pimelodella spelaea: a new cave catfish from central brazil, with data on ecology and evolutionary...
TRANSCRIPT

q 2004 by the American Society of Ichthyologists and Herpetologists
Copeia, 2004(2), pp. 315–325
Pimelodella spelaea: A New Cave Catfish from Central Brazil, with Data onEcology and Evolutionary Considerations (Siluriformes: Heptapteridae)
ELEONORA TRAJANO, ROBERTO E. REIS, AND MARIA ELINA BICHUETTE
Pimelodella spelaea, new species, is described from a subterranean stream tributaryto the Sao Bernardo River inside the Sao Bernardo Cave, in the Sao Domingos karstarea, upper Tocantins River basin, central Brazil. The new species can be distin-guished from its epigean (surface) congeners by derived troglomorphic featuressuch as smaller eyes and fainter coloration, which are nevertheless less reduced thanin the other known troglobitic (exclusively subterranean) species, Pimelodella kronei,from southeast Brazil. Ecological data such as habitat characteristics, populationdata based on mark-recapture techniques, and distributions of frequencies of stan-dard length, weight, and condition factor, were obtained during the dry season of2000 (May to September). A relatively high population density (about one individualper m22) was recorded for the accessible habitat, which is probably much lower inthe nonaccessible, phreatic area of distribution. The low condition factor recordedfor P. spelaea, which tended to decrease along the study period, indicates a regimeof severe food limitation, intensifying along the dry season. An allopatric model ofdifferentiation is hypothesized, either because of topographic isolation or becauseof local extinction of epigean relatives, or both factors combined. The conservationstatus of the new species is also discussed.
Pimelodella spelaea, especie nova, e descrita de um riacho subterraneo tributariodo rio Sao Bernardo dentro da caverna Sao Bernardo, na area carstica de SaoDomingos, alto rio Tocantins, Brasil. A nova especie pode ser distinguida dos seuscongeneres epıgeos (de superfıcie) por caracterısticas troglomorficas derivadas taiscomo olhos e pigmentacao reduzidos. Estes sao, entretanto, menos reduzidos quena outra especie troglobia (exclusivamente subterranea) conhecida, Pimelodella kro-nei, do sudeste do Brasil. Dados ecologicos tais como caracterısticas do habitat,dados de populacao baseados em tecnicas de marcacao e recaptura, e distribuicaode frequencias de comprimento-padrao, peso e fator de condicao, foram obtidosdurante a estacao seca de 2000 (maio a setembro). Uma densidade populacionarelativamente alta (cerca de 1 ind. m22) foi registrada na porcao acessıvel do habitatda especie, que e possivelmente muito menor na area freatica, nao acessıvel da suadistribuicao. Os baixos fatores de condicao registrados para P. spelaea, que tende-rem a diminuir ao longo do perıodo de estudo, indicam um regime de severa li-mitacao alimentar, intensificando-se ao longo da estacao seca. Um modelo alopatricode diferenciacao e sugerido, seja pelo isolamento topografico ou pela extincao localde parente epıgeos, ou ainda por ambos os fatores combinados. O estado de con-servacao da nova especie tambem e discutido.
THE Neotropical region is remarkable for itsrich subterranean ichthyofauna, only com-
parable to that from China and southeasternAsia. More than 35 among approximately 100troglobitic (exclusively subterranean) fish spe-cies occurring around the world are knownfrom Mexico, Caribbean, Venezuela, Ecuador,Peru, Bolivia and Brazil, including several un-described taxa (Weber et al., 1998; Romero andPaulson, 2001; ET and MEB, unpubl. data). Thegreat majority are siluriforms belonging to theHeptapteridae (Rhamdia and Pimelodella, amongothers), Trichomycteridae (Trichomycterus andItuglanis), and Loricariidae (Ancistrus). The
prevalence of these groups in the subterraneanbiotope is an evidence of high potential for col-onization of the subterranean habitats, includ-ing caves, probably facilitated by their predom-inantly nocturnal activity and generalized car-nivorous (heptapterids, trichomycterids) or de-tritivorous (Ancistrus) diet (Trajano, 2001).Troglobitic species are generally characterizedby autapomorphies related to the isolation inthe perpetually dark subterranean habitat (tro-glomorphisms), the most widespread being thereduction or even loss of eyes and pigmenta-tion.
The Sao Domingos karst area, a limestone re-

316 COPEIA, 2004, NO. 2
gion in Central Brazil crossed by tributaries ofthe upper Tocantins River, Amazon basin, pre-sents the most diversified cave ichthyofauna inthe country. In the course of our studies in thisarea, we discovered a new cave species of hep-tapterid catfish, genus Pimelodella, living in slow-moving pools in an upper conduit of the SaoBernardo Cave. Herein we describe this newspecies and present data on population ecologygathered during the dry season of 2002. This isthe second troglobitic species known for the ge-nus, the first one being Pimelodella kronei (Ribei-ro, 1907), from southeastern Brazil.
The genus Pimelodella, with 71 valid speciesand a cis- and trans-Andean distribution fromPanama to Paraguay and southern Brazil, is thelargest among the Heptapteridae (Bockmannand Guazelli, 2003). Pimelodella is currently inneed of a taxonomic revision being last re-viewed by Eigenmann (1917) who diagnosedthe genus based on several not unique featuresbut including a few that provide a diagnosiswhen in combination. These features are ‘‘. . .a long, adnate, adipose fin; caudal fin deeplyforked; a frontal and a parietal fontanel, the lat-ter reaching the base of the occipital process,which is narrow and reaches, or nearly reachesthe plate in front of the dorsal; and humeralprocess spine-like . . . ’’ (Eigenmann, 1917:229).More recently, however, Bockmann (1998) pre-sented two characters that may represent syna-pomorphies for Pimelodella or to a subgroup ofspecies within Pimelodella. These are (1) the twoinnermost caudal-fin rays not directly articulat-ing with hypural bones (character 220), and (2)those same rays united by an incomplete caudal-fin membrane (character 223). The new speciesdescribed below shares the second characterbut not the first, because the two innermost cau-dal-fin rays directly articulate with the hypuralbones.
MATERIALS AND METHODS
The fieldwork was conducted in May, July,and September 2000 and was restricted to thedry season because of the danger of beingcaught by floods in the rainy period (Octoberto March). Collections were made with handnets in May and with funnel-type minnow trapsbaited with chicken liver in July and September.Baits were kept in small nylon bags to avoid be-ing swallowed by the fish and interfering withweight measurements. Fish were anesthetized ina solution of MS-222 (tricaine-methanesulpho-nate), measured (standard length, SL) to thenearest 0.1 mm, and weighed with a Pesola dy-namometer, 0.1 g precision. Most catfishes were
marked by subcutaneous injection of fluores-cent pigments (‘‘Photonics marking,’’ fromNew West Co.), and released. Association ofmark color and position, and fish size, pro-duced individual marks. Some specimens weresacrificed by overanesthesia and preserved informalin (later transferred to alcohol 70GL) fordescription. Only eight specimens were pre-served to avoid unnecessary mortality in thepopulation, whose total size was unknown.
Mark-recapture data from May to July andfrom July to September were used separately toestimate the number of individuals in the studysection using the Lincoln-Petersen Index (Be-gon, 1979; Blower et al., 1981). We did not usethe whole set of data for a single estimation us-ing a multiple-recapture method (such as thetriple-catch) because there is evidence of someloss of marks by pigment reabsorption duringour four-month observation period.
Standard length and weight data were usedto calculate the allometric condition factors (K5 100.W.SL2b; Le Cren, 1951). For these cal-culations, units of millimeters and grams wereused. The power coefficient (b) was estimatedfrom the slope of the regression of log weighton log standard length using the pooled data.Dice-Leraas diagrams (Dice and Leraas, 1936)were used to show tendencies along the studyperiod. Graphs with the distributions of fre-quencies of size (SL), weight and condition fac-tor (K) in July and in September were built; Maywas not included here because of the low sam-ple size. July and September samples were com-pared by two-sample t-test, with a 5 5%.
For comparison, we estimated the averagecondition factor for P. kronei from the Areiascave, upper Ribeira karst area, in the Atlanticrain forest in southeastern Brazil, based on datagathered between May and September of 1984and 1985 (ET, unpubl. data), a period of lowrainfall locally corresponding to the dry season.
Measurements of preserved fish, taken withelectronic calipers (0.1 mm precision), followedbasically Lundberg et al. (1991) and Trajanoand Britski (1992). In the description, meristicdata are followed by number of specimens dis-playing each count in parentheses. Density ofchromatophores was estimated by direct countsunder stereomicroscope on a 1-mm2 area locat-ed immediately posterior of the dorsal-fin ori-gin, midway between the lateral line and thedorsal fin. A cleared-and-stained specimen wasprepared according to Taylor and Van Dyke(1985).
No Pimelodella species has been describedfrom the upper Tocantins River to date. How-ever, watersheds around the upper Tocantins in-

317TRAJANO ET AL.—NEW CAVE PIMELODELLA FROM BRAZIL
Fig. 1. Holotype of Pimelodella spelaea, MZUSP 81726, 80.5 mm SL; Brazil, State of Goias, Sao Domingos,upper Tocantins River basin, subterranean stream tributary to Sao Bernardo River inside Sao Bernardo Cave.Photograph by Pablo Lehmann.
clude the Araguaia, the Sao Francisco, and theupper Parana River basins. Five species were de-scribed from these areas: Pimelodella boschmai, Pi-melodella meeki, and Pimelodella avanhandavaefrom the upper Parana River, and Pimelodellalaurenti and Pimelodella vittata from the Sao Fran-cisco River. A sixth, unidentified, probably un-described, species from the upper TocantinsRiver (MCP 16056) is also included here andlisted below as Pimelodella sp. 5. Pimelodella spe-laea is compared to these species below, but be-cause of the confused state of the taxonomy ofPimelodella, it was not practical to compare morebroadly across the genus.
Pimelodella spelaea new speciesFigure 1
Holotype.—MZUSP 81726, 80.5 mm SL, Brazil,State of Goias, Sao Domingos, upper TocantinsRiver basin, subterranean stream tributary toSao Bernardo River inside Sao Bernardo Cave,138499S 468219W, 26 July 2000, E. Trajano andM. E. Bichuette.
Paratypes.—Seven specimens, same locality ofholotype. MZUSP 81727, 42.8 mm SL, and MCP31482, 1 1 1 c&s, 47.5–66.7 mm SL, same dataof holotype. MZUSP 81728, 47.3 mm SL and
318 COPEIA, 2004, NO. 2
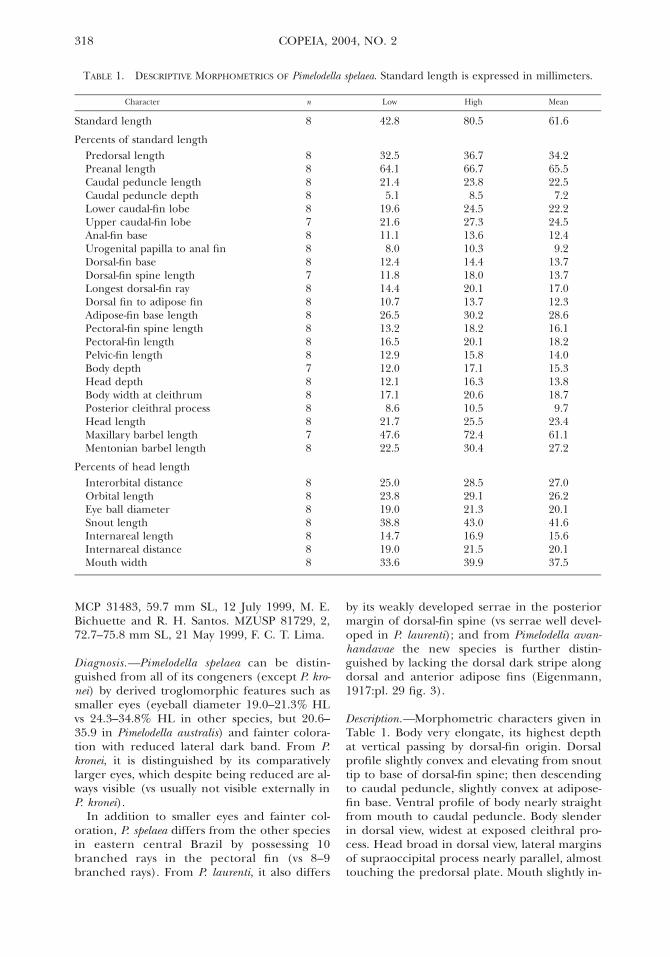
TABLE 1. DESCRIPTIVE MORPHOMETRICS OF Pimelodella spelaea. Standard length is expressed in millimeters.
Character n Low High Mean
Standard length 8 42.8 80.5 61.6
Percents of standard lengthPredorsal lengthPreanal lengthCaudal peduncle lengthCaudal peduncle depthLower caudal-fin lobe
88888
32.564.121.45.1
19.6
36.766.723.88.5
24.5
34.265.522.57.2
22.2Upper caudal-fin lobeAnal-fin baseUrogenital papilla to anal finDorsal-fin baseDorsal-fin spine length
78887
21.611.18.0
12.411.8
27.313.610.314.418.0
24.512.49.2
13.713.7
Longest dorsal-fin rayDorsal fin to adipose finAdipose-fin base lengthPectoral-fin spine lengthPectoral-fin length
88888
14.410.726.513.216.5
20.113.730.218.220.1
17.012.328.616.118.2
Pelvic-fin lengthBody depthHead depthBody width at cleithrumPosterior cleithral processHead lengthMaxillary barbel lengthMentonian barbel length
87888878
12.912.012.117.18.6
21.747.622.5
15.817.116.320.610.525.572.430.4
14.015.313.818.79.7
23.461.127.2
Percents of head lengthInterorbital distanceOrbital lengthEye ball diameterSnout lengthInternareal lengthInternareal distanceMouth width
8888888
25.023.819.038.814.719.033.6
28.529.121.343.016.921.539.9
27.026.220.141.615.620.137.5
MCP 31483, 59.7 mm SL, 12 July 1999, M. E.Bichuette and R. H. Santos. MZUSP 81729, 2,72.7–75.8 mm SL, 21 May 1999, F. C. T. Lima.
Diagnosis.—Pimelodella spelaea can be distin-guished from all of its congeners (except P. kro-nei) by derived troglomorphic features such assmaller eyes (eyeball diameter 19.0–21.3% HLvs 24.3–34.8% HL in other species, but 20.6–35.9 in Pimelodella australis) and fainter colora-tion with reduced lateral dark band. From P.kronei, it is distinguished by its comparativelylarger eyes, which despite being reduced are al-ways visible (vs usually not visible externally inP. kronei).
In addition to smaller eyes and fainter col-oration, P. spelaea differs from the other speciesin eastern central Brazil by possessing 10branched rays in the pectoral fin (vs 8–9branched rays). From P. laurenti, it also differs
by its weakly developed serrae in the posteriormargin of dorsal-fin spine (vs serrae well devel-oped in P. laurenti); and from Pimelodella avan-handavae the new species is further distin-guished by lacking the dorsal dark stripe alongdorsal and anterior adipose fins (Eigenmann,1917:pl. 29 fig. 3).
Description.—Morphometric characters given inTable 1. Body very elongate, its highest depthat vertical passing by dorsal-fin origin. Dorsalprofile slightly convex and elevating from snouttip to base of dorsal-fin spine; then descendingto caudal peduncle, slightly convex at adipose-fin base. Ventral profile of body nearly straightfrom mouth to caudal peduncle. Body slenderin dorsal view, widest at exposed cleithral pro-cess. Head broad in dorsal view, lateral marginsof supraoccipital process nearly parallel, almosttouching the predorsal plate. Mouth slightly in-

319TRAJANO ET AL.—NEW CAVE PIMELODELLA FROM BRAZIL
Fig. 2. Pimelodella spelaea, pectoral-fin spine, dorsalview. Scale 2 mm.
ferior; small, conic teeth irregularly arranged inplates on dentaries and premaxillae. Eyes re-duced in size (19.0–21.3% HL) but apparentlyfunctional. Maxillary barbels comparativelyshort (47.6–72.4% SL), reaching between pel-vic-fin insertion and origin of anal fin. Mentalbarbels short, outer slightly longer, reachingend of pectoral-fin base.
Dorsal fin with one spinelet, one spine and 6(7) or 7 (1) branched rays, originating approx-imately above end of pseudotympanum. Dorsal-fin spine smooth, without serrae and compara-tively short, its length about three-fourths offirst branched ray. Distal margin of dorsal finslightly convex. Adipose fin comparatively short(26.5–30.2% SL), its origin on vertical throughurogenital papilla, and its end slightly posteriorto end of depressed anal fin. Adipose fin withslightly convex dorsal margin and very smallposterior free lobe. Anal fin short and with con-vex posterior margin, with 5 (7) or 6 (1) simpleand 8 (1), 9 (6), or 10 (1) branched rays. Cau-dal fin deeply forked, dorsal lobe slightly longerthan ventral. Upper lobe with i17 (8) and lowerlobe with i18 (8) principal rays.
Pectoral fins with one spine and 10 (8)branched rays (one specimen with 9 branchedrays on one side), reaching vertical throughthird or fourth dorsal-fin ray when depressed.Pectoral-fin spine with weak retrorse serrae oc-cupying up to about two-thirds of posterior mar-gin and very small thorns on anterior margin,restricted to proximal portion (Fig. 2). Serraeon posterior margin of pectoral-fin spine in-creasing with size, 5–6 in smaller to 11–13 inlarger specimens. Bony, pungent portion ofspine shorter than first branched ray. Distalmargin of dorsal fin slightly convex. Posteriorprocess of cleithrum roughly triangular, con-cave posterodorsally, ornamented with a seriesof longitudinal, low bony ridges. Pelvic finshort, reaching midway between urogenital pa-pilla and anal-fin origin, with one unbranchedand 5 (8) branched rays. Pelvic fin originatingon or slightly posterior of vertical through endof dorsal-fin base.
Color in alcohol.—Overall body coloration paleyellowish, lighter ventrally. Dorsum of body,from top of head to behind adipose fin, slightly
darkened. Eyes dark grey. Concentration ofchromatophores on supraoccipital process andpredorsal plate smaller than in neighboringskin, making bones lighter than fleshy dorsum.Skin covering dorsal-fin spine and dorsal mar-gin of adipose fin slightly duskier. Chromato-phores more concentrated on pseudotympa-num and continuing to caudal pedunclearound lateral line, forming faint lateral stripe,hardly distinguishable in some specimens.
Color in life.—Dorsal portions of body light graywith a faint lateral stripe. Fins and most of body,especially caudal peduncle highly translucent.When removed from water, skin has slight pur-ple or greenish iridescence.
Etymology.—Specific epithet spelaea from the Lat-in speleum, cave, cavern, in allusion to its habitat.An adjective.
Distribution.—Known from its type-locality only,a small upper tributary of the Sao Bernardo Riv-er inside the Sao Bernardo Cave, upper Tocan-tins River, Sao Domingos, State of Goias, Brazil.
Habitat and ecology.—The type-locality of P. spe-laea is situated within the limits of the TerraRonca State Park (138309–138509S, 468109–468309W), in Sao Domingos, eastern State ofGoias. Sao Domingos is a carbonatic karst areacharacterized by the presence of continuouslimestone outcrops belonging to the BambuıFormation and crossed by several parallel clearwater streams running westward to join the Pa-rana River, a tributary of the upper TocantinsRiver. It lies in the Cerrado (the savannah-likeBrazilian vegetation) phytogeographic domainand is characterized by a tropical semi-humidclimate (Nimer, 1989). This is a seasonal climatewith a well-defined dry season from March andApril to September and October, and possibilityof strong rains in December to February, whencaves or cave conduits may be partially or com-pletely flooded depending on conduit dimen-sions and topography. The Sao Bernardo Cave(138499S, 0468219W) is part of a cave system(Sao Bernardo-Palmeiras) with 8000 linear m ofdevelopment. This particular cave has about3500 m of linear development and is crossed bya subterranean river with mean width anddepth of 5 m and 0.60 m, respectively. There isa small tributary in the left margin where thefishes occur 700 m downstream from the caveentrance. This vadose tributary is in a higherlevel compared with the main stream and emp-ties into it through a small waterfall. The fishesoccur mainly in soft-bottomed pools with very

320 COPEIA, 2004, NO. 2
slow current. About 1500 m downstream fromthis tributary, the Sao Bernardo River joins thePalmeiras River inside the cave.
The accessible fish habitat is a slow moving,soft-bottomed pool, 5 m long, 2 m wide and av-eraging 1.5 m deep, situated 5 m above themainstream level. Water from the pool forms ashallow water course flowing over a rocky bedalong 50 m and discharging into the main-stream through a small waterfall. Access to thefish habitat upstream of the pool requires scubagear, and the total extent of this habitat is un-known. Just a few Pimelodella catfishes were seenin the shallow reach. Four tetra characins (Crea-grutus sp.) and a small armored catfish (Hypos-tomus sp.) were found along with P. spelaea.
Physico-chemical variables in the P. spelaeapool, measured with a digital HORIBA appara-tus in May 1999/May 2000/September 2000, arethe following: temperature (C) 5 24.5/24.8/24.6; pH 5 7.31/7.15/7.30; dissolved oxygen(mg l21) 5 7.45/7.65/–; conductivity (ms cm21)5 0.588/0.607/0.546; salinity (%) 5 0.02/0.02/0.02. In the main subterranean river, the envi-ronmental variables in May/1999/May/2000/August/2000 were temperature (C) 5 23.6/25.7/24.5; pH 5 6.36/7.35/7.40; dissolved oxy-gen (mg l21) 5 8.66/7.67/7.99; conductivity(ms cm21) 5 0.013/0.038/0.024; salinity (%) 50/0/0. These variables measured in May 1999in the epigean reach before the main cavestream sinkhole were temperature (C) 5 24.8; pH5 5.63; dissolved oxygen (mg l21) 5 8.03; con-ductivity (ms cm21) 5 0.010; salinity (%) 5 0.
The physico-chemical conditions of water(e.g., pH higher than in base-level streams andtypical of slow moving karst waters) and the ab-sence of small epigean tributaries to the SaoBernardo River basin indicate that the hypoge-an upper tributary where P. spelaea lives origi-nates from epikarst waters (the uppermost layerof a karstified rock in which a large proportionof fissures have been enlarged by solutional ero-sion allowing for a rapid infiltration and storageof large quantities of water; Drew, unpubl.),forming a perched aquifer.
Data on number of captured, marked and re-captured specimens in each date are as follows:1 May: 13 captured, marked and released (1 8–9 observed), SL 5 49.4–81.7 mm, weight 5 1.4–4.5 g; 26 July: 57 captured (including three re-captures), 53 marked and released, SL 5 43.7–87.3 mm; weight 5 0.9–5.4 g; 8 September: 43captured (including 24 recaptures, one dou-ble), SL 5 56.7–77.0 mm; weight 5 1.6–4.4 g.The number of individuals in the study section,estimated by Lincoln-Petersen method, is 188.5(SE 5 81.3) for beginning of May, and 95 (SE
5 12) for the end of July (assuming losses butno gains along the study period).
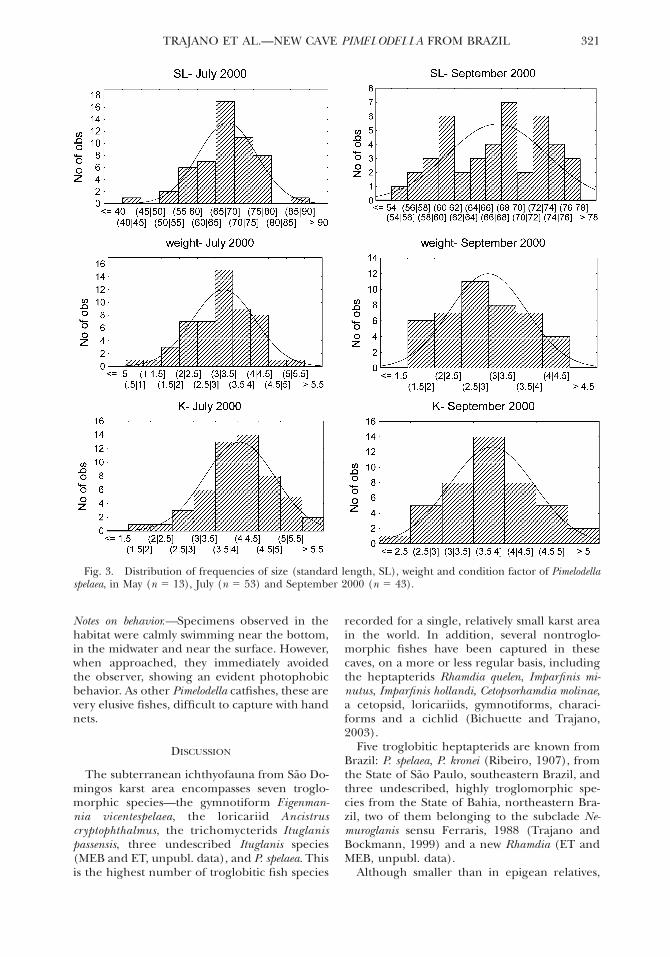
The distributions of frequencies of standardlength, weight and condition factor (K) in Julyand September in the cave habitat are shown inFigure 3. Most specimens examined were veryskinny and some were severely underfed. Formost specimens recaptured in September (16 of24 captured fish), the measured values of SLand weight were lower than in July, representingcases of weight loss associated with negativegrowth in length, that is, these fish shrank (con-ditions during the measurements, including thecaliper used, the person taking the measure-ments and the degree of fish anesthesia, weresimilar at all occasions). After a period of 44days, the differences in standard length (SL inSeptember, SL in May) ranged from 21.0 to10.3 mm (mean 5 20.25 mm), and all but onerecaptured fish lost weight (differences inweight ranged from 0 to 20.7 g). No specimenshowed developed gonads (that would be seenby transparency, if present).
The power coefficient (b) for the pooled datawas 2.351. Mean (and standard deviation) forstandard length, weight and condition factorwere, respectively, May (n 5 13)264.7 (8.09),3.17 (0.86), 4.16 (0.78); July (n 5 53)268.1(7.87), 3.24 (0.88), 4.06 (0.82); and August (n5 43)267.4 (6.30), 2.98 (0.71), 3.79 (0.68).Tendencies in monthly means depicted as Dice-Leraas diagrams are shown in Figure 4A–C. Atendency of decrease in weight and conditionfactor from July to September is apparent in thegraphs (Figs. 3–4), which is in accordance withdata from recaptured specimens. Nevertheless,t-test failed to show a significant difference be-tween these months: for standard length, t 50.423, P 5 0.673; for weight, t 5 1.520, P 50.132; for condition factor (K), t 5 1.713, P 50.090 (marginally significant). The latter is closeto the adopted critical significance value (P 50.05), supporting the idea of a tendency of de-crease in condition factor toward the end of thedry season.
For P. kronei, the power coefficient (b) was2.768 for the pooled data (1984 plus 1985; n 576), and 2.806 for 1984 alone (n 5 55). No dif-ference was observed between consecutive years(Fig. 4D).
Comparison of the length measurements inthe field (under anesthesia) and after preser-vation in alcohol of six specimens of P. spelaearevealed differences of 1.2–6.5% of standardlength, with values always lower after preserva-tion. Such differences should be taken into ac-count when comparing size distributions of fishsamples before and after preservation.
321TRAJANO ET AL.—NEW CAVE PIMELODELLA FROM BRAZIL
Fig. 3. Distribution of frequencies of size (standard length, SL), weight and condition factor of Pimelodellaspelaea, in May (n 5 13), July (n 5 53) and September 2000 (n 5 43).
Notes on behavior.—Specimens observed in thehabitat were calmly swimming near the bottom,in the midwater and near the surface. However,when approached, they immediately avoidedthe observer, showing an evident photophobicbehavior. As other Pimelodella catfishes, these arevery elusive fishes, difficult to capture with handnets.
DISCUSSION
The subterranean ichthyofauna from Sao Do-mingos karst area encompasses seven troglo-morphic species—the gymnotiform Eigenman-nia vicentespelaea, the loricariid Ancistruscryptophthalmus, the trichomycterids Ituglanispassensis, three undescribed Ituglanis species(MEB and ET, unpubl. data), and P. spelaea. Thisis the highest number of troglobitic fish species
recorded for a single, relatively small karst areain the world. In addition, several nontroglo-morphic fishes have been captured in thesecaves, on a more or less regular basis, includingthe heptapterids Rhamdia quelen, Imparfinis mi-nutus, Imparfinis hollandi, Cetopsorhamdia molinae,a cetopsid, loricariids, gymnotiforms, characi-forms and a cichlid (Bichuette and Trajano,2003).
Five troglobitic heptapterids are known fromBrazil: P. spelaea, P. kronei (Ribeiro, 1907), fromthe State of Sao Paulo, southeastern Brazil, andthree undescribed, highly troglomorphic spe-cies from the State of Bahia, northeastern Bra-zil, two of them belonging to the subclade Ne-muroglanis sensu Ferraris, 1988 (Trajano andBockmann, 1999) and a new Rhamdia (ET andMEB, unpubl. data).
Although smaller than in epigean relatives,

322 COPEIA, 2004, NO. 2
Fig. 4. Dice-Leraas diagrams showing tendencies in standard length (A), weight (B), and condition factor(C) of Pimelodella spelaea, from Central Brazil, during the dry season of 2000 (May, n 5 13; July, n 5 53;September, n 5 43), and condition factor of Pimelodella kronei (D), from southeastern Brazil, during the dryseasons of 1984 and 1985. Central squares 5 means; vertical bars 5 6 standard errors.
the eyes of P. spelaea show a low degree of re-duction in comparison to other troglobitic hep-tapterids, such as P. kronei (the great majority ofspecimens with eyes not externally visible; Tra-jano and Britski, 1992), and the heptapteridsfrom Bahia, which are completely eyeless andunpigmented. A great deal of intra- and inter-specific variation in eyes development is ob-served among cave Rhamdia species (Weber,1996), but most have eyes far more reducedthan P. spelaea.
Pigmentation is also not greatly reduced in P.spelaea. Although it is clearly paler than mostepigean relatives, the color pattern typical of Pi-melodella catfishes is distinguishable in mostspecimens, with a faint, but generally visible lat-eral stripe. Paler color may be attributed to thedensity of chromatophores in P. spelaea, that is,lower than in epigean species. A comparisonwith an unidentified epigean species of Pimelo-
della (MCP 16056) from the same river basin ofP. spelaea, the upper Tocantins River, revealed asignificant lower density of chromatophores persquare millimeter in the latter (P. spelaea: 71–110 [n 5 8, mean 91.5, SD 17.5], Pimelodella sp.:110–220 [n 5 10, mean 154.8, SD 38.6], P 50.008).
The high recapture rates in P. spelaea (53%from July to September) indicate permanencein the study site at least for short periods (44days). Preference for slow-moving waters withsoft-bottom and tendency to sedentariness wasalso observed for the troglobitic P. kronei (Tra-jano, 1991) and its epigean putative sister-spe-cies, Pimelodella transitoria (P. Gerhard, unpubl.data).
Comparison with another cave Pimelodella cat-fish, P. kronei, studied in seasons correspondingwith the study period for P. spelaea, emphasizesthe low average condition factor observed for

323TRAJANO ET AL.—NEW CAVE PIMELODELLA FROM BRAZIL
the latter, at least during the dry season. Pime-lodella kronei clearly is a more robust and betterfed cave catfish. This may be partly a species-specific difference, with P. spelaea descendingfrom an epigean taxon with a more delicateconstitution, and partly caused by differences infood availability along the year in their respec-tive cave habitats. Pimelodella kronei lives in base-level cave streams that flow continuously duringthe dry season, which is not as accentuated andwell defined as in P. spelaea locality. Therefore,a reasonable amount of food items would beavailable throughout the year for P. kronei.
The low condition factor recorded for P. spe-laea, which tended to decrease along the studyperiod, indicates a regime of severe food limi-tation, intensifying along the dry season. Thesame was reported for the troglobitic trichomyc-terid Trichomycterus itacarambiensis, from easternBrazil, also with cases of negative growth(shrinkage in length; Trajano, 1997). Thecleared-and-stained P. spelaea specimen, a verythin, severely underfed individual, showed noskeletal abnormalities, including the vertebrae.Therefore, apparently the shrinkage in lengthrecorded in some of the marked and subse-quently recaptured fishes is not explained by al-terations in the axial skeleton.
Feeding stress increasing along the dry sea-son is consistent with the decrease in the num-ber of specimens estimated for the study sectionfrom May to July, possibly a consequence of losscaused by mortality. However, the density ofspecimens in the study site (about one individ-ual per m22) may be considered high for cave(Trajano, 2001) and for epigean standards aswell. The higher availability of prey in large, va-dose (aerated) conduits, represented by terres-trial food items, explains the concentration ofP. spelaea in the accessible study area. A muchlower population density is expected to occur inthe phreatic area of distribution.
The actual extent of P. spelaea habitat is notknown, but the cave topography, the absence ofrecords in other localities in spite of the inten-sive collecting effort in the area, and the lackof adaptations to life in small phreatic spaces,indicate a geographically restricted habitat.Therefore, we estimated the total populationsize of P. spelaea in hundreds, possibly reaching1000 individuals, which is a small populationsize even for cave fish standards (Trajano,2001).
The low degree of troglomorphism affectingeyes and melanic pigmentation of P. spelaea isconsistent with the observed for the other cavefishes from Sao Domingos karst area. Eigenman-nia vicentespelaea, from a different cave system,
exhibits a similar low degree of reduction ofeyes and pigmentation (MEB, unpubl. data),and a great deal of intra- and interpopulationvariation occurs in Ancistrus cryptophthalmus(Bessa and Trajano, 2002). The four species ofItuglanis have more reduced eyes and pigmen-tation, but there is still some individual variationand none of these populations is homogeneous-ly anopthalmic and unpigmented (MEB, un-publ. data), unlike troglobitic catfishes found insome other Brazilian karst areas (e.g., the newheptapterids from Bahia, and the loricariid An-cistrus formoso, from Mato Grosso do Sul). Thisindicates relatively short times of isolation in thesubterranean habitat for the cave fishes fromSao Domingos.
The vadose tributary where P spelaea lives isnowadays mostly isolated from the main, base-level drainage, by differences in topographiclevel, as a consequence of alluvial downcuttingresulting in a regional progressive lowering ofthe water table. Epigean Pimelodella ancestorsprobably lived in water courses at the preteritebase level, directly flowing into caves and allow-ing for the establishment of cave populations.As a consequence of alluvial downcutting, suchformerly running-water streams would becomeperched and fed by infiltration water from theepikarst. Therefore, differences in topographiclevel may account for (at least partial) isolationof fish populations surviving in perched waterbodies. Topographic isolation has been pro-posed by Borowsky and Mertz (2001) to explainthe genetic differentiation between populationsof the cave balitorid, Schistura oedipus, fromThailand.
However, in spite of intensive collecting ef-forts in the epigean stream reaches of the area,where other heptapterids (Imparfinis hollandi,Cetopsorhamdia molinae, Phenacorhamdia tenebrosa)have been sampled, no Pimelodella catfish wasfound (Bichuette and Trajano, 2003). It is pos-sible that epigean populations also descendingfrom the ancestor of the troglobitic Pimelodelladisappeared from this area. We hypothesize thatthe morphological differentiation resulting inthe presently described taxon occurred in allop-atry, either because of topographic isolation orbecause of local extinction of epigean relatives,or both factors combined.
This also applies to the three undescribed tro-globitic species of Ituglanis from Sao Domingosregion, respectively, from upper vadose tributar-ies in Angelica and in Sao Mateus caves andfrom a side pool in Sao Bernardo Cave (notsyntopic with P. spelaea), for which no epigeanrelatives have been found in the area. However,the model does not fit A. cryptophthalmus and E.

324 COPEIA, 2004, NO. 2
vicentespelaea, that live in subterranean base-levelstreams continuous with epigean reaches whereepigean relatives are currently found. There-fore, the expressive diversity of cave fishes inSao Domingos karst area seems to be the resultof a combination of different, but all relativelyrecent, events and processes.
Conservation remarks.—It is well known that sub-terranean ecosystems are fragile, very vulnera-ble to disturbance, and several troglobitic fishesaround the world are currently endangered bythreats such as habitat degradation, hydrologi-cal manipulations, pollution, and human activ-ities (Proudlove, 2001). In the case of P. spelaea,no immediate threat has been identified. Nev-ertheless, the fact that, in spite of intensive studyin the caves of Sao Domingos karst area, P. spe-laea was found in only one restricted site insidea single cave, the Sao Bernardo Cave (high de-gree of endemism), and the periods of severefood stress to which this species is subject, pointto an intrinsic fragility calling for protectivemeasures. The type locality of P. spelaea is situ-ated within the boundaries of a State Park, theParque Estadual de Terra Ronca. However, vis-itation to caves is poorly controlled, and nomanagement plan has been enforced so far. En-forcement of control of visitation to Sao Ber-nardo Cave, with interdiction of access to P. spe-laea site, is therefore recommended for effectiveprotection of this species.
COMPARATIVE MATERIAL EXAMINED
All localities in Brazil: Pimelodella australis,MCP 9349 (48 specimens), Rio Saica near Ca-cequi, Rio Grande do Sul. Pimelodella boschmai,MZUSP 22932 (8 specimens), Rio Mogi-Guacuat Emas, Sao Paulo. Pimelodella gracilis, MCP23140 (10 specimens), Rio Jaguarı near SaoFrancisco de Assis, Rio Grande do Sul. Pimelo-della kronei, MCP 18312 (2 specimens) Areias deCima Cave, Iporanga, Sao Paulo. Pimelodella la-teristriga, MCP 12574 (10 specimens), Corregoda Posse near Carmo, Rio de Janeiro. Pimelodellalaurenti, ANSP 69380 (holotype) and ANSP69381 (2 specimens), Rio Sao Francisco at Ja-toba, Pernambuco, and MZUSP 46454 (5 spec-imens), Rio Verde at Montes Claros, Minas Ge-rais. Pimelodella meeki, MCP 28519 (1 specimen),Ribeirao Cachoeira near Piracicaba, Sao Paulo,MCP 28520 (3 specimens), Rio Piracicaba at Pi-racicaba, Sao Paulo, and MZUSP 22485 (8 spec-imens) Rio Corumbataı at Corumbataı, SaoPaulo. Pimelodella pappenhaimi, MCP 16526 (13specimens), Ribeirao Sao Luıs near Apiuna,Santa Catarina. Pimelodella transitoria, MCP
28513 (6 specimens), Rio Ipiranga, near SeteBarras, Sao Paulo. Pimelodella vittata, MCP 16695(46 specimens), tributary to Rio Sao Francisconear Bocaiuva, Minas Gerais and MZUSP 47310(5 specimens), Rio Jequitaı, between Buenopo-lis e Engenheiro Dolabela, Minas Gerais. Pime-lodella sp. 1, MCP 25003 (10 specimens), RioGuama near Sao Miguel do Guama, Para. Pi-meldella sp. 2, MCP 15783 (6 specimens), Rio doBugre, near Porto Esperidiao, Mato Grosso. Pi-melodella sp. 3, MCP 15708 (5 specimens), Riodo Brugre at Barra do Bugre, Mato Grosso. Pi-melodella sp. 4, MCP 15775 (62 specimens), RioParaguay at Caceres, Mato Grosso. Pimelodellasp. 5, MCP 16056 (15 specimens), Riacho Araranear Rosariana, Niquelandia, Goias.
ACKNOWLEDGMENTS
We are greatly indebted to the colleagues andspeleologists who helped us during the field-work, especially to F. C. T. Lima, A. Chagas Jr.,E. P. D. Costa Jr., L. F. Duboc and R. Borowsky,and to R. H. Santos, our local guide. We alsothank R. Schulz, manager of the Parque Esta-dual Terra Ronca, who allowed us to use thePark facilities, providing all the necessary sup-port during the field trips. The photograph ofthe holotype is by P. Lehmann. The Fundacaode Amparo a Pesquisa do Estado de Sao Pauloprovided grants for fieldwork (FAPESP 99/0376-1). Two authors are partially supported bythe Conselho Nacional de DesenvolvimentoCientıfico e Tecnologico (CNPq fellowship n.306066/88-2 [ET] and 305344/87-0 [RER]).Permission for collecting species was granted byIBAMA.
LITERATURE CITED
BEGON, M. 1979. Estimating animal abundance. Ed-ward Arnold, London.
BESSA, E., AND E. TRAJANO. 2002. Light reaction andcryptobiotic habits in armoured catfishes, genusAncistrus, from caves in Central and Northwest Bra-zil (Siluriformes: Loricariidae). Mem. Biospeol. 28:29–37.
BICHUETTE, M. E., AND E. TRAJANO. 2003. Epigean andsubterranean ichthyofauna from Sao Domingoskarst area, Upper Tocantins River basin, CentralBrazil. J. Fish Biol. 63:1100–1121.
BLOWER, J. G., L. M. COOK, AND J. A. BISHOP. 1981.Estimating the size of animal populations. GeorgeAllen and Unwin, London.
BOCKMANN, F. A. 1998. Analise filogenetica da famıliaHeptapteridae (Teleostei, Ostariophysi, Silurifor-mes) e redefinicao de seus generos. Unpubl. Ph.D.diss., Univ. of Sao Paulo, Sao Paulo, Brazil.
———, AND G. M. GUAZZELLI. 2003. Family Heptap-teridae, p. 406–431. In: Check list of the freshwater

325TRAJANO ET AL.—NEW CAVE PIMELODELLA FROM BRAZIL
fishes of South and Central America. R. E. Reis, S.O. Kullander, and C. J. Ferraris Jr. (eds.). Edi-pucrs,Porto Alegre, Brazil.
BOROWSKY, R., AND L. MERTZ. 2001. Genetic differen-tiation among populations of the Cave Fish Schis-tura oedipus (Cypriniformes: Balitoridae). Environ.Biol. Fish. 62:225–231.
DICE, L. R., AND H. J. LERAAS. 1936. A graphic methodfor comparing several sets of measurements. Con-trib. Lab. Vert. Genet. Univ. Mich. 3:1–3.
EIGENMANN, C. H. 1917. Pimelodella and Typhlobagrus.Mem. Carnegie Mus. 7:229–258.
FERRARIS, C. 1988. Relationships of the Neotropicalcatfish genus Nemuroglanis, with a description of anew species (Osteichthyes, Siluriformes, Pimelodi-dae). Proc. Biol. Soc. Wash. 101:509–516.
LE CREN, E. D. 1951. The length-weight relationshipsand seasonal cycle in gonad weight and conditionin the perch (Perca fluviatilis). J. Anim. Ecol. 20:201–219.
LUNDBERG, J. G., A. L. BORNBUSCH, AND F. MAGO-LEC-CIA. 1991. Gladioglanis conquistador n. sp. form Ec-uador with diagnoses of the subfamilies Rhamdii-nae Bleeker and Pseudopimelodinae N. Subf. (Si-luriformes: Pimelodidae). Copeia 1991:190–209.
NIMER, E. 1989. Climatologia do Brasil. SUPREN, Riode Janeiro, Brazil.
PROUDLOVE, G. S. 2001. The conservation status ofhypogean fishes. Environ. Biol. Fish. 62:201–213[special volume ‘‘The biology of hypogean fishes’’].
ROMERO, A., AND K. M. PAULSON. 2001. It’s a wonder-ful hypogean life: a guide to the troglomorphic fish-es of the world. Ibid. 62:13–41 [special volume ‘‘Thebiology of hypogean fishes’’].
TAYLOR, W. R., AND G. C. VAN DYKE. 1985. Revisedprocedures for staining and clearing small fishesand other vertebrates for bone and cartilage study.Cybium 9:107–119.
TRAJANO, E. 1991. Populational ecology of Pimelodellakronei, troglobitic catfish from southeastern Brazil(Siluriformes, Pimelodidae). Environ. Biol. Fish.30:407–421.
———. 1997. Population ecology of Trichomycterus ita-carambiensis, a cave catfish from eastern Brazil (Si-luriformes, Trichomycteridae). Ibid. 50:357–369.
———. 2001. Ecology of subterranean fishes: an over-view. Ibid. 62:133–160 [special volume ‘‘The biologyof hypogean fishes’’].
———, AND H. A. BRITSKI. 1992. Pimelodella kronei (Ri-beiro, 1907) e seu sinonimo Caecorhamdella brasilien-sis Borodin, 1927: morfologia externa, taxonomia eevolucao (Teleostomi, Siluriformes). Bolm. Zool.,Sao Paulo 12:53–89.
———, AND F. A. BOCKMANN. 1999. Evolution of ecol-ogy and behaviour in Brazilian cave Heptapterinaecatfishes, based on cladistic analysis (Teleostei: Si-luriformes). Mem. Biospeol. 26:123–129.
WEBER, A. 1996. Cave dwelling catfish populations ofthe genus Rhamdia (Pimelodidae, Siluroidei, teleos-tei) in Mexico. Mem. Biospeol. 23:73–85.
———, G. S. PROUDLOVE, J. PARZEFALL, H. WILKENS,AND T. T. NALBANT. 1998. Pisces (Teleostei), p.1179–1190. In: Encyclopaedia Biospeologica, TomeII. C. Juberthie and V. Decu (eds.). Societe de Bios-peologie,Moulis, France.
(ET, MEB) DEPARTAMENTO DE ZOOLOGIA, INS-TITUTO DE BIOCIENCIAS DA USP; P.O. BOX
11461, 05422-970 SAO PAULO, BRAZIL; AND
(RER) LABORATORIO DE ICTIOLOGIA, PONTIF-ıCIA UNIVERSIDADE CATOLICA DO RIO GRANDE
DO SUL; P.O. BOX 1429, 90619-900 PORTO
ALEGRE, RS, BRAZIL. E-mail: (ET) [email protected]. Send reprints request to ET. Submit-ted: 30 May 2003. Accepted: 19 Oct. 2003.Section editor: J. W. Armbruster.