on vegetation recovery, goats and giant tortoises on pinta island, galápagos, ecuador
TRANSCRIPT

Biodiversity and Conservation 2, 138-151 (1993)
On vegetation recovery, goats and giant tortoises on Pinta Island, Gakipagos, Ecuador': OLE H A M A N N Botanic Garden, University of Copenhagen, Oster Farimagsgade 2 B, DK-1353 Copenhagen K, Denmark
Received 10 September 1992; accepted 7 October 1992
Data on vegetational changes on Pinta Island, Gal~ipagos, collected during the period 1970-1988, show that feral goats have had a destructive impact on the vegetation. Stands of Scalesia baurii ssp. hopkinsii (Asteraceae) and Opuntia galapageia var. galapageia (Cactaceae) were close to a complete collapse, while stands of Bursera graveolens (Burseraceae) apparently were able to persist for a longer time. After the elimination of large numbers of goats, Scalesia, Opuntia and Bursera were recovering rapidly, but the patterns of regeneration differed in accordance with differences in longevity, growth and seedling establishment.
It is suggested that the absence of herbivores (the Pinta subspecies of giant tortoise is practically extinct, and feral goats were finally eradicated in 1990) in the long term may lead to decreased diversity in the vegetation of Pinta. As a management experiment aimed at conserving the biological diversity of Pinta, it is proposed to reintroduce giant tortoises to the island, preceded and accompanied by a careful long-term monitoring.of vegetational changes.
Keywords: Gal~ipagos; Pinta island; regeneration of woody plants: feral goats; giant tortoises: ecological diversity
Introduction
The flora of the uninhabited island of Pinta is extraordinarily rich compared to other small islands in the Gal~pagos archipelago. In the 1950s goats were introduced to Pinta, where they multiplied extremely rapidly and soon had a very negative impact on the vegetation. Closed forest and scrub were opened, soil erosion started and natural regeneration of several plant species was prevented. In the early 1970s the Gahipagos National Park Service (GNPS) initiated a goat eradication campaign; when the campaign was concluded successfully in 1990 and the last goats eliminated, an estimated total of 40 000 goats had been killed since the start of the campaign.
During the period 1972 to 1981 a notable regeneration of the vegetation on Pinta was recorded (Hamann, 1984), concurrent with the eradication of large numbers of feral goats. In 1988, when only a handful r emainedn the island, new data were compiled on the regeneration of the vegetation. The present paper deals in particular with the recovery of three woody species, viz. the endemics Opuntia galapageia var. galapageid (Cactaceae) and Scalesia baurii ssp. hopkinsii (Asteraceae) and the indigenous Bursera
*Contribution No. 486 of the Charles Darwin Foundation for the Galapagos Isles. *Plant names are in accordance with Wiggins and Porter (1971) and Eliasson (1974), unless otherwise indicated.
0960-3115 © 1993 Chapman & Hall

Vegetation recovery on Pinta Island, Galfpagos 139
graveolens (Burseraceae). The structure of stands according to size (and age) and the patterns of regeneration of the three species are presented and discussed.
On Pinta there is still a recognizable pattern of mosaic vegetation, in which forested areas are interspersed with open, meadow-like areas. Such a mosaic pattern is in other places in the archipelago associated with tortoise grazing and is presumably a result of a very long process of coevolution and adaptation. In former times, the Pinta subspecies of giant tortoise, Geochelone elephantopus abmgdoni, was the only indigenous herbivorous species on the island. The population of G. e. abingdonii collapsed already during the last Century, and today only one male individual, known as 'Lonesome George', is surviving. The moasic pattern of vegetation on Pinta might reflect the former presence of tortoises, even though it is more than 100 years ago that tortoises occurred there in any great numbers, but the pattern may also have been greatly accentuated by the impact of goats.
Since both indigenous and introduced herbivores now have been removed from Pinta, the rapid regeneration among the woody plants, which is occurring even in open meadow-like areas, may eventually lead to a reduction in the overall ecological diversity. It is proposed that it is both desirable and possible to reintroduce tortoises to Pinta in a long-term management experiment aiming at maintaining the biological diversity on the island. Such a reintroduction should be preceded and over the years accompanied by careful monitoring of the plant life on the island.
Profile of Pinta
Pinta is one of the northern islands in the Gal~ipagos archipelago, rarely visited by scientists and not at all by tourists. It covers about 59 km 2 and reaches an altitude of 65(I m. Due to the high elevation of the central part, Pinta displays similar features as other high altitude Gal~ipagos islands, which, however, generally are much larger. There is a great variety of habitats, several vegetation zones can be distinguished corresponding to the local variation in climatic conditions, and the diversity of plants and animals is exceptionally high compared to other islands in the archipelago (Hamann, 1981). This is also related to the long history of biological isolation and the former presence of a population of giant tortoises.
With only 59 km 2 Pinta covers about 0.75% of the total land surface of the Gal~ipagos archipelago, but it has 32% of the native land plants. No less than 180 taxa of higher plants are known from Pinta, of which 59 are endemic to Gal~ipagos, 117 are indigenous, and only four can be considered as pantropical weeds representing early accidental introductions. Of the 176 native taxa 19 are rare on an archipelago scale, while six are indeterminate or unknown with respect to conservation status. Two plant taxa are Pinta endemics, viz. Scalesia baurii ssp. hopkinsii (the Pinta Sunflower-tree) and Alternanthera filifolia spp. pintensis (Amaranthaceae), while Acalypha abingdonii O. Seberg (Euphorbiaceae) is only known from Pinta and Marchena islands; two indigenous species, Sida veronicaefolia Lam. (Malvaceae) and Eragrostis montufari HBK (Poaceae), are in the Gal~ipagos only known from Pinta ~.
~The occurrence of Eragrostis montufari HBK on Pinta was first noted by Dr Ole Seberg, University of Copenhagen, during an expedition in 1981. Later it turned out that it had been collected by earlier visitors, but it had not been correctly identified before (O. Seberg, personal communication). In the Galtipagos it is only known from Pinta, where it is common or in places even dominant.

140 Hamann
Numerous indigenous and endemic birds live on Pinta, as for example 9 out of 13 species of Darwin's finches. The three indigenous taxa of reptiles on the island are all Pinta endemics, viz. the giant tortoise, the marine iguana (Amblyrhynchus cristatus sielmanni) and the lava lizard (Tropidurus pacificus). There are no native land mammals on Pinta, but feral goats were present from the 1950s to 1990.
Feral goats on Pinta
It is generally believed that goats first were introduced to Pinta in 1959, but it may have happened 5 years previously (Hoeck, 1984). Originally 1 male and 2 female goats were set ashore, presumably by a local fisherman, but in a few years the population grew dramatically. In the early 1970s the Gal~ipagos National Park Service (GNPS) estimated that about 10 000 goats were present on Pinta. The goats had a very destructive impact on the ecosystems: closed scrub and forest were opened, soil erosion started and several plant species, including Scalesia baurii ssp. hopkinsii, were severely reduced in numbers. The goats particularly prevented natural regeneration and turn-over in the plant cover, so it could be foreseen that unless the goats were eliminated or at least controlled, the rich and varied ecosystems of Pinta would suffer perhaps irreversible changes within a very short time (Weber, 1971; Hamann, 1975; Adsersen, 1976). The GNPS then initiated a major goat eradication campaign, which continued until 1990, when finally all goats had been eliminated (Evans, 1990). In total, the park wardens have shot about 40000 goats during the 20 year period. The recovery of vegetation taking place as the goats were reduced in numbers has been rapid and spectacular (Hamann, 1975, 1979a, 1984).
Tortoises on Pinta
During the 17th, 18th and 19th Centuries buccaneers and whalers took a heavy toll on the tortoises of Gal~ipagos. Two of the originally existing 15 subspecies were hunted and collected to extinction. A conservative estimate gives a total figure of more than 100000 tortoises taken away from Gal~ipagos, particularly by whalers who used the animals as a supply of fresh meat on the long hauls in the Pacific (de Vries, 1984).
The Pinta giant tortoise is today practically extinct. Only the famous 'Lonesome George' is known to be alive (Fig. 1). He is kept at the Charles Darwin Research Station (CDRS) on Santa Cruz island, where he has been since he was caught on Pinta in 1972. 'Lonesome George' is the last of his kind, which presumably numbered hundreds or thousands of individuals in former times (Reynolds and Marlow, 1983). The tortoise population on Pinta collapsed during the 19th Century. Records from American whalers document that 455 tortoises were removed from Pinta just during the period 1831-68, to which figure should be added an unknown number taken by whalers from other nations (Reynolds and Marlow, 1983). Since then, very few tortoises have been recorded or taken on Pinta, and most have been males. This is related to the fact that females are smaller and easier to carry, and that they during the nesting season are found closer to the coast, wherefore the whalers predominantly must have removed females. The last known living female tortoise was taken on Pinta in 1901 (Pritchard, 1984)1 In 1906, three male specimens were collected by the California Academy of Sciences expedition (van Denburgh. 1914). Since then, no tortoises were seen on Pinta until 'Lonesome George' was discovered in 1971 and caught in 1972.

Vegetation recovery on Pinta Island, Galfpagos 141
Fig. 1. 'Lonesome George' on Pinta Island in March 1972, on the day he was caught by National Park wardens. The next day he was sailed to Santa Cruz Island, where he since has been kept at the CDRS. 'Lonesome George' is the last known living individual of the Pinta subspecies of giant tortoises, Geochelone elephantopus abingdonii.
During subsequent expeditions to Pinta in the 1970s and 1980s researchers and national park wardens have been looking for tortoises or signs of them. In 1981 herpetologist Linda Cayot and I found a dried tortoise dropping, which was intact but apparently quite old. It was assumed that if it had been deposited by 'Lonesome George' before he was taken away from Pinta in 1972, it would most likely have been broken down by 10 years of wind, rain and sun. So visitors are still looking for a tortoise on Pinta, but all searches since then have been in vain. The likelihood that there may be one or a few tortoises left on the island must be almost nill.
Vegetation studies on Pinta
Study areas and methods
In 1966 the CDRS initiated a study of plant successions in the Gal~pagos by establishing permanent sample plots (quadrats of 10 x 10 m) in selected areas and starting periodic analyses of the vegetation in these quadrats. In 1971-72 1 was staff scientist at the CDRS, at which time I enlarged the study by establishing a number of new permanent quadrats in selected areas. During several subsequent visits and expeditions I have continued the studies on vegetational changes in the GaMpagos. This has yielded information on the impact of introduced grazers, on regeneration trends in various vegetation types, and on

142 Hamann
the population structure and survival strategies of some of the dominant plant species (e.g. Hamann, 1975, 1979a,b, 1981, 1984, 1985). An overview of plant vegetation studies in the Gal/tpagos by means of permanent quadrats during 1966 to 1987, carried out by various researchers, was given by Adsersen (1990).
In 1988 an expedition was made to Pinta in order to monitor the vegetational changes taking place after goats had been almost totally eradicated. In 1988 no more goats were present in the vegetated parts on Pinta, where the permanent quadrats are located. The few goats that remained on the island at that time were living close to the barren lava fields in the north-western part of the island (Whelan and Hamann, 1989).
Four of the five existing permanent quadrats on Pinta were found and analysed; species composition, density and coverage were recorded, and the individuals of woody plants were measured, new individuals were tagged and dead or absent ones recorded, so that growth, recruitment and mortality of woody species could be calculated. Notes on phenology were taken.
In the present paper, data on Scalesia baurii ssp. hopkinsii generated from permanent quadrat No. 2 is presented. This quadrat was established in 1970 by Tj. de Vries of the CDRS in typical Arid zone vegetation at 125 m (above sea level). The vegetation can be classified as dry season deciduous steppe forest (Hamann, 1981), in which Bursera graveolens and Opuntia galapageia var. galapageia formed an open-canopied tree layer; an equally open shrub layer was composed of Castela galapageia (Simaroubaceae), Scalesia baurii ssp. hopkinsii, Lantana peduncularis (Verbenaceae) and Croton scouleri
Fig. 2. Study area at 125 m above sea level. Open dry season deciduous steppe forest with Bursera graveolens and Opuntia galapageia var. galapageia as the most conspicuous species. Among the shrubby species~ Castela galapageia was the most abundant.

Vegetation recovery on Pinta Island, Gal@agos 143
Fig. 3. Study area at 45 m above sea level. Very open dry season deciduous steppe forest with scattered Bursera graveolens and many young Opuntia galapageia var. galapageia. On the rocky ridge in the background several large Scalesia baurii ssp. hopkinsii.
(Euphorbiaceae). The rather open herb layer was composed of many species. In this quadrat, a notable regeneration of woody plants had already been recorded by 1981 (Hamann, 1984).
In addition, two study areas were selected for measuring populations of Scalesia baurii ssp. hopkinsii, Opuntia galapageia var. galapageia and Bursera graveolens. One study area was the dry season deciduous steppe forest adjacent to permanent quadrat No. 2 at 125 m a.s.l. (Fig. 2). The other study area was situated at 45 m a.s.l., and was characterized by a more open dry season deciduous steppe forest vegetation with scattered Bursera and Opuntia, fewer shrubs, very little ground cover and large stretches of lava rock without vegetation (Fig. 3). In each study area, size was measured on 100 individuals selected at random, and notes were taken on the phenology of the individuals.
Results
Scalesia baurii spp. hopkinsii
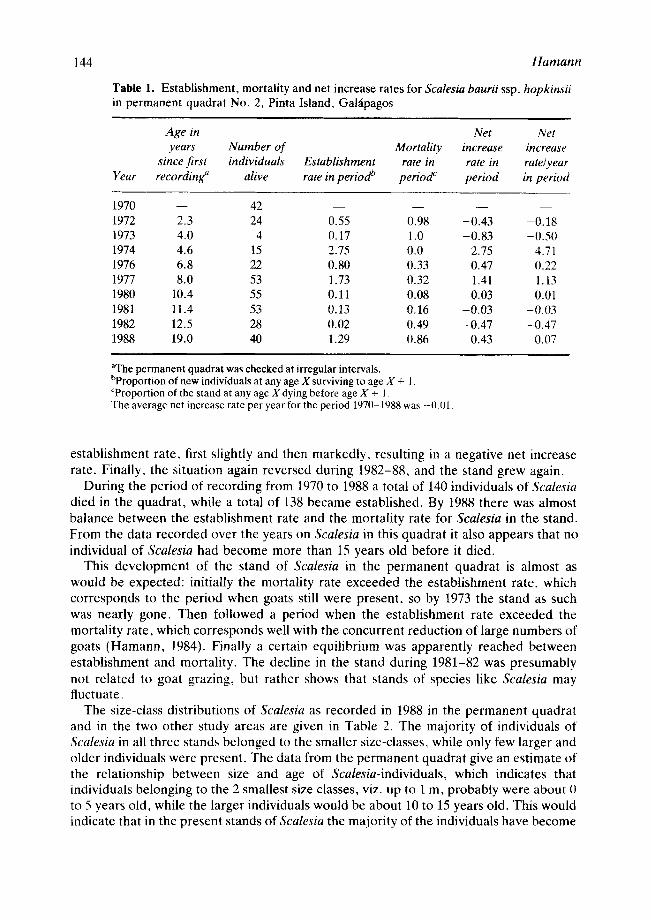
Based on the data collected from permanent quadrat No. 2 since 1970, the establish- ment, mortality and net increase rates of Scalesia were calculated (Table 1).
The Scalesia stand declined from 1970 to an all time low in 1973. From 1973 to 1980 the stand built up again, but during the next two years the mortality rate exceeded the
144 Hamann
Table 1. Establishment, mortality and net increase rates for Scalesia baurii ssp. hopkinsii in permanent quadrat No. 2, Pinta Island, Gal~ipagos
Age in Net Net years Number of Mortality increase increase
since first individuals Establishment rate in rate in rate/year Year recording" alive rate in period b perioa ~ period in period
1970 - - 42 . . . . 1972 2.3 24 0.55 0.98 -0.43 -0.18 1973 4.0 4 O. 17 1.0 -0.83 -0.50 1974 4.6 15 2.75 0.0 2.75 4.71 1976 6.8 22 O. 80 0.33 O. 47 0.22 1977 8,0 53 1.73 0.32 1.41 1.13 1980 10.4 55 O. 11 0.08 0,03 0.01 1981 11.4 53 0.13 0.16 -0.03 -0.03 1982 12.5 28 0.02 0.49 -0.47 -0.47 1988 19.0 40 1.29 O. 86 O. 43 0.07
aThe permanent quadrat was checked at irregular intervals. bProportion of new individuals at any age X surviving to age X + 1. CProportion of the stand at any age X dying before age X + 1. The average net increase rate per year for the period 197[)-1988 was --0.1H.
establishment rate, first slightly and then markedly, resulting in a negative net increase rate. Finally, the situation again reversed during 1982-88, and the stand grew again.
During the period of recording from 1970 to 1988 a total of 140 individuals of Scalesia died in the quadrat , while a total of 138 became established. By 1988 there was almost balance between the establishment rate and the mortality rate for Scalesia in the stand. From the data recorded over the years on Scalesia in this quadrat it also appears that no individual of Scalesia had become more than 15 years old before it died.
This development of the stand of Scalesia in the permanent quadrat is almost as would be expected: initially the mortality rate exceeded the establishment rate, which corresponds to the period when goats still were present, so by 1973 the stand as such was nearly gone. Then followed a period when the establishment rate exceeded the mortality rate, which corresponds well with the concurrent reduction of large numbers of goats (Hamann , 1984). Finally a certain equilibrium was apparently reached between establishment and mortality. The decline in the stand during 1981-82 was presumably not related to goat grazing, but rather shows that stands of species like Scalesia may fluctuate.
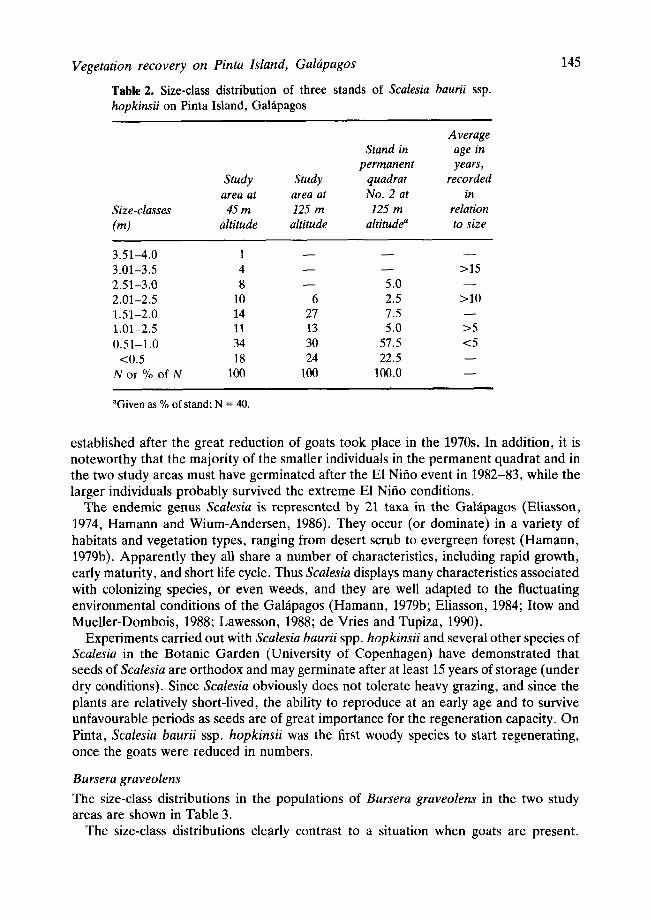
The size-class distributions of Scalesia as recorded in 1988 in the permanent quadrat and in the two other study areas are given in Table 2. The majority of individuals of Scalesia in all three stands belonged to the smaller size-classes, while only few larger and older individuals were present. The data from the permanent quadrat give an estimate of the relationship between size and age of Scalesia-individuals, which indicates that individuals belonging to the 2 smallest size classes, viz. up to 1 m, probably were about 0 to 5 years old, while the larger individuals would be about 10 to 15 years old. This would indicate that in the present stands of Scalesia the majority of the individuals have become
Vegetation recovery on Pinta Island, Galdpagos
T a b l e 2. Size-class distribution of three stands of Scalesia baurii ssp. hopkinsii on Pinta Island, Gahipagos
Average Stand in age in
permanent years, Study Study quadrat recorded area at area at No. 2 at in
Size-classes 45 m 125 m 125 m relation (m) altitude altitude altitude a to size
145
3.51-4.0 1 - - - - - -
3.01-3.5 4 - - - - >15 2.51-3.0 8 - - 5.0 - - 2.01-2.5 10 6 2.5 >10 1.51-2.0 14 27 7.5 - - 1.01-2.5 11 13 5.0 >5 0.51-1.0 34 30 57.5 <5 <0.5 18 24 22.5 - -
N or % of N 100 100 100.0 - -
aGiven as % of stand; N = 40.
established after the great reduction of goats took place in the 1970s. In addition, it is noteworthy that the majority of the smaller individuals in the permanent quadrat and in the two study areas must have germinated after the El Nifio event in 1982-83, while the larger individuals probably survived the extreme El Nifio conditions.
The endemic genus Scalesia is represented by 21 taxa in the Gal~ipagos (Eliasson, 1974, Hamann and Wium-Andersen, 1986). They occur (or dominate) in a variety of habitats and vegetation types, ranging from desert scrub to evergreen forest (Hamann, 1979b). Apparently they all share a number of characteristics, including rapid growth, early maturity, and short life cycle. Thus Scalesia displays many characteristics associated with colonizing species, or even weeds, and they are well adapted to the fluctuating environmental conditions of the Gal~pagos (Hamann, 1979b; Eliasson, 1984; Itow and Mueller-Dombois, 1988; Lawesson, 1988; de Vries and Tupiza, 1990).
Experiments carried out with Scalesia baurii spp. hopkinsii and several other species of Scalesia in the Botanic Garden (University of Copenhagen) have demonstrated that seeds of Scalesia are orthodox and may germinate after at least 15 years of storage (under dry conditions). Since Scalesia obviously does not tolerate heavy grazing, and since the plants are relatively short-lived, the ability to reproduce at an early age and to survive unfavourable periods as seeds are of great importance for the regeneration capacity. On Pinta, Scalesia baurii ssp. hopkinsii was the first woody species to start regenerating, once the goats were reduced in numbers.
Bursera graveolens
The size-class distributions in the populations of Bursera graveolens in the two study areas are shown in Table 3.
The size-class distributions clearly contrast to a situation when goats are present.

146
Table 3. Size-class distribution of two stands of Bursera graveolens on Pinta Island, Galfipagos
Size-classes Study area at Study area at (m) 45 m altitude 125 rn altitude
8.01-9.0 -- l 7.01-8.0 -- 8 6.01-7.0 2 13 5.01-6.0 7 29 4.01-5.0 23 21 3.01-4.0 8 8 2.01-3.0 -- - - 1.01-2.0 14 1 <1.0 46 19
N 100 100
H a m a n n
Grant (1981) illustrated the size-class distribution in three Bursera-populations, one growing in an area without goats and two in areas infested with goats. Those two goat-damaged populations (on Santa Cruz and Isabela islands) comprised no small Bursera-individuals and the populations had an unstable top-heavy age-structure (Grant, 1981).
In the Bursera-populations in the two study areas on Pinta the distribution in the size- classes from about 3 m to the maximum of 9 m shows that large and older individuals had survived until they started dying off naturally; this part of the pattern corresponds to the goat-damaged populations analyzed by Grant (1981). However, evidently a strong regeneration was under way on Pinta, with a large proportion of the individuals being less than 1 m tall.
On Pinta, some size-classes were completely lacking or poorly represented (Table 3). which corresponds to a gap in recruitment. This may partly he ascribed directly to goat grazing, and partly to the altered conditions for germination and establishment, which resulted from goat grazing. Observations on the island of Santa Fe indicate that germination and establishment of Bursera seedlings are facilitated by a moderate vegetation cover (Hamann, 1985). So the altered ecological conditions in the lower parts of Pinta, where the vegetation was opened and the ground cover to a large extent removed by goat grazing, may for a time have prevented the re-establishment of Bursera, even after the removal of goats. On the other hand, the E1 Nifio conditions of 1982-83 may have accelerated the regeneration: the El Nifio brought an uninterrupted 9 months of heavy rains and high temperatures to the Gahipagos, which resulted in a spectacular appearance of annual herbs, grasses and vines, particularly in the usually arid regions (Hamann, 1985). It is quite possible that all the Bursera individuals recorded in the two smaller size classes were post-E1 Nifio in origin, viz. individuals the germination and establishment of which were facilitated by the extreme El Nifio conditions.
In his Bursera study Grant estimated the oldest tree to be 68 years, and he found it unlikely that there would be any Bursera-trees still standing " ' . . . which Charles Darwin

Vegetation recovery on Pinta Island, Galfipagos 147
gazed upon 146 years ago" (Grant, 1981). Even so, it has not been possible to accurately determine the age of Bursera trees, but the data recorded from several permanent quadrats and from observing seedlings tagged by Tj. de Vries on Santa Fe island show that Bursera-trees are slow-growing and live much longer than, for example, Scalesia trees. As the typical arid zone dominant in the Galfipagos Bursera is well adapted to survive long periods of drought. One element in this would be seed dormancy, which McMullen (1986) demonstrated for about 67% of the species in the basically arid zone of Galfipagos (but Bursera was not included in McMullen's study). In the Botanical Garden, University of Copenhagen, seeds of Bursera have been germinated after several years of dry storage, and the regeneration pattern of Bursera recorded in the permanent quadrats in the islands implies that the seeds are dormant.
Opuntia galapageia var. galapageia
The size-class distribution of Opuntia galapageia var. galapageia (Table 4) parallels more or less that of Bursera. A majority of the populations fell within the lower size-classes, viz. the majority of recorded individuals were less than about 2.5 m tall. There was no gap in the representation of individuals in the size classes. However, in both study areas the largest size classes contained those Opuntia-individuals of a size and age, where the spine-less, bark-covered trunk and many branches with pads (a 'crown') had been developed, while all the individuals in the lower size-classes had spiny trunks, only few branches and no 'crowns'.
The recovery of Opuntia megasperma var. orientalis on Espafiola island was recently investigated by Grant and Grant (1989). After the elimination of feral goats on Espafiola in 1978 (Hoeck, 1984), an investigation in 1979 suggested that regeneration had started. However, the recovery of Opuntia recorded by 1988 had been remarkably poor. Grant and Grant (1989) concluded that either the two recent E1 Nifio events, especially the first
Table 4. Size-class distribution of two stands of Opuntia galapageia var. galapageia on Pinta Island, Galfipagos
Size-classes Study area at Study area at (rn) 45 m altitude 125 m altitude
3.51-4.0 - - 2 a 3.01-3.5 2 a 2 a 2.51-3.0 3 a 5 2.01-2.5 8 15 1.51-2.0 19 29 1.01-2.5 23 16 0.51-1.0 21 20 <0.5 24 11
N 100 100
aMature individuals, which had a well developed, spineless trunk, many branches and a 'crown'. All smaller individuals had spiny stems and no 'crown'.

148 H amann
in 1982-83, had delayed the expected regeneration, or other factors not yet identified had been responsible for the weak recruitment and the low adult density of Opuntia on Espanola.
In contrast, the regeneration on Pinta by Opuntia galapageia var. galapageia was well under way by 1988, as showed by the strong recruitment in the two study areas, but as on Espanola, the Pinta populations of Opuntia had a very low density of adult individuals. The distribution on size-classes indicates that almost all old and mature individuals of Opuntia were gone in 1988, but also that a tremendous regeneration was under way. That so few old individuals were present may first of all be related to the fact that goats are able to destroy even mature trees of Opuntia by eating through the spineless bark of the trunk. So mature Opuntia individuals are more vulnerable to goat 'grazing' than older individuals of, for example, Bursera. Secondly, the El Nifio conditions in 1982-83 may well have killed a number of old Opuntia plants, as it was observed on other islands (Hamann, 1985; Grant and Grant, 1989). Thus, the populations of Opuntia in the two localities investigated on Pinta were probably closer to a complete breakdown than the populations of Bursera and Scalesia were, as a result of first the impact of goats and later the El Nifio. However, the large representation of Opuntia individuals in all the smaller size classes indicates that regeneration now had been under way for a number of years.
Discussion
The largest individuals of Scalesia were encountered in the study area at 45 m elevation, while the largest individuals of Bursera and Opuntia were recorded at 125 m elevation. With respect to Scalesia this difference appears to be a result of a few individuals surviving in a sheltered place below a rocky ridge at the lower elevation, while the preponderance of large Bursera and Opuntia at the higher elevation reflects the general trend in vegetational changes along the altitudinal gradient. On the south slope of Pinta, the density, structural complexity and development of individuals of woody plants in the vegetation increase with altitude at the same time as the dry season deciduous steppe forest gradually changes into a taller, dry season deciduous forest, which then at higher elevation is replaced by evergreen steppe forest and forest (Hamann, 1981).
The data on Scalesia, Bursera and Opuntia on Pinta show that goats had a serious, .negative impact on the populations. The stands of Scalesia and Opuntia had probably been close to a complete collapse, while the stands of Bursera apparently were able to survive longer without new recruitment. After the removal of goats, all three species were able to recover. However, the regeneration patterns were different, which reflects the differences between the species in longevity, growth form and habit, and in seed germination and seedling establishment patterns.
Now, with both indigenous and introduced herbivores gone from the island, how will the vegetation react to the complete absence of grazers? Evidently, a very rapid and notable regeneration of vegetation is under way in all vegetation zones, including the high altitude vegetation close to the summit (Hamann, 1984; Whelan and Hamann, 1989). Even though the tortoise population on Pinta collapsed more than a hundred years ago, the former presence of tortoises apparently influenced the vegetation in the same way as on Santa Cruz for example; there 'tortoise country'-vegetation typically consists of a mosaic of open areas dominated by grasses and herbs and more closed areas dominated by woody plants. Such mosaic vegetation is still occurring on Pinta, but it may

Vegetation recovery on Pinta Island, Galdpagos 149
also, at least partly, be a result of or have been accentuated by the presence of large numbers of goats from the 1950s to 1990. In this mosaic vegetation on Pinta, which mostly is found at intermediate altitudes, the herb-grass meadows occur interspersed in evergreen forest or steppe forest; some of the characteristic meadow species are Justicia galapagana (Acanthaceae), Acalypha abingdonii (Euphorbiaceae) and Salvia occidentalis (Lamiaceae) (Hamann, 1981). By 1988, the herb-grass meadows were being colonized by woody plants such as Darwiniothamnus tenuifolius (Asteraceae), Lippia rosmarinifolia (Verbenaceae) and Zanthoxylum fagara (Rutaceae). Of the species in the herb-grass meadows, both Justicia and Salvia are common in other vegetation types as well, but the endemic Acalypha abingdonii appears to have a restricted occurrence in this vegetation type. It may not be able to survive in very close vegetation (Seberg, 1984). So absence of grazers will perhaps in the long term lead to both the disappearance of herb- grass meadows as a vegetation type on Pinta and to the disappearance from Pinta of an endemic species, which is only known from this island and Marchena.
Aiming at conserving the ecological diversity of Pinta, it may therefore be argued that tortoises should be reintroduced to the island. Man has disturbed the evolutionary and ecological processes, particularly with respect to coevolution and balance between native plants and animals on the island. But this may be rectified, and Pinta presents a case for a large-scale and long-term experiment in conservation management.
A key question is whether it will be possible to save the Pinta subspecies of tortoise, or at least preserve the genes of 'Lonesome George'. The various options were discussed by Reynolds and Marlow (1983) and Pritchard (1984), but since then the Charles Darwin Research Station (CDRS) and the Gal~ipagos National Park Service (GNPS) have initiated an attempt of mating 'Lonesome George' with the morphologically closest relative, Geochelone elephantopus becki, from Volcan Wolf, Isabela (CDRS, personal communication). Given the nature of tortoises, their long lives, and the ease by which they generally are located in the field (or removed if it is so wished), reintroducing tortoises to Pinta would be rather easy. The crucial factor would be to introduce a known set of tortoises to Pinta. Depending on the outcome of the current attempts to preserve the genes of 'Lonesome George', such a set could consist either of offspring resulting from successfully mating 'Lonesome George' with G. e. becki, or of pure G. e. becki. If the first option becomes possible, this would also open the attractive possibility for setting 'Lonesome George' back on his home island one day, to live out his days among a stock of his own offspring. The safeguard for such a management experiment would be that if, after 10, 25 or 100 years, it should be considered non-desirable to have 'foreign' tortoises on Pinta, the animals could fairly easily be removed from the island again.
The experiment should be accompanied by a careful monitoring of the vegetation on the island, beginning with a detailed study before the reintroduction takes place, and followed by regular monitoring. This will then generate data directly related to the influence of grazing tortoises on plant diversity, ecological diversity, and on the successional processes on Pinta. The already existing permanent quadrats on Pinta may serve as a nucleus for monitoring the changes over the years.
Acknowledgements
It is a great pleasure to acknowledge the support received during many years from the Ecuadorian authorities, the TAME airline, the Gal~ipagos National Park Service and the

150 Hamann
Charles Darwin Research Station. I also wish to thank Ra61 Salazar, Fionnuala Walsh, Pat Whelan and Andy Wilson for inspiring company and assistance during the 1988 expedition to Pinta. The financial support to the 1988 expedition from the Danish Natural Science Research Council (SNF Grant No. 81-6623) is gratefully acknowledged.
References
Adsersen, H. (1976) A Botanist's Notes on Pinta. Noticias de Galdpagos 24, 26-8. Adsersen, H. (1990) Permanent vegetation quadrats in Galfipagos. Monogr. Syst. Bot. Missouri
Bot. Gard. 32, 149-52. de Vries, Tj. (1984) The giant tortoises: A natural history disturbed by Man. In Key Environ-
ments. Galapagos (R. Perry, ed.), pp. 145-56. Oxford: Pergamon Press. de Vries, Tj. and Tupiza, A. (1990) The dynamics of Scalesia cordata (Asteraceae) in different
habitats in Sierra Negra, Isabela, Gal~pagos. Monogr. Syst. Bot. Missouri Bot. Gard. 32,137 47.
Eliasson, U. (1974) Studies in Gal~pagos Plants XIV. The genus Scalesia Arn. Opera Botanica 36, 1-117.
Eliasson, U. (1984) Native Climax Forest. In Key Environments. Galapagos (R. Perry, ed.), pp. 101-14. Oxford: Pergamon Press.
Evans, D. (1990) Informe del Director. 56ta. Reuni6n del Consejo Ejecutivo de la Fundacion Charles Darwin. Octubre 25 de 1990. Quito, Ecuador.
Grant, P.R. (1981) Population fluctutations, tree rings and climate. Noticias de Gahtpagos 33. 12-6.
Grant, P.R. and Grant, B.R. (1989) The slow recovery of Opuntia megasperma on Espanola. Noticias de Gal(lpagos 48, 13-15.
Hamann, O. (1975) Vegetational changes in the Gafftpagos Islands during the period 1966-73. Biol. Conserv. 7, 37-59.
Hamann, O. (1979a) Regeneration of vegetation on Santa Fe and Pinta Islands, Gal~pagos, after the eradication of Goats. Biol. Conserv. 15, 215-36.
Hamann, O (1979b) Dynamics of a stand of Scalesia pedunculata Hooker ill., Santa Cruz Island, Gal~pagos. Bot. J. Linn. Soc. 78, 67-84.
Hamann, O. (1981) Plant Communities of the Gal~tpagos Islands. Dansk Bot. Ark& 34, 1-163. Hamann, O. (1984) Changes and threats to the Vegetation. In Key Environments, Gal{~pagos (R.
Perry, ed.), pp. 115-31. Oxford: Pergamon Press. Hamann, O. (1985) The E1 Nifio influence on the Galfipagos vegetation. In El Nino en Las lslas
Galdpagos. E1 Evento de 1982-1983 (G. Robinson and E.M. del Pino, eds), pp. 299-330. Quito: Impressiones Gr~ficas Isalpro.
Hamann, O. and Wium-Andersen, S. (1986) Scalesia gordilloi sp. nov. (Asteraceae) from the Gal~tpagos Islands, Ecuador. Nordic. J. Bot., 6, 35-8.
Hoeck, H.N. (1984) Introduced Fauna. In Key Environments. Gal(~pagos (R. Perry, ed.), pp. 233-45. Oxford: Pergamon Press.
Itow, S. and Mueller-Dombois, D. (1988) Population structure, stand-level dieback and recovery of Scalesia pedunculata forest in the Gal~ipagos Islands. Ecol, Res. 3, 333-9.
Lawesson, J.E. (1988) Stand-level dieback and regeneration of forests in the Gal~pagos Islands. Vegetatio 77, 87-93,
McMullen, C.K. (1986) Seed germination studies of selected Galfipagos Islands angiosperms. Noticias de Galdpagos 44, 13-15.
Pritchard, P.C.H. (1984) Further thoughts on 'Lonesome George'. Noticias de Gal6pagos 39, 20-3.

Vegetation recovery on Pinta Island, Gal(~pagos 151
Reynolds, R.P. and Marlow, R.W. (1983) Lonesome George, the Pinta Island tortoise: a case of limited alternatives. Noticias de Gal6pagos 37, 14-17.
Seberg, O. (1984) Taxonomy and phylogeny of the genus Acalypha (Euphorbiaceae) in the Gal~pagos Archipelago. Nord. J. Bot. 4, 159-90.
van Denburgh, J. (1914) The gigantic land tortoises of the Gal~ipagos Islands. Proc. Calif. Acad. Sci. 4, 203-374.
Weber, D. (1971) Pinta, Gal~ipagos: Une ile ~ sauver. Biol. Conserv. 4, 8-12. Whelan, P.M. and Hamann, O. (1989) Vegetation regrowth on Isla Pinta: a success story.
Noticias de Gal~pagos 48, 11-13. Wiggins, I.R. and Porter, D.M. (1971) Flora of the Galdpagos Islands. Stanford, California:
Stanford University Press.