pericentric inversion in natural populations of oligoryzomys nigripes (rodentia: sigmodontinae)
TRANSCRIPT

Pericentric inversion in natural populations ofOligoryzomys nigripes (Rodentia: Sigmodontinae)
Cibele R. Bonvicino, Paulo S. D’Andrea, and Pavel M. Borodin
Abstract: We analysed polymorphism for pericentric inversion in chromosome 3 ofOligoryzomys nigripes(Rodentia:Sigmodontinae) in several populations in Brazil and examined the meiotic behaviour of this chromosome in heterozy-gotes. We observed an orderly pairing of all chromosomes at pachytene in heterozygotes for the inverted chromosome3. No indication of meiotic arrest and germ-cell death was found. Electron microscopy of synaptonemal complexes andconventional meiotic analysis indicated strictly nonhomologous synapsis and crossing-over suppression in the invertedregion in the heterozygotes, which prevent the formation of unbalanced gametes. Thus, the pericentric inversion inchromosome 3 does not apparently result in any selective disadvantages in heterozygous carriers. In the majority of thepopulations studied, the frequencies of acrocentric homozygotes, metacentric homozygotes, and heterozygotes were inHardy–Weinberg equilibrium. However, in some populations, we detected an excess of heterozygotes and a deficiencyof acrocentric homozygotes.
Key words: chromosome rearrangements, inversion, meiosis,Oligoryzomys nigripes.
Résumé: Les auteurs ont examiné le polymorphisme pour une inversion péricentrique chez le chromosome 3 de l’Oli-goryzomys nigripes(Rodentia : Sigmodontinae) au sein de diverses populations brésiliennes de même que le comporte-ment méiotique de ce chromosome chez des hétérozygotes. Ils ont observé un appariement ordonné de tous leschromosomes au stade pachytène chez les hétérozygotes pour l’inversion du chromosome 3. Aucune indication d’arrêtméiotique ou de mort des cellules germinales n’a été relevée. Des analyses du complexe synaptonémique en micros-copie électronique ainsi que des analyses méiotiques conventionnelles ont montré un appariement non-homologue cou-plé à une suppression des enjambements au sein de la région inversée chez les hétérozygotes. Ceci prévient laformation de gamètes au bagage génétique déséquilibré. Ainsi, l’inversion péricentrique du chromosome 3 n’entraîneapparemment aucun désavantage sélectif chez les porteurs hétérozygotes. La fréquence des homozygotes acrocentriques,des homozygotes métacentriques et des hétérozygotes chez la majorité des populations étudiées respectait l’équilibre deHardy–Weinberg. Cependant, chez certaines populations, les auteurs ont détecté un excès d’hétérozygotes et un déficitd’homozygotes acrocentriques.
Mots clés: réarrangements chromosomiques, inversions, méiose,Oligoryzomys nigripes.
[Traduit par la Rédaction] Bonvicino et al. 796
Introduction
Pericentric inversions and Robertsonian translocationsplay an important role in the karyotypic evolution of mam-mals (for review see Searle 1993; Qumsiyeh 1994). WhileRobertsonian translocations have been well studied in manyspecies, the effects of polymorphisms for pericentric inver-sions in natural populations are less clearly understood.
Species of the genusOligoryzomys (Sigmodontinae,Rodentia) show inter- and intra-population polymorphism
for pericentric inversions affecting one or more chromosomepairs (Zanchin 1988; Almeida and Yonenaga-Yassuda 1991;Espinoza and Reig 1991). The most polymorphic and widelydistributed of these species isOligoryzomys nigripes, whichis found in Paraguay, Uruguay, and Brazil. In Brazil, itsrange extends from Bahia in the north to Rio Grande do Sulin the south. It occurs in various habitats, including Cerradoand Atlantic rainforest. The pericentric inversion polymor-phism detected in various populations ofO. nigripesinvolves four chromosome pairs, viz., pairs 2, 3, 4, and 8
Genome44: 791–796 (2001) © 2001 NRC Canada
791
DOI: 10.1139/gen-44-5-791
Received November 2, 2000. Accepted June 15, 2001. Published on the NRC Research Press Web site at http://genome.nrc.ca onAugust 16, 2001.
Corresponding Editor: C.B. Gillies.
C.R. Bonvicino.1 Department of Genetics, National Institute of Cancer, and Department of Tropical Medicine, Oswaldo CruzInstitute, Rio de Janeiro, Brazil.P.S. D’Andrea. Department of Tropical Medicine, Oswaldo Cruz Institute, Rio de Janeiro, Brazil.P.M. Borodin. Institute of Biophysics, Federal University of Rio de Janeiro, Brazil, and Institute of Cytology and Genetics,Novosibirsk, Russia.
1Corresponding author at Department of Genetics, National Institute of Cancer, Praça da Cruz Vermelha, 23, 6 andar, Rio de Janeiro20230-130, Brazil (email: [email protected]).
I:\gen\gen44\gen-05\G01-080.vpThursday, August 09, 2001 3:11:14 PM
Color profile: Generic CMYK printer profileComposite Default screen

(Myers and Carleton 1981; Brum-Zorilla et al. 1988;Zanchin 1988; Almeida and Yonenaga-Yassuda 1991).
We examined chromosome pairing and recombination inO. nigripesheterozygous for pericentric inversion in chro-mosome 3 and analysed polymorphism for this inversion inseveral populations in Brazil.
Materials and methods
We karyotyped 98 specimens ofO. nigripes trapped in severallocalities (see Fig. 1) and 24 specimens of a captive bred colony ofO. nigripesmaintained at the Oswaldo Cruz Institute (Rio de Ja-neiro).
Mitotic chromosome preparations were obtained from short-term bone marrow cell cultures incubated for 2 h at37°C in RPMI1640 culture medium (Sigma, Brazil) supplemented with 20% foe-tal calf serum, ethidium bromide (5µg/mL), and colchicine (10–6
M) or from primary cultures of kidney epithelium maintained inDulbecco MEM medium (Sigma, Brazil) supplemented with 10%foetal calf serum. G-banding was performed following Seabright(1971).
Conventional meiotic chromosome spreads were prepared fromtestes of five adult males according to Chandley (1989) and stainedwith Giemsa. Surface spreads of primary spermatocytes from thesame males were prepared as described by Chandley (1989) andstained with silver nitrate (Howell and Black 1980). After light mi-croscopic examination, good spreads were transferred to specimengrids and examined and photographed with a JEM-1250 (JEOL)electron microscope at 80 kV.
Results
Meiosis in inversion heterozygotesKaryological analyses of the collected specimens ofO.
nigripes showed that they all had 2n = 62 chromosomes,while fundamental number (FN) varied from 80 to 82 as aresult of floating structural polymorphism for chromosome 3(Table 1). Figure 2 shows the G-banded karyotype ofO.nigripeswith FN = 81. Heteromorphic pair 3 consists of onemetacentric chromosome and one acrocentric counterpart.The pericentric inversion involves approximately 40% of thechromosome.
We analysed chromosome pairing at meiotic prophase inthree individuals heterozygous and two individuals homozy-gous for metacentric chromosome 3, using electron micros-copy. Figure 3 shows an electron micrograph of the surfacespread synaptonemal complex (SC) of male LBCE 550,which was shown to be heterozygous by G-band analysis.Centromeres were not visible in silver-stained preparationsand, therefore, we could not identify bivalent 3 unequivo-cally on the basis of centromere position. A total of 528pachytene nuclei from three of the inversion heterozygoteswere scored for loop-like, eye-like, or Y-like configurations,which are expected inside the inverted segment in cases ofhomologous or delayed synapsis. All autosomal bivalentswere straight paired. This indicated that the inverted regionof chromosome 3 proceeded directly to heterosynapsis, with-out an intervening homosynaptic phase, and the hetero-
© 2001 NRC Canada
792 Genome Vol. 44, 2001
Fig. 1. Map showing the sites whereOligoryzomys nigripeswere collected.
I:\gen\gen44\gen-05\G01-080.vpThursday, August 09, 2001 3:11:22 PM
Color profile: Generic CMYK printer profileComposite Default screen
©2
00
1N
RC
Ca
na
da
Bonvicino
etal.
793
No. of:
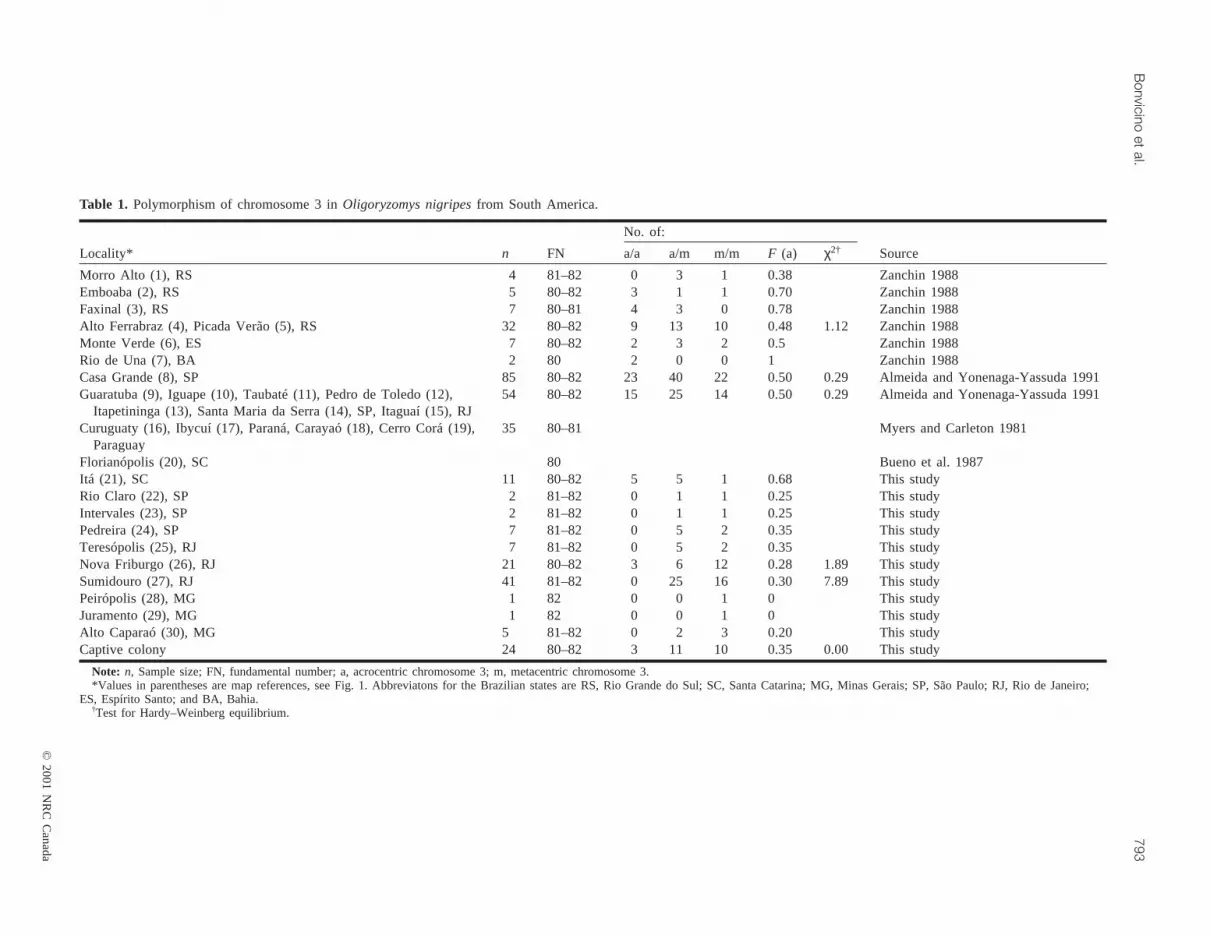
Locality* n FN a/a a/m m/m F (a) χ2† Source
Morro Alto (1), RS 4 81–82 0 3 1 0.38 Zanchin 1988Emboaba (2), RS 5 80–82 3 1 1 0.70 Zanchin 1988Faxinal (3), RS 7 80–81 4 3 0 0.78 Zanchin 1988Alto Ferrabraz (4), Picada Verão (5), RS 32 80–82 9 13 10 0.48 1.12 Zanchin 1988Monte Verde (6), ES 7 80–82 2 3 2 0.5 Zanchin 1988Rio de Una (7), BA 2 80 2 0 0 1 Zanchin 1988Casa Grande (8), SP 85 80–82 23 40 22 0.50 0.29 Almeida and Yonenaga-Yassuda 1991Guaratuba (9), Iguape (10), Taubaté (11), Pedro de Toledo (12),
Itapetininga (13), Santa Maria da Serra (14), SP, Itaguaí (15), RJ54 80–82 15 25 14 0.50 0.29 Almeida and Yonenaga-Yassuda 1991
Curuguaty (16), Ibycuí (17), Paraná, Carayaó (18), Cerro Corá (19),Paraguay
35 80–81 Myers and Carleton 1981
Florianópolis (20), SC 80 Bueno et al. 1987Itá (21), SC 11 80–82 5 5 1 0.68 This studyRio Claro (22), SP 2 81–82 0 1 1 0.25 This studyIntervales (23), SP 2 81–82 0 1 1 0.25 This studyPedreira (24), SP 7 81–82 0 5 2 0.35 This studyTeresópolis (25), RJ 7 81–82 0 5 2 0.35 This studyNova Friburgo (26), RJ 21 80–82 3 6 12 0.28 1.89 This studySumidouro (27), RJ 41 81–82 0 25 16 0.30 7.89 This studyPeirópolis (28), MG 1 82 0 0 1 0 This studyJuramento (29), MG 1 82 0 0 1 0 This studyAlto Caparaó (30), MG 5 81–82 0 2 3 0.20 This studyCaptive colony 24 80–82 3 11 10 0.35 0.00 This study
Note: n, Sample size; FN, fundamental number; a, acrocentric chromosome 3; m, metacentric chromosome 3.*Values in parentheses are map references, see Fig. 1. Abbreviatons for the Brazilian states are RS, Rio Grande do Sul; SC, Santa Catarina; MG, Minas Gerais; SP, São Paulo; RJ, Rio de Janeiro;
ES, Espírito Santo; and BA, Bahia.†Test for Hardy–Weinberg equilibrium.
Table 1. Polymorphism of chromosome 3 inOligoryzomys nigripesfrom South America.
I:\gen\gen44\gen-05\G01-080.vp
Thursday, August 09, 2001 3:11:23 PM
Color profile: Generic CMYK printer profile
Composite Default screen
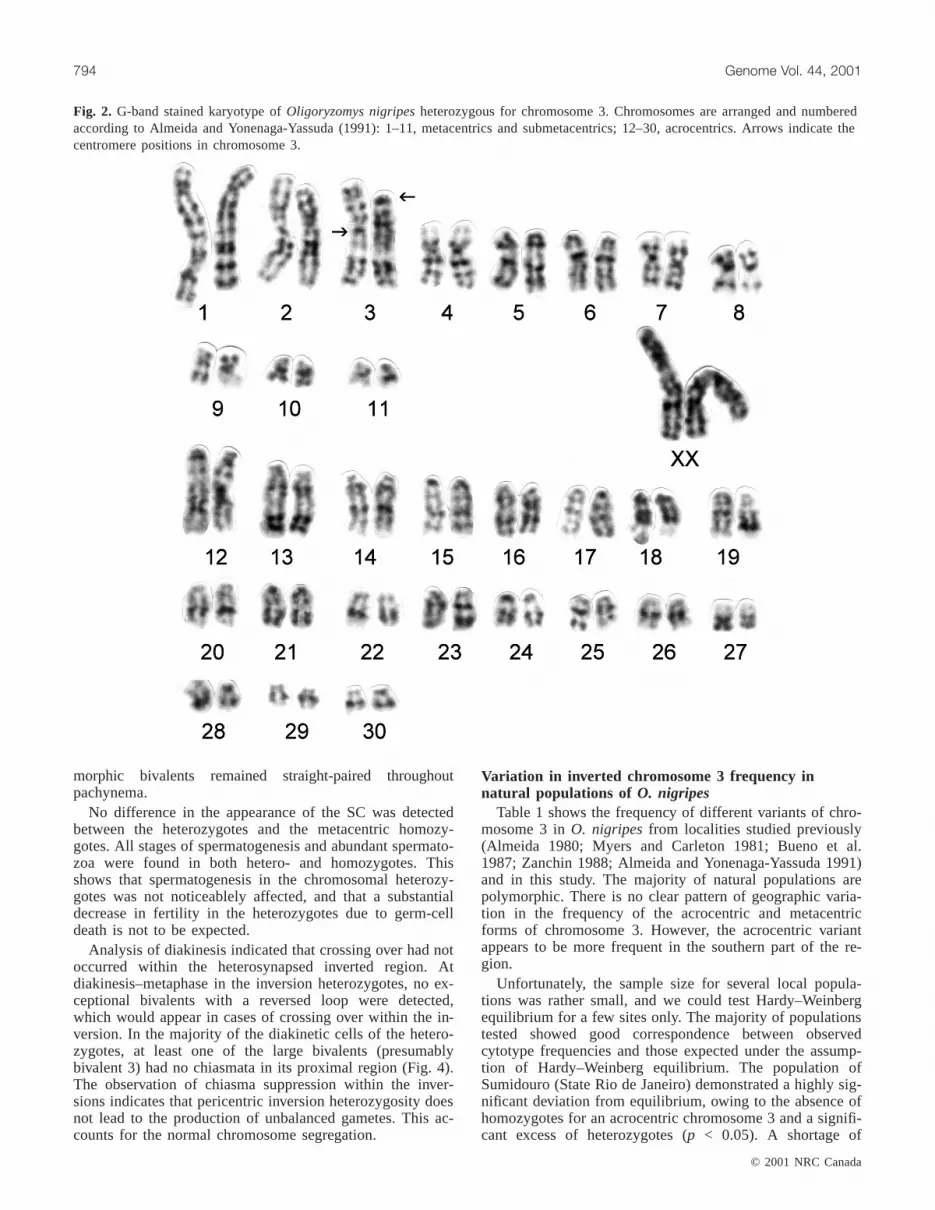
morphic bivalents remained straight-paired throughoutpachynema.
No difference in the appearance of the SC was detectedbetween the heterozygotes and the metacentric homozy-gotes. All stages of spermatogenesis and abundant spermato-zoa were found in both hetero- and homozygotes. Thisshows that spermatogenesis in the chromosomal heterozy-gotes was not noticeablely affected, and that a substantialdecrease in fertility in the heterozygotes due to germ-celldeath is not to be expected.
Analysis of diakinesis indicated that crossing over had notoccurred within the heterosynapsed inverted region. Atdiakinesis–metaphase in the inversion heterozygotes, no ex-ceptional bivalents with a reversed loop were detected,which would appear in cases of crossing over within the in-version. In the majority of the diakinetic cells of the hetero-zygotes, at least one of the large bivalents (presumablybivalent 3) had no chiasmata in its proximal region (Fig. 4).The observation of chiasma suppression within the inver-sions indicates that pericentric inversion heterozygosity doesnot lead to the production of unbalanced gametes. This ac-counts for the normal chromosome segregation.
Variation in inverted chromosome 3 frequency innatural populations of O. nigripes
Table 1 shows the frequency of different variants of chro-mosome 3 inO. nigripes from localities studied previously(Almeida 1980; Myers and Carleton 1981; Bueno et al.1987; Zanchin 1988; Almeida and Yonenaga-Yassuda 1991)and in this study. The majority of natural populations arepolymorphic. There is no clear pattern of geographic varia-tion in the frequency of the acrocentric and metacentricforms of chromosome 3. However, the acrocentric variantappears to be more frequent in the southern part of the re-gion.
Unfortunately, the sample size for several local popula-tions was rather small, and we could test Hardy–Weinbergequilibrium for a few sites only. The majority of populationstested showed good correspondence between observedcytotype frequencies and those expected under the assump-tion of Hardy–Weinberg equilibrium. The population ofSumidouro (State Rio de Janeiro) demonstrated a highly sig-nificant deviation from equilibrium, owing to the absence ofhomozygotes for an acrocentric chromosome 3 and a signifi-cant excess of heterozygotes (p < 0.05). A shortage of
© 2001 NRC Canada
794 Genome Vol. 44, 2001
Fig. 2. G-band stained karyotype ofOligoryzomys nigripesheterozygous for chromosome 3. Chromosomes are arranged and numberedaccording to Almeida and Yonenaga-Yassuda (1991): 1–11, metacentrics and submetacentrics; 12–30, acrocentrics. Arrows indicate thecentromere positions in chromosome 3.
I:\gen\gen44\gen-05\G01-080.vpThursday, August 09, 2001 3:11:34 PM
Color profile: Generic CMYK printer profileComposite Default screen

acrocentric homozygotes and an excess of heterozygoteswere also detected in several other populations from StatesRio de Janeiro and Minas Gerais and from the northern partof State São Paulo (sites 22–25 and 28–30; Table 1), al-though sample sizes were too small to test each populationseparately. On the other hand, the population of NovaFriburgo, located 62 km southeast of Sumidouro, was foundto be in Hardy–Weinberg equilibrium. The distribution ofthe variant chromosomes in a captive bred population de-rived from the population of Sumidouro also showed goodcorrespondence between observed and expected cytotypefrequencies.
Discussion
Chromosome rearrangements that are polymorphic in nat-ural populations rarely affect meiosis or reduce fertility inheterozygous carriers. The meiotic mechanism of “taming”the rearrangements is similar in different species and fordifferent types of rearrangements. It usually involvesnonhomologous pairing of the segments, which differ incontent or position in a heterozygous combination. In thecase of pericentric inversions, nonhomologous synapsis pre-cludes crossing over inside the inverted segments and theformation of unbalanced gametes. Prevalence of nonhomo-logous pairing over homologous pairing has been demon-strated in several species of mammals that are polymorphicfor an inversion (as in, for example, the sand rat (Ashley etal. 1981) and the deer mouse (Greenbaum and Reed 1984;Hale 1986; Hale and Greenbaum 1988)). Suppression ofchiasma formation within the inverted segments (presumably
resulting from heterosynapsis) has been found in pericentric-inversion heterozygotes of the white-throated sparrow(Thorneycroft 1975), the slate-colored junco (Shields 1976),and the spiny lizard (Sites 1983).
The same mechanism is involved in meiotic taming of thepericentric inversion inO. nigripes. We found an orderlypairing of all chromosomes at pachytene in the heterozy-gotes for the inversion. Both SC and conventional meiotic
© 2001 NRC Canada
Bonvicino et al. 795
Fig. 3. Silver-stained surface-spread synaptonemal complex (SC) of a maleOligoryzomys nigripesheterozygous for chromosome 3. Allautosomal bivalents demonstrate straight pairing. The arrowhead indicates the XY bivalent. Scale bar = 10µm.
Fig. 4. Diakinesis in a maleO. nigripes heterozygous for chro-mosome 3. The arrow shows a bivalent with one terminal andone interstitial chiasma. The arrowhead indicates the XY biva-lent. Scale bar = 5µm.
I:\gen\gen44\gen-05\G01-080.vpThursday, August 09, 2001 3:11:48 PM
Color profile: Generic CMYK printer profileComposite Default screen

analysis indicated nonhomologous synapsis in the invertedregion in the heterozygotes. This prevents recombinationwithin the inversion and, therefore, nondisjunction and theformation of unbalanced gametes. Hence, a substantial de-crease in fertility in heterozygous carriers of the inversion isnot to be expected.
Thus, the pericentric inversion in chromosome 3 appar-ently does not result in any selective disadvantages for itsheterozygous carriers. Once it appeared, it should have be-haved as a common neutral mutation, which is the case: thefrequencies of acrocentric homozygotes, metacentric homo-zygotes, and heterozygotes are in accordance with Hardy–Weinberg expectations in the majority of the populationsstudied.
The excess of heterozygotes and the deficiency of acro-centric homozygotes in the Sumidouro population (and per-haps in some other populations from States Rio de Janeiroand Minas Gerais and from the northern part of State SãoPaulo) may be due merely to chance, because the samplesize was relatively small. However, this finding may also beinterpreted in terms of balanced polymorphism. Since re-combination in the inverted region is blocked, owing tononhomologous synapsis, it segregates as a Mendelian unitor supergene, sensu Dobzhansky (1970). Any mutation thatmight have appeared within the inverted region of theacrocentric chromosome would have affected all carriers ofthe inversion. To explain why a deviation from Hardy–Wein-berg equilibrium was observed in a native population ofO.nigripes in Sumidouro but not in the captive bred colony de-rived from it, we hypothesize that this presumed mutationmanifests itself in the particular environment in which theSumidouro population lives, but not under laboratory condi-tions.
Acknowledgements
We are grateful to E.M. Vieira, L. Geise, R.T. Santori, andthe technical staff of the National Health Foundation (FNS)for help in specimen collection and to the Brazilian Instituteof Environment (IBAMA) for granting us permission to col-lect the animals. This work was supported by research grantsfrom the Brazilian National Research Council (CNPq),PRONEX, and the Russian Foundation for Basic Research(RFBR).
References
Almeida, E.J.C. 1980. Variabilidade cromossômica dos roedoresOryzomyse Thomasomys(Cricetidae, Rodentia). Ph.D. thesis,University of São Paulo, São Paulo, Brazil.
Almeida, E.J.C., and Yonenaga-Yassuda, Y. 1991. Pericentric in-version and sexual chromosome heteromorphisms inOryzomysnigripes (Rodentia, Cricetidae). Caryologia,44: 63–73.
Ashley, T., Moses, M.J., and Solari, A.J. 1981. Fine structure andbehaviour of a pericentric inversion in the sand rat,Psammomysobesus. J. Cell Sci.50: 105–119.
Brum-Zorilla, N., de Fronza, T.G., Wainberg, R., Vidal, L., andZwirner, N. 1988.Oryzomys flavescensand O. delticola chro-mosomes (Rodentia, Cricetidae) from Uruguay and Argentina.Caryologia,41: 275–288.
Bueno, A.M., Agostini, J.M.S., Moraes, J., and Ramos, A.P.D.1987. Estudos cromossômicos preliminares de roedores dafamília Cricetidae coletados na Ilha de Santa Catarina. Cienc.Cult. (São Paulo),39(Suppl.): 746.
Chandley, A.C. 1989. The handling and analysis of meiotic cells indomestic and laboratory animals.In Cytogenetics of animals.Edited byC.R.E. Halnan. Commonwealth Agricultural BureauxInternational, Wallingford, U.K. pp. 41–56.
Dobzhansky, T.G. 1970. Genetics of the evolutionary process.Columbia University Press, New York.
Espinoza, M.B., and Reig, O.A. 1991. Cytogenetics andkaryosystematics of South America oryzomyine rodents(Cricetidae, Sigmodontinae). III. Banding karyotypes of Argen-tineanOligoryzomys. Z. Saeugetierkd.56: 306–317.
Greenbaum, I.F., and Reed, M.J. 1984. Evidence for heterosynapticpairing of the inverted segment in pericentric inversion hetero-zygotes of the deer mouse (Peromyscus maniculatus).Cytogenet. Cell Genet.38: 106–111.
Hale, D.W. 1986. Heterosynapsis and suppression of chiasmatawithin heterozygous pericentric inversions of the Sitka deermouse. Chromosoma (Berlin),94: 425–432.
Hale, D.W., and Greenbaum, I.F. 1988. Synapsis of a chromosomalpair heterozygous for a pericentric inversion and the presence ofa heterochromatic short arm. Cytogenet. Cell Genet.48: 55–57.
Howell, W.M., and Black, D.A. 1980. Controlled silver-staining ofnucleolus organizer regions with protective colloidal developer:a 1-step method. Experientia (Basel),36: 1014–1015.
Myers, P., and Carleton, M.D. 1981. The species ofOryzomys(Oligoryzomys) in Paraguay and the identity of Azara’s “ratsixieme ou rat à tarse noir.” Misc. Publ. Zool. No. 161 of theUniversity of Michigan, Ann Arbor. pp. 1–41.
Qumsiyeh, M.B. 1994. Evolution of number and morphology ofmammalian chromosomes. J. Hered.85: 455–465.
Seabright, M. 1971. A rapid technique for human chromosomes.Lancet,2: 971–972.
Searle, J.B. 1993. Chromosomal hybrid zones in eutherian mam-mals. In Hybrid zones and the evolutionary process.Edited byR.G. Harrison. Oxford University Press, New York. pp. 309–351.
Shields, G.F. 1976. Meiotic evidence for pericentric inversion poly-morphism inJunco(Aves). Can. J. Genet. Cytol.18: 747–751.
Sites, J.W. 1983. Chromosome evolution in iguanid lizardsSceloporus grammicus. I. Chromosome polymorphisms. Evolu-tion, 37: 38–53.
Thorneycroft, H.B. 1975. A cytogenetic study of the white-throated sparrow,Zonotrichia albicollis(Gmelin). Evolution,29:611–621.
Zanchin, N. 1988. Estudos cromossômicos em orizominos eequimídeos da Mata Atlântica. M. Sc. thesis, University of SãoPaulo, São Paulo, Brazil.
© 2001 NRC Canada
796 Genome Vol. 44, 2001
I:\gen\gen44\gen-05\G01-080.vpThursday, August 09, 2001 3:11:49 PM
Color profile: Generic CMYK printer profileComposite Default screen