nutrient deficiencies in greater yam and their effects on leaf nutrient concentrations
TRANSCRIPT

Journal of Plant Nutrition, 29: 1663–1674, 2006
Copyright © Taylor & Francis Group, LLC
ISSN: 0190-4167 print / 1532-4087 online
DOI: 10.1080/01904160600851569
Nutrient Deficiencies in Greater Yam and TheirEffects on Leaf Nutrient Concentrations
Jane N. O’Sullivan and Rachel Jenner
School of Land and Food Sciences, University of Queensland, Brisbane, Australia
ABSTRACT
This paper describes the first systematic study of nutritional deficiencies of greater yam(Dioscorea alata). Yam plants (cv. ‘Mahoa’a’) were propagated from tuber discs andgrown in nutrient solution, with nutrients supplied following a modified programmednutrient-addition method. After an establishment period of four weeks, deficiencies ofnitrogen (N), phosphorus (P), potassium (K), calcium (Ca), magnesium (Mg), sulfur (S),iron (Fe), boron (B), manganese (Mn), copper (Cu), zinc (Zn), and molybdenum (Mo)were induced by omitting the relevant nutrient from the solution. Foliar symptoms wererecorded photographically. Notably, deficiencies of the mobile macronutrients failedto induce senescence of oldest leaves, while vine growth and younger leaves were af-fected. Leaf blades of the main stem were sampled in sequence and analyzed chemically,providing the distribution of each nutrient from youngest to oldest leaves in both ade-quately supplied and deficient plants. The nutrient-concentration profiles, together withthe visible symptoms, indicated that little remobilization of mobile macronutrients hadoccurred. For both macro- and micronutrients, young leaves gave the best separation ofnutrient concentrations between well-nourished and deficient plants.
Keywords: index leaf, nutrient mobility, nutrient omission, solution culture, symptoms,tissue analysis
INTRODUCTION
Yams are generally regarded as demanding on soil fertility. In shifting agri-cultural systems, they are almost invariably planted first in the cropping cycle,immediately after clearing the fallow. In areas where fallow length has beenreduced due to land pressure, yams are grown less frequently, as the soil is
Received 15 April 2005; accepted 11 January 2006.Address correspondence to J. N. O’Sullivan, School of Land and Food Sciences,
University of Queensland, Brisbane 4072, Australia. E-mail: [email protected]
1663

1664 J. N. O’Sullivan and R. Jenner
regarded as insufficiently fertile for them. Yet very little research has beendone to determine which nutrients most limit yam production, and how specificnutritional disorders can be diagnosed and corrected.
Gaztambide and Cibes (1975) induced several nutrient deficiencies inDioscorea rotundata cv. ‘Habanero,’ grown in sand culture, and described leafsymptoms and relative tuber production. Critical tissue concentrations were notestablished in their study.
In a review of early research on yam nutrition, Obigbesan (1981) foundthat such research has focused primarily on fertilizer responses to the macronu-trients nitrogen (N), phosphorus (P), and potassium (K). He expressed a needfor a wider understanding of other yield-limiting nutrients and for more predic-tive diagnostic procedures. The focus on major nutrients has continued sincethen, although a number of studies have extended the knowledge of fertilizer re-sponses by examining fertilizer-use efficiency (Castillo- Rodriguez et al., 1989),nutrient removal (Irizarry et al., 1995), and optimum timing and placement offertilizers (Nwinyi and Enwezor, 1985). However, none of these studies hasattempted to relate fertilizer responses to plant tissue or soil-nutrient levels ina way that can be used to predict nutrient requirements at other sites.
This paper presents preliminary findings of a study of nutritional disordersof greater yam (Dioscorea alata). The experiment aimed to generate symptomsof a range of nutrient deficiencies, and to examine associated changes in leaf-nutrient concentrations in leaves of differing age. These data were intended toassist the selection of appropriate index tissues for diagnostic tissue analysis.Subsequent work is under way to define a critical concentration in the selectedindex tissue, indicative of deficiency of each nutrient.
MATERIALS AND METHODS
Tuber discs (approx. 300 g) of yam (D. alata cv. ‘Mahoa’a’) were sproutedunder intermittent mist [200 µM calcium (Ca) and 3 µM boron (B) in deion-ized water]. Rooted shoots were excised with approximately 30 g of tuber andtransferred to solution culture. Each plant was supported over 20 L of nutrientsolution, using a plug of polyurethane foam inside a polyethylene bag. Ini-tial nutrient concentrations were (µM): nitrogen (N), 103; phosphorus (P), 6;potassium (K), 51; Ca, 521; magnesium (Mg), 100; sulfur (S), 6; iron (Fe),10; and B, 0.3. Other micronutrients [manganese (Mn), zinc (Zn), copper (Cu),and molybdenum (Mo)] were withheld during the establishment period. Nutri-ents were added weekly according to anticipated demand over the followingweek (Asher and Blamey, 1987), adjusted by actual removal (K measured bypotassium electrode, and other nutrients added proportionally). Solution pHwas adjusted weekly to maintain pH in the range of 5.5–6.0. After an establish-ment period of 32 d, the original tuber piece was removed, plants were blockedaccording to their vine growth over the establishment period, treatments wereallocated randomly within blocks, and solutions were replaced. Treatments

Nutrient Deficiencies in Yam 1665
consisted of the omission of one nutrient (N, P, K, Ca, Mg, S, Fe, B, Mn, Zn,Cu or Mo) from the nutrient solution, and were replicated in five randomizedcomplete blocks, each block also containing two control pots receiving all nu-trients. Weekly measurements of vine height, solution K concentration, andsolution P concentration were made, and weekly nutrient additions adjustedsuch that nutrient concentrations were similar for each pot within a block, andsufficient to meet the demands of the fastest-growing plant within that block.As growth varied considerably between the blocks, blocks were harvested se-quentially from 37 d to 65 d after application of treatments for blocks 1 to5, respectively, as sufficient symptom development was achieved. Symptomswere recorded and photographed as they developed. The experiment was con-ducted at 27.5◦ S from February 2 to May 10, day length 12 h, with mean dailymaximum/minimum temperatures in the greenhouse of 42◦C/32◦C during theestablishment period and 38◦C/27◦C during the treatment period.
Harvested plants were separated into roots, tubers, and vines, and leafblades on the dominant vine were sampled in groups of four nodes, fromthe youngest open leaf. The groupings were necessary to ensure large enoughsamples for analysis, especially of immature and severely stunted leaves. Dryweights were recorded and leaf tissue analyzed by inductively coupled plasmaatomic emission spectrometer (ICP-AES) following nitric and perchloric aciddigestion (for P, K, Ca, Mg, S, Fe, Mn, Zn, Cu, Mo) or following dry ashing (forB). Copper and Mo concentrations were checked by additional analysis by in-ductively coupled plasma mass spectrometer (ICP-MS). Nitrogen was analyzedusing a Leco CNS combustion analyzer. Data were analyzed using the Gen-eral Linear Means procedure of the SAS software; and least squares means arequoted. For leaf-nutrient concentrations, statistical procedures were performedon log-transformed data.
RESULTS AND DISCUSSION
Growth and Yield Data
Plant establishment in the solution culture system was slow and variable. Fun-gal root infections were a frequent problem. Plants were found to be highlydependent on the remaining tuber piece in the initial weeks, despite consider-able root growth. The small size of the tuber piece was necessary to minimizethe plant’s nutrient reserves, but slow establishment was a consequence.
Vines of D. alata may have nodes with either one or two leaves (i.e., ineither alternate or opposite arrangement), and may change leaf arrangementintermittently. In general, vines were initially alternate and changed to oppositeas they matured, but the timing was highly variable and temporary reversionswere common. Some nutritional stresses may have delayed the transition, butsample sizes in this experiment were insufficient to characterize the effectsof specific disorders. These changes altered the relationships between vine

1666 J. N. O’Sullivan and R. Jenner
height and dry-matter production, the rate of leaf area acquisition, and rateof production of new nodes. A vine with alternate leaves tended to producemore nodes but fewer leaves over a period of time than an equivalent vine withopposite leaves. Thus, the node number assigned to leaves for analysis is animperfect surrogate for their actual or physiological age, contributing to thevariability of leaf analysis data. To have sampled on the basis of leaf numberwould have caused similar errors in the opposite direction.
Table 1 summarizes the dry-matter yield from each treatment at harvest.All omission treatments except −Fe and −Mo significantly reduced vine dryweight, although for −Mg the total weight reduction was significant only atP< 0.1. Variability at the time of imposing treatments may have affected finalweights. However, analysis of the increase in vine length after treatments wereinitiated gave a statistical separation of treatments very similar to that of vinedry weight. Blocking minimized the effects of initial size, and analysis of thevine height at treatment initiation revealed no significant differences amongtreatments; hence, the initial variability was unbiased.
The experimental period was too short to allow relative tuber yield toreflect the field situation. Notably, control plants had a relatively low harvestindex. This may be expected if nutritional stress promoted early initiation of
Table 1Dry-matter yield of D. alata plants grown in full nutrient solution or in solutions omittingone nutrient
Dry weight (g)
Treatment Total Vines Tubers Roots Harvest index
All 26.27 a 17.58 a 5.93 a b 2.76 a 0.22 bminus N 16.70 bc 6.14 c d 8.55 a 2.01 a b c 0.52 aminus P 14.46 bc 8.89 c d 3.40 b c 2.17 a b c 0.22 bminus K 11.94 c 7.30 c d 3.48 b c 1.16 cd 0.30 bminus Ca 10.42 c 3.51 d 5.77 a b c 1.15 cd 0.56 aminus Mg 17.73 a b c 10.32 b c 5.34 a b c 2.07 a b c 0.32 bminus S 15.87 b c 8.68 b c d 4.59 a b c 2.61 a b 0.30 bminus Fe 25.11 a b 15.21 a b 7.69 a b 2.21 a b c 0.32 bminus B 15.63 b c 8.70 c d 5.25 a b c 1.68 b c d 0.29 bminus Mn 8.94 c 6.42 c d 1.64 c 0.89 d 0.18 bminus Zn 14.83 b c 9.65 b c d 3.52 b c 1.66 b c d 0.27 bminus Cu 14.44 b c 9.92 b c 2.93 b c 1.60 b c d 0.18 bminus Mo 19.88 a b c 12.35 a b c 5.69 a b c 1.84 a b c d 0.26 b
Harvest index is the weight of the tubers divided by the total plant weight. Numbers arethe mean of eight replicates (“all”) or four replicates (other treatments); those followedby the same letter are not significantly different at P < 0.05.

Nutrient Deficiencies in Yam 1667
tubers, while control plants may have had greater potential for tuber fillinglater in the season. However, the relatively high harvest indices for N- andCa-deficient plants suggest a shift in root:shoot sink strength. A similar effecthas been commonly reported for N nutrition in other root crops, in whichgenerous use of N fertilizers tends to cause yield decline while promoting topgrowth (Harper and Walker, 1985). Likewise, Ca deficiency in ginger (Zingiberofficinale L.) has been reported to restrict shoot production severely, whilerhizome growth was increased (Islam, 1980).
Symptoms
Symptom development in D. alata was strikingly different from that in otherspecies in our experience. Severe symptoms of deficiency were induced forseveral micronutrients (particularly Mn and Cu) that are difficult to achieve inmany species, especially those that are vegetatively propagated. In contrast, anddespite conspicuous growth retardation, deficiencies of the mobile macronutri-ents (N, P, K, and Mg) caused little or none of the typical older-leaf symptomsthat are readily produced in most species grown in solution culture. Most de-ficiencies increased the chlorosis of young leaves, but as control plants alsodisplayed a gradual darkening of green pigmentation from young to matureleaves, the relative chlorosis associated with deficiency is difficult to define asa useful diagnostic feature.
Deficiencies of N, S, and Ca produced similar symptoms on the vines(Plate 1A, B, and C respectively). Youngest leaves were pale green to cream incolor, darkening gradually to dark green in the oldest leaves. Calcium-deficientplants tended to have small leaves and thin vines, and some plants developedsmall necrotic lesions on young mature leaves (Plate 1D). In both Ca-deficientand S-deficient plants, mid-leaves occasionally developed necrosis along themain veins, extending from the petiole attachment on the lower leaf surface(Plate 1C, lower leaves). Plants deficient in N and S produced long, taperingtubers similar to those on control plants, while Ca-deficient plants producedshort, turnip-shaped tubers.
Phosphorus deficiency was associated with reduced size of young leaves,usually with some chlorosis (Plate 1E). Leaf tips often curled under and weresometimes necrotic. Leaves of all ages were generally thickened and stiff. Youngmature leaves often exhibited diffuse interveinal chlorosis and frequently de-veloped dark necrotic spots scattered in the interveinal tissue (Plate 1F).
Leaves of both K-deficient and Mg-deficient plants developed a mild inter-veinal chlorosis. All mature leaves were affected by Mg deficiency, but underK deficiency, leaves of intermediate age were most affected. The mild symp-toms associated with K deficiency were surprising given the extent of growthreduction. Under K deficiency, dark green persisted adjacent to both majorand secondary veins, with a fairly abrupt transition, giving the veins a feath-ered appearance (Plate 1G). Under Mg deficiency, the transition was more

1668 J. N. O’Sullivan and R. Jenner
Plate 1. A. N-deficient shoot. B. S-deficient shoot. C. Ca-deficient shoot. D. Youngmature leaf on Ca-deficient plant, showing necrotic spots. E. Young leaves of P-deficientplant showing chlorosis, size reduction, stiffening, and downward curling of tips. F.Young mature P-deficient leaves with interveinal chlorosis and necrotic spots. G. Midleafon a K-deficient plant showing interveinal chlorosis. H. Mature leaf on a Mg-deficientplant showing diffuse interveinal chlorosis and tip necrosis. I. Fe-deficient young matureleaf showing chlorosis between fine network of veins. J. Severe chlorosis and necrosison a young leaf due to Fe deficiency. K. Young B-deficient leaves showing stunting,mild chlorosis and bronzing, and tip deflection. L. B-deficient leaves showing smallsize, gathering along main veins, and interveinal chlorotic and necrotic patches.
gradual, producing a diffuse interveinal mottle (Plate 1H). Leaf tips often be-came necrotic, but the necrosis did not spread. On only one Mg-deficient plantdid interveinal chlorosis progress to yellowing on two older leaves, and light-brown necrotic lesions formed on the yellow tissue.

Nutrient Deficiencies in Yam 1669
Symptoms of micronutrient deficiencies were generally more typical ofthose reported for other species. Iron deficiency resulted in a finely delineatedinterveinal chlorosis on mature leaves (Plate 1I), and pale green to whitishyoung leaves, usually with light green on main veins. In the most severe cases,light-brown necrotic lesions spread over the young leaf blades (Plate 1J).
Boron-deficient plants developed small, broad young leaves that werethickened and brittle, and were sometimes mildly chlorotic or bronzed. Of-ten the leaf tip was bent either up or down (Plate 1K). On some leaves, thetissue was puckered or gathered along the veins (Plate 1L). In the most se-vere cases, the apical meristem died, and subsequent axillary shoots emerg-ing near the base of the plant produced deformed leaves of very reducedsize.
Manganese deficiency was the most severe of the disorders induced.Young leaves were pale and somewhat translucent, and developed small in-terveinal pale spots that were slightly sunken and soon became necrotic(Plate 2A). Leaves were reduced in size, and expanding leaves were con-stricted by the necrotic lesions (Plate 2B). Vine wings also became necroticin patches. Mature leaves developed numerous dark necrotic spots scatteredover the entire leaf blade (Plate 2C). Leaf painting of newly open leaves with1% MnSO4 prevented lesion development and increased leaf expansion andgreening (Plate 2D). In the most severe cases, the apical meristem died, andsecondary shoots that erupted near the tip failed to develop beyond a fewcentimeters.
Copper deficiency caused distinctive deformity and size reduction of youngleaf blades (Plate 2E). Initially, leaf blades became short and broad, often withnecrotic tips, and cupped upward, with puckering adjacent to the main veins(Plate 2F). These leaves were often very shiny, and some developed a diffusewhitish bleaching of interveinal tissue. As the symptoms intensified, new leavesfailed to expand beyond 1–2 cm. The apical tip became inactive, prompting aprofusion of axillary shoot growth from the uppermost nodes, all producingshoots with normal vine thickness and internode length but very reduced leaves(Plate 2G).
Zinc deficiency was relatively mild in this experiment, as reflected bythe modest growth reduction of vines. Most plants grew healthy foliage forsome time, but eventually the youngest leaves became pale and elongated inshape (Plate 2H). Mature leaves developed a mild general chlorosis, whichwas reversed by painting with 0.5% ZnSO4 (Plate 2I). On axillary shoots,leaf blades were markedly reduced in size, and the elongated leaf tip becametwisted.
The −Mo treatment showed little sign of deficiency in plant growth. Latein the experiment, a fine interveinal chlorotic mottle was observed on olderleaves (Plate 2J). In one case, necrotic lesions developed in the interveinaltissue (Plate 2K). These began as small speckles, and expanded and coalescedto form irregularly shaped, russet-brown lesions of 2–5 mm.

1670 J. N. O’Sullivan and R. Jenner
Plate 2. A. Young leaf showing early signs of Mn deficiency, with numerous pale,sunken, interveinal spots, some becoming necrotic. B. Tip of Mn-deficient plant show-ing the development of symptoms as leaves expand, and necrosis on the stem wings.C. Mature leaves (upper and lower surfaces) showing dark necrotic spots due to Mndeficiency. D. A young Mn-deficient leaf, painted on the left side with a solution of 1%MnSO4 when newly opened. Symptom development has been greatly reduced on thepainted side. E. Young leaves on a Cu-deficient leaves showing shortened shape withnecrotic tips and gathering along the main veins. G. Upper part of a Cu-deficient vine,showing proliferation of axillary shoots following shoot-tip death, but greatly reducedleaf blades. F. Young Cu-deficient leaves showing shortened shape with necrotic tipsand gathering along the main veins. G. Upper part of a Cu-deficient vine, showing pro-liferation of axillary shoots following shoot-tip death, but greatly reduced leaf blades onthe new shoots. H. Young expanding leaves showing narrowing and chlorosis due to Zndeficiency. I. A young mature leaf on which the right half was painted with a solution of0.5% ZnSO4, demonstrating reversal of chlorosis due to Zn deficiency. J. An older leafon a Mo-deficient plant, showing chlorosis between minor veins. K. Oldest leaves on aMo-deficient plant, with necrotic spots developing in interveinal chlorotic patches.

Nutrient Deficiencies in Yam 1671
Leaf-Nutrient Concentrations
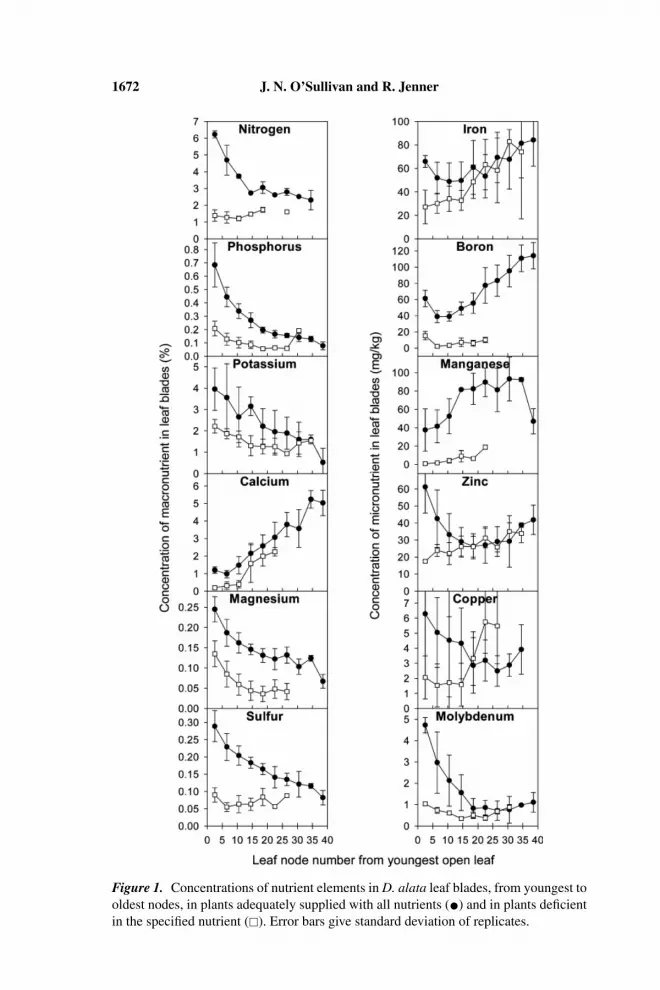
The concentrations of nutrient elements in leaf blades are summarized inFigure 1. In general, in well-nourished plants, the mobile macronutrientsshowed a steep decline in concentration from the youngest to the oldest leavesIn deficient plants, the concentration profile was usually flatter (with the ex-ception of Mg deficiency) such that the greatest difference between the controland deficient plants was found in the youngest leaves. The oldest leaves hadconcentrations similar to those in the well-nourished plants. This result is inmarked contrast to the pattern observed in sweet potato (O’Sullivan et al.,1996), in which mobile nutrients tended to have a relatively uniform concentra-tion across all leaf ages in healthy plants, but to decline markedly from youngestto mature leaves in deficient plants.
The relatively flat age profile of these nutrients in deficient yam plantssupports the idea that remobilization of nutrients is inefficient in this species.This result is consistent with the observed lack of older-leaf symptoms in plantsdeficient in these nutrients.
Concentrations of Ca, Fe, B, and Mn all tended to increase with leafage in control plants, although they may have been slightly elevated in theyoungest leaf sample, where incompletely expanded blades may have had aconcentrating effect. In contrast, concentrations of Zn, Cu, and Mo tendedto decline with leaf age. The shape of the profile in deficient plants may re-flect the development of deficiency under that treatment. Zinc and Fe divergedfrom the healthy curve only in young leaves, corresponding to the relativelylate and mild development of deficiency in these nutrients. The B concen-trations were consistently low, despite the variability in producing acute B-deficiency symptoms. This result might suggest relative mobility of B in thisspecies.
The high variability in Cu data prevents a confident separation of healthyand deficient concentrations. Only a small proportion of this variation can beattributed to measurement error, as standard leaf material, digested with eachbatch of leaf samples, consistently tested close to certified Cu concentrations.Thus, both healthy and deficient plants appeared to be highly variable in Cuconcentration, which was somewhat surprising given the consistently severedevelopment of Cu-deficiency symptoms.
Overall, young leaves showed the best separation between healthy anddeficient plants. However, the very youngest sample, containing expandingleaves from nodes 1 to 4, diverged from the nutrient profiles of subsequentsamples, indicating that their nutrient concentrations depended considerablyon the degree of leaf expansion. Node number may be only a crude indicatorof the degree of expansion under variable field conditions. For the purposesof diagnostic sampling, a less variable tissue is preferred. Very small leavesare also undesirable due to the number needed to make a sample. From ourobservations, leaf expansion may continue until between the sixth and tenth
1672 J. N. O’Sullivan and R. Jenner
Figure 1. Concentrations of nutrient elements in D. alata leaf blades, from youngest tooldest nodes, in plants adequately supplied with all nutrients (•) and in plants deficientin the specified nutrient (�). Error bars give standard deviation of replicates.

Nutrient Deficiencies in Yam 1673
nodes. However, sampling from the tenth node in the field is quite difficult, asit is generally more than a meter from the vine tip and well entangled in themass of vines typical of field-grown yams. As an index leaf, a sampling fromabout the sixth node may provide a suitable compromise between maturity andaccessibility. Further investigation of variation among and within the first 10nodes is to be undertaken before final selection of an index tissue.
ACKNOWLEDGMENTS
This study was funded by the Australian Centre for International AgriculturalResearch (ACIAR), as part of the project “Diagnosis and Correction of Nutri-tional Disorders in Yam.” Janette Mercer and David Appleton are thanked forassistance with tissue and solution analyses.
REFERENCES
Asher, C. J., and F. P. C. Blamey. 1987. Experimental control of plant nutrientstatus using programmed nutrient addition. Journal of Plant Nutrition 10:1371–1380.
Castillo-Rodriguez, J., L. A. Ruiz Martinez, J. M. Portieles Rodriguez, and J.O. Milian Morales. 1989. Nutrient uptake and utilization coefficients forfertilizers and soil nutrients in a yam (Dioscorea alata) crop. Cientia yTecnica en la Agricultura, Viandeas Tropicales 12(1): 59–73.
Gaztambide, S., and H. R. Cibes. 1975. Nutritional deficiencies of yams(Dioscorea sp.) and related effects on yield and leaf composition. Jour-nal of Agriculture of the University of Puerto Rico 59: 264–272.
Harper, T. W., and I. R. Walker. 1985. Response of a sweet potato variety tolevels of nitrogen and potassium applied at planting. Queensland Journalof Agricultural and Animal Sciences 42: 23–27.
Irizarry, H., R. Goenaga, and U. Chardon. 1995. Nutrient uptake and dry matteryield in the ‘Gunung’ yam (Dioscorea alata) grown on an Ultisol withoutvine support. Journal of Agriculture of the University of Puerto Rico 79(3-4): 121–130.
Islam, A. K. M. S. 1980. Effects of soil pH on yield and mineral nutrition ofginger (Zingiber officinale Roscoe), PhD diss., University of Queensland.
Nwinyi, S. C. O., and W. O. Enwezor. 1985. Evaluation of fertilizer placementmethods for white yam (Dioscorea rotundata). Experimental Agriculture21(1): 73–78.
Obigbesan, G. O. 1981. Nutrient requirements of yams (Dioscorea species).Agricultural Research Bulletin Vol. 2, No. 1. Ibadan, Nigeria: Ibadan Uni-versity Press.

1674 J. N. O’Sullivan and R. Jenner
O’Sullivan, J. N., F. P. C. Blamey, and C. J. Asher. 1996. Diagnostic criteria fornutrition disorders of sweet potato: II. Critical nutrient concentrations inleaves. In Mineral nutrient disorders of root crops in the Pacific, ACIARProceedings No. 65, eds. E. T. Craswell, C. J. Asher, and J. N. O’Sullivan,39–43. Canberra, Australia: Australian Center for International Agricul-tural Research.