opportunities to manipulate nutrient-by-water interactions in rainfed lowland rice systems
TRANSCRIPT

ELSEVIER Field Crops Research 56 (1998) 93-112
F i e l d Crops R e s e a r c h
Opportunities to manipulate nutrient-by-water interactions in rainfed lowland rice systems
L.J. Wade *, T. George, J.K. Ladha, U. Singh, S.I. Bhuiyan, S. Pandey International Rice Research Institute, P.O. Box 933, 1099 Manila, Philippines
Abstract
Water stress, accompanied by changes in soil aeration, severely limits rice productivity in rainfed systems. These factors affect nutrient availability. Nitrate (NO 3) that accumulates in aerobic soil is rapidly lost through leaching or denitrification in flooded soil. Green manures can act as NO 3 catch crops and legumes may gain additional N from biological N fixation. Direct seeding permits additional crops to be grown. Roots are commonly shallow in rainfed lowlands. It is not clear to what extent rice yields in rainfed lowlands are limited by water, nutrients, and the interactions between them, over diverse soil types, cultural practices and seasonal conditions. Research must determine what really limits root growth, water extraction and nutrient uptake. Some evidence suggests that manipulation of controlled-release fertilizer and root system development may be the key to optimizing nutrient release and capture in fluctuating environments. The potential for using strategic application of nutrients to buffer water limitation and stabilize yields must be examined. Models such as QUEFTS (Quantitative Evaluation of the Fertility of Tropical Soils) provide a potential framework for analyzing the effects of soil fertility and water availability on growth and yield of rice. © 1998 Elsevier Science B.V.
Keywords: Cropping systems; Genotype × environment interaction; Green manure; Nutrient X water interaction; Nutrient-use efficiency; Rice; Water deficit
1. Introduction
Rainfed r ice-growing areas cover approximately 69 M ha, representing 45% of the total area planted to rice worldwide (IRRI, 1993). In rainfed agricul- ture, crop performance is variable and unpredictable. Yields are strongly influenced by seasonal character- istics as well as by spatial heterogeneity over soil types, topographic sequences and agrohydrologic conditions. Water stress is commonly the most se- vere limitation to productivity of rice in rainfed rice
* Corresponding author.
systems (Widawsky and O'Toole , 1990), and grain yields average 1.0, 2.3 and 1.6 t ha-1 under upland, rainfed lowland and f lood-prone conditions, respec- tively (IRRI, 1993). In the uplands, soils remain aerobic throughout the season. In rainfed lowlands and flood-prone ecosystems, ponding and submer- gence may occur (Garrity et al., 1986). These chang- ing soil conditions have enormous consequences for nutrient availabili ty and the adaptive strategies that the plant may need for successful performance. Spa- tial heterogeneity and seasonal variabili ty in the tim- ing, duration and intensity of water stress, and its interactions with other factors, further complicate the development of strategies l ikely to be higher yielding
0378-4290/98/$19.00 © 1998 Elsevier Science B.V. All rights reserved. PII S0378-4290(97)00142- 1

94 L.J. Wade et aL / Field Crops Research 56 (1998) 93-112
on average, with less probability of crop failure. Developing these strategies is especially important in subsistence agriculture, where survival of the farmer and the family is largely dependent on the season's harvest (Ceccarelli, 1996).
Over 200 million of the world's poorest people depend upon the 40 M ha of rainfed lowland rice for their livelihood (IRRI, 1993). In the humid subtrop- ics of South and Southeast Asia, where rice systems are dominant, over half the population lives in poverty and average daily food intake is less than 2200 kcal per person. Because the lands are flooded during the rainy season, rice is the only crop that can be grown on most of the land. Any increase in rice productivity under such conditions would, therefore, have a large impact on rural poverty in Asia. Rainfed lowland rice must contribute substantially to meeting the projected 70% increase in demand for rice by the year 2030. Indeed, it is estimated that rainfed rice must contribute about 30% of this increase (Scobie et al., 1993). To do so, yields in the rainfed lowlands must increase from 2.3 t ha- 1 to just 3.5 t ha- 1. The challenge is how to increase yields in a system that until now has defied modest attempts at productivity increases.
Rainfed lowland rice encounters an environment more complex and unpredictable than most crops. It is grown in bunded fields without water control, and it, therefore, experiences hydrologic conditions fluc- tuating from complete submergence of the crop to drought, often during the same growing season. Such severe changes have marked effects on soil condi- tions and the availability of nutrients and water. The alternating periods of soil oxidation and reduction resulting from changing hydrologic conditions lead to gaseous loss of N and immobilization of other nutrients, together with changes in soil acidity and the concentrations of toxic Fe and A1. Periods with- out rain may be more damaging than expected be- cause water infiltration and storage at depth in pud- dled soils may be low, as may be depth of rooting. The effects of drought may be compounded by poor nutrition and soil chemical toxicities (Burford et al., 1989).
The traditional rice cultivars that predominate in these areas can tolerate these stresses to some de- gree, but they generally have low yield potentials (Mackill, 1986). Farmers growing unresponsive cul-
tivars and faced with the risk of crop loss are reluc- tant to invest in inputs. The rice crop must, therefore, commonly subsist on native soil nutrients and water, or residual nutrients from fertilizer applied to a previous cash crop. Nutrient availability is unlikely to be synchronous with crop demand, however, and much of the available nutrients that accumulate in fallow periods may be lost from the system (Buresh et al., 1989; George et al., 1993).
Loss of resources not only limits productivity in the rainfed lowland system, but also harms ecosys- tems elsewhere. For example, nitrate (NO 3) derived from the decomposition of soil organic matter in the fallow period before the rice crop and subsequently leached off the land with the early rains may accu- mulate in groundwater or surface water. Runoff may lead to flooding in low-lying areas, and if accompa- nied by soil erosion, may lead to siltation and salin- ization. Thus, greater efficiency of resource capture would improve productivity, enhance system sustain- ability and, at the same time, reduce environmental damage.
There is a trend in the rainfed lowlands to replace transplanting of rice with direct seeding on moist soil (Fujisaka et al., 1993). Direct seeding requires less labor and permits earlier sowing because less rain is needed than for flooding. This may enable critical stages of rice development to be synchronized more closely with periods of more reliable rainfall, thus improving the stability of rice production. Direct seeding may also permit a second crop to be grown before or after the main crop of rice, where other- wise it may not be possible. If the additional crop is a legume, this may result in a net input of biologi- cally fixed N into the system (George et al., 1994). Earlier sowing may improve the chances that bio- logically fixed N or N accumulated in the fallow soil will be taken up by the crop before it is lost through leaching and denitrification when the soil is flooded. Direct seeding, therefore, opens up a number of opportunities for improving resource capture, and it is expected that direct seeding will become increas- ingly important. Success will be dependent, how- ever, on improving the reliability of crop establish- ment, and on reducing losses due to greater weed competition. Thus, integrated research is needed to link soil preparation, fertilizer application, weed con- trol and crop duration to ensure establishment of a

L.J. Wade et al./ Field Crops Research 56 (1998) 93-112 95
vigorous, competitive stand of rice. We will require a much better understanding of the underlying pro- cesses of water extraction and nutrient uptake under the fluctuating conditions of the rainfed lowlands than we have at present.
In the past, the problems of a difficult and vari- able environment coupled with extreme rural poverty dictated that few research resources were targeted to the rainfed lowlands. This was a correct decision at the time, in that probabilities of success were then low. Now, however, with the rapid growth of agri- cultural research technologies, new research tools are available, and these promise to help raise productiv- ity substantially in the less favored environments. Time-domain reflectometry (TDR) for measurements of soil water content and capacitance measurements for root system volume (Chloupek, 1972, 1977; J. van Beem and M. Smith, Cornell University, per- sonal communication, 1995) are two examples. Ad- vances in microelectronics allow sophisticated mea- surements and experiments to be conducted in often-remote ralnfed environments. Nutrient uptake by rice roots may now be simulated using mixed-bed ion-exchange resins. Pattern analysis, computer sim- ulation models and geographic information systems permit extrapolation of experimental results and sim- ulation of technology performance over sites and years that previously would have required pro- hibitive replication of experiments. New polymer- coated controlled-release fertilizers offer the oppor- tunity to provide a steady supply of nutrients to the rice crop, buffering environmental fluctuations through the season. The rapid progress in biotech- nology, particularly in rice, promises to simplify breeding for complex traits, and offers the possibility of bringing specific stress tolerance genes to rice from other Oryza species.
A cropping system that aims to combine maxi- mum capture of water and nutrient resources with minimum use of external inputs is only sustainable if the inputs and outputs of water and nutrients are balanced at the level of a particular target yield. In general, technologies to improve productivity in rain- fed lowland rice-based systems would have several components: (a) cultivars that are generally adapted to the seasonal pattern of the ralnfed lowland system, by combining an ability to escape or tolerate the principal stresses of submergence and drought; (b)
management options that increase the availability of existing soil and water resources to the rice crop and that open the possibility of intensifying and diversi- fying rainfed lowland agriculture through use of other crops; and (c) cultivars that are able to capture more available soil nutrients and water, and are responsive to inputs, so that there would be a good return on farmer investment.
For nutrient-use efficiency in rainfed lowland ecosystems, the special challenge is the changeability of the water regime. Many of the soils are inherently low in fertility. Fertilizer applied close to the soil surface may not be available when the soil is dry, or may be lost by leaching or denitriflcation in subse- quent wetting and drying cycles. Conversely, if wa- ter is available but nutrients are limiting, the crop may not be able to utilize the water efficiently. Root development may be critical in this regard. These issues are reviewed in the following sections. Nutri- ent and water issues at the cropping systems level are considered first, before exploring nutrient-by- water interactions, nutrient management, nutrient re- lease and capture, and root ecophysiology in more detail. Finally, issues for further research attention are identified.
2. Soil nitrogen dynamics and the role of legumes
Water, either from rainfall or supplemental irriga- tion, determines the type of rice culture and prospects for other crops in the rotation. George et al. (1992) distinguished three broad classes of tropical lowland rice culture in terms of the annual period in which the soil is not flooded (continuously flooded, short non-flooded and long non-flooded), and how this influences soil N dynamics. A change from a flooded to a non-flooded soil brings about a transition in soil aeration status from anaerobic to aerobic. The dura- tion of the flooded (or non-flooded) period and the transition from aerobic to anaerobic aeration status has a major impact on accumulation and dissipation of soil mineral N (Buresh and De Datta, 1991; George et al., 1992). Ammonium (NH 4) is the pre- dominant soil mineral N form in continuously flooded or saturated rice systems, while NO 3 is the major form in non-flooded systems. The longer the dura-
96 L.J. Wade et al. / Field Crops Research 56 (1998) 93-112
©
<
o
e~
6
b
g m
b
el)
g
e~
e
z ~
m
Z ~
e~
[--,
I I + I [ I J I I I I I
~ ~ ~ i : ~
~D ~ z
z ~ , - .~ ©,..~
~~
z ~
~, ~ .
~ = ~ . ~
Z r ~
z = ~
~._~ = az,
L.J. Wade et al. / Field Crops Research 56 (1998) 93-112 97
tion of flood or non-flood, the more would be the likely accumulation of the respective mineral N form, provided there is no loss or plant removal from the soil.
Pre-rice NO3-N levels in Philippine lowland rice fields measured by Buresh et al. (1989) ranged from 39 to 91 kg NO3-N ha -1 in the top 60 cm soil. Buresh and De Datta (1991) proposed a potential role for plants in conserving pre-rice soil N O 3. Sub- sequently, George et al. (1992) developed a concep- tual understanding of soil N dynamics and the role of plants, particularly legumes, in conserving soil and biologically fixed N (BNF-N) in rainfed lowlands. According to that scheme, NO 3 that accumulates in aerobic soil during the dry season is lost during the aerobic to anaerobic transition by denitrification and leaching unless N O 3 is assimilated and held in plant or microbial biomass. When legumes are grown before rice, additional N input may be possible through BNF. Any net BNF-N contribution is depen- dent, however, on the facultative nature of legume systems; the legume will scavenge available soil mineral N first and supplement it with BNF-N if needed. Shortly after soil flooding, the amount of mineral N (NO 3 and NH 4) in the puddled soil layer would be negligible, since by then, NO 3 is lost by denitrification and leaching but NH 4 is yet to accu- mulate as aerobic transformation to N O 3 has just ceased. Ponding of water in the field was not essen- tial for NO3-N losses to occur (Singh et al., 1995a). When water-filled porosity exceeded 70%, NO3-N was lost from all soil layers. Since N did not accu- mulate in deeper soil layers, losses of NO3-N were attributed to denitrification. Although not measured here, immobilization of NO 3 was expected to be a minor cause of N loss, and this was subsequently verified by the groundwater studies of Gumtang et al. (1998).
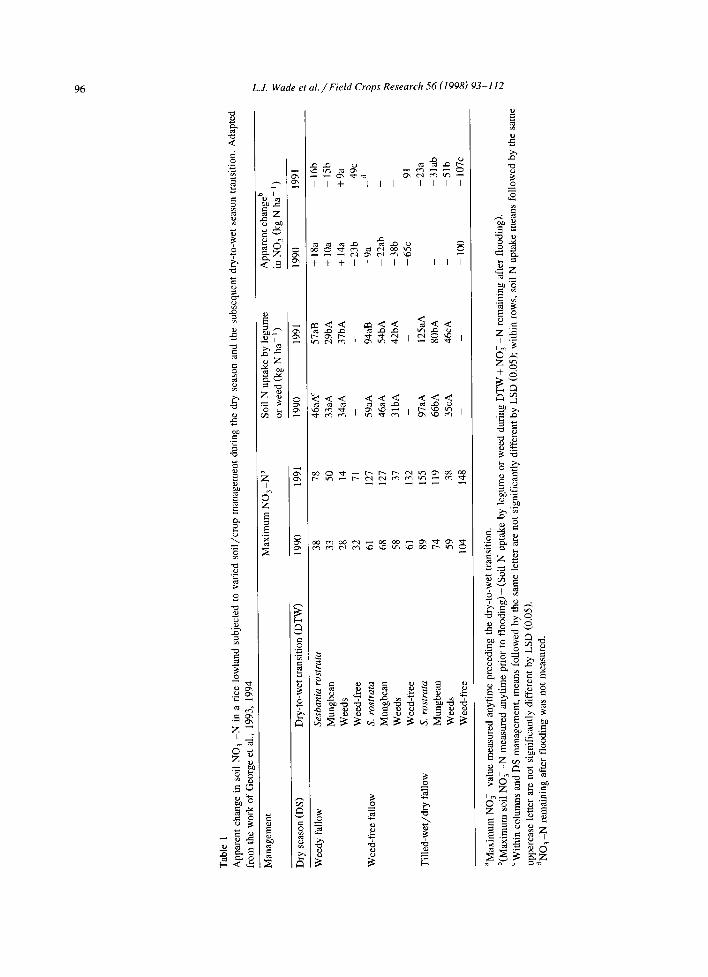
Lowland soils have immense capacity to accumu- late soil NO 3 during the aerobic soil phase but the amount accumulated varies with soil management (Buresh et al., 1993). George et al. (1993) measured soil NO 3 in excess of 150 kg N ha -1 in treatments where the impact of intensive dry-season cropping on N mineralization was simulated by repeated tillage and water application (Table 1). Although the magni- tude and range of N O 3 buildup observed in this particular soil may not represent all lowland rice
growing soils, the results demonstrate clearly the large potential of management to influence N O 3
buildup. Buresh et al. (1993) were able to show the potential for reduction in NO 3 loss if flooded rice was preceded by either weedy fallow or S. ros tra ta
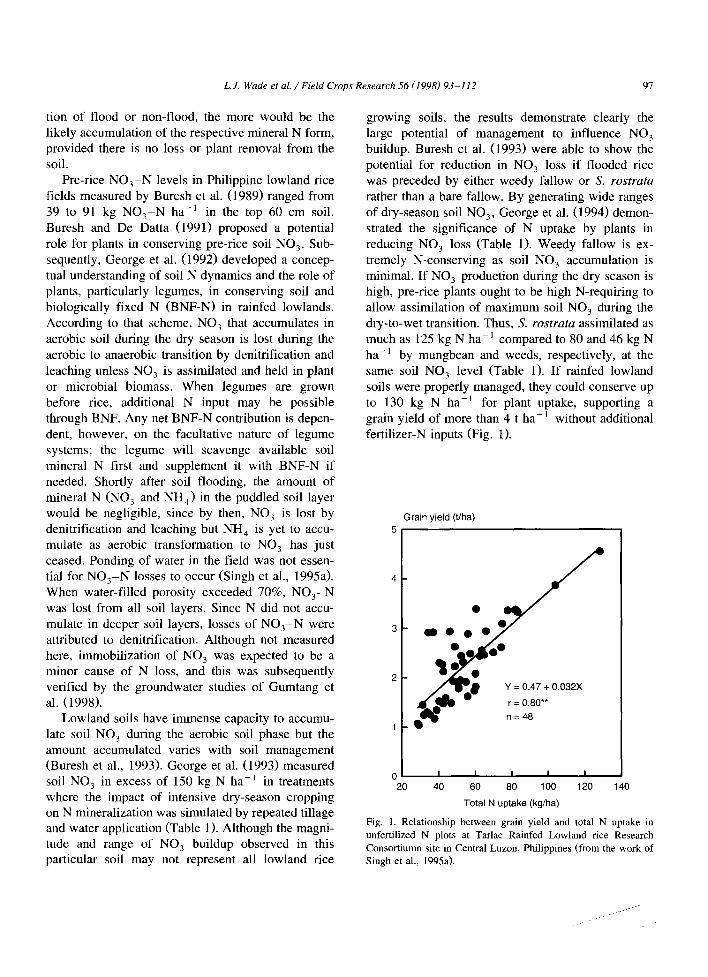
rather than a bare fallow. By generating wide ranges of dry-season soil NO3, George et al. (1994) demon- strated the significance of N uptake by plants in reducing NO 3 loss (Table 1). Weedy fallow is ex- tremely N-conserving as soil NO 3 accumulation is minimal. If N O 3 production during the dry season is high, pre-rice plants ought to be high N-requiring to allow assimilation of maximum soil N O 3 during the dry-to-wet transition. Thus, S. ros tra ta assimilated as much as 125 kg N ha -1 compared to 80 and 46 kg N ha-1 by mungbean and weeds, respectively, at the same soil NO 3 level (Table 1). If rainfed lowland soils were properly managed, they could conserve up to 130 kg N ha -~ for plant uptake, supporting a grain yield of more than 4 t ha- l without additional fertilizer-N inputs (Fig. 1).
Grain yield (t/ha) 5
Y -- 0.47 + 0.032X
r = 0.80**
n =48
0 I I I I ! 20 40 60 80 100 120 140
Total N uptake (kg/ha)
Fig. 1. Relationship between grain yield and total N uptake in unfertilized N plots at Tarlac Rainfed Lowland rice Research Consortiumn site in Central Luzon, Philippines (from the work of Singh et al., 1995a).
98 L J. Wade et al. / Field Crops Research 56 (1998) 93 - 112
o
,fl .=.
o z
o 2,
~..~ o
r~
<.~ ~
~ z
- Z E
N N z ~
"7 ! ""
~ ~
o .~ I = ~
z
,..o
< ~
=.~
o ~
N N N N N N ~
= ~
Z ~ ~"~
< ~ . ~ . Z . ~
~ -
~ . ~
.~ ~.~ ~
m ~
og
? o o
,~ ~ ~.=- .
£ ' " ~ o
v ~ o "'=z
. ~
II ~6 ~ . ~ ~ c~,.~
. = ~ < ~ . _

L.J. Wade et al. / Field Crops Research 56 (1998) 93-112 99
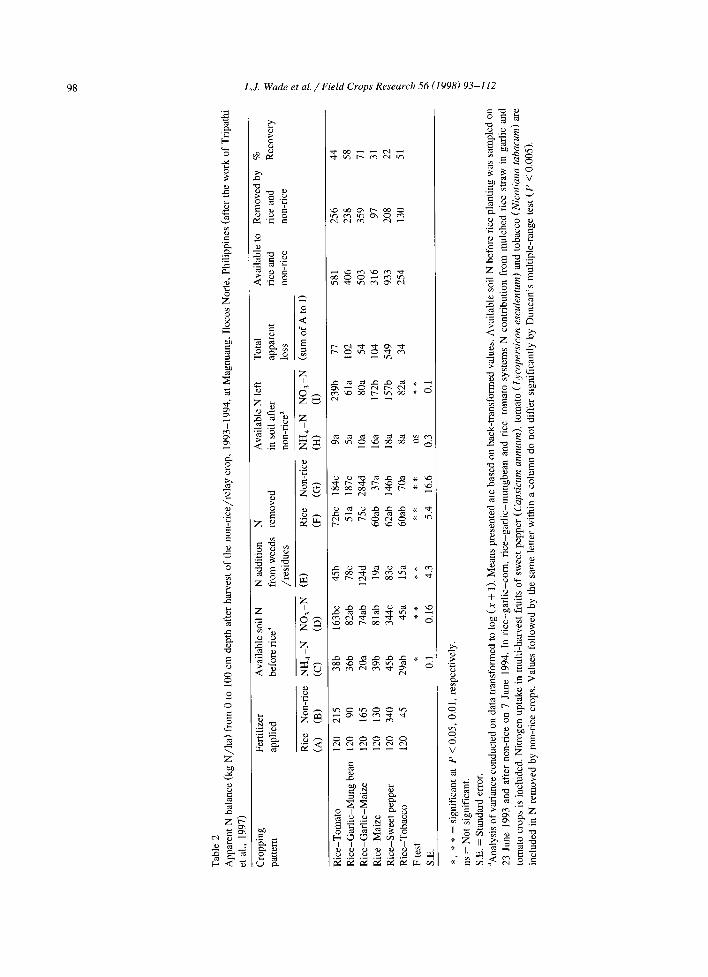
Where supplemental irrigation water is available, the rainfed lowlands are undergoing rapid crop inten- sification, with greater additions of fertilizer N and other inputs applied to high value cash crops such as vegetables. Tripathi et al. (1997) examined N dy- namics and N balance in intensified rice-upland cropping systems at three sites in Ilocos None, Philippines, where high rates of N are applied to the high-value, supplementary-irrigated, dry-season crop (Table 2). Consistent with previous studies (Buresh et al., 1989; George et al., 1993), pre-rice NO 3 accumulation was substantial in these systems amounting to as much as 344 kg ha-1 in the case of a heavily N-fertilized sweet-pepper crop (Table 2). The total amount of N recovered by rice and non-rice crops ranged from 20 to 71% of the 254 to 933 kg N ha-1 estimated to be the minimum N supply avail- able from all N sources (fertilizer, soil and weed /c rop residues. Minimum estimated N loss un- der various cropping systems ranged from 13 to 78% of that N supply. Most of this loss occurred during a brief period following soil flooding for rice produc- tion.
The variations in N available and N lost were largely due to differences in fertilizer N inputs and crop N uptake during the dry and the dry-to-wet transition seasons. Nitrogen loss was the greatest when high fertilizer N application relative to crop N uptake was also associated with the absence of suffi- cient weed or crop vegetation during the dry-to-wet transition. In one instance, it was estimated that the apparent N loss would have been only 54 kg ha - instead of 303 kg ha-~ if a third crop of maize had been included in a rice-garlic system. Sowing of a third crop after a high-value, high-input main dry season crop seems to be a good strategy to catch and recycle residual mineral N from the main crop. This
is consistent with the 'nitrate catch crop' concept developed by George et al. (1994).
Recent research has established the previously undocumented role of pre-rice green manure legumes in conserving soil N while gaining additional N from biological N fixation (George et al., 1994). Legumes, because of their greater N requirements per unit of dry matter, were better than weeds and other crops in capturing soil N when large amounts of soil NO 3 were carried over from the dry season. High soil N uptake reduced the amount of legume BNF, but did not lead to negative N gain as soil NO 3 would have been lost from the system otherwise. Nitrogen de- rived from BNF accounted for a major portion of the recycled N only when a legume followed a weedy fallow, because NO 3 was depleted by the weeds (George et al., 1994).
Regardless of whether soil N O 3 - N or BNF con- tributed to green manure N, the application of large quantities of it to rice is unlikely to increase N uptake (Furoc and Morris, 1989; Becker et al., 1990). In experiments of George et al. (1994), S. r o s t r a m
green manure contained 134 to 222 kg N ha -1 compared to 31 to 46 kg N ha- i in weed biomass and 30 to 77 kg N ha -1 in mungbean harvest residues. Residue N recovered by rice was dispropor- tionately less than what it could have been at the level of N applied (Table 3). Across the 2 years, only 20% of S. ros t ra ta N, 24% of mungbean residue N, and 16% of weed N were recovered by the rice crop. A substantial portion of residue N was found to remain below-ground in soil, rice roots and unde- composed residue fragments. Based on 15N estimates from labelled microplots, an average of 32% of the S. ros t ra ta N was unaccounted for, presumably lost as ammonia from the flood water, thus only postpon- ing N loss from the legume crop period to the rice
Table 3 Nitrogen-15 based estimates of disposition of in situ-produced residue N applied to flooded rice (T. George et al., unpublished)
Pre-rice crop Residue N Rice N Residue N Residue N Residue N (kg N ha 1) (kg N ha -l) in rice a remaining in soil b unaccoun~dfor
Sesban~ 208 c 109 20 47 67 Mungbean 62 78 24 62 85 Weeds 41 58 16 107 123
aNitrogen-15 isotope based estimates. bEstimates based on below-ground N in rice + undecomposed residue materials in the top 20 cm soil layer. c Values averaged across 2 years.

100 L.J. Wade et al. / Field Crops Research 56 (1998) 93-112
crop period. Most of the mungbean and weed N, however, was found to remain in soil.
While yield gains are possible with legume green manures as an N source for rice, they have gained only limited acceptance by farmers, as N contribu- tion to rice alone is not a sufficient incentive to commit land, labor and resources for green manure production (Ladha and Garrity, 1994). Legumes with an immediate economic return are more attractive to farmers and, therefore, grain legumes are often pre- ferred to green manures. With grain legumes, only the harvest residues are available for recycling. Re- cycling of legume residues should be the norm un- less there are alternative uses for them, but harvest residues are less efficient sources of N for rice than green manures, because of wider C:N ratios (George et al., 1995). Further, grain legumes would probably serve the farmer better if management seeks to maxi- mize grain yield rather than residues. Nitrogen re- quirement of grain legumes is greater when grain yield is maximized, and as a result, greater acquisi- tion of soil NO3-N and greater BNF-N would result (George et al., 1992).
Forage legumes are also an option, as they are economically feasible both as animal feed and as an N source to rice. Effects of forage legumes grown during the dry season on soil N dynamics and crop productivity were studied by Ladha et al. (1996).
Forage biomass of pigeon pea (Cajanus cajan), sira- tro (Macropti l ium atropurpureum), desmanthus (Desmanthus indica), and clitoria (Clitoria ter- matea), harvested in several clippings amounted to 5 and 9 t ha-1 during a 5-month growth period (Table 4). This biomass corresponded to N accumulations of about 130 to 300 kg ha -1 with 100 to 200 kg N ha -1 derived from BNF. The last cut of forage, amounting to 3 to 5 t ha-1 and containing about 80 to 160 kg N ha - l , was recycled to rice as green manure. Average increase in rice grain yield was 1 t ha -1 , but this yield increase was achieved even with 3 t ha-1 biomass containing 90 kg N ha-1 in the
case of siratro, indicating that even more of the biomass could have been diverted to forage purposes without a negative impact on rice grain yield. The yield increase due to biomass incorporation was equivalent to yields obtained in weedy or weed-free fallow plots receiving 50 kg fertilizer N ha -1, again indicating that N substitution cannot be large even with excessive green manure application. There was, however, an apparent positive soil N balance of 50 kg N ha -1, including an estimated legume root
contribution of 23 kg N ha -1. Whether such a positive soil N balance is beneficial in the long term remains to be researched. It is unlikely that a positive N balance can be maintained if NO 3 loss during the aerobic to anaerobic transition is high. Measures are
Table 4 Biomass and N accumulation by dry season forage legumes and response of wet season rice to incorporation of last cuttings of forage legumes, IRRI Farm, Philippines (Ladha et al., 1996)
Legume No. of cuttings Total biomass Total N N in cutting Grain yield Total rice N Increase in rice N due to residue
1991 Pigeonpea 3 6.5b 154a 113ab 5.2ab 98ab 28 * * Crotalaria 6 7.6bc 277b 118b 5.6a 90ab 27 * * Clitoria 4 6.9bc 256b 114ab 5.2ab 110a 33 * * Desmanthus 4 8.0c 251b 137b 5.lab 89ab 25. * Siratro 3 4.9a 132a 81a 4.8ab 82ab 21 * *
1992 Pigeonpea 3 9.0c 235b 143bc 5.0ab 92ab 21 * * Crotalaria 6 7.8b 279b 117ab 5.lab 87b 36 * * Clitofia 4 7.7b 306c 162ab 5.3a 101a 44 * * Desmanthus 3 9.1c 283c 161c 4.8bc 82bc 35. * Siratro 3 5.5 178a 99a 4.8bc 72c 18 * *
Values in a column followed by a common letter are not significantly different by DMRT. * * Significant at 1% level.

L.J. Wade et aL /Field Crops Research 56 (1998) 93-112 101
needed that will convert dry season soil NO 3 into usable products including green manure in moderate amounts.
3. Water balance and movement in rainfed low- land systems
The water status in the ricefield, whether rainfed or irrigated, greatly influences nutrient-use efficiency as well as nutrient balances in crop-soil systems. A part of the water input in excess of the amount evapotranspired by the soil-crop system is retained in the soil profile within the root zone, while the remaining part percolates downwards and eventually reaches the groundwater. The process of percolation or leaching to the subsoil is accelerated as more water accumulates. Through convection or mass flow, all dissolved chemicals can thus be transported from various depths in the soil to deeper layers and ulti- mately to the groundwater. Some of the N applied to rice fields is rapidly transformed into NO 3 under normal soil conditions and, being very mobile in most soils, is moved downwards with the percolating water. When nitrogenous fertilizers are used at a high rate and there is excess water on the soil, more NO 3 is expected deeper in the soil profile (Ladha et al., 1996).
As cropping intensity increases and more N fertil- izers are used to obtain higher yields, contamination of fresh groundwater by NO3-N may increase. The possibility of excessive NO3-N in shallow ground- water in rural areas must be a concern because shallow aquifers are the source of water for domestic use in the rural areas. Drinking water containing in excess of 0.70 mM NO3-N is considered hazardous for human health (Viets and Hageman, 1971) in many countries. The European Community adopted a standard for acceptable limit of NO 3 in drinking water that is equivalent to 0.80 mM NO3-N (Coun- cil of European Communities, 1980). High concen- trations of NO3-N can cause methaemoglobinaemia (blue baby syndrome) in infants and carcinogenic effects, especially stomach cancer, in adults (Mercado, 1976; Follet and Walker, 1989; Addiscott et al., 1991).
An IRRI study conducted from 1989 to 1991 determined the NO3-N concentrations in shallow groundwater within two irrigated rice areas that have been intensively cultivated (rice-rice system) for more than 30 years and for two adjacent rainfed areas (rice-fallow system) in Laguna and Nueva Ecija provinces, Philippines. Farmers pumped groundwater for domestic consumption from shallow aquifers using manually operated tubewells that were only 6 to 12 m deep. Water samples for N O 3 analy- sis were drawn from water pumped from these tube- wells. Field surveys indicated that farmers' average N use was 81 kg ha- l in dry and 111 kg ha- l in wet seasons in Laguna, and 78 and 100 kg ha -1 in dry and wet seasons in Nueva Ecija.
The NO3-N analysis revealed that 61 to 100% of the groundwater samples in the dry season in both irrigated areas had NO3-N present in them. In the wet season, 10 to 31% of the samples had NO3-N. The average concentrations of NO3-N were 0.05 mM in the wet season and 0.04 mM in the dry season in Laguna, and between 0.00 and 0.03 mM in both seasons in Nueva Ecija, which are very low relative to the recommended upper limit of 0.70 mM. In the rainfed areas, groundwater samples had com- parably low average concentrations. The highest con- centrations were 0.19 mM in Laguna irrigated area, 0.16 mM in Nueva Ecija irrigated area, and 0.06 and 0.11 in their respective rainfed counterparts. Be- tween the two irrigated areas, Laguna had lighter- textured soils (loam and clay) and consequently higher percolation rates (ave. 7.5 mm day -1) than Nueva Ecija (silty clay loam to clay; ave. percola- tion: 4.9 mm day- l ) . This may be the reason for the slightly higher concentrations of NO3-N in Laguna (Castaneda and Bhuiyan, 1991).
The results indicated that current levels of NO 3 in the groundwater of the study area are much lower than the recommended maximum for human con- sumption. The NO 3 leaching processes in rice-based cropping systems need to be clearly understood if we are to predict future concentrations of the contami- nant in the groundwater resource under various alter- native water and nutrient management options. Simu- lation modeling may be an appropriate tool for pre- diction. Nevertheless, these concentrations are ex- tremely low and NO3-N concentrations are not ex- pected to be a problem where rainfall is high.
102 L.J. Wade et &/Field Crops Research 56 (19981 93-112
4. Analysis of risk for alternative nutrient strate- gies
The seasonal variability and spatial heterogeneity of the rainfed lowlands necessitates use of crop models and geographic information systems (GIS) to examine the likely consequences of change in cul- tural practices or cultivar characteristics. It is neces- sary to identify strategies that are less subject to crop failure, but are able to exploit the favorable season when it occurs. Risk assessment, economic analysis and socioeconomic impact should also be consid- ered. Probability distributions of grain yield relative to long-term meteorological data, or extrapolation domains for a proposed technology, are likely targets for such research. Work is currently in progress to utilize these tools to understand genotype-by-en- vironment interaction in the rainfed lowlands (Wade et al., 1995, 1996). Comparable activity is proposed for nutrient interactions in these complex environ- ments (Fukai et al., 1995; Singh et al., 1995a,b).
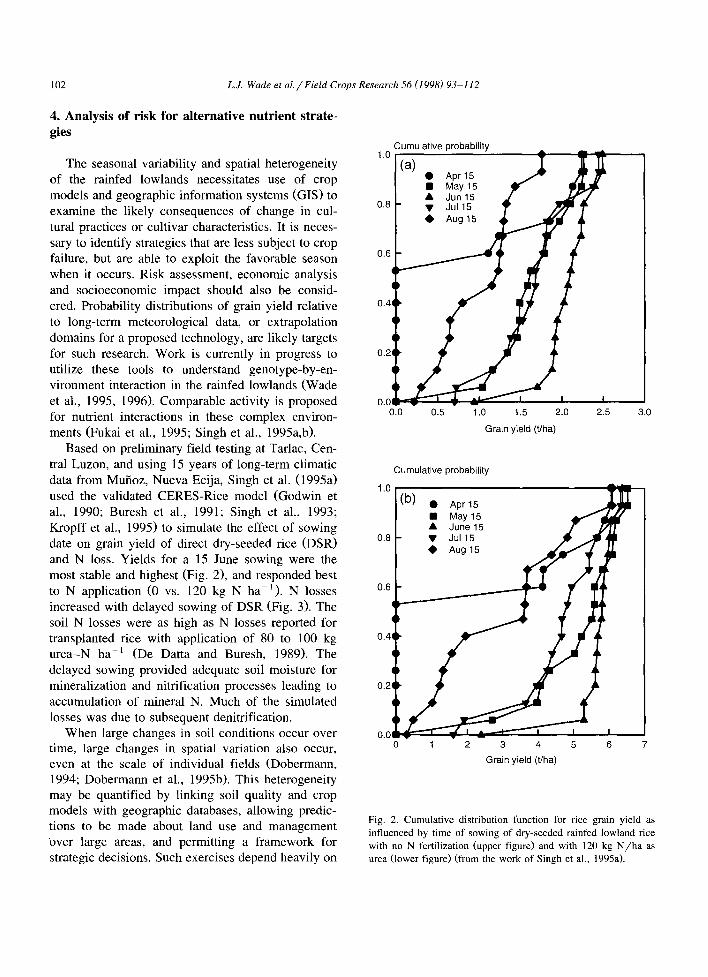
Based on preliminary field testing at Tarlac, Cen- tral Luzon, and using 15 years of long-term climatic data from Muiioz, Nueva Ecija, Singh et al. (1995a) used the validated CERES-Rice model (Godwin et al., 1990; Buresh et al., 1991; Singh et al., 1993; Kropff et al., 1995) to simulate the effect of sowing date on grain yield of direct dry-seeded rice (DSR) and N loss. Yields for a 15 June sowing were the most stable and highest (Fig. 2) and responded best to N application (0 vs. 120 kg N ha-’ ). N losses increased with delayed sowing of DSR (Fig. 3). The soil N losses were as high as N losses reported for transplanted rice with application of 80 to 100 kg urea-N ha-’ (De Datta and Buresh, 1989). The delayed sowing provided adequate soil moisture for mineralization and nitrification processes leading to accumulation of mineral N. Much of the simulated losses was due to subsequent denitrification.
When large changes in soil conditions occur over time, large changes in spatial variation also occur, even at the scale of individual fields (Dobermann, 1994; Dobermann et al., 1995b). This heterogeneity may be quantified by linking soil quality and crop models with geographic databases, allowing predic- tions to be made about land use and management ‘over large areas, and permitting a framework for strategic decisions. Such exercises depend heavily on
0.6
0.4
0.2
0.0 (
Cumulative probability
1 .o 1.5 2.0 2.5 3.0
Grain yield (t/ha)
Cumulative probability
8 May15 A June15
0.8 - v Jul15
0.0 0 1 2 3 4 5 6 7
Grain yield (t/ha)
Fig. 2. Cumulative distribution function for rice grain yield as
influenced by time of sowing of dry-seeded rainfed lowland rice
with no N fertilization (upper figure) and with 120 kg N/ha as urea (lower figure) (from the work of Singh et al., 1995a).
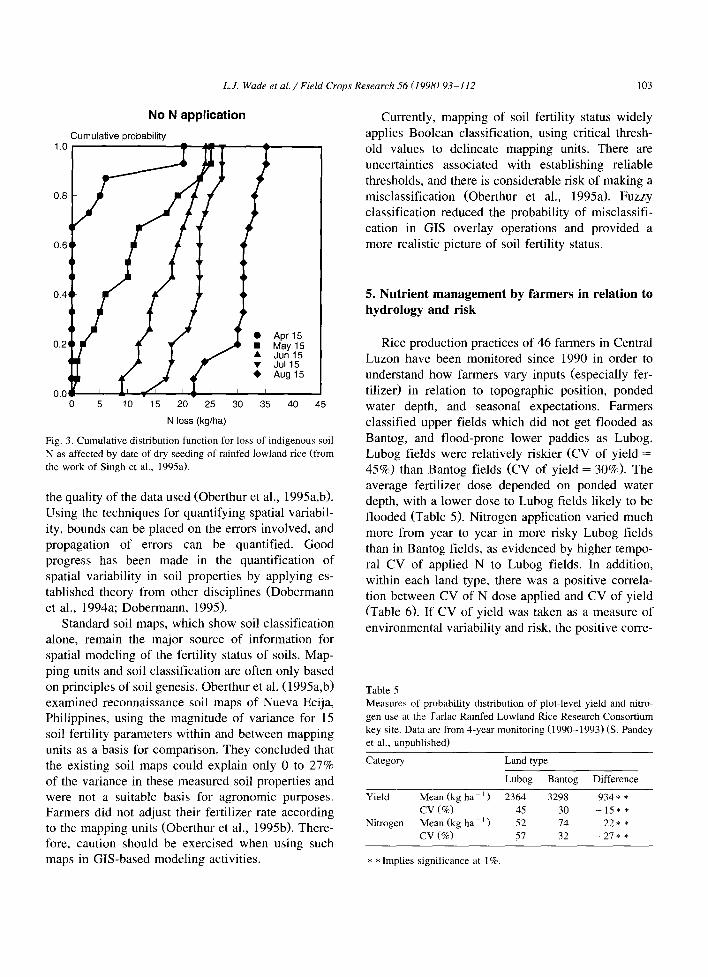
LJ. Wade et a l . / Field Crops Research 56 (1998) 93-112 103
No N appl icat ion
Cumulative probability 1.0
0.8
qr 0.6q~
0.4
0.2 ~ ~ • ~ • May 15 ( 1 _1_ ~ • Jun 15 ' • F Y • Jul 15
0.0 5 10 15 20 25 30 35 40 45
N loss (kg/ha)
Fig. 3. Cumulative distribution function for loss of indigenous soil N as affected by date of dry seeding of rainfed lowland rice (from the work of Singh et al., 1995a).
the quality of the data used (Oberthur et al., 1995a,b). Using the techniques for quantifying spatial variabil- ity, bounds can be placed on the errors involved, and propagation of errors can be quantified. Good progress has been made in the quantification of spatial variabili ty in soil properties by applying es- tablished theory from other disciplines (Dobermann et al., 1994a; Dobermann, 1995).
Standard soil maps, which show soil classification alone, remain the major source of information for spatial modeling of the fertility status of soils. Map- ping units and soil classification are often only based on principles of soil genesis. Oberthur et al. (1995a,b) examined reconnaissance soil maps of Nueva Ecija, Philippines, using the magnitude of variance for 15 soil fertility parameters within and between mapping units as a basis for comparison. They concluded that the existing soil maps could explain only 0 to 27% of the variance in these measured soil properties and were not a suitable basis for agronomic purposes. Farmers did not adjust their fertilizer rate according to the mapping units (Oberthur et al., 1995b). There- fore, caution should be exercised when using such maps in GIS-based modeling activities.
Currently, mapping of soil fertility status widely applies Boolean classification, using critical thresh- old values to delineate mapping units. There are uncertainties associated with establishing reliable thresholds, and there is considerable risk of making a misclassification (Oberthur et al., 1995a). Fuzzy classification reduced the probabil i ty of misclassifi- cation in GIS overlay operations and provided a more realistic picture of soil fertility status.
5. Nutrient management by farmers in relation to hydrology and risk
Rice production practices of 46 farmers in Central Luzon have been monitored since 1990 in order to understand how farmers vary inputs (especially fer- tilizer) in relation to topographic position, ponded water depth, and seasonal expectations. Farmers classified upper fields which did not get f looded as Bantog, and flood-prone lower paddies as Lubog. Lubog fields were relatively riskier (CV of yield = 45%) than Bantog fields (CV of yield = 30%). The average fertilizer dose depended on ponded water depth, with a lower dose to Lubog fields l ikely to be flooded (Table 5). Nitrogen application varied much more from year to year in more risky Lubog fields than in Bantog fields, as evidenced by higher tempo- ral CV of applied N to Lubog fields. In addition, within each land type, there was a positive correla- tion between CV of N dose applied and CV of yield (Table 6). I f CV of yield was taken as a measure of environmental variabili ty and risk, the positive corre-
Table 5 Measures of probability distribution of plot-level yield and nitro- gen use at the Tarlac Rainfed Lowland Rice Research Consortium key site. Data are from 4-year monitoring (1990-1993) (S. Pandey et al., unpublished)
Category Land type
Lubog Bantog Difference
Yield Mean (kg ha 1 ) 2364 3298 934, * CV (%) 45 30 - 15 * *
Nitrogen Mean (kg ha l ) 52 74 22. * CV (%) 57 32 - 27 * *
* * Implies significance at 1%.

104 L.J. Wade et a l . / Field Crops Research 56 (1998) 93-112
Table 6 Regression coefficients reflecting the effects of yield variability on variability in nitrogen use (S. Pandey et al. unpublished)
Land type Intercept CV of R 2 Sample yield size
Lubog (flood-prone) 14.06 1.24. * 0.35 25 (20.10) (0.35)
Bantog (not flood-prone) 17.73. * 0.25. 0.07 76 (3.14) (0.10)
lation indicated that farmers adjusted N application by a greater amount in more risky fields.
In-depth interviews indicated that factors that farmers considered most critical to N management were the field hydrology and crop growth stage. Very often, the depth of standing water was used as a guide for deciding the date of application. If the depth of water was too low or too high (flooded), farmers reported that they would reduce fertilizer dose or skip it altogether. Farmers also adjusted fertilizer dosage to the condition of the crop. Some- times an additional quantity was applied in an at- tempt to encourage tillering to compensate for a poor stand. Thus, although farmers may not base N appli- cation on the N-supplying capacity of the soil (Cass- man et al., 1993), they make conscious attempts to adjust N application to a perceived demand by the crop. Crop models may be used to assist this process by providing probability distributions of yield expec- tations for alternative nutrient strategies at different locations.
6. Nutrient-by-water interactions in rainfed low- land systems
In the past, most research effort has been placed on N as the limiting nutrient in rice systems. This was largely because, in favorable irrigated systems that have received external nutrient inputs for many years, responses to nutrients other than N have been less dramatic. But, in the rainfed lowlands where soils are less fertile and more permeable and nutrient inputs are smaller, deficiencies of other nutrients are widespread, particularly those of P and Zn. The mobility of sparingly soluble nutrients such as P is much reduced in dry soils, so that their supply is critical in crop establishment under dry-seeded con-
ditions. Further nutrient constraints may compound the effects of drought, and conversely, a constant nutrient supply may alleviate the effects of drought in some circumstances (Biswas et al., 1982; Tan- guilig and De Datta, 1988; Otoo et al., 1989; Zaman et al., 1990).
Efficient nutrient use by the plant presupposes timely availability of adequate and appropriate nutri- ents and has two components: the efficiency of uptake and the efficiency of internal cycling and use. Uptake efficiency (the capture of limiting resources) is likely to be more important in the rainfed low- lands, given low rates of soil amendments. Only when adequate nutrients have been captured does physiological efficiency (the production of dry mat- ter per unit of uptake and its partitioning between stem and grain) become more important. Uptake efficiency has a number of components: root geome- try effects, owing to differences in rooting depth and density, and the proportion of fine roots and root hairs; solubilization effects, due to root-induced changes in soil chemical conditions and consequent increases in the solubility and hence plant-availabil- ity of particular nutrients; and microbial effects, such as infection with mycorrhizal fungi and other symbi- otic associations.
There is evidence for genotypic variation among rice cultivars in the uptake efficiency of various nutrients (Kirk et al., 1993; Hedley et al., 1994; Ladha et al., 1996). Recent research has demon- strated the importance of sobulization effects in the acquisition of phosphate (Kirk and Saleque, 1995; Saleque and Kirk, 1995) and zinc (Kirk and Bajita, 1995) by rice. Different solubilization mechanisms operate under moist, aerobic soil conditions com- pared with water-saturated, anaerobic conditions. If cultivar differences in uptake efficiency could be explained in terms of such mechanisms, simple screening techniques may be developed for breeding nutrient-efficient germplasm. Also, an understanding of these mechanisms would help in developing im- proved fertilizer management strategies.
7. Replaceability of nutrients and water in rainfed lowland systems
For rainfed lowlands, compared to other agricul- tural systems, it is not clear to what extent rice yields

L.J. Wade et al. / Field Crops Research 56 (1998) 93-112 105
are limited by nutrients, water, and the interactions between them, over the diverse soil types, cultural practices and seasonal conditions. Few of the many experiments have obtained the data essential for thorough interpretation, and none has attempted to develop an understanding of prospects for manipulat- ing nutrient X water interactions across the varied combinations of flood, drought, soil type and cultural practices. Experiments to address these issues com- menced during the 1995 wet season at a series of sites associated with the Rainfed Lowland Rice Re- search Consortium. The objectives were: (a) to quan- tify the effects of indigenous and applied nutrients on the efficiency of water use and the stability of grain yield in diverse rainfed lowland rice environ- ments, (b) to determine the relative importance of nutrients and water as yield constraints in these environments, (c) to examine prospects for using strategic applications of nutrients to stabilize yields against water limitation, and (d) to provide data for model validation and risk analysis.
Thirty sites were examined in 1995, including eight in Tarlac, Nueva Ecija, Philippines (15°N, 121°E), 10 in Ubon Ratchathani-Khon Kaen, Thai- land (16°N, 103-105°E), nine in Raipur-Faizabad, India (21-27°N, 82°E), and three in Jakenan-Re- mbang, Indonesia (4°N, I l l°E) . At each location, three replicates of each of six treatments were estab- lished: control (NIL); farmyard manure (FYM); PK; NPK; a controlled-release formulation of N, together with P and K (CR-NPK); and a complete nutrient dressing that included micronutrients (ALL). Fertil- izer rates were adjusted for different locations, but within each location any common element was ap- plied at the same rate over treatments. Typical appli- cation rates were 10 t ha- ~ of FYM; and 140, 40, 40 kg ha-1 of N, P and K, respectively. Data were obtained on: soil characteristics (description, particle size analysis, water retention properties, and nutrient status by layer); weather data (daily maximum and minimum temperature, radiation, evaporation, rain- fall); depth to the water table and depth of ponded water; crop phenology, height, and leaf rolling and leaf death scores; specific leaf N at mid-tillering, panicle initiation and flowering; and final dry weight, grain yield and grain N, P, and K content.
This research is still in progress, but some pat- terns of response have been revealed by cluster
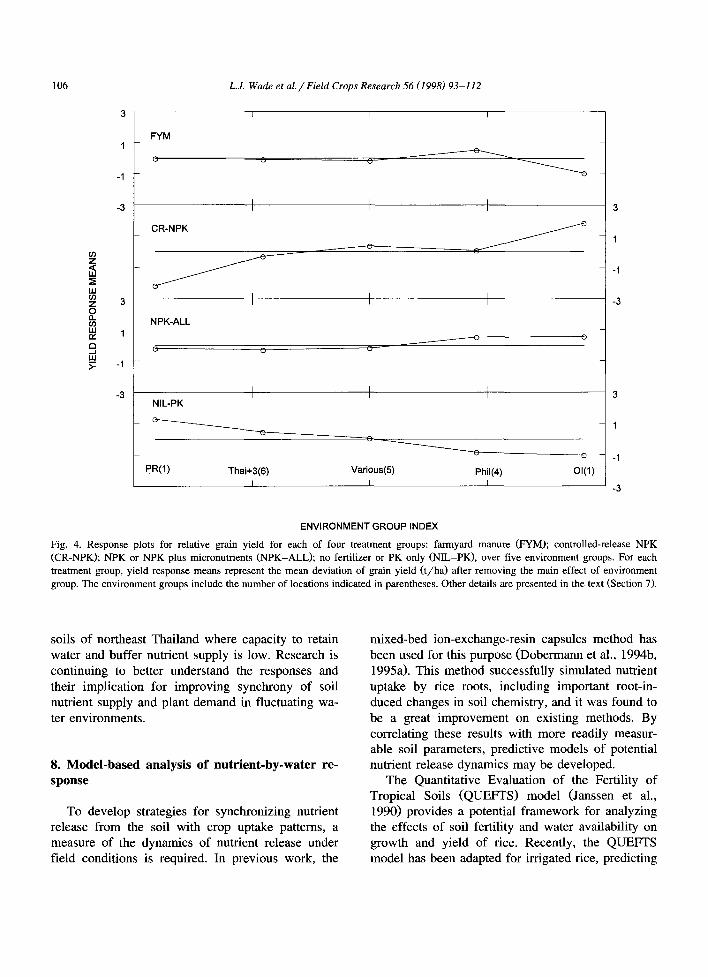
analyses of data from 17 of the sites sown in 1995. The treatment by location (T X L) interaction ac- counted for 24% of the total sum of squares (SS), with the main effects of treatments and locations responsible for 28 and 48%, respectively. More than 80% of T X L SS was captured in four treatment groups and five environment groups. Four treatment groups were identified, because (NIL and PK) and (NPK and ALL) had similar patterns of response over environments. The effect of adding micronutri- ents was thus similar across locations. Environment groups were differentiated by the patterns of yield response to the treatments imposed, and comprised a group of one location in Nueve Ecija, Philippines (PR), three sites in Thailand plus three other sites (Thai + 3), five sites in various locations (Various), four Philippine sites (Phil), and one Indonesian site (OI). Yield responses of the four treatment groups across the five environment groups are shown in Fig. 4.
The first three principal components, interpreted as representing inherent soil fertility (66%), respon- siveness to controlled-release fertilizer (21%) and responsiveness to farmyard manure (7%), accounted for 94% of the total variation. The importance of soil in defining environment groups is demonstrated by responses to NIL-PK in the bottom cell of Fig. 4, where environment groups PR and Thai + 3 had relatively higher yields (positive values), and groups Phil and OI had relatively lower yields (negative values). Response to controlled-release fertilizer was the next discriminant, with yields of environment group PR being strongly depressed, and of group OI being strongly increased by controlled-release fertil- izer. Finally, farmyard manure increased relative yield for the Phil group, but decreased relative yield for group OI. The general response to NPK-ALL did not further assist pattern definition, providing an opposite but smaller response relative to NIL-PK.
The importance of nutrient-by-water interactions is inferred from these results. Controlled-release fer- tilizer N should be less influenced by excess water than conventional fertilizers when floods occur, and may be expected to again release N with the next rain after a dry spell. Controlled-release NPK was observed to enhance crop survival after submergence at Ghagharaghat near Faizabad, India. Farmyard ma- nure was especially beneficial on coarse-textured
106 L.J. Wade et a l . / Field Crops Research 56 (1998) 93-112
-1
-3
z
I i i O3 z 3 o Q. O3 LU 1 n," a
~. -1
-3
FYM
,3
I I
G G
I CR-NPK
I I
J
NPK-ALL
0 0
f O
3
1
-1
-3
3 I I I NIL-PK
"O
PR(1 ) Thai+3(6) Various(5) Phil(4) I I
olo)
1
-1
-3
ENVIRONMENT GROUP INDEX
Fig. 4. Response plots for relative grain yield for each of four treatment groups: farmyard manure (FYM); controlled-release NPK (CR-NPK); NPK or NPK plus micronutrients (NPK-ALL); no fertilizer or PK only (NIL-PK), over five environment groups. For each treatment group, yield response means represent the mean deviation of grain yield ( t /ha) after removing the main effect of environment group. The environment groups include the number of locations indicated in parentheses. Other details are presented in the text (Section 7).
soils of northeast Thailand where capacity to retain water and buffer nutrient supply is low. Research is continuing to better understand the responses and their implication for improving synchrony of soil nutrient supply and plant demand in fluctuating wa- ter environments.
8. Model-based analysis of nutrient-by-water re- sponse
To develop strategies for synchronizing nutrient release from the soil with crop uptake patterns, a measure of the dynamics of nutrient release under field conditions is required. In previous work, the
mixed-bed ion-exchange-resin capsules method has been used for this purpose (Dobermann et al., 1994b, 1995a). This method successfully simulated nutrient uptake by rice roots, including important root-in- duced changes in soil chemistry, and it was found to be a great improvement on existing methods. By correlating these results with more readily measur- able soil parameters, predictive models of potential nutrient release dynamics may be developed.
The Quantitative Evaluation of the Fertility of Tropical Soils (QUEFTS) model (Janssen et al., 1990) provides a potential framework for analyzing the effects of soil fertility and water availability on growth and yield of rice. Recently, the QUEFTS model has been adapted for irrigated rice, predicting

L.J. Wade et aL / Field Crops Research 56 (1998) 93-112 107
the uptake of N, P and K and rice yield for actual site conditions with limited supplies of N, P and K (Dobermann et al., 1996). The model has been cali- brated with data from long-term fertility experiments in five countries. The impact of a change in soil fertility or in use of fertilizer may be simulated, facilitating more effective decisions about fertilizer rates. To provide a general framework for analyzing rainfed lowland environments, the model needs to be expanded to include the effects of water stress, and to allow for deficiencies and toxicities of other ele- ments.
To expand QUEFTS for rainfed lowland rice, a series of field experiments across a wide range of soil and climatic conditions is required. Using the QUEFFS model (Janssen et al., 1990), empirical relationships may be developed between soil nutrient supply, plant nutrient uptake, and grain yield of rice. These relationships, together with rice crop growth and development models (ORYZA-1, Kropff et al., 1993; ORYZA-W, Wopereis et al., 1995; and CERES-RICE, Ritchie et al., 1987), may then be used to simulate potential, water-limited, and soil fertility-limited rice yields for a range of sites. This approach would permit analysis of different fertilizer application scenarios, such as the use of controlled- release fertilizer to better match soil nutrient supply with crop demand in fluctuating water environments. Incorporation of sensitivity and risk analysis compo- nents would be necessary to account for the uncer- tainties associated with highly variable rainfed low- land environments. The nutrient × water experiments described previously would be suitable for this pur- pose, especially with additional data collection at key sites.
9. Root ecophysiology in relation to nutrients and water
Drought resistance may be conferred by three strategies: escape, avoidance and tolerance (Ludlow and Muchow, 1990). Plant breeders have been suc- cessful in exploiting drought escape by adjusting crop phenology to minimize coincidence of sensitive growth stages with expected periods of drought.
Breeders have been less successful in utilizing the more complex strategies of drought avoidance and drought tolerance, however, and are seeking help from physiologists and biotechnologists. Plants may avoid drought by being able to extract more water from deeper layers of soil, or by reducing losses during periods of stress. Plants may tolerate drought by physicochemical adjustments to offset the adverse effects of partial tissue dehydration.
Rice is commonly regarded as having a poor root system, often failing to extract water from deeper layers of the soil profile, relative to other crops (Fukai and Inthapan, 1988; Inthapan and Fukai, 1988). Yoshida and Hasegawa (1982) reported sig- nificant variation in root length density in rice below 30 cm. Upland cultivars with large deep root systems were able to extract more water, and tended to be taller and have fewer tillers. Similarly, Puckridge and O'Toole (1981) found that deep-rooted cultivars extracted more water from 40 to 70 cm than shallow-rooted cultivars. Consequently, several re- views have concluded that a drought avoidance strat- egy (a greater maximum rooting depth, a greater root length density at depth, and a greater capacity to conduct water from depth) would be helpful for upland conditions (O'Toole, 1982; Fukai and Cooper, 1995).
Understanding of root traits is less clear for other rainfed ecosystems (Fukai and Cooper, 1995). Roots are commonly quite shallow in rainfed lowlands, where conditions may change quickly from aerobic to anaerobic when rainfall is adequate to pond water in bunded fields (Sharma et al., 1995). While a deeper root system may be helpful for accessing resources from the soil profile, the ability of roots to penetrate non-puddled or compacted layers may be more important (Yu et al., 1995).
In many fields, subsoils are low in fertility and poor in water-holding capacity, so the quantity of resources available for capture from deeper layers may also be in question. Other strategies may also be helpful in these conditions. For example, a number of lines from rainfed lowland and flood-prone sys- tems were found to possess a capacity to tolerate partial tissue dehydration (Lilley and Ludlow, 1996). Thus, a suite of traits could confer an adaptive advantage in rainfed lowland and flood-prone condi- tions. Fukai and Cooper (1995) concluded that root
108 L.J. Wade et al. / Field Crops Research 56 (1998) 93-112
traits important for rainfed lowland conditions in- clude capacity to penetrate a hardpan, to explore greater soil depth, to effectively extract water and nutrients, and to conduct water and nutrients from depth in time of stress. They noted it is important to consider the characteristics of the target population of environments when defining traits for selection. There remains a need to demonstrate the benefits of incorporating these putative root traits for drought resistance into cultivars for rainfed lowland systems (Samson et al., 1995).
Roots are commonly in the surface layer in these systems (Sharma et al., 1994). Oxygen supply may limit root growth and function in deeper layers (Pradhan et al., 1973), and physical barriers may also prevent access of roots to deeper soil layers. The
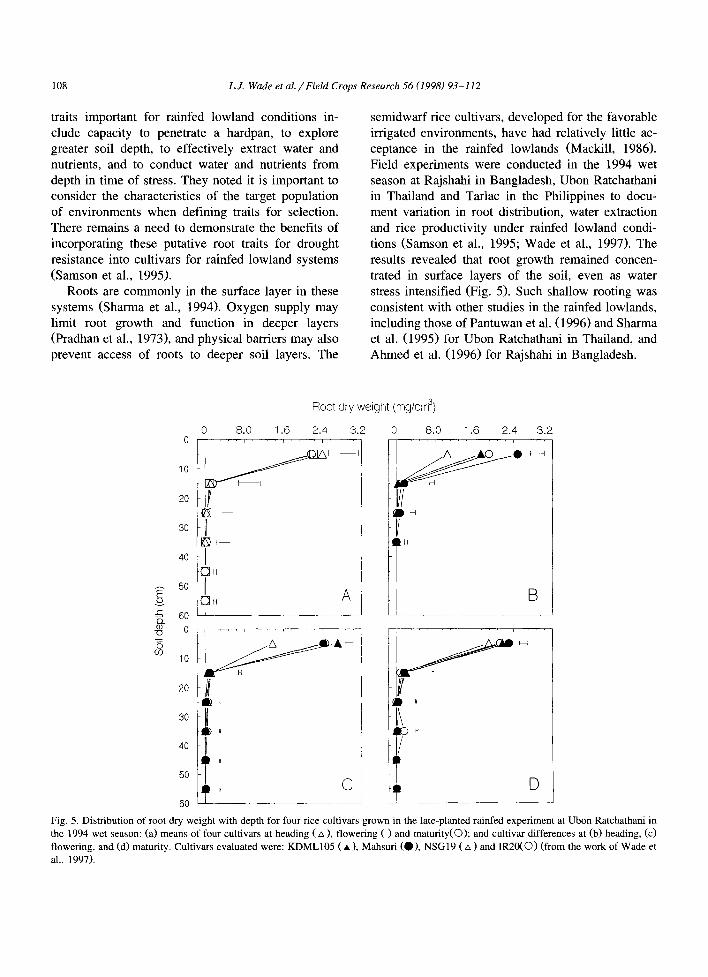
semidwarf rice cultivars, developed for the favorable irrigated environments, have had relatively little ac- ceptance in the rainfed lowlands (Mackill, 1986). Field experiments were conducted in the 1994 wet season at Rajshahi in Bangladesh, Ubon Ratchathani in Thailand and Tarlac in the Philippines to docu- ment variation in root distribution, water extraction and rice productivity under rainfed lowland condi- tions (Samson et al., 1995; Wade et al., 1997). The results revealed that root growth remained concen- trated in surface layers of the soil, even as water stress intensified (Fig. 5). Such shallow rooting was consistent with other studies in the rainfed lowlands, including those of Pantuwan et al. (1996) and Sharma et al. (1995) for Ubon Ratchathani in Thailand, and Ahmed et al. (1996) for Rajshahi in Bangladesh.
0
10
Root dry weight (mg/cm 3)
20
30
40
50 E
J~ 60 o_
0
o O9
10
20
30
4O
5O
6O
0 8.0 1.6 2.4 3.2
1 EH
EIH A
. . . . i . . . . F . . . . . .
H C
0 8.0 1.6 2 .4 3 .2
r
B t . . . .
D
Fig. 5. Distribution of root dry weight with depth for four rice cultivars grown in the late-planted rainfed experiment at Ubon Ratchathani in the 1994 wet season: (a) means of four cultivars at heading (zx), flowering ( ) and maturity(O); and cultivar differences at (b) heading, (c) flowering, and (d) maturity. Cultivars evaluated were: KDML 105 ( • ), Mahsuri (O), NSG 19 ( z~ ) and IR20(O ) (from the work of Wade et al., 1997).

L.J. Wade et al. / Field Crops Research 56 (1998) 93-112 109
Root exploration or efficiency could be limited by low soil fertility at depths greater than 10 cm on these soils (Herrera et al., 1996), an effect that may be further complicated by pH changes with wetting and drying cycles on soils of low organic matter and low cation exchange capacity (Willett, 1994). Rapid onset of drought may preclude the root system from exploring deeper soil layers on soils of low water- holding capacity. Roots of rainfed lowland rice may be poorly adapted to a period of drought late in the life cycle, if all of the roots were developed during anaerobic conditions, and thus are short and well supplied with aerenchyma. Do roots developed dur- ing subsequent aerobic conditions differ anatomi- cally, with a stronger adaptation to water uptake and water transport? Is plant response confused by root signals (Ludlow et al., 1989) from those roots in the drying surface layer, even though water may be available in deeper layers of the profile? It is not clear to what extent these factors may exist or oper- ate, either alone or in combination.
In a recent review, Wade (1996) contrasted cul- tural practices for the coarse-textured, permeable soils of northeast Thailand with those useful on the silty, compact ion-prone soils of northwest Bangladesh. At the former, research has indicated a benefit to yield of rainfed lowland rice by compact- ing the subsoil and restricting the root zone while increasing the store of available resources for the crop (Sharma et al., 1995). In contrast, use of deep cultivation or a tap-rooted legume to perforate the hardpan resulted in higher yields of rainfed lowland rice on compaction-prone soils (Ahmed et al., 1996). The results of Ahmed et al. and Sharma et al. indicate that making or breaking a hardpan may be beneficial in some situations. Determining an appro- priate strategy in each circumstance requires an un- derstanding of the dynamics of storage and capture of nutrient and water resources over different loca- tions and seasons. Defining these conditions and quantifying the associated probabilities may be as- sisted by crop simulation modeling (Singh et al., 1995a; Fukai et al., 1995).
Research is currently examining variation in the capacity among rice lines to penetrate a hardpan (Champoux et al., 1995; Yu et al., 1995; Ray et al., 1996). If the roots do penetrate, however, would resources be available for capture from deeper layers
of the soil profile below the hardpan? Conversely, when soils are permeable, would development of a hardpan be beneficial for reducing loss of resources? Again, target environments must be carefully defined when undertaking cultivar improvement in root traits for the variable rainfed lowland ecosystem.
10. Conclusions
This review demonstrates that not enough is known about the rainfed lowland rice production systems. The most critical needs are to: (a) learn how to predict variation in indigenous nutrient supply among fields at key sites, and between sites; (b) determine expected yields when nutrients and water are adequate, when nutrients are adequate but the crop is rainfed, and when neither nutrients nor water are adequate, for representative sites in the rainfed lowlands; (c) understand the relative global impor- tance of nutrients and water as limitations in the rainfed lowlands; (d) examine the potential for using strategic application of nutrients to buffer water limi- tation and stabilize yields; and (e) provide a database on soil properties, fertilizer nutrient uptake effi- ciency, climate, and crop performance, and define the relationships between these variables. This infor- mation will permit clarification of research priorities for the rainfed lowlands. Multilocation nutrient-by- water experiments provide a suitable framework for this research. Linking these experiments to the QUEFTS model and various crop models aids ex- trapolation of results to other soil types and seasonal conditions. More understanding is required of the processes of nutrient release and capture under the fluctuating moisture regimes of the rainfed lowlands. There is some evidence that manipulation of con- trolled-release fertilizer and root system development may be the key to optimizing these processes.
References
Addiscott, T.M., Whitmore, A.P., Powlson, D.S., 1991. Farming, Fertilizers and the Nitrate Problem. CAB International, Oxon, Wallingford, UK.
Ahmed, H.U., Ali, M.L., Zaman, S.K., Kabir, M.A., Miah, N.M., 1996. Varietal characteristics and soil management to reduce drought stress. In: V.P. Singh et al. (Eds.), Physiology of

110 L.J. Wade et al. / Field Crops Research 56 (1998) 93-112
Stress Tolerance in Rice, Proceedings of the International Conference on Stress Physiology of Rice, February 28-March 5, 1994. Lucknow, India, pp. 150-167.
Becket, M., Ladha, J.K., Ottow, J.C.G., 1990. Growth and N 2 fixation of two stem-nodulating legumes and their effect as green manure on lowland rice. Soil Biol. Biochem. 22, 1109- 1119.
Biswas, A.K. Jr., Nayek, B., Choudhuri, M.A., 1982. Effect of calcium on the response of a field-grown rice plant to water stress. Proc. Indian Natl. Sci. Acad. B 48, 669-705.
Buresh, R.J., De Datta, S.K., 1991. Nitrogen dynamics and man- agement in rice-legume cropping systems. Adv. Agron. 45, 1-59.
Buresh, R.J., Woodhead, T., Shepherd, K.D., Hordelis, E., Caban- gon, R.C., 1989. Nitrate accumulation and loss in a mung- bean/lowland rice cropping system. Soil Sci. Soc. Am. J. 53, 477-482.
Buresh, R.J., Singh, U., Godwin, D.C., Ritchie, J.T., De Datta, S.K., 1991. Simulating soil nitrogen transformations with CERES-Rice. Agrotechnol. Transfer 13, 7-10.
Buresh, R.J., Chua, T.T., Castillo, E.G., Liboon, S.P., Garfity, D.P., 1993. Fallow and Sesbania effects on soil nitrogen dynamics in lowland rice-based cropping systems. Agron. J. 85, 316-321.
Burford, J.R., Sahrawat, K.L., Singh, R.P., 1989. Nutrient man- agement in vertisols in the Indian semi-add tropics. Manage- ment of Vertisols for Improved Agricultural Production, Pro- ceedings of an IBSRAM Inaugural Workshop, 18-22 Febru- ary 1985. ICRISAT Center, Patanchern, A.P., India. pp. 147- 159.
Cassman, K.G., Kropff, M.J., Gaunt, J., Peng, S., 1993. Nitrogen use efficiency of rice resonsidered: what are the key con- straints?. Plant Soil 155-156, 359-362.
Castaneda, A.R., Bhuiyan, S.I., 1991. Nitrate-nitrogen concentra- tions in shallow groundwater underneath ricefields. Phil. J. Crop Sci. 16, 57-62.
Ceccarelli, S., 1996. Positive interpretation of genotype by envi- ronment interactions in relation to sustainability and biodiver- sity. In: Cooper, M., Hammer, G.L. (Eds.), Plant Adaptation and Crop Improvement. CAB International, Wallingford, UK.
Champoux, M.C., Wang, G., Sarkarung, S., Mackill, D.J., O'Toole, J.C., Huang, N., McCouch, S.R., 1995. Locating genes associated with root morphology and drought avoidance in rice via linkage to molecular markers. Theor. Appl, Gen. 90, 969-981.
Chloupek, O., 1972. The relationship between electric capacitance and some other parameters of plant roots. Biol. Plant. 14, 227-230.
Chloupek, O., 1977. Evaluation of the size of a plant's root system using its electrical capacitance. Plant Soil 48, 525-532.
Council of European Communities, 1980. Relating to the quality of water intended for human consumption. Official J. Eur. Communities L 229, 11-29.
De Datta, S.K., Buresh, R.J., 1989. Integrated nitrogen manage- ment in irrigated rice. Adv. Soil Sci. 10, 143-169.
Dobermann, A., 1994. Factors causing field variation of direct- seeded flooded rice. Geoderma 62, 125-150.
Dobermann, A., 1995. Options for classifying soils in Cambodia.
In: White, P.F., Oberthuer, T., Domingo, L.V. (Eds.), Classify- ing Cambodia Soil from an Agronomic Perspective, Proceed- ings of a Workshop May 8-12, 1995, Phnom Penh, Cambo- dia. Cambodia-IRRI-Australia Project, Phnom Penh, Cambo- dia, pp. 16-40.
Dobermann, A., Gaunt, J.L., Neue, H.U., Grant, I.F., Adviento, M.A., Pampolino, M.F., 1994a. Spatial and temporal variabil- ity of ammonium in flooded ricefields. Soil Sci. Soc. Am. J. 58, 1708-1717.
Dobermann, A., Langner, H., Mutscher, H., Skogley, E.O., Neue, H.U., Yang, J.E., Adviento, M.A., Pampolino, M.F., 1994b. Nutrient absorption kinetics of ion exchange resin capsules: a study with soils of international origin. Commun. Soil Sci. Plant Anal. 25, 1329-1353.
Dobermann, A., Cassman, K.G., Sta. Cruz, P.C., Neue, H.U., Skogley, E.O., Pampolino, M.F., Adviento, M.A., 1995a. Dy- namic soil tests for rice. In: Kirk, G.J.D., Ladha, J.K. (Eds.), Fragile Lives in Fragile Ecosystems, Proceedings of the Inter- national Rice Research Conference, February 13-17, 1995. International Rice Research Institute, Los Bafios, Philippines, pp. 343-365.
Dobermann, A., Pampolino, M.F., Neue, H.U., 1995b. Spatial and temporal variability of transplanted rice at the field scale. Agron. J. 87, 712-720.
Dobermann, A., Cassman, C.G., Peng, S., Pham, S.T., Cao, V.P., Sta. Cruz, P.C., Bajita, J.B., Adviento, M.A.A., Olk, D.C., 1996. Precision nutrient management in intensive irrigated rice systems. In: Attanandana, T., Kheoruenromne, I., Oongakul P., Vearaslip, T. (Eds.), Maximizing Sustainable Rice Yields Through Improved Soil and Environmental Management. Symposium of the Paddy Soil Fertility Working Group of the International Soil Science Society, Khon Kaen, Bangkok, 11- 17 November, 1996. DOA/SFS/DOD Thailand and ISSS, Bangkok, pp. 133-154.
Follet, R.F., Walker, D.J., 1989. Groundwater Quality Concerns About Nitrogen. Nitrogen Management and Groundwater Pro- tection. Elsevier, Amsterdam.
Fujisaka, J.S., Moody, K., Ingram, K.T., 1993. A descriptive study of farming practices for dry-seeded rainfed lowland rice in India, Indonesia and Myanmar. Agric. Ecosyst. Environ. 45, 115-128.
Fukai, S., Cooper, M., 1995. Review. Development of drought-re- sistant cultivars using physio-morphological traits in rice. Field Crops Res. 40, 67-86.
Fukai, S., Inthapan, P., 1988. Growth and yield of rice cultivars under sprinkler irrigation in southeastern Queensland: III. Water extraction and plant water relations--comparison with maize and grain sorghum. Aust. J. Exp. Agric. 28, 249-252.
Fukai, S., Rajatsasereekul, S., Boonjung, H., Skulkhu, E., 1995, Simulation modeling to quantify the effect of drought for rainfed lowland rice in northeast Thailand. In: Tuong, T.P. (Ed.), Fragile Lives in Fragile Ecosystems, Proceedings of the International Rice Research Conference, February 13-17, 1995. International Rice Research Institute, Los Bafios, Philip- pines, pp. 657-674.
Furoc, R., Morris, R.A., 1989. Apparent recovery and physio- logical efficiency of nitrogen in Sesbania incorporated before rice. Agron. J. 8 (5), 797-802.

L.J. Wade et al. / Field Crops Research 56 (1998) 93-112 111
Garrity, D.P., Oldeman, L.R., Morris, R.A., 1986. Rainfed low- land rice ecosystems: characterization and distribution. Progress in Rainfed Lowland Rice. International Rice Re- search Institute, Los Bafios, Philippines, pp. 3-23.
George, T., Ladha, J.K., Buresh, R.L, Garrity, D.P., 1992. Manag- ing native and legume-fixed nitrogen in lowland rice-based cropping systems. Plant Soil 141, 69-91.
George, T., Ladha, J.K., Buresh, R.J., Garrity, D.P., 1993. Nitrate dynamics during the aerobic soil phase in lowland rice-based cropping systems. Soil Sci. Soc. Am. J. 57, 1526-1532.
George, T., Ladha, J.K., Garrity, D.P., Buresh, R.J., 1994. Legumes as nitrate catch crops during the dry-to-wet season transition in lowland rice-based cropping systems. Agron. J. 86, 267-273.
George, T., Ladha, J.K., Garrity, D.P., Tortes, R.O., 1995. Nitro- gen dynamics of grain legume-weedy fallow-flooded rice se- quences in the tropics. Agron. J. 87, 1-6.
Godwin, D.C., Singh, U., Buresh, R.J., De Datta, S.K., 1990. Modeling of nitrogen dynamics in relation to rice growth and yield. Trans. 14th Int. Cong. Soil Sci., Kyoto, Japan, IV, pp. 320-325.
Gumtang, R.J., Pampolino, M.F., Tuong, T.P., 1998. Groundwater dynamics and quality under intensive cropping systems in Batac, Philippines. Exp. Agric (In press).
Hedley, M.J., Kirk, G.J.D., Santos, M.B., 1994. Phosphorus effi- ciency and the forms of soil phosphorus utilized by upland rice cultivars. Plant Soil 158, 53-62.
Herrera, W.A.T., Garrity, D.P., Vejpas, C., 1996. Management of Sesbania rostrata green manure crops grown and its effect prior to rainfed lowland rice on sandy soils. Field Crops Res. 49, 259-268.
Inthapan, P., Fukai, S., 1988. Growth and yield of rice cultivars under sprinkler irrigation in southeastern Queensland: II. Com- parison with maize and grain sorghum under wet and dry conditions. Aust. J. Exp. Agric. 28, 243-248.
IRRI, 1993. 1993-1995 IRRI Rice Almanac. International Rice Research Institute, Los Bafios, Philippines.
Janssen, B.H., Guiking, F.C.T., Van der Eijk, D., Smaling, E.M.A., Wolf, J., Van Reuler, H., 1990. A system for Quantitative Evaluation of the Fertility of Tropical Soils (QUEFTS). Get- derma 46, 299-318.
Kirk, G.J.D., Bajita, J.B., 1995. Root induced iron oxidation, pH changes and zinc solubilization in the rhizosphere of lowland rice. New Phytol. 131, 129-137.
Kirk, G.J.D., Saleque, M.A., 1995. Solubilization of phosphate by rice plants growing in reduced soil: prediction of the amount solubilized and the resultant increase in uptake. Eur. J. Soil Sci. 46, 247-255.
Kirk, G.J.D., Hedley, M.J., Bouldin, D.R., 1993. Phosphorus efficiency in upland rice cultivars. Reports and papers on the management of acid soils (IBSRAM/ASIALAND). Network Document No. 6, IBSRAM, Bangkok, pp. 279-296.
Kropff, M.J., Van Laar, H.H., Ten-Berge, H.F.M., 1993. ORYZAI- -a basic model for irrigated lowland rice produc- tion. International Rice Research Institute, Los Bafios, Philip- pines.
Kropff, M.J., Williams, R.J., Horie, T., Angus, J.F., Singh, U.,
Centeno, H.G., Cassman, K.G., 1995. Predicting the yield potential of rice in different environments. Aust. J. Agric. Res., pp. 657-663.
Ladha, J.K., Garrity, D.P., 1994. Green Manure Production Sys- tems for Asian Riceland: Selected Papers from the Interna- tional Rice Research Conference. International Rice Research Institute, Los Bafios, Philippines.
Ladha, J.K., Kundu, D.K., Van Coppenolle, G., Peoples, M.B., Carangal, V.R., Dent, P.J., 1996. Legume productivity and soil nitrogen dynamics in lowland rice-based cropping systems. Soil Sci. Soc. Am. J. 60, 183-192.
Lilley, J.M., Ludlow, M.M., 1996. Expression of osmotic adjust- ment and dehydration tolerance in diverse rice lines. Field Crops Res. 48, 185-197.
Ludlow, M.M., Muchow, R.C., 1990. A critical evaluation of the traits for improving crop yields in water limited environments. Adv. Agron. 43, 107-153.
Ludlow, M.M., Sommer, K.J., Flower, D.J., Ferraris, R., So, H.B., 1989. Influence of root signals resulting from soil dehydration and high soil strength on the growth of crop plants. Current Top. Plant Biochem. Physiol. 8, 81-99.
Mackill, D.J., 1986. Varietal improvement for rainfed lowland rice in South and Southeast Asia: results of a survey. Progress in Rainfed Lowland Rice. International Rice Research Insti- tute, Los Bafios, Philippines, pp. 115-144.
Mercado, A., 1976. Nitrate and chloride pollution of aquifers. A regional study with the aid of a single-cell model. Water Res. Res. 12, 731-747.
Oberthur, T., Dobermann, A., Neue, H.U., 1995a. Spatial model- ing of soil fertility--a case study in the Nueva Ecija province, Philippines. Proceedings of the International Rice Research Conference, February 13-17, 1995. International Rice Re- search Institute, Los Bafios, Philippines, pp. 689-705.
Oberthur, T., Dobermann, A., Neue, H.U., 1995b. How good is a reconnaisance soil map for agronomic purposes. Soil Use Manage. 12, 33-43.
Otto, E., Ishii, R., Kumura, A., 1989. Interaction of nitrogen supply and soil water stress on photosynthesis and transpira- tion in rice. Jpn. J. Crop Sci. 58, 424-429.
O'Toole, J.C., 1982. Adaptation of rice to drought-prone environ- ments. Drought Resistance in Crops with Emphasis on Rice. International Rice Research Institute, Los Bafios, Philippines, pp. 195-213.
Pantuwan, G., Ingram, K.T., Sharma, P.K., 1996. Rice root systems development under rainfed conditions. In: V.P. Singh et al. (Eds.), Physiology of Stress Tolerance in Rice, Proceed- ings of the International Conference on Stress Physiology of Rice, February 28-March 5, 1994, Lucknow, India. Interna- tional Rice Research Institute, Los Bafios, Philippines pp. 198-206.
Pradhan, S.K., Varade, S.B., Kar, S., 1973. Influence of soil water conditions on growth and root porosity of rice. Plant Soil 38, 501-507.
Puckridge, D.W., O'Toole, J.C., 1981. Dry matter and grain production of rice, using a line source sprinkler in drought studies. Field Crops Res. 3, 303-319.
Ray, J.D., Yu, L., McCouch, S.R., Champoux, M., Wang, G.,

112 LJ. Wade et al. / Field Crops Research 56 (1998) 93-112
Nguyen, H.T., 1996. Mapping quantitative trait loci associated with root penetration ability in rice (Oryza sativa L.). Theor. Appl. Gen. 92, 627-636.
Ritchie, J.T., Alocilja, E.C., Singh, U., Uehara, G., 1987. IB- SNAT and the CERES-RICE model. Weather and Rice, Pro- ceedings of the International Workshop on the Impact of Weather Parameters on the Growth and Yield of Rice. Interna- tional Rice Research Institute, Los Bafios, Philippines, pp. 271-283.
Saleque, M.A., Kirk, G.J.D., 1995. Root-induced solubilization of phosphate in the rhizosphere of lowland rice. New Phytol. 129, 325-336.
Samson, B.K., Wade, L.J., Sarkarung, S., Hasan, M., Amin, R., Harnpichitvitaya, D., Pantuwan, G., Rodriguez, R., Sigari, T., Calendacion, A.N., 1995. Examining genotypic variation in root traits for drought resistance. In: Tuong, T.P. (Ed.), Fragile Lives in Fragile Ecosystems, Proceedings of the International Rice Research Conference, February 13-17, 1995. Interna- tional Rice Research Institute, Los Bafios, Philippines, pp. 521-534.
Scobie, G.M., McDonald, D., Willey, R., 1993. Investment in Rice Research in the CGIAR: A Global Perspective. TAC Secretariat, FAO, Rome.
Sharma, P.K., Pantuwan, G., Ingram, K.T., De Datta, S.K., 1994. Rainfed lowland rice roots: soil and hydrologic effects. In: Kirk, G.J.D. (Ed.), Rice Roots: Nutrient and Water Use. Selected Papers from the International Rice Research Confer- ence. International Rice Research Institute, Los Bafios, Philip- pines, pp. 55-66.
Sharma, P.K., Ingram, K.T., Harnpichitvitaya, D., 1995. Subsoil compaction to improve water use efficiency and yields of rainfed lowland rice in coarse-textured soils. Soil Tillage Res. 36, 33-44.
Singh, U., Ritchie, J.T., Godwin, D.C., 1993. A user's guide to CERES-Rice V2.10. Simulation Manual IFDC-SM-4. Int. Felt. Dev. Center, Muscle Shoals, AL, 132 pp.
Singh, U., Cassman, K.G., Ladha, J.K., Bronson, K.F., 1995a. Innovative nitrogen management strategies for lowland rice systems. In: Kirk, G.J.D., Ladba, J.K. (Eds.), Proceedings of the International Rice Research Conference, February 13-17, 1995. International Rice Research Institute, Los Bafios, Philip- pines, pp. 507-519.
Singh, A.K., Tuong, T.P., Wopereis, M.C.S., Boling, A.A., Kropff, M.J., 1995b. Quantifying lowland rice responses to soil-water deficit. In: Kirk, G.J.D., Ladha, J.K. (Eds.), Proceedings of the International Rice Research Conference, February 13-17, 1995. International Rice Research Institute, P.O. Box 933, Manila, Philippines.
Tanguilig, U.C., De Datta, S.K., 1988. Potassium effects on root growth and yield of upland rice (Oryza sativa L.) under drought and compacted soil conditions. Phil. J. Crop Sci. 13, $33.
Tripathi, B.P., Ladha, J.K., Timsina, J., Pascua, S.R., 1997.
Nitrogen dynamics and balance in intensified rainfed lowland rice-based cropping systems. Soil Sci. Soc. Am. J. 61, 812- 821.
Viets, F.G. Jr., Hageman, R.H., 1971. Factors affecting the accu- mulation of nitrate in soil, water, and plants. Agriculture Handbook No. 413, ARS-USDA, Washington, DC.
Wade, L.J., 1996. Soil compaction--make it or break it for rainfed lowland ecosystem. In: So, H.B., Kirchhof, G. (Eds.), Management of Clay Soils in Rainfed Lowland Ecosystem. ACIAR Proceedings Number 70. ACIAR, Canberra, Australia, pp. 243-247.
Wade, L.J., Sarkarung, S., McLaren, C.G., Guhey., A., Quader, B., Boonvite, C., Amarante, S.T., Sarawgi, A.K., Haque, A., Harnpichitvitaya, D., Pamplona, A., Bhamri, M., 1995. Geno- type by environment interaction and selection methods for identifying improved rainfed lowland rice genotypes. In: Ben- nett, J., Courtois, B., Khush, G.S., Senadhira, D. (Eds.), Fragile Lives in Fragile Ecosystems, Proceedings of the Inter- national Rice Research Conference, February 13-15, 1995. International Rice Research Institute, Los Bafios, Philippines, pp. 885-900.
Wade, L.J., McLaren, C.G., Samson, B.K., Regmi, K.R., Sarkarung, S., 1996. The importance of site characterization for understanding genotype by environment interactions. In: Cooper, M., Hammer, G.L. (Eds.), Plant Adaptation and Crop Improvement, CAB International, Wallingford, UK, pp. 549- 562.
Wade, L.J., Moya, T.B., Pantuwan, G., Regmi, K.R., Samson, B.K., 1997. Research on root systems of rice at IRRI. In: Morita, S., Yamauchi, A. (Eds.), Ideotype of Root System of Rice. World Scientific Publications, Singapore.
Widawsky, D.A., O'Toole, J.C., 1990. Prioritizing the rice bio- technology research agenda for Eastern India. The Rockefeller Foundation, New York.
Willett, I.R., 1994. Role of organic matter in controlling chemical properties and fertility of sandy soils used for lowland rice in northeast Thailand. Management of Soil Organic Matter for Sustainable Agriculture in Asia. Proceedings of the ACIAR Conference, Ubon Ratchathani, Thailand, August 24-26, 1994.
Wopereis, M.C.S., Kropff, M.J., Maligaya, A.R., Tuong, T.P., 1995. Drought stress responses of two lowland rice cultivars to soil-water status. Field Crops Res. 46, 21-39.
Yoshida, S., Hasegawa, S., 1982. The rice root system: its devel- opment and function. Drought Resistance in Crops with Em- phasis on Rice. International Rice Research Institute, Los Bafios, Philippines, pp. 97-114.
Yu, L.U., Ray, J.D., O'Toole, J.C., Nguyen, H.T., 1995. Use of wax-petrolatum layers for screening rice root penetration. Crop Sci. 35, 684-687.
Zaman, A., Roy, G.B., Mallick, S., Maiti, A., 1990. Response of direct seeded rice to irrigation and nitrogen in drought prone laterite tract of West Bengal. Environ. Ecol. 8, 311-314.