plant respiration: krebs cycle, fermentation, and electron transport system intended to complete...
TRANSCRIPT

MANUSCRIPT
PLANT RESPIRATION KREBS CYCLE FERMENTATION AND
ELECTRON TRANSPORT SYSTEM
Intended to Complete Plant Physiology Subject
ANNISA ADELINA FAJRI
1201304 2012
BIOLOGY EDUCATION (ISTE)
BIOLOGY DEPARTMENT
MATH AND SCIENCE FACULTY
PADANG STATE UNIVERSITY
2013
2
PREFACE
ldquoPlant Respiration Krebs Cycle Fermentation and
Transport Systemrdquo in this manuscript is intended to
complete the task from our lecturer Dr Azwir Anhar
MSi and to meet the needs of university students as
the guidersquos book in learning and practice activity for
the member of International Standard Teacher Education
(ISTE) program in Public University of Padang
My greatest wish is that the readers find this
manuscript helpful in gaining the knowledge they need
for effective learning and practice and in preparing
for examination designed to test that knowledge Reader
comments and suggestions are welcome and have again
been helpful in improving our manuscript Your
contribution may assist many other students in the
coming years
Writer
September 2013
i
__________________________
CONTENT
PREFACE i
CONTENT ii
CHAPTER I INTRODUCTION
A Background 1
B Purposes 2
C Benefits 2
CHAPTER II PLANT RESPIRATION
A Krebs Cycle 3
B Fermentation 8
C Electron Transport System
11
CHAPTER III CLOSING
ii
A Conclution
20
B Suggestion
20
REFERENCE
iii
CHAPTER I
INTRODUCTION
A Background
Connected with photosynthesis a metabolic
process takes place in chloroplast-containing plant
cells which like respiration takes up O2 and
releases CO2 in the light but which contrary to
respiration ceases in the dark This O2CO2 gas
exchange has been called right respiration or
photorespiration The substrate for the
photorespiratory metabolism is again ribulose
biphosphate which can be an acceptor not only for
CO2 but also for O2 By taking up oxygen RuBP is
split into PGA and phospoglycolate The supply of O2
and CO2 regulates the relationship between acceptor
oxidation (photorespiration) and acceptor
carboxylation (photosynthesis) via the enzyme
complex RuBP carboxylaseoxygenase High partial
pressure of O2 favors photorespiration A large
supply of CO2 favors photosynthesis The formation
of phospoglycolate is dependent on the supply of
RuBP via the Calvin cycle the photorespiratory O2
uptake and CO2 release increase with light
intensity
1
The process of glycolate metabolism has not
yet been understood in all details The glycolate is
transported out of the chloroplasts phospoglycolate
and phospate The glycolate is transported out of
the chloroplast into peroxisomes cell compartments
about the size of mitochondria which contain
glycolate oxidase catalase and transaminases In
the peroxysomes when O2 is taken up glycolate is
oxidized to glyoxylate and the peroxide thus
produced is detoxified by catalase Glycolate can
either be completely reduced via oxalate by further
O2 uptake or transformed to glycine by
transamination Glycine is transported from the
peroxisomes into the mitochondria where two
molecules of glycine are coupled to form one
molecule of serine with the release of CO2 Serine
is taken over by the amino acid metabolism or
2
converted to glycerate after deamination by
hydropyruvate This can be photophosporylated in the
chloroplasts and returned to the Calvin cycle or
used elsewhere (Larcher 1980 81-82)
B Purposes
This manuscript is intended to complete the
task from our lecturer Dr Azwir Anhar MSi and
to meet the needs of university students as the
guidersquos book in learning and practice activity for
the member of International Standard Teacher
Education (ISTE) program in Public University of
Padang
C Benefits
Benefits that we gained from this manuscript
are
1 As reference in learning process of plant
phisiology subject especially in plant
respiration title
2 As information about steps in plant
respiration
3
CHAPTER II
PLANT RESPIRATION
A Krebs Cycle
The Krebs cycle was named in honor of the
English biochemist Hans A Krebs who in 1937
proposed a cycle of reactions to explain how pyruvate
breakdown takes place in the breast muscle of
pigeons He called his proposed pathway the citric
acid cycle because citric acid is an important
intermediet Another common name for the same group
of reactions is the Tricarboxuylic Acid (TCA) cycle
a term used because citric and isocitric acids have
three carboxyl groups It was not until the early
1950s that mitochondria capable of carrying out this
cycle were isolated from plant cells
The initial step leading to the Krebs cycle
involves the oxidation and loss of CO2 from pyruvate
and the combination of the remaining 2-carbon acetate
unit with a sulfur-containing compound co-enzyme A
(CoA) forming acetyl CoA This and another
comparable role of CoA in the Krebs cycle are
important reasons why sulfur is an essential element
4
The reaction of pyruvate decarboxylation also
involves a phosporylated form of thiamine (vitamin
B1) as a prostethic group Participation of thiamine
in this section partially explains the essential
function of vitamin B1 in plants and animals Besides
the loss of CO2 two hydrogen atoms are removed from
pyruvic acid during the formation of Acetyl CoA The
enzyme catalyzing the complete reaction is called
pyruvic acid dehydrogenase but it is actually an
organized complex containing numerous copies of three
or four different enzymes The hydrogen atoms removed
are finally accepted by NAD+ yielding NADH
The Krebs cycle accomplishes removal of some of
the electrons from organic acid intermediates and
transfer of these electrons to NAD+ or FAD Notice
that none of dehydrogenase enzyme of the cycle uses
NADP+ as an electron acceptor In fact NADP+ is
usually undetectable in plant mitochondria a
situation opposite to that of chloroplasts where
NADP+ is abundant but where there is much less NAD+
Not only are NADH and FADH2 important products of the
Krebs cycle but one molecule of ATP is formed from
ADP and Pi during the conversion of succinyl coenzyme
A to succinic acid Two additional CO2 molecules are
released in these Krebs cycle reactions so there is
a net loss of both carbon atoms from the incoming
5
acetate of acetyl CoA The release of CO2 in the
Krebs cycle accounts for the product of CO2 in the
summary equation for resspiration but no O2 is
absorbed during any Kreb cycle reaction (Lakitan
2011)
During metabolism the synthesis and breakdown
of different organic compounds takes place through
various pathways like the breakdown and synthesis of
proteins carbohydrates fats and nucleic acids
These different pathways and intermediates are also
responsible for the production of energyKrebrsquos
cycle named after Hans Krebs who began working out
its details in 1930s is a series of reactions in
which the pyruvate from glycolysis is oxidized to Co2
under aerobic conditions Kreb cycle is also known as
citric acid cycle or Tri-carboxylic acid cycle (TCA)
( Galston 1968 76)
6
Steps involved in the process of Kreb cycle are
1 Fate of pyruvic acid
Pyruvic acid can form different compounds by
different pathways ie it can be converted into
lactic acid It can be converted into acetyl
coenzyme A as a result of oxidation
2 Fate of Acetyl CoA
It can either undergo condensation with itself or
its derivatives to form fatty acids having 14 ndash 20
carbons Acetyl CoA can also go through a series
of reactions in Krebs cycle
3 Formation of citrate
Acetyl CoA enzyme condenses with oxaloacetate by
an enzyme citrate synthatase to form citrate with
the release of acetyl CoA If the amount of
7
oxaloacetate is very small then small number of
acetyl CoA would be reacting with oxaloacetate
leaving surplus acetyl CoA to go through another
pathway for the formation of long chain fatty
acids
4 Formation of C is ndash acotinate and iso-citrate
Citrate is changed first into C is ndash acotinate and
then to iso-citrate under the enzyme acotinase
Equilibrium is established between citrate C is
aconitate and iso-citrate It has been observed
that most of the time this equilibrium is shifted
towards the iso-citrate If the concentration of
iso-citrate in increased the formation of citrate
will result which also indicates that the
equilibrium also shift in the reserve direction
5 Formation of oxalosucinate
Iso-citrate is acted upon by an enzyme iso-citrate
dehydrogenase using nicotinamide adenine
dinucleotide (NAD) as coenzyme As a result iso-
citrate is converted into oxalosucciate and NAD is
reduced to NADH2 Similar reaction is carried out
by same enzyme using nicotindmide adenine
dinucleotide phosphate (NADP) as coenzyme which is
reduced to NDAPH2
6 Formation of α-Ketoglutarate
8
Oxalosucciante is changed into α-ketoglutrate by
iso-citrate dehydrogenase with the help of
coenzyme NAD or NADP In this reaction carbon
dioxide and NADH2 or NADPH2 are also released
7 Formation of succinyl CoA
α-ketoglutarate combines with acetyl CoA in the
presence of coenzyme NAD and enzyme α-
keoglutaratedehyrogenase to form succinyl coenzyme
a carbon dioxide and NADH2
8 Formation of Succinate
Later on coenzyme a is removed from succinyl
coenzyme A in the presence of guanosinediphosphate
(GDP) and inorganic phosphate to form succinate
and guanosine triphosphate (GTP) This reaction is
carried out by an enzyme called succinyl CoA
synthatase
9 Formation of Funarate
An enzyme succinic dehydrogenase removes hydrogen
from succinate to form funarate
10 Formation of Malate
Fumarate reacts with water in the presence of
enzyme fumarase to form malate
11 Regeneration of oxaloacetate
Malate is oxidized by malic dehydrogenase and NAD
forming oxaloacetate and NADH2 Thus oxalo acetate
is again available to start another cycle
9
(httpkrebs-cycle-or-citric-acid-cycle-or-
trihtml)
Importance of CTA Cycle
1 Source of energy
In addition to routine organic compounds described
above by products like nicotinanide adenine
dinucleotide (NADH2) and guanosine triphosphate
(GTP) are the source of biological energy NADH2
after oxidation produce energy whereas GTP is
itself high energy phosphate compound
2 Oxidation of organic compounds taken as food
Oxidation of fats carbohydrates and proteins take
place through it or in other words it can be said
that oxidation of all compounds having carbon
atoms can take place through TCA cycle Some of
amino acids like alanine glutamic acid and
aspartic acid at one stage or the other enters
into TCA cycle eg glutamic acid enters cycle
after its transformation into α-ketoglutarate
Similarly alanine enters the cycle after its
conversion into pyruvate
3 Intermediate compounds
TCA cycle is also involved in synthesis of
intermediate compounds leading to the formation of
larger molecules (Ray 1963 17)
10
Verification of Krebs Cycle
It was done by radioactive traces like C14 as
radioactive carbon dioxide of different levels and
reactions After addition of radioactive carbon
dioxide different chemical compounds produced like
glucose fats amino acids were isolated and looked
for radioactive carbon In this way whole of
metabolic reaction were verified including individual
reactions alternative metabolic pathways
intermediates of fats carbohydrates and amino acids
etc in the body cells as well as in test tubes
(Wilson 1962 99)
The primary function of the Krebs cycle are as
follows
1 Reduction of NAD+ and FAD to the electron donors
NADH and FADH2 that are subsequently oxidized to
yield ATP
2 Direct synthesis of a limited amount of ATP (1 ATP
for each pyruvate oxidized)
3 Formation of carbon skeletons that can be used to
synthesize certain amino acids that in turn are
converted into larger molecules
11
B Fermentation
Although glycolysis can function well without
O2 the further oxidation of pyruvate and NADH by
mitochondria requires this gas Thus when O2 is
limiting NADH and pyruvate begin to accumulate
Under this condition plants carry out fermentation
(anaerobic respiration) forming either ethanol or
lactic acid (usually ethanol) As shown in the
picture below the two top reactions consists of a
decarboxylation to form acetaldehide then rapid
reduction of acetaldehide by NADH to form ethanol
These reactions are catalyzed by alcohol
dehydrogenase Some cells contain lactic acid
12
dehydrogenase which uses NADH to reduce pyruvic to
lactic acid Thus either ethanol or lactic acid or
both are fermentation products depending on the
activities of each dehydrogenase present In each
case NADH is the reductant and only under
anaerobic conditions is it abundant anough to cause
reduction Furthermore in some plants NADH is used
to cause accumulation of the other compounds when O2
is limiting especially malate and glycerol
(Salisbury 1969 233)
The fate of pyruvate produced by glycolysis
depends primarily on whether or not oxygen is
present Under normal aerobic conditions pyruvate
is transported into the mitochondrion where it is
further oxidized to CO2 and water transferring its
electrons ultimately to molecular oxygen
13
Although higher plants are obligate aerobs and
are able to tolerate anoxia for only short periods
tissues or organs are occasionally subjected to
anaerobic conditions A typical siuation is that of
roots when the soil is saturated with water When
there is no oxygen to serve as the terminal electron
acceptor mitochondrial respiration will shut down
and metabolism will shift over to fermentation
Fermentation converts pyruvate to either ethanol
through the action of the enzyme alcohol
dehydrogenase (ADH) or lactate via lactate
dehydrogenase (LDH) In most plants the principal
products of fermentation are CO2 and ethanol Some
lactate may be formed primarily in the early stages
of anoxia However lactate lowers he pH of the
cytosol which in turn activates pyruvate
decarboxylase and initiates the production of
ethanol
Either one of the fermentation reactions
consumes the NADH produced earlier in glycolysis by
the oxidation of glyceraldehide-3-P Although this
means there is no net gain of reducing potential in
fermentation This recycling of NADH is still
important to the cell The poolof NADH plus NAD+ in
the cell is relatively small and if the NADH is not
recycled there will be no supply of NAD+ to support
14
the continued oxidation of glyceraldehyde-3-P if
this were the case glycolysis and the producton of
even the small quantities of ATP necessary to
maintain the cells under anaerobic conditions would
then grind to a halt (Hopkins 2009 241)
Soil waterlogging has long been identified as a
major abiotic stress and the constraints that it
provides the roots have a significant effect on the
growth and development of plants When this event
occurs in the spring then these puddles can reduce
seed germination and seedling development Thus a
pool of water is an important factor that affects
the growth development and survival of plant
species not only in natural ecosystems but also
the agricultural and horticultural systems (Dat et
al 2006)
After flooding rapid changes in soil
properties At the time of pore water meets the
land the air pushed out gas diffusion is reduced
and toxic compounds accumulate due to anaerobic
conditions All of these changes greatly affect the
ability of plants to survive In response increased
stomatal resistance photosynthesis and root
hydraulic conductivity decreases and reduced
translocation fotoassimilat Nevertheless one of
15
the best adaptations of plants to hypoxia anoxia
is transitional biochemical and metabolic processes
common to current limited availability of O2 (Dat et
al 2004)
Selective synthesis of a set of about 20
anaerobic stress proteins (ANPS) allows the process
of producing energy without oxygen metabolism under
anaerobic conditions (Subbaiah and Sachs 2003)
Another adaptation is the observed morphological
changes consisting of the formation of hypertrophic
lenticels adventitious root initiation and or
progression of aerenchyma (Vartapetian and Jackson
1997 Jackson and Colmer 2005 Folzer et al 2006)
This review details the diverse plant stress
response to hypoxia anoxia caused by soil
waterlogging flooding and examines some of the key
features of metabolic adaptation physiological and
morphological
16
C Electron
Transport System
We noted earlier that one of the principal
functions of the respiration is to retrieve in
useful form some of the energy initially stored in
assimilates Our traditional measure of useful
energy in most processes is the number of ATP
molecules gained or consumed By this measure alone
the yield from both glycolysis and the citric acid
cycle is quite low After two complete turns of the
cycle one molecule of glucose has been completely
oxidized to six molecules of CO2 but only four
molecules of ATP have been produced (a net of two
ATP from glycolysis plus one for each turn of the
cycle) At this point most of the energy associated
with the glucose molecule has been conserved in the
17
form of electron pairs generated by the oxidation of
glycolytic and citric acid cycle intermediates
In this section we will discuss the third stage
of cellular respirationmdashthe transfer of electrons
fromNADH and FADH2 to oxygen and the accompanying
conversion of redox energy to ATP The transfer of
electrons fromNADHand FADH2 to oxygen involves a
sequence of electron carriers arranged in an
electron transport chainMembrane fractionation
studies have shown that the enzymes and electron
carriers making up the electron transport chain are
organized predominantly into four large
multimolecular complexes (complexes ImdashIV) and two
mobile carriers located in the inner mitochondrial
membrane (Figure 109) In this sense there are a
great number of similarities between the
mitochondrial inner membrane and the thylakoid
membranes of the chloroplast (compare Figure 109
with Figure 76) This is not unexpected since the
principal function of each membrane is energy
transformation and many of the same or similar
components are involved
The Components of the Electron Transport Chain
The electron transport chain of the
mitochondria is the means by which electrons are
18
removed from the reduced carrier NADH and
transferred to oxygen to yield H2O
1) NADH
NADH is generated in the matrix by the reactions
of pyruvate dehydrogenase isocitrate
dehydrogenase α-ketoglutarate dehydrogenase and
malate dehyrogenase The electron transport chain
begins with reoxidizing NADH to form NAD+ and
channeling the electrons into the formation of
reduced coenzymes Important to note that NADH
transfers 2 electrons at a time in the form of a
hydride
NAD+ + 2e- + H+ 1048581 NADH Eorsquo = minus0315 V
2) Flavoproteins
Flavoproteins have either a FAD (flavin adenosine
dinucleotide) or a FMN (flavin mononucleotide)
prosthetic group Flavoproteins can accept or
donate electrons one at time or two at a time
Thus they are often intermediaries between two
electron acceptorsdonors and one electron
acceptorsdonors For flavoproteins the typical
standard reduction potentials are around 0 V
FAD + 2e- + 2H+ 1048581 FADH2 FMN + 2e- + 2H+ 1048581 FMNH2
Eorsquo asymp 0 V
19
3) Coenzyme Q (CoQ)
aka ubiquinone (UQ) Shown below CoQ has ten
repeating isoprene units which make it
insoluble in water but soluble in the
hydrophobic lipid bylayer Coenzyme Q is a
versatile cofactor because it is a soluble
electron carrier in the hydrophobic bilipid layer
of the
inner mitochondrial membrane Like flavoproteins
CoQ can acceptdonate electrons one at a time or
two at a time
Q + 2e- + 2H+ 1048581 QH2 Eorsquo =0060 V
Q + e- + H+ 1048581 1048581QH Eorsquo =0030 V
1048581QH + e- + H+ 1048581 QH2 Eorsquo =0190 V
4) Cytochromes
Cytochromes are proteins that contain heme
prosthetic groups which function as
one electron carriers The heme iron is involved
in one electron transfers involving the Fe2+ and
Fe3+ oxidation states Cytochromes are named by
their absorption spectra which depends
on the porphyrin structure and environment The
example shown is the heme prosthetic group of
cytochrome b Cytochrome b contains the same iron
porphyrin found in hemoglobin and myoglobin
20
Other cytochromes we will encounter in the
electron transport complexes are cytochromes b
c c1 a and a3
5) Iron-Sulfur Proteins
In the electron transport chain we will encounter
many iron-sulfer proteins which participate in
one electron transfers involving the the Fe2+ and
Fe3+ oxidation states These are non-heme iron-
sulfur proteinsThe simplest iron-sulfer protein
is FeS in which iron is tetrahedrally coordinated
by four cysteines The second form is Fe2S2 which
contains two irons complexed to 2 cysteine
residues and two inorganic sulfides The third
form is Fe3S4 which contains 3 iron atoms
coordinated to three cysteine residues and 4
inorganic sulfides The last form is the most
complicated Fe4S4 which contains 4 iron atoms
coordinated to 4 cysteine residues and 4
inorganic sulfides
6) Copper Proteins
Copper bound proteins participate in one electron
transfers involving the Cu+ and Cu2+ oxidation
states
21
Overview of the Electron Transport Chain
Electrons move along the electron transport
chain going from donor to acceptor until they reach
oxygen the ultimate electron acceptor The standard
reduction potentials of the electron carriers are
between the NADHNAD+ couple (-0315 V) and the
oxygenH2O couple (0816 V) as on the next page The
components of the electron transport chain are
organized into 4 complexes Each complex contains
several different electron carriers
1 Complex I also known as the NADH-coenzyme Q
reductase or NADH dehydrogenase
2 Complex II also known as succinate-coenzyme Q
reductase or succinate dehydrogenase
3 Complex III also known as coenzyme Q reductase
4 Complex IV also known as cytochrome c reductase
The free energy necessary to generate ATP
is extracted from the oxidation of NADH and
FADH2 by the electron transport chain It
consists of four protein complexes and single
protein chain of cytochrome c through which
electrons pass from lower to higher standard
redox potentials Electrons are carried from complex
I and II to complex III by the membrane (lipid)
soluble coenzyme Q (CoQ or ubiquinone) and between
22
complexes III and IV by the peripheral membrane
protein cytochrome c
In the following I give the overall
reactions catalysed by corresponding protein
complexes in the mitochondrial electron transport
chain
1 Complex I
Complex I is also called NADH-Coenzyme Q
reductase because this large protein complex
transfers 2 electrons from NADH to coenzyme Q
Complex I was formerly known as NADH dehydrogenase
Complex I is huge 850000 kD and is composed of more
than thirty subunits It contains a FMN prosthetic
group and seven or more Fe-S clusters This complex
has between 20-26 iron atoms boundThe prosthetic
group FMN is absolutely required for activity
Therefore this complex is a flavoprotein This
complex binds NADH transfers two electrons in the
form of a hydride to FMN to produce NAD+ and FMNH2
23
The subsequent steps involve the transfer of
electrons one at a time to a series of iron-sulfer
complexes that includes both 2Fe-2S and 4Fe-4S
clusters Note the importance of FMN First it
functions as a 2 electron acceptor in the hydride
transfer from NADH Second it functions as a 1
electron donor to the series of iron sulfur clusters
FMN and FAD often play crucial links between 2
electron transfer agents and 1 electron transfer
agents The final step of this complex is the
transfer of 2 electrons one at a time to coenzyme Q
CoQ like FMN and FAD can function as a 2 electron
donoracceptor and as a 1 electron donoracceptor
CoQ is a mobile electron carrier because its
isoprenoid tail makes it highly hydrophobic and
lipophillic It diffuses freely in the bilipid layer
of the inner mitochondrial membrane
The process of transferring electrons from NADH
to CoQ by complex I results in the net transport of
protons from the matrix side of the inner
mitochondrial membrane to the inter membrane space
where the H+ ions accumulate generating a proton
motive force The intermembrane space side of the
inner membrane is referred to as the P face (P
standing for positive) The matrix side of the inner
membrane is referred to as the the N face The
24
transport of electrons from NADH to CoQ is coupled to
the transport of protons across the membrane This is
an example of active transport The stiochiometry is
4 H+ transported per 2 electrons
2 Complex II
Believe it or not you are already familiar
with Complex II It is none other than succinate
dehydrogenase The only enzyme of the citric acid
cycle that is an integral membrane protein This
complex is composed of four subunits 2 of which are
iron-sulfur proteins and the other two subunits
together bind FAD through a covalent link to a
histidine residue These two subunits are called
flavoprotein 2 or FP2 Complex II contains 3 Fe-S
centers 1 4Fe-4S cluster 1 3Fe-4S cluster and 1
2Fe- 2S cluster In the first step of this complex
succinate is bound and a hydride is transferred to
FAD to generate FADH2 and fumarate FADH2 then
transfers its electrons one at a time to the Fe-S
centers Thus once again FAD functions as 2 electron
acceptor and a 1 electron donor The final step of
this complex is the transfer of 2 electrons one at a
time to coenzyme Q to produce CoQH2 For complex II
the standard free energy change of the overall
reaction is too small to drive the transport of
protons across the inner mitochondrial membrane This
25
accounts for the 15 ATPrsquos generated per FADH2
compared with the 25 ATPrsquos generated per NADH
3 Complex III
This complex is also known as coenzyme Q-
cytochrome c reductase because it passes the
electrons form CoQH2 to cyt c through a very unique
electron transport pathway called the Q-cycle Shown
to the left are the porphyrins found in cytochromes
Cytochrome b contains the same iron protoporphyrin as
hemoglobin and myoglobin The c cytochromes contain
heme c through covalent attachment by cysteine
residues Cytochrome a is found in two forms in
complex IV In complex III we find two b-type
cytochromes and one c-type cytochrome Complex III is
complex and we have a crystal structure
4 Complex IV
Complex IV is also known as cytochrome c oxidase
because it accepts the electrons from cytochrome c
and directs them towards the four electron reduction
of O2 to form 2 molecules of H2O
4 cyt c (Fe2+) + 4 H+ + O2 4 cyt c (Fe3+) +
2H2O
26
Cytochrome c oxidase contains 2 heme
centerscytochrome a and cytochrome a3 and two copper
proteins Each of the protein bound coppers are
associated with one of the cytochromes The copper
sites are called CuA and CuB CuA is associated with
cytochrome a and is shown to the left CuB is
associated with cytochrome a3 The copper sites
function as 1 electron carriers cycling between the
cuprous state Cu+ and the cupric state Cu2+ Just
like iron containing proteins they transfer
electrons one at a time Cytochrome c is bound on the
P-face of the membrane and transfers its electron to
CuA The oxidized cytochrome c dissociates CuA then
transfers the electron to cytochrome a The protein
bound CuA and the iron bound in cytochrome a are 15 Aring
apart In contrast the CuB and the iron bound in
cytochrome a3 are very close to each other forming a
binuclear metal center shown below
27
Electron transport chains are associated with
membranes such as the mitochondrial membrane in
eukaryotic organisms the thylakoid membrane of
photosynthetic organisms and the plasma membrane of
prokaryotic organisms While some of the details
differ electron transport chains in all organisms
operate on the same basic principles
Excited electrons are brought to the electron
transport chain by electron carriers such as NADH
and FADH2 In photosynthetic organisms the electrons
are donated by water and excited by light energy
The electron donor is oxidized as the electrons are
passed to the first protein complex in the chain As
it accepts the electrons the protein complex is
reduced Next the electrons are passed from the
28
first protein complex which becomes oxidized to
the second protein complex which becomes reduced
Each of these redox reactions is slightly exergonic
or energy releasing Some of this energy is used to
move protons across the membrane against their
concentration gradient With each successive
transfer of electrons the original excited
electrons lose some of their energy After several
successive redox reactions the electrons are
donated to a terminal electron acceptor In aerobic
respiration the terminal electron acceptor is
oxygen In photosynthesis the terminal electron
acceptor is a compound called NADP
The energy in the excited electrons has been
transformed into an electrochemical gradient
represented by the high concentration of protons on
one side of the membrane These protons can diffuse
back across the membrane by passing through a
protein complex called ATP synthase As these
protons pass through ATP synthase down their
concentration gradient they provide the energy
needed to make ATP from ADP and inorganic phosphate
within the cell
29
CHAPTER III
CLOSING
A Conclution
When a hexose is completely oxidized to CO2
and H2O using these three processes Glycolysis
yields two ATP and two NADH per hexose used Each
such NADH oxidized by the electron transport system
30
yields two ATP as dscribed above so glycolysis
contributes a total of six ATP per hexose The Krebs
cycle contributes two ATP per hexose or per two
pyruvates when succinyl CoA is cleaved to succinate
and CoASH This cycle also produces eight NADH per
hexose within the mitochondrial matrix by oxidative
phosporylation each of these NADH yields three ATP
or 24 per hexose Each FADH2 from the Krebs cycle
yields two ATP by oxydative phosporylation or four
per hexose (two pyruvates) The total contribution of
the Krebs cycle is then 30 Adding these 30 to the 6
from glycolysis leads to a total of 36 per hexose
completely respired by these process
B Suggestion
My greatest wish is that the readers find
this manuscript helpful in gaining the knowledge they
need for effective learning and practice and in
preparing for examination designed to test that
knowledge Reader comments and suggestions are
welcome and would be very helpful in improving our
manuscript Your contribution may assist many other
students in the coming years
31
REFERENCE
Anonim 2013 Anaerobic Respiration
httpwwwscoolcoukgcsebiologyrespiration
revise-itanaerobic-respiration Accessed
September 10th 2013
Anonim 2013 What is the difference between aerobic
and anaerobic respiration in plants
httpwwwbiologylifeeasyorg4010difference
-between-aerobic-anaerobic-respiration-plants
Accessed September 10th 2013
Campbell Mary K 2006 Biochemistry New York Thomson
Brooks Cole Inc
Dat J Capelli N Folzer H Bourgeade P Badot P-M
(2004) Sensing and signaling during
plantflooding Plant Physiology and
Biochemistry 42 273-282
Dat J Folzer H Parent C Badot P-M Capelli N
(2006) HypoxiastressCurrent Understanding and
Perspectives In Teixeira da Silva JA (Ed)
Floriculture Ornamental and Plant
Biotechnology Advances and Topical Issues (Vol
3) Global Science Books Isleworth United
Kingdompp 664-674
Hatta Muhammad 2011 Dunia Pertanian
httpemhattawordpresscomcategorystress
Accessed September 10th 2013
Hopkins William G 2009 Introduction to Plant Physiology
New York John Wiley amp Sons Inc
Lakitan Benyamin 2011 Dasar-Dasar Fisiologi Tumbuhan
Jakarta PT Raja Grafindo Persada
Larcher W 1980 Physiological Plant Ecology New York
Springer-Verlag
Raven 2013 Biology Fifth Edition
httphigheredmcgraw-hillcomsites983409233
9student_view0chapter39
photosynthetic_electron_transport_and_atp_synth
esishtml Accessed September 10th 2013
Salisbury Frank B 1969 Plant Physiology Third Edition
California Wadsworth Publishing Company
Subbaiah C Sachs M (2003) Molecular and cellular
adaptations of maize to flooding stress Annals
of Botany 91 119-127
Wilson CarlL1962 Botany Third EditionUnited State of
America Holt Rinch Inc

PADANG STATE UNIVERSITY
2013
2
PREFACE
ldquoPlant Respiration Krebs Cycle Fermentation and
Transport Systemrdquo in this manuscript is intended to
complete the task from our lecturer Dr Azwir Anhar
MSi and to meet the needs of university students as
the guidersquos book in learning and practice activity for
the member of International Standard Teacher Education
(ISTE) program in Public University of Padang
My greatest wish is that the readers find this
manuscript helpful in gaining the knowledge they need
for effective learning and practice and in preparing
for examination designed to test that knowledge Reader
comments and suggestions are welcome and have again
been helpful in improving our manuscript Your
contribution may assist many other students in the
coming years
Writer
September 2013
i
__________________________
CONTENT
PREFACE i
CONTENT ii
CHAPTER I INTRODUCTION
A Background 1
B Purposes 2
C Benefits 2
CHAPTER II PLANT RESPIRATION
A Krebs Cycle 3
B Fermentation 8
C Electron Transport System
11
CHAPTER III CLOSING
ii
A Conclution
20
B Suggestion
20
REFERENCE
iii
CHAPTER I
INTRODUCTION
A Background
Connected with photosynthesis a metabolic
process takes place in chloroplast-containing plant
cells which like respiration takes up O2 and
releases CO2 in the light but which contrary to
respiration ceases in the dark This O2CO2 gas
exchange has been called right respiration or
photorespiration The substrate for the
photorespiratory metabolism is again ribulose
biphosphate which can be an acceptor not only for
CO2 but also for O2 By taking up oxygen RuBP is
split into PGA and phospoglycolate The supply of O2
and CO2 regulates the relationship between acceptor
oxidation (photorespiration) and acceptor
carboxylation (photosynthesis) via the enzyme
complex RuBP carboxylaseoxygenase High partial
pressure of O2 favors photorespiration A large
supply of CO2 favors photosynthesis The formation
of phospoglycolate is dependent on the supply of
RuBP via the Calvin cycle the photorespiratory O2
uptake and CO2 release increase with light
intensity
1
The process of glycolate metabolism has not
yet been understood in all details The glycolate is
transported out of the chloroplasts phospoglycolate
and phospate The glycolate is transported out of
the chloroplast into peroxisomes cell compartments
about the size of mitochondria which contain
glycolate oxidase catalase and transaminases In
the peroxysomes when O2 is taken up glycolate is
oxidized to glyoxylate and the peroxide thus
produced is detoxified by catalase Glycolate can
either be completely reduced via oxalate by further
O2 uptake or transformed to glycine by
transamination Glycine is transported from the
peroxisomes into the mitochondria where two
molecules of glycine are coupled to form one
molecule of serine with the release of CO2 Serine
is taken over by the amino acid metabolism or
2
converted to glycerate after deamination by
hydropyruvate This can be photophosporylated in the
chloroplasts and returned to the Calvin cycle or
used elsewhere (Larcher 1980 81-82)
B Purposes
This manuscript is intended to complete the
task from our lecturer Dr Azwir Anhar MSi and
to meet the needs of university students as the
guidersquos book in learning and practice activity for
the member of International Standard Teacher
Education (ISTE) program in Public University of
Padang
C Benefits
Benefits that we gained from this manuscript
are
1 As reference in learning process of plant
phisiology subject especially in plant
respiration title
2 As information about steps in plant
respiration
3
CHAPTER II
PLANT RESPIRATION
A Krebs Cycle
The Krebs cycle was named in honor of the
English biochemist Hans A Krebs who in 1937
proposed a cycle of reactions to explain how pyruvate
breakdown takes place in the breast muscle of
pigeons He called his proposed pathway the citric
acid cycle because citric acid is an important
intermediet Another common name for the same group
of reactions is the Tricarboxuylic Acid (TCA) cycle
a term used because citric and isocitric acids have
three carboxyl groups It was not until the early
1950s that mitochondria capable of carrying out this
cycle were isolated from plant cells
The initial step leading to the Krebs cycle
involves the oxidation and loss of CO2 from pyruvate
and the combination of the remaining 2-carbon acetate
unit with a sulfur-containing compound co-enzyme A
(CoA) forming acetyl CoA This and another
comparable role of CoA in the Krebs cycle are
important reasons why sulfur is an essential element
4
The reaction of pyruvate decarboxylation also
involves a phosporylated form of thiamine (vitamin
B1) as a prostethic group Participation of thiamine
in this section partially explains the essential
function of vitamin B1 in plants and animals Besides
the loss of CO2 two hydrogen atoms are removed from
pyruvic acid during the formation of Acetyl CoA The
enzyme catalyzing the complete reaction is called
pyruvic acid dehydrogenase but it is actually an
organized complex containing numerous copies of three
or four different enzymes The hydrogen atoms removed
are finally accepted by NAD+ yielding NADH
The Krebs cycle accomplishes removal of some of
the electrons from organic acid intermediates and
transfer of these electrons to NAD+ or FAD Notice
that none of dehydrogenase enzyme of the cycle uses
NADP+ as an electron acceptor In fact NADP+ is
usually undetectable in plant mitochondria a
situation opposite to that of chloroplasts where
NADP+ is abundant but where there is much less NAD+
Not only are NADH and FADH2 important products of the
Krebs cycle but one molecule of ATP is formed from
ADP and Pi during the conversion of succinyl coenzyme
A to succinic acid Two additional CO2 molecules are
released in these Krebs cycle reactions so there is
a net loss of both carbon atoms from the incoming
5
acetate of acetyl CoA The release of CO2 in the
Krebs cycle accounts for the product of CO2 in the
summary equation for resspiration but no O2 is
absorbed during any Kreb cycle reaction (Lakitan
2011)
During metabolism the synthesis and breakdown
of different organic compounds takes place through
various pathways like the breakdown and synthesis of
proteins carbohydrates fats and nucleic acids
These different pathways and intermediates are also
responsible for the production of energyKrebrsquos
cycle named after Hans Krebs who began working out
its details in 1930s is a series of reactions in
which the pyruvate from glycolysis is oxidized to Co2
under aerobic conditions Kreb cycle is also known as
citric acid cycle or Tri-carboxylic acid cycle (TCA)
( Galston 1968 76)
6
Steps involved in the process of Kreb cycle are
1 Fate of pyruvic acid
Pyruvic acid can form different compounds by
different pathways ie it can be converted into
lactic acid It can be converted into acetyl
coenzyme A as a result of oxidation
2 Fate of Acetyl CoA
It can either undergo condensation with itself or
its derivatives to form fatty acids having 14 ndash 20
carbons Acetyl CoA can also go through a series
of reactions in Krebs cycle
3 Formation of citrate
Acetyl CoA enzyme condenses with oxaloacetate by
an enzyme citrate synthatase to form citrate with
the release of acetyl CoA If the amount of
7
oxaloacetate is very small then small number of
acetyl CoA would be reacting with oxaloacetate
leaving surplus acetyl CoA to go through another
pathway for the formation of long chain fatty
acids
4 Formation of C is ndash acotinate and iso-citrate
Citrate is changed first into C is ndash acotinate and
then to iso-citrate under the enzyme acotinase
Equilibrium is established between citrate C is
aconitate and iso-citrate It has been observed
that most of the time this equilibrium is shifted
towards the iso-citrate If the concentration of
iso-citrate in increased the formation of citrate
will result which also indicates that the
equilibrium also shift in the reserve direction
5 Formation of oxalosucinate
Iso-citrate is acted upon by an enzyme iso-citrate
dehydrogenase using nicotinamide adenine
dinucleotide (NAD) as coenzyme As a result iso-
citrate is converted into oxalosucciate and NAD is
reduced to NADH2 Similar reaction is carried out
by same enzyme using nicotindmide adenine
dinucleotide phosphate (NADP) as coenzyme which is
reduced to NDAPH2
6 Formation of α-Ketoglutarate
8
Oxalosucciante is changed into α-ketoglutrate by
iso-citrate dehydrogenase with the help of
coenzyme NAD or NADP In this reaction carbon
dioxide and NADH2 or NADPH2 are also released
7 Formation of succinyl CoA
α-ketoglutarate combines with acetyl CoA in the
presence of coenzyme NAD and enzyme α-
keoglutaratedehyrogenase to form succinyl coenzyme
a carbon dioxide and NADH2
8 Formation of Succinate
Later on coenzyme a is removed from succinyl
coenzyme A in the presence of guanosinediphosphate
(GDP) and inorganic phosphate to form succinate
and guanosine triphosphate (GTP) This reaction is
carried out by an enzyme called succinyl CoA
synthatase
9 Formation of Funarate
An enzyme succinic dehydrogenase removes hydrogen
from succinate to form funarate
10 Formation of Malate
Fumarate reacts with water in the presence of
enzyme fumarase to form malate
11 Regeneration of oxaloacetate
Malate is oxidized by malic dehydrogenase and NAD
forming oxaloacetate and NADH2 Thus oxalo acetate
is again available to start another cycle
9
(httpkrebs-cycle-or-citric-acid-cycle-or-
trihtml)
Importance of CTA Cycle
1 Source of energy
In addition to routine organic compounds described
above by products like nicotinanide adenine
dinucleotide (NADH2) and guanosine triphosphate
(GTP) are the source of biological energy NADH2
after oxidation produce energy whereas GTP is
itself high energy phosphate compound
2 Oxidation of organic compounds taken as food
Oxidation of fats carbohydrates and proteins take
place through it or in other words it can be said
that oxidation of all compounds having carbon
atoms can take place through TCA cycle Some of
amino acids like alanine glutamic acid and
aspartic acid at one stage or the other enters
into TCA cycle eg glutamic acid enters cycle
after its transformation into α-ketoglutarate
Similarly alanine enters the cycle after its
conversion into pyruvate
3 Intermediate compounds
TCA cycle is also involved in synthesis of
intermediate compounds leading to the formation of
larger molecules (Ray 1963 17)
10
Verification of Krebs Cycle
It was done by radioactive traces like C14 as
radioactive carbon dioxide of different levels and
reactions After addition of radioactive carbon
dioxide different chemical compounds produced like
glucose fats amino acids were isolated and looked
for radioactive carbon In this way whole of
metabolic reaction were verified including individual
reactions alternative metabolic pathways
intermediates of fats carbohydrates and amino acids
etc in the body cells as well as in test tubes
(Wilson 1962 99)
The primary function of the Krebs cycle are as
follows
1 Reduction of NAD+ and FAD to the electron donors
NADH and FADH2 that are subsequently oxidized to
yield ATP
2 Direct synthesis of a limited amount of ATP (1 ATP
for each pyruvate oxidized)
3 Formation of carbon skeletons that can be used to
synthesize certain amino acids that in turn are
converted into larger molecules
11
B Fermentation
Although glycolysis can function well without
O2 the further oxidation of pyruvate and NADH by
mitochondria requires this gas Thus when O2 is
limiting NADH and pyruvate begin to accumulate
Under this condition plants carry out fermentation
(anaerobic respiration) forming either ethanol or
lactic acid (usually ethanol) As shown in the
picture below the two top reactions consists of a
decarboxylation to form acetaldehide then rapid
reduction of acetaldehide by NADH to form ethanol
These reactions are catalyzed by alcohol
dehydrogenase Some cells contain lactic acid
12
dehydrogenase which uses NADH to reduce pyruvic to
lactic acid Thus either ethanol or lactic acid or
both are fermentation products depending on the
activities of each dehydrogenase present In each
case NADH is the reductant and only under
anaerobic conditions is it abundant anough to cause
reduction Furthermore in some plants NADH is used
to cause accumulation of the other compounds when O2
is limiting especially malate and glycerol
(Salisbury 1969 233)
The fate of pyruvate produced by glycolysis
depends primarily on whether or not oxygen is
present Under normal aerobic conditions pyruvate
is transported into the mitochondrion where it is
further oxidized to CO2 and water transferring its
electrons ultimately to molecular oxygen
13
Although higher plants are obligate aerobs and
are able to tolerate anoxia for only short periods
tissues or organs are occasionally subjected to
anaerobic conditions A typical siuation is that of
roots when the soil is saturated with water When
there is no oxygen to serve as the terminal electron
acceptor mitochondrial respiration will shut down
and metabolism will shift over to fermentation
Fermentation converts pyruvate to either ethanol
through the action of the enzyme alcohol
dehydrogenase (ADH) or lactate via lactate
dehydrogenase (LDH) In most plants the principal
products of fermentation are CO2 and ethanol Some
lactate may be formed primarily in the early stages
of anoxia However lactate lowers he pH of the
cytosol which in turn activates pyruvate
decarboxylase and initiates the production of
ethanol
Either one of the fermentation reactions
consumes the NADH produced earlier in glycolysis by
the oxidation of glyceraldehide-3-P Although this
means there is no net gain of reducing potential in
fermentation This recycling of NADH is still
important to the cell The poolof NADH plus NAD+ in
the cell is relatively small and if the NADH is not
recycled there will be no supply of NAD+ to support
14
the continued oxidation of glyceraldehyde-3-P if
this were the case glycolysis and the producton of
even the small quantities of ATP necessary to
maintain the cells under anaerobic conditions would
then grind to a halt (Hopkins 2009 241)
Soil waterlogging has long been identified as a
major abiotic stress and the constraints that it
provides the roots have a significant effect on the
growth and development of plants When this event
occurs in the spring then these puddles can reduce
seed germination and seedling development Thus a
pool of water is an important factor that affects
the growth development and survival of plant
species not only in natural ecosystems but also
the agricultural and horticultural systems (Dat et
al 2006)
After flooding rapid changes in soil
properties At the time of pore water meets the
land the air pushed out gas diffusion is reduced
and toxic compounds accumulate due to anaerobic
conditions All of these changes greatly affect the
ability of plants to survive In response increased
stomatal resistance photosynthesis and root
hydraulic conductivity decreases and reduced
translocation fotoassimilat Nevertheless one of
15
the best adaptations of plants to hypoxia anoxia
is transitional biochemical and metabolic processes
common to current limited availability of O2 (Dat et
al 2004)
Selective synthesis of a set of about 20
anaerobic stress proteins (ANPS) allows the process
of producing energy without oxygen metabolism under
anaerobic conditions (Subbaiah and Sachs 2003)
Another adaptation is the observed morphological
changes consisting of the formation of hypertrophic
lenticels adventitious root initiation and or
progression of aerenchyma (Vartapetian and Jackson
1997 Jackson and Colmer 2005 Folzer et al 2006)
This review details the diverse plant stress
response to hypoxia anoxia caused by soil
waterlogging flooding and examines some of the key
features of metabolic adaptation physiological and
morphological
16
C Electron
Transport System
We noted earlier that one of the principal
functions of the respiration is to retrieve in
useful form some of the energy initially stored in
assimilates Our traditional measure of useful
energy in most processes is the number of ATP
molecules gained or consumed By this measure alone
the yield from both glycolysis and the citric acid
cycle is quite low After two complete turns of the
cycle one molecule of glucose has been completely
oxidized to six molecules of CO2 but only four
molecules of ATP have been produced (a net of two
ATP from glycolysis plus one for each turn of the
cycle) At this point most of the energy associated
with the glucose molecule has been conserved in the
17
form of electron pairs generated by the oxidation of
glycolytic and citric acid cycle intermediates
In this section we will discuss the third stage
of cellular respirationmdashthe transfer of electrons
fromNADH and FADH2 to oxygen and the accompanying
conversion of redox energy to ATP The transfer of
electrons fromNADHand FADH2 to oxygen involves a
sequence of electron carriers arranged in an
electron transport chainMembrane fractionation
studies have shown that the enzymes and electron
carriers making up the electron transport chain are
organized predominantly into four large
multimolecular complexes (complexes ImdashIV) and two
mobile carriers located in the inner mitochondrial
membrane (Figure 109) In this sense there are a
great number of similarities between the
mitochondrial inner membrane and the thylakoid
membranes of the chloroplast (compare Figure 109
with Figure 76) This is not unexpected since the
principal function of each membrane is energy
transformation and many of the same or similar
components are involved
The Components of the Electron Transport Chain
The electron transport chain of the
mitochondria is the means by which electrons are
18
removed from the reduced carrier NADH and
transferred to oxygen to yield H2O
1) NADH
NADH is generated in the matrix by the reactions
of pyruvate dehydrogenase isocitrate
dehydrogenase α-ketoglutarate dehydrogenase and
malate dehyrogenase The electron transport chain
begins with reoxidizing NADH to form NAD+ and
channeling the electrons into the formation of
reduced coenzymes Important to note that NADH
transfers 2 electrons at a time in the form of a
hydride
NAD+ + 2e- + H+ 1048581 NADH Eorsquo = minus0315 V
2) Flavoproteins
Flavoproteins have either a FAD (flavin adenosine
dinucleotide) or a FMN (flavin mononucleotide)
prosthetic group Flavoproteins can accept or
donate electrons one at time or two at a time
Thus they are often intermediaries between two
electron acceptorsdonors and one electron
acceptorsdonors For flavoproteins the typical
standard reduction potentials are around 0 V
FAD + 2e- + 2H+ 1048581 FADH2 FMN + 2e- + 2H+ 1048581 FMNH2
Eorsquo asymp 0 V
19
3) Coenzyme Q (CoQ)
aka ubiquinone (UQ) Shown below CoQ has ten
repeating isoprene units which make it
insoluble in water but soluble in the
hydrophobic lipid bylayer Coenzyme Q is a
versatile cofactor because it is a soluble
electron carrier in the hydrophobic bilipid layer
of the
inner mitochondrial membrane Like flavoproteins
CoQ can acceptdonate electrons one at a time or
two at a time
Q + 2e- + 2H+ 1048581 QH2 Eorsquo =0060 V
Q + e- + H+ 1048581 1048581QH Eorsquo =0030 V
1048581QH + e- + H+ 1048581 QH2 Eorsquo =0190 V
4) Cytochromes
Cytochromes are proteins that contain heme
prosthetic groups which function as
one electron carriers The heme iron is involved
in one electron transfers involving the Fe2+ and
Fe3+ oxidation states Cytochromes are named by
their absorption spectra which depends
on the porphyrin structure and environment The
example shown is the heme prosthetic group of
cytochrome b Cytochrome b contains the same iron
porphyrin found in hemoglobin and myoglobin
20
Other cytochromes we will encounter in the
electron transport complexes are cytochromes b
c c1 a and a3
5) Iron-Sulfur Proteins
In the electron transport chain we will encounter
many iron-sulfer proteins which participate in
one electron transfers involving the the Fe2+ and
Fe3+ oxidation states These are non-heme iron-
sulfur proteinsThe simplest iron-sulfer protein
is FeS in which iron is tetrahedrally coordinated
by four cysteines The second form is Fe2S2 which
contains two irons complexed to 2 cysteine
residues and two inorganic sulfides The third
form is Fe3S4 which contains 3 iron atoms
coordinated to three cysteine residues and 4
inorganic sulfides The last form is the most
complicated Fe4S4 which contains 4 iron atoms
coordinated to 4 cysteine residues and 4
inorganic sulfides
6) Copper Proteins
Copper bound proteins participate in one electron
transfers involving the Cu+ and Cu2+ oxidation
states
21
Overview of the Electron Transport Chain
Electrons move along the electron transport
chain going from donor to acceptor until they reach
oxygen the ultimate electron acceptor The standard
reduction potentials of the electron carriers are
between the NADHNAD+ couple (-0315 V) and the
oxygenH2O couple (0816 V) as on the next page The
components of the electron transport chain are
organized into 4 complexes Each complex contains
several different electron carriers
1 Complex I also known as the NADH-coenzyme Q
reductase or NADH dehydrogenase
2 Complex II also known as succinate-coenzyme Q
reductase or succinate dehydrogenase
3 Complex III also known as coenzyme Q reductase
4 Complex IV also known as cytochrome c reductase
The free energy necessary to generate ATP
is extracted from the oxidation of NADH and
FADH2 by the electron transport chain It
consists of four protein complexes and single
protein chain of cytochrome c through which
electrons pass from lower to higher standard
redox potentials Electrons are carried from complex
I and II to complex III by the membrane (lipid)
soluble coenzyme Q (CoQ or ubiquinone) and between
22
complexes III and IV by the peripheral membrane
protein cytochrome c
In the following I give the overall
reactions catalysed by corresponding protein
complexes in the mitochondrial electron transport
chain
1 Complex I
Complex I is also called NADH-Coenzyme Q
reductase because this large protein complex
transfers 2 electrons from NADH to coenzyme Q
Complex I was formerly known as NADH dehydrogenase
Complex I is huge 850000 kD and is composed of more
than thirty subunits It contains a FMN prosthetic
group and seven or more Fe-S clusters This complex
has between 20-26 iron atoms boundThe prosthetic
group FMN is absolutely required for activity
Therefore this complex is a flavoprotein This
complex binds NADH transfers two electrons in the
form of a hydride to FMN to produce NAD+ and FMNH2
23
The subsequent steps involve the transfer of
electrons one at a time to a series of iron-sulfer
complexes that includes both 2Fe-2S and 4Fe-4S
clusters Note the importance of FMN First it
functions as a 2 electron acceptor in the hydride
transfer from NADH Second it functions as a 1
electron donor to the series of iron sulfur clusters
FMN and FAD often play crucial links between 2
electron transfer agents and 1 electron transfer
agents The final step of this complex is the
transfer of 2 electrons one at a time to coenzyme Q
CoQ like FMN and FAD can function as a 2 electron
donoracceptor and as a 1 electron donoracceptor
CoQ is a mobile electron carrier because its
isoprenoid tail makes it highly hydrophobic and
lipophillic It diffuses freely in the bilipid layer
of the inner mitochondrial membrane
The process of transferring electrons from NADH
to CoQ by complex I results in the net transport of
protons from the matrix side of the inner
mitochondrial membrane to the inter membrane space
where the H+ ions accumulate generating a proton
motive force The intermembrane space side of the
inner membrane is referred to as the P face (P
standing for positive) The matrix side of the inner
membrane is referred to as the the N face The
24
transport of electrons from NADH to CoQ is coupled to
the transport of protons across the membrane This is
an example of active transport The stiochiometry is
4 H+ transported per 2 electrons
2 Complex II
Believe it or not you are already familiar
with Complex II It is none other than succinate
dehydrogenase The only enzyme of the citric acid
cycle that is an integral membrane protein This
complex is composed of four subunits 2 of which are
iron-sulfur proteins and the other two subunits
together bind FAD through a covalent link to a
histidine residue These two subunits are called
flavoprotein 2 or FP2 Complex II contains 3 Fe-S
centers 1 4Fe-4S cluster 1 3Fe-4S cluster and 1
2Fe- 2S cluster In the first step of this complex
succinate is bound and a hydride is transferred to
FAD to generate FADH2 and fumarate FADH2 then
transfers its electrons one at a time to the Fe-S
centers Thus once again FAD functions as 2 electron
acceptor and a 1 electron donor The final step of
this complex is the transfer of 2 electrons one at a
time to coenzyme Q to produce CoQH2 For complex II
the standard free energy change of the overall
reaction is too small to drive the transport of
protons across the inner mitochondrial membrane This
25
accounts for the 15 ATPrsquos generated per FADH2
compared with the 25 ATPrsquos generated per NADH
3 Complex III
This complex is also known as coenzyme Q-
cytochrome c reductase because it passes the
electrons form CoQH2 to cyt c through a very unique
electron transport pathway called the Q-cycle Shown
to the left are the porphyrins found in cytochromes
Cytochrome b contains the same iron protoporphyrin as
hemoglobin and myoglobin The c cytochromes contain
heme c through covalent attachment by cysteine
residues Cytochrome a is found in two forms in
complex IV In complex III we find two b-type
cytochromes and one c-type cytochrome Complex III is
complex and we have a crystal structure
4 Complex IV
Complex IV is also known as cytochrome c oxidase
because it accepts the electrons from cytochrome c
and directs them towards the four electron reduction
of O2 to form 2 molecules of H2O
4 cyt c (Fe2+) + 4 H+ + O2 4 cyt c (Fe3+) +
2H2O
26
Cytochrome c oxidase contains 2 heme
centerscytochrome a and cytochrome a3 and two copper
proteins Each of the protein bound coppers are
associated with one of the cytochromes The copper
sites are called CuA and CuB CuA is associated with
cytochrome a and is shown to the left CuB is
associated with cytochrome a3 The copper sites
function as 1 electron carriers cycling between the
cuprous state Cu+ and the cupric state Cu2+ Just
like iron containing proteins they transfer
electrons one at a time Cytochrome c is bound on the
P-face of the membrane and transfers its electron to
CuA The oxidized cytochrome c dissociates CuA then
transfers the electron to cytochrome a The protein
bound CuA and the iron bound in cytochrome a are 15 Aring
apart In contrast the CuB and the iron bound in
cytochrome a3 are very close to each other forming a
binuclear metal center shown below
27
Electron transport chains are associated with
membranes such as the mitochondrial membrane in
eukaryotic organisms the thylakoid membrane of
photosynthetic organisms and the plasma membrane of
prokaryotic organisms While some of the details
differ electron transport chains in all organisms
operate on the same basic principles
Excited electrons are brought to the electron
transport chain by electron carriers such as NADH
and FADH2 In photosynthetic organisms the electrons
are donated by water and excited by light energy
The electron donor is oxidized as the electrons are
passed to the first protein complex in the chain As
it accepts the electrons the protein complex is
reduced Next the electrons are passed from the
28
first protein complex which becomes oxidized to
the second protein complex which becomes reduced
Each of these redox reactions is slightly exergonic
or energy releasing Some of this energy is used to
move protons across the membrane against their
concentration gradient With each successive
transfer of electrons the original excited
electrons lose some of their energy After several
successive redox reactions the electrons are
donated to a terminal electron acceptor In aerobic
respiration the terminal electron acceptor is
oxygen In photosynthesis the terminal electron
acceptor is a compound called NADP
The energy in the excited electrons has been
transformed into an electrochemical gradient
represented by the high concentration of protons on
one side of the membrane These protons can diffuse
back across the membrane by passing through a
protein complex called ATP synthase As these
protons pass through ATP synthase down their
concentration gradient they provide the energy
needed to make ATP from ADP and inorganic phosphate
within the cell
29
CHAPTER III
CLOSING
A Conclution
When a hexose is completely oxidized to CO2
and H2O using these three processes Glycolysis
yields two ATP and two NADH per hexose used Each
such NADH oxidized by the electron transport system
30
yields two ATP as dscribed above so glycolysis
contributes a total of six ATP per hexose The Krebs
cycle contributes two ATP per hexose or per two
pyruvates when succinyl CoA is cleaved to succinate
and CoASH This cycle also produces eight NADH per
hexose within the mitochondrial matrix by oxidative
phosporylation each of these NADH yields three ATP
or 24 per hexose Each FADH2 from the Krebs cycle
yields two ATP by oxydative phosporylation or four
per hexose (two pyruvates) The total contribution of
the Krebs cycle is then 30 Adding these 30 to the 6
from glycolysis leads to a total of 36 per hexose
completely respired by these process
B Suggestion
My greatest wish is that the readers find
this manuscript helpful in gaining the knowledge they
need for effective learning and practice and in
preparing for examination designed to test that
knowledge Reader comments and suggestions are
welcome and would be very helpful in improving our
manuscript Your contribution may assist many other
students in the coming years
31
REFERENCE
Anonim 2013 Anaerobic Respiration
httpwwwscoolcoukgcsebiologyrespiration
revise-itanaerobic-respiration Accessed
September 10th 2013
Anonim 2013 What is the difference between aerobic
and anaerobic respiration in plants
httpwwwbiologylifeeasyorg4010difference
-between-aerobic-anaerobic-respiration-plants
Accessed September 10th 2013
Campbell Mary K 2006 Biochemistry New York Thomson
Brooks Cole Inc
Dat J Capelli N Folzer H Bourgeade P Badot P-M
(2004) Sensing and signaling during
plantflooding Plant Physiology and
Biochemistry 42 273-282
Dat J Folzer H Parent C Badot P-M Capelli N
(2006) HypoxiastressCurrent Understanding and
Perspectives In Teixeira da Silva JA (Ed)
Floriculture Ornamental and Plant
Biotechnology Advances and Topical Issues (Vol
3) Global Science Books Isleworth United
Kingdompp 664-674
Hatta Muhammad 2011 Dunia Pertanian
httpemhattawordpresscomcategorystress
Accessed September 10th 2013
Hopkins William G 2009 Introduction to Plant Physiology
New York John Wiley amp Sons Inc
Lakitan Benyamin 2011 Dasar-Dasar Fisiologi Tumbuhan
Jakarta PT Raja Grafindo Persada
Larcher W 1980 Physiological Plant Ecology New York
Springer-Verlag
Raven 2013 Biology Fifth Edition
httphigheredmcgraw-hillcomsites983409233
9student_view0chapter39
photosynthetic_electron_transport_and_atp_synth
esishtml Accessed September 10th 2013
Salisbury Frank B 1969 Plant Physiology Third Edition
California Wadsworth Publishing Company
Subbaiah C Sachs M (2003) Molecular and cellular
adaptations of maize to flooding stress Annals
of Botany 91 119-127
Wilson CarlL1962 Botany Third EditionUnited State of
America Holt Rinch Inc

PREFACE
ldquoPlant Respiration Krebs Cycle Fermentation and
Transport Systemrdquo in this manuscript is intended to
complete the task from our lecturer Dr Azwir Anhar
MSi and to meet the needs of university students as
the guidersquos book in learning and practice activity for
the member of International Standard Teacher Education
(ISTE) program in Public University of Padang
My greatest wish is that the readers find this
manuscript helpful in gaining the knowledge they need
for effective learning and practice and in preparing
for examination designed to test that knowledge Reader
comments and suggestions are welcome and have again
been helpful in improving our manuscript Your
contribution may assist many other students in the
coming years
Writer
September 2013
i
__________________________
CONTENT
PREFACE i
CONTENT ii
CHAPTER I INTRODUCTION
A Background 1
B Purposes 2
C Benefits 2
CHAPTER II PLANT RESPIRATION
A Krebs Cycle 3
B Fermentation 8
C Electron Transport System
11
CHAPTER III CLOSING
ii
A Conclution
20
B Suggestion
20
REFERENCE
iii
CHAPTER I
INTRODUCTION
A Background
Connected with photosynthesis a metabolic
process takes place in chloroplast-containing plant
cells which like respiration takes up O2 and
releases CO2 in the light but which contrary to
respiration ceases in the dark This O2CO2 gas
exchange has been called right respiration or
photorespiration The substrate for the
photorespiratory metabolism is again ribulose
biphosphate which can be an acceptor not only for
CO2 but also for O2 By taking up oxygen RuBP is
split into PGA and phospoglycolate The supply of O2
and CO2 regulates the relationship between acceptor
oxidation (photorespiration) and acceptor
carboxylation (photosynthesis) via the enzyme
complex RuBP carboxylaseoxygenase High partial
pressure of O2 favors photorespiration A large
supply of CO2 favors photosynthesis The formation
of phospoglycolate is dependent on the supply of
RuBP via the Calvin cycle the photorespiratory O2
uptake and CO2 release increase with light
intensity
1
The process of glycolate metabolism has not
yet been understood in all details The glycolate is
transported out of the chloroplasts phospoglycolate
and phospate The glycolate is transported out of
the chloroplast into peroxisomes cell compartments
about the size of mitochondria which contain
glycolate oxidase catalase and transaminases In
the peroxysomes when O2 is taken up glycolate is
oxidized to glyoxylate and the peroxide thus
produced is detoxified by catalase Glycolate can
either be completely reduced via oxalate by further
O2 uptake or transformed to glycine by
transamination Glycine is transported from the
peroxisomes into the mitochondria where two
molecules of glycine are coupled to form one
molecule of serine with the release of CO2 Serine
is taken over by the amino acid metabolism or
2
converted to glycerate after deamination by
hydropyruvate This can be photophosporylated in the
chloroplasts and returned to the Calvin cycle or
used elsewhere (Larcher 1980 81-82)
B Purposes
This manuscript is intended to complete the
task from our lecturer Dr Azwir Anhar MSi and
to meet the needs of university students as the
guidersquos book in learning and practice activity for
the member of International Standard Teacher
Education (ISTE) program in Public University of
Padang
C Benefits
Benefits that we gained from this manuscript
are
1 As reference in learning process of plant
phisiology subject especially in plant
respiration title
2 As information about steps in plant
respiration
3
CHAPTER II
PLANT RESPIRATION
A Krebs Cycle
The Krebs cycle was named in honor of the
English biochemist Hans A Krebs who in 1937
proposed a cycle of reactions to explain how pyruvate
breakdown takes place in the breast muscle of
pigeons He called his proposed pathway the citric
acid cycle because citric acid is an important
intermediet Another common name for the same group
of reactions is the Tricarboxuylic Acid (TCA) cycle
a term used because citric and isocitric acids have
three carboxyl groups It was not until the early
1950s that mitochondria capable of carrying out this
cycle were isolated from plant cells
The initial step leading to the Krebs cycle
involves the oxidation and loss of CO2 from pyruvate
and the combination of the remaining 2-carbon acetate
unit with a sulfur-containing compound co-enzyme A
(CoA) forming acetyl CoA This and another
comparable role of CoA in the Krebs cycle are
important reasons why sulfur is an essential element
4
The reaction of pyruvate decarboxylation also
involves a phosporylated form of thiamine (vitamin
B1) as a prostethic group Participation of thiamine
in this section partially explains the essential
function of vitamin B1 in plants and animals Besides
the loss of CO2 two hydrogen atoms are removed from
pyruvic acid during the formation of Acetyl CoA The
enzyme catalyzing the complete reaction is called
pyruvic acid dehydrogenase but it is actually an
organized complex containing numerous copies of three
or four different enzymes The hydrogen atoms removed
are finally accepted by NAD+ yielding NADH
The Krebs cycle accomplishes removal of some of
the electrons from organic acid intermediates and
transfer of these electrons to NAD+ or FAD Notice
that none of dehydrogenase enzyme of the cycle uses
NADP+ as an electron acceptor In fact NADP+ is
usually undetectable in plant mitochondria a
situation opposite to that of chloroplasts where
NADP+ is abundant but where there is much less NAD+
Not only are NADH and FADH2 important products of the
Krebs cycle but one molecule of ATP is formed from
ADP and Pi during the conversion of succinyl coenzyme
A to succinic acid Two additional CO2 molecules are
released in these Krebs cycle reactions so there is
a net loss of both carbon atoms from the incoming
5
acetate of acetyl CoA The release of CO2 in the
Krebs cycle accounts for the product of CO2 in the
summary equation for resspiration but no O2 is
absorbed during any Kreb cycle reaction (Lakitan
2011)
During metabolism the synthesis and breakdown
of different organic compounds takes place through
various pathways like the breakdown and synthesis of
proteins carbohydrates fats and nucleic acids
These different pathways and intermediates are also
responsible for the production of energyKrebrsquos
cycle named after Hans Krebs who began working out
its details in 1930s is a series of reactions in
which the pyruvate from glycolysis is oxidized to Co2
under aerobic conditions Kreb cycle is also known as
citric acid cycle or Tri-carboxylic acid cycle (TCA)
( Galston 1968 76)
6
Steps involved in the process of Kreb cycle are
1 Fate of pyruvic acid
Pyruvic acid can form different compounds by
different pathways ie it can be converted into
lactic acid It can be converted into acetyl
coenzyme A as a result of oxidation
2 Fate of Acetyl CoA
It can either undergo condensation with itself or
its derivatives to form fatty acids having 14 ndash 20
carbons Acetyl CoA can also go through a series
of reactions in Krebs cycle
3 Formation of citrate
Acetyl CoA enzyme condenses with oxaloacetate by
an enzyme citrate synthatase to form citrate with
the release of acetyl CoA If the amount of
7
oxaloacetate is very small then small number of
acetyl CoA would be reacting with oxaloacetate
leaving surplus acetyl CoA to go through another
pathway for the formation of long chain fatty
acids
4 Formation of C is ndash acotinate and iso-citrate
Citrate is changed first into C is ndash acotinate and
then to iso-citrate under the enzyme acotinase
Equilibrium is established between citrate C is
aconitate and iso-citrate It has been observed
that most of the time this equilibrium is shifted
towards the iso-citrate If the concentration of
iso-citrate in increased the formation of citrate
will result which also indicates that the
equilibrium also shift in the reserve direction
5 Formation of oxalosucinate
Iso-citrate is acted upon by an enzyme iso-citrate
dehydrogenase using nicotinamide adenine
dinucleotide (NAD) as coenzyme As a result iso-
citrate is converted into oxalosucciate and NAD is
reduced to NADH2 Similar reaction is carried out
by same enzyme using nicotindmide adenine
dinucleotide phosphate (NADP) as coenzyme which is
reduced to NDAPH2
6 Formation of α-Ketoglutarate
8
Oxalosucciante is changed into α-ketoglutrate by
iso-citrate dehydrogenase with the help of
coenzyme NAD or NADP In this reaction carbon
dioxide and NADH2 or NADPH2 are also released
7 Formation of succinyl CoA
α-ketoglutarate combines with acetyl CoA in the
presence of coenzyme NAD and enzyme α-
keoglutaratedehyrogenase to form succinyl coenzyme
a carbon dioxide and NADH2
8 Formation of Succinate
Later on coenzyme a is removed from succinyl
coenzyme A in the presence of guanosinediphosphate
(GDP) and inorganic phosphate to form succinate
and guanosine triphosphate (GTP) This reaction is
carried out by an enzyme called succinyl CoA
synthatase
9 Formation of Funarate
An enzyme succinic dehydrogenase removes hydrogen
from succinate to form funarate
10 Formation of Malate
Fumarate reacts with water in the presence of
enzyme fumarase to form malate
11 Regeneration of oxaloacetate
Malate is oxidized by malic dehydrogenase and NAD
forming oxaloacetate and NADH2 Thus oxalo acetate
is again available to start another cycle
9
(httpkrebs-cycle-or-citric-acid-cycle-or-
trihtml)
Importance of CTA Cycle
1 Source of energy
In addition to routine organic compounds described
above by products like nicotinanide adenine
dinucleotide (NADH2) and guanosine triphosphate
(GTP) are the source of biological energy NADH2
after oxidation produce energy whereas GTP is
itself high energy phosphate compound
2 Oxidation of organic compounds taken as food
Oxidation of fats carbohydrates and proteins take
place through it or in other words it can be said
that oxidation of all compounds having carbon
atoms can take place through TCA cycle Some of
amino acids like alanine glutamic acid and
aspartic acid at one stage or the other enters
into TCA cycle eg glutamic acid enters cycle
after its transformation into α-ketoglutarate
Similarly alanine enters the cycle after its
conversion into pyruvate
3 Intermediate compounds
TCA cycle is also involved in synthesis of
intermediate compounds leading to the formation of
larger molecules (Ray 1963 17)
10
Verification of Krebs Cycle
It was done by radioactive traces like C14 as
radioactive carbon dioxide of different levels and
reactions After addition of radioactive carbon
dioxide different chemical compounds produced like
glucose fats amino acids were isolated and looked
for radioactive carbon In this way whole of
metabolic reaction were verified including individual
reactions alternative metabolic pathways
intermediates of fats carbohydrates and amino acids
etc in the body cells as well as in test tubes
(Wilson 1962 99)
The primary function of the Krebs cycle are as
follows
1 Reduction of NAD+ and FAD to the electron donors
NADH and FADH2 that are subsequently oxidized to
yield ATP
2 Direct synthesis of a limited amount of ATP (1 ATP
for each pyruvate oxidized)
3 Formation of carbon skeletons that can be used to
synthesize certain amino acids that in turn are
converted into larger molecules
11
B Fermentation
Although glycolysis can function well without
O2 the further oxidation of pyruvate and NADH by
mitochondria requires this gas Thus when O2 is
limiting NADH and pyruvate begin to accumulate
Under this condition plants carry out fermentation
(anaerobic respiration) forming either ethanol or
lactic acid (usually ethanol) As shown in the
picture below the two top reactions consists of a
decarboxylation to form acetaldehide then rapid
reduction of acetaldehide by NADH to form ethanol
These reactions are catalyzed by alcohol
dehydrogenase Some cells contain lactic acid
12
dehydrogenase which uses NADH to reduce pyruvic to
lactic acid Thus either ethanol or lactic acid or
both are fermentation products depending on the
activities of each dehydrogenase present In each
case NADH is the reductant and only under
anaerobic conditions is it abundant anough to cause
reduction Furthermore in some plants NADH is used
to cause accumulation of the other compounds when O2
is limiting especially malate and glycerol
(Salisbury 1969 233)
The fate of pyruvate produced by glycolysis
depends primarily on whether or not oxygen is
present Under normal aerobic conditions pyruvate
is transported into the mitochondrion where it is
further oxidized to CO2 and water transferring its
electrons ultimately to molecular oxygen
13
Although higher plants are obligate aerobs and
are able to tolerate anoxia for only short periods
tissues or organs are occasionally subjected to
anaerobic conditions A typical siuation is that of
roots when the soil is saturated with water When
there is no oxygen to serve as the terminal electron
acceptor mitochondrial respiration will shut down
and metabolism will shift over to fermentation
Fermentation converts pyruvate to either ethanol
through the action of the enzyme alcohol
dehydrogenase (ADH) or lactate via lactate
dehydrogenase (LDH) In most plants the principal
products of fermentation are CO2 and ethanol Some
lactate may be formed primarily in the early stages
of anoxia However lactate lowers he pH of the
cytosol which in turn activates pyruvate
decarboxylase and initiates the production of
ethanol
Either one of the fermentation reactions
consumes the NADH produced earlier in glycolysis by
the oxidation of glyceraldehide-3-P Although this
means there is no net gain of reducing potential in
fermentation This recycling of NADH is still
important to the cell The poolof NADH plus NAD+ in
the cell is relatively small and if the NADH is not
recycled there will be no supply of NAD+ to support
14
the continued oxidation of glyceraldehyde-3-P if
this were the case glycolysis and the producton of
even the small quantities of ATP necessary to
maintain the cells under anaerobic conditions would
then grind to a halt (Hopkins 2009 241)
Soil waterlogging has long been identified as a
major abiotic stress and the constraints that it
provides the roots have a significant effect on the
growth and development of plants When this event
occurs in the spring then these puddles can reduce
seed germination and seedling development Thus a
pool of water is an important factor that affects
the growth development and survival of plant
species not only in natural ecosystems but also
the agricultural and horticultural systems (Dat et
al 2006)
After flooding rapid changes in soil
properties At the time of pore water meets the
land the air pushed out gas diffusion is reduced
and toxic compounds accumulate due to anaerobic
conditions All of these changes greatly affect the
ability of plants to survive In response increased
stomatal resistance photosynthesis and root
hydraulic conductivity decreases and reduced
translocation fotoassimilat Nevertheless one of
15
the best adaptations of plants to hypoxia anoxia
is transitional biochemical and metabolic processes
common to current limited availability of O2 (Dat et
al 2004)
Selective synthesis of a set of about 20
anaerobic stress proteins (ANPS) allows the process
of producing energy without oxygen metabolism under
anaerobic conditions (Subbaiah and Sachs 2003)
Another adaptation is the observed morphological
changes consisting of the formation of hypertrophic
lenticels adventitious root initiation and or
progression of aerenchyma (Vartapetian and Jackson
1997 Jackson and Colmer 2005 Folzer et al 2006)
This review details the diverse plant stress
response to hypoxia anoxia caused by soil
waterlogging flooding and examines some of the key
features of metabolic adaptation physiological and
morphological
16
C Electron
Transport System
We noted earlier that one of the principal
functions of the respiration is to retrieve in
useful form some of the energy initially stored in
assimilates Our traditional measure of useful
energy in most processes is the number of ATP
molecules gained or consumed By this measure alone
the yield from both glycolysis and the citric acid
cycle is quite low After two complete turns of the
cycle one molecule of glucose has been completely
oxidized to six molecules of CO2 but only four
molecules of ATP have been produced (a net of two
ATP from glycolysis plus one for each turn of the
cycle) At this point most of the energy associated
with the glucose molecule has been conserved in the
17
form of electron pairs generated by the oxidation of
glycolytic and citric acid cycle intermediates
In this section we will discuss the third stage
of cellular respirationmdashthe transfer of electrons
fromNADH and FADH2 to oxygen and the accompanying
conversion of redox energy to ATP The transfer of
electrons fromNADHand FADH2 to oxygen involves a
sequence of electron carriers arranged in an
electron transport chainMembrane fractionation
studies have shown that the enzymes and electron
carriers making up the electron transport chain are
organized predominantly into four large
multimolecular complexes (complexes ImdashIV) and two
mobile carriers located in the inner mitochondrial
membrane (Figure 109) In this sense there are a
great number of similarities between the
mitochondrial inner membrane and the thylakoid
membranes of the chloroplast (compare Figure 109
with Figure 76) This is not unexpected since the
principal function of each membrane is energy
transformation and many of the same or similar
components are involved
The Components of the Electron Transport Chain
The electron transport chain of the
mitochondria is the means by which electrons are
18
removed from the reduced carrier NADH and
transferred to oxygen to yield H2O
1) NADH
NADH is generated in the matrix by the reactions
of pyruvate dehydrogenase isocitrate
dehydrogenase α-ketoglutarate dehydrogenase and
malate dehyrogenase The electron transport chain
begins with reoxidizing NADH to form NAD+ and
channeling the electrons into the formation of
reduced coenzymes Important to note that NADH
transfers 2 electrons at a time in the form of a
hydride
NAD+ + 2e- + H+ 1048581 NADH Eorsquo = minus0315 V
2) Flavoproteins
Flavoproteins have either a FAD (flavin adenosine
dinucleotide) or a FMN (flavin mononucleotide)
prosthetic group Flavoproteins can accept or
donate electrons one at time or two at a time
Thus they are often intermediaries between two
electron acceptorsdonors and one electron
acceptorsdonors For flavoproteins the typical
standard reduction potentials are around 0 V
FAD + 2e- + 2H+ 1048581 FADH2 FMN + 2e- + 2H+ 1048581 FMNH2
Eorsquo asymp 0 V
19
3) Coenzyme Q (CoQ)
aka ubiquinone (UQ) Shown below CoQ has ten
repeating isoprene units which make it
insoluble in water but soluble in the
hydrophobic lipid bylayer Coenzyme Q is a
versatile cofactor because it is a soluble
electron carrier in the hydrophobic bilipid layer
of the
inner mitochondrial membrane Like flavoproteins
CoQ can acceptdonate electrons one at a time or
two at a time
Q + 2e- + 2H+ 1048581 QH2 Eorsquo =0060 V
Q + e- + H+ 1048581 1048581QH Eorsquo =0030 V
1048581QH + e- + H+ 1048581 QH2 Eorsquo =0190 V
4) Cytochromes
Cytochromes are proteins that contain heme
prosthetic groups which function as
one electron carriers The heme iron is involved
in one electron transfers involving the Fe2+ and
Fe3+ oxidation states Cytochromes are named by
their absorption spectra which depends
on the porphyrin structure and environment The
example shown is the heme prosthetic group of
cytochrome b Cytochrome b contains the same iron
porphyrin found in hemoglobin and myoglobin
20
Other cytochromes we will encounter in the
electron transport complexes are cytochromes b
c c1 a and a3
5) Iron-Sulfur Proteins
In the electron transport chain we will encounter
many iron-sulfer proteins which participate in
one electron transfers involving the the Fe2+ and
Fe3+ oxidation states These are non-heme iron-
sulfur proteinsThe simplest iron-sulfer protein
is FeS in which iron is tetrahedrally coordinated
by four cysteines The second form is Fe2S2 which
contains two irons complexed to 2 cysteine
residues and two inorganic sulfides The third
form is Fe3S4 which contains 3 iron atoms
coordinated to three cysteine residues and 4
inorganic sulfides The last form is the most
complicated Fe4S4 which contains 4 iron atoms
coordinated to 4 cysteine residues and 4
inorganic sulfides
6) Copper Proteins
Copper bound proteins participate in one electron
transfers involving the Cu+ and Cu2+ oxidation
states
21
Overview of the Electron Transport Chain
Electrons move along the electron transport
chain going from donor to acceptor until they reach
oxygen the ultimate electron acceptor The standard
reduction potentials of the electron carriers are
between the NADHNAD+ couple (-0315 V) and the
oxygenH2O couple (0816 V) as on the next page The
components of the electron transport chain are
organized into 4 complexes Each complex contains
several different electron carriers
1 Complex I also known as the NADH-coenzyme Q
reductase or NADH dehydrogenase
2 Complex II also known as succinate-coenzyme Q
reductase or succinate dehydrogenase
3 Complex III also known as coenzyme Q reductase
4 Complex IV also known as cytochrome c reductase
The free energy necessary to generate ATP
is extracted from the oxidation of NADH and
FADH2 by the electron transport chain It
consists of four protein complexes and single
protein chain of cytochrome c through which
electrons pass from lower to higher standard
redox potentials Electrons are carried from complex
I and II to complex III by the membrane (lipid)
soluble coenzyme Q (CoQ or ubiquinone) and between
22
complexes III and IV by the peripheral membrane
protein cytochrome c
In the following I give the overall
reactions catalysed by corresponding protein
complexes in the mitochondrial electron transport
chain
1 Complex I
Complex I is also called NADH-Coenzyme Q
reductase because this large protein complex
transfers 2 electrons from NADH to coenzyme Q
Complex I was formerly known as NADH dehydrogenase
Complex I is huge 850000 kD and is composed of more
than thirty subunits It contains a FMN prosthetic
group and seven or more Fe-S clusters This complex
has between 20-26 iron atoms boundThe prosthetic
group FMN is absolutely required for activity
Therefore this complex is a flavoprotein This
complex binds NADH transfers two electrons in the
form of a hydride to FMN to produce NAD+ and FMNH2
23
The subsequent steps involve the transfer of
electrons one at a time to a series of iron-sulfer
complexes that includes both 2Fe-2S and 4Fe-4S
clusters Note the importance of FMN First it
functions as a 2 electron acceptor in the hydride
transfer from NADH Second it functions as a 1
electron donor to the series of iron sulfur clusters
FMN and FAD often play crucial links between 2
electron transfer agents and 1 electron transfer
agents The final step of this complex is the
transfer of 2 electrons one at a time to coenzyme Q
CoQ like FMN and FAD can function as a 2 electron
donoracceptor and as a 1 electron donoracceptor
CoQ is a mobile electron carrier because its
isoprenoid tail makes it highly hydrophobic and
lipophillic It diffuses freely in the bilipid layer
of the inner mitochondrial membrane
The process of transferring electrons from NADH
to CoQ by complex I results in the net transport of
protons from the matrix side of the inner
mitochondrial membrane to the inter membrane space
where the H+ ions accumulate generating a proton
motive force The intermembrane space side of the
inner membrane is referred to as the P face (P
standing for positive) The matrix side of the inner
membrane is referred to as the the N face The
24
transport of electrons from NADH to CoQ is coupled to
the transport of protons across the membrane This is
an example of active transport The stiochiometry is
4 H+ transported per 2 electrons
2 Complex II
Believe it or not you are already familiar
with Complex II It is none other than succinate
dehydrogenase The only enzyme of the citric acid
cycle that is an integral membrane protein This
complex is composed of four subunits 2 of which are
iron-sulfur proteins and the other two subunits
together bind FAD through a covalent link to a
histidine residue These two subunits are called
flavoprotein 2 or FP2 Complex II contains 3 Fe-S
centers 1 4Fe-4S cluster 1 3Fe-4S cluster and 1
2Fe- 2S cluster In the first step of this complex
succinate is bound and a hydride is transferred to
FAD to generate FADH2 and fumarate FADH2 then
transfers its electrons one at a time to the Fe-S
centers Thus once again FAD functions as 2 electron
acceptor and a 1 electron donor The final step of
this complex is the transfer of 2 electrons one at a
time to coenzyme Q to produce CoQH2 For complex II
the standard free energy change of the overall
reaction is too small to drive the transport of
protons across the inner mitochondrial membrane This
25
accounts for the 15 ATPrsquos generated per FADH2
compared with the 25 ATPrsquos generated per NADH
3 Complex III
This complex is also known as coenzyme Q-
cytochrome c reductase because it passes the
electrons form CoQH2 to cyt c through a very unique
electron transport pathway called the Q-cycle Shown
to the left are the porphyrins found in cytochromes
Cytochrome b contains the same iron protoporphyrin as
hemoglobin and myoglobin The c cytochromes contain
heme c through covalent attachment by cysteine
residues Cytochrome a is found in two forms in
complex IV In complex III we find two b-type
cytochromes and one c-type cytochrome Complex III is
complex and we have a crystal structure
4 Complex IV
Complex IV is also known as cytochrome c oxidase
because it accepts the electrons from cytochrome c
and directs them towards the four electron reduction
of O2 to form 2 molecules of H2O
4 cyt c (Fe2+) + 4 H+ + O2 4 cyt c (Fe3+) +
2H2O
26
Cytochrome c oxidase contains 2 heme
centerscytochrome a and cytochrome a3 and two copper
proteins Each of the protein bound coppers are
associated with one of the cytochromes The copper
sites are called CuA and CuB CuA is associated with
cytochrome a and is shown to the left CuB is
associated with cytochrome a3 The copper sites
function as 1 electron carriers cycling between the
cuprous state Cu+ and the cupric state Cu2+ Just
like iron containing proteins they transfer
electrons one at a time Cytochrome c is bound on the
P-face of the membrane and transfers its electron to
CuA The oxidized cytochrome c dissociates CuA then
transfers the electron to cytochrome a The protein
bound CuA and the iron bound in cytochrome a are 15 Aring
apart In contrast the CuB and the iron bound in
cytochrome a3 are very close to each other forming a
binuclear metal center shown below
27
Electron transport chains are associated with
membranes such as the mitochondrial membrane in
eukaryotic organisms the thylakoid membrane of
photosynthetic organisms and the plasma membrane of
prokaryotic organisms While some of the details
differ electron transport chains in all organisms
operate on the same basic principles
Excited electrons are brought to the electron
transport chain by electron carriers such as NADH
and FADH2 In photosynthetic organisms the electrons
are donated by water and excited by light energy
The electron donor is oxidized as the electrons are
passed to the first protein complex in the chain As
it accepts the electrons the protein complex is
reduced Next the electrons are passed from the
28
first protein complex which becomes oxidized to
the second protein complex which becomes reduced
Each of these redox reactions is slightly exergonic
or energy releasing Some of this energy is used to
move protons across the membrane against their
concentration gradient With each successive
transfer of electrons the original excited
electrons lose some of their energy After several
successive redox reactions the electrons are
donated to a terminal electron acceptor In aerobic
respiration the terminal electron acceptor is
oxygen In photosynthesis the terminal electron
acceptor is a compound called NADP
The energy in the excited electrons has been
transformed into an electrochemical gradient
represented by the high concentration of protons on
one side of the membrane These protons can diffuse
back across the membrane by passing through a
protein complex called ATP synthase As these
protons pass through ATP synthase down their
concentration gradient they provide the energy
needed to make ATP from ADP and inorganic phosphate
within the cell
29
CHAPTER III
CLOSING
A Conclution
When a hexose is completely oxidized to CO2
and H2O using these three processes Glycolysis
yields two ATP and two NADH per hexose used Each
such NADH oxidized by the electron transport system
30
yields two ATP as dscribed above so glycolysis
contributes a total of six ATP per hexose The Krebs
cycle contributes two ATP per hexose or per two
pyruvates when succinyl CoA is cleaved to succinate
and CoASH This cycle also produces eight NADH per
hexose within the mitochondrial matrix by oxidative
phosporylation each of these NADH yields three ATP
or 24 per hexose Each FADH2 from the Krebs cycle
yields two ATP by oxydative phosporylation or four
per hexose (two pyruvates) The total contribution of
the Krebs cycle is then 30 Adding these 30 to the 6
from glycolysis leads to a total of 36 per hexose
completely respired by these process
B Suggestion
My greatest wish is that the readers find
this manuscript helpful in gaining the knowledge they
need for effective learning and practice and in
preparing for examination designed to test that
knowledge Reader comments and suggestions are
welcome and would be very helpful in improving our
manuscript Your contribution may assist many other
students in the coming years
31
REFERENCE
Anonim 2013 Anaerobic Respiration
httpwwwscoolcoukgcsebiologyrespiration
revise-itanaerobic-respiration Accessed
September 10th 2013
Anonim 2013 What is the difference between aerobic
and anaerobic respiration in plants
httpwwwbiologylifeeasyorg4010difference
-between-aerobic-anaerobic-respiration-plants
Accessed September 10th 2013
Campbell Mary K 2006 Biochemistry New York Thomson
Brooks Cole Inc
Dat J Capelli N Folzer H Bourgeade P Badot P-M
(2004) Sensing and signaling during
plantflooding Plant Physiology and
Biochemistry 42 273-282
Dat J Folzer H Parent C Badot P-M Capelli N
(2006) HypoxiastressCurrent Understanding and
Perspectives In Teixeira da Silva JA (Ed)
Floriculture Ornamental and Plant
Biotechnology Advances and Topical Issues (Vol
3) Global Science Books Isleworth United
Kingdompp 664-674
Hatta Muhammad 2011 Dunia Pertanian
httpemhattawordpresscomcategorystress
Accessed September 10th 2013
Hopkins William G 2009 Introduction to Plant Physiology
New York John Wiley amp Sons Inc
Lakitan Benyamin 2011 Dasar-Dasar Fisiologi Tumbuhan
Jakarta PT Raja Grafindo Persada
Larcher W 1980 Physiological Plant Ecology New York
Springer-Verlag
Raven 2013 Biology Fifth Edition
httphigheredmcgraw-hillcomsites983409233
9student_view0chapter39
photosynthetic_electron_transport_and_atp_synth
esishtml Accessed September 10th 2013
Salisbury Frank B 1969 Plant Physiology Third Edition
California Wadsworth Publishing Company
Subbaiah C Sachs M (2003) Molecular and cellular
adaptations of maize to flooding stress Annals
of Botany 91 119-127
Wilson CarlL1962 Botany Third EditionUnited State of
America Holt Rinch Inc

__________________________
CONTENT
PREFACE i
CONTENT ii
CHAPTER I INTRODUCTION
A Background 1
B Purposes 2
C Benefits 2
CHAPTER II PLANT RESPIRATION
A Krebs Cycle 3
B Fermentation 8
C Electron Transport System
11
CHAPTER III CLOSING
ii
A Conclution
20
B Suggestion
20
REFERENCE
iii
CHAPTER I
INTRODUCTION
A Background
Connected with photosynthesis a metabolic
process takes place in chloroplast-containing plant
cells which like respiration takes up O2 and
releases CO2 in the light but which contrary to
respiration ceases in the dark This O2CO2 gas
exchange has been called right respiration or
photorespiration The substrate for the
photorespiratory metabolism is again ribulose
biphosphate which can be an acceptor not only for
CO2 but also for O2 By taking up oxygen RuBP is
split into PGA and phospoglycolate The supply of O2
and CO2 regulates the relationship between acceptor
oxidation (photorespiration) and acceptor
carboxylation (photosynthesis) via the enzyme
complex RuBP carboxylaseoxygenase High partial
pressure of O2 favors photorespiration A large
supply of CO2 favors photosynthesis The formation
of phospoglycolate is dependent on the supply of
RuBP via the Calvin cycle the photorespiratory O2
uptake and CO2 release increase with light
intensity
1
The process of glycolate metabolism has not
yet been understood in all details The glycolate is
transported out of the chloroplasts phospoglycolate
and phospate The glycolate is transported out of
the chloroplast into peroxisomes cell compartments
about the size of mitochondria which contain
glycolate oxidase catalase and transaminases In
the peroxysomes when O2 is taken up glycolate is
oxidized to glyoxylate and the peroxide thus
produced is detoxified by catalase Glycolate can
either be completely reduced via oxalate by further
O2 uptake or transformed to glycine by
transamination Glycine is transported from the
peroxisomes into the mitochondria where two
molecules of glycine are coupled to form one
molecule of serine with the release of CO2 Serine
is taken over by the amino acid metabolism or
2
converted to glycerate after deamination by
hydropyruvate This can be photophosporylated in the
chloroplasts and returned to the Calvin cycle or
used elsewhere (Larcher 1980 81-82)
B Purposes
This manuscript is intended to complete the
task from our lecturer Dr Azwir Anhar MSi and
to meet the needs of university students as the
guidersquos book in learning and practice activity for
the member of International Standard Teacher
Education (ISTE) program in Public University of
Padang
C Benefits
Benefits that we gained from this manuscript
are
1 As reference in learning process of plant
phisiology subject especially in plant
respiration title
2 As information about steps in plant
respiration
3
CHAPTER II
PLANT RESPIRATION
A Krebs Cycle
The Krebs cycle was named in honor of the
English biochemist Hans A Krebs who in 1937
proposed a cycle of reactions to explain how pyruvate
breakdown takes place in the breast muscle of
pigeons He called his proposed pathway the citric
acid cycle because citric acid is an important
intermediet Another common name for the same group
of reactions is the Tricarboxuylic Acid (TCA) cycle
a term used because citric and isocitric acids have
three carboxyl groups It was not until the early
1950s that mitochondria capable of carrying out this
cycle were isolated from plant cells
The initial step leading to the Krebs cycle
involves the oxidation and loss of CO2 from pyruvate
and the combination of the remaining 2-carbon acetate
unit with a sulfur-containing compound co-enzyme A
(CoA) forming acetyl CoA This and another
comparable role of CoA in the Krebs cycle are
important reasons why sulfur is an essential element
4
The reaction of pyruvate decarboxylation also
involves a phosporylated form of thiamine (vitamin
B1) as a prostethic group Participation of thiamine
in this section partially explains the essential
function of vitamin B1 in plants and animals Besides
the loss of CO2 two hydrogen atoms are removed from
pyruvic acid during the formation of Acetyl CoA The
enzyme catalyzing the complete reaction is called
pyruvic acid dehydrogenase but it is actually an
organized complex containing numerous copies of three
or four different enzymes The hydrogen atoms removed
are finally accepted by NAD+ yielding NADH
The Krebs cycle accomplishes removal of some of
the electrons from organic acid intermediates and
transfer of these electrons to NAD+ or FAD Notice
that none of dehydrogenase enzyme of the cycle uses
NADP+ as an electron acceptor In fact NADP+ is
usually undetectable in plant mitochondria a
situation opposite to that of chloroplasts where
NADP+ is abundant but where there is much less NAD+
Not only are NADH and FADH2 important products of the
Krebs cycle but one molecule of ATP is formed from
ADP and Pi during the conversion of succinyl coenzyme
A to succinic acid Two additional CO2 molecules are
released in these Krebs cycle reactions so there is
a net loss of both carbon atoms from the incoming
5
acetate of acetyl CoA The release of CO2 in the
Krebs cycle accounts for the product of CO2 in the
summary equation for resspiration but no O2 is
absorbed during any Kreb cycle reaction (Lakitan
2011)
During metabolism the synthesis and breakdown
of different organic compounds takes place through
various pathways like the breakdown and synthesis of
proteins carbohydrates fats and nucleic acids
These different pathways and intermediates are also
responsible for the production of energyKrebrsquos
cycle named after Hans Krebs who began working out
its details in 1930s is a series of reactions in
which the pyruvate from glycolysis is oxidized to Co2
under aerobic conditions Kreb cycle is also known as
citric acid cycle or Tri-carboxylic acid cycle (TCA)
( Galston 1968 76)
6
Steps involved in the process of Kreb cycle are
1 Fate of pyruvic acid
Pyruvic acid can form different compounds by
different pathways ie it can be converted into
lactic acid It can be converted into acetyl
coenzyme A as a result of oxidation
2 Fate of Acetyl CoA
It can either undergo condensation with itself or
its derivatives to form fatty acids having 14 ndash 20
carbons Acetyl CoA can also go through a series
of reactions in Krebs cycle
3 Formation of citrate
Acetyl CoA enzyme condenses with oxaloacetate by
an enzyme citrate synthatase to form citrate with
the release of acetyl CoA If the amount of
7
oxaloacetate is very small then small number of
acetyl CoA would be reacting with oxaloacetate
leaving surplus acetyl CoA to go through another
pathway for the formation of long chain fatty
acids
4 Formation of C is ndash acotinate and iso-citrate
Citrate is changed first into C is ndash acotinate and
then to iso-citrate under the enzyme acotinase
Equilibrium is established between citrate C is
aconitate and iso-citrate It has been observed
that most of the time this equilibrium is shifted
towards the iso-citrate If the concentration of
iso-citrate in increased the formation of citrate
will result which also indicates that the
equilibrium also shift in the reserve direction
5 Formation of oxalosucinate
Iso-citrate is acted upon by an enzyme iso-citrate
dehydrogenase using nicotinamide adenine
dinucleotide (NAD) as coenzyme As a result iso-
citrate is converted into oxalosucciate and NAD is
reduced to NADH2 Similar reaction is carried out
by same enzyme using nicotindmide adenine
dinucleotide phosphate (NADP) as coenzyme which is
reduced to NDAPH2
6 Formation of α-Ketoglutarate
8
Oxalosucciante is changed into α-ketoglutrate by
iso-citrate dehydrogenase with the help of
coenzyme NAD or NADP In this reaction carbon
dioxide and NADH2 or NADPH2 are also released
7 Formation of succinyl CoA
α-ketoglutarate combines with acetyl CoA in the
presence of coenzyme NAD and enzyme α-
keoglutaratedehyrogenase to form succinyl coenzyme
a carbon dioxide and NADH2
8 Formation of Succinate
Later on coenzyme a is removed from succinyl
coenzyme A in the presence of guanosinediphosphate
(GDP) and inorganic phosphate to form succinate
and guanosine triphosphate (GTP) This reaction is
carried out by an enzyme called succinyl CoA
synthatase
9 Formation of Funarate
An enzyme succinic dehydrogenase removes hydrogen
from succinate to form funarate
10 Formation of Malate
Fumarate reacts with water in the presence of
enzyme fumarase to form malate
11 Regeneration of oxaloacetate
Malate is oxidized by malic dehydrogenase and NAD
forming oxaloacetate and NADH2 Thus oxalo acetate
is again available to start another cycle
9
(httpkrebs-cycle-or-citric-acid-cycle-or-
trihtml)
Importance of CTA Cycle
1 Source of energy
In addition to routine organic compounds described
above by products like nicotinanide adenine
dinucleotide (NADH2) and guanosine triphosphate
(GTP) are the source of biological energy NADH2
after oxidation produce energy whereas GTP is
itself high energy phosphate compound
2 Oxidation of organic compounds taken as food
Oxidation of fats carbohydrates and proteins take
place through it or in other words it can be said
that oxidation of all compounds having carbon
atoms can take place through TCA cycle Some of
amino acids like alanine glutamic acid and
aspartic acid at one stage or the other enters
into TCA cycle eg glutamic acid enters cycle
after its transformation into α-ketoglutarate
Similarly alanine enters the cycle after its
conversion into pyruvate
3 Intermediate compounds
TCA cycle is also involved in synthesis of
intermediate compounds leading to the formation of
larger molecules (Ray 1963 17)
10
Verification of Krebs Cycle
It was done by radioactive traces like C14 as
radioactive carbon dioxide of different levels and
reactions After addition of radioactive carbon
dioxide different chemical compounds produced like
glucose fats amino acids were isolated and looked
for radioactive carbon In this way whole of
metabolic reaction were verified including individual
reactions alternative metabolic pathways
intermediates of fats carbohydrates and amino acids
etc in the body cells as well as in test tubes
(Wilson 1962 99)
The primary function of the Krebs cycle are as
follows
1 Reduction of NAD+ and FAD to the electron donors
NADH and FADH2 that are subsequently oxidized to
yield ATP
2 Direct synthesis of a limited amount of ATP (1 ATP
for each pyruvate oxidized)
3 Formation of carbon skeletons that can be used to
synthesize certain amino acids that in turn are
converted into larger molecules
11
B Fermentation
Although glycolysis can function well without
O2 the further oxidation of pyruvate and NADH by
mitochondria requires this gas Thus when O2 is
limiting NADH and pyruvate begin to accumulate
Under this condition plants carry out fermentation
(anaerobic respiration) forming either ethanol or
lactic acid (usually ethanol) As shown in the
picture below the two top reactions consists of a
decarboxylation to form acetaldehide then rapid
reduction of acetaldehide by NADH to form ethanol
These reactions are catalyzed by alcohol
dehydrogenase Some cells contain lactic acid
12
dehydrogenase which uses NADH to reduce pyruvic to
lactic acid Thus either ethanol or lactic acid or
both are fermentation products depending on the
activities of each dehydrogenase present In each
case NADH is the reductant and only under
anaerobic conditions is it abundant anough to cause
reduction Furthermore in some plants NADH is used
to cause accumulation of the other compounds when O2
is limiting especially malate and glycerol
(Salisbury 1969 233)
The fate of pyruvate produced by glycolysis
depends primarily on whether or not oxygen is
present Under normal aerobic conditions pyruvate
is transported into the mitochondrion where it is
further oxidized to CO2 and water transferring its
electrons ultimately to molecular oxygen
13
Although higher plants are obligate aerobs and
are able to tolerate anoxia for only short periods
tissues or organs are occasionally subjected to
anaerobic conditions A typical siuation is that of
roots when the soil is saturated with water When
there is no oxygen to serve as the terminal electron
acceptor mitochondrial respiration will shut down
and metabolism will shift over to fermentation
Fermentation converts pyruvate to either ethanol
through the action of the enzyme alcohol
dehydrogenase (ADH) or lactate via lactate
dehydrogenase (LDH) In most plants the principal
products of fermentation are CO2 and ethanol Some
lactate may be formed primarily in the early stages
of anoxia However lactate lowers he pH of the
cytosol which in turn activates pyruvate
decarboxylase and initiates the production of
ethanol
Either one of the fermentation reactions
consumes the NADH produced earlier in glycolysis by
the oxidation of glyceraldehide-3-P Although this
means there is no net gain of reducing potential in
fermentation This recycling of NADH is still
important to the cell The poolof NADH plus NAD+ in
the cell is relatively small and if the NADH is not
recycled there will be no supply of NAD+ to support
14
the continued oxidation of glyceraldehyde-3-P if
this were the case glycolysis and the producton of
even the small quantities of ATP necessary to
maintain the cells under anaerobic conditions would
then grind to a halt (Hopkins 2009 241)
Soil waterlogging has long been identified as a
major abiotic stress and the constraints that it
provides the roots have a significant effect on the
growth and development of plants When this event
occurs in the spring then these puddles can reduce
seed germination and seedling development Thus a
pool of water is an important factor that affects
the growth development and survival of plant
species not only in natural ecosystems but also
the agricultural and horticultural systems (Dat et
al 2006)
After flooding rapid changes in soil
properties At the time of pore water meets the
land the air pushed out gas diffusion is reduced
and toxic compounds accumulate due to anaerobic
conditions All of these changes greatly affect the
ability of plants to survive In response increased
stomatal resistance photosynthesis and root
hydraulic conductivity decreases and reduced
translocation fotoassimilat Nevertheless one of
15
the best adaptations of plants to hypoxia anoxia
is transitional biochemical and metabolic processes
common to current limited availability of O2 (Dat et
al 2004)
Selective synthesis of a set of about 20
anaerobic stress proteins (ANPS) allows the process
of producing energy without oxygen metabolism under
anaerobic conditions (Subbaiah and Sachs 2003)
Another adaptation is the observed morphological
changes consisting of the formation of hypertrophic
lenticels adventitious root initiation and or
progression of aerenchyma (Vartapetian and Jackson
1997 Jackson and Colmer 2005 Folzer et al 2006)
This review details the diverse plant stress
response to hypoxia anoxia caused by soil
waterlogging flooding and examines some of the key
features of metabolic adaptation physiological and
morphological
16
C Electron
Transport System
We noted earlier that one of the principal
functions of the respiration is to retrieve in
useful form some of the energy initially stored in
assimilates Our traditional measure of useful
energy in most processes is the number of ATP
molecules gained or consumed By this measure alone
the yield from both glycolysis and the citric acid
cycle is quite low After two complete turns of the
cycle one molecule of glucose has been completely
oxidized to six molecules of CO2 but only four
molecules of ATP have been produced (a net of two
ATP from glycolysis plus one for each turn of the
cycle) At this point most of the energy associated
with the glucose molecule has been conserved in the
17
form of electron pairs generated by the oxidation of
glycolytic and citric acid cycle intermediates
In this section we will discuss the third stage
of cellular respirationmdashthe transfer of electrons
fromNADH and FADH2 to oxygen and the accompanying
conversion of redox energy to ATP The transfer of
electrons fromNADHand FADH2 to oxygen involves a
sequence of electron carriers arranged in an
electron transport chainMembrane fractionation
studies have shown that the enzymes and electron
carriers making up the electron transport chain are
organized predominantly into four large
multimolecular complexes (complexes ImdashIV) and two
mobile carriers located in the inner mitochondrial
membrane (Figure 109) In this sense there are a
great number of similarities between the
mitochondrial inner membrane and the thylakoid
membranes of the chloroplast (compare Figure 109
with Figure 76) This is not unexpected since the
principal function of each membrane is energy
transformation and many of the same or similar
components are involved
The Components of the Electron Transport Chain
The electron transport chain of the
mitochondria is the means by which electrons are
18
removed from the reduced carrier NADH and
transferred to oxygen to yield H2O
1) NADH
NADH is generated in the matrix by the reactions
of pyruvate dehydrogenase isocitrate
dehydrogenase α-ketoglutarate dehydrogenase and
malate dehyrogenase The electron transport chain
begins with reoxidizing NADH to form NAD+ and
channeling the electrons into the formation of
reduced coenzymes Important to note that NADH
transfers 2 electrons at a time in the form of a
hydride
NAD+ + 2e- + H+ 1048581 NADH Eorsquo = minus0315 V
2) Flavoproteins
Flavoproteins have either a FAD (flavin adenosine
dinucleotide) or a FMN (flavin mononucleotide)
prosthetic group Flavoproteins can accept or
donate electrons one at time or two at a time
Thus they are often intermediaries between two
electron acceptorsdonors and one electron
acceptorsdonors For flavoproteins the typical
standard reduction potentials are around 0 V
FAD + 2e- + 2H+ 1048581 FADH2 FMN + 2e- + 2H+ 1048581 FMNH2
Eorsquo asymp 0 V
19
3) Coenzyme Q (CoQ)
aka ubiquinone (UQ) Shown below CoQ has ten
repeating isoprene units which make it
insoluble in water but soluble in the
hydrophobic lipid bylayer Coenzyme Q is a
versatile cofactor because it is a soluble
electron carrier in the hydrophobic bilipid layer
of the
inner mitochondrial membrane Like flavoproteins
CoQ can acceptdonate electrons one at a time or
two at a time
Q + 2e- + 2H+ 1048581 QH2 Eorsquo =0060 V
Q + e- + H+ 1048581 1048581QH Eorsquo =0030 V
1048581QH + e- + H+ 1048581 QH2 Eorsquo =0190 V
4) Cytochromes
Cytochromes are proteins that contain heme
prosthetic groups which function as
one electron carriers The heme iron is involved
in one electron transfers involving the Fe2+ and
Fe3+ oxidation states Cytochromes are named by
their absorption spectra which depends
on the porphyrin structure and environment The
example shown is the heme prosthetic group of
cytochrome b Cytochrome b contains the same iron
porphyrin found in hemoglobin and myoglobin
20
Other cytochromes we will encounter in the
electron transport complexes are cytochromes b
c c1 a and a3
5) Iron-Sulfur Proteins
In the electron transport chain we will encounter
many iron-sulfer proteins which participate in
one electron transfers involving the the Fe2+ and
Fe3+ oxidation states These are non-heme iron-
sulfur proteinsThe simplest iron-sulfer protein
is FeS in which iron is tetrahedrally coordinated
by four cysteines The second form is Fe2S2 which
contains two irons complexed to 2 cysteine
residues and two inorganic sulfides The third
form is Fe3S4 which contains 3 iron atoms
coordinated to three cysteine residues and 4
inorganic sulfides The last form is the most
complicated Fe4S4 which contains 4 iron atoms
coordinated to 4 cysteine residues and 4
inorganic sulfides
6) Copper Proteins
Copper bound proteins participate in one electron
transfers involving the Cu+ and Cu2+ oxidation
states
21
Overview of the Electron Transport Chain
Electrons move along the electron transport
chain going from donor to acceptor until they reach
oxygen the ultimate electron acceptor The standard
reduction potentials of the electron carriers are
between the NADHNAD+ couple (-0315 V) and the
oxygenH2O couple (0816 V) as on the next page The
components of the electron transport chain are
organized into 4 complexes Each complex contains
several different electron carriers
1 Complex I also known as the NADH-coenzyme Q
reductase or NADH dehydrogenase
2 Complex II also known as succinate-coenzyme Q
reductase or succinate dehydrogenase
3 Complex III also known as coenzyme Q reductase
4 Complex IV also known as cytochrome c reductase
The free energy necessary to generate ATP
is extracted from the oxidation of NADH and
FADH2 by the electron transport chain It
consists of four protein complexes and single
protein chain of cytochrome c through which
electrons pass from lower to higher standard
redox potentials Electrons are carried from complex
I and II to complex III by the membrane (lipid)
soluble coenzyme Q (CoQ or ubiquinone) and between
22
complexes III and IV by the peripheral membrane
protein cytochrome c
In the following I give the overall
reactions catalysed by corresponding protein
complexes in the mitochondrial electron transport
chain
1 Complex I
Complex I is also called NADH-Coenzyme Q
reductase because this large protein complex
transfers 2 electrons from NADH to coenzyme Q
Complex I was formerly known as NADH dehydrogenase
Complex I is huge 850000 kD and is composed of more
than thirty subunits It contains a FMN prosthetic
group and seven or more Fe-S clusters This complex
has between 20-26 iron atoms boundThe prosthetic
group FMN is absolutely required for activity
Therefore this complex is a flavoprotein This
complex binds NADH transfers two electrons in the
form of a hydride to FMN to produce NAD+ and FMNH2
23
The subsequent steps involve the transfer of
electrons one at a time to a series of iron-sulfer
complexes that includes both 2Fe-2S and 4Fe-4S
clusters Note the importance of FMN First it
functions as a 2 electron acceptor in the hydride
transfer from NADH Second it functions as a 1
electron donor to the series of iron sulfur clusters
FMN and FAD often play crucial links between 2
electron transfer agents and 1 electron transfer
agents The final step of this complex is the
transfer of 2 electrons one at a time to coenzyme Q
CoQ like FMN and FAD can function as a 2 electron
donoracceptor and as a 1 electron donoracceptor
CoQ is a mobile electron carrier because its
isoprenoid tail makes it highly hydrophobic and
lipophillic It diffuses freely in the bilipid layer
of the inner mitochondrial membrane
The process of transferring electrons from NADH
to CoQ by complex I results in the net transport of
protons from the matrix side of the inner
mitochondrial membrane to the inter membrane space
where the H+ ions accumulate generating a proton
motive force The intermembrane space side of the
inner membrane is referred to as the P face (P
standing for positive) The matrix side of the inner
membrane is referred to as the the N face The
24
transport of electrons from NADH to CoQ is coupled to
the transport of protons across the membrane This is
an example of active transport The stiochiometry is
4 H+ transported per 2 electrons
2 Complex II
Believe it or not you are already familiar
with Complex II It is none other than succinate
dehydrogenase The only enzyme of the citric acid
cycle that is an integral membrane protein This
complex is composed of four subunits 2 of which are
iron-sulfur proteins and the other two subunits
together bind FAD through a covalent link to a
histidine residue These two subunits are called
flavoprotein 2 or FP2 Complex II contains 3 Fe-S
centers 1 4Fe-4S cluster 1 3Fe-4S cluster and 1
2Fe- 2S cluster In the first step of this complex
succinate is bound and a hydride is transferred to
FAD to generate FADH2 and fumarate FADH2 then
transfers its electrons one at a time to the Fe-S
centers Thus once again FAD functions as 2 electron
acceptor and a 1 electron donor The final step of
this complex is the transfer of 2 electrons one at a
time to coenzyme Q to produce CoQH2 For complex II
the standard free energy change of the overall
reaction is too small to drive the transport of
protons across the inner mitochondrial membrane This
25
accounts for the 15 ATPrsquos generated per FADH2
compared with the 25 ATPrsquos generated per NADH
3 Complex III
This complex is also known as coenzyme Q-
cytochrome c reductase because it passes the
electrons form CoQH2 to cyt c through a very unique
electron transport pathway called the Q-cycle Shown
to the left are the porphyrins found in cytochromes
Cytochrome b contains the same iron protoporphyrin as
hemoglobin and myoglobin The c cytochromes contain
heme c through covalent attachment by cysteine
residues Cytochrome a is found in two forms in
complex IV In complex III we find two b-type
cytochromes and one c-type cytochrome Complex III is
complex and we have a crystal structure
4 Complex IV
Complex IV is also known as cytochrome c oxidase
because it accepts the electrons from cytochrome c
and directs them towards the four electron reduction
of O2 to form 2 molecules of H2O
4 cyt c (Fe2+) + 4 H+ + O2 4 cyt c (Fe3+) +
2H2O
26
Cytochrome c oxidase contains 2 heme
centerscytochrome a and cytochrome a3 and two copper
proteins Each of the protein bound coppers are
associated with one of the cytochromes The copper
sites are called CuA and CuB CuA is associated with
cytochrome a and is shown to the left CuB is
associated with cytochrome a3 The copper sites
function as 1 electron carriers cycling between the
cuprous state Cu+ and the cupric state Cu2+ Just
like iron containing proteins they transfer
electrons one at a time Cytochrome c is bound on the
P-face of the membrane and transfers its electron to
CuA The oxidized cytochrome c dissociates CuA then
transfers the electron to cytochrome a The protein
bound CuA and the iron bound in cytochrome a are 15 Aring
apart In contrast the CuB and the iron bound in
cytochrome a3 are very close to each other forming a
binuclear metal center shown below
27
Electron transport chains are associated with
membranes such as the mitochondrial membrane in
eukaryotic organisms the thylakoid membrane of
photosynthetic organisms and the plasma membrane of
prokaryotic organisms While some of the details
differ electron transport chains in all organisms
operate on the same basic principles
Excited electrons are brought to the electron
transport chain by electron carriers such as NADH
and FADH2 In photosynthetic organisms the electrons
are donated by water and excited by light energy
The electron donor is oxidized as the electrons are
passed to the first protein complex in the chain As
it accepts the electrons the protein complex is
reduced Next the electrons are passed from the
28
first protein complex which becomes oxidized to
the second protein complex which becomes reduced
Each of these redox reactions is slightly exergonic
or energy releasing Some of this energy is used to
move protons across the membrane against their
concentration gradient With each successive
transfer of electrons the original excited
electrons lose some of their energy After several
successive redox reactions the electrons are
donated to a terminal electron acceptor In aerobic
respiration the terminal electron acceptor is
oxygen In photosynthesis the terminal electron
acceptor is a compound called NADP
The energy in the excited electrons has been
transformed into an electrochemical gradient
represented by the high concentration of protons on
one side of the membrane These protons can diffuse
back across the membrane by passing through a
protein complex called ATP synthase As these
protons pass through ATP synthase down their
concentration gradient they provide the energy
needed to make ATP from ADP and inorganic phosphate
within the cell
29
CHAPTER III
CLOSING
A Conclution
When a hexose is completely oxidized to CO2
and H2O using these three processes Glycolysis
yields two ATP and two NADH per hexose used Each
such NADH oxidized by the electron transport system
30
yields two ATP as dscribed above so glycolysis
contributes a total of six ATP per hexose The Krebs
cycle contributes two ATP per hexose or per two
pyruvates when succinyl CoA is cleaved to succinate
and CoASH This cycle also produces eight NADH per
hexose within the mitochondrial matrix by oxidative
phosporylation each of these NADH yields three ATP
or 24 per hexose Each FADH2 from the Krebs cycle
yields two ATP by oxydative phosporylation or four
per hexose (two pyruvates) The total contribution of
the Krebs cycle is then 30 Adding these 30 to the 6
from glycolysis leads to a total of 36 per hexose
completely respired by these process
B Suggestion
My greatest wish is that the readers find
this manuscript helpful in gaining the knowledge they
need for effective learning and practice and in
preparing for examination designed to test that
knowledge Reader comments and suggestions are
welcome and would be very helpful in improving our
manuscript Your contribution may assist many other
students in the coming years
31
REFERENCE
Anonim 2013 Anaerobic Respiration
httpwwwscoolcoukgcsebiologyrespiration
revise-itanaerobic-respiration Accessed
September 10th 2013
Anonim 2013 What is the difference between aerobic
and anaerobic respiration in plants
httpwwwbiologylifeeasyorg4010difference
-between-aerobic-anaerobic-respiration-plants
Accessed September 10th 2013
Campbell Mary K 2006 Biochemistry New York Thomson
Brooks Cole Inc
Dat J Capelli N Folzer H Bourgeade P Badot P-M
(2004) Sensing and signaling during
plantflooding Plant Physiology and
Biochemistry 42 273-282
Dat J Folzer H Parent C Badot P-M Capelli N
(2006) HypoxiastressCurrent Understanding and
Perspectives In Teixeira da Silva JA (Ed)
Floriculture Ornamental and Plant
Biotechnology Advances and Topical Issues (Vol
3) Global Science Books Isleworth United
Kingdompp 664-674
Hatta Muhammad 2011 Dunia Pertanian
httpemhattawordpresscomcategorystress
Accessed September 10th 2013
Hopkins William G 2009 Introduction to Plant Physiology
New York John Wiley amp Sons Inc
Lakitan Benyamin 2011 Dasar-Dasar Fisiologi Tumbuhan
Jakarta PT Raja Grafindo Persada
Larcher W 1980 Physiological Plant Ecology New York
Springer-Verlag
Raven 2013 Biology Fifth Edition
httphigheredmcgraw-hillcomsites983409233
9student_view0chapter39
photosynthetic_electron_transport_and_atp_synth
esishtml Accessed September 10th 2013
Salisbury Frank B 1969 Plant Physiology Third Edition
California Wadsworth Publishing Company
Subbaiah C Sachs M (2003) Molecular and cellular
adaptations of maize to flooding stress Annals
of Botany 91 119-127
Wilson CarlL1962 Botany Third EditionUnited State of
America Holt Rinch Inc

A Conclution
20
B Suggestion
20
REFERENCE
iii
CHAPTER I
INTRODUCTION
A Background
Connected with photosynthesis a metabolic
process takes place in chloroplast-containing plant
cells which like respiration takes up O2 and
releases CO2 in the light but which contrary to
respiration ceases in the dark This O2CO2 gas
exchange has been called right respiration or
photorespiration The substrate for the
photorespiratory metabolism is again ribulose
biphosphate which can be an acceptor not only for
CO2 but also for O2 By taking up oxygen RuBP is
split into PGA and phospoglycolate The supply of O2
and CO2 regulates the relationship between acceptor
oxidation (photorespiration) and acceptor
carboxylation (photosynthesis) via the enzyme
complex RuBP carboxylaseoxygenase High partial
pressure of O2 favors photorespiration A large
supply of CO2 favors photosynthesis The formation
of phospoglycolate is dependent on the supply of
RuBP via the Calvin cycle the photorespiratory O2
uptake and CO2 release increase with light
intensity
1
The process of glycolate metabolism has not
yet been understood in all details The glycolate is
transported out of the chloroplasts phospoglycolate
and phospate The glycolate is transported out of
the chloroplast into peroxisomes cell compartments
about the size of mitochondria which contain
glycolate oxidase catalase and transaminases In
the peroxysomes when O2 is taken up glycolate is
oxidized to glyoxylate and the peroxide thus
produced is detoxified by catalase Glycolate can
either be completely reduced via oxalate by further
O2 uptake or transformed to glycine by
transamination Glycine is transported from the
peroxisomes into the mitochondria where two
molecules of glycine are coupled to form one
molecule of serine with the release of CO2 Serine
is taken over by the amino acid metabolism or
2
converted to glycerate after deamination by
hydropyruvate This can be photophosporylated in the
chloroplasts and returned to the Calvin cycle or
used elsewhere (Larcher 1980 81-82)
B Purposes
This manuscript is intended to complete the
task from our lecturer Dr Azwir Anhar MSi and
to meet the needs of university students as the
guidersquos book in learning and practice activity for
the member of International Standard Teacher
Education (ISTE) program in Public University of
Padang
C Benefits
Benefits that we gained from this manuscript
are
1 As reference in learning process of plant
phisiology subject especially in plant
respiration title
2 As information about steps in plant
respiration
3
CHAPTER II
PLANT RESPIRATION
A Krebs Cycle
The Krebs cycle was named in honor of the
English biochemist Hans A Krebs who in 1937
proposed a cycle of reactions to explain how pyruvate
breakdown takes place in the breast muscle of
pigeons He called his proposed pathway the citric
acid cycle because citric acid is an important
intermediet Another common name for the same group
of reactions is the Tricarboxuylic Acid (TCA) cycle
a term used because citric and isocitric acids have
three carboxyl groups It was not until the early
1950s that mitochondria capable of carrying out this
cycle were isolated from plant cells
The initial step leading to the Krebs cycle
involves the oxidation and loss of CO2 from pyruvate
and the combination of the remaining 2-carbon acetate
unit with a sulfur-containing compound co-enzyme A
(CoA) forming acetyl CoA This and another
comparable role of CoA in the Krebs cycle are
important reasons why sulfur is an essential element
4
The reaction of pyruvate decarboxylation also
involves a phosporylated form of thiamine (vitamin
B1) as a prostethic group Participation of thiamine
in this section partially explains the essential
function of vitamin B1 in plants and animals Besides
the loss of CO2 two hydrogen atoms are removed from
pyruvic acid during the formation of Acetyl CoA The
enzyme catalyzing the complete reaction is called
pyruvic acid dehydrogenase but it is actually an
organized complex containing numerous copies of three
or four different enzymes The hydrogen atoms removed
are finally accepted by NAD+ yielding NADH
The Krebs cycle accomplishes removal of some of
the electrons from organic acid intermediates and
transfer of these electrons to NAD+ or FAD Notice
that none of dehydrogenase enzyme of the cycle uses
NADP+ as an electron acceptor In fact NADP+ is
usually undetectable in plant mitochondria a
situation opposite to that of chloroplasts where
NADP+ is abundant but where there is much less NAD+
Not only are NADH and FADH2 important products of the
Krebs cycle but one molecule of ATP is formed from
ADP and Pi during the conversion of succinyl coenzyme
A to succinic acid Two additional CO2 molecules are
released in these Krebs cycle reactions so there is
a net loss of both carbon atoms from the incoming
5
acetate of acetyl CoA The release of CO2 in the
Krebs cycle accounts for the product of CO2 in the
summary equation for resspiration but no O2 is
absorbed during any Kreb cycle reaction (Lakitan
2011)
During metabolism the synthesis and breakdown
of different organic compounds takes place through
various pathways like the breakdown and synthesis of
proteins carbohydrates fats and nucleic acids
These different pathways and intermediates are also
responsible for the production of energyKrebrsquos
cycle named after Hans Krebs who began working out
its details in 1930s is a series of reactions in
which the pyruvate from glycolysis is oxidized to Co2
under aerobic conditions Kreb cycle is also known as
citric acid cycle or Tri-carboxylic acid cycle (TCA)
( Galston 1968 76)
6
Steps involved in the process of Kreb cycle are
1 Fate of pyruvic acid
Pyruvic acid can form different compounds by
different pathways ie it can be converted into
lactic acid It can be converted into acetyl
coenzyme A as a result of oxidation
2 Fate of Acetyl CoA
It can either undergo condensation with itself or
its derivatives to form fatty acids having 14 ndash 20
carbons Acetyl CoA can also go through a series
of reactions in Krebs cycle
3 Formation of citrate
Acetyl CoA enzyme condenses with oxaloacetate by
an enzyme citrate synthatase to form citrate with
the release of acetyl CoA If the amount of
7
oxaloacetate is very small then small number of
acetyl CoA would be reacting with oxaloacetate
leaving surplus acetyl CoA to go through another
pathway for the formation of long chain fatty
acids
4 Formation of C is ndash acotinate and iso-citrate
Citrate is changed first into C is ndash acotinate and
then to iso-citrate under the enzyme acotinase
Equilibrium is established between citrate C is
aconitate and iso-citrate It has been observed
that most of the time this equilibrium is shifted
towards the iso-citrate If the concentration of
iso-citrate in increased the formation of citrate
will result which also indicates that the
equilibrium also shift in the reserve direction
5 Formation of oxalosucinate
Iso-citrate is acted upon by an enzyme iso-citrate
dehydrogenase using nicotinamide adenine
dinucleotide (NAD) as coenzyme As a result iso-
citrate is converted into oxalosucciate and NAD is
reduced to NADH2 Similar reaction is carried out
by same enzyme using nicotindmide adenine
dinucleotide phosphate (NADP) as coenzyme which is
reduced to NDAPH2
6 Formation of α-Ketoglutarate
8
Oxalosucciante is changed into α-ketoglutrate by
iso-citrate dehydrogenase with the help of
coenzyme NAD or NADP In this reaction carbon
dioxide and NADH2 or NADPH2 are also released
7 Formation of succinyl CoA
α-ketoglutarate combines with acetyl CoA in the
presence of coenzyme NAD and enzyme α-
keoglutaratedehyrogenase to form succinyl coenzyme
a carbon dioxide and NADH2
8 Formation of Succinate
Later on coenzyme a is removed from succinyl
coenzyme A in the presence of guanosinediphosphate
(GDP) and inorganic phosphate to form succinate
and guanosine triphosphate (GTP) This reaction is
carried out by an enzyme called succinyl CoA
synthatase
9 Formation of Funarate
An enzyme succinic dehydrogenase removes hydrogen
from succinate to form funarate
10 Formation of Malate
Fumarate reacts with water in the presence of
enzyme fumarase to form malate
11 Regeneration of oxaloacetate
Malate is oxidized by malic dehydrogenase and NAD
forming oxaloacetate and NADH2 Thus oxalo acetate
is again available to start another cycle
9
(httpkrebs-cycle-or-citric-acid-cycle-or-
trihtml)
Importance of CTA Cycle
1 Source of energy
In addition to routine organic compounds described
above by products like nicotinanide adenine
dinucleotide (NADH2) and guanosine triphosphate
(GTP) are the source of biological energy NADH2
after oxidation produce energy whereas GTP is
itself high energy phosphate compound
2 Oxidation of organic compounds taken as food
Oxidation of fats carbohydrates and proteins take
place through it or in other words it can be said
that oxidation of all compounds having carbon
atoms can take place through TCA cycle Some of
amino acids like alanine glutamic acid and
aspartic acid at one stage or the other enters
into TCA cycle eg glutamic acid enters cycle
after its transformation into α-ketoglutarate
Similarly alanine enters the cycle after its
conversion into pyruvate
3 Intermediate compounds
TCA cycle is also involved in synthesis of
intermediate compounds leading to the formation of
larger molecules (Ray 1963 17)
10
Verification of Krebs Cycle
It was done by radioactive traces like C14 as
radioactive carbon dioxide of different levels and
reactions After addition of radioactive carbon
dioxide different chemical compounds produced like
glucose fats amino acids were isolated and looked
for radioactive carbon In this way whole of
metabolic reaction were verified including individual
reactions alternative metabolic pathways
intermediates of fats carbohydrates and amino acids
etc in the body cells as well as in test tubes
(Wilson 1962 99)
The primary function of the Krebs cycle are as
follows
1 Reduction of NAD+ and FAD to the electron donors
NADH and FADH2 that are subsequently oxidized to
yield ATP
2 Direct synthesis of a limited amount of ATP (1 ATP
for each pyruvate oxidized)
3 Formation of carbon skeletons that can be used to
synthesize certain amino acids that in turn are
converted into larger molecules
11
B Fermentation
Although glycolysis can function well without
O2 the further oxidation of pyruvate and NADH by
mitochondria requires this gas Thus when O2 is
limiting NADH and pyruvate begin to accumulate
Under this condition plants carry out fermentation
(anaerobic respiration) forming either ethanol or
lactic acid (usually ethanol) As shown in the
picture below the two top reactions consists of a
decarboxylation to form acetaldehide then rapid
reduction of acetaldehide by NADH to form ethanol
These reactions are catalyzed by alcohol
dehydrogenase Some cells contain lactic acid
12
dehydrogenase which uses NADH to reduce pyruvic to
lactic acid Thus either ethanol or lactic acid or
both are fermentation products depending on the
activities of each dehydrogenase present In each
case NADH is the reductant and only under
anaerobic conditions is it abundant anough to cause
reduction Furthermore in some plants NADH is used
to cause accumulation of the other compounds when O2
is limiting especially malate and glycerol
(Salisbury 1969 233)
The fate of pyruvate produced by glycolysis
depends primarily on whether or not oxygen is
present Under normal aerobic conditions pyruvate
is transported into the mitochondrion where it is
further oxidized to CO2 and water transferring its
electrons ultimately to molecular oxygen
13
Although higher plants are obligate aerobs and
are able to tolerate anoxia for only short periods
tissues or organs are occasionally subjected to
anaerobic conditions A typical siuation is that of
roots when the soil is saturated with water When
there is no oxygen to serve as the terminal electron
acceptor mitochondrial respiration will shut down
and metabolism will shift over to fermentation
Fermentation converts pyruvate to either ethanol
through the action of the enzyme alcohol
dehydrogenase (ADH) or lactate via lactate
dehydrogenase (LDH) In most plants the principal
products of fermentation are CO2 and ethanol Some
lactate may be formed primarily in the early stages
of anoxia However lactate lowers he pH of the
cytosol which in turn activates pyruvate
decarboxylase and initiates the production of
ethanol
Either one of the fermentation reactions
consumes the NADH produced earlier in glycolysis by
the oxidation of glyceraldehide-3-P Although this
means there is no net gain of reducing potential in
fermentation This recycling of NADH is still
important to the cell The poolof NADH plus NAD+ in
the cell is relatively small and if the NADH is not
recycled there will be no supply of NAD+ to support
14
the continued oxidation of glyceraldehyde-3-P if
this were the case glycolysis and the producton of
even the small quantities of ATP necessary to
maintain the cells under anaerobic conditions would
then grind to a halt (Hopkins 2009 241)
Soil waterlogging has long been identified as a
major abiotic stress and the constraints that it
provides the roots have a significant effect on the
growth and development of plants When this event
occurs in the spring then these puddles can reduce
seed germination and seedling development Thus a
pool of water is an important factor that affects
the growth development and survival of plant
species not only in natural ecosystems but also
the agricultural and horticultural systems (Dat et
al 2006)
After flooding rapid changes in soil
properties At the time of pore water meets the
land the air pushed out gas diffusion is reduced
and toxic compounds accumulate due to anaerobic
conditions All of these changes greatly affect the
ability of plants to survive In response increased
stomatal resistance photosynthesis and root
hydraulic conductivity decreases and reduced
translocation fotoassimilat Nevertheless one of
15
the best adaptations of plants to hypoxia anoxia
is transitional biochemical and metabolic processes
common to current limited availability of O2 (Dat et
al 2004)
Selective synthesis of a set of about 20
anaerobic stress proteins (ANPS) allows the process
of producing energy without oxygen metabolism under
anaerobic conditions (Subbaiah and Sachs 2003)
Another adaptation is the observed morphological
changes consisting of the formation of hypertrophic
lenticels adventitious root initiation and or
progression of aerenchyma (Vartapetian and Jackson
1997 Jackson and Colmer 2005 Folzer et al 2006)
This review details the diverse plant stress
response to hypoxia anoxia caused by soil
waterlogging flooding and examines some of the key
features of metabolic adaptation physiological and
morphological
16
C Electron
Transport System
We noted earlier that one of the principal
functions of the respiration is to retrieve in
useful form some of the energy initially stored in
assimilates Our traditional measure of useful
energy in most processes is the number of ATP
molecules gained or consumed By this measure alone
the yield from both glycolysis and the citric acid
cycle is quite low After two complete turns of the
cycle one molecule of glucose has been completely
oxidized to six molecules of CO2 but only four
molecules of ATP have been produced (a net of two
ATP from glycolysis plus one for each turn of the
cycle) At this point most of the energy associated
with the glucose molecule has been conserved in the
17
form of electron pairs generated by the oxidation of
glycolytic and citric acid cycle intermediates
In this section we will discuss the third stage
of cellular respirationmdashthe transfer of electrons
fromNADH and FADH2 to oxygen and the accompanying
conversion of redox energy to ATP The transfer of
electrons fromNADHand FADH2 to oxygen involves a
sequence of electron carriers arranged in an
electron transport chainMembrane fractionation
studies have shown that the enzymes and electron
carriers making up the electron transport chain are
organized predominantly into four large
multimolecular complexes (complexes ImdashIV) and two
mobile carriers located in the inner mitochondrial
membrane (Figure 109) In this sense there are a
great number of similarities between the
mitochondrial inner membrane and the thylakoid
membranes of the chloroplast (compare Figure 109
with Figure 76) This is not unexpected since the
principal function of each membrane is energy
transformation and many of the same or similar
components are involved
The Components of the Electron Transport Chain
The electron transport chain of the
mitochondria is the means by which electrons are
18
removed from the reduced carrier NADH and
transferred to oxygen to yield H2O
1) NADH
NADH is generated in the matrix by the reactions
of pyruvate dehydrogenase isocitrate
dehydrogenase α-ketoglutarate dehydrogenase and
malate dehyrogenase The electron transport chain
begins with reoxidizing NADH to form NAD+ and
channeling the electrons into the formation of
reduced coenzymes Important to note that NADH
transfers 2 electrons at a time in the form of a
hydride
NAD+ + 2e- + H+ 1048581 NADH Eorsquo = minus0315 V
2) Flavoproteins
Flavoproteins have either a FAD (flavin adenosine
dinucleotide) or a FMN (flavin mononucleotide)
prosthetic group Flavoproteins can accept or
donate electrons one at time or two at a time
Thus they are often intermediaries between two
electron acceptorsdonors and one electron
acceptorsdonors For flavoproteins the typical
standard reduction potentials are around 0 V
FAD + 2e- + 2H+ 1048581 FADH2 FMN + 2e- + 2H+ 1048581 FMNH2
Eorsquo asymp 0 V
19
3) Coenzyme Q (CoQ)
aka ubiquinone (UQ) Shown below CoQ has ten
repeating isoprene units which make it
insoluble in water but soluble in the
hydrophobic lipid bylayer Coenzyme Q is a
versatile cofactor because it is a soluble
electron carrier in the hydrophobic bilipid layer
of the
inner mitochondrial membrane Like flavoproteins
CoQ can acceptdonate electrons one at a time or
two at a time
Q + 2e- + 2H+ 1048581 QH2 Eorsquo =0060 V
Q + e- + H+ 1048581 1048581QH Eorsquo =0030 V
1048581QH + e- + H+ 1048581 QH2 Eorsquo =0190 V
4) Cytochromes
Cytochromes are proteins that contain heme
prosthetic groups which function as
one electron carriers The heme iron is involved
in one electron transfers involving the Fe2+ and
Fe3+ oxidation states Cytochromes are named by
their absorption spectra which depends
on the porphyrin structure and environment The
example shown is the heme prosthetic group of
cytochrome b Cytochrome b contains the same iron
porphyrin found in hemoglobin and myoglobin
20
Other cytochromes we will encounter in the
electron transport complexes are cytochromes b
c c1 a and a3
5) Iron-Sulfur Proteins
In the electron transport chain we will encounter
many iron-sulfer proteins which participate in
one electron transfers involving the the Fe2+ and
Fe3+ oxidation states These are non-heme iron-
sulfur proteinsThe simplest iron-sulfer protein
is FeS in which iron is tetrahedrally coordinated
by four cysteines The second form is Fe2S2 which
contains two irons complexed to 2 cysteine
residues and two inorganic sulfides The third
form is Fe3S4 which contains 3 iron atoms
coordinated to three cysteine residues and 4
inorganic sulfides The last form is the most
complicated Fe4S4 which contains 4 iron atoms
coordinated to 4 cysteine residues and 4
inorganic sulfides
6) Copper Proteins
Copper bound proteins participate in one electron
transfers involving the Cu+ and Cu2+ oxidation
states
21
Overview of the Electron Transport Chain
Electrons move along the electron transport
chain going from donor to acceptor until they reach
oxygen the ultimate electron acceptor The standard
reduction potentials of the electron carriers are
between the NADHNAD+ couple (-0315 V) and the
oxygenH2O couple (0816 V) as on the next page The
components of the electron transport chain are
organized into 4 complexes Each complex contains
several different electron carriers
1 Complex I also known as the NADH-coenzyme Q
reductase or NADH dehydrogenase
2 Complex II also known as succinate-coenzyme Q
reductase or succinate dehydrogenase
3 Complex III also known as coenzyme Q reductase
4 Complex IV also known as cytochrome c reductase
The free energy necessary to generate ATP
is extracted from the oxidation of NADH and
FADH2 by the electron transport chain It
consists of four protein complexes and single
protein chain of cytochrome c through which
electrons pass from lower to higher standard
redox potentials Electrons are carried from complex
I and II to complex III by the membrane (lipid)
soluble coenzyme Q (CoQ or ubiquinone) and between
22
complexes III and IV by the peripheral membrane
protein cytochrome c
In the following I give the overall
reactions catalysed by corresponding protein
complexes in the mitochondrial electron transport
chain
1 Complex I
Complex I is also called NADH-Coenzyme Q
reductase because this large protein complex
transfers 2 electrons from NADH to coenzyme Q
Complex I was formerly known as NADH dehydrogenase
Complex I is huge 850000 kD and is composed of more
than thirty subunits It contains a FMN prosthetic
group and seven or more Fe-S clusters This complex
has between 20-26 iron atoms boundThe prosthetic
group FMN is absolutely required for activity
Therefore this complex is a flavoprotein This
complex binds NADH transfers two electrons in the
form of a hydride to FMN to produce NAD+ and FMNH2
23
The subsequent steps involve the transfer of
electrons one at a time to a series of iron-sulfer
complexes that includes both 2Fe-2S and 4Fe-4S
clusters Note the importance of FMN First it
functions as a 2 electron acceptor in the hydride
transfer from NADH Second it functions as a 1
electron donor to the series of iron sulfur clusters
FMN and FAD often play crucial links between 2
electron transfer agents and 1 electron transfer
agents The final step of this complex is the
transfer of 2 electrons one at a time to coenzyme Q
CoQ like FMN and FAD can function as a 2 electron
donoracceptor and as a 1 electron donoracceptor
CoQ is a mobile electron carrier because its
isoprenoid tail makes it highly hydrophobic and
lipophillic It diffuses freely in the bilipid layer
of the inner mitochondrial membrane
The process of transferring electrons from NADH
to CoQ by complex I results in the net transport of
protons from the matrix side of the inner
mitochondrial membrane to the inter membrane space
where the H+ ions accumulate generating a proton
motive force The intermembrane space side of the
inner membrane is referred to as the P face (P
standing for positive) The matrix side of the inner
membrane is referred to as the the N face The
24
transport of electrons from NADH to CoQ is coupled to
the transport of protons across the membrane This is
an example of active transport The stiochiometry is
4 H+ transported per 2 electrons
2 Complex II
Believe it or not you are already familiar
with Complex II It is none other than succinate
dehydrogenase The only enzyme of the citric acid
cycle that is an integral membrane protein This
complex is composed of four subunits 2 of which are
iron-sulfur proteins and the other two subunits
together bind FAD through a covalent link to a
histidine residue These two subunits are called
flavoprotein 2 or FP2 Complex II contains 3 Fe-S
centers 1 4Fe-4S cluster 1 3Fe-4S cluster and 1
2Fe- 2S cluster In the first step of this complex
succinate is bound and a hydride is transferred to
FAD to generate FADH2 and fumarate FADH2 then
transfers its electrons one at a time to the Fe-S
centers Thus once again FAD functions as 2 electron
acceptor and a 1 electron donor The final step of
this complex is the transfer of 2 electrons one at a
time to coenzyme Q to produce CoQH2 For complex II
the standard free energy change of the overall
reaction is too small to drive the transport of
protons across the inner mitochondrial membrane This
25
accounts for the 15 ATPrsquos generated per FADH2
compared with the 25 ATPrsquos generated per NADH
3 Complex III
This complex is also known as coenzyme Q-
cytochrome c reductase because it passes the
electrons form CoQH2 to cyt c through a very unique
electron transport pathway called the Q-cycle Shown
to the left are the porphyrins found in cytochromes
Cytochrome b contains the same iron protoporphyrin as
hemoglobin and myoglobin The c cytochromes contain
heme c through covalent attachment by cysteine
residues Cytochrome a is found in two forms in
complex IV In complex III we find two b-type
cytochromes and one c-type cytochrome Complex III is
complex and we have a crystal structure
4 Complex IV
Complex IV is also known as cytochrome c oxidase
because it accepts the electrons from cytochrome c
and directs them towards the four electron reduction
of O2 to form 2 molecules of H2O
4 cyt c (Fe2+) + 4 H+ + O2 4 cyt c (Fe3+) +
2H2O
26
Cytochrome c oxidase contains 2 heme
centerscytochrome a and cytochrome a3 and two copper
proteins Each of the protein bound coppers are
associated with one of the cytochromes The copper
sites are called CuA and CuB CuA is associated with
cytochrome a and is shown to the left CuB is
associated with cytochrome a3 The copper sites
function as 1 electron carriers cycling between the
cuprous state Cu+ and the cupric state Cu2+ Just
like iron containing proteins they transfer
electrons one at a time Cytochrome c is bound on the
P-face of the membrane and transfers its electron to
CuA The oxidized cytochrome c dissociates CuA then
transfers the electron to cytochrome a The protein
bound CuA and the iron bound in cytochrome a are 15 Aring
apart In contrast the CuB and the iron bound in
cytochrome a3 are very close to each other forming a
binuclear metal center shown below
27
Electron transport chains are associated with
membranes such as the mitochondrial membrane in
eukaryotic organisms the thylakoid membrane of
photosynthetic organisms and the plasma membrane of
prokaryotic organisms While some of the details
differ electron transport chains in all organisms
operate on the same basic principles
Excited electrons are brought to the electron
transport chain by electron carriers such as NADH
and FADH2 In photosynthetic organisms the electrons
are donated by water and excited by light energy
The electron donor is oxidized as the electrons are
passed to the first protein complex in the chain As
it accepts the electrons the protein complex is
reduced Next the electrons are passed from the
28
first protein complex which becomes oxidized to
the second protein complex which becomes reduced
Each of these redox reactions is slightly exergonic
or energy releasing Some of this energy is used to
move protons across the membrane against their
concentration gradient With each successive
transfer of electrons the original excited
electrons lose some of their energy After several
successive redox reactions the electrons are
donated to a terminal electron acceptor In aerobic
respiration the terminal electron acceptor is
oxygen In photosynthesis the terminal electron
acceptor is a compound called NADP
The energy in the excited electrons has been
transformed into an electrochemical gradient
represented by the high concentration of protons on
one side of the membrane These protons can diffuse
back across the membrane by passing through a
protein complex called ATP synthase As these
protons pass through ATP synthase down their
concentration gradient they provide the energy
needed to make ATP from ADP and inorganic phosphate
within the cell
29
CHAPTER III
CLOSING
A Conclution
When a hexose is completely oxidized to CO2
and H2O using these three processes Glycolysis
yields two ATP and two NADH per hexose used Each
such NADH oxidized by the electron transport system
30
yields two ATP as dscribed above so glycolysis
contributes a total of six ATP per hexose The Krebs
cycle contributes two ATP per hexose or per two
pyruvates when succinyl CoA is cleaved to succinate
and CoASH This cycle also produces eight NADH per
hexose within the mitochondrial matrix by oxidative
phosporylation each of these NADH yields three ATP
or 24 per hexose Each FADH2 from the Krebs cycle
yields two ATP by oxydative phosporylation or four
per hexose (two pyruvates) The total contribution of
the Krebs cycle is then 30 Adding these 30 to the 6
from glycolysis leads to a total of 36 per hexose
completely respired by these process
B Suggestion
My greatest wish is that the readers find
this manuscript helpful in gaining the knowledge they
need for effective learning and practice and in
preparing for examination designed to test that
knowledge Reader comments and suggestions are
welcome and would be very helpful in improving our
manuscript Your contribution may assist many other
students in the coming years
31
REFERENCE
Anonim 2013 Anaerobic Respiration
httpwwwscoolcoukgcsebiologyrespiration
revise-itanaerobic-respiration Accessed
September 10th 2013
Anonim 2013 What is the difference between aerobic
and anaerobic respiration in plants
httpwwwbiologylifeeasyorg4010difference
-between-aerobic-anaerobic-respiration-plants
Accessed September 10th 2013
Campbell Mary K 2006 Biochemistry New York Thomson
Brooks Cole Inc
Dat J Capelli N Folzer H Bourgeade P Badot P-M
(2004) Sensing and signaling during
plantflooding Plant Physiology and
Biochemistry 42 273-282
Dat J Folzer H Parent C Badot P-M Capelli N
(2006) HypoxiastressCurrent Understanding and
Perspectives In Teixeira da Silva JA (Ed)
Floriculture Ornamental and Plant
Biotechnology Advances and Topical Issues (Vol
3) Global Science Books Isleworth United
Kingdompp 664-674
Hatta Muhammad 2011 Dunia Pertanian
httpemhattawordpresscomcategorystress
Accessed September 10th 2013
Hopkins William G 2009 Introduction to Plant Physiology
New York John Wiley amp Sons Inc
Lakitan Benyamin 2011 Dasar-Dasar Fisiologi Tumbuhan
Jakarta PT Raja Grafindo Persada
Larcher W 1980 Physiological Plant Ecology New York
Springer-Verlag
Raven 2013 Biology Fifth Edition
httphigheredmcgraw-hillcomsites983409233
9student_view0chapter39
photosynthetic_electron_transport_and_atp_synth
esishtml Accessed September 10th 2013
Salisbury Frank B 1969 Plant Physiology Third Edition
California Wadsworth Publishing Company
Subbaiah C Sachs M (2003) Molecular and cellular
adaptations of maize to flooding stress Annals
of Botany 91 119-127
Wilson CarlL1962 Botany Third EditionUnited State of
America Holt Rinch Inc

CHAPTER I
INTRODUCTION
A Background
Connected with photosynthesis a metabolic
process takes place in chloroplast-containing plant
cells which like respiration takes up O2 and
releases CO2 in the light but which contrary to
respiration ceases in the dark This O2CO2 gas
exchange has been called right respiration or
photorespiration The substrate for the
photorespiratory metabolism is again ribulose
biphosphate which can be an acceptor not only for
CO2 but also for O2 By taking up oxygen RuBP is
split into PGA and phospoglycolate The supply of O2
and CO2 regulates the relationship between acceptor
oxidation (photorespiration) and acceptor
carboxylation (photosynthesis) via the enzyme
complex RuBP carboxylaseoxygenase High partial
pressure of O2 favors photorespiration A large
supply of CO2 favors photosynthesis The formation
of phospoglycolate is dependent on the supply of
RuBP via the Calvin cycle the photorespiratory O2
uptake and CO2 release increase with light
intensity
1
The process of glycolate metabolism has not
yet been understood in all details The glycolate is
transported out of the chloroplasts phospoglycolate
and phospate The glycolate is transported out of
the chloroplast into peroxisomes cell compartments
about the size of mitochondria which contain
glycolate oxidase catalase and transaminases In
the peroxysomes when O2 is taken up glycolate is
oxidized to glyoxylate and the peroxide thus
produced is detoxified by catalase Glycolate can
either be completely reduced via oxalate by further
O2 uptake or transformed to glycine by
transamination Glycine is transported from the
peroxisomes into the mitochondria where two
molecules of glycine are coupled to form one
molecule of serine with the release of CO2 Serine
is taken over by the amino acid metabolism or
2
converted to glycerate after deamination by
hydropyruvate This can be photophosporylated in the
chloroplasts and returned to the Calvin cycle or
used elsewhere (Larcher 1980 81-82)
B Purposes
This manuscript is intended to complete the
task from our lecturer Dr Azwir Anhar MSi and
to meet the needs of university students as the
guidersquos book in learning and practice activity for
the member of International Standard Teacher
Education (ISTE) program in Public University of
Padang
C Benefits
Benefits that we gained from this manuscript
are
1 As reference in learning process of plant
phisiology subject especially in plant
respiration title
2 As information about steps in plant
respiration
3
CHAPTER II
PLANT RESPIRATION
A Krebs Cycle
The Krebs cycle was named in honor of the
English biochemist Hans A Krebs who in 1937
proposed a cycle of reactions to explain how pyruvate
breakdown takes place in the breast muscle of
pigeons He called his proposed pathway the citric
acid cycle because citric acid is an important
intermediet Another common name for the same group
of reactions is the Tricarboxuylic Acid (TCA) cycle
a term used because citric and isocitric acids have
three carboxyl groups It was not until the early
1950s that mitochondria capable of carrying out this
cycle were isolated from plant cells
The initial step leading to the Krebs cycle
involves the oxidation and loss of CO2 from pyruvate
and the combination of the remaining 2-carbon acetate
unit with a sulfur-containing compound co-enzyme A
(CoA) forming acetyl CoA This and another
comparable role of CoA in the Krebs cycle are
important reasons why sulfur is an essential element
4
The reaction of pyruvate decarboxylation also
involves a phosporylated form of thiamine (vitamin
B1) as a prostethic group Participation of thiamine
in this section partially explains the essential
function of vitamin B1 in plants and animals Besides
the loss of CO2 two hydrogen atoms are removed from
pyruvic acid during the formation of Acetyl CoA The
enzyme catalyzing the complete reaction is called
pyruvic acid dehydrogenase but it is actually an
organized complex containing numerous copies of three
or four different enzymes The hydrogen atoms removed
are finally accepted by NAD+ yielding NADH
The Krebs cycle accomplishes removal of some of
the electrons from organic acid intermediates and
transfer of these electrons to NAD+ or FAD Notice
that none of dehydrogenase enzyme of the cycle uses
NADP+ as an electron acceptor In fact NADP+ is
usually undetectable in plant mitochondria a
situation opposite to that of chloroplasts where
NADP+ is abundant but where there is much less NAD+
Not only are NADH and FADH2 important products of the
Krebs cycle but one molecule of ATP is formed from
ADP and Pi during the conversion of succinyl coenzyme
A to succinic acid Two additional CO2 molecules are
released in these Krebs cycle reactions so there is
a net loss of both carbon atoms from the incoming
5
acetate of acetyl CoA The release of CO2 in the
Krebs cycle accounts for the product of CO2 in the
summary equation for resspiration but no O2 is
absorbed during any Kreb cycle reaction (Lakitan
2011)
During metabolism the synthesis and breakdown
of different organic compounds takes place through
various pathways like the breakdown and synthesis of
proteins carbohydrates fats and nucleic acids
These different pathways and intermediates are also
responsible for the production of energyKrebrsquos
cycle named after Hans Krebs who began working out
its details in 1930s is a series of reactions in
which the pyruvate from glycolysis is oxidized to Co2
under aerobic conditions Kreb cycle is also known as
citric acid cycle or Tri-carboxylic acid cycle (TCA)
( Galston 1968 76)
6
Steps involved in the process of Kreb cycle are
1 Fate of pyruvic acid
Pyruvic acid can form different compounds by
different pathways ie it can be converted into
lactic acid It can be converted into acetyl
coenzyme A as a result of oxidation
2 Fate of Acetyl CoA
It can either undergo condensation with itself or
its derivatives to form fatty acids having 14 ndash 20
carbons Acetyl CoA can also go through a series
of reactions in Krebs cycle
3 Formation of citrate
Acetyl CoA enzyme condenses with oxaloacetate by
an enzyme citrate synthatase to form citrate with
the release of acetyl CoA If the amount of
7
oxaloacetate is very small then small number of
acetyl CoA would be reacting with oxaloacetate
leaving surplus acetyl CoA to go through another
pathway for the formation of long chain fatty
acids
4 Formation of C is ndash acotinate and iso-citrate
Citrate is changed first into C is ndash acotinate and
then to iso-citrate under the enzyme acotinase
Equilibrium is established between citrate C is
aconitate and iso-citrate It has been observed
that most of the time this equilibrium is shifted
towards the iso-citrate If the concentration of
iso-citrate in increased the formation of citrate
will result which also indicates that the
equilibrium also shift in the reserve direction
5 Formation of oxalosucinate
Iso-citrate is acted upon by an enzyme iso-citrate
dehydrogenase using nicotinamide adenine
dinucleotide (NAD) as coenzyme As a result iso-
citrate is converted into oxalosucciate and NAD is
reduced to NADH2 Similar reaction is carried out
by same enzyme using nicotindmide adenine
dinucleotide phosphate (NADP) as coenzyme which is
reduced to NDAPH2
6 Formation of α-Ketoglutarate
8
Oxalosucciante is changed into α-ketoglutrate by
iso-citrate dehydrogenase with the help of
coenzyme NAD or NADP In this reaction carbon
dioxide and NADH2 or NADPH2 are also released
7 Formation of succinyl CoA
α-ketoglutarate combines with acetyl CoA in the
presence of coenzyme NAD and enzyme α-
keoglutaratedehyrogenase to form succinyl coenzyme
a carbon dioxide and NADH2
8 Formation of Succinate
Later on coenzyme a is removed from succinyl
coenzyme A in the presence of guanosinediphosphate
(GDP) and inorganic phosphate to form succinate
and guanosine triphosphate (GTP) This reaction is
carried out by an enzyme called succinyl CoA
synthatase
9 Formation of Funarate
An enzyme succinic dehydrogenase removes hydrogen
from succinate to form funarate
10 Formation of Malate
Fumarate reacts with water in the presence of
enzyme fumarase to form malate
11 Regeneration of oxaloacetate
Malate is oxidized by malic dehydrogenase and NAD
forming oxaloacetate and NADH2 Thus oxalo acetate
is again available to start another cycle
9
(httpkrebs-cycle-or-citric-acid-cycle-or-
trihtml)
Importance of CTA Cycle
1 Source of energy
In addition to routine organic compounds described
above by products like nicotinanide adenine
dinucleotide (NADH2) and guanosine triphosphate
(GTP) are the source of biological energy NADH2
after oxidation produce energy whereas GTP is
itself high energy phosphate compound
2 Oxidation of organic compounds taken as food
Oxidation of fats carbohydrates and proteins take
place through it or in other words it can be said
that oxidation of all compounds having carbon
atoms can take place through TCA cycle Some of
amino acids like alanine glutamic acid and
aspartic acid at one stage or the other enters
into TCA cycle eg glutamic acid enters cycle
after its transformation into α-ketoglutarate
Similarly alanine enters the cycle after its
conversion into pyruvate
3 Intermediate compounds
TCA cycle is also involved in synthesis of
intermediate compounds leading to the formation of
larger molecules (Ray 1963 17)
10
Verification of Krebs Cycle
It was done by radioactive traces like C14 as
radioactive carbon dioxide of different levels and
reactions After addition of radioactive carbon
dioxide different chemical compounds produced like
glucose fats amino acids were isolated and looked
for radioactive carbon In this way whole of
metabolic reaction were verified including individual
reactions alternative metabolic pathways
intermediates of fats carbohydrates and amino acids
etc in the body cells as well as in test tubes
(Wilson 1962 99)
The primary function of the Krebs cycle are as
follows
1 Reduction of NAD+ and FAD to the electron donors
NADH and FADH2 that are subsequently oxidized to
yield ATP
2 Direct synthesis of a limited amount of ATP (1 ATP
for each pyruvate oxidized)
3 Formation of carbon skeletons that can be used to
synthesize certain amino acids that in turn are
converted into larger molecules
11
B Fermentation
Although glycolysis can function well without
O2 the further oxidation of pyruvate and NADH by
mitochondria requires this gas Thus when O2 is
limiting NADH and pyruvate begin to accumulate
Under this condition plants carry out fermentation
(anaerobic respiration) forming either ethanol or
lactic acid (usually ethanol) As shown in the
picture below the two top reactions consists of a
decarboxylation to form acetaldehide then rapid
reduction of acetaldehide by NADH to form ethanol
These reactions are catalyzed by alcohol
dehydrogenase Some cells contain lactic acid
12
dehydrogenase which uses NADH to reduce pyruvic to
lactic acid Thus either ethanol or lactic acid or
both are fermentation products depending on the
activities of each dehydrogenase present In each
case NADH is the reductant and only under
anaerobic conditions is it abundant anough to cause
reduction Furthermore in some plants NADH is used
to cause accumulation of the other compounds when O2
is limiting especially malate and glycerol
(Salisbury 1969 233)
The fate of pyruvate produced by glycolysis
depends primarily on whether or not oxygen is
present Under normal aerobic conditions pyruvate
is transported into the mitochondrion where it is
further oxidized to CO2 and water transferring its
electrons ultimately to molecular oxygen
13
Although higher plants are obligate aerobs and
are able to tolerate anoxia for only short periods
tissues or organs are occasionally subjected to
anaerobic conditions A typical siuation is that of
roots when the soil is saturated with water When
there is no oxygen to serve as the terminal electron
acceptor mitochondrial respiration will shut down
and metabolism will shift over to fermentation
Fermentation converts pyruvate to either ethanol
through the action of the enzyme alcohol
dehydrogenase (ADH) or lactate via lactate
dehydrogenase (LDH) In most plants the principal
products of fermentation are CO2 and ethanol Some
lactate may be formed primarily in the early stages
of anoxia However lactate lowers he pH of the
cytosol which in turn activates pyruvate
decarboxylase and initiates the production of
ethanol
Either one of the fermentation reactions
consumes the NADH produced earlier in glycolysis by
the oxidation of glyceraldehide-3-P Although this
means there is no net gain of reducing potential in
fermentation This recycling of NADH is still
important to the cell The poolof NADH plus NAD+ in
the cell is relatively small and if the NADH is not
recycled there will be no supply of NAD+ to support
14
the continued oxidation of glyceraldehyde-3-P if
this were the case glycolysis and the producton of
even the small quantities of ATP necessary to
maintain the cells under anaerobic conditions would
then grind to a halt (Hopkins 2009 241)
Soil waterlogging has long been identified as a
major abiotic stress and the constraints that it
provides the roots have a significant effect on the
growth and development of plants When this event
occurs in the spring then these puddles can reduce
seed germination and seedling development Thus a
pool of water is an important factor that affects
the growth development and survival of plant
species not only in natural ecosystems but also
the agricultural and horticultural systems (Dat et
al 2006)
After flooding rapid changes in soil
properties At the time of pore water meets the
land the air pushed out gas diffusion is reduced
and toxic compounds accumulate due to anaerobic
conditions All of these changes greatly affect the
ability of plants to survive In response increased
stomatal resistance photosynthesis and root
hydraulic conductivity decreases and reduced
translocation fotoassimilat Nevertheless one of
15
the best adaptations of plants to hypoxia anoxia
is transitional biochemical and metabolic processes
common to current limited availability of O2 (Dat et
al 2004)
Selective synthesis of a set of about 20
anaerobic stress proteins (ANPS) allows the process
of producing energy without oxygen metabolism under
anaerobic conditions (Subbaiah and Sachs 2003)
Another adaptation is the observed morphological
changes consisting of the formation of hypertrophic
lenticels adventitious root initiation and or
progression of aerenchyma (Vartapetian and Jackson
1997 Jackson and Colmer 2005 Folzer et al 2006)
This review details the diverse plant stress
response to hypoxia anoxia caused by soil
waterlogging flooding and examines some of the key
features of metabolic adaptation physiological and
morphological
16
C Electron
Transport System
We noted earlier that one of the principal
functions of the respiration is to retrieve in
useful form some of the energy initially stored in
assimilates Our traditional measure of useful
energy in most processes is the number of ATP
molecules gained or consumed By this measure alone
the yield from both glycolysis and the citric acid
cycle is quite low After two complete turns of the
cycle one molecule of glucose has been completely
oxidized to six molecules of CO2 but only four
molecules of ATP have been produced (a net of two
ATP from glycolysis plus one for each turn of the
cycle) At this point most of the energy associated
with the glucose molecule has been conserved in the
17
form of electron pairs generated by the oxidation of
glycolytic and citric acid cycle intermediates
In this section we will discuss the third stage
of cellular respirationmdashthe transfer of electrons
fromNADH and FADH2 to oxygen and the accompanying
conversion of redox energy to ATP The transfer of
electrons fromNADHand FADH2 to oxygen involves a
sequence of electron carriers arranged in an
electron transport chainMembrane fractionation
studies have shown that the enzymes and electron
carriers making up the electron transport chain are
organized predominantly into four large
multimolecular complexes (complexes ImdashIV) and two
mobile carriers located in the inner mitochondrial
membrane (Figure 109) In this sense there are a
great number of similarities between the
mitochondrial inner membrane and the thylakoid
membranes of the chloroplast (compare Figure 109
with Figure 76) This is not unexpected since the
principal function of each membrane is energy
transformation and many of the same or similar
components are involved
The Components of the Electron Transport Chain
The electron transport chain of the
mitochondria is the means by which electrons are
18
removed from the reduced carrier NADH and
transferred to oxygen to yield H2O
1) NADH
NADH is generated in the matrix by the reactions
of pyruvate dehydrogenase isocitrate
dehydrogenase α-ketoglutarate dehydrogenase and
malate dehyrogenase The electron transport chain
begins with reoxidizing NADH to form NAD+ and
channeling the electrons into the formation of
reduced coenzymes Important to note that NADH
transfers 2 electrons at a time in the form of a
hydride
NAD+ + 2e- + H+ 1048581 NADH Eorsquo = minus0315 V
2) Flavoproteins
Flavoproteins have either a FAD (flavin adenosine
dinucleotide) or a FMN (flavin mononucleotide)
prosthetic group Flavoproteins can accept or
donate electrons one at time or two at a time
Thus they are often intermediaries between two
electron acceptorsdonors and one electron
acceptorsdonors For flavoproteins the typical
standard reduction potentials are around 0 V
FAD + 2e- + 2H+ 1048581 FADH2 FMN + 2e- + 2H+ 1048581 FMNH2
Eorsquo asymp 0 V
19
3) Coenzyme Q (CoQ)
aka ubiquinone (UQ) Shown below CoQ has ten
repeating isoprene units which make it
insoluble in water but soluble in the
hydrophobic lipid bylayer Coenzyme Q is a
versatile cofactor because it is a soluble
electron carrier in the hydrophobic bilipid layer
of the
inner mitochondrial membrane Like flavoproteins
CoQ can acceptdonate electrons one at a time or
two at a time
Q + 2e- + 2H+ 1048581 QH2 Eorsquo =0060 V
Q + e- + H+ 1048581 1048581QH Eorsquo =0030 V
1048581QH + e- + H+ 1048581 QH2 Eorsquo =0190 V
4) Cytochromes
Cytochromes are proteins that contain heme
prosthetic groups which function as
one electron carriers The heme iron is involved
in one electron transfers involving the Fe2+ and
Fe3+ oxidation states Cytochromes are named by
their absorption spectra which depends
on the porphyrin structure and environment The
example shown is the heme prosthetic group of
cytochrome b Cytochrome b contains the same iron
porphyrin found in hemoglobin and myoglobin
20
Other cytochromes we will encounter in the
electron transport complexes are cytochromes b
c c1 a and a3
5) Iron-Sulfur Proteins
In the electron transport chain we will encounter
many iron-sulfer proteins which participate in
one electron transfers involving the the Fe2+ and
Fe3+ oxidation states These are non-heme iron-
sulfur proteinsThe simplest iron-sulfer protein
is FeS in which iron is tetrahedrally coordinated
by four cysteines The second form is Fe2S2 which
contains two irons complexed to 2 cysteine
residues and two inorganic sulfides The third
form is Fe3S4 which contains 3 iron atoms
coordinated to three cysteine residues and 4
inorganic sulfides The last form is the most
complicated Fe4S4 which contains 4 iron atoms
coordinated to 4 cysteine residues and 4
inorganic sulfides
6) Copper Proteins
Copper bound proteins participate in one electron
transfers involving the Cu+ and Cu2+ oxidation
states
21
Overview of the Electron Transport Chain
Electrons move along the electron transport
chain going from donor to acceptor until they reach
oxygen the ultimate electron acceptor The standard
reduction potentials of the electron carriers are
between the NADHNAD+ couple (-0315 V) and the
oxygenH2O couple (0816 V) as on the next page The
components of the electron transport chain are
organized into 4 complexes Each complex contains
several different electron carriers
1 Complex I also known as the NADH-coenzyme Q
reductase or NADH dehydrogenase
2 Complex II also known as succinate-coenzyme Q
reductase or succinate dehydrogenase
3 Complex III also known as coenzyme Q reductase
4 Complex IV also known as cytochrome c reductase
The free energy necessary to generate ATP
is extracted from the oxidation of NADH and
FADH2 by the electron transport chain It
consists of four protein complexes and single
protein chain of cytochrome c through which
electrons pass from lower to higher standard
redox potentials Electrons are carried from complex
I and II to complex III by the membrane (lipid)
soluble coenzyme Q (CoQ or ubiquinone) and between
22
complexes III and IV by the peripheral membrane
protein cytochrome c
In the following I give the overall
reactions catalysed by corresponding protein
complexes in the mitochondrial electron transport
chain
1 Complex I
Complex I is also called NADH-Coenzyme Q
reductase because this large protein complex
transfers 2 electrons from NADH to coenzyme Q
Complex I was formerly known as NADH dehydrogenase
Complex I is huge 850000 kD and is composed of more
than thirty subunits It contains a FMN prosthetic
group and seven or more Fe-S clusters This complex
has between 20-26 iron atoms boundThe prosthetic
group FMN is absolutely required for activity
Therefore this complex is a flavoprotein This
complex binds NADH transfers two electrons in the
form of a hydride to FMN to produce NAD+ and FMNH2
23
The subsequent steps involve the transfer of
electrons one at a time to a series of iron-sulfer
complexes that includes both 2Fe-2S and 4Fe-4S
clusters Note the importance of FMN First it
functions as a 2 electron acceptor in the hydride
transfer from NADH Second it functions as a 1
electron donor to the series of iron sulfur clusters
FMN and FAD often play crucial links between 2
electron transfer agents and 1 electron transfer
agents The final step of this complex is the
transfer of 2 electrons one at a time to coenzyme Q
CoQ like FMN and FAD can function as a 2 electron
donoracceptor and as a 1 electron donoracceptor
CoQ is a mobile electron carrier because its
isoprenoid tail makes it highly hydrophobic and
lipophillic It diffuses freely in the bilipid layer
of the inner mitochondrial membrane
The process of transferring electrons from NADH
to CoQ by complex I results in the net transport of
protons from the matrix side of the inner
mitochondrial membrane to the inter membrane space
where the H+ ions accumulate generating a proton
motive force The intermembrane space side of the
inner membrane is referred to as the P face (P
standing for positive) The matrix side of the inner
membrane is referred to as the the N face The
24
transport of electrons from NADH to CoQ is coupled to
the transport of protons across the membrane This is
an example of active transport The stiochiometry is
4 H+ transported per 2 electrons
2 Complex II
Believe it or not you are already familiar
with Complex II It is none other than succinate
dehydrogenase The only enzyme of the citric acid
cycle that is an integral membrane protein This
complex is composed of four subunits 2 of which are
iron-sulfur proteins and the other two subunits
together bind FAD through a covalent link to a
histidine residue These two subunits are called
flavoprotein 2 or FP2 Complex II contains 3 Fe-S
centers 1 4Fe-4S cluster 1 3Fe-4S cluster and 1
2Fe- 2S cluster In the first step of this complex
succinate is bound and a hydride is transferred to
FAD to generate FADH2 and fumarate FADH2 then
transfers its electrons one at a time to the Fe-S
centers Thus once again FAD functions as 2 electron
acceptor and a 1 electron donor The final step of
this complex is the transfer of 2 electrons one at a
time to coenzyme Q to produce CoQH2 For complex II
the standard free energy change of the overall
reaction is too small to drive the transport of
protons across the inner mitochondrial membrane This
25
accounts for the 15 ATPrsquos generated per FADH2
compared with the 25 ATPrsquos generated per NADH
3 Complex III
This complex is also known as coenzyme Q-
cytochrome c reductase because it passes the
electrons form CoQH2 to cyt c through a very unique
electron transport pathway called the Q-cycle Shown
to the left are the porphyrins found in cytochromes
Cytochrome b contains the same iron protoporphyrin as
hemoglobin and myoglobin The c cytochromes contain
heme c through covalent attachment by cysteine
residues Cytochrome a is found in two forms in
complex IV In complex III we find two b-type
cytochromes and one c-type cytochrome Complex III is
complex and we have a crystal structure
4 Complex IV
Complex IV is also known as cytochrome c oxidase
because it accepts the electrons from cytochrome c
and directs them towards the four electron reduction
of O2 to form 2 molecules of H2O
4 cyt c (Fe2+) + 4 H+ + O2 4 cyt c (Fe3+) +
2H2O
26
Cytochrome c oxidase contains 2 heme
centerscytochrome a and cytochrome a3 and two copper
proteins Each of the protein bound coppers are
associated with one of the cytochromes The copper
sites are called CuA and CuB CuA is associated with
cytochrome a and is shown to the left CuB is
associated with cytochrome a3 The copper sites
function as 1 electron carriers cycling between the
cuprous state Cu+ and the cupric state Cu2+ Just
like iron containing proteins they transfer
electrons one at a time Cytochrome c is bound on the
P-face of the membrane and transfers its electron to
CuA The oxidized cytochrome c dissociates CuA then
transfers the electron to cytochrome a The protein
bound CuA and the iron bound in cytochrome a are 15 Aring
apart In contrast the CuB and the iron bound in
cytochrome a3 are very close to each other forming a
binuclear metal center shown below
27
Electron transport chains are associated with
membranes such as the mitochondrial membrane in
eukaryotic organisms the thylakoid membrane of
photosynthetic organisms and the plasma membrane of
prokaryotic organisms While some of the details
differ electron transport chains in all organisms
operate on the same basic principles
Excited electrons are brought to the electron
transport chain by electron carriers such as NADH
and FADH2 In photosynthetic organisms the electrons
are donated by water and excited by light energy
The electron donor is oxidized as the electrons are
passed to the first protein complex in the chain As
it accepts the electrons the protein complex is
reduced Next the electrons are passed from the
28
first protein complex which becomes oxidized to
the second protein complex which becomes reduced
Each of these redox reactions is slightly exergonic
or energy releasing Some of this energy is used to
move protons across the membrane against their
concentration gradient With each successive
transfer of electrons the original excited
electrons lose some of their energy After several
successive redox reactions the electrons are
donated to a terminal electron acceptor In aerobic
respiration the terminal electron acceptor is
oxygen In photosynthesis the terminal electron
acceptor is a compound called NADP
The energy in the excited electrons has been
transformed into an electrochemical gradient
represented by the high concentration of protons on
one side of the membrane These protons can diffuse
back across the membrane by passing through a
protein complex called ATP synthase As these
protons pass through ATP synthase down their
concentration gradient they provide the energy
needed to make ATP from ADP and inorganic phosphate
within the cell
29
CHAPTER III
CLOSING
A Conclution
When a hexose is completely oxidized to CO2
and H2O using these three processes Glycolysis
yields two ATP and two NADH per hexose used Each
such NADH oxidized by the electron transport system
30
yields two ATP as dscribed above so glycolysis
contributes a total of six ATP per hexose The Krebs
cycle contributes two ATP per hexose or per two
pyruvates when succinyl CoA is cleaved to succinate
and CoASH This cycle also produces eight NADH per
hexose within the mitochondrial matrix by oxidative
phosporylation each of these NADH yields three ATP
or 24 per hexose Each FADH2 from the Krebs cycle
yields two ATP by oxydative phosporylation or four
per hexose (two pyruvates) The total contribution of
the Krebs cycle is then 30 Adding these 30 to the 6
from glycolysis leads to a total of 36 per hexose
completely respired by these process
B Suggestion
My greatest wish is that the readers find
this manuscript helpful in gaining the knowledge they
need for effective learning and practice and in
preparing for examination designed to test that
knowledge Reader comments and suggestions are
welcome and would be very helpful in improving our
manuscript Your contribution may assist many other
students in the coming years
31
REFERENCE
Anonim 2013 Anaerobic Respiration
httpwwwscoolcoukgcsebiologyrespiration
revise-itanaerobic-respiration Accessed
September 10th 2013
Anonim 2013 What is the difference between aerobic
and anaerobic respiration in plants
httpwwwbiologylifeeasyorg4010difference
-between-aerobic-anaerobic-respiration-plants
Accessed September 10th 2013
Campbell Mary K 2006 Biochemistry New York Thomson
Brooks Cole Inc
Dat J Capelli N Folzer H Bourgeade P Badot P-M
(2004) Sensing and signaling during
plantflooding Plant Physiology and
Biochemistry 42 273-282
Dat J Folzer H Parent C Badot P-M Capelli N
(2006) HypoxiastressCurrent Understanding and
Perspectives In Teixeira da Silva JA (Ed)
Floriculture Ornamental and Plant
Biotechnology Advances and Topical Issues (Vol
3) Global Science Books Isleworth United
Kingdompp 664-674
Hatta Muhammad 2011 Dunia Pertanian
httpemhattawordpresscomcategorystress
Accessed September 10th 2013
Hopkins William G 2009 Introduction to Plant Physiology
New York John Wiley amp Sons Inc
Lakitan Benyamin 2011 Dasar-Dasar Fisiologi Tumbuhan
Jakarta PT Raja Grafindo Persada
Larcher W 1980 Physiological Plant Ecology New York
Springer-Verlag
Raven 2013 Biology Fifth Edition
httphigheredmcgraw-hillcomsites983409233
9student_view0chapter39
photosynthetic_electron_transport_and_atp_synth
esishtml Accessed September 10th 2013
Salisbury Frank B 1969 Plant Physiology Third Edition
California Wadsworth Publishing Company
Subbaiah C Sachs M (2003) Molecular and cellular
adaptations of maize to flooding stress Annals
of Botany 91 119-127
Wilson CarlL1962 Botany Third EditionUnited State of
America Holt Rinch Inc

The process of glycolate metabolism has not
yet been understood in all details The glycolate is
transported out of the chloroplasts phospoglycolate
and phospate The glycolate is transported out of
the chloroplast into peroxisomes cell compartments
about the size of mitochondria which contain
glycolate oxidase catalase and transaminases In
the peroxysomes when O2 is taken up glycolate is
oxidized to glyoxylate and the peroxide thus
produced is detoxified by catalase Glycolate can
either be completely reduced via oxalate by further
O2 uptake or transformed to glycine by
transamination Glycine is transported from the
peroxisomes into the mitochondria where two
molecules of glycine are coupled to form one
molecule of serine with the release of CO2 Serine
is taken over by the amino acid metabolism or
2
converted to glycerate after deamination by
hydropyruvate This can be photophosporylated in the
chloroplasts and returned to the Calvin cycle or
used elsewhere (Larcher 1980 81-82)
B Purposes
This manuscript is intended to complete the
task from our lecturer Dr Azwir Anhar MSi and
to meet the needs of university students as the
guidersquos book in learning and practice activity for
the member of International Standard Teacher
Education (ISTE) program in Public University of
Padang
C Benefits
Benefits that we gained from this manuscript
are
1 As reference in learning process of plant
phisiology subject especially in plant
respiration title
2 As information about steps in plant
respiration
3
CHAPTER II
PLANT RESPIRATION
A Krebs Cycle
The Krebs cycle was named in honor of the
English biochemist Hans A Krebs who in 1937
proposed a cycle of reactions to explain how pyruvate
breakdown takes place in the breast muscle of
pigeons He called his proposed pathway the citric
acid cycle because citric acid is an important
intermediet Another common name for the same group
of reactions is the Tricarboxuylic Acid (TCA) cycle
a term used because citric and isocitric acids have
three carboxyl groups It was not until the early
1950s that mitochondria capable of carrying out this
cycle were isolated from plant cells
The initial step leading to the Krebs cycle
involves the oxidation and loss of CO2 from pyruvate
and the combination of the remaining 2-carbon acetate
unit with a sulfur-containing compound co-enzyme A
(CoA) forming acetyl CoA This and another
comparable role of CoA in the Krebs cycle are
important reasons why sulfur is an essential element
4
The reaction of pyruvate decarboxylation also
involves a phosporylated form of thiamine (vitamin
B1) as a prostethic group Participation of thiamine
in this section partially explains the essential
function of vitamin B1 in plants and animals Besides
the loss of CO2 two hydrogen atoms are removed from
pyruvic acid during the formation of Acetyl CoA The
enzyme catalyzing the complete reaction is called
pyruvic acid dehydrogenase but it is actually an
organized complex containing numerous copies of three
or four different enzymes The hydrogen atoms removed
are finally accepted by NAD+ yielding NADH
The Krebs cycle accomplishes removal of some of
the electrons from organic acid intermediates and
transfer of these electrons to NAD+ or FAD Notice
that none of dehydrogenase enzyme of the cycle uses
NADP+ as an electron acceptor In fact NADP+ is
usually undetectable in plant mitochondria a
situation opposite to that of chloroplasts where
NADP+ is abundant but where there is much less NAD+
Not only are NADH and FADH2 important products of the
Krebs cycle but one molecule of ATP is formed from
ADP and Pi during the conversion of succinyl coenzyme
A to succinic acid Two additional CO2 molecules are
released in these Krebs cycle reactions so there is
a net loss of both carbon atoms from the incoming
5
acetate of acetyl CoA The release of CO2 in the
Krebs cycle accounts for the product of CO2 in the
summary equation for resspiration but no O2 is
absorbed during any Kreb cycle reaction (Lakitan
2011)
During metabolism the synthesis and breakdown
of different organic compounds takes place through
various pathways like the breakdown and synthesis of
proteins carbohydrates fats and nucleic acids
These different pathways and intermediates are also
responsible for the production of energyKrebrsquos
cycle named after Hans Krebs who began working out
its details in 1930s is a series of reactions in
which the pyruvate from glycolysis is oxidized to Co2
under aerobic conditions Kreb cycle is also known as
citric acid cycle or Tri-carboxylic acid cycle (TCA)
( Galston 1968 76)
6
Steps involved in the process of Kreb cycle are
1 Fate of pyruvic acid
Pyruvic acid can form different compounds by
different pathways ie it can be converted into
lactic acid It can be converted into acetyl
coenzyme A as a result of oxidation
2 Fate of Acetyl CoA
It can either undergo condensation with itself or
its derivatives to form fatty acids having 14 ndash 20
carbons Acetyl CoA can also go through a series
of reactions in Krebs cycle
3 Formation of citrate
Acetyl CoA enzyme condenses with oxaloacetate by
an enzyme citrate synthatase to form citrate with
the release of acetyl CoA If the amount of
7
oxaloacetate is very small then small number of
acetyl CoA would be reacting with oxaloacetate
leaving surplus acetyl CoA to go through another
pathway for the formation of long chain fatty
acids
4 Formation of C is ndash acotinate and iso-citrate
Citrate is changed first into C is ndash acotinate and
then to iso-citrate under the enzyme acotinase
Equilibrium is established between citrate C is
aconitate and iso-citrate It has been observed
that most of the time this equilibrium is shifted
towards the iso-citrate If the concentration of
iso-citrate in increased the formation of citrate
will result which also indicates that the
equilibrium also shift in the reserve direction
5 Formation of oxalosucinate
Iso-citrate is acted upon by an enzyme iso-citrate
dehydrogenase using nicotinamide adenine
dinucleotide (NAD) as coenzyme As a result iso-
citrate is converted into oxalosucciate and NAD is
reduced to NADH2 Similar reaction is carried out
by same enzyme using nicotindmide adenine
dinucleotide phosphate (NADP) as coenzyme which is
reduced to NDAPH2
6 Formation of α-Ketoglutarate
8
Oxalosucciante is changed into α-ketoglutrate by
iso-citrate dehydrogenase with the help of
coenzyme NAD or NADP In this reaction carbon
dioxide and NADH2 or NADPH2 are also released
7 Formation of succinyl CoA
α-ketoglutarate combines with acetyl CoA in the
presence of coenzyme NAD and enzyme α-
keoglutaratedehyrogenase to form succinyl coenzyme
a carbon dioxide and NADH2
8 Formation of Succinate
Later on coenzyme a is removed from succinyl
coenzyme A in the presence of guanosinediphosphate
(GDP) and inorganic phosphate to form succinate
and guanosine triphosphate (GTP) This reaction is
carried out by an enzyme called succinyl CoA
synthatase
9 Formation of Funarate
An enzyme succinic dehydrogenase removes hydrogen
from succinate to form funarate
10 Formation of Malate
Fumarate reacts with water in the presence of
enzyme fumarase to form malate
11 Regeneration of oxaloacetate
Malate is oxidized by malic dehydrogenase and NAD
forming oxaloacetate and NADH2 Thus oxalo acetate
is again available to start another cycle
9
(httpkrebs-cycle-or-citric-acid-cycle-or-
trihtml)
Importance of CTA Cycle
1 Source of energy
In addition to routine organic compounds described
above by products like nicotinanide adenine
dinucleotide (NADH2) and guanosine triphosphate
(GTP) are the source of biological energy NADH2
after oxidation produce energy whereas GTP is
itself high energy phosphate compound
2 Oxidation of organic compounds taken as food
Oxidation of fats carbohydrates and proteins take
place through it or in other words it can be said
that oxidation of all compounds having carbon
atoms can take place through TCA cycle Some of
amino acids like alanine glutamic acid and
aspartic acid at one stage or the other enters
into TCA cycle eg glutamic acid enters cycle
after its transformation into α-ketoglutarate
Similarly alanine enters the cycle after its
conversion into pyruvate
3 Intermediate compounds
TCA cycle is also involved in synthesis of
intermediate compounds leading to the formation of
larger molecules (Ray 1963 17)
10
Verification of Krebs Cycle
It was done by radioactive traces like C14 as
radioactive carbon dioxide of different levels and
reactions After addition of radioactive carbon
dioxide different chemical compounds produced like
glucose fats amino acids were isolated and looked
for radioactive carbon In this way whole of
metabolic reaction were verified including individual
reactions alternative metabolic pathways
intermediates of fats carbohydrates and amino acids
etc in the body cells as well as in test tubes
(Wilson 1962 99)
The primary function of the Krebs cycle are as
follows
1 Reduction of NAD+ and FAD to the electron donors
NADH and FADH2 that are subsequently oxidized to
yield ATP
2 Direct synthesis of a limited amount of ATP (1 ATP
for each pyruvate oxidized)
3 Formation of carbon skeletons that can be used to
synthesize certain amino acids that in turn are
converted into larger molecules
11
B Fermentation
Although glycolysis can function well without
O2 the further oxidation of pyruvate and NADH by
mitochondria requires this gas Thus when O2 is
limiting NADH and pyruvate begin to accumulate
Under this condition plants carry out fermentation
(anaerobic respiration) forming either ethanol or
lactic acid (usually ethanol) As shown in the
picture below the two top reactions consists of a
decarboxylation to form acetaldehide then rapid
reduction of acetaldehide by NADH to form ethanol
These reactions are catalyzed by alcohol
dehydrogenase Some cells contain lactic acid
12
dehydrogenase which uses NADH to reduce pyruvic to
lactic acid Thus either ethanol or lactic acid or
both are fermentation products depending on the
activities of each dehydrogenase present In each
case NADH is the reductant and only under
anaerobic conditions is it abundant anough to cause
reduction Furthermore in some plants NADH is used
to cause accumulation of the other compounds when O2
is limiting especially malate and glycerol
(Salisbury 1969 233)
The fate of pyruvate produced by glycolysis
depends primarily on whether or not oxygen is
present Under normal aerobic conditions pyruvate
is transported into the mitochondrion where it is
further oxidized to CO2 and water transferring its
electrons ultimately to molecular oxygen
13
Although higher plants are obligate aerobs and
are able to tolerate anoxia for only short periods
tissues or organs are occasionally subjected to
anaerobic conditions A typical siuation is that of
roots when the soil is saturated with water When
there is no oxygen to serve as the terminal electron
acceptor mitochondrial respiration will shut down
and metabolism will shift over to fermentation
Fermentation converts pyruvate to either ethanol
through the action of the enzyme alcohol
dehydrogenase (ADH) or lactate via lactate
dehydrogenase (LDH) In most plants the principal
products of fermentation are CO2 and ethanol Some
lactate may be formed primarily in the early stages
of anoxia However lactate lowers he pH of the
cytosol which in turn activates pyruvate
decarboxylase and initiates the production of
ethanol
Either one of the fermentation reactions
consumes the NADH produced earlier in glycolysis by
the oxidation of glyceraldehide-3-P Although this
means there is no net gain of reducing potential in
fermentation This recycling of NADH is still
important to the cell The poolof NADH plus NAD+ in
the cell is relatively small and if the NADH is not
recycled there will be no supply of NAD+ to support
14
the continued oxidation of glyceraldehyde-3-P if
this were the case glycolysis and the producton of
even the small quantities of ATP necessary to
maintain the cells under anaerobic conditions would
then grind to a halt (Hopkins 2009 241)
Soil waterlogging has long been identified as a
major abiotic stress and the constraints that it
provides the roots have a significant effect on the
growth and development of plants When this event
occurs in the spring then these puddles can reduce
seed germination and seedling development Thus a
pool of water is an important factor that affects
the growth development and survival of plant
species not only in natural ecosystems but also
the agricultural and horticultural systems (Dat et
al 2006)
After flooding rapid changes in soil
properties At the time of pore water meets the
land the air pushed out gas diffusion is reduced
and toxic compounds accumulate due to anaerobic
conditions All of these changes greatly affect the
ability of plants to survive In response increased
stomatal resistance photosynthesis and root
hydraulic conductivity decreases and reduced
translocation fotoassimilat Nevertheless one of
15
the best adaptations of plants to hypoxia anoxia
is transitional biochemical and metabolic processes
common to current limited availability of O2 (Dat et
al 2004)
Selective synthesis of a set of about 20
anaerobic stress proteins (ANPS) allows the process
of producing energy without oxygen metabolism under
anaerobic conditions (Subbaiah and Sachs 2003)
Another adaptation is the observed morphological
changes consisting of the formation of hypertrophic
lenticels adventitious root initiation and or
progression of aerenchyma (Vartapetian and Jackson
1997 Jackson and Colmer 2005 Folzer et al 2006)
This review details the diverse plant stress
response to hypoxia anoxia caused by soil
waterlogging flooding and examines some of the key
features of metabolic adaptation physiological and
morphological
16
C Electron
Transport System
We noted earlier that one of the principal
functions of the respiration is to retrieve in
useful form some of the energy initially stored in
assimilates Our traditional measure of useful
energy in most processes is the number of ATP
molecules gained or consumed By this measure alone
the yield from both glycolysis and the citric acid
cycle is quite low After two complete turns of the
cycle one molecule of glucose has been completely
oxidized to six molecules of CO2 but only four
molecules of ATP have been produced (a net of two
ATP from glycolysis plus one for each turn of the
cycle) At this point most of the energy associated
with the glucose molecule has been conserved in the
17
form of electron pairs generated by the oxidation of
glycolytic and citric acid cycle intermediates
In this section we will discuss the third stage
of cellular respirationmdashthe transfer of electrons
fromNADH and FADH2 to oxygen and the accompanying
conversion of redox energy to ATP The transfer of
electrons fromNADHand FADH2 to oxygen involves a
sequence of electron carriers arranged in an
electron transport chainMembrane fractionation
studies have shown that the enzymes and electron
carriers making up the electron transport chain are
organized predominantly into four large
multimolecular complexes (complexes ImdashIV) and two
mobile carriers located in the inner mitochondrial
membrane (Figure 109) In this sense there are a
great number of similarities between the
mitochondrial inner membrane and the thylakoid
membranes of the chloroplast (compare Figure 109
with Figure 76) This is not unexpected since the
principal function of each membrane is energy
transformation and many of the same or similar
components are involved
The Components of the Electron Transport Chain
The electron transport chain of the
mitochondria is the means by which electrons are
18
removed from the reduced carrier NADH and
transferred to oxygen to yield H2O
1) NADH
NADH is generated in the matrix by the reactions
of pyruvate dehydrogenase isocitrate
dehydrogenase α-ketoglutarate dehydrogenase and
malate dehyrogenase The electron transport chain
begins with reoxidizing NADH to form NAD+ and
channeling the electrons into the formation of
reduced coenzymes Important to note that NADH
transfers 2 electrons at a time in the form of a
hydride
NAD+ + 2e- + H+ 1048581 NADH Eorsquo = minus0315 V
2) Flavoproteins
Flavoproteins have either a FAD (flavin adenosine
dinucleotide) or a FMN (flavin mononucleotide)
prosthetic group Flavoproteins can accept or
donate electrons one at time or two at a time
Thus they are often intermediaries between two
electron acceptorsdonors and one electron
acceptorsdonors For flavoproteins the typical
standard reduction potentials are around 0 V
FAD + 2e- + 2H+ 1048581 FADH2 FMN + 2e- + 2H+ 1048581 FMNH2
Eorsquo asymp 0 V
19
3) Coenzyme Q (CoQ)
aka ubiquinone (UQ) Shown below CoQ has ten
repeating isoprene units which make it
insoluble in water but soluble in the
hydrophobic lipid bylayer Coenzyme Q is a
versatile cofactor because it is a soluble
electron carrier in the hydrophobic bilipid layer
of the
inner mitochondrial membrane Like flavoproteins
CoQ can acceptdonate electrons one at a time or
two at a time
Q + 2e- + 2H+ 1048581 QH2 Eorsquo =0060 V
Q + e- + H+ 1048581 1048581QH Eorsquo =0030 V
1048581QH + e- + H+ 1048581 QH2 Eorsquo =0190 V
4) Cytochromes
Cytochromes are proteins that contain heme
prosthetic groups which function as
one electron carriers The heme iron is involved
in one electron transfers involving the Fe2+ and
Fe3+ oxidation states Cytochromes are named by
their absorption spectra which depends
on the porphyrin structure and environment The
example shown is the heme prosthetic group of
cytochrome b Cytochrome b contains the same iron
porphyrin found in hemoglobin and myoglobin
20
Other cytochromes we will encounter in the
electron transport complexes are cytochromes b
c c1 a and a3
5) Iron-Sulfur Proteins
In the electron transport chain we will encounter
many iron-sulfer proteins which participate in
one electron transfers involving the the Fe2+ and
Fe3+ oxidation states These are non-heme iron-
sulfur proteinsThe simplest iron-sulfer protein
is FeS in which iron is tetrahedrally coordinated
by four cysteines The second form is Fe2S2 which
contains two irons complexed to 2 cysteine
residues and two inorganic sulfides The third
form is Fe3S4 which contains 3 iron atoms
coordinated to three cysteine residues and 4
inorganic sulfides The last form is the most
complicated Fe4S4 which contains 4 iron atoms
coordinated to 4 cysteine residues and 4
inorganic sulfides
6) Copper Proteins
Copper bound proteins participate in one electron
transfers involving the Cu+ and Cu2+ oxidation
states
21
Overview of the Electron Transport Chain
Electrons move along the electron transport
chain going from donor to acceptor until they reach
oxygen the ultimate electron acceptor The standard
reduction potentials of the electron carriers are
between the NADHNAD+ couple (-0315 V) and the
oxygenH2O couple (0816 V) as on the next page The
components of the electron transport chain are
organized into 4 complexes Each complex contains
several different electron carriers
1 Complex I also known as the NADH-coenzyme Q
reductase or NADH dehydrogenase
2 Complex II also known as succinate-coenzyme Q
reductase or succinate dehydrogenase
3 Complex III also known as coenzyme Q reductase
4 Complex IV also known as cytochrome c reductase
The free energy necessary to generate ATP
is extracted from the oxidation of NADH and
FADH2 by the electron transport chain It
consists of four protein complexes and single
protein chain of cytochrome c through which
electrons pass from lower to higher standard
redox potentials Electrons are carried from complex
I and II to complex III by the membrane (lipid)
soluble coenzyme Q (CoQ or ubiquinone) and between
22
complexes III and IV by the peripheral membrane
protein cytochrome c
In the following I give the overall
reactions catalysed by corresponding protein
complexes in the mitochondrial electron transport
chain
1 Complex I
Complex I is also called NADH-Coenzyme Q
reductase because this large protein complex
transfers 2 electrons from NADH to coenzyme Q
Complex I was formerly known as NADH dehydrogenase
Complex I is huge 850000 kD and is composed of more
than thirty subunits It contains a FMN prosthetic
group and seven or more Fe-S clusters This complex
has between 20-26 iron atoms boundThe prosthetic
group FMN is absolutely required for activity
Therefore this complex is a flavoprotein This
complex binds NADH transfers two electrons in the
form of a hydride to FMN to produce NAD+ and FMNH2
23
The subsequent steps involve the transfer of
electrons one at a time to a series of iron-sulfer
complexes that includes both 2Fe-2S and 4Fe-4S
clusters Note the importance of FMN First it
functions as a 2 electron acceptor in the hydride
transfer from NADH Second it functions as a 1
electron donor to the series of iron sulfur clusters
FMN and FAD often play crucial links between 2
electron transfer agents and 1 electron transfer
agents The final step of this complex is the
transfer of 2 electrons one at a time to coenzyme Q
CoQ like FMN and FAD can function as a 2 electron
donoracceptor and as a 1 electron donoracceptor
CoQ is a mobile electron carrier because its
isoprenoid tail makes it highly hydrophobic and
lipophillic It diffuses freely in the bilipid layer
of the inner mitochondrial membrane
The process of transferring electrons from NADH
to CoQ by complex I results in the net transport of
protons from the matrix side of the inner
mitochondrial membrane to the inter membrane space
where the H+ ions accumulate generating a proton
motive force The intermembrane space side of the
inner membrane is referred to as the P face (P
standing for positive) The matrix side of the inner
membrane is referred to as the the N face The
24
transport of electrons from NADH to CoQ is coupled to
the transport of protons across the membrane This is
an example of active transport The stiochiometry is
4 H+ transported per 2 electrons
2 Complex II
Believe it or not you are already familiar
with Complex II It is none other than succinate
dehydrogenase The only enzyme of the citric acid
cycle that is an integral membrane protein This
complex is composed of four subunits 2 of which are
iron-sulfur proteins and the other two subunits
together bind FAD through a covalent link to a
histidine residue These two subunits are called
flavoprotein 2 or FP2 Complex II contains 3 Fe-S
centers 1 4Fe-4S cluster 1 3Fe-4S cluster and 1
2Fe- 2S cluster In the first step of this complex
succinate is bound and a hydride is transferred to
FAD to generate FADH2 and fumarate FADH2 then
transfers its electrons one at a time to the Fe-S
centers Thus once again FAD functions as 2 electron
acceptor and a 1 electron donor The final step of
this complex is the transfer of 2 electrons one at a
time to coenzyme Q to produce CoQH2 For complex II
the standard free energy change of the overall
reaction is too small to drive the transport of
protons across the inner mitochondrial membrane This
25
accounts for the 15 ATPrsquos generated per FADH2
compared with the 25 ATPrsquos generated per NADH
3 Complex III
This complex is also known as coenzyme Q-
cytochrome c reductase because it passes the
electrons form CoQH2 to cyt c through a very unique
electron transport pathway called the Q-cycle Shown
to the left are the porphyrins found in cytochromes
Cytochrome b contains the same iron protoporphyrin as
hemoglobin and myoglobin The c cytochromes contain
heme c through covalent attachment by cysteine
residues Cytochrome a is found in two forms in
complex IV In complex III we find two b-type
cytochromes and one c-type cytochrome Complex III is
complex and we have a crystal structure
4 Complex IV
Complex IV is also known as cytochrome c oxidase
because it accepts the electrons from cytochrome c
and directs them towards the four electron reduction
of O2 to form 2 molecules of H2O
4 cyt c (Fe2+) + 4 H+ + O2 4 cyt c (Fe3+) +
2H2O
26
Cytochrome c oxidase contains 2 heme
centerscytochrome a and cytochrome a3 and two copper
proteins Each of the protein bound coppers are
associated with one of the cytochromes The copper
sites are called CuA and CuB CuA is associated with
cytochrome a and is shown to the left CuB is
associated with cytochrome a3 The copper sites
function as 1 electron carriers cycling between the
cuprous state Cu+ and the cupric state Cu2+ Just
like iron containing proteins they transfer
electrons one at a time Cytochrome c is bound on the
P-face of the membrane and transfers its electron to
CuA The oxidized cytochrome c dissociates CuA then
transfers the electron to cytochrome a The protein
bound CuA and the iron bound in cytochrome a are 15 Aring
apart In contrast the CuB and the iron bound in
cytochrome a3 are very close to each other forming a
binuclear metal center shown below
27
Electron transport chains are associated with
membranes such as the mitochondrial membrane in
eukaryotic organisms the thylakoid membrane of
photosynthetic organisms and the plasma membrane of
prokaryotic organisms While some of the details
differ electron transport chains in all organisms
operate on the same basic principles
Excited electrons are brought to the electron
transport chain by electron carriers such as NADH
and FADH2 In photosynthetic organisms the electrons
are donated by water and excited by light energy
The electron donor is oxidized as the electrons are
passed to the first protein complex in the chain As
it accepts the electrons the protein complex is
reduced Next the electrons are passed from the
28
first protein complex which becomes oxidized to
the second protein complex which becomes reduced
Each of these redox reactions is slightly exergonic
or energy releasing Some of this energy is used to
move protons across the membrane against their
concentration gradient With each successive
transfer of electrons the original excited
electrons lose some of their energy After several
successive redox reactions the electrons are
donated to a terminal electron acceptor In aerobic
respiration the terminal electron acceptor is
oxygen In photosynthesis the terminal electron
acceptor is a compound called NADP
The energy in the excited electrons has been
transformed into an electrochemical gradient
represented by the high concentration of protons on
one side of the membrane These protons can diffuse
back across the membrane by passing through a
protein complex called ATP synthase As these
protons pass through ATP synthase down their
concentration gradient they provide the energy
needed to make ATP from ADP and inorganic phosphate
within the cell
29
CHAPTER III
CLOSING
A Conclution
When a hexose is completely oxidized to CO2
and H2O using these three processes Glycolysis
yields two ATP and two NADH per hexose used Each
such NADH oxidized by the electron transport system
30
yields two ATP as dscribed above so glycolysis
contributes a total of six ATP per hexose The Krebs
cycle contributes two ATP per hexose or per two
pyruvates when succinyl CoA is cleaved to succinate
and CoASH This cycle also produces eight NADH per
hexose within the mitochondrial matrix by oxidative
phosporylation each of these NADH yields three ATP
or 24 per hexose Each FADH2 from the Krebs cycle
yields two ATP by oxydative phosporylation or four
per hexose (two pyruvates) The total contribution of
the Krebs cycle is then 30 Adding these 30 to the 6
from glycolysis leads to a total of 36 per hexose
completely respired by these process
B Suggestion
My greatest wish is that the readers find
this manuscript helpful in gaining the knowledge they
need for effective learning and practice and in
preparing for examination designed to test that
knowledge Reader comments and suggestions are
welcome and would be very helpful in improving our
manuscript Your contribution may assist many other
students in the coming years
31
REFERENCE
Anonim 2013 Anaerobic Respiration
httpwwwscoolcoukgcsebiologyrespiration
revise-itanaerobic-respiration Accessed
September 10th 2013
Anonim 2013 What is the difference between aerobic
and anaerobic respiration in plants
httpwwwbiologylifeeasyorg4010difference
-between-aerobic-anaerobic-respiration-plants
Accessed September 10th 2013
Campbell Mary K 2006 Biochemistry New York Thomson
Brooks Cole Inc
Dat J Capelli N Folzer H Bourgeade P Badot P-M
(2004) Sensing and signaling during
plantflooding Plant Physiology and
Biochemistry 42 273-282
Dat J Folzer H Parent C Badot P-M Capelli N
(2006) HypoxiastressCurrent Understanding and
Perspectives In Teixeira da Silva JA (Ed)
Floriculture Ornamental and Plant
Biotechnology Advances and Topical Issues (Vol
3) Global Science Books Isleworth United
Kingdompp 664-674
Hatta Muhammad 2011 Dunia Pertanian
httpemhattawordpresscomcategorystress
Accessed September 10th 2013
Hopkins William G 2009 Introduction to Plant Physiology
New York John Wiley amp Sons Inc
Lakitan Benyamin 2011 Dasar-Dasar Fisiologi Tumbuhan
Jakarta PT Raja Grafindo Persada
Larcher W 1980 Physiological Plant Ecology New York
Springer-Verlag
Raven 2013 Biology Fifth Edition
httphigheredmcgraw-hillcomsites983409233
9student_view0chapter39
photosynthetic_electron_transport_and_atp_synth
esishtml Accessed September 10th 2013
Salisbury Frank B 1969 Plant Physiology Third Edition
California Wadsworth Publishing Company
Subbaiah C Sachs M (2003) Molecular and cellular
adaptations of maize to flooding stress Annals
of Botany 91 119-127
Wilson CarlL1962 Botany Third EditionUnited State of
America Holt Rinch Inc

converted to glycerate after deamination by
hydropyruvate This can be photophosporylated in the
chloroplasts and returned to the Calvin cycle or
used elsewhere (Larcher 1980 81-82)
B Purposes
This manuscript is intended to complete the
task from our lecturer Dr Azwir Anhar MSi and
to meet the needs of university students as the
guidersquos book in learning and practice activity for
the member of International Standard Teacher
Education (ISTE) program in Public University of
Padang
C Benefits
Benefits that we gained from this manuscript
are
1 As reference in learning process of plant
phisiology subject especially in plant
respiration title
2 As information about steps in plant
respiration
3
CHAPTER II
PLANT RESPIRATION
A Krebs Cycle
The Krebs cycle was named in honor of the
English biochemist Hans A Krebs who in 1937
proposed a cycle of reactions to explain how pyruvate
breakdown takes place in the breast muscle of
pigeons He called his proposed pathway the citric
acid cycle because citric acid is an important
intermediet Another common name for the same group
of reactions is the Tricarboxuylic Acid (TCA) cycle
a term used because citric and isocitric acids have
three carboxyl groups It was not until the early
1950s that mitochondria capable of carrying out this
cycle were isolated from plant cells
The initial step leading to the Krebs cycle
involves the oxidation and loss of CO2 from pyruvate
and the combination of the remaining 2-carbon acetate
unit with a sulfur-containing compound co-enzyme A
(CoA) forming acetyl CoA This and another
comparable role of CoA in the Krebs cycle are
important reasons why sulfur is an essential element
4
The reaction of pyruvate decarboxylation also
involves a phosporylated form of thiamine (vitamin
B1) as a prostethic group Participation of thiamine
in this section partially explains the essential
function of vitamin B1 in plants and animals Besides
the loss of CO2 two hydrogen atoms are removed from
pyruvic acid during the formation of Acetyl CoA The
enzyme catalyzing the complete reaction is called
pyruvic acid dehydrogenase but it is actually an
organized complex containing numerous copies of three
or four different enzymes The hydrogen atoms removed
are finally accepted by NAD+ yielding NADH
The Krebs cycle accomplishes removal of some of
the electrons from organic acid intermediates and
transfer of these electrons to NAD+ or FAD Notice
that none of dehydrogenase enzyme of the cycle uses
NADP+ as an electron acceptor In fact NADP+ is
usually undetectable in plant mitochondria a
situation opposite to that of chloroplasts where
NADP+ is abundant but where there is much less NAD+
Not only are NADH and FADH2 important products of the
Krebs cycle but one molecule of ATP is formed from
ADP and Pi during the conversion of succinyl coenzyme
A to succinic acid Two additional CO2 molecules are
released in these Krebs cycle reactions so there is
a net loss of both carbon atoms from the incoming
5
acetate of acetyl CoA The release of CO2 in the
Krebs cycle accounts for the product of CO2 in the
summary equation for resspiration but no O2 is
absorbed during any Kreb cycle reaction (Lakitan
2011)
During metabolism the synthesis and breakdown
of different organic compounds takes place through
various pathways like the breakdown and synthesis of
proteins carbohydrates fats and nucleic acids
These different pathways and intermediates are also
responsible for the production of energyKrebrsquos
cycle named after Hans Krebs who began working out
its details in 1930s is a series of reactions in
which the pyruvate from glycolysis is oxidized to Co2
under aerobic conditions Kreb cycle is also known as
citric acid cycle or Tri-carboxylic acid cycle (TCA)
( Galston 1968 76)
6
Steps involved in the process of Kreb cycle are
1 Fate of pyruvic acid
Pyruvic acid can form different compounds by
different pathways ie it can be converted into
lactic acid It can be converted into acetyl
coenzyme A as a result of oxidation
2 Fate of Acetyl CoA
It can either undergo condensation with itself or
its derivatives to form fatty acids having 14 ndash 20
carbons Acetyl CoA can also go through a series
of reactions in Krebs cycle
3 Formation of citrate
Acetyl CoA enzyme condenses with oxaloacetate by
an enzyme citrate synthatase to form citrate with
the release of acetyl CoA If the amount of
7
oxaloacetate is very small then small number of
acetyl CoA would be reacting with oxaloacetate
leaving surplus acetyl CoA to go through another
pathway for the formation of long chain fatty
acids
4 Formation of C is ndash acotinate and iso-citrate
Citrate is changed first into C is ndash acotinate and
then to iso-citrate under the enzyme acotinase
Equilibrium is established between citrate C is
aconitate and iso-citrate It has been observed
that most of the time this equilibrium is shifted
towards the iso-citrate If the concentration of
iso-citrate in increased the formation of citrate
will result which also indicates that the
equilibrium also shift in the reserve direction
5 Formation of oxalosucinate
Iso-citrate is acted upon by an enzyme iso-citrate
dehydrogenase using nicotinamide adenine
dinucleotide (NAD) as coenzyme As a result iso-
citrate is converted into oxalosucciate and NAD is
reduced to NADH2 Similar reaction is carried out
by same enzyme using nicotindmide adenine
dinucleotide phosphate (NADP) as coenzyme which is
reduced to NDAPH2
6 Formation of α-Ketoglutarate
8
Oxalosucciante is changed into α-ketoglutrate by
iso-citrate dehydrogenase with the help of
coenzyme NAD or NADP In this reaction carbon
dioxide and NADH2 or NADPH2 are also released
7 Formation of succinyl CoA
α-ketoglutarate combines with acetyl CoA in the
presence of coenzyme NAD and enzyme α-
keoglutaratedehyrogenase to form succinyl coenzyme
a carbon dioxide and NADH2
8 Formation of Succinate
Later on coenzyme a is removed from succinyl
coenzyme A in the presence of guanosinediphosphate
(GDP) and inorganic phosphate to form succinate
and guanosine triphosphate (GTP) This reaction is
carried out by an enzyme called succinyl CoA
synthatase
9 Formation of Funarate
An enzyme succinic dehydrogenase removes hydrogen
from succinate to form funarate
10 Formation of Malate
Fumarate reacts with water in the presence of
enzyme fumarase to form malate
11 Regeneration of oxaloacetate
Malate is oxidized by malic dehydrogenase and NAD
forming oxaloacetate and NADH2 Thus oxalo acetate
is again available to start another cycle
9
(httpkrebs-cycle-or-citric-acid-cycle-or-
trihtml)
Importance of CTA Cycle
1 Source of energy
In addition to routine organic compounds described
above by products like nicotinanide adenine
dinucleotide (NADH2) and guanosine triphosphate
(GTP) are the source of biological energy NADH2
after oxidation produce energy whereas GTP is
itself high energy phosphate compound
2 Oxidation of organic compounds taken as food
Oxidation of fats carbohydrates and proteins take
place through it or in other words it can be said
that oxidation of all compounds having carbon
atoms can take place through TCA cycle Some of
amino acids like alanine glutamic acid and
aspartic acid at one stage or the other enters
into TCA cycle eg glutamic acid enters cycle
after its transformation into α-ketoglutarate
Similarly alanine enters the cycle after its
conversion into pyruvate
3 Intermediate compounds
TCA cycle is also involved in synthesis of
intermediate compounds leading to the formation of
larger molecules (Ray 1963 17)
10
Verification of Krebs Cycle
It was done by radioactive traces like C14 as
radioactive carbon dioxide of different levels and
reactions After addition of radioactive carbon
dioxide different chemical compounds produced like
glucose fats amino acids were isolated and looked
for radioactive carbon In this way whole of
metabolic reaction were verified including individual
reactions alternative metabolic pathways
intermediates of fats carbohydrates and amino acids
etc in the body cells as well as in test tubes
(Wilson 1962 99)
The primary function of the Krebs cycle are as
follows
1 Reduction of NAD+ and FAD to the electron donors
NADH and FADH2 that are subsequently oxidized to
yield ATP
2 Direct synthesis of a limited amount of ATP (1 ATP
for each pyruvate oxidized)
3 Formation of carbon skeletons that can be used to
synthesize certain amino acids that in turn are
converted into larger molecules
11
B Fermentation
Although glycolysis can function well without
O2 the further oxidation of pyruvate and NADH by
mitochondria requires this gas Thus when O2 is
limiting NADH and pyruvate begin to accumulate
Under this condition plants carry out fermentation
(anaerobic respiration) forming either ethanol or
lactic acid (usually ethanol) As shown in the
picture below the two top reactions consists of a
decarboxylation to form acetaldehide then rapid
reduction of acetaldehide by NADH to form ethanol
These reactions are catalyzed by alcohol
dehydrogenase Some cells contain lactic acid
12
dehydrogenase which uses NADH to reduce pyruvic to
lactic acid Thus either ethanol or lactic acid or
both are fermentation products depending on the
activities of each dehydrogenase present In each
case NADH is the reductant and only under
anaerobic conditions is it abundant anough to cause
reduction Furthermore in some plants NADH is used
to cause accumulation of the other compounds when O2
is limiting especially malate and glycerol
(Salisbury 1969 233)
The fate of pyruvate produced by glycolysis
depends primarily on whether or not oxygen is
present Under normal aerobic conditions pyruvate
is transported into the mitochondrion where it is
further oxidized to CO2 and water transferring its
electrons ultimately to molecular oxygen
13
Although higher plants are obligate aerobs and
are able to tolerate anoxia for only short periods
tissues or organs are occasionally subjected to
anaerobic conditions A typical siuation is that of
roots when the soil is saturated with water When
there is no oxygen to serve as the terminal electron
acceptor mitochondrial respiration will shut down
and metabolism will shift over to fermentation
Fermentation converts pyruvate to either ethanol
through the action of the enzyme alcohol
dehydrogenase (ADH) or lactate via lactate
dehydrogenase (LDH) In most plants the principal
products of fermentation are CO2 and ethanol Some
lactate may be formed primarily in the early stages
of anoxia However lactate lowers he pH of the
cytosol which in turn activates pyruvate
decarboxylase and initiates the production of
ethanol
Either one of the fermentation reactions
consumes the NADH produced earlier in glycolysis by
the oxidation of glyceraldehide-3-P Although this
means there is no net gain of reducing potential in
fermentation This recycling of NADH is still
important to the cell The poolof NADH plus NAD+ in
the cell is relatively small and if the NADH is not
recycled there will be no supply of NAD+ to support
14
the continued oxidation of glyceraldehyde-3-P if
this were the case glycolysis and the producton of
even the small quantities of ATP necessary to
maintain the cells under anaerobic conditions would
then grind to a halt (Hopkins 2009 241)
Soil waterlogging has long been identified as a
major abiotic stress and the constraints that it
provides the roots have a significant effect on the
growth and development of plants When this event
occurs in the spring then these puddles can reduce
seed germination and seedling development Thus a
pool of water is an important factor that affects
the growth development and survival of plant
species not only in natural ecosystems but also
the agricultural and horticultural systems (Dat et
al 2006)
After flooding rapid changes in soil
properties At the time of pore water meets the
land the air pushed out gas diffusion is reduced
and toxic compounds accumulate due to anaerobic
conditions All of these changes greatly affect the
ability of plants to survive In response increased
stomatal resistance photosynthesis and root
hydraulic conductivity decreases and reduced
translocation fotoassimilat Nevertheless one of
15
the best adaptations of plants to hypoxia anoxia
is transitional biochemical and metabolic processes
common to current limited availability of O2 (Dat et
al 2004)
Selective synthesis of a set of about 20
anaerobic stress proteins (ANPS) allows the process
of producing energy without oxygen metabolism under
anaerobic conditions (Subbaiah and Sachs 2003)
Another adaptation is the observed morphological
changes consisting of the formation of hypertrophic
lenticels adventitious root initiation and or
progression of aerenchyma (Vartapetian and Jackson
1997 Jackson and Colmer 2005 Folzer et al 2006)
This review details the diverse plant stress
response to hypoxia anoxia caused by soil
waterlogging flooding and examines some of the key
features of metabolic adaptation physiological and
morphological
16
C Electron
Transport System
We noted earlier that one of the principal
functions of the respiration is to retrieve in
useful form some of the energy initially stored in
assimilates Our traditional measure of useful
energy in most processes is the number of ATP
molecules gained or consumed By this measure alone
the yield from both glycolysis and the citric acid
cycle is quite low After two complete turns of the
cycle one molecule of glucose has been completely
oxidized to six molecules of CO2 but only four
molecules of ATP have been produced (a net of two
ATP from glycolysis plus one for each turn of the
cycle) At this point most of the energy associated
with the glucose molecule has been conserved in the
17
form of electron pairs generated by the oxidation of
glycolytic and citric acid cycle intermediates
In this section we will discuss the third stage
of cellular respirationmdashthe transfer of electrons
fromNADH and FADH2 to oxygen and the accompanying
conversion of redox energy to ATP The transfer of
electrons fromNADHand FADH2 to oxygen involves a
sequence of electron carriers arranged in an
electron transport chainMembrane fractionation
studies have shown that the enzymes and electron
carriers making up the electron transport chain are
organized predominantly into four large
multimolecular complexes (complexes ImdashIV) and two
mobile carriers located in the inner mitochondrial
membrane (Figure 109) In this sense there are a
great number of similarities between the
mitochondrial inner membrane and the thylakoid
membranes of the chloroplast (compare Figure 109
with Figure 76) This is not unexpected since the
principal function of each membrane is energy
transformation and many of the same or similar
components are involved
The Components of the Electron Transport Chain
The electron transport chain of the
mitochondria is the means by which electrons are
18
removed from the reduced carrier NADH and
transferred to oxygen to yield H2O
1) NADH
NADH is generated in the matrix by the reactions
of pyruvate dehydrogenase isocitrate
dehydrogenase α-ketoglutarate dehydrogenase and
malate dehyrogenase The electron transport chain
begins with reoxidizing NADH to form NAD+ and
channeling the electrons into the formation of
reduced coenzymes Important to note that NADH
transfers 2 electrons at a time in the form of a
hydride
NAD+ + 2e- + H+ 1048581 NADH Eorsquo = minus0315 V
2) Flavoproteins
Flavoproteins have either a FAD (flavin adenosine
dinucleotide) or a FMN (flavin mononucleotide)
prosthetic group Flavoproteins can accept or
donate electrons one at time or two at a time
Thus they are often intermediaries between two
electron acceptorsdonors and one electron
acceptorsdonors For flavoproteins the typical
standard reduction potentials are around 0 V
FAD + 2e- + 2H+ 1048581 FADH2 FMN + 2e- + 2H+ 1048581 FMNH2
Eorsquo asymp 0 V
19
3) Coenzyme Q (CoQ)
aka ubiquinone (UQ) Shown below CoQ has ten
repeating isoprene units which make it
insoluble in water but soluble in the
hydrophobic lipid bylayer Coenzyme Q is a
versatile cofactor because it is a soluble
electron carrier in the hydrophobic bilipid layer
of the
inner mitochondrial membrane Like flavoproteins
CoQ can acceptdonate electrons one at a time or
two at a time
Q + 2e- + 2H+ 1048581 QH2 Eorsquo =0060 V
Q + e- + H+ 1048581 1048581QH Eorsquo =0030 V
1048581QH + e- + H+ 1048581 QH2 Eorsquo =0190 V
4) Cytochromes
Cytochromes are proteins that contain heme
prosthetic groups which function as
one electron carriers The heme iron is involved
in one electron transfers involving the Fe2+ and
Fe3+ oxidation states Cytochromes are named by
their absorption spectra which depends
on the porphyrin structure and environment The
example shown is the heme prosthetic group of
cytochrome b Cytochrome b contains the same iron
porphyrin found in hemoglobin and myoglobin
20
Other cytochromes we will encounter in the
electron transport complexes are cytochromes b
c c1 a and a3
5) Iron-Sulfur Proteins
In the electron transport chain we will encounter
many iron-sulfer proteins which participate in
one electron transfers involving the the Fe2+ and
Fe3+ oxidation states These are non-heme iron-
sulfur proteinsThe simplest iron-sulfer protein
is FeS in which iron is tetrahedrally coordinated
by four cysteines The second form is Fe2S2 which
contains two irons complexed to 2 cysteine
residues and two inorganic sulfides The third
form is Fe3S4 which contains 3 iron atoms
coordinated to three cysteine residues and 4
inorganic sulfides The last form is the most
complicated Fe4S4 which contains 4 iron atoms
coordinated to 4 cysteine residues and 4
inorganic sulfides
6) Copper Proteins
Copper bound proteins participate in one electron
transfers involving the Cu+ and Cu2+ oxidation
states
21
Overview of the Electron Transport Chain
Electrons move along the electron transport
chain going from donor to acceptor until they reach
oxygen the ultimate electron acceptor The standard
reduction potentials of the electron carriers are
between the NADHNAD+ couple (-0315 V) and the
oxygenH2O couple (0816 V) as on the next page The
components of the electron transport chain are
organized into 4 complexes Each complex contains
several different electron carriers
1 Complex I also known as the NADH-coenzyme Q
reductase or NADH dehydrogenase
2 Complex II also known as succinate-coenzyme Q
reductase or succinate dehydrogenase
3 Complex III also known as coenzyme Q reductase
4 Complex IV also known as cytochrome c reductase
The free energy necessary to generate ATP
is extracted from the oxidation of NADH and
FADH2 by the electron transport chain It
consists of four protein complexes and single
protein chain of cytochrome c through which
electrons pass from lower to higher standard
redox potentials Electrons are carried from complex
I and II to complex III by the membrane (lipid)
soluble coenzyme Q (CoQ or ubiquinone) and between
22
complexes III and IV by the peripheral membrane
protein cytochrome c
In the following I give the overall
reactions catalysed by corresponding protein
complexes in the mitochondrial electron transport
chain
1 Complex I
Complex I is also called NADH-Coenzyme Q
reductase because this large protein complex
transfers 2 electrons from NADH to coenzyme Q
Complex I was formerly known as NADH dehydrogenase
Complex I is huge 850000 kD and is composed of more
than thirty subunits It contains a FMN prosthetic
group and seven or more Fe-S clusters This complex
has between 20-26 iron atoms boundThe prosthetic
group FMN is absolutely required for activity
Therefore this complex is a flavoprotein This
complex binds NADH transfers two electrons in the
form of a hydride to FMN to produce NAD+ and FMNH2
23
The subsequent steps involve the transfer of
electrons one at a time to a series of iron-sulfer
complexes that includes both 2Fe-2S and 4Fe-4S
clusters Note the importance of FMN First it
functions as a 2 electron acceptor in the hydride
transfer from NADH Second it functions as a 1
electron donor to the series of iron sulfur clusters
FMN and FAD often play crucial links between 2
electron transfer agents and 1 electron transfer
agents The final step of this complex is the
transfer of 2 electrons one at a time to coenzyme Q
CoQ like FMN and FAD can function as a 2 electron
donoracceptor and as a 1 electron donoracceptor
CoQ is a mobile electron carrier because its
isoprenoid tail makes it highly hydrophobic and
lipophillic It diffuses freely in the bilipid layer
of the inner mitochondrial membrane
The process of transferring electrons from NADH
to CoQ by complex I results in the net transport of
protons from the matrix side of the inner
mitochondrial membrane to the inter membrane space
where the H+ ions accumulate generating a proton
motive force The intermembrane space side of the
inner membrane is referred to as the P face (P
standing for positive) The matrix side of the inner
membrane is referred to as the the N face The
24
transport of electrons from NADH to CoQ is coupled to
the transport of protons across the membrane This is
an example of active transport The stiochiometry is
4 H+ transported per 2 electrons
2 Complex II
Believe it or not you are already familiar
with Complex II It is none other than succinate
dehydrogenase The only enzyme of the citric acid
cycle that is an integral membrane protein This
complex is composed of four subunits 2 of which are
iron-sulfur proteins and the other two subunits
together bind FAD through a covalent link to a
histidine residue These two subunits are called
flavoprotein 2 or FP2 Complex II contains 3 Fe-S
centers 1 4Fe-4S cluster 1 3Fe-4S cluster and 1
2Fe- 2S cluster In the first step of this complex
succinate is bound and a hydride is transferred to
FAD to generate FADH2 and fumarate FADH2 then
transfers its electrons one at a time to the Fe-S
centers Thus once again FAD functions as 2 electron
acceptor and a 1 electron donor The final step of
this complex is the transfer of 2 electrons one at a
time to coenzyme Q to produce CoQH2 For complex II
the standard free energy change of the overall
reaction is too small to drive the transport of
protons across the inner mitochondrial membrane This
25
accounts for the 15 ATPrsquos generated per FADH2
compared with the 25 ATPrsquos generated per NADH
3 Complex III
This complex is also known as coenzyme Q-
cytochrome c reductase because it passes the
electrons form CoQH2 to cyt c through a very unique
electron transport pathway called the Q-cycle Shown
to the left are the porphyrins found in cytochromes
Cytochrome b contains the same iron protoporphyrin as
hemoglobin and myoglobin The c cytochromes contain
heme c through covalent attachment by cysteine
residues Cytochrome a is found in two forms in
complex IV In complex III we find two b-type
cytochromes and one c-type cytochrome Complex III is
complex and we have a crystal structure
4 Complex IV
Complex IV is also known as cytochrome c oxidase
because it accepts the electrons from cytochrome c
and directs them towards the four electron reduction
of O2 to form 2 molecules of H2O
4 cyt c (Fe2+) + 4 H+ + O2 4 cyt c (Fe3+) +
2H2O
26
Cytochrome c oxidase contains 2 heme
centerscytochrome a and cytochrome a3 and two copper
proteins Each of the protein bound coppers are
associated with one of the cytochromes The copper
sites are called CuA and CuB CuA is associated with
cytochrome a and is shown to the left CuB is
associated with cytochrome a3 The copper sites
function as 1 electron carriers cycling between the
cuprous state Cu+ and the cupric state Cu2+ Just
like iron containing proteins they transfer
electrons one at a time Cytochrome c is bound on the
P-face of the membrane and transfers its electron to
CuA The oxidized cytochrome c dissociates CuA then
transfers the electron to cytochrome a The protein
bound CuA and the iron bound in cytochrome a are 15 Aring
apart In contrast the CuB and the iron bound in
cytochrome a3 are very close to each other forming a
binuclear metal center shown below
27
Electron transport chains are associated with
membranes such as the mitochondrial membrane in
eukaryotic organisms the thylakoid membrane of
photosynthetic organisms and the plasma membrane of
prokaryotic organisms While some of the details
differ electron transport chains in all organisms
operate on the same basic principles
Excited electrons are brought to the electron
transport chain by electron carriers such as NADH
and FADH2 In photosynthetic organisms the electrons
are donated by water and excited by light energy
The electron donor is oxidized as the electrons are
passed to the first protein complex in the chain As
it accepts the electrons the protein complex is
reduced Next the electrons are passed from the
28
first protein complex which becomes oxidized to
the second protein complex which becomes reduced
Each of these redox reactions is slightly exergonic
or energy releasing Some of this energy is used to
move protons across the membrane against their
concentration gradient With each successive
transfer of electrons the original excited
electrons lose some of their energy After several
successive redox reactions the electrons are
donated to a terminal electron acceptor In aerobic
respiration the terminal electron acceptor is
oxygen In photosynthesis the terminal electron
acceptor is a compound called NADP
The energy in the excited electrons has been
transformed into an electrochemical gradient
represented by the high concentration of protons on
one side of the membrane These protons can diffuse
back across the membrane by passing through a
protein complex called ATP synthase As these
protons pass through ATP synthase down their
concentration gradient they provide the energy
needed to make ATP from ADP and inorganic phosphate
within the cell
29
CHAPTER III
CLOSING
A Conclution
When a hexose is completely oxidized to CO2
and H2O using these three processes Glycolysis
yields two ATP and two NADH per hexose used Each
such NADH oxidized by the electron transport system
30
yields two ATP as dscribed above so glycolysis
contributes a total of six ATP per hexose The Krebs
cycle contributes two ATP per hexose or per two
pyruvates when succinyl CoA is cleaved to succinate
and CoASH This cycle also produces eight NADH per
hexose within the mitochondrial matrix by oxidative
phosporylation each of these NADH yields three ATP
or 24 per hexose Each FADH2 from the Krebs cycle
yields two ATP by oxydative phosporylation or four
per hexose (two pyruvates) The total contribution of
the Krebs cycle is then 30 Adding these 30 to the 6
from glycolysis leads to a total of 36 per hexose
completely respired by these process
B Suggestion
My greatest wish is that the readers find
this manuscript helpful in gaining the knowledge they
need for effective learning and practice and in
preparing for examination designed to test that
knowledge Reader comments and suggestions are
welcome and would be very helpful in improving our
manuscript Your contribution may assist many other
students in the coming years
31
REFERENCE
Anonim 2013 Anaerobic Respiration
httpwwwscoolcoukgcsebiologyrespiration
revise-itanaerobic-respiration Accessed
September 10th 2013
Anonim 2013 What is the difference between aerobic
and anaerobic respiration in plants
httpwwwbiologylifeeasyorg4010difference
-between-aerobic-anaerobic-respiration-plants
Accessed September 10th 2013
Campbell Mary K 2006 Biochemistry New York Thomson
Brooks Cole Inc
Dat J Capelli N Folzer H Bourgeade P Badot P-M
(2004) Sensing and signaling during
plantflooding Plant Physiology and
Biochemistry 42 273-282
Dat J Folzer H Parent C Badot P-M Capelli N
(2006) HypoxiastressCurrent Understanding and
Perspectives In Teixeira da Silva JA (Ed)
Floriculture Ornamental and Plant
Biotechnology Advances and Topical Issues (Vol
3) Global Science Books Isleworth United
Kingdompp 664-674
Hatta Muhammad 2011 Dunia Pertanian
httpemhattawordpresscomcategorystress
Accessed September 10th 2013
Hopkins William G 2009 Introduction to Plant Physiology
New York John Wiley amp Sons Inc
Lakitan Benyamin 2011 Dasar-Dasar Fisiologi Tumbuhan
Jakarta PT Raja Grafindo Persada
Larcher W 1980 Physiological Plant Ecology New York
Springer-Verlag
Raven 2013 Biology Fifth Edition
httphigheredmcgraw-hillcomsites983409233
9student_view0chapter39
photosynthetic_electron_transport_and_atp_synth
esishtml Accessed September 10th 2013
Salisbury Frank B 1969 Plant Physiology Third Edition
California Wadsworth Publishing Company
Subbaiah C Sachs M (2003) Molecular and cellular
adaptations of maize to flooding stress Annals
of Botany 91 119-127
Wilson CarlL1962 Botany Third EditionUnited State of
America Holt Rinch Inc

CHAPTER II
PLANT RESPIRATION
A Krebs Cycle
The Krebs cycle was named in honor of the
English biochemist Hans A Krebs who in 1937
proposed a cycle of reactions to explain how pyruvate
breakdown takes place in the breast muscle of
pigeons He called his proposed pathway the citric
acid cycle because citric acid is an important
intermediet Another common name for the same group
of reactions is the Tricarboxuylic Acid (TCA) cycle
a term used because citric and isocitric acids have
three carboxyl groups It was not until the early
1950s that mitochondria capable of carrying out this
cycle were isolated from plant cells
The initial step leading to the Krebs cycle
involves the oxidation and loss of CO2 from pyruvate
and the combination of the remaining 2-carbon acetate
unit with a sulfur-containing compound co-enzyme A
(CoA) forming acetyl CoA This and another
comparable role of CoA in the Krebs cycle are
important reasons why sulfur is an essential element
4
The reaction of pyruvate decarboxylation also
involves a phosporylated form of thiamine (vitamin
B1) as a prostethic group Participation of thiamine
in this section partially explains the essential
function of vitamin B1 in plants and animals Besides
the loss of CO2 two hydrogen atoms are removed from
pyruvic acid during the formation of Acetyl CoA The
enzyme catalyzing the complete reaction is called
pyruvic acid dehydrogenase but it is actually an
organized complex containing numerous copies of three
or four different enzymes The hydrogen atoms removed
are finally accepted by NAD+ yielding NADH
The Krebs cycle accomplishes removal of some of
the electrons from organic acid intermediates and
transfer of these electrons to NAD+ or FAD Notice
that none of dehydrogenase enzyme of the cycle uses
NADP+ as an electron acceptor In fact NADP+ is
usually undetectable in plant mitochondria a
situation opposite to that of chloroplasts where
NADP+ is abundant but where there is much less NAD+
Not only are NADH and FADH2 important products of the
Krebs cycle but one molecule of ATP is formed from
ADP and Pi during the conversion of succinyl coenzyme
A to succinic acid Two additional CO2 molecules are
released in these Krebs cycle reactions so there is
a net loss of both carbon atoms from the incoming
5
acetate of acetyl CoA The release of CO2 in the
Krebs cycle accounts for the product of CO2 in the
summary equation for resspiration but no O2 is
absorbed during any Kreb cycle reaction (Lakitan
2011)
During metabolism the synthesis and breakdown
of different organic compounds takes place through
various pathways like the breakdown and synthesis of
proteins carbohydrates fats and nucleic acids
These different pathways and intermediates are also
responsible for the production of energyKrebrsquos
cycle named after Hans Krebs who began working out
its details in 1930s is a series of reactions in
which the pyruvate from glycolysis is oxidized to Co2
under aerobic conditions Kreb cycle is also known as
citric acid cycle or Tri-carboxylic acid cycle (TCA)
( Galston 1968 76)
6
Steps involved in the process of Kreb cycle are
1 Fate of pyruvic acid
Pyruvic acid can form different compounds by
different pathways ie it can be converted into
lactic acid It can be converted into acetyl
coenzyme A as a result of oxidation
2 Fate of Acetyl CoA
It can either undergo condensation with itself or
its derivatives to form fatty acids having 14 ndash 20
carbons Acetyl CoA can also go through a series
of reactions in Krebs cycle
3 Formation of citrate
Acetyl CoA enzyme condenses with oxaloacetate by
an enzyme citrate synthatase to form citrate with
the release of acetyl CoA If the amount of
7
oxaloacetate is very small then small number of
acetyl CoA would be reacting with oxaloacetate
leaving surplus acetyl CoA to go through another
pathway for the formation of long chain fatty
acids
4 Formation of C is ndash acotinate and iso-citrate
Citrate is changed first into C is ndash acotinate and
then to iso-citrate under the enzyme acotinase
Equilibrium is established between citrate C is
aconitate and iso-citrate It has been observed
that most of the time this equilibrium is shifted
towards the iso-citrate If the concentration of
iso-citrate in increased the formation of citrate
will result which also indicates that the
equilibrium also shift in the reserve direction
5 Formation of oxalosucinate
Iso-citrate is acted upon by an enzyme iso-citrate
dehydrogenase using nicotinamide adenine
dinucleotide (NAD) as coenzyme As a result iso-
citrate is converted into oxalosucciate and NAD is
reduced to NADH2 Similar reaction is carried out
by same enzyme using nicotindmide adenine
dinucleotide phosphate (NADP) as coenzyme which is
reduced to NDAPH2
6 Formation of α-Ketoglutarate
8
Oxalosucciante is changed into α-ketoglutrate by
iso-citrate dehydrogenase with the help of
coenzyme NAD or NADP In this reaction carbon
dioxide and NADH2 or NADPH2 are also released
7 Formation of succinyl CoA
α-ketoglutarate combines with acetyl CoA in the
presence of coenzyme NAD and enzyme α-
keoglutaratedehyrogenase to form succinyl coenzyme
a carbon dioxide and NADH2
8 Formation of Succinate
Later on coenzyme a is removed from succinyl
coenzyme A in the presence of guanosinediphosphate
(GDP) and inorganic phosphate to form succinate
and guanosine triphosphate (GTP) This reaction is
carried out by an enzyme called succinyl CoA
synthatase
9 Formation of Funarate
An enzyme succinic dehydrogenase removes hydrogen
from succinate to form funarate
10 Formation of Malate
Fumarate reacts with water in the presence of
enzyme fumarase to form malate
11 Regeneration of oxaloacetate
Malate is oxidized by malic dehydrogenase and NAD
forming oxaloacetate and NADH2 Thus oxalo acetate
is again available to start another cycle
9
(httpkrebs-cycle-or-citric-acid-cycle-or-
trihtml)
Importance of CTA Cycle
1 Source of energy
In addition to routine organic compounds described
above by products like nicotinanide adenine
dinucleotide (NADH2) and guanosine triphosphate
(GTP) are the source of biological energy NADH2
after oxidation produce energy whereas GTP is
itself high energy phosphate compound
2 Oxidation of organic compounds taken as food
Oxidation of fats carbohydrates and proteins take
place through it or in other words it can be said
that oxidation of all compounds having carbon
atoms can take place through TCA cycle Some of
amino acids like alanine glutamic acid and
aspartic acid at one stage or the other enters
into TCA cycle eg glutamic acid enters cycle
after its transformation into α-ketoglutarate
Similarly alanine enters the cycle after its
conversion into pyruvate
3 Intermediate compounds
TCA cycle is also involved in synthesis of
intermediate compounds leading to the formation of
larger molecules (Ray 1963 17)
10
Verification of Krebs Cycle
It was done by radioactive traces like C14 as
radioactive carbon dioxide of different levels and
reactions After addition of radioactive carbon
dioxide different chemical compounds produced like
glucose fats amino acids were isolated and looked
for radioactive carbon In this way whole of
metabolic reaction were verified including individual
reactions alternative metabolic pathways
intermediates of fats carbohydrates and amino acids
etc in the body cells as well as in test tubes
(Wilson 1962 99)
The primary function of the Krebs cycle are as
follows
1 Reduction of NAD+ and FAD to the electron donors
NADH and FADH2 that are subsequently oxidized to
yield ATP
2 Direct synthesis of a limited amount of ATP (1 ATP
for each pyruvate oxidized)
3 Formation of carbon skeletons that can be used to
synthesize certain amino acids that in turn are
converted into larger molecules
11
B Fermentation
Although glycolysis can function well without
O2 the further oxidation of pyruvate and NADH by
mitochondria requires this gas Thus when O2 is
limiting NADH and pyruvate begin to accumulate
Under this condition plants carry out fermentation
(anaerobic respiration) forming either ethanol or
lactic acid (usually ethanol) As shown in the
picture below the two top reactions consists of a
decarboxylation to form acetaldehide then rapid
reduction of acetaldehide by NADH to form ethanol
These reactions are catalyzed by alcohol
dehydrogenase Some cells contain lactic acid
12
dehydrogenase which uses NADH to reduce pyruvic to
lactic acid Thus either ethanol or lactic acid or
both are fermentation products depending on the
activities of each dehydrogenase present In each
case NADH is the reductant and only under
anaerobic conditions is it abundant anough to cause
reduction Furthermore in some plants NADH is used
to cause accumulation of the other compounds when O2
is limiting especially malate and glycerol
(Salisbury 1969 233)
The fate of pyruvate produced by glycolysis
depends primarily on whether or not oxygen is
present Under normal aerobic conditions pyruvate
is transported into the mitochondrion where it is
further oxidized to CO2 and water transferring its
electrons ultimately to molecular oxygen
13
Although higher plants are obligate aerobs and
are able to tolerate anoxia for only short periods
tissues or organs are occasionally subjected to
anaerobic conditions A typical siuation is that of
roots when the soil is saturated with water When
there is no oxygen to serve as the terminal electron
acceptor mitochondrial respiration will shut down
and metabolism will shift over to fermentation
Fermentation converts pyruvate to either ethanol
through the action of the enzyme alcohol
dehydrogenase (ADH) or lactate via lactate
dehydrogenase (LDH) In most plants the principal
products of fermentation are CO2 and ethanol Some
lactate may be formed primarily in the early stages
of anoxia However lactate lowers he pH of the
cytosol which in turn activates pyruvate
decarboxylase and initiates the production of
ethanol
Either one of the fermentation reactions
consumes the NADH produced earlier in glycolysis by
the oxidation of glyceraldehide-3-P Although this
means there is no net gain of reducing potential in
fermentation This recycling of NADH is still
important to the cell The poolof NADH plus NAD+ in
the cell is relatively small and if the NADH is not
recycled there will be no supply of NAD+ to support
14
the continued oxidation of glyceraldehyde-3-P if
this were the case glycolysis and the producton of
even the small quantities of ATP necessary to
maintain the cells under anaerobic conditions would
then grind to a halt (Hopkins 2009 241)
Soil waterlogging has long been identified as a
major abiotic stress and the constraints that it
provides the roots have a significant effect on the
growth and development of plants When this event
occurs in the spring then these puddles can reduce
seed germination and seedling development Thus a
pool of water is an important factor that affects
the growth development and survival of plant
species not only in natural ecosystems but also
the agricultural and horticultural systems (Dat et
al 2006)
After flooding rapid changes in soil
properties At the time of pore water meets the
land the air pushed out gas diffusion is reduced
and toxic compounds accumulate due to anaerobic
conditions All of these changes greatly affect the
ability of plants to survive In response increased
stomatal resistance photosynthesis and root
hydraulic conductivity decreases and reduced
translocation fotoassimilat Nevertheless one of
15
the best adaptations of plants to hypoxia anoxia
is transitional biochemical and metabolic processes
common to current limited availability of O2 (Dat et
al 2004)
Selective synthesis of a set of about 20
anaerobic stress proteins (ANPS) allows the process
of producing energy without oxygen metabolism under
anaerobic conditions (Subbaiah and Sachs 2003)
Another adaptation is the observed morphological
changes consisting of the formation of hypertrophic
lenticels adventitious root initiation and or
progression of aerenchyma (Vartapetian and Jackson
1997 Jackson and Colmer 2005 Folzer et al 2006)
This review details the diverse plant stress
response to hypoxia anoxia caused by soil
waterlogging flooding and examines some of the key
features of metabolic adaptation physiological and
morphological
16
C Electron
Transport System
We noted earlier that one of the principal
functions of the respiration is to retrieve in
useful form some of the energy initially stored in
assimilates Our traditional measure of useful
energy in most processes is the number of ATP
molecules gained or consumed By this measure alone
the yield from both glycolysis and the citric acid
cycle is quite low After two complete turns of the
cycle one molecule of glucose has been completely
oxidized to six molecules of CO2 but only four
molecules of ATP have been produced (a net of two
ATP from glycolysis plus one for each turn of the
cycle) At this point most of the energy associated
with the glucose molecule has been conserved in the
17
form of electron pairs generated by the oxidation of
glycolytic and citric acid cycle intermediates
In this section we will discuss the third stage
of cellular respirationmdashthe transfer of electrons
fromNADH and FADH2 to oxygen and the accompanying
conversion of redox energy to ATP The transfer of
electrons fromNADHand FADH2 to oxygen involves a
sequence of electron carriers arranged in an
electron transport chainMembrane fractionation
studies have shown that the enzymes and electron
carriers making up the electron transport chain are
organized predominantly into four large
multimolecular complexes (complexes ImdashIV) and two
mobile carriers located in the inner mitochondrial
membrane (Figure 109) In this sense there are a
great number of similarities between the
mitochondrial inner membrane and the thylakoid
membranes of the chloroplast (compare Figure 109
with Figure 76) This is not unexpected since the
principal function of each membrane is energy
transformation and many of the same or similar
components are involved
The Components of the Electron Transport Chain
The electron transport chain of the
mitochondria is the means by which electrons are
18
removed from the reduced carrier NADH and
transferred to oxygen to yield H2O
1) NADH
NADH is generated in the matrix by the reactions
of pyruvate dehydrogenase isocitrate
dehydrogenase α-ketoglutarate dehydrogenase and
malate dehyrogenase The electron transport chain
begins with reoxidizing NADH to form NAD+ and
channeling the electrons into the formation of
reduced coenzymes Important to note that NADH
transfers 2 electrons at a time in the form of a
hydride
NAD+ + 2e- + H+ 1048581 NADH Eorsquo = minus0315 V
2) Flavoproteins
Flavoproteins have either a FAD (flavin adenosine
dinucleotide) or a FMN (flavin mononucleotide)
prosthetic group Flavoproteins can accept or
donate electrons one at time or two at a time
Thus they are often intermediaries between two
electron acceptorsdonors and one electron
acceptorsdonors For flavoproteins the typical
standard reduction potentials are around 0 V
FAD + 2e- + 2H+ 1048581 FADH2 FMN + 2e- + 2H+ 1048581 FMNH2
Eorsquo asymp 0 V
19
3) Coenzyme Q (CoQ)
aka ubiquinone (UQ) Shown below CoQ has ten
repeating isoprene units which make it
insoluble in water but soluble in the
hydrophobic lipid bylayer Coenzyme Q is a
versatile cofactor because it is a soluble
electron carrier in the hydrophobic bilipid layer
of the
inner mitochondrial membrane Like flavoproteins
CoQ can acceptdonate electrons one at a time or
two at a time
Q + 2e- + 2H+ 1048581 QH2 Eorsquo =0060 V
Q + e- + H+ 1048581 1048581QH Eorsquo =0030 V
1048581QH + e- + H+ 1048581 QH2 Eorsquo =0190 V
4) Cytochromes
Cytochromes are proteins that contain heme
prosthetic groups which function as
one electron carriers The heme iron is involved
in one electron transfers involving the Fe2+ and
Fe3+ oxidation states Cytochromes are named by
their absorption spectra which depends
on the porphyrin structure and environment The
example shown is the heme prosthetic group of
cytochrome b Cytochrome b contains the same iron
porphyrin found in hemoglobin and myoglobin
20
Other cytochromes we will encounter in the
electron transport complexes are cytochromes b
c c1 a and a3
5) Iron-Sulfur Proteins
In the electron transport chain we will encounter
many iron-sulfer proteins which participate in
one electron transfers involving the the Fe2+ and
Fe3+ oxidation states These are non-heme iron-
sulfur proteinsThe simplest iron-sulfer protein
is FeS in which iron is tetrahedrally coordinated
by four cysteines The second form is Fe2S2 which
contains two irons complexed to 2 cysteine
residues and two inorganic sulfides The third
form is Fe3S4 which contains 3 iron atoms
coordinated to three cysteine residues and 4
inorganic sulfides The last form is the most
complicated Fe4S4 which contains 4 iron atoms
coordinated to 4 cysteine residues and 4
inorganic sulfides
6) Copper Proteins
Copper bound proteins participate in one electron
transfers involving the Cu+ and Cu2+ oxidation
states
21
Overview of the Electron Transport Chain
Electrons move along the electron transport
chain going from donor to acceptor until they reach
oxygen the ultimate electron acceptor The standard
reduction potentials of the electron carriers are
between the NADHNAD+ couple (-0315 V) and the
oxygenH2O couple (0816 V) as on the next page The
components of the electron transport chain are
organized into 4 complexes Each complex contains
several different electron carriers
1 Complex I also known as the NADH-coenzyme Q
reductase or NADH dehydrogenase
2 Complex II also known as succinate-coenzyme Q
reductase or succinate dehydrogenase
3 Complex III also known as coenzyme Q reductase
4 Complex IV also known as cytochrome c reductase
The free energy necessary to generate ATP
is extracted from the oxidation of NADH and
FADH2 by the electron transport chain It
consists of four protein complexes and single
protein chain of cytochrome c through which
electrons pass from lower to higher standard
redox potentials Electrons are carried from complex
I and II to complex III by the membrane (lipid)
soluble coenzyme Q (CoQ or ubiquinone) and between
22
complexes III and IV by the peripheral membrane
protein cytochrome c
In the following I give the overall
reactions catalysed by corresponding protein
complexes in the mitochondrial electron transport
chain
1 Complex I
Complex I is also called NADH-Coenzyme Q
reductase because this large protein complex
transfers 2 electrons from NADH to coenzyme Q
Complex I was formerly known as NADH dehydrogenase
Complex I is huge 850000 kD and is composed of more
than thirty subunits It contains a FMN prosthetic
group and seven or more Fe-S clusters This complex
has between 20-26 iron atoms boundThe prosthetic
group FMN is absolutely required for activity
Therefore this complex is a flavoprotein This
complex binds NADH transfers two electrons in the
form of a hydride to FMN to produce NAD+ and FMNH2
23
The subsequent steps involve the transfer of
electrons one at a time to a series of iron-sulfer
complexes that includes both 2Fe-2S and 4Fe-4S
clusters Note the importance of FMN First it
functions as a 2 electron acceptor in the hydride
transfer from NADH Second it functions as a 1
electron donor to the series of iron sulfur clusters
FMN and FAD often play crucial links between 2
electron transfer agents and 1 electron transfer
agents The final step of this complex is the
transfer of 2 electrons one at a time to coenzyme Q
CoQ like FMN and FAD can function as a 2 electron
donoracceptor and as a 1 electron donoracceptor
CoQ is a mobile electron carrier because its
isoprenoid tail makes it highly hydrophobic and
lipophillic It diffuses freely in the bilipid layer
of the inner mitochondrial membrane
The process of transferring electrons from NADH
to CoQ by complex I results in the net transport of
protons from the matrix side of the inner
mitochondrial membrane to the inter membrane space
where the H+ ions accumulate generating a proton
motive force The intermembrane space side of the
inner membrane is referred to as the P face (P
standing for positive) The matrix side of the inner
membrane is referred to as the the N face The
24
transport of electrons from NADH to CoQ is coupled to
the transport of protons across the membrane This is
an example of active transport The stiochiometry is
4 H+ transported per 2 electrons
2 Complex II
Believe it or not you are already familiar
with Complex II It is none other than succinate
dehydrogenase The only enzyme of the citric acid
cycle that is an integral membrane protein This
complex is composed of four subunits 2 of which are
iron-sulfur proteins and the other two subunits
together bind FAD through a covalent link to a
histidine residue These two subunits are called
flavoprotein 2 or FP2 Complex II contains 3 Fe-S
centers 1 4Fe-4S cluster 1 3Fe-4S cluster and 1
2Fe- 2S cluster In the first step of this complex
succinate is bound and a hydride is transferred to
FAD to generate FADH2 and fumarate FADH2 then
transfers its electrons one at a time to the Fe-S
centers Thus once again FAD functions as 2 electron
acceptor and a 1 electron donor The final step of
this complex is the transfer of 2 electrons one at a
time to coenzyme Q to produce CoQH2 For complex II
the standard free energy change of the overall
reaction is too small to drive the transport of
protons across the inner mitochondrial membrane This
25
accounts for the 15 ATPrsquos generated per FADH2
compared with the 25 ATPrsquos generated per NADH
3 Complex III
This complex is also known as coenzyme Q-
cytochrome c reductase because it passes the
electrons form CoQH2 to cyt c through a very unique
electron transport pathway called the Q-cycle Shown
to the left are the porphyrins found in cytochromes
Cytochrome b contains the same iron protoporphyrin as
hemoglobin and myoglobin The c cytochromes contain
heme c through covalent attachment by cysteine
residues Cytochrome a is found in two forms in
complex IV In complex III we find two b-type
cytochromes and one c-type cytochrome Complex III is
complex and we have a crystal structure
4 Complex IV
Complex IV is also known as cytochrome c oxidase
because it accepts the electrons from cytochrome c
and directs them towards the four electron reduction
of O2 to form 2 molecules of H2O
4 cyt c (Fe2+) + 4 H+ + O2 4 cyt c (Fe3+) +
2H2O
26
Cytochrome c oxidase contains 2 heme
centerscytochrome a and cytochrome a3 and two copper
proteins Each of the protein bound coppers are
associated with one of the cytochromes The copper
sites are called CuA and CuB CuA is associated with
cytochrome a and is shown to the left CuB is
associated with cytochrome a3 The copper sites
function as 1 electron carriers cycling between the
cuprous state Cu+ and the cupric state Cu2+ Just
like iron containing proteins they transfer
electrons one at a time Cytochrome c is bound on the
P-face of the membrane and transfers its electron to
CuA The oxidized cytochrome c dissociates CuA then
transfers the electron to cytochrome a The protein
bound CuA and the iron bound in cytochrome a are 15 Aring
apart In contrast the CuB and the iron bound in
cytochrome a3 are very close to each other forming a
binuclear metal center shown below
27
Electron transport chains are associated with
membranes such as the mitochondrial membrane in
eukaryotic organisms the thylakoid membrane of
photosynthetic organisms and the plasma membrane of
prokaryotic organisms While some of the details
differ electron transport chains in all organisms
operate on the same basic principles
Excited electrons are brought to the electron
transport chain by electron carriers such as NADH
and FADH2 In photosynthetic organisms the electrons
are donated by water and excited by light energy
The electron donor is oxidized as the electrons are
passed to the first protein complex in the chain As
it accepts the electrons the protein complex is
reduced Next the electrons are passed from the
28
first protein complex which becomes oxidized to
the second protein complex which becomes reduced
Each of these redox reactions is slightly exergonic
or energy releasing Some of this energy is used to
move protons across the membrane against their
concentration gradient With each successive
transfer of electrons the original excited
electrons lose some of their energy After several
successive redox reactions the electrons are
donated to a terminal electron acceptor In aerobic
respiration the terminal electron acceptor is
oxygen In photosynthesis the terminal electron
acceptor is a compound called NADP
The energy in the excited electrons has been
transformed into an electrochemical gradient
represented by the high concentration of protons on
one side of the membrane These protons can diffuse
back across the membrane by passing through a
protein complex called ATP synthase As these
protons pass through ATP synthase down their
concentration gradient they provide the energy
needed to make ATP from ADP and inorganic phosphate
within the cell
29
CHAPTER III
CLOSING
A Conclution
When a hexose is completely oxidized to CO2
and H2O using these three processes Glycolysis
yields two ATP and two NADH per hexose used Each
such NADH oxidized by the electron transport system
30
yields two ATP as dscribed above so glycolysis
contributes a total of six ATP per hexose The Krebs
cycle contributes two ATP per hexose or per two
pyruvates when succinyl CoA is cleaved to succinate
and CoASH This cycle also produces eight NADH per
hexose within the mitochondrial matrix by oxidative
phosporylation each of these NADH yields three ATP
or 24 per hexose Each FADH2 from the Krebs cycle
yields two ATP by oxydative phosporylation or four
per hexose (two pyruvates) The total contribution of
the Krebs cycle is then 30 Adding these 30 to the 6
from glycolysis leads to a total of 36 per hexose
completely respired by these process
B Suggestion
My greatest wish is that the readers find
this manuscript helpful in gaining the knowledge they
need for effective learning and practice and in
preparing for examination designed to test that
knowledge Reader comments and suggestions are
welcome and would be very helpful in improving our
manuscript Your contribution may assist many other
students in the coming years
31
REFERENCE
Anonim 2013 Anaerobic Respiration
httpwwwscoolcoukgcsebiologyrespiration
revise-itanaerobic-respiration Accessed
September 10th 2013
Anonim 2013 What is the difference between aerobic
and anaerobic respiration in plants
httpwwwbiologylifeeasyorg4010difference
-between-aerobic-anaerobic-respiration-plants
Accessed September 10th 2013
Campbell Mary K 2006 Biochemistry New York Thomson
Brooks Cole Inc
Dat J Capelli N Folzer H Bourgeade P Badot P-M
(2004) Sensing and signaling during
plantflooding Plant Physiology and
Biochemistry 42 273-282
Dat J Folzer H Parent C Badot P-M Capelli N
(2006) HypoxiastressCurrent Understanding and
Perspectives In Teixeira da Silva JA (Ed)
Floriculture Ornamental and Plant
Biotechnology Advances and Topical Issues (Vol
3) Global Science Books Isleworth United
Kingdompp 664-674
Hatta Muhammad 2011 Dunia Pertanian
httpemhattawordpresscomcategorystress
Accessed September 10th 2013
Hopkins William G 2009 Introduction to Plant Physiology
New York John Wiley amp Sons Inc
Lakitan Benyamin 2011 Dasar-Dasar Fisiologi Tumbuhan
Jakarta PT Raja Grafindo Persada
Larcher W 1980 Physiological Plant Ecology New York
Springer-Verlag
Raven 2013 Biology Fifth Edition
httphigheredmcgraw-hillcomsites983409233
9student_view0chapter39
photosynthetic_electron_transport_and_atp_synth
esishtml Accessed September 10th 2013
Salisbury Frank B 1969 Plant Physiology Third Edition
California Wadsworth Publishing Company
Subbaiah C Sachs M (2003) Molecular and cellular
adaptations of maize to flooding stress Annals
of Botany 91 119-127
Wilson CarlL1962 Botany Third EditionUnited State of
America Holt Rinch Inc

The reaction of pyruvate decarboxylation also
involves a phosporylated form of thiamine (vitamin
B1) as a prostethic group Participation of thiamine
in this section partially explains the essential
function of vitamin B1 in plants and animals Besides
the loss of CO2 two hydrogen atoms are removed from
pyruvic acid during the formation of Acetyl CoA The
enzyme catalyzing the complete reaction is called
pyruvic acid dehydrogenase but it is actually an
organized complex containing numerous copies of three
or four different enzymes The hydrogen atoms removed
are finally accepted by NAD+ yielding NADH
The Krebs cycle accomplishes removal of some of
the electrons from organic acid intermediates and
transfer of these electrons to NAD+ or FAD Notice
that none of dehydrogenase enzyme of the cycle uses
NADP+ as an electron acceptor In fact NADP+ is
usually undetectable in plant mitochondria a
situation opposite to that of chloroplasts where
NADP+ is abundant but where there is much less NAD+
Not only are NADH and FADH2 important products of the
Krebs cycle but one molecule of ATP is formed from
ADP and Pi during the conversion of succinyl coenzyme
A to succinic acid Two additional CO2 molecules are
released in these Krebs cycle reactions so there is
a net loss of both carbon atoms from the incoming
5
acetate of acetyl CoA The release of CO2 in the
Krebs cycle accounts for the product of CO2 in the
summary equation for resspiration but no O2 is
absorbed during any Kreb cycle reaction (Lakitan
2011)
During metabolism the synthesis and breakdown
of different organic compounds takes place through
various pathways like the breakdown and synthesis of
proteins carbohydrates fats and nucleic acids
These different pathways and intermediates are also
responsible for the production of energyKrebrsquos
cycle named after Hans Krebs who began working out
its details in 1930s is a series of reactions in
which the pyruvate from glycolysis is oxidized to Co2
under aerobic conditions Kreb cycle is also known as
citric acid cycle or Tri-carboxylic acid cycle (TCA)
( Galston 1968 76)
6
Steps involved in the process of Kreb cycle are
1 Fate of pyruvic acid
Pyruvic acid can form different compounds by
different pathways ie it can be converted into
lactic acid It can be converted into acetyl
coenzyme A as a result of oxidation
2 Fate of Acetyl CoA
It can either undergo condensation with itself or
its derivatives to form fatty acids having 14 ndash 20
carbons Acetyl CoA can also go through a series
of reactions in Krebs cycle
3 Formation of citrate
Acetyl CoA enzyme condenses with oxaloacetate by
an enzyme citrate synthatase to form citrate with
the release of acetyl CoA If the amount of
7
oxaloacetate is very small then small number of
acetyl CoA would be reacting with oxaloacetate
leaving surplus acetyl CoA to go through another
pathway for the formation of long chain fatty
acids
4 Formation of C is ndash acotinate and iso-citrate
Citrate is changed first into C is ndash acotinate and
then to iso-citrate under the enzyme acotinase
Equilibrium is established between citrate C is
aconitate and iso-citrate It has been observed
that most of the time this equilibrium is shifted
towards the iso-citrate If the concentration of
iso-citrate in increased the formation of citrate
will result which also indicates that the
equilibrium also shift in the reserve direction
5 Formation of oxalosucinate
Iso-citrate is acted upon by an enzyme iso-citrate
dehydrogenase using nicotinamide adenine
dinucleotide (NAD) as coenzyme As a result iso-
citrate is converted into oxalosucciate and NAD is
reduced to NADH2 Similar reaction is carried out
by same enzyme using nicotindmide adenine
dinucleotide phosphate (NADP) as coenzyme which is
reduced to NDAPH2
6 Formation of α-Ketoglutarate
8
Oxalosucciante is changed into α-ketoglutrate by
iso-citrate dehydrogenase with the help of
coenzyme NAD or NADP In this reaction carbon
dioxide and NADH2 or NADPH2 are also released
7 Formation of succinyl CoA
α-ketoglutarate combines with acetyl CoA in the
presence of coenzyme NAD and enzyme α-
keoglutaratedehyrogenase to form succinyl coenzyme
a carbon dioxide and NADH2
8 Formation of Succinate
Later on coenzyme a is removed from succinyl
coenzyme A in the presence of guanosinediphosphate
(GDP) and inorganic phosphate to form succinate
and guanosine triphosphate (GTP) This reaction is
carried out by an enzyme called succinyl CoA
synthatase
9 Formation of Funarate
An enzyme succinic dehydrogenase removes hydrogen
from succinate to form funarate
10 Formation of Malate
Fumarate reacts with water in the presence of
enzyme fumarase to form malate
11 Regeneration of oxaloacetate
Malate is oxidized by malic dehydrogenase and NAD
forming oxaloacetate and NADH2 Thus oxalo acetate
is again available to start another cycle
9
(httpkrebs-cycle-or-citric-acid-cycle-or-
trihtml)
Importance of CTA Cycle
1 Source of energy
In addition to routine organic compounds described
above by products like nicotinanide adenine
dinucleotide (NADH2) and guanosine triphosphate
(GTP) are the source of biological energy NADH2
after oxidation produce energy whereas GTP is
itself high energy phosphate compound
2 Oxidation of organic compounds taken as food
Oxidation of fats carbohydrates and proteins take
place through it or in other words it can be said
that oxidation of all compounds having carbon
atoms can take place through TCA cycle Some of
amino acids like alanine glutamic acid and
aspartic acid at one stage or the other enters
into TCA cycle eg glutamic acid enters cycle
after its transformation into α-ketoglutarate
Similarly alanine enters the cycle after its
conversion into pyruvate
3 Intermediate compounds
TCA cycle is also involved in synthesis of
intermediate compounds leading to the formation of
larger molecules (Ray 1963 17)
10
Verification of Krebs Cycle
It was done by radioactive traces like C14 as
radioactive carbon dioxide of different levels and
reactions After addition of radioactive carbon
dioxide different chemical compounds produced like
glucose fats amino acids were isolated and looked
for radioactive carbon In this way whole of
metabolic reaction were verified including individual
reactions alternative metabolic pathways
intermediates of fats carbohydrates and amino acids
etc in the body cells as well as in test tubes
(Wilson 1962 99)
The primary function of the Krebs cycle are as
follows
1 Reduction of NAD+ and FAD to the electron donors
NADH and FADH2 that are subsequently oxidized to
yield ATP
2 Direct synthesis of a limited amount of ATP (1 ATP
for each pyruvate oxidized)
3 Formation of carbon skeletons that can be used to
synthesize certain amino acids that in turn are
converted into larger molecules
11
B Fermentation
Although glycolysis can function well without
O2 the further oxidation of pyruvate and NADH by
mitochondria requires this gas Thus when O2 is
limiting NADH and pyruvate begin to accumulate
Under this condition plants carry out fermentation
(anaerobic respiration) forming either ethanol or
lactic acid (usually ethanol) As shown in the
picture below the two top reactions consists of a
decarboxylation to form acetaldehide then rapid
reduction of acetaldehide by NADH to form ethanol
These reactions are catalyzed by alcohol
dehydrogenase Some cells contain lactic acid
12
dehydrogenase which uses NADH to reduce pyruvic to
lactic acid Thus either ethanol or lactic acid or
both are fermentation products depending on the
activities of each dehydrogenase present In each
case NADH is the reductant and only under
anaerobic conditions is it abundant anough to cause
reduction Furthermore in some plants NADH is used
to cause accumulation of the other compounds when O2
is limiting especially malate and glycerol
(Salisbury 1969 233)
The fate of pyruvate produced by glycolysis
depends primarily on whether or not oxygen is
present Under normal aerobic conditions pyruvate
is transported into the mitochondrion where it is
further oxidized to CO2 and water transferring its
electrons ultimately to molecular oxygen
13
Although higher plants are obligate aerobs and
are able to tolerate anoxia for only short periods
tissues or organs are occasionally subjected to
anaerobic conditions A typical siuation is that of
roots when the soil is saturated with water When
there is no oxygen to serve as the terminal electron
acceptor mitochondrial respiration will shut down
and metabolism will shift over to fermentation
Fermentation converts pyruvate to either ethanol
through the action of the enzyme alcohol
dehydrogenase (ADH) or lactate via lactate
dehydrogenase (LDH) In most plants the principal
products of fermentation are CO2 and ethanol Some
lactate may be formed primarily in the early stages
of anoxia However lactate lowers he pH of the
cytosol which in turn activates pyruvate
decarboxylase and initiates the production of
ethanol
Either one of the fermentation reactions
consumes the NADH produced earlier in glycolysis by
the oxidation of glyceraldehide-3-P Although this
means there is no net gain of reducing potential in
fermentation This recycling of NADH is still
important to the cell The poolof NADH plus NAD+ in
the cell is relatively small and if the NADH is not
recycled there will be no supply of NAD+ to support
14
the continued oxidation of glyceraldehyde-3-P if
this were the case glycolysis and the producton of
even the small quantities of ATP necessary to
maintain the cells under anaerobic conditions would
then grind to a halt (Hopkins 2009 241)
Soil waterlogging has long been identified as a
major abiotic stress and the constraints that it
provides the roots have a significant effect on the
growth and development of plants When this event
occurs in the spring then these puddles can reduce
seed germination and seedling development Thus a
pool of water is an important factor that affects
the growth development and survival of plant
species not only in natural ecosystems but also
the agricultural and horticultural systems (Dat et
al 2006)
After flooding rapid changes in soil
properties At the time of pore water meets the
land the air pushed out gas diffusion is reduced
and toxic compounds accumulate due to anaerobic
conditions All of these changes greatly affect the
ability of plants to survive In response increased
stomatal resistance photosynthesis and root
hydraulic conductivity decreases and reduced
translocation fotoassimilat Nevertheless one of
15
the best adaptations of plants to hypoxia anoxia
is transitional biochemical and metabolic processes
common to current limited availability of O2 (Dat et
al 2004)
Selective synthesis of a set of about 20
anaerobic stress proteins (ANPS) allows the process
of producing energy without oxygen metabolism under
anaerobic conditions (Subbaiah and Sachs 2003)
Another adaptation is the observed morphological
changes consisting of the formation of hypertrophic
lenticels adventitious root initiation and or
progression of aerenchyma (Vartapetian and Jackson
1997 Jackson and Colmer 2005 Folzer et al 2006)
This review details the diverse plant stress
response to hypoxia anoxia caused by soil
waterlogging flooding and examines some of the key
features of metabolic adaptation physiological and
morphological
16
C Electron
Transport System
We noted earlier that one of the principal
functions of the respiration is to retrieve in
useful form some of the energy initially stored in
assimilates Our traditional measure of useful
energy in most processes is the number of ATP
molecules gained or consumed By this measure alone
the yield from both glycolysis and the citric acid
cycle is quite low After two complete turns of the
cycle one molecule of glucose has been completely
oxidized to six molecules of CO2 but only four
molecules of ATP have been produced (a net of two
ATP from glycolysis plus one for each turn of the
cycle) At this point most of the energy associated
with the glucose molecule has been conserved in the
17
form of electron pairs generated by the oxidation of
glycolytic and citric acid cycle intermediates
In this section we will discuss the third stage
of cellular respirationmdashthe transfer of electrons
fromNADH and FADH2 to oxygen and the accompanying
conversion of redox energy to ATP The transfer of
electrons fromNADHand FADH2 to oxygen involves a
sequence of electron carriers arranged in an
electron transport chainMembrane fractionation
studies have shown that the enzymes and electron
carriers making up the electron transport chain are
organized predominantly into four large
multimolecular complexes (complexes ImdashIV) and two
mobile carriers located in the inner mitochondrial
membrane (Figure 109) In this sense there are a
great number of similarities between the
mitochondrial inner membrane and the thylakoid
membranes of the chloroplast (compare Figure 109
with Figure 76) This is not unexpected since the
principal function of each membrane is energy
transformation and many of the same or similar
components are involved
The Components of the Electron Transport Chain
The electron transport chain of the
mitochondria is the means by which electrons are
18
removed from the reduced carrier NADH and
transferred to oxygen to yield H2O
1) NADH
NADH is generated in the matrix by the reactions
of pyruvate dehydrogenase isocitrate
dehydrogenase α-ketoglutarate dehydrogenase and
malate dehyrogenase The electron transport chain
begins with reoxidizing NADH to form NAD+ and
channeling the electrons into the formation of
reduced coenzymes Important to note that NADH
transfers 2 electrons at a time in the form of a
hydride
NAD+ + 2e- + H+ 1048581 NADH Eorsquo = minus0315 V
2) Flavoproteins
Flavoproteins have either a FAD (flavin adenosine
dinucleotide) or a FMN (flavin mononucleotide)
prosthetic group Flavoproteins can accept or
donate electrons one at time or two at a time
Thus they are often intermediaries between two
electron acceptorsdonors and one electron
acceptorsdonors For flavoproteins the typical
standard reduction potentials are around 0 V
FAD + 2e- + 2H+ 1048581 FADH2 FMN + 2e- + 2H+ 1048581 FMNH2
Eorsquo asymp 0 V
19
3) Coenzyme Q (CoQ)
aka ubiquinone (UQ) Shown below CoQ has ten
repeating isoprene units which make it
insoluble in water but soluble in the
hydrophobic lipid bylayer Coenzyme Q is a
versatile cofactor because it is a soluble
electron carrier in the hydrophobic bilipid layer
of the
inner mitochondrial membrane Like flavoproteins
CoQ can acceptdonate electrons one at a time or
two at a time
Q + 2e- + 2H+ 1048581 QH2 Eorsquo =0060 V
Q + e- + H+ 1048581 1048581QH Eorsquo =0030 V
1048581QH + e- + H+ 1048581 QH2 Eorsquo =0190 V
4) Cytochromes
Cytochromes are proteins that contain heme
prosthetic groups which function as
one electron carriers The heme iron is involved
in one electron transfers involving the Fe2+ and
Fe3+ oxidation states Cytochromes are named by
their absorption spectra which depends
on the porphyrin structure and environment The
example shown is the heme prosthetic group of
cytochrome b Cytochrome b contains the same iron
porphyrin found in hemoglobin and myoglobin
20
Other cytochromes we will encounter in the
electron transport complexes are cytochromes b
c c1 a and a3
5) Iron-Sulfur Proteins
In the electron transport chain we will encounter
many iron-sulfer proteins which participate in
one electron transfers involving the the Fe2+ and
Fe3+ oxidation states These are non-heme iron-
sulfur proteinsThe simplest iron-sulfer protein
is FeS in which iron is tetrahedrally coordinated
by four cysteines The second form is Fe2S2 which
contains two irons complexed to 2 cysteine
residues and two inorganic sulfides The third
form is Fe3S4 which contains 3 iron atoms
coordinated to three cysteine residues and 4
inorganic sulfides The last form is the most
complicated Fe4S4 which contains 4 iron atoms
coordinated to 4 cysteine residues and 4
inorganic sulfides
6) Copper Proteins
Copper bound proteins participate in one electron
transfers involving the Cu+ and Cu2+ oxidation
states
21
Overview of the Electron Transport Chain
Electrons move along the electron transport
chain going from donor to acceptor until they reach
oxygen the ultimate electron acceptor The standard
reduction potentials of the electron carriers are
between the NADHNAD+ couple (-0315 V) and the
oxygenH2O couple (0816 V) as on the next page The
components of the electron transport chain are
organized into 4 complexes Each complex contains
several different electron carriers
1 Complex I also known as the NADH-coenzyme Q
reductase or NADH dehydrogenase
2 Complex II also known as succinate-coenzyme Q
reductase or succinate dehydrogenase
3 Complex III also known as coenzyme Q reductase
4 Complex IV also known as cytochrome c reductase
The free energy necessary to generate ATP
is extracted from the oxidation of NADH and
FADH2 by the electron transport chain It
consists of four protein complexes and single
protein chain of cytochrome c through which
electrons pass from lower to higher standard
redox potentials Electrons are carried from complex
I and II to complex III by the membrane (lipid)
soluble coenzyme Q (CoQ or ubiquinone) and between
22
complexes III and IV by the peripheral membrane
protein cytochrome c
In the following I give the overall
reactions catalysed by corresponding protein
complexes in the mitochondrial electron transport
chain
1 Complex I
Complex I is also called NADH-Coenzyme Q
reductase because this large protein complex
transfers 2 electrons from NADH to coenzyme Q
Complex I was formerly known as NADH dehydrogenase
Complex I is huge 850000 kD and is composed of more
than thirty subunits It contains a FMN prosthetic
group and seven or more Fe-S clusters This complex
has between 20-26 iron atoms boundThe prosthetic
group FMN is absolutely required for activity
Therefore this complex is a flavoprotein This
complex binds NADH transfers two electrons in the
form of a hydride to FMN to produce NAD+ and FMNH2
23
The subsequent steps involve the transfer of
electrons one at a time to a series of iron-sulfer
complexes that includes both 2Fe-2S and 4Fe-4S
clusters Note the importance of FMN First it
functions as a 2 electron acceptor in the hydride
transfer from NADH Second it functions as a 1
electron donor to the series of iron sulfur clusters
FMN and FAD often play crucial links between 2
electron transfer agents and 1 electron transfer
agents The final step of this complex is the
transfer of 2 electrons one at a time to coenzyme Q
CoQ like FMN and FAD can function as a 2 electron
donoracceptor and as a 1 electron donoracceptor
CoQ is a mobile electron carrier because its
isoprenoid tail makes it highly hydrophobic and
lipophillic It diffuses freely in the bilipid layer
of the inner mitochondrial membrane
The process of transferring electrons from NADH
to CoQ by complex I results in the net transport of
protons from the matrix side of the inner
mitochondrial membrane to the inter membrane space
where the H+ ions accumulate generating a proton
motive force The intermembrane space side of the
inner membrane is referred to as the P face (P
standing for positive) The matrix side of the inner
membrane is referred to as the the N face The
24
transport of electrons from NADH to CoQ is coupled to
the transport of protons across the membrane This is
an example of active transport The stiochiometry is
4 H+ transported per 2 electrons
2 Complex II
Believe it or not you are already familiar
with Complex II It is none other than succinate
dehydrogenase The only enzyme of the citric acid
cycle that is an integral membrane protein This
complex is composed of four subunits 2 of which are
iron-sulfur proteins and the other two subunits
together bind FAD through a covalent link to a
histidine residue These two subunits are called
flavoprotein 2 or FP2 Complex II contains 3 Fe-S
centers 1 4Fe-4S cluster 1 3Fe-4S cluster and 1
2Fe- 2S cluster In the first step of this complex
succinate is bound and a hydride is transferred to
FAD to generate FADH2 and fumarate FADH2 then
transfers its electrons one at a time to the Fe-S
centers Thus once again FAD functions as 2 electron
acceptor and a 1 electron donor The final step of
this complex is the transfer of 2 electrons one at a
time to coenzyme Q to produce CoQH2 For complex II
the standard free energy change of the overall
reaction is too small to drive the transport of
protons across the inner mitochondrial membrane This
25
accounts for the 15 ATPrsquos generated per FADH2
compared with the 25 ATPrsquos generated per NADH
3 Complex III
This complex is also known as coenzyme Q-
cytochrome c reductase because it passes the
electrons form CoQH2 to cyt c through a very unique
electron transport pathway called the Q-cycle Shown
to the left are the porphyrins found in cytochromes
Cytochrome b contains the same iron protoporphyrin as
hemoglobin and myoglobin The c cytochromes contain
heme c through covalent attachment by cysteine
residues Cytochrome a is found in two forms in
complex IV In complex III we find two b-type
cytochromes and one c-type cytochrome Complex III is
complex and we have a crystal structure
4 Complex IV
Complex IV is also known as cytochrome c oxidase
because it accepts the electrons from cytochrome c
and directs them towards the four electron reduction
of O2 to form 2 molecules of H2O
4 cyt c (Fe2+) + 4 H+ + O2 4 cyt c (Fe3+) +
2H2O
26
Cytochrome c oxidase contains 2 heme
centerscytochrome a and cytochrome a3 and two copper
proteins Each of the protein bound coppers are
associated with one of the cytochromes The copper
sites are called CuA and CuB CuA is associated with
cytochrome a and is shown to the left CuB is
associated with cytochrome a3 The copper sites
function as 1 electron carriers cycling between the
cuprous state Cu+ and the cupric state Cu2+ Just
like iron containing proteins they transfer
electrons one at a time Cytochrome c is bound on the
P-face of the membrane and transfers its electron to
CuA The oxidized cytochrome c dissociates CuA then
transfers the electron to cytochrome a The protein
bound CuA and the iron bound in cytochrome a are 15 Aring
apart In contrast the CuB and the iron bound in
cytochrome a3 are very close to each other forming a
binuclear metal center shown below
27
Electron transport chains are associated with
membranes such as the mitochondrial membrane in
eukaryotic organisms the thylakoid membrane of
photosynthetic organisms and the plasma membrane of
prokaryotic organisms While some of the details
differ electron transport chains in all organisms
operate on the same basic principles
Excited electrons are brought to the electron
transport chain by electron carriers such as NADH
and FADH2 In photosynthetic organisms the electrons
are donated by water and excited by light energy
The electron donor is oxidized as the electrons are
passed to the first protein complex in the chain As
it accepts the electrons the protein complex is
reduced Next the electrons are passed from the
28
first protein complex which becomes oxidized to
the second protein complex which becomes reduced
Each of these redox reactions is slightly exergonic
or energy releasing Some of this energy is used to
move protons across the membrane against their
concentration gradient With each successive
transfer of electrons the original excited
electrons lose some of their energy After several
successive redox reactions the electrons are
donated to a terminal electron acceptor In aerobic
respiration the terminal electron acceptor is
oxygen In photosynthesis the terminal electron
acceptor is a compound called NADP
The energy in the excited electrons has been
transformed into an electrochemical gradient
represented by the high concentration of protons on
one side of the membrane These protons can diffuse
back across the membrane by passing through a
protein complex called ATP synthase As these
protons pass through ATP synthase down their
concentration gradient they provide the energy
needed to make ATP from ADP and inorganic phosphate
within the cell
29
CHAPTER III
CLOSING
A Conclution
When a hexose is completely oxidized to CO2
and H2O using these three processes Glycolysis
yields two ATP and two NADH per hexose used Each
such NADH oxidized by the electron transport system
30
yields two ATP as dscribed above so glycolysis
contributes a total of six ATP per hexose The Krebs
cycle contributes two ATP per hexose or per two
pyruvates when succinyl CoA is cleaved to succinate
and CoASH This cycle also produces eight NADH per
hexose within the mitochondrial matrix by oxidative
phosporylation each of these NADH yields three ATP
or 24 per hexose Each FADH2 from the Krebs cycle
yields two ATP by oxydative phosporylation or four
per hexose (two pyruvates) The total contribution of
the Krebs cycle is then 30 Adding these 30 to the 6
from glycolysis leads to a total of 36 per hexose
completely respired by these process
B Suggestion
My greatest wish is that the readers find
this manuscript helpful in gaining the knowledge they
need for effective learning and practice and in
preparing for examination designed to test that
knowledge Reader comments and suggestions are
welcome and would be very helpful in improving our
manuscript Your contribution may assist many other
students in the coming years
31
REFERENCE
Anonim 2013 Anaerobic Respiration
httpwwwscoolcoukgcsebiologyrespiration
revise-itanaerobic-respiration Accessed
September 10th 2013
Anonim 2013 What is the difference between aerobic
and anaerobic respiration in plants
httpwwwbiologylifeeasyorg4010difference
-between-aerobic-anaerobic-respiration-plants
Accessed September 10th 2013
Campbell Mary K 2006 Biochemistry New York Thomson
Brooks Cole Inc
Dat J Capelli N Folzer H Bourgeade P Badot P-M
(2004) Sensing and signaling during
plantflooding Plant Physiology and
Biochemistry 42 273-282
Dat J Folzer H Parent C Badot P-M Capelli N
(2006) HypoxiastressCurrent Understanding and
Perspectives In Teixeira da Silva JA (Ed)
Floriculture Ornamental and Plant
Biotechnology Advances and Topical Issues (Vol
3) Global Science Books Isleworth United
Kingdompp 664-674
Hatta Muhammad 2011 Dunia Pertanian
httpemhattawordpresscomcategorystress
Accessed September 10th 2013
Hopkins William G 2009 Introduction to Plant Physiology
New York John Wiley amp Sons Inc
Lakitan Benyamin 2011 Dasar-Dasar Fisiologi Tumbuhan
Jakarta PT Raja Grafindo Persada
Larcher W 1980 Physiological Plant Ecology New York
Springer-Verlag
Raven 2013 Biology Fifth Edition
httphigheredmcgraw-hillcomsites983409233
9student_view0chapter39
photosynthetic_electron_transport_and_atp_synth
esishtml Accessed September 10th 2013
Salisbury Frank B 1969 Plant Physiology Third Edition
California Wadsworth Publishing Company
Subbaiah C Sachs M (2003) Molecular and cellular
adaptations of maize to flooding stress Annals
of Botany 91 119-127
Wilson CarlL1962 Botany Third EditionUnited State of
America Holt Rinch Inc

acetate of acetyl CoA The release of CO2 in the
Krebs cycle accounts for the product of CO2 in the
summary equation for resspiration but no O2 is
absorbed during any Kreb cycle reaction (Lakitan
2011)
During metabolism the synthesis and breakdown
of different organic compounds takes place through
various pathways like the breakdown and synthesis of
proteins carbohydrates fats and nucleic acids
These different pathways and intermediates are also
responsible for the production of energyKrebrsquos
cycle named after Hans Krebs who began working out
its details in 1930s is a series of reactions in
which the pyruvate from glycolysis is oxidized to Co2
under aerobic conditions Kreb cycle is also known as
citric acid cycle or Tri-carboxylic acid cycle (TCA)
( Galston 1968 76)
6
Steps involved in the process of Kreb cycle are
1 Fate of pyruvic acid
Pyruvic acid can form different compounds by
different pathways ie it can be converted into
lactic acid It can be converted into acetyl
coenzyme A as a result of oxidation
2 Fate of Acetyl CoA
It can either undergo condensation with itself or
its derivatives to form fatty acids having 14 ndash 20
carbons Acetyl CoA can also go through a series
of reactions in Krebs cycle
3 Formation of citrate
Acetyl CoA enzyme condenses with oxaloacetate by
an enzyme citrate synthatase to form citrate with
the release of acetyl CoA If the amount of
7
oxaloacetate is very small then small number of
acetyl CoA would be reacting with oxaloacetate
leaving surplus acetyl CoA to go through another
pathway for the formation of long chain fatty
acids
4 Formation of C is ndash acotinate and iso-citrate
Citrate is changed first into C is ndash acotinate and
then to iso-citrate under the enzyme acotinase
Equilibrium is established between citrate C is
aconitate and iso-citrate It has been observed
that most of the time this equilibrium is shifted
towards the iso-citrate If the concentration of
iso-citrate in increased the formation of citrate
will result which also indicates that the
equilibrium also shift in the reserve direction
5 Formation of oxalosucinate
Iso-citrate is acted upon by an enzyme iso-citrate
dehydrogenase using nicotinamide adenine
dinucleotide (NAD) as coenzyme As a result iso-
citrate is converted into oxalosucciate and NAD is
reduced to NADH2 Similar reaction is carried out
by same enzyme using nicotindmide adenine
dinucleotide phosphate (NADP) as coenzyme which is
reduced to NDAPH2
6 Formation of α-Ketoglutarate
8
Oxalosucciante is changed into α-ketoglutrate by
iso-citrate dehydrogenase with the help of
coenzyme NAD or NADP In this reaction carbon
dioxide and NADH2 or NADPH2 are also released
7 Formation of succinyl CoA
α-ketoglutarate combines with acetyl CoA in the
presence of coenzyme NAD and enzyme α-
keoglutaratedehyrogenase to form succinyl coenzyme
a carbon dioxide and NADH2
8 Formation of Succinate
Later on coenzyme a is removed from succinyl
coenzyme A in the presence of guanosinediphosphate
(GDP) and inorganic phosphate to form succinate
and guanosine triphosphate (GTP) This reaction is
carried out by an enzyme called succinyl CoA
synthatase
9 Formation of Funarate
An enzyme succinic dehydrogenase removes hydrogen
from succinate to form funarate
10 Formation of Malate
Fumarate reacts with water in the presence of
enzyme fumarase to form malate
11 Regeneration of oxaloacetate
Malate is oxidized by malic dehydrogenase and NAD
forming oxaloacetate and NADH2 Thus oxalo acetate
is again available to start another cycle
9
(httpkrebs-cycle-or-citric-acid-cycle-or-
trihtml)
Importance of CTA Cycle
1 Source of energy
In addition to routine organic compounds described
above by products like nicotinanide adenine
dinucleotide (NADH2) and guanosine triphosphate
(GTP) are the source of biological energy NADH2
after oxidation produce energy whereas GTP is
itself high energy phosphate compound
2 Oxidation of organic compounds taken as food
Oxidation of fats carbohydrates and proteins take
place through it or in other words it can be said
that oxidation of all compounds having carbon
atoms can take place through TCA cycle Some of
amino acids like alanine glutamic acid and
aspartic acid at one stage or the other enters
into TCA cycle eg glutamic acid enters cycle
after its transformation into α-ketoglutarate
Similarly alanine enters the cycle after its
conversion into pyruvate
3 Intermediate compounds
TCA cycle is also involved in synthesis of
intermediate compounds leading to the formation of
larger molecules (Ray 1963 17)
10
Verification of Krebs Cycle
It was done by radioactive traces like C14 as
radioactive carbon dioxide of different levels and
reactions After addition of radioactive carbon
dioxide different chemical compounds produced like
glucose fats amino acids were isolated and looked
for radioactive carbon In this way whole of
metabolic reaction were verified including individual
reactions alternative metabolic pathways
intermediates of fats carbohydrates and amino acids
etc in the body cells as well as in test tubes
(Wilson 1962 99)
The primary function of the Krebs cycle are as
follows
1 Reduction of NAD+ and FAD to the electron donors
NADH and FADH2 that are subsequently oxidized to
yield ATP
2 Direct synthesis of a limited amount of ATP (1 ATP
for each pyruvate oxidized)
3 Formation of carbon skeletons that can be used to
synthesize certain amino acids that in turn are
converted into larger molecules
11
B Fermentation
Although glycolysis can function well without
O2 the further oxidation of pyruvate and NADH by
mitochondria requires this gas Thus when O2 is
limiting NADH and pyruvate begin to accumulate
Under this condition plants carry out fermentation
(anaerobic respiration) forming either ethanol or
lactic acid (usually ethanol) As shown in the
picture below the two top reactions consists of a
decarboxylation to form acetaldehide then rapid
reduction of acetaldehide by NADH to form ethanol
These reactions are catalyzed by alcohol
dehydrogenase Some cells contain lactic acid
12
dehydrogenase which uses NADH to reduce pyruvic to
lactic acid Thus either ethanol or lactic acid or
both are fermentation products depending on the
activities of each dehydrogenase present In each
case NADH is the reductant and only under
anaerobic conditions is it abundant anough to cause
reduction Furthermore in some plants NADH is used
to cause accumulation of the other compounds when O2
is limiting especially malate and glycerol
(Salisbury 1969 233)
The fate of pyruvate produced by glycolysis
depends primarily on whether or not oxygen is
present Under normal aerobic conditions pyruvate
is transported into the mitochondrion where it is
further oxidized to CO2 and water transferring its
electrons ultimately to molecular oxygen
13
Although higher plants are obligate aerobs and
are able to tolerate anoxia for only short periods
tissues or organs are occasionally subjected to
anaerobic conditions A typical siuation is that of
roots when the soil is saturated with water When
there is no oxygen to serve as the terminal electron
acceptor mitochondrial respiration will shut down
and metabolism will shift over to fermentation
Fermentation converts pyruvate to either ethanol
through the action of the enzyme alcohol
dehydrogenase (ADH) or lactate via lactate
dehydrogenase (LDH) In most plants the principal
products of fermentation are CO2 and ethanol Some
lactate may be formed primarily in the early stages
of anoxia However lactate lowers he pH of the
cytosol which in turn activates pyruvate
decarboxylase and initiates the production of
ethanol
Either one of the fermentation reactions
consumes the NADH produced earlier in glycolysis by
the oxidation of glyceraldehide-3-P Although this
means there is no net gain of reducing potential in
fermentation This recycling of NADH is still
important to the cell The poolof NADH plus NAD+ in
the cell is relatively small and if the NADH is not
recycled there will be no supply of NAD+ to support
14
the continued oxidation of glyceraldehyde-3-P if
this were the case glycolysis and the producton of
even the small quantities of ATP necessary to
maintain the cells under anaerobic conditions would
then grind to a halt (Hopkins 2009 241)
Soil waterlogging has long been identified as a
major abiotic stress and the constraints that it
provides the roots have a significant effect on the
growth and development of plants When this event
occurs in the spring then these puddles can reduce
seed germination and seedling development Thus a
pool of water is an important factor that affects
the growth development and survival of plant
species not only in natural ecosystems but also
the agricultural and horticultural systems (Dat et
al 2006)
After flooding rapid changes in soil
properties At the time of pore water meets the
land the air pushed out gas diffusion is reduced
and toxic compounds accumulate due to anaerobic
conditions All of these changes greatly affect the
ability of plants to survive In response increased
stomatal resistance photosynthesis and root
hydraulic conductivity decreases and reduced
translocation fotoassimilat Nevertheless one of
15
the best adaptations of plants to hypoxia anoxia
is transitional biochemical and metabolic processes
common to current limited availability of O2 (Dat et
al 2004)
Selective synthesis of a set of about 20
anaerobic stress proteins (ANPS) allows the process
of producing energy without oxygen metabolism under
anaerobic conditions (Subbaiah and Sachs 2003)
Another adaptation is the observed morphological
changes consisting of the formation of hypertrophic
lenticels adventitious root initiation and or
progression of aerenchyma (Vartapetian and Jackson
1997 Jackson and Colmer 2005 Folzer et al 2006)
This review details the diverse plant stress
response to hypoxia anoxia caused by soil
waterlogging flooding and examines some of the key
features of metabolic adaptation physiological and
morphological
16
C Electron
Transport System
We noted earlier that one of the principal
functions of the respiration is to retrieve in
useful form some of the energy initially stored in
assimilates Our traditional measure of useful
energy in most processes is the number of ATP
molecules gained or consumed By this measure alone
the yield from both glycolysis and the citric acid
cycle is quite low After two complete turns of the
cycle one molecule of glucose has been completely
oxidized to six molecules of CO2 but only four
molecules of ATP have been produced (a net of two
ATP from glycolysis plus one for each turn of the
cycle) At this point most of the energy associated
with the glucose molecule has been conserved in the
17
form of electron pairs generated by the oxidation of
glycolytic and citric acid cycle intermediates
In this section we will discuss the third stage
of cellular respirationmdashthe transfer of electrons
fromNADH and FADH2 to oxygen and the accompanying
conversion of redox energy to ATP The transfer of
electrons fromNADHand FADH2 to oxygen involves a
sequence of electron carriers arranged in an
electron transport chainMembrane fractionation
studies have shown that the enzymes and electron
carriers making up the electron transport chain are
organized predominantly into four large
multimolecular complexes (complexes ImdashIV) and two
mobile carriers located in the inner mitochondrial
membrane (Figure 109) In this sense there are a
great number of similarities between the
mitochondrial inner membrane and the thylakoid
membranes of the chloroplast (compare Figure 109
with Figure 76) This is not unexpected since the
principal function of each membrane is energy
transformation and many of the same or similar
components are involved
The Components of the Electron Transport Chain
The electron transport chain of the
mitochondria is the means by which electrons are
18
removed from the reduced carrier NADH and
transferred to oxygen to yield H2O
1) NADH
NADH is generated in the matrix by the reactions
of pyruvate dehydrogenase isocitrate
dehydrogenase α-ketoglutarate dehydrogenase and
malate dehyrogenase The electron transport chain
begins with reoxidizing NADH to form NAD+ and
channeling the electrons into the formation of
reduced coenzymes Important to note that NADH
transfers 2 electrons at a time in the form of a
hydride
NAD+ + 2e- + H+ 1048581 NADH Eorsquo = minus0315 V
2) Flavoproteins
Flavoproteins have either a FAD (flavin adenosine
dinucleotide) or a FMN (flavin mononucleotide)
prosthetic group Flavoproteins can accept or
donate electrons one at time or two at a time
Thus they are often intermediaries between two
electron acceptorsdonors and one electron
acceptorsdonors For flavoproteins the typical
standard reduction potentials are around 0 V
FAD + 2e- + 2H+ 1048581 FADH2 FMN + 2e- + 2H+ 1048581 FMNH2
Eorsquo asymp 0 V
19
3) Coenzyme Q (CoQ)
aka ubiquinone (UQ) Shown below CoQ has ten
repeating isoprene units which make it
insoluble in water but soluble in the
hydrophobic lipid bylayer Coenzyme Q is a
versatile cofactor because it is a soluble
electron carrier in the hydrophobic bilipid layer
of the
inner mitochondrial membrane Like flavoproteins
CoQ can acceptdonate electrons one at a time or
two at a time
Q + 2e- + 2H+ 1048581 QH2 Eorsquo =0060 V
Q + e- + H+ 1048581 1048581QH Eorsquo =0030 V
1048581QH + e- + H+ 1048581 QH2 Eorsquo =0190 V
4) Cytochromes
Cytochromes are proteins that contain heme
prosthetic groups which function as
one electron carriers The heme iron is involved
in one electron transfers involving the Fe2+ and
Fe3+ oxidation states Cytochromes are named by
their absorption spectra which depends
on the porphyrin structure and environment The
example shown is the heme prosthetic group of
cytochrome b Cytochrome b contains the same iron
porphyrin found in hemoglobin and myoglobin
20
Other cytochromes we will encounter in the
electron transport complexes are cytochromes b
c c1 a and a3
5) Iron-Sulfur Proteins
In the electron transport chain we will encounter
many iron-sulfer proteins which participate in
one electron transfers involving the the Fe2+ and
Fe3+ oxidation states These are non-heme iron-
sulfur proteinsThe simplest iron-sulfer protein
is FeS in which iron is tetrahedrally coordinated
by four cysteines The second form is Fe2S2 which
contains two irons complexed to 2 cysteine
residues and two inorganic sulfides The third
form is Fe3S4 which contains 3 iron atoms
coordinated to three cysteine residues and 4
inorganic sulfides The last form is the most
complicated Fe4S4 which contains 4 iron atoms
coordinated to 4 cysteine residues and 4
inorganic sulfides
6) Copper Proteins
Copper bound proteins participate in one electron
transfers involving the Cu+ and Cu2+ oxidation
states
21
Overview of the Electron Transport Chain
Electrons move along the electron transport
chain going from donor to acceptor until they reach
oxygen the ultimate electron acceptor The standard
reduction potentials of the electron carriers are
between the NADHNAD+ couple (-0315 V) and the
oxygenH2O couple (0816 V) as on the next page The
components of the electron transport chain are
organized into 4 complexes Each complex contains
several different electron carriers
1 Complex I also known as the NADH-coenzyme Q
reductase or NADH dehydrogenase
2 Complex II also known as succinate-coenzyme Q
reductase or succinate dehydrogenase
3 Complex III also known as coenzyme Q reductase
4 Complex IV also known as cytochrome c reductase
The free energy necessary to generate ATP
is extracted from the oxidation of NADH and
FADH2 by the electron transport chain It
consists of four protein complexes and single
protein chain of cytochrome c through which
electrons pass from lower to higher standard
redox potentials Electrons are carried from complex
I and II to complex III by the membrane (lipid)
soluble coenzyme Q (CoQ or ubiquinone) and between
22
complexes III and IV by the peripheral membrane
protein cytochrome c
In the following I give the overall
reactions catalysed by corresponding protein
complexes in the mitochondrial electron transport
chain
1 Complex I
Complex I is also called NADH-Coenzyme Q
reductase because this large protein complex
transfers 2 electrons from NADH to coenzyme Q
Complex I was formerly known as NADH dehydrogenase
Complex I is huge 850000 kD and is composed of more
than thirty subunits It contains a FMN prosthetic
group and seven or more Fe-S clusters This complex
has between 20-26 iron atoms boundThe prosthetic
group FMN is absolutely required for activity
Therefore this complex is a flavoprotein This
complex binds NADH transfers two electrons in the
form of a hydride to FMN to produce NAD+ and FMNH2
23
The subsequent steps involve the transfer of
electrons one at a time to a series of iron-sulfer
complexes that includes both 2Fe-2S and 4Fe-4S
clusters Note the importance of FMN First it
functions as a 2 electron acceptor in the hydride
transfer from NADH Second it functions as a 1
electron donor to the series of iron sulfur clusters
FMN and FAD often play crucial links between 2
electron transfer agents and 1 electron transfer
agents The final step of this complex is the
transfer of 2 electrons one at a time to coenzyme Q
CoQ like FMN and FAD can function as a 2 electron
donoracceptor and as a 1 electron donoracceptor
CoQ is a mobile electron carrier because its
isoprenoid tail makes it highly hydrophobic and
lipophillic It diffuses freely in the bilipid layer
of the inner mitochondrial membrane
The process of transferring electrons from NADH
to CoQ by complex I results in the net transport of
protons from the matrix side of the inner
mitochondrial membrane to the inter membrane space
where the H+ ions accumulate generating a proton
motive force The intermembrane space side of the
inner membrane is referred to as the P face (P
standing for positive) The matrix side of the inner
membrane is referred to as the the N face The
24
transport of electrons from NADH to CoQ is coupled to
the transport of protons across the membrane This is
an example of active transport The stiochiometry is
4 H+ transported per 2 electrons
2 Complex II
Believe it or not you are already familiar
with Complex II It is none other than succinate
dehydrogenase The only enzyme of the citric acid
cycle that is an integral membrane protein This
complex is composed of four subunits 2 of which are
iron-sulfur proteins and the other two subunits
together bind FAD through a covalent link to a
histidine residue These two subunits are called
flavoprotein 2 or FP2 Complex II contains 3 Fe-S
centers 1 4Fe-4S cluster 1 3Fe-4S cluster and 1
2Fe- 2S cluster In the first step of this complex
succinate is bound and a hydride is transferred to
FAD to generate FADH2 and fumarate FADH2 then
transfers its electrons one at a time to the Fe-S
centers Thus once again FAD functions as 2 electron
acceptor and a 1 electron donor The final step of
this complex is the transfer of 2 electrons one at a
time to coenzyme Q to produce CoQH2 For complex II
the standard free energy change of the overall
reaction is too small to drive the transport of
protons across the inner mitochondrial membrane This
25
accounts for the 15 ATPrsquos generated per FADH2
compared with the 25 ATPrsquos generated per NADH
3 Complex III
This complex is also known as coenzyme Q-
cytochrome c reductase because it passes the
electrons form CoQH2 to cyt c through a very unique
electron transport pathway called the Q-cycle Shown
to the left are the porphyrins found in cytochromes
Cytochrome b contains the same iron protoporphyrin as
hemoglobin and myoglobin The c cytochromes contain
heme c through covalent attachment by cysteine
residues Cytochrome a is found in two forms in
complex IV In complex III we find two b-type
cytochromes and one c-type cytochrome Complex III is
complex and we have a crystal structure
4 Complex IV
Complex IV is also known as cytochrome c oxidase
because it accepts the electrons from cytochrome c
and directs them towards the four electron reduction
of O2 to form 2 molecules of H2O
4 cyt c (Fe2+) + 4 H+ + O2 4 cyt c (Fe3+) +
2H2O
26
Cytochrome c oxidase contains 2 heme
centerscytochrome a and cytochrome a3 and two copper
proteins Each of the protein bound coppers are
associated with one of the cytochromes The copper
sites are called CuA and CuB CuA is associated with
cytochrome a and is shown to the left CuB is
associated with cytochrome a3 The copper sites
function as 1 electron carriers cycling between the
cuprous state Cu+ and the cupric state Cu2+ Just
like iron containing proteins they transfer
electrons one at a time Cytochrome c is bound on the
P-face of the membrane and transfers its electron to
CuA The oxidized cytochrome c dissociates CuA then
transfers the electron to cytochrome a The protein
bound CuA and the iron bound in cytochrome a are 15 Aring
apart In contrast the CuB and the iron bound in
cytochrome a3 are very close to each other forming a
binuclear metal center shown below
27
Electron transport chains are associated with
membranes such as the mitochondrial membrane in
eukaryotic organisms the thylakoid membrane of
photosynthetic organisms and the plasma membrane of
prokaryotic organisms While some of the details
differ electron transport chains in all organisms
operate on the same basic principles
Excited electrons are brought to the electron
transport chain by electron carriers such as NADH
and FADH2 In photosynthetic organisms the electrons
are donated by water and excited by light energy
The electron donor is oxidized as the electrons are
passed to the first protein complex in the chain As
it accepts the electrons the protein complex is
reduced Next the electrons are passed from the
28
first protein complex which becomes oxidized to
the second protein complex which becomes reduced
Each of these redox reactions is slightly exergonic
or energy releasing Some of this energy is used to
move protons across the membrane against their
concentration gradient With each successive
transfer of electrons the original excited
electrons lose some of their energy After several
successive redox reactions the electrons are
donated to a terminal electron acceptor In aerobic
respiration the terminal electron acceptor is
oxygen In photosynthesis the terminal electron
acceptor is a compound called NADP
The energy in the excited electrons has been
transformed into an electrochemical gradient
represented by the high concentration of protons on
one side of the membrane These protons can diffuse
back across the membrane by passing through a
protein complex called ATP synthase As these
protons pass through ATP synthase down their
concentration gradient they provide the energy
needed to make ATP from ADP and inorganic phosphate
within the cell
29
CHAPTER III
CLOSING
A Conclution
When a hexose is completely oxidized to CO2
and H2O using these three processes Glycolysis
yields two ATP and two NADH per hexose used Each
such NADH oxidized by the electron transport system
30
yields two ATP as dscribed above so glycolysis
contributes a total of six ATP per hexose The Krebs
cycle contributes two ATP per hexose or per two
pyruvates when succinyl CoA is cleaved to succinate
and CoASH This cycle also produces eight NADH per
hexose within the mitochondrial matrix by oxidative
phosporylation each of these NADH yields three ATP
or 24 per hexose Each FADH2 from the Krebs cycle
yields two ATP by oxydative phosporylation or four
per hexose (two pyruvates) The total contribution of
the Krebs cycle is then 30 Adding these 30 to the 6
from glycolysis leads to a total of 36 per hexose
completely respired by these process
B Suggestion
My greatest wish is that the readers find
this manuscript helpful in gaining the knowledge they
need for effective learning and practice and in
preparing for examination designed to test that
knowledge Reader comments and suggestions are
welcome and would be very helpful in improving our
manuscript Your contribution may assist many other
students in the coming years
31
REFERENCE
Anonim 2013 Anaerobic Respiration
httpwwwscoolcoukgcsebiologyrespiration
revise-itanaerobic-respiration Accessed
September 10th 2013
Anonim 2013 What is the difference between aerobic
and anaerobic respiration in plants
httpwwwbiologylifeeasyorg4010difference
-between-aerobic-anaerobic-respiration-plants
Accessed September 10th 2013
Campbell Mary K 2006 Biochemistry New York Thomson
Brooks Cole Inc
Dat J Capelli N Folzer H Bourgeade P Badot P-M
(2004) Sensing and signaling during
plantflooding Plant Physiology and
Biochemistry 42 273-282
Dat J Folzer H Parent C Badot P-M Capelli N
(2006) HypoxiastressCurrent Understanding and
Perspectives In Teixeira da Silva JA (Ed)
Floriculture Ornamental and Plant
Biotechnology Advances and Topical Issues (Vol
3) Global Science Books Isleworth United
Kingdompp 664-674
Hatta Muhammad 2011 Dunia Pertanian
httpemhattawordpresscomcategorystress
Accessed September 10th 2013
Hopkins William G 2009 Introduction to Plant Physiology
New York John Wiley amp Sons Inc
Lakitan Benyamin 2011 Dasar-Dasar Fisiologi Tumbuhan
Jakarta PT Raja Grafindo Persada
Larcher W 1980 Physiological Plant Ecology New York
Springer-Verlag
Raven 2013 Biology Fifth Edition
httphigheredmcgraw-hillcomsites983409233
9student_view0chapter39
photosynthetic_electron_transport_and_atp_synth
esishtml Accessed September 10th 2013
Salisbury Frank B 1969 Plant Physiology Third Edition
California Wadsworth Publishing Company
Subbaiah C Sachs M (2003) Molecular and cellular
adaptations of maize to flooding stress Annals
of Botany 91 119-127
Wilson CarlL1962 Botany Third EditionUnited State of
America Holt Rinch Inc

Steps involved in the process of Kreb cycle are
1 Fate of pyruvic acid
Pyruvic acid can form different compounds by
different pathways ie it can be converted into
lactic acid It can be converted into acetyl
coenzyme A as a result of oxidation
2 Fate of Acetyl CoA
It can either undergo condensation with itself or
its derivatives to form fatty acids having 14 ndash 20
carbons Acetyl CoA can also go through a series
of reactions in Krebs cycle
3 Formation of citrate
Acetyl CoA enzyme condenses with oxaloacetate by
an enzyme citrate synthatase to form citrate with
the release of acetyl CoA If the amount of
7
oxaloacetate is very small then small number of
acetyl CoA would be reacting with oxaloacetate
leaving surplus acetyl CoA to go through another
pathway for the formation of long chain fatty
acids
4 Formation of C is ndash acotinate and iso-citrate
Citrate is changed first into C is ndash acotinate and
then to iso-citrate under the enzyme acotinase
Equilibrium is established between citrate C is
aconitate and iso-citrate It has been observed
that most of the time this equilibrium is shifted
towards the iso-citrate If the concentration of
iso-citrate in increased the formation of citrate
will result which also indicates that the
equilibrium also shift in the reserve direction
5 Formation of oxalosucinate
Iso-citrate is acted upon by an enzyme iso-citrate
dehydrogenase using nicotinamide adenine
dinucleotide (NAD) as coenzyme As a result iso-
citrate is converted into oxalosucciate and NAD is
reduced to NADH2 Similar reaction is carried out
by same enzyme using nicotindmide adenine
dinucleotide phosphate (NADP) as coenzyme which is
reduced to NDAPH2
6 Formation of α-Ketoglutarate
8
Oxalosucciante is changed into α-ketoglutrate by
iso-citrate dehydrogenase with the help of
coenzyme NAD or NADP In this reaction carbon
dioxide and NADH2 or NADPH2 are also released
7 Formation of succinyl CoA
α-ketoglutarate combines with acetyl CoA in the
presence of coenzyme NAD and enzyme α-
keoglutaratedehyrogenase to form succinyl coenzyme
a carbon dioxide and NADH2
8 Formation of Succinate
Later on coenzyme a is removed from succinyl
coenzyme A in the presence of guanosinediphosphate
(GDP) and inorganic phosphate to form succinate
and guanosine triphosphate (GTP) This reaction is
carried out by an enzyme called succinyl CoA
synthatase
9 Formation of Funarate
An enzyme succinic dehydrogenase removes hydrogen
from succinate to form funarate
10 Formation of Malate
Fumarate reacts with water in the presence of
enzyme fumarase to form malate
11 Regeneration of oxaloacetate
Malate is oxidized by malic dehydrogenase and NAD
forming oxaloacetate and NADH2 Thus oxalo acetate
is again available to start another cycle
9
(httpkrebs-cycle-or-citric-acid-cycle-or-
trihtml)
Importance of CTA Cycle
1 Source of energy
In addition to routine organic compounds described
above by products like nicotinanide adenine
dinucleotide (NADH2) and guanosine triphosphate
(GTP) are the source of biological energy NADH2
after oxidation produce energy whereas GTP is
itself high energy phosphate compound
2 Oxidation of organic compounds taken as food
Oxidation of fats carbohydrates and proteins take
place through it or in other words it can be said
that oxidation of all compounds having carbon
atoms can take place through TCA cycle Some of
amino acids like alanine glutamic acid and
aspartic acid at one stage or the other enters
into TCA cycle eg glutamic acid enters cycle
after its transformation into α-ketoglutarate
Similarly alanine enters the cycle after its
conversion into pyruvate
3 Intermediate compounds
TCA cycle is also involved in synthesis of
intermediate compounds leading to the formation of
larger molecules (Ray 1963 17)
10
Verification of Krebs Cycle
It was done by radioactive traces like C14 as
radioactive carbon dioxide of different levels and
reactions After addition of radioactive carbon
dioxide different chemical compounds produced like
glucose fats amino acids were isolated and looked
for radioactive carbon In this way whole of
metabolic reaction were verified including individual
reactions alternative metabolic pathways
intermediates of fats carbohydrates and amino acids
etc in the body cells as well as in test tubes
(Wilson 1962 99)
The primary function of the Krebs cycle are as
follows
1 Reduction of NAD+ and FAD to the electron donors
NADH and FADH2 that are subsequently oxidized to
yield ATP
2 Direct synthesis of a limited amount of ATP (1 ATP
for each pyruvate oxidized)
3 Formation of carbon skeletons that can be used to
synthesize certain amino acids that in turn are
converted into larger molecules
11
B Fermentation
Although glycolysis can function well without
O2 the further oxidation of pyruvate and NADH by
mitochondria requires this gas Thus when O2 is
limiting NADH and pyruvate begin to accumulate
Under this condition plants carry out fermentation
(anaerobic respiration) forming either ethanol or
lactic acid (usually ethanol) As shown in the
picture below the two top reactions consists of a
decarboxylation to form acetaldehide then rapid
reduction of acetaldehide by NADH to form ethanol
These reactions are catalyzed by alcohol
dehydrogenase Some cells contain lactic acid
12
dehydrogenase which uses NADH to reduce pyruvic to
lactic acid Thus either ethanol or lactic acid or
both are fermentation products depending on the
activities of each dehydrogenase present In each
case NADH is the reductant and only under
anaerobic conditions is it abundant anough to cause
reduction Furthermore in some plants NADH is used
to cause accumulation of the other compounds when O2
is limiting especially malate and glycerol
(Salisbury 1969 233)
The fate of pyruvate produced by glycolysis
depends primarily on whether or not oxygen is
present Under normal aerobic conditions pyruvate
is transported into the mitochondrion where it is
further oxidized to CO2 and water transferring its
electrons ultimately to molecular oxygen
13
Although higher plants are obligate aerobs and
are able to tolerate anoxia for only short periods
tissues or organs are occasionally subjected to
anaerobic conditions A typical siuation is that of
roots when the soil is saturated with water When
there is no oxygen to serve as the terminal electron
acceptor mitochondrial respiration will shut down
and metabolism will shift over to fermentation
Fermentation converts pyruvate to either ethanol
through the action of the enzyme alcohol
dehydrogenase (ADH) or lactate via lactate
dehydrogenase (LDH) In most plants the principal
products of fermentation are CO2 and ethanol Some
lactate may be formed primarily in the early stages
of anoxia However lactate lowers he pH of the
cytosol which in turn activates pyruvate
decarboxylase and initiates the production of
ethanol
Either one of the fermentation reactions
consumes the NADH produced earlier in glycolysis by
the oxidation of glyceraldehide-3-P Although this
means there is no net gain of reducing potential in
fermentation This recycling of NADH is still
important to the cell The poolof NADH plus NAD+ in
the cell is relatively small and if the NADH is not
recycled there will be no supply of NAD+ to support
14
the continued oxidation of glyceraldehyde-3-P if
this were the case glycolysis and the producton of
even the small quantities of ATP necessary to
maintain the cells under anaerobic conditions would
then grind to a halt (Hopkins 2009 241)
Soil waterlogging has long been identified as a
major abiotic stress and the constraints that it
provides the roots have a significant effect on the
growth and development of plants When this event
occurs in the spring then these puddles can reduce
seed germination and seedling development Thus a
pool of water is an important factor that affects
the growth development and survival of plant
species not only in natural ecosystems but also
the agricultural and horticultural systems (Dat et
al 2006)
After flooding rapid changes in soil
properties At the time of pore water meets the
land the air pushed out gas diffusion is reduced
and toxic compounds accumulate due to anaerobic
conditions All of these changes greatly affect the
ability of plants to survive In response increased
stomatal resistance photosynthesis and root
hydraulic conductivity decreases and reduced
translocation fotoassimilat Nevertheless one of
15
the best adaptations of plants to hypoxia anoxia
is transitional biochemical and metabolic processes
common to current limited availability of O2 (Dat et
al 2004)
Selective synthesis of a set of about 20
anaerobic stress proteins (ANPS) allows the process
of producing energy without oxygen metabolism under
anaerobic conditions (Subbaiah and Sachs 2003)
Another adaptation is the observed morphological
changes consisting of the formation of hypertrophic
lenticels adventitious root initiation and or
progression of aerenchyma (Vartapetian and Jackson
1997 Jackson and Colmer 2005 Folzer et al 2006)
This review details the diverse plant stress
response to hypoxia anoxia caused by soil
waterlogging flooding and examines some of the key
features of metabolic adaptation physiological and
morphological
16
C Electron
Transport System
We noted earlier that one of the principal
functions of the respiration is to retrieve in
useful form some of the energy initially stored in
assimilates Our traditional measure of useful
energy in most processes is the number of ATP
molecules gained or consumed By this measure alone
the yield from both glycolysis and the citric acid
cycle is quite low After two complete turns of the
cycle one molecule of glucose has been completely
oxidized to six molecules of CO2 but only four
molecules of ATP have been produced (a net of two
ATP from glycolysis plus one for each turn of the
cycle) At this point most of the energy associated
with the glucose molecule has been conserved in the
17
form of electron pairs generated by the oxidation of
glycolytic and citric acid cycle intermediates
In this section we will discuss the third stage
of cellular respirationmdashthe transfer of electrons
fromNADH and FADH2 to oxygen and the accompanying
conversion of redox energy to ATP The transfer of
electrons fromNADHand FADH2 to oxygen involves a
sequence of electron carriers arranged in an
electron transport chainMembrane fractionation
studies have shown that the enzymes and electron
carriers making up the electron transport chain are
organized predominantly into four large
multimolecular complexes (complexes ImdashIV) and two
mobile carriers located in the inner mitochondrial
membrane (Figure 109) In this sense there are a
great number of similarities between the
mitochondrial inner membrane and the thylakoid
membranes of the chloroplast (compare Figure 109
with Figure 76) This is not unexpected since the
principal function of each membrane is energy
transformation and many of the same or similar
components are involved
The Components of the Electron Transport Chain
The electron transport chain of the
mitochondria is the means by which electrons are
18
removed from the reduced carrier NADH and
transferred to oxygen to yield H2O
1) NADH
NADH is generated in the matrix by the reactions
of pyruvate dehydrogenase isocitrate
dehydrogenase α-ketoglutarate dehydrogenase and
malate dehyrogenase The electron transport chain
begins with reoxidizing NADH to form NAD+ and
channeling the electrons into the formation of
reduced coenzymes Important to note that NADH
transfers 2 electrons at a time in the form of a
hydride
NAD+ + 2e- + H+ 1048581 NADH Eorsquo = minus0315 V
2) Flavoproteins
Flavoproteins have either a FAD (flavin adenosine
dinucleotide) or a FMN (flavin mononucleotide)
prosthetic group Flavoproteins can accept or
donate electrons one at time or two at a time
Thus they are often intermediaries between two
electron acceptorsdonors and one electron
acceptorsdonors For flavoproteins the typical
standard reduction potentials are around 0 V
FAD + 2e- + 2H+ 1048581 FADH2 FMN + 2e- + 2H+ 1048581 FMNH2
Eorsquo asymp 0 V
19
3) Coenzyme Q (CoQ)
aka ubiquinone (UQ) Shown below CoQ has ten
repeating isoprene units which make it
insoluble in water but soluble in the
hydrophobic lipid bylayer Coenzyme Q is a
versatile cofactor because it is a soluble
electron carrier in the hydrophobic bilipid layer
of the
inner mitochondrial membrane Like flavoproteins
CoQ can acceptdonate electrons one at a time or
two at a time
Q + 2e- + 2H+ 1048581 QH2 Eorsquo =0060 V
Q + e- + H+ 1048581 1048581QH Eorsquo =0030 V
1048581QH + e- + H+ 1048581 QH2 Eorsquo =0190 V
4) Cytochromes
Cytochromes are proteins that contain heme
prosthetic groups which function as
one electron carriers The heme iron is involved
in one electron transfers involving the Fe2+ and
Fe3+ oxidation states Cytochromes are named by
their absorption spectra which depends
on the porphyrin structure and environment The
example shown is the heme prosthetic group of
cytochrome b Cytochrome b contains the same iron
porphyrin found in hemoglobin and myoglobin
20
Other cytochromes we will encounter in the
electron transport complexes are cytochromes b
c c1 a and a3
5) Iron-Sulfur Proteins
In the electron transport chain we will encounter
many iron-sulfer proteins which participate in
one electron transfers involving the the Fe2+ and
Fe3+ oxidation states These are non-heme iron-
sulfur proteinsThe simplest iron-sulfer protein
is FeS in which iron is tetrahedrally coordinated
by four cysteines The second form is Fe2S2 which
contains two irons complexed to 2 cysteine
residues and two inorganic sulfides The third
form is Fe3S4 which contains 3 iron atoms
coordinated to three cysteine residues and 4
inorganic sulfides The last form is the most
complicated Fe4S4 which contains 4 iron atoms
coordinated to 4 cysteine residues and 4
inorganic sulfides
6) Copper Proteins
Copper bound proteins participate in one electron
transfers involving the Cu+ and Cu2+ oxidation
states
21
Overview of the Electron Transport Chain
Electrons move along the electron transport
chain going from donor to acceptor until they reach
oxygen the ultimate electron acceptor The standard
reduction potentials of the electron carriers are
between the NADHNAD+ couple (-0315 V) and the
oxygenH2O couple (0816 V) as on the next page The
components of the electron transport chain are
organized into 4 complexes Each complex contains
several different electron carriers
1 Complex I also known as the NADH-coenzyme Q
reductase or NADH dehydrogenase
2 Complex II also known as succinate-coenzyme Q
reductase or succinate dehydrogenase
3 Complex III also known as coenzyme Q reductase
4 Complex IV also known as cytochrome c reductase
The free energy necessary to generate ATP
is extracted from the oxidation of NADH and
FADH2 by the electron transport chain It
consists of four protein complexes and single
protein chain of cytochrome c through which
electrons pass from lower to higher standard
redox potentials Electrons are carried from complex
I and II to complex III by the membrane (lipid)
soluble coenzyme Q (CoQ or ubiquinone) and between
22
complexes III and IV by the peripheral membrane
protein cytochrome c
In the following I give the overall
reactions catalysed by corresponding protein
complexes in the mitochondrial electron transport
chain
1 Complex I
Complex I is also called NADH-Coenzyme Q
reductase because this large protein complex
transfers 2 electrons from NADH to coenzyme Q
Complex I was formerly known as NADH dehydrogenase
Complex I is huge 850000 kD and is composed of more
than thirty subunits It contains a FMN prosthetic
group and seven or more Fe-S clusters This complex
has between 20-26 iron atoms boundThe prosthetic
group FMN is absolutely required for activity
Therefore this complex is a flavoprotein This
complex binds NADH transfers two electrons in the
form of a hydride to FMN to produce NAD+ and FMNH2
23
The subsequent steps involve the transfer of
electrons one at a time to a series of iron-sulfer
complexes that includes both 2Fe-2S and 4Fe-4S
clusters Note the importance of FMN First it
functions as a 2 electron acceptor in the hydride
transfer from NADH Second it functions as a 1
electron donor to the series of iron sulfur clusters
FMN and FAD often play crucial links between 2
electron transfer agents and 1 electron transfer
agents The final step of this complex is the
transfer of 2 electrons one at a time to coenzyme Q
CoQ like FMN and FAD can function as a 2 electron
donoracceptor and as a 1 electron donoracceptor
CoQ is a mobile electron carrier because its
isoprenoid tail makes it highly hydrophobic and
lipophillic It diffuses freely in the bilipid layer
of the inner mitochondrial membrane
The process of transferring electrons from NADH
to CoQ by complex I results in the net transport of
protons from the matrix side of the inner
mitochondrial membrane to the inter membrane space
where the H+ ions accumulate generating a proton
motive force The intermembrane space side of the
inner membrane is referred to as the P face (P
standing for positive) The matrix side of the inner
membrane is referred to as the the N face The
24
transport of electrons from NADH to CoQ is coupled to
the transport of protons across the membrane This is
an example of active transport The stiochiometry is
4 H+ transported per 2 electrons
2 Complex II
Believe it or not you are already familiar
with Complex II It is none other than succinate
dehydrogenase The only enzyme of the citric acid
cycle that is an integral membrane protein This
complex is composed of four subunits 2 of which are
iron-sulfur proteins and the other two subunits
together bind FAD through a covalent link to a
histidine residue These two subunits are called
flavoprotein 2 or FP2 Complex II contains 3 Fe-S
centers 1 4Fe-4S cluster 1 3Fe-4S cluster and 1
2Fe- 2S cluster In the first step of this complex
succinate is bound and a hydride is transferred to
FAD to generate FADH2 and fumarate FADH2 then
transfers its electrons one at a time to the Fe-S
centers Thus once again FAD functions as 2 electron
acceptor and a 1 electron donor The final step of
this complex is the transfer of 2 electrons one at a
time to coenzyme Q to produce CoQH2 For complex II
the standard free energy change of the overall
reaction is too small to drive the transport of
protons across the inner mitochondrial membrane This
25
accounts for the 15 ATPrsquos generated per FADH2
compared with the 25 ATPrsquos generated per NADH
3 Complex III
This complex is also known as coenzyme Q-
cytochrome c reductase because it passes the
electrons form CoQH2 to cyt c through a very unique
electron transport pathway called the Q-cycle Shown
to the left are the porphyrins found in cytochromes
Cytochrome b contains the same iron protoporphyrin as
hemoglobin and myoglobin The c cytochromes contain
heme c through covalent attachment by cysteine
residues Cytochrome a is found in two forms in
complex IV In complex III we find two b-type
cytochromes and one c-type cytochrome Complex III is
complex and we have a crystal structure
4 Complex IV
Complex IV is also known as cytochrome c oxidase
because it accepts the electrons from cytochrome c
and directs them towards the four electron reduction
of O2 to form 2 molecules of H2O
4 cyt c (Fe2+) + 4 H+ + O2 4 cyt c (Fe3+) +
2H2O
26
Cytochrome c oxidase contains 2 heme
centerscytochrome a and cytochrome a3 and two copper
proteins Each of the protein bound coppers are
associated with one of the cytochromes The copper
sites are called CuA and CuB CuA is associated with
cytochrome a and is shown to the left CuB is
associated with cytochrome a3 The copper sites
function as 1 electron carriers cycling between the
cuprous state Cu+ and the cupric state Cu2+ Just
like iron containing proteins they transfer
electrons one at a time Cytochrome c is bound on the
P-face of the membrane and transfers its electron to
CuA The oxidized cytochrome c dissociates CuA then
transfers the electron to cytochrome a The protein
bound CuA and the iron bound in cytochrome a are 15 Aring
apart In contrast the CuB and the iron bound in
cytochrome a3 are very close to each other forming a
binuclear metal center shown below
27
Electron transport chains are associated with
membranes such as the mitochondrial membrane in
eukaryotic organisms the thylakoid membrane of
photosynthetic organisms and the plasma membrane of
prokaryotic organisms While some of the details
differ electron transport chains in all organisms
operate on the same basic principles
Excited electrons are brought to the electron
transport chain by electron carriers such as NADH
and FADH2 In photosynthetic organisms the electrons
are donated by water and excited by light energy
The electron donor is oxidized as the electrons are
passed to the first protein complex in the chain As
it accepts the electrons the protein complex is
reduced Next the electrons are passed from the
28
first protein complex which becomes oxidized to
the second protein complex which becomes reduced
Each of these redox reactions is slightly exergonic
or energy releasing Some of this energy is used to
move protons across the membrane against their
concentration gradient With each successive
transfer of electrons the original excited
electrons lose some of their energy After several
successive redox reactions the electrons are
donated to a terminal electron acceptor In aerobic
respiration the terminal electron acceptor is
oxygen In photosynthesis the terminal electron
acceptor is a compound called NADP
The energy in the excited electrons has been
transformed into an electrochemical gradient
represented by the high concentration of protons on
one side of the membrane These protons can diffuse
back across the membrane by passing through a
protein complex called ATP synthase As these
protons pass through ATP synthase down their
concentration gradient they provide the energy
needed to make ATP from ADP and inorganic phosphate
within the cell
29
CHAPTER III
CLOSING
A Conclution
When a hexose is completely oxidized to CO2
and H2O using these three processes Glycolysis
yields two ATP and two NADH per hexose used Each
such NADH oxidized by the electron transport system
30
yields two ATP as dscribed above so glycolysis
contributes a total of six ATP per hexose The Krebs
cycle contributes two ATP per hexose or per two
pyruvates when succinyl CoA is cleaved to succinate
and CoASH This cycle also produces eight NADH per
hexose within the mitochondrial matrix by oxidative
phosporylation each of these NADH yields three ATP
or 24 per hexose Each FADH2 from the Krebs cycle
yields two ATP by oxydative phosporylation or four
per hexose (two pyruvates) The total contribution of
the Krebs cycle is then 30 Adding these 30 to the 6
from glycolysis leads to a total of 36 per hexose
completely respired by these process
B Suggestion
My greatest wish is that the readers find
this manuscript helpful in gaining the knowledge they
need for effective learning and practice and in
preparing for examination designed to test that
knowledge Reader comments and suggestions are
welcome and would be very helpful in improving our
manuscript Your contribution may assist many other
students in the coming years
31
REFERENCE
Anonim 2013 Anaerobic Respiration
httpwwwscoolcoukgcsebiologyrespiration
revise-itanaerobic-respiration Accessed
September 10th 2013
Anonim 2013 What is the difference between aerobic
and anaerobic respiration in plants
httpwwwbiologylifeeasyorg4010difference
-between-aerobic-anaerobic-respiration-plants
Accessed September 10th 2013
Campbell Mary K 2006 Biochemistry New York Thomson
Brooks Cole Inc
Dat J Capelli N Folzer H Bourgeade P Badot P-M
(2004) Sensing and signaling during
plantflooding Plant Physiology and
Biochemistry 42 273-282
Dat J Folzer H Parent C Badot P-M Capelli N
(2006) HypoxiastressCurrent Understanding and
Perspectives In Teixeira da Silva JA (Ed)
Floriculture Ornamental and Plant
Biotechnology Advances and Topical Issues (Vol
3) Global Science Books Isleworth United
Kingdompp 664-674
Hatta Muhammad 2011 Dunia Pertanian
httpemhattawordpresscomcategorystress
Accessed September 10th 2013
Hopkins William G 2009 Introduction to Plant Physiology
New York John Wiley amp Sons Inc
Lakitan Benyamin 2011 Dasar-Dasar Fisiologi Tumbuhan
Jakarta PT Raja Grafindo Persada
Larcher W 1980 Physiological Plant Ecology New York
Springer-Verlag
Raven 2013 Biology Fifth Edition
httphigheredmcgraw-hillcomsites983409233
9student_view0chapter39
photosynthetic_electron_transport_and_atp_synth
esishtml Accessed September 10th 2013
Salisbury Frank B 1969 Plant Physiology Third Edition
California Wadsworth Publishing Company
Subbaiah C Sachs M (2003) Molecular and cellular
adaptations of maize to flooding stress Annals
of Botany 91 119-127
Wilson CarlL1962 Botany Third EditionUnited State of
America Holt Rinch Inc

oxaloacetate is very small then small number of
acetyl CoA would be reacting with oxaloacetate
leaving surplus acetyl CoA to go through another
pathway for the formation of long chain fatty
acids
4 Formation of C is ndash acotinate and iso-citrate
Citrate is changed first into C is ndash acotinate and
then to iso-citrate under the enzyme acotinase
Equilibrium is established between citrate C is
aconitate and iso-citrate It has been observed
that most of the time this equilibrium is shifted
towards the iso-citrate If the concentration of
iso-citrate in increased the formation of citrate
will result which also indicates that the
equilibrium also shift in the reserve direction
5 Formation of oxalosucinate
Iso-citrate is acted upon by an enzyme iso-citrate
dehydrogenase using nicotinamide adenine
dinucleotide (NAD) as coenzyme As a result iso-
citrate is converted into oxalosucciate and NAD is
reduced to NADH2 Similar reaction is carried out
by same enzyme using nicotindmide adenine
dinucleotide phosphate (NADP) as coenzyme which is
reduced to NDAPH2
6 Formation of α-Ketoglutarate
8
Oxalosucciante is changed into α-ketoglutrate by
iso-citrate dehydrogenase with the help of
coenzyme NAD or NADP In this reaction carbon
dioxide and NADH2 or NADPH2 are also released
7 Formation of succinyl CoA
α-ketoglutarate combines with acetyl CoA in the
presence of coenzyme NAD and enzyme α-
keoglutaratedehyrogenase to form succinyl coenzyme
a carbon dioxide and NADH2
8 Formation of Succinate
Later on coenzyme a is removed from succinyl
coenzyme A in the presence of guanosinediphosphate
(GDP) and inorganic phosphate to form succinate
and guanosine triphosphate (GTP) This reaction is
carried out by an enzyme called succinyl CoA
synthatase
9 Formation of Funarate
An enzyme succinic dehydrogenase removes hydrogen
from succinate to form funarate
10 Formation of Malate
Fumarate reacts with water in the presence of
enzyme fumarase to form malate
11 Regeneration of oxaloacetate
Malate is oxidized by malic dehydrogenase and NAD
forming oxaloacetate and NADH2 Thus oxalo acetate
is again available to start another cycle
9
(httpkrebs-cycle-or-citric-acid-cycle-or-
trihtml)
Importance of CTA Cycle
1 Source of energy
In addition to routine organic compounds described
above by products like nicotinanide adenine
dinucleotide (NADH2) and guanosine triphosphate
(GTP) are the source of biological energy NADH2
after oxidation produce energy whereas GTP is
itself high energy phosphate compound
2 Oxidation of organic compounds taken as food
Oxidation of fats carbohydrates and proteins take
place through it or in other words it can be said
that oxidation of all compounds having carbon
atoms can take place through TCA cycle Some of
amino acids like alanine glutamic acid and
aspartic acid at one stage or the other enters
into TCA cycle eg glutamic acid enters cycle
after its transformation into α-ketoglutarate
Similarly alanine enters the cycle after its
conversion into pyruvate
3 Intermediate compounds
TCA cycle is also involved in synthesis of
intermediate compounds leading to the formation of
larger molecules (Ray 1963 17)
10
Verification of Krebs Cycle
It was done by radioactive traces like C14 as
radioactive carbon dioxide of different levels and
reactions After addition of radioactive carbon
dioxide different chemical compounds produced like
glucose fats amino acids were isolated and looked
for radioactive carbon In this way whole of
metabolic reaction were verified including individual
reactions alternative metabolic pathways
intermediates of fats carbohydrates and amino acids
etc in the body cells as well as in test tubes
(Wilson 1962 99)
The primary function of the Krebs cycle are as
follows
1 Reduction of NAD+ and FAD to the electron donors
NADH and FADH2 that are subsequently oxidized to
yield ATP
2 Direct synthesis of a limited amount of ATP (1 ATP
for each pyruvate oxidized)
3 Formation of carbon skeletons that can be used to
synthesize certain amino acids that in turn are
converted into larger molecules
11
B Fermentation
Although glycolysis can function well without
O2 the further oxidation of pyruvate and NADH by
mitochondria requires this gas Thus when O2 is
limiting NADH and pyruvate begin to accumulate
Under this condition plants carry out fermentation
(anaerobic respiration) forming either ethanol or
lactic acid (usually ethanol) As shown in the
picture below the two top reactions consists of a
decarboxylation to form acetaldehide then rapid
reduction of acetaldehide by NADH to form ethanol
These reactions are catalyzed by alcohol
dehydrogenase Some cells contain lactic acid
12
dehydrogenase which uses NADH to reduce pyruvic to
lactic acid Thus either ethanol or lactic acid or
both are fermentation products depending on the
activities of each dehydrogenase present In each
case NADH is the reductant and only under
anaerobic conditions is it abundant anough to cause
reduction Furthermore in some plants NADH is used
to cause accumulation of the other compounds when O2
is limiting especially malate and glycerol
(Salisbury 1969 233)
The fate of pyruvate produced by glycolysis
depends primarily on whether or not oxygen is
present Under normal aerobic conditions pyruvate
is transported into the mitochondrion where it is
further oxidized to CO2 and water transferring its
electrons ultimately to molecular oxygen
13
Although higher plants are obligate aerobs and
are able to tolerate anoxia for only short periods
tissues or organs are occasionally subjected to
anaerobic conditions A typical siuation is that of
roots when the soil is saturated with water When
there is no oxygen to serve as the terminal electron
acceptor mitochondrial respiration will shut down
and metabolism will shift over to fermentation
Fermentation converts pyruvate to either ethanol
through the action of the enzyme alcohol
dehydrogenase (ADH) or lactate via lactate
dehydrogenase (LDH) In most plants the principal
products of fermentation are CO2 and ethanol Some
lactate may be formed primarily in the early stages
of anoxia However lactate lowers he pH of the
cytosol which in turn activates pyruvate
decarboxylase and initiates the production of
ethanol
Either one of the fermentation reactions
consumes the NADH produced earlier in glycolysis by
the oxidation of glyceraldehide-3-P Although this
means there is no net gain of reducing potential in
fermentation This recycling of NADH is still
important to the cell The poolof NADH plus NAD+ in
the cell is relatively small and if the NADH is not
recycled there will be no supply of NAD+ to support
14
the continued oxidation of glyceraldehyde-3-P if
this were the case glycolysis and the producton of
even the small quantities of ATP necessary to
maintain the cells under anaerobic conditions would
then grind to a halt (Hopkins 2009 241)
Soil waterlogging has long been identified as a
major abiotic stress and the constraints that it
provides the roots have a significant effect on the
growth and development of plants When this event
occurs in the spring then these puddles can reduce
seed germination and seedling development Thus a
pool of water is an important factor that affects
the growth development and survival of plant
species not only in natural ecosystems but also
the agricultural and horticultural systems (Dat et
al 2006)
After flooding rapid changes in soil
properties At the time of pore water meets the
land the air pushed out gas diffusion is reduced
and toxic compounds accumulate due to anaerobic
conditions All of these changes greatly affect the
ability of plants to survive In response increased
stomatal resistance photosynthesis and root
hydraulic conductivity decreases and reduced
translocation fotoassimilat Nevertheless one of
15
the best adaptations of plants to hypoxia anoxia
is transitional biochemical and metabolic processes
common to current limited availability of O2 (Dat et
al 2004)
Selective synthesis of a set of about 20
anaerobic stress proteins (ANPS) allows the process
of producing energy without oxygen metabolism under
anaerobic conditions (Subbaiah and Sachs 2003)
Another adaptation is the observed morphological
changes consisting of the formation of hypertrophic
lenticels adventitious root initiation and or
progression of aerenchyma (Vartapetian and Jackson
1997 Jackson and Colmer 2005 Folzer et al 2006)
This review details the diverse plant stress
response to hypoxia anoxia caused by soil
waterlogging flooding and examines some of the key
features of metabolic adaptation physiological and
morphological
16
C Electron
Transport System
We noted earlier that one of the principal
functions of the respiration is to retrieve in
useful form some of the energy initially stored in
assimilates Our traditional measure of useful
energy in most processes is the number of ATP
molecules gained or consumed By this measure alone
the yield from both glycolysis and the citric acid
cycle is quite low After two complete turns of the
cycle one molecule of glucose has been completely
oxidized to six molecules of CO2 but only four
molecules of ATP have been produced (a net of two
ATP from glycolysis plus one for each turn of the
cycle) At this point most of the energy associated
with the glucose molecule has been conserved in the
17
form of electron pairs generated by the oxidation of
glycolytic and citric acid cycle intermediates
In this section we will discuss the third stage
of cellular respirationmdashthe transfer of electrons
fromNADH and FADH2 to oxygen and the accompanying
conversion of redox energy to ATP The transfer of
electrons fromNADHand FADH2 to oxygen involves a
sequence of electron carriers arranged in an
electron transport chainMembrane fractionation
studies have shown that the enzymes and electron
carriers making up the electron transport chain are
organized predominantly into four large
multimolecular complexes (complexes ImdashIV) and two
mobile carriers located in the inner mitochondrial
membrane (Figure 109) In this sense there are a
great number of similarities between the
mitochondrial inner membrane and the thylakoid
membranes of the chloroplast (compare Figure 109
with Figure 76) This is not unexpected since the
principal function of each membrane is energy
transformation and many of the same or similar
components are involved
The Components of the Electron Transport Chain
The electron transport chain of the
mitochondria is the means by which electrons are
18
removed from the reduced carrier NADH and
transferred to oxygen to yield H2O
1) NADH
NADH is generated in the matrix by the reactions
of pyruvate dehydrogenase isocitrate
dehydrogenase α-ketoglutarate dehydrogenase and
malate dehyrogenase The electron transport chain
begins with reoxidizing NADH to form NAD+ and
channeling the electrons into the formation of
reduced coenzymes Important to note that NADH
transfers 2 electrons at a time in the form of a
hydride
NAD+ + 2e- + H+ 1048581 NADH Eorsquo = minus0315 V
2) Flavoproteins
Flavoproteins have either a FAD (flavin adenosine
dinucleotide) or a FMN (flavin mononucleotide)
prosthetic group Flavoproteins can accept or
donate electrons one at time or two at a time
Thus they are often intermediaries between two
electron acceptorsdonors and one electron
acceptorsdonors For flavoproteins the typical
standard reduction potentials are around 0 V
FAD + 2e- + 2H+ 1048581 FADH2 FMN + 2e- + 2H+ 1048581 FMNH2
Eorsquo asymp 0 V
19
3) Coenzyme Q (CoQ)
aka ubiquinone (UQ) Shown below CoQ has ten
repeating isoprene units which make it
insoluble in water but soluble in the
hydrophobic lipid bylayer Coenzyme Q is a
versatile cofactor because it is a soluble
electron carrier in the hydrophobic bilipid layer
of the
inner mitochondrial membrane Like flavoproteins
CoQ can acceptdonate electrons one at a time or
two at a time
Q + 2e- + 2H+ 1048581 QH2 Eorsquo =0060 V
Q + e- + H+ 1048581 1048581QH Eorsquo =0030 V
1048581QH + e- + H+ 1048581 QH2 Eorsquo =0190 V
4) Cytochromes
Cytochromes are proteins that contain heme
prosthetic groups which function as
one electron carriers The heme iron is involved
in one electron transfers involving the Fe2+ and
Fe3+ oxidation states Cytochromes are named by
their absorption spectra which depends
on the porphyrin structure and environment The
example shown is the heme prosthetic group of
cytochrome b Cytochrome b contains the same iron
porphyrin found in hemoglobin and myoglobin
20
Other cytochromes we will encounter in the
electron transport complexes are cytochromes b
c c1 a and a3
5) Iron-Sulfur Proteins
In the electron transport chain we will encounter
many iron-sulfer proteins which participate in
one electron transfers involving the the Fe2+ and
Fe3+ oxidation states These are non-heme iron-
sulfur proteinsThe simplest iron-sulfer protein
is FeS in which iron is tetrahedrally coordinated
by four cysteines The second form is Fe2S2 which
contains two irons complexed to 2 cysteine
residues and two inorganic sulfides The third
form is Fe3S4 which contains 3 iron atoms
coordinated to three cysteine residues and 4
inorganic sulfides The last form is the most
complicated Fe4S4 which contains 4 iron atoms
coordinated to 4 cysteine residues and 4
inorganic sulfides
6) Copper Proteins
Copper bound proteins participate in one electron
transfers involving the Cu+ and Cu2+ oxidation
states
21
Overview of the Electron Transport Chain
Electrons move along the electron transport
chain going from donor to acceptor until they reach
oxygen the ultimate electron acceptor The standard
reduction potentials of the electron carriers are
between the NADHNAD+ couple (-0315 V) and the
oxygenH2O couple (0816 V) as on the next page The
components of the electron transport chain are
organized into 4 complexes Each complex contains
several different electron carriers
1 Complex I also known as the NADH-coenzyme Q
reductase or NADH dehydrogenase
2 Complex II also known as succinate-coenzyme Q
reductase or succinate dehydrogenase
3 Complex III also known as coenzyme Q reductase
4 Complex IV also known as cytochrome c reductase
The free energy necessary to generate ATP
is extracted from the oxidation of NADH and
FADH2 by the electron transport chain It
consists of four protein complexes and single
protein chain of cytochrome c through which
electrons pass from lower to higher standard
redox potentials Electrons are carried from complex
I and II to complex III by the membrane (lipid)
soluble coenzyme Q (CoQ or ubiquinone) and between
22
complexes III and IV by the peripheral membrane
protein cytochrome c
In the following I give the overall
reactions catalysed by corresponding protein
complexes in the mitochondrial electron transport
chain
1 Complex I
Complex I is also called NADH-Coenzyme Q
reductase because this large protein complex
transfers 2 electrons from NADH to coenzyme Q
Complex I was formerly known as NADH dehydrogenase
Complex I is huge 850000 kD and is composed of more
than thirty subunits It contains a FMN prosthetic
group and seven or more Fe-S clusters This complex
has between 20-26 iron atoms boundThe prosthetic
group FMN is absolutely required for activity
Therefore this complex is a flavoprotein This
complex binds NADH transfers two electrons in the
form of a hydride to FMN to produce NAD+ and FMNH2
23
The subsequent steps involve the transfer of
electrons one at a time to a series of iron-sulfer
complexes that includes both 2Fe-2S and 4Fe-4S
clusters Note the importance of FMN First it
functions as a 2 electron acceptor in the hydride
transfer from NADH Second it functions as a 1
electron donor to the series of iron sulfur clusters
FMN and FAD often play crucial links between 2
electron transfer agents and 1 electron transfer
agents The final step of this complex is the
transfer of 2 electrons one at a time to coenzyme Q
CoQ like FMN and FAD can function as a 2 electron
donoracceptor and as a 1 electron donoracceptor
CoQ is a mobile electron carrier because its
isoprenoid tail makes it highly hydrophobic and
lipophillic It diffuses freely in the bilipid layer
of the inner mitochondrial membrane
The process of transferring electrons from NADH
to CoQ by complex I results in the net transport of
protons from the matrix side of the inner
mitochondrial membrane to the inter membrane space
where the H+ ions accumulate generating a proton
motive force The intermembrane space side of the
inner membrane is referred to as the P face (P
standing for positive) The matrix side of the inner
membrane is referred to as the the N face The
24
transport of electrons from NADH to CoQ is coupled to
the transport of protons across the membrane This is
an example of active transport The stiochiometry is
4 H+ transported per 2 electrons
2 Complex II
Believe it or not you are already familiar
with Complex II It is none other than succinate
dehydrogenase The only enzyme of the citric acid
cycle that is an integral membrane protein This
complex is composed of four subunits 2 of which are
iron-sulfur proteins and the other two subunits
together bind FAD through a covalent link to a
histidine residue These two subunits are called
flavoprotein 2 or FP2 Complex II contains 3 Fe-S
centers 1 4Fe-4S cluster 1 3Fe-4S cluster and 1
2Fe- 2S cluster In the first step of this complex
succinate is bound and a hydride is transferred to
FAD to generate FADH2 and fumarate FADH2 then
transfers its electrons one at a time to the Fe-S
centers Thus once again FAD functions as 2 electron
acceptor and a 1 electron donor The final step of
this complex is the transfer of 2 electrons one at a
time to coenzyme Q to produce CoQH2 For complex II
the standard free energy change of the overall
reaction is too small to drive the transport of
protons across the inner mitochondrial membrane This
25
accounts for the 15 ATPrsquos generated per FADH2
compared with the 25 ATPrsquos generated per NADH
3 Complex III
This complex is also known as coenzyme Q-
cytochrome c reductase because it passes the
electrons form CoQH2 to cyt c through a very unique
electron transport pathway called the Q-cycle Shown
to the left are the porphyrins found in cytochromes
Cytochrome b contains the same iron protoporphyrin as
hemoglobin and myoglobin The c cytochromes contain
heme c through covalent attachment by cysteine
residues Cytochrome a is found in two forms in
complex IV In complex III we find two b-type
cytochromes and one c-type cytochrome Complex III is
complex and we have a crystal structure
4 Complex IV
Complex IV is also known as cytochrome c oxidase
because it accepts the electrons from cytochrome c
and directs them towards the four electron reduction
of O2 to form 2 molecules of H2O
4 cyt c (Fe2+) + 4 H+ + O2 4 cyt c (Fe3+) +
2H2O
26
Cytochrome c oxidase contains 2 heme
centerscytochrome a and cytochrome a3 and two copper
proteins Each of the protein bound coppers are
associated with one of the cytochromes The copper
sites are called CuA and CuB CuA is associated with
cytochrome a and is shown to the left CuB is
associated with cytochrome a3 The copper sites
function as 1 electron carriers cycling between the
cuprous state Cu+ and the cupric state Cu2+ Just
like iron containing proteins they transfer
electrons one at a time Cytochrome c is bound on the
P-face of the membrane and transfers its electron to
CuA The oxidized cytochrome c dissociates CuA then
transfers the electron to cytochrome a The protein
bound CuA and the iron bound in cytochrome a are 15 Aring
apart In contrast the CuB and the iron bound in
cytochrome a3 are very close to each other forming a
binuclear metal center shown below
27
Electron transport chains are associated with
membranes such as the mitochondrial membrane in
eukaryotic organisms the thylakoid membrane of
photosynthetic organisms and the plasma membrane of
prokaryotic organisms While some of the details
differ electron transport chains in all organisms
operate on the same basic principles
Excited electrons are brought to the electron
transport chain by electron carriers such as NADH
and FADH2 In photosynthetic organisms the electrons
are donated by water and excited by light energy
The electron donor is oxidized as the electrons are
passed to the first protein complex in the chain As
it accepts the electrons the protein complex is
reduced Next the electrons are passed from the
28
first protein complex which becomes oxidized to
the second protein complex which becomes reduced
Each of these redox reactions is slightly exergonic
or energy releasing Some of this energy is used to
move protons across the membrane against their
concentration gradient With each successive
transfer of electrons the original excited
electrons lose some of their energy After several
successive redox reactions the electrons are
donated to a terminal electron acceptor In aerobic
respiration the terminal electron acceptor is
oxygen In photosynthesis the terminal electron
acceptor is a compound called NADP
The energy in the excited electrons has been
transformed into an electrochemical gradient
represented by the high concentration of protons on
one side of the membrane These protons can diffuse
back across the membrane by passing through a
protein complex called ATP synthase As these
protons pass through ATP synthase down their
concentration gradient they provide the energy
needed to make ATP from ADP and inorganic phosphate
within the cell
29
CHAPTER III
CLOSING
A Conclution
When a hexose is completely oxidized to CO2
and H2O using these three processes Glycolysis
yields two ATP and two NADH per hexose used Each
such NADH oxidized by the electron transport system
30
yields two ATP as dscribed above so glycolysis
contributes a total of six ATP per hexose The Krebs
cycle contributes two ATP per hexose or per two
pyruvates when succinyl CoA is cleaved to succinate
and CoASH This cycle also produces eight NADH per
hexose within the mitochondrial matrix by oxidative
phosporylation each of these NADH yields three ATP
or 24 per hexose Each FADH2 from the Krebs cycle
yields two ATP by oxydative phosporylation or four
per hexose (two pyruvates) The total contribution of
the Krebs cycle is then 30 Adding these 30 to the 6
from glycolysis leads to a total of 36 per hexose
completely respired by these process
B Suggestion
My greatest wish is that the readers find
this manuscript helpful in gaining the knowledge they
need for effective learning and practice and in
preparing for examination designed to test that
knowledge Reader comments and suggestions are
welcome and would be very helpful in improving our
manuscript Your contribution may assist many other
students in the coming years
31
REFERENCE
Anonim 2013 Anaerobic Respiration
httpwwwscoolcoukgcsebiologyrespiration
revise-itanaerobic-respiration Accessed
September 10th 2013
Anonim 2013 What is the difference between aerobic
and anaerobic respiration in plants
httpwwwbiologylifeeasyorg4010difference
-between-aerobic-anaerobic-respiration-plants
Accessed September 10th 2013
Campbell Mary K 2006 Biochemistry New York Thomson
Brooks Cole Inc
Dat J Capelli N Folzer H Bourgeade P Badot P-M
(2004) Sensing and signaling during
plantflooding Plant Physiology and
Biochemistry 42 273-282
Dat J Folzer H Parent C Badot P-M Capelli N
(2006) HypoxiastressCurrent Understanding and
Perspectives In Teixeira da Silva JA (Ed)
Floriculture Ornamental and Plant
Biotechnology Advances and Topical Issues (Vol
3) Global Science Books Isleworth United
Kingdompp 664-674
Hatta Muhammad 2011 Dunia Pertanian
httpemhattawordpresscomcategorystress
Accessed September 10th 2013
Hopkins William G 2009 Introduction to Plant Physiology
New York John Wiley amp Sons Inc
Lakitan Benyamin 2011 Dasar-Dasar Fisiologi Tumbuhan
Jakarta PT Raja Grafindo Persada
Larcher W 1980 Physiological Plant Ecology New York
Springer-Verlag
Raven 2013 Biology Fifth Edition
httphigheredmcgraw-hillcomsites983409233
9student_view0chapter39
photosynthetic_electron_transport_and_atp_synth
esishtml Accessed September 10th 2013
Salisbury Frank B 1969 Plant Physiology Third Edition
California Wadsworth Publishing Company
Subbaiah C Sachs M (2003) Molecular and cellular
adaptations of maize to flooding stress Annals
of Botany 91 119-127
Wilson CarlL1962 Botany Third EditionUnited State of
America Holt Rinch Inc

Oxalosucciante is changed into α-ketoglutrate by
iso-citrate dehydrogenase with the help of
coenzyme NAD or NADP In this reaction carbon
dioxide and NADH2 or NADPH2 are also released
7 Formation of succinyl CoA
α-ketoglutarate combines with acetyl CoA in the
presence of coenzyme NAD and enzyme α-
keoglutaratedehyrogenase to form succinyl coenzyme
a carbon dioxide and NADH2
8 Formation of Succinate
Later on coenzyme a is removed from succinyl
coenzyme A in the presence of guanosinediphosphate
(GDP) and inorganic phosphate to form succinate
and guanosine triphosphate (GTP) This reaction is
carried out by an enzyme called succinyl CoA
synthatase
9 Formation of Funarate
An enzyme succinic dehydrogenase removes hydrogen
from succinate to form funarate
10 Formation of Malate
Fumarate reacts with water in the presence of
enzyme fumarase to form malate
11 Regeneration of oxaloacetate
Malate is oxidized by malic dehydrogenase and NAD
forming oxaloacetate and NADH2 Thus oxalo acetate
is again available to start another cycle
9
(httpkrebs-cycle-or-citric-acid-cycle-or-
trihtml)
Importance of CTA Cycle
1 Source of energy
In addition to routine organic compounds described
above by products like nicotinanide adenine
dinucleotide (NADH2) and guanosine triphosphate
(GTP) are the source of biological energy NADH2
after oxidation produce energy whereas GTP is
itself high energy phosphate compound
2 Oxidation of organic compounds taken as food
Oxidation of fats carbohydrates and proteins take
place through it or in other words it can be said
that oxidation of all compounds having carbon
atoms can take place through TCA cycle Some of
amino acids like alanine glutamic acid and
aspartic acid at one stage or the other enters
into TCA cycle eg glutamic acid enters cycle
after its transformation into α-ketoglutarate
Similarly alanine enters the cycle after its
conversion into pyruvate
3 Intermediate compounds
TCA cycle is also involved in synthesis of
intermediate compounds leading to the formation of
larger molecules (Ray 1963 17)
10
Verification of Krebs Cycle
It was done by radioactive traces like C14 as
radioactive carbon dioxide of different levels and
reactions After addition of radioactive carbon
dioxide different chemical compounds produced like
glucose fats amino acids were isolated and looked
for radioactive carbon In this way whole of
metabolic reaction were verified including individual
reactions alternative metabolic pathways
intermediates of fats carbohydrates and amino acids
etc in the body cells as well as in test tubes
(Wilson 1962 99)
The primary function of the Krebs cycle are as
follows
1 Reduction of NAD+ and FAD to the electron donors
NADH and FADH2 that are subsequently oxidized to
yield ATP
2 Direct synthesis of a limited amount of ATP (1 ATP
for each pyruvate oxidized)
3 Formation of carbon skeletons that can be used to
synthesize certain amino acids that in turn are
converted into larger molecules
11
B Fermentation
Although glycolysis can function well without
O2 the further oxidation of pyruvate and NADH by
mitochondria requires this gas Thus when O2 is
limiting NADH and pyruvate begin to accumulate
Under this condition plants carry out fermentation
(anaerobic respiration) forming either ethanol or
lactic acid (usually ethanol) As shown in the
picture below the two top reactions consists of a
decarboxylation to form acetaldehide then rapid
reduction of acetaldehide by NADH to form ethanol
These reactions are catalyzed by alcohol
dehydrogenase Some cells contain lactic acid
12
dehydrogenase which uses NADH to reduce pyruvic to
lactic acid Thus either ethanol or lactic acid or
both are fermentation products depending on the
activities of each dehydrogenase present In each
case NADH is the reductant and only under
anaerobic conditions is it abundant anough to cause
reduction Furthermore in some plants NADH is used
to cause accumulation of the other compounds when O2
is limiting especially malate and glycerol
(Salisbury 1969 233)
The fate of pyruvate produced by glycolysis
depends primarily on whether or not oxygen is
present Under normal aerobic conditions pyruvate
is transported into the mitochondrion where it is
further oxidized to CO2 and water transferring its
electrons ultimately to molecular oxygen
13
Although higher plants are obligate aerobs and
are able to tolerate anoxia for only short periods
tissues or organs are occasionally subjected to
anaerobic conditions A typical siuation is that of
roots when the soil is saturated with water When
there is no oxygen to serve as the terminal electron
acceptor mitochondrial respiration will shut down
and metabolism will shift over to fermentation
Fermentation converts pyruvate to either ethanol
through the action of the enzyme alcohol
dehydrogenase (ADH) or lactate via lactate
dehydrogenase (LDH) In most plants the principal
products of fermentation are CO2 and ethanol Some
lactate may be formed primarily in the early stages
of anoxia However lactate lowers he pH of the
cytosol which in turn activates pyruvate
decarboxylase and initiates the production of
ethanol
Either one of the fermentation reactions
consumes the NADH produced earlier in glycolysis by
the oxidation of glyceraldehide-3-P Although this
means there is no net gain of reducing potential in
fermentation This recycling of NADH is still
important to the cell The poolof NADH plus NAD+ in
the cell is relatively small and if the NADH is not
recycled there will be no supply of NAD+ to support
14
the continued oxidation of glyceraldehyde-3-P if
this were the case glycolysis and the producton of
even the small quantities of ATP necessary to
maintain the cells under anaerobic conditions would
then grind to a halt (Hopkins 2009 241)
Soil waterlogging has long been identified as a
major abiotic stress and the constraints that it
provides the roots have a significant effect on the
growth and development of plants When this event
occurs in the spring then these puddles can reduce
seed germination and seedling development Thus a
pool of water is an important factor that affects
the growth development and survival of plant
species not only in natural ecosystems but also
the agricultural and horticultural systems (Dat et
al 2006)
After flooding rapid changes in soil
properties At the time of pore water meets the
land the air pushed out gas diffusion is reduced
and toxic compounds accumulate due to anaerobic
conditions All of these changes greatly affect the
ability of plants to survive In response increased
stomatal resistance photosynthesis and root
hydraulic conductivity decreases and reduced
translocation fotoassimilat Nevertheless one of
15
the best adaptations of plants to hypoxia anoxia
is transitional biochemical and metabolic processes
common to current limited availability of O2 (Dat et
al 2004)
Selective synthesis of a set of about 20
anaerobic stress proteins (ANPS) allows the process
of producing energy without oxygen metabolism under
anaerobic conditions (Subbaiah and Sachs 2003)
Another adaptation is the observed morphological
changes consisting of the formation of hypertrophic
lenticels adventitious root initiation and or
progression of aerenchyma (Vartapetian and Jackson
1997 Jackson and Colmer 2005 Folzer et al 2006)
This review details the diverse plant stress
response to hypoxia anoxia caused by soil
waterlogging flooding and examines some of the key
features of metabolic adaptation physiological and
morphological
16
C Electron
Transport System
We noted earlier that one of the principal
functions of the respiration is to retrieve in
useful form some of the energy initially stored in
assimilates Our traditional measure of useful
energy in most processes is the number of ATP
molecules gained or consumed By this measure alone
the yield from both glycolysis and the citric acid
cycle is quite low After two complete turns of the
cycle one molecule of glucose has been completely
oxidized to six molecules of CO2 but only four
molecules of ATP have been produced (a net of two
ATP from glycolysis plus one for each turn of the
cycle) At this point most of the energy associated
with the glucose molecule has been conserved in the
17
form of electron pairs generated by the oxidation of
glycolytic and citric acid cycle intermediates
In this section we will discuss the third stage
of cellular respirationmdashthe transfer of electrons
fromNADH and FADH2 to oxygen and the accompanying
conversion of redox energy to ATP The transfer of
electrons fromNADHand FADH2 to oxygen involves a
sequence of electron carriers arranged in an
electron transport chainMembrane fractionation
studies have shown that the enzymes and electron
carriers making up the electron transport chain are
organized predominantly into four large
multimolecular complexes (complexes ImdashIV) and two
mobile carriers located in the inner mitochondrial
membrane (Figure 109) In this sense there are a
great number of similarities between the
mitochondrial inner membrane and the thylakoid
membranes of the chloroplast (compare Figure 109
with Figure 76) This is not unexpected since the
principal function of each membrane is energy
transformation and many of the same or similar
components are involved
The Components of the Electron Transport Chain
The electron transport chain of the
mitochondria is the means by which electrons are
18
removed from the reduced carrier NADH and
transferred to oxygen to yield H2O
1) NADH
NADH is generated in the matrix by the reactions
of pyruvate dehydrogenase isocitrate
dehydrogenase α-ketoglutarate dehydrogenase and
malate dehyrogenase The electron transport chain
begins with reoxidizing NADH to form NAD+ and
channeling the electrons into the formation of
reduced coenzymes Important to note that NADH
transfers 2 electrons at a time in the form of a
hydride
NAD+ + 2e- + H+ 1048581 NADH Eorsquo = minus0315 V
2) Flavoproteins
Flavoproteins have either a FAD (flavin adenosine
dinucleotide) or a FMN (flavin mononucleotide)
prosthetic group Flavoproteins can accept or
donate electrons one at time or two at a time
Thus they are often intermediaries between two
electron acceptorsdonors and one electron
acceptorsdonors For flavoproteins the typical
standard reduction potentials are around 0 V
FAD + 2e- + 2H+ 1048581 FADH2 FMN + 2e- + 2H+ 1048581 FMNH2
Eorsquo asymp 0 V
19
3) Coenzyme Q (CoQ)
aka ubiquinone (UQ) Shown below CoQ has ten
repeating isoprene units which make it
insoluble in water but soluble in the
hydrophobic lipid bylayer Coenzyme Q is a
versatile cofactor because it is a soluble
electron carrier in the hydrophobic bilipid layer
of the
inner mitochondrial membrane Like flavoproteins
CoQ can acceptdonate electrons one at a time or
two at a time
Q + 2e- + 2H+ 1048581 QH2 Eorsquo =0060 V
Q + e- + H+ 1048581 1048581QH Eorsquo =0030 V
1048581QH + e- + H+ 1048581 QH2 Eorsquo =0190 V
4) Cytochromes
Cytochromes are proteins that contain heme
prosthetic groups which function as
one electron carriers The heme iron is involved
in one electron transfers involving the Fe2+ and
Fe3+ oxidation states Cytochromes are named by
their absorption spectra which depends
on the porphyrin structure and environment The
example shown is the heme prosthetic group of
cytochrome b Cytochrome b contains the same iron
porphyrin found in hemoglobin and myoglobin
20
Other cytochromes we will encounter in the
electron transport complexes are cytochromes b
c c1 a and a3
5) Iron-Sulfur Proteins
In the electron transport chain we will encounter
many iron-sulfer proteins which participate in
one electron transfers involving the the Fe2+ and
Fe3+ oxidation states These are non-heme iron-
sulfur proteinsThe simplest iron-sulfer protein
is FeS in which iron is tetrahedrally coordinated
by four cysteines The second form is Fe2S2 which
contains two irons complexed to 2 cysteine
residues and two inorganic sulfides The third
form is Fe3S4 which contains 3 iron atoms
coordinated to three cysteine residues and 4
inorganic sulfides The last form is the most
complicated Fe4S4 which contains 4 iron atoms
coordinated to 4 cysteine residues and 4
inorganic sulfides
6) Copper Proteins
Copper bound proteins participate in one electron
transfers involving the Cu+ and Cu2+ oxidation
states
21
Overview of the Electron Transport Chain
Electrons move along the electron transport
chain going from donor to acceptor until they reach
oxygen the ultimate electron acceptor The standard
reduction potentials of the electron carriers are
between the NADHNAD+ couple (-0315 V) and the
oxygenH2O couple (0816 V) as on the next page The
components of the electron transport chain are
organized into 4 complexes Each complex contains
several different electron carriers
1 Complex I also known as the NADH-coenzyme Q
reductase or NADH dehydrogenase
2 Complex II also known as succinate-coenzyme Q
reductase or succinate dehydrogenase
3 Complex III also known as coenzyme Q reductase
4 Complex IV also known as cytochrome c reductase
The free energy necessary to generate ATP
is extracted from the oxidation of NADH and
FADH2 by the electron transport chain It
consists of four protein complexes and single
protein chain of cytochrome c through which
electrons pass from lower to higher standard
redox potentials Electrons are carried from complex
I and II to complex III by the membrane (lipid)
soluble coenzyme Q (CoQ or ubiquinone) and between
22
complexes III and IV by the peripheral membrane
protein cytochrome c
In the following I give the overall
reactions catalysed by corresponding protein
complexes in the mitochondrial electron transport
chain
1 Complex I
Complex I is also called NADH-Coenzyme Q
reductase because this large protein complex
transfers 2 electrons from NADH to coenzyme Q
Complex I was formerly known as NADH dehydrogenase
Complex I is huge 850000 kD and is composed of more
than thirty subunits It contains a FMN prosthetic
group and seven or more Fe-S clusters This complex
has between 20-26 iron atoms boundThe prosthetic
group FMN is absolutely required for activity
Therefore this complex is a flavoprotein This
complex binds NADH transfers two electrons in the
form of a hydride to FMN to produce NAD+ and FMNH2
23
The subsequent steps involve the transfer of
electrons one at a time to a series of iron-sulfer
complexes that includes both 2Fe-2S and 4Fe-4S
clusters Note the importance of FMN First it
functions as a 2 electron acceptor in the hydride
transfer from NADH Second it functions as a 1
electron donor to the series of iron sulfur clusters
FMN and FAD often play crucial links between 2
electron transfer agents and 1 electron transfer
agents The final step of this complex is the
transfer of 2 electrons one at a time to coenzyme Q
CoQ like FMN and FAD can function as a 2 electron
donoracceptor and as a 1 electron donoracceptor
CoQ is a mobile electron carrier because its
isoprenoid tail makes it highly hydrophobic and
lipophillic It diffuses freely in the bilipid layer
of the inner mitochondrial membrane
The process of transferring electrons from NADH
to CoQ by complex I results in the net transport of
protons from the matrix side of the inner
mitochondrial membrane to the inter membrane space
where the H+ ions accumulate generating a proton
motive force The intermembrane space side of the
inner membrane is referred to as the P face (P
standing for positive) The matrix side of the inner
membrane is referred to as the the N face The
24
transport of electrons from NADH to CoQ is coupled to
the transport of protons across the membrane This is
an example of active transport The stiochiometry is
4 H+ transported per 2 electrons
2 Complex II
Believe it or not you are already familiar
with Complex II It is none other than succinate
dehydrogenase The only enzyme of the citric acid
cycle that is an integral membrane protein This
complex is composed of four subunits 2 of which are
iron-sulfur proteins and the other two subunits
together bind FAD through a covalent link to a
histidine residue These two subunits are called
flavoprotein 2 or FP2 Complex II contains 3 Fe-S
centers 1 4Fe-4S cluster 1 3Fe-4S cluster and 1
2Fe- 2S cluster In the first step of this complex
succinate is bound and a hydride is transferred to
FAD to generate FADH2 and fumarate FADH2 then
transfers its electrons one at a time to the Fe-S
centers Thus once again FAD functions as 2 electron
acceptor and a 1 electron donor The final step of
this complex is the transfer of 2 electrons one at a
time to coenzyme Q to produce CoQH2 For complex II
the standard free energy change of the overall
reaction is too small to drive the transport of
protons across the inner mitochondrial membrane This
25
accounts for the 15 ATPrsquos generated per FADH2
compared with the 25 ATPrsquos generated per NADH
3 Complex III
This complex is also known as coenzyme Q-
cytochrome c reductase because it passes the
electrons form CoQH2 to cyt c through a very unique
electron transport pathway called the Q-cycle Shown
to the left are the porphyrins found in cytochromes
Cytochrome b contains the same iron protoporphyrin as
hemoglobin and myoglobin The c cytochromes contain
heme c through covalent attachment by cysteine
residues Cytochrome a is found in two forms in
complex IV In complex III we find two b-type
cytochromes and one c-type cytochrome Complex III is
complex and we have a crystal structure
4 Complex IV
Complex IV is also known as cytochrome c oxidase
because it accepts the electrons from cytochrome c
and directs them towards the four electron reduction
of O2 to form 2 molecules of H2O
4 cyt c (Fe2+) + 4 H+ + O2 4 cyt c (Fe3+) +
2H2O
26
Cytochrome c oxidase contains 2 heme
centerscytochrome a and cytochrome a3 and two copper
proteins Each of the protein bound coppers are
associated with one of the cytochromes The copper
sites are called CuA and CuB CuA is associated with
cytochrome a and is shown to the left CuB is
associated with cytochrome a3 The copper sites
function as 1 electron carriers cycling between the
cuprous state Cu+ and the cupric state Cu2+ Just
like iron containing proteins they transfer
electrons one at a time Cytochrome c is bound on the
P-face of the membrane and transfers its electron to
CuA The oxidized cytochrome c dissociates CuA then
transfers the electron to cytochrome a The protein
bound CuA and the iron bound in cytochrome a are 15 Aring
apart In contrast the CuB and the iron bound in
cytochrome a3 are very close to each other forming a
binuclear metal center shown below
27
Electron transport chains are associated with
membranes such as the mitochondrial membrane in
eukaryotic organisms the thylakoid membrane of
photosynthetic organisms and the plasma membrane of
prokaryotic organisms While some of the details
differ electron transport chains in all organisms
operate on the same basic principles
Excited electrons are brought to the electron
transport chain by electron carriers such as NADH
and FADH2 In photosynthetic organisms the electrons
are donated by water and excited by light energy
The electron donor is oxidized as the electrons are
passed to the first protein complex in the chain As
it accepts the electrons the protein complex is
reduced Next the electrons are passed from the
28
first protein complex which becomes oxidized to
the second protein complex which becomes reduced
Each of these redox reactions is slightly exergonic
or energy releasing Some of this energy is used to
move protons across the membrane against their
concentration gradient With each successive
transfer of electrons the original excited
electrons lose some of their energy After several
successive redox reactions the electrons are
donated to a terminal electron acceptor In aerobic
respiration the terminal electron acceptor is
oxygen In photosynthesis the terminal electron
acceptor is a compound called NADP
The energy in the excited electrons has been
transformed into an electrochemical gradient
represented by the high concentration of protons on
one side of the membrane These protons can diffuse
back across the membrane by passing through a
protein complex called ATP synthase As these
protons pass through ATP synthase down their
concentration gradient they provide the energy
needed to make ATP from ADP and inorganic phosphate
within the cell
29
CHAPTER III
CLOSING
A Conclution
When a hexose is completely oxidized to CO2
and H2O using these three processes Glycolysis
yields two ATP and two NADH per hexose used Each
such NADH oxidized by the electron transport system
30
yields two ATP as dscribed above so glycolysis
contributes a total of six ATP per hexose The Krebs
cycle contributes two ATP per hexose or per two
pyruvates when succinyl CoA is cleaved to succinate
and CoASH This cycle also produces eight NADH per
hexose within the mitochondrial matrix by oxidative
phosporylation each of these NADH yields three ATP
or 24 per hexose Each FADH2 from the Krebs cycle
yields two ATP by oxydative phosporylation or four
per hexose (two pyruvates) The total contribution of
the Krebs cycle is then 30 Adding these 30 to the 6
from glycolysis leads to a total of 36 per hexose
completely respired by these process
B Suggestion
My greatest wish is that the readers find
this manuscript helpful in gaining the knowledge they
need for effective learning and practice and in
preparing for examination designed to test that
knowledge Reader comments and suggestions are
welcome and would be very helpful in improving our
manuscript Your contribution may assist many other
students in the coming years
31
REFERENCE
Anonim 2013 Anaerobic Respiration
httpwwwscoolcoukgcsebiologyrespiration
revise-itanaerobic-respiration Accessed
September 10th 2013
Anonim 2013 What is the difference between aerobic
and anaerobic respiration in plants
httpwwwbiologylifeeasyorg4010difference
-between-aerobic-anaerobic-respiration-plants
Accessed September 10th 2013
Campbell Mary K 2006 Biochemistry New York Thomson
Brooks Cole Inc
Dat J Capelli N Folzer H Bourgeade P Badot P-M
(2004) Sensing and signaling during
plantflooding Plant Physiology and
Biochemistry 42 273-282
Dat J Folzer H Parent C Badot P-M Capelli N
(2006) HypoxiastressCurrent Understanding and
Perspectives In Teixeira da Silva JA (Ed)
Floriculture Ornamental and Plant
Biotechnology Advances and Topical Issues (Vol
3) Global Science Books Isleworth United
Kingdompp 664-674
Hatta Muhammad 2011 Dunia Pertanian
httpemhattawordpresscomcategorystress
Accessed September 10th 2013
Hopkins William G 2009 Introduction to Plant Physiology
New York John Wiley amp Sons Inc
Lakitan Benyamin 2011 Dasar-Dasar Fisiologi Tumbuhan
Jakarta PT Raja Grafindo Persada
Larcher W 1980 Physiological Plant Ecology New York
Springer-Verlag
Raven 2013 Biology Fifth Edition
httphigheredmcgraw-hillcomsites983409233
9student_view0chapter39
photosynthetic_electron_transport_and_atp_synth
esishtml Accessed September 10th 2013
Salisbury Frank B 1969 Plant Physiology Third Edition
California Wadsworth Publishing Company
Subbaiah C Sachs M (2003) Molecular and cellular
adaptations of maize to flooding stress Annals
of Botany 91 119-127
Wilson CarlL1962 Botany Third EditionUnited State of
America Holt Rinch Inc

(httpkrebs-cycle-or-citric-acid-cycle-or-
trihtml)
Importance of CTA Cycle
1 Source of energy
In addition to routine organic compounds described
above by products like nicotinanide adenine
dinucleotide (NADH2) and guanosine triphosphate
(GTP) are the source of biological energy NADH2
after oxidation produce energy whereas GTP is
itself high energy phosphate compound
2 Oxidation of organic compounds taken as food
Oxidation of fats carbohydrates and proteins take
place through it or in other words it can be said
that oxidation of all compounds having carbon
atoms can take place through TCA cycle Some of
amino acids like alanine glutamic acid and
aspartic acid at one stage or the other enters
into TCA cycle eg glutamic acid enters cycle
after its transformation into α-ketoglutarate
Similarly alanine enters the cycle after its
conversion into pyruvate
3 Intermediate compounds
TCA cycle is also involved in synthesis of
intermediate compounds leading to the formation of
larger molecules (Ray 1963 17)
10
Verification of Krebs Cycle
It was done by radioactive traces like C14 as
radioactive carbon dioxide of different levels and
reactions After addition of radioactive carbon
dioxide different chemical compounds produced like
glucose fats amino acids were isolated and looked
for radioactive carbon In this way whole of
metabolic reaction were verified including individual
reactions alternative metabolic pathways
intermediates of fats carbohydrates and amino acids
etc in the body cells as well as in test tubes
(Wilson 1962 99)
The primary function of the Krebs cycle are as
follows
1 Reduction of NAD+ and FAD to the electron donors
NADH and FADH2 that are subsequently oxidized to
yield ATP
2 Direct synthesis of a limited amount of ATP (1 ATP
for each pyruvate oxidized)
3 Formation of carbon skeletons that can be used to
synthesize certain amino acids that in turn are
converted into larger molecules
11
B Fermentation
Although glycolysis can function well without
O2 the further oxidation of pyruvate and NADH by
mitochondria requires this gas Thus when O2 is
limiting NADH and pyruvate begin to accumulate
Under this condition plants carry out fermentation
(anaerobic respiration) forming either ethanol or
lactic acid (usually ethanol) As shown in the
picture below the two top reactions consists of a
decarboxylation to form acetaldehide then rapid
reduction of acetaldehide by NADH to form ethanol
These reactions are catalyzed by alcohol
dehydrogenase Some cells contain lactic acid
12
dehydrogenase which uses NADH to reduce pyruvic to
lactic acid Thus either ethanol or lactic acid or
both are fermentation products depending on the
activities of each dehydrogenase present In each
case NADH is the reductant and only under
anaerobic conditions is it abundant anough to cause
reduction Furthermore in some plants NADH is used
to cause accumulation of the other compounds when O2
is limiting especially malate and glycerol
(Salisbury 1969 233)
The fate of pyruvate produced by glycolysis
depends primarily on whether or not oxygen is
present Under normal aerobic conditions pyruvate
is transported into the mitochondrion where it is
further oxidized to CO2 and water transferring its
electrons ultimately to molecular oxygen
13
Although higher plants are obligate aerobs and
are able to tolerate anoxia for only short periods
tissues or organs are occasionally subjected to
anaerobic conditions A typical siuation is that of
roots when the soil is saturated with water When
there is no oxygen to serve as the terminal electron
acceptor mitochondrial respiration will shut down
and metabolism will shift over to fermentation
Fermentation converts pyruvate to either ethanol
through the action of the enzyme alcohol
dehydrogenase (ADH) or lactate via lactate
dehydrogenase (LDH) In most plants the principal
products of fermentation are CO2 and ethanol Some
lactate may be formed primarily in the early stages
of anoxia However lactate lowers he pH of the
cytosol which in turn activates pyruvate
decarboxylase and initiates the production of
ethanol
Either one of the fermentation reactions
consumes the NADH produced earlier in glycolysis by
the oxidation of glyceraldehide-3-P Although this
means there is no net gain of reducing potential in
fermentation This recycling of NADH is still
important to the cell The poolof NADH plus NAD+ in
the cell is relatively small and if the NADH is not
recycled there will be no supply of NAD+ to support
14
the continued oxidation of glyceraldehyde-3-P if
this were the case glycolysis and the producton of
even the small quantities of ATP necessary to
maintain the cells under anaerobic conditions would
then grind to a halt (Hopkins 2009 241)
Soil waterlogging has long been identified as a
major abiotic stress and the constraints that it
provides the roots have a significant effect on the
growth and development of plants When this event
occurs in the spring then these puddles can reduce
seed germination and seedling development Thus a
pool of water is an important factor that affects
the growth development and survival of plant
species not only in natural ecosystems but also
the agricultural and horticultural systems (Dat et
al 2006)
After flooding rapid changes in soil
properties At the time of pore water meets the
land the air pushed out gas diffusion is reduced
and toxic compounds accumulate due to anaerobic
conditions All of these changes greatly affect the
ability of plants to survive In response increased
stomatal resistance photosynthesis and root
hydraulic conductivity decreases and reduced
translocation fotoassimilat Nevertheless one of
15
the best adaptations of plants to hypoxia anoxia
is transitional biochemical and metabolic processes
common to current limited availability of O2 (Dat et
al 2004)
Selective synthesis of a set of about 20
anaerobic stress proteins (ANPS) allows the process
of producing energy without oxygen metabolism under
anaerobic conditions (Subbaiah and Sachs 2003)
Another adaptation is the observed morphological
changes consisting of the formation of hypertrophic
lenticels adventitious root initiation and or
progression of aerenchyma (Vartapetian and Jackson
1997 Jackson and Colmer 2005 Folzer et al 2006)
This review details the diverse plant stress
response to hypoxia anoxia caused by soil
waterlogging flooding and examines some of the key
features of metabolic adaptation physiological and
morphological
16
C Electron
Transport System
We noted earlier that one of the principal
functions of the respiration is to retrieve in
useful form some of the energy initially stored in
assimilates Our traditional measure of useful
energy in most processes is the number of ATP
molecules gained or consumed By this measure alone
the yield from both glycolysis and the citric acid
cycle is quite low After two complete turns of the
cycle one molecule of glucose has been completely
oxidized to six molecules of CO2 but only four
molecules of ATP have been produced (a net of two
ATP from glycolysis plus one for each turn of the
cycle) At this point most of the energy associated
with the glucose molecule has been conserved in the
17
form of electron pairs generated by the oxidation of
glycolytic and citric acid cycle intermediates
In this section we will discuss the third stage
of cellular respirationmdashthe transfer of electrons
fromNADH and FADH2 to oxygen and the accompanying
conversion of redox energy to ATP The transfer of
electrons fromNADHand FADH2 to oxygen involves a
sequence of electron carriers arranged in an
electron transport chainMembrane fractionation
studies have shown that the enzymes and electron
carriers making up the electron transport chain are
organized predominantly into four large
multimolecular complexes (complexes ImdashIV) and two
mobile carriers located in the inner mitochondrial
membrane (Figure 109) In this sense there are a
great number of similarities between the
mitochondrial inner membrane and the thylakoid
membranes of the chloroplast (compare Figure 109
with Figure 76) This is not unexpected since the
principal function of each membrane is energy
transformation and many of the same or similar
components are involved
The Components of the Electron Transport Chain
The electron transport chain of the
mitochondria is the means by which electrons are
18
removed from the reduced carrier NADH and
transferred to oxygen to yield H2O
1) NADH
NADH is generated in the matrix by the reactions
of pyruvate dehydrogenase isocitrate
dehydrogenase α-ketoglutarate dehydrogenase and
malate dehyrogenase The electron transport chain
begins with reoxidizing NADH to form NAD+ and
channeling the electrons into the formation of
reduced coenzymes Important to note that NADH
transfers 2 electrons at a time in the form of a
hydride
NAD+ + 2e- + H+ 1048581 NADH Eorsquo = minus0315 V
2) Flavoproteins
Flavoproteins have either a FAD (flavin adenosine
dinucleotide) or a FMN (flavin mononucleotide)
prosthetic group Flavoproteins can accept or
donate electrons one at time or two at a time
Thus they are often intermediaries between two
electron acceptorsdonors and one electron
acceptorsdonors For flavoproteins the typical
standard reduction potentials are around 0 V
FAD + 2e- + 2H+ 1048581 FADH2 FMN + 2e- + 2H+ 1048581 FMNH2
Eorsquo asymp 0 V
19
3) Coenzyme Q (CoQ)
aka ubiquinone (UQ) Shown below CoQ has ten
repeating isoprene units which make it
insoluble in water but soluble in the
hydrophobic lipid bylayer Coenzyme Q is a
versatile cofactor because it is a soluble
electron carrier in the hydrophobic bilipid layer
of the
inner mitochondrial membrane Like flavoproteins
CoQ can acceptdonate electrons one at a time or
two at a time
Q + 2e- + 2H+ 1048581 QH2 Eorsquo =0060 V
Q + e- + H+ 1048581 1048581QH Eorsquo =0030 V
1048581QH + e- + H+ 1048581 QH2 Eorsquo =0190 V
4) Cytochromes
Cytochromes are proteins that contain heme
prosthetic groups which function as
one electron carriers The heme iron is involved
in one electron transfers involving the Fe2+ and
Fe3+ oxidation states Cytochromes are named by
their absorption spectra which depends
on the porphyrin structure and environment The
example shown is the heme prosthetic group of
cytochrome b Cytochrome b contains the same iron
porphyrin found in hemoglobin and myoglobin
20
Other cytochromes we will encounter in the
electron transport complexes are cytochromes b
c c1 a and a3
5) Iron-Sulfur Proteins
In the electron transport chain we will encounter
many iron-sulfer proteins which participate in
one electron transfers involving the the Fe2+ and
Fe3+ oxidation states These are non-heme iron-
sulfur proteinsThe simplest iron-sulfer protein
is FeS in which iron is tetrahedrally coordinated
by four cysteines The second form is Fe2S2 which
contains two irons complexed to 2 cysteine
residues and two inorganic sulfides The third
form is Fe3S4 which contains 3 iron atoms
coordinated to three cysteine residues and 4
inorganic sulfides The last form is the most
complicated Fe4S4 which contains 4 iron atoms
coordinated to 4 cysteine residues and 4
inorganic sulfides
6) Copper Proteins
Copper bound proteins participate in one electron
transfers involving the Cu+ and Cu2+ oxidation
states
21
Overview of the Electron Transport Chain
Electrons move along the electron transport
chain going from donor to acceptor until they reach
oxygen the ultimate electron acceptor The standard
reduction potentials of the electron carriers are
between the NADHNAD+ couple (-0315 V) and the
oxygenH2O couple (0816 V) as on the next page The
components of the electron transport chain are
organized into 4 complexes Each complex contains
several different electron carriers
1 Complex I also known as the NADH-coenzyme Q
reductase or NADH dehydrogenase
2 Complex II also known as succinate-coenzyme Q
reductase or succinate dehydrogenase
3 Complex III also known as coenzyme Q reductase
4 Complex IV also known as cytochrome c reductase
The free energy necessary to generate ATP
is extracted from the oxidation of NADH and
FADH2 by the electron transport chain It
consists of four protein complexes and single
protein chain of cytochrome c through which
electrons pass from lower to higher standard
redox potentials Electrons are carried from complex
I and II to complex III by the membrane (lipid)
soluble coenzyme Q (CoQ or ubiquinone) and between
22
complexes III and IV by the peripheral membrane
protein cytochrome c
In the following I give the overall
reactions catalysed by corresponding protein
complexes in the mitochondrial electron transport
chain
1 Complex I
Complex I is also called NADH-Coenzyme Q
reductase because this large protein complex
transfers 2 electrons from NADH to coenzyme Q
Complex I was formerly known as NADH dehydrogenase
Complex I is huge 850000 kD and is composed of more
than thirty subunits It contains a FMN prosthetic
group and seven or more Fe-S clusters This complex
has between 20-26 iron atoms boundThe prosthetic
group FMN is absolutely required for activity
Therefore this complex is a flavoprotein This
complex binds NADH transfers two electrons in the
form of a hydride to FMN to produce NAD+ and FMNH2
23
The subsequent steps involve the transfer of
electrons one at a time to a series of iron-sulfer
complexes that includes both 2Fe-2S and 4Fe-4S
clusters Note the importance of FMN First it
functions as a 2 electron acceptor in the hydride
transfer from NADH Second it functions as a 1
electron donor to the series of iron sulfur clusters
FMN and FAD often play crucial links between 2
electron transfer agents and 1 electron transfer
agents The final step of this complex is the
transfer of 2 electrons one at a time to coenzyme Q
CoQ like FMN and FAD can function as a 2 electron
donoracceptor and as a 1 electron donoracceptor
CoQ is a mobile electron carrier because its
isoprenoid tail makes it highly hydrophobic and
lipophillic It diffuses freely in the bilipid layer
of the inner mitochondrial membrane
The process of transferring electrons from NADH
to CoQ by complex I results in the net transport of
protons from the matrix side of the inner
mitochondrial membrane to the inter membrane space
where the H+ ions accumulate generating a proton
motive force The intermembrane space side of the
inner membrane is referred to as the P face (P
standing for positive) The matrix side of the inner
membrane is referred to as the the N face The
24
transport of electrons from NADH to CoQ is coupled to
the transport of protons across the membrane This is
an example of active transport The stiochiometry is
4 H+ transported per 2 electrons
2 Complex II
Believe it or not you are already familiar
with Complex II It is none other than succinate
dehydrogenase The only enzyme of the citric acid
cycle that is an integral membrane protein This
complex is composed of four subunits 2 of which are
iron-sulfur proteins and the other two subunits
together bind FAD through a covalent link to a
histidine residue These two subunits are called
flavoprotein 2 or FP2 Complex II contains 3 Fe-S
centers 1 4Fe-4S cluster 1 3Fe-4S cluster and 1
2Fe- 2S cluster In the first step of this complex
succinate is bound and a hydride is transferred to
FAD to generate FADH2 and fumarate FADH2 then
transfers its electrons one at a time to the Fe-S
centers Thus once again FAD functions as 2 electron
acceptor and a 1 electron donor The final step of
this complex is the transfer of 2 electrons one at a
time to coenzyme Q to produce CoQH2 For complex II
the standard free energy change of the overall
reaction is too small to drive the transport of
protons across the inner mitochondrial membrane This
25
accounts for the 15 ATPrsquos generated per FADH2
compared with the 25 ATPrsquos generated per NADH
3 Complex III
This complex is also known as coenzyme Q-
cytochrome c reductase because it passes the
electrons form CoQH2 to cyt c through a very unique
electron transport pathway called the Q-cycle Shown
to the left are the porphyrins found in cytochromes
Cytochrome b contains the same iron protoporphyrin as
hemoglobin and myoglobin The c cytochromes contain
heme c through covalent attachment by cysteine
residues Cytochrome a is found in two forms in
complex IV In complex III we find two b-type
cytochromes and one c-type cytochrome Complex III is
complex and we have a crystal structure
4 Complex IV
Complex IV is also known as cytochrome c oxidase
because it accepts the electrons from cytochrome c
and directs them towards the four electron reduction
of O2 to form 2 molecules of H2O
4 cyt c (Fe2+) + 4 H+ + O2 4 cyt c (Fe3+) +
2H2O
26
Cytochrome c oxidase contains 2 heme
centerscytochrome a and cytochrome a3 and two copper
proteins Each of the protein bound coppers are
associated with one of the cytochromes The copper
sites are called CuA and CuB CuA is associated with
cytochrome a and is shown to the left CuB is
associated with cytochrome a3 The copper sites
function as 1 electron carriers cycling between the
cuprous state Cu+ and the cupric state Cu2+ Just
like iron containing proteins they transfer
electrons one at a time Cytochrome c is bound on the
P-face of the membrane and transfers its electron to
CuA The oxidized cytochrome c dissociates CuA then
transfers the electron to cytochrome a The protein
bound CuA and the iron bound in cytochrome a are 15 Aring
apart In contrast the CuB and the iron bound in
cytochrome a3 are very close to each other forming a
binuclear metal center shown below
27
Electron transport chains are associated with
membranes such as the mitochondrial membrane in
eukaryotic organisms the thylakoid membrane of
photosynthetic organisms and the plasma membrane of
prokaryotic organisms While some of the details
differ electron transport chains in all organisms
operate on the same basic principles
Excited electrons are brought to the electron
transport chain by electron carriers such as NADH
and FADH2 In photosynthetic organisms the electrons
are donated by water and excited by light energy
The electron donor is oxidized as the electrons are
passed to the first protein complex in the chain As
it accepts the electrons the protein complex is
reduced Next the electrons are passed from the
28
first protein complex which becomes oxidized to
the second protein complex which becomes reduced
Each of these redox reactions is slightly exergonic
or energy releasing Some of this energy is used to
move protons across the membrane against their
concentration gradient With each successive
transfer of electrons the original excited
electrons lose some of their energy After several
successive redox reactions the electrons are
donated to a terminal electron acceptor In aerobic
respiration the terminal electron acceptor is
oxygen In photosynthesis the terminal electron
acceptor is a compound called NADP
The energy in the excited electrons has been
transformed into an electrochemical gradient
represented by the high concentration of protons on
one side of the membrane These protons can diffuse
back across the membrane by passing through a
protein complex called ATP synthase As these
protons pass through ATP synthase down their
concentration gradient they provide the energy
needed to make ATP from ADP and inorganic phosphate
within the cell
29
CHAPTER III
CLOSING
A Conclution
When a hexose is completely oxidized to CO2
and H2O using these three processes Glycolysis
yields two ATP and two NADH per hexose used Each
such NADH oxidized by the electron transport system
30
yields two ATP as dscribed above so glycolysis
contributes a total of six ATP per hexose The Krebs
cycle contributes two ATP per hexose or per two
pyruvates when succinyl CoA is cleaved to succinate
and CoASH This cycle also produces eight NADH per
hexose within the mitochondrial matrix by oxidative
phosporylation each of these NADH yields three ATP
or 24 per hexose Each FADH2 from the Krebs cycle
yields two ATP by oxydative phosporylation or four
per hexose (two pyruvates) The total contribution of
the Krebs cycle is then 30 Adding these 30 to the 6
from glycolysis leads to a total of 36 per hexose
completely respired by these process
B Suggestion
My greatest wish is that the readers find
this manuscript helpful in gaining the knowledge they
need for effective learning and practice and in
preparing for examination designed to test that
knowledge Reader comments and suggestions are
welcome and would be very helpful in improving our
manuscript Your contribution may assist many other
students in the coming years
31
REFERENCE
Anonim 2013 Anaerobic Respiration
httpwwwscoolcoukgcsebiologyrespiration
revise-itanaerobic-respiration Accessed
September 10th 2013
Anonim 2013 What is the difference between aerobic
and anaerobic respiration in plants
httpwwwbiologylifeeasyorg4010difference
-between-aerobic-anaerobic-respiration-plants
Accessed September 10th 2013
Campbell Mary K 2006 Biochemistry New York Thomson
Brooks Cole Inc
Dat J Capelli N Folzer H Bourgeade P Badot P-M
(2004) Sensing and signaling during
plantflooding Plant Physiology and
Biochemistry 42 273-282
Dat J Folzer H Parent C Badot P-M Capelli N
(2006) HypoxiastressCurrent Understanding and
Perspectives In Teixeira da Silva JA (Ed)
Floriculture Ornamental and Plant
Biotechnology Advances and Topical Issues (Vol
3) Global Science Books Isleworth United
Kingdompp 664-674
Hatta Muhammad 2011 Dunia Pertanian
httpemhattawordpresscomcategorystress
Accessed September 10th 2013
Hopkins William G 2009 Introduction to Plant Physiology
New York John Wiley amp Sons Inc
Lakitan Benyamin 2011 Dasar-Dasar Fisiologi Tumbuhan
Jakarta PT Raja Grafindo Persada
Larcher W 1980 Physiological Plant Ecology New York
Springer-Verlag
Raven 2013 Biology Fifth Edition
httphigheredmcgraw-hillcomsites983409233
9student_view0chapter39
photosynthetic_electron_transport_and_atp_synth
esishtml Accessed September 10th 2013
Salisbury Frank B 1969 Plant Physiology Third Edition
California Wadsworth Publishing Company
Subbaiah C Sachs M (2003) Molecular and cellular
adaptations of maize to flooding stress Annals
of Botany 91 119-127
Wilson CarlL1962 Botany Third EditionUnited State of
America Holt Rinch Inc

Verification of Krebs Cycle
It was done by radioactive traces like C14 as
radioactive carbon dioxide of different levels and
reactions After addition of radioactive carbon
dioxide different chemical compounds produced like
glucose fats amino acids were isolated and looked
for radioactive carbon In this way whole of
metabolic reaction were verified including individual
reactions alternative metabolic pathways
intermediates of fats carbohydrates and amino acids
etc in the body cells as well as in test tubes
(Wilson 1962 99)
The primary function of the Krebs cycle are as
follows
1 Reduction of NAD+ and FAD to the electron donors
NADH and FADH2 that are subsequently oxidized to
yield ATP
2 Direct synthesis of a limited amount of ATP (1 ATP
for each pyruvate oxidized)
3 Formation of carbon skeletons that can be used to
synthesize certain amino acids that in turn are
converted into larger molecules
11
B Fermentation
Although glycolysis can function well without
O2 the further oxidation of pyruvate and NADH by
mitochondria requires this gas Thus when O2 is
limiting NADH and pyruvate begin to accumulate
Under this condition plants carry out fermentation
(anaerobic respiration) forming either ethanol or
lactic acid (usually ethanol) As shown in the
picture below the two top reactions consists of a
decarboxylation to form acetaldehide then rapid
reduction of acetaldehide by NADH to form ethanol
These reactions are catalyzed by alcohol
dehydrogenase Some cells contain lactic acid
12
dehydrogenase which uses NADH to reduce pyruvic to
lactic acid Thus either ethanol or lactic acid or
both are fermentation products depending on the
activities of each dehydrogenase present In each
case NADH is the reductant and only under
anaerobic conditions is it abundant anough to cause
reduction Furthermore in some plants NADH is used
to cause accumulation of the other compounds when O2
is limiting especially malate and glycerol
(Salisbury 1969 233)
The fate of pyruvate produced by glycolysis
depends primarily on whether or not oxygen is
present Under normal aerobic conditions pyruvate
is transported into the mitochondrion where it is
further oxidized to CO2 and water transferring its
electrons ultimately to molecular oxygen
13
Although higher plants are obligate aerobs and
are able to tolerate anoxia for only short periods
tissues or organs are occasionally subjected to
anaerobic conditions A typical siuation is that of
roots when the soil is saturated with water When
there is no oxygen to serve as the terminal electron
acceptor mitochondrial respiration will shut down
and metabolism will shift over to fermentation
Fermentation converts pyruvate to either ethanol
through the action of the enzyme alcohol
dehydrogenase (ADH) or lactate via lactate
dehydrogenase (LDH) In most plants the principal
products of fermentation are CO2 and ethanol Some
lactate may be formed primarily in the early stages
of anoxia However lactate lowers he pH of the
cytosol which in turn activates pyruvate
decarboxylase and initiates the production of
ethanol
Either one of the fermentation reactions
consumes the NADH produced earlier in glycolysis by
the oxidation of glyceraldehide-3-P Although this
means there is no net gain of reducing potential in
fermentation This recycling of NADH is still
important to the cell The poolof NADH plus NAD+ in
the cell is relatively small and if the NADH is not
recycled there will be no supply of NAD+ to support
14
the continued oxidation of glyceraldehyde-3-P if
this were the case glycolysis and the producton of
even the small quantities of ATP necessary to
maintain the cells under anaerobic conditions would
then grind to a halt (Hopkins 2009 241)
Soil waterlogging has long been identified as a
major abiotic stress and the constraints that it
provides the roots have a significant effect on the
growth and development of plants When this event
occurs in the spring then these puddles can reduce
seed germination and seedling development Thus a
pool of water is an important factor that affects
the growth development and survival of plant
species not only in natural ecosystems but also
the agricultural and horticultural systems (Dat et
al 2006)
After flooding rapid changes in soil
properties At the time of pore water meets the
land the air pushed out gas diffusion is reduced
and toxic compounds accumulate due to anaerobic
conditions All of these changes greatly affect the
ability of plants to survive In response increased
stomatal resistance photosynthesis and root
hydraulic conductivity decreases and reduced
translocation fotoassimilat Nevertheless one of
15
the best adaptations of plants to hypoxia anoxia
is transitional biochemical and metabolic processes
common to current limited availability of O2 (Dat et
al 2004)
Selective synthesis of a set of about 20
anaerobic stress proteins (ANPS) allows the process
of producing energy without oxygen metabolism under
anaerobic conditions (Subbaiah and Sachs 2003)
Another adaptation is the observed morphological
changes consisting of the formation of hypertrophic
lenticels adventitious root initiation and or
progression of aerenchyma (Vartapetian and Jackson
1997 Jackson and Colmer 2005 Folzer et al 2006)
This review details the diverse plant stress
response to hypoxia anoxia caused by soil
waterlogging flooding and examines some of the key
features of metabolic adaptation physiological and
morphological
16
C Electron
Transport System
We noted earlier that one of the principal
functions of the respiration is to retrieve in
useful form some of the energy initially stored in
assimilates Our traditional measure of useful
energy in most processes is the number of ATP
molecules gained or consumed By this measure alone
the yield from both glycolysis and the citric acid
cycle is quite low After two complete turns of the
cycle one molecule of glucose has been completely
oxidized to six molecules of CO2 but only four
molecules of ATP have been produced (a net of two
ATP from glycolysis plus one for each turn of the
cycle) At this point most of the energy associated
with the glucose molecule has been conserved in the
17
form of electron pairs generated by the oxidation of
glycolytic and citric acid cycle intermediates
In this section we will discuss the third stage
of cellular respirationmdashthe transfer of electrons
fromNADH and FADH2 to oxygen and the accompanying
conversion of redox energy to ATP The transfer of
electrons fromNADHand FADH2 to oxygen involves a
sequence of electron carriers arranged in an
electron transport chainMembrane fractionation
studies have shown that the enzymes and electron
carriers making up the electron transport chain are
organized predominantly into four large
multimolecular complexes (complexes ImdashIV) and two
mobile carriers located in the inner mitochondrial
membrane (Figure 109) In this sense there are a
great number of similarities between the
mitochondrial inner membrane and the thylakoid
membranes of the chloroplast (compare Figure 109
with Figure 76) This is not unexpected since the
principal function of each membrane is energy
transformation and many of the same or similar
components are involved
The Components of the Electron Transport Chain
The electron transport chain of the
mitochondria is the means by which electrons are
18
removed from the reduced carrier NADH and
transferred to oxygen to yield H2O
1) NADH
NADH is generated in the matrix by the reactions
of pyruvate dehydrogenase isocitrate
dehydrogenase α-ketoglutarate dehydrogenase and
malate dehyrogenase The electron transport chain
begins with reoxidizing NADH to form NAD+ and
channeling the electrons into the formation of
reduced coenzymes Important to note that NADH
transfers 2 electrons at a time in the form of a
hydride
NAD+ + 2e- + H+ 1048581 NADH Eorsquo = minus0315 V
2) Flavoproteins
Flavoproteins have either a FAD (flavin adenosine
dinucleotide) or a FMN (flavin mononucleotide)
prosthetic group Flavoproteins can accept or
donate electrons one at time or two at a time
Thus they are often intermediaries between two
electron acceptorsdonors and one electron
acceptorsdonors For flavoproteins the typical
standard reduction potentials are around 0 V
FAD + 2e- + 2H+ 1048581 FADH2 FMN + 2e- + 2H+ 1048581 FMNH2
Eorsquo asymp 0 V
19
3) Coenzyme Q (CoQ)
aka ubiquinone (UQ) Shown below CoQ has ten
repeating isoprene units which make it
insoluble in water but soluble in the
hydrophobic lipid bylayer Coenzyme Q is a
versatile cofactor because it is a soluble
electron carrier in the hydrophobic bilipid layer
of the
inner mitochondrial membrane Like flavoproteins
CoQ can acceptdonate electrons one at a time or
two at a time
Q + 2e- + 2H+ 1048581 QH2 Eorsquo =0060 V
Q + e- + H+ 1048581 1048581QH Eorsquo =0030 V
1048581QH + e- + H+ 1048581 QH2 Eorsquo =0190 V
4) Cytochromes
Cytochromes are proteins that contain heme
prosthetic groups which function as
one electron carriers The heme iron is involved
in one electron transfers involving the Fe2+ and
Fe3+ oxidation states Cytochromes are named by
their absorption spectra which depends
on the porphyrin structure and environment The
example shown is the heme prosthetic group of
cytochrome b Cytochrome b contains the same iron
porphyrin found in hemoglobin and myoglobin
20
Other cytochromes we will encounter in the
electron transport complexes are cytochromes b
c c1 a and a3
5) Iron-Sulfur Proteins
In the electron transport chain we will encounter
many iron-sulfer proteins which participate in
one electron transfers involving the the Fe2+ and
Fe3+ oxidation states These are non-heme iron-
sulfur proteinsThe simplest iron-sulfer protein
is FeS in which iron is tetrahedrally coordinated
by four cysteines The second form is Fe2S2 which
contains two irons complexed to 2 cysteine
residues and two inorganic sulfides The third
form is Fe3S4 which contains 3 iron atoms
coordinated to three cysteine residues and 4
inorganic sulfides The last form is the most
complicated Fe4S4 which contains 4 iron atoms
coordinated to 4 cysteine residues and 4
inorganic sulfides
6) Copper Proteins
Copper bound proteins participate in one electron
transfers involving the Cu+ and Cu2+ oxidation
states
21
Overview of the Electron Transport Chain
Electrons move along the electron transport
chain going from donor to acceptor until they reach
oxygen the ultimate electron acceptor The standard
reduction potentials of the electron carriers are
between the NADHNAD+ couple (-0315 V) and the
oxygenH2O couple (0816 V) as on the next page The
components of the electron transport chain are
organized into 4 complexes Each complex contains
several different electron carriers
1 Complex I also known as the NADH-coenzyme Q
reductase or NADH dehydrogenase
2 Complex II also known as succinate-coenzyme Q
reductase or succinate dehydrogenase
3 Complex III also known as coenzyme Q reductase
4 Complex IV also known as cytochrome c reductase
The free energy necessary to generate ATP
is extracted from the oxidation of NADH and
FADH2 by the electron transport chain It
consists of four protein complexes and single
protein chain of cytochrome c through which
electrons pass from lower to higher standard
redox potentials Electrons are carried from complex
I and II to complex III by the membrane (lipid)
soluble coenzyme Q (CoQ or ubiquinone) and between
22
complexes III and IV by the peripheral membrane
protein cytochrome c
In the following I give the overall
reactions catalysed by corresponding protein
complexes in the mitochondrial electron transport
chain
1 Complex I
Complex I is also called NADH-Coenzyme Q
reductase because this large protein complex
transfers 2 electrons from NADH to coenzyme Q
Complex I was formerly known as NADH dehydrogenase
Complex I is huge 850000 kD and is composed of more
than thirty subunits It contains a FMN prosthetic
group and seven or more Fe-S clusters This complex
has between 20-26 iron atoms boundThe prosthetic
group FMN is absolutely required for activity
Therefore this complex is a flavoprotein This
complex binds NADH transfers two electrons in the
form of a hydride to FMN to produce NAD+ and FMNH2
23
The subsequent steps involve the transfer of
electrons one at a time to a series of iron-sulfer
complexes that includes both 2Fe-2S and 4Fe-4S
clusters Note the importance of FMN First it
functions as a 2 electron acceptor in the hydride
transfer from NADH Second it functions as a 1
electron donor to the series of iron sulfur clusters
FMN and FAD often play crucial links between 2
electron transfer agents and 1 electron transfer
agents The final step of this complex is the
transfer of 2 electrons one at a time to coenzyme Q
CoQ like FMN and FAD can function as a 2 electron
donoracceptor and as a 1 electron donoracceptor
CoQ is a mobile electron carrier because its
isoprenoid tail makes it highly hydrophobic and
lipophillic It diffuses freely in the bilipid layer
of the inner mitochondrial membrane
The process of transferring electrons from NADH
to CoQ by complex I results in the net transport of
protons from the matrix side of the inner
mitochondrial membrane to the inter membrane space
where the H+ ions accumulate generating a proton
motive force The intermembrane space side of the
inner membrane is referred to as the P face (P
standing for positive) The matrix side of the inner
membrane is referred to as the the N face The
24
transport of electrons from NADH to CoQ is coupled to
the transport of protons across the membrane This is
an example of active transport The stiochiometry is
4 H+ transported per 2 electrons
2 Complex II
Believe it or not you are already familiar
with Complex II It is none other than succinate
dehydrogenase The only enzyme of the citric acid
cycle that is an integral membrane protein This
complex is composed of four subunits 2 of which are
iron-sulfur proteins and the other two subunits
together bind FAD through a covalent link to a
histidine residue These two subunits are called
flavoprotein 2 or FP2 Complex II contains 3 Fe-S
centers 1 4Fe-4S cluster 1 3Fe-4S cluster and 1
2Fe- 2S cluster In the first step of this complex
succinate is bound and a hydride is transferred to
FAD to generate FADH2 and fumarate FADH2 then
transfers its electrons one at a time to the Fe-S
centers Thus once again FAD functions as 2 electron
acceptor and a 1 electron donor The final step of
this complex is the transfer of 2 electrons one at a
time to coenzyme Q to produce CoQH2 For complex II
the standard free energy change of the overall
reaction is too small to drive the transport of
protons across the inner mitochondrial membrane This
25
accounts for the 15 ATPrsquos generated per FADH2
compared with the 25 ATPrsquos generated per NADH
3 Complex III
This complex is also known as coenzyme Q-
cytochrome c reductase because it passes the
electrons form CoQH2 to cyt c through a very unique
electron transport pathway called the Q-cycle Shown
to the left are the porphyrins found in cytochromes
Cytochrome b contains the same iron protoporphyrin as
hemoglobin and myoglobin The c cytochromes contain
heme c through covalent attachment by cysteine
residues Cytochrome a is found in two forms in
complex IV In complex III we find two b-type
cytochromes and one c-type cytochrome Complex III is
complex and we have a crystal structure
4 Complex IV
Complex IV is also known as cytochrome c oxidase
because it accepts the electrons from cytochrome c
and directs them towards the four electron reduction
of O2 to form 2 molecules of H2O
4 cyt c (Fe2+) + 4 H+ + O2 4 cyt c (Fe3+) +
2H2O
26
Cytochrome c oxidase contains 2 heme
centerscytochrome a and cytochrome a3 and two copper
proteins Each of the protein bound coppers are
associated with one of the cytochromes The copper
sites are called CuA and CuB CuA is associated with
cytochrome a and is shown to the left CuB is
associated with cytochrome a3 The copper sites
function as 1 electron carriers cycling between the
cuprous state Cu+ and the cupric state Cu2+ Just
like iron containing proteins they transfer
electrons one at a time Cytochrome c is bound on the
P-face of the membrane and transfers its electron to
CuA The oxidized cytochrome c dissociates CuA then
transfers the electron to cytochrome a The protein
bound CuA and the iron bound in cytochrome a are 15 Aring
apart In contrast the CuB and the iron bound in
cytochrome a3 are very close to each other forming a
binuclear metal center shown below
27
Electron transport chains are associated with
membranes such as the mitochondrial membrane in
eukaryotic organisms the thylakoid membrane of
photosynthetic organisms and the plasma membrane of
prokaryotic organisms While some of the details
differ electron transport chains in all organisms
operate on the same basic principles
Excited electrons are brought to the electron
transport chain by electron carriers such as NADH
and FADH2 In photosynthetic organisms the electrons
are donated by water and excited by light energy
The electron donor is oxidized as the electrons are
passed to the first protein complex in the chain As
it accepts the electrons the protein complex is
reduced Next the electrons are passed from the
28
first protein complex which becomes oxidized to
the second protein complex which becomes reduced
Each of these redox reactions is slightly exergonic
or energy releasing Some of this energy is used to
move protons across the membrane against their
concentration gradient With each successive
transfer of electrons the original excited
electrons lose some of their energy After several
successive redox reactions the electrons are
donated to a terminal electron acceptor In aerobic
respiration the terminal electron acceptor is
oxygen In photosynthesis the terminal electron
acceptor is a compound called NADP
The energy in the excited electrons has been
transformed into an electrochemical gradient
represented by the high concentration of protons on
one side of the membrane These protons can diffuse
back across the membrane by passing through a
protein complex called ATP synthase As these
protons pass through ATP synthase down their
concentration gradient they provide the energy
needed to make ATP from ADP and inorganic phosphate
within the cell
29
CHAPTER III
CLOSING
A Conclution
When a hexose is completely oxidized to CO2
and H2O using these three processes Glycolysis
yields two ATP and two NADH per hexose used Each
such NADH oxidized by the electron transport system
30
yields two ATP as dscribed above so glycolysis
contributes a total of six ATP per hexose The Krebs
cycle contributes two ATP per hexose or per two
pyruvates when succinyl CoA is cleaved to succinate
and CoASH This cycle also produces eight NADH per
hexose within the mitochondrial matrix by oxidative
phosporylation each of these NADH yields three ATP
or 24 per hexose Each FADH2 from the Krebs cycle
yields two ATP by oxydative phosporylation or four
per hexose (two pyruvates) The total contribution of
the Krebs cycle is then 30 Adding these 30 to the 6
from glycolysis leads to a total of 36 per hexose
completely respired by these process
B Suggestion
My greatest wish is that the readers find
this manuscript helpful in gaining the knowledge they
need for effective learning and practice and in
preparing for examination designed to test that
knowledge Reader comments and suggestions are
welcome and would be very helpful in improving our
manuscript Your contribution may assist many other
students in the coming years
31
REFERENCE
Anonim 2013 Anaerobic Respiration
httpwwwscoolcoukgcsebiologyrespiration
revise-itanaerobic-respiration Accessed
September 10th 2013
Anonim 2013 What is the difference between aerobic
and anaerobic respiration in plants
httpwwwbiologylifeeasyorg4010difference
-between-aerobic-anaerobic-respiration-plants
Accessed September 10th 2013
Campbell Mary K 2006 Biochemistry New York Thomson
Brooks Cole Inc
Dat J Capelli N Folzer H Bourgeade P Badot P-M
(2004) Sensing and signaling during
plantflooding Plant Physiology and
Biochemistry 42 273-282
Dat J Folzer H Parent C Badot P-M Capelli N
(2006) HypoxiastressCurrent Understanding and
Perspectives In Teixeira da Silva JA (Ed)
Floriculture Ornamental and Plant
Biotechnology Advances and Topical Issues (Vol
3) Global Science Books Isleworth United
Kingdompp 664-674
Hatta Muhammad 2011 Dunia Pertanian
httpemhattawordpresscomcategorystress
Accessed September 10th 2013
Hopkins William G 2009 Introduction to Plant Physiology
New York John Wiley amp Sons Inc
Lakitan Benyamin 2011 Dasar-Dasar Fisiologi Tumbuhan
Jakarta PT Raja Grafindo Persada
Larcher W 1980 Physiological Plant Ecology New York
Springer-Verlag
Raven 2013 Biology Fifth Edition
httphigheredmcgraw-hillcomsites983409233
9student_view0chapter39
photosynthetic_electron_transport_and_atp_synth
esishtml Accessed September 10th 2013
Salisbury Frank B 1969 Plant Physiology Third Edition
California Wadsworth Publishing Company
Subbaiah C Sachs M (2003) Molecular and cellular
adaptations of maize to flooding stress Annals
of Botany 91 119-127
Wilson CarlL1962 Botany Third EditionUnited State of
America Holt Rinch Inc
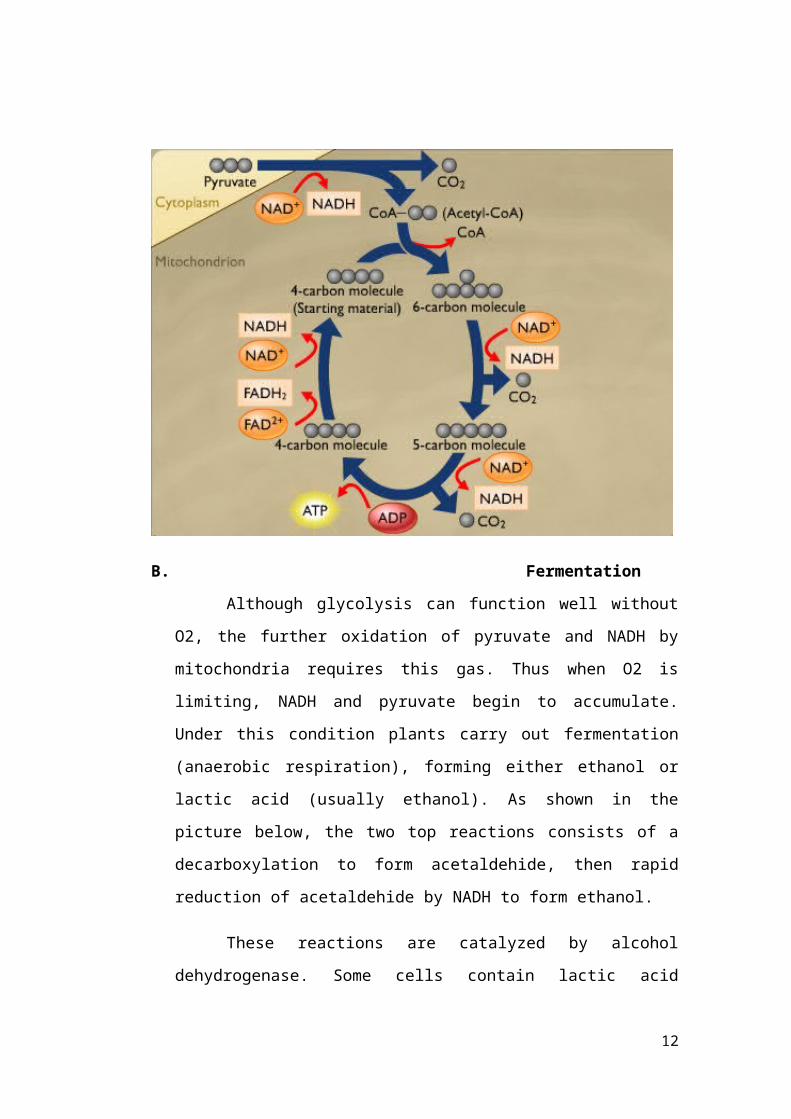
B Fermentation
Although glycolysis can function well without
O2 the further oxidation of pyruvate and NADH by
mitochondria requires this gas Thus when O2 is
limiting NADH and pyruvate begin to accumulate
Under this condition plants carry out fermentation
(anaerobic respiration) forming either ethanol or
lactic acid (usually ethanol) As shown in the
picture below the two top reactions consists of a
decarboxylation to form acetaldehide then rapid
reduction of acetaldehide by NADH to form ethanol
These reactions are catalyzed by alcohol
dehydrogenase Some cells contain lactic acid
12
dehydrogenase which uses NADH to reduce pyruvic to
lactic acid Thus either ethanol or lactic acid or
both are fermentation products depending on the
activities of each dehydrogenase present In each
case NADH is the reductant and only under
anaerobic conditions is it abundant anough to cause
reduction Furthermore in some plants NADH is used
to cause accumulation of the other compounds when O2
is limiting especially malate and glycerol
(Salisbury 1969 233)
The fate of pyruvate produced by glycolysis
depends primarily on whether or not oxygen is
present Under normal aerobic conditions pyruvate
is transported into the mitochondrion where it is
further oxidized to CO2 and water transferring its
electrons ultimately to molecular oxygen
13
Although higher plants are obligate aerobs and
are able to tolerate anoxia for only short periods
tissues or organs are occasionally subjected to
anaerobic conditions A typical siuation is that of
roots when the soil is saturated with water When
there is no oxygen to serve as the terminal electron
acceptor mitochondrial respiration will shut down
and metabolism will shift over to fermentation
Fermentation converts pyruvate to either ethanol
through the action of the enzyme alcohol
dehydrogenase (ADH) or lactate via lactate
dehydrogenase (LDH) In most plants the principal
products of fermentation are CO2 and ethanol Some
lactate may be formed primarily in the early stages
of anoxia However lactate lowers he pH of the
cytosol which in turn activates pyruvate
decarboxylase and initiates the production of
ethanol
Either one of the fermentation reactions
consumes the NADH produced earlier in glycolysis by
the oxidation of glyceraldehide-3-P Although this
means there is no net gain of reducing potential in
fermentation This recycling of NADH is still
important to the cell The poolof NADH plus NAD+ in
the cell is relatively small and if the NADH is not
recycled there will be no supply of NAD+ to support
14
the continued oxidation of glyceraldehyde-3-P if
this were the case glycolysis and the producton of
even the small quantities of ATP necessary to
maintain the cells under anaerobic conditions would
then grind to a halt (Hopkins 2009 241)
Soil waterlogging has long been identified as a
major abiotic stress and the constraints that it
provides the roots have a significant effect on the
growth and development of plants When this event
occurs in the spring then these puddles can reduce
seed germination and seedling development Thus a
pool of water is an important factor that affects
the growth development and survival of plant
species not only in natural ecosystems but also
the agricultural and horticultural systems (Dat et
al 2006)
After flooding rapid changes in soil
properties At the time of pore water meets the
land the air pushed out gas diffusion is reduced
and toxic compounds accumulate due to anaerobic
conditions All of these changes greatly affect the
ability of plants to survive In response increased
stomatal resistance photosynthesis and root
hydraulic conductivity decreases and reduced
translocation fotoassimilat Nevertheless one of
15
the best adaptations of plants to hypoxia anoxia
is transitional biochemical and metabolic processes
common to current limited availability of O2 (Dat et
al 2004)
Selective synthesis of a set of about 20
anaerobic stress proteins (ANPS) allows the process
of producing energy without oxygen metabolism under
anaerobic conditions (Subbaiah and Sachs 2003)
Another adaptation is the observed morphological
changes consisting of the formation of hypertrophic
lenticels adventitious root initiation and or
progression of aerenchyma (Vartapetian and Jackson
1997 Jackson and Colmer 2005 Folzer et al 2006)
This review details the diverse plant stress
response to hypoxia anoxia caused by soil
waterlogging flooding and examines some of the key
features of metabolic adaptation physiological and
morphological
16
C Electron
Transport System
We noted earlier that one of the principal
functions of the respiration is to retrieve in
useful form some of the energy initially stored in
assimilates Our traditional measure of useful
energy in most processes is the number of ATP
molecules gained or consumed By this measure alone
the yield from both glycolysis and the citric acid
cycle is quite low After two complete turns of the
cycle one molecule of glucose has been completely
oxidized to six molecules of CO2 but only four
molecules of ATP have been produced (a net of two
ATP from glycolysis plus one for each turn of the
cycle) At this point most of the energy associated
with the glucose molecule has been conserved in the
17
form of electron pairs generated by the oxidation of
glycolytic and citric acid cycle intermediates
In this section we will discuss the third stage
of cellular respirationmdashthe transfer of electrons
fromNADH and FADH2 to oxygen and the accompanying
conversion of redox energy to ATP The transfer of
electrons fromNADHand FADH2 to oxygen involves a
sequence of electron carriers arranged in an
electron transport chainMembrane fractionation
studies have shown that the enzymes and electron
carriers making up the electron transport chain are
organized predominantly into four large
multimolecular complexes (complexes ImdashIV) and two
mobile carriers located in the inner mitochondrial
membrane (Figure 109) In this sense there are a
great number of similarities between the
mitochondrial inner membrane and the thylakoid
membranes of the chloroplast (compare Figure 109
with Figure 76) This is not unexpected since the
principal function of each membrane is energy
transformation and many of the same or similar
components are involved
The Components of the Electron Transport Chain
The electron transport chain of the
mitochondria is the means by which electrons are
18
removed from the reduced carrier NADH and
transferred to oxygen to yield H2O
1) NADH
NADH is generated in the matrix by the reactions
of pyruvate dehydrogenase isocitrate
dehydrogenase α-ketoglutarate dehydrogenase and
malate dehyrogenase The electron transport chain
begins with reoxidizing NADH to form NAD+ and
channeling the electrons into the formation of
reduced coenzymes Important to note that NADH
transfers 2 electrons at a time in the form of a
hydride
NAD+ + 2e- + H+ 1048581 NADH Eorsquo = minus0315 V
2) Flavoproteins
Flavoproteins have either a FAD (flavin adenosine
dinucleotide) or a FMN (flavin mononucleotide)
prosthetic group Flavoproteins can accept or
donate electrons one at time or two at a time
Thus they are often intermediaries between two
electron acceptorsdonors and one electron
acceptorsdonors For flavoproteins the typical
standard reduction potentials are around 0 V
FAD + 2e- + 2H+ 1048581 FADH2 FMN + 2e- + 2H+ 1048581 FMNH2
Eorsquo asymp 0 V
19
3) Coenzyme Q (CoQ)
aka ubiquinone (UQ) Shown below CoQ has ten
repeating isoprene units which make it
insoluble in water but soluble in the
hydrophobic lipid bylayer Coenzyme Q is a
versatile cofactor because it is a soluble
electron carrier in the hydrophobic bilipid layer
of the
inner mitochondrial membrane Like flavoproteins
CoQ can acceptdonate electrons one at a time or
two at a time
Q + 2e- + 2H+ 1048581 QH2 Eorsquo =0060 V
Q + e- + H+ 1048581 1048581QH Eorsquo =0030 V
1048581QH + e- + H+ 1048581 QH2 Eorsquo =0190 V
4) Cytochromes
Cytochromes are proteins that contain heme
prosthetic groups which function as
one electron carriers The heme iron is involved
in one electron transfers involving the Fe2+ and
Fe3+ oxidation states Cytochromes are named by
their absorption spectra which depends
on the porphyrin structure and environment The
example shown is the heme prosthetic group of
cytochrome b Cytochrome b contains the same iron
porphyrin found in hemoglobin and myoglobin
20
Other cytochromes we will encounter in the
electron transport complexes are cytochromes b
c c1 a and a3
5) Iron-Sulfur Proteins
In the electron transport chain we will encounter
many iron-sulfer proteins which participate in
one electron transfers involving the the Fe2+ and
Fe3+ oxidation states These are non-heme iron-
sulfur proteinsThe simplest iron-sulfer protein
is FeS in which iron is tetrahedrally coordinated
by four cysteines The second form is Fe2S2 which
contains two irons complexed to 2 cysteine
residues and two inorganic sulfides The third
form is Fe3S4 which contains 3 iron atoms
coordinated to three cysteine residues and 4
inorganic sulfides The last form is the most
complicated Fe4S4 which contains 4 iron atoms
coordinated to 4 cysteine residues and 4
inorganic sulfides
6) Copper Proteins
Copper bound proteins participate in one electron
transfers involving the Cu+ and Cu2+ oxidation
states
21
Overview of the Electron Transport Chain
Electrons move along the electron transport
chain going from donor to acceptor until they reach
oxygen the ultimate electron acceptor The standard
reduction potentials of the electron carriers are
between the NADHNAD+ couple (-0315 V) and the
oxygenH2O couple (0816 V) as on the next page The
components of the electron transport chain are
organized into 4 complexes Each complex contains
several different electron carriers
1 Complex I also known as the NADH-coenzyme Q
reductase or NADH dehydrogenase
2 Complex II also known as succinate-coenzyme Q
reductase or succinate dehydrogenase
3 Complex III also known as coenzyme Q reductase
4 Complex IV also known as cytochrome c reductase
The free energy necessary to generate ATP
is extracted from the oxidation of NADH and
FADH2 by the electron transport chain It
consists of four protein complexes and single
protein chain of cytochrome c through which
electrons pass from lower to higher standard
redox potentials Electrons are carried from complex
I and II to complex III by the membrane (lipid)
soluble coenzyme Q (CoQ or ubiquinone) and between
22
complexes III and IV by the peripheral membrane
protein cytochrome c
In the following I give the overall
reactions catalysed by corresponding protein
complexes in the mitochondrial electron transport
chain
1 Complex I
Complex I is also called NADH-Coenzyme Q
reductase because this large protein complex
transfers 2 electrons from NADH to coenzyme Q
Complex I was formerly known as NADH dehydrogenase
Complex I is huge 850000 kD and is composed of more
than thirty subunits It contains a FMN prosthetic
group and seven or more Fe-S clusters This complex
has between 20-26 iron atoms boundThe prosthetic
group FMN is absolutely required for activity
Therefore this complex is a flavoprotein This
complex binds NADH transfers two electrons in the
form of a hydride to FMN to produce NAD+ and FMNH2
23
The subsequent steps involve the transfer of
electrons one at a time to a series of iron-sulfer
complexes that includes both 2Fe-2S and 4Fe-4S
clusters Note the importance of FMN First it
functions as a 2 electron acceptor in the hydride
transfer from NADH Second it functions as a 1
electron donor to the series of iron sulfur clusters
FMN and FAD often play crucial links between 2
electron transfer agents and 1 electron transfer
agents The final step of this complex is the
transfer of 2 electrons one at a time to coenzyme Q
CoQ like FMN and FAD can function as a 2 electron
donoracceptor and as a 1 electron donoracceptor
CoQ is a mobile electron carrier because its
isoprenoid tail makes it highly hydrophobic and
lipophillic It diffuses freely in the bilipid layer
of the inner mitochondrial membrane
The process of transferring electrons from NADH
to CoQ by complex I results in the net transport of
protons from the matrix side of the inner
mitochondrial membrane to the inter membrane space
where the H+ ions accumulate generating a proton
motive force The intermembrane space side of the
inner membrane is referred to as the P face (P
standing for positive) The matrix side of the inner
membrane is referred to as the the N face The
24
transport of electrons from NADH to CoQ is coupled to
the transport of protons across the membrane This is
an example of active transport The stiochiometry is
4 H+ transported per 2 electrons
2 Complex II
Believe it or not you are already familiar
with Complex II It is none other than succinate
dehydrogenase The only enzyme of the citric acid
cycle that is an integral membrane protein This
complex is composed of four subunits 2 of which are
iron-sulfur proteins and the other two subunits
together bind FAD through a covalent link to a
histidine residue These two subunits are called
flavoprotein 2 or FP2 Complex II contains 3 Fe-S
centers 1 4Fe-4S cluster 1 3Fe-4S cluster and 1
2Fe- 2S cluster In the first step of this complex
succinate is bound and a hydride is transferred to
FAD to generate FADH2 and fumarate FADH2 then
transfers its electrons one at a time to the Fe-S
centers Thus once again FAD functions as 2 electron
acceptor and a 1 electron donor The final step of
this complex is the transfer of 2 electrons one at a
time to coenzyme Q to produce CoQH2 For complex II
the standard free energy change of the overall
reaction is too small to drive the transport of
protons across the inner mitochondrial membrane This
25
accounts for the 15 ATPrsquos generated per FADH2
compared with the 25 ATPrsquos generated per NADH
3 Complex III
This complex is also known as coenzyme Q-
cytochrome c reductase because it passes the
electrons form CoQH2 to cyt c through a very unique
electron transport pathway called the Q-cycle Shown
to the left are the porphyrins found in cytochromes
Cytochrome b contains the same iron protoporphyrin as
hemoglobin and myoglobin The c cytochromes contain
heme c through covalent attachment by cysteine
residues Cytochrome a is found in two forms in
complex IV In complex III we find two b-type
cytochromes and one c-type cytochrome Complex III is
complex and we have a crystal structure
4 Complex IV
Complex IV is also known as cytochrome c oxidase
because it accepts the electrons from cytochrome c
and directs them towards the four electron reduction
of O2 to form 2 molecules of H2O
4 cyt c (Fe2+) + 4 H+ + O2 4 cyt c (Fe3+) +
2H2O
26
Cytochrome c oxidase contains 2 heme
centerscytochrome a and cytochrome a3 and two copper
proteins Each of the protein bound coppers are
associated with one of the cytochromes The copper
sites are called CuA and CuB CuA is associated with
cytochrome a and is shown to the left CuB is
associated with cytochrome a3 The copper sites
function as 1 electron carriers cycling between the
cuprous state Cu+ and the cupric state Cu2+ Just
like iron containing proteins they transfer
electrons one at a time Cytochrome c is bound on the
P-face of the membrane and transfers its electron to
CuA The oxidized cytochrome c dissociates CuA then
transfers the electron to cytochrome a The protein
bound CuA and the iron bound in cytochrome a are 15 Aring
apart In contrast the CuB and the iron bound in
cytochrome a3 are very close to each other forming a
binuclear metal center shown below
27
Electron transport chains are associated with
membranes such as the mitochondrial membrane in
eukaryotic organisms the thylakoid membrane of
photosynthetic organisms and the plasma membrane of
prokaryotic organisms While some of the details
differ electron transport chains in all organisms
operate on the same basic principles
Excited electrons are brought to the electron
transport chain by electron carriers such as NADH
and FADH2 In photosynthetic organisms the electrons
are donated by water and excited by light energy
The electron donor is oxidized as the electrons are
passed to the first protein complex in the chain As
it accepts the electrons the protein complex is
reduced Next the electrons are passed from the
28
first protein complex which becomes oxidized to
the second protein complex which becomes reduced
Each of these redox reactions is slightly exergonic
or energy releasing Some of this energy is used to
move protons across the membrane against their
concentration gradient With each successive
transfer of electrons the original excited
electrons lose some of their energy After several
successive redox reactions the electrons are
donated to a terminal electron acceptor In aerobic
respiration the terminal electron acceptor is
oxygen In photosynthesis the terminal electron
acceptor is a compound called NADP
The energy in the excited electrons has been
transformed into an electrochemical gradient
represented by the high concentration of protons on
one side of the membrane These protons can diffuse
back across the membrane by passing through a
protein complex called ATP synthase As these
protons pass through ATP synthase down their
concentration gradient they provide the energy
needed to make ATP from ADP and inorganic phosphate
within the cell
29
CHAPTER III
CLOSING
A Conclution
When a hexose is completely oxidized to CO2
and H2O using these three processes Glycolysis
yields two ATP and two NADH per hexose used Each
such NADH oxidized by the electron transport system
30
yields two ATP as dscribed above so glycolysis
contributes a total of six ATP per hexose The Krebs
cycle contributes two ATP per hexose or per two
pyruvates when succinyl CoA is cleaved to succinate
and CoASH This cycle also produces eight NADH per
hexose within the mitochondrial matrix by oxidative
phosporylation each of these NADH yields three ATP
or 24 per hexose Each FADH2 from the Krebs cycle
yields two ATP by oxydative phosporylation or four
per hexose (two pyruvates) The total contribution of
the Krebs cycle is then 30 Adding these 30 to the 6
from glycolysis leads to a total of 36 per hexose
completely respired by these process
B Suggestion
My greatest wish is that the readers find
this manuscript helpful in gaining the knowledge they
need for effective learning and practice and in
preparing for examination designed to test that
knowledge Reader comments and suggestions are
welcome and would be very helpful in improving our
manuscript Your contribution may assist many other
students in the coming years
31
REFERENCE
Anonim 2013 Anaerobic Respiration
httpwwwscoolcoukgcsebiologyrespiration
revise-itanaerobic-respiration Accessed
September 10th 2013
Anonim 2013 What is the difference between aerobic
and anaerobic respiration in plants
httpwwwbiologylifeeasyorg4010difference
-between-aerobic-anaerobic-respiration-plants
Accessed September 10th 2013
Campbell Mary K 2006 Biochemistry New York Thomson
Brooks Cole Inc
Dat J Capelli N Folzer H Bourgeade P Badot P-M
(2004) Sensing and signaling during
plantflooding Plant Physiology and
Biochemistry 42 273-282
Dat J Folzer H Parent C Badot P-M Capelli N
(2006) HypoxiastressCurrent Understanding and
Perspectives In Teixeira da Silva JA (Ed)
Floriculture Ornamental and Plant
Biotechnology Advances and Topical Issues (Vol
3) Global Science Books Isleworth United
Kingdompp 664-674
Hatta Muhammad 2011 Dunia Pertanian
httpemhattawordpresscomcategorystress
Accessed September 10th 2013
Hopkins William G 2009 Introduction to Plant Physiology
New York John Wiley amp Sons Inc
Lakitan Benyamin 2011 Dasar-Dasar Fisiologi Tumbuhan
Jakarta PT Raja Grafindo Persada
Larcher W 1980 Physiological Plant Ecology New York
Springer-Verlag
Raven 2013 Biology Fifth Edition
httphigheredmcgraw-hillcomsites983409233
9student_view0chapter39
photosynthetic_electron_transport_and_atp_synth
esishtml Accessed September 10th 2013
Salisbury Frank B 1969 Plant Physiology Third Edition
California Wadsworth Publishing Company
Subbaiah C Sachs M (2003) Molecular and cellular
adaptations of maize to flooding stress Annals
of Botany 91 119-127
Wilson CarlL1962 Botany Third EditionUnited State of
America Holt Rinch Inc

dehydrogenase which uses NADH to reduce pyruvic to
lactic acid Thus either ethanol or lactic acid or
both are fermentation products depending on the
activities of each dehydrogenase present In each
case NADH is the reductant and only under
anaerobic conditions is it abundant anough to cause
reduction Furthermore in some plants NADH is used
to cause accumulation of the other compounds when O2
is limiting especially malate and glycerol
(Salisbury 1969 233)
The fate of pyruvate produced by glycolysis
depends primarily on whether or not oxygen is
present Under normal aerobic conditions pyruvate
is transported into the mitochondrion where it is
further oxidized to CO2 and water transferring its
electrons ultimately to molecular oxygen
13
Although higher plants are obligate aerobs and
are able to tolerate anoxia for only short periods
tissues or organs are occasionally subjected to
anaerobic conditions A typical siuation is that of
roots when the soil is saturated with water When
there is no oxygen to serve as the terminal electron
acceptor mitochondrial respiration will shut down
and metabolism will shift over to fermentation
Fermentation converts pyruvate to either ethanol
through the action of the enzyme alcohol
dehydrogenase (ADH) or lactate via lactate
dehydrogenase (LDH) In most plants the principal
products of fermentation are CO2 and ethanol Some
lactate may be formed primarily in the early stages
of anoxia However lactate lowers he pH of the
cytosol which in turn activates pyruvate
decarboxylase and initiates the production of
ethanol
Either one of the fermentation reactions
consumes the NADH produced earlier in glycolysis by
the oxidation of glyceraldehide-3-P Although this
means there is no net gain of reducing potential in
fermentation This recycling of NADH is still
important to the cell The poolof NADH plus NAD+ in
the cell is relatively small and if the NADH is not
recycled there will be no supply of NAD+ to support
14
the continued oxidation of glyceraldehyde-3-P if
this were the case glycolysis and the producton of
even the small quantities of ATP necessary to
maintain the cells under anaerobic conditions would
then grind to a halt (Hopkins 2009 241)
Soil waterlogging has long been identified as a
major abiotic stress and the constraints that it
provides the roots have a significant effect on the
growth and development of plants When this event
occurs in the spring then these puddles can reduce
seed germination and seedling development Thus a
pool of water is an important factor that affects
the growth development and survival of plant
species not only in natural ecosystems but also
the agricultural and horticultural systems (Dat et
al 2006)
After flooding rapid changes in soil
properties At the time of pore water meets the
land the air pushed out gas diffusion is reduced
and toxic compounds accumulate due to anaerobic
conditions All of these changes greatly affect the
ability of plants to survive In response increased
stomatal resistance photosynthesis and root
hydraulic conductivity decreases and reduced
translocation fotoassimilat Nevertheless one of
15
the best adaptations of plants to hypoxia anoxia
is transitional biochemical and metabolic processes
common to current limited availability of O2 (Dat et
al 2004)
Selective synthesis of a set of about 20
anaerobic stress proteins (ANPS) allows the process
of producing energy without oxygen metabolism under
anaerobic conditions (Subbaiah and Sachs 2003)
Another adaptation is the observed morphological
changes consisting of the formation of hypertrophic
lenticels adventitious root initiation and or
progression of aerenchyma (Vartapetian and Jackson
1997 Jackson and Colmer 2005 Folzer et al 2006)
This review details the diverse plant stress
response to hypoxia anoxia caused by soil
waterlogging flooding and examines some of the key
features of metabolic adaptation physiological and
morphological
16
C Electron
Transport System
We noted earlier that one of the principal
functions of the respiration is to retrieve in
useful form some of the energy initially stored in
assimilates Our traditional measure of useful
energy in most processes is the number of ATP
molecules gained or consumed By this measure alone
the yield from both glycolysis and the citric acid
cycle is quite low After two complete turns of the
cycle one molecule of glucose has been completely
oxidized to six molecules of CO2 but only four
molecules of ATP have been produced (a net of two
ATP from glycolysis plus one for each turn of the
cycle) At this point most of the energy associated
with the glucose molecule has been conserved in the
17
form of electron pairs generated by the oxidation of
glycolytic and citric acid cycle intermediates
In this section we will discuss the third stage
of cellular respirationmdashthe transfer of electrons
fromNADH and FADH2 to oxygen and the accompanying
conversion of redox energy to ATP The transfer of
electrons fromNADHand FADH2 to oxygen involves a
sequence of electron carriers arranged in an
electron transport chainMembrane fractionation
studies have shown that the enzymes and electron
carriers making up the electron transport chain are
organized predominantly into four large
multimolecular complexes (complexes ImdashIV) and two
mobile carriers located in the inner mitochondrial
membrane (Figure 109) In this sense there are a
great number of similarities between the
mitochondrial inner membrane and the thylakoid
membranes of the chloroplast (compare Figure 109
with Figure 76) This is not unexpected since the
principal function of each membrane is energy
transformation and many of the same or similar
components are involved
The Components of the Electron Transport Chain
The electron transport chain of the
mitochondria is the means by which electrons are
18
removed from the reduced carrier NADH and
transferred to oxygen to yield H2O
1) NADH
NADH is generated in the matrix by the reactions
of pyruvate dehydrogenase isocitrate
dehydrogenase α-ketoglutarate dehydrogenase and
malate dehyrogenase The electron transport chain
begins with reoxidizing NADH to form NAD+ and
channeling the electrons into the formation of
reduced coenzymes Important to note that NADH
transfers 2 electrons at a time in the form of a
hydride
NAD+ + 2e- + H+ 1048581 NADH Eorsquo = minus0315 V
2) Flavoproteins
Flavoproteins have either a FAD (flavin adenosine
dinucleotide) or a FMN (flavin mononucleotide)
prosthetic group Flavoproteins can accept or
donate electrons one at time or two at a time
Thus they are often intermediaries between two
electron acceptorsdonors and one electron
acceptorsdonors For flavoproteins the typical
standard reduction potentials are around 0 V
FAD + 2e- + 2H+ 1048581 FADH2 FMN + 2e- + 2H+ 1048581 FMNH2
Eorsquo asymp 0 V
19
3) Coenzyme Q (CoQ)
aka ubiquinone (UQ) Shown below CoQ has ten
repeating isoprene units which make it
insoluble in water but soluble in the
hydrophobic lipid bylayer Coenzyme Q is a
versatile cofactor because it is a soluble
electron carrier in the hydrophobic bilipid layer
of the
inner mitochondrial membrane Like flavoproteins
CoQ can acceptdonate electrons one at a time or
two at a time
Q + 2e- + 2H+ 1048581 QH2 Eorsquo =0060 V
Q + e- + H+ 1048581 1048581QH Eorsquo =0030 V
1048581QH + e- + H+ 1048581 QH2 Eorsquo =0190 V
4) Cytochromes
Cytochromes are proteins that contain heme
prosthetic groups which function as
one electron carriers The heme iron is involved
in one electron transfers involving the Fe2+ and
Fe3+ oxidation states Cytochromes are named by
their absorption spectra which depends
on the porphyrin structure and environment The
example shown is the heme prosthetic group of
cytochrome b Cytochrome b contains the same iron
porphyrin found in hemoglobin and myoglobin
20
Other cytochromes we will encounter in the
electron transport complexes are cytochromes b
c c1 a and a3
5) Iron-Sulfur Proteins
In the electron transport chain we will encounter
many iron-sulfer proteins which participate in
one electron transfers involving the the Fe2+ and
Fe3+ oxidation states These are non-heme iron-
sulfur proteinsThe simplest iron-sulfer protein
is FeS in which iron is tetrahedrally coordinated
by four cysteines The second form is Fe2S2 which
contains two irons complexed to 2 cysteine
residues and two inorganic sulfides The third
form is Fe3S4 which contains 3 iron atoms
coordinated to three cysteine residues and 4
inorganic sulfides The last form is the most
complicated Fe4S4 which contains 4 iron atoms
coordinated to 4 cysteine residues and 4
inorganic sulfides
6) Copper Proteins
Copper bound proteins participate in one electron
transfers involving the Cu+ and Cu2+ oxidation
states
21
Overview of the Electron Transport Chain
Electrons move along the electron transport
chain going from donor to acceptor until they reach
oxygen the ultimate electron acceptor The standard
reduction potentials of the electron carriers are
between the NADHNAD+ couple (-0315 V) and the
oxygenH2O couple (0816 V) as on the next page The
components of the electron transport chain are
organized into 4 complexes Each complex contains
several different electron carriers
1 Complex I also known as the NADH-coenzyme Q
reductase or NADH dehydrogenase
2 Complex II also known as succinate-coenzyme Q
reductase or succinate dehydrogenase
3 Complex III also known as coenzyme Q reductase
4 Complex IV also known as cytochrome c reductase
The free energy necessary to generate ATP
is extracted from the oxidation of NADH and
FADH2 by the electron transport chain It
consists of four protein complexes and single
protein chain of cytochrome c through which
electrons pass from lower to higher standard
redox potentials Electrons are carried from complex
I and II to complex III by the membrane (lipid)
soluble coenzyme Q (CoQ or ubiquinone) and between
22
complexes III and IV by the peripheral membrane
protein cytochrome c
In the following I give the overall
reactions catalysed by corresponding protein
complexes in the mitochondrial electron transport
chain
1 Complex I
Complex I is also called NADH-Coenzyme Q
reductase because this large protein complex
transfers 2 electrons from NADH to coenzyme Q
Complex I was formerly known as NADH dehydrogenase
Complex I is huge 850000 kD and is composed of more
than thirty subunits It contains a FMN prosthetic
group and seven or more Fe-S clusters This complex
has between 20-26 iron atoms boundThe prosthetic
group FMN is absolutely required for activity
Therefore this complex is a flavoprotein This
complex binds NADH transfers two electrons in the
form of a hydride to FMN to produce NAD+ and FMNH2
23
The subsequent steps involve the transfer of
electrons one at a time to a series of iron-sulfer
complexes that includes both 2Fe-2S and 4Fe-4S
clusters Note the importance of FMN First it
functions as a 2 electron acceptor in the hydride
transfer from NADH Second it functions as a 1
electron donor to the series of iron sulfur clusters
FMN and FAD often play crucial links between 2
electron transfer agents and 1 electron transfer
agents The final step of this complex is the
transfer of 2 electrons one at a time to coenzyme Q
CoQ like FMN and FAD can function as a 2 electron
donoracceptor and as a 1 electron donoracceptor
CoQ is a mobile electron carrier because its
isoprenoid tail makes it highly hydrophobic and
lipophillic It diffuses freely in the bilipid layer
of the inner mitochondrial membrane
The process of transferring electrons from NADH
to CoQ by complex I results in the net transport of
protons from the matrix side of the inner
mitochondrial membrane to the inter membrane space
where the H+ ions accumulate generating a proton
motive force The intermembrane space side of the
inner membrane is referred to as the P face (P
standing for positive) The matrix side of the inner
membrane is referred to as the the N face The
24
transport of electrons from NADH to CoQ is coupled to
the transport of protons across the membrane This is
an example of active transport The stiochiometry is
4 H+ transported per 2 electrons
2 Complex II
Believe it or not you are already familiar
with Complex II It is none other than succinate
dehydrogenase The only enzyme of the citric acid
cycle that is an integral membrane protein This
complex is composed of four subunits 2 of which are
iron-sulfur proteins and the other two subunits
together bind FAD through a covalent link to a
histidine residue These two subunits are called
flavoprotein 2 or FP2 Complex II contains 3 Fe-S
centers 1 4Fe-4S cluster 1 3Fe-4S cluster and 1
2Fe- 2S cluster In the first step of this complex
succinate is bound and a hydride is transferred to
FAD to generate FADH2 and fumarate FADH2 then
transfers its electrons one at a time to the Fe-S
centers Thus once again FAD functions as 2 electron
acceptor and a 1 electron donor The final step of
this complex is the transfer of 2 electrons one at a
time to coenzyme Q to produce CoQH2 For complex II
the standard free energy change of the overall
reaction is too small to drive the transport of
protons across the inner mitochondrial membrane This
25
accounts for the 15 ATPrsquos generated per FADH2
compared with the 25 ATPrsquos generated per NADH
3 Complex III
This complex is also known as coenzyme Q-
cytochrome c reductase because it passes the
electrons form CoQH2 to cyt c through a very unique
electron transport pathway called the Q-cycle Shown
to the left are the porphyrins found in cytochromes
Cytochrome b contains the same iron protoporphyrin as
hemoglobin and myoglobin The c cytochromes contain
heme c through covalent attachment by cysteine
residues Cytochrome a is found in two forms in
complex IV In complex III we find two b-type
cytochromes and one c-type cytochrome Complex III is
complex and we have a crystal structure
4 Complex IV
Complex IV is also known as cytochrome c oxidase
because it accepts the electrons from cytochrome c
and directs them towards the four electron reduction
of O2 to form 2 molecules of H2O
4 cyt c (Fe2+) + 4 H+ + O2 4 cyt c (Fe3+) +
2H2O
26
Cytochrome c oxidase contains 2 heme
centerscytochrome a and cytochrome a3 and two copper
proteins Each of the protein bound coppers are
associated with one of the cytochromes The copper
sites are called CuA and CuB CuA is associated with
cytochrome a and is shown to the left CuB is
associated with cytochrome a3 The copper sites
function as 1 electron carriers cycling between the
cuprous state Cu+ and the cupric state Cu2+ Just
like iron containing proteins they transfer
electrons one at a time Cytochrome c is bound on the
P-face of the membrane and transfers its electron to
CuA The oxidized cytochrome c dissociates CuA then
transfers the electron to cytochrome a The protein
bound CuA and the iron bound in cytochrome a are 15 Aring
apart In contrast the CuB and the iron bound in
cytochrome a3 are very close to each other forming a
binuclear metal center shown below
27
Electron transport chains are associated with
membranes such as the mitochondrial membrane in
eukaryotic organisms the thylakoid membrane of
photosynthetic organisms and the plasma membrane of
prokaryotic organisms While some of the details
differ electron transport chains in all organisms
operate on the same basic principles
Excited electrons are brought to the electron
transport chain by electron carriers such as NADH
and FADH2 In photosynthetic organisms the electrons
are donated by water and excited by light energy
The electron donor is oxidized as the electrons are
passed to the first protein complex in the chain As
it accepts the electrons the protein complex is
reduced Next the electrons are passed from the
28
first protein complex which becomes oxidized to
the second protein complex which becomes reduced
Each of these redox reactions is slightly exergonic
or energy releasing Some of this energy is used to
move protons across the membrane against their
concentration gradient With each successive
transfer of electrons the original excited
electrons lose some of their energy After several
successive redox reactions the electrons are
donated to a terminal electron acceptor In aerobic
respiration the terminal electron acceptor is
oxygen In photosynthesis the terminal electron
acceptor is a compound called NADP
The energy in the excited electrons has been
transformed into an electrochemical gradient
represented by the high concentration of protons on
one side of the membrane These protons can diffuse
back across the membrane by passing through a
protein complex called ATP synthase As these
protons pass through ATP synthase down their
concentration gradient they provide the energy
needed to make ATP from ADP and inorganic phosphate
within the cell
29
CHAPTER III
CLOSING
A Conclution
When a hexose is completely oxidized to CO2
and H2O using these three processes Glycolysis
yields two ATP and two NADH per hexose used Each
such NADH oxidized by the electron transport system
30
yields two ATP as dscribed above so glycolysis
contributes a total of six ATP per hexose The Krebs
cycle contributes two ATP per hexose or per two
pyruvates when succinyl CoA is cleaved to succinate
and CoASH This cycle also produces eight NADH per
hexose within the mitochondrial matrix by oxidative
phosporylation each of these NADH yields three ATP
or 24 per hexose Each FADH2 from the Krebs cycle
yields two ATP by oxydative phosporylation or four
per hexose (two pyruvates) The total contribution of
the Krebs cycle is then 30 Adding these 30 to the 6
from glycolysis leads to a total of 36 per hexose
completely respired by these process
B Suggestion
My greatest wish is that the readers find
this manuscript helpful in gaining the knowledge they
need for effective learning and practice and in
preparing for examination designed to test that
knowledge Reader comments and suggestions are
welcome and would be very helpful in improving our
manuscript Your contribution may assist many other
students in the coming years
31
REFERENCE
Anonim 2013 Anaerobic Respiration
httpwwwscoolcoukgcsebiologyrespiration
revise-itanaerobic-respiration Accessed
September 10th 2013
Anonim 2013 What is the difference between aerobic
and anaerobic respiration in plants
httpwwwbiologylifeeasyorg4010difference
-between-aerobic-anaerobic-respiration-plants
Accessed September 10th 2013
Campbell Mary K 2006 Biochemistry New York Thomson
Brooks Cole Inc
Dat J Capelli N Folzer H Bourgeade P Badot P-M
(2004) Sensing and signaling during
plantflooding Plant Physiology and
Biochemistry 42 273-282
Dat J Folzer H Parent C Badot P-M Capelli N
(2006) HypoxiastressCurrent Understanding and
Perspectives In Teixeira da Silva JA (Ed)
Floriculture Ornamental and Plant
Biotechnology Advances and Topical Issues (Vol
3) Global Science Books Isleworth United
Kingdompp 664-674
Hatta Muhammad 2011 Dunia Pertanian
httpemhattawordpresscomcategorystress
Accessed September 10th 2013
Hopkins William G 2009 Introduction to Plant Physiology
New York John Wiley amp Sons Inc
Lakitan Benyamin 2011 Dasar-Dasar Fisiologi Tumbuhan
Jakarta PT Raja Grafindo Persada
Larcher W 1980 Physiological Plant Ecology New York
Springer-Verlag
Raven 2013 Biology Fifth Edition
httphigheredmcgraw-hillcomsites983409233
9student_view0chapter39
photosynthetic_electron_transport_and_atp_synth
esishtml Accessed September 10th 2013
Salisbury Frank B 1969 Plant Physiology Third Edition
California Wadsworth Publishing Company
Subbaiah C Sachs M (2003) Molecular and cellular
adaptations of maize to flooding stress Annals
of Botany 91 119-127
Wilson CarlL1962 Botany Third EditionUnited State of
America Holt Rinch Inc

Although higher plants are obligate aerobs and
are able to tolerate anoxia for only short periods
tissues or organs are occasionally subjected to
anaerobic conditions A typical siuation is that of
roots when the soil is saturated with water When
there is no oxygen to serve as the terminal electron
acceptor mitochondrial respiration will shut down
and metabolism will shift over to fermentation
Fermentation converts pyruvate to either ethanol
through the action of the enzyme alcohol
dehydrogenase (ADH) or lactate via lactate
dehydrogenase (LDH) In most plants the principal
products of fermentation are CO2 and ethanol Some
lactate may be formed primarily in the early stages
of anoxia However lactate lowers he pH of the
cytosol which in turn activates pyruvate
decarboxylase and initiates the production of
ethanol
Either one of the fermentation reactions
consumes the NADH produced earlier in glycolysis by
the oxidation of glyceraldehide-3-P Although this
means there is no net gain of reducing potential in
fermentation This recycling of NADH is still
important to the cell The poolof NADH plus NAD+ in
the cell is relatively small and if the NADH is not
recycled there will be no supply of NAD+ to support
14
the continued oxidation of glyceraldehyde-3-P if
this were the case glycolysis and the producton of
even the small quantities of ATP necessary to
maintain the cells under anaerobic conditions would
then grind to a halt (Hopkins 2009 241)
Soil waterlogging has long been identified as a
major abiotic stress and the constraints that it
provides the roots have a significant effect on the
growth and development of plants When this event
occurs in the spring then these puddles can reduce
seed germination and seedling development Thus a
pool of water is an important factor that affects
the growth development and survival of plant
species not only in natural ecosystems but also
the agricultural and horticultural systems (Dat et
al 2006)
After flooding rapid changes in soil
properties At the time of pore water meets the
land the air pushed out gas diffusion is reduced
and toxic compounds accumulate due to anaerobic
conditions All of these changes greatly affect the
ability of plants to survive In response increased
stomatal resistance photosynthesis and root
hydraulic conductivity decreases and reduced
translocation fotoassimilat Nevertheless one of
15
the best adaptations of plants to hypoxia anoxia
is transitional biochemical and metabolic processes
common to current limited availability of O2 (Dat et
al 2004)
Selective synthesis of a set of about 20
anaerobic stress proteins (ANPS) allows the process
of producing energy without oxygen metabolism under
anaerobic conditions (Subbaiah and Sachs 2003)
Another adaptation is the observed morphological
changes consisting of the formation of hypertrophic
lenticels adventitious root initiation and or
progression of aerenchyma (Vartapetian and Jackson
1997 Jackson and Colmer 2005 Folzer et al 2006)
This review details the diverse plant stress
response to hypoxia anoxia caused by soil
waterlogging flooding and examines some of the key
features of metabolic adaptation physiological and
morphological
16
C Electron
Transport System
We noted earlier that one of the principal
functions of the respiration is to retrieve in
useful form some of the energy initially stored in
assimilates Our traditional measure of useful
energy in most processes is the number of ATP
molecules gained or consumed By this measure alone
the yield from both glycolysis and the citric acid
cycle is quite low After two complete turns of the
cycle one molecule of glucose has been completely
oxidized to six molecules of CO2 but only four
molecules of ATP have been produced (a net of two
ATP from glycolysis plus one for each turn of the
cycle) At this point most of the energy associated
with the glucose molecule has been conserved in the
17
form of electron pairs generated by the oxidation of
glycolytic and citric acid cycle intermediates
In this section we will discuss the third stage
of cellular respirationmdashthe transfer of electrons
fromNADH and FADH2 to oxygen and the accompanying
conversion of redox energy to ATP The transfer of
electrons fromNADHand FADH2 to oxygen involves a
sequence of electron carriers arranged in an
electron transport chainMembrane fractionation
studies have shown that the enzymes and electron
carriers making up the electron transport chain are
organized predominantly into four large
multimolecular complexes (complexes ImdashIV) and two
mobile carriers located in the inner mitochondrial
membrane (Figure 109) In this sense there are a
great number of similarities between the
mitochondrial inner membrane and the thylakoid
membranes of the chloroplast (compare Figure 109
with Figure 76) This is not unexpected since the
principal function of each membrane is energy
transformation and many of the same or similar
components are involved
The Components of the Electron Transport Chain
The electron transport chain of the
mitochondria is the means by which electrons are
18
removed from the reduced carrier NADH and
transferred to oxygen to yield H2O
1) NADH
NADH is generated in the matrix by the reactions
of pyruvate dehydrogenase isocitrate
dehydrogenase α-ketoglutarate dehydrogenase and
malate dehyrogenase The electron transport chain
begins with reoxidizing NADH to form NAD+ and
channeling the electrons into the formation of
reduced coenzymes Important to note that NADH
transfers 2 electrons at a time in the form of a
hydride
NAD+ + 2e- + H+ 1048581 NADH Eorsquo = minus0315 V
2) Flavoproteins
Flavoproteins have either a FAD (flavin adenosine
dinucleotide) or a FMN (flavin mononucleotide)
prosthetic group Flavoproteins can accept or
donate electrons one at time or two at a time
Thus they are often intermediaries between two
electron acceptorsdonors and one electron
acceptorsdonors For flavoproteins the typical
standard reduction potentials are around 0 V
FAD + 2e- + 2H+ 1048581 FADH2 FMN + 2e- + 2H+ 1048581 FMNH2
Eorsquo asymp 0 V
19
3) Coenzyme Q (CoQ)
aka ubiquinone (UQ) Shown below CoQ has ten
repeating isoprene units which make it
insoluble in water but soluble in the
hydrophobic lipid bylayer Coenzyme Q is a
versatile cofactor because it is a soluble
electron carrier in the hydrophobic bilipid layer
of the
inner mitochondrial membrane Like flavoproteins
CoQ can acceptdonate electrons one at a time or
two at a time
Q + 2e- + 2H+ 1048581 QH2 Eorsquo =0060 V
Q + e- + H+ 1048581 1048581QH Eorsquo =0030 V
1048581QH + e- + H+ 1048581 QH2 Eorsquo =0190 V
4) Cytochromes
Cytochromes are proteins that contain heme
prosthetic groups which function as
one electron carriers The heme iron is involved
in one electron transfers involving the Fe2+ and
Fe3+ oxidation states Cytochromes are named by
their absorption spectra which depends
on the porphyrin structure and environment The
example shown is the heme prosthetic group of
cytochrome b Cytochrome b contains the same iron
porphyrin found in hemoglobin and myoglobin
20
Other cytochromes we will encounter in the
electron transport complexes are cytochromes b
c c1 a and a3
5) Iron-Sulfur Proteins
In the electron transport chain we will encounter
many iron-sulfer proteins which participate in
one electron transfers involving the the Fe2+ and
Fe3+ oxidation states These are non-heme iron-
sulfur proteinsThe simplest iron-sulfer protein
is FeS in which iron is tetrahedrally coordinated
by four cysteines The second form is Fe2S2 which
contains two irons complexed to 2 cysteine
residues and two inorganic sulfides The third
form is Fe3S4 which contains 3 iron atoms
coordinated to three cysteine residues and 4
inorganic sulfides The last form is the most
complicated Fe4S4 which contains 4 iron atoms
coordinated to 4 cysteine residues and 4
inorganic sulfides
6) Copper Proteins
Copper bound proteins participate in one electron
transfers involving the Cu+ and Cu2+ oxidation
states
21
Overview of the Electron Transport Chain
Electrons move along the electron transport
chain going from donor to acceptor until they reach
oxygen the ultimate electron acceptor The standard
reduction potentials of the electron carriers are
between the NADHNAD+ couple (-0315 V) and the
oxygenH2O couple (0816 V) as on the next page The
components of the electron transport chain are
organized into 4 complexes Each complex contains
several different electron carriers
1 Complex I also known as the NADH-coenzyme Q
reductase or NADH dehydrogenase
2 Complex II also known as succinate-coenzyme Q
reductase or succinate dehydrogenase
3 Complex III also known as coenzyme Q reductase
4 Complex IV also known as cytochrome c reductase
The free energy necessary to generate ATP
is extracted from the oxidation of NADH and
FADH2 by the electron transport chain It
consists of four protein complexes and single
protein chain of cytochrome c through which
electrons pass from lower to higher standard
redox potentials Electrons are carried from complex
I and II to complex III by the membrane (lipid)
soluble coenzyme Q (CoQ or ubiquinone) and between
22
complexes III and IV by the peripheral membrane
protein cytochrome c
In the following I give the overall
reactions catalysed by corresponding protein
complexes in the mitochondrial electron transport
chain
1 Complex I
Complex I is also called NADH-Coenzyme Q
reductase because this large protein complex
transfers 2 electrons from NADH to coenzyme Q
Complex I was formerly known as NADH dehydrogenase
Complex I is huge 850000 kD and is composed of more
than thirty subunits It contains a FMN prosthetic
group and seven or more Fe-S clusters This complex
has between 20-26 iron atoms boundThe prosthetic
group FMN is absolutely required for activity
Therefore this complex is a flavoprotein This
complex binds NADH transfers two electrons in the
form of a hydride to FMN to produce NAD+ and FMNH2
23
The subsequent steps involve the transfer of
electrons one at a time to a series of iron-sulfer
complexes that includes both 2Fe-2S and 4Fe-4S
clusters Note the importance of FMN First it
functions as a 2 electron acceptor in the hydride
transfer from NADH Second it functions as a 1
electron donor to the series of iron sulfur clusters
FMN and FAD often play crucial links between 2
electron transfer agents and 1 electron transfer
agents The final step of this complex is the
transfer of 2 electrons one at a time to coenzyme Q
CoQ like FMN and FAD can function as a 2 electron
donoracceptor and as a 1 electron donoracceptor
CoQ is a mobile electron carrier because its
isoprenoid tail makes it highly hydrophobic and
lipophillic It diffuses freely in the bilipid layer
of the inner mitochondrial membrane
The process of transferring electrons from NADH
to CoQ by complex I results in the net transport of
protons from the matrix side of the inner
mitochondrial membrane to the inter membrane space
where the H+ ions accumulate generating a proton
motive force The intermembrane space side of the
inner membrane is referred to as the P face (P
standing for positive) The matrix side of the inner
membrane is referred to as the the N face The
24
transport of electrons from NADH to CoQ is coupled to
the transport of protons across the membrane This is
an example of active transport The stiochiometry is
4 H+ transported per 2 electrons
2 Complex II
Believe it or not you are already familiar
with Complex II It is none other than succinate
dehydrogenase The only enzyme of the citric acid
cycle that is an integral membrane protein This
complex is composed of four subunits 2 of which are
iron-sulfur proteins and the other two subunits
together bind FAD through a covalent link to a
histidine residue These two subunits are called
flavoprotein 2 or FP2 Complex II contains 3 Fe-S
centers 1 4Fe-4S cluster 1 3Fe-4S cluster and 1
2Fe- 2S cluster In the first step of this complex
succinate is bound and a hydride is transferred to
FAD to generate FADH2 and fumarate FADH2 then
transfers its electrons one at a time to the Fe-S
centers Thus once again FAD functions as 2 electron
acceptor and a 1 electron donor The final step of
this complex is the transfer of 2 electrons one at a
time to coenzyme Q to produce CoQH2 For complex II
the standard free energy change of the overall
reaction is too small to drive the transport of
protons across the inner mitochondrial membrane This
25
accounts for the 15 ATPrsquos generated per FADH2
compared with the 25 ATPrsquos generated per NADH
3 Complex III
This complex is also known as coenzyme Q-
cytochrome c reductase because it passes the
electrons form CoQH2 to cyt c through a very unique
electron transport pathway called the Q-cycle Shown
to the left are the porphyrins found in cytochromes
Cytochrome b contains the same iron protoporphyrin as
hemoglobin and myoglobin The c cytochromes contain
heme c through covalent attachment by cysteine
residues Cytochrome a is found in two forms in
complex IV In complex III we find two b-type
cytochromes and one c-type cytochrome Complex III is
complex and we have a crystal structure
4 Complex IV
Complex IV is also known as cytochrome c oxidase
because it accepts the electrons from cytochrome c
and directs them towards the four electron reduction
of O2 to form 2 molecules of H2O
4 cyt c (Fe2+) + 4 H+ + O2 4 cyt c (Fe3+) +
2H2O
26
Cytochrome c oxidase contains 2 heme
centerscytochrome a and cytochrome a3 and two copper
proteins Each of the protein bound coppers are
associated with one of the cytochromes The copper
sites are called CuA and CuB CuA is associated with
cytochrome a and is shown to the left CuB is
associated with cytochrome a3 The copper sites
function as 1 electron carriers cycling between the
cuprous state Cu+ and the cupric state Cu2+ Just
like iron containing proteins they transfer
electrons one at a time Cytochrome c is bound on the
P-face of the membrane and transfers its electron to
CuA The oxidized cytochrome c dissociates CuA then
transfers the electron to cytochrome a The protein
bound CuA and the iron bound in cytochrome a are 15 Aring
apart In contrast the CuB and the iron bound in
cytochrome a3 are very close to each other forming a
binuclear metal center shown below
27
Electron transport chains are associated with
membranes such as the mitochondrial membrane in
eukaryotic organisms the thylakoid membrane of
photosynthetic organisms and the plasma membrane of
prokaryotic organisms While some of the details
differ electron transport chains in all organisms
operate on the same basic principles
Excited electrons are brought to the electron
transport chain by electron carriers such as NADH
and FADH2 In photosynthetic organisms the electrons
are donated by water and excited by light energy
The electron donor is oxidized as the electrons are
passed to the first protein complex in the chain As
it accepts the electrons the protein complex is
reduced Next the electrons are passed from the
28
first protein complex which becomes oxidized to
the second protein complex which becomes reduced
Each of these redox reactions is slightly exergonic
or energy releasing Some of this energy is used to
move protons across the membrane against their
concentration gradient With each successive
transfer of electrons the original excited
electrons lose some of their energy After several
successive redox reactions the electrons are
donated to a terminal electron acceptor In aerobic
respiration the terminal electron acceptor is
oxygen In photosynthesis the terminal electron
acceptor is a compound called NADP
The energy in the excited electrons has been
transformed into an electrochemical gradient
represented by the high concentration of protons on
one side of the membrane These protons can diffuse
back across the membrane by passing through a
protein complex called ATP synthase As these
protons pass through ATP synthase down their
concentration gradient they provide the energy
needed to make ATP from ADP and inorganic phosphate
within the cell
29
CHAPTER III
CLOSING
A Conclution
When a hexose is completely oxidized to CO2
and H2O using these three processes Glycolysis
yields two ATP and two NADH per hexose used Each
such NADH oxidized by the electron transport system
30
yields two ATP as dscribed above so glycolysis
contributes a total of six ATP per hexose The Krebs
cycle contributes two ATP per hexose or per two
pyruvates when succinyl CoA is cleaved to succinate
and CoASH This cycle also produces eight NADH per
hexose within the mitochondrial matrix by oxidative
phosporylation each of these NADH yields three ATP
or 24 per hexose Each FADH2 from the Krebs cycle
yields two ATP by oxydative phosporylation or four
per hexose (two pyruvates) The total contribution of
the Krebs cycle is then 30 Adding these 30 to the 6
from glycolysis leads to a total of 36 per hexose
completely respired by these process
B Suggestion
My greatest wish is that the readers find
this manuscript helpful in gaining the knowledge they
need for effective learning and practice and in
preparing for examination designed to test that
knowledge Reader comments and suggestions are
welcome and would be very helpful in improving our
manuscript Your contribution may assist many other
students in the coming years
31
REFERENCE
Anonim 2013 Anaerobic Respiration
httpwwwscoolcoukgcsebiologyrespiration
revise-itanaerobic-respiration Accessed
September 10th 2013
Anonim 2013 What is the difference between aerobic
and anaerobic respiration in plants
httpwwwbiologylifeeasyorg4010difference
-between-aerobic-anaerobic-respiration-plants
Accessed September 10th 2013
Campbell Mary K 2006 Biochemistry New York Thomson
Brooks Cole Inc
Dat J Capelli N Folzer H Bourgeade P Badot P-M
(2004) Sensing and signaling during
plantflooding Plant Physiology and
Biochemistry 42 273-282
Dat J Folzer H Parent C Badot P-M Capelli N
(2006) HypoxiastressCurrent Understanding and
Perspectives In Teixeira da Silva JA (Ed)
Floriculture Ornamental and Plant
Biotechnology Advances and Topical Issues (Vol
3) Global Science Books Isleworth United
Kingdompp 664-674
Hatta Muhammad 2011 Dunia Pertanian
httpemhattawordpresscomcategorystress
Accessed September 10th 2013
Hopkins William G 2009 Introduction to Plant Physiology
New York John Wiley amp Sons Inc
Lakitan Benyamin 2011 Dasar-Dasar Fisiologi Tumbuhan
Jakarta PT Raja Grafindo Persada
Larcher W 1980 Physiological Plant Ecology New York
Springer-Verlag
Raven 2013 Biology Fifth Edition
httphigheredmcgraw-hillcomsites983409233
9student_view0chapter39
photosynthetic_electron_transport_and_atp_synth
esishtml Accessed September 10th 2013
Salisbury Frank B 1969 Plant Physiology Third Edition
California Wadsworth Publishing Company
Subbaiah C Sachs M (2003) Molecular and cellular
adaptations of maize to flooding stress Annals
of Botany 91 119-127
Wilson CarlL1962 Botany Third EditionUnited State of
America Holt Rinch Inc

the continued oxidation of glyceraldehyde-3-P if
this were the case glycolysis and the producton of
even the small quantities of ATP necessary to
maintain the cells under anaerobic conditions would
then grind to a halt (Hopkins 2009 241)
Soil waterlogging has long been identified as a
major abiotic stress and the constraints that it
provides the roots have a significant effect on the
growth and development of plants When this event
occurs in the spring then these puddles can reduce
seed germination and seedling development Thus a
pool of water is an important factor that affects
the growth development and survival of plant
species not only in natural ecosystems but also
the agricultural and horticultural systems (Dat et
al 2006)
After flooding rapid changes in soil
properties At the time of pore water meets the
land the air pushed out gas diffusion is reduced
and toxic compounds accumulate due to anaerobic
conditions All of these changes greatly affect the
ability of plants to survive In response increased
stomatal resistance photosynthesis and root
hydraulic conductivity decreases and reduced
translocation fotoassimilat Nevertheless one of
15
the best adaptations of plants to hypoxia anoxia
is transitional biochemical and metabolic processes
common to current limited availability of O2 (Dat et
al 2004)
Selective synthesis of a set of about 20
anaerobic stress proteins (ANPS) allows the process
of producing energy without oxygen metabolism under
anaerobic conditions (Subbaiah and Sachs 2003)
Another adaptation is the observed morphological
changes consisting of the formation of hypertrophic
lenticels adventitious root initiation and or
progression of aerenchyma (Vartapetian and Jackson
1997 Jackson and Colmer 2005 Folzer et al 2006)
This review details the diverse plant stress
response to hypoxia anoxia caused by soil
waterlogging flooding and examines some of the key
features of metabolic adaptation physiological and
morphological
16
C Electron
Transport System
We noted earlier that one of the principal
functions of the respiration is to retrieve in
useful form some of the energy initially stored in
assimilates Our traditional measure of useful
energy in most processes is the number of ATP
molecules gained or consumed By this measure alone
the yield from both glycolysis and the citric acid
cycle is quite low After two complete turns of the
cycle one molecule of glucose has been completely
oxidized to six molecules of CO2 but only four
molecules of ATP have been produced (a net of two
ATP from glycolysis plus one for each turn of the
cycle) At this point most of the energy associated
with the glucose molecule has been conserved in the
17
form of electron pairs generated by the oxidation of
glycolytic and citric acid cycle intermediates
In this section we will discuss the third stage
of cellular respirationmdashthe transfer of electrons
fromNADH and FADH2 to oxygen and the accompanying
conversion of redox energy to ATP The transfer of
electrons fromNADHand FADH2 to oxygen involves a
sequence of electron carriers arranged in an
electron transport chainMembrane fractionation
studies have shown that the enzymes and electron
carriers making up the electron transport chain are
organized predominantly into four large
multimolecular complexes (complexes ImdashIV) and two
mobile carriers located in the inner mitochondrial
membrane (Figure 109) In this sense there are a
great number of similarities between the
mitochondrial inner membrane and the thylakoid
membranes of the chloroplast (compare Figure 109
with Figure 76) This is not unexpected since the
principal function of each membrane is energy
transformation and many of the same or similar
components are involved
The Components of the Electron Transport Chain
The electron transport chain of the
mitochondria is the means by which electrons are
18
removed from the reduced carrier NADH and
transferred to oxygen to yield H2O
1) NADH
NADH is generated in the matrix by the reactions
of pyruvate dehydrogenase isocitrate
dehydrogenase α-ketoglutarate dehydrogenase and
malate dehyrogenase The electron transport chain
begins with reoxidizing NADH to form NAD+ and
channeling the electrons into the formation of
reduced coenzymes Important to note that NADH
transfers 2 electrons at a time in the form of a
hydride
NAD+ + 2e- + H+ 1048581 NADH Eorsquo = minus0315 V
2) Flavoproteins
Flavoproteins have either a FAD (flavin adenosine
dinucleotide) or a FMN (flavin mononucleotide)
prosthetic group Flavoproteins can accept or
donate electrons one at time or two at a time
Thus they are often intermediaries between two
electron acceptorsdonors and one electron
acceptorsdonors For flavoproteins the typical
standard reduction potentials are around 0 V
FAD + 2e- + 2H+ 1048581 FADH2 FMN + 2e- + 2H+ 1048581 FMNH2
Eorsquo asymp 0 V
19
3) Coenzyme Q (CoQ)
aka ubiquinone (UQ) Shown below CoQ has ten
repeating isoprene units which make it
insoluble in water but soluble in the
hydrophobic lipid bylayer Coenzyme Q is a
versatile cofactor because it is a soluble
electron carrier in the hydrophobic bilipid layer
of the
inner mitochondrial membrane Like flavoproteins
CoQ can acceptdonate electrons one at a time or
two at a time
Q + 2e- + 2H+ 1048581 QH2 Eorsquo =0060 V
Q + e- + H+ 1048581 1048581QH Eorsquo =0030 V
1048581QH + e- + H+ 1048581 QH2 Eorsquo =0190 V
4) Cytochromes
Cytochromes are proteins that contain heme
prosthetic groups which function as
one electron carriers The heme iron is involved
in one electron transfers involving the Fe2+ and
Fe3+ oxidation states Cytochromes are named by
their absorption spectra which depends
on the porphyrin structure and environment The
example shown is the heme prosthetic group of
cytochrome b Cytochrome b contains the same iron
porphyrin found in hemoglobin and myoglobin
20
Other cytochromes we will encounter in the
electron transport complexes are cytochromes b
c c1 a and a3
5) Iron-Sulfur Proteins
In the electron transport chain we will encounter
many iron-sulfer proteins which participate in
one electron transfers involving the the Fe2+ and
Fe3+ oxidation states These are non-heme iron-
sulfur proteinsThe simplest iron-sulfer protein
is FeS in which iron is tetrahedrally coordinated
by four cysteines The second form is Fe2S2 which
contains two irons complexed to 2 cysteine
residues and two inorganic sulfides The third
form is Fe3S4 which contains 3 iron atoms
coordinated to three cysteine residues and 4
inorganic sulfides The last form is the most
complicated Fe4S4 which contains 4 iron atoms
coordinated to 4 cysteine residues and 4
inorganic sulfides
6) Copper Proteins
Copper bound proteins participate in one electron
transfers involving the Cu+ and Cu2+ oxidation
states
21
Overview of the Electron Transport Chain
Electrons move along the electron transport
chain going from donor to acceptor until they reach
oxygen the ultimate electron acceptor The standard
reduction potentials of the electron carriers are
between the NADHNAD+ couple (-0315 V) and the
oxygenH2O couple (0816 V) as on the next page The
components of the electron transport chain are
organized into 4 complexes Each complex contains
several different electron carriers
1 Complex I also known as the NADH-coenzyme Q
reductase or NADH dehydrogenase
2 Complex II also known as succinate-coenzyme Q
reductase or succinate dehydrogenase
3 Complex III also known as coenzyme Q reductase
4 Complex IV also known as cytochrome c reductase
The free energy necessary to generate ATP
is extracted from the oxidation of NADH and
FADH2 by the electron transport chain It
consists of four protein complexes and single
protein chain of cytochrome c through which
electrons pass from lower to higher standard
redox potentials Electrons are carried from complex
I and II to complex III by the membrane (lipid)
soluble coenzyme Q (CoQ or ubiquinone) and between
22
complexes III and IV by the peripheral membrane
protein cytochrome c
In the following I give the overall
reactions catalysed by corresponding protein
complexes in the mitochondrial electron transport
chain
1 Complex I
Complex I is also called NADH-Coenzyme Q
reductase because this large protein complex
transfers 2 electrons from NADH to coenzyme Q
Complex I was formerly known as NADH dehydrogenase
Complex I is huge 850000 kD and is composed of more
than thirty subunits It contains a FMN prosthetic
group and seven or more Fe-S clusters This complex
has between 20-26 iron atoms boundThe prosthetic
group FMN is absolutely required for activity
Therefore this complex is a flavoprotein This
complex binds NADH transfers two electrons in the
form of a hydride to FMN to produce NAD+ and FMNH2
23
The subsequent steps involve the transfer of
electrons one at a time to a series of iron-sulfer
complexes that includes both 2Fe-2S and 4Fe-4S
clusters Note the importance of FMN First it
functions as a 2 electron acceptor in the hydride
transfer from NADH Second it functions as a 1
electron donor to the series of iron sulfur clusters
FMN and FAD often play crucial links between 2
electron transfer agents and 1 electron transfer
agents The final step of this complex is the
transfer of 2 electrons one at a time to coenzyme Q
CoQ like FMN and FAD can function as a 2 electron
donoracceptor and as a 1 electron donoracceptor
CoQ is a mobile electron carrier because its
isoprenoid tail makes it highly hydrophobic and
lipophillic It diffuses freely in the bilipid layer
of the inner mitochondrial membrane
The process of transferring electrons from NADH
to CoQ by complex I results in the net transport of
protons from the matrix side of the inner
mitochondrial membrane to the inter membrane space
where the H+ ions accumulate generating a proton
motive force The intermembrane space side of the
inner membrane is referred to as the P face (P
standing for positive) The matrix side of the inner
membrane is referred to as the the N face The
24
transport of electrons from NADH to CoQ is coupled to
the transport of protons across the membrane This is
an example of active transport The stiochiometry is
4 H+ transported per 2 electrons
2 Complex II
Believe it or not you are already familiar
with Complex II It is none other than succinate
dehydrogenase The only enzyme of the citric acid
cycle that is an integral membrane protein This
complex is composed of four subunits 2 of which are
iron-sulfur proteins and the other two subunits
together bind FAD through a covalent link to a
histidine residue These two subunits are called
flavoprotein 2 or FP2 Complex II contains 3 Fe-S
centers 1 4Fe-4S cluster 1 3Fe-4S cluster and 1
2Fe- 2S cluster In the first step of this complex
succinate is bound and a hydride is transferred to
FAD to generate FADH2 and fumarate FADH2 then
transfers its electrons one at a time to the Fe-S
centers Thus once again FAD functions as 2 electron
acceptor and a 1 electron donor The final step of
this complex is the transfer of 2 electrons one at a
time to coenzyme Q to produce CoQH2 For complex II
the standard free energy change of the overall
reaction is too small to drive the transport of
protons across the inner mitochondrial membrane This
25
accounts for the 15 ATPrsquos generated per FADH2
compared with the 25 ATPrsquos generated per NADH
3 Complex III
This complex is also known as coenzyme Q-
cytochrome c reductase because it passes the
electrons form CoQH2 to cyt c through a very unique
electron transport pathway called the Q-cycle Shown
to the left are the porphyrins found in cytochromes
Cytochrome b contains the same iron protoporphyrin as
hemoglobin and myoglobin The c cytochromes contain
heme c through covalent attachment by cysteine
residues Cytochrome a is found in two forms in
complex IV In complex III we find two b-type
cytochromes and one c-type cytochrome Complex III is
complex and we have a crystal structure
4 Complex IV
Complex IV is also known as cytochrome c oxidase
because it accepts the electrons from cytochrome c
and directs them towards the four electron reduction
of O2 to form 2 molecules of H2O
4 cyt c (Fe2+) + 4 H+ + O2 4 cyt c (Fe3+) +
2H2O
26
Cytochrome c oxidase contains 2 heme
centerscytochrome a and cytochrome a3 and two copper
proteins Each of the protein bound coppers are
associated with one of the cytochromes The copper
sites are called CuA and CuB CuA is associated with
cytochrome a and is shown to the left CuB is
associated with cytochrome a3 The copper sites
function as 1 electron carriers cycling between the
cuprous state Cu+ and the cupric state Cu2+ Just
like iron containing proteins they transfer
electrons one at a time Cytochrome c is bound on the
P-face of the membrane and transfers its electron to
CuA The oxidized cytochrome c dissociates CuA then
transfers the electron to cytochrome a The protein
bound CuA and the iron bound in cytochrome a are 15 Aring
apart In contrast the CuB and the iron bound in
cytochrome a3 are very close to each other forming a
binuclear metal center shown below
27
Electron transport chains are associated with
membranes such as the mitochondrial membrane in
eukaryotic organisms the thylakoid membrane of
photosynthetic organisms and the plasma membrane of
prokaryotic organisms While some of the details
differ electron transport chains in all organisms
operate on the same basic principles
Excited electrons are brought to the electron
transport chain by electron carriers such as NADH
and FADH2 In photosynthetic organisms the electrons
are donated by water and excited by light energy
The electron donor is oxidized as the electrons are
passed to the first protein complex in the chain As
it accepts the electrons the protein complex is
reduced Next the electrons are passed from the
28
first protein complex which becomes oxidized to
the second protein complex which becomes reduced
Each of these redox reactions is slightly exergonic
or energy releasing Some of this energy is used to
move protons across the membrane against their
concentration gradient With each successive
transfer of electrons the original excited
electrons lose some of their energy After several
successive redox reactions the electrons are
donated to a terminal electron acceptor In aerobic
respiration the terminal electron acceptor is
oxygen In photosynthesis the terminal electron
acceptor is a compound called NADP
The energy in the excited electrons has been
transformed into an electrochemical gradient
represented by the high concentration of protons on
one side of the membrane These protons can diffuse
back across the membrane by passing through a
protein complex called ATP synthase As these
protons pass through ATP synthase down their
concentration gradient they provide the energy
needed to make ATP from ADP and inorganic phosphate
within the cell
29
CHAPTER III
CLOSING
A Conclution
When a hexose is completely oxidized to CO2
and H2O using these three processes Glycolysis
yields two ATP and two NADH per hexose used Each
such NADH oxidized by the electron transport system
30
yields two ATP as dscribed above so glycolysis
contributes a total of six ATP per hexose The Krebs
cycle contributes two ATP per hexose or per two
pyruvates when succinyl CoA is cleaved to succinate
and CoASH This cycle also produces eight NADH per
hexose within the mitochondrial matrix by oxidative
phosporylation each of these NADH yields three ATP
or 24 per hexose Each FADH2 from the Krebs cycle
yields two ATP by oxydative phosporylation or four
per hexose (two pyruvates) The total contribution of
the Krebs cycle is then 30 Adding these 30 to the 6
from glycolysis leads to a total of 36 per hexose
completely respired by these process
B Suggestion
My greatest wish is that the readers find
this manuscript helpful in gaining the knowledge they
need for effective learning and practice and in
preparing for examination designed to test that
knowledge Reader comments and suggestions are
welcome and would be very helpful in improving our
manuscript Your contribution may assist many other
students in the coming years
31
REFERENCE
Anonim 2013 Anaerobic Respiration
httpwwwscoolcoukgcsebiologyrespiration
revise-itanaerobic-respiration Accessed
September 10th 2013
Anonim 2013 What is the difference between aerobic
and anaerobic respiration in plants
httpwwwbiologylifeeasyorg4010difference
-between-aerobic-anaerobic-respiration-plants
Accessed September 10th 2013
Campbell Mary K 2006 Biochemistry New York Thomson
Brooks Cole Inc
Dat J Capelli N Folzer H Bourgeade P Badot P-M
(2004) Sensing and signaling during
plantflooding Plant Physiology and
Biochemistry 42 273-282
Dat J Folzer H Parent C Badot P-M Capelli N
(2006) HypoxiastressCurrent Understanding and
Perspectives In Teixeira da Silva JA (Ed)
Floriculture Ornamental and Plant
Biotechnology Advances and Topical Issues (Vol
3) Global Science Books Isleworth United
Kingdompp 664-674
Hatta Muhammad 2011 Dunia Pertanian
httpemhattawordpresscomcategorystress
Accessed September 10th 2013
Hopkins William G 2009 Introduction to Plant Physiology
New York John Wiley amp Sons Inc
Lakitan Benyamin 2011 Dasar-Dasar Fisiologi Tumbuhan
Jakarta PT Raja Grafindo Persada
Larcher W 1980 Physiological Plant Ecology New York
Springer-Verlag
Raven 2013 Biology Fifth Edition
httphigheredmcgraw-hillcomsites983409233
9student_view0chapter39
photosynthetic_electron_transport_and_atp_synth
esishtml Accessed September 10th 2013
Salisbury Frank B 1969 Plant Physiology Third Edition
California Wadsworth Publishing Company
Subbaiah C Sachs M (2003) Molecular and cellular
adaptations of maize to flooding stress Annals
of Botany 91 119-127
Wilson CarlL1962 Botany Third EditionUnited State of
America Holt Rinch Inc

the best adaptations of plants to hypoxia anoxia
is transitional biochemical and metabolic processes
common to current limited availability of O2 (Dat et
al 2004)
Selective synthesis of a set of about 20
anaerobic stress proteins (ANPS) allows the process
of producing energy without oxygen metabolism under
anaerobic conditions (Subbaiah and Sachs 2003)
Another adaptation is the observed morphological
changes consisting of the formation of hypertrophic
lenticels adventitious root initiation and or
progression of aerenchyma (Vartapetian and Jackson
1997 Jackson and Colmer 2005 Folzer et al 2006)
This review details the diverse plant stress
response to hypoxia anoxia caused by soil
waterlogging flooding and examines some of the key
features of metabolic adaptation physiological and
morphological
16
C Electron
Transport System
We noted earlier that one of the principal
functions of the respiration is to retrieve in
useful form some of the energy initially stored in
assimilates Our traditional measure of useful
energy in most processes is the number of ATP
molecules gained or consumed By this measure alone
the yield from both glycolysis and the citric acid
cycle is quite low After two complete turns of the
cycle one molecule of glucose has been completely
oxidized to six molecules of CO2 but only four
molecules of ATP have been produced (a net of two
ATP from glycolysis plus one for each turn of the
cycle) At this point most of the energy associated
with the glucose molecule has been conserved in the
17
form of electron pairs generated by the oxidation of
glycolytic and citric acid cycle intermediates
In this section we will discuss the third stage
of cellular respirationmdashthe transfer of electrons
fromNADH and FADH2 to oxygen and the accompanying
conversion of redox energy to ATP The transfer of
electrons fromNADHand FADH2 to oxygen involves a
sequence of electron carriers arranged in an
electron transport chainMembrane fractionation
studies have shown that the enzymes and electron
carriers making up the electron transport chain are
organized predominantly into four large
multimolecular complexes (complexes ImdashIV) and two
mobile carriers located in the inner mitochondrial
membrane (Figure 109) In this sense there are a
great number of similarities between the
mitochondrial inner membrane and the thylakoid
membranes of the chloroplast (compare Figure 109
with Figure 76) This is not unexpected since the
principal function of each membrane is energy
transformation and many of the same or similar
components are involved
The Components of the Electron Transport Chain
The electron transport chain of the
mitochondria is the means by which electrons are
18
removed from the reduced carrier NADH and
transferred to oxygen to yield H2O
1) NADH
NADH is generated in the matrix by the reactions
of pyruvate dehydrogenase isocitrate
dehydrogenase α-ketoglutarate dehydrogenase and
malate dehyrogenase The electron transport chain
begins with reoxidizing NADH to form NAD+ and
channeling the electrons into the formation of
reduced coenzymes Important to note that NADH
transfers 2 electrons at a time in the form of a
hydride
NAD+ + 2e- + H+ 1048581 NADH Eorsquo = minus0315 V
2) Flavoproteins
Flavoproteins have either a FAD (flavin adenosine
dinucleotide) or a FMN (flavin mononucleotide)
prosthetic group Flavoproteins can accept or
donate electrons one at time or two at a time
Thus they are often intermediaries between two
electron acceptorsdonors and one electron
acceptorsdonors For flavoproteins the typical
standard reduction potentials are around 0 V
FAD + 2e- + 2H+ 1048581 FADH2 FMN + 2e- + 2H+ 1048581 FMNH2
Eorsquo asymp 0 V
19
3) Coenzyme Q (CoQ)
aka ubiquinone (UQ) Shown below CoQ has ten
repeating isoprene units which make it
insoluble in water but soluble in the
hydrophobic lipid bylayer Coenzyme Q is a
versatile cofactor because it is a soluble
electron carrier in the hydrophobic bilipid layer
of the
inner mitochondrial membrane Like flavoproteins
CoQ can acceptdonate electrons one at a time or
two at a time
Q + 2e- + 2H+ 1048581 QH2 Eorsquo =0060 V
Q + e- + H+ 1048581 1048581QH Eorsquo =0030 V
1048581QH + e- + H+ 1048581 QH2 Eorsquo =0190 V
4) Cytochromes
Cytochromes are proteins that contain heme
prosthetic groups which function as
one electron carriers The heme iron is involved
in one electron transfers involving the Fe2+ and
Fe3+ oxidation states Cytochromes are named by
their absorption spectra which depends
on the porphyrin structure and environment The
example shown is the heme prosthetic group of
cytochrome b Cytochrome b contains the same iron
porphyrin found in hemoglobin and myoglobin
20
Other cytochromes we will encounter in the
electron transport complexes are cytochromes b
c c1 a and a3
5) Iron-Sulfur Proteins
In the electron transport chain we will encounter
many iron-sulfer proteins which participate in
one electron transfers involving the the Fe2+ and
Fe3+ oxidation states These are non-heme iron-
sulfur proteinsThe simplest iron-sulfer protein
is FeS in which iron is tetrahedrally coordinated
by four cysteines The second form is Fe2S2 which
contains two irons complexed to 2 cysteine
residues and two inorganic sulfides The third
form is Fe3S4 which contains 3 iron atoms
coordinated to three cysteine residues and 4
inorganic sulfides The last form is the most
complicated Fe4S4 which contains 4 iron atoms
coordinated to 4 cysteine residues and 4
inorganic sulfides
6) Copper Proteins
Copper bound proteins participate in one electron
transfers involving the Cu+ and Cu2+ oxidation
states
21
Overview of the Electron Transport Chain
Electrons move along the electron transport
chain going from donor to acceptor until they reach
oxygen the ultimate electron acceptor The standard
reduction potentials of the electron carriers are
between the NADHNAD+ couple (-0315 V) and the
oxygenH2O couple (0816 V) as on the next page The
components of the electron transport chain are
organized into 4 complexes Each complex contains
several different electron carriers
1 Complex I also known as the NADH-coenzyme Q
reductase or NADH dehydrogenase
2 Complex II also known as succinate-coenzyme Q
reductase or succinate dehydrogenase
3 Complex III also known as coenzyme Q reductase
4 Complex IV also known as cytochrome c reductase
The free energy necessary to generate ATP
is extracted from the oxidation of NADH and
FADH2 by the electron transport chain It
consists of four protein complexes and single
protein chain of cytochrome c through which
electrons pass from lower to higher standard
redox potentials Electrons are carried from complex
I and II to complex III by the membrane (lipid)
soluble coenzyme Q (CoQ or ubiquinone) and between
22
complexes III and IV by the peripheral membrane
protein cytochrome c
In the following I give the overall
reactions catalysed by corresponding protein
complexes in the mitochondrial electron transport
chain
1 Complex I
Complex I is also called NADH-Coenzyme Q
reductase because this large protein complex
transfers 2 electrons from NADH to coenzyme Q
Complex I was formerly known as NADH dehydrogenase
Complex I is huge 850000 kD and is composed of more
than thirty subunits It contains a FMN prosthetic
group and seven or more Fe-S clusters This complex
has between 20-26 iron atoms boundThe prosthetic
group FMN is absolutely required for activity
Therefore this complex is a flavoprotein This
complex binds NADH transfers two electrons in the
form of a hydride to FMN to produce NAD+ and FMNH2
23
The subsequent steps involve the transfer of
electrons one at a time to a series of iron-sulfer
complexes that includes both 2Fe-2S and 4Fe-4S
clusters Note the importance of FMN First it
functions as a 2 electron acceptor in the hydride
transfer from NADH Second it functions as a 1
electron donor to the series of iron sulfur clusters
FMN and FAD often play crucial links between 2
electron transfer agents and 1 electron transfer
agents The final step of this complex is the
transfer of 2 electrons one at a time to coenzyme Q
CoQ like FMN and FAD can function as a 2 electron
donoracceptor and as a 1 electron donoracceptor
CoQ is a mobile electron carrier because its
isoprenoid tail makes it highly hydrophobic and
lipophillic It diffuses freely in the bilipid layer
of the inner mitochondrial membrane
The process of transferring electrons from NADH
to CoQ by complex I results in the net transport of
protons from the matrix side of the inner
mitochondrial membrane to the inter membrane space
where the H+ ions accumulate generating a proton
motive force The intermembrane space side of the
inner membrane is referred to as the P face (P
standing for positive) The matrix side of the inner
membrane is referred to as the the N face The
24
transport of electrons from NADH to CoQ is coupled to
the transport of protons across the membrane This is
an example of active transport The stiochiometry is
4 H+ transported per 2 electrons
2 Complex II
Believe it or not you are already familiar
with Complex II It is none other than succinate
dehydrogenase The only enzyme of the citric acid
cycle that is an integral membrane protein This
complex is composed of four subunits 2 of which are
iron-sulfur proteins and the other two subunits
together bind FAD through a covalent link to a
histidine residue These two subunits are called
flavoprotein 2 or FP2 Complex II contains 3 Fe-S
centers 1 4Fe-4S cluster 1 3Fe-4S cluster and 1
2Fe- 2S cluster In the first step of this complex
succinate is bound and a hydride is transferred to
FAD to generate FADH2 and fumarate FADH2 then
transfers its electrons one at a time to the Fe-S
centers Thus once again FAD functions as 2 electron
acceptor and a 1 electron donor The final step of
this complex is the transfer of 2 electrons one at a
time to coenzyme Q to produce CoQH2 For complex II
the standard free energy change of the overall
reaction is too small to drive the transport of
protons across the inner mitochondrial membrane This
25
accounts for the 15 ATPrsquos generated per FADH2
compared with the 25 ATPrsquos generated per NADH
3 Complex III
This complex is also known as coenzyme Q-
cytochrome c reductase because it passes the
electrons form CoQH2 to cyt c through a very unique
electron transport pathway called the Q-cycle Shown
to the left are the porphyrins found in cytochromes
Cytochrome b contains the same iron protoporphyrin as
hemoglobin and myoglobin The c cytochromes contain
heme c through covalent attachment by cysteine
residues Cytochrome a is found in two forms in
complex IV In complex III we find two b-type
cytochromes and one c-type cytochrome Complex III is
complex and we have a crystal structure
4 Complex IV
Complex IV is also known as cytochrome c oxidase
because it accepts the electrons from cytochrome c
and directs them towards the four electron reduction
of O2 to form 2 molecules of H2O
4 cyt c (Fe2+) + 4 H+ + O2 4 cyt c (Fe3+) +
2H2O
26
Cytochrome c oxidase contains 2 heme
centerscytochrome a and cytochrome a3 and two copper
proteins Each of the protein bound coppers are
associated with one of the cytochromes The copper
sites are called CuA and CuB CuA is associated with
cytochrome a and is shown to the left CuB is
associated with cytochrome a3 The copper sites
function as 1 electron carriers cycling between the
cuprous state Cu+ and the cupric state Cu2+ Just
like iron containing proteins they transfer
electrons one at a time Cytochrome c is bound on the
P-face of the membrane and transfers its electron to
CuA The oxidized cytochrome c dissociates CuA then
transfers the electron to cytochrome a The protein
bound CuA and the iron bound in cytochrome a are 15 Aring
apart In contrast the CuB and the iron bound in
cytochrome a3 are very close to each other forming a
binuclear metal center shown below
27
Electron transport chains are associated with
membranes such as the mitochondrial membrane in
eukaryotic organisms the thylakoid membrane of
photosynthetic organisms and the plasma membrane of
prokaryotic organisms While some of the details
differ electron transport chains in all organisms
operate on the same basic principles
Excited electrons are brought to the electron
transport chain by electron carriers such as NADH
and FADH2 In photosynthetic organisms the electrons
are donated by water and excited by light energy
The electron donor is oxidized as the electrons are
passed to the first protein complex in the chain As
it accepts the electrons the protein complex is
reduced Next the electrons are passed from the
28
first protein complex which becomes oxidized to
the second protein complex which becomes reduced
Each of these redox reactions is slightly exergonic
or energy releasing Some of this energy is used to
move protons across the membrane against their
concentration gradient With each successive
transfer of electrons the original excited
electrons lose some of their energy After several
successive redox reactions the electrons are
donated to a terminal electron acceptor In aerobic
respiration the terminal electron acceptor is
oxygen In photosynthesis the terminal electron
acceptor is a compound called NADP
The energy in the excited electrons has been
transformed into an electrochemical gradient
represented by the high concentration of protons on
one side of the membrane These protons can diffuse
back across the membrane by passing through a
protein complex called ATP synthase As these
protons pass through ATP synthase down their
concentration gradient they provide the energy
needed to make ATP from ADP and inorganic phosphate
within the cell
29
CHAPTER III
CLOSING
A Conclution
When a hexose is completely oxidized to CO2
and H2O using these three processes Glycolysis
yields two ATP and two NADH per hexose used Each
such NADH oxidized by the electron transport system
30
yields two ATP as dscribed above so glycolysis
contributes a total of six ATP per hexose The Krebs
cycle contributes two ATP per hexose or per two
pyruvates when succinyl CoA is cleaved to succinate
and CoASH This cycle also produces eight NADH per
hexose within the mitochondrial matrix by oxidative
phosporylation each of these NADH yields three ATP
or 24 per hexose Each FADH2 from the Krebs cycle
yields two ATP by oxydative phosporylation or four
per hexose (two pyruvates) The total contribution of
the Krebs cycle is then 30 Adding these 30 to the 6
from glycolysis leads to a total of 36 per hexose
completely respired by these process
B Suggestion
My greatest wish is that the readers find
this manuscript helpful in gaining the knowledge they
need for effective learning and practice and in
preparing for examination designed to test that
knowledge Reader comments and suggestions are
welcome and would be very helpful in improving our
manuscript Your contribution may assist many other
students in the coming years
31
REFERENCE
Anonim 2013 Anaerobic Respiration
httpwwwscoolcoukgcsebiologyrespiration
revise-itanaerobic-respiration Accessed
September 10th 2013
Anonim 2013 What is the difference between aerobic
and anaerobic respiration in plants
httpwwwbiologylifeeasyorg4010difference
-between-aerobic-anaerobic-respiration-plants
Accessed September 10th 2013
Campbell Mary K 2006 Biochemistry New York Thomson
Brooks Cole Inc
Dat J Capelli N Folzer H Bourgeade P Badot P-M
(2004) Sensing and signaling during
plantflooding Plant Physiology and
Biochemistry 42 273-282
Dat J Folzer H Parent C Badot P-M Capelli N
(2006) HypoxiastressCurrent Understanding and
Perspectives In Teixeira da Silva JA (Ed)
Floriculture Ornamental and Plant
Biotechnology Advances and Topical Issues (Vol
3) Global Science Books Isleworth United
Kingdompp 664-674
Hatta Muhammad 2011 Dunia Pertanian
httpemhattawordpresscomcategorystress
Accessed September 10th 2013
Hopkins William G 2009 Introduction to Plant Physiology
New York John Wiley amp Sons Inc
Lakitan Benyamin 2011 Dasar-Dasar Fisiologi Tumbuhan
Jakarta PT Raja Grafindo Persada
Larcher W 1980 Physiological Plant Ecology New York
Springer-Verlag
Raven 2013 Biology Fifth Edition
httphigheredmcgraw-hillcomsites983409233
9student_view0chapter39
photosynthetic_electron_transport_and_atp_synth
esishtml Accessed September 10th 2013
Salisbury Frank B 1969 Plant Physiology Third Edition
California Wadsworth Publishing Company
Subbaiah C Sachs M (2003) Molecular and cellular
adaptations of maize to flooding stress Annals
of Botany 91 119-127
Wilson CarlL1962 Botany Third EditionUnited State of
America Holt Rinch Inc
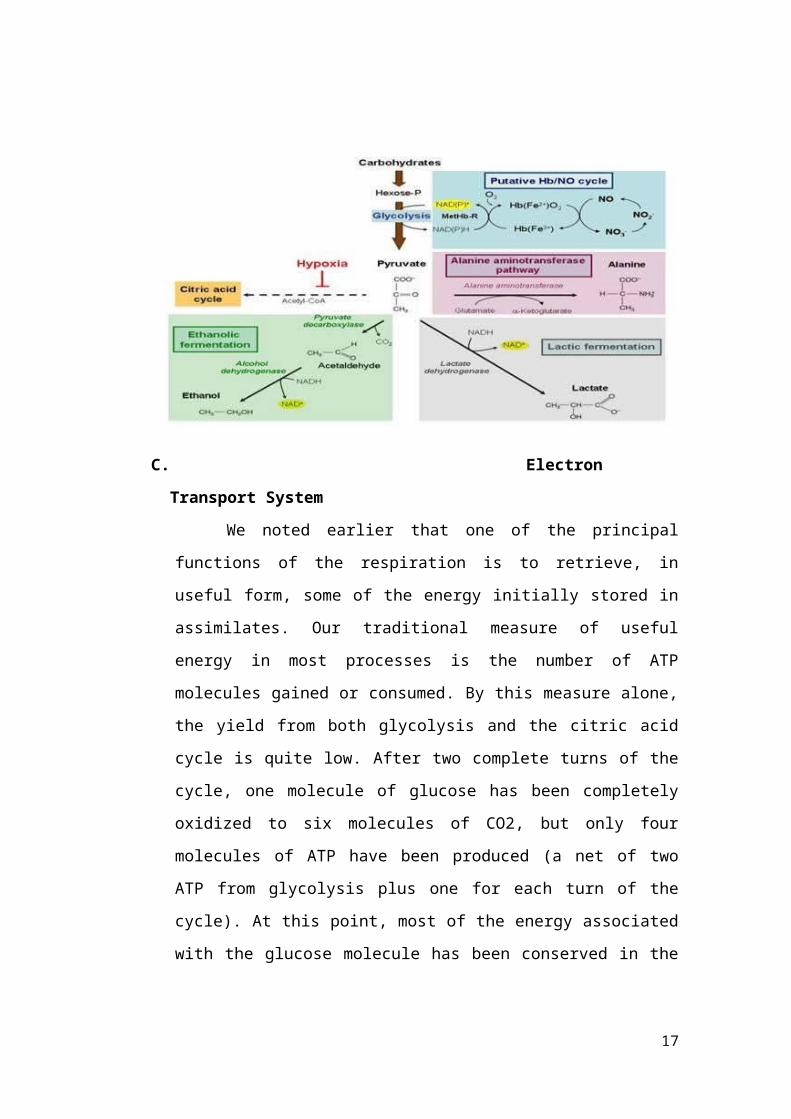
C Electron
Transport System
We noted earlier that one of the principal
functions of the respiration is to retrieve in
useful form some of the energy initially stored in
assimilates Our traditional measure of useful
energy in most processes is the number of ATP
molecules gained or consumed By this measure alone
the yield from both glycolysis and the citric acid
cycle is quite low After two complete turns of the
cycle one molecule of glucose has been completely
oxidized to six molecules of CO2 but only four
molecules of ATP have been produced (a net of two
ATP from glycolysis plus one for each turn of the
cycle) At this point most of the energy associated
with the glucose molecule has been conserved in the
17
form of electron pairs generated by the oxidation of
glycolytic and citric acid cycle intermediates
In this section we will discuss the third stage
of cellular respirationmdashthe transfer of electrons
fromNADH and FADH2 to oxygen and the accompanying
conversion of redox energy to ATP The transfer of
electrons fromNADHand FADH2 to oxygen involves a
sequence of electron carriers arranged in an
electron transport chainMembrane fractionation
studies have shown that the enzymes and electron
carriers making up the electron transport chain are
organized predominantly into four large
multimolecular complexes (complexes ImdashIV) and two
mobile carriers located in the inner mitochondrial
membrane (Figure 109) In this sense there are a
great number of similarities between the
mitochondrial inner membrane and the thylakoid
membranes of the chloroplast (compare Figure 109
with Figure 76) This is not unexpected since the
principal function of each membrane is energy
transformation and many of the same or similar
components are involved
The Components of the Electron Transport Chain
The electron transport chain of the
mitochondria is the means by which electrons are
18
removed from the reduced carrier NADH and
transferred to oxygen to yield H2O
1) NADH
NADH is generated in the matrix by the reactions
of pyruvate dehydrogenase isocitrate
dehydrogenase α-ketoglutarate dehydrogenase and
malate dehyrogenase The electron transport chain
begins with reoxidizing NADH to form NAD+ and
channeling the electrons into the formation of
reduced coenzymes Important to note that NADH
transfers 2 electrons at a time in the form of a
hydride
NAD+ + 2e- + H+ 1048581 NADH Eorsquo = minus0315 V
2) Flavoproteins
Flavoproteins have either a FAD (flavin adenosine
dinucleotide) or a FMN (flavin mononucleotide)
prosthetic group Flavoproteins can accept or
donate electrons one at time or two at a time
Thus they are often intermediaries between two
electron acceptorsdonors and one electron
acceptorsdonors For flavoproteins the typical
standard reduction potentials are around 0 V
FAD + 2e- + 2H+ 1048581 FADH2 FMN + 2e- + 2H+ 1048581 FMNH2
Eorsquo asymp 0 V
19
3) Coenzyme Q (CoQ)
aka ubiquinone (UQ) Shown below CoQ has ten
repeating isoprene units which make it
insoluble in water but soluble in the
hydrophobic lipid bylayer Coenzyme Q is a
versatile cofactor because it is a soluble
electron carrier in the hydrophobic bilipid layer
of the
inner mitochondrial membrane Like flavoproteins
CoQ can acceptdonate electrons one at a time or
two at a time
Q + 2e- + 2H+ 1048581 QH2 Eorsquo =0060 V
Q + e- + H+ 1048581 1048581QH Eorsquo =0030 V
1048581QH + e- + H+ 1048581 QH2 Eorsquo =0190 V
4) Cytochromes
Cytochromes are proteins that contain heme
prosthetic groups which function as
one electron carriers The heme iron is involved
in one electron transfers involving the Fe2+ and
Fe3+ oxidation states Cytochromes are named by
their absorption spectra which depends
on the porphyrin structure and environment The
example shown is the heme prosthetic group of
cytochrome b Cytochrome b contains the same iron
porphyrin found in hemoglobin and myoglobin
20
Other cytochromes we will encounter in the
electron transport complexes are cytochromes b
c c1 a and a3
5) Iron-Sulfur Proteins
In the electron transport chain we will encounter
many iron-sulfer proteins which participate in
one electron transfers involving the the Fe2+ and
Fe3+ oxidation states These are non-heme iron-
sulfur proteinsThe simplest iron-sulfer protein
is FeS in which iron is tetrahedrally coordinated
by four cysteines The second form is Fe2S2 which
contains two irons complexed to 2 cysteine
residues and two inorganic sulfides The third
form is Fe3S4 which contains 3 iron atoms
coordinated to three cysteine residues and 4
inorganic sulfides The last form is the most
complicated Fe4S4 which contains 4 iron atoms
coordinated to 4 cysteine residues and 4
inorganic sulfides
6) Copper Proteins
Copper bound proteins participate in one electron
transfers involving the Cu+ and Cu2+ oxidation
states
21
Overview of the Electron Transport Chain
Electrons move along the electron transport
chain going from donor to acceptor until they reach
oxygen the ultimate electron acceptor The standard
reduction potentials of the electron carriers are
between the NADHNAD+ couple (-0315 V) and the
oxygenH2O couple (0816 V) as on the next page The
components of the electron transport chain are
organized into 4 complexes Each complex contains
several different electron carriers
1 Complex I also known as the NADH-coenzyme Q
reductase or NADH dehydrogenase
2 Complex II also known as succinate-coenzyme Q
reductase or succinate dehydrogenase
3 Complex III also known as coenzyme Q reductase
4 Complex IV also known as cytochrome c reductase
The free energy necessary to generate ATP
is extracted from the oxidation of NADH and
FADH2 by the electron transport chain It
consists of four protein complexes and single
protein chain of cytochrome c through which
electrons pass from lower to higher standard
redox potentials Electrons are carried from complex
I and II to complex III by the membrane (lipid)
soluble coenzyme Q (CoQ or ubiquinone) and between
22
complexes III and IV by the peripheral membrane
protein cytochrome c
In the following I give the overall
reactions catalysed by corresponding protein
complexes in the mitochondrial electron transport
chain
1 Complex I
Complex I is also called NADH-Coenzyme Q
reductase because this large protein complex
transfers 2 electrons from NADH to coenzyme Q
Complex I was formerly known as NADH dehydrogenase
Complex I is huge 850000 kD and is composed of more
than thirty subunits It contains a FMN prosthetic
group and seven or more Fe-S clusters This complex
has between 20-26 iron atoms boundThe prosthetic
group FMN is absolutely required for activity
Therefore this complex is a flavoprotein This
complex binds NADH transfers two electrons in the
form of a hydride to FMN to produce NAD+ and FMNH2
23
The subsequent steps involve the transfer of
electrons one at a time to a series of iron-sulfer
complexes that includes both 2Fe-2S and 4Fe-4S
clusters Note the importance of FMN First it
functions as a 2 electron acceptor in the hydride
transfer from NADH Second it functions as a 1
electron donor to the series of iron sulfur clusters
FMN and FAD often play crucial links between 2
electron transfer agents and 1 electron transfer
agents The final step of this complex is the
transfer of 2 electrons one at a time to coenzyme Q
CoQ like FMN and FAD can function as a 2 electron
donoracceptor and as a 1 electron donoracceptor
CoQ is a mobile electron carrier because its
isoprenoid tail makes it highly hydrophobic and
lipophillic It diffuses freely in the bilipid layer
of the inner mitochondrial membrane
The process of transferring electrons from NADH
to CoQ by complex I results in the net transport of
protons from the matrix side of the inner
mitochondrial membrane to the inter membrane space
where the H+ ions accumulate generating a proton
motive force The intermembrane space side of the
inner membrane is referred to as the P face (P
standing for positive) The matrix side of the inner
membrane is referred to as the the N face The
24
transport of electrons from NADH to CoQ is coupled to
the transport of protons across the membrane This is
an example of active transport The stiochiometry is
4 H+ transported per 2 electrons
2 Complex II
Believe it or not you are already familiar
with Complex II It is none other than succinate
dehydrogenase The only enzyme of the citric acid
cycle that is an integral membrane protein This
complex is composed of four subunits 2 of which are
iron-sulfur proteins and the other two subunits
together bind FAD through a covalent link to a
histidine residue These two subunits are called
flavoprotein 2 or FP2 Complex II contains 3 Fe-S
centers 1 4Fe-4S cluster 1 3Fe-4S cluster and 1
2Fe- 2S cluster In the first step of this complex
succinate is bound and a hydride is transferred to
FAD to generate FADH2 and fumarate FADH2 then
transfers its electrons one at a time to the Fe-S
centers Thus once again FAD functions as 2 electron
acceptor and a 1 electron donor The final step of
this complex is the transfer of 2 electrons one at a
time to coenzyme Q to produce CoQH2 For complex II
the standard free energy change of the overall
reaction is too small to drive the transport of
protons across the inner mitochondrial membrane This
25
accounts for the 15 ATPrsquos generated per FADH2
compared with the 25 ATPrsquos generated per NADH
3 Complex III
This complex is also known as coenzyme Q-
cytochrome c reductase because it passes the
electrons form CoQH2 to cyt c through a very unique
electron transport pathway called the Q-cycle Shown
to the left are the porphyrins found in cytochromes
Cytochrome b contains the same iron protoporphyrin as
hemoglobin and myoglobin The c cytochromes contain
heme c through covalent attachment by cysteine
residues Cytochrome a is found in two forms in
complex IV In complex III we find two b-type
cytochromes and one c-type cytochrome Complex III is
complex and we have a crystal structure
4 Complex IV
Complex IV is also known as cytochrome c oxidase
because it accepts the electrons from cytochrome c
and directs them towards the four electron reduction
of O2 to form 2 molecules of H2O
4 cyt c (Fe2+) + 4 H+ + O2 4 cyt c (Fe3+) +
2H2O
26
Cytochrome c oxidase contains 2 heme
centerscytochrome a and cytochrome a3 and two copper
proteins Each of the protein bound coppers are
associated with one of the cytochromes The copper
sites are called CuA and CuB CuA is associated with
cytochrome a and is shown to the left CuB is
associated with cytochrome a3 The copper sites
function as 1 electron carriers cycling between the
cuprous state Cu+ and the cupric state Cu2+ Just
like iron containing proteins they transfer
electrons one at a time Cytochrome c is bound on the
P-face of the membrane and transfers its electron to
CuA The oxidized cytochrome c dissociates CuA then
transfers the electron to cytochrome a The protein
bound CuA and the iron bound in cytochrome a are 15 Aring
apart In contrast the CuB and the iron bound in
cytochrome a3 are very close to each other forming a
binuclear metal center shown below
27
Electron transport chains are associated with
membranes such as the mitochondrial membrane in
eukaryotic organisms the thylakoid membrane of
photosynthetic organisms and the plasma membrane of
prokaryotic organisms While some of the details
differ electron transport chains in all organisms
operate on the same basic principles
Excited electrons are brought to the electron
transport chain by electron carriers such as NADH
and FADH2 In photosynthetic organisms the electrons
are donated by water and excited by light energy
The electron donor is oxidized as the electrons are
passed to the first protein complex in the chain As
it accepts the electrons the protein complex is
reduced Next the electrons are passed from the
28
first protein complex which becomes oxidized to
the second protein complex which becomes reduced
Each of these redox reactions is slightly exergonic
or energy releasing Some of this energy is used to
move protons across the membrane against their
concentration gradient With each successive
transfer of electrons the original excited
electrons lose some of their energy After several
successive redox reactions the electrons are
donated to a terminal electron acceptor In aerobic
respiration the terminal electron acceptor is
oxygen In photosynthesis the terminal electron
acceptor is a compound called NADP
The energy in the excited electrons has been
transformed into an electrochemical gradient
represented by the high concentration of protons on
one side of the membrane These protons can diffuse
back across the membrane by passing through a
protein complex called ATP synthase As these
protons pass through ATP synthase down their
concentration gradient they provide the energy
needed to make ATP from ADP and inorganic phosphate
within the cell
29
CHAPTER III
CLOSING
A Conclution
When a hexose is completely oxidized to CO2
and H2O using these three processes Glycolysis
yields two ATP and two NADH per hexose used Each
such NADH oxidized by the electron transport system
30
yields two ATP as dscribed above so glycolysis
contributes a total of six ATP per hexose The Krebs
cycle contributes two ATP per hexose or per two
pyruvates when succinyl CoA is cleaved to succinate
and CoASH This cycle also produces eight NADH per
hexose within the mitochondrial matrix by oxidative
phosporylation each of these NADH yields three ATP
or 24 per hexose Each FADH2 from the Krebs cycle
yields two ATP by oxydative phosporylation or four
per hexose (two pyruvates) The total contribution of
the Krebs cycle is then 30 Adding these 30 to the 6
from glycolysis leads to a total of 36 per hexose
completely respired by these process
B Suggestion
My greatest wish is that the readers find
this manuscript helpful in gaining the knowledge they
need for effective learning and practice and in
preparing for examination designed to test that
knowledge Reader comments and suggestions are
welcome and would be very helpful in improving our
manuscript Your contribution may assist many other
students in the coming years
31
REFERENCE
Anonim 2013 Anaerobic Respiration
httpwwwscoolcoukgcsebiologyrespiration
revise-itanaerobic-respiration Accessed
September 10th 2013
Anonim 2013 What is the difference between aerobic
and anaerobic respiration in plants
httpwwwbiologylifeeasyorg4010difference
-between-aerobic-anaerobic-respiration-plants
Accessed September 10th 2013
Campbell Mary K 2006 Biochemistry New York Thomson
Brooks Cole Inc
Dat J Capelli N Folzer H Bourgeade P Badot P-M
(2004) Sensing and signaling during
plantflooding Plant Physiology and
Biochemistry 42 273-282
Dat J Folzer H Parent C Badot P-M Capelli N
(2006) HypoxiastressCurrent Understanding and
Perspectives In Teixeira da Silva JA (Ed)
Floriculture Ornamental and Plant
Biotechnology Advances and Topical Issues (Vol
3) Global Science Books Isleworth United
Kingdompp 664-674
Hatta Muhammad 2011 Dunia Pertanian
httpemhattawordpresscomcategorystress
Accessed September 10th 2013
Hopkins William G 2009 Introduction to Plant Physiology
New York John Wiley amp Sons Inc
Lakitan Benyamin 2011 Dasar-Dasar Fisiologi Tumbuhan
Jakarta PT Raja Grafindo Persada
Larcher W 1980 Physiological Plant Ecology New York
Springer-Verlag
Raven 2013 Biology Fifth Edition
httphigheredmcgraw-hillcomsites983409233
9student_view0chapter39
photosynthetic_electron_transport_and_atp_synth
esishtml Accessed September 10th 2013
Salisbury Frank B 1969 Plant Physiology Third Edition
California Wadsworth Publishing Company
Subbaiah C Sachs M (2003) Molecular and cellular
adaptations of maize to flooding stress Annals
of Botany 91 119-127
Wilson CarlL1962 Botany Third EditionUnited State of
America Holt Rinch Inc

form of electron pairs generated by the oxidation of
glycolytic and citric acid cycle intermediates
In this section we will discuss the third stage
of cellular respirationmdashthe transfer of electrons
fromNADH and FADH2 to oxygen and the accompanying
conversion of redox energy to ATP The transfer of
electrons fromNADHand FADH2 to oxygen involves a
sequence of electron carriers arranged in an
electron transport chainMembrane fractionation
studies have shown that the enzymes and electron
carriers making up the electron transport chain are
organized predominantly into four large
multimolecular complexes (complexes ImdashIV) and two
mobile carriers located in the inner mitochondrial
membrane (Figure 109) In this sense there are a
great number of similarities between the
mitochondrial inner membrane and the thylakoid
membranes of the chloroplast (compare Figure 109
with Figure 76) This is not unexpected since the
principal function of each membrane is energy
transformation and many of the same or similar
components are involved
The Components of the Electron Transport Chain
The electron transport chain of the
mitochondria is the means by which electrons are
18
removed from the reduced carrier NADH and
transferred to oxygen to yield H2O
1) NADH
NADH is generated in the matrix by the reactions
of pyruvate dehydrogenase isocitrate
dehydrogenase α-ketoglutarate dehydrogenase and
malate dehyrogenase The electron transport chain
begins with reoxidizing NADH to form NAD+ and
channeling the electrons into the formation of
reduced coenzymes Important to note that NADH
transfers 2 electrons at a time in the form of a
hydride
NAD+ + 2e- + H+ 1048581 NADH Eorsquo = minus0315 V
2) Flavoproteins
Flavoproteins have either a FAD (flavin adenosine
dinucleotide) or a FMN (flavin mononucleotide)
prosthetic group Flavoproteins can accept or
donate electrons one at time or two at a time
Thus they are often intermediaries between two
electron acceptorsdonors and one electron
acceptorsdonors For flavoproteins the typical
standard reduction potentials are around 0 V
FAD + 2e- + 2H+ 1048581 FADH2 FMN + 2e- + 2H+ 1048581 FMNH2
Eorsquo asymp 0 V
19
3) Coenzyme Q (CoQ)
aka ubiquinone (UQ) Shown below CoQ has ten
repeating isoprene units which make it
insoluble in water but soluble in the
hydrophobic lipid bylayer Coenzyme Q is a
versatile cofactor because it is a soluble
electron carrier in the hydrophobic bilipid layer
of the
inner mitochondrial membrane Like flavoproteins
CoQ can acceptdonate electrons one at a time or
two at a time
Q + 2e- + 2H+ 1048581 QH2 Eorsquo =0060 V
Q + e- + H+ 1048581 1048581QH Eorsquo =0030 V
1048581QH + e- + H+ 1048581 QH2 Eorsquo =0190 V
4) Cytochromes
Cytochromes are proteins that contain heme
prosthetic groups which function as
one electron carriers The heme iron is involved
in one electron transfers involving the Fe2+ and
Fe3+ oxidation states Cytochromes are named by
their absorption spectra which depends
on the porphyrin structure and environment The
example shown is the heme prosthetic group of
cytochrome b Cytochrome b contains the same iron
porphyrin found in hemoglobin and myoglobin
20
Other cytochromes we will encounter in the
electron transport complexes are cytochromes b
c c1 a and a3
5) Iron-Sulfur Proteins
In the electron transport chain we will encounter
many iron-sulfer proteins which participate in
one electron transfers involving the the Fe2+ and
Fe3+ oxidation states These are non-heme iron-
sulfur proteinsThe simplest iron-sulfer protein
is FeS in which iron is tetrahedrally coordinated
by four cysteines The second form is Fe2S2 which
contains two irons complexed to 2 cysteine
residues and two inorganic sulfides The third
form is Fe3S4 which contains 3 iron atoms
coordinated to three cysteine residues and 4
inorganic sulfides The last form is the most
complicated Fe4S4 which contains 4 iron atoms
coordinated to 4 cysteine residues and 4
inorganic sulfides
6) Copper Proteins
Copper bound proteins participate in one electron
transfers involving the Cu+ and Cu2+ oxidation
states
21
Overview of the Electron Transport Chain
Electrons move along the electron transport
chain going from donor to acceptor until they reach
oxygen the ultimate electron acceptor The standard
reduction potentials of the electron carriers are
between the NADHNAD+ couple (-0315 V) and the
oxygenH2O couple (0816 V) as on the next page The
components of the electron transport chain are
organized into 4 complexes Each complex contains
several different electron carriers
1 Complex I also known as the NADH-coenzyme Q
reductase or NADH dehydrogenase
2 Complex II also known as succinate-coenzyme Q
reductase or succinate dehydrogenase
3 Complex III also known as coenzyme Q reductase
4 Complex IV also known as cytochrome c reductase
The free energy necessary to generate ATP
is extracted from the oxidation of NADH and
FADH2 by the electron transport chain It
consists of four protein complexes and single
protein chain of cytochrome c through which
electrons pass from lower to higher standard
redox potentials Electrons are carried from complex
I and II to complex III by the membrane (lipid)
soluble coenzyme Q (CoQ or ubiquinone) and between
22
complexes III and IV by the peripheral membrane
protein cytochrome c
In the following I give the overall
reactions catalysed by corresponding protein
complexes in the mitochondrial electron transport
chain
1 Complex I
Complex I is also called NADH-Coenzyme Q
reductase because this large protein complex
transfers 2 electrons from NADH to coenzyme Q
Complex I was formerly known as NADH dehydrogenase
Complex I is huge 850000 kD and is composed of more
than thirty subunits It contains a FMN prosthetic
group and seven or more Fe-S clusters This complex
has between 20-26 iron atoms boundThe prosthetic
group FMN is absolutely required for activity
Therefore this complex is a flavoprotein This
complex binds NADH transfers two electrons in the
form of a hydride to FMN to produce NAD+ and FMNH2
23
The subsequent steps involve the transfer of
electrons one at a time to a series of iron-sulfer
complexes that includes both 2Fe-2S and 4Fe-4S
clusters Note the importance of FMN First it
functions as a 2 electron acceptor in the hydride
transfer from NADH Second it functions as a 1
electron donor to the series of iron sulfur clusters
FMN and FAD often play crucial links between 2
electron transfer agents and 1 electron transfer
agents The final step of this complex is the
transfer of 2 electrons one at a time to coenzyme Q
CoQ like FMN and FAD can function as a 2 electron
donoracceptor and as a 1 electron donoracceptor
CoQ is a mobile electron carrier because its
isoprenoid tail makes it highly hydrophobic and
lipophillic It diffuses freely in the bilipid layer
of the inner mitochondrial membrane
The process of transferring electrons from NADH
to CoQ by complex I results in the net transport of
protons from the matrix side of the inner
mitochondrial membrane to the inter membrane space
where the H+ ions accumulate generating a proton
motive force The intermembrane space side of the
inner membrane is referred to as the P face (P
standing for positive) The matrix side of the inner
membrane is referred to as the the N face The
24
transport of electrons from NADH to CoQ is coupled to
the transport of protons across the membrane This is
an example of active transport The stiochiometry is
4 H+ transported per 2 electrons
2 Complex II
Believe it or not you are already familiar
with Complex II It is none other than succinate
dehydrogenase The only enzyme of the citric acid
cycle that is an integral membrane protein This
complex is composed of four subunits 2 of which are
iron-sulfur proteins and the other two subunits
together bind FAD through a covalent link to a
histidine residue These two subunits are called
flavoprotein 2 or FP2 Complex II contains 3 Fe-S
centers 1 4Fe-4S cluster 1 3Fe-4S cluster and 1
2Fe- 2S cluster In the first step of this complex
succinate is bound and a hydride is transferred to
FAD to generate FADH2 and fumarate FADH2 then
transfers its electrons one at a time to the Fe-S
centers Thus once again FAD functions as 2 electron
acceptor and a 1 electron donor The final step of
this complex is the transfer of 2 electrons one at a
time to coenzyme Q to produce CoQH2 For complex II
the standard free energy change of the overall
reaction is too small to drive the transport of
protons across the inner mitochondrial membrane This
25
accounts for the 15 ATPrsquos generated per FADH2
compared with the 25 ATPrsquos generated per NADH
3 Complex III
This complex is also known as coenzyme Q-
cytochrome c reductase because it passes the
electrons form CoQH2 to cyt c through a very unique
electron transport pathway called the Q-cycle Shown
to the left are the porphyrins found in cytochromes
Cytochrome b contains the same iron protoporphyrin as
hemoglobin and myoglobin The c cytochromes contain
heme c through covalent attachment by cysteine
residues Cytochrome a is found in two forms in
complex IV In complex III we find two b-type
cytochromes and one c-type cytochrome Complex III is
complex and we have a crystal structure
4 Complex IV
Complex IV is also known as cytochrome c oxidase
because it accepts the electrons from cytochrome c
and directs them towards the four electron reduction
of O2 to form 2 molecules of H2O
4 cyt c (Fe2+) + 4 H+ + O2 4 cyt c (Fe3+) +
2H2O
26
Cytochrome c oxidase contains 2 heme
centerscytochrome a and cytochrome a3 and two copper
proteins Each of the protein bound coppers are
associated with one of the cytochromes The copper
sites are called CuA and CuB CuA is associated with
cytochrome a and is shown to the left CuB is
associated with cytochrome a3 The copper sites
function as 1 electron carriers cycling between the
cuprous state Cu+ and the cupric state Cu2+ Just
like iron containing proteins they transfer
electrons one at a time Cytochrome c is bound on the
P-face of the membrane and transfers its electron to
CuA The oxidized cytochrome c dissociates CuA then
transfers the electron to cytochrome a The protein
bound CuA and the iron bound in cytochrome a are 15 Aring
apart In contrast the CuB and the iron bound in
cytochrome a3 are very close to each other forming a
binuclear metal center shown below
27
Electron transport chains are associated with
membranes such as the mitochondrial membrane in
eukaryotic organisms the thylakoid membrane of
photosynthetic organisms and the plasma membrane of
prokaryotic organisms While some of the details
differ electron transport chains in all organisms
operate on the same basic principles
Excited electrons are brought to the electron
transport chain by electron carriers such as NADH
and FADH2 In photosynthetic organisms the electrons
are donated by water and excited by light energy
The electron donor is oxidized as the electrons are
passed to the first protein complex in the chain As
it accepts the electrons the protein complex is
reduced Next the electrons are passed from the
28
first protein complex which becomes oxidized to
the second protein complex which becomes reduced
Each of these redox reactions is slightly exergonic
or energy releasing Some of this energy is used to
move protons across the membrane against their
concentration gradient With each successive
transfer of electrons the original excited
electrons lose some of their energy After several
successive redox reactions the electrons are
donated to a terminal electron acceptor In aerobic
respiration the terminal electron acceptor is
oxygen In photosynthesis the terminal electron
acceptor is a compound called NADP
The energy in the excited electrons has been
transformed into an electrochemical gradient
represented by the high concentration of protons on
one side of the membrane These protons can diffuse
back across the membrane by passing through a
protein complex called ATP synthase As these
protons pass through ATP synthase down their
concentration gradient they provide the energy
needed to make ATP from ADP and inorganic phosphate
within the cell
29
CHAPTER III
CLOSING
A Conclution
When a hexose is completely oxidized to CO2
and H2O using these three processes Glycolysis
yields two ATP and two NADH per hexose used Each
such NADH oxidized by the electron transport system
30
yields two ATP as dscribed above so glycolysis
contributes a total of six ATP per hexose The Krebs
cycle contributes two ATP per hexose or per two
pyruvates when succinyl CoA is cleaved to succinate
and CoASH This cycle also produces eight NADH per
hexose within the mitochondrial matrix by oxidative
phosporylation each of these NADH yields three ATP
or 24 per hexose Each FADH2 from the Krebs cycle
yields two ATP by oxydative phosporylation or four
per hexose (two pyruvates) The total contribution of
the Krebs cycle is then 30 Adding these 30 to the 6
from glycolysis leads to a total of 36 per hexose
completely respired by these process
B Suggestion
My greatest wish is that the readers find
this manuscript helpful in gaining the knowledge they
need for effective learning and practice and in
preparing for examination designed to test that
knowledge Reader comments and suggestions are
welcome and would be very helpful in improving our
manuscript Your contribution may assist many other
students in the coming years
31
REFERENCE
Anonim 2013 Anaerobic Respiration
httpwwwscoolcoukgcsebiologyrespiration
revise-itanaerobic-respiration Accessed
September 10th 2013
Anonim 2013 What is the difference between aerobic
and anaerobic respiration in plants
httpwwwbiologylifeeasyorg4010difference
-between-aerobic-anaerobic-respiration-plants
Accessed September 10th 2013
Campbell Mary K 2006 Biochemistry New York Thomson
Brooks Cole Inc
Dat J Capelli N Folzer H Bourgeade P Badot P-M
(2004) Sensing and signaling during
plantflooding Plant Physiology and
Biochemistry 42 273-282
Dat J Folzer H Parent C Badot P-M Capelli N
(2006) HypoxiastressCurrent Understanding and
Perspectives In Teixeira da Silva JA (Ed)
Floriculture Ornamental and Plant
Biotechnology Advances and Topical Issues (Vol
3) Global Science Books Isleworth United
Kingdompp 664-674
Hatta Muhammad 2011 Dunia Pertanian
httpemhattawordpresscomcategorystress
Accessed September 10th 2013
Hopkins William G 2009 Introduction to Plant Physiology
New York John Wiley amp Sons Inc
Lakitan Benyamin 2011 Dasar-Dasar Fisiologi Tumbuhan
Jakarta PT Raja Grafindo Persada
Larcher W 1980 Physiological Plant Ecology New York
Springer-Verlag
Raven 2013 Biology Fifth Edition
httphigheredmcgraw-hillcomsites983409233
9student_view0chapter39
photosynthetic_electron_transport_and_atp_synth
esishtml Accessed September 10th 2013
Salisbury Frank B 1969 Plant Physiology Third Edition
California Wadsworth Publishing Company
Subbaiah C Sachs M (2003) Molecular and cellular
adaptations of maize to flooding stress Annals
of Botany 91 119-127
Wilson CarlL1962 Botany Third EditionUnited State of
America Holt Rinch Inc

removed from the reduced carrier NADH and
transferred to oxygen to yield H2O
1) NADH
NADH is generated in the matrix by the reactions
of pyruvate dehydrogenase isocitrate
dehydrogenase α-ketoglutarate dehydrogenase and
malate dehyrogenase The electron transport chain
begins with reoxidizing NADH to form NAD+ and
channeling the electrons into the formation of
reduced coenzymes Important to note that NADH
transfers 2 electrons at a time in the form of a
hydride
NAD+ + 2e- + H+ 1048581 NADH Eorsquo = minus0315 V
2) Flavoproteins
Flavoproteins have either a FAD (flavin adenosine
dinucleotide) or a FMN (flavin mononucleotide)
prosthetic group Flavoproteins can accept or
donate electrons one at time or two at a time
Thus they are often intermediaries between two
electron acceptorsdonors and one electron
acceptorsdonors For flavoproteins the typical
standard reduction potentials are around 0 V
FAD + 2e- + 2H+ 1048581 FADH2 FMN + 2e- + 2H+ 1048581 FMNH2
Eorsquo asymp 0 V
19
3) Coenzyme Q (CoQ)
aka ubiquinone (UQ) Shown below CoQ has ten
repeating isoprene units which make it
insoluble in water but soluble in the
hydrophobic lipid bylayer Coenzyme Q is a
versatile cofactor because it is a soluble
electron carrier in the hydrophobic bilipid layer
of the
inner mitochondrial membrane Like flavoproteins
CoQ can acceptdonate electrons one at a time or
two at a time
Q + 2e- + 2H+ 1048581 QH2 Eorsquo =0060 V
Q + e- + H+ 1048581 1048581QH Eorsquo =0030 V
1048581QH + e- + H+ 1048581 QH2 Eorsquo =0190 V
4) Cytochromes
Cytochromes are proteins that contain heme
prosthetic groups which function as
one electron carriers The heme iron is involved
in one electron transfers involving the Fe2+ and
Fe3+ oxidation states Cytochromes are named by
their absorption spectra which depends
on the porphyrin structure and environment The
example shown is the heme prosthetic group of
cytochrome b Cytochrome b contains the same iron
porphyrin found in hemoglobin and myoglobin
20
Other cytochromes we will encounter in the
electron transport complexes are cytochromes b
c c1 a and a3
5) Iron-Sulfur Proteins
In the electron transport chain we will encounter
many iron-sulfer proteins which participate in
one electron transfers involving the the Fe2+ and
Fe3+ oxidation states These are non-heme iron-
sulfur proteinsThe simplest iron-sulfer protein
is FeS in which iron is tetrahedrally coordinated
by four cysteines The second form is Fe2S2 which
contains two irons complexed to 2 cysteine
residues and two inorganic sulfides The third
form is Fe3S4 which contains 3 iron atoms
coordinated to three cysteine residues and 4
inorganic sulfides The last form is the most
complicated Fe4S4 which contains 4 iron atoms
coordinated to 4 cysteine residues and 4
inorganic sulfides
6) Copper Proteins
Copper bound proteins participate in one electron
transfers involving the Cu+ and Cu2+ oxidation
states
21
Overview of the Electron Transport Chain
Electrons move along the electron transport
chain going from donor to acceptor until they reach
oxygen the ultimate electron acceptor The standard
reduction potentials of the electron carriers are
between the NADHNAD+ couple (-0315 V) and the
oxygenH2O couple (0816 V) as on the next page The
components of the electron transport chain are
organized into 4 complexes Each complex contains
several different electron carriers
1 Complex I also known as the NADH-coenzyme Q
reductase or NADH dehydrogenase
2 Complex II also known as succinate-coenzyme Q
reductase or succinate dehydrogenase
3 Complex III also known as coenzyme Q reductase
4 Complex IV also known as cytochrome c reductase
The free energy necessary to generate ATP
is extracted from the oxidation of NADH and
FADH2 by the electron transport chain It
consists of four protein complexes and single
protein chain of cytochrome c through which
electrons pass from lower to higher standard
redox potentials Electrons are carried from complex
I and II to complex III by the membrane (lipid)
soluble coenzyme Q (CoQ or ubiquinone) and between
22
complexes III and IV by the peripheral membrane
protein cytochrome c
In the following I give the overall
reactions catalysed by corresponding protein
complexes in the mitochondrial electron transport
chain
1 Complex I
Complex I is also called NADH-Coenzyme Q
reductase because this large protein complex
transfers 2 electrons from NADH to coenzyme Q
Complex I was formerly known as NADH dehydrogenase
Complex I is huge 850000 kD and is composed of more
than thirty subunits It contains a FMN prosthetic
group and seven or more Fe-S clusters This complex
has between 20-26 iron atoms boundThe prosthetic
group FMN is absolutely required for activity
Therefore this complex is a flavoprotein This
complex binds NADH transfers two electrons in the
form of a hydride to FMN to produce NAD+ and FMNH2
23
The subsequent steps involve the transfer of
electrons one at a time to a series of iron-sulfer
complexes that includes both 2Fe-2S and 4Fe-4S
clusters Note the importance of FMN First it
functions as a 2 electron acceptor in the hydride
transfer from NADH Second it functions as a 1
electron donor to the series of iron sulfur clusters
FMN and FAD often play crucial links between 2
electron transfer agents and 1 electron transfer
agents The final step of this complex is the
transfer of 2 electrons one at a time to coenzyme Q
CoQ like FMN and FAD can function as a 2 electron
donoracceptor and as a 1 electron donoracceptor
CoQ is a mobile electron carrier because its
isoprenoid tail makes it highly hydrophobic and
lipophillic It diffuses freely in the bilipid layer
of the inner mitochondrial membrane
The process of transferring electrons from NADH
to CoQ by complex I results in the net transport of
protons from the matrix side of the inner
mitochondrial membrane to the inter membrane space
where the H+ ions accumulate generating a proton
motive force The intermembrane space side of the
inner membrane is referred to as the P face (P
standing for positive) The matrix side of the inner
membrane is referred to as the the N face The
24
transport of electrons from NADH to CoQ is coupled to
the transport of protons across the membrane This is
an example of active transport The stiochiometry is
4 H+ transported per 2 electrons
2 Complex II
Believe it or not you are already familiar
with Complex II It is none other than succinate
dehydrogenase The only enzyme of the citric acid
cycle that is an integral membrane protein This
complex is composed of four subunits 2 of which are
iron-sulfur proteins and the other two subunits
together bind FAD through a covalent link to a
histidine residue These two subunits are called
flavoprotein 2 or FP2 Complex II contains 3 Fe-S
centers 1 4Fe-4S cluster 1 3Fe-4S cluster and 1
2Fe- 2S cluster In the first step of this complex
succinate is bound and a hydride is transferred to
FAD to generate FADH2 and fumarate FADH2 then
transfers its electrons one at a time to the Fe-S
centers Thus once again FAD functions as 2 electron
acceptor and a 1 electron donor The final step of
this complex is the transfer of 2 electrons one at a
time to coenzyme Q to produce CoQH2 For complex II
the standard free energy change of the overall
reaction is too small to drive the transport of
protons across the inner mitochondrial membrane This
25
accounts for the 15 ATPrsquos generated per FADH2
compared with the 25 ATPrsquos generated per NADH
3 Complex III
This complex is also known as coenzyme Q-
cytochrome c reductase because it passes the
electrons form CoQH2 to cyt c through a very unique
electron transport pathway called the Q-cycle Shown
to the left are the porphyrins found in cytochromes
Cytochrome b contains the same iron protoporphyrin as
hemoglobin and myoglobin The c cytochromes contain
heme c through covalent attachment by cysteine
residues Cytochrome a is found in two forms in
complex IV In complex III we find two b-type
cytochromes and one c-type cytochrome Complex III is
complex and we have a crystal structure
4 Complex IV
Complex IV is also known as cytochrome c oxidase
because it accepts the electrons from cytochrome c
and directs them towards the four electron reduction
of O2 to form 2 molecules of H2O
4 cyt c (Fe2+) + 4 H+ + O2 4 cyt c (Fe3+) +
2H2O
26
Cytochrome c oxidase contains 2 heme
centerscytochrome a and cytochrome a3 and two copper
proteins Each of the protein bound coppers are
associated with one of the cytochromes The copper
sites are called CuA and CuB CuA is associated with
cytochrome a and is shown to the left CuB is
associated with cytochrome a3 The copper sites
function as 1 electron carriers cycling between the
cuprous state Cu+ and the cupric state Cu2+ Just
like iron containing proteins they transfer
electrons one at a time Cytochrome c is bound on the
P-face of the membrane and transfers its electron to
CuA The oxidized cytochrome c dissociates CuA then
transfers the electron to cytochrome a The protein
bound CuA and the iron bound in cytochrome a are 15 Aring
apart In contrast the CuB and the iron bound in
cytochrome a3 are very close to each other forming a
binuclear metal center shown below
27
Electron transport chains are associated with
membranes such as the mitochondrial membrane in
eukaryotic organisms the thylakoid membrane of
photosynthetic organisms and the plasma membrane of
prokaryotic organisms While some of the details
differ electron transport chains in all organisms
operate on the same basic principles
Excited electrons are brought to the electron
transport chain by electron carriers such as NADH
and FADH2 In photosynthetic organisms the electrons
are donated by water and excited by light energy
The electron donor is oxidized as the electrons are
passed to the first protein complex in the chain As
it accepts the electrons the protein complex is
reduced Next the electrons are passed from the
28
first protein complex which becomes oxidized to
the second protein complex which becomes reduced
Each of these redox reactions is slightly exergonic
or energy releasing Some of this energy is used to
move protons across the membrane against their
concentration gradient With each successive
transfer of electrons the original excited
electrons lose some of their energy After several
successive redox reactions the electrons are
donated to a terminal electron acceptor In aerobic
respiration the terminal electron acceptor is
oxygen In photosynthesis the terminal electron
acceptor is a compound called NADP
The energy in the excited electrons has been
transformed into an electrochemical gradient
represented by the high concentration of protons on
one side of the membrane These protons can diffuse
back across the membrane by passing through a
protein complex called ATP synthase As these
protons pass through ATP synthase down their
concentration gradient they provide the energy
needed to make ATP from ADP and inorganic phosphate
within the cell
29
CHAPTER III
CLOSING
A Conclution
When a hexose is completely oxidized to CO2
and H2O using these three processes Glycolysis
yields two ATP and two NADH per hexose used Each
such NADH oxidized by the electron transport system
30
yields two ATP as dscribed above so glycolysis
contributes a total of six ATP per hexose The Krebs
cycle contributes two ATP per hexose or per two
pyruvates when succinyl CoA is cleaved to succinate
and CoASH This cycle also produces eight NADH per
hexose within the mitochondrial matrix by oxidative
phosporylation each of these NADH yields three ATP
or 24 per hexose Each FADH2 from the Krebs cycle
yields two ATP by oxydative phosporylation or four
per hexose (two pyruvates) The total contribution of
the Krebs cycle is then 30 Adding these 30 to the 6
from glycolysis leads to a total of 36 per hexose
completely respired by these process
B Suggestion
My greatest wish is that the readers find
this manuscript helpful in gaining the knowledge they
need for effective learning and practice and in
preparing for examination designed to test that
knowledge Reader comments and suggestions are
welcome and would be very helpful in improving our
manuscript Your contribution may assist many other
students in the coming years
31
REFERENCE
Anonim 2013 Anaerobic Respiration
httpwwwscoolcoukgcsebiologyrespiration
revise-itanaerobic-respiration Accessed
September 10th 2013
Anonim 2013 What is the difference between aerobic
and anaerobic respiration in plants
httpwwwbiologylifeeasyorg4010difference
-between-aerobic-anaerobic-respiration-plants
Accessed September 10th 2013
Campbell Mary K 2006 Biochemistry New York Thomson
Brooks Cole Inc
Dat J Capelli N Folzer H Bourgeade P Badot P-M
(2004) Sensing and signaling during
plantflooding Plant Physiology and
Biochemistry 42 273-282
Dat J Folzer H Parent C Badot P-M Capelli N
(2006) HypoxiastressCurrent Understanding and
Perspectives In Teixeira da Silva JA (Ed)
Floriculture Ornamental and Plant
Biotechnology Advances and Topical Issues (Vol
3) Global Science Books Isleworth United
Kingdompp 664-674
Hatta Muhammad 2011 Dunia Pertanian
httpemhattawordpresscomcategorystress
Accessed September 10th 2013
Hopkins William G 2009 Introduction to Plant Physiology
New York John Wiley amp Sons Inc
Lakitan Benyamin 2011 Dasar-Dasar Fisiologi Tumbuhan
Jakarta PT Raja Grafindo Persada
Larcher W 1980 Physiological Plant Ecology New York
Springer-Verlag
Raven 2013 Biology Fifth Edition
httphigheredmcgraw-hillcomsites983409233
9student_view0chapter39
photosynthetic_electron_transport_and_atp_synth
esishtml Accessed September 10th 2013
Salisbury Frank B 1969 Plant Physiology Third Edition
California Wadsworth Publishing Company
Subbaiah C Sachs M (2003) Molecular and cellular
adaptations of maize to flooding stress Annals
of Botany 91 119-127
Wilson CarlL1962 Botany Third EditionUnited State of
America Holt Rinch Inc

3) Coenzyme Q (CoQ)
aka ubiquinone (UQ) Shown below CoQ has ten
repeating isoprene units which make it
insoluble in water but soluble in the
hydrophobic lipid bylayer Coenzyme Q is a
versatile cofactor because it is a soluble
electron carrier in the hydrophobic bilipid layer
of the
inner mitochondrial membrane Like flavoproteins
CoQ can acceptdonate electrons one at a time or
two at a time
Q + 2e- + 2H+ 1048581 QH2 Eorsquo =0060 V
Q + e- + H+ 1048581 1048581QH Eorsquo =0030 V
1048581QH + e- + H+ 1048581 QH2 Eorsquo =0190 V
4) Cytochromes
Cytochromes are proteins that contain heme
prosthetic groups which function as
one electron carriers The heme iron is involved
in one electron transfers involving the Fe2+ and
Fe3+ oxidation states Cytochromes are named by
their absorption spectra which depends
on the porphyrin structure and environment The
example shown is the heme prosthetic group of
cytochrome b Cytochrome b contains the same iron
porphyrin found in hemoglobin and myoglobin
20
Other cytochromes we will encounter in the
electron transport complexes are cytochromes b
c c1 a and a3
5) Iron-Sulfur Proteins
In the electron transport chain we will encounter
many iron-sulfer proteins which participate in
one electron transfers involving the the Fe2+ and
Fe3+ oxidation states These are non-heme iron-
sulfur proteinsThe simplest iron-sulfer protein
is FeS in which iron is tetrahedrally coordinated
by four cysteines The second form is Fe2S2 which
contains two irons complexed to 2 cysteine
residues and two inorganic sulfides The third
form is Fe3S4 which contains 3 iron atoms
coordinated to three cysteine residues and 4
inorganic sulfides The last form is the most
complicated Fe4S4 which contains 4 iron atoms
coordinated to 4 cysteine residues and 4
inorganic sulfides
6) Copper Proteins
Copper bound proteins participate in one electron
transfers involving the Cu+ and Cu2+ oxidation
states
21
Overview of the Electron Transport Chain
Electrons move along the electron transport
chain going from donor to acceptor until they reach
oxygen the ultimate electron acceptor The standard
reduction potentials of the electron carriers are
between the NADHNAD+ couple (-0315 V) and the
oxygenH2O couple (0816 V) as on the next page The
components of the electron transport chain are
organized into 4 complexes Each complex contains
several different electron carriers
1 Complex I also known as the NADH-coenzyme Q
reductase or NADH dehydrogenase
2 Complex II also known as succinate-coenzyme Q
reductase or succinate dehydrogenase
3 Complex III also known as coenzyme Q reductase
4 Complex IV also known as cytochrome c reductase
The free energy necessary to generate ATP
is extracted from the oxidation of NADH and
FADH2 by the electron transport chain It
consists of four protein complexes and single
protein chain of cytochrome c through which
electrons pass from lower to higher standard
redox potentials Electrons are carried from complex
I and II to complex III by the membrane (lipid)
soluble coenzyme Q (CoQ or ubiquinone) and between
22
complexes III and IV by the peripheral membrane
protein cytochrome c
In the following I give the overall
reactions catalysed by corresponding protein
complexes in the mitochondrial electron transport
chain
1 Complex I
Complex I is also called NADH-Coenzyme Q
reductase because this large protein complex
transfers 2 electrons from NADH to coenzyme Q
Complex I was formerly known as NADH dehydrogenase
Complex I is huge 850000 kD and is composed of more
than thirty subunits It contains a FMN prosthetic
group and seven or more Fe-S clusters This complex
has between 20-26 iron atoms boundThe prosthetic
group FMN is absolutely required for activity
Therefore this complex is a flavoprotein This
complex binds NADH transfers two electrons in the
form of a hydride to FMN to produce NAD+ and FMNH2
23
The subsequent steps involve the transfer of
electrons one at a time to a series of iron-sulfer
complexes that includes both 2Fe-2S and 4Fe-4S
clusters Note the importance of FMN First it
functions as a 2 electron acceptor in the hydride
transfer from NADH Second it functions as a 1
electron donor to the series of iron sulfur clusters
FMN and FAD often play crucial links between 2
electron transfer agents and 1 electron transfer
agents The final step of this complex is the
transfer of 2 electrons one at a time to coenzyme Q
CoQ like FMN and FAD can function as a 2 electron
donoracceptor and as a 1 electron donoracceptor
CoQ is a mobile electron carrier because its
isoprenoid tail makes it highly hydrophobic and
lipophillic It diffuses freely in the bilipid layer
of the inner mitochondrial membrane
The process of transferring electrons from NADH
to CoQ by complex I results in the net transport of
protons from the matrix side of the inner
mitochondrial membrane to the inter membrane space
where the H+ ions accumulate generating a proton
motive force The intermembrane space side of the
inner membrane is referred to as the P face (P
standing for positive) The matrix side of the inner
membrane is referred to as the the N face The
24
transport of electrons from NADH to CoQ is coupled to
the transport of protons across the membrane This is
an example of active transport The stiochiometry is
4 H+ transported per 2 electrons
2 Complex II
Believe it or not you are already familiar
with Complex II It is none other than succinate
dehydrogenase The only enzyme of the citric acid
cycle that is an integral membrane protein This
complex is composed of four subunits 2 of which are
iron-sulfur proteins and the other two subunits
together bind FAD through a covalent link to a
histidine residue These two subunits are called
flavoprotein 2 or FP2 Complex II contains 3 Fe-S
centers 1 4Fe-4S cluster 1 3Fe-4S cluster and 1
2Fe- 2S cluster In the first step of this complex
succinate is bound and a hydride is transferred to
FAD to generate FADH2 and fumarate FADH2 then
transfers its electrons one at a time to the Fe-S
centers Thus once again FAD functions as 2 electron
acceptor and a 1 electron donor The final step of
this complex is the transfer of 2 electrons one at a
time to coenzyme Q to produce CoQH2 For complex II
the standard free energy change of the overall
reaction is too small to drive the transport of
protons across the inner mitochondrial membrane This
25
accounts for the 15 ATPrsquos generated per FADH2
compared with the 25 ATPrsquos generated per NADH
3 Complex III
This complex is also known as coenzyme Q-
cytochrome c reductase because it passes the
electrons form CoQH2 to cyt c through a very unique
electron transport pathway called the Q-cycle Shown
to the left are the porphyrins found in cytochromes
Cytochrome b contains the same iron protoporphyrin as
hemoglobin and myoglobin The c cytochromes contain
heme c through covalent attachment by cysteine
residues Cytochrome a is found in two forms in
complex IV In complex III we find two b-type
cytochromes and one c-type cytochrome Complex III is
complex and we have a crystal structure
4 Complex IV
Complex IV is also known as cytochrome c oxidase
because it accepts the electrons from cytochrome c
and directs them towards the four electron reduction
of O2 to form 2 molecules of H2O
4 cyt c (Fe2+) + 4 H+ + O2 4 cyt c (Fe3+) +
2H2O
26
Cytochrome c oxidase contains 2 heme
centerscytochrome a and cytochrome a3 and two copper
proteins Each of the protein bound coppers are
associated with one of the cytochromes The copper
sites are called CuA and CuB CuA is associated with
cytochrome a and is shown to the left CuB is
associated with cytochrome a3 The copper sites
function as 1 electron carriers cycling between the
cuprous state Cu+ and the cupric state Cu2+ Just
like iron containing proteins they transfer
electrons one at a time Cytochrome c is bound on the
P-face of the membrane and transfers its electron to
CuA The oxidized cytochrome c dissociates CuA then
transfers the electron to cytochrome a The protein
bound CuA and the iron bound in cytochrome a are 15 Aring
apart In contrast the CuB and the iron bound in
cytochrome a3 are very close to each other forming a
binuclear metal center shown below
27
Electron transport chains are associated with
membranes such as the mitochondrial membrane in
eukaryotic organisms the thylakoid membrane of
photosynthetic organisms and the plasma membrane of
prokaryotic organisms While some of the details
differ electron transport chains in all organisms
operate on the same basic principles
Excited electrons are brought to the electron
transport chain by electron carriers such as NADH
and FADH2 In photosynthetic organisms the electrons
are donated by water and excited by light energy
The electron donor is oxidized as the electrons are
passed to the first protein complex in the chain As
it accepts the electrons the protein complex is
reduced Next the electrons are passed from the
28
first protein complex which becomes oxidized to
the second protein complex which becomes reduced
Each of these redox reactions is slightly exergonic
or energy releasing Some of this energy is used to
move protons across the membrane against their
concentration gradient With each successive
transfer of electrons the original excited
electrons lose some of their energy After several
successive redox reactions the electrons are
donated to a terminal electron acceptor In aerobic
respiration the terminal electron acceptor is
oxygen In photosynthesis the terminal electron
acceptor is a compound called NADP
The energy in the excited electrons has been
transformed into an electrochemical gradient
represented by the high concentration of protons on
one side of the membrane These protons can diffuse
back across the membrane by passing through a
protein complex called ATP synthase As these
protons pass through ATP synthase down their
concentration gradient they provide the energy
needed to make ATP from ADP and inorganic phosphate
within the cell
29
CHAPTER III
CLOSING
A Conclution
When a hexose is completely oxidized to CO2
and H2O using these three processes Glycolysis
yields two ATP and two NADH per hexose used Each
such NADH oxidized by the electron transport system
30
yields two ATP as dscribed above so glycolysis
contributes a total of six ATP per hexose The Krebs
cycle contributes two ATP per hexose or per two
pyruvates when succinyl CoA is cleaved to succinate
and CoASH This cycle also produces eight NADH per
hexose within the mitochondrial matrix by oxidative
phosporylation each of these NADH yields three ATP
or 24 per hexose Each FADH2 from the Krebs cycle
yields two ATP by oxydative phosporylation or four
per hexose (two pyruvates) The total contribution of
the Krebs cycle is then 30 Adding these 30 to the 6
from glycolysis leads to a total of 36 per hexose
completely respired by these process
B Suggestion
My greatest wish is that the readers find
this manuscript helpful in gaining the knowledge they
need for effective learning and practice and in
preparing for examination designed to test that
knowledge Reader comments and suggestions are
welcome and would be very helpful in improving our
manuscript Your contribution may assist many other
students in the coming years
31
REFERENCE
Anonim 2013 Anaerobic Respiration
httpwwwscoolcoukgcsebiologyrespiration
revise-itanaerobic-respiration Accessed
September 10th 2013
Anonim 2013 What is the difference between aerobic
and anaerobic respiration in plants
httpwwwbiologylifeeasyorg4010difference
-between-aerobic-anaerobic-respiration-plants
Accessed September 10th 2013
Campbell Mary K 2006 Biochemistry New York Thomson
Brooks Cole Inc
Dat J Capelli N Folzer H Bourgeade P Badot P-M
(2004) Sensing and signaling during
plantflooding Plant Physiology and
Biochemistry 42 273-282
Dat J Folzer H Parent C Badot P-M Capelli N
(2006) HypoxiastressCurrent Understanding and
Perspectives In Teixeira da Silva JA (Ed)
Floriculture Ornamental and Plant
Biotechnology Advances and Topical Issues (Vol
3) Global Science Books Isleworth United
Kingdompp 664-674
Hatta Muhammad 2011 Dunia Pertanian
httpemhattawordpresscomcategorystress
Accessed September 10th 2013
Hopkins William G 2009 Introduction to Plant Physiology
New York John Wiley amp Sons Inc
Lakitan Benyamin 2011 Dasar-Dasar Fisiologi Tumbuhan
Jakarta PT Raja Grafindo Persada
Larcher W 1980 Physiological Plant Ecology New York
Springer-Verlag
Raven 2013 Biology Fifth Edition
httphigheredmcgraw-hillcomsites983409233
9student_view0chapter39
photosynthetic_electron_transport_and_atp_synth
esishtml Accessed September 10th 2013
Salisbury Frank B 1969 Plant Physiology Third Edition
California Wadsworth Publishing Company
Subbaiah C Sachs M (2003) Molecular and cellular
adaptations of maize to flooding stress Annals
of Botany 91 119-127
Wilson CarlL1962 Botany Third EditionUnited State of
America Holt Rinch Inc

Other cytochromes we will encounter in the
electron transport complexes are cytochromes b
c c1 a and a3
5) Iron-Sulfur Proteins
In the electron transport chain we will encounter
many iron-sulfer proteins which participate in
one electron transfers involving the the Fe2+ and
Fe3+ oxidation states These are non-heme iron-
sulfur proteinsThe simplest iron-sulfer protein
is FeS in which iron is tetrahedrally coordinated
by four cysteines The second form is Fe2S2 which
contains two irons complexed to 2 cysteine
residues and two inorganic sulfides The third
form is Fe3S4 which contains 3 iron atoms
coordinated to three cysteine residues and 4
inorganic sulfides The last form is the most
complicated Fe4S4 which contains 4 iron atoms
coordinated to 4 cysteine residues and 4
inorganic sulfides
6) Copper Proteins
Copper bound proteins participate in one electron
transfers involving the Cu+ and Cu2+ oxidation
states
21
Overview of the Electron Transport Chain
Electrons move along the electron transport
chain going from donor to acceptor until they reach
oxygen the ultimate electron acceptor The standard
reduction potentials of the electron carriers are
between the NADHNAD+ couple (-0315 V) and the
oxygenH2O couple (0816 V) as on the next page The
components of the electron transport chain are
organized into 4 complexes Each complex contains
several different electron carriers
1 Complex I also known as the NADH-coenzyme Q
reductase or NADH dehydrogenase
2 Complex II also known as succinate-coenzyme Q
reductase or succinate dehydrogenase
3 Complex III also known as coenzyme Q reductase
4 Complex IV also known as cytochrome c reductase
The free energy necessary to generate ATP
is extracted from the oxidation of NADH and
FADH2 by the electron transport chain It
consists of four protein complexes and single
protein chain of cytochrome c through which
electrons pass from lower to higher standard
redox potentials Electrons are carried from complex
I and II to complex III by the membrane (lipid)
soluble coenzyme Q (CoQ or ubiquinone) and between
22
complexes III and IV by the peripheral membrane
protein cytochrome c
In the following I give the overall
reactions catalysed by corresponding protein
complexes in the mitochondrial electron transport
chain
1 Complex I
Complex I is also called NADH-Coenzyme Q
reductase because this large protein complex
transfers 2 electrons from NADH to coenzyme Q
Complex I was formerly known as NADH dehydrogenase
Complex I is huge 850000 kD and is composed of more
than thirty subunits It contains a FMN prosthetic
group and seven or more Fe-S clusters This complex
has between 20-26 iron atoms boundThe prosthetic
group FMN is absolutely required for activity
Therefore this complex is a flavoprotein This
complex binds NADH transfers two electrons in the
form of a hydride to FMN to produce NAD+ and FMNH2
23
The subsequent steps involve the transfer of
electrons one at a time to a series of iron-sulfer
complexes that includes both 2Fe-2S and 4Fe-4S
clusters Note the importance of FMN First it
functions as a 2 electron acceptor in the hydride
transfer from NADH Second it functions as a 1
electron donor to the series of iron sulfur clusters
FMN and FAD often play crucial links between 2
electron transfer agents and 1 electron transfer
agents The final step of this complex is the
transfer of 2 electrons one at a time to coenzyme Q
CoQ like FMN and FAD can function as a 2 electron
donoracceptor and as a 1 electron donoracceptor
CoQ is a mobile electron carrier because its
isoprenoid tail makes it highly hydrophobic and
lipophillic It diffuses freely in the bilipid layer
of the inner mitochondrial membrane
The process of transferring electrons from NADH
to CoQ by complex I results in the net transport of
protons from the matrix side of the inner
mitochondrial membrane to the inter membrane space
where the H+ ions accumulate generating a proton
motive force The intermembrane space side of the
inner membrane is referred to as the P face (P
standing for positive) The matrix side of the inner
membrane is referred to as the the N face The
24
transport of electrons from NADH to CoQ is coupled to
the transport of protons across the membrane This is
an example of active transport The stiochiometry is
4 H+ transported per 2 electrons
2 Complex II
Believe it or not you are already familiar
with Complex II It is none other than succinate
dehydrogenase The only enzyme of the citric acid
cycle that is an integral membrane protein This
complex is composed of four subunits 2 of which are
iron-sulfur proteins and the other two subunits
together bind FAD through a covalent link to a
histidine residue These two subunits are called
flavoprotein 2 or FP2 Complex II contains 3 Fe-S
centers 1 4Fe-4S cluster 1 3Fe-4S cluster and 1
2Fe- 2S cluster In the first step of this complex
succinate is bound and a hydride is transferred to
FAD to generate FADH2 and fumarate FADH2 then
transfers its electrons one at a time to the Fe-S
centers Thus once again FAD functions as 2 electron
acceptor and a 1 electron donor The final step of
this complex is the transfer of 2 electrons one at a
time to coenzyme Q to produce CoQH2 For complex II
the standard free energy change of the overall
reaction is too small to drive the transport of
protons across the inner mitochondrial membrane This
25
accounts for the 15 ATPrsquos generated per FADH2
compared with the 25 ATPrsquos generated per NADH
3 Complex III
This complex is also known as coenzyme Q-
cytochrome c reductase because it passes the
electrons form CoQH2 to cyt c through a very unique
electron transport pathway called the Q-cycle Shown
to the left are the porphyrins found in cytochromes
Cytochrome b contains the same iron protoporphyrin as
hemoglobin and myoglobin The c cytochromes contain
heme c through covalent attachment by cysteine
residues Cytochrome a is found in two forms in
complex IV In complex III we find two b-type
cytochromes and one c-type cytochrome Complex III is
complex and we have a crystal structure
4 Complex IV
Complex IV is also known as cytochrome c oxidase
because it accepts the electrons from cytochrome c
and directs them towards the four electron reduction
of O2 to form 2 molecules of H2O
4 cyt c (Fe2+) + 4 H+ + O2 4 cyt c (Fe3+) +
2H2O
26
Cytochrome c oxidase contains 2 heme
centerscytochrome a and cytochrome a3 and two copper
proteins Each of the protein bound coppers are
associated with one of the cytochromes The copper
sites are called CuA and CuB CuA is associated with
cytochrome a and is shown to the left CuB is
associated with cytochrome a3 The copper sites
function as 1 electron carriers cycling between the
cuprous state Cu+ and the cupric state Cu2+ Just
like iron containing proteins they transfer
electrons one at a time Cytochrome c is bound on the
P-face of the membrane and transfers its electron to
CuA The oxidized cytochrome c dissociates CuA then
transfers the electron to cytochrome a The protein
bound CuA and the iron bound in cytochrome a are 15 Aring
apart In contrast the CuB and the iron bound in
cytochrome a3 are very close to each other forming a
binuclear metal center shown below
27
Electron transport chains are associated with
membranes such as the mitochondrial membrane in
eukaryotic organisms the thylakoid membrane of
photosynthetic organisms and the plasma membrane of
prokaryotic organisms While some of the details
differ electron transport chains in all organisms
operate on the same basic principles
Excited electrons are brought to the electron
transport chain by electron carriers such as NADH
and FADH2 In photosynthetic organisms the electrons
are donated by water and excited by light energy
The electron donor is oxidized as the electrons are
passed to the first protein complex in the chain As
it accepts the electrons the protein complex is
reduced Next the electrons are passed from the
28
first protein complex which becomes oxidized to
the second protein complex which becomes reduced
Each of these redox reactions is slightly exergonic
or energy releasing Some of this energy is used to
move protons across the membrane against their
concentration gradient With each successive
transfer of electrons the original excited
electrons lose some of their energy After several
successive redox reactions the electrons are
donated to a terminal electron acceptor In aerobic
respiration the terminal electron acceptor is
oxygen In photosynthesis the terminal electron
acceptor is a compound called NADP
The energy in the excited electrons has been
transformed into an electrochemical gradient
represented by the high concentration of protons on
one side of the membrane These protons can diffuse
back across the membrane by passing through a
protein complex called ATP synthase As these
protons pass through ATP synthase down their
concentration gradient they provide the energy
needed to make ATP from ADP and inorganic phosphate
within the cell
29
CHAPTER III
CLOSING
A Conclution
When a hexose is completely oxidized to CO2
and H2O using these three processes Glycolysis
yields two ATP and two NADH per hexose used Each
such NADH oxidized by the electron transport system
30
yields two ATP as dscribed above so glycolysis
contributes a total of six ATP per hexose The Krebs
cycle contributes two ATP per hexose or per two
pyruvates when succinyl CoA is cleaved to succinate
and CoASH This cycle also produces eight NADH per
hexose within the mitochondrial matrix by oxidative
phosporylation each of these NADH yields three ATP
or 24 per hexose Each FADH2 from the Krebs cycle
yields two ATP by oxydative phosporylation or four
per hexose (two pyruvates) The total contribution of
the Krebs cycle is then 30 Adding these 30 to the 6
from glycolysis leads to a total of 36 per hexose
completely respired by these process
B Suggestion
My greatest wish is that the readers find
this manuscript helpful in gaining the knowledge they
need for effective learning and practice and in
preparing for examination designed to test that
knowledge Reader comments and suggestions are
welcome and would be very helpful in improving our
manuscript Your contribution may assist many other
students in the coming years
31
REFERENCE
Anonim 2013 Anaerobic Respiration
httpwwwscoolcoukgcsebiologyrespiration
revise-itanaerobic-respiration Accessed
September 10th 2013
Anonim 2013 What is the difference between aerobic
and anaerobic respiration in plants
httpwwwbiologylifeeasyorg4010difference
-between-aerobic-anaerobic-respiration-plants
Accessed September 10th 2013
Campbell Mary K 2006 Biochemistry New York Thomson
Brooks Cole Inc
Dat J Capelli N Folzer H Bourgeade P Badot P-M
(2004) Sensing and signaling during
plantflooding Plant Physiology and
Biochemistry 42 273-282
Dat J Folzer H Parent C Badot P-M Capelli N
(2006) HypoxiastressCurrent Understanding and
Perspectives In Teixeira da Silva JA (Ed)
Floriculture Ornamental and Plant
Biotechnology Advances and Topical Issues (Vol
3) Global Science Books Isleworth United
Kingdompp 664-674
Hatta Muhammad 2011 Dunia Pertanian
httpemhattawordpresscomcategorystress
Accessed September 10th 2013
Hopkins William G 2009 Introduction to Plant Physiology
New York John Wiley amp Sons Inc
Lakitan Benyamin 2011 Dasar-Dasar Fisiologi Tumbuhan
Jakarta PT Raja Grafindo Persada
Larcher W 1980 Physiological Plant Ecology New York
Springer-Verlag
Raven 2013 Biology Fifth Edition
httphigheredmcgraw-hillcomsites983409233
9student_view0chapter39
photosynthetic_electron_transport_and_atp_synth
esishtml Accessed September 10th 2013
Salisbury Frank B 1969 Plant Physiology Third Edition
California Wadsworth Publishing Company
Subbaiah C Sachs M (2003) Molecular and cellular
adaptations of maize to flooding stress Annals
of Botany 91 119-127
Wilson CarlL1962 Botany Third EditionUnited State of
America Holt Rinch Inc

Overview of the Electron Transport Chain
Electrons move along the electron transport
chain going from donor to acceptor until they reach
oxygen the ultimate electron acceptor The standard
reduction potentials of the electron carriers are
between the NADHNAD+ couple (-0315 V) and the
oxygenH2O couple (0816 V) as on the next page The
components of the electron transport chain are
organized into 4 complexes Each complex contains
several different electron carriers
1 Complex I also known as the NADH-coenzyme Q
reductase or NADH dehydrogenase
2 Complex II also known as succinate-coenzyme Q
reductase or succinate dehydrogenase
3 Complex III also known as coenzyme Q reductase
4 Complex IV also known as cytochrome c reductase
The free energy necessary to generate ATP
is extracted from the oxidation of NADH and
FADH2 by the electron transport chain It
consists of four protein complexes and single
protein chain of cytochrome c through which
electrons pass from lower to higher standard
redox potentials Electrons are carried from complex
I and II to complex III by the membrane (lipid)
soluble coenzyme Q (CoQ or ubiquinone) and between
22
complexes III and IV by the peripheral membrane
protein cytochrome c
In the following I give the overall
reactions catalysed by corresponding protein
complexes in the mitochondrial electron transport
chain
1 Complex I
Complex I is also called NADH-Coenzyme Q
reductase because this large protein complex
transfers 2 electrons from NADH to coenzyme Q
Complex I was formerly known as NADH dehydrogenase
Complex I is huge 850000 kD and is composed of more
than thirty subunits It contains a FMN prosthetic
group and seven or more Fe-S clusters This complex
has between 20-26 iron atoms boundThe prosthetic
group FMN is absolutely required for activity
Therefore this complex is a flavoprotein This
complex binds NADH transfers two electrons in the
form of a hydride to FMN to produce NAD+ and FMNH2
23
The subsequent steps involve the transfer of
electrons one at a time to a series of iron-sulfer
complexes that includes both 2Fe-2S and 4Fe-4S
clusters Note the importance of FMN First it
functions as a 2 electron acceptor in the hydride
transfer from NADH Second it functions as a 1
electron donor to the series of iron sulfur clusters
FMN and FAD often play crucial links between 2
electron transfer agents and 1 electron transfer
agents The final step of this complex is the
transfer of 2 electrons one at a time to coenzyme Q
CoQ like FMN and FAD can function as a 2 electron
donoracceptor and as a 1 electron donoracceptor
CoQ is a mobile electron carrier because its
isoprenoid tail makes it highly hydrophobic and
lipophillic It diffuses freely in the bilipid layer
of the inner mitochondrial membrane
The process of transferring electrons from NADH
to CoQ by complex I results in the net transport of
protons from the matrix side of the inner
mitochondrial membrane to the inter membrane space
where the H+ ions accumulate generating a proton
motive force The intermembrane space side of the
inner membrane is referred to as the P face (P
standing for positive) The matrix side of the inner
membrane is referred to as the the N face The
24
transport of electrons from NADH to CoQ is coupled to
the transport of protons across the membrane This is
an example of active transport The stiochiometry is
4 H+ transported per 2 electrons
2 Complex II
Believe it or not you are already familiar
with Complex II It is none other than succinate
dehydrogenase The only enzyme of the citric acid
cycle that is an integral membrane protein This
complex is composed of four subunits 2 of which are
iron-sulfur proteins and the other two subunits
together bind FAD through a covalent link to a
histidine residue These two subunits are called
flavoprotein 2 or FP2 Complex II contains 3 Fe-S
centers 1 4Fe-4S cluster 1 3Fe-4S cluster and 1
2Fe- 2S cluster In the first step of this complex
succinate is bound and a hydride is transferred to
FAD to generate FADH2 and fumarate FADH2 then
transfers its electrons one at a time to the Fe-S
centers Thus once again FAD functions as 2 electron
acceptor and a 1 electron donor The final step of
this complex is the transfer of 2 electrons one at a
time to coenzyme Q to produce CoQH2 For complex II
the standard free energy change of the overall
reaction is too small to drive the transport of
protons across the inner mitochondrial membrane This
25
accounts for the 15 ATPrsquos generated per FADH2
compared with the 25 ATPrsquos generated per NADH
3 Complex III
This complex is also known as coenzyme Q-
cytochrome c reductase because it passes the
electrons form CoQH2 to cyt c through a very unique
electron transport pathway called the Q-cycle Shown
to the left are the porphyrins found in cytochromes
Cytochrome b contains the same iron protoporphyrin as
hemoglobin and myoglobin The c cytochromes contain
heme c through covalent attachment by cysteine
residues Cytochrome a is found in two forms in
complex IV In complex III we find two b-type
cytochromes and one c-type cytochrome Complex III is
complex and we have a crystal structure
4 Complex IV
Complex IV is also known as cytochrome c oxidase
because it accepts the electrons from cytochrome c
and directs them towards the four electron reduction
of O2 to form 2 molecules of H2O
4 cyt c (Fe2+) + 4 H+ + O2 4 cyt c (Fe3+) +
2H2O
26
Cytochrome c oxidase contains 2 heme
centerscytochrome a and cytochrome a3 and two copper
proteins Each of the protein bound coppers are
associated with one of the cytochromes The copper
sites are called CuA and CuB CuA is associated with
cytochrome a and is shown to the left CuB is
associated with cytochrome a3 The copper sites
function as 1 electron carriers cycling between the
cuprous state Cu+ and the cupric state Cu2+ Just
like iron containing proteins they transfer
electrons one at a time Cytochrome c is bound on the
P-face of the membrane and transfers its electron to
CuA The oxidized cytochrome c dissociates CuA then
transfers the electron to cytochrome a The protein
bound CuA and the iron bound in cytochrome a are 15 Aring
apart In contrast the CuB and the iron bound in
cytochrome a3 are very close to each other forming a
binuclear metal center shown below
27
Electron transport chains are associated with
membranes such as the mitochondrial membrane in
eukaryotic organisms the thylakoid membrane of
photosynthetic organisms and the plasma membrane of
prokaryotic organisms While some of the details
differ electron transport chains in all organisms
operate on the same basic principles
Excited electrons are brought to the electron
transport chain by electron carriers such as NADH
and FADH2 In photosynthetic organisms the electrons
are donated by water and excited by light energy
The electron donor is oxidized as the electrons are
passed to the first protein complex in the chain As
it accepts the electrons the protein complex is
reduced Next the electrons are passed from the
28
first protein complex which becomes oxidized to
the second protein complex which becomes reduced
Each of these redox reactions is slightly exergonic
or energy releasing Some of this energy is used to
move protons across the membrane against their
concentration gradient With each successive
transfer of electrons the original excited
electrons lose some of their energy After several
successive redox reactions the electrons are
donated to a terminal electron acceptor In aerobic
respiration the terminal electron acceptor is
oxygen In photosynthesis the terminal electron
acceptor is a compound called NADP
The energy in the excited electrons has been
transformed into an electrochemical gradient
represented by the high concentration of protons on
one side of the membrane These protons can diffuse
back across the membrane by passing through a
protein complex called ATP synthase As these
protons pass through ATP synthase down their
concentration gradient they provide the energy
needed to make ATP from ADP and inorganic phosphate
within the cell
29
CHAPTER III
CLOSING
A Conclution
When a hexose is completely oxidized to CO2
and H2O using these three processes Glycolysis
yields two ATP and two NADH per hexose used Each
such NADH oxidized by the electron transport system
30
yields two ATP as dscribed above so glycolysis
contributes a total of six ATP per hexose The Krebs
cycle contributes two ATP per hexose or per two
pyruvates when succinyl CoA is cleaved to succinate
and CoASH This cycle also produces eight NADH per
hexose within the mitochondrial matrix by oxidative
phosporylation each of these NADH yields three ATP
or 24 per hexose Each FADH2 from the Krebs cycle
yields two ATP by oxydative phosporylation or four
per hexose (two pyruvates) The total contribution of
the Krebs cycle is then 30 Adding these 30 to the 6
from glycolysis leads to a total of 36 per hexose
completely respired by these process
B Suggestion
My greatest wish is that the readers find
this manuscript helpful in gaining the knowledge they
need for effective learning and practice and in
preparing for examination designed to test that
knowledge Reader comments and suggestions are
welcome and would be very helpful in improving our
manuscript Your contribution may assist many other
students in the coming years
31
REFERENCE
Anonim 2013 Anaerobic Respiration
httpwwwscoolcoukgcsebiologyrespiration
revise-itanaerobic-respiration Accessed
September 10th 2013
Anonim 2013 What is the difference between aerobic
and anaerobic respiration in plants
httpwwwbiologylifeeasyorg4010difference
-between-aerobic-anaerobic-respiration-plants
Accessed September 10th 2013
Campbell Mary K 2006 Biochemistry New York Thomson
Brooks Cole Inc
Dat J Capelli N Folzer H Bourgeade P Badot P-M
(2004) Sensing and signaling during
plantflooding Plant Physiology and
Biochemistry 42 273-282
Dat J Folzer H Parent C Badot P-M Capelli N
(2006) HypoxiastressCurrent Understanding and
Perspectives In Teixeira da Silva JA (Ed)
Floriculture Ornamental and Plant
Biotechnology Advances and Topical Issues (Vol
3) Global Science Books Isleworth United
Kingdompp 664-674
Hatta Muhammad 2011 Dunia Pertanian
httpemhattawordpresscomcategorystress
Accessed September 10th 2013
Hopkins William G 2009 Introduction to Plant Physiology
New York John Wiley amp Sons Inc
Lakitan Benyamin 2011 Dasar-Dasar Fisiologi Tumbuhan
Jakarta PT Raja Grafindo Persada
Larcher W 1980 Physiological Plant Ecology New York
Springer-Verlag
Raven 2013 Biology Fifth Edition
httphigheredmcgraw-hillcomsites983409233
9student_view0chapter39
photosynthetic_electron_transport_and_atp_synth
esishtml Accessed September 10th 2013
Salisbury Frank B 1969 Plant Physiology Third Edition
California Wadsworth Publishing Company
Subbaiah C Sachs M (2003) Molecular and cellular
adaptations of maize to flooding stress Annals
of Botany 91 119-127
Wilson CarlL1962 Botany Third EditionUnited State of
America Holt Rinch Inc

complexes III and IV by the peripheral membrane
protein cytochrome c
In the following I give the overall
reactions catalysed by corresponding protein
complexes in the mitochondrial electron transport
chain
1 Complex I
Complex I is also called NADH-Coenzyme Q
reductase because this large protein complex
transfers 2 electrons from NADH to coenzyme Q
Complex I was formerly known as NADH dehydrogenase
Complex I is huge 850000 kD and is composed of more
than thirty subunits It contains a FMN prosthetic
group and seven or more Fe-S clusters This complex
has between 20-26 iron atoms boundThe prosthetic
group FMN is absolutely required for activity
Therefore this complex is a flavoprotein This
complex binds NADH transfers two electrons in the
form of a hydride to FMN to produce NAD+ and FMNH2
23
The subsequent steps involve the transfer of
electrons one at a time to a series of iron-sulfer
complexes that includes both 2Fe-2S and 4Fe-4S
clusters Note the importance of FMN First it
functions as a 2 electron acceptor in the hydride
transfer from NADH Second it functions as a 1
electron donor to the series of iron sulfur clusters
FMN and FAD often play crucial links between 2
electron transfer agents and 1 electron transfer
agents The final step of this complex is the
transfer of 2 electrons one at a time to coenzyme Q
CoQ like FMN and FAD can function as a 2 electron
donoracceptor and as a 1 electron donoracceptor
CoQ is a mobile electron carrier because its
isoprenoid tail makes it highly hydrophobic and
lipophillic It diffuses freely in the bilipid layer
of the inner mitochondrial membrane
The process of transferring electrons from NADH
to CoQ by complex I results in the net transport of
protons from the matrix side of the inner
mitochondrial membrane to the inter membrane space
where the H+ ions accumulate generating a proton
motive force The intermembrane space side of the
inner membrane is referred to as the P face (P
standing for positive) The matrix side of the inner
membrane is referred to as the the N face The
24
transport of electrons from NADH to CoQ is coupled to
the transport of protons across the membrane This is
an example of active transport The stiochiometry is
4 H+ transported per 2 electrons
2 Complex II
Believe it or not you are already familiar
with Complex II It is none other than succinate
dehydrogenase The only enzyme of the citric acid
cycle that is an integral membrane protein This
complex is composed of four subunits 2 of which are
iron-sulfur proteins and the other two subunits
together bind FAD through a covalent link to a
histidine residue These two subunits are called
flavoprotein 2 or FP2 Complex II contains 3 Fe-S
centers 1 4Fe-4S cluster 1 3Fe-4S cluster and 1
2Fe- 2S cluster In the first step of this complex
succinate is bound and a hydride is transferred to
FAD to generate FADH2 and fumarate FADH2 then
transfers its electrons one at a time to the Fe-S
centers Thus once again FAD functions as 2 electron
acceptor and a 1 electron donor The final step of
this complex is the transfer of 2 electrons one at a
time to coenzyme Q to produce CoQH2 For complex II
the standard free energy change of the overall
reaction is too small to drive the transport of
protons across the inner mitochondrial membrane This
25
accounts for the 15 ATPrsquos generated per FADH2
compared with the 25 ATPrsquos generated per NADH
3 Complex III
This complex is also known as coenzyme Q-
cytochrome c reductase because it passes the
electrons form CoQH2 to cyt c through a very unique
electron transport pathway called the Q-cycle Shown
to the left are the porphyrins found in cytochromes
Cytochrome b contains the same iron protoporphyrin as
hemoglobin and myoglobin The c cytochromes contain
heme c through covalent attachment by cysteine
residues Cytochrome a is found in two forms in
complex IV In complex III we find two b-type
cytochromes and one c-type cytochrome Complex III is
complex and we have a crystal structure
4 Complex IV
Complex IV is also known as cytochrome c oxidase
because it accepts the electrons from cytochrome c
and directs them towards the four electron reduction
of O2 to form 2 molecules of H2O
4 cyt c (Fe2+) + 4 H+ + O2 4 cyt c (Fe3+) +
2H2O
26
Cytochrome c oxidase contains 2 heme
centerscytochrome a and cytochrome a3 and two copper
proteins Each of the protein bound coppers are
associated with one of the cytochromes The copper
sites are called CuA and CuB CuA is associated with
cytochrome a and is shown to the left CuB is
associated with cytochrome a3 The copper sites
function as 1 electron carriers cycling between the
cuprous state Cu+ and the cupric state Cu2+ Just
like iron containing proteins they transfer
electrons one at a time Cytochrome c is bound on the
P-face of the membrane and transfers its electron to
CuA The oxidized cytochrome c dissociates CuA then
transfers the electron to cytochrome a The protein
bound CuA and the iron bound in cytochrome a are 15 Aring
apart In contrast the CuB and the iron bound in
cytochrome a3 are very close to each other forming a
binuclear metal center shown below
27
Electron transport chains are associated with
membranes such as the mitochondrial membrane in
eukaryotic organisms the thylakoid membrane of
photosynthetic organisms and the plasma membrane of
prokaryotic organisms While some of the details
differ electron transport chains in all organisms
operate on the same basic principles
Excited electrons are brought to the electron
transport chain by electron carriers such as NADH
and FADH2 In photosynthetic organisms the electrons
are donated by water and excited by light energy
The electron donor is oxidized as the electrons are
passed to the first protein complex in the chain As
it accepts the electrons the protein complex is
reduced Next the electrons are passed from the
28
first protein complex which becomes oxidized to
the second protein complex which becomes reduced
Each of these redox reactions is slightly exergonic
or energy releasing Some of this energy is used to
move protons across the membrane against their
concentration gradient With each successive
transfer of electrons the original excited
electrons lose some of their energy After several
successive redox reactions the electrons are
donated to a terminal electron acceptor In aerobic
respiration the terminal electron acceptor is
oxygen In photosynthesis the terminal electron
acceptor is a compound called NADP
The energy in the excited electrons has been
transformed into an electrochemical gradient
represented by the high concentration of protons on
one side of the membrane These protons can diffuse
back across the membrane by passing through a
protein complex called ATP synthase As these
protons pass through ATP synthase down their
concentration gradient they provide the energy
needed to make ATP from ADP and inorganic phosphate
within the cell
29
CHAPTER III
CLOSING
A Conclution
When a hexose is completely oxidized to CO2
and H2O using these three processes Glycolysis
yields two ATP and two NADH per hexose used Each
such NADH oxidized by the electron transport system
30
yields two ATP as dscribed above so glycolysis
contributes a total of six ATP per hexose The Krebs
cycle contributes two ATP per hexose or per two
pyruvates when succinyl CoA is cleaved to succinate
and CoASH This cycle also produces eight NADH per
hexose within the mitochondrial matrix by oxidative
phosporylation each of these NADH yields three ATP
or 24 per hexose Each FADH2 from the Krebs cycle
yields two ATP by oxydative phosporylation or four
per hexose (two pyruvates) The total contribution of
the Krebs cycle is then 30 Adding these 30 to the 6
from glycolysis leads to a total of 36 per hexose
completely respired by these process
B Suggestion
My greatest wish is that the readers find
this manuscript helpful in gaining the knowledge they
need for effective learning and practice and in
preparing for examination designed to test that
knowledge Reader comments and suggestions are
welcome and would be very helpful in improving our
manuscript Your contribution may assist many other
students in the coming years
31
REFERENCE
Anonim 2013 Anaerobic Respiration
httpwwwscoolcoukgcsebiologyrespiration
revise-itanaerobic-respiration Accessed
September 10th 2013
Anonim 2013 What is the difference between aerobic
and anaerobic respiration in plants
httpwwwbiologylifeeasyorg4010difference
-between-aerobic-anaerobic-respiration-plants
Accessed September 10th 2013
Campbell Mary K 2006 Biochemistry New York Thomson
Brooks Cole Inc
Dat J Capelli N Folzer H Bourgeade P Badot P-M
(2004) Sensing and signaling during
plantflooding Plant Physiology and
Biochemistry 42 273-282
Dat J Folzer H Parent C Badot P-M Capelli N
(2006) HypoxiastressCurrent Understanding and
Perspectives In Teixeira da Silva JA (Ed)
Floriculture Ornamental and Plant
Biotechnology Advances and Topical Issues (Vol
3) Global Science Books Isleworth United
Kingdompp 664-674
Hatta Muhammad 2011 Dunia Pertanian
httpemhattawordpresscomcategorystress
Accessed September 10th 2013
Hopkins William G 2009 Introduction to Plant Physiology
New York John Wiley amp Sons Inc
Lakitan Benyamin 2011 Dasar-Dasar Fisiologi Tumbuhan
Jakarta PT Raja Grafindo Persada
Larcher W 1980 Physiological Plant Ecology New York
Springer-Verlag
Raven 2013 Biology Fifth Edition
httphigheredmcgraw-hillcomsites983409233
9student_view0chapter39
photosynthetic_electron_transport_and_atp_synth
esishtml Accessed September 10th 2013
Salisbury Frank B 1969 Plant Physiology Third Edition
California Wadsworth Publishing Company
Subbaiah C Sachs M (2003) Molecular and cellular
adaptations of maize to flooding stress Annals
of Botany 91 119-127
Wilson CarlL1962 Botany Third EditionUnited State of
America Holt Rinch Inc

The subsequent steps involve the transfer of
electrons one at a time to a series of iron-sulfer
complexes that includes both 2Fe-2S and 4Fe-4S
clusters Note the importance of FMN First it
functions as a 2 electron acceptor in the hydride
transfer from NADH Second it functions as a 1
electron donor to the series of iron sulfur clusters
FMN and FAD often play crucial links between 2
electron transfer agents and 1 electron transfer
agents The final step of this complex is the
transfer of 2 electrons one at a time to coenzyme Q
CoQ like FMN and FAD can function as a 2 electron
donoracceptor and as a 1 electron donoracceptor
CoQ is a mobile electron carrier because its
isoprenoid tail makes it highly hydrophobic and
lipophillic It diffuses freely in the bilipid layer
of the inner mitochondrial membrane
The process of transferring electrons from NADH
to CoQ by complex I results in the net transport of
protons from the matrix side of the inner
mitochondrial membrane to the inter membrane space
where the H+ ions accumulate generating a proton
motive force The intermembrane space side of the
inner membrane is referred to as the P face (P
standing for positive) The matrix side of the inner
membrane is referred to as the the N face The
24
transport of electrons from NADH to CoQ is coupled to
the transport of protons across the membrane This is
an example of active transport The stiochiometry is
4 H+ transported per 2 electrons
2 Complex II
Believe it or not you are already familiar
with Complex II It is none other than succinate
dehydrogenase The only enzyme of the citric acid
cycle that is an integral membrane protein This
complex is composed of four subunits 2 of which are
iron-sulfur proteins and the other two subunits
together bind FAD through a covalent link to a
histidine residue These two subunits are called
flavoprotein 2 or FP2 Complex II contains 3 Fe-S
centers 1 4Fe-4S cluster 1 3Fe-4S cluster and 1
2Fe- 2S cluster In the first step of this complex
succinate is bound and a hydride is transferred to
FAD to generate FADH2 and fumarate FADH2 then
transfers its electrons one at a time to the Fe-S
centers Thus once again FAD functions as 2 electron
acceptor and a 1 electron donor The final step of
this complex is the transfer of 2 electrons one at a
time to coenzyme Q to produce CoQH2 For complex II
the standard free energy change of the overall
reaction is too small to drive the transport of
protons across the inner mitochondrial membrane This
25
accounts for the 15 ATPrsquos generated per FADH2
compared with the 25 ATPrsquos generated per NADH
3 Complex III
This complex is also known as coenzyme Q-
cytochrome c reductase because it passes the
electrons form CoQH2 to cyt c through a very unique
electron transport pathway called the Q-cycle Shown
to the left are the porphyrins found in cytochromes
Cytochrome b contains the same iron protoporphyrin as
hemoglobin and myoglobin The c cytochromes contain
heme c through covalent attachment by cysteine
residues Cytochrome a is found in two forms in
complex IV In complex III we find two b-type
cytochromes and one c-type cytochrome Complex III is
complex and we have a crystal structure
4 Complex IV
Complex IV is also known as cytochrome c oxidase
because it accepts the electrons from cytochrome c
and directs them towards the four electron reduction
of O2 to form 2 molecules of H2O
4 cyt c (Fe2+) + 4 H+ + O2 4 cyt c (Fe3+) +
2H2O
26
Cytochrome c oxidase contains 2 heme
centerscytochrome a and cytochrome a3 and two copper
proteins Each of the protein bound coppers are
associated with one of the cytochromes The copper
sites are called CuA and CuB CuA is associated with
cytochrome a and is shown to the left CuB is
associated with cytochrome a3 The copper sites
function as 1 electron carriers cycling between the
cuprous state Cu+ and the cupric state Cu2+ Just
like iron containing proteins they transfer
electrons one at a time Cytochrome c is bound on the
P-face of the membrane and transfers its electron to
CuA The oxidized cytochrome c dissociates CuA then
transfers the electron to cytochrome a The protein
bound CuA and the iron bound in cytochrome a are 15 Aring
apart In contrast the CuB and the iron bound in
cytochrome a3 are very close to each other forming a
binuclear metal center shown below
27
Electron transport chains are associated with
membranes such as the mitochondrial membrane in
eukaryotic organisms the thylakoid membrane of
photosynthetic organisms and the plasma membrane of
prokaryotic organisms While some of the details
differ electron transport chains in all organisms
operate on the same basic principles
Excited electrons are brought to the electron
transport chain by electron carriers such as NADH
and FADH2 In photosynthetic organisms the electrons
are donated by water and excited by light energy
The electron donor is oxidized as the electrons are
passed to the first protein complex in the chain As
it accepts the electrons the protein complex is
reduced Next the electrons are passed from the
28
first protein complex which becomes oxidized to
the second protein complex which becomes reduced
Each of these redox reactions is slightly exergonic
or energy releasing Some of this energy is used to
move protons across the membrane against their
concentration gradient With each successive
transfer of electrons the original excited
electrons lose some of their energy After several
successive redox reactions the electrons are
donated to a terminal electron acceptor In aerobic
respiration the terminal electron acceptor is
oxygen In photosynthesis the terminal electron
acceptor is a compound called NADP
The energy in the excited electrons has been
transformed into an electrochemical gradient
represented by the high concentration of protons on
one side of the membrane These protons can diffuse
back across the membrane by passing through a
protein complex called ATP synthase As these
protons pass through ATP synthase down their
concentration gradient they provide the energy
needed to make ATP from ADP and inorganic phosphate
within the cell
29
CHAPTER III
CLOSING
A Conclution
When a hexose is completely oxidized to CO2
and H2O using these three processes Glycolysis
yields two ATP and two NADH per hexose used Each
such NADH oxidized by the electron transport system
30
yields two ATP as dscribed above so glycolysis
contributes a total of six ATP per hexose The Krebs
cycle contributes two ATP per hexose or per two
pyruvates when succinyl CoA is cleaved to succinate
and CoASH This cycle also produces eight NADH per
hexose within the mitochondrial matrix by oxidative
phosporylation each of these NADH yields three ATP
or 24 per hexose Each FADH2 from the Krebs cycle
yields two ATP by oxydative phosporylation or four
per hexose (two pyruvates) The total contribution of
the Krebs cycle is then 30 Adding these 30 to the 6
from glycolysis leads to a total of 36 per hexose
completely respired by these process
B Suggestion
My greatest wish is that the readers find
this manuscript helpful in gaining the knowledge they
need for effective learning and practice and in
preparing for examination designed to test that
knowledge Reader comments and suggestions are
welcome and would be very helpful in improving our
manuscript Your contribution may assist many other
students in the coming years
31
REFERENCE
Anonim 2013 Anaerobic Respiration
httpwwwscoolcoukgcsebiologyrespiration
revise-itanaerobic-respiration Accessed
September 10th 2013
Anonim 2013 What is the difference between aerobic
and anaerobic respiration in plants
httpwwwbiologylifeeasyorg4010difference
-between-aerobic-anaerobic-respiration-plants
Accessed September 10th 2013
Campbell Mary K 2006 Biochemistry New York Thomson
Brooks Cole Inc
Dat J Capelli N Folzer H Bourgeade P Badot P-M
(2004) Sensing and signaling during
plantflooding Plant Physiology and
Biochemistry 42 273-282
Dat J Folzer H Parent C Badot P-M Capelli N
(2006) HypoxiastressCurrent Understanding and
Perspectives In Teixeira da Silva JA (Ed)
Floriculture Ornamental and Plant
Biotechnology Advances and Topical Issues (Vol
3) Global Science Books Isleworth United
Kingdompp 664-674
Hatta Muhammad 2011 Dunia Pertanian
httpemhattawordpresscomcategorystress
Accessed September 10th 2013
Hopkins William G 2009 Introduction to Plant Physiology
New York John Wiley amp Sons Inc
Lakitan Benyamin 2011 Dasar-Dasar Fisiologi Tumbuhan
Jakarta PT Raja Grafindo Persada
Larcher W 1980 Physiological Plant Ecology New York
Springer-Verlag
Raven 2013 Biology Fifth Edition
httphigheredmcgraw-hillcomsites983409233
9student_view0chapter39
photosynthetic_electron_transport_and_atp_synth
esishtml Accessed September 10th 2013
Salisbury Frank B 1969 Plant Physiology Third Edition
California Wadsworth Publishing Company
Subbaiah C Sachs M (2003) Molecular and cellular
adaptations of maize to flooding stress Annals
of Botany 91 119-127
Wilson CarlL1962 Botany Third EditionUnited State of
America Holt Rinch Inc

transport of electrons from NADH to CoQ is coupled to
the transport of protons across the membrane This is
an example of active transport The stiochiometry is
4 H+ transported per 2 electrons
2 Complex II
Believe it or not you are already familiar
with Complex II It is none other than succinate
dehydrogenase The only enzyme of the citric acid
cycle that is an integral membrane protein This
complex is composed of four subunits 2 of which are
iron-sulfur proteins and the other two subunits
together bind FAD through a covalent link to a
histidine residue These two subunits are called
flavoprotein 2 or FP2 Complex II contains 3 Fe-S
centers 1 4Fe-4S cluster 1 3Fe-4S cluster and 1
2Fe- 2S cluster In the first step of this complex
succinate is bound and a hydride is transferred to
FAD to generate FADH2 and fumarate FADH2 then
transfers its electrons one at a time to the Fe-S
centers Thus once again FAD functions as 2 electron
acceptor and a 1 electron donor The final step of
this complex is the transfer of 2 electrons one at a
time to coenzyme Q to produce CoQH2 For complex II
the standard free energy change of the overall
reaction is too small to drive the transport of
protons across the inner mitochondrial membrane This
25
accounts for the 15 ATPrsquos generated per FADH2
compared with the 25 ATPrsquos generated per NADH
3 Complex III
This complex is also known as coenzyme Q-
cytochrome c reductase because it passes the
electrons form CoQH2 to cyt c through a very unique
electron transport pathway called the Q-cycle Shown
to the left are the porphyrins found in cytochromes
Cytochrome b contains the same iron protoporphyrin as
hemoglobin and myoglobin The c cytochromes contain
heme c through covalent attachment by cysteine
residues Cytochrome a is found in two forms in
complex IV In complex III we find two b-type
cytochromes and one c-type cytochrome Complex III is
complex and we have a crystal structure
4 Complex IV
Complex IV is also known as cytochrome c oxidase
because it accepts the electrons from cytochrome c
and directs them towards the four electron reduction
of O2 to form 2 molecules of H2O
4 cyt c (Fe2+) + 4 H+ + O2 4 cyt c (Fe3+) +
2H2O
26
Cytochrome c oxidase contains 2 heme
centerscytochrome a and cytochrome a3 and two copper
proteins Each of the protein bound coppers are
associated with one of the cytochromes The copper
sites are called CuA and CuB CuA is associated with
cytochrome a and is shown to the left CuB is
associated with cytochrome a3 The copper sites
function as 1 electron carriers cycling between the
cuprous state Cu+ and the cupric state Cu2+ Just
like iron containing proteins they transfer
electrons one at a time Cytochrome c is bound on the
P-face of the membrane and transfers its electron to
CuA The oxidized cytochrome c dissociates CuA then
transfers the electron to cytochrome a The protein
bound CuA and the iron bound in cytochrome a are 15 Aring
apart In contrast the CuB and the iron bound in
cytochrome a3 are very close to each other forming a
binuclear metal center shown below
27
Electron transport chains are associated with
membranes such as the mitochondrial membrane in
eukaryotic organisms the thylakoid membrane of
photosynthetic organisms and the plasma membrane of
prokaryotic organisms While some of the details
differ electron transport chains in all organisms
operate on the same basic principles
Excited electrons are brought to the electron
transport chain by electron carriers such as NADH
and FADH2 In photosynthetic organisms the electrons
are donated by water and excited by light energy
The electron donor is oxidized as the electrons are
passed to the first protein complex in the chain As
it accepts the electrons the protein complex is
reduced Next the electrons are passed from the
28
first protein complex which becomes oxidized to
the second protein complex which becomes reduced
Each of these redox reactions is slightly exergonic
or energy releasing Some of this energy is used to
move protons across the membrane against their
concentration gradient With each successive
transfer of electrons the original excited
electrons lose some of their energy After several
successive redox reactions the electrons are
donated to a terminal electron acceptor In aerobic
respiration the terminal electron acceptor is
oxygen In photosynthesis the terminal electron
acceptor is a compound called NADP
The energy in the excited electrons has been
transformed into an electrochemical gradient
represented by the high concentration of protons on
one side of the membrane These protons can diffuse
back across the membrane by passing through a
protein complex called ATP synthase As these
protons pass through ATP synthase down their
concentration gradient they provide the energy
needed to make ATP from ADP and inorganic phosphate
within the cell
29
CHAPTER III
CLOSING
A Conclution
When a hexose is completely oxidized to CO2
and H2O using these three processes Glycolysis
yields two ATP and two NADH per hexose used Each
such NADH oxidized by the electron transport system
30
yields two ATP as dscribed above so glycolysis
contributes a total of six ATP per hexose The Krebs
cycle contributes two ATP per hexose or per two
pyruvates when succinyl CoA is cleaved to succinate
and CoASH This cycle also produces eight NADH per
hexose within the mitochondrial matrix by oxidative
phosporylation each of these NADH yields three ATP
or 24 per hexose Each FADH2 from the Krebs cycle
yields two ATP by oxydative phosporylation or four
per hexose (two pyruvates) The total contribution of
the Krebs cycle is then 30 Adding these 30 to the 6
from glycolysis leads to a total of 36 per hexose
completely respired by these process
B Suggestion
My greatest wish is that the readers find
this manuscript helpful in gaining the knowledge they
need for effective learning and practice and in
preparing for examination designed to test that
knowledge Reader comments and suggestions are
welcome and would be very helpful in improving our
manuscript Your contribution may assist many other
students in the coming years
31
REFERENCE
Anonim 2013 Anaerobic Respiration
httpwwwscoolcoukgcsebiologyrespiration
revise-itanaerobic-respiration Accessed
September 10th 2013
Anonim 2013 What is the difference between aerobic
and anaerobic respiration in plants
httpwwwbiologylifeeasyorg4010difference
-between-aerobic-anaerobic-respiration-plants
Accessed September 10th 2013
Campbell Mary K 2006 Biochemistry New York Thomson
Brooks Cole Inc
Dat J Capelli N Folzer H Bourgeade P Badot P-M
(2004) Sensing and signaling during
plantflooding Plant Physiology and
Biochemistry 42 273-282
Dat J Folzer H Parent C Badot P-M Capelli N
(2006) HypoxiastressCurrent Understanding and
Perspectives In Teixeira da Silva JA (Ed)
Floriculture Ornamental and Plant
Biotechnology Advances and Topical Issues (Vol
3) Global Science Books Isleworth United
Kingdompp 664-674
Hatta Muhammad 2011 Dunia Pertanian
httpemhattawordpresscomcategorystress
Accessed September 10th 2013
Hopkins William G 2009 Introduction to Plant Physiology
New York John Wiley amp Sons Inc
Lakitan Benyamin 2011 Dasar-Dasar Fisiologi Tumbuhan
Jakarta PT Raja Grafindo Persada
Larcher W 1980 Physiological Plant Ecology New York
Springer-Verlag
Raven 2013 Biology Fifth Edition
httphigheredmcgraw-hillcomsites983409233
9student_view0chapter39
photosynthetic_electron_transport_and_atp_synth
esishtml Accessed September 10th 2013
Salisbury Frank B 1969 Plant Physiology Third Edition
California Wadsworth Publishing Company
Subbaiah C Sachs M (2003) Molecular and cellular
adaptations of maize to flooding stress Annals
of Botany 91 119-127
Wilson CarlL1962 Botany Third EditionUnited State of
America Holt Rinch Inc

accounts for the 15 ATPrsquos generated per FADH2
compared with the 25 ATPrsquos generated per NADH
3 Complex III
This complex is also known as coenzyme Q-
cytochrome c reductase because it passes the
electrons form CoQH2 to cyt c through a very unique
electron transport pathway called the Q-cycle Shown
to the left are the porphyrins found in cytochromes
Cytochrome b contains the same iron protoporphyrin as
hemoglobin and myoglobin The c cytochromes contain
heme c through covalent attachment by cysteine
residues Cytochrome a is found in two forms in
complex IV In complex III we find two b-type
cytochromes and one c-type cytochrome Complex III is
complex and we have a crystal structure
4 Complex IV
Complex IV is also known as cytochrome c oxidase
because it accepts the electrons from cytochrome c
and directs them towards the four electron reduction
of O2 to form 2 molecules of H2O
4 cyt c (Fe2+) + 4 H+ + O2 4 cyt c (Fe3+) +
2H2O
26
Cytochrome c oxidase contains 2 heme
centerscytochrome a and cytochrome a3 and two copper
proteins Each of the protein bound coppers are
associated with one of the cytochromes The copper
sites are called CuA and CuB CuA is associated with
cytochrome a and is shown to the left CuB is
associated with cytochrome a3 The copper sites
function as 1 electron carriers cycling between the
cuprous state Cu+ and the cupric state Cu2+ Just
like iron containing proteins they transfer
electrons one at a time Cytochrome c is bound on the
P-face of the membrane and transfers its electron to
CuA The oxidized cytochrome c dissociates CuA then
transfers the electron to cytochrome a The protein
bound CuA and the iron bound in cytochrome a are 15 Aring
apart In contrast the CuB and the iron bound in
cytochrome a3 are very close to each other forming a
binuclear metal center shown below
27
Electron transport chains are associated with
membranes such as the mitochondrial membrane in
eukaryotic organisms the thylakoid membrane of
photosynthetic organisms and the plasma membrane of
prokaryotic organisms While some of the details
differ electron transport chains in all organisms
operate on the same basic principles
Excited electrons are brought to the electron
transport chain by electron carriers such as NADH
and FADH2 In photosynthetic organisms the electrons
are donated by water and excited by light energy
The electron donor is oxidized as the electrons are
passed to the first protein complex in the chain As
it accepts the electrons the protein complex is
reduced Next the electrons are passed from the
28
first protein complex which becomes oxidized to
the second protein complex which becomes reduced
Each of these redox reactions is slightly exergonic
or energy releasing Some of this energy is used to
move protons across the membrane against their
concentration gradient With each successive
transfer of electrons the original excited
electrons lose some of their energy After several
successive redox reactions the electrons are
donated to a terminal electron acceptor In aerobic
respiration the terminal electron acceptor is
oxygen In photosynthesis the terminal electron
acceptor is a compound called NADP
The energy in the excited electrons has been
transformed into an electrochemical gradient
represented by the high concentration of protons on
one side of the membrane These protons can diffuse
back across the membrane by passing through a
protein complex called ATP synthase As these
protons pass through ATP synthase down their
concentration gradient they provide the energy
needed to make ATP from ADP and inorganic phosphate
within the cell
29
CHAPTER III
CLOSING
A Conclution
When a hexose is completely oxidized to CO2
and H2O using these three processes Glycolysis
yields two ATP and two NADH per hexose used Each
such NADH oxidized by the electron transport system
30
yields two ATP as dscribed above so glycolysis
contributes a total of six ATP per hexose The Krebs
cycle contributes two ATP per hexose or per two
pyruvates when succinyl CoA is cleaved to succinate
and CoASH This cycle also produces eight NADH per
hexose within the mitochondrial matrix by oxidative
phosporylation each of these NADH yields three ATP
or 24 per hexose Each FADH2 from the Krebs cycle
yields two ATP by oxydative phosporylation or four
per hexose (two pyruvates) The total contribution of
the Krebs cycle is then 30 Adding these 30 to the 6
from glycolysis leads to a total of 36 per hexose
completely respired by these process
B Suggestion
My greatest wish is that the readers find
this manuscript helpful in gaining the knowledge they
need for effective learning and practice and in
preparing for examination designed to test that
knowledge Reader comments and suggestions are
welcome and would be very helpful in improving our
manuscript Your contribution may assist many other
students in the coming years
31
REFERENCE
Anonim 2013 Anaerobic Respiration
httpwwwscoolcoukgcsebiologyrespiration
revise-itanaerobic-respiration Accessed
September 10th 2013
Anonim 2013 What is the difference between aerobic
and anaerobic respiration in plants
httpwwwbiologylifeeasyorg4010difference
-between-aerobic-anaerobic-respiration-plants
Accessed September 10th 2013
Campbell Mary K 2006 Biochemistry New York Thomson
Brooks Cole Inc
Dat J Capelli N Folzer H Bourgeade P Badot P-M
(2004) Sensing and signaling during
plantflooding Plant Physiology and
Biochemistry 42 273-282
Dat J Folzer H Parent C Badot P-M Capelli N
(2006) HypoxiastressCurrent Understanding and
Perspectives In Teixeira da Silva JA (Ed)
Floriculture Ornamental and Plant
Biotechnology Advances and Topical Issues (Vol
3) Global Science Books Isleworth United
Kingdompp 664-674
Hatta Muhammad 2011 Dunia Pertanian
httpemhattawordpresscomcategorystress
Accessed September 10th 2013
Hopkins William G 2009 Introduction to Plant Physiology
New York John Wiley amp Sons Inc
Lakitan Benyamin 2011 Dasar-Dasar Fisiologi Tumbuhan
Jakarta PT Raja Grafindo Persada
Larcher W 1980 Physiological Plant Ecology New York
Springer-Verlag
Raven 2013 Biology Fifth Edition
httphigheredmcgraw-hillcomsites983409233
9student_view0chapter39
photosynthetic_electron_transport_and_atp_synth
esishtml Accessed September 10th 2013
Salisbury Frank B 1969 Plant Physiology Third Edition
California Wadsworth Publishing Company
Subbaiah C Sachs M (2003) Molecular and cellular
adaptations of maize to flooding stress Annals
of Botany 91 119-127
Wilson CarlL1962 Botany Third EditionUnited State of
America Holt Rinch Inc

Cytochrome c oxidase contains 2 heme
centerscytochrome a and cytochrome a3 and two copper
proteins Each of the protein bound coppers are
associated with one of the cytochromes The copper
sites are called CuA and CuB CuA is associated with
cytochrome a and is shown to the left CuB is
associated with cytochrome a3 The copper sites
function as 1 electron carriers cycling between the
cuprous state Cu+ and the cupric state Cu2+ Just
like iron containing proteins they transfer
electrons one at a time Cytochrome c is bound on the
P-face of the membrane and transfers its electron to
CuA The oxidized cytochrome c dissociates CuA then
transfers the electron to cytochrome a The protein
bound CuA and the iron bound in cytochrome a are 15 Aring
apart In contrast the CuB and the iron bound in
cytochrome a3 are very close to each other forming a
binuclear metal center shown below
27
Electron transport chains are associated with
membranes such as the mitochondrial membrane in
eukaryotic organisms the thylakoid membrane of
photosynthetic organisms and the plasma membrane of
prokaryotic organisms While some of the details
differ electron transport chains in all organisms
operate on the same basic principles
Excited electrons are brought to the electron
transport chain by electron carriers such as NADH
and FADH2 In photosynthetic organisms the electrons
are donated by water and excited by light energy
The electron donor is oxidized as the electrons are
passed to the first protein complex in the chain As
it accepts the electrons the protein complex is
reduced Next the electrons are passed from the
28
first protein complex which becomes oxidized to
the second protein complex which becomes reduced
Each of these redox reactions is slightly exergonic
or energy releasing Some of this energy is used to
move protons across the membrane against their
concentration gradient With each successive
transfer of electrons the original excited
electrons lose some of their energy After several
successive redox reactions the electrons are
donated to a terminal electron acceptor In aerobic
respiration the terminal electron acceptor is
oxygen In photosynthesis the terminal electron
acceptor is a compound called NADP
The energy in the excited electrons has been
transformed into an electrochemical gradient
represented by the high concentration of protons on
one side of the membrane These protons can diffuse
back across the membrane by passing through a
protein complex called ATP synthase As these
protons pass through ATP synthase down their
concentration gradient they provide the energy
needed to make ATP from ADP and inorganic phosphate
within the cell
29
CHAPTER III
CLOSING
A Conclution
When a hexose is completely oxidized to CO2
and H2O using these three processes Glycolysis
yields two ATP and two NADH per hexose used Each
such NADH oxidized by the electron transport system
30
yields two ATP as dscribed above so glycolysis
contributes a total of six ATP per hexose The Krebs
cycle contributes two ATP per hexose or per two
pyruvates when succinyl CoA is cleaved to succinate
and CoASH This cycle also produces eight NADH per
hexose within the mitochondrial matrix by oxidative
phosporylation each of these NADH yields three ATP
or 24 per hexose Each FADH2 from the Krebs cycle
yields two ATP by oxydative phosporylation or four
per hexose (two pyruvates) The total contribution of
the Krebs cycle is then 30 Adding these 30 to the 6
from glycolysis leads to a total of 36 per hexose
completely respired by these process
B Suggestion
My greatest wish is that the readers find
this manuscript helpful in gaining the knowledge they
need for effective learning and practice and in
preparing for examination designed to test that
knowledge Reader comments and suggestions are
welcome and would be very helpful in improving our
manuscript Your contribution may assist many other
students in the coming years
31
REFERENCE
Anonim 2013 Anaerobic Respiration
httpwwwscoolcoukgcsebiologyrespiration
revise-itanaerobic-respiration Accessed
September 10th 2013
Anonim 2013 What is the difference between aerobic
and anaerobic respiration in plants
httpwwwbiologylifeeasyorg4010difference
-between-aerobic-anaerobic-respiration-plants
Accessed September 10th 2013
Campbell Mary K 2006 Biochemistry New York Thomson
Brooks Cole Inc
Dat J Capelli N Folzer H Bourgeade P Badot P-M
(2004) Sensing and signaling during
plantflooding Plant Physiology and
Biochemistry 42 273-282
Dat J Folzer H Parent C Badot P-M Capelli N
(2006) HypoxiastressCurrent Understanding and
Perspectives In Teixeira da Silva JA (Ed)
Floriculture Ornamental and Plant
Biotechnology Advances and Topical Issues (Vol
3) Global Science Books Isleworth United
Kingdompp 664-674
Hatta Muhammad 2011 Dunia Pertanian
httpemhattawordpresscomcategorystress
Accessed September 10th 2013
Hopkins William G 2009 Introduction to Plant Physiology
New York John Wiley amp Sons Inc
Lakitan Benyamin 2011 Dasar-Dasar Fisiologi Tumbuhan
Jakarta PT Raja Grafindo Persada
Larcher W 1980 Physiological Plant Ecology New York
Springer-Verlag
Raven 2013 Biology Fifth Edition
httphigheredmcgraw-hillcomsites983409233
9student_view0chapter39
photosynthetic_electron_transport_and_atp_synth
esishtml Accessed September 10th 2013
Salisbury Frank B 1969 Plant Physiology Third Edition
California Wadsworth Publishing Company
Subbaiah C Sachs M (2003) Molecular and cellular
adaptations of maize to flooding stress Annals
of Botany 91 119-127
Wilson CarlL1962 Botany Third EditionUnited State of
America Holt Rinch Inc

Electron transport chains are associated with
membranes such as the mitochondrial membrane in
eukaryotic organisms the thylakoid membrane of
photosynthetic organisms and the plasma membrane of
prokaryotic organisms While some of the details
differ electron transport chains in all organisms
operate on the same basic principles
Excited electrons are brought to the electron
transport chain by electron carriers such as NADH
and FADH2 In photosynthetic organisms the electrons
are donated by water and excited by light energy
The electron donor is oxidized as the electrons are
passed to the first protein complex in the chain As
it accepts the electrons the protein complex is
reduced Next the electrons are passed from the
28
first protein complex which becomes oxidized to
the second protein complex which becomes reduced
Each of these redox reactions is slightly exergonic
or energy releasing Some of this energy is used to
move protons across the membrane against their
concentration gradient With each successive
transfer of electrons the original excited
electrons lose some of their energy After several
successive redox reactions the electrons are
donated to a terminal electron acceptor In aerobic
respiration the terminal electron acceptor is
oxygen In photosynthesis the terminal electron
acceptor is a compound called NADP
The energy in the excited electrons has been
transformed into an electrochemical gradient
represented by the high concentration of protons on
one side of the membrane These protons can diffuse
back across the membrane by passing through a
protein complex called ATP synthase As these
protons pass through ATP synthase down their
concentration gradient they provide the energy
needed to make ATP from ADP and inorganic phosphate
within the cell
29
CHAPTER III
CLOSING
A Conclution
When a hexose is completely oxidized to CO2
and H2O using these three processes Glycolysis
yields two ATP and two NADH per hexose used Each
such NADH oxidized by the electron transport system
30
yields two ATP as dscribed above so glycolysis
contributes a total of six ATP per hexose The Krebs
cycle contributes two ATP per hexose or per two
pyruvates when succinyl CoA is cleaved to succinate
and CoASH This cycle also produces eight NADH per
hexose within the mitochondrial matrix by oxidative
phosporylation each of these NADH yields three ATP
or 24 per hexose Each FADH2 from the Krebs cycle
yields two ATP by oxydative phosporylation or four
per hexose (two pyruvates) The total contribution of
the Krebs cycle is then 30 Adding these 30 to the 6
from glycolysis leads to a total of 36 per hexose
completely respired by these process
B Suggestion
My greatest wish is that the readers find
this manuscript helpful in gaining the knowledge they
need for effective learning and practice and in
preparing for examination designed to test that
knowledge Reader comments and suggestions are
welcome and would be very helpful in improving our
manuscript Your contribution may assist many other
students in the coming years
31
REFERENCE
Anonim 2013 Anaerobic Respiration
httpwwwscoolcoukgcsebiologyrespiration
revise-itanaerobic-respiration Accessed
September 10th 2013
Anonim 2013 What is the difference between aerobic
and anaerobic respiration in plants
httpwwwbiologylifeeasyorg4010difference
-between-aerobic-anaerobic-respiration-plants
Accessed September 10th 2013
Campbell Mary K 2006 Biochemistry New York Thomson
Brooks Cole Inc
Dat J Capelli N Folzer H Bourgeade P Badot P-M
(2004) Sensing and signaling during
plantflooding Plant Physiology and
Biochemistry 42 273-282
Dat J Folzer H Parent C Badot P-M Capelli N
(2006) HypoxiastressCurrent Understanding and
Perspectives In Teixeira da Silva JA (Ed)
Floriculture Ornamental and Plant
Biotechnology Advances and Topical Issues (Vol
3) Global Science Books Isleworth United
Kingdompp 664-674
Hatta Muhammad 2011 Dunia Pertanian
httpemhattawordpresscomcategorystress
Accessed September 10th 2013
Hopkins William G 2009 Introduction to Plant Physiology
New York John Wiley amp Sons Inc
Lakitan Benyamin 2011 Dasar-Dasar Fisiologi Tumbuhan
Jakarta PT Raja Grafindo Persada
Larcher W 1980 Physiological Plant Ecology New York
Springer-Verlag
Raven 2013 Biology Fifth Edition
httphigheredmcgraw-hillcomsites983409233
9student_view0chapter39
photosynthetic_electron_transport_and_atp_synth
esishtml Accessed September 10th 2013
Salisbury Frank B 1969 Plant Physiology Third Edition
California Wadsworth Publishing Company
Subbaiah C Sachs M (2003) Molecular and cellular
adaptations of maize to flooding stress Annals
of Botany 91 119-127
Wilson CarlL1962 Botany Third EditionUnited State of
America Holt Rinch Inc

first protein complex which becomes oxidized to
the second protein complex which becomes reduced
Each of these redox reactions is slightly exergonic
or energy releasing Some of this energy is used to
move protons across the membrane against their
concentration gradient With each successive
transfer of electrons the original excited
electrons lose some of their energy After several
successive redox reactions the electrons are
donated to a terminal electron acceptor In aerobic
respiration the terminal electron acceptor is
oxygen In photosynthesis the terminal electron
acceptor is a compound called NADP
The energy in the excited electrons has been
transformed into an electrochemical gradient
represented by the high concentration of protons on
one side of the membrane These protons can diffuse
back across the membrane by passing through a
protein complex called ATP synthase As these
protons pass through ATP synthase down their
concentration gradient they provide the energy
needed to make ATP from ADP and inorganic phosphate
within the cell
29
CHAPTER III
CLOSING
A Conclution
When a hexose is completely oxidized to CO2
and H2O using these three processes Glycolysis
yields two ATP and two NADH per hexose used Each
such NADH oxidized by the electron transport system
30
yields two ATP as dscribed above so glycolysis
contributes a total of six ATP per hexose The Krebs
cycle contributes two ATP per hexose or per two
pyruvates when succinyl CoA is cleaved to succinate
and CoASH This cycle also produces eight NADH per
hexose within the mitochondrial matrix by oxidative
phosporylation each of these NADH yields three ATP
or 24 per hexose Each FADH2 from the Krebs cycle
yields two ATP by oxydative phosporylation or four
per hexose (two pyruvates) The total contribution of
the Krebs cycle is then 30 Adding these 30 to the 6
from glycolysis leads to a total of 36 per hexose
completely respired by these process
B Suggestion
My greatest wish is that the readers find
this manuscript helpful in gaining the knowledge they
need for effective learning and practice and in
preparing for examination designed to test that
knowledge Reader comments and suggestions are
welcome and would be very helpful in improving our
manuscript Your contribution may assist many other
students in the coming years
31
REFERENCE
Anonim 2013 Anaerobic Respiration
httpwwwscoolcoukgcsebiologyrespiration
revise-itanaerobic-respiration Accessed
September 10th 2013
Anonim 2013 What is the difference between aerobic
and anaerobic respiration in plants
httpwwwbiologylifeeasyorg4010difference
-between-aerobic-anaerobic-respiration-plants
Accessed September 10th 2013
Campbell Mary K 2006 Biochemistry New York Thomson
Brooks Cole Inc
Dat J Capelli N Folzer H Bourgeade P Badot P-M
(2004) Sensing and signaling during
plantflooding Plant Physiology and
Biochemistry 42 273-282
Dat J Folzer H Parent C Badot P-M Capelli N
(2006) HypoxiastressCurrent Understanding and
Perspectives In Teixeira da Silva JA (Ed)
Floriculture Ornamental and Plant
Biotechnology Advances and Topical Issues (Vol
3) Global Science Books Isleworth United
Kingdompp 664-674
Hatta Muhammad 2011 Dunia Pertanian
httpemhattawordpresscomcategorystress
Accessed September 10th 2013
Hopkins William G 2009 Introduction to Plant Physiology
New York John Wiley amp Sons Inc
Lakitan Benyamin 2011 Dasar-Dasar Fisiologi Tumbuhan
Jakarta PT Raja Grafindo Persada
Larcher W 1980 Physiological Plant Ecology New York
Springer-Verlag
Raven 2013 Biology Fifth Edition
httphigheredmcgraw-hillcomsites983409233
9student_view0chapter39
photosynthetic_electron_transport_and_atp_synth
esishtml Accessed September 10th 2013
Salisbury Frank B 1969 Plant Physiology Third Edition
California Wadsworth Publishing Company
Subbaiah C Sachs M (2003) Molecular and cellular
adaptations of maize to flooding stress Annals
of Botany 91 119-127
Wilson CarlL1962 Botany Third EditionUnited State of
America Holt Rinch Inc

CHAPTER III
CLOSING
A Conclution
When a hexose is completely oxidized to CO2
and H2O using these three processes Glycolysis
yields two ATP and two NADH per hexose used Each
such NADH oxidized by the electron transport system
30
yields two ATP as dscribed above so glycolysis
contributes a total of six ATP per hexose The Krebs
cycle contributes two ATP per hexose or per two
pyruvates when succinyl CoA is cleaved to succinate
and CoASH This cycle also produces eight NADH per
hexose within the mitochondrial matrix by oxidative
phosporylation each of these NADH yields three ATP
or 24 per hexose Each FADH2 from the Krebs cycle
yields two ATP by oxydative phosporylation or four
per hexose (two pyruvates) The total contribution of
the Krebs cycle is then 30 Adding these 30 to the 6
from glycolysis leads to a total of 36 per hexose
completely respired by these process
B Suggestion
My greatest wish is that the readers find
this manuscript helpful in gaining the knowledge they
need for effective learning and practice and in
preparing for examination designed to test that
knowledge Reader comments and suggestions are
welcome and would be very helpful in improving our
manuscript Your contribution may assist many other
students in the coming years
31
REFERENCE
Anonim 2013 Anaerobic Respiration
httpwwwscoolcoukgcsebiologyrespiration
revise-itanaerobic-respiration Accessed
September 10th 2013
Anonim 2013 What is the difference between aerobic
and anaerobic respiration in plants
httpwwwbiologylifeeasyorg4010difference
-between-aerobic-anaerobic-respiration-plants
Accessed September 10th 2013
Campbell Mary K 2006 Biochemistry New York Thomson
Brooks Cole Inc
Dat J Capelli N Folzer H Bourgeade P Badot P-M
(2004) Sensing and signaling during
plantflooding Plant Physiology and
Biochemistry 42 273-282
Dat J Folzer H Parent C Badot P-M Capelli N
(2006) HypoxiastressCurrent Understanding and
Perspectives In Teixeira da Silva JA (Ed)
Floriculture Ornamental and Plant
Biotechnology Advances and Topical Issues (Vol
3) Global Science Books Isleworth United
Kingdompp 664-674
Hatta Muhammad 2011 Dunia Pertanian
httpemhattawordpresscomcategorystress
Accessed September 10th 2013
Hopkins William G 2009 Introduction to Plant Physiology
New York John Wiley amp Sons Inc
Lakitan Benyamin 2011 Dasar-Dasar Fisiologi Tumbuhan
Jakarta PT Raja Grafindo Persada
Larcher W 1980 Physiological Plant Ecology New York
Springer-Verlag
Raven 2013 Biology Fifth Edition
httphigheredmcgraw-hillcomsites983409233
9student_view0chapter39
photosynthetic_electron_transport_and_atp_synth
esishtml Accessed September 10th 2013
Salisbury Frank B 1969 Plant Physiology Third Edition
California Wadsworth Publishing Company
Subbaiah C Sachs M (2003) Molecular and cellular
adaptations of maize to flooding stress Annals
of Botany 91 119-127
Wilson CarlL1962 Botany Third EditionUnited State of
America Holt Rinch Inc

yields two ATP as dscribed above so glycolysis
contributes a total of six ATP per hexose The Krebs
cycle contributes two ATP per hexose or per two
pyruvates when succinyl CoA is cleaved to succinate
and CoASH This cycle also produces eight NADH per
hexose within the mitochondrial matrix by oxidative
phosporylation each of these NADH yields three ATP
or 24 per hexose Each FADH2 from the Krebs cycle
yields two ATP by oxydative phosporylation or four
per hexose (two pyruvates) The total contribution of
the Krebs cycle is then 30 Adding these 30 to the 6
from glycolysis leads to a total of 36 per hexose
completely respired by these process
B Suggestion
My greatest wish is that the readers find
this manuscript helpful in gaining the knowledge they
need for effective learning and practice and in
preparing for examination designed to test that
knowledge Reader comments and suggestions are
welcome and would be very helpful in improving our
manuscript Your contribution may assist many other
students in the coming years
31
REFERENCE
Anonim 2013 Anaerobic Respiration
httpwwwscoolcoukgcsebiologyrespiration
revise-itanaerobic-respiration Accessed
September 10th 2013
Anonim 2013 What is the difference between aerobic
and anaerobic respiration in plants
httpwwwbiologylifeeasyorg4010difference
-between-aerobic-anaerobic-respiration-plants
Accessed September 10th 2013
Campbell Mary K 2006 Biochemistry New York Thomson
Brooks Cole Inc
Dat J Capelli N Folzer H Bourgeade P Badot P-M
(2004) Sensing and signaling during
plantflooding Plant Physiology and
Biochemistry 42 273-282
Dat J Folzer H Parent C Badot P-M Capelli N
(2006) HypoxiastressCurrent Understanding and
Perspectives In Teixeira da Silva JA (Ed)
Floriculture Ornamental and Plant
Biotechnology Advances and Topical Issues (Vol
3) Global Science Books Isleworth United
Kingdompp 664-674
Hatta Muhammad 2011 Dunia Pertanian
httpemhattawordpresscomcategorystress
Accessed September 10th 2013
Hopkins William G 2009 Introduction to Plant Physiology
New York John Wiley amp Sons Inc
Lakitan Benyamin 2011 Dasar-Dasar Fisiologi Tumbuhan
Jakarta PT Raja Grafindo Persada
Larcher W 1980 Physiological Plant Ecology New York
Springer-Verlag
Raven 2013 Biology Fifth Edition
httphigheredmcgraw-hillcomsites983409233
9student_view0chapter39
photosynthetic_electron_transport_and_atp_synth
esishtml Accessed September 10th 2013
Salisbury Frank B 1969 Plant Physiology Third Edition
California Wadsworth Publishing Company
Subbaiah C Sachs M (2003) Molecular and cellular
adaptations of maize to flooding stress Annals
of Botany 91 119-127
Wilson CarlL1962 Botany Third EditionUnited State of
America Holt Rinch Inc

REFERENCE
Anonim 2013 Anaerobic Respiration
httpwwwscoolcoukgcsebiologyrespiration
revise-itanaerobic-respiration Accessed
September 10th 2013
Anonim 2013 What is the difference between aerobic
and anaerobic respiration in plants
httpwwwbiologylifeeasyorg4010difference
-between-aerobic-anaerobic-respiration-plants
Accessed September 10th 2013
Campbell Mary K 2006 Biochemistry New York Thomson
Brooks Cole Inc
Dat J Capelli N Folzer H Bourgeade P Badot P-M
(2004) Sensing and signaling during
plantflooding Plant Physiology and
Biochemistry 42 273-282
Dat J Folzer H Parent C Badot P-M Capelli N
(2006) HypoxiastressCurrent Understanding and
Perspectives In Teixeira da Silva JA (Ed)
Floriculture Ornamental and Plant
Biotechnology Advances and Topical Issues (Vol
3) Global Science Books Isleworth United
Kingdompp 664-674
Hatta Muhammad 2011 Dunia Pertanian
httpemhattawordpresscomcategorystress
Accessed September 10th 2013
Hopkins William G 2009 Introduction to Plant Physiology
New York John Wiley amp Sons Inc
Lakitan Benyamin 2011 Dasar-Dasar Fisiologi Tumbuhan
Jakarta PT Raja Grafindo Persada
Larcher W 1980 Physiological Plant Ecology New York
Springer-Verlag
Raven 2013 Biology Fifth Edition
httphigheredmcgraw-hillcomsites983409233
9student_view0chapter39
photosynthetic_electron_transport_and_atp_synth
esishtml Accessed September 10th 2013
Salisbury Frank B 1969 Plant Physiology Third Edition
California Wadsworth Publishing Company
Subbaiah C Sachs M (2003) Molecular and cellular
adaptations of maize to flooding stress Annals
of Botany 91 119-127
Wilson CarlL1962 Botany Third EditionUnited State of
America Holt Rinch Inc

Hatta Muhammad 2011 Dunia Pertanian
httpemhattawordpresscomcategorystress
Accessed September 10th 2013
Hopkins William G 2009 Introduction to Plant Physiology
New York John Wiley amp Sons Inc
Lakitan Benyamin 2011 Dasar-Dasar Fisiologi Tumbuhan
Jakarta PT Raja Grafindo Persada
Larcher W 1980 Physiological Plant Ecology New York
Springer-Verlag
Raven 2013 Biology Fifth Edition
httphigheredmcgraw-hillcomsites983409233
9student_view0chapter39
photosynthetic_electron_transport_and_atp_synth
esishtml Accessed September 10th 2013
Salisbury Frank B 1969 Plant Physiology Third Edition
California Wadsworth Publishing Company
Subbaiah C Sachs M (2003) Molecular and cellular
adaptations of maize to flooding stress Annals
of Botany 91 119-127
Wilson CarlL1962 Botany Third EditionUnited State of
America Holt Rinch Inc