peptidomics of the pars intercerebralis-corpus cardiacum complex of the migratory locust, locusta...
TRANSCRIPT

Eur. J. Biochem. 268, 1929±1939 (2001) q FEBS 2001
Peptidomics of the pars intercerebralis±corpus cardiacum complex ofthe migratory locust, Locusta migratoria
Elke Clynen1, Geert Baggerman1, Dirk Veelaert1, Anja Cerstiaens1, Dick Van der Horst2, Lucien Harthoorn2,Rita Derua3, Etienne Waelkens3, Arnold De Loof1 and Liliane Schoofs1
1Zoological Institute, Catholic University Leuven, Belgium; 2Department of Biochemical Physiology, Utrecht University, the Netherlands;3Laboratory of Biochemistry, Catholic University Leuven, Belgium
The pars intercerebralis±corpora cardiaca system (PI±CC)
of insects is the endocrinological equivalent of the hypo-
thalamus±pituitary system of vertebrates. Peptide profiles
of the pars intercerebralis and the corpora cardiaca were
characterized using simple sampling protocols in combina-
tion with MALDI-TOF and electrospray ionization double
quadrupole time of flight (ESI-Qq-TOF) mass spectro-
metric technologies. The results were compared with earlier
results of conventional sequencing methods and immuno-
cytochemical methods. In addition to many known peptides,
several m/z signals corresponding to putative novel peptides
were observed in the corpora cardiaca and/or pars inter-
cerebralis. Furthermore, for a number of peptides evidence
was provided about their localization and MALDI-TOF
analysis of the released material from the corpora cardiaca
yielded information on the hormonal status of particular
brain peptides.
Keywords: Q-TOF mass spectrometric analysis; MALDI-
TOF imaging; Locusta migratoria; pars intercerebralis;
corpora cardiaca.
The corpora cardiaca (CC) of insects are important neuro-endocrine organs of dual anatomical composition, namely astorage lobe (CCs) and a glandular lobe (CCg). In the CCsneurosecretory products originating from neurosecretorycells located in the brain are stored. The CCg consists ofintrinsic secretory cells, which synthesize and release anumber of other neuropeptides. The CCs can be seen asthe physiological equivalent of the posterior lobe of thepituitary and the CCg of the anterior lobe. Because of thesimilarity between the hypothalamo±hypophysial system ofmammals and the pars intercerebralis±corpora cardiaca(PI±CC) system of insects, the latter has attracted con-siderable interest among endocrinologists. Three types ofpeptides have been isolated and fully identified from thelocust PI±CCs system, namely ovary maturating parsin [1],insulin-related peptide [2] and the group of neuroparsins[3,4]. From the CCg the adipokinetic hormones (AKHs)together with the AKH-precursor related peptides (APRPs)
have been isolated [5±10]. Furthermore, at least 14 neuro-peptides (including FLRF-amide peptides, tachykinins,myotropins, crustacean cardioactive peptide, corazonin,accessory gland myotropin, diuretic hormone, myoinhibit-ing peptide) have been immunolocalized in the CC [11].However, the limitations of this technique, including cross-immunoreactivity with structurally similar peptides, preventthe unequivocal identification of a particular neuropeptide.Not much is known about the hormonal status of all thesepeptides, i.e. whether they are released into the hemolymphor not. So far, this has only been proven for the AKHs [12]and the diuretic hormone [13].
With the advent of ESI and MALDI in the late 1980s,selective and sensitive detection of biomolecules by massspectrometry has become possible. Peptides and proteinscan be detected at very low levels, allowing mass spectro-metric detection of certain compounds present in singleorgans or even in single cells and cell organelles [14]. In thepast, there have been several reports on single organsdirectly analyzed with MALDI-TOF MS without priorpreparation steps [15±19]. This MALDI-TOF method wasmodified by Predel et al. [20] who used a methanolextraction prior to analysis of the neuropeptide content ofthe abdominal perisympathetic organs in cockroaches. Inthe present study, MALDI-TOF and the more sensitive andaccurate ESI-Qq-TOF mass spectrometric methods wereapplied and compared for direct peptide profiling of thelocust PI±CC complex and of the neurohormones releasedfrom the CC. The innovative mass technology ESI-Qq-TOFnot only allows more accurate detection of known andunknown peptides, but also for low molecular mass ionmasses (below 3 kDa), sequence information can beobtained in a tandem MS-analysis from peptide ionsdirectly desorbed from the tissue supernatant. With ourprocedure we bypass the time-consuming chromatographicseparation steps and the mass spectrometric analysis is
Correspondence to E. Clynen, Zoological Institute, Naamsestraat 59,
B-3000 Leuven, Belgium. Fax: 1 32 016 323902,
Tel.: 1 32 016 324260, E-mail: [email protected]
Abbreviations: AKH, adipokinetic hormone; APRP, adipokinetic
hormone precursor related peptide; CC, corpora cardiaca; CCAP,
crustacean cardioactive peptide; CCg, glandular part of the corpus
cardiacum; CCs, storage part of the corpus cardiacum; DH, diuretic
hormone; ESI-Qq-TOF MS, electrospray ionization double quadrupole
time of flight mass spectrometry; F, arginine vasopressine-like insect
diuretic hormone; HrTH, hypertrehalosaemic hormone; LMCI, Locusta
migratoria chymotrypsin inhibitor; Lom, Locusta migratoria; MT,
myotropin; NPA, neuroparsin A; NPB, neuroparsin B; OMP, ovary
maturating parsin; PI, pars intercerebralis.
Note: web page available at http://www.kuleuven.ac.be/bio/fysio
(Received 3 October 2000, revised 5 December 2000, accepted
29 January 2001)

performed with minor sample pretreatment. This speeds upthe total peptide identification procedure significantly. Inaddition, we show that chromatographic analysis followedby MALDI-TOF MS imaging of all collected fractions isalso a relatively fast and elegant technique to screen for themost prominent peptides released from the CC.
M A T E R I A L S A N D M E T H O D S
Investigation of single tissue by means of MALDI-TOFmass spectrometry
Locusta migratoria was raised under laboratory conditions[21]. For all experiments adult locusts were used. The CCand the PI were dissected. After extensive rinsing withNaCl solution (0.1 m), the dissected organ was placed in amicrotube containing 3 mL methanol and was gently shakenfor 10 min. Sample solution (2 mL) was mixed with 0.5 mLof a 50-mm solution of a-cyano-4-hydroxycinnamic acid inacetonitrile/ethanol/trifluoroacetic acid (50 : 49.9 : 0.1) andapplied on a multisample target. This mixture was air-dried,and the target was then introduced in the instrument, aVG Tofspec (Micromass, UK), equipped with a N2-laser(337 nm) and operating with continuous extraction. Thesamples were measured either in the linear (accelerationvoltage 24 kV) or in the reflectron mode (accelerationvoltage 24 kV, reflectron voltage 28.8 kV). In either case,the laser energy was reduced until an optimal resolution andsignal/noise ratio was obtained. The results of 10±20 shotswere averaged to obtain the final spectrum.
In a second experiment the dissected organ was directlyput on the multisample target after rinsing and mixed withthe matrix solution.
Further investigation of tissues by means of Q-TOFmass spectrometry
The CC and PI of Locusta migratoria were dissected,rinsed with a 0.1-m NaCl solution and placed in amicrotube containing 5 mL methanol. After gently shakingfor 10 min, 50 mL milliQ-water was added and the samplewas concentrated and desalted using a ziptipC18 (Millipore,15 mm). First the ziptipC18 was pre-equilibrated for samplebinding using a acetonitrile/water/trifluoroacetic acid(50 : 49.9 : 0.1) solution and subsequently washed with0.1% trifluoroacetic acid in water. Then the sample wasloaded on the ziptipC18 and after washing with 0.1%trifluoroacetic acid in water, eluted with 3 mL acetonitrile/water/formic acid (80 : 19.9 : 0.1). Nanoflow ESI doublequadrupole orthogonal acceleration (Qqoa) TOF massspectrometry was performed on a Q-TOF system (Micro-mass, UK). Two microliter of the acetonitrile/water/formicacid (50 : 49.9 : 0.1) solution containing the sample wasloaded into a gold coated capillary (Protana L/Q nanoflowneedle). This sample was sprayed at a flow rate of30 nL´min21 giving extended analysis time in which MSspectra as well as several MS/MS spectra were acquired.During MS/MS or tandem mass spectrometry, fragment ionsare generated from a selected precursor ion by collisioninduced dissociation. As not all peptide ions fragment withthe same efficiency, the collision energy is typically variedbetween 20 and 35 kV so that the parent ion is fragmentedin a satisfying number of different daughter ions. The
amino-acid sequence is determined by calculating the m/zdifference (which corresponds to the mass of an aminoacid) between the adjacent y-ion peaks and/or b-ion peaks.
Corpora cardiaca release assay
Based on the AKH release assay [12] we developed a moregeneral release assay for peptides released from the CC. CCwere excised and collected on watchglasses in insect salinebuffer (150 mm NaCl, 10 mm KCl, 4 mm CaCl2, 2 mmMgCl2, 10 mm Hepes, pH 7.0). After rinsing three timeswith insect saline buffer, the pooled glands were incubatedin insect saline buffer in control conditions or in insectsaline buffer with a high potassium concentration (100 mmKCl), known to induce depolarization, in experimentalconditions. Incubation took place under continuous andmoderate shaking for 1 h at 21 8C. Incubation media werecollected in microtubes and diluted 1:1 with methanol/water/acetic acid (90:9:1) to prevent sticking of the peptidesto the wall of the tubes. For the same reason the microtubesused were siliconized with 1% diethylchlorosilane dilutedin chloroform.
The incubation media of 120 CC were fractionated bymeans of high performance liquid chromatography (HPLC).Column and operating conditions for HPLC on a Gilson 712system were Waters Symmetry C18 (4.6 � 250 mm), solventA: 0.1% trifluoroacetic acid in water; solvent B: 80%acetonitrile in 0.1% aqueous trifluoroacetic acid. Columnelution conditions: 100% A for 10 min, linear gradient to100% B in 60 min, flow rate 1 mL´min21, detector range:0.2 AUFS (absorption unit full scale). Absorbance wasrecorded at 214 nm and fractions were collected per minute.
One tenth of all collected fractions was concentrated andsubjected to MALDI-TOF analysis. The concentratedsample was mixed with 0.5 mL of a 50-mm solution ofa-cyano-4-hydroxycinnamic acid in acetonitrile/ethanol/trifluoroacetic acid (50 : 49.9 : 0.1), applied on a multi-sample target and analyzed as described before.
N-Terminal amino-acid sequencing of 1/10th of thesamples was performed on a Perkin Elmer/Applied Bio-systems Procise 492 microsequencer operating in thepulsed liquid mode [22].
In another experimental approach the incubation mediaof 5 CC were, after concentrating and desalting with aziptipC18, directly analyzed by MALDI-TOF MS.
R E S U L T S
Comparison of MALDI-TOF and Q-TOF spectra fromsingle tissues
In the locust, Locusta migratoria, 44 neuropeptides havebeen purified and sequenced from whole or parts of centralnervous systems.
To get an idea of the peptide content of, respectively, thePI and the CC, a single organ (CC or PI) was dissected anddirectly subjected to MALDI-TOF mass analysis with orwithout preceding methanol extraction. The best resultswere achieved following methanol extraction. The samespectra were obtained in a very repetitive manner. A fewdifferences, most likely attributable to differences in physio-logical conditions and age, were found in the neuropeptidepattern of individual locusts. The same organs were also
1930 E. Clynen et al. (Eur. J. Biochem. 268) q FEBS 2001

analyzed on Q-TOF, demonstrating a higher sensitivity anda better resolution than on our MALDI-TOF instrument.We have to keep in mind however, that a MALDI-TOFinstrument working with `delayed or time-lag extraction'(this is a small variation on the way ions are extracted fromthe matrix) would have resulted in a better mass accuracythan our MALDI-TOF instrument operating with constantelectrical fields.
Table 1 presents an overview of the peptides found in thePI and/or CC and a comparison between the observed andthe theoretical mass.
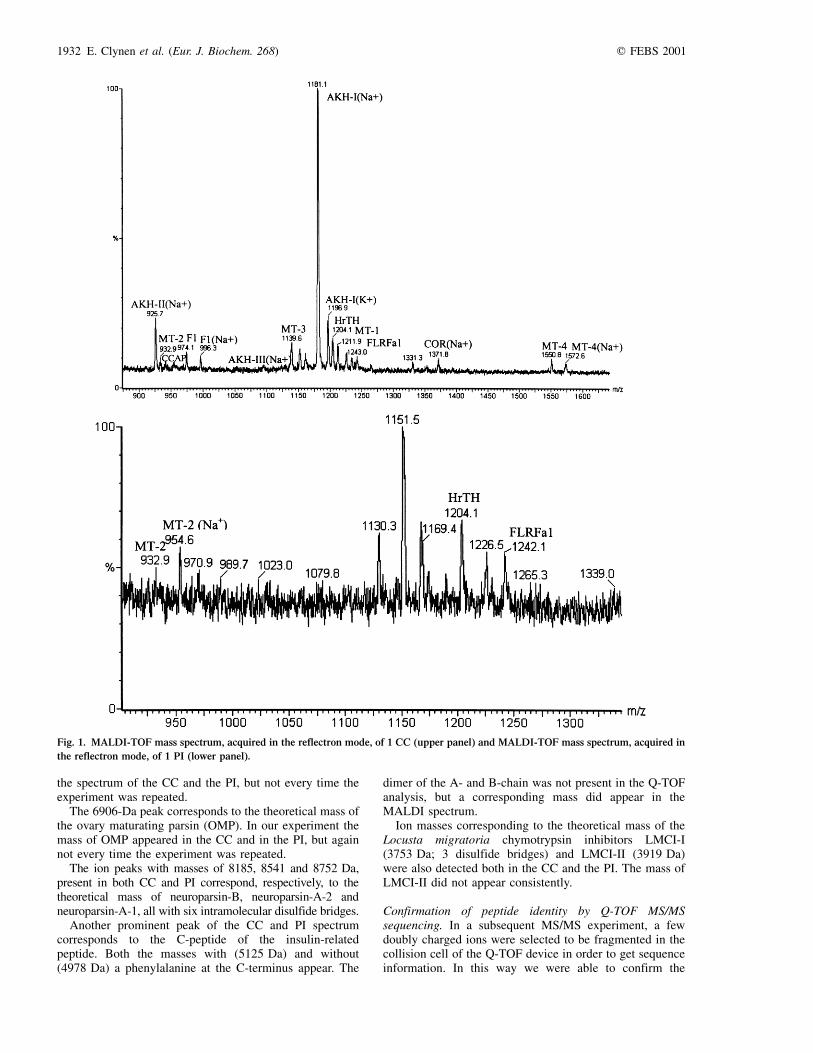
MALDI-TOF analysis of masses in the low molecular massrange (, 3 kDa). The MALDI-TOF mass spectra (Fig. 1)clearly reveal the presence of molecules with massescorresponding to previously sequenced peptides. The mostaccurate data were obtained in the reflectron mode. In theCC, the most prominent MS peaks were 1181 and 1197 Da.These masses perfectly match to the masses, respectively,the sodiated and potassiated ionic forms of AKH-I. Theother two AKHs were also present [AKH-II: 925 Da (Na1)and 942 Da (K1); AKH-III: 1095 Da (Na1)]. The massesof the AKHs were not present in the PI. The mass of therecently discovered Locusta migratoria hypertrehalosaemichormone or Lom-HrTH (1204 Da), was detected in the CCand the PI. Eight other ion mass signals in the CCcorresponded to the theoretical mass of a peptide, namelythe four locustamyotropins [MT-1: 1212 Da; MT-2:933 Da; MT-3: 1140 Da; MT-4: 1551 Da], His-corazonin[1372 Da (Na1)], FLRFamide-1 (1243 Da), crustaceancardioactive peptide (957 Da) and `arginine vasopressine-like diuretic hormone' Lom-F1 (974 Da).
In the PI, the only ion masses matching the theoreticalmass of a peptide were that of HrTH, MT-2 andFLRFamide-1.
Other evident ion peaks in the MS-readout of the CC(for example 1225, 1331 and 1769 Da) and of the PI (forexample 1130, 1151, 1168 and 1225 Da) could not becorrelated with known peptides.
Q-TOF analysis of masses between 3000 and 9000 Da.Because the mass accuracy obtained with our MALDI-TOF instrument in the linear mode is not satisfying,peptides in the higher mass range that could not beseen with MALDI in the reflectron mode were analyzed byQ-TOF.
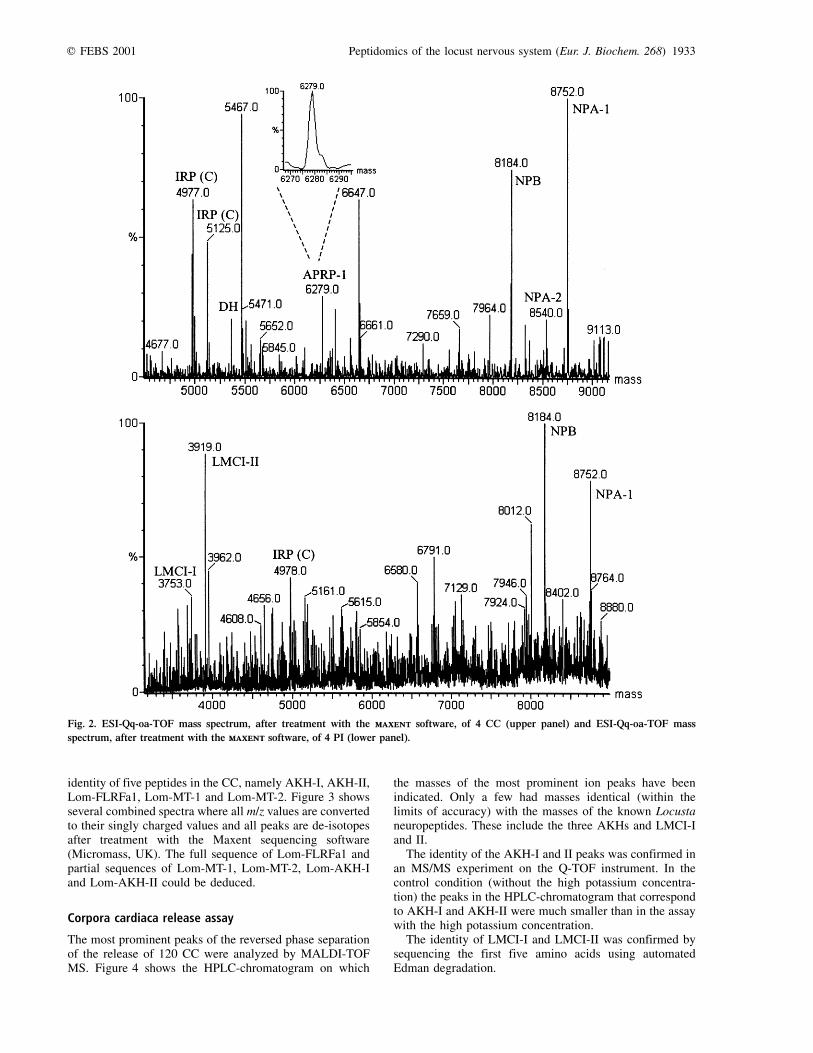
From the multiply charged ions in the Q-TOF massspectra we deduced the masses of the peptides between3000±9000 Da using the maxent software (Micromass,UK) (Fig. 2). A number of ion masses corresponded,within the limits of accuracy inherent to the technique used,to the mass of already sequenced peptides. One of theprominent peaks in the CC profile corresponds to thetheoretical mass of APRP-1 or the AKH-precursor relatedpeptide. This peak was only present in the MS profilereadout of the CC and not the PI. The measured molecularmass (6279 Da) is in agreement with the theoreticallycalculated mass having an intermolecular disulfide bridge.A shoulder of this peak (above 6280 Da) may be anindication for the presence of additional types of dimers(APRP-2 and APRP-3).
The 5363-Da peak corresponds to the theoretical mass ofthe diuretic hormone (DH). The mass of DH was found in
Table 1. List of peptides observed in the CC and the PI. The masses
were obtained by MALDI-TOF MS (reflectron) for peptides below
3000 Da and by ESI-TOF MS (after treatment with the MaxEnt
software) for peptides larger than 3000 Da. The measured mass and the
theoretical mass are given.
Peptides Measured mass
Theoretical mass
(average mass)
Peptides observed only in the CC
Locustamyotropins
Lom-MT-1 1211�.9 Da 1213�.4 Da
Lom-MT-3 1139�.6 Da 1140�.4 Da
Lom-MT-4 1550�.8 Da 1552�.8 Da
1572�.6 Da (Na1) 1574�.8 Da (Na1)
Adipokinetic hormones
Lom-AKH-1 1181�.1 Da (Na1) 1182�.3 Da (Na1)
1196�.9 Da (K1) 1198�.3 Da (K1)
Lom-AKH-2 925�.7 Da (Na1) 927�.0 Da (Na1)
941�.7 Da (K1) 943�.0 Da (K1)
Lom-AKH-3 1095�.4 Da (Na1) 1096�.2 Da (Na1)
AKH-precursor related peptides
APRP-1 (A-A) 6279 Da (1 S-S) 6279�.0 Da (1 S-S)
Corazonin
Lom-COR 1371�.8 Da (Na1) 1373�.4 Da (Na1)
Crustacean cardioactive peptide
Lom-CCAP 954�.6 Da 957�.1 Da
Arginin vasopressin-like diuretic hormone
Lom-F1 974�.1 Da 976�.2 Da
996�.3 Da (Na1) 998�.2 Da (Na1)
Peptides observed in both the CC and the PI
Locustamyotropins
Lom-MT-2 932�.9 Da 934�.0 Da
954�.6 Da (Na1) 956�.0 Da (Na1)
FMRFamides
Lom-FLRFa1 1243�.0 Da 1244�.4 Da
Hypertrehalosaemic hormone
Lom-HrTH 1204�.1 Da 1205�.3. Da
Neuroparsins
Lom-NPB 8184 Da (6 S-S) 8185�.4 Da (6 S-S)
Lom-NPA-1 8752 Da (6 S-S) 8753�.1 Da (6 S-S)
Lom-NPA-2 8540 Da (6 S-S) 8541�.8 Da (6 S-S)
Ovary maturating parsin
Lom-OMP 6906 Da 6906�.5 Da
Insulin-related peptide
Lom-IRP
Chain C 5125 Da 5125�.5 Da
4978 Da (± Phe) 4978�.5 Da (± Phe)
Diuretic peptide
Lom-DH 5363 Da 5363�.4 Da
Protease inhibitors
Lom-LMCI-I 3753 Da (3S-S) 3753�.3 Da (3S-S)
Lom-LMCI-II 3919 Da 3918�.4 Da
q FEBS 2001 Peptidomics of the locust nervous system (Eur. J. Biochem. 268) 1931
the spectrum of the CC and the PI, but not every time theexperiment was repeated.
The 6906-Da peak corresponds to the theoretical mass ofthe ovary maturating parsin (OMP). In our experiment themass of OMP appeared in the CC and in the PI, but againnot every time the experiment was repeated.
The ion peaks with masses of 8185, 8541 and 8752 Da,present in both CC and PI correspond, respectively, to thetheoretical mass of neuroparsin-B, neuroparsin-A-2 andneuroparsin-A-1, all with six intramolecular disulfide bridges.
Another prominent peak of the CC and PI spectrumcorresponds to the C-peptide of the insulin-relatedpeptide. Both the masses with (5125 Da) and without(4978 Da) a phenylalanine at the C-terminus appear. The
dimer of the A- and B-chain was not present in the Q-TOFanalysis, but a corresponding mass did appear in theMALDI spectrum.
Ion masses corresponding to the theoretical mass of theLocusta migratoria chymotrypsin inhibitors LMCI-I(3753 Da; 3 disulfide bridges) and LMCI-II (3919 Da)were also detected both in the CC and the PI. The mass ofLMCI-II did not appear consistently.
Confirmation of peptide identity by Q-TOF MS/MSsequencing. In a subsequent MS/MS experiment, a fewdoubly charged ions were selected to be fragmented in thecollision cell of the Q-TOF device in order to get sequenceinformation. In this way we were able to confirm the
Fig. 1. MALDI-TOF mass spectrum, acquired in the reflectron mode, of 1 CC (upper panel) and MALDI-TOF mass spectrum, acquired in
the reflectron mode, of 1 PI (lower panel).
1932 E. Clynen et al. (Eur. J. Biochem. 268) q FEBS 2001
identity of five peptides in the CC, namely AKH-I, AKH-II,Lom-FLRFa1, Lom-MT-1 and Lom-MT-2. Figure 3 showsseveral combined spectra where all m/z values are convertedto their singly charged values and all peaks are de-isotopesafter treatment with the Maxent sequencing software(Micromass, UK). The full sequence of Lom-FLRFa1 andpartial sequences of Lom-MT-1, Lom-MT-2, Lom-AKH-Iand Lom-AKH-II could be deduced.
Corpora cardiaca release assay
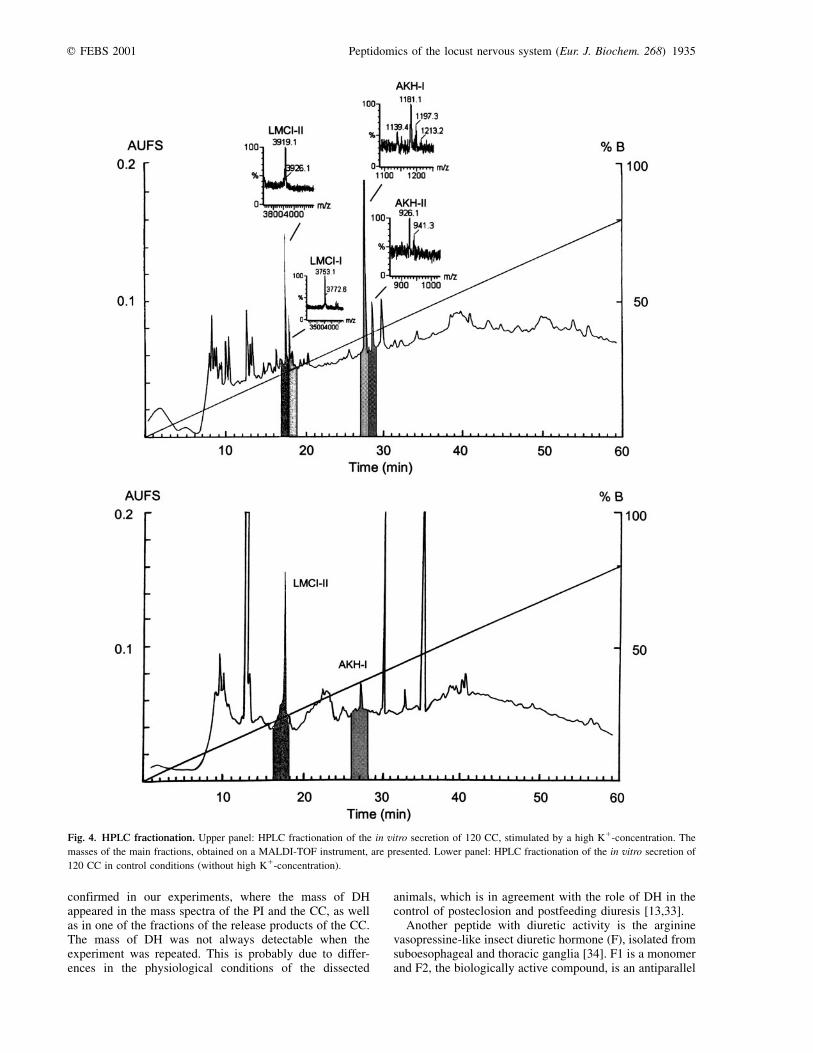
The most prominent peaks of the reversed phase separationof the release of 120 CC were analyzed by MALDI-TOFMS. Figure 4 shows the HPLC-chromatogram on which
the masses of the most prominent ion peaks have beenindicated. Only a few had masses identical (within thelimits of accuracy) with the masses of the known Locustaneuropeptides. These include the three AKHs and LMCI-Iand II.
The identity of the AKH-I and II peaks was confirmed inan MS/MS experiment on the Q-TOF instrument. In thecontrol condition (without the high potassium concentra-tion) the peaks in the HPLC-chromatogram that correspondto AKH-I and AKH-II were much smaller than in the assaywith the high potassium concentration.
The identity of LMCI-I and LMCI-II was confirmed bysequencing the first five amino acids using automatedEdman degradation.
Fig. 2. ESI-Qq-oa-TOF mass spectrum, after treatment with the maxentmaxent software, of 4 CC (upper panel) and ESI-Qq-oa-TOF mass
spectrum, after treatment with the maxentmaxent software, of 4 PI (lower panel).
q FEBS 2001 Peptidomics of the locust nervous system (Eur. J. Biochem. 268) 1933

Direct profiling of the secretion of the CC by MALDI-TOF MS immediately gives an idea of the most abundantpeptides. Among the masses found are those of AKH-I,DH, the three neuroparsins, the C-peptide of the insulin-related peptide and the protease inhibitors LMCI-I andLMCI-II.
D I S C U S S I O N
MALDI-TOF as well as ESI-Qq-TOF MS peptide profilingproves to be an extremely powerful and elegant tool for thefast imaging of prominent neuropeptides present in neuro-endocrine organs, such as the PI±CC system of insects. Inboth the PI and the CC of the migratory locust, Locustamigratoria, we found ion mass signals corresponding to thetheoretical mass of the diuretic hormone, the proteaseinhibitors LMCI-I and II, the neuroparsins A1-2 and B,ovary maturating parsin, insulin related peptide, the hyper-trehalosaemic peptide and the myotropic peptides MT-2and Lom-FLRFa1. AKH I-III and APRP-1 were only con-fined to the CC. More than 20 ion masses did not corre-spond to the theoretical mass of an already identifiedpeptide. We are currently trying to identify them byperforming MS/MS experiments on the Q-TOF (for lowmolecular ion masses) or by Edman sequencing (for ionmasses . 3000 Da).
AKHs are insect neurohormones, which regulate themobilization of lipids and carbohydrates stored in the fatbody to serve as energy substrates for the flight muscles[23]. The synthesis and release of AKHs by the intrinsicneuroendocrine cells of the CC has been known for sometime, and a single short HPLC-run also allows the iden-tification of AKH-I and II in a single CC [24]. Using ourmass spectrometric methods, ion mass signals correspond-ing to the theoretical mass of all three AKHs were detectedin the CC, but not in the PI, which is in agreement withpreviously reported immunocytochemical data [25].
The recently discovered Lom-HrTH was identified bySiegert in the CCs [26]. The site of synthesis was so farunknown. In agreement with Siegert we found the mass ofHrTH in the CC. In addition, we provide evidence that this
peptide is produced by the PI, as the mass was also presentin the PI. Lom-HrTH is structurally related to the AKHsand increases hemolymph trehalose levels in cockroaches.Its biological role in locusts remains to be established.
Three other peptides, APRP 1±3, are cosynthesized withthe AKHs by the neurosecretory cells of the CCg. Both theAPRPs and AKH-I and II are derived from two AKHprecursors (A and B), forming three dimers (A±A, A±Band B±B) [27]. The APRPs consist of three structurallyrelated dimers, two homodimers [respectively a-a chains(APRP-1) and b-b chains (APRP-3)] and a heterodimer[a-b chains (APRP-2)] [28,29]. Their physiological func-tion remains to be clarified [30]. The present mass spec-trometric profiling reveals a mass signal corresponding tothe theoretical mass of the APRP-1. Because of the smalldifference in mass between the three APRPs and the limitedresolution of our mass spectrometric method we cannotconclude with certainty that the other two APRPs are alsopresent. The mass of the APRP g-chain (derived from theAKH-III precursor) was not present in our mass spectra.
Interestingly, in contrast to the AKHs, the APRPs werenot always present in the MS-spectrum of the CC secretion.Hence, the corelease of AKHs and APRPs as a consequenceof their colocalization as proposed by Hekimi and O'Shea[10] and the presence of all three AKH-associated peptidesthat can form the different APRPs in secretory granules asdemonstrated by immunocytochemistry [25] is not evidentfrom our MS analyses.
The presented peptidomic analyses of the master neuro-endocrine organs of locusts not only allows the identifica-tion of the very abundant peptides in the CC, such as theAKHs, but also of less prominent neuropeptides such asLom-DH, Lom-FLRFamide, Lom-myotropins, the parsinsand the serine protease inhibitor peptides.
An ion mass signal corresponding to the theoretical massof Lom-DH was detected in the PI, the CC and in the CCsecretion. DH (46 amino acids) is a corticotropin-releasingfactor-related diuretic peptide that stimulates fluid secretionand cyclic AMP production by the Malpighian tubules. It issynthesized in neurosecretory cells of the PI and is releasedfrom the CC into the hemolymph [31,32]. This was
Fig. 3. The CID (collision induced dissociation) MS/MS fragmentation spectrum of the doubly charged ion at m/z 622.29 Da showing b and
y 0 0 sequence ions obtained by a Q-TOF with a nano-ES (electrospray) source. The obtained sequence corresponds to FLRFa1.
1934 E. Clynen et al. (Eur. J. Biochem. 268) q FEBS 2001
confirmed in our experiments, where the mass of DHappeared in the mass spectra of the PI and the CC, as wellas in one of the fractions of the release products of the CC.The mass of DH was not always detectable when theexperiment was repeated. This is probably due to differ-ences in the physiological conditions of the dissected
animals, which is in agreement with the role of DH in thecontrol of posteclosion and postfeeding diuresis [13,33].
Another peptide with diuretic activity is the argininevasopressine-like insect diuretic hormone (F), isolated fromsuboesophageal and thoracic ganglia [34]. F1 is a monomerand F2, the biologically active compound, is an antiparallel
Fig. 4. HPLC fractionation. Upper panel: HPLC fractionation of the in vitro secretion of 120 CC, stimulated by a high K1-concentration. The
masses of the main fractions, obtained on a MALDI-TOF instrument, are presented. Lower panel: HPLC fractionation of the in vitro secretion of
120 CC in control conditions (without high K1-concentration).
q FEBS 2001 Peptidomics of the locust nervous system (Eur. J. Biochem. 268) 1935

dimer of F1. F1 and F2 had not been demonstrated in theCC yet. In our MALDI-analysis F1 is clearly present in theCC and not in the PI.
6906 Da is the average mass of OMP or the ovarymaturating parsin. OMP has been purified from locust CCextracts and is the first identified gonadotropic hormone ininsects [1,35]. The peptide was immunolocalized in theB-type neurosecretory cells of the PI±CC system fromembryo to adult in both sexes [36,37]. In our experiment amass corresponding to OMP appeared in most of the PI andthe CC MS spectra. However, in a few CC and PI, no massfor OMP could be detected. OMP is known to play agonadotropic role in females by stimulating ecdyson bio-synthesis by the ovary [38,39]. This ecdysiotropic functionis consistent with a temporarily enriched presence in thePI±CC system.
Other prominent ion peaks (8184, 8541 and 8752 Da)corresponded to the masses of the neuroparsins B, A2 andA1. For each neuroparsin a difference of 12 mass unitsbetween the theoretical and the measured mass is observed.This is in agreement with the previously by Hietter andcoworkers suggested six intramolecular disulfide bridges[40] and in disagreement with the dimer structure pre-viously proposed by Girardie and colleagues [4]. The fivemembers of the neuroparsin group, neuroparsin-A 1±4 andneuroparsin-B, were isolated from the CCs. It wasdescribed earlier that NPA is synthesized in the A1-typeneurosecretory cells of the PI and transported via the nervuscorpus cardiacum I (NCCI) to the CC, whereas NPB is notformed in the brain, but only occurs in the CC [41]. Thepresent MALDI imaging experiment unambiguouslyreveals the presence of both NPA and NPB in the PI.The absence of NPA-3 and NPA-4 both in the CC and PIMS-spectra could indicate a differential expression of theNPAs in the PI.
The third parsin is a peptide structurally related to insulin.The 145-residue insulin precursor is post-translationallyprocessed in a 21-residue A chain, a 31-residue B chain anda C-peptide. In our mass spectra of the CC and the PI masssignals corresponding to two forms of the C-peptide (4978and 5125 Da) appeared, but not always together. In the PI,the 4978-Da peak was usually dominant, whereas in theCC, the 5125-Da form was dominant. They are the result ofa C-terminal truncation of a phenylalanine residue. HeÂtruand coworkers isolated the peptide with C-terminal phenyl-alanine [2], while Hietter and coworkers had previouslycharacterized the form without phenylalanine, using adifferent extraction procedure [42]. A mass correspondingto the dimer of the A-and B-chain could be detected inthe MALDI-spectra (^ 5720 Da), but not in the Q-TOFspectra. It is known that the insulin related peptide is poorlysoluble at neutral and slightly acidic pH, as opposed to theC-peptide.
Lom-FLRFa1 and Lom-FLRFa2 have been isolated frombrain tissue of Locusta migratoria [43]. Both peptidespotently inhibit the heart rhythm [44] and inhibit or reducespontaneous contractions of the oviduct of Locustamigratoria [43]. FMRFamide-like immunoreactivity wasdemonstrated in many cells of the PI [45]. However,FMRFamide-antisera cannot differentiate between all locustpeptides ending in RFamide, including locustasulfakinin(with a HMRFamide carboxyterminus). The present ana-lysis clearly identifies Lom-FLRFa1 in the CC and the PI.
Synaptic contacts on the adipokinetic cells in the CCg alsocontain FMRFamide-like material [44], which, according toour present study, is most likely due to the presence ofLom-FLRFa1. The absence of an ion peak in the MALDI-TOF analysis of the CC-secrete is consistent with theinhibitory transmitter function of Lom-FLRFa1 on AKHrelease by the CC [46,47].
Locustamyotropins 1±4 (Lom-MTs) belong to theFXPRLamide peptide family [48,49]. Members of thispeptide family in insects are involved in the control ofvisceral muscles, diapause, pheromone production, pupari-ation and cuticular melanization [50]. The six members ofLocusta migratoria, locustamyotropins 1±4 and locusta-pyrokinins 1±2 were purified from brain-CC complexes.They all stimulate contractions of the locust oviduct [11,50].The detection of Lom-MT ion peaks in the CC spectrum is inagreement with the strong Lom-MT immunoreactivity inthe CCs [51]. However, these Lom-MT-immunoreactivefibers were suggested to originate only from the immuno-reactive cells of the lateral tritocerebrum of the brain.Staining in the PI cells that project to the CC was doubtfuldue to unspecific staining of the nonimmune sera in thesecells. Our MALDI and Q-TOF mass spectra demonstratethe presence of Lom-MT 2 in the PI. Four out of sixisoforms were detected in the CC by mass spectrometry,suggesting a differential expression of the FXPRLamidefamily in the brain, or at least indicating that the myotropins1±4 are the most abundant ones.
Another detected mass in the MALDI-spectrum of theCC was that of the crustacean cardioactive peptide (CCAP)[52]. This peptide which activates heartbeat during flightalso stimulates the release of AKH in a dose-dependentmanner, similar to the hypothalamic releasing factors ofvertebrates [53]. Although CCAP meets the requirementsfor a hormonal status, it was undetectable in the spectra ofthe in vitro secretion assay, which might be explained by itsvery low concentration.
The dark-color inducing neuropeptide [His7]corazoninwas also present in the MALDI-spectrum of the CC [54],but not in the PI, which is in agreement with the recentlyreported immunolocalization of [His7]corazonin in thelocust pars lateralis [55].
Finally, an ion mass corresponding to the mass of theserine protease inhibitor LMCI-I (Locusta migratoriachymotrypsin inhibitor-I) was detected in both PI and CCspectra. The mass of LMCI-II occurred sporadically.LMCI-I and LMCI-II are, respectively, 35 and 36 amino-acid residue peptides, LMCI-I with three intramoleculardisulfide bridges (observed mass: 3753 Da; calculated mass:3759 Da). Boigegrain and associates isolated LMCI-I andII from the plasma of Locusta migratoria [56,57]. At thesame time another group isolated two peptides with thesame characteristics from the PI on the basis of their highUV-absorbance peaks [58]. The peptides were also found tobe present in the fat body and hemolymph and this in higherconcentrations. This all raised the question about theirexact site of biosynthesis and whether the presence in the PIcould have been due to a contamination during micro-dissection. Harding and associates showed the presence ofLMCI-I in the neurosecretory cells of the PI by immuno-cytochemistry [59]. This, however, is no guarantee for thereal occurrence of LMCI-I because false positive stainingcould have occurred. No preimmunesera were used in their
1936 E. Clynen et al. (Eur. J. Biochem. 268) q FEBS 2001

experiment and it is known that a large number of pre-immunesera raised in rabbits against peptides isolated fromlocust brains, stain cell populations and fibers in the centralnervous system due to preexisting antibodies of theIgG class [60]. Our experiments, however, unequivocallyprove the presence of LMCI-I in the neurosecretory cellsof the PI and in the CC. The masses of LMCI-I andLMCI-II were also detected in two fractions of the release-assay of the CC and their identity was confirmed by partialN-terminal sequencing. This indicates that the peptides areproduced by the neurosecretory cells of the PI, transportedto the CC and subsequently released into the hemolymph,similar to neurohormones. The peaks of LMCI-I andLMCI-II were about the same size in both control andexperimental conditions, indicating that their release is notpotassium-dependent.
Some neuropeptides, including locustasulfakinin, access-ory gland myotropins I-II, locustatachykinins, locustapyro-kinins and locustamyoinhibiting peptide, which were purifiedfrom locust brain-suboesophageal ganglion-CC extracts, werenot detected in the MS spectra of the PI and the CC. So it islikely that they are synthesized in other parts of the brain, andnot released by the CC. Another possibility is that theirconcentration in the PI or the CC is below the detection limitof our mass spectrometric methods.
Locusta migratoria is an ideal model insect to study thePI±CC system because not only the CC but also the PI caneasily and cleanly be isolated from other parts of the brain.In addition, the full sequence of numerous Locusta neuro-peptides is already available. With the recently introducedtechnology using isotope coded affinity tags which allowquantitative analysis of MS-peaks, we envisage that in thefuture differences in peptide profiles of the PI±CC systemas visualized by MALDI and ESI-Qq-TOF imaging can becorrelated with physiological events, such as reproduction,molting, feeding, etc. Such differential peptide displaystudies would give information on how the titer of theexpressed peptides of the PI±CC system changes in time.We will be able to look at all peptides at once and thereforesuch peptidomic analysis will provide answers to a varietyof neuro-endocrinological questions.
A C K N O W L E D G E M E N T S
This project was sponsored by the Research Foundation of the K. U.
Leuven (GOA /2000/04) and by the Flemish Science Foundation
(FWO) (G.0356.98). E. C. benefits from a scholarship from the FWO.
Special thanks to Mrs. Julie Puttemans for drawings.
R E F E R E N C E S
1. Girardie, J., Richard, O., Huet, J.C., Nespoulous, C., Van
Dorsselaer, A. & Pernollet, J.C. (1991) Physical characterization
and sequence identification of the ovary maturating parsin. A new
neurohormone purified from the nervous corpora cardiaca of the
African locust (Locusta migratoria migratorioides). Eur. J.
Biochem. 202, 1121±1126.
2. HeÂtru, C., Li, K.W., Bulet, P., Lagueux, M. & Hoffmann, J.A.
(1991) Isolation and structural characterization of an insulin-
related molecule, a pre-dominant neuropeptide from Locusta
migratoria. Eur. J. Biochem. 201, 495±499.
3. Girardie, J., Girardie, A., Huet, J.C. & Pernollet, J.C. (1989)
Amino acid sequence of locust neuroparsins. FEBS Lett. 245, 4±8.
4. Girardie, J., Huet, J.C. & Pernollet, J.C. (1990) The locust
neuroparsin A: sequence and similarities with vertebrate and insect
polypeptide hormones. Insect Biochem. 20, 659±666.
5. Stone, E.V. & Mordue, W. (1976) Structure of locust adipokinetic
hormone, a neurohormone that regulates lipid utilisation during
flight. Nature 263, 207±211.
6. Siegert, K., Morgan, P. & Mordue, W. (1985) Primary structures of
locust adipokinetic hormones II. Biol. Chem. Hoppe Seyler 366,
723±727.
7. Oudejans, R.C.H.M., Kooiman, F.P., Heerma, W., Versluis, C.,
Slotboom, A.J. & Beenakkers, A.M.T. (1991) Isolation and struc-
ture elucidation of a novel adipokinetic hormone (Lom-AKH-III)
from the glandular lobes of the corpora cardiaca of the migratory
locust, Locusta migratoria. Eur. J. Biochem. 195, 351±359.
8. Schoofs, L., Holman, G.M., Proost, P., Van Damme, J., Neven, H.,
Oudejans, R.C.H.M. & De Loof, A. (1993) Isolation, identification
and synthesis of AKH I4210 from Locusta migratoria. Gen. Comp.
Endocrinol. 90, 364±371.
9. Hekimi, S. & O'Shea, M. (1987) Identification and purification
of two precursors of the insect neuropeptide adipokinetic hormone.
J. Neurosci. 7, 2773±2784.
10. Hekimi, S. & O'Shea, M. (1989) Biosynthesis of adipokinetic
hormones (AKHs): Further characterization of precursors and
identification of novel products of processing. J. Neurosci. 9,
996±1003.
11. Veelaert, D., Schoofs, L. & De Loof, A. (1998) Peptidergic control
of the corpus cardiacum-corpora allata complex of locusts. Int.
Rev. Cytol. 182, 249±302.
12. Passier, P.C.C.M., Vullings, H.G.B., Diederen, J.H.B. & Van der
Horst, D.J. (1995) Modulatory effects of biogenic amines on
adipokinetic hormone secretion from locust corpora cardiaca in
vitro. Gen. Comp. Endocrinol. 97, 231±238.
13. Audsley, N., Goldsworthy, G.J. & Coast, G.M. (1997) Quantifica-
tion of Locusta diuretic hormone in the central nervous system and
corpora cardiaca: influence of age and feeding status, and
mechanism of release. Regul. Pept. 69, 25±32.
14. Rubakhin, S.S., Garden, R.W., Fuller, R.R. & Sweedler, J.V.
(2000) Measuring the peptides in individual organelles with mass
spectrometry. Nat. Biotechnol. 18, 172±175.
15. van Veelen, P.A., JimeÂnez, C.R., Li, K.W., Wildering, W.C.,
Geraerts, W.P.M., Tjaden, U.R. & van der Greef, J. (1993) Direct
peptide profiling of single neurons by matrix-assisted laser
desorption-ionization mass spectrometry. Org. Mass Spectrom.
28, 1542±1546.
16. JimeÂnez, C.R., van Veelen, P.A., Li, K.W., Wildering, W.C.,
Geraerts, W.P., Tjaden, U.R. & van der Greef, J. (1994) Neuro-
peptide expression and processing as revealed by direct matrix-
assisted laser desorption ionization mass spectrometry of single
neurons. J. Neurochem. 62, 404±407.
17. JimeÂnez, C.R., Li, K.W., Dreisewerd, K., Spijker, S., Kingston, R.,
Batem, R.H., Burlingame, A.L., Smit, A.B., van Minnen, J. &
Geraerts, W.P. (1998) Direct mass spectrometric peptide profiling
and sequencing of single neurons reveals differential peptide
patterns in a small neuronal network. Biochemistry 37, 2070±2076.
18. Garden, R.W., Moroz, L.L., Moroz, T.P., Shippy, S.A. & Sweedler,
J.V. (1996) Excess salt removal with matrix rinsing: direct peptide
profiling of neurons from marine invertebrates using matrix
assisted laser desorption/ionization time-of-flight mass spectro-
metry. J. Mass Spectrom. 31, 1126±1130.
19. Li, L., Garden, R.W. & Sweedler, J.V. (2000) Single-cell MALDI:
a new tool for direct peptide profiling. Trends Biotechnol. 18,
151±160.
20. Predel, R., Eckert, M. & Holman, G.M. (1999) The unique neuro-
peptide pattern in abdominal perisympathetic organs of insects.
Ann. NY Acad. Sci. 897, 282±290.
21. Ashby, G.J. (1972) Locusts. In UFAW Handbook on the Care and
Management of Laboratory Animals (Poole, T., ed.), pp. 582±587.
Churchill Livingstone, Edinburgh, UK.
q FEBS 2001 Peptidomics of the locust nervous system (Eur. J. Biochem. 268) 1937

22. Hunkapiller, M.W., Hewick, R.M., Dreyer, W.J. & Hood, L. (1983)
High-sensitivity sequencing with a gas-phase sequenator. Methods
Enzymol. 91, 399±412.
23. Oudejans, R.C.H.M., Harthoorn, L.F., Diederen, J.H.B. & Van der
Horst, D.J. (1999) Adipokinetic hormones. Coupling between
biosynthesis and release. Ann. NY Acad. Sci. 897, 291±299.
24. GaÈde, G., Goldsworthy, G.J., Kegel, G. & Keller, R. (1984) Single
step purification of locust adipokinetic hormones I and II by
reversed-phase high-performance liquid chromatography, and
amino acid composition of the hormone II. Hoppe Seylers Z.
Physiol. Chem. 365, 393±398.
25. Harthoorn, L.F., Diederen, J.H.B., Oudejans, R.C.H.M. & Van der
Horst, D.J. (1999) Differential location of peptide hormones in the
secretory pathway of insect adipokinetic cells. Cell Tissue Res.
298, 361±369.
26. Siegert, K.J. (1999) Locust corpora cardiaca contain an inactive
adipokinetic hormone. FEBS Lett. 447, 237±240.
27. Schulz-Aellen, M.F., Roulet, E., Fischer-Lougheed, J. & O'Shea,
M. (1989) Synthesis of a homodimer neurohormone precursor of
locust adipokinetic hormone studied by in vitro translation and
cDNA cloning. Neuron 2, 1369±1373.
28. Hietter, H., Luu, B., GoltzeneÂ, F., Zachary, D., Hoffmann, J. & Van
Dorsselaer, A. (1989) Isolation and structure of two 6-kDa dimeric
peptides from the corpora cardiaca of the insect Locusta
migratoria. Molecular mass determination by mass spectrometry.
Eur. J. Biochem. 182, 77±84.
29. Hekimi, S., Fischer-Lougheed, J. & O'Shea, M. (1991) Regulation
of neuropeptide stoichiometry in neurosecretory cells. J. Neurosci.
11, 3246±3256.
30. Hatle, J.D. & Spring, J.H. (1999) Tests of potential adipokinetic
hormone precursor related peptide (APRP) functions: lack of
responses. Arch. Insect Biochem. Physiol. 42, 163±166.
31. Kay, I., Wheeler, C.H., Coast, G.M., Totty, N.F., Cusinato, O.,
Patel, M. & Goldsworthy, G.J. (1991) Characterization of a
diuretic peptide from Locusta migratoria. Biol. Chem. Hoppe
Seyler 372, 929±934.
32. Lehmberg, E., Ota, R.B. & Furuya, K. (1991) Identification of a
diuretic hormone of Locusta migratoria. Biochem. Biophys. Res.
Commun. 179, 1036±1041.
33. Patel, M., Chung, J.S., Kay, I., Mallet, A.I., Gibbon, C.R.,
Thompson, K.S.J., Bacon, J.P. & Coast, G.M. (1994) Localization
of Locusta-DP in locust CNS and hemolymph satisfies initial
hormonal criteria. Peptides 15, 591±602.
34. Proux, J.P., Miller, C.A., Li, J.P., Carney, R.L., Girardie, A.,
Delaage, M. & Schooley, D.A. (1987) Identification of an arginine
vasopressine-like diuretic hormone from Locusta migratoria.
Biochem. Biophys. Res. Commun. 149, 180±186.
35. Girardie, J., Richard, O. & Girardie, A. (1996) Detection of
vitellogenin in the haemolymph of larval female locusts (Locusta
migratoria) treated with the neurohormone, Lom-OMP. J. Insect
Physiol. 42, 107±113.
36. Richard, O., Tamarelle, M., Geoffre, S. & Girardie, J. (1994)
Restricted occurrence of Locusta migratoria ovary maturating
parsin in the brain-corpora cardiaca complex of various insect
species. Histochemistry 102, 233±239.
37. Richard, O. & Girardie, J. (1992) Immunochemical analysis of the
distribution of the new ovary maturating neurohormone during
development of the African locust, Locusta migratoria. Cell.
Tissue Res. 270, 587±596.
38. Girardie, J., Richard, O. & Girardie, A. (1992) Time-dependent
variations in the activity of a novel ovary maturating neuro-
hormone from the nervous corpora cardiaca during oogenesis in
the locust, Locusta migratoria migratorioides. J. Insect Physiol.
38, 215±221.
39. Girardie, J. & Girardie, A. (1996) Lom OMP, a putative
ecdysiotropic factor for the ovary in Locusta migratoria. J. Insect
Physiol. 42, 215±221.
40. Hietter, H., Van Dorsselaer, A. & Luu, B. (1991) Characterization
of three structurally-related 8±9 kDa monomeric peptides present
in the corpora cardiaca of Locusta: a revised structure for the
neuroparsins. Insect Biochem. 21, 259±264.
41. Girardie, J., Boureme, D. & Girardie, A. (1987) Production sites
of the three neurosecretory proteins characterized in the
corpora cardiaca of the migratory locust. Insect Biochem. 17,
29±36.
42. Hietter, H., Van Dorsselaer, A., Green, B., Denoroy, L., Hoffmann,
J. & Lu, B. (1990) Isolation and structure elucidation of a novel
5-kDa peptide from neurohaemal lobes of the corpora cardiaca of
Locusta migratoria (Insecta, Orthoptera). Eur. J. Biochem. 187,
241±247.
43. Schoofs, L., Holman, G.M., Paemen, L., Veelaert, D.,
Amelinckx, M. & De Loof, A. (1993) Isolation, identification
and synthesis of PDVDHVFLRFamide (SchistoFLRFamide) in
Locusta migratoria and its association with the male accessory
glands, the salivary glands, the heart and the oviduct. Peptides 14,
409±421.
44. Robb, S. & Evans, P.D. (1990) FMRF-like peptides in the locust:
distribution, partial characterization and bioactivity. J. Exp Biol.
149, 335±360.
45. Myers, C.M. & Evans, P.D. (1987) An FMRFamide antiserum
differentiates between populations of antigens in the brain
retrocerebral complex of the locust, Schistocerca gregaria. Cell
Tissue Res. 250, 93±99.
46. Passier, P.C.C.M., Van der Jagt, E.M., Vullings, H.G.B.,
Diederen, J.H.B. & Van der Horst, D.J. (1994) The innervation of
the adipokinetic cells in Locusta migratoria: Involvement of
FMRFamide-immunopositive nerve fibers. In Sensory Transduction
(Elsner, N., & Breer, H., eds), pp. 683. Thieme-Verlag, Stuttgart,
Germany.
47. Vullings, H.G., Ten Voorde, S.E., Passier, P.C., Diederen, J.H.,
Van der Horst, D.J. & NaÈssel, D.R. (1998) A possible role
of SchistoFLRFamide in inhibition of adipokinetic hormone
release from locust corpora cardiaca. J. Neurocytol. 27, 901±913.
48. Schoofs, L., Holman, G.M., Hayes, T.K., Tips, A., Nachman, R.J.,
Vandesande, F. & De Loof, A. (1990) Isolation and identification
of locustamyotropin (Lom-MT), a novel biologically active insect
neuropeptide. Peptides 11, 427±433.
49. Schoofs, L., Holman, G.M., Hayes, T.K., Nachman, R.J. &
De Loof, A. (1990) Isolation, identification and synthesis of
locustamyotropin II, an additional neuropeptide of Locusta
migratoria: Member of the cephalomyotropic peptide family.
Insect Biochem. 20, 479±484.
50. Schoofs, L., Veelaert, D., Vanden Broeck, J. & De Loof, A. (1997)
Peptides in the locusts, Locusta migratoria and Schistocerca
gregaria. Peptides 18, 145±156.
51. Schoofs, L., Tips, A., Holman, G.M., Nachman, R.J. & De Loof,
A. (1992) Distribution of locustamyotropin-like immunoreactivity
in the nervous system of Locusta migratoria. Regul. Pept. 37,
237±254.
52. Veelaert, D., Passier, P., Devreese, B., Vanden Broeck, J.,
Van Beeumen, J., Vullings, H.G.B., Diederen, J.H.B.,
Schoofs, L. & De Loof, A. (1997) Isolation and characterization
of an adipokinetic hormone release-inducing factor in
locusts: the crustacean cardioactive peptide. Endocrinology 138,
138±142.
53. Stangier, J., Hilbich, C. & Keller, R. (1989) Occurrence of
crustacean cardioactive peptide (CCAP) in the nervous system of
an insect. J. Comp. Physiol. B. 159, 5±11.
54. Tawfik, A.I., Tanaka, S., De Loof, A., Schoofs, L., Baggerman, G.,
Waelkens, E., Derua, R., Milner, Y., Yerushalmi, Y. & Pener, M.P.
(1999) Identification of the gregarization-associated dark-pigmen-
totropin in locusts through an albino mutant. Proc. Natl Acad. Sci.
USA 96, 7083±7087.
1938 E. Clynen et al. (Eur. J. Biochem. 268) q FEBS 2001

55. Schoofs, L., Baggerman, G., Veelaert, D., Breuer, M., Tanaka, S.
& De Loof, A. (2000) The pigmentotropic hormone [His7]-
corazonin, absent in a Locusta migratoria albino strain, occurs in
an albino strain of Schistocerca gregaria. Mol. Cell. Endocrinol.
168, 101±109.
56. Boigegrain, R.A., Mattras, H., BreheÂlin, M., Paroutaud, P. &
Coletti-Previero, M.A. (1992) Insect immunity: two proteinase
inhibitors from hemolymph of Locusta migratoria. Biochem.
Biophys. Res. Commun. 189, 790±793.
57. Boigegrain, R.A., PugnieÁre, M., Paroutaud, P., Castro, B. &
BreheÂlin, M. (2000) Low molecular weight serine protease
inhibitors from insects are proteins with highly conserved
sequences. Insect Biochem. Mol Biol. 30, 145±152.
58. Nakakura, N., Hietter, H., Van Dorsselaer, A. & Luu, B. (1992)
Isolation and structural determination of three peptides from the
insect Locusta migratoria. Identification of a deoxyhexose-linked
peptide. Eur. J. Biochem. 204, 147±153.
59. Harding, L., Scott, R.H., Kellenberger, C., Hietter, H., Luu, B.,
Beadle, D.J. & Bermudez, I. (1995) Inhibition of high voltage-
activated Ca21 currents from cultured sensory neurones by a
novel insect peptide. J. Recept. Signal Transduct. Res. 15,
355±364.
60. Tips, A., Schoofs, L., Paemen, L., Hendrickx, K. & De Loof, A.
(1997) False positive immunostaining of Locusta neurosecretory
cells with a variety of preimmune sera. Gen. Comp. Endocrinol.
106, 231±240.
q FEBS 2001 Peptidomics of the locust nervous system (Eur. J. Biochem. 268) 1939