orbital and ventromedial prefrontal cortex functioning in parkinson’s disease: neuropsychological...
TRANSCRIPT

Brain and Cognition 79 (2012) 23–33
Contents lists available at SciVerse ScienceDirect
Brain and Cognition
journal homepage: www.elsevier .com/ locate /b&c
Orbital and ventromedial prefrontal cortex functioning in Parkinson’s disease:Neuropsychological evidence
Michele Poletti, Ubaldo Bonuccelli ⇑Department of Neuroscience, University of Pisa, Pisa, Italy
a r t i c l e i n f o
Article history:Accepted 3 February 2012Available online 3 March 2012
Keywords:Parkinson’s diseaseOrbital prefrontal cortexVentromedial prefrontal cortexReversal learning taskIowa Gambling TaskAffective Theory of Mind
0278-2626/$ - see front matter � 2012 Elsevier Inc. Adoi:10.1016/j.bandc.2012.02.002
Abbreviations: GPi, internal segment of globus papars reticulata; VP, ventral pallidum; MD, medialdorsalis pars parvocellularis; MDmc, medialis dorsalisventralis anterior pars magnocellularis; VApc, ventralaris; VLo, ventralis lateralis pars oralis; VLm, ventracaudolateral; ldm, lateral dorsalmedial; mdm, mediamedial; rd, rostrodorsal; rl, rostrolateral; rm, rostrom⇑ Corresponding author. Address: Dipartimento di
Pisa, Via Savi 10, 56100 Pisa, Italy.E-mail address: [email protected] (U. Bon
a b s t r a c t
A recent paper (Zald & Andreotti, 2010) reviewed neuropsychological tasks that assess the function of theorbital and ventromedial portions of the prefrontal cortex (OMPFC). Neuropathological studies haveshown that the function of the OMPFC should be preserved in the early stages of Parkinson’s disease(PD) but becomes affected in the advanced stages of PD. This pattern has also been suggested by studiesthat have shown that dopaminergic drugs impair the performance of early PD patients in OMPFC tasksthat involve reinforcement learning but enhance the performance of advanced PD patients. Based onthese empirical findings, we reviewed the neuropsychological evidence of OMPFC functions in PDpatients to test two hypotheses regarding the following: (1) OMPFC functions at different stages of PD;(2) different effects of dopaminergic drugs on OMPFC functions based on PD stage and task demand.We focused our review only on the neuropsychological tasks that were specific and sensitive to the func-tions of the OMPFC and that were adopted at different stages of PD, such as reversal learning tasks, theIowa Gambling Task and the affective Theory of Mind task. We found robust empirical evidence that inearly PD, OMPFC functions are preserved and dopaminergic drugs result in a detrimental effect when thetask involves reinforcement learning. Further studies are needed to verify the status of OMPFC functionsin non-demented, advanced PD and to describe the longitudinal course of OMPFC functions in this clinicalpopulation.
� 2012 Elsevier Inc. All rights reserved.
1. Introduction
A recent paper (Zald & Andreotti, 2010) reviewed the neuropsy-chological tasks that permit assessment of cognitive functions ofthe orbital prefrontal cortex (OFC) and of the ventromedial pre-frontal cortex (VMPFC). The following cognitive domains were re-viewed: (1) the ability to change prepotent or previouslyacquired stimulus–reinforcer associations; (2) olfactory process-ing; (3) social processing; (4) autobiographical memory; and (5)emotional characteristics of personality. Different tasks were ana-lyzed within each functional domain, which demonstrated that dif-ferent tasks are supported by varying levels of neuropsychological
ll rights reserved.
llidus; SNr, substantia nigrais dorsalis; MDpc, medialispars magnocellularis; VAmc,lis anterior pars parvocellu-
lis lateralis pars medialis; cl,l dorsomedial; pm, postero-edial.Neuroscienze, Università di
uccelli).
evidence with regards to their sensitivity and specificity to dam-ages to the OFC and VMPFC.
Parkinson’s disease (PD) is a progressive neurodegenerative dis-order diagnosed based on characteristic motor disturbances (bra-dykinesia, resting tremor, rigidity and postural instability),asymmetrical motor onset and good response to levodopa (Litvanet al., 2003). As a synucleinopathy, PD is linked to the pathogenicfibrillization of the unstructured soluble protein, a-synuclein. Itis also characterized by the formation of Lewy bodies in nigral re-gions, limbic and brainstem nuclei and neocortical regions (Kala-itzakis & Pearce, 2009), although neurofibrillary tangles andplaques are also commonly present in these areas (Braak et al.,2003). The resulting neuronal degeneration directly affects cate-cholaminergic (i.e., dopamine and norepinephrine) and cholinergic(acetylcholine) neurotransmission (Bohnen et al., 2006; Brooks &Piccini, 2006; Calabresi, Picconi, Parnetti, & Di Filippo, 2006).
Cognitive impairments are frequently reported in patients withPD, beginning in the untreated early clinical stages (Aarsland et al.,2010). They principally involve executive functions, based on theDorsolateral Prefrontal Cortex (DLPFC) (for review, see Cools,2006; Koerts, Leenders, & Brouwer, 2009, or Poletti, Emre, & Bonuc-celli, 2011). Cognitive impairment in early PD is related to the pro-gressive loss of dopaminergic neurons in the substantia nigra,

24 M. Poletti, U. Bonuccelli / Brain and Cognition 79 (2012) 23–33
which is an essential component of the basal ganglia circuitry(Kehagia, Barker, & Robbins, 2010). On the other hand, dementiais a common feature in the advanced stages of PD and is relatedto the cortical diffusion of Lewy bodies (Emre et al., 2007; Kalaitza-kis & Pearce, 2009). While the impairment of DLPFC functions is awell-established feature of PD patients, the status of OFC andVMPFC functions in this clinical population has been investigatedless thoroughly.
This paper aims at providing an up-to-date review of currentneuropsychological evidence about functions related to the OFCand VMPFC in patients with PD. Here, we first present the anatomyof the OFC and VMPFC in healthy subjects and in PD patients (inSection 2), as well as show models of the interaction betweendopamine depletion (PD stage) and the dopaminergic replacementtherapy (in Section 3). Secondly, we formulate working hypotheseson the functioning of the OFC and VMPFC and on the effect of dopa-minergic therapies on these functions in PD (in Section 4). Then,we present a critical literature review (in Section 5), and we pres-ent empirical findings on OMPFC functions in PD (in Sections 6 and7). Finally, in Section 8 we discuss the results of the literature re-view to test working hypotheses on OMPFC functions in PD andsuggest future research directions.
2. Anatomy of the OFC and the VMPFC
The OFC and VMPFC include the medial portions of Brodmann’sareas 9, 10, 11, and 12, areas 13 and 25 and the inferior portion ofarea 47 (Damasio, 1996). The OFC comprises the ventral surface ofthe PFC, while the VMPFC comprises the inferior portion of themedial wall of the frontal lobe. Although there are cyto-architec-tural and connectional differences between the OFC and theVMPFC (Carmichael & Price, 1996; Price, 2006), pathologies that af-fect the PFC usually damage both of these areas. In fact, in studiesdescribing patients with OFC damages, VMPFC damages are alsooften present and vice versa. For this reason, while electrophysio-logical methods in monkeys may detect differences of activation inthese areas (e.g., Bouret & Richmond, 2010), neuropsychologicalstudies in brain-damaged patients failed to find clear functionaldifferences between these areas and to disentangle their relativecontributions in isolation. Therefore, the term OMPFC could beadopted to refer to the combination of the OFC and VMPFC (Zald& Andreotti, 2010). For the purpose of this review, we would liketo emphasize that the OMPFC is connected to limbic structures,such as the amygdala and the hypothalamus. The DLPFC, on theother hand, is an important source of cortico-striatal inputs tothe nucleus accumbens, which reciprocally modulates the activityof the midbrain dopaminergic neurons and is itself an importantnode in the circuit that processes reinforcement (Damasio, 1996;Price, 2006; Rolls, 2004).
2.1. The OMPFC in Parkinson’s disease
Four frontostriatal loops are involved in the motor, cognitive,affective and motivational aspects of behavior (Fig. 1) (Alexander,Delong, & Strick, 1986; Middleton & Strick, 2001; Yeterian & Pand-ya, 1991) as follows: the ‘‘dorsolateral’’ loop includes the Dorsolat-eral Prefrontal Cortex (DLPFC), the striatum (dorsolateral caudatenucleus and dorsolateral putamen), the globus pallidus (dorsome-dial) and the thalamus; the ‘‘orbital’’ loop includes the orbitofron-tal cortex (OFC), the striatum (ventromedial caudate nucleus andventral putamen), the globus pallidus (dorsomedial) and the thal-amus; the ‘‘anterior cingulate’’ loop includes the anterior cingulatecortex (ACC), the striatum (ventromedial caudate nucleus and ven-tral putamen), the nucleus accumbens, the olfactory tubercle, theglobus pallidus (rostromedial) and the thalamus; and the ‘‘motor’’loop includes the supplementary motor area, the putamen, the glo-
bus pallidus (ventrolateral) and the thalamus. Additionally, withineach circuit, two loops connect the striatum with the PFC: a directexcitatory loop and an indirect inhibitory loop (Alexander, Crut-cher, & DeLong, 1990).
With regards to the neuropathological staging of PD, we refer toBraak et al. (2003). In PD patients, the progressive striatal dopa-mine depletion has different effects on the functioning of thefrontostriatal loops. In early PD (Braak stages 3 and 4), the dopa-mine depletion is greatest in the ventrolateral tier of the substantianigra pars compacta, which projects primarily to the dorsal stria-tum (i.e., the dorsolateral putamen and the dorsal parts of the cau-date nucleus), an area involved in the dorsolateral loop. The ventralstriatum and the related orbital loop, however, are mostly pre-served (Kish, Shannak, & Hornykiewicz, 1988), as Lewy bodies donot impair the PFC in these stages of PD (Braak et al., 2003). Withthe progression of PD, dopamine depletion at the striatal level alsoimpairs the functioning of the orbital loop. Moreover, in thesestages of illness, the PFC is directly damaged by the cortical diffu-sion of Lewy bodies (Braak et al., 2003; Scatton, Rouquier, Javoy-Agid, & Agid, 1982). Thereafter, in advanced PD (Braak stages 5and 6), OMPFC functions can be impaired by two mechanisms:(1) indirectly, by the reduced striatal dopaminergic stimulationand (2) directly, by the cortical diffusion of Lewy bodies.
3. The interaction between dopamine depletion anddopaminergic therapy in PD
Levodopa and dopamine agonists represent the gold-standardtherapy for treating the motor symptoms of PD patients (Bonuccel-li & Pavese, 2006; Poewe, Antonini, Zijlmans, Burkhard, & Vinge-rhoets, 2010). How does the dopaminergic therapy interact withthe progressive striatal dopamine depletion? Different models onthis topic have been proposed. The inverted U-shape curve model(Cools, 2006) proposed that, in early PD patients, the withdrawalof dopaminergic medication has a detrimental effect on cognitivefunctions associated with the dorsolateral loop, and a beneficial ef-fect on the cognitive functions associated with the orbital loop. Aslevodopa mainly elevates dopamine levels in the striatum (Horn-ykiewicz, 1974), these differential effects are likely due to opposingeffects of levodopa in the dorsal and the ventral striatum, whichare connected to different cortical areas via segregated frontostri-atal loops (Alexander et al., 1986). This double dissociation is evi-dent when directly comparing patients ‘‘on’’ and ‘‘off’’dopaminergic medication and represents an empirical confirma-tion of the ‘‘dopamine overdose hypothesis’’ (Gotham, Brown, &Marsden, 1986; Gotham, Brown, & Marsden, 1988). This hypothe-sis states that the administration of dopaminergic medication toearly PD patients may replete dopamine-depleted circuits (includ-ing the dorsal striatum), thus improving performances in tasks re-lated to the dorsolateral loop while ‘‘overdosing’’ relatively intactcircuits (including the orbital loop).
In the neurocomputational model of frontostriatal circuitry inPD proposed by Frank and colleagues (2004), the basal gangliamodulate the selection of actions under consideration in the PFC.Two main projection pathways from the striatum travel up tothe cortex through the thalamus via different basal ganglia outputstructures. The subthalamic nucleus provides a self-adaptive, dy-namic control signal that temporarily prevents the execution ofany response, depending on decision conflict (Frank, 2006). The di-rect frontostriatal ‘‘orbital’’ pathway is excitatory and the indirectfrontostriatal ‘‘orbital’’ pathway is inhibitory. Transient changesin dopamine levels that occur during positive and negative feed-back loops have opposite effects on the D1 and D2 (dopamine)receptors, which are relatively segregated in the direct and indirectpathways, respectively (Hernandez-Lopez et al., 2000). Dopamine

Fig. 1. Parallel organization of functionally segregated circuits connecting the frontal cortex and striatum, based on the original scheme by Alexander et al. (1986) andadapted by Chudasama and Robbins (2006). Four frontostriatal ‘loops’ are shown. Each ‘loop’ starts in a specific region of the frontal cortex and innervates different levels ofthe striatum before the signal is relayed back to its cortical origin, via the thalamus. NB: short, thin, black arrows indicate dopaminergic (DA) innervation of the cortex andstriatum. The diagram to the left of the dashed line indicates the general organization of the cortico-striatal-pallidal-thalamic loop.
M. Poletti, U. Bonuccelli / Brain and Cognition 79 (2012) 23–33 25
bursts during positive reinforcement activate the direct pathwayand deactivate the indirect pathway, driving learning so that rein-forced responses are subsequently facilitated. Conversely, de-creases in dopamine result in negative feedback, or deactivation,of the direct pathway and activation of the indirect pathway. Thus,unreinforced responses are subsequently suppressed or avoided.This model predicts a stronger processing of positive rewards inmedicated PD patients, since levodopa increases dopaminergicbursts and facilitates an excitatory activity in the direct pathwayof the cortico–striato–thalamo–cortical loops. Otherwise, medi-cated PD patients should show a decreased ability to learn throughthe mechanism of reward omission. This is because levodopa pre-vents dips in dopaminergic systems, which disturbs the inhibitoryactivity of the indirect pathway in the cortico–striato–thalamo–cortical loop. Unmedicated patients should show the opposite pat-tern, learning sufficiently from negative feedback to avoid harm,while showing impairment in learning from positive reinforce-ment. This neurocomputational model has been empirically con-firmed by administrating a probabilistic selection task to PDpatients ‘‘on’’ and ‘‘off’’ dopaminergic medication (Frank, Samanta,Moustafa, & Sherman, 2007). Levodopa altered the patients’ ten-dency to learn from positive versus negative outcomes, withoutmodifying conflict-induced slowing.
In the tonic/phasic model of dopamine system regulation (Goto,Otani, & Grace, 2007; Grace, 2000), the nucleus accumbens (NAccs)is believed to regulate goal-directed behavior because it receivesconvergent synaptic inputs from limbic structures and the PFC. Thus,the NAcc is located such that contextual information from the hippo-campus and emotional information from the amygdala, could beintegrated with actions programmed in the PFC (Grace, 2000). Elec-trophysiological experiments in rats showed that tonic and phasicdopamine release selectively modulates hippocampal and prefron-tal cortical inputs through the D1 and D2 receptors, respectively.In addition, D1 activation and D2 inactivation in the NAcc producesbehaviorally selective effects (learning versus set-shifting of re-
sponse strategy) that correspond to specific afferents. These resultssuggest that the dynamics of dopamine release regulate the balancebetween the limbic and cortical drives through activation and inac-tivation of specific dopamine receptor subtypes in the NAcc, and thisregulates goal-directed behavior (Goto & Grace, 2005a; Goto &Grace, 2005b). For a recent review of the role of the dopaminergicsystem in reinforcement learning see Glimcher (2011).
These results are also consistent with the results of Cools, Alta-mirano, and D’Esposito (2006), who demonstrated at a behaviorallevel that dopaminergic medication impairs reversal learning inpatients with mild PD. The same group clarified the neural mech-anism underlying this impaired reversal learning caused by dopa-minergic therapy in PD patients in a fMRI study (Cools, Lewis,Clark, Barker, & Robbins, 2007). PD patients who were ‘‘on’’ or‘‘off’’ L-dopa medication had their brain activity measured by fMRIwhile performing a probabilistic reversal learning task able to acti-vate the ventral striatum and the orbital frontostriatal circuit(Cools, Clark, Owen, & Robbins, 2002). fMRI data showed a roleof the NAcc in the dopaminergic modulation of reversal learningin patients with mild PD. Reversal learning was accompanied byan increased NAcc activity only when patients were ‘‘off’’ theirdopaminergic therapy. Upon resuming therapy, reversal learningwas disrupted due to changes in the functioning of the NAcc. Assuggested by the authors, (Cools et al., 2007) future studies arenecessary to address the pharmacological mechanisms underlyingthe medication-induced reversal impairment. In particular, studiesin patients with severe PD accompanied by a loss of dopamine inthe NAcc, will reveal whether the L-dopa-induced deficits in pa-tients with mild PD depend on the level of dopamine depletionin the NAcc. Whereas other accounts of the medication-inducedimpairment do not require the NAcc to be intact (Frank, Seeberger,& O’Reilly, 2004), it is possible that the impairment could be abol-ished during progression of the disease. Therefore, in an overdosedorbital loop, the dopaminergic replacement therapy prevents thedips in those dopaminergic systems that support the ‘‘No Go’’

Table 1Modified Hoehn And Yahr staging of Parkinson’s disease.
Stage 0 – no signs of diseaseStage 1 – unilateral diseaseStage 1.5 – unilateral and axial involvementStage 2 – bilateral disease, without impairment of balanceStage 2.5 – mild bilateral disease with recovery on pull testStage 3 – mild to moderate bilateral disease; some postural instability;
physically independentStage 4 – severe disability; still able to walk or stand unassistedStage 5 – wheelchair bound or bedridden unless aided
26 M. Poletti, U. Bonuccelli / Brain and Cognition 79 (2012) 23–33
learning through the indirect pathway of the cortico–striato–thal-amo–cortical loop. This phenomenon likely causes dysfunctionalreward processing, which impairs learning from reward omission(Frank et al., 2007). Moreover, considering that the phasic-actingL-dopa needed to restore dopaminergic bursts effaces dopaminer-gic dips during reinforcement learning, while tonic-acting dopa-mine agonists should impair both dopaminergic bursts and dips,the question remains as to whether L-dopa and dopamine agonistshave different effects on reinforcement learning. Further studieswith the manipulation of dopaminergic treatments (L-dopa versusdopamine agonists; comparison of different dopamine agonists)are needed on this topic.
Among tasks that assess OMPFC functions (Zald & Andreotti,2010), some involve reinforcement learning processes (reversallearning tasks and decision making tasks), while others do not(olfactory processing; social processing; autobiographical mem-ory; emotional characteristics of personality). Considering the ef-fects of dopaminergic therapies on reinforcement learning, it ispossible that performances in tasks that involve this process aremore influenced by dopaminergic therapies than tasks that do not.
4. OMPFC functioning in PD: working hypotheses
Evidence from anatomical and neuropathological studies sug-gests that OMPFC functions could be preserved in the early clinicalstages of PD but might be impaired in the advanced stages of PD.Moreover, dopaminergic therapy could impair OMPFC functionsin the early clinical stages of PD but may improve OMPFC functionsin the advanced stages of PD. Therefore, we hypothesize that per-formances on neuropsychological tasks that assess OMPFC func-tions will worsen with the progression of PD (Hypothesis 1). Inparticular, we believe that these functions are preserved in theearly stages of PD (Sub-Hypothesis 1A) but become impaired inthe advanced stages of PD (Sub-Hypothesis 1B). We also hypothe-size that dopaminergic drugs have different effects on OMPFCfunctions, specifically in those tasks that involve reinforcementlearning, depending on the PD stage (Hypothesis 2). We alsohypothesize that dopaminergic drugs impair OMPFC functions inthe early stages of PD (Sub-Hypothesis 2A) but enhance OMPFCfunctions in the advanced stages of PD (SubHypothesis 2B).
We adopted the Hoehn and Yahr Staging Scale (H&Y: Hoehn &Yahr, 1967) to define the operative criteria that distinguish theclinical stages of PD. The modified version of this scale describesthe progression of PD in seven stages (Table 1). Following sugges-tions of the Movement Disorder Society Task Force Report on theH&Y (Goetz et al., 2004), we defined the clinical stages of PD asearly (H&Y Stages 1, 1.5 and 2), moderate (H&Y Stage 2.5 and 3)and advanced (H&Y Stages 4 and 5). Early PD stages and advancedPD stages are well established, although, as the authors noted, ‘‘it isless clear that Stage 3 operationally defines moderate PD, because thehallmark of the Stage 3, the occurrence of balance difficulties, hasnever been defined in a standardized manner and no systematic appli-cation of the objective pull test and teaching tape have been devel-oped’’ (Goetz et al., 2004, pp. 1026).
5. Criteria of the literature review
Before performing a search in any electronic databases, we se-lected those neuropsychological tasks that were more specificand sensitive to OMPFC dysfunctions (Zald & Andreotti, 2010).The Wisconsin Card Sort Task (WCST: Grant & Berg, 1948) andthe response inhibition task were excluded because they are notclearly related to the OMPFC, but rather, they relate to other pre-frontal regions, specifically the DLPFC for the WCST (Dias, Robbins,& Roberts, 1996) and the dorsomedial and ventrolateral PFC for re-
sponse inhibition tasks (Simmonds, Pekar, & Mostofsky, 2008). Inregards to social cognition tasks, only the Theory of Mind taskswere considered, while emotion recognition tasks were excluded.In fact, although fMRI studies show that the OMPFC may be acti-vated by emotion recognition tasks (Dougherty, Shin, & Rauch,2006), the importance of the OMPFC to emotional recognition ofstimuli remains unresolved (Zald & Andreotti, 2010). Moreover,emotion recognition tasks could be differentially sensitive to thelocation of the OMPFC lesions, although the methodological differ-ences across existing studies have precluded firm conclusionsabout sub-regional localization of emotional recognition(Heberlein, Padon, Gillihan, Farah, & Fellows, 2008). With regardto PD patients, although they present specific deficits in emotionrecognition (Gray & Tickle-Degnen, 2010), a recent review sug-gested that these deficits, especially for disgust, are more relatedto subcortical structures, such as the amygdala and basal ganglia,than to the OMPFC (Assogna, Pontieri, Caltagirone, & Spalletta,2008).
Moreover, olfactory processing tasks were also excluded fromour review, since they are not specific in the detection of OMPFCdysfunctions in PD patients. As a matter of fact, odor identificationand odor discrimination deficits occur at the earliest stages of PD,often preceding the clinical motor onset, due to the presence ofLewy bodies in the olfactory bulb (for review see Benarroch,2010; Haehner, Hummel, & Reichmann, 2009). Therefore, earlydeficits of PD patients in these tasks may not be attributed to theOMPFC, but rather, to the dysfunction of other subcortical and cor-tical structures that precede the OMPFC in the bottom-up neuralnetworks involved in olfactory processing. Overall, we excludedmany tasks from our review that were not specific to OMPFC dys-functions (WCST, response inhibition tasks, emotion recognitiontasks) or because they were not suitable for the detection ofOMPFC dysfunctions in PD (olfactory processing tasks). This choicemay have reduced the extent of this review but hopefully will re-sult in more robust conclusions on OMPFC functioning in PD.
Next, we performed a search on the following electronic dat-abases: Medline, PsychInfo and ISI Web of Knowledge. We combedfor the term ‘‘Parkinson’s disease’’ with each of the other neuropsy-chological tasks proposed for the assessment of OMPFC functions(Zald & Andreotti, 2010). The last search was carried out in Decem-ber 2010. Reversal learning tasks, the Iowa Gambling Task and theTheory of Mind tasks were widely adopted in PD studies, whileonly one study adopted the Object Alternation task (Freedman,1990), and only one study investigated autobiographical memory(Smith, Souchay, & Conway, 2010) in PD. No studies utilized ObjectReversal Learning tasks or interviews and questionnaires about theemotional characteristics of personality.
To test our working hypotheses, we needed to compare the per-formance of patients at different stages of PD. Thus, we decided to re-view only the empirical evidence derived from tasks adopted atdifferent stages of PD, including reversal learning tasks, the IowaGambling Task and the Theory of Mind tasks. Among studies identi-fied in electronic databases, we included only those reporting dataon the performance of PD patients compared to healthy controls.

M. Poletti, U. Bonuccelli / Brain and Cognition 79 (2012) 23–33 27
6. Learning and adapting to changing reinforcementcontingencies
The broadest group of cognitive tasks that showed sensitivity toOMPFC lesions were tasks in which the individual must learn a re-ward contingency that diverges from expectations or that is chang-ing over time (Zald & Andreotti, 2010). Within this group of tasks,reversal learning tasks and the Iowa Gambling Task were used totest PD patients.
6.1. Reversal learning tasks
In reversal learning tasks (RLTs), the subject is rewarded forselecting a stimulus until reaching a certain level of performance,after which the reward contingency reverses. In detail, upon rever-sal, the previous reward stimulus is no longer rewarded, and thepreviously non-reward stimulus becomes the reward stimulus.Depending upon the specific paradigm, the reward contingencymay undergo a single reversal or multiple reversals. In probabilisticRLTs, the stimulus probability and reward value slowly reverseover the course of the task. Lesion studies have shown that OMPFCdamages are associated with deficient RLT performances, espe-cially if the lesions are bilateral (Zald & Andreotti, 2010). A fMRIstudy (Cools et al., 2002) confirmed that RLTs activate the orbitalloop circuit, which includes the ventral striatum and the OMPFC.Specifically, the ventral striatum is necessary for learning a new re-sponse when a previously learned response is no longer rewarding(Shohamy, Myers, Hopkins, Sage, & Gluck, 2009).
Swainson and colleagues (2000) provided the first empiricalevidence of deficient performance on RLTs by medicated early PDpatients, whereas unmedicated (‘‘off’’ therapy) early PD patientshad normal performances on RLTs, as compared to Healthy Con-trols (HCs). The detrimental effect of the dopaminergic therapyon RLT performances of early PD patients was subsequently con-firmed by other studies (Cools, Barker, Sahakian, & Robbins,2001; Czernecki et al., 2002; Graef et al., 2010). This medication ef-fect is particularly evident when reversal is signaled by unexpectedpunishments (Cools, Altamirano, & D’Esposito, 2006) and is relatedto the disruption of activity in the NAcc, a subcortical structurestrongly connected with the OMPFC (Cools et al., 2007). A recentstudy (Peterson et al., 2009) used RLTs in patients at early/moder-ate stages of PD (mean disease duration 10.4 years; H&Y MeanStage 2.4 ± 0.5). Patients were assessed ‘‘off’’ therapy and reporteddeficient performances, as compared to HCs. This finding couldpreliminarily confirm that, in moderate PD, the withdrawal ofdopaminergic therapies has a detrimental effect on reversallearning.
6.2. The Iowa Gambling Task
The Iowa Gambling Task (IGT; Bechara, 2007; Bechara, Damasio,Damasio, & Anderson, 1994) is said to measure decision makingunder uncertainty, since successful performance requires subjectsto use the feedback gained throughout the task to identify strate-gies that maximize the initial bet.
The IGT requires 100 card selections from four decks of cardsidentical in appearance. Subjects are asked to maximize their profitstarting from a $2000 loan of play money. After turning over sev-eral cards, subjects are both given money and sometimes askedto pay a penalty, according to a pre-programmed schedule of re-wards and punishments. Gains and losses vary for each card se-lected from the four decks: decks A and B are ‘‘disadvantageous’’;whilst they pay $100, the penalty amounts are higher, so they costmore in the long run. Decks C and D are ‘‘advantageous’’; they payonly $50, but the penalty amounts are lower, resulting in an overall
gain over time. In summary, decks A and B are equivalent in termsof overall net loss over the trials, as are decks C and D; the differ-ence is that in decks A and C punishment is more frequent, but ofsmaller magnitude, while in decks B and D punishment is less fre-quent but of larger magnitude.
The goal of the game is to win as much money as possible, or toavoid losing money for as long as possible. To attain this goal, sub-jects must find out the most advantageous decks and predomi-nantly pick up cards from those decks. Successful strategies relyon sampling more from decks C and D than from decks A and B;there is no advantage for participants in selecting more from thefrequent punishment (A and C) versus infrequent punishment (Band D) decks, or vice versa. Healthy subjects tend to predominantlychoose cards from the advantageous decks (C and D) after about 40card choices, revealing that also reversal learning is a crucial com-ponent of IGT (Fellows & Farah, 2005). The IGT score is calculatedby subtracting the number of disadvantageous choices (cards cho-sen from decks A and B) from the number of advantageous ones(decks C and D); therefore, successful performances are indicatedby a positive score (>0) and evidence an advantageous decisionmaking style. Moreover, unsuccessful performances are indicatedby a negative score (60), with lower scores indicating a more dis-advantageous decision-making style (Bechara, 2007).
Neuroimaging studies (i.e., Li, Lu, D’Argembeau, Ng, & Bechara,2009) and lesion studies (i.e., Bechara, Tranel, & Damasio, 2000)indicated that IGT performances are a measure of OMPFC function-ing. For example, only patients with damages to this area, but notto dorsolateral or dorsomedial sectors of the PFC, persist in draw-ing cards from the high payout/high penalty decks, despite the ulti-mately punishing consequences of this behavior. Moreover,neuropsychological studies showed that IGT performance accountsfor variances in decision-making performance not currently attrib-utable to either the intellectual level or classic measures of execu-tive functioning (inhibition, working memory, set-shifting (Toplak,Sorge, Benoit, West, & Stanovich, 2010).
A search in electronic databases found 13 studies that utilizedthe IGT to test samples of PD patients (see Table 2). Of these, sixstudies included only early PD patients, while seven studies in-cluded both early and moderate PD patients. One study assessedPD patients with dementia (Delazer et al., 2009), and another studyassessed PD patients who displayed pathological gambling (Rossiet al., 2010).
With regard to studies including only early PD patients, 3 of the6 studies reported a disadvantageous decision-making style (TotalIGT Score 6 0) (Kobayakawa, Koyama, Mimura, & Kawamura,2008; Kobayakawa, Tsuruya, & Kawamura, 2010; Perretta, Pari, &Beninger, 2005), whereas three studies reported an advantageousdecision-making style (Total IGT Score > 0) (Czernecki et al.,2002; Ibarretxe-Bilbao et al., 2009; Poletti et al., 2010). Moreover,although only half of the IGT studies reported a disadvantageousdecision-making style of PD patients, 5 of these six studies re-ported significantly worse IGT performances in comparison toHCs (Czernecki et al., 2002; Ibarretxe-Bilbao et al., 2009; Kobayak-awa et al., 2008; Kobayakawa et al., 2010; Perretta et al., 2005).Only the study that assessed newly diagnosed, drug-naïve PD pa-tients reported similar IGT performances in comparison to HCs(Poletti et al., 2010).
In those studies that included both early PD patients and mod-erate PD patients, four studies reported a disadvantageous deci-sion-making style (Delazer et al., 2009; Euteneuer et al., 2009;Pagonabarraga et al., 2007; Thiel et al., 2003), while three studiesreported an advantageous decision-making style (Mimura, Oeda,& Kawamura, 2006; Perretta et al., 2005; Rossi et al., 2010; Stout,Rodawalt, & Siemers, 2001). Of these studies, three reported signif-icantly worse IGT performances in comparison to HCs (Delazeret al., 2009; Pagonabarraga et al., 2007; Stout et al., 2001), whereas
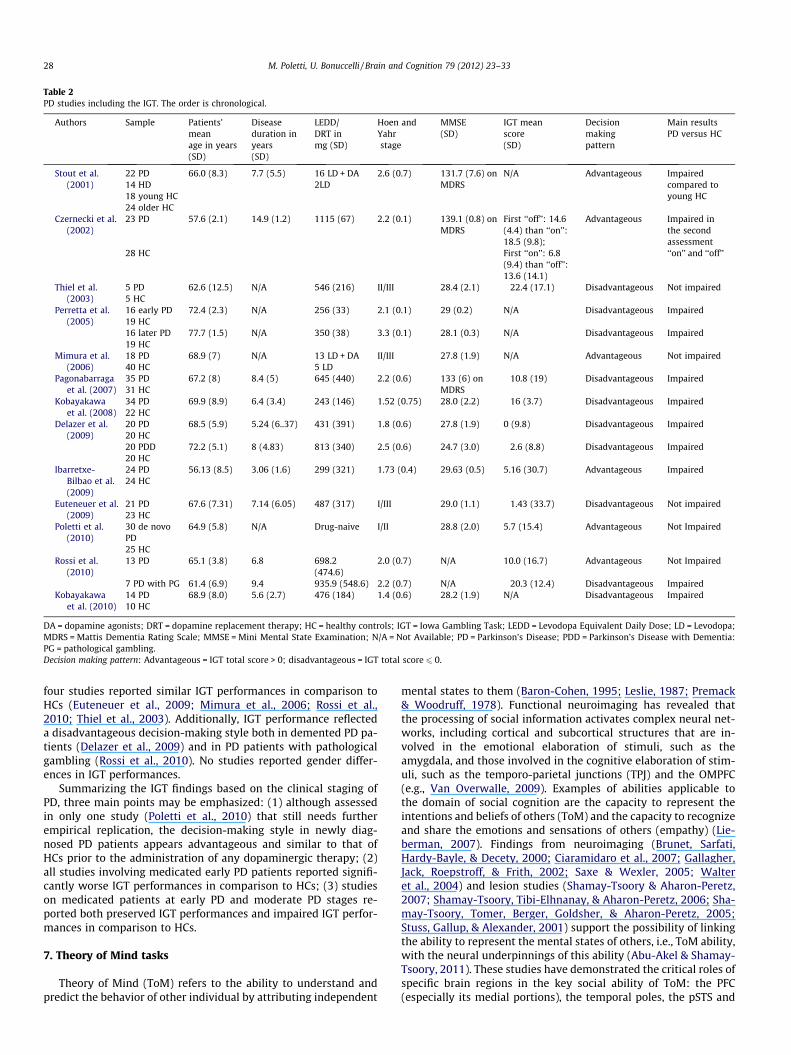
Table 2PD studies including the IGT. The order is chronological.
Authors Sample Patients’meanage in years(SD)
Diseaseduration inyears(SD)
LEDD/DRT inmg (SD)
Hoen andYahrstage
MMSE(SD)
IGT meanscore(SD)
Decisionmakingpattern
Main resultsPD versus HC
Stout et al.(2001)
22 PD 66.0 (8.3) 7.7 (5.5) 16 LD + DA 2.6 (0.7) 131.7 (7.6) onMDRS
N/A Advantageous Impairedcompared toyoung HC
14 HD 2LD18 young HC24 older HC
Czernecki et al.(2002)
23 PD 57.6 (2.1) 14.9 (1.2) 1115 (67) 2.2 (0.1) 139.1 (0.8) onMDRS
First ‘‘off’’: 14.6(4.4) than ‘‘on’’:18.5 (9.8);
Advantageous Impaired inthe secondassessment‘‘on’’ and ‘‘off’’28 HC First ‘‘on’’: 6.8
(9.4) than ‘‘off’’:13.6 (14.1)
Thiel et al.(2003)
5 PD 62.6 (12.5) N/A 546 (216) II/III 28.4 (2.1) �22.4 (17.1) Disadvantageous Not impaired5 HC
Perretta et al.(2005)
16 early PD 72.4 (2.3) N/A 256 (33) 2.1 (0.1) 29 (0.2) N/A Disadvantageous Impaired19 HC16 later PD 77.7 (1.5) N/A 350 (38) 3.3 (0.1) 28.1 (0.3) N/A Disadvantageous Impaired19 HC
Mimura et al.(2006)
18 PD 68.9 (7) N/A 13 LD + DA II/III 27.8 (1.9) N/A Advantageous Not impaired40 HC 5 LD
Pagonabarragaet al. (2007)
35 PD 67.2 (8) 8.4 (5) 645 (440) 2.2 (0.6) 133 (6) onMDRS
�10.8 (19) Disadvantageous Impaired31 HC
Kobayakawaet al. (2008)
34 PD 69.9 (8.9) 6.4 (3.4) 243 (146) 1.52 (0.75) 28.0 (2.2) �16 (3.7) Disadvantageous Impaired22 HC
Delazer et al.(2009)
20 PD 68.5 (5.9) 5.24 (6..37) 431 (391) 1.8 (0.6) 27.8 (1.9) 0 (9.8) Disadvantageous Impaired20 HC20 PDD 72.2 (5.1) 8 (4.83) 813 (340) 2.5 (0.6) 24.7 (3.0) �2.6 (8.8) Disadvantageous Impaired20 HC
Ibarretxe-Bilbao et al.(2009)
24 PD 56.13 (8.5) 3.06 (1.6) 299 (321) 1.73 (0.4) 29.63 (0.5) 5.16 (30.7) Advantageous Impaired24 HC
Euteneuer et al.(2009)
21 PD 67.6 (7.31) 7.14 (6.05) 487 (317) I/III 29.0 (1.1) �1.43 (33.7) Disadvantageous Not impaired23 HC
Poletti et al.(2010)
30 de novoPD
64.9 (5.8) N/A Drug-naive I/II 28.8 (2.0) 5.7 (15.4) Advantageous Not Impaired
25 HCRossi et al.
(2010)13 PD 65.1 (3.8) 6.8 698.2 2.0 (0.7) N/A 10.0 (16.7) Advantageous Not Impaired
(474.6)7 PD with PG 61.4 (6.9) 9.4 935.9 (548.6) 2.2 (0.7) N/A �20.3 (12.4) Disadvantageous Impaired
Kobayakawaet al. (2010)
14 PD 68.9 (8.0) 5.6 (2.7) 476 (184) 1.4 (0.6) 28.2 (1.9) N/A Disadvantageous Impaired10 HC
DA = dopamine agonists; DRT = dopamine replacement therapy; HC = healthy controls; IGT = Iowa Gambling Task; LEDD = Levodopa Equivalent Daily Dose; LD = Levodopa;MDRS = Mattis Dementia Rating Scale; MMSE = Mini Mental State Examination; N/A = Not Available; PD = Parkinson’s Disease; PDD = Parkinson’s Disease with Dementia:PG = pathological gambling.Decision making pattern: Advantageous = IGT total score > 0; disadvantageous = IGT total score 6 0.
28 M. Poletti, U. Bonuccelli / Brain and Cognition 79 (2012) 23–33
four studies reported similar IGT performances in comparison toHCs (Euteneuer et al., 2009; Mimura et al., 2006; Rossi et al.,2010; Thiel et al., 2003). Additionally, IGT performance reflecteda disadvantageous decision-making style both in demented PD pa-tients (Delazer et al., 2009) and in PD patients with pathologicalgambling (Rossi et al., 2010). No studies reported gender differ-ences in IGT performances.
Summarizing the IGT findings based on the clinical staging ofPD, three main points may be emphasized: (1) although assessedin only one study (Poletti et al., 2010) that still needs furtherempirical replication, the decision-making style in newly diag-nosed PD patients appears advantageous and similar to that ofHCs prior to the administration of any dopaminergic therapy; (2)all studies involving medicated early PD patients reported signifi-cantly worse IGT performances in comparison to HCs; (3) studieson medicated patients at early PD and moderate PD stages re-ported both preserved IGT performances and impaired IGT perfor-mances in comparison to HCs.
7. Theory of Mind tasks
Theory of Mind (ToM) refers to the ability to understand andpredict the behavior of other individual by attributing independent
mental states to them (Baron-Cohen, 1995; Leslie, 1987; Premack& Woodruff, 1978). Functional neuroimaging has revealed thatthe processing of social information activates complex neural net-works, including cortical and subcortical structures that are in-volved in the emotional elaboration of stimuli, such as theamygdala, and those involved in the cognitive elaboration of stim-uli, such as the temporo-parietal junctions (TPJ) and the OMPFC(e.g., Van Overwalle, 2009). Examples of abilities applicable tothe domain of social cognition are the capacity to represent theintentions and beliefs of others (ToM) and the capacity to recognizeand share the emotions and sensations of others (empathy) (Lie-berman, 2007). Findings from neuroimaging (Brunet, Sarfati,Hardy-Bayle, & Decety, 2000; Ciaramidaro et al., 2007; Gallagher,Jack, Roepstroff, & Frith, 2002; Saxe & Wexler, 2005; Walteret al., 2004) and lesion studies (Shamay-Tsoory & Aharon-Peretz,2007; Shamay-Tsoory, Tibi-Elhnanay, & Aharon-Peretz, 2006; Sha-may-Tsoory, Tomer, Berger, Goldsher, & Aharon-Peretz, 2005;Stuss, Gallup, & Alexander, 2001) support the possibility of linkingthe ability to represent the mental states of others, i.e., ToM ability,with the neural underpinnings of this ability (Abu-Akel & Shamay-Tsoory, 2011). These studies have demonstrated the critical roles ofspecific brain regions in the key social ability of ToM: the PFC(especially its medial portions), the temporal poles, the pSTS and

M. Poletti, U. Bonuccelli / Brain and Cognition 79 (2012) 23–33 29
a neighboring but distinct region of the TPJ. Furthermore, it isworth noting that the ACC, a crucial area in the anterior cingulatecircuit affected in PD, was listed as one of the areas involved inToM processing (Amodio & Frith, 2006).
With regard to the ToM tasks, Zald and Andreotti (2010) princi-pally focused their review on the Faux Pas Recognition task (FPR;Stone, Baron-Cohen, & Knight, 1998). However, lesion studies alsoshowed that other ToM tasks permit assessment of OMPFC func-tions. In fact, it may be appropriate to distinguish between anaffective ToM subcomponent (belief about feelings) related to theOMPFC and to the orbital loop and a cognitive ToM subcomponent(belief about belief) related to the DLPFC and to the dorsolateralloop (for review Abu-Akel & Shamay-Tsoory, 2011; Kalbe et al.,2007; Kalbe et al., 2010; Shamay-Tsoory et al., 2005; Shamay-Tso-ory et al., 2006). Therefore, we reviewed the empirical evidence de-rived from other affective ToM tasks: the Reading the Mind in theEyes (RME) test and the Yoni test.
The RME test is an advanced affective ToM task consisting of thepresentation of photographs of the eye region of human faces (Bar-on-Cohen, Jolliffe, Mortimore, & Robertson, 1997; Baron-Cohen,Wheelwright, Hill, Raste, & Plumb, 2001). Participants are requiredto choose which word best describes what the individual in thephotograph is thinking or feeling (e.g., which of the followingwords best describes the eye region shown: excited, relieved,shy, or despondent). In the FPR test, the participant hears 10 storiesread aloud containing a social faux pas, as well as 10 control storiesreporting a minor conflict in which no faux pas is committed(Stone et al., 1998). After each story, participants are askedwhether anyone said anything that they should not have said,i.e., to correctly identify the stories containing a faux pas. Whena faux pas is detected, further questions are posed to evaluatethe participant’s understanding of the situation (e.g., a cognitivecomponent: ‘‘why do you think the character said it?’’ and an affec-tive component: ‘‘how do you think the character felt?’’). Thus, FPRwas designed to differentially assess both the cognitive and affec-tive aspects of the ToM. Finally, in the Yoni test (Kalbe et al., 2010;Shamay-Tsoory & Aharon-Peretz, 2007), a face named ‘Yoni’ isshown in the middle of a computer screen with four colored pic-tures in the corners that show either faces and/or examples of asemantic category (e.g., animals, fruits). Participants have to eval-uate which of these four pictures best corresponds to a sentencecontemporaneously presented on each screen about the image towhich Yoni is referring. The items can be subdivided into 3 typesof categories corresponding to: affective ToM, cognitive ToM, andcontrol conditions. While the answers for the control conditiononly require an analysis of the character’s physical attributes,choices in the affective and cognitive ToM items involve an analy-sis of mental processes based on verbal cues contained in the sen-tences, eye gaze, and/or facial expression (e.g., cognitivecomponent: ‘‘Yoni is thinking of. . .’’, affective component: ‘‘Yoniloves. . .’’, and control condition: ‘‘Yoni is close to. . .’’).
A search through the electronic databases found nine studiesthat assessed affective ToM in PD patients (see Table 3). A study(Rossi et al., 2010) was excluded from our analysis due to the lackof a comparison with HCs.
7.1. Reading the Mind in the Eyes test
The RME test was utilized in seven studies involving PD pa-tients. Only early PD patients were included in 2 of the studies(Roca et al., 2010; Tusruya, Kobayakawa, & Kawamura, 2011),while four studies included both early PD patients and moderatePD patients (Bodden et al., 2010; Euteneuer et al., 2009; Mimuraet al., 2006; Peron et al., 2010) and one study distinguished be-tween early and moderate PD patients (Peron et al., 2009). No stud-ies reported gender differences in RME performances. The majority
of these studies showed that the performance of medicated pa-tients in the RME test might be preserved in the early stages ofPD (Mimura et al., 2006; Peron et al., 2009; Roca et al., 2010), upto a mean disease duration of 7.1 years (Euteneuer et al., 2009)or 10.2 years (Peron et al., 2009). Only two studies reported lowerRME performances in medicated PD patients in comparison to HCs(Bodden et al., 2010; Tusruya et al., 2011). In the study of Tsuruyaand colleagues (2011), the patients had a mean PD duration of5.1 ± 0.7 years and were in the early stages (H&Y Mean Stage1.5 ± 0.7), while in the study of Bodden and colleagues (2010),the patients had a mean PD duration of 5.1 ± 2.8 years. However,in the Bodden study, the clinical staging of PD was heterogeneous,with both early PD and moderate PD patients (H&Y Mean Stage 2.5,Range 1–3). Moreover, in the latter study, HCs were younger thanPD patients (mean age, respectively 58.7 years and 63.7 years).Although this difference was not statistically significant(p = 0.10), the mean age of the HCs is particularly important con-sidering that the RME performances have been reported to declinewith age in healthy subjects (Pardini & Nichelli, 2009). In anotherstudy (Mimura et al., 2006), medicated patients in early PD andmoderate PD stages (Range of H&Y Stages 1–3) obtained poorerscores than HCs, but the authors highlighted that the RME perfor-mances by both PD patients and HCs were relatively high, althoughnot at the ceiling. In summary, the majority of studies reportedthat early PD patients might have preserved RME performances,even after 10 years of disease, but that mild difficulties mightemerge at least 5 years after the onset of PD.
With regards to the influence of dopaminergic therapy, a study(Peron et al., 2009) that compared RME performances of early PDpatients tested both ‘on’ and ‘off’ dopaminergic therapy did not re-port any differences. In the study by Roca et al. (2010), the RMEtest was administered to both medicated early PD patients andnewly diagnosed, drug-naive PD patients (with similar PD stages:mean H&Y score 1.42 ± 0.57 and 1.33 ± 0.54, respectively). They re-ported no differences between the performances of the medicatedPD patients, unmedicated PD patients and HCs. Finally, only thestudy of Peron and colleagues (2009) compared RME performancesof patients at different stages of PD. No differences were found be-tween RME performances of early PD patients with a short meandisease duration (2.5 years) ‘‘on’’ and ‘‘off’’ therapy and early/mod-erate PD patients with a longer mean disease duration (10.2 years).In conclusion, RME studies showed that: (1) early PD patients maypresent preserved performances in RME tasks independent of thedopaminergic therapy, even after 10 years of disease duration;(2) medicated PD patients in moderate stages of PD with at least5 years of disease duration may have difficulties, compared toHCs (Bodden et al., 2010).
7.2. The Faux Pas Recognition test and the Yoni test
Of the three studies that assessed the detection of faux pas insocial situations in PD patients, none reported gender differencesin FPR performances. These studies converged on a preservedaffective ToM component and on an impaired cognitive ToM com-ponent. Kawamura and Koyama (2007) reported that PD patientswere as able as HCs to detect the inappropriate remarks in the sto-ries (the affective component of the task) but had more difficultyinferring the reason why the person in the story had made an inap-propriate remark (the cognitive component of the task). Similarfindings were reported by Peron and colleagues (2009). In theirstudy, the FPR test was administered, as previously described, toearly medicated PD patients ‘on’ and ‘off’ medication and to med-icated moderate PD patients. No differences were found in theemotion attribution score (the affective component of the task) be-tween controls, early PD patients (‘on’ and ‘off’ dopaminergic ther-apy), and moderate PD patients. On the one hand, in terms of the
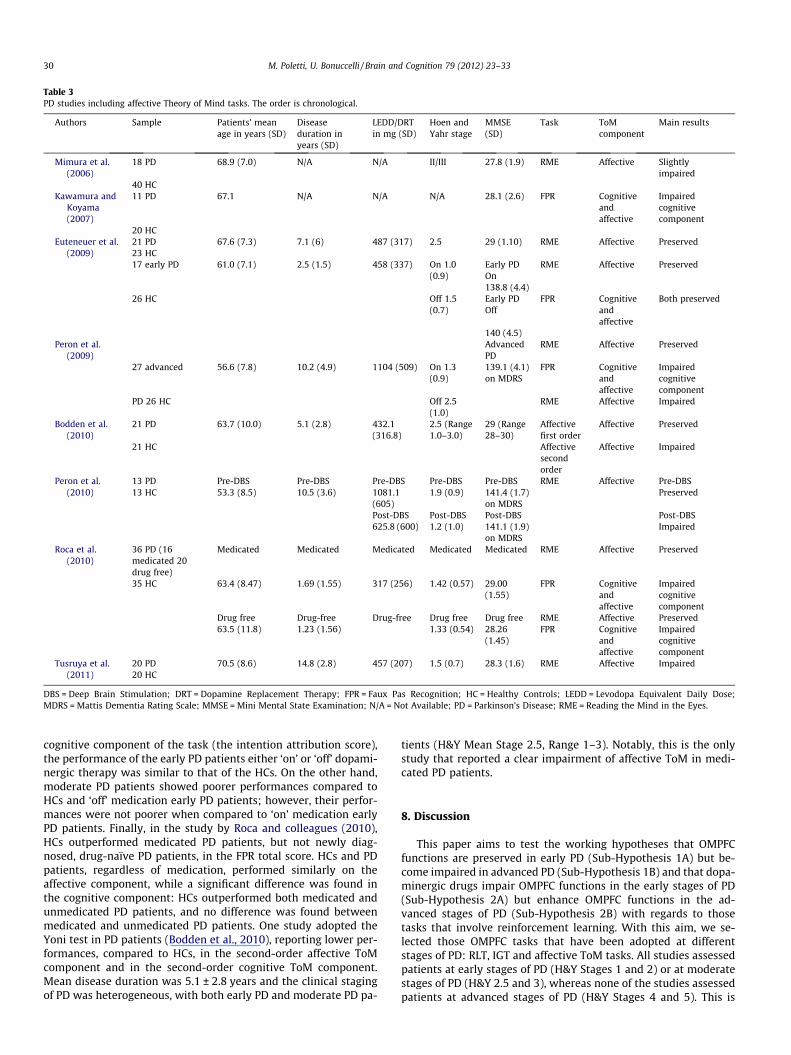
Table 3PD studies including affective Theory of Mind tasks. The order is chronological.
Authors Sample Patients’ meanage in years (SD)
Diseaseduration inyears (SD)
LEDD/DRTin mg (SD)
Hoen andYahr stage
MMSE(SD)
Task ToMcomponent
Main results
Mimura et al.(2006)
18 PD 68.9 (7.0) N/A N/A II/III 27.8 (1.9) RME Affective Slightlyimpaired
40 HCKawamura and
Koyama(2007)
11 PD 67.1 N/A N/A N/A 28.1 (2.6) FPR Cognitiveandaffective
Impairedcognitivecomponent
20 HCEuteneuer et al.
(2009)21 PD 67.6 (7.3) 7.1 (6) 487 (317) 2.5 29 (1.10) RME Affective Preserved23 HC17 early PD 61.0 (7.1) 2.5 (1.5) 458 (337) On 1.0
(0.9)Early PDOn
RME Affective Preserved
138.8 (4.4)26 HC Off 1.5
(0.7)Early PDOff
FPR Cognitiveandaffective
Both preserved
140 (4.5)Peron et al.
(2009)AdvancedPD
RME Affective Preserved
27 advanced 56.6 (7.8) 10.2 (4.9) 1104 (509) On 1.3(0.9)
139.1 (4.1)on MDRS
FPR Cognitiveandaffective
Impairedcognitivecomponent
PD 26 HC Off 2.5(1.0)
RME Affective Impaired
Bodden et al.(2010)
21 PD 63.7 (10.0) 5.1 (2.8) 432.1(316.8)
2.5 (Range1.0–3.0)
29 (Range28–30)
Affectivefirst order
Affective Preserved
21 HC Affectivesecondorder
Affective Impaired
Peron et al.(2010)
13 PD Pre-DBS Pre-DBS Pre-DBS Pre-DBS Pre-DBS RME Affective Pre-DBS13 HC 53.3 (8.5) 10.5 (3.6) 1081.1
(605)1.9 (0.9) 141.4 (1.7)
on MDRSPreserved
Post-DBS Post-DBS Post-DBS Post-DBS625.8 (600) 1.2 (1.0) 141.1 (1.9)
on MDRSImpaired
Roca et al.(2010)
36 PD (16medicated 20drug free)
Medicated Medicated Medicated Medicated Medicated RME Affective Preserved
35 HC 63.4 (8.47) 1.69 (1.55) 317 (256) 1.42 (0.57) 29.00(1.55)
FPR Cognitiveandaffective
Impairedcognitivecomponent
Drug free Drug-free Drug-free Drug free Drug free RME Affective Preserved63.5 (11.8) 1.23 (1.56) 1.33 (0.54) 28.26
(1.45)FPR Cognitive
andaffective
Impairedcognitivecomponent
Tusruya et al.(2011)
20 PD 70.5 (8.6) 14.8 (2.8) 457 (207) 1.5 (0.7) 28.3 (1.6) RME Affective Impaired20 HC
DBS = Deep Brain Stimulation; DRT = Dopamine Replacement Therapy; FPR = Faux Pas Recognition; HC = Healthy Controls; LEDD = Levodopa Equivalent Daily Dose;MDRS = Mattis Dementia Rating Scale; MMSE = Mini Mental State Examination; N/A = Not Available; PD = Parkinson’s Disease; RME = Reading the Mind in the Eyes.
30 M. Poletti, U. Bonuccelli / Brain and Cognition 79 (2012) 23–33
cognitive component of the task (the intention attribution score),the performance of the early PD patients either ‘on’ or ‘off’ dopami-nergic therapy was similar to that of the HCs. On the other hand,moderate PD patients showed poorer performances compared toHCs and ‘off’ medication early PD patients; however, their perfor-mances were not poorer when compared to ‘on’ medication earlyPD patients. Finally, in the study by Roca and colleagues (2010),HCs outperformed medicated PD patients, but not newly diag-nosed, drug-naïve PD patients, in the FPR total score. HCs and PDpatients, regardless of medication, performed similarly on theaffective component, while a significant difference was found inthe cognitive component: HCs outperformed both medicated andunmedicated PD patients, and no difference was found betweenmedicated and unmedicated PD patients. One study adopted theYoni test in PD patients (Bodden et al., 2010), reporting lower per-formances, compared to HCs, in the second-order affective ToMcomponent and in the second-order cognitive ToM component.Mean disease duration was 5.1 ± 2.8 years and the clinical stagingof PD was heterogeneous, with both early PD and moderate PD pa-
tients (H&Y Mean Stage 2.5, Range 1–3). Notably, this is the onlystudy that reported a clear impairment of affective ToM in medi-cated PD patients.
8. Discussion
This paper aims to test the working hypotheses that OMPFCfunctions are preserved in early PD (Sub-Hypothesis 1A) but be-come impaired in advanced PD (Sub-Hypothesis 1B) and that dopa-minergic drugs impair OMPFC functions in the early stages of PD(Sub-Hypothesis 2A) but enhance OMPFC functions in the ad-vanced stages of PD (Sub-Hypothesis 2B) with regards to thosetasks that involve reinforcement learning. With this aim, we se-lected those OMPFC tasks that have been adopted at differentstages of PD: RLT, IGT and affective ToM tasks. All studies assessedpatients at early stages of PD (H&Y Stages 1 and 2) or at moderatestages of PD (H&Y 2.5 and 3), whereas none of the studies assessedpatients at advanced stages of PD (H&Y Stages 4 and 5). This is

M. Poletti, U. Bonuccelli / Brain and Cognition 79 (2012) 23–33 31
likely due to the frequent association with dementia in these laterstages, and since severe motor impairment often hampers the neu-ropsychological assessment and the identification of specific cogni-tive deficits in these patients. This pattern of findings permitted usto test the Sub-Hypotheses 1A and 2A, while further studies areneeded to test the sub-Hypotheses 1B and 2B.
With regards to OMPFC functions in early PD, the studies we re-viewed showed that patients have preserved RLT performances’’off’’ therapy and impaired RLT performances ‘‘on’’ therapy, sug-gesting that the dopaminergic therapy impairs reversal learningin early PD. Moreover, drug-naïve patients have preserved IGT per-formances (although this was assessed in only one study), while allstudies with medicated early PD patients reported impaired IGTperformances, suggesting that the dopaminergic therapy impairsdecision-making in early PD. In sum, the preliminary findings sug-gest that early PD patients have preserved RLT and IGT perfor-mances when they are not medicated, but medication impairstheir performances in the same tasks. These findings are consistentwith the dopamine overdose hypothesis (Gotham et al., 1986,1988) and with the inverted U-shape curve model (Cools, 2006),which describe the relationship between the administration ofdopaminergic therapies and cognitive functioning in PD patients.Moreover, these findings suggest that the orbital loop, includingthe OMPFC, is preserved in early PD when it is not overdosed bythe dopaminergic therapy, which supports the validity of theSub-Hypotheses 1A. Additionally, studies with affective ToM taskssupport this conclusion, when one considers that preserved perfor-mances have been reported in the RME test and in the affectivecomponent of the FPR.
From the literature review, a different pattern emerged in rela-tion to the effects of the dopaminergic therapy between tasks thatevaluated the ability to learn and adapt to changing reinforcementcontingencies (RLT and IGT) and affective ToM tasks. Especially instudies with RLT, in comparing performances of early PD patients‘‘on’’ and ‘‘off’’ dopaminergic therapy, there was a significant differ-ence between these conditions, with the ‘‘off’’ patients outperform-ing the ‘‘on’’ patients (Cools et al., 2001; Czernecki et al., 2002;Graef et al., 2010; Swainson et al., 2000). On the contrary, studiesthat assessed affective ToM in early PD did not report any differ-ence between performances ‘‘on’’ and ‘‘off’’ dopaminergic therapy(Peron et al., 2009; Roca et al., 2010). These findings suggested thatthe dopaminergic therapy has a detrimental effect on reversallearning and on adapting to changing reinforcement contingenciesin early PD, while the effect of therapy on affective ToM is attenu-ated. This conclusion supports the validity of the Sub-Hypothesis2A.
The Sub-Hypotheses 1B and 2B could be supported by findingsshowing that advanced PD patients have impaired performances inOMPFC tasks when they are ‘‘off’’ therapy, but they have improvedperformances when they are ‘‘on’’ therapy. No studies using theRLT or IGT assessed patients at this stage of PD. However, theRLT and IGT performances of moderate PD patients show thatOMPFC functions worsen with the progression of the disease fromearly to moderate stages. Peterson and colleagues (2009) reportedthat non-demented, unmedicated (‘‘off’’ therapy) patients with amean PD duration of 10.4 years had deficient performances whencompared to HCs in a probabilistic RLT. This finding suggested thatRLT performances decline with the progression of PD ‘‘off’’ therapy.Moreover, while all IGT studies with early, medicated PD patientsreported a decision-making impairment, most studies with medi-cated, moderate PD patients reported preservation of decision-making. Following the model of Cools (2006), this pattern indi-rectly and preliminarily suggests that OMPFC functions declinefrom early to moderate PD stages, as evidenced by the different ef-fect of dopaminergic drugs on decision-making (detrimental atearly stages, enhancing at moderate stages).
8.1. The relationship between the different OMPFC functions
In this review, we discussed studies adopting the IGT and affec-tive ToM tasks. Thus, it is interesting to report that two of thesestudies adopted these tasks in the same samples of PD patients.In the study led by Mimura and colleagues (2006), PD patients per-formed similarly to HCs both in the IGT and the RME, and the TotalIGT Score correlated significantly with the RME score (r = 0.59,p < 0.01). On the other hand, in the study by Euteneuer and col-leagues (2009), PD patients performed similarly to HCs both inthe IGT and RME, but the performances in these tasks were not sig-nificantly correlated. The lack of correlation between the ToM tasksand the IGT, despite the acknowledged involvement of the OMPFCin both these tasks (Zald & Andreotti, 2010), could be because theneural substrates for these two domains are independent. Cogni-tively, the development of a successful strategy on the IGT doesnot require representation of the intentions, beliefs or emotionsof others; likewise, ToM task deficits are not clearly dependenton decision-making ability. Anatomically, this dissociation couldbe within the OFC (i.e., lateral versus medial aspects: see Section 2).The resolution of the lesion and functional imaging studies, how-ever, may be insufficient to dissect such anatomical heterogeneitywithin the OMPFC using these tasks. Moreover, while both the IGTand affective ToM tasks may share a neural substrate within theOMPFC, they may be differentially affected by the involvement ofother subcortical and cortical regions outside this region (Torralvaet al., 2007). Interactions with areas, such as the amygdala or theDLPFC, could disproportionately bias the performance on the IGT(or ToM tasks), effectively obscuring any underlying correlation re-lated to the OMPFC (Lawrence, Jollant, O’Daly, Zelaya, & Phillips,2009; Xi et al., 2011). Further studies analyzing both of these pro-cesses in PD patients must to clarify if deficits in decision-makingcan occur without deficits in ToM, or if deficits in ToM usually co-occur with deficits in decision-making.
9. Conclusions and future directions
We examined studies that assessed OMPFC functions in pa-tients with PD. We found empirical evidence that in early PD,OMPFC functions are preserved (Sub-Hypothesis 1A), and dopami-nergic drugs have a detrimental effect on these functions when thetask involves reinforcement learning (Sub-Hypothesis 2A). The lackof studies in non-demented, advanced PD patients impedes thetesting of the sub-hypotheses on OMPFC functions in advancedPD (1B; 2B), but this topic deserves further neuropsychologicalinvestigation.
A limit of this paper is that the reviewed empirical evidence isderived from only three types of OMPFC tasks: reversal learningtasks, decision-making tasks and affective ToM tasks. Therefore,further studies that utilize other OMPFC tasks are needed to clarifythe course of OMPFC functions during the progression of PD. Withregards to other OMPFC tasks reviewed by Zald and Andreotti(2010) and by other reviews on OMPFC functions (Clark, Cools, &Robbins, 2004; Rolls, 2004), olfactory processing tasks and emotionrecognition tasks appeared not to be suited to detect specificOMPFC dysfunctions in PD. As a matter of fact, other neural struc-tures involved in these processes (the olfactory bulb for olfactoryprocessing, the amygdala and basal ganglia for emotion recogni-tion) are affected earlier by PD-related neuropathology, accordingto the Braak stages (Braak et al., 2003). Future studies should uti-lize OMPFC tasks that are scarcely or not ever used in PD assess-ment, including the following: (1) reinforcement learning tasks,such as the Object Alternation and Object Reversal Learning tasks;(2) autobiographical memory tasks; and (3) questionnaires aboutthe emotional characteristics of personality. Moreover, recentpapers have focused on other functions of the OMPFC and related

32 M. Poletti, U. Bonuccelli / Brain and Cognition 79 (2012) 23–33
tasks that could be adopted to test its function in PD patients,including n-back tasks of the coordination of working memory(Barbey, Koenigs, & Grafman, 2011) and moral judgment tasks(Forbes & Grafman, 2010; Moll, Zahn, de Oliveira-Souza, Krueger,& Grafman, 2005).
References
Aarsland, D., Bronnick, K., Williams-Gray, C., Weintraub, D., Marder, K., Kulisevsky,J., et al. (2010). Mild cognitive impairment in Parkinson disease: A multicenterpooled analysis. Neurology, 75, 1062–1069.
Abu-Akel, A., & Shamay-Tsoory, S. (2011). Neuroanatomical and neurochemicalbases of theory of mind. Neuropsychologia, 49, 2971–2984.
Alexander, G. E., Crutcher, M. D., & DeLong, M. R. (1990). Basal ganglia-thalamocortical circuits: Parallel substrates for motor, oculomotor,‘‘prefrontal’’ and ‘‘limbic’’ functions. Progress in Brain Research, 85, 119–146.
Alexander, G. E., Delong, M. R., & Strick, P. L. (1986). Parallel organisation offunctionally segregated circuits linking basal ganglia and cortex. Annual Reviewsof Neuroscience, 9, 357–381.
Amodio, D. M., & Frith, C. D. (2006). Meeting of minds: The medial frontal cortex andsocial cognition. Nature Reviews Neuroscience, 7, 268–277.
Assogna, F., Pontieri, F. E., Caltagirone, C., & Spalletta, G. (2008). The recognition offacial emotional expression in Parkinson’s disease. EuropeanNeuropsychopharmacology, 18, 835–848.
Barbey, A. K., Koenigs, M., & Grafman, J. (2011). Orbitofrontal contributions tohuman working memory. Cerebral Cortex, 21, 789–795.
Baron-Cohen, S. (1995). Mindblindness: An essay on autism and Theory of Mind.Cambridge: MIT Press.
Baron-Cohen, S., Jolliffe, T., Mortimore, C., & Robertson, M. (1997). Anotheradvanced test of Theory of Mind: Evidence from very high functioning adultswith autism or Asperger syndrome. Journal of Child Psychology and Psychiatry,38, 813–822.
Baron-Cohen, S., Wheelwright, S., Hill, J., Raste, Y., & Plumb, I. (2001). The ‘‘Readingthe Mind in the Eyes’’ test revised version: A study with normal adults, andadults with Asperger syndrome or high-functioning autism. Journal of ChildPsychology and Psychiatry, 42, 241–251.
Bechara, A. (2007). Iowa gambling task professional manual. Lutz, FL: PsychologicalAssessment Resources.
Bechara, A., Damasio, A. R., Damasio, H., & Anderson, S. W. (1994). Insensitivity tofuture consequences following damage to human prefrontal cortex. Cognition,50, 7–15.
Bechara, A., Tranel, D., & Damasio, H. (2000). Characterization of decision makingdeficits in patients with ventromedial prefrontal cortex lesions. Brain, 123,2189–2202.
Benarroch, E. (2010). Olfactory system: Functional organization and involvement inneurodegenerative disease. Neurology, 75, 1104–1109.
Bodden, M. E., Mollenhauer, B., Trenkwalder, C., Cabanel, C., Eggert, K. M., Unger, M.M., et al. (2010). Affective and cognitive Theory of Mind in patients withParkinson’s disease. Parkinsonism and Related Disorders, 16, 466–470.
Bohnen, N. I., Kaufer, D. I., Hendrickson, R., Ivanco, L. S., Lopresti, B. J., Constantine, G.M., et al. (2006). Cognitive correlates of cortical cholinergic denervation inParkinson’s disease and parkinsonian dementias. Journal of Neurology, 253,242–247.
Bonuccelli, U., & Pavese, N. (2006). Dopamine agonists in the treatment ofParkinson’s disease. Expert Review of Neurotherapeutics, 6, 81–89.
Bouret, S., & Richmond, B. J. (2010). Ventromedial and orbitofrontal neuronsdifferentially encode internally and externally driven motivational values inmonkeys. Journal of Neuroscience, 30, 8591–8601.
Braak, H., Del Tredici, K., Rub, U., de Vos, R. A., Jansen Steur, E. N., & Braak, E. (2003).Staging of brain pathology related to sporadic Parkinson’s disease. Neurobiologyof Aging, 24, 197–211.
Brooks, D. J., & Piccini, P. (2006). Imaging in Parkinson’s disease: The role ofmonoamines in behaviour. Biological Psychiatry, 59, 908–918.
Brunet, E., Sarfati, Y., Hardy-Bayle, M. C., & Decety, J. (2000). A PET investigation ofthe attribution of intentions with a nonverbal task. NeuroImage, 11, 157–166.
Calabresi, P., Picconi, B., Parnetti, L., & Di Filippo, M. (2006). A convergent model forcognitive dysfunctions in Parkinson’s disease: The critical dopamine–acetylcholine synaptic balance. Lancet Neurology, 5, 974–983.
Carmichael, S. T., & Price, J. L. (1996). Connectional networks within the orbital andmedial prefrontal cortex of macaque monkeys. Journal of ComparativeNeurology, 346, 179–207.
Chudasama, Y., & Robbins, T. W. (2006). Functions of frontostriatal systems incognition: Comparative neuropsychopharmacological studies in rats, monkeysand humans. Biological Psychology, 73, 19–38.
Ciaramidaro, A., Adenzato, M., Enrici, I., Erk, S., Pia, L., Bara, B. G., et al. (2007). Theintentional network: How the brain reads varieties of intentions.Neuropsychologia, 45, 3105–3113.
Clark, L., Cools, R., & Robbins, T. W. (2004). The neuropsychology of ventralprefrontal cortex: Decision making and reversal learning. Brain and Cognition,55, 41–53.
Cools, R. (2006). Dopaminergic modulation of cognitive function-implications for L-DOPA treatment in Parkinson’s disease. Neuroscience & Biobehavioral Reviews,30, 1–23.
Cools, R., Altamirano, L., & D’Esposito, M. (2006). Reversal learning in Parkinson’sdisease depends of medication status and outcome valence. Neuropsychologia,44, 1663–1673.
Cools, R., Barker, R. A., Sahakian, B. J., & Robbins, T. W. (2001). Enhanced or impairedcognitive function in Parkinson’s disease as a function of dopaminergicmedication and task demands. Cerebral Cortex, 11, 1136–1143.
Cools, R., Clark, L., Owen, A. M., & Robbins, T. W. (2002). Defining the neuralmechanism of probabilistic reversal learning using event-related functionalmagnetic resonance imaging. Journal of Neuroscience, 22, 4583–4587.
Cools, R., Lewis, S. J., Clark, L., Barker, R. A., & Robbins, T. W. (2007). L-DOPA disruptsactivity in the nucleus accumbens during reversal learning in Parkinson’sdisease. Neuropsychopharmachology, 32, 180–189.
Czernecki, V., Pillon, B., Houeto, J. L., Pochon, J. B., Levy, R., & Dubois, B. (2002).Motivation, reward, and Parkinson’s disease: Influence of dopatherapy.Neuropsychologia, 40, 2257–2267.
Damasio, H. (1996). Human neuroanatomy relevant to decision-making. In A. R.Damasio, H. Damasio, Y. Christen (Eds.), Neurobiology of decision-making. Berlin:Springer-Verlag.
Delazer, M., Sinz, H., Zamarian, L., Stockner, H., Seppi, K., Wenning, G. K., et al.(2009). Decision making under risk and under ambiguity in Parkinson’s disease.Neuropsychologia, 47, 1901–1908.
Dias, R., Robbins, T. W., & Roberts, A. C. (1996). Dissociation in prefrontal cortex ofaffective and attentional shifts. Nature, 380, 69–72.
Dougherty, D., Shin, L., & Rauch, S. L. (2006). Orbitofrontal cortex activation duringfunctional neuroimaging studies of emotion induction in humans. In D. H., Zald,& S. L. Rauch (Eds.), Orbitofrontal cortex (pp. 377–392). Oxford: University Press.
Emre, M., Aarsland, D., Brown, R., Burn, D. J., Duyckaerts, C., Mizuno, Y., et al. (2007).Clinical diagnostic criteria for dementia associated with Parkinson’s Disease.Movement Disorders, 22, 1689–1707.
Euteneuer, F., Schaefer, F., Stuermer, R., Boucsein, W., Timmermann, L., Barbe, M. T.,et al. (2009). Dissociation of decision making under ambiguity and decisionmaking under risk in patients with Parkinson’s disease: A neuropsychologicaland psychophysiological study. Neuropsychologia, 47, 2882–2890.
Fellows, L. K., & Farah, M. J. (2005). Different underlying impairments in decision-making following ventromedial and dorsolateral frontal lobe damage. CerebralCortex, 15, 58–63.
Forbes, C. E., & Grafman, J. (2010). The role of the human prefrontal cortex in socialcognition and moral judgements. Annual Review of Neuroscience, 33, 299–324.
Frank, M. J. (2006). Hold your horses: A dynamic neurocomputational role of thesubthalamic nucleus in decision making. Neural Networks, 19, 1120–1136.
Frank, M. J., Samanta, J., Moustafa, A. A., & Sherman, S. J. (2007). Hold your horses:Impulsivity, deep brain stimulation and medication in parkinsonism. Science,318, 1309–1312.
Frank, M. J., Seeberger, L. C., & O’Reilly, R. C. (2004). By carrot or by stick: Cognitivereinforcement learning in parkinsonism. Science, 306, 1940–1943.
Freedman, M. (1990). Object alternation and orbitofrontal system in Alzheimer’sand Parkinson’s disease. Brain and Cognition, 14, 134–143.
Gallagher, H. L., Jack, A. I., Roepstroff, A., & Frith, C. D. (2002). Imaging theintentional stance in a competitive game. Neuroimage, 16, 814–821.
Glimcher, P. W. (2011). Understanding dopamine and reinforcement learning: Thedopamine reward prediction error. Proceedings of the National Academy ofSciences of the United States of America, 108, S3, 15647–15654.
Goetz, C. G., Poewe, W., Rascol, O., Sampaio, C., Stebbins, G. T., Counsell, C., et al.(2004). Movement disorder society task force report on the Hoehn and Yahrstaging scale: Status and recommendations. The movement disorder societytask force on rating scales for Parkinson’s disease. Movement Disorders, 19,1020–1028.
Gotham, A., Brown, R., & Marsden, C. (1986). Levodopa treatment may benefit orimpair frontal function in Parkinson’s disease. The Lancet, 2, 970–971.
Gotham, A. M., Brown, R. G., & Marsden, C. D. (1988). Frontal cognitive function inpatients with Parkinson’s disease ‘on’ and ‘off’ levodopa. Brain, 111, 299–321.
Goto, Y., & Grace, A. E. (2005a). Dopaminergic modulation of limbic and corticaldrive of nucleus accumbens in goal-directed behaviour. Nature Neuroscience, 8,805–812.
Goto, Y., & Grace, A. E. (2005b). Dopamine-dependent interactions between limbicand prefrontal cortical plasticity in the nucleus accumbens: Disruption bycocaine sensitization. Neuron, 47, 255–266.
Goto, Y., Otani, S., & Grace, A. E. (2007). The Yin and Yang of dopamine release: Anew perspective. Neuropharmacology, 53, 583–587.
Grace, A. A. (2000). The tonic/phasic model of dopamine system regulation and itsimplications for understanding alcohol and psychostimulant craving. Addiction,95, S2, S119–S128.
Graef, S., Biele, G., Krugel, L. K., Marzinzik, F., Wahl, M., Wotka, J., et al. (2010).Differential influence of levodopa on reward-based learning in Parkinson’sdisease. Frontiers in Human Neuroscience, 4, 169.
Grant, D. A., & Berg, E. A. (1948). A behavioral analysis of degree of reinforcementand ease of shifting to new responses in a Weigl-type card sorting problem.Journal of Experimental Psychology, 34, 404–411.
Gray, H. M., & Tickle-Degnen, L. (2010). A meta-analysis of performance on emotionrecognition tasks in Parkinson’s disease. Neuropsychology, 24, 176–191.
Haehner, A., Hummel, T., & Reichmann, H. (2009). Olfactory dysfunction as adiagnostic marker for Parkinson’s disease. Expert Review of Neurotherapeutics, 9,1773–1779.
Heberlein, A. S., Padon, A. A., Gillihan, S. J., Farah, M. J., & Fellows, L. K. (2008).Ventromedial frontal lobe plays a critical role in facial emotion recognition.Journal of Cognitive Neuroscience, 20, 721–733.

M. Poletti, U. Bonuccelli / Brain and Cognition 79 (2012) 23–33 33
Hernandez-Lopez, S., Tkatch, T., Perez-Garci, E., Galarraga, E., Bargas, J., Hamm, H.,et al. (2000). D2 dopamine receptors in striatal medium spiny neurons reduce L-type Ca2+ currents and excitability via a novel PLC (Beta)1-IP3 calcineurinsignalling cascade. Journal of Neuroscience, 20, 8987–8995.
Hoehn, M. M., & Yahr, M. D. (1967). Parkinsonism: Onset, progression and mortality.Neurology, 17, 427–442.
Hornykiewicz, O. (1974). The mechanism of action of L-DOPA in Parkinson’s disease.Life Sciences, 15, 1249–1259.
Ibarretxe-Bilbao, N., Junque, C., Tolosa, E., Marti, M. J., Valldeoriola, F., Bargallo, N.,et al. (2009). Neuroanatomical correlates of impaired decision making and facialemotion recognition in early Parkinson’s disease. European Journal ofNeuroscience, 30, 1162–1170.
Kalaitzakis, M. E., & Pearce, R. K. (2009). The morbid anatomy of dementia inParkinson’s disease. Acta Neuropathologica, 118, 587–598.
Kalbe, E., Grabenhorst, F., Brand, M., Kessler, J., Hilker, R., & Markowitsch, H. J.(2007). Elevated emotional reactivity in affective but not cognitive componentsof theory of mind: A psychophysiological study. Journal of Neuropsychology, 1,27–38.
Kalbe, E., Schlegel, M., Sack, A. T., Nowak, D. A., Dafotakis, M., Bangard, C., et al.(2010). Dissociating cognitive from affective theory of mind: A TMS study.Cortex, 46, 769–780.
Kawamura, M., & Koyama, S. (2007). Social cognitive impairment in Parkinson’sdisease. Journal of Neurology, 254, 49–53.
Kehagia, A. A., Barker, R. A., & Robbins, T. W. (2010). Neuropsychological and clinicalheterogeneity of cognitive impairment and dementia in patients withParkinson’s disease. Lancet Neurology, 9, 1200–1213.
Kish, S. J., Shannak, K., & Hornykiewicz, O. (1988). Uneven pattern of dopamine lossin the striatum of patients with idiopathic Parkinson’s disease. New EnglandJournal of Medicine, 318, 876–880.
Kobayakawa, M., Koyama, S., Mimura, M., & Kawamura, M. (2008). Decision makingin Parkinson’s disease: Analysis of behavioral and physiological patterns in theIowa gambling task. Movement Disorders, 23, 547–552.
Kobayakawa, M., Tsuruya, N., & Kawamura, M. (2010). Sensitivity to reward andpunishment in Parkinson’s disease: An analysis of behavioral patterns using amodified version of the Iowa Gambling Task. Parkinsonism & Related Disorders,16, 453–457.
Koerts, J., Leenders, K. L., & Brouwer, W. H. (2009). Cognitive dysfunction in non-demented Parkinson’s patients: Controlled and automatic behavior. Cortex, 45,922–999.
Lawrence, N. S., Jollant, F., O’Daly, O., Zelaya, F., & Phillips, M. L. (2009). Distinct rolesof prefrontal cortical subregions in the Iowa Gambling Task. Cerebral Cortex, 19,1134–1143.
Leslie, A. M. (1987). Pretense and representation: The origins of ‘Theory of Mind’.Psychological Review, 94, 412–426.
Li, X., Lu, Z. L., D’Argembeau, A., Ng, M., & Bechara, A. (2009). The Iowa GamblingTask in fMRI images. Human Brain Mapping, 31, 410–423.
Lieberman, M. D. (2007). Social cognitive neuroscience. A review of core processes.Annual Review of Psychology, 58, 259–279.
Litvan, I., Bathia, K. P., Burn, D. J., Goetz, C. G., Lang, A. E., McKeith, I., et al. (2003).Movement Disorders Society Scientific Issues Committee report: SIC task forceappraisal of clinical diagnostic criteria for Parkinsonian disorders. MovementDisorders, 18, 467–486.
Middleton, F. A., & Strick, P. L. (2001). Revised neuroanatomy of frontal–subcorticalcircuits. In D. G. Lichter & J. L. Cummings (Eds.), Frontal–subcortical circuits inpsychiatric and neurological disorders (pp. 44–58). New York, NY: Guilford Press.
Mimura, M., Oeda, R., & Kawamura, M. (2006). Impaired decision-making inParkinson’s disease. Parkinsonism and Related Disorders, 12, 169–175.
Moll, J., Zahn, R., de Oliveira-Souza, R., Krueger, F., & Grafman, J. (2005). Opinion:The neural basis of human moral cognition. Nature Reviews Neuroscience, 6,799–809.
Pagonabarraga, J., Garcia-Sanchez, C., Llebaria, G., Pascual-Sedano, B., Gironell, A., &Kulisevsky, J. (2007). Controlled study of decision making and cognitiveimpairment in Parkinson’s disease. Movement Disorders, 22, 1430–1435.
Pardini, M., & Nichelli, P. F. (2009). Age-related decline in mentalizing skills acrossadult life span. Experimental Aging Research, 35, 98–106.
Peron, J., Le Jeune, F., Haegelen, C., Dondaine, T., Drapier, D., Sauleau, P., et al. (2010).Subthalamic nucleus stimulation affects Theory of Mind network: A PET studyin Parkinson’s disease. PLoS One, 5, e9919.
Peron, J., Vicente, S., Leray, E., Drapier, S., Drapier, D., Cohen, R., et al. (2009). Aredopaminergic pathways involved in Theory of Mind? A study in Parkinson’sdisease. Neuropsychologia, 47, 406–414.
Perretta, J. G., Pari, G., & Beninger, R. J. (2005). Effects of Parkinson Disease on twoputative nondeclarative learning tasks: Probabilistic classification andgambling. Cognitive & Behavioural Neurology, 18, 185–192.
Peterson, D. A., Elliott, C., Song, D. D., Makeig, S., Sejnowski, T. J., et al. (2009).Probabilistic reversal learning is impaired in Parkinson’s disease. Neuroscience,163, 1092–1101.
Poewe, W., Antonini, A., Zijlmans, J. C., Burkhard, P. R., & Vingerhoets, F. (2010).Levodopa in the treatment of Parkinson’s disease: An old drug still going strong.Clinical Interventions in Aging, 7, 229–238.
Poletti, M., Emre, M., & Bonuccelli, U. (2011). Mild Cognitive Impairment andcognitive reserve in Parkinson’s disease. Epub ahead of print (11 April).
Poletti, M., Frosini, D., Lucetti, C., Del Dotto, P., Ceravolo, P., & Bonuccelli, U. (2010).Decision making in de novo Parkinson’s disease. Movement Disorders, 25,1432–1436.
Premack, D. G., & Woodruff, G. (1978). Does the chimpanzee have a Theory of Mind?Behavioral and Brain Sciences, 1, 515–526.
Price, J. L. (2006). Connections of the orbital cortex. In D. H. Zald & S. L. Rauch (Eds.),Orbitofrontal cortex (pp. 39–56). Oxford, UK: Oxford University Press.
Roca, M., Torralva, T., Gleichgerrercht, E., Chade, A., Arevalo, G. G., Gershanik, O.,et al. (2010). Impairments in social cognition in early medicated andunmedicated Parkinson’s disease. Cognitive & Behavioral Neurology, 23,152–158.
Rolls, E. T. (2004). The functions of the orbitofrontal cortex. Brain and Cognition, 55,11–29.
Rossi, M., Gerschovich, E. R., de Achaval, D., Peretz-Lloret, S., Cerquetti, D.,Cammarota, A., et al. (2010). Decision making in Parkinson’s disease patientswith and without pathological gambling. European Journal of Neurology, 17,97–102.
Saxe, R., & Wexler, A. (2005). Making sense of another mind: The role of the righttemporoparietal junction. Neuropsychologia, 43, 1391–1399.
Scatton, B., Rouquier, L., Javoy-Agid, F., & Agid, Y. (1982). Dopamine deficiency in thecerebral-cortex in Parkinson disease. Neurology, 32, 1039–1040.
Shamay-Tsoory, S. G., & Aharon-Peretz, J. (2007). Dissociable prefrontal networksfor cognitive and affective Theory of Mind: A lesion study. Neuropsychologia, 45,3054–3067.
Shamay-Tsoory, S. G., Tibi-Elhnanay, Y., & Aharon-Peretz, J. (2006). Theventromedial prefrontal cortex is involved in understanding affective but notcognitive Theory of Mind stories. Social Neuroscience, 1, 149–166.
Shamay-Tsoory, S. G., Tomer, R., Berger, B. D., Goldsher, D., & Aharon-Peretz, J.(2005). Impaired affective Theory of Mind is associated with right ventromedialprefrontal damage. Cognitive & Behavioral Neurology, 18, 55–67.
Shohamy, D., Myers, C. E., Hopkins, R. O., Sage, J., & Gluck, M. A. (2009). Distincthippocampal and basal ganglia contributions to probabilistic learning andreversal. Journal of Cognitive Neuroscience, 21, 1821–1833.
Simmonds, D. J., Pekar, J. J., & Mostofsky, S. H. (2008). Meta-analysis of Go/No-gotasks demonstrating that fMRI activation associated with response inhibition istask-dependent. Neuropsychologia, 46, 224–232.
Smith, S. J., Souchay, C., & Conway, M. A. (2010). Overgeneral autobiographicalmemory in Parkinson’s disease. Cortex, 46, 787–793.
Stone, V. E., Baron-Cohen, S., & Knight, R. T. (1998). Frontal lobe contributions totheory of mind. Journal of Cognitive Neuroscience, 10, 640–656.
Stout, J. C., Rodawalt, W. C., & Siemers, E. R. (2001). Risky decision making inHuntington disease. Journal of the International Neuropsychological Society, 7,92–101.
Stuss, D. T., Gallup, G. G., Jr., & Alexander, M. P. (2001). The frontal lobes arenecessary for ‘Theory of Mind’. Brain, 124, 279–286.
Swainson, R., Rogers, R. D., Sahakian, B. J., Summers, B. A., Polkey, C. E., & Robbins, T.W. (2000). Probabilistic learning and reversal deficits in patients withParkinson’s disease or frontal or temporal lobe lesions: Possible adverseeffects of dopaminergic medication. Neuropsychologia, 38, 596–612.
Thiel, A., Hilker, R., Kessler, J., Habedank, B., Herholz, K., & Heiss, W. D. (2003).Activation of basal ganglia loops in idiopathic Parkinson’s disease: A PET study.Journal of Neural Transmission, 110, 1289–1301.
Toplak, M. E., Sorge, G. B., Benoit, A., West, R. F., & Stanovich, K. E. (2010). Decisionmaking and cognitive abilities: A review of associations between IowaGambling Task performance, executive functions and intelligence. ClinicalPsychology Review, 30, 562–581.
Torralva, T., Kipps, C. M., Hodges, J. R., Clark, L., Bekinschtein, T., Roca, M., et al.(2007). The relation between affective decision making and theory of mind infrontotemporal dementia. Neuropsychologia, 45, 342–349.
Tusruya, N., Kobayakawa, M., & Kawamura, M. (2011). Is ‘‘reading mind in the eyes’’impaired in Parkinson’s disease? Parkinsonism and Related Disorders, 17,246–248.
Van Overwalle, F. (2009). Social cognition and the brain: A meta-analysis. HumanBrain Mapping, 30, 829–858.
Walter, H., Adenzato, M., Ciaramidaro, A., Enrici, I., Pia, L., & Bara, B. G. (2004).Understanding intentions in social interaction: The role of the anteriorparacingulate cortex. Journal of Cognitive Neuroscience, 16, 1854–1863.
Xi, C., Zhu, Y., Niu, C., Lee, T. M., Tian, Y., & Wang, K. (2011). Contributions ofsubregions of the prefrontal cortex to theory of mind and decision making.Behavioral Brain Research, 221, 587–593.
Yeterian, E. H., & Pandya, D. N. (1991). Prefrontostriatal connections in relation tocortical architectonic organisation in rhesus monkeys. Journal of ComparativeNeurology, 312, 43–67.
Zald, D. H., & Andreotti, C. (2010). Neuropsychological assessment of the orbital andventromedial prefrontal cortex. Neuropsychologia, 48, 3377–3391.