phases of polyoma virus infection t cells differ for acute and
TRANSCRIPT

of February 9, 2022.This information is current as
Phases of Polyoma Virus Infection T Cells Differ for Acute and Persistent+CD8
Costimulation Requirements for Antiviral
and Aron E. LukacherSzomolanyi-Tsuda, Thomas C. Pearson, Christian P. Larsen Christopher C. Kemball, Eun D. Han Lee, Eva
http://www.jimmunol.org/content/176/3/1814doi: 10.4049/jimmunol.176.3.1814
2006; 176:1814-1824; ;J Immunol
Referenceshttp://www.jimmunol.org/content/176/3/1814.full#ref-list-1
, 46 of which you can access for free at: cites 66 articlesThis article
average*
4 weeks from acceptance to publicationFast Publication! •
Every submission reviewed by practicing scientistsNo Triage! •
from submission to initial decisionRapid Reviews! 30 days* •
Submit online. ?The JIWhy
Subscriptionhttp://jimmunol.org/subscription
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/About/Publications/JI/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/alertsReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists All rights reserved.Copyright © 2006 by The American Association of1451 Rockville Pike, Suite 650, Rockville, MD 20852The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on February 9, 2022http://w
ww
.jimm
unol.org/D
ownloaded from
by guest on February 9, 2022
http://ww
w.jim
munol.org/
Dow
nloaded from

Costimulation Requirements for Antiviral CD8� T Cells Differfor Acute and Persistent Phases of Polyoma Virus Infection1
Christopher C. Kemball,2* Eun D. Han Lee,2† Eva Szomolanyi-Tsuda,‡ Thomas C. Pearson,†
Christian P. Larsen,† and Aron E. Lukacher3*
The requirement for costimulation in antiviral CD8� T cell responses has been actively investigated for acutely resolved viralinfections, but it is less defined for CD8� T cell responses to persistent virus infection. Using mouse polyoma virus (PyV) as a modelof low-level persistent virus infection, we asked whether blockade of the CD40 ligand (CD40L) and CD28 costimulatory pathwaysimpacts the magnitude and function of the PyV-specific CD8� T response, as well as the humoral response and viral control duringacute and persistent phases of infection. Costimulation blockade or gene knockout of either CD28 or CD40L substantially damp-ened the magnitude of the acute CD8� T cell response; simultaneous CD28 and CD40L blockade severely depressed the acute Tcell response, altered the cell surface phenotype of PyV-specific CD8� T cells, decreased PyV VP1-specific serum IgG titers, andresulted in an increase in viral DNA levels in multiple organs. CD28 and CD40L costimulation blockade during acute infectionalso diminished the memory PyV-specific CD8� T cell response and serum IgG titer, but control of viral persistence variedbetween mouse strains and among organs. Interestingly, we found that CD28 and CD40L costimulation is dispensable for gen-erating and/or maintaining PyV-specific CD8� T cells during persistent infection; however, blockade of CD27 and CD28 co-stimulation in persistently infected mice caused a reduction in PyV-specific CD8� T cells. Taken together, these data indicate thatCD8� T cells primed within the distinct microenvironments of acute vs persistent virus infection differ in their costimulationrequirements. The Journal of Immunology, 2006, 176: 1814–1824.
C omplete Ag-driven T cell differentiation generally re-quires the contribution of multiple costimulatory signalsthat amplify the signal initiated by TCR engagement with
a cognate peptide-MHC ligand. Optimal CD8� T cell responses toacute viral infections require costimulation through at least one ofseveral pathways, including B7-CD28, CD40-CD40 ligand(CD40L),4 41BB-41BB ligand, and CD27-CD70 (1, 2). The rela-tive importance of each of these costimulatory signals is oftenvirus dependent. The generation and maintenance of lymphocyticchoriomeningitis virus (LCMV)-specific CD8� effector and mem-ory T cells is largely CD28 independent (3–5), whereas vesicularstomatitis virus, vaccinia virus, and influenza virus (IV)-specificCD8� T cell responses are CD28 dependent (5–9). Acute, but notmemory, LCMV-specific CD8� T cell responses are also CD40-CD40L independent, whereas vesicular stomatitis virus-specificCD8� T cell responses are CD40-CD40L dependent (10–12). Fur-
thermore, 41BB-41BB ligand costimulation is necessary for opti-mal primary, memory, and secondary CD8� T cell responses toLCMV and IV infections (4, 13–16). CD27 costimulation pro-motes the survival and accumulation of IV-specific CD8� T cellsat effector sites (17, 18).
Less is known about the requirement for costimulation of anongoing CD8� T cell response to a persistent virus infection. Inthe setting where CD8� T cells are buffeted by Ag long-term,costimulation may be required to sustain protective immunity andkeep the virus in check. The most thoroughly characterized mousemodels of persistent virus infection include gammaherpesvirus 68(�HV-68) and highly replicating strains of LCMV (e.g., clone 13or t1b). The magnitude of the �HV-68-specific CD8� T cell re-sponse is normal in CD40L�/� mice, although CD40�/� andCD40L�/� mice lack long-term control of the virus (19–21).CD28�/� mice, in contrast, maintain long-term viral control butmount a delayed CD8� T cell IFN-� response, despite normalCTL activity (20). Virus-specific CD8� T cell responses inCD40L�/� mice infected by LCMV clone 13 or t1b undergo pro-gressive functional deterioration (22). In addition, Ab-mediatedblockade of the B7-CD28 and CD40-CD40L pathways during per-sistent infection by LCMV clone 13 significantly decreases virus-specific CD8� T cell IFN-� production and prolongs viremia (23).These data suggest that antiviral CD8� T cells require continualCD28 and/or CD40L costimulation for protective immunity in thesettings of latent (e.g., �HV-68) and high-level (e.g., LCMV clone13) persistent infection. In humans infected by HIV and CMV, alarge fraction of virus-specific CD8� T cells express CD27 (24,25). Little information is available on the contribution of CD27costimulation to maintaining antiviral CD8� T cells in persistentinfection.
The requirements for costimulation to elicit and maintain CD8�
T cell responses against viruses that establish low-level, systemicpersistent infection are not well defined. Mouse polyoma virus
*Department of Pathology and †Department of Surgery, Emory University School ofMedicine, Atlanta, GA 30322; and ‡Department of Pathology, University of Massa-chusetts Medical School, Worcester, MA 01655
Received for publication July 18, 2005. Accepted for publication November 10, 2005.
The costs of publication of this article were defrayed in part by the payment of pagecharges. This article must therefore be hereby marked advertisement in accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.1 This work was supported by National Institutes of Health Grants R01 CA71971 (toA.E.L.) and P01 AI44644 (to C.P.L.), Public Health Service Grant R01 CA66644 (toE.S.-T.) from the National Cancer Institute, and a Novartis Basic Science Fellowshipfrom the American Society of Transplantation (to E.D.H.L.).2 C.C.K. and E.D.H.L. contributed equally to this study.3 Address correspondence and reprint requests to Dr. Aron E. Lukacher, Departmentof Pathology, Woodruff Memorial Research Building, Room 7307, Emory UniversitySchool of Medicine, 101 Woodruff Circle, Atlanta, GA 30322. E-mail address:[email protected] Abbreviations used in this paper: CD40L, CD40 ligand; LCMV, lymphocytic cho-riomeningitis virus; IV, influenza virus; �HV-68, gammaherpesvirus 68; PyV, poly-oma virus; CD62L, L-selectin; p.i., postinfection.
The Journal of Immunology
Copyright © 2006 by The American Association of Immunologists, Inc. 0022-1767/06/$02.00
by guest on February 9, 2022http://w
ww
.jimm
unol.org/D
ownloaded from

(PyV) establishes a low-level persistent infection in immunocom-petent inbred strains of mice, which require a PyV-specific CD8�
T cell response for control of viral persistence and protectionagainst tumor development (26–28). PyV epitope-specific CD8�
T cell populations vary in rates of expansion and contraction, hi-erarchy, phenotype, and cytokine profile in acute and persistentinfection (29). Importantly, we found that new effector PyV-spe-cific CD8� T cells are primed during the persistent phase of in-fection (29). Antiviral CD8� T cells newly recruited during per-sistent infection may be particularly dependent on costimulationfor optimal priming because reduced or absent virus-induced in-flammatory signals during persistent infection might elevate theneed for costimulation. In addition, CD40-CD40L interaction is amajor component of CD4� T cell help provided to B cells (30).Thus, blockade of the CD40-CD40L pathway decreases virus-spe-cific IgG responses to a number of virus infections, such asLCMV, vaccinia virus, and PyV, but it does not completely abolishthem (10, 31). The contribution of the B7-CD28 costimulatorypathway to IgG responses in PyV infection has not been previouslyexamined.
In this study, we used costimulation blockade and homozygousgene knockout mice to determine how obstruction of several co-stimulatory pathways impacts the magnitude and functionality ofthe PyV-specific CD8� T cell response, humoral response, andviral clearance. Costimulation blockade or gene knockout of eitherCD28 or CD40L substantially dampened the magnitude of theacute PyV-specific response. Simultaneous blockade of these path-ways severely depressed the acute PyV-specific CD8� T cell re-sponse, altered the cell surface phenotype of PyV-specific CD8� Tcells, decreased PyV VP1-specific serum IgG titers, and was as-sociated with an increase in viral DNA levels in multiple organs.Combined CD28 and CD40L blockade during acute infection alsodampened the memory PyV-specific CD8� T cell response andserum IgG titer, but control of viral persistence varied betweenmouse strains and among organs. We further demonstrate thatPyV-specific CD8� T cells generated de novo during persistentinfection do not require CD28 and CD40L costimulation. PyV-specific CD8� T cells declined in number when CD27 and CD28costimulatory pathways were blocked during persistent infection.Taken together, these findings show that the costimulation require-ments of PyV-specific CD8� T cells differ between acute and per-sistent phases of infection.
Materials and MethodsMice
C57BL/6NCr (B6) and C3H/HeNCr female mice were purchased from theFrederick Cancer Research and Development Center of the National Can-cer Institute. B6.129S2-Tnfsf5tm1Imx (CD40L�/� on a B6 background) andB6.129S2-Cd28tm1Mak (CD28�/� on a B6 background) mice were pur-chased from The Jackson Laboratory. B6.SJL-Ptprca/BoAiTac (CD45.1)mice were purchased from Taconic. Mice were housed and bred in accor-dance with the guidelines of the Institutional Animal Care and Use Com-mittee and the Department of Animal Resources at Emory University.
Viruses and virus inoculation
PyV strain A2 was molecularly cloned and plaque purified, and virusstocks were prepared on primary baby mouse kidney cells as previouslydescribed (28). Each mouse was inoculated s.c. in each hind footpad with2 � 106 PFU of virus. Mice were infected at 6–12 wk of age.
Peptides and class I MHC tetramers
MT245-253, LT638-646, and MT389-397 peptides were synthesizedand stored as described (29). LT359-368C7Abu (SAVKNY[Abu]SKL),in which the cysteine residue at position 7 was replaced with �-amino-butyric acid, a thiol-less cysteine analog residue, was synthesized by theEmory University Microchemical Core Facility. Recently, we deter-mined that H-2Db monomers refolded with LT359-368C7Abu with
greater efficiency than did monomers with the LT360-368C6Abu pep-tide (AVKNY[Abu]SKL) (data not shown), which was previously usedto construct tetramers (29). At peptide concentrations �1 �M, LT359-368C7Abu stimulated IFN-� production by PyV-specific CD8� T cellsfrom the spleens of infected mice equivalent to that from the LT360-368C6Abu peptide (data not shown). In addition, RMA/S class I MHCpeptide stabilization assays revealed that LT359-368C7Abu boundH-2Db with �10-fold greater affinity than did LT360-368C6Abu(data not shown). These data confirm that an asparagine residue atposition 5 optimizes peptide binding to Db, as previously described forDb-restricted LCMV CD8� T cell epitopes (32). For simplicity, werefer to this population of Ag-specific T cells as LT359-specific CD8�
T cells. Db LT359C7Abu and Dk MT389 tetramers were generated asdescribed (29).
Flow cytometry
Lymphocyte populations were isolated from the blood, spleen, and lungs aspreviously described (29). Cells were stained in PBS containing 2% FBSand 0.1% sodium azide (FACS buffer) for 45 min at 4°C or room temper-ature, followed by two washes in FACS buffer. Samples were immediatelyacquired on a FACSCalibur (BD Biosciences) or were fixed in PBS con-taining 1% paraformaldehyde overnight. Cells were stained with allophy-cocyanin-conjugated tetramers. CD8�, CD45.1, CD122, NKG2A/C/E(clone 20d5), and Armenian hamster isotype control Abs were purchasedfrom BD Pharmingen. CD27, CD28, CD122, NKG2A (clone 16a11), andGolden Syrian hamster isotype control Abs were purchased from eBio-science. CD11a and L-selectin (CD62L) Abs were purchased from CaltagLaboratories. Data were analyzed using CellQuest software (BDBiosciences).
Intracellular IFN-� staining
Cells were stimulated directly ex vivo with 10 �M synthetic peptides andstained for CD8� and IFN-� as described elsewhere (29). The absolutenumber of Ag-specific IFN-��CD8� T cells was determined by subtract-ing unstimulated IFN-��CD8� cells (no peptide) from peptide-stimulatedIFN-��CD8� cells.
CD28 and CD40L costimulation blockade and bone marrowchimeras
The costimulation blockade protocol was performed as previously de-scribed (33). For acute CD28 and CD40L costimulation blockade, micereceived 500 �g of hamster anti-mouse CD40L Ab (clone MR1; Bio Ex-press) and human CTLA4-Ig (Bristol-Myers Squibb), separately or com-bined, i.p. on days �2, 0, and �2 relative to the day of PyV infection. Thegeneration of persistently infected or naive CD45 congenic bone marrowchimeras has been previously described (29). Persistently infected, busul-fan-treated B6 (CD45.2�) mice received bone marrow from naive B6.SJL(CD45.1�) mice, with or without 500 �g of MR1 Ab and CTLA4-Ig ondays 0, 2, and 4 post-transplant and weekly thereafter for a 7-wk duration.Naive busulfan-treated B6 (CD45.2�) mice given bone marrow from naiveB6.SJL (CD45.1�) mice were infected by PyV 48 days later with or with-out 500 �g of MR1 Ab and CTLA4-Ig on days �2, 0, and �2 of infection.
CD70 blockade
For blockade during acute infection, mice received 500 �g of rat anti-mouse CD70 (clone FR70; Bio Express) � 500 �g of CTLA4-Ig (or 500�g of ChromPure rat IgG (Jackson ImmunoResearch) alone) i.p. 2 daysbefore PyV infection and 250 �g of FR70 � 500 �g CTLA4-Ig (or 250 �gof rat IgG alone) on days 0 and �2 of infection. For blockade duringpersistent infection, mice received 250 �g of FR70 � 500 �g of CTLA4-Ig(or 250 �g of rat IgG alone) i.p. on days 40, 42, 44, 50, 57, 64, and 71postinfection (p.i.).
Taqman real-time PCR
DNA isolation and Taqman PCR were performed as described (29). ThePyV DNA quantity is expressed in genome copies per milligram of tissueor per milliliter of blood and is calculated based on a standard curve ofknown PyV genome copy number vs threshold cycle of detection. Thedetection limit with this assay is 10 copies of genomic viral DNA.
ELISAs
PyV major capsid protein VP1-specific ELISAs were performed in 96-wellmicrotiter plates coated with purified recombinant VP1 protein (50 ng/wellin carbonate buffer) that was produced in Escherichia coli (34) and waskindly provided by R. L. Garcea (University of Colorado Health Sciences
1815The Journal of Immunology
by guest on February 9, 2022http://w
ww
.jimm
unol.org/D
ownloaded from

Center, Denver, CO). The serum samples were serially diluted (2-fold) andtested in duplicates. Biotinylated goat anti-mouse IgG and streptavidin-HRP were used (Vector Laboratories) to detect IgG with 3,3� 5,5�-tetra-methyl-benzidine tablet substrates (Sigma-Aldrich). The plates were readat 450 nm by a THERMOMAX plate reader and Softmax software (Molec-ular Devices). Two-fold serial dilutions of a positive control serum sampleharvested from PyV-infected C57BL/6 mice on day 21 p.i. were used toobtain a standard reference curve on each 96-well plate, and the VP1-specific IgG concentrations of the test samples were expressed in arbitraryunits based on comparison with this standard curve.
Statistics
Statistical significance was determined by the unpaired Student’s t test,assuming unequal variances. A p value of �0.05 was considered statisti-cally significant.
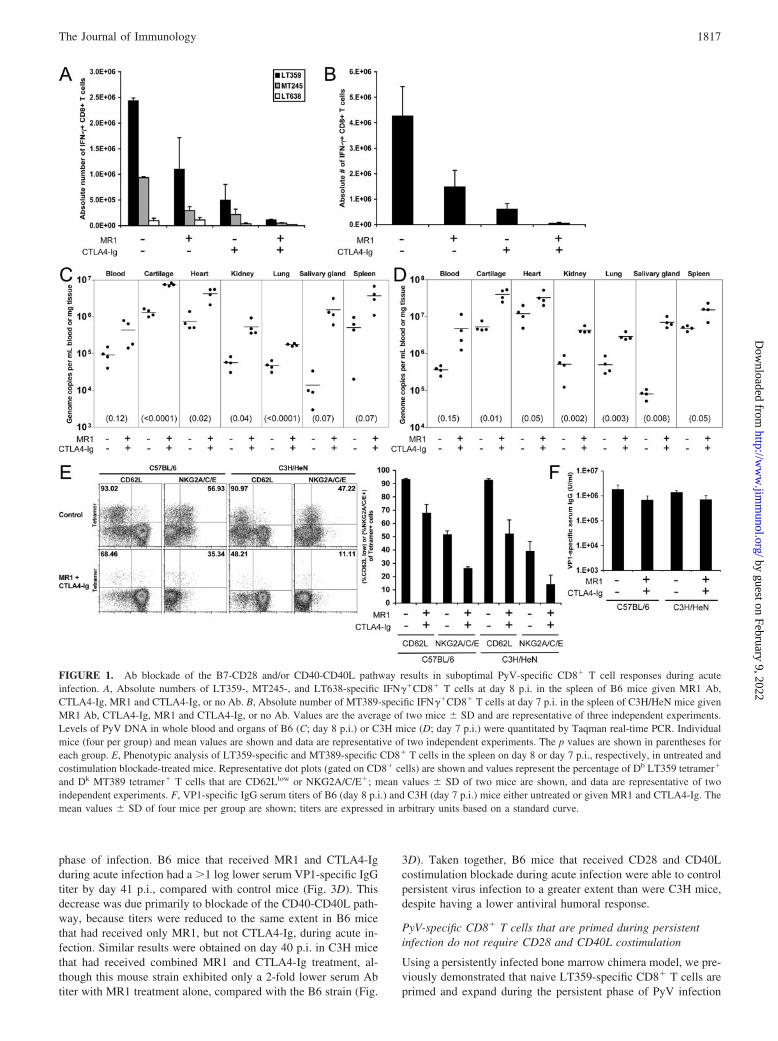
ResultsCD28 and CD40L costimulation blockade diminishes acute PyV-specific cellular and humoral responses and is associated withelevated viral levels
C57BL/6 (B6) mice, but not C3H mice, have been reported togenerate allospecific T cells in the absence of both B7-CD28 andCD40-CD40L costimulation (33). Therefore, we asked whetherthese mouse strains differ in the PyV-specific immune responsesafter blockade of each or both of these costimulation pathways. Wepreviously showed that B6 mice mount a potent CD8� T cell re-sponse that peaks at day 8 p.i., with a viral epitope-specific dom-inance hierarchy of LT359�MT245�LT638 (29). In C3H mice,PyV-specific CD8� T cells are predominantly focused on a singleepitope, MT389, and peak in magnitude on day 7–8 p.i (28, 35).Ab-mediated costimulation blockade of either the B7-CD28 orCD40-CD40L pathways substantially dampened the magnitude ofthe acute PyV-specific response in both B6 and C3H mousestrains, resulting in a decrease in the absolute number of splenic Db
LT359- and Dk MT389-specific CD8� T cells, respectively, asdetected by intracellular IFN-� production (Fig. 1, A and B). Sim-ilar results were obtained by enumerating Db LT359 or Dk MT389tetramer�CD8� T cells in the spleen directly ex vivo (data notshown). In B6 mice, the subdominant MT245- and LT638-specificCD8� T cell responses were likewise reduced (Fig. 1A). Simulta-neous blockade of both pathways almost entirely eliminated theacute PyV-specific CD8� T cell response and was associated witha �1 log increase in PyV genomes in the blood and multiple or-gans, including the cartilage, heart, kidney, lungs, salivary gland,and spleen (Fig. 1, A–D). Blockade of either the B7-CD28 orCD40-CD40L pathway individually led to only modest increase inviral DNA levels (data not shown), presumably because sufficientnumbers of antiviral CD8� T cells were elicited to control PyVinfection.
We next asked whether the PyV-specific CD8� T cells primedin the face of costimulation blockade were phenotypically altered.Of the small population of tetramer�CD8� T cells primed duringacute infection in the presence of CD28 and CD40L blockade,there was a substantially reduced percentage of CD62Llow cells(compared with controls) in both B6 and C3H mice (Fig. 1E). Inaddition, a smaller percentage of tetramer�CD8� T cells ex-pressed the NK cell inhibitory receptor CD94/NKG2A (Fig. 1E),which is up-regulated on PyV-specific CD8� T cells during thecourse of infection (36). Despite these phenotypic differences,PyV-specific CD8� T cells primed in the presence or absence ofCD28 and CD40L costimulation blockade were uniformlyCD11ahigh (data not shown). Blockade of either the B7-CD28 orCD40-CD40L pathway individually did not lead to phenotypic dif-ferences of antiviral CD8� T cells (data not shown). These datasuggest that combined CD28 and CD40L costimulation blockadeimpeded differentiation of PyV-specific CD8� T cells.
Unlike many viruses that require T cell help to generate protec-tive, isotype-switched antiviral Ab responses, PyV infection elicitsa virus-neutralizing, isotype-switched, T-cell-independent type 2Ab response (37, 38). A reduced PyV-specific Ab response againstthe major PyV capsid protein, VP1, could account for the elevatedPyV DNA levels in mice that received costimulation blockade. Asshown in Fig. 1F, VP1-specific serum IgG levels were reduced2-fold in B6 and C3H mice that received costimulation blockadecompared with wild-type mice at day 8 p.i., similar to that reportedat day 14 p.i. in B6 mice given CD40L Ab (31).
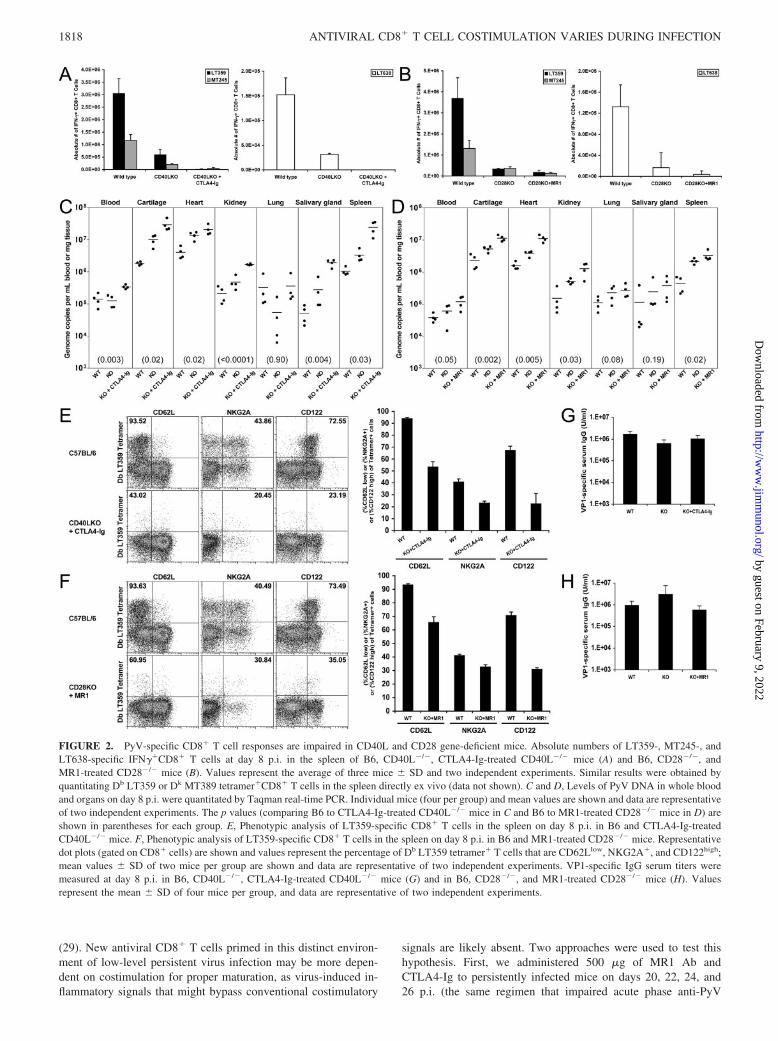
Acute PyV-specific CD8� T cell and humoral responses inCD40L�/� and CD28�/� mice
We sought to confirm the above results using homozygous CD28and CD40L gene knockout mice. When compared with wild-typeB6 mice, both CD28�/� and CD40L�/� mice generated reducedacute dominant and subdominant PyV-specific CD8� T cell re-sponses, which were further diminished with Ab blockade of thealternate pathway (Fig. 2, A and B). In CD28�/� mice, the LT359-specific response was reduced to a greater extent than was theMT245-specific response, suggesting that the dominant populationwas more dependent on CD28 signaling than a subdominant pop-ulation (Fig. 2B). This reduction in the magnitude of the CD8� Tcell response was associated with significantly higher levels ofPyV DNA in the blood and in nearly all organs examined (Fig. 2,C and D). Furthermore, in CD40L�/� mice that receivedCTLA4-Ig and in CD28�/� mice that received MR1 Ab, there wasa substantial reduction in the percentage of tetramer�CD8� T cellsthat were CD62Llow, NKG2A�, and CD122high, compared withwild-type mice (Fig. 2, E and F); LT359-specific CD8� T cellsfrom all groups of mice were uniformly CD11ahigh (data notshown). In agreement with previous data (Fig. 1F), VP1-specificserum IgG titers were modestly reduced in CD40L�/� mice (3-fold, compared with wild-type; Fig. 2G). CD28KO mice mountedan equivalent VP1-specific response to wild-type mice, which wasreduced 2-fold if CD40-CD40L interactions were blocked withMR1 Ab (Fig. 2H).
Acute CD28 and CD40L costimulation blockade dampens PyV-specific cellular and humoral responses during the persistentphase of infection
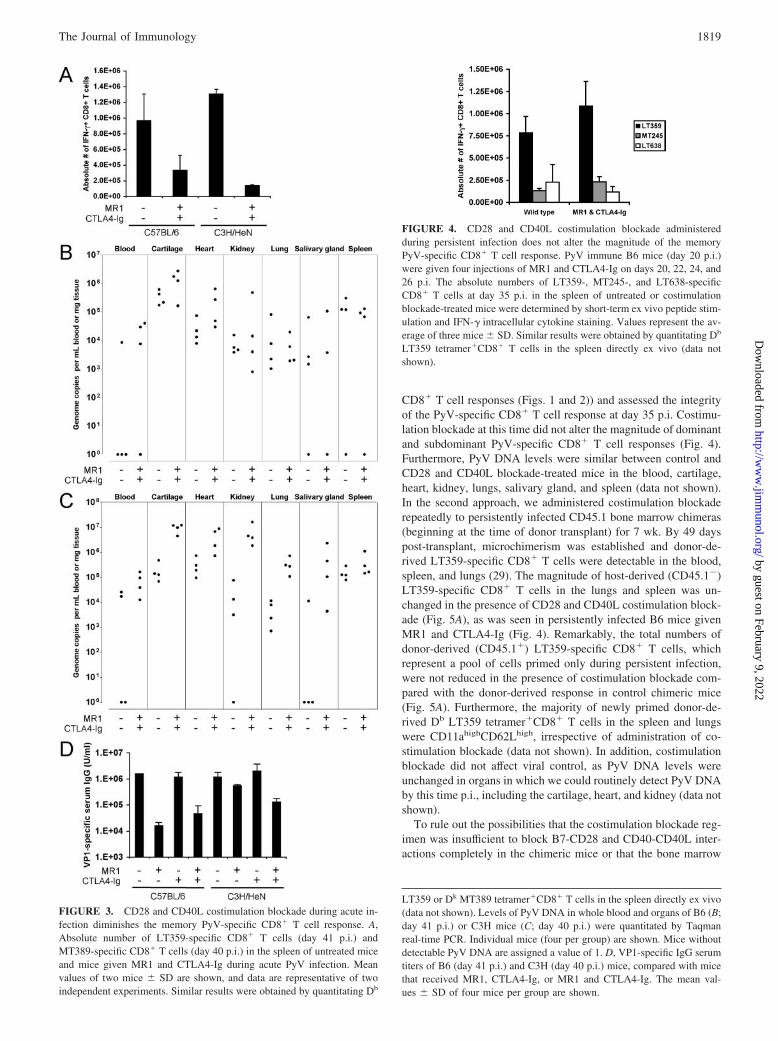
Given the profound effect of CD28 and CD40L costimulationblockade on the acute PyV-specific CD8� T cell response, we nextexamined the long-term consequences of costimulation blockade(administered only during acute infection) on the magnitude andfunction of the PyV-specific CD8� T cell response. The magnitudeof LT359-specific (B6 mice) and MT389-specific (C3H) CD8� Tcell responses in persistently infected mice were each reduced afteracute CD28 and CD40L costimulation blockade but were de-pressed to a greater extent in C3H mice (Fig. 3A). PyV DNA levelswere not significantly different between control B6 and costimu-lation blockade groups by 41 days p.i. in multiple organs, althougha greater proportion of costimulation blockade-treated mice dis-played viremia (Fig. 3B). In contrast, C3H mice that had receivedacute costimulation blockade remained viremic by 40 days p.i. andaveraged �1 log increase in viral DNA in the cartilage, heart,kidney, lungs, and salivary gland (with a significant increase ( p 0.01) observed in the cartilage), compared with control mice (Fig.3C). In summary, acute CD28 and CD40L costimulation blockadedampened the PyV-specific CD8� T cell response long-term, withcontrol of viral persistence after acute costimulation blockadevarying between strains of mice and among organs.
Finally, we asked whether acute costimulation blockade alsodampened the humoral VP1-specific response during the persistent
1816 ANTIVIRAL CD8� T CELL COSTIMULATION VARIES DURING INFECTION
by guest on February 9, 2022http://w
ww
.jimm
unol.org/D
ownloaded from
phase of infection. B6 mice that received MR1 and CTLA4-Igduring acute infection had a �1 log lower serum VP1-specific IgGtiter by day 41 p.i., compared with control mice (Fig. 3D). Thisdecrease was due primarily to blockade of the CD40-CD40L path-way, because titers were reduced to the same extent in B6 micethat had received only MR1, but not CTLA4-Ig, during acute in-fection. Similar results were obtained on day 40 p.i. in C3H micethat had received combined MR1 and CTLA4-Ig treatment, al-though this mouse strain exhibited only a 2-fold lower serum Abtiter with MR1 treatment alone, compared with the B6 strain (Fig.
3D). Taken together, B6 mice that received CD28 and CD40Lcostimulation blockade during acute infection were able to controlpersistent virus infection to a greater extent than were C3H mice,despite having a lower antiviral humoral response.
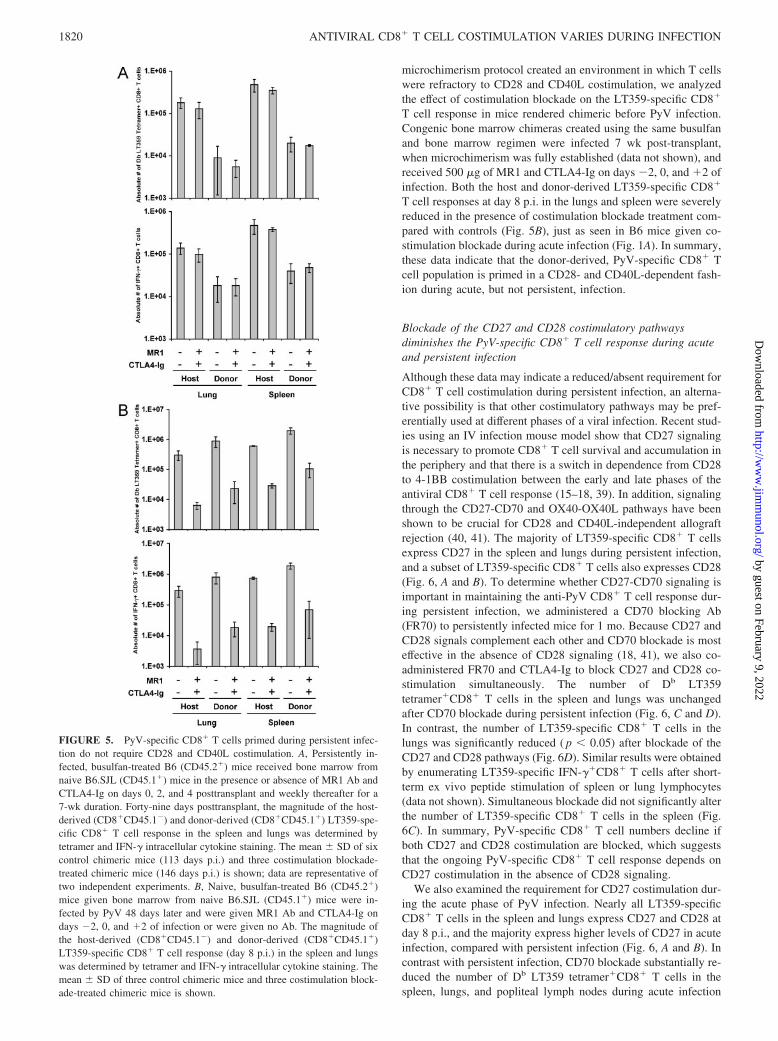
PyV-specific CD8� T cells that are primed during persistentinfection do not require CD28 and CD40L costimulation
Using a persistently infected bone marrow chimera model, we pre-viously demonstrated that naive LT359-specific CD8� T cells areprimed and expand during the persistent phase of PyV infection
FIGURE 1. Ab blockade of the B7-CD28 and/or CD40-CD40L pathway results in suboptimal PyV-specific CD8� T cell responses during acuteinfection. A, Absolute numbers of LT359-, MT245-, and LT638-specific IFN��CD8� T cells at day 8 p.i. in the spleen of B6 mice given MR1 Ab,CTLA4-Ig, MR1 and CTLA4-Ig, or no Ab. B, Absolute number of MT389-specific IFN��CD8� T cells at day 7 p.i. in the spleen of C3H/HeN mice givenMR1 Ab, CTLA4-Ig, MR1 and CTLA4-Ig, or no Ab. Values are the average of two mice � SD and are representative of three independent experiments.Levels of PyV DNA in whole blood and organs of B6 (C; day 8 p.i.) or C3H mice (D; day 7 p.i.) were quantitated by Taqman real-time PCR. Individualmice (four per group) and mean values are shown and data are representative of two independent experiments. The p values are shown in parentheses foreach group. E, Phenotypic analysis of LT359-specific and MT389-specific CD8� T cells in the spleen on day 8 or day 7 p.i., respectively, in untreated andcostimulation blockade-treated mice. Representative dot plots (gated on CD8� cells) are shown and values represent the percentage of Db LT359 tetramer�
and Dk MT389 tetramer� T cells that are CD62Llow or NKG2A/C/E�; mean values � SD of two mice are shown, and data are representative of twoindependent experiments. F, VP1-specific IgG serum titers of B6 (day 8 p.i.) and C3H (day 7 p.i.) mice either untreated or given MR1 and CTLA4-Ig. Themean values � SD of four mice per group are shown; titers are expressed in arbitrary units based on a standard curve.
1817The Journal of Immunology
by guest on February 9, 2022http://w
ww
.jimm
unol.org/D
ownloaded from
(29). New antiviral CD8� T cells primed in this distinct environ-ment of low-level persistent virus infection may be more depen-dent on costimulation for proper maturation, as virus-induced in-flammatory signals that might bypass conventional costimulatory
signals are likely absent. Two approaches were used to test thishypothesis. First, we administered 500 �g of MR1 Ab andCTLA4-Ig to persistently infected mice on days 20, 22, 24, and26 p.i. (the same regimen that impaired acute phase anti-PyV
FIGURE 2. PyV-specific CD8� T cell responses are impaired in CD40L and CD28 gene-deficient mice. Absolute numbers of LT359-, MT245-, andLT638-specific IFN��CD8� T cells at day 8 p.i. in the spleen of B6, CD40L�/�, CTLA4-Ig-treated CD40L�/� mice (A) and B6, CD28�/�, andMR1-treated CD28�/� mice (B). Values represent the average of three mice � SD and two independent experiments. Similar results were obtained byquantitating Db LT359 or Dk MT389 tetramer�CD8� T cells in the spleen directly ex vivo (data not shown). C and D, Levels of PyV DNA in whole bloodand organs on day 8 p.i. were quantitated by Taqman real-time PCR. Individual mice (four per group) and mean values are shown and data are representativeof two independent experiments. The p values (comparing B6 to CTLA4-Ig-treated CD40L�/� mice in C and B6 to MR1-treated CD28�/� mice in D) areshown in parentheses for each group. E, Phenotypic analysis of LT359-specific CD8� T cells in the spleen on day 8 p.i. in B6 and CTLA4-Ig-treatedCD40L�/� mice. F, Phenotypic analysis of LT359-specific CD8� T cells in the spleen on day 8 p.i. in B6 and MR1-treated CD28�/� mice. Representativedot plots (gated on CD8� cells) are shown and values represent the percentage of Db LT359 tetramer� T cells that are CD62Llow, NKG2A�, and CD122high;mean values � SD of two mice per group are shown and data are representative of two independent experiments. VP1-specific IgG serum titers weremeasured at day 8 p.i. in B6, CD40L�/�, CTLA4-Ig-treated CD40L�/� mice (G) and in B6, CD28�/�, and MR1-treated CD28�/� mice (H). Valuesrepresent the mean � SD of four mice per group, and data are representative of two independent experiments.
1818 ANTIVIRAL CD8� T CELL COSTIMULATION VARIES DURING INFECTION
by guest on February 9, 2022http://w
ww
.jimm
unol.org/D
ownloaded from
CD8� T cell responses (Figs. 1 and 2)) and assessed the integrityof the PyV-specific CD8� T cell response at day 35 p.i. Costimu-lation blockade at this time did not alter the magnitude of dominantand subdominant PyV-specific CD8� T cell responses (Fig. 4).Furthermore, PyV DNA levels were similar between control andCD28 and CD40L blockade-treated mice in the blood, cartilage,heart, kidney, lungs, salivary gland, and spleen (data not shown).In the second approach, we administered costimulation blockaderepeatedly to persistently infected CD45.1 bone marrow chimeras(beginning at the time of donor transplant) for 7 wk. By 49 dayspost-transplant, microchimerism was established and donor-de-rived LT359-specific CD8� T cells were detectable in the blood,spleen, and lungs (29). The magnitude of host-derived (CD45.1�)LT359-specific CD8� T cells in the lungs and spleen was un-changed in the presence of CD28 and CD40L costimulation block-ade (Fig. 5A), as was seen in persistently infected B6 mice givenMR1 and CTLA4-Ig (Fig. 4). Remarkably, the total numbers ofdonor-derived (CD45.1�) LT359-specific CD8� T cells, whichrepresent a pool of cells primed only during persistent infection,were not reduced in the presence of costimulation blockade com-pared with the donor-derived response in control chimeric mice(Fig. 5A). Furthermore, the majority of newly primed donor-de-rived Db LT359 tetramer�CD8� T cells in the spleen and lungswere CD11ahighCD62Lhigh, irrespective of administration of co-stimulation blockade (data not shown). In addition, costimulationblockade did not affect viral control, as PyV DNA levels wereunchanged in organs in which we could routinely detect PyV DNAby this time p.i., including the cartilage, heart, and kidney (data notshown).
To rule out the possibilities that the costimulation blockade reg-imen was insufficient to block B7-CD28 and CD40-CD40L inter-actions completely in the chimeric mice or that the bone marrow
FIGURE 4. CD28 and CD40L costimulation blockade administeredduring persistent infection does not alter the magnitude of the memoryPyV-specific CD8� T cell response. PyV immune B6 mice (day 20 p.i.)were given four injections of MR1 and CTLA4-Ig on days 20, 22, 24, and26 p.i. The absolute numbers of LT359-, MT245-, and LT638-specificCD8� T cells at day 35 p.i. in the spleen of untreated or costimulationblockade-treated mice were determined by short-term ex vivo peptide stim-ulation and IFN-� intracellular cytokine staining. Values represent the av-erage of three mice � SD. Similar results were obtained by quantitating Db
LT359 tetramer�CD8� T cells in the spleen directly ex vivo (data notshown).
LT359 or Dk MT389 tetramer�CD8� T cells in the spleen directly ex vivo(data not shown). Levels of PyV DNA in whole blood and organs of B6 (B;day 41 p.i.) or C3H mice (C; day 40 p.i.) were quantitated by Taqmanreal-time PCR. Individual mice (four per group) are shown. Mice withoutdetectable PyV DNA are assigned a value of 1. D, VP1-specific IgG serumtiters of B6 (day 41 p.i.) and C3H (day 40 p.i.) mice, compared with micethat received MR1, CTLA4-Ig, or MR1 and CTLA4-Ig. The mean val-ues � SD of four mice per group are shown.
FIGURE 3. CD28 and CD40L costimulation blockade during acute in-fection diminishes the memory PyV-specific CD8� T cell response. A,Absolute number of LT359-specific CD8� T cells (day 41 p.i.) andMT389-specific CD8� T cells (day 40 p.i.) in the spleen of untreated miceand mice given MR1 and CTLA4-Ig during acute PyV infection. Meanvalues of two mice � SD are shown, and data are representative of twoindependent experiments. Similar results were obtained by quantitating Db
1819The Journal of Immunology
by guest on February 9, 2022http://w
ww
.jimm
unol.org/D
ownloaded from
microchimerism protocol created an environment in which T cellswere refractory to CD28 and CD40L costimulation, we analyzedthe effect of costimulation blockade on the LT359-specific CD8�
T cell response in mice rendered chimeric before PyV infection.Congenic bone marrow chimeras created using the same busulfanand bone marrow regimen were infected 7 wk post-transplant,when microchimerism was fully established (data not shown), andreceived 500 �g of MR1 and CTLA4-Ig on days �2, 0, and �2 ofinfection. Both the host and donor-derived LT359-specific CD8�
T cell responses at day 8 p.i. in the lungs and spleen were severelyreduced in the presence of costimulation blockade treatment com-pared with controls (Fig. 5B), just as seen in B6 mice given co-stimulation blockade during acute infection (Fig. 1A). In summary,these data indicate that the donor-derived, PyV-specific CD8� Tcell population is primed in a CD28- and CD40L-dependent fash-ion during acute, but not persistent, infection.
Blockade of the CD27 and CD28 costimulatory pathwaysdiminishes the PyV-specific CD8� T cell response during acuteand persistent infection
Although these data may indicate a reduced/absent requirement forCD8� T cell costimulation during persistent infection, an alterna-tive possibility is that other costimulatory pathways may be pref-erentially used at different phases of a viral infection. Recent stud-ies using an IV infection mouse model show that CD27 signalingis necessary to promote CD8� T cell survival and accumulation inthe periphery and that there is a switch in dependence from CD28to 4-1BB costimulation between the early and late phases of theantiviral CD8� T cell response (15–18, 39). In addition, signalingthrough the CD27-CD70 and OX40-OX40L pathways have beenshown to be crucial for CD28 and CD40L-independent allograftrejection (40, 41). The majority of LT359-specific CD8� T cellsexpress CD27 in the spleen and lungs during persistent infection,and a subset of LT359-specific CD8� T cells also expresses CD28(Fig. 6, A and B). To determine whether CD27-CD70 signaling isimportant in maintaining the anti-PyV CD8� T cell response dur-ing persistent infection, we administered a CD70 blocking Ab(FR70) to persistently infected mice for 1 mo. Because CD27 andCD28 signals complement each other and CD70 blockade is mosteffective in the absence of CD28 signaling (18, 41), we also co-administered FR70 and CTLA4-Ig to block CD27 and CD28 co-stimulation simultaneously. The number of Db LT359tetramer�CD8� T cells in the spleen and lungs was unchangedafter CD70 blockade during persistent infection (Fig. 6, C and D).In contrast, the number of LT359-specific CD8� T cells in thelungs was significantly reduced ( p � 0.05) after blockade of theCD27 and CD28 pathways (Fig. 6D). Similar results were obtainedby enumerating LT359-specific IFN-��CD8� T cells after short-term ex vivo peptide stimulation of spleen or lung lymphocytes(data not shown). Simultaneous blockade did not significantly alterthe number of LT359-specific CD8� T cells in the spleen (Fig.6C). In summary, PyV-specific CD8� T cell numbers decline ifboth CD27 and CD28 costimulation are blocked, which suggeststhat the ongoing PyV-specific CD8� T cell response depends onCD27 costimulation in the absence of CD28 signaling.
We also examined the requirement for CD27 costimulation dur-ing the acute phase of PyV infection. Nearly all LT359-specificCD8� T cells in the spleen and lungs express CD27 and CD28 atday 8 p.i., and the majority express higher levels of CD27 in acuteinfection, compared with persistent infection (Fig. 6, A and B). Incontrast with persistent infection, CD70 blockade substantially re-duced the number of Db LT359 tetramer�CD8� T cells in thespleen, lungs, and popliteal lymph nodes during acute infection
FIGURE 5. PyV-specific CD8� T cells primed during persistent infec-tion do not require CD28 and CD40L costimulation. A, Persistently in-fected, busulfan-treated B6 (CD45.2�) mice received bone marrow fromnaive B6.SJL (CD45.1�) mice in the presence or absence of MR1 Ab andCTLA4-Ig on days 0, 2, and 4 posttransplant and weekly thereafter for a7-wk duration. Forty-nine days posttransplant, the magnitude of the host-derived (CD8�CD45.1�) and donor-derived (CD8�CD45.1�) LT359-spe-cific CD8� T cell response in the spleen and lungs was determined bytetramer and IFN-� intracellular cytokine staining. The mean � SD of sixcontrol chimeric mice (113 days p.i.) and three costimulation blockade-treated chimeric mice (146 days p.i.) is shown; data are representative oftwo independent experiments. B, Naive, busulfan-treated B6 (CD45.2�)mice given bone marrow from naive B6.SJL (CD45.1�) mice were in-fected by PyV 48 days later and were given MR1 Ab and CTLA4-Ig ondays �2, 0, and �2 of infection or were given no Ab. The magnitude ofthe host-derived (CD8�CD45.1�) and donor-derived (CD8�CD45.1�)LT359-specific CD8� T cell response (day 8 p.i.) in the spleen and lungswas determined by tetramer and IFN-� intracellular cytokine staining. Themean � SD of three control chimeric mice and three costimulation block-ade-treated chimeric mice is shown.
1820 ANTIVIRAL CD8� T CELL COSTIMULATION VARIES DURING INFECTION
by guest on February 9, 2022http://w
ww
.jimm
unol.org/D
ownloaded from
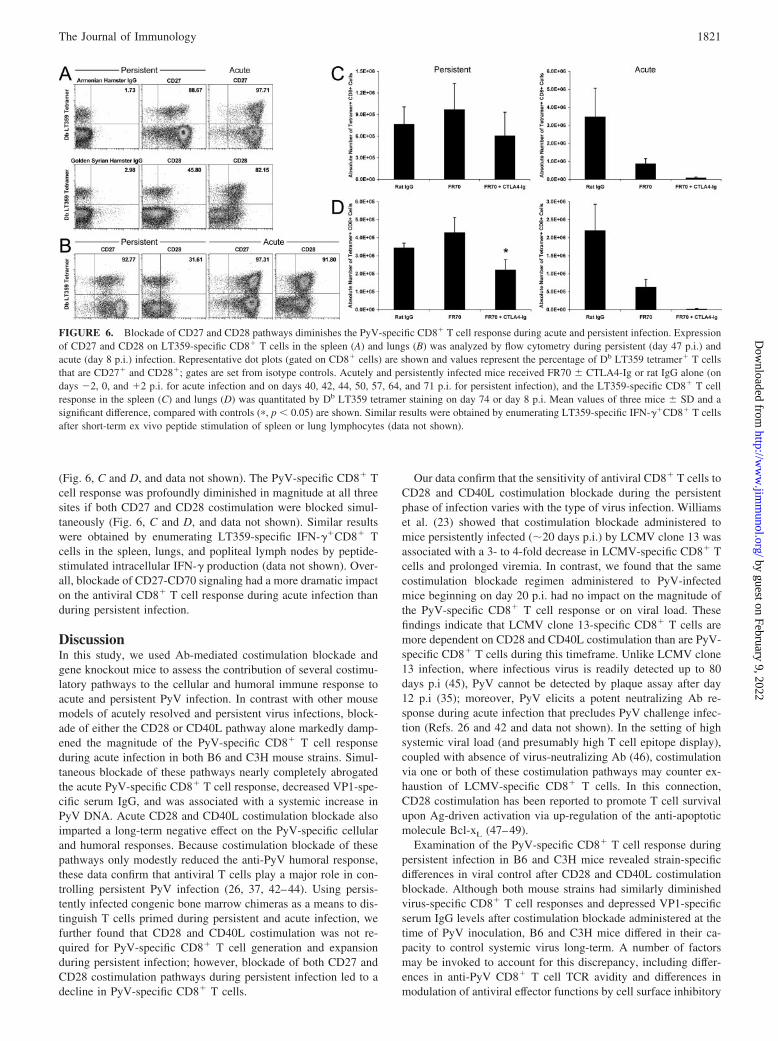
(Fig. 6, C and D, and data not shown). The PyV-specific CD8� Tcell response was profoundly diminished in magnitude at all threesites if both CD27 and CD28 costimulation were blocked simul-taneously (Fig. 6, C and D, and data not shown). Similar resultswere obtained by enumerating LT359-specific IFN-��CD8� Tcells in the spleen, lungs, and popliteal lymph nodes by peptide-stimulated intracellular IFN-� production (data not shown). Over-all, blockade of CD27-CD70 signaling had a more dramatic impacton the antiviral CD8� T cell response during acute infection thanduring persistent infection.
DiscussionIn this study, we used Ab-mediated costimulation blockade andgene knockout mice to assess the contribution of several costimu-latory pathways to the cellular and humoral immune response toacute and persistent PyV infection. In contrast with other mousemodels of acutely resolved and persistent virus infections, block-ade of either the CD28 or CD40L pathway alone markedly damp-ened the magnitude of the PyV-specific CD8� T cell responseduring acute infection in both B6 and C3H mouse strains. Simul-taneous blockade of these pathways nearly completely abrogatedthe acute PyV-specific CD8� T cell response, decreased VP1-spe-cific serum IgG, and was associated with a systemic increase inPyV DNA. Acute CD28 and CD40L costimulation blockade alsoimparted a long-term negative effect on the PyV-specific cellularand humoral responses. Because costimulation blockade of thesepathways only modestly reduced the anti-PyV humoral response,these data confirm that antiviral T cells play a major role in con-trolling persistent PyV infection (26, 37, 42–44). Using persis-tently infected congenic bone marrow chimeras as a means to dis-tinguish T cells primed during persistent and acute infection, wefurther found that CD28 and CD40L costimulation was not re-quired for PyV-specific CD8� T cell generation and expansionduring persistent infection; however, blockade of both CD27 andCD28 costimulation pathways during persistent infection led to adecline in PyV-specific CD8� T cells.
Our data confirm that the sensitivity of antiviral CD8� T cells toCD28 and CD40L costimulation blockade during the persistentphase of infection varies with the type of virus infection. Williamset al. (23) showed that costimulation blockade administered tomice persistently infected (�20 days p.i.) by LCMV clone 13 wasassociated with a 3- to 4-fold decrease in LCMV-specific CD8� Tcells and prolonged viremia. In contrast, we found that the samecostimulation blockade regimen administered to PyV-infectedmice beginning on day 20 p.i. had no impact on the magnitude ofthe PyV-specific CD8� T cell response or on viral load. Thesefindings indicate that LCMV clone 13-specific CD8� T cells aremore dependent on CD28 and CD40L costimulation than are PyV-specific CD8� T cells during this timeframe. Unlike LCMV clone13 infection, where infectious virus is readily detected up to 80days p.i (45), PyV cannot be detected by plaque assay after day12 p.i (35); moreover, PyV elicits a potent neutralizing Ab re-sponse during acute infection that precludes PyV challenge infec-tion (Refs. 26 and 42 and data not shown). In the setting of highsystemic viral load (and presumably high T cell epitope display),coupled with absence of virus-neutralizing Ab (46), costimulationvia one or both of these costimulation pathways may counter ex-haustion of LCMV-specific CD8� T cells. In this connection,CD28 costimulation has been reported to promote T cell survivalupon Ag-driven activation via up-regulation of the anti-apoptoticmolecule Bcl-xL (47–49).
Examination of the PyV-specific CD8� T cell response duringpersistent infection in B6 and C3H mice revealed strain-specificdifferences in viral control after CD28 and CD40L costimulationblockade. Although both mouse strains had similarly diminishedvirus-specific CD8� T cell responses and depressed VP1-specificserum IgG levels after costimulation blockade administered at thetime of PyV inoculation, B6 and C3H mice differed in their ca-pacity to control systemic virus long-term. A number of factorsmay be invoked to account for this discrepancy, including differ-ences in anti-PyV CD8� T cell TCR avidity and differences inmodulation of antiviral effector functions by cell surface inhibitory
FIGURE 6. Blockade of CD27 and CD28 pathways diminishes the PyV-specific CD8� T cell response during acute and persistent infection. Expressionof CD27 and CD28 on LT359-specific CD8� T cells in the spleen (A) and lungs (B) was analyzed by flow cytometry during persistent (day 47 p.i.) andacute (day 8 p.i.) infection. Representative dot plots (gated on CD8� cells) are shown and values represent the percentage of Db LT359 tetramer� T cellsthat are CD27� and CD28�; gates are set from isotype controls. Acutely and persistently infected mice received FR70 � CTLA4-Ig or rat IgG alone (ondays �2, 0, and �2 p.i. for acute infection and on days 40, 42, 44, 50, 57, 64, and 71 p.i. for persistent infection), and the LT359-specific CD8� T cellresponse in the spleen (C) and lungs (D) was quantitated by Db LT359 tetramer staining on day 74 or day 8 p.i. Mean values of three mice � SD and asignificant difference, compared with controls (�, p � 0.05) are shown. Similar results were obtained by enumerating LT359-specific IFN-��CD8� T cellsafter short-term ex vivo peptide stimulation of spleen or lung lymphocytes (data not shown).
1821The Journal of Immunology
by guest on February 9, 2022http://w
ww
.jimm
unol.org/D
ownloaded from

receptors (50) or by regulatory T cells (51). Recent studies onCD8� T cell responses and viremia in resolved and chronichepatitis C virus and cytomegalovirus infection in humans havegiven rise to the concept that a broad multispecific CD8� T cellresponse, rather than one narrowly focused on a limited set ofepitopes, favors control of persistent viral infection (52–54).Such a multi-pronged CD8� T cell response would clearly limitemergence of variant epitope-loss viruses, a common immuneescape ploy used by highly mutable RNA viruses, such as hep-atitis C virus and HIV. Additionally, a multispecificity antiviralCD8� T cell response would facilitate clearance of infectedcells that vary in density of different epitopes (55) or expressviral proteins at different points in the viral lifecycle (56). In thePyV mouse model, extensive epitope mapping using overlap-ping synthetic peptides uncovered three CD8� T cell epitopesderived by the viral MT and LT proteins in B6 mice, but onlya strong dominant and a very weak subdominant epitope fromMT in C3H mice (Refs. 29 and 35 and our unpublished obser-vations). The importance of early control for persistently in-fecting viruses is highlighted by a recent study showing that�HV-68 latency is established during acute infection (57). Fi-nally, the superior control of persistent PyV infection in B6 thanin C3H mice may stem from more efficient priming of naivePyV-specific CD8� T cells during persistent infection in B6mice. Given the potent oncogenic potential of PyV and the as-sociation of high genome copy number with PyV-induced tu-mors (58), the higher systemic viral burden in C3H mice givencostimulation blockade during acute infection should predis-pose them to PyV tumorigenesis.
CD28 and CD40L costimulation blockade altered the surfacephenotype of PyV-specific CD8� T cells during acute infection,specifically by decreasing the proportion of LT359- andMT389-specific CD8� T cells that were CD62Llow, CD94/NKG2A�, and CD122high. These phenotypic changes may re-flect a priming environment that provides suboptimal signals forfull T cell activation and differentiation. This possibility is con-sistent with a report demonstrating that maintenance of CD94/NKG2A expression by CD8� T cells may require chronic TCRstimulation (59). Reduced T cell stimulation due to costimula-tion blockade could also translate into either a short-liveddown-regulation of surface CD62L or preferential differentia-tion into a CD62Lhigh subset (60 – 62); enhanced CD62L levelsmay, in turn, alter the trafficking patterns of costimulation-blocked anti-PyV CD8� T cells.
A central finding of this study is that the requirement forcostimulation differs between acute and persistent phases of in-fection. Within the priming microenvironment of persistent in-fection and CD28 and CD40L costimulation blockade, PyV-specific CD8� T cells receive sufficient signals for expansionand acquisition of effector function, despite lower levels of Agand virus-induced inflammation than in acute infection. En-gagement of other costimulatory molecules on PyV-specificCD8� T cells, such as CD27, could provide survival signalssufficient to maintain the antiviral CD8� T cell response in theabsence of CD28 and CD40L signaling. Blockade of eitherCD27 or CD28 costimulation alone during the persistent phaseof infection did not lead to a deterioration of the PyV-specificT cell response (Figs. 4 and 6). Interestingly, combined CD27and CD28 costimulation blockade during persistent infectionled to a significant decline in PyV-specific CD8� T cell num-bers in the lungs, but not in the spleen (Fig. 6, C and D). In thisconnection, CD27 supports IV-specific CD8� T cell survivaland accumulation in the lungs, particularly in the absence ofCD28 (17, 18). Taken together, these data suggest that the co-
stimulation requirements for PyV-specific CD8� T cells duringpersistent infection are less dependent on any single pathway;the ongoing antiviral T cell response wanes only after multiplecostimulatory signals are obstructed.
Multiple factors may contribute to the differential requirementfor CD27 and CD28 costimulation to maintain PyV-specific CD8�
T cells in the lungs vs spleen during persistent infection, includingAPC diversity among tissues and variability in their presentationof T cell epitopes, as well as tissue-specific dependence on par-ticular costimulatory signals. PyV DNA levels are 10- to 20-foldlower in the lungs than in the spleen during persistent infection(Fig. 3B), which could translate into reduced presentation of viralT cell epitopes by resident APCs in the lungs. It has been appre-ciated that acute viral replication in vivo sustains a high level ofAg display that heightens TCR signaling and reduces the require-ment for T cell costimulation (63, 64). With decreased TCR en-gagement by cognate Ag, PyV-specific CD8� T cells may requiremore costimulation to achieve sufficient TCR signal strength togenerate and/or maintain antiviral CD8� T cells in persistentlyinfected hosts.
The finding that PyV-specific CD8� T cells are primed in-dependently of CD28 and CD40L signaling during persistentinfection has clinical implications for development of costimu-lation blockade therapies to promote transplantation tolerance.Our observations address a continuing concern that CD28and/or CD40 blockade-based strategies, while promoting graftacceptance, may disrupt protective immunity to preexisting in-fections. Reactivation of BK virus, a human PyV, is a majorcause of renal allograft dysfunction and failure (65). PrimaryBK virus infection of renal allografts or reactivation of latentBK virus in the host may be due to the use of strong immuno-suppressants after transplantation (66). Tolerance induction viatargeting of the B7-CD28 and CD40-CD40L pathways is a po-tential alternative therapy, but the effects of costimulationblockade have not been completely characterized during per-sistent infection. Our findings suggest that kidney graft toler-ance induction via CD28 and CD40L costimulation blockademay be considered as a therapy that does not dampen the on-going and emerging anti-PyV-specific CD8� T cell response,thereby preventing viral resurgence and allograft failure.
AcknowledgmentsWe thank Dr. Robert L. Garcea for providing recombinant VP1 protein,and Annette Hadley, Thuvan Dinh, Tatyana Andreyeva, and Dale Staplerfor expert technical assistance.
DisclosuresThe authors have no financial conflict of interest.
References1. Whitmire, J. K., and R. Ahmed. 2000. Costimulation in antiviral immunity: dif-
ferential requirements for CD4� and CD8� T cell responses. Curr. Opin. Im-munol. 12: 448–455.
2. Borst, J., J. Hendriks, and Y. Xiao. 2005. CD27 and CD70 in T cell and B cellactivation. Curr. Opin. Immunol. 17: 275–281.
3. Suresh, M., J. K. Whitmire, L. E. Harrington, C. P. Larsen, T. C. Pearson,J. D. Altman, and R. Ahmed. 2001. Role of CD28–B7 interactions in generationand maintenance of CD8 T cell memory. J. Immunol. 167: 5565–5573.
4. Tan, J. T., J. K. Whitmire, R. Ahmed, T. C. Pearson, and C. P. Larsen. 1999.4-1BB ligand, a member of the TNF family, is important for the generation ofantiviral CD8 T cell responses. J. Immunol. 163: 4859–4868.
5. Shahinian, A., K. Pfeffer, K. P. Lee, T. M. Kundig, K. Kishihara,A. Wakeham, K. Kawai, P. S. Ohashi, C. B. Thompson, and T. W. Mak. 1993.Differential T cell costimulatory requirements in CD28-deficient mice. Sci-ence 261: 609 – 612.
6. Kundig, T. M., A. Shahinian, K. Kawai, H. W. Mittrucker, E. Sebzda,M. F. Bachmann, T. W. Mak, and P. S. Ohashi. 1996. Duration of TCR stimu-lation determines costimulatory requirement of T cells. Immunity 5: 41–52.
1822 ANTIVIRAL CD8� T CELL COSTIMULATION VARIES DURING INFECTION
by guest on February 9, 2022http://w
ww
.jimm
unol.org/D
ownloaded from

7. McAdam, A. J., E. A. Farkash, B. E. Gewurz, and A. H. Sharpe. 2000. B7costimulation is critical for antibody class switching and CD8� cytotoxic T-lymphocyte generation in the host response to vesicular stomatitis virus. J. Virol.74: 203–208.
8. Sigal, L. J., H. Reiser, and K. L. Rock. 1998. The role of B7-1 and B7-2 co-stimulation for the generation of CTL responses in vivo. J. Immunol. 161:2740–2745.
9. Lumsden, J. M., J. M. Roberts, N. L. Harris, R. J. Peach, and F. Ronchese. 2000.Differential requirement for CD80 and CD80/CD86-dependent costimulation inthe lung immune response to an influenza virus infection. J. Immunol. 164:79–85.
10. Borrow, P., A. Tishon, S. Lee, J. Xu, I. S. Grewal, M. B. Oldstone, andR. A. Flavell. 1996. CD40L-deficient mice show deficits in antiviral immunityand have an impaired memory CD8� CTL response. J. Exp. Med. 183:2129–2142.
11. Whitmire, J. K., M. K. Slifka, I. S. Grewal, R. A. Flavell, and R. Ahmed. 1996.CD40 ligand-deficient mice generate a normal primary cytotoxic T-lymphocyteresponse but a defective humoral response to a viral infection. J. Virol. 70:8375–8381.
12. Andreasen, S. O., J. E. Christensen, O. Marker, and A. R. Thomsen. 2000. Roleof CD40 ligand and CD28 in induction and maintenance of antiviral CD8� ef-fector T cell responses. J. Immunol. 164: 3689–3697.
13. Tan, J. T., J. K. Whitmire, K. Murali-Krishna, R. Ahmed, J. D. Altman,R. S. Mittler, A. Sette, T. C. Pearson, and C. P. Larsen. 2000. 4-1BB costimu-lation is required for protective anti-viral immunity after peptide vaccination.J. Immunol. 164: 2320–2325.
14. DeBenedette, M. A., T. Wen, M. F. Bachmann, P. S. Ohashi, B. H. Barber,K. L. Stocking, J. J. Peschon, and T. H. Watts. 1999. Analysis of 4-1BB ligand(4-1BBL)-deficient mice and of mice lacking both 4-1BBL and CD28 reveals arole for 4-1BBL in skin allograft rejection and in the cytotoxic T cell response toinfluenza virus. J. Immunol. 163: 4833–4841.
15. Bertram, E. M., P. Lau, and T. H. Watts. 2002. Temporal segregation of 4-1BBversus CD28-mediated costimulation: 4-1BB ligand influences T cell numberslate in the primary response and regulates the size of the T cell memory responsefollowing influenza infection. J. Immunol. 168: 3777–3785.
16. Bertram, E. M., W. Dawicki, B. Sedgmen, J. L. Bramson, D. H. Lynch, andT. H. Watts. 2004. A switch in costimulation from CD28 to 4-1BB during pri-mary versus secondary CD8 T cell response to influenza in vivo. J. Immunol. 172:981–988.
17. Hendriks, J., L. A. Gravestein, K. Tesselaar, R. A. van Lier, T. N. Schumacher,and J. Borst. 2000. CD27 is required for generation and long-term maintenanceof T cell immunity. Nat. Immunol. 1: 433–440.
18. Hendriks, J., Y. Xiao, and J. Borst. 2003. CD27 promotes survival of activated Tcells and complements CD28 in generation and establishment of the effector Tcell pool. J. Exp. Med. 198: 1369–1380.
19. Brooks, J. W., A. M. Hamilton-Easton, J. P. Christensen, R. D. Cardin,C. L. Hardy, and P. C. Doherty. 1999. Requirement for CD40 ligand, CD4� Tcells, and B cells in an infectious mononucleosis-like syndrome. J. Virol. 73:9650–9654.
20. Lee, B. J., S. K. Reiter, M. Anderson, and S. R. Sarawar. 2002. CD28�/� miceshow defects in cellular and humoral immunity but are able to control infectionwith murine gammaherpesvirus 68. J. Virol. 76: 3049–3053.
21. Sarawar, S. R., B. J. Lee, and F. Giannoni. 2004. Cytokines and costimulatorymolecules in the immune response to murine gammaherpesvirus-68. Viral Im-munol. 17: 3–11.
22. Whitmire, J. K., R. A. Flavell, I. S. Grewal, C. P. Larsen, T. C. Pearson, andR. Ahmed. 1999. CD40-CD40 ligand costimulation is required for generatingantiviral CD4 T cell responses but is dispensable for CD8 T cell responses.J. Immunol. 163: 3194–3201.
23. Williams, M. A., T. M. Onami, A. B. Adams, M. M. Durham, T. C. Pearson,R. Ahmed, and C. P. Larsen. 2002. Cutting edge: persistent viral infection pre-vents tolerance induction and escapes immune control following CD28/CD40blockade-based regimen. J. Immunol. 169: 5387–5391.
24. Ochsenbein, A. F., S. R. Riddell, M. Brown, L. Corey, G. M. Baerlocher,P. M. Lansdorp, and P. D. Greenberg. 2004. CD27 expression promotes long-term survival of functional effector-memory CD8� cytotoxic T lymphocytes inHIV-infected patients. J. Exp. Med. 200: 1407–1417.
25. Gamadia, L. E., E. M. van Leeuwen, E. B. Remmerswaal, S. L. Yong,S. Surachno, P. M. Wertheim-van Dillen, I. J. Ten Berge, and R. A. Van Lier.2004. The size and phenotype of virus-specific T cell populations is determinedby repetitive antigenic stimulation and environmental cytokines. J. Immunol. 172:6107–6114.
26. Drake, D. R., III, and A. E. Lukacher. 1998. �2-microglobulin knockout mice arehighly susceptible to polyoma virus tumorigenesis. Virology 252: 275–284.
27. Moser, J. M., J. D. Altman, and A. E. Lukacher. 2001. Antiviral CD8� T cellresponses in neonatal mice: susceptibility to polyoma virus-induced tumors isassociated with lack of cytotoxic function by viral antigen-specific T cells. J. Exp.Med. 193: 595–606.
28. Lukacher, A. E., and C. S. Wilson. 1998. Resistance to polyoma virus-inducedtumors correlates with CTL recognition of an immunodominant H-2Dk-restrictedepitope in the middle T protein. J. Immunol. 160: 1724–1734.
29. Kemball, C. C., E. D. Lee, V. Vezys, T. C. Pearson, C. P. Larsen, andA. E. Lukacher. 2005. Late priming and variability of epitope-specific CD8� Tcell responses during a persistent virus infection. J. Immunol. 174: 7950–7960.
30. Grammer, A. C., and P. E. Lipsky. 2000. CD40-mediated regulation of immuneresponses by TRAF-dependent and TRAF-independent signaling mechanisms.Adv. Immunol. 76: 61–178.
31. Szomolanyi-Tsuda, E., J. D. Brien, J. E. Dorgan, R. M. Welsh, and R. L. Garcea.2000. The role of CD40-CD154 interaction in antiviral T cell-independent IgGresponses. J. Immunol. 164: 5877–5882.
32. Gairin, J. E., H. Mazarguil, D. Hudrisier, and M. B. Oldstone. 1995. Optimallymphocytic choriomeningitis virus sequences restricted by H-2Db major histo-compatibility complex class I molecules and presented to cytotoxic T lympho-cytes. J. Virol. 69: 2297–2305.
33. Williams, M. A., J. Trambley, J. Ha, A. B. Adams, M. M. Durham, P. Rees,S. R. Cowan, T. C. Pearson, and C. P. Larsen. 2000. Genetic characterization ofstrain differences in the ability to mediate CD40/CD28-independent rejection ofskin allografts. J. Immunol. 165: 6849–6857.
34. Leavitt, A. D., T. M. Roberts, and R. L. Garcea. 1985. Polyoma virus majorcapsid protein, VP1: purification after high level expression in Escherichia coli.J. Biol. Chem. 260: 12803–12809.
35. Lukacher, A. E., J. M. Moser, A. Hadley, and J. D. Altman. 1999. Visualizationof polyoma virus-specific CD8� T cells in vivo during infection and tumor re-jection. J. Immunol. 163: 3369–3378.
36. Moser, J. M., J. Gibbs, P. E. Jensen, and A. E. Lukacher. 2002. CD94-NKG2A receptors regulate antiviral CD8� T cell responses. Nat. Immunol. 3:189 –195.
37. Szomolanyi-Tsuda, E., and R. M. Welsh. 1996. T cell-independent antibody-mediated clearance of polyoma virus in T cell-deficient mice. J. Exp. Med. 183:403–411.
38. Szomolanyi-Tsuda, E., J. D. Brien, J. E. Dorgan, R. L. Garcea,R. T. Woodland, and R. M. Welsh. 2001. Antiviral T-cell-independent type 2antibody responses induced in vivo in the absence of T and NK cells. Virology280: 160 –168.
39. Dawicki, W., and T. H. Watts. 2004. Expression and function of 4-1BB duringCD4 versus CD8 T cell responses in vivo. Eur. J. Immunol. 34: 743–751.
40. Demirci, G., F. Amanullah, R. Kewalaramani, H. Yagita, T. B. Strom,M. H. Sayegh, and X. C. Li. 2004. Critical role of OX40 in CD28 and CD154-independent rejection. J. Immunol. 172: 1691–1698.
41. Yamada, A., A. D. Salama, M. Sho, N. Najafian, T. Ito, J. P. Forman,R. Kewalramani, S. Sandner, H. Harada, M. R. Clarkson, et al. 2005. CD70signaling is critical for CD28-independent CD8� T cell-mediated alloimmuneresponses in vivo. J. Immunol. 174: 1357–1364.
42. Lukacher, A. E., R. Freund, J. P. Carroll, R. T. Bronson, and T. L. Benjamin.1993. PyvS: a dominantly acting gene in C3H/BiDa mice conferring susceptibil-ity to tumor induction by polyoma virus. Virology 196: 241–248.
43. Heidari, S., Z. Berke, A. Berglof, A. Woldegiorgis, C. I. Smith, andT. Dalianis. 1999. Susceptibility to polyoma virus tumorigenesis in X-linkedimmunodeficient (XID) and B-cell deficient (�MT) mice is not increased. InVivo 13: 439 – 444.
44. Berke, Z., and T. Dalianis. 2000. Studies on polyomavirus persistence and poly-omavirus-induced tumor development in relation to the immune system. Adv.Cancer Res. 79: 249–276.
45. Zajac, A. J., J. N. Blattman, K. Murali-Krishna, D. J. Sourdive, M. Suresh,J. D. Altman, and R. Ahmed. 1998. Viral immune evasion due to persistence ofactivated T cells without effector function. J. Exp. Med. 188: 2205–2213.
46. Battegay, M., D. Moskophidis, H. Waldner, M. A. Brundler, W. P. Fung-Leung,T. W. Mak, H. Hengartner, and R. M. Zinkernagel. 1993. Impairment and delayof neutralizing antiviral antibody responses by virus-specific cytotoxic T cells.J. Immunol. 151: 5408–5415.
47. Boise, L. H., A. J. Minn, P. J. Noel, C. H. June, M. A. Accavitti, T. Lindsten, andC. B. Thompson. 1995. CD28 costimulation can promote T cell survival byenhancing the expression of Bcl-xL. Immunity 3: 87–98.
48. Noel, P. J., L. H. Boise, J. M. Green, and C. B. Thompson. 1996. CD28 co-stimulation prevents cell death during primary T cell activation. J. Immunol. 157:636–642.
49. Radvanyi, L. G., Y. Shi, H. Vaziri, A. Sharma, R. Dhala, G. B. Mills, andR. G. Miller. 1996. CD28 costimulation inhibits TCR-induced apoptosis duringa primary T cell response. J. Immunol. 156: 1788–1798.
50. Byers, A. M., C. C. Kemball, N. P. Andrews, and A. E. Lukacher. 2003. Regu-lation of antiviral CD8� T cells by inhibitory natural killer cell receptors. Mi-crobes Infect. 5: 169–177.
51. Rouse, B. T., and S. Suvas. 2004. Regulatory cells and infectious agents: detentescordiale and contraire. J. Immunol. 173: 2211–2215.
52. Lauer, G. M., E. Barnes, M. Lucas, J. Timm, K. Ouchi, A. Y. Kim, C. L. Day,G. K. Robbins, D. R. Casson, M. Reiser, et al. 2004. High resolution analysis ofcellular immune responses in resolved and persistent hepatitis C virus infection.Gastroenterology 127: 924–936.
53. Sacre, K., G. Carcelain, N. Cassoux, A. M. Fillet, D. Costagliola, D. Vittecoq,D. Salmon, Z. Amoura, C. Katlama, and B. Autran. 2005. Repertoire, diver-sity, and differentiation of specific CD8 T cells are associated with immuneprotection against human cytomegalovirus disease. J. Exp. Med. 201:1999 –2010.
54. Posnett, D. N., M. E. Engelhorn, and A. N. Houghton. 2005. Antiviral T cellresponses: phalanx or multipronged attack? J. Exp. Med. 201: 1881–1884.
55. Crowe, S. R., S. J. Turner, S. C. Miller, A. D. Roberts, R. A. Rappolo,P. C. Doherty, K. H. Ely, and D. L. Woodland. 2003. Differential antigenpresentation regulates the changing patterns of CD8� T cell immunodomi-nance in primary and secondary influenza virus infections. J. Exp. Med. 198:399 – 410.
56. Liu, L., E. Flano, E. J. Usherwood, S. Surman, M. A. Blackman, andD. L. Woodland. 1999. Lytic cycle T cell epitopes are expressed in two distinctphases during MHV-68 infection. J. Immunol. 163: 868–874.
1823The Journal of Immunology
by guest on February 9, 2022http://w
ww
.jimm
unol.org/D
ownloaded from

57. Flano, E., Q. Jia, J. Moore, D. L. Woodland, R. Sun, and M. A. Blackman. 2005.Early establishment of �-herpesvirus latency: implications for immune control.J. Immunol. 174: 4972–4978.
58. Wirth, J. J., L. Chen, and M. M. Fluck. 2000. Systemic polyomavirus genomeincrease and dissemination of capsid-defective genomes in mammary gland tu-mor-bearing mice. J. Virol. 74: 6975–6983.
59. McMahon, C. W., A. J. Zajac, A. M. Jamieson, L. Corral, G. E. Hammer,R. Ahmed, and D. H. Raulet. 2002. Viral and bacterial infections induce expres-sion of multiple NK cell receptors in responding CD8� T cells. J. Immunol. 169:1444–1452.
60. Tewari, K., J. Sacha, X. Gao, and M. Suresh. 2004. Effect of chronic viral in-fection on epitope selection, cytokine production, and surface phenotype of CD8T cells and the role of IFN-� receptor in immune regulation. J. Immunol. 172:1491–1500.
61. Wherry, E. J., V. Teichgraber, T. C. Becker, D. Masopust, S. M. Kaech, R. Antia,U. H. von Andrian, and R. Ahmed. 2003. Lineage relationship and protectiveimmunity of memory CD8 T cell subsets. Nat. Immunol. 4: 225–234.
62. van Faassen, H., M. Saldanha, D. Gilbertson, R. Dudani, L. Krishnan, and S. Sad.2005. Reducing the stimulation of CD8� T cells during infection with intracel-lular bacteria promotes differentiation primarily into a central(CD62LhighCD44high) subset. J. Immunol. 174: 5341–5350.
63. Bachmann, M. F., R. M. Zinkernagel, and A. Oxenius. 1998. Immune responsesin the absence of costimulation: viruses know the trick. J. Immunol. 161:5791–5794.
64. Bertram, E. M., W. Dawicki, and T. H. Watts. 2004. Role of T cell costimulationin anti-viral immunity. Semin. Immunol. 16: 185–196.
65. Hirsch, H. H., W. Knowles, M. Dickenmann, J. Passweg, T. Klimkait,M. J. Mihatsch, and J. Steiger. 2002. Prospective study of polyomavirus type BKreplication and nephropathy in renal-transplant recipients. N. Engl. J. Med. 347:488–496.
66. Barri, Y. M., I. Ahmad, B. L. Ketel, G. W. Barone, P. D. Walker,S. M. Bonsib, and S. R. Abul-Ezz. 2001. Polyoma viral infection in renaltransplantation: the role of immunosuppressive therapy. Clin. Transplant. 15:240 –246.
1824 ANTIVIRAL CD8� T CELL COSTIMULATION VARIES DURING INFECTION
by guest on February 9, 2022http://w
ww
.jimm
unol.org/D
ownloaded from