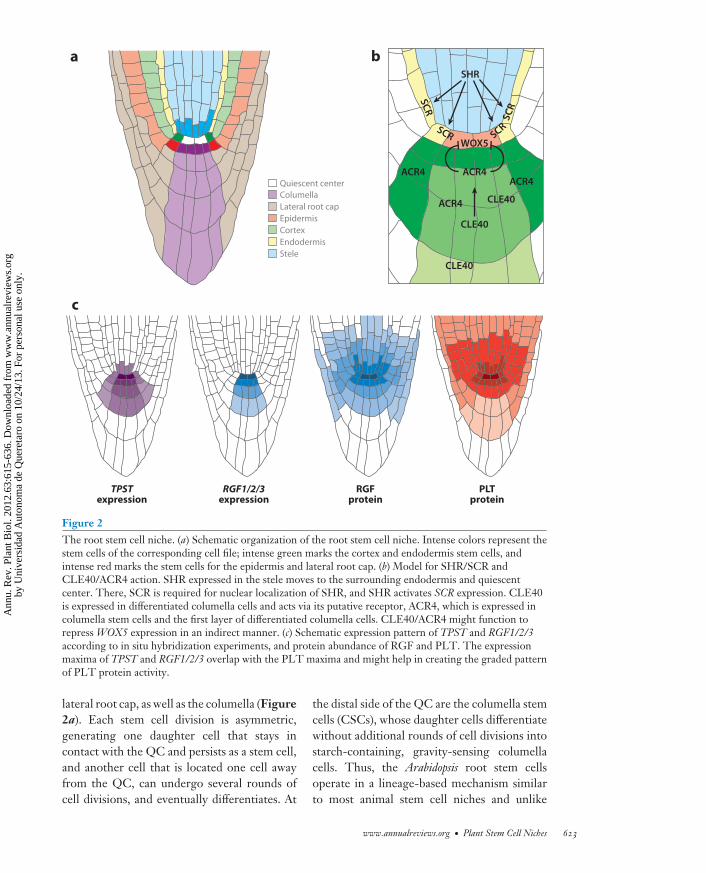
Figure 2The root stem cell niche. (a) Schematic organization of the root stem cell niche. Intense colors represent thestem cells of the corresponding cell file; intense green marks the cortex and endodermis stem cells, andintense red marks the stem cells for the epidermis and lateral root cap. (b) Model for SHR/SCR andCLE40/ACR4 action. SHR expressed in the stele moves to the surrounding endodermis and quiescentcenter. There, SCR is required for nuclear localization of SHR, and SHR activates SCR expression. CLE40is expressed in differentiated columella cells and acts via its putative receptor, ACR4, which is expressed incolumella stem cells and the first layer of differentiated columella cells. CLE40/ACR4 might function torepress WOX5 expression in an indirect manner. (c) Schematic expression pattern of TPST and RGF1/2/3according to in situ hybridization experiments, and protein abundance of RGF and PLT. The expressionmaxima of TPST and RGF1/2/3 overlap with the PLT maxima and might help in creating the graded patternof PLT protein activity.
lateral root cap, as well as the columella (Figure2a). Each stem cell division is asymmetric,generating one daughter cell that stays incontact with the QC and persists as a stem cell,and another cell that is located one cell awayfrom the QC, can undergo several rounds ofcell divisions, and eventually differentiates. At
the distal side of the QC are the columella stemcells (CSCs), whose daughter cells differentiatewithout additional rounds of cell divisions intostarch-containing, gravity-sensing columellacells. Thus, the Arabidopsis root stem cellsoperate in a lineage-based mechanism similarto most animal stem cell niches and unlike
www.annualreviews.org • Plant Stem Cell Niches 623
Ann
u. R
ev. P
lant
Bio
l. 20
12.6
3:61
5-63
6. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by U
nive
rsid
ad A
uton
oma
de Q
uere
taro
on
10/2
4/13
. For
per
sona
l use
onl
y.

PP63CH25-Laux ARI 27 March 2012 11:48
the shoot meristem (73). Notably, each stemcell in the root meristem gives rise to onlyone tissue, raising the question of whether thestem cell potential is limited. Elegant ablationstudies have shown, however, that the daughtercells differentiate according to signals fromolder differentiated cells and have the abilityto switch fates if displaced to a new position(128).
The simple structure of the Arabidopsis rootmakes it an ideal model for developmental stud-ies, but compared with other species this is morean exception than the rule. For example, the QCin maize root consists of up to 1,000 cells andshows a gradient of quiescence with increasingmitotic activity toward the proximal meristem(28, 37). Although species with similarly largeroot meristems have been used for initial histo-logical studies, the stereotypic and simple orga-nization of Arabidopsis roots has aided its use as amodel for stem cell research in plants. Here, wediscuss how QC cells and stem cells are spec-ified and how the specific pattern of the rootstem cell niche is regulated.
The Quiescent Center Functionsas Organizer of the Root StemCell Niche
Direct evidence that the QC plays a role incontrolling stem cells came from laser ablationof individual Arabidopsis QC cells (129). CSCsabutting the ablated QC cells ceased prolifer-ation and differentiated into starch-containingcolumella cells, whereas abutting cortex and en-dodermis initials (CEIs) differentiated into CEIdaughter cells. The fact that only cells in directcontact with the QC are maintained as stemcells might suggest short-range or contact-based signaling. That this is not the case wasshown in an elegant experiment by Wildwateret al. (135) in which differentiation of stemcell daughters was blocked by RNAi-mediateddownregulation of RETINOBLASTOMA-RELATED (RBR) activity, resulting in severallayers of undifferentiated cells next to the QC.These extra cells lost their undifferentiated
state when the QC was ablated, indicating thatthe stem-cell-promoting signals from the QCcan work over several cell diameters but nor-mally are counteracted in cells without directcontact to the QC. Although the QC-bornesignal molecules are still undiscovered, geneticand molecular studies have identified pathwaysthat are essential for stem cell maintenance inthe Arabidopsis root.
WOX5 Expression in the QuiescentCenter Is Required to MaintainStem Cells
The WUS homolog WOX5 is specifically ex-pressed in the QC, and loss of WOX5 functionleads to differentiation of the CSCs, similarto what was observed upon QC ablation (49,112). Furthermore, overexpression of WOX5in the columella blocks differentiation andgenerates stem-cell-like cells. In contrast tothe excessive undifferentiated cells caused byRBR downregulation, QC ablation does notsuppress the effects of WOX5 overexpression,consistent with the hypothesis that no QCsignal other than the one(s) generated byWOX5 is required for stem cell maintenance.In addition to its effect on CSCs, WOX5 is alsorequired for maintaining proximal stem cells,but here WOX5 acts redundantly with theSHORTROOT (SHR)/SCARECROW (SCR)and PLETHORA (PLT ) pathways (112).
Several CLE peptides can cause differen-tiation of stem cells and/or reduce the size ofthe root meristem (16, 22, 54, 57, 124). TheCLE40 gene is expressed in the differentiatedcolumella cells and promotes differentiationvia the receptor-like kinase ARABIDOPSISCRINKLY 4 (ACR4) (124). Both acr4 andcle40 loss-of-function mutants display anadditional layer of distal stem cells and a lateralexpansion of WOX5 expression (31, 124).Thus, the CLE40/ACR4 module provides asignal that counteracts stem-cell-promotingQC signals and allows distant columella cellsto differentiate (Figure 2b). As the expressionpattern of the putative CLE40p receptor ACR4
624 Aichinger et al.
Ann
u. R
ev. P
lant
Bio
l. 20
12.6
3:61
5-63
6. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by U
nive
rsid
ad A
uton
oma
de Q
uere
taro
on
10/2
4/13
. For
per
sona
l use
onl
y.

PP63CH25-Laux ARI 27 March 2012 11:48
does not overlap with the WOX5 expressiondomain, the effect on WOX5 expression mightbe indirect, e.g., owing to misspecified CSCs.
The Stele-Borne SHR SignalIs Required for QuiescentCenter Function
The GRAS (GAI, RGA, SCR) transcriptionfactor SHR is expressed in the stele and movesto the surrounding cells, including the QC,where it activates expression of the related SCRgene (Figure 2b) (51, 97). SCR itself is re-quired for the nuclear localization of SHR, andmutations in either SCR or SHR result in irreg-ular morphology of the stem cell niche, lack ofQC-specific marker expression, and ultimatelythe collapse of the root meristem (30, 78, 110).QC-specific expression of SCR in the shr mu-tant background cannot rescue the QC defectsof shr, suggesting that both proteins must bepresent in the QC to maintain stem cell activity(110).
Long-Range Control of the Stem CellNiche via Auxin and PLT Genes
The root stem cell niche is marked by an auxinmaximum at the position of the QC (104,109). Computer modeling and the discoveryof polar localization of PIN auxin transportfacilitators to one side of a cell suggest thatauxin accumulates in the QC region by arootward-directed auxin transport in thevasculature and a shootward-directed transportin the lateral root cap and epidermis (46, 137).After excision of the root tip or ablation ofthe QC, a new auxin maximum is established afew cell layers apically from the new tip and anew stem cell niche is formed (109, 118, 138),suggesting that the auxin maximum and thestem cell niche are functionally linked.
Auxin function in the stem cell niche ismediated by PLT transcription factors (3, 42).The activity of the different PLT proteinsis additive, and manipulating the expressionlevels suggests a dose-dependent readout,
reminiscent of animal morphogens. The high-est PLT levels are in the QC, and seem to be re-quired for specifying and maintaining the stemcell niche; intermediate PLT levels in the prox-imal meristem are required for mitotic activity;and low levels correlate with differentiation(Figure 2c). Because the response of PLTexpression to auxin occurs later than forother known auxin response genes, it hasbeen postulated to be rather indirect (3).Tyrosylprotein sulfotransferase (TPST) andthe ROOT GROWTH FACTOR (RGF)tyrosine-sulfated peptides might link auxinto PLT protein levels (90, 150). The currentmodel holds that auxin induces the expressionof TPST and several RGF genes. TPST in turnsulfates the RGF peptides, which by an un-known mechanism induce transcriptional andposttranscriptional upregulation of the PLTprotein levels (Figure 2c). The direct targetsof PLT proteins are still unknown. Notably,PLT activity enhances PIN expression, whichmight create a positive-feedback loop thatstabilizes the auxin maximum at the root tip(14).
As in many other examples, the readout ofauxin in the root stem cell system depends onthe cellular context. Unlike in the QC, in thecolumella, auxin restricts stem cell identity andpromotes differentiation. This occurs via theAUXIN RESPONSE FACTOR 10 (ARF10)and ARF16, which are activated by auxin (133).ARF16 is expressed in differentiated columellacells and CSCs, and in the double mutant arf10arf16, the CSC daughter cells fail to undergodifferentiation. Furthermore, increased ARF16activity reduces the levels of WOX5 expression(32). A similar repressive effect on WOX5 can beobtained after application of auxin, which mightact via auxin-mediated upregulation of ARF16.As WOX5 and ARF16 do not have overlappingexpression domains, ARF16 might be involvedin restricting the WOX5 domain to the QC andthus allowing CSC daughter cells to differenti-ate. In summary, the hormone auxin not onlypromotes the root stem cell niche, but is alsoinvolved in restricting it.
www.annualreviews.org • Plant Stem Cell Niches 625
Ann
u. R
ev. P
lant
Bio
l. 20
12.6
3:61
5-63
6. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by U
nive
rsid
ad A
uton
oma
de Q
uere
taro
on
10/2
4/13
. For
per
sona
l use
onl
y.

PP63CH25-Laux ARI 27 March 2012 11:48
Quiescence of the Quiescent Center
The low mitotic activity of QC cells is causedby a prolonged G1 phase. For example, maizeQC cells divide every 180–200 h, comparedwith a 20-h cell cycle in the proximal meristem(7, 28). Recent reports provide insight into howthe quiescence of QC cells is controlled. First,QC-specific knockout of RBR function indi-cates that RBR suppresses cell divisions in theQC (131). Second, the oxidized redox statusof the QC has been proposed to cause arrestat the G1/S transition (for a review, see 59).As this oxidized status is postulated to be dueto auxin degradation, how auxin, redox regula-tion, and cell cycle control are linked will be aninteresting question.
The biological significance of a low celldivision rate in the QC is unknown. Notably,under certain conditions, cell divisions of QCcells do occur and QC derivatives can replacestem cells. In Arabidopsis thaliana, the QC ismitotically more active in older roots, andmitosis is induced by stress conditions, alteredhormone levels, or a reduced redox status(10, 32, 60, 63, 102, 146). In other species withlarger stem cell niches, such as maize, there isno clear boundary between mitotically almostinactive QC cells and the dividing stem cellsof the proximal meristem (28, 37). Therefore,the QC can be seen as a flexible and responsiveorganizer that is competent to replenish stemcells when necessary.
THE VASCULAR STEMCELL NICHE
The plant vasculature is the main route forlong-distance transport of water and min-erals upward in the xylem and of organiccompounds downward in the phloem; it alsogenerates mechanical support for the growingstem. Although the cell types are the same inall vascular plants, the architecture of the vas-culature varies, even between organs. Despiteits herbaceous nature, Arabidopsis has proven tobe an excellent model system for vascular de-velopment (147). In the Arabidopsis root, thexylem is located in a central row of cells, withthe protoxylem located on the marginal posi-tions and the metaxylem in a central position(Figure 3a). On the perpendicular axis, twopoles of phloem are present and the interveningprocambium consists of pluripotent stem cells.These tissues are surrounded by the pericy-cle and together form the vasculature (or stele)(Figure 3a). In the stem, a ring of vascularbundles is present with phloem on the outside,the procambium (or fascicular cambium) in themiddle and the xylem on the inside (Figure 3b).During secondary growth, the fascicular cam-bium and the interfascicular cambium (IC) forma closed cambium ring. The current opinion,supported by transcriptional profiling in Popu-lus (116), is that the cambium contains stem cellswith phloem mother cells on one side and xylemmother cells on the other (reviewed in 33).
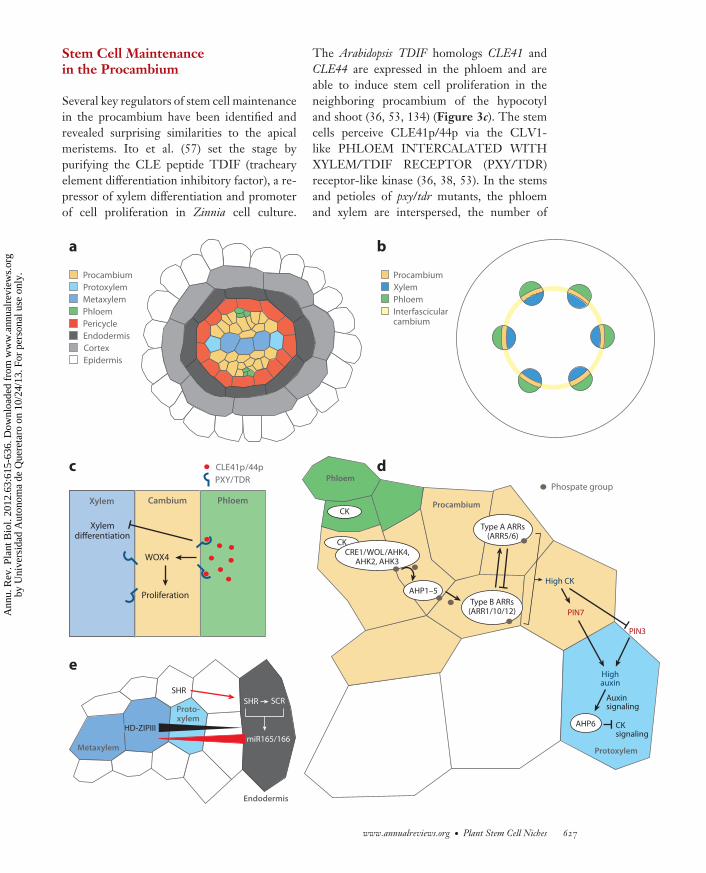
−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−→Figure 3The vascular stem cell niche, with involved cell types shown in color. (a) Organization of vasculature in the Arabidopsis postembryonicroot. (b) Organization of vasculature in the Arabidopsis postembryonic stem. During secondary growth, the intervening procambial cells(or fascicular cambium) and the interfascicular cambium files start to undergo periclinal divisions to initiate radial growth, continuouslyproducing the xylem (inward) and phloem (outward) in concentric rings. (c) The CLE–PXY/TDR–WOX4 pathway is involved in stemcell proliferation and possibly cell division orientation. (d ) In the root, cytokinin (CK) and auxin determine boundary formationbetween the procambium and protoxylem. CK is transported from the phloem and binds to hybrid histidine kinase receptors (CRE1/WOL/AHK4, AHK2, AHK3), inducing autophosphorylation. The phosphate group is then transferred to Arabidopsis histidinephosphotransfer proteins (AHP1–5), which move into the nucleus and phosphorylate Arabidopsis response regulators (ARRs). Type BARRs act as activators and directly induce type A ARRs, which act as repressors (reviewed in 4, 103, 126). The CK signal influenceslocalization of PIN3 and PIN7, creating an auxin maximum in the xylem. There, auxin induces pseudophosphotransfer protein AHP6,which acts as an inhibitor of cytokinin signaling and promotes protoxylem formation. (e) In the root, positional information from theendodermis specifies protoxylem and metaxylem differentiation through HD-ZIPIII protein-level regulation. The SHR gene isexpressed in the stele and the SHR protein moves to the endodermis (red arrow), where it activates SCR gene expression. SHR and SCRdirectly activate miR165/166 gene expression, and miR165/166 subsequently moves toward the center of the vasculature.Downregulation of HD-ZIPIII mRNA by miR165/166 in turn creates a reverse HD-ZIPIII gradient, which specifies dosage-dependentxylem differentiation.
626 Aichinger et al.
Ann
u. R
ev. P
lant
Bio
l. 20
12.6
3:61
5-63
6. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by U
nive
rsid
ad A
uton
oma
de Q
uere
taro
on
10/2
4/13
. For
per
sona
l use
onl
y.
PP63CH25-Laux ARI 27 March 2012 11:48
Stem Cell Maintenancein the Procambium
Several key regulators of stem cell maintenancein the procambium have been identified andrevealed surprising similarities to the apicalmeristems. Ito et al. (57) set the stage bypurifying the CLE peptide TDIF (trachearyelement differentiation inhibitory factor), a re-pressor of xylem differentiation and promoterof cell proliferation in Zinnia cell culture.
The Arabidopsis TDIF homologs CLE41 andCLE44 are expressed in the phloem and areable to induce stem cell proliferation in theneighboring procambium of the hypocotyland shoot (36, 53, 134) (Figure 3c). The stemcells perceive CLE41p/44p via the CLV1-like PHLOEM INTERCALATED WITHXYLEM/TDIF RECEPTOR (PXY/TDR)receptor-like kinase (36, 38, 53). In the stemsand petioles of pxy/tdr mutants, the phloemand xylem are interspersed, the number of
a
e
c
WOX4
CLE41p/44pPXY/TDR
Proliferation
PhloemCambiumXylem
Xylemdifferentiation
SHRSHR SCR
miR165/166HD-ZIPIIIHD-ZIPIIIHD-ZIPIII
Endodermis
Proto-xylem
Metaxylem
d
AHP6
Phospate group
Highauxin
High CK
PIN3
PIN7
Auxinsignaling
CKsignaling
Phloem
Procambium
Protoxylem
Pericycle
MetaxylemProtoxylem
Phloem
Procambium
EndodermisCortexEpidermis
b
PhloemXylem
Interfascicularcambium
Procambium
CKCRE1/WOL/AHK4,
AHK2, AHK3
AHP1–5
Type A ARRs(ARR5/6)
Type B ARRs(ARR1/10/12)
CK
www.annualreviews.org • Plant Stem Cell Niches 627
Ann
u. R
ev. P
lant
Bio
l. 20
12.6
3:61
5-63
6. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by U
nive
rsid
ad A
uton
oma
de Q
uere
taro
on
10/2
4/13
. For
per
sona
l use
onl
y.

PP63CH25-Laux ARI 27 March 2012 11:48
procambium cells is reduced, and the orien-tation of procambium cell division is lost (38,53). Thus, the differentiating phloem daughtercells provide stem-cell-promoting signals andact like niche cells, similar to the OC and QC.
Ubiquitous overexpression of the ligand andthe receptor represses xylem differentiation andcauses more cells to accumulate in the vascularbundle and in the interfascicular region, rem-iniscent of secondary growth initiation (36).However, overexpression of CLE41 specificallyin the phloem and ubiquitous overexpressionof PXY/TDR do not repress xylem differenti-ation, suggesting that the range of CLE41p islimited by an unknown mechanism (36). There-fore, the CLE41p–PXY/TDR module definesthe boundary between vascular cell types andregulates the size of the vascular stem cellpopulation. Furthermore, the position of theCLE-peptide-producing cells relative to thestem cells has been reported to correlate withthe orientation of stem cell divisions, becauseubiquitous and xylem-specific expression ofCLE41 induces disoriented procambial cell di-visions, whereas overexpression of CLE41 in thephloem (where it is normally expressed) inducesonly correctly oriented cell divisions (36).
CLE41/44 signaling stimulates stem celldivisions in the procambium by promotingexpression of the WOX4 gene there (52)(Figure 3c). However, unlike in the pxy/tdrmutant, loss of WOX4 function does notresult in complete loss of the interveningprocambial cell layer and does not suppressdiscontinuous xylem strand formation uponapplication of TDIF. This shows that WOX4mediates only stem cell divisions regulated byPXY/TDR, whereas another still-unidentifiedpathway must mediate repression of xylemdifferentiation by PXY/TDR (52) (Figure 3c).
In addition to CLE41p/44p, treatment ofplants with combinations of CLE peptidesresults in cell proliferation in the vasculature(134). Furthermore, the two receptor-likekinases [MORE LATERAL GROWTH1(MOL1) and REDUCED IN LATERALGROWTH1 (RUL1)] seem to affect cambiumactivity in an opposite manner (1). Finally,
transcript profiling of Arabidopsis and Populuscambiums suggests that the known shootmeristem regulators CLV1 (expressed in thephloem and cambium) and STM (expressed inthe cambium) might also play a role in vascularstem cell maintenance (65, 116, 148). Howall these factors regulate vascular stem cellsrequires further investigation.
More Boundaries Within theVascular Stem Cell Niche
How do the cell types in the vasculature be-come organized within distinct boundaries? Inthe root vasculature, cytokinin moves throughsymplastic connections in the phloem from theshoot to the root (13). Cytokinin depletion orreduced cytokinin signaling results in fewercells in the vasculature and in all cells havingprotoxylem identity (5, 56, 67, 70, 85–87, 89,143) (Figure 3d ), suggesting that cytokininpromotes proliferation and maintenance of theprocambium stem cells. Cytokinin signalingis active in the intervening procambial cellfiles adjacent to the xylem axis (12) and affectslocalization of PIN3 (expressed in the peri-cycle adjacent to the protoxylem) and PIN7(expressed in the procambium and phloem) (12)(Figure 3d ). This bisymmetric PIN localiza-tion could channel auxin to the central xylemaxis, where it would induce expression of the cy-tokinin signaling repressor gene ARABIDOP-SIS HISTIDINE PHOSPHOTRANSFERPROTEIN 6 (AHP6) at the protoxylem posi-tion and consequently protoxylem formation(12). Therefore, a mutually inhibitory interac-tion between cytokinin and auxin determinesboundary formation between procambiumstem cells and the protoxylem (12).
The HD-ZIPIII proteins have recently beenshown to restrict the number of procambiumcells by promoting xylem differentiation (6, 55).SHR and SCR transcription factors directlyactivate miR165a/166b gene expression in theendodermis, and miR165/166 moves towardthe stele center and restricts expression of theHD-ZIPIII genes [PHB, PHV, REV, CORONA(CNA), ARABIDOPSIS HOMEOBOX GENE
628 Aichinger et al.
Ann
u. R
ev. P
lant
Bio
l. 20
12.6
3:61
5-63
6. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by U
nive
rsid
ad A
uton
oma
de Q
uere
taro
on
10/2
4/13
. For
per
sona
l use
onl
y.

PP63CH25-Laux ARI 27 March 2012 11:48
8 (ATHB8)] (Figure 3e) (21). The resultingHD-ZIPIII protein gradient determines xylemdevelopment in a dosage-dependent manner:High levels specify metaxylem and lowerlevels protoxylem (21). In conclusion, theneighboring endodermis provides positionalinformation for different cell fates along thexylem axis.
Coordination of Longitudinaland Lateral Growth
With increasing height, plants must ensure thattheir stems can carry their weight. Interestingly,plant body weight can induce secondary growthin Arabidopsis stems (Figure 3b), because the ad-dition of artificial weights (placing a 2.5-g tubeon the top of an immature plant) can induceIC imitation, probably through auxin signaling(66).
However, a recent study suggests thatthere is no linear correlation between plantheight/weight and IC initiation (117). The au-thors instead found that manipulation of JAsignaling affects secondary growth. Because thetouch-inducible JA signaling gene JAZ10 is ex-pressed at the base of the stem in the xylemand IC, it was hypothesized that intratissuetension might play a role. This tension mayarise from divisions in the fascicular cambiumor from xylem formation, which pushes thecambium outward, inducing JA signaling andthereby causing IC initiation (117). Thus, bodyweight and tension might provide input into thecambium stem cell niche.
MOLECULAR SIGNATUREOF PLANT STEM CELLS
The striking similarities between the shoot androot niches in both regulation and develop-ment have been interpreted as an indicationof an evolutionary relationship (112), in linewith paleobotanical views that the root evolvedfrom a shoot (106). CLE/WOX modules havebeen identified for all three stem cell niches dis-cussed in this review, but each acts distinctlyat the molecular level. First, CLE peptide sig-naling negatively regulates WUS and WOX5
CHROMATIN STATES IN ANIMALSTEM CELLS
In animals, the Oct4/POU5F1, Sox2, and Nanog transcriptionfactors are found in stem cells and are sufficient to reprogramdifferentiated cells into stem cells. This transcriptional programis implemented in the context of a distinct chromatin state instem cells (reviewed in 43, 101, 145). Pluripotent stem cellscontain open chromatin compared with differentiated cells: lessheterochromatin, more loosely bound (or hyperdynamic) archi-tectural chromatin proteins, less H3K9 methylation, and globaltranscriptional hyperactivity. Upon differentiation, the transcrip-tional program needs to be rapidly switched—possibly mediatedby the presence of both activating and repressive chromatin marks(so-called bivalent domains) on lineage-specific developmentalregulators—in a process whereby these regulators are silencedand at the same time poised for activation. In addition, embry-onic stem cells are sensitive to reduced levels of key structuralcomponents of chromatin (cohesin and condensing complexes).
expression in the shoot and root, respectively,whereas CLE41p positively affects WOX4 ex-pression in the vasculature (52, 91, 112, 115,124). Second, the sources and sinks of the CLEpeptides differ: In the shoot meristem, CLV3psignals from the stem cells to the OC, whereasin the root and in the vasculature, differenti-ated stem cell daughters signal back to the nicheor stem cells (36, 96, 115, 124). Thus, withoutfurther studies, we cannot determine whetherthe repeated use of CLE/WOX modules is co-incidental or reflects adaptations of an ancientstem-cell-regulating mechanism.
There are more striking differences in thestem cell niches. Cytokinin signaling is requiredin the shoot meristem and vascular stem cells,whereas auxin is important for root stem cellmaintenance, as discussed above. In addition,HD-ZIPIII activity needs to be kept high in theshoot meristem (34, 121, 136), but it promotesvascular xylem differentiation (6, 21, 55) andrepresses root development (47).
A yet unanswered question is whetherstem cells have specific stemness factors thatmake them pluripotent, or are simply any kind
www.annualreviews.org • Plant Stem Cell Niches 629
Ann
u. R
ev. P
lant
Bio
l. 20
12.6
3:61
5-63
6. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by U
nive
rsid
ad A
uton
oma
de Q
uere
taro
on
10/2
4/13
. For
per
sona
l use
onl
y.

PP63CH25-Laux ARI 27 March 2012 11:48
of cell that divides and is blocked from thenext step of differentiation. Mutant analysishighlighted that chromatin factors and genomeorganization factors are pivotal for stem cellmaintenance, and for most of them, it has beenshown that they regulate key stem cell regula-tors (like WUS, WOX5, or PLT) (2, 8, 11, 45,62, 68, 71). Thus, shoot and root plant stemcells might contain a chromatin state distinctfrom that of differentiated cells, a situationthat is similar to animal stem cells (see sidebar,Chromatin States in Animal Stem Cells). In-depth understanding of the molecular nature ofplant stem cells also profits from transcriptionalprofiling experiments. The cambial stem cells
in Populus are characterized by signal trans-duction and transcriptional regulation factors(116). The transcription profile of the rootcortex/endodermis stem cells shows high ex-pression of the G2/M-phase-specific genes, andit was shown that cyclinD6;1 has a role in theasymmetric division of these stem cells (122).Shoot stem cells have an overrepresentation oftranscripts encoding factors involved in DNAmetabolism, DNA replication and repair, chro-mosome organization, and biogenesis (139). Toelucidate whether there is a signature commonto all plant stem cells, we will have to awaita systematic comparison of transcriptionalprofiles.
DISCLOSURE STATEMENT
The authors are not aware of any affiliations, memberships, funding, or financial holdings thatmight be perceived as affecting the objectivity of this review.
ACKNOWLEDGMENTS
We thank Leron Katsir for critical comments on the manuscript. We apologize to colleagueswhose work could not be cited owing to space constraints. Work in our laboratory is supportedby grants from the DFG to T.L. as part of the SFB592 program, from the European Union aspart of the ERA-PG program, and from the BMBF as part of the FRISYS program, as well as anEMBO postdoctoral fellowship to N.K.
LITERATURE CITED
1. Agusti J, Lichtenberger R, Schwarz M, Nehlin L, Greb T. 2011. Characterization of transcriptomeremodeling during cambium formation identifies MOL1 and RUL1 as opposing regulators of secondarygrowth. PLoS Genet. 7:e1001312
2. Aichinger E, Villar CB, Di Mambro R, Sabatini S, Kohler C. 2011. The CHD3 chromatin remodelerPICKLE and Polycomb group proteins antagonistically regulate meristem activity in the Arabidopsisroot. Plant Cell 23:1047–60
3. Aida M, Beis D, Heidstra R, Willemsen V, Blilou I, et al. 2004. The PLETHORA genes mediatepatterning of the Arabidopsis root stem cell niche. Cell 119:109–20
4. Argueso CT, Raines T, Kieber JJ. 2010. Cytokinin signaling and transcriptional networks. Curr. Opin.Plant Biol. 13:533–39
5. Argyros RD, Mathews DE, Chiang YH, Palmer CM, Thibault DM, et al. 2008. Type B responseregulators of Arabidopsis play key roles in cytokinin signaling and plant development. Plant Cell 20:2102–16
6. Baima S, Possenti M, Matteucci A, Wisman E, Altamura MM, et al. 2001. The Arabidopsis ATHB-8HD-zip protein acts as a differentiation-promoting transcription factor of the vascular meristems. PlantPhysiol. 126:643–55
7. Barlow PW, Macdonald PDM. 1973. An analysis of the mitotic cell cycle in the root meristem of Zeamays. Proc. R. Soc. Lond. B. 183:385–98
630 Aichinger et al.
Ann
u. R
ev. P
lant
Bio
l. 20
12.6
3:61
5-63
6. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by U
nive
rsid
ad A
uton
oma
de Q
uere
taro
on
10/2
4/13
. For
per
sona
l use
onl
y.

PP63CH25-Laux ARI 27 March 2012 11:48
8. Barrero JM, Gonzalez-Bayon R, del Pozo JC, Ponce MR, Micol JL. 2007. INCURVATA2 encodes thecatalytic subunit of DNA Polymerase alpha and interacts with genes involved in chromatin-mediatedcellular memory in Arabidopsis thaliana. Plant Cell 19:2822–38
9. Barton MK. 2010. Twenty years on: the inner workings of the shoot apical meristem, a developmentaldynamo. Dev. Biol. 341:95–113
10. Baum SF, Dubrovsky JG, Rost TL. 2002. Apical organization and maturation of the cortex and vascularcylinder in Arabidopsis thaliana (Brassicaceae) roots. Am. J. Bot. 89:908–20
11. Bertrand C, Bergounioux C, Domenichini S, Delarue M, Zhou DX. 2003. Arabidopsis histoneacetyltransferase AtGCN5 regulates the floral meristem activity through the WUSCHEL/AGAMOUSpathway. J. Biol. Chem. 278:28246–51
12. Bishopp A, Help H, El-Showk S, Weijers D, Scheres B, et al. 2011. A mutually inhibitory interactionbetween auxin and cytokinin specifies vascular pattern in roots. Curr. Biol. 21:917–26
13. Bishopp A, Lehesranta S, Vaten A, Help H, El-Showk S, et al. 2011. Phloem-transported cytokininregulates polar auxin transport and maintains vascular pattern in the root meristem. Curr. Biol. 21:927–32
14. Blilou I, Xu J, Wildwater M, Willemsen V, Paponov I, et al. 2005. The PIN auxin efflux facilitatornetwork controls growth and patterning in Arabidopsis roots. Nature 433:39–44
15. Bosca S, Knauer S, Laux T. 2011. Embryonic development in Arabidopsis thaliana: from the zygotedivision to the shoot meristem. Front. Plant Physiol. 2:93
16. Brand U, Fletcher JC, Hobe M, Meyerowitz EM, Simon R. 2000. Dependence of stem cell fate inArabidopsis on a feedback loop regulated by CLV3 activity. Science 289:617–19
17. Brand U, Grunewald M, Hobe M, Simon R. 2002. Regulation of CLV3 expression by two homeoboxgenes in Arabidopsis. Plant Physiol. 129:565–75
18. Braybrook SA, Kuhlemeier C. 2010. How a plant builds leaves. Plant Cell 22:1006–1819. Busch W, Miotk A, Ariel FD, Zhao Z, Forner J, et al. 2010. Transcriptional control of a plant stem cell
niche. Dev. Cell 18:849–6120. Byrne ME, Barley R, Curtis M, Arroyo JM, Dunham M, et al. 2000. Asymmetric leaves1 mediates leaf
patterning and stem cell function in Arabidopsis. Nature 408:967–7121. Carlsbecker A, Lee JY, Roberts CJ, Dettmer J, Lehesranta S, et al. 2010. Cell signalling by mi-
croRNA165/6 directs gene dose-dependent root cell fate. Nature 465:316–2122. Casamitjana-Martinez E, Hofhuis HF, Xu J, Liu CM, Heidstra R, Scheres B. 2003. Root-specific CLE19
overexpression and the sol1/2 suppressors implicate a CLV-like pathway in the control of Arabidopsis rootmeristem maintenance. Curr. Biol. 13:1435–41
23. Chen H, Banerjee AK, Hannapel DJ. 2004. The tandem complex of BEL and KNOX partners is requiredfor transcriptional repression of ga20ox1. Plant J. 38:276–84
24. Chitwood DH, Timmermans MC. 2010. Small RNAs are on the move. Nature 467:415–1925. Clark SE. 1997. Organ formation at the vegetative shoot meristem. Plant Cell 9:1067–7626. Clark SE, Running MP, Meyerowitz EM. 1993. CLAVATA1, a regulator of meristem and flower devel-
opment in Arabidopsis. Development 119:397–41827. Clark SE, Running MP, Meyerowitz EM. 1995. CLAVATA3 is a specific regulator of shoot and floral
meristem development affecting the same processes as CLAVATA1. Development 121:2057–6728. Clowes FAL. 1971. The proportion of cells that divide in root meristems of Zea mays L. Ann. Bot.
35:249–6129. Cock JM, McCormick S. 2001. A large family of genes that share homology with CLAVATA3. Plant
Physiol. 126:939–4230. Cui H, Levesque MP, Vernoux T, Jung JW, Paquette AJ, et al. 2007. An evolutionarily conserved
mechanism delimiting SHR movement defines a single layer of endodermis in plants. Science 316:421–2531. De Smet I, Vassileva V, De Rybel B, Levesque MP, Grunewald W, et al. 2008. Receptor-like kinase
ACR4 restricts formative cell divisions in the Arabidopsis root. Science 322:594–9732. Ding Z, Friml J. 2010. Auxin regulates distal stem cell differentiation in Arabidopsis roots. Proc. Natl.
Acad. Sci. USA 107:12046–5133. Elo A, Immanen J, Nieminen K, Helariutta Y. 2009. Stem cell function during plant vascular develop-
ment. Semin. Cell Dev. Biol. 20:1097–106
www.annualreviews.org • Plant Stem Cell Niches 631
Ann
u. R
ev. P
lant
Bio
l. 20
12.6
3:61
5-63
6. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by U
nive
rsid
ad A
uton
oma
de Q
uere
taro
on
10/2
4/13
. For
per
sona
l use
onl
y.

PP63CH25-Laux ARI 27 March 2012 11:48
34. Emery JF, Floyd SK, Alvarez J, Eshed Y, Hawker NP, et al. 2003. Radial patterning of Arabidopsis shootsby class III HD-ZIP and KANADI genes. Curr. Biol. 13:1768–74
35. Endrizzi K, Moussian B, Haecker A, Levin J, Laux T. 1996. The SHOOT MERISTEMLESS gene isrequired for maintenance of undifferentiated cells in Arabidopsis shoot and floral meristems and acts at adifferent regulatory level than the meristem genes WUSCHEL and ZWILLE. Plant J. 10:967–79
36. Etchells JP, Turner SR. 2010. The PXY-CLE41 receptor ligand pair defines a multifunctional pathwaythat controls the rate and orientation of vascular cell division. Development 137:767–74
37. Feldman LJ, Torrey JG. 1976. The isolation and culture in vitro of the quiescent center of Zea mays.Am. J. Bot. 63:345–55
38. Fisher K, Turner S. 2007. PXY, a receptor-like kinase essential for maintaining polarity during plantvascular-tissue development. Curr. Biol. 17:1061–66
39. Fletcher JC, Brand U, Running MP, Simon R, Meyerowitz EM. 1999. Signaling of cell fate decisionsby CLAVATA3 in Arabidopsis shoot meristems. Science 283:1911–14
40. Frugis G, Giannino D, Mele G, Nicolodi C, Chiappetta A, et al. 2001. Overexpression of KNAT1 in let-tuce shifts leaf determinate growth to a shoot-like indeterminate growth associated with an accumulationof isopentenyl-type cytokinins. Plant Physiol. 126:1370–80
41. Fulcher N, Sablowski R. 2009. Hypersensitivity to DNA damage in plant stem cell niches. Proc. Natl.Acad. Sci. USA 106:20984–88
42. Galinha C, Hofhuis H, Luijten M, Willemsen V, Blilou I, et al. 2007. PLETHORA proteins as dose-dependent master regulators of Arabidopsis root development. Nature 449:1053–57
43. Gaspar-Maia A, Alajem A, Meshorer E, Ramalho-Santos M. 2011. Open chromatin in pluripotency andreprogramming. Nat. Rev. Mol. Cell Biol. 12:36–47
44. Gordon SP, Chickarmane VS, Ohno C, Meyerowitz EM. 2009. Multiple feedback loops through cy-tokinin signaling control stem cell number within the Arabidopsis shoot meristem. Proc. Natl. Acad. Sci.USA 106:16529–34
45. Graf P, Dolzblasz A, Wurschum T, Lenhard M, Pfreundt U, Laux T. 2010. MGOUN1 encodes anArabidopsis type IB DNA topoisomerase required in stem cell regulation and to maintain developmentallyregulated gene silencing. Plant Cell 22:716–28
46. Grieneisen VA, Xu J, Maree AF, Hogeweg P, Scheres B. 2007. Auxin transport is sufficient to generatea maximum and gradient guiding root growth. Nature 449:1008–13
47. Grigg SP, Galinha C, Kornet N, Canales C, Scheres B, Tsiantis M. 2009. Repression of apical homeoboxgenes is required for embryonic root development in Arabidopsis. Curr. Biol. 19:1485–90
48. Guo Y, Han L, Hymes M, Denver R, Clark SE. 2010. CLAVATA2 forms a distinct CLE-bindingreceptor complex regulating Arabidopsis stem cell specification. Plant J. 63:889–900
49. Haecker A, Gross-Hardt R, Geiges B, Sarkar A, Breuninger H, et al. 2004. Expression dynamics of WOXgenes mark cell fate decisions during early embryonic patterning in Arabidopsis thaliana. Development131:657–68
50. Hake S, Vollbrecht E, Freeling M. 1989. Cloning Knotted, the dominant morphological mutant in maizeusing Ds2 as a transposon tag. EMBO J. 8:15–22
51. Helariutta Y, Fukaki H, Wysocka-Diller J, Nakajima K, Jung J, et al. 2000. The SHORT-ROOT genecontrols radial patterning of the Arabidopsis root through radial signaling. Cell 101:555–67
52. Hirakawa Y, Kondo Y, Fukuda H. 2010. TDIF peptide signaling regulates vascular stem cell proliferationvia the WOX4 homeobox gene in Arabidopsis. Plant Cell 22:2618–29
53. Hirakawa Y, Shinohara H, Kondo Y, Inoue A, Nakanomyo I, et al. 2008. Non-cell-autonomous controlof vascular stem cell fate by a CLE peptide/receptor system. Proc. Natl. Acad. Sci. USA 105:15208–13
54. Hobe M, Muller R, Grunewald M, Brand U, Simon R. 2003. Loss of CLE40, a protein functionallyequivalent to the stem cell restricting signal CLV3, enhances root waving in Arabidopsis. Dev. Genes Evol.213:371–81
55. Ilegems M, Douet V, Meylan-Bettex M, Uyttewaal M, Brand L, et al. 2010. Interplay of auxin, KANADIand Class III HD-ZIP transcription factors in vascular tissue formation. Development 137:975–84
56. Ishida K, Yamashino T, Yokoyama A, Mizuno T. 2008. Three type-B response regulators, ARR1, ARR10and ARR12, play essential but redundant roles in cytokinin signal transduction throughout the life cycleof Arabidopsis thaliana. Plant Cell Physiol. 49:47–57
632 Aichinger et al.
Ann
u. R
ev. P
lant
Bio
l. 20
12.6
3:61
5-63
6. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by U
nive
rsid
ad A
uton
oma
de Q
uere
taro
on
10/2
4/13
. For
per
sona
l use
onl
y.

PP63CH25-Laux ARI 27 March 2012 11:48
57. Ito Y, Nakanomyo I, Motose H, Iwamoto K, Sawa S, et al. 2006. Dodeca-CLE peptides as suppressorsof plant stem cell differentiation. Science 313:842–45
58. Jasinski S, Piazza P, Craft J, Hay A, Woolley L, et al. 2005. KNOX action in Arabidopsis is mediated bycoordinate regulation of cytokinin and gibberellin activities. Curr. Biol. 15:1560–65
59. Jiang K, Feldman LJ. 2005. Regulation of root apical meristem development. Annu. Rev. Cell Dev. Biol.21:485–509
60. Jiang K, Meng YL, Feldman LJ. 2003. Quiescent center formation in maize roots is associated with anauxin-regulated oxidizing environment. Development 130:1429–38
61. Katsir L, Davies KA, Bergmann DC, Laux T. 2011. Peptide signaling in plant development. Curr. Biol.21:R356–64
62. Kaya H, Shibahara KI, Taoka KI, Iwabuchi M, Stillman B, Araki T. 2001. FASCIATA genes for chromatinassembly factor-1 in Arabidopsis maintain the cellular organization of apical meristems. Cell 104:131–42
63. Kidner C, Sundaresan V, Roberts K, Dolan L. 2000. Clonal analysis of the Arabidopsis root confirms thatposition, not lineage, determines cell fate. Planta 211:191–99
64. Kinoshita A, Betsuyaku S, Osakabe Y, Mizuno S, Nagawa S, et al. 2010. RPK2 is an essential receptor-likekinase that transmits the CLV3 signal in Arabidopsis. Development 137:3911–20
65. Ko JH, Han KH. 2004. Arabidopsis whole-transcriptome profiling defines the features of coordinatedregulations that occur during secondary growth. Plant Mol. Biol. 55:433–53
66. Ko JH, Han KH, Park S, Yang J. 2004. Plant body weight-induced secondary growth in Arabidopsis andits transcription phenotype revealed by whole-transcriptome profiling. Plant Physiol. 135:1069–83
67. Kondo Y, Hirakawa Y, Kieber JJ, Fukuda H. 2011. CLE peptides can negatively regulate protoxylemvessel formation via cytokinin signaling. Plant Cell Physiol. 52:37–48
68. Kornet N, Scheres B. 2009. Members of the GCN5 histone acetyltransferase complex regulatePLETHORA-mediated root stem cell niche maintenance and transit amplifying cell proliferation inArabidopsis. Plant Cell 21:1070–79
69. Kurakawa T, Ueda N, Maekawa M, Kobayashi K, Kojima M, et al. 2007. Direct control of shoot meristemactivity by a cytokinin-activating enzyme. Nature 445:652–55
70. Kuroha T, Ueguchi C, Sakakibara H, Satoh S. 2006. Cytokinin receptors are required for normaldevelopment of auxin-transporting vascular tissues in the hypocotyl but not in adventitious roots.Plant Cell Physiol. 47:234–43
71. Kwon CS, Chen C, Wagner D. 2005. WUSCHEL is a primary target for transcriptional regulation bySPLAYED in dynamic control of stem cell fate in Arabidopsis. Genes Dev. 19:992–1003
72. Lau S, Slane D, Herud O, Kong J, Jurgens G. 2012. Early embryogenesis in flowering plants: setting upthe basic body pattern. Annu. Rev. Plant Biol. 63:483–506
73. Laux T. 2003. The stem cell concept in plants: a matter of debate. Cell 113:281–8374. Laux T, Mayer KF, Berger J, Jurgens G. 1996. The WUSCHEL gene is required for shoot and floral
meristem integrity in Arabidopsis. Development 122:87–9675. Leibfried A, To JP, Busch W, Stehling S, Kehle A, et al. 2005. WUSCHEL controls meristem function
by direct regulation of cytokinin-inducible response regulators. Nature 438:1172–7576. Lenhard M, Jurgens G, Laux T. 2002. The WUSCHEL and SHOOTMERISTEMLESS genes fulfil
complementary roles in Arabidopsis shoot meristem regulation. Development 129:3195–20677. Lenhard M, Laux T. 2003. Stem cell homeostasis in the Arabidopsis shoot meristem is regulated by
intercellular movement of CLAVATA3 and its sequestration by CLAVATA1. Development 130:3163–7378. Levesque MP, Vernoux T, Busch W, Cui H, Wang JY, et al. 2006. Whole-genome analysis of the
SHORT-ROOT developmental pathway in Arabidopsis. PLoS Biol. 4:e14379. Liu C, Thong Z, Yu H. 2009. Coming into bloom: the specification of floral meristems. Development
136:3379–9180. Liu Q, Yao X, Pi L, Wang H, Cui X, Huang H. 2009. The ARGONAUTE10 gene modulates shoot
apical meristem maintenance and leaf polarity establishment by repressing miR165/166 in Arabidopsis.Plant J. 58:27–40
81. Loiseau JE. 1959. Observation et experimentation sur la phyllotaxie et le fonctionnement du sommetvegetatif chez quelques Balsaminacees. Ann. Sci. Nat. Bot. Ser. 11:201–14
www.annualreviews.org • Plant Stem Cell Niches 633
Ann
u. R
ev. P
lant
Bio
l. 20
12.6
3:61
5-63
6. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by U
nive
rsid
ad A
uton
oma
de Q
uere
taro
on
10/2
4/13
. For
per
sona
l use
onl
y.

PP63CH25-Laux ARI 27 March 2012 11:48
82. Long JA, Barton MK. 1998. The development of apical embryonic pattern in Arabidopsis. Development125:3027–35
83. Long JA, Moan EI, Medford JI, Barton MK. 1996. A member of the KNOTTED class of homeodomainproteins encoded by the STM gene of Arabidopsis. Nature 379:66–69
84. Lyndon RF. 1998. The Shoot Apical Meristem: Its Growth and Development. Cambridge: Cambridge Univ.Press
85. Mahonen AP, Bishopp A, Higuchi M, Nieminen KM, Kinoshita K, et al. 2006. Cytokinin signaling andits inhibitor AHP6 regulate cell fate during vascular development. Science 311:94–98
86. Mahonen AP, Bonke M, Kauppinen L, Riikonen M, Benfey PN, Helariutta Y. 2000. A novel two-component hybrid molecule regulates vascular morphogenesis of the Arabidopsis root. Genes Dev.14:2938–43
87. Mahonen AP, Higuchi M, Tormakangas K, Miyawaki K, Pischke MS, et al. 2006. Cytokinins regulatea bidirectional phosphorelay network in Arabidopsis. Curr. Biol. 16:1116–22
88. Mallory AC, Hinze A, Tucker MR, Bouche N, Gasciolli V, et al. 2009. Redundant and specific roles ofthe ARGONAUTE proteins AGO1 and ZLL in development and small RNA-directed gene silencing.PLoS Genet. 5:e1000646
89. Matsumoto-Kitano M, Kusumoto T, Tarkowski P, Kinoshita-Tsujimura K, Vaclavikova K, et al. 2008.Cytokinins are central regulators of cambial activity. Proc. Natl. Acad. Sci. USA 105:20027–31
90. Matsuzaki Y, Ogawa-Ohnishi M, Mori A, Matsubayashi Y. 2010. Secreted peptide signals required formaintenance of root stem cell niche in Arabidopsis. Science 329:1065–67
91. Mayer KF, Schoof H, Haecker A, Lenhard M, Jurgens G, Laux T. 1998. Role of WUSCHEL inregulating stem cell fate in the Arabidopsis shoot meristem. Cell 95:805–15
92. McConnell JR, Barton MK. 1995. Effects of mutations in the PINHEAD gene of Arabidopsis on theformation of shoot apical meristems. Dev. Genet. 16:358–66
93. McConnell JR, Emery J, Eshed Y, Bao N, Bowman J, Barton MK. 2001. Role of PHABULOSA andPHAVOLUTA in determining radial patterning in shoots. Nature 411:709–13
94. Mordhorst AP, Hartog MV, El Tamer MK, Laux T, de Vries SC. 2002. Somatic embryogenesis fromArabidopsis shoot apical meristem mutants. Planta 214:829–36
95. Moussian B, Schoof H, Haecker A, Jurgens G, Laux T. 1998. Role of the ZWILLE gene in the regulationof central shoot meristem cell fate during Arabidopsis embryogenesis. EMBO J. 17:1799–809
96. Muller R, Borghi L, Kwiatkowska D, Laufs P, Simon R. 2006. Dynamic and compensatory responses ofArabidopsis shoot and floral meristems to CLV3 signaling. Plant Cell 18:1188–98
97. Nakajima K, Sena G, Nawy T, Benfey P. 2001. Intercellular movement of the putative transcriptionfactor SHR in root patterning. Nature 413:307–11
98. Newman IV. 1965. Patterns in the meristems of vascular plants. III. Pursuing the patterns where no cellis a permanent cell. J. Linn. Soc. Bot. 59:185–214
99. Nimchuk ZL, Tarr PT, Meyerowitz EM. 2011. An evolutionarily conserved pseudokinase mediates stemcell production in plants. Plant Cell 23:851–54
100. Nimchuk ZL, Tarr PT, Ohno C, Qu X, Meyerowitz EM. 2011. Plant stem cell signaling involvesligand-dependent trafficking of the CLAVATA1 receptor kinase. Curr. Biol. 21:345–52
101. Orkin SH, Hochedlinger K. 2011. Chromatin connections to pluripotency and cellular reprogramming.Cell 145:835–50
102. Ortega-Martinez O, Pernas M, Carol RJ, Dolan L. 2007. Ethylene modulates stem cell division in theArabidopsis thaliana root. Science 317:507–10
103. Perilli S, Moubayidin L, Sabatini S. 2010. The molecular basis of cytokinin function. Curr. Opin. PlantBiol. 13:21–26
104. Petersson SV, Johansson AI, Kowalczyk M, Makoveychuk A, Wang JY, et al. 2009. An auxin gradientand maximum in the Arabidopsis root apex shown by high-resolution cell-specific analysis of IAA distri-bution and synthesis. Plant Cell 21:1659–68
105. Prigge MJ, Otsuga D, Alonso JM, Ecker JR, Drews GN, Clark SE. 2005. Class III homeodomain-leucinezipper gene family members have overlapping, antagonistic, and distinct roles in Arabidopsis development.Plant Cell 17:61–76
634 Aichinger et al.
Ann
u. R
ev. P
lant
Bio
l. 20
12.6
3:61
5-63
6. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by U
nive
rsid
ad A
uton
oma
de Q
uere
taro
on
10/2
4/13
. For
per
sona
l use
onl
y.

PP63CH25-Laux ARI 27 March 2012 11:48
106. Raven JA, Edwards D. 2001. Roots: evolutionary origins and biogeochemical significance. J. Exp. Bot.52:381–401
107. Reddy GV, Meyerowitz EM. 2005. Stem-cell homeostasis and growth dynamics can be uncoupled inthe Arabidopsis shoot apex. Science 310:663–67
108. Reinhardt D, Frenz M, Mandel T, Kuhlemeier C. 2003. Microsurgical and laser ablation analysis ofinteractions between the zones and layers of the tomato shoot apical meristem. Development 130:4073–83
109. Sabatini S, Beis D, Wolkenfelt H, Murfett J, Guilfoyle T, et al. 1999. An auxin-dependent distal organizerof pattern and polarity in the Arabidopsis root. Cell 99:463–72
110. Sabatini S, Heidstra R, Wildwater M, Scheres B. 2003. SCARECROW is involved in positioning thestem cell niche in the Arabidopsis root meristem. Genes Dev. 17:354–58
111. Sakamoto T, Kamiya N, Ueguchi-Tanaka M, Iwahori S, Matsuoka M. 2001. KNOX homeodomainprotein directly suppresses the expression of a gibberellin biosynthetic gene in the tobacco shoot apicalmeristem. Genes Dev. 15:581–90
112. Sarkar A, Luijten M, Miyashima S, Lenhard M, Hashimoto T, et al. 2007. Conserved factors regulatesignalling in Arabidopsis thaliana shoot and root stem cell organizers. Nature 446:811–14
113. Satina S, Blakeslee AF, Avery A. 1940. Demonstration of three germ layers in the shoot apex of Daturaby means of induced polyploidy in periclinal chimeras. Am. J. Bot. 27:895–905
114. Schofield R. 1978. The relationship between the spleen colony-forming cell and the haemopoietic stemcell. Blood Cells 4:7–25
115. Schoof H, Lenhard M, Haecker A, Mayer KF, Jurgens G, Laux T. 2000. The stem cell population ofArabidopsis shoot meristems is maintained by a regulatory loop between the CLAVATA and WUSCHELgenes. Cell 100:635–44
116. Schrader J, Moyle R, Bhalerao R, Hertzberg M, Lundeberg J, et al. 2004. Cambial meristem dormancyin trees involves extensive remodelling of the transcriptome. Plant J. 40:173–87
117. Sehr EM, Agusti J, Lehner R, Farmer EE, Schwarz M, Greb T. 2010. Analysis of secondary growthin the Arabidopsis shoot reveals a positive role of jasmonate signalling in cambium formation. Plant J.63:811–22
118. Sena G, Wang X, Liu HY, Hofhuis H, Birnbaum KD. 2009. Organ regeneration does not require afunctional stem cell niche in plants. Nature 457:1150–53
119. Shani E, Yanai O, Ori N. 2006. The role of hormones in shoot apical meristem function. Curr. Opin.Plant Biol. 9:484–89
120. Smith LG, Breene B, Veit B, Hake S. 1992. A dominant mutation in the maize homeobox gene, Knotted-1,causes its ectopic expression in leaf cells with altered fates. Development 116:21–30
121. Smith ZR, Long JA. 2010. Control of Arabidopsis apical-basal embryo polarity by antagonistic transcrip-tion factors. Nature 464:423–26
122. Sozzani R, Cui H, Moreno-Risueno MA, Busch W, Van Norman JM, et al. 2010. Spatiotemporalregulation of cell-cycle genes by SHORTROOT links patterning and growth. Nature 466:128–32
123. Spradling A, Drummond-Barbosa D, Kai T. 2001. Stem cells find their niche. Nature 414:98–104124. Stahl Y, Wink RH, Ingram GC, Simon R. 2009. A signaling module controlling the stem cell niche in
Arabidopsis root meristems. Curr. Biol. 19:909–14125. Stewart RN, Dermen H. 1970. Determination of number and mitotic activity of shoot apical initial cells
by analysis of mericlinal chimeras. Am. J. Bot. 57:816–26126. To JP, Kieber JJ. 2008. Cytokinin signaling: two-components and more. Trends Plant Sci. 13:85–92127. Tucker MR, Hinze A, Tucker EJ, Takada S, Jurgens G, Laux T. 2008. Vascular signalling mediated
by ZWILLE potentiates WUSCHEL function during shoot meristem stem cell development in theArabidopsis embryo. Development 135:2839–43
128. van den Berg C, Willemsen V, Hage W, Weisbeek P, Scheres B. 1995. Cell fate in the Arabidopsis rootmeristem determined by directional signalling. Nature 378:62–65
129. van den Berg C, Willemsen V, Hendriks G, Weisbeek P, Scheres B. 1997. Short-range control of celldifferentiation in the Arabidopsis root meristem. Nature 390:287–89
130. van der Graaff E, Laux T, Rensing SA. 2009. The WUS homeobox-containing (WOX) protein family.Genome Biol. 10:248
www.annualreviews.org • Plant Stem Cell Niches 635
Ann
u. R
ev. P
lant
Bio
l. 20
12.6
3:61
5-63
6. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by U
nive
rsid
ad A
uton
oma
de Q
uere
taro
on
10/2
4/13
. For
per
sona
l use
onl
y.

PP63CH25-Laux ARI 27 March 2012 11:48
131. Wachsman G, Heidstra R, Scheres B. 2011. Distinct cell-autonomous functions ofRETINOBLASTOMA-RELATED in Arabidopsis stem cells revealed by the Brother of Brainbowclonal analysis system. Plant Cell 23:2581–91
132. Waites R, Selvadurai HR, Oliver IR, Hudson A. 1998. The PHANTASTICA gene encodes a MYBtranscription factor involved in growth and dorsoventrality of lateral organs in Antirrhinum. Cell 93:779–89
133. Wang JW, Wang LJ, Mao YB, Cai WJ, Xue HW, Chen XY. 2005. Control of root cap formation bymicroRNA-targeted auxin response factors in Arabidopsis. Plant Cell 17:2204–16
134. Whitford R, Fernandez A, De Groodt R, Ortega E, Hilson P. 2008. Plant CLE peptides from two distinctfunctional classes synergistically induce division of vascular cells. Proc. Natl. Acad. Sci. USA 105:18625–30
135. Wildwater M, Campilho A, Perez-Perez JM, Heidstra R, Blilou I, et al. 2005. The RETINOBLASTOMA-RELATED gene regulates stem cell maintenance in Arabidopsis roots. Cell 123:1337–49
136. Williams L, Fletcher JC. 2005. Stem cell regulation in the Arabidopsis shoot apical meristem. Curr. Opin.Plant Biol. 8:582–86
137. Wisniewska J, Xu J, Seifertova D, Brewer PB, Ruzicka K, et al. 2006. Polar PIN localization directs auxinflow in plants. Science 312:883
138. Xu J, Hofhuis H, Heidstra R, Sauer M, Friml J, Scheres B. 2006. A molecular framework for plantregeneration. Science 311:385–88
139. Yadav RK, Girke T, Pasala S, Xie M, Reddy GV. 2009. Gene expression map of the Arabidopsis shootapical meristem stem cell niche. Proc. Natl. Acad. Sci. USA 106:4941–46
140. Yadav RK, Perales M, Gruel J, Girke T, Jonsson H, Reddy GV. 2011. WUSCHEL protein movementmediates stem cell homeostasis in the Arabidopsis shoot apex. Genes Dev. 25:2025–30
141. Yadav RK, Tavakkoli M, Reddy GV. 2010. WUSCHEL mediates stem cell homeostasis by regulatingstem cell number and patterns of cell division and differentiation of stem cell progenitors. Development137:3581–89
142. Yanai O, Shani E, Dolezal K, Tarkowski P, Sablowski R, et al. 2005. Arabidopsis KNOXI proteins activatecytokinin biosynthesis. Curr. Biol. 15:1566–71
143. Yokoyama A, Yamashino T, Amano Y, Tajima Y, Imamura A, et al. 2007. Type-B ARR transcription fac-tors, ARR10 and ARR12, are implicated in cytokinin-mediated regulation of protoxylem differentiationin roots of Arabidopsis thaliana. Plant Cell Physiol. 48:84–96
144. Yoshida S, Mandel T, Kuhlemeier C. 2011. Stem cell activation by light guides plant organogenesis.Genes Dev. 25:1439–50
145. Young RA. 2011. Control of the embryonic stem cell state. Cell 144:940–54146. Zhang H, Han W, De Smet I, Talboys P, Loya R, et al. 2010. ABA promotes quiescence of the quiescent
centre and suppresses stem cell differentiation in the Arabidopsis primary root meristem. Plant J. 64:764–74
147. Zhang J, Elo A, Helariutta Y. 2011. Arabidopsis as a model for wood formation. Curr. Opin. Biotechnol.22:293–99
148. Zhao C, Craig JC, Petzold HE, Dickerman AW, Beers EP. 2005. The xylem and phloem transcriptomesfrom secondary tissues of the Arabidopsis root-hypocotyl. Plant Physiol. 138:803–18
149. Zhao Z, Andersen SU, Ljung K, Dolezal K, Miotk A, et al. 2010. Hormonal control of the shoot stem-cellniche. Nature 465:1089–92
150. Zhou W, Wei L, Xu J, Zhai Q, Jiang H, et al. 2010. Arabidopsis tyrosylprotein sulfotransferase acts inthe auxin/PLETHORA pathway in regulating postembryonic maintenance of the root stem cell niche.Plant Cell 22:3692–709
151. Zhu H, Hu F, Wang R, Zhou X, Sze SH, et al. 2011. Arabidopsis Argonaute10 specifically sequestersmiR166/165 to regulate shoot apical meristem development. Cell 145:242–56
636 Aichinger et al.
Ann
u. R
ev. P
lant
Bio
l. 20
12.6
3:61
5-63
6. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by U
nive
rsid
ad A
uton
oma
de Q
uere
taro
on
10/2
4/13
. For
per
sona
l use
onl
y.

PP63-FrontMatter ARI 26 March 2012 18:10
Annual Review ofPlant Biology
Volume 63, 2012Contents
There Ought to Be an Equation for ThatJoseph A. Berry � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 1
Photorespiration and the Evolution of C4 PhotosynthesisRowan F. Sage, Tammy L. Sage, and Ferit Kocacinar � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � �19
The Evolution of Flavin-Binding Photoreceptors: An AncientChromophore Serving Trendy Blue-Light SensorsAba Losi and Wolfgang Gartner � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � �49
The Shikimate Pathway and Aromatic Amino Acid Biosynthesisin PlantsHiroshi Maeda and Natalia Dudareva � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � �73
Regulation of Seed Germination and Seedling Growth by ChemicalSignals from Burning VegetationDavid C. Nelson, Gavin R. Flematti, Emilio L. Ghisalberti, Kingsley W. Dixon,
and Steven M. Smith � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 107
Iron Uptake, Translocation, and Regulation in Higher PlantsTakanori Kobayashi and Naoko K. Nishizawa � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 131
Plant Nitrogen Assimilation and Use EfficiencyGuohua Xu, Xiaorong Fan, and Anthony J. Miller � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 153
Vacuolar Transporters in Their Physiological ContextEnrico Martinoia, Stefan Meyer, Alexis De Angeli, and Reka Nagy � � � � � � � � � � � � � � � � � � � � 183
Autophagy: Pathways for Self-Eating in Plant CellsYimo Liu and Diane C. Bassham � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 215
Plasmodesmata Paradigm Shift: Regulation from WithoutVersus WithinTessa M. Burch-Smith and Patricia C. Zambryski � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 239
Small Molecules Present Large Opportunities in Plant BiologyGlenn R. Hicks and Natasha V. Raikhel � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 261
Genome-Enabled Insights into Legume BiologyNevin D. Young and Arvind K. Bharti � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 283
v
Ann
u. R
ev. P
lant
Bio
l. 20
12.6
3:61
5-63
6. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by U
nive
rsid
ad A
uton
oma
de Q
uere
taro
on
10/2
4/13
. For
per
sona
l use
onl
y.

PP63-FrontMatter ARI 26 March 2012 18:10
Synthetic Chromosome Platforms in PlantsRobert T. Gaeta, Rick E. Masonbrink, Lakshminarasimhan Krishnaswamy,
Changzeng Zhao, and James A. Birchler � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 307
Epigenetic Mechanisms Underlying Genomic Imprinting in PlantsClaudia Kohler, Philip Wolff, and Charles Spillane � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 331
Cytokinin Signaling NetworksIldoo Hwang, Jen Sheen, and Bruno Muller � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 353
Growth Control and Cell Wall Signaling in PlantsSebastian Wolf, Kian Hematy, and Herman Hofte � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 381
Phosphoinositide SignalingWendy F. Boss and Yang Ju Im � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 409
Plant Defense Against Herbivores: Chemical AspectsAxel Mithofer and Wilhelm Boland � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 431
Plant Innate Immunity: Perception of Conserved Microbial SignaturesBenjamin Schwessinger and Pamela C. Ronald � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 451
Early Embryogenesis in Flowering Plants: Setting Upthe Basic Body PatternSteffen Lau, Daniel Slane, Ole Herud, Jixiang Kong, and Gerd Jurgens � � � � � � � � � � � � � � 483
Seed Germination and VigorLoıc Rajjou, Manuel Duval, Karine Gallardo, Julie Catusse, Julia Bally,
Claudette Job, and Dominique Job � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 507
A New Development: Evolving Concepts in Leaf OntogenyBrad T. Townsley and Neelima R. Sinha � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 535
Control of Arabidopsis Root DevelopmentJalean J. Petricka, Cara M. Winter, and Philip N. Benfey � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 563
Mechanisms of Stomatal DevelopmentLynn Jo Pillitteri and Keiko U. Torii � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 591
Plant Stem Cell NichesErnst Aichinger, Noortje Kornet, Thomas Friedrich, and Thomas Laux � � � � � � � � � � � � � � � � 615
The Effects of Tropospheric Ozone on Net Primary Productivityand Implications for Climate ChangeElizabeth A. Ainsworth, Craig R. Yendrek, Stephen Sitch, William J. Collins,
and Lisa D. Emberson � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 637
Quantitative Imaging with Fluorescent BiosensorsSakiko Okumoto, Alexander Jones, and Wolf B. Frommer � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 663
vi Contents
Ann
u. R
ev. P
lant
Bio
l. 20
12.6
3:61
5-63
6. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by U
nive
rsid
ad A
uton
oma
de Q
uere
taro
on
10/2
4/13
. For
per
sona
l use
onl
y.