past vegetation changes in the brazilian pantanal arboreal-grassy savanna ecotone by using carbon...
TRANSCRIPT

Global Change Biology (1995) 1,165-171
Past vegetation changes in the Brazilian Pantanalarboreal-grassy savanna ecotone by using carbonisotopes in the soil organic matter
REYNALDO LUIZ VICTORIA*, FERNANDO F E R N A N D E S t , LUIZ A N T O N I OMARTINELLI*, MARISA DE CASSIA PICCOLO*, PLINIO BARBOSA DE C A M A R G O 'andSUSAN TRUMBOREJ"Centra de Eneri;;ia Nuclear na Agriatltura. Av. Centend rio 303. CEP 13416-000. Piracicaba. SP. Brazil, iCetitro dc PesqitisaAgropeciidria do Pautanal, Etnbrapa, Corumbd. MT. Brazil, JUnwers/h/ of California at Irvine, Department of Earth SystemScience, Irvine, CA, USA
Abstract
Measurements of the organic carbon inventory, its stable isotopic composition andradiocarbon content were used to deduce vegetation history from two soil profiles inarboreal and grassy savanna ecotones in the Brazilian Pantanal. The Pantanal is a largefloodplain area with grass-dominated lowlands subject to seasonal flooding, and arborealsavanna uplands which are only rarely flooded. Organic carbon inventories were lowerin the grassy savanna site than in the upland arboreal savanna site, with carbondecreasing exponentially with depth from the surface in both profiles. Changes in ^ Cof soil organic matter (SOM) with depth differed markedly between the two sites.Differences in surface SOM "C values reflect the change from C3 to C4 plants betweenthe sites, as confirmed by measurements of ' C of vegetation and the soil surface alonga transect between the upland closed-canopy forest and lowland grassy savanna. Changesof ^ C in SOM with depth at both sites are larger than the 3-4 per mil increases expectedfrom fractionation associated with organic matter decomposition. We interpret these asrecording past changes in the relative abundance of C3 and C4 plants at these sites.Mass balances with '*C and "C suggest that past vegetational changes from C3 to C4plants in the grassy savanna, and in the deeper part of the arboreal savanna, occurredbetween 4600 and 11 400 BP, when major climatic changes were also observed in severalplaces of the South American Continent. The change from C4 to C3, observed only inthe upper part of the arboreal savanna, was much more recent (1400 BP), and wasprobably caused by a local change in the flooding regime.
Keywords: Brazil, Pantanal, radiocarbon, wetland, savanna, soil organic matter, stable carbonisotope, vegetation change
Received 14 November 1994; revision received 28 February 1995; accepted 6 May 1995
Introduction
The Pantanal, with an area of c. 140 000 km , is one ofthe largest fioodplains in the world. It is a large depressionformed at the end of the Cretaceous and filled by sedi-ments deposited mainly by the Paraguay river and itsmajor tributaries, especially during the Quaternary(Moreira 1977; Moreira Franco & Pinheiro 1982; Ab'Saber1988). It is located at the centre of South America,bordering the main physiographic zones of the continent.
Correspondence: Reynaldo Luiz Victoria, fax +55-194-228339,e-mail [email protected]
with the Amazon region to the north, the Cerrado to theeast and the Chaco to the west. Due to the alternation ofdry and wet climates that occurred at the end of thePleistocene and during the Quaternary (Alvarenga et al.1984), a mixed vegetation pattern was established, withspecies characteristic of the border areas (Amazon,Cerrado and Chaco) and with very few endemic speciesOunk 1993). Drier periods also fostered the invasion ofxeric species from the Caatinga of NE Brazil (Ab'Saber1988). The distribution of the vegetation in the landscapeis a function of the topography, which also governs thepattern of flooding. Consequently, small changes in the
1995 Blackwell Science Ltd. 165

166 R.L. VICTORIA et al.
topography lead to a mosaic of habitats (Junk 1993),supporting numerous species of plants and animals, inone of the most biodiverse ecosystems of the world. Themajor vegetation type in the Pantanal is the savanna, inwhich different sub-units are established as a function ofthe local topography/hydrology (Loureiro et al. 1982).Plant communities may be classified based on the relativeproportion of C4 (grasses) and C3 plants. The low-landareas are dominated by C4 grasses forming the so-calledgrassy savanna. Higher elevation areas have an increasingproportion of C3 plants with the progression from parksavanna to open canopy arboreal savanna. Closed canopyarboreal savanna, formed almost exclusively by C3 plants,dominates the highest elevations.
Spatial distributions of these vegetation are expectedto be dynamic in a system like the Pantanal, both becauseof periodic flooding, and past climate changes.
In this paper we deduce present and past changes inthe vegetation composition of a typical savanna of thesouth-eastern part of the Pantanal, using '•'C isotopiccomposition and radiocarbon dating of soil organic matter(SOM). The bases for the use of the '•'C technique are: (i)the isotopic composition of the SOM is similar to theisotopic composition of the vegetation cover from whichit is derived, and (ii) there is a large difference in theisotopic composition of C3 and C4 plants (Vitorello t'f al.1989; Martin et al. 1990; Ambrose & Sikes 1991; Mariotti& Peterschmitt 1994). Generally, the SOM in deeper layersof a soil is enriched in -C (Flexor & Volkoff 1977; Volkoff& Cerd 1987; GuiUet et al. 1988; Desjardins et al. 1991).Depending on the degree of this enrichment two basicprocesses can explain the observed trends: (i) increasessmaller than 3-4%o are likely due to organic matterdecomposition, which preferentially removes '•^,enriching deeper, older soil layers in '•'C; and (ii) largervalues are likely the result of changes from C4 to C3vegetation. Radiocarbon dating is used here to identifypast vegetation change.
We collected samples from soil pits dug in grass andin closed canopy arboreal savannas. Virtually no grasseswere present in the arboreal savanna. The absence ofsignificant change in the stable carbon isotopic composi-tion with depth, for any of the pits, would indicate thatthe system has been stable for the period over which thesoil has formed. This period is determined from theradiocarbon age of SOM in the profile. A '^C enrichmentwith depth in the arboreal savanna, larger than theenrichment of 3-4 %o that typically results indecomposition, would indicate that the system evolvedfrom a grassy savanna. Alternatively, a '' C depletionwith depth in the grassy savanna, would suggest that anarboreal savanna was present before being replaced byC4 grassy savanna.
Material and methods
Study area
The Pantanal may be divided into 10 different geomor-phological regions (Adamoli 1982). The Fazenda Nhumi-rim (18° 59'S and 56'' 39'W), our sampling station, issituated in the Nhecolandia region. The climate of theregion is classified Aw, after the Koppen system. Theannual average temperature is 25" C, and the annualaverage rainfall is 1280 mm. The dry season extends fromMay to September and the rainy season from October toApril. The soil was classified as Areia Quartzoza accordingto the Brazilian classification, equivalent to an Inceptisolin the American classification system.
The samples were collected in a typical savanna com-plex. Grassy savanna, formed almost exclusively by C4grasses, dominated the lower elevation terrain, which isperiodically inundated. C3 plants as a proportion of thevegetation increased with elevation, to the exclusively C3dosed arboreal savanna, situated in higher terrain c. 1.5 mabove the grassy savanna and so avoiding periodicalflooding. Two soil sampling procedures were used. Thefirst was a 950 m transect from the grassy to the arborealsavanna, where surface soil samples (0-20 cm) werecollected every 50 m. The second was performed in soilpits dug in both the grassy and the arboreal savanna.Samples were collected at 5 cm intervals from the surfaceto 20 cm deep, and then at 10 cm intervals to the bottomof the pits (110 cm in the grassy savanna and 170 cm inthe arboreal savanna). Following a botanical survey ofthe area (Pott ct al 1986) leaves from the most commonplants in the grassy savanna, in the forest and in theborder of the ecotone were collected for '-*C determina-tion. In the arboreal savanna leaves were collected fromthe upper canopy (10-15 m) and also from the groundflora.
Analytical methods
Large roots and debris were separated from the soil usinga 2 mm sieve. The sandy texture of the soil allowedmanual separation of small roots by hand. Carbon concen-trations were determined in a CHN analyser (PerkinElmer, 2400 CHN), Sample treatment for isotope analysiswas done through combustion with CuO in evacuatedPyrex tubes. Isotope measurements were performed witha Finnigan DELTA-E mass spectrometer fitted with doubleinlet and double collector systems. Results are expressedin 5 C relative to the PDB standard, defined as:
\ ^standard
where Rgampie and Rstdnd.ird ^rt' the ratio '-'C:'^C of the
© 1995 Blackwell Science Ltd., Global Change Biology, 1,165-171

PAST VEGETATION C H A N G E IN THE PANTANAL 167
sample and standard, respectively. Samples were ana-lysed at least in duplicate with a maximum difference of0.3 %o between replicates.
'*C activity was measured in an accelerator massspectrometer at the Lawrence Livermore Laboratory, USA(Southon ct al. 1992; Trumbore 1993). The conventionalradiocarbon age, expressed as years before present (BP)± standard-deviation, called Apparent Mean ResidenceTime (AMRT), was estimated using the following equa-tion, assuming a '"'C half-life of 5568 years (Stuiver &Pearson 1986):
t = -80331n'sample
^standard(2)
where / is the radiocarbon age expressed in years BI', and- siimple 3nd ' stand.ud ^ -e the radiocarbon activity of thesample and standard, respectively.
For the '•*C mass balance presented in the Discussionsection, the '*C activity was expressed as A ' C per mil,estimated according to the following equation (Stuiver &Polach 1977):
/ "sample \1000, (3)
Carbon stocks in soils were calculated by multiplyingthe respective concentrations by soil density and by soildepth. Soil density was calculated by collecting a massof soil in a small cylinder of known volume. After dryingto constant mass, the density was calculated.
Results
Soil organic carbon concentration and stock
Organic carbon concentrations were very tow in bothareas, and showed the typical decrease with depth. Forany particular depth the concentration was higher in thearboreal than in the grassy savanna (Fig. 1). The highercarbon stocks in the arboreal savanna, resulted from ahigher carbon concentration, since the soil densities fromboth areas were not significantly different. The carbonstock to 1 m depth was equal to 3.4 kgC m~^ in the arborealsavanna and equal to 2.0 kgC m" in the grassy savanna.
in the forest floor (-31.1 ±1.5 %o, n = 6) vegetation. Thisenrichment was also present in the tropical environment(van der Merwe & Medina 1989; Stemberg et al. 1989;Kapos et ai 1993). The probable causes are, in order ofimportance, lower irradiance inside the forest, increasingthe isotopic fractionation during photosynthesis, and toa minor extent, the use of isotopically depleted biogenicCO2 (Farquhar et al. 1982; Schleser & Jayasekera 1985;Sternberg et al. 1989). Surprisingly, the difference betweenthe upper canopy and ground flora of the arborealsavanna was not statistically different, as usually foundin tropical forests (van der Merwe & Medina 1989;Martinelli et al. 1991). The small number of forest floorvegetation samples, together with the high variance ofthe data, may explain this lack of significance. Also, thearboreal savanna at this site was more open with onlytwo strata, in comparison to forest of higher rainfall areas,with less potential for recycling of C within the canopy.
Carbon isotopic composition of soil organic matteralong the transect
The variation of 5 C along the transect followed theanticipated pattern: as the proportion of C3 to C4 plantsincreased, the values became more negative, reflectingthe increasing contribution of C3 derived organic matterin the SOM (Fig. 2). Two major changes were observedin the transect, the first after 350 m, where the 5'' C valuesdropped from -17 to -20 %o, and the second at 600 m,where a drop from -20 to —27 %o occurred. Thesechanges are the results of changes in vegetation cover.The first drop indicates the fransition from grassy savannato park savanna, and the second the transition from parkto arboreal savanna.
The average 5' C values for soil surface samples col-lected in the grassy savanna (-17.2i0.6 '%»>, n = 7) arewithin the normal range for this type of vegetation cover.The average value of 8 - C found in the literature forsamples collected from surface soils covered with C4grasses was -16.1 ±2.2 %o (Dzurec et al. 1985; Mondenesiet al. 1986; Schwartz et al. 1986; Volkoff & Cerri 1987;Martin et al. 1990; Desjardins et al. 1991; McPherson et al.1993; Wang et al. 1993; Mariotti & Peterschmitt 1994;Trouve et al 1994).
Carbon composition of plant species
The h^^C values of the plant species collected along thetransect are shown in Table 1. The dominant grass in thegrassy savanna, Elyonurus miticus, showed a value typicalof C4 plants (-12.4 %o). The trees from the border areashowed an average 5''*C value of -28.8±0.9 %o (n = 10),which is significantly less negative than the averagefound in the upper canopy (-30.3±0.9 %<,, n = 11) and
Variation in carbon isotopic composition with soildepth and radiocarbon dating of the soil
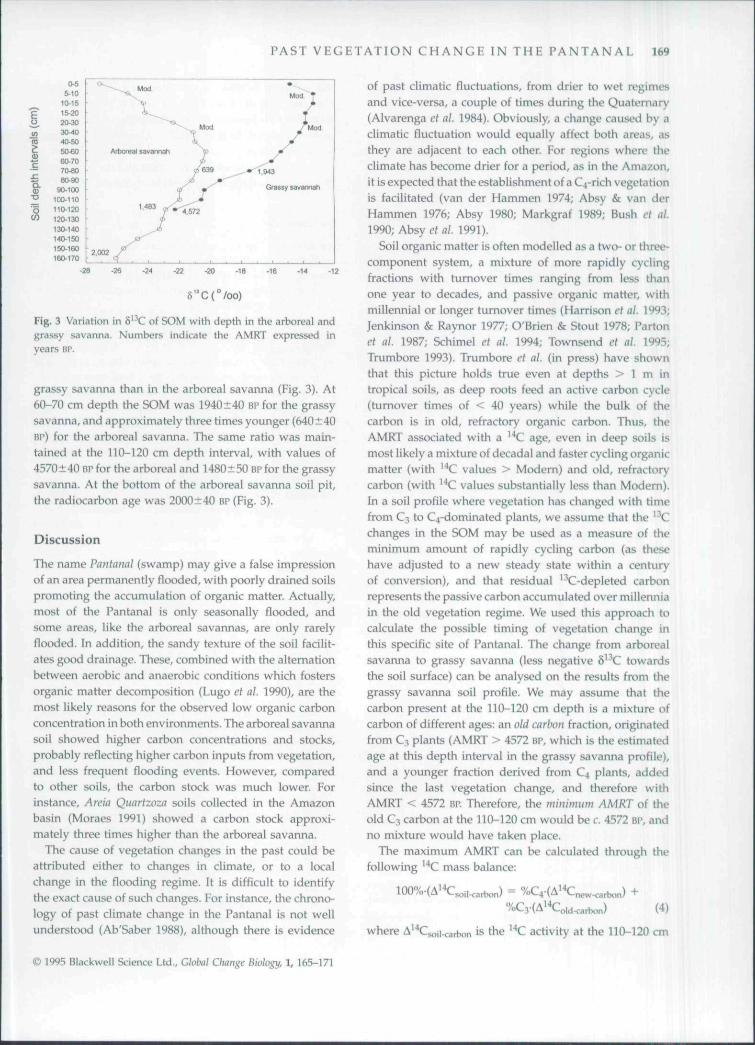
The variations in d^Hl with depth, for both areas, werelarger than the expected from fractionation during decom-position, suggesting that vegetation changes occurred inthe past (Fig. 3). In the grassy savanna the S' 'C becameprogressively more negative with depth, suggesting that aprogressive replacement of C3 by C4 plants had occurred.
1995 Blackweil Science Ltd., Clobal Change Biology, 1. 165-171
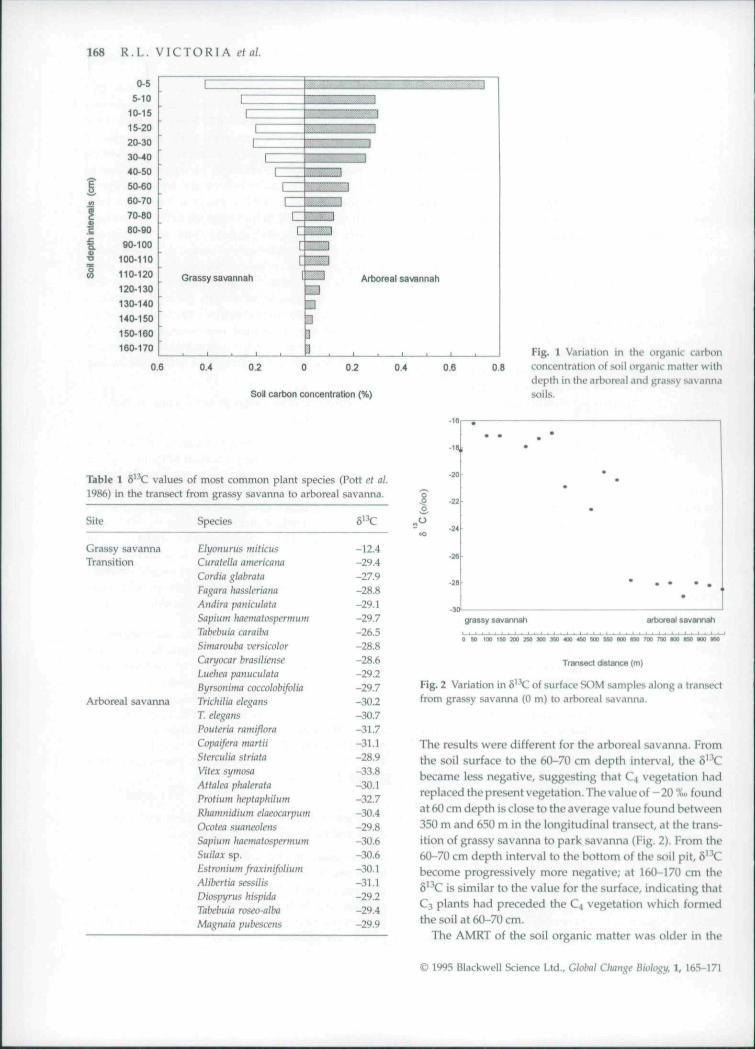
168 R,L. VICTORIA et al.
0-5
5-10
10-1515-2020-3030-4040-5050-6060-7070-8080-30
90-100
100-110
110-120
120-130130-140
140-150150-160
160-170
0,6
Grassy savannah Arboreal savannah
0.4
Soil carbon concentration (%}
Table 1 5' C values of most common plant species (Pott et ai1986) in the transect from grassy savanna to arboreal savanna.
Site Species
Grassy savannaTransition
Arboreal savanna
Elyonurus miticusCuratella americamCordia glabralaFagara hasslirianaAndirn paiiiculalaSapium haeninfosperniumTabehuia caraibaSimarouba versicohrCaryocar brasilienseLuehea pamiculataByrsonima coccolobifoliaTrichilia elegansT. elegansPouteria ramifloraCopaifera martiiSterculia striataVite.\ symosaAttak'a phalerataProtium heptapluhimRhamnidiuni elacocarpumOcotea suaneolensSapium haematospermumSuilax sp.Estroitium fraxinifoUumAlibertia sessilisDiospyrus hispidaTabebuia rosea-albaMagnaia pubescens
-12.4-29.4-27.9-28.8-29.1-29.7-26.5-28.8-28.6-29.2-29.7-30.2-30.7-31.7-31.1-28.9-33.8-30,1-32.7-30.4-29.8-30,6-30.6-30,1-31,1-29.2-29.4-29.9
0,6 0.8
Fig. 1 Variation in the organic carbonconcentration of soil organic matter withdepth in the arboreal and grassy savannasoils.
grassy savannah arboreal savannah
Transect distance (m)
Fig. 2 Variation in 6'Hr of surface SOM samples along a transectfrom grassy savanna (0 m) to arboreal savanna.
The results were different for the arboreal savanna. From
the soil surface to the 60-70 cm depth inlerval, the 6' C
became less negative, suggesting that C4 vegetation had
replaced the present vegetation. The value of -20 %•• found
at 60 cm depth is close to the average value found between
350 m and 650 m in the longitudinal transect, at the trans-
ition of grassy savanna to park savanna (Fig, 2), From the
60-70 cm depth interval to the bottom of the soil pit, S'-'C
become progressively more negative; at 160-170 cm the
S'-'C is similar to the value for the surface, indicating that
C3 plants had preceded the C4 vegetation which formed
the soil at 60-70 cm.
The AMRT of the soil organic matter was older in the
1995 Blackwell Science Ltd., Global Change Biology, t, 165-171
PAST VEGETATION CHANGE IN THE PANTANAL 169
a.<t>•o
o
Fig. 3 Variation in S -'C of SOM with depth in the arboreal andgrassy savanna. Numbers indicate the AMRT expressed inyears ar.
grassy savanna than in the arboreal savanna (Fig. 3). At60-70 cm depth the SOM was 1940±40 BP for the grassysavanna, and approximately three times younger (640±40BP) for the arboreal savanna. The same ratio was main-tained at the 110-120 cm depth interval, with values of4570+40 BP for the arboreal and 1480±50 BP for the grassysavanna. At the bottom of the arboreal savanna soil pit,the radiocarbon age was 2000±40 BP(Fig. 3).
Discussion
The name Patitatml (swamp) may give a false impressionof an area permanently flooded, with poorly drained soilspromoting the accumulation of organic matter. Actually,most of the Pantanal is only seasonally flooded, andsome areas, like the arboreal savannas, are only rarelyflooded. In addition, the sandy texture of the soil facilit-ates good drainage. These, combined with the alternationbetween aerobic and anaerobic conditions which fostersorganic matter decomposition (Lugo et ai 1990), are themost likely reasons for the observed low organic carbonconcentration in both environments. The arboreal savannasoil showed higher carbon concentrations and stocks,probably reflecting higher carbon inputs from vegetation,and less frequent flooding events. However, comparedto other soils, the carbon stock was much lower. Eorinstance, Areia Quartzoza soils collected in the Amazonbasin (Moraes 1991) showed a carbon stock approxi-mately three times higher than the arboreal savanna.
The cause of vegetation changes in the past could beattributed either to changes in climate, or to a localchange in the flooding regime. It is difficult to identifythe exact cause of such changes. For Instance, the chrono-logy of past climate change in the Pantanal is not wellunderstood (Ab'Saber 1988), although there is evidence
of past climatic fluctuations, from drier to wet regimesand vice-versa, a couple of times during the Quaternary(Alvarenga cl al. 1984). Obviously, a change caused by aclimatic fluctuation would equally affect both areas, asthey are adjacent to each other. Eor regions where theclimate has become drier for a period, as in the Amazon,it is expected that the establishment of a C4-rich vegetationis facilitated (van der Hammen 1974; Absy & van derHammen 1976; Absy 1980; Markgraf 1989; Bush ct al.1990; Absy et al. 1991).
Soil organic matter is often modelled as a two- or three-component system, a mixture of more rapidly cyclingfractions with turnover times ranging from less thanone year to decades, and passive organic matter, withmillennial or longer turnover times (Harrison et al. 1993;Jenkinson & Raynor 1977; O'Brien & Stout 1978; Partonel al. 1987; Schimel et al. 1994; Townsend cl al. 1995;Trumbore 1993). Trumbore et al. (in press) have shownthat this picture holds true even at depths > 1 m intropical soils, as deep roots feed an active carbon cycle(turnover times of < 40 years) while the bulk of thecarbon is in old, refractory organic carbon. Thus, theAMRT associated with a ^ C age, even in deep soils ismost likely a mixture of decadal and faster cycling organicmatter (with '^C values > Modern) and old, refractorycarbon (with '"'C values substantially less than Modern).In a soil profile where vegetation has changed with timefrom CJ to C4-dominated plants, we assume that the ' \l!changes in the SOM may be used as a measure of theminimum amount of rapidly cycling carbon (as thesehave adjusted to a new steady state within a centuryof conversion), and that residual '^C-depleted carbonrepresents the passive carbon accumulated over millenniain the old vegetation regime. We used this approach tocalculate the possible timing of vegetation change inthis specific site of Pantanai. The change from arborealsavanna to grassy savanna (less negative S' C towardsthe soil surface) can be analysed on the results from thegrassy savanna soil profile. We may assume that thecarbon present at the 110-120 cm depth is a mixture ofcarbon of different ages: an old carbon fraction, originatedfrom C3 plants (AMRT > 4572 BP, which is the estimatedage at this depth interval in the grassy savanna profile),and a younger fraction derived from C4 plants, addedsince the last vegetation change, and therefore withAMRT < 4572 BP. Therefore, the minimum AMRT of theold C3 carbon at the 110-120 cm would be c. 4572 BP, andno mixture would have taken place.
The maximum AMRT can be calculated through thefollowing ''C mass balance:
where
%C3-(Al^old,,rt,on) (4)
i|.carbon is the *C activity at the 110-120 cm
1995 Blackwell Science Ltd., Global Change Biology, 1, 165-171

170 R.L. VICTORIA ct al.
soil depth interval in the grassy savarma profile (-435 %o),''* new-carbon is the '*C activity of the new C4 carbon,
taken as the '' C activity at the soil surface in the grassysavanna (+ 141 %«), and A' 'Coij .j-arbon is the ""C activityof the old C3 carbon that we want to estimate. The %C4and %C3, the relative percentage of C4 and C3 carbon inthe soil profile, were estimated by '''C mass balance,through the following equation:
100, (5)
where S Igampie is the carbon isotopic composition ofthe soil at the 110-120 cm depth in the grassy savanna(-22.3%i>), 8^^c4 is the carbon isotopic composition ofthe C4 material that is being added to the soil (-12.4 'Hm,or the * C isotopic composition of the C4 vegetationcover), and 5' Cc3 is the carbon isotopic composition ofthe C3 material in the soil (-27.9 %o, assumed to be 2 %oless negative than the average isotopic composition of C3plants,-29.9±1.4 '%«>, n = 29). The proportion of C4 plantsis 100 -%C3.
Using (4) and (5) we calculated a maximum AMRTequal to 11 420 BP. Therefore, the vegetation change fromarboreal to grassy savanna probably occurred between4572 and 11 420 BP. The two major dry periods reportedto have occurred in tropical and sub-tropical regions ofSouth America were from 4000 to 5000 BP, and from10 400 to 10 500 BP (Absy & van der Hammen 1976; Absy1980; Absy et ai 1991; Servant el ai 1993), which agreewith the range found in the Pantanal soil.
The same approach was used to estimate the timingof the change from grassy savanna to arboreal savanna,indicated by the results of the 50-60 cm depth intervalin the arboreal savanna soil profile (Fig. 3). In this case,however, as the C3 carbon increased 50-60 cm depthtowards the soil surface, old carbon was assumed to be ofC4 origin, and new carbon of C3 origin. In this case thedata was analysed based on the results of the 30-40 cmdepth interval in the arboreal savanna soil profile.The following values were used in (4): + 51 %c. for
S'*Cs,,jiH:aHx.n. + 141 %o for 5^ *Cnew-carbon- 3 " ^ 5'*Co Id-carbon
is the ''*C activity of the old C4 carbon that we want toestimate. In (5), the same values of S^^C for vegetationtypes were used, and the S C of the soil samples at 50-60 cm depth inten.'al was equal to -21.1 'J(» The maximumAMRT was then estimated to be equal to 1390 BP, sug-gesting that the change from grassy savanna to arborealsavanna was a recent local event.
In parallel with suggestions for other areas in the world(e.g. MacPerson et al. 1993), wtxKiland is expanding overthe grassland at this specific site of the Pantanal. However,we also observed that the highly dynamic characteristicof the flooding regime may cause large variations in the
stable carbon isotopic composition, and in the radiocar-bon age as well. Therefore, the results presented hereshould not be extrapolated to other areas of the Pantanalregion, without further studies. Overall, the findingsreveal the dynamic nature of the spatial distribution ofC3 and C4 savanna sub-communities and the coincidenceof vegetation change with periods ot climate change.
References
Ab'Saber AN (1988) O Pantanal Mato-Grossense ea Teoria dosReftigios. Revista Brasiteira de Geografia, 50, 9-57.
Absy ML, Van der Hammen T (1976) Some paleo-ecological datafrom Rondonia, southern part of Amazonian Basin. ActaAniaztmiai, 6, 29^299.
Absy ML (1980) Dados sobre as mudan^as do clima e davegetaijao da Amaz6nia durante o Quatemdrio, ActaArmzonica, 10, 929-.
Absy ML, Cleef A, Fournier M cf al. (19^1) Mise en evidence dequatre phases d ouverture de la foret dense dans Ie sud-estde 1 Amazonie au cours des 60000 demi^res antics. Premierecomparaison avec d'autres regions tropicales. Comptes Rendusde I'Aaademie des Sciences de Paris Serie ll. 312, 675-678.
Ad moli JM (1982) O Pantanal e suas relaq6es fiotogeograficascom OS cerrados, Discussao sobre o conceito 'ComplexoPantanal'. In: Proceedings of Cotigresso Brasiteiro de Bot^ttica,pp. 109-119. Sociedade Brasileira de Botanica, Teresina, Brazil.
Alvarenga SM, Brasil AE, Pinheini R, Kux HJH (1984) Kstudogeomorfolcgico aplicado a Bacia do Alto Paraguai e PantanaisMato-Grossenses. Boletim Tecnico no. 1 - ProjetoRadamBrasil - S^rie Geomorfologia.
Ambrose SH, Sikes, NE (1991) Soil carbon isotope evidence forHolocene habitat change in the Kenya Rift Valey. Science, 253,1402-1405.
Bush MB, Coiinvaux PA, Wiemann C, Pipemo DR, Liu KB (!*«())Latt' Pleistocene Temperature Depression and VegetationChange in Ecuadorian Amazonia. Quaternary Research, 34,330-345.
Desjardins T, Volkoff B, Andreux F, Cerri C (1991) Distributiondu carbone total et de I'isotope '•'C dans des sols ferrallitiquesdu Bresil, Sciaice du Sol, 29, 175-187,
Dzurec RS, Boutton TW. Caldwell MM, Smith BN (1985) Carbonisotope ratios of soil organic matter and their use in assessingcommunity composition changes in Curlew Valley, Utah.Oecologia, 66, 17-24.
Farquhar C, O'l,eary M, Berry ) (1982) On the relationshipbetween carbon isotope discrimination and the intercellularcarbon dioxide concentration in leaves. Australian fournal ofPlant Physiology, 9, 121-137.
Flexor JM, Volkoff B (1977) Distribution de I'isotopt' stable ' Cdans la matiere organique d'un sol ferralitique de I'etat deBahia (Bresil). Comptes Retidus df lAcademie des Sciences deParis. 284 s*rie D, 1655-1657.
Guillet B, Faivre P, MariotH A, Khobzi J (1988) The ''•C datesand ' • ' C / ' ^ ratios of soil organic matter as a means ofstudying the past vegetation in intertropical regions:examples from Colombia (South America). Palaeogeography,Pataeoclimatology. Palaeoecology, 65, 51-58.
1995 Blackwell Science Ltd., Global Change Biology, 1,165-171

PAST VEGETATION CHANGE IN THE PANTANAL 171
Harrison K, Broecker W, Bonani G (1993) A strategy forestimating the impact of COj fertilization on soil carbonstorage. Global Biogeochemical Cycles, 7. 69-80,
Jenkinson DJ, Raynor JH (1977) The turnover of soil organicmatter in st)me of the Rothamsted classical experiments. SoilScifijci; 123, 298-305.
Junk WJ (1993) Wetlands of Tropical South America. In: Wetlandsof the World I (ed. Whigham DF), pp. 679-739. Kluwer,Dordrecht.
Kapos V, Ganade G, Matsui E, Victoria RL (1993) 5 - C as anindicator of edge effects in tropical rainforest reserves, journalof Ecology, 81. 425-43,2.
Loureiro RL, Souza Lima JP, Fonzar BC, Oliveira Filho, LC (1982)Vegeta^ao, As Regi6es Fitoecologicas, sua Natureza e seusRecursos Economicos. in: Projeto RadamBrasil. Vol. 27, FolhaSE.2L Corumb, pp. 329-364. Ministerio das Minas eEnergia, Brasilia.
Lugo AE, Brown S, Brinson MM (1990) Concepts in WetlandEcology. In: Forested Wetlands - Ecosystems of the World 15(eds Lugo AE, Brinson MM, Brown S), pp. 53-85. Elsevier,Amsterdam.
Mariotti A, Petershmitt E (1994) Forest savanna ecotonedynamics in India as revealed by carbon isotope ratios ofsoil organic matter. Oecohgia, 97, 475-480.
Markgraf V (1989) Paleoclimates in Central and South AmericaSince, 18, 000 BP Based on Pollen and Lake-Level Records.Quaternary Science Reviews, 8, 1-24.
Martin A, Mariotti A, Balesdent J, Lavelle P, Vuattoux V (1990)Estimate of Organic Matter Turnover Rate in Savanna Soilby -'C Natural Abundance Measurements. Soil Biology andBiochemistry, 22, 517-523.
MarHnelli LA, DeVol. AH, Victoria RL, Richey JE (1991) Stablecarbon isotope variation in Ci and C4 plants along tbeAmazon River. Nature, 353, 57-59.
McPherson GR, Boutton TW, Midwood AJ (1993) Stable carbonisotope analysis of soil organic matter illustrates vegetationchange at the grassland/wot^idland boundary in southeasternArizona, USA. Oecohgia, 93, 95-101.
Mondenesi MC, Matsui E, Volkoff B (1986) RelaijSo ' ^ C / ' ^ nosliorizontes humiferos superficiais e nos horizontes escurosprofundos dos solos de campo e mata da regi3o de Camposdo Jordao, Sao Paulo, Brasil, In; Regional Colloquium onSoil Organic Matter Studies, pp, 155-160. Centro de EnergiaNuclear na Agricultura, Piracicaba, Brazil.
Moraes JF (1991) Conteudo de Carbono e Uitrog^nio e Tipohgiade Horizontes nos Solos da Bacia Amazonica. Master thesis,Universidade de Sao Paulo, Piracicaba, Brazil, 84 pp.
Moreira A AN (1977) Relevo da RegiSo CentroOeste. In: Geografiado Brasil - Regiao CentroOeste (ed, Galvao MV), pp. 1-33.IBGE, Rio de Janeiro,
Moreira Franco MS, Pinheiro R (1982) Geomorfologia, In: ProjetoRadamBrasil, Vol, 27, Folha SE.21, Corumba, pp, 161-224.Ministerio das Minas e Energia, Brasilia.
O'Brien BJ, Stout JD (1978) Movement and turnover of soilorganic matteras indicated by carbon isotopic measurements.Soil Biology ami Biochemistry, 10, 309-317,
Parton WJ, Schimel DS, Coie CV, qima DS (1987) Analysis offactors controlling soil organic matter levels in Great Plainsgrasslands. Soi7SdewcfS(ifit*(yoM"'fnV-rt/()»n-((j/, 51,1173-1179,
Pott V, Pott A, Ratter JA, Walls JFM (1986) Rora da FazendaNhumirim, Nbecolandia, Pantanal. Rela^ao Preliminar. In:Serie Pesquisa em Andamento n 5, Embrapa/CPAP, Corumba,26 pp.
Schimel DS, Braswell BH, Holland EA, McKeown R, OjimaDS, Painter TH, Parton WJ, Townsend AR (1994) Climatic,edaphic and biotic controls over storage and turnover ofcarbon in soils. Global BiogetKhemical Cycles, 8, 279-294,
Schleser GH, Jayasekera R (1985) 6'^C-variations of leaves inforests as an indication of reassimilated CO2 from the soil.Oecohgia, 65, 536-542.
Schwartz D, Mariotti A, Lanfranchi R, Guillet B (1986) ' ^ / ^ ^ratios of soil organic matter as indicators of ecosystemschanges in the Congo. Geoderma, 39, 97-103,
Servant M, Maley J, Turcq B, Absy ML, Brenac P, Foumier M,Ledru MP (1993) Tropical forest changes during the LateQuaternary in African and South American lowlands.Paleogeography, Paleoclimatohgy, Palaeoecology, 7, 1--16.
Southon J, Vogel JS, Trumbore SE (1992) Progress in AMSmeasurements at the LLNL spectrometer, Radiocarbon, 34,473-477,
Stemberg LSL, Mulkey SS, Wright SJ (1989) Ecologicalinterpretation of leaf carbon istitope ratios: influence ofrespired carbon dioxide. Ecology, 70, 1317-1324.
Stuiver M, Pearson GW (1986) High-precision calibration of theraditKarbon timescale, AD 1950-500 BC, Radiocarbon, 28,805-838,
Stuiver M, Polach H (1977) Reporting of 14C data. Radiocarbon,19, 355-363.
Townsend AR, Vitousek PM, Trumbore SE (in press) Soil andorganic matter dynamics along gradients of temperature andland-use. Ecology.
Trouve C, Mariotti A, Scwartz D, Guillet B (1994) Soil organiccarbon dynamics under Eucalyptus and Pinus planted onsavannas in the Congo, Soil Biology and Biochemistry, 26,287-296.
Trumbore, S.E. (1993) Comparison of carbon dynamics in twosoils using measurements of radiocarbon in pre-and post-bomb soils. Global Biogeochemicat Cycles, 7, 275-290.
Trumbore SE, Davidson EA, Camargo P, Nepstad D, MartinelliLA (in press) Below-ground cycling of carbon in forests andpastures of eastern Amazonia. Global Biogeochewical Cycles
van der Metwe NJ, Medina E (1989) Photosynthesis and ' ^ C / ' ^rations in Amazonian rain forests. Geochimica et CosmochimicaActa, 53, 1091-1094.
van der Hammen T (1974) The Pleistocen changes of vegetationand climate in tropical South America, Joumal of Bhgeography,1, 3-26,
Vitorello VA, Cerri CC, Andreaux F, Feller C, Victoria RL(1989) Organic Matter and Natural Carbon-13 Distributionin Forested and Cultivated Oxisols, American Soil ScienceSociety lourtial, 53, 773-778,
Volkoff B, Cerri CC (1987) Carbon isptopic fractionation insubtropical Brazilian grassland soils. Comparison withtropical forest soil. Plant and Soil, 102, 27-31.
Wang Y, Cerling TE, Effand WR (1993) Stable isotope ratios ofsoil carbonate and soil organic matter as indicators of forestinvasion of prairie near Ames, Iowa. Oecohgia, 95, 365-369,
© 1995 Blackwell Science Ltd., Global Change Biology, 1, 165-171
