nocardiopsis species: incidence, ecological roles and adaptations
TRANSCRIPT

R
N
Ta
b
a
ARRAA
KNSMAE
C
h0
Microbiological Research 174 (2015) 33–47
Contents lists available at ScienceDirect
Microbiological Research
j ourna l h om epage: www.elsev ier .com/ locate /micres
eview
ocardiopsis species: Incidence, ecological roles and adaptations
ahsin Bennura, Ameeta Ravi Kumara, Smita Zinjardea,∗, Vaishali Javdekarb,∗∗
Institute of Bioinformatics and Biotechnology, Savitribai Phule Pune University, Pune 411007, IndiaDepartment of Biotechnology, Abasaheb Garware College, Pune 411004, India
r t i c l e i n f o
rticle history:eceived 11 November 2014eceived in revised form 13 March 2015ccepted 16 March 2015vailable online 21 March 2015
eywords:ocardiopsisoilarine
ntibioticsnzymes
a b s t r a c t
Members of the genus Nocardiopsis are ecologically versatile and biotechnologically important. Theyproduce a variety of bioactive compounds such as antimicrobial agents, anticancer substances, tumorinducers, toxins and immunomodulators. They also secrete novel extracellular enzymes such as amylases,chitinases, cellulases, �-glucanases, inulinases, xylanases and proteases. Nocardiopsis species are aerobic,Gram-positive, non-acid-fast, catalase-positive actinomycetes with nocardioform substrate mycelia andtheir aerial mycelia bear long chains of spores. Their DNA possesses high contents of guanine and cyto-sine. There is a marked variation in properties of the isolates obtained from different ecological nichesand their products. An important feature of several species is their halophilic or halotolerant nature. Theyare associated with a variety of marine and terrestrial biological forms wherein they produce antibioticsand toxins that help their hosts in evading pathogens and predators. Two Nocardiopsis species, namely,N. dassonvillei and N. synnemataformans (among the thirty nine reported ones) are opportunistic humanpathogens and cause mycetoma, suppurative infections and abscesses. Nocardiopsis species are present insome plants (as endophytes or surface microflora) and their rhizospheres. Here, they are reported to pro-duce enzymes such as �-amylases and antifungal agents that are effective in warding-off plant pathogens.They are prevalent as free-living entities in terrestrial locales, indoor locations, marine ecosystems and
hypersaline habitats on account of their salt-, alkali- and desiccation-resistant behavior. In such naturallocations, Nocardiopsis species mainly help in recycling organic compounds. Survival under these diverseconditions is mediated by the production of extracellular enzymes, antibiotics, surfactants, and the accu-mulation of compatible solutes. The accommodative genomic features of Nocardiopsis species supporttheir existence under the diverse conditions where they prevail.© 2015 Elsevier GmbH. All rights reserved.
ontents
1. Introduction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 342. Nocardiopsis species and other living forms: incidence, isolation, and their role therein . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 35
2.1. Ecological perspectives of Nocardiopsis species associated with biological forms . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 352.2. The role of Nocardiopsis species associated with animal forms in addressing future societal challenges . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 362.3. Nocardiopsis species associated with plants . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 38
3. Occurrence of Nocardiopsis species in terrestrial, indoor, marine, hypersaline, cold locations and their ecological role . . . . . . . . . . . . . . . . . . . . . . . . . . 384. Adaptations and survival modes . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 41
4.1. Enzymes produced by Nocardiopsis species . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 414.2. Production of antimicrobial agents and biosurfactants . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 414.3. Compatible solutes in halophilic Nocardiopsis species . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 42
5. Genomic and proteomic features related to survival . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 426. Conclusion . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 44
Conflict of interest . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .Acknowledgements . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .References . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
∗ Corresponding author. Tel.: +91 020 25601385; fax: +91 020 25690087.∗∗ Corresponding author. Tel.: +91 020 41038244.
E-mail addresses: [email protected] (S. Zinjarde), [email protected] (V.
ttp://dx.doi.org/10.1016/j.micres.2015.03.010944-5013/© 2015 Elsevier GmbH. All rights reserved.
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 45 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 45
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 45
Javdekar).
3 gical R
1
tdfdaotofea‘
cptMttap5hTnnP
F(
4 T. Bennur et al. / Microbiolo
. Introduction
The genus Nocardiopsis was first described by Meyer in 1976. Onhe basis of morphological and biochemical criteria, ‘Actinomaduraassonvillei (Brocq-Rousseu) Lechevalier and Lechevalier’ and theormer ‘Nocardia dassonvillei (Brocq-Rousseu) Liegard and Lan-rieu’, were transferred from the genus ‘Actinomadura Lechevaliernd Lechevalier’ to a new genus, ‘Nocardiopsis’ (Nocardia – a genusf the order Actinomycetales; opsis – appearance). On the basis ofhe phylogenetic position, morphological features and chemotax-nomic properties displayed by Nocardiopsis species, a new familyor the genus referred to as ‘Nocardiopsaceae’ was created (Raineyt al. 1996). The genus ‘Nocardiopsis’ was phylogenetically coherentnd represented a distinct lineage within the radiation of the order
Actinomycetales’.This genus includes aerobic, Gram-positive, non-acid-fast,
atalase-positive actinomycetes. Colonies on organic media agarlates are coarsely wrinkled or folded. Their colonies are similaro members of the genera Nocardia and Actinomadura (Cook and
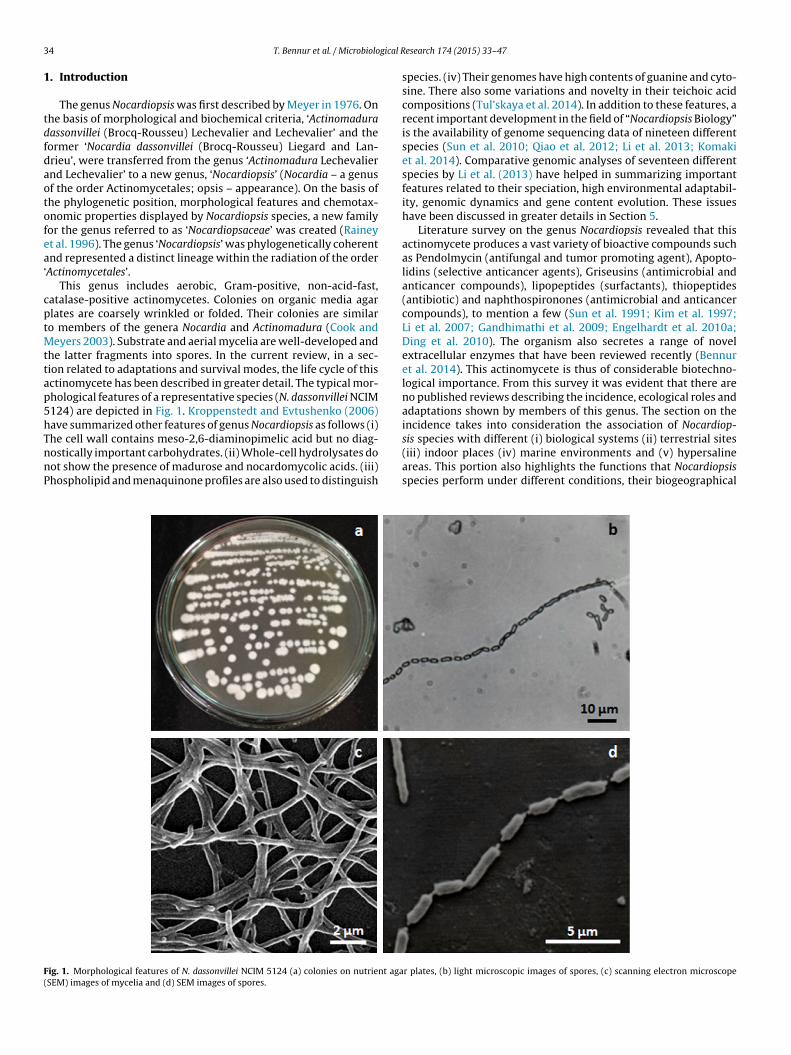
eyers 2003). Substrate and aerial mycelia are well-developed andhe latter fragments into spores. In the current review, in a sec-ion related to adaptations and survival modes, the life cycle of thisctinomycete has been described in greater detail. The typical mor-hological features of a representative species (N. dassonvillei NCIM124) are depicted in Fig. 1. Kroppenstedt and Evtushenko (2006)ave summarized other features of genus Nocardiopsis as follows (i)
he cell wall contains meso-2,6-diaminopimelic acid but no diag-ostically important carbohydrates. (ii) Whole-cell hydrolysates doot show the presence of madurose and nocardomycolic acids. (iii)hospholipid and menaquinone profiles are also used to distinguishig. 1. Morphological features of N. dassonvillei NCIM 5124 (a) colonies on nutrient agaSEM) images of mycelia and (d) SEM images of spores.
esearch 174 (2015) 33–47
species. (iv) Their genomes have high contents of guanine and cyto-sine. There also some variations and novelty in their teichoic acidcompositions (Tul’skaya et al. 2014). In addition to these features, arecent important development in the field of “Nocardiopsis Biology”is the availability of genome sequencing data of nineteen differentspecies (Sun et al. 2010; Qiao et al. 2012; Li et al. 2013; Komakiet al. 2014). Comparative genomic analyses of seventeen differentspecies by Li et al. (2013) have helped in summarizing importantfeatures related to their speciation, high environmental adaptabil-ity, genomic dynamics and gene content evolution. These issueshave been discussed in greater details in Section 5.
Literature survey on the genus Nocardiopsis revealed that thisactinomycete produces a vast variety of bioactive compounds suchas Pendolmycin (antifungal and tumor promoting agent), Apopto-lidins (selective anticancer agents), Griseusins (antimicrobial andanticancer compounds), lipopeptides (surfactants), thiopeptides(antibiotic) and naphthospironones (antimicrobial and anticancercompounds), to mention a few (Sun et al. 1991; Kim et al. 1997;Li et al. 2007; Gandhimathi et al. 2009; Engelhardt et al. 2010a;Ding et al. 2010). The organism also secretes a range of novelextracellular enzymes that have been reviewed recently (Bennuret al. 2014). This actinomycete is thus of considerable biotechno-logical importance. From this survey it was evident that there areno published reviews describing the incidence, ecological roles andadaptations shown by members of this genus. The section on theincidence takes into consideration the association of Nocardiop-
sis species with different (i) biological systems (ii) terrestrial sites(iii) indoor places (iv) marine environments and (v) hypersalineareas. This portion also highlights the functions that Nocardiopsisspecies perform under different conditions, their biogeographicalr plates, (b) light microscopic images of spores, (c) scanning electron microscope
T. Bennur et al. / Microbiological Research 174 (2015) 33–47 35
Table 1Features of Nocardiopsis species associated with different animal forms.
Nocardiopsis sp. Host animal Role therein/other features Reference
Nocardiopsis sp. Marine sponge (Hymeniacidon perleve);Yellow Sea, China
Nocardiopsis sp. recovered as second most dominant genusafter Streptomyces; role: not described
Zhang et al. (2006)
N. dassonvillei Marine sponge (Haliclona sp.); South ChinaSea
One N. dassonvillei isolate recovered; Streptomyces mostpredominant; genes for PKS and NRPS present
Jiang et al. (2007)
N. alba strain MSA10 Marine sponge (Fasciopongia cavernosa);Southwest Coast, India
Produced lipopeptide biosurfactant with antimicrobialproperties
Gandhimathi et al. (2009)
Nocardiopsis strainHB383
Sponge (Halichondria panacea); Baltic Sea,Germany
One Nocardiopsis sp. recovered; Streptomyces mostpredominant; displayed antibacterial activity; had PKSgene cluster; produced cytotoxic dicyclopeptide
Schneemann et al. (2010a,b)
Nocardiopsis sp. Five Gorgonian corals species investigated;South China Sea
Two Nocardiopsis sp. isolated from Melitodes squamata andSubergorgia suberosa; Streptomyces predominant;Nocardiopsis sp. display no antimicrobial activity
Zhang et al. (2013)
N. coralliicola DSM45611
Gorgonian coral (Menella praelonga);Weizhou Island, China
Novel species; coralliicola: inhabitant of corals; notinvestigated for bioactive compounds
Li et al. (2012)
N. litoralis DSM 45168 Sea anemone; South China Sea Novel species; litoralis: of the shore; not investigated forbioactive compounds
Chen et al. (2009)
N. alba CR167 Thick shelled mollusk (Conus rolani);MactanIsland, Philippines
N. alba present as a casual symbiont; producedNocapyrones with neuroactive and cytotoxic effects; onetype II and two type I PKS gene clusters present
Lin et al. (2013)
N. dassonvillei Puffer fish (Fugu rubripes); Bohai Sea, China N. dassonvillei isolated from ovaries produced Tetrodotoxin Wu et al. (2005a)Nocardiopsis EWC9 Earthworm (Pheretima posthuma);
Uttarakhand, IndiaStreptomyces (60%) predominant; Nocardiopsis sp. showedantimicrobial activity toward Staphylocccus aureus
Kumar et al. (2012)
Nocardiopsis sp. Mosquito (Culex quinquefasciatus);Pondicherry, India
Minor gut flora of larvae along with Escherichia, Proteus,Aspergillus and Streptomyces; incubation of Nocardiopsis sp.with larvae caused 100% mortality
Vasanthi and Hoti (1992)
N. alba IM1-3 Honeybee (Apis mellifera); Thailand Streptomyces predominant; N. alba showed antimicrobialactivity toward causative agents of American andEuropean foulbrood diseases; may be involved in negativeantagonism
Promnuan et al. (2009)
N. alba BE74 (ATCCBAA-2165)
Honeybee; Southeastern Ohio, USA Streptomyces predominant; most actinomycetes inhibitedBacillus marisflavi; Showed the presence of phenazinebiosynthetic cluster; whole genome sequenced; 8 geneclusters for secondary metabolite synthesis identified
Patil et al. (2010) and Qiaoet al. (2012)
Nocardiopsis S1 Goat; Henan Province, China Oerskovia and Nocardiopsis sp. predominant; may beprobiotic actinomycetes
Tan et al. (2009)
N. dassonvillei Man (Homo sapiens); USA, Poland, France,Spain, India
Opportunistic pathogen causing infection of nasalvestibule, lungs, bile duct and skin
Sindhuphak et al. (1985),Mordarska et al. (1998), Beauet al. (1999), González-Lópezet al. (2011), Rudramurthy
d from
dtpod
2i
vroapia
2w
stsee
N. synnemataformans Man (Homo sapiens), Turkey Obtaine
istribution and phylogenetic relationships. In the section relatedo the adaptations, (i) the secretion of different enzymes (ii) theroduction of various antibiotics and surfactants (iii) accumulationf compatible solutes and (iv) relevance of genomic features areiscussed.
. Nocardiopsis species and other living forms: incidence,solation, and their role therein
Nocardiopsis species have been found in association with aariety of invertebrate and vertebrate animals from marine and ter-estrial locations (Table 1). As summarized in Fig. 2, the significancef Nocardiopsis species isolated from animal sources can be gener-lly categorized with respect to (i) the ecological roles that theyerform during such associations and (ii) the futuristic use of these
solates in addressing societal challenges. In addition, a sub-sectionlso summarizes the association of Nocardiopsis species with plants.
.1. Ecological perspectives of Nocardiopsis species associatedith biological forms
There are reports on the isolation of Nocardiopsis species fromponges present in China, India and Germany (Table 1). From
he ecological point of view, it is observed that sponges showymbiotic associations with bacteria (Lee et al. 2001). In gen-ral, the sessile, filter-feeding animals offer bacteria with a safenvironment and better nutrition (sponges hold more nutrientset al. (2012) the sputum of a kidney transplant Yassin et al. (1997)
than sea water or sediments). Sponges in turn, are said to benefitfrom the defense chemicals (toxins, antibiotics and other bioac-tive compounds) synthesized by bacteria that protect them frompathogens and predators. In marine environments, there is also apositive correlation between antimicrobial activity and the absenceof epibiosis. Such activities prevent the establishment of bacterialfilms that are regarded as the initial step in succession (Wahl et al.2012). The production of such antimicrobial compounds conferson the host harboring them, survival advantages particularly, inwarding-off pathogenic bacteria (Zhang et al. 2013). In additionto sponges, Gorgonian corals and sea anemones (obtained fromChina) are also reported to harbor Nocardiopsis species. Zhang et al.(2013) isolated two coral-associated Nocardiopsis isolates that didnot produce bioactive compounds. These biological forms havealso yielded some new species. For example, two novel species N.litoralis and N. coralliicola have been isolated from sea anemone andcoral specimens, respectively (Chen et al. 2009; Li et al. 2012). Sofar, these cultures have not been investigated for the production ofbioactive compounds.
In some cases, Nocardiopsis species symbiotically existing indifferent marine forms produce neuroactive and cytotoxic com-pounds. For example, N. alba CR167 was found to exist as a casualsymbiont in the mucus generating cells and the venom duct of the
thick-shelled mollusc, Conus rolani present in Phillipines. StrainCR167 produced neuroactive and cytotoxic polyketide pyronesreferred to as Nocapyrones (Lin et al. 2013). In addition to theheavy shells and toxic venom that protect these molluscs, such
36 T. Bennur et al. / Microbiological Research 174 (2015) 33–47
ciated
bdaibdc
dmloIbflogwihmadMfaNistctabe
Fig. 2. Highlights of the ecological relevance of Nocardiopsis species asso
ioactive small molecules were proposed to be significant inefense. Another relevant toxin is the tetrodotoxin (TTX) associ-ted with the puffer fish, Fugu rubripes. The toxicity of this fishs attributed to the presence of certain tetrodotoxin producingacteria (Wu et al. 2005b). One bacterial culture (identified as N.assonvillei) associated with the specimens of this fish in Chinaontributed toward its toxicity (Wu et al. 2005a).
Among the terrestrial forms, the members of genus Nocar-iopsis have been found in association with invertebrates andammals (Table 1 and Fig. 2). In one such investigation, an iso-
ate of Nocardiopsis displaying antimicrobial properties (along withther actinomycetes) was obtained from castings of earthworms inndia (Kumar et al. 2012). Insects such as mosquitoes and honey-ees in particular, are reported to host Nocardiopsis species as gutora. During a study on the gut flora of Culex uinquefasciatus larvaef Indian origin, an isolate of Nocardiopsis was obtained althoughenera such as Escherichia, Proteus, Aspergillus and Streptomycesere more abundant (Vasanthi and Hoti 1992). As summarized
n Table 1, there are two reports on the presence of N. alba inoneybees. It has been suggested that actinomycetes in insectsay play a role in defensive antagonism (Kaltenpoth 2009). N.
lba strain IM1-3 associated with honeybee hives in Thailand pro-uced antibiotics against the pathogens Paenibacillus larvae andelisococcus plutonius, causative agents of American and European
oulbrood diseases, respectively (Promnuan et al. 2009). Duringnother study conducted in USA, it was observed that a strain of. alba BE74 effectively controlled the growth of a Bacillus mar-
sflavi isolate also obtained from honeybee gut during the sametudy (Patil et al. 2010). A representative gene (phzD) involved inhe biosynthesis of phenazines could also be amplified from theulture. Such phenazines in nature, are significant as (i) respira-
ory electron acceptors (allowing the bacterium to survive undernaerobic environments prevalent in the gut) and (ii) as antimicro-ial agents that inhibit the growth of competing bacteria (Wangt al. 2009; Gao et al. 2012).with animals and their futuristic use for addressing societal challenges.
As depicted in Fig. 3, there are thirty nine validly named species(and some sub-species) in this genus including the new species thathas been recently isolated (Bouras et al. 2015). Out of these, twoare reported to be opportunistic human pathogens. N. dassonvilleihas been described since long to be the main pathogenic speciesassociated with human beings from patients belonging to differentgeographical locations (USA, Poland, France, Spain and India). It hasbeen (i) implicated in causing Actinomycosis or mycetoma wheremultiple nodules and draining sinuses were noticed (Sindhuphaket al. 1985), (ii) linked to a rare suppurative pulmonary infection(Mordarska et al. 1998), (iii) isolated from the blood of a cholan-gitis patient wherein an infection of the bile duct was observed(Beau et al. 1999), (iv) related to a sporotrichoid cutaneous infectionwhere skin lesions progressed along the dermal and subcutaneouslymphatics (González-López et al. 2011) and (v) associated withnasal vestibular abscess (Rudramurthy et al. 2012). Apart fromN. dassonvillei, N. synnemataformans (isolated from a patient inTurkey) is the only other pathogenic species (Yassin et al. 1997).Phylogenetic analysis shows that N. dassonvillei (and its subspecies,dassonvillei, alborubida, antarctica) and N. synnemataformans areclosely related (Fig. 3). With the exception of these two speciesthat are rarely infectious other members of this genus are non-pathogenic.
2.2. The role of Nocardiopsis species associated with animalforms in addressing future societal challenges
From the biotechnological point of view, sponges have beenexploited for the production of a variety of bioactive metabolites(Thomas et al. 2010). It is often hypothesized that these compoundsmay be derived partially, if not completely, from the abundant
bacteria associated with them. For example, sponge-associated Vib-rio species and Pseudomonas aeruginosa are reported to producebioactive compounds (Oclarit et al. 1994; Jayatilake et al. 1996).As summarized in Table 1, Nocardiopsis species have been isolated
T. Bennur et al. / Microbiological Research 174 (2015) 33–47 37
Fig. 3. Phylogenetic relationship between the validly published Nocardiopsis species on the basis of 16S rRNA gene sequences mainly derived fromh sing NJ e. Anac
fp2pct(
aIotat
ttp://www.bacterio.net/nocardiopsis.html. Phylogenetic tree has been drawn uukes–Cantor method. The scale bar represents the number of substitutions per sitonducted in MEGA6.
rom certain sponges and some of these produce bioactive com-ounds such as lipopetptides and Nocapyrones (Gandhimathi et al.009; Schneemann et al. 2010a,b). Others have the potential toroduce them since they harbor the necessary biosynthetic genelusters (Jiang et al. 2007). Such Nocardiopsis sp. can serve as poten-ial sources of natural products and new metabolites in the futureFig. 2).
The product TTX is biotechnologically relevant as a potentialnalgesic and local anesthetic agent and is currently under phaseI clinic trials (Wu et al. 2005a). It must be noted that a variety
f bacteria including Vibrio, Pseudomonas, Alteromonas, Flavobac-erium and Micrococcus produce TTX (Simidu et al. 1987; Thuesennd Kogure 1989; Miyazawa and Noguchi 2001). The report onhe isolation of N. dassonvillei culture in puffer fish has extendedeighbor-Joining method and the evolutionary distances were computed usinglysis involved forty one nucleotide sequences and the evolutionary analyses were
the range of bacteria producing this potent toxin. Conventionally,TTX is purified from the ovaries and livers of puffer fish and theyield is limited (Yu et al. 2004). Isolation of TTX producing bacteriaand scale-up of fermentation procedures would make it possible toproduce large amounts of TTX without destroying the puffer fishpopulation.
An interesting biotechnological application of the Nocardiopsisspecies isolated from the gut of C. uinquefasciatus mosquito larvaehas been reported (Vasanthi and Hoti 1992). Incubation of purecultures of this species with mosquito larvae caused complete mor-
tality of the latter. Such results indicate the possibility of usingthis species in the biocontrol of C. uinquefasciatus that are impor-tant vectors in transmitting bancroftian filariasis, avian malaria andsome arbovirus-mediated diseases.
3 gical R
eiPwwasagdF
2
NpgietrrfpAefspscp
3m
goab(pnuTNtptNtpM3astatilf
8 T. Bennur et al. / Microbiolo
N. alba BE74 isolated from honeybee gut showed the pres-nce of a representative gene of the phz gene cluster that isnvolved in the biosynthesis of phenazines (Patil et al. 2010).henazines are biotechnologically important bioactive productsith several applications (Pierson and Pierson 2010). Moreover,ith the whole genome sequence of N. alba BE74 (ATCC BAA-2165)
nd other species being available, genetic manipulation and muta-ynthetic approaches can be used to improve the production ofnd increase range of secondary metabolites derived from thisenus. A summary of the futuristic applications of such Nocar-iopsis species associated with animal forms are summarized inig. 2.
.3. Nocardiopsis species associated with plants
There are some preliminary reports on the isolation of theocardiopsis sp. in association with plants and their rhizospheresarticularly from tropical regions. All these reports only describeeneric information and detailed identification of species is lack-ng. In one case, an isolate obtained from the yam bean Pachyrhizusrosus (from Brazil) as an endophyte was studied for the produc-ion of a thermostable �-amylase (Stamford et al. 2001). In anothereport, Nocardiopsis sp. 94N10-1 (EU196477.1) isolated from thehizosphere of the Cumin plant (from Thailand) produced anti-ungal agents that were effective against Colletotrichum, a plantathogen causing anthracnose disease (Intra et al. 2011). Whenustralian endemic trees were screened for the potential produc-rs of new drug candidates, a Nocardiopsis species (PIP 172) wasound to be associated with Pittosporum phylliraeoides (apricot)tem (Kaewkla and Franco 2013). Although the bioactive com-ounds have not been identified in the latter two reports, in nature,uch bacteria could be providing additional advantages to the asso-iated plants by producing antimicrobial compounds that keep-offlant pathogens.
. Occurrence of Nocardiopsis species in terrestrial, indoor,arine, hypersaline, cold locations and their ecological role
Nocardiopsis species like most of the other Actinomycetes areenerally associated with soils. There are several reports on therganism being incident in terrestrial locations along with otherctinomycetes. In some desert soils (Sahara in Algerian), mem-ers of the genus Nocardiopsis were found to be predominantZitouni et al. 2005; Meklat et al. 2011). Different types of soil sam-les (saline, hypersaline, desert, and alkaline) have yielded severalovel isolates. Table 2 summarizes important features and prod-cts obtained from such isolates obtained from different locations.he most obvious trait of these isolates is their requirement foraCl or tolerance toward it (Hamedi et al. 2013). As discussed later,
he presence of a set of genes permitting the synthesis and trans-ort of compatible solutes may be allowing them to survive underhese conditions. Some species such as N. alkaliphila, N. valliformis,. nikkonensis, N. ganjiahuensis, N. arvandica and the recently iden-
ified species N. algeriensis are alkalophilic in nature (Table 2). Thehylogenetic relation between these species is depicted in Fig. 3.ost of the species have an optimum temperature between 28 and
0 ◦C. One isolate obtained from compost (N. yanglingensis) wasble to grow at 45 ◦C and another one from Antarctica (N. filde-ensis) was psychrotolerant and grew at 10 ◦C. In a recent study,his genus has been implicated to be present in ice core sampleslong with other microflora (Miteva et al. 2014). Different soil
ypes (mainly from China) have yielded a vast variety of novelsolates. Iran and Egypt are other regions from where such iso-ates have been obtained. A few isolates have also been obtainedrom Algeria, Germany, Seychelles and Antarctica (Table 2). Theesearch 174 (2015) 33–47
range of alkali tolerant, thermostable and cold adapted enzymesproduced by some members of this genus (described in a latersection), maybe permitting their existence under these circum-stances.
On some occasions, Nocardiopsis species have been isolated fromindoor environments such as dust samples in schools and day carecenters, pulp used in making recycled fibers and water-damagedbuilding material (Andersson et al. 1999; Suihko et al. 2006; Suihkoet al. 2009; Schäfer et al. 2010). Indoor environments (mainly fromdifferent locations in Europe) have also yielded some novel species(Table 2). The survival of these bacteria in indoor environments hasbeen attributed to their xerotolerant nature and their ability to pro-duce different extracellular enzymes (Peltola et al. 2001a). Somesuch species have been obtained from indoor buildings whereinthe occupants were reported to be suffering from serious building-related ill-health symptoms (Peltola et al. 2001b). These isolatesproduced toxic substances that dissipated the mitochondrial mem-brane potential and depleted cells of ATP. Such compounds may beresponsible for the observed ill-health symptoms.
Marine ecosystems provide a repertoire of rare Actinomycetesthat can be potential source of drugs (Subramani and Aalbersberg2012, 2013). Several studies on the diversity of actinomycetes inmarine environments have resulted in the isolation of Nocardiop-sis species (Bredholdt et al. 2007; Becerril-Espinosa et al. 2013;Romanenko et al. 2013; Zhang et al. 2013; Meena et al. 2013). Inaddition to the species associated with different marine biologi-cal forms (Section 2.1), the above mentioned species and thosedescribed in Table 2 exist in the free-living form. Hypersalinelocales such as soda lakes and salterns are other habitats thatharbor Nocardiopsis species along with other actinomycetes. Forexample, a study on the bacterial diversity from the haloalka-line Lake Elmenteita resulted in the isolation of Streptomyces andNocardiopsis species (Mwirichia et al. 2010). Microbial analysis ofsolar salterns has also shown the presence of the members of thisgenus along with other actinomycetes (Jose and Jebakumar 2012).Some marine sediments, salterns and salt marshes located in Egypt,China, Korea and Kuwait have also yielded some novel species thatare detailed in Table 2. As expected, the marine and the hypersalineisolates were also halotolerant or halophilic in nature.
From the above sections it is evident that Nocardiopsis speciesare prevalent in a variety of terrestrial, marine and hypersalineenvironments (Table 2). Under these conditions, the members ofthis genus may be playing a degradative role in mediating thebreakdown of naturally occurring complex polymers. As detailedin a later section and Table 3, a range of enzymes aid this activity.Several isolates obtained from soil, marine forms, marine sedi-ments and mine tailings also produce different types of antibiotics.This microbial genus has the gene clusters for polyketide syn-thases (PKSs) and nonribosomal peptide synthetases (NRPSs) andmany of the natural isolates may be producing antibiotics (Zitouniet al. 2005; Meklat et al. 2011; Becerril-Espinosa et al. 2013;Romanenko et al. 2013). On several occasions, bioactive compoundshave been isolated and characterized. For example, a marine Nocar-diopsis species produced cyclic hexapeptides that displayed weakantimicrobial activity (Wu et al. 2013). In some cases, novel bioac-tive compounds have been obtained. For example, soil-derived N.trehalosei produced a novel antibiotic 3-trehalosamine; N muta-bilis produced dopsisamine and Nocardiopsis species TFS65-07isolated from marine sediments secreted a thiopeptide antibioticwith a rare aminoacetone moiety (Dolak et al. 1980; Takahashiet al. 1986; Engelhardt et al. 2010a). Other antibiotics producedby the members of this genus are elucidated in Table 4. It is
thus evident that this versatile actinomycete performs differ-ent functions in nature. The wide-ranging adaptations and otherimportant features it needs to do so, are detailed in the followingsections.
T. Bennur
et al.
/ M
icrobiological R
esearch 174
(2015) 33–47
39
Table 2Features of the novel Nocardiopsis sp. isolated from soil, indoor, marine and hypersaline environments.
Nocardiopsis sp.; site; location Optimum conditions Nomenclature details and other features Reference
SoilN. halophila DSM 44494; saline soil; Iraq NaCl: 20%; temperature: 30 ◦C halophila: salt loving Al-Tai and Ruan (1994)N. tropica DSM 44381; rhizosphere; Seychelles NaCl: 10%; pH: 7.4; temperature: 28 ◦C tropica: isolated from tropical region Evtushenko et al. (2000)N. metallicus DSM 44598; alkaline slag dump; Germany NaCl: 10%; pH: 7.0–10.5; temperature: 30 ◦C metallicus: able to mobilize metals from slag Schippers et al. (2002)N. xinjiangensis DSM 44589; saline soil; Xinjiang, China NaCl: 10%; pH: 7.2; temperature: 28 ◦C xinjiangensis: pertaining to the site of isolation (Xinjiang) Li et al. (2003)N. salina YIM 90010; hypersaline soil; Xinjiang, China NaCl: 10%; pH: 7.2; temperature: 28 ◦C salina: from salty environment Li et al. (2004)N. alkaliphila DSM 44657; desert soil; Egypt NaCl: 10%; pH: 9.5–10; temperature: 28 ◦C alkaliphila: loving alkaline environments;
produces Nocardiopyrones A and B, pyridinols 3–5, and1-acetyl-�-carboline
Hozzein et al. (2004)Wang et al. (2013)
N. gilva DSM 44841; hypersaline soil; Xinjiang, China NaCl: 5–8%; pH: 7.2; temperature: 28–30 ◦C gilva: pale yellow mycelium; produces p-terphenylderivatives, antibiotic novobiocin, cyclodipeptides
Li et al. (2006)Tian et al. (2013)
N. baichengensis DSM 44845; hypersaline soil;Xinjiang, China
NaCl: 5–8%; pH: 7.2; temperature: 37–40 ◦C baichengensis: pertaining to the site of isolation (Baicheng) Li et al. (2006)
N. rosea DSM 44842; hypersaline soil; Xinjiang, China NaCl: 5–8%; pH: 7.2; temperature: 37–40 ◦C rosea: rose colored mycelium Li et al. (2006)N. rhodophaea DSM 44843; hypersaline soil; Xinjiang,
ChinaNaCl: 5–8%; pH: 7.2; temperature: 37–40 ◦C rhodophaea: rose-brown mycelium Li et al. (2006)
N. chromatogenes DSM 44844; hypersaline soil;Xinjiang, China
NaCl: 5–8%; pH: 7.2; temperature: 37–40 ◦C chromatogenes: producing color Li et al. (2006)
N. quinghaiensis DSM 44739; saline soil; Qinghai, China NaCl: 3%; pH: 7; temperature: 28 ◦C quinghaiensis: pertaining to the site of isolation (Qinghai) Chen et al. (2008)N. arabia DSM 45083; Sand-dune soil, Egypt NaCl: 5%; pH: 7.8; temperature: 28–30 ◦C arabia: pertaining to Arabia Hozzein and Goodfellow
(2008)N. ganjiahuensis DSM 45031; Soil, Xinjiang, China NaCl: 0–5%; pH: 8.5–9.5; temperature:
28–32 ◦Cganjiahuensis: pertaining to the site of isolation (GanjiahuNatural Reserve)
Zhang et al. (2008)
N. valliformis DSM 45023; Alkali lake soil, XinjiangChina
NaCl: 0–5%; pH: 9.5–13; temperature: 28 ◦C valliformis: referring to the characteristic of myceliumarranged in a shape like a palisade
Yang et al. (2008)
N. terrae YIM 90022; Saline soil, Qaidam Basin, China NaCl: 3–5%; pH: 8.5; temperature: 30 ◦C terrae: isolated from soil; produces antimicrobialcompounds
Chen et al. (2010)Tian et al. (2014)
N. sinuspersici DSM 45277; Sandy soil, IranN. nikkonensis KCTC 19666; Compost soil, Japan
NaCl: 2.5%; pH: 7; temperature: 28 ◦CNaCl: 0–10%; pH: 6–11; temperature: 30 ◦C
sinuspersici: Isolated from Persian Gulfnikkonensis: pertaining to the site of isolation (Nikko,Japan)
Hamedi et al. (2010)Yamamura et al. (2010)
N. yanglingensis KCTC 19723; compost soil, China NaCl: 5%; pH: 7.5; temperature: 25–45 ◦C yanglingensis: pertaining to the site of isolation (Yanglingcity)
Yan et al. (2011)
N. arvandica DSM 45278; sandy soil, Iran NaCl: 0–5%; pH: 6–11; temperature: 25–30 ◦C arvandica: belonging to the Arvand River Hamedi et al. (2011)N. fildesensis DSM 45699; soil, Antarctica NaCl: 0–12%; pH: 9–11; temperature: 10–37 ◦C fildesensis: pertaining to the site of isolation (Fildes
Peninsula)Xu et al. (2014)
N. algeriensis DSM 45462; soil, Algeria NaCl:0–5%; pH: 7–10; temperature: 25–35 ◦C algeriensis: referring to Algeria (the country of isolation) Bouras et al. (2015)
Indoor environmentsN. exhalans DSM 44407; indoor air; building basement NaCl: 10%; temperature: 10–37 ◦C exhalans: related to emitting odors, fumes toxins;
xerotolerant; produces phosphatase and �-glucosidasePeltola et al. (2001a)
N. umidischolae DSM 44362; indoor air school NaCl: 7.5%; temperature: 10–37 ◦C umidischolae: related to moist school environment;xerotolerant; produces �- and �-glucosidases,phosphatase, and peptidases
Peltola et al. (2001a)
N. compostus DSM 44551; air; composting facility,Germany
NaCl: 15%; pH: 6.5–9.5; temperature:>10–50 ◦C
compostus: originating from compost Kampfer et al. (2002)
N. potens DSM 45234; household waste NaCl: 0–12%; temperature: 20–37 ◦C potens: powerful, pertaining to the metabolic activities ofthe organism
Yassin et al. (2009)
Marine sediments and salternsN. aegyptia DSM 44442; marine sediment, Egypt NaCl: 5%; temperature: 10 ◦C aegyptia: from Egypt
Degrades poly(3-hydroxybutyrate)Sabry et al. (2004)Ghanem et al. (2005)
N. flavescens CGMCC 4.5723; marine sediment, China NaCl: 0–3%; pH: 7.2–7.5; temperature: 35 ◦C flavescens: becoming golden yellow in color Fang et al. (2011)N. kunsanensis KCTC 9831; saltern, Korea NaCl: 10%; pH: 9 temperature: 37 ◦C kunsanensis: pertaining to the site of isolation (Kunsan) Chun et al. (2000)N. halotolerans DSM 44410; salt marsh soil, Kuwait NaCl: 0–10%; temperature: 28–35 ◦C halotolerans: ability to tolerate high salt concentrations Al-Zarban et al. (2002)
40 T. Bennur et al. / Microbiological Research 174 (2015) 33–47
Table 3Summary of the enzymes produced by Nocardiopsis species that may aid existence under different conditions.
Enzyme; strain; location Important features Reference
Cold-adapted enzymes�-Amylase; Nocardiopsis strain 7326; deep sediment,
AntarcticaOptimum pH: 8.0; pH stability: 5–10; optimum temperature: 35 ◦C;inactivated above 45 ◦C
Zhang and Zeng (2008)
�-Amylase; N. aegyptia; Egypt Statistical design used to optimize conditions; cell immobilizationyielded better production
Abou-Elela et al. (2009)
Thermotolerant enzymes�-Amylase; Nocardiopsis sp.; Brazil Optimum pH: 5.0; optimum temperature: 70 ◦C Stamford et al. (2001)Xylanases (X-I, X-II, X-III); N. dassonvillei subsp. alba
OPC-18; JapanOptimum pH: X-I and X-II:7.0; optimum temperature: 60 ◦C; X-III:optimum temperature: 50 ◦C
Tsujibo et al. (1990b, 1991)
Thermostable enzymes�-Amylase; Nocardiopsis sp. B2; India Optimum pH: 9.0; optimum temperature: 45 ◦C; maximum enzyme
activity with 11% (w/v) NaCl; retained 75% and 69% activity afterincubation for 1 h at 75 and 85 ◦C, respectively; effectiveimmobilization in gellan gum microspheres
Chakraborty et al. (2014)
Thermoalkalotolerant enzymesCellulase; Nocardiopsis sp. KNU; Korea Optimum pH: 5.0; optimum temperature: 40 ◦C Saratale and Oh (2011)�-1,3-glucanase; Nocardiopsis sp. F96; Japan Optimum pH: 9.0; optimum temperature: 70 ◦C; hydrolyzed insoluble
�-1,3-glucans; �-1,3–1,4-glucans preferred over �-1,3-glucans;cloning and expression; crystal structure; construction andcharacterization of chimeras
Masuda et al. (2006),Fibriansah et al. (2006,2007), Koizumi et al. (2007,2009)
Alkalotolerant enzymeCellulase; Nocardiopsis sp. SES28; Argentina Optimum pH: 8.0; optimum temperature: 40 ◦C; application in the
detergent industryWalker et al. (2006)
Alkalitolerant thermostable enzymesInulinase; Nocardiopsis sp. DN-K15; China Optimum pH: 8.0; optimum temperature: 60 ◦C; exoinulinase
(fructose as hydrolysis product)Lu et al. (2014)
Chitinases; N. prasina OPC-131; Japan Optimum temperature: ChiA and ChiB: 60; optimum pH: ChiA:7.0ChiB: 6.0; at pH 9.0, 50% of optimal activities observed
Tsujibo et al. (2003)
Alkaline proteasesAlkaline serine proteases; N. dassonvillei sub species
prasina OPC-210; JapanProtease I: Optimum pH: 10.0–12.0; optimum temperature: 70 ◦CProtease II: Optimum pH: 10.5; optimum temperature: 60 ◦C
Tsujibo et al. (1990a)
Alkaline serine proteases; N. dassonvillei NCIM 5124;India
Protease I and II Optimum pH: 10.0–11.0; optimum temperature: 60 ◦C Dixit and Pant (2000a) andRohamare et al. (2013)
Alkaline proteases; N. prasina HA-4; India Optimum pH: 7.0 and 10.0; optimum temperature: 55 ◦C Ningthoujam et al. (2009)Alkaline serine protease; N. alba OK-5; India Optimum pH: 10.0; optimum temperature: 80 ◦C Gohel and Singh (2012)Alkaline serine protease; Nocardiopsis species; Brazil Optimum pH: 10.5; optimum temperature: 50 ◦C Moreira et al. (2002)Milk-clotting protease; Nocardiopsis species; Brazil Optimum pH: 7.5; optimum temperature: 55 ◦C Cavalcanti et al. (2004)Keratinolytic protease; Nocardiopsis sp. TOA-1; Japan Optimum pH: 11–11.5; optimum temperature: 70–75 ◦C Mitsuiki et al. (2002)Keratinase and protease; Nocardiopsis species SD5;
IndiaOptimum pH: 9.0; optimum temperature: 50 ◦C; degrades feathers Saha et al. (2013)
Table 4Summary of the antibiotics produced by Nocardiopsis species.
Nocardiopsis sp. Location of isolation Chemical nature ofantibiotic
Target cultures Reference
N. trehalosei sp.NRRL12026
Soil, USA 3-Trehalosamine Staphylococcus aureus, Bacillus subtilis, Streptococcuspneumonia
Dolak et al. (1980)
N. mutabilis subsp.cryophilis TS-1980
Cold soil, Japan Polyamine dopsisamine S. aureus, Micrococcus luteus, B. subtilis, Escherichia coli,Proteus vulgaris, Klebsiella pneumoniae, Mycobaceriumsmegmatis, Candida albicans, Saccharomyces cerevisiae,Pyricularia oryzae, Xanthomonas oryzae, Aspergillus niger,Trichophyton mentagrophytes
Takahashi et al.(1986)
N. dassonvillei MAD08 Marine sponge Dendrillanigra, India
87 kDa protein C. albicans Selvin et al. (2009)
Nocardiopsis sp.TFS65-07
Marine sediment, Norway Thiopeptide TP-1161 M. luteus, C. albicans Engelhardt et al.(2010a)
N. dassonvillei HR10-5 Marine sediment, China �-Pyrones, nocapyronesE-G
B. subtilis Fu et al. (2011)
Nocardiopsis sp. YIMDT266
Tin mineTailings, China
Griseusins (F and G) S. aureus, M. luteus, B. subtilis Ding et al. (2012)
N. alkaliphila DSM44657
Desert soilEgypt
Nocardiopyrone ANocardiopyrone B
Pseudomonas aeruginosa, Enterobacter aerogenes, E. coliC. albicans, S. aureus
Wang et al. (2013)
Nocardiopsis sp. 236 Marine sediments, China 6-Phenazinediol and6-methoxy-1-phenazinol
M. smegmatis Lu et al. (2013)
N. gilva DSM 44841 Saline soil, China p-Terphenyl derivativesNovobiocin
Fusarium avenaceum, Fusarium graminearum, FusariumculmorumC. albicans, B. subtilis
Tian et al. (2013)
N. terrae YIM 90022 Saline soil, China Quinoline alkaloid,N-acetyl-anthranilic acid
S. aureus, B. subtilis, E. coli, Pyricularia oryzae Tian et al. (2014)

gical R
4
dNmotpriAmltdtfwtmb
dfdttetafsTa
4
neaeablplotfIabaawttmom
iwf
T. Bennur et al. / Microbiolo
. Adaptations and survival modes
The life cycle of Streptomyces species has been discussed inetail (Chater 2006). In similar manner, members of the genusocardiopsis also appear to follow a complex life cycle that cul-inates in the formation of sporulating aerial mycelia; the release
f extracellular enzymes for solubilization of natural polymers andhe production of antibiotics. In general, spores germinate in theresence of soluble nutrients and early mycelial growth occursapidly. When soluble nutrients are exhausted, these microorgan-sms secrete enzymes for degrading insoluble available polymers.
concomitant release of antibiotics disallows the growth of othericroorganisms thereby ensuring an exclusive use of the solubi-
ized nutrients by the mycelium. It has also been suggested thathis could be an ambush strategy whereby the would-be invadersie and provide additional nutrition (Shi and Zusman 1993). Whenhe readily assimilable nutrients get exhausted, aerial mycelia areormed. Aerial growth is partially parasitic on the mycelium fromhich it emerges, some of which breaks down to provide nutrition
o the aerial hyphae (Wildermuth 1970). Spores borne on the aerialycelium are resistant to harsh conditions and are disseminated
y different means.From the earlier sections in this review, it is evident that Nocar-
iopsis species have been isolated from varied habitats rangingrom soil and marine samples, to extreme environments such aseserts, alkaline or hypersaline soils. Their capacity to inhabit mul-iple niches and interact with other organisms appears to be linkedo the abovementioned life cycle that involves the production ofnzymes and other secondary metabolites. Literature survey showshat Nocardiopsis species produce a vast array of compounds suchs (i) extracellular enzymes (ii) antimicrobial agents and biosur-actants (iii) compatible solutes (particularly in case of halophilicpecies) that may aid their existence under different conditions.heir proteomic and genomic features are also the basis of theirdaptability to the diverse milieu where they exist.
.1. Enzymes produced by Nocardiopsis species
Nocardiopsis species are a potential source of diverse andovel extracellular enzymes (Bennur et al. 2014). Some of thesenzymes are able to withstand harsh conditions that are usu-lly thought to be destructive to proteins and are referred to asxtremozymes. Such extreme enzymes have several importantpplications (Elleuche et al. 2014). The major enzymes producedy different Nocardiopsis species isolated from a variety of eco-
ogical niches are summarized in Table 3. Most of the isolatesroducing these enzymes have not been identified to the species
evel and are named in an informal manner. Among the identifiednes, N. dassonvillei, N. alba, N. prasina and N. aegytpia are impor-ant. Most of the isolates producing enzymes have been obtainedrom Japan, India, Brazil, Argentina, China, Korea and Antarctica.n general, cold-adapted, thermotolerant, thermoalklotolerant andlkalotolerant enzymes acting on different naturally occurring car-ohydrates such as cellulose, chitin, �-1,3-glucans, inulin, starch,nd xylan are produced. Alkaline proteases, milk clotting enzymesnd keratinases are also secreted by Nocardiopsis species and theseould help in utilizing proteinaceous matter in nature. The utiliza-
ion of keratinolytic substrates may offer an additional advantageo these actinomycetes since keratin is amply found in the environ-
ent. This vast range of extracellular enzymes equips the membersf this genus to degrade complex vegetal, animal and fungal poly-ers and acquire nutrition.
On three occasions, the structural features that confer stabil-ty to proteases have been elucidated. In two cases, the enzymesere found to be kinetically stable proteins. They displayed high
ree energy kinetic barriers separating the folded and unfolded
esearch 174 (2015) 33–47 41
states and preferably existed in the folded state under harsh condi-tions. The kinetically stable protease (NAPase) derived from N. albashowed multiple salt-bridges on the outer regions that conferredon the protein the observed acid stability (Kelch et al. 2007). Thealkaline protease NprotI derived from N. dassonvillei NCIM 5124showed the presence of a distinctive polyproline II helix (PPII) thatmade the protease kinetically stable thereby, resisting denaturationand proteolytic digestion (Rohamare et al. 2013). The stability ofthe serine protease derived from N. prasina was attributed to twoamino acid residues (Asn47 and Pro124) situated in its loop regions(Farrell et al. 2012).
In some instances, genomic and metaexoproteomic approacheshave been used to demonstrate the relevance of Nocardiopsis-derived enzymes. For example, the putative cellulose degradingenzyme sequences in actinobacterial genomes have been identi-fied (Anderson et al. 2012). Among the microorganisms evaluated,N. dassonvillei DSM 43111 produced cellulases when provided withappropriate substrates and the genome also showed sequences forsix predicted cellulose degrading enzymes. In another report, therole of the chitinases associated with Nocardiopsis species havebeen recently demonstrated by employing the metaproteomicsapproach (Johnson-Rollings et al. 2014). It must be noted that themetaexoproteome is the subset of proteins found outside the celland studies related to this allow one to assess cellular functions andinteractions within an environment. Nocardiopsis-like chitinaseswere found to be solely responsible for the chitinolytic activity inthe soil that was being investigated by the authors. The findingshave thus justified the predominance and adaptability of Nocar-diopsis species in soil environments. On account of the frequentoccurrence of Nocardiopsis species in different types of soils andwater bodies and their ability to produce a wide range of hydrolyticenzymes, one can assume that this bacterial species may be playingan important role in the carbon cycle.
4.2. Production of antimicrobial agents and biosurfactants
It is a well-established fact that certain bacteria (mainly Acti-nomycetes) produce antibiotics that mediate the destruction orinhibition of other microorganisms (Waksman et al. 2010). Most ofthe studies related to isolation of Actinomycete populations fromnature (including some references covered in this review, Table 1)indicate the predominance of the members of the genus Strepto-myces. This prevalence is attributed to the vast range of antibioticsproduced by the members of this genus. It must be noted that about80% of the antibiotics today are sourced from the genus Strepto-myces (de Lima Procópio et al. 2012). Secondary metabolites usuallyfunction like weapons, serve as molecular signals in cellular com-munication and interfere with gene transfer phenomena (Chater2006; Linares et al. 2006; Davies 2006). These metabolites thus playa significant role in structuring soil microbial communities (Tarkkaet al. 2008).
The plethora of Nocardiopsis sp. obtained from different ecologi-cal niches (soils, marine forms, marine sediments and mine tailings)produce a variety of antibiotics (Table 4). These include quino-line alkaloids, p-terphenyl derivatives, novobiocin, phenazinediol,cyclic tetrapeptides, nocardiopyrones, thiopepetides and proteins.Most of these antibiotics share structural details with com-pounds produced by other microorganisms and may not be novel.However, as discussed earlier, three antibiotics (3-trehalosmaine,dopsisamine and thiopeptide) were found to have novel structures(Dolak et al. 1980; Takahashi et al. 1986; Engelhardt et al. 2010a).Nocardiopsis species producing antibiotics may benefit as these
compounds would help in eliminating competing microorganisms.The inhibition of different Gram positive and Gram negative bacte-ria as well as fungal isolates by the range of antibiotics producedby different strains under laboratory conditions is summarized
4 gical R
idNafJeiaitdaedsanct
wgiarewassoob2tfttto(shbs
4
bom2wlupseiNnb1
2 T. Bennur et al. / Microbiolo
n Table 4. Important species producing antibiotics include N.assonvillei, N. trehalosei, N. mutabilis, N. alkaliphila, N. gilva and. terrae. Some unidentified species also produce antimicrobialgents. The antibiotic producing isolates have mainly been isolatedrom China. Other locations include USA, Egypt, Norway, India andapan. This highlights the possibility of these antibiotics in beingffective under natural conditions as well. It must be noted thatn nature, microorganisms face stiff competition for food, waters well as other resources and the production of antibiotics is anmportant survival strategy. The presence of antibiotics enableshem to ward-off competing species and helps them to exist underifferent conditions. Moreover, it also allows the nutrients madevailable (by secretion of extracellular enzymes) to be used morexclusively by the producer (Chater 2006). Like the antibioticserived from Streptomyces species, they may also be involved in cellignaling and communication, gene transfer related phenomenand in the development of soil microbial communities. The sig-ificance of different gene clusters mediating the synthesis of suchompounds has been discussed in a later section that deals withhe genomic features of this microorganism.
Microbial biosurfactants are a class of amphiphilic compoundsith varied chemical structures and surface properties. They affect
rowth of the microorganisms producing them in varied ways andn general, aid their growth in different ecological niches (Ronnd Rosenberg 2001). Some such compounds display antibacte-ial or antifungal activities (Singh and Cameotra 2004; Rodriguest al. 2006). Others enhance the growth of bacteria on hydrophobicater-insoluble substrates by increasing their bioavailability and
pparent solubility (Cameotra and Bollag 2003). Like other bacteriaynthesizing surfactants, some members of the genus Nocardiop-is also produce such compounds. For example, there are reportsn the production of a lipopeptide by a sponge-associated strainf N. alba MSA10 and biosurfactant (chemically not characterized)y Nocardiopsis strain B4 (Gandhimathi et al. 2009; Khopade et al.012). In addition, some members (N. dassonvillei and N. lucen-ensis MSA04) make glycolipids that have been used as markersor identification of strains and for bioremedial purposes, respec-ively (Mordarska et al. 1983; Seghal et al. 2010). Although none ofhese biosurfactants showed antimicrobial or antibiofilm proper-ies, one of them displayed good emulsification activity and anotherne could be used in the bioremediation of marine environmentsKhopade et al. 2012; Seghal et al. 2010). It must be noted thatome species such as N. dassonvillei NCIM 5124 effectively degradeydrocarbons (Dixit and Pant 2000b) and the presence of suchiosurfactants may aid the effective utilization of this class of sub-trates.
.3. Compatible solutes in halophilic Nocardiopsis species
Several species isolated from sea water, saline soils, salt lakes,rines and alkaline saline habitats tolerate high concentrationsf NaCl (Table 2). Nocardiopsis species are credited to be theost abundant halophilic actinomycete species (Hamedi et al.
013). In general, halophiles and halotolerant bacteria cope upith the challenge of environmental osmotic stress by regu-
ating the intracellular concentrations of low molecular weightncharged or zwitterionic organic molecules referred to as com-atible solutes. Such solutes are stress-protecting agents thattabilize biomacromolecules against harsh environments (Pastort al. 2010). There are a few reports on the accumulation of compat-ble solutes by some halophilic species of Nocardiopsis. For example,
ocardiopsis sp. A5-1 amassed three types of compatible solutesamely, ectoine (1,4,5,6-tetrahydro-2-methyl-4-pyrimidine car-oxylic acid), hydroxyectoine and trehalose (Severin et al.992). N. halophila showed the presence of hydroxyectoine andesearch 174 (2015) 33–47
alpha-glutamate and N. lucentensis accumulated ectoine as com-patible solutes (Yassin et al. 1993; DasSarma and Arora 2002).
N. dassonvillei is another species that is present in differentsalt containing environments (Dixit and Pant 2000b; Wu et al.2005a; Jiang et al. 2007; Selvin et al. 2009; Fu et al. 2011).Although there are no reports detailing the production of com-patible solutes from this species, from the available genomedata, we could identify the predicted coding sequences (CDS)that are involved in the biosynthesis and transport of ectoin,hydroxyectoine and trehalose. The predicted coding sequencesinvolved in the synthesis and transport of ectoine and hydrox-yectoine are detailed in Fig. 4. In addition, some predictedsequences coding for enzymes involved in the biosynthesis oftrehalose from maltooligosaccharides [maltooligosyl trehalosesynthase (Ndas 2269); malto-oligosyltrehalose trehalohydrolase(Ndas 2267); glycogen debranching enzyme (Ndas 2820)] andmaltose [trehalose synthase (Ndas 0248)] have also been demon-strated in the N. dassonvillei genome. This genetic makeup wouldenable such species to synthesize compatible solutes when exposedto saline environments thereby permitting them to cope up withosmotic stress encountered under such conditions. These compati-ble solutes would also be important as “general stress metabolites”permitting the existence of Nocardiopsis sp. under hot (desert soils,salterns), dry (indoor environments) and cold (frozen soils) condi-tions where they are often present (Table 2).
The ecological significance of such compatible solutes in generalhas been discussed earlier (Welsh 2000). These solutes in additionto conferring osmotic stability, serve as intracellular reserves of car-bon, nitrogen and energy. They are also regarded as common stressmetabolites that protect cells against other harsh environmentalconditions inflicted by heat, desiccation and freezing. The release ofthese compatible solutes in the environment has important ecolog-ical implications. They act as carbon, nitrogen and energy sourcesfor other microflora. Although there are several recent reports onhalophilic species of Nocardiopsis being isolated from salt contain-ing environments, studies on the production of compatible solutesis something that needs to be studied in greater detail in the future.This will allow the exploitation of these isolates for large scaleproduction of these biotechnologically relevant products.
5. Genomic and proteomic features related to survival
From Sections 2 and 3, it is evident that Nocardiopsis species(i) are associated with different biological systems and (ii) existas free-living forms in a variety of soils, marine systems andhypersaline locations. As stated earlier, there are thirty nineidentified species of Nocardiopsis. 16S rRNA gene sequences ofthese species have been used to derive the phylogenetic rela-tionship between them as detailed in Fig. 3. Although there areforty five species generally reported to be present in this genus,(http://www.bacterio.net/nocardiopsis.html; Parte 2014) litera-ture survey shows that six of them have been reclassified. N.longispora, N. mutabilis, N. coeruleofusca and N. syringae have beentransferred to the genus Saccharothrix; N. flava has been includedin the genus Lechevalieria and N. africana now belongs to thegenus Microtetraspora (Kroppenstedt and Evtushenko, 2006). Thereare forty one entries in Fig. 3 including the three subspecies ofN. dassonvillei (subsp. dassonvillei, alborubida and antarctica). Therelatedness between these species is evident from this tree. Asstated earlier, the pathogenic forms are clustered phylogenetically.In addition, novel species isolated from a particular region are found
to be closely related. For example, hypersaline soil from the Xin-jiang region in China has yielded five species N. gilva, N. rosea, N.rhodophaea, N. chromatogenes and N. baichengensis that are closelyclustered in the phylogenetic tree (Fig. 3).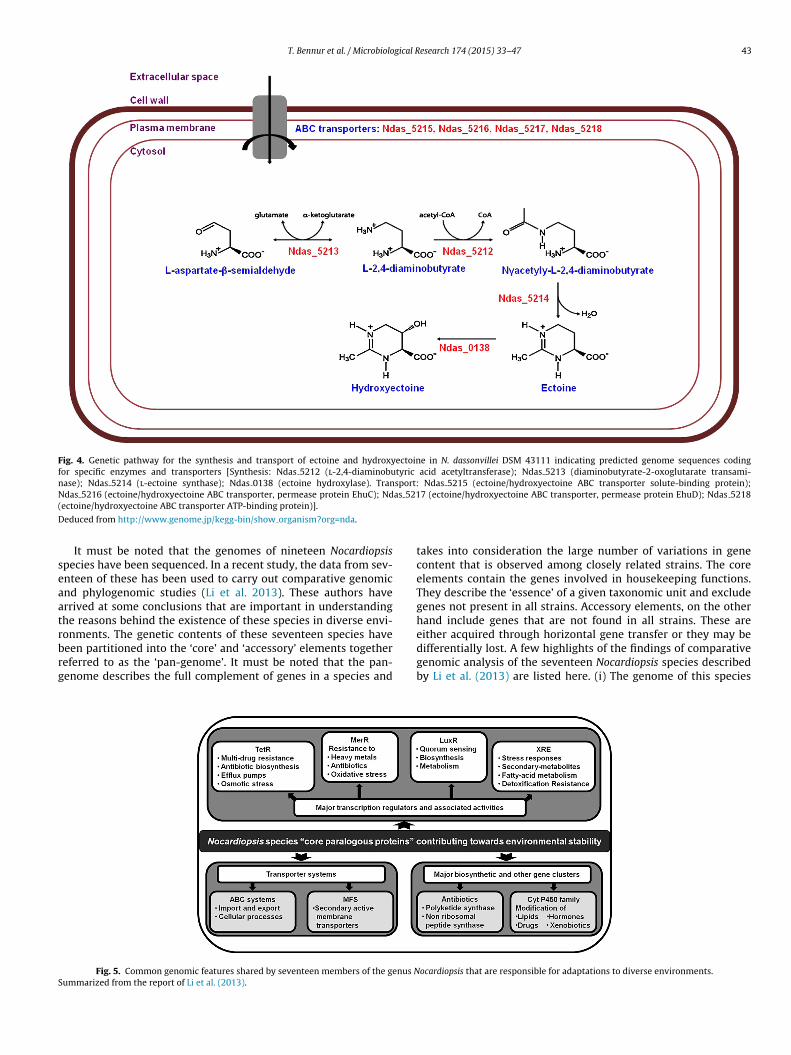
T. Bennur et al. / Microbiological Research 174 (2015) 33–47 43
Fig. 4. Genetic pathway for the synthesis and transport of ectoine and hydroxyectoine in N. dassonvillei DSM 43111 indicating predicted genome sequences codingfor specific enzymes and transporters [Synthesis: Ndas 5212 (l-2,4-diaminobutyric acid acetyltransferase); Ndas 5213 (diaminobutyrate-2-oxoglutarate transami-nase); Ndas 5214 (l-ectoine synthase); Ndas 0138 (ectoine hydroxylase). Transport: Ndas 5215 (ectoine/hydroxyectoine ABC transporter solute-binding protein);N as 521(
D
seaatrbrg
S
das 5216 (ectoine/hydroxyectoine ABC transporter, permease protein EhuC); Ndectoine/hydroxyectoine ABC transporter ATP-binding protein)].
educed from http://www.genome.jp/kegg-bin/show organism?org=nda.
It must be noted that the genomes of nineteen Nocardiopsispecies have been sequenced. In a recent study, the data from sev-nteen of these has been used to carry out comparative genomicnd phylogenomic studies (Li et al. 2013). These authors haverrived at some conclusions that are important in understandinghe reasons behind the existence of these species in diverse envi-onments. The genetic contents of these seventeen species have
een partitioned into the ‘core’ and ‘accessory’ elements togethereferred to as the ‘pan-genome’. It must be noted that the pan-enome describes the full complement of genes in a species andFig. 5. Common genomic features shared by seventeen members of the genus Nummarized from the report of Li et al. (2013).
7 (ectoine/hydroxyectoine ABC transporter, permease protein EhuD); Ndas 5218
takes into consideration the large number of variations in genecontent that is observed among closely related strains. The coreelements contain the genes involved in housekeeping functions.They describe the ‘essence’ of a given taxonomic unit and excludegenes not present in all strains. Accessory elements, on the otherhand include genes that are not found in all strains. These areeither acquired through horizontal gene transfer or they may be
differentially lost. A few highlights of the findings of comparativegenomic analysis of the seventeen Nocardiopsis species describedby Li et al. (2013) are listed here. (i) The genome of this speciesocardiopsis that are responsible for adaptations to diverse environments.

4 gical R
c2wicTisctwwtapotsibcuifmaadih
fadotstrghptbaa
amhNpt2itesseaaaIts
4 T. Bennur et al. / Microbiolo
ontained, on an average, 5864 genes. (ii) The core genome included517 genes. (iii) The pan-genome displayed 22,143 genes. (iv) Thereere 99,684 protein coding sequences with a majority of them hav-
ng homologous counterparts. (v) Some proteins were unique andould not be matched to any homologs in the pan-genome. (vi)he highest percentage of unique proteins (25.1%) was observedn the proteome of N. gilva and the lowest (3.4%) in that of N. das-onvillei. (vii) On an average, 13.9% of the Nocardiopsis proteomesomprised unique proteins. The authors have also suggested thathe inherent features of Nocardiopsis species; a dynamic genomeith core and accessory proteins could be responsible for theiride distribution and their adaptability to diverse environmen-
al conditions. In a manner similar to most other bacteria thatre encountered in related environments, some “core” paralogousroteins (present in all the species) were significant in conferringnto the genus its remarkable adaptability. Functional classifica-ion of these homologous clusters (that are common to all thepecies) has aided in identifying some protein families relevantn this regard as summarized in Fig. 5. Many of these may alsoe significant in other microorganisms that exist under similaronditions. TetR, XRE, LuxR, MerR and some other transcription reg-lators were particularly significant. Other important gene families
n Nocardiopsis code for ABC transporters, major facilitator super-amily (MFS) group of secondary active membrane transporters,
embers of the cytochrome P450 superfamily. All these featuresllow bacteria to communicate effectively, maintain viability, anddapt to the external environment. The key genomic characteristicsisplayed by Nocardiopsis species are quite similar to those present
n other microorganisms that are encountered in related ecologicalabitats.
Apart from the aforementioned summary of the comparativeeatures of the seventeen species described by Li et al. (2013), therere certain other characteristics related to the adaptability of Nocar-iopsis species as discussed in Section 4. In general, the membersf this genus are able to prevail under different conditions becausehey (i) secrete enzymes (ii) produce antibiotics and (iii) synthe-ize compatible solutes. The relevant genetic factors involved inhe synthesis of different enzymes have already been covered in aecent review (Bennur et al. 2014) and are not detailed here. Theenetic elements involved in the synthesis of compatible solutesave been described in Section 4.3 and Fig. 4. Considered in theresent section, are some specific features regarding the gene clus-ers involved in the biosynthesis of antibiotics, a topic that has noteen dealt with in earlier sections. In particular, such clusters medi-ting synthesis of polyketides, phenazines and thiopeptide type ofntibiotics have been identified in different species.
Nocardiopsis species produce some polyketide structures thatre antimicrobial in nature. The synthesis of such molecules isediated via a variety of polyketide synthetase (PKS) gene clusters
arbored in their genomes. The draft genome sequence of a marineocardiopsis sp. (TP-A0876) that was recently published shows theresence of two type-I PKS, one type-II PKS gene clusters in additiono one hybrid PKS/NRPS and one NRPS gene clusters (Komaki et al.014). Type-I PKSs are multifunctional enzymes that are structured
nto modules (Olano et al. 2010). Each module executes a set of dis-inct activities eventually catalyzing one cycle of polyketide chainlongation. Methyltransferases and oxidoreductases modify thesekeletons to form the final products. The role of PKSs in the synthe-is of Nocapyrones in N. alba strain CR167 has been described (Lint al. 2013). The “ncpA-D” gene cluster via their products (ncpD:cyl transferase; ncpC: complex containing ˇ-keto acyl synthase,cyl transferase and acyl carrier protein; ncpB: methyltransferase
nd ncpA: oxidoreductase) mediated the synthesis of Nocapyrones.ncidentally, this “ncpA-D” gene cluster in N. alba CR167 was foundo be homologous to a type-I PKS gene cluster in another marinetrain Nocardiopsis sp. TP-A0876 (Komaki et al. 2014).esearch 174 (2015) 33–47
In general, “phz” gene clusters mediate the synthesis ofphenazines (Pierson and Pierson 2010). As discussed earlier, somephenazine derivatives displaying antimicrobial activity have beenobtained from Nocardiopsis species and the existence of the pheDgene has been demonstrated in a strain of N. alba obtainedfrom honey bees (Patil et al. 2010). The genome sequence ofN. dassonvillei DSM 43111, shows entries related to phenazinebiosynthesis as follows: Ndas 5028: phzA/B (phenazine biosynthe-sis proteins); Ndas 5029: phzC (3-deoxy-7-phosphoheptulonatesynthase); Ndas 5030: phzD (isochorismatase); Ndas 5031: phzE(2-amino-4-deoxychorismate synthase) and Ndas 5032: phzF(trans-2,3-dihydro-3-hydroxyanthranilate isomerase). Differentphenazine derivatives are generated by the expression of thesegenes.
The core structures of thiopeptides (including the novelthiopeptide TP-1161 derived from Nocardiopsis sp. TFS65-07) arechromosomally encoded precursor peptides (Engelhardt et al.2010a). These skeletons are further modified by a series of posttranslational enzymatic reactions into the final macrocyclic struc-ture. In Nocardiopsis TFS65-07, TpaA was found to be the precursorpeptide. The 147-bp-long open reading frame mediating its synthe-sis was also identified (Engelhardt et al. 2010b). The genes encodingthe modification enzymes (cyclodehydratases, dehydratases, dehy-drogenases, peptidases, methyl transferases) and those necessaryfor conferring resistance to the thiopeptide were also documented.Such gene clusters present in Nocardiopsis species mediate the syn-thesis of an array of natural products. Moreover, identification ofthese clusters would allow subsequent genetic manipulation andincreased yields of different bioactive compounds.
The genetic elements involved in the synthesis of differentenzymes, compatible solutes and antibiotics thus permit membersof the genus Nocardiopsis to exist under different conditions wherethey are prevalent.
6. Conclusion
The current review analyzes relevant details related to the inci-dence, ecological roles and adaptations of Nocardiopsis species andhighlights some key messages. One important fact that needs to benoted is that on several occasions, the isolates of Nocardiopsis havenot been identified to the species level and bear informal labels(e.g. HB383, EWC9, S1; Table 1). Such unidentified species cannot beconsidered for phylogenetic analysis. With respect to the incidenceof Nocardiopsis sp. in nature, some general observations are as fol-lows. There are reports on the association of the members of thisgenus with different living forms. Among the identified species, N.dassonvillei and N. alba are the major species associated with marineforms (sponges, corals, anemones, thick shelled molluscs, pufferfish) and terrestrial forms such as insects and mammals (Table 1).Under these conditions, they are either (i) endo or casual symbionts(most marine forms) (ii) gut flora (in terrestrial forms exhibitingdefensive antagonism or producing enzymes) or (iii) opportunis-tic pathogens (human beings). The Nocardiopsis isolates associatedwith plants (as endophytes or surface flora) and rhizospheres havenot been identified to the species level thus making it difficult todraw any conclusions regarding their phylogentic relationships.
In addition to their associations with different biological forms,several novel Nocardiopsis species exist as free-living entities in avariety of soils and other saline or hypersaline habitats. A major-ity of the soil isolates have been obtained from different regionsin China (Table 2). Other geographical locales that have yielded
novel species include Iran, Egypt, Algeria, Seychelles Germany andAntarctica. Some indoor places in Europe and hypersaline locationsin Egypt, China, Korea and Kuwait have also been the sources of newspecies. Most of the isolates listed in Table 2 have a common feature,
gical R
trtvfmob
hlvetmftTostinmdctds
C
A
ff
R
A
A
A
A
A
B
B
B
B
B
C
T. Bennur et al. / Microbiolo
heir requirement for or tolerance toward NaCl. Their phylogeneticelationships are depicted in (Fig 3). Another obvious conclusionhat can be drawn from literature is that although there are aast variety of halotolerant and halophilic species, there are veryew reports on the characterization of compatible solutes from the
embers of this genus. In the future, with strain selection, processptimization and genetic manipulation, Nocardiopsis species coulde a major source of these biotechnologically important products.
The ecological diversity and adaptability of Nocardiopsis speciesas been the focus of our discussion in this review. In the free-
iving form, the members of this genus appear to produce a wideariety of unusual and robust enzymes that are functional underxtreme conditions such as alkaline pH, low and high tempera-ures and increased salt contents. Under natural conditions they
ay be aiding the degradation of complex substrates. From theuturistic point of view, Nocardiopsis species could be an impor-ant source of such biotechnologically significant extremozymes.he members of this genus are also equipped with a broad rangef bioactive and antimicrobial compounds that confer on them aelective advantage over their competitors. An understanding onhe gene clusters involved in their biosynthesis would allow one toncrease their production by following genetic manipulation tech-iques and their diversity by using mutasynthetic approaches. Itust be noted that to date, some products derived from Nocar-
iopsis species (e.g. K-252a, protein kinase and NGF inhibitors) areommercially available. With further understanding on the biosyn-hetic capabilities of the several novel species, the range of productserived from the members of this genus and the field of ‘Nocardiop-is Biotechnology’ is projected to grow in the coming years.
onflict of interest
The authors declare that they have no conflict of interest.
cknowledgements
The authors at IBB thank University Grants Commission forunding under UPE Phase II and Savitribai Phule Pune Universityor funding. TB thanks CSIR, India for Junior Research Fellowship.
eferences
bou-Elela GM, El-Sersy NA, Wefky SH. Statistical optimization of cold adapted �-amylase production by free and immobilized cells of Nocardiopsis aegyptia. J ApplSci Res 2009;5:286–92.
l-Tai AM, Ruan JS. Nocardiopsis halophila sp. nov., a new halophilic actinomyceteisolated from soil. Int J Syst Bacteriol 1994;44:474–8.
l-Zarban SS, Abbas I, Al-Musallam AA, Steiner U, Stackebrandt E, Kroppenstedt RM.Nocardiopsis halotolerans sp. nov., isolated from salt marsh soil in Kuwait. Int JSyst Evol Microbiol 2002;52:525–9.
nderson I, Abt B, Lykidis A, Klenk HP, Kyrpides N, Ivanova N. Genomics of aerobiccellulose utilization systems in actinobacteria. PLoS ONE 2012;7(6):e39331.
ndersson AM, Weiss N, Rainey F, Salkinoja-Salonen MS. Dust-borne bacteriain animal sheds, schools and children’s day care centres. J Appl Microbiol1999;86:622–34.
eau F, Bollet C, Coton T, Garnotel E, Drancourt M. Molecular identification of aNocardiopsis dassonvillei blood isolate. Clin Microbiol 1999;37:3366–8.
ecerril-Espinosa A, Freel KC, Jensen PR, Soria-Mercado IE. Marine actinobacte-ria from the Gulf of California: diversity, abundance and secondary metabolitebiosynthetic potential. Antonie Van Leeuwenhoek 2013;103:809–19.
ennur T, Kumar AR, Zinjarde S, Javdekar V. Nocardiopsis species as potentialsources of diverse and novel extracellular enzymes. Appl Microbiol Biotechnol2014;98:9173–85.
ouras N, Meklat A, Zitouni A, Mathieu F, Schumann P, Spröer C, et al. Nocardiopsisalgeriensis sp. nov., an alkalitolerant actinomycete isolated from Saharan soil.Antonie Van Leeuwenhoek 2015;107:313–20.
redholdt H, Galatenko OA, Engelhardt K, Fjaervik E, Terekhova LP, Zotchev SB.
Rare actinomycete bacteria from the shallow water sediments of the Trond-heim fjord Norway: isolation, diversity and biological activity. Environ Microbiol2007;9:2756–64.ameotra SS, Bollag JM. Biosurfactant-enhanced bioremediation of polycyclic aro-matic hydrocarbons. Crit Rev Environ Sci Technol 2003;33:111–26.
esearch 174 (2015) 33–47 45
Cavalcanti MTH, Teixeira MFS, Lima Filho JL, Porto ALF. Partial purification ofnew milk-clotting enzyme produced by Nocardiopsis sp. Bioresour Technol2004;93:29–35.
Chakraborty S, Jana S, Gandhi A, Sen KK, Zhiang W, Kokare C. Gellan gum micro-spheres containing a novel alpha-amylase from marine Nocardiopsis sp. strainB2 for immobilization. Int J Biol Macromol 2014;70:292–9.
Chater KF. Streptomyces inside-out: a new perspective on the bacteria that provideus with antibiotics. Phil Trans Roy Soc London B Biol Sci 2006;361:761–8.
Chen YG, Cui XL, Kroppenstedt RM, Stackebrandt E, Wen ML, Xu LH, et al. Nocardiopsisquinghaiensis sp. nov., isolated from saline soil in China. Int J Syst Evol Microbiol2008;58:699–705.
Chen YG, Wang YX, Zhang YQ, Tang SK, Liu ZX, Xiao HD, et al. Nocardiopsis litoralissp. nov., a halophilic marine actinomycete isolated from a sea anemone. Int JSyst Evol Microbiol 2009;59:2708–13.
Chen YG, Zhang YQ, Tang SK, Liu ZX, Xu LH, Zhang LX, et al. Nocardiopsis terrae sp. nov.,a halophilic actinomycete isolated from saline soil. Antonie Van Leeuwenhoek2010;98:31–8.
Chun J, Bae KS, Moon EY, Jung SO, Lee HK, Kim SJ. Nocardiopsis kunsanensis sp. nov.,a moderately halophilic actinomycete isolated from a saltern. Int J Syst EvolMicrobiol 2000;50:1909–13.
Cook AE, Meyers PR. Rapid identification of filamentous actinomycetes to the genuslevel using genus-specific 16S rRNA gene restriction fragment patterns. Int J SystEvol Microbiol 2003;53:1907–15.
DasSarma S, Arora P. Halophiles Encyclopedia of Life Sciences, vol. 8. London: NaturePublishing Group; 2002. p. 458–66.
Davies J. Are antibiotics naturally antibiotics? J Ind Microbiol Biotechnol2006;33:496–9.
de Lima Procópio RE, da Silva IR, Martins MK, de Azevedo JL, de Araújo JM. Antibioticsproduced by Streptomyces. Braz J Infect Dis 2012;16:466–71.
Ding ZG, Li MG, Zhao JY, Ren J, Huang R, Xie MJ, et al. Naphthospironone A: anunprecedented and highly functionalized polycyclic metabolite from an alkalinemine waste extremophile. Chem Eur J 2010;16:3902–5.
Ding ZG, Zhao JY, Li MG, Huang R, Li QM, Cui XL, et al. Griseusins F and G, spiro-naphthoquinones from a tin mine tailings-derived alkalophilic Nocardiopsisspecies. J Nat Prod 2012;75:1994–8.
Dixit VS, Pant A. Comparative characterization of two serine endopeptidases fromNocardiopsis sp. NCIM 5124. Biochim Biophys Acta 2000a;1523:261–8.
Dixit VS, Pant A. Hydrocarbon degradation and protease production by Nocardiopsissp. NCIM 5124. Lett Appl Microbiol 2000b;30:67–9.
Dolak LA, Castle TM, Laborde AL. 3-Trehalosamine, a new disaccharide antibiotic. JAntibiot 1980;7:690–4.
Elleuche S, Schröder C, Sahm K, Antranikian G. Extremozymes-biocatalysts withunique properties from extremophilic microorganisms. Curr Opin Biotechnol2014;29:116–23.
Engelhardt K, Degnes KF, Kemmler M, Bredholt H, Fjaervik E, Klinkenberg G, et al.Production of a new thiopeptide antibiotic TP-1161, by a marine Nocardiopsisspecies. Appl Environ Microbiol 2010a;76:4969–76.
Engelhardt K, Degnes KF, Zotchev SB. Isolation and characterization of the genecluster for biosynthesis of the thiopeptide antibiotic TP-1161. Appl EnvironMicrobiol 2010b;76:7093–101.
Evtushenko LI, Taran VV, Akimov VN, Kroppenstedt RM, Tiedje JM, StackebrandtE. Nocardiopsis tropica sp. nov., Nocardiopsis trehalosi sp. nov., nom. rev. andNocardiopsis dassonvillei subsp. albirubida subsp. nov., comb. nov. Int J Syst EvolMicrobiol 2000;50:73–81.
Fang C, Zhang J, Pang H, Li Y, Xin Y, Zhang Y. Nocardiopsis flavescens sp. nov.,an actinomycete isolated from marine sediment. Int J Syst Evol Microbiol2011;61:2640–5.
Farrell D, Webb H, Johnston MA, Poulsen TA, O’Meara F, Christensen LL, et al.Toward fast determination of protein stability maps: experimental and theoret-ical analysis of mutants of a Nocardiopsis prasina serine protease. Biochemistry2012;51:5339–47.
Fibriansah G, Masuda S, Hirose R, Hamada K, Tanaka N, Nakamura S, et al. Crystal-lization and preliminary crystallographic analysis of endo-1,3-beta-glucanasefrom alkaliphilic Nocardiopsis sp. strain F96. Acta Crystallogr F: Struct Biol CrystCommun 2006;62:20–2.
Fibriansah G, Masuda S, Koizumi N, Nakamura S, Kumasaka T. The 1.3 A crystalstructure of a novel endo-beta-1,3-glucanase of glycoside hydrolase family 16from alkaliphilic Nocardiopsis sp. strain F96. Proteins 2007;69:683–90.
Fu P, Liu P, Qu H, Wang Y, Chen D, Wang H, et al. A-pyrones and diketopiperazinederivatives from the marine-derived actinomycete Nocardiopsis dassonvilleiHR10-5. J Nat Prod 2011;74:2219–23.
Gandhimathi R, Seghal KG, Hema TA, Selvin J, Rajeetha RT, Shanmughapriya S.Production and characterization of lipopeptide biosurfactant by a sponge-associated marine actinomycetes Nocardiopsis alba MSA10. Bioprocess BiosystEng 2009;32:825–35.
Gao X, Lua Y, Xing Y, Ma Y, Lu J, Bao W, et al. A novel anticancer and antifun-gus phenazine derivative from a marine actinomycete BM-17. Microbiol Res2012;167:616–22.
Ghanem NB, Mabrouk ME, Sabry SA, El-Badan DE. Degradation of polyesters by anovel marine Nocardiopsis aegyptia sp. nov.: application of Plackett–Burmanexperimental design for the improvement of PHB depolymerase activity. J Gen
Appl Microbiol 2005;51:151–8.Gohel SD, Singh SP. Purification strategies, characteristics and thermodynamic anal-ysis of a highly thermostable alkaline protease from a salt-tolerant alkaliphilicactinomycete, Nocardiopsis alba OK-5. J Chromatogr B: Analyt Technol BiomedLife Sci 2012;889-890:61–8.

4 gical R
G
H
H
H
H
H
I
J
J
J
J
K
K
K
K
K
K
K
K
K
K
K
L
L
L
L
L
L
L
L
L
6 T. Bennur et al. / Microbiolo
onzález-López MA, González-Vela MC, Salas-Venero CA, Conde R, Val-BernalJF. Cutaneous infection caused by Nocardiopsis dassonvillei presenting withsporotrichoid spread. J Am Acad Dermatol 2011;65:e90–1.
amedi J, Mohammadipanah F, Pötter G, Spröer C, Schumann P, Göker M, et al.Nocardiopsis arvandica sp. nov., isolated from sandy soil. Int J Syst Evol Microbiol2011;61:1189–94.
amedi J, Mohammadipanah F, von Jan M, Pötter G, Schumann P, Spröer C, et al.Nocardiopsis sinuspersici sp. nov., isolated from sandy rhizospheric soil. Int J SystEvol Microbiol 2010;60:2346–52.
amedi J, Mohammadipanah F, Ventosa A. Systematic and biotechnological aspectsof halophilic and halotolerant actinomycetes. Extremophiles 2013;17:1–13.
ozzein WN, Goodfellow M. Nocardiopsis arabia sp. nov., a halotolerant actino-mycete isolated from a sand-dune soil. Int J Syst Evol Microbiol 2008;58:2520–4.
ozzein WN, Li WJ, Ali MI, Hammouda O, Mousa AS, Xu LH, et al. Nocardiopsis alka-liphila sp. nov., a novel alkaliphilic actinomycete isolated from desert soil inEgypt. Int J Syst Evol Microbiol 2004;54:247–52.
ntra B, Mungsuntisuk I, Nihira T, Igarashi Y, Panbangred W. Identification ofactinomycetes from plant rhizospheric soils with inhibitory activity against Col-letotrichum spp., the causative agent of anthracnose disease. BMC Res Notes2011;4:98.
ayatilake GS, Thornton MP, Leonard AC, Grimwade JE, Baker BJ. Metabolites froman Antarctic sponge associated bacterium Pseudomonas aeruginosa. J Nat Prod1996;59:293–6.
iang S, Sun W, Chen M, Dai S, Zhang L, Liu Y, et al. Diversity of culturable acti-nobacteria isolated from marine sponge Haliclona sp. Antonie Van Leeuwenhoek2007;92:405–16.
ohnson-Rollings AS, Wright H, Masciandaro G, Macci C, Doni S, Calvo-Bado LA, et al.Exploring the functional soil–microbe interface and exoenzymes through soilmetaexoproteomics. ISME J 2014., http://dx.doi.org/10.1038/ismej.2014.130.
ose PA, Jebakumar SR. Phylogenetic diversity of actinomycetes cultured fromcoastal multipond solar saltern in Tuticorin, India. Aquat Biosyst 2012;8:23.
aewkla O, Franco CM. Rational approaches to improving the isolation of endophyticactinobacteria from Australian native trees. Microb Ecol 2013;65:384–93.
altenpoth M. Actinobacteria as mutualists: general healthcare for insects? TrendsMicrobiol 2009;17:529–35.
ampfer P, Busse HJ, Rainey FA. Nocardiopsis compostus sp. nov., from the atmo-sphere of a composting facility. Int J Syst Evol Microbiol 2002;52:621–7.
elch BA, Eagen KP, Erciyas FP, Humphris EL, Thomason AR, Mitsuiki S, et al. Struc-tural and mechanistic exploration of acid resistance: kinetic stability facilitatesevolution of extremophilic behavior. J Mol Biol 2007;68:870–83.
hopade A, Biao R, Liu X, Mahadik K, Zhang L, Kokare C. Production and stability stud-ies of the biosurfactant isolated from marine Nocardiopsis sp. B4. Desalination2012;285:198–204.
im JW, Adachi H, Shin-ya K, Hayakawa Y, Seto H. Apoptolidin, a new apoptosisinducer in transformed cells from Nocardiopsis sp. J Antibiot 1997;50:628–30.
oizumi N, Isoda Y, Maeda K, Masuda S, Fibriansah G, Kumasaka T, et al. Characteri-zation of Nocardiopsis beta-1,3-glucanase with additional carbohydrate-bindingdomains. Nucleic Acids Symp Ser (Oxf) 2007;51:459–60.
oizumi N, Masuda S, Maeda K, Isoda Y, Yatsunami R, Fukui T, et al. Additionalcarbohydrate-binding modules enhance the insoluble substrate-hydrolyticactivity of beta-1,3-glucanase from alkaliphilic Nocardiopsis sp. F96. Biosci Bio-technol Biochem 2009;73:1078–82.
omaki H, Ichikawa N, Hosoyama A, Fujita N, Igarashi Y. Draft genome sequenceof marine-derived actinomycete Nocardiopsis sp. strain TP-A0876, a producer ofpolyketide pyrones. Genome Announc 2014;2(4):e00665–714.
roppenstedt RM, Evtushenko LI. The family Nocardiopsaceae. In: Dworkin M,Falkow S, Rosenberg E, Schleifer KH, Stackebrandt E, editors. The Prokaryotes: AHandbook on the Biology of Bacteria, vol. 3, 3rd ed. New York: Springer-Verlag;2006. p. 754–95.
umar V, Bharti A, Negi YK, Gusain O, Pandey P, Bisht GS. Screening of actinomycetesfrom earthworm castings for their antimicrobial activity and industrial enzymes.Braz J Microbiol 2012;43:205–14.
ee YK, Lee JH, Lee HK. Microbial symbiosis in marine sponges. J Microbiol2001;39:254–64.
i HW, Zhi XY, Yao JC, Zhou Y, Tang SK, Klenk HP, et al. Comparative genomic analysisof the genus Nocardiopsis provides new insights into its genetic mechanisms ofenvironmental adaptability. PLOS ONE 2013;8(4):e61528.
i J, Yang J, Zhu WY, He J, Tian XP, Xie Q, et al. Nocardiopsis coralliicola sp. nov.,isolated from the gorgonian coral, Menella praelonga. Int J Syst Evol Microbiol2012;62:1653–8.
i MG, Li WJ, Xu P, Cui XL, Xu LH, Jiang CL. Nocardiopsis xinjiangensis sp. nov., ahalophilic actinomycete isolated from a saline soil sample in China. Int J SystEvol Microbiol 2003;53:317–21.
i WJ, Kroppenstedt RM, Wang D, Tang SK, Lee JC, Park DJ, et al. Five novel species ofthe genus Nocardiopsis isolated from hypersaline soils and emended descriptionof Nocardiopsis salina Li et al 2004. Int J Syst Evol Microbiol 2006;56:1089–96.
i WJ, Park DJ, Tang SK, Wang D, Lee JC, Xu LH, et al. Nocardiopsis salina sp. nov., anovel halophilic actinomycete isolated from saline soil in China. Int J Syst EvolMicrobiol 2004;54:1805–9.
i YQ, Li MG, Li W, Zhao JY, Ding ZG, Cui XL, et al. Griseusin D, a new pyranonaphtho-quinone derivative from a alkaphilic Nocardiopsis sp. J Antibiot 2007;60:757–61.
in Z, Torres JP, Ammon MA, Marett L, Teichert RW, Reilly CA, et al. A bacterial sourcefor mollusk pyrone polyketides. Chem Biol 2013;20:73–81.
inares JF, Gustafsson I, Baquero F, Martinez JL. Antibiotics as intermicrobialsignaling agents instead of weapons. Proc Natl Acad Sci U S A 2006;103:19484–9.
esearch 174 (2015) 33–47
Lu CH, Li YY, Wang HX, Wang BM, Shen YM. A new phenoxazine derivative isolatedfrom marine sediment actinomycetes Nocardiopsis sp. 236. Drug Discov Ther2013;7:101–4.
Lu WD, Li AX, Guo QL. Production of novel alkalitolerant and thermostable inulinasefrom marine actinomycete Nocardiopsis sp. DN-K15 and inulin hydrolysis by theenzyme. Ann Microbiol 2014;64:441–9.
Masuda S, Endo K, Koizumi N, Hayami T, Fukazawa T, Yatsunami R, et al. Molecularidentification of a novel beta-1,3-glucanase from alkaliphilic Nocardiopsis sp.strain F96. Extremophiles 2006;10:251–5.
Meena B, Rajan LA, Vinithkumar NV, Kirubagaran R. Novel marine actinobacteriafrom emerald Andaman and Nicobar Islands: a prospective source for industrialand pharmaceutical byproducts. BMC Microbiol 2013;13:145.
Meklat A, Sabaou N, Zitouni A, Mathieu F, Lebrihi A. Isolation, taxonomy, and antag-onistic properties of halophilic actinomycetes in Saharan soils of Algeria. ApplEnviron Microbiol 2011;77:6710–4.
Meyer J. Nocardiopsis, a new genus of the order Actinomycetales. Int J Syst Bacteriol1976;26:487–93.
Miteva V, Burlingame C, Sowers T, Brenchley J. Comparative evaluation of the indige-nous microbial diversity vs. drilling fluid contaminants in the NEEM Greenlandice core. FEMS Microbiol Ecol 2014;89:238–56.
Mitsuiki S, Sakai M, Moriyama Y, Goto M, Furukawa K. Purification and some proper-ties of a keratinolytic enzyme from an alkaliphilic Nocardiopsis sp TOA-1. BiosciBiotechnol Biochem 2002;66:164–7.
Miyazawa K, Noguchi T. Distribution and origin of tetrodotoxin. J Toxicol – ToxiconRev 2001;20:11–33.
Mordarska H, Gamian A, Carrasco J. Sugar-containing lipids in the classification ofrepresentative Actinomadura and Nocardiopsis species. Arch Immunol Ther Exp1983;31:135–43.
Mordarska H, Zakrzewska-Czerwinska J, Pasciak M, Szponar B, Rowinski S. Rare,suppurative pulmonary infection caused by Nocardiopsis dassonvillei recognizedby glycolipid markers. FEMS Immunol Med Microbiol 1998;21:47–55.
Moreira KA, Albuquerque BF, Teixeira MFS, Porto ALF, Lima-Filho JL. Application ofprotease from Nocardiospis sp. as a laundry detergent additive. World J MicrobiolBiotechnol 2002;18:307–12.
Mwirichia R, Muigai AW, Tindall B, Boga HI, Stackebrandt E. Isolation and character-isation of bacteria from the haloalkaline Lake Elmenteita, Kenya. Extremophiles2010;14:339–48.
Ningthoujam DS, Kshetri P, Sanasam S, Nimaichand S. Screening, identification ofbest producers and optimization of extracellular proteases from moderatelyhalophilic alkalithermotolerant indigenous actinomycetes. World Appl Sci J2009;7:907–16.
Oclarit JM, Okada H, Ohta S, Kaminura K, Yamaoka Y, Iizuka T, et al. Anti-bacillussubstance in the marine sponge Hyatella species, produced by an associatedVibrio species bacterium. Microbios 1994;78:7–16.
Olano C, Mendez C, Salas JA. Post-PKS tailoring steps in natural product-producingactinomycetes from the perspective of combinatorial biosynthesis. Nat Prod Rep2010;27:571–616.
Parte AC. LPSN-list of prokaryotic names with standing in nomenclature. NucleicAcid Res 2014;42:D613–6.
Pastor JM, Salvador M, Argandona M, Bernal V, Reina-Bueno M, Csonka LN, et al.Ectoines in cell stress protection: uses and biotechnological production. Bio-technol Adv 2010;28:782–801.
Patil PB, Zeng Y, Coursey T, Houston P, Miller I, Chen S. Isolation and characterizationof a Nocardiopsis sp. from honeybee guts. FEMS Microbiol Lett 2010;312:110–8.
Peltola J, Andersson MA, Haahtela T, Mussalo-Rauhamaa H, Rainey FA, Kroppen-stedt RM, et al. Toxic-metabolite-producing bacteria and fungus in an indoorenvironment. Appl Environ Microbiol 2001a;67:3269–74.
Peltola JS, Andersson MA, Kämpfer P, Auling G, Kroppenstedt RM, Busse HJ,et al. Isolation of toxigenic Nocardiopsis strains from indoor environmentsand description of two new Nocardiopsis Species N. exhalans sp. nov. and N.umidischolae sp. nov. Appl Environ Microbiol 2001b;67:4293–304.
Pierson LS III, Pierson EA. Metabolism and function of phenazines in bacteria:impacts on the behavior of bacteria in the environment and biotechnologicalprocesses. Appl Microbiol Biotechnol 2010;86:1659–70.
Promnuan Y, Kudo T, Chantawannakul P. Actinomycetes isolated from beehives inThailand. World J Microbiol Biotechnol 2009;25:1685–9.
Qiao J, Chen L, Li Y, Wang J, Zhang W, Chen S. Whole-genome sequence of Nocar-diopsis alba strain ATCC BAA-2165, associated with honeybees. J Bacteriol2012;194:6358–9.
Rainey FA, Rainey NW, Kroppenstedt RM, Stackebrandt E. The Genus Nocardiopsisrepresents a phylogenetically coherent taxon and a distinct Actinomycete lin-eage: proposal of Nocardiomaceae fam. nov. Int J Syst Bacteriol 1996;46:1088–92.
Rodrigues L, Banat IM, Teixeira J, Oliveira R. Biosurfactants: potential applicationsin medicine. J Antimicrob Chemother 2006;57:609–18.
Rohamare SB, Dixit V, Nareddy PK, Sivaramakrishna D, Swamy MJ, Gaikwad SM.Polyproline fold in imparting kinetic stability to an alkaline serine endopepti-dase. Biochim Biophys Acta 2013;1834:708–16.
Romanenko LA, Tanaka N, Kalinovskaya NI, Mikhailov VV. Antimicrobial potentialof deep surface sediment associated bacteria from the Sea of Japan. World JMicrobiol Biotechnol 2013;29:1169–77.
Ron EZ, Rosenberg E. Natural roles of biosurfactants. Environ Microbiol
2001;3:229–36.Rudramurthy M, Sumangala B, Honnavar P, Madhav YB, Munegowda KC, Ravi D,et al. Nasal vestibulitis due to Nocardiopsis dassonvillei in a diabetic patient. JMed Microbiol 2012;61:1168–73.

gical R
S
S
S
S
S
S
S
S
S
S
SS
S
S
S
S
S
S
S
S
S
T
T
T
T
T
T
T
T
T
T. Bennur et al. / Microbiolo
abry SA, Ghanem NB, Abu-Ella GA, Schumann P, Stackebrandt E, Kroppenstedt RM.Nocardiopsis aegyptia sp. nov., isolated from marine sediment. Int J Syst EvolMicrobiol 2004;54:453–6.
aha S, Dhanasekaran D, Shanmugapriya S, Latha S. Nocardiopsis sp. SD5: a potentfeather degrading rare actinobacterium isolated from feather waste in TamilNadu. India J Basic Microbiol 2013;53:608–16.
aratale GD, Oh SE. Production of thermotolerant and alkalotolerant cellulolyticenzymes by isolated Nocardiopsis sp. KNU. Biodegradation 2011;22:905–19.
chäfer J, Jäckel U, Kämpfer P. Analysis of actinobacteria from mould-colonizedwater damaged building material. Syst Appl Microbiol 2010;33:260–8.
chippers A, Bosecker K, Willscher S, Spröer C, Schumann P, Kroppenstedt RM.Nocardiopsis metallicus sp. nov., a metal-leaching actinomycete isolated froman alkaline slag dump. Int J Syst Evol Microbiol 2002;52:2291–5.
chneemann I, Nagel K, Kajahn I, Labes A, Wiese J, Imhoff JF. Comprehensive investi-gation of marine actinobacteria associated with the sponge Halichondria panicea.Appl Environ Microbiol 2010a;76:3702–14.
chneemann I, Ohlendorf B, Zinecker H, Nagel K, Wiese J, Imhoff JF. Nocapy-rones A–D, gamma-pyrones from a Nocardiopsis strain isolated from the marinesponge Halichondria panicea. Nat Prod 2010b;73:1444–7.
eghal KG, Thomas TA, Selvin J. Production of a new glycolipid biosurfactant frommarine Nocardiopsis lucentensis MSA04 in solid-state cultivation. Colloids SurfB: Biointerfaces 2010;78:8–16.
elvin J, Shanmughapriya S, Gandhimathi R, Seghal KG, Rajeetha RT, Natarajaseeni-vasan K, et al. Optimization and production of novel antimicrobial agents fromsponge associated marine actinomycetes Nocardiopsis dassonvillei MAD08. ApplMicrobiol Biotechnol 2009;83:435–45.
everin J, Wohlfarth A, Galinski EA. The predominant role of recently discoveredtetrahydropyrimidines for the osmoadaptation of halophilic eubacteria. J GenMicrobiol 1992;138:1629–38.
hi W, Zusman D. Fatal attraction. Nature 1993;366:414–5.imidu U, Noguchi T, Hwang DF, Shida Y, Hashimoto K. Marine bacteria which pro-
duce tetrodotoxin. Appl Environ Microbiol 1987;53:1714–5.indhuphak W, Macdonald E, Head E. Actinomycetoma caused by Nocardiopsis das-
sonvillei. Arch Dermatol 1985;121:1332–4.ingh P, Cameotra SS. Potential applications of microbial surfactants in biomedical
sciences. Trends Biotechnol 2004;22:142–6.tamford TL, Stamford NP, Coelho LC, Araújo JM. Production and characterization
of a thermostable alpha-amylase from Nocardiopsis sp. endophyte of yam bean.Bioresour Technol 2001;76:137–41.
ubramani R, Aalbersberg W. Culturable rare Actinomycetes: diversity, iso-lation and marine natural product discovery. Appl Microbiol Biotechnol2013;97:9291–321.
ubramani R, Aalbersberg W. Marine actinomycetes: an ongoing source of novelbioactive metabolites. Microbiol Res 2012;167:571–80.
uihko ML, Kroppenstedt RM, Stackebrandt E. Occurrence and characterization ofactinobacteria and thermoactinomycetes isolated from pulp and board samplescontaining recycled fibres. J Ind Microbiol Biotechnol 2006;33:183–91.
uihko ML, Priha O, Alakomi HL, Thompson P, Mälarstig B, Stott R, et al.Detection and molecular characterization of filamentous actinobacteria andthermoactinomycetes present in water-damaged building materials. Indoor Air2009;19:268–77.
un H, Lapidus A, Nolan M, Lucas S, Rio TGD, Tice H, et al. Complete genomesequence of Nocardiopsis dassonvillei type strain (IMRU 509T). Stand GenomicSci 2010;3:325–36.
un HH, White CB, Dedinas J, Cooper R, Sedlock DM, Methylpendolmycin. an indolac-tam from a Nocardiopsis sp. J Nat Prod 1991;54:1440–3.
akahashi A, Hotta K, Saito N, Morioka M, Okami Y, Umezawa H. Production ofnovel antibiotic, dopsisamine, by a new subspecies of Nocardiopsis mutabilis withmultiple antibiotic resistance. J Antibiotics 1986;39:175–83.
an H, Deng Z, Cao L. Isolation and characterization of actinomycetes from healthygoat faeces. Lett Appl Microbiol 2009;49:248–53.
arkka MT, Lehr NA, Hampp R, Schrey SD. Plant behavior upon contact with Strep-tomycetes. Plant Signal Behav 2008;3:917–9.
homas TRA, Kavlekar DP, LokaBharathi PA. Marine drugs from sponge-microbeassociation – a review. Mar Drugs 2010;8:1417–68.
huesen E, Kogure K. Bacterial production of tetrodotoxin in four species of Chaetog-natha. Biol Bull 1989;176:191–4.
ian S, Yang Y, Liu K, Xiong Z, Xu L, Zhao L. Antimicrobial metabolites from anovel halophilic actinomycete Nocardiopsis terrae YIM 90022. Nat Prod Res2014;28:344–6.
ian SZ, Pu X, Luo G, Zhao LX, Xu LH, Li WJ, et al. Isolation and characterizationof new p-Terphenyls with antifungal, antibacterial, and antioxidant activitiesfrom halophilic actinomycete Nocardiopsis gilva YIM 90087. J Agric Food Chem2013;61:3006–12.
sujibo H, Kubota T, Yamamoto M, Miyamoto K, Inamori Y. Characterization of chiti-
nase genes from an alkaliphilic actinomycete Nocardiopsis prasina OPC-131. ApplEnviron Microbiol 2003;69:894–900.sujibo H, Miyamoto K, Hasegawa T, Inamori Y. Purification and characterization oftwo types of alkaline serine proteases produced by an alkalophilic actinomycete.J Appl Bacteriol 1990a;69:520–9.
esearch 174 (2015) 33–47 47
Tsujibo H, Sakamoto T, Miyamoto K, Hasegawa T, Fujimoto M, Inamori Y. Aminoacid compositions and partial sequences of xylanases from a new subspeciesNocardiopsis dassonvillei subsp. alba OPC-18. Agric Biol Chem 1991;55:2173–4.
Tsujibo H, Sakamoto T, Nishino N, Hasegawa T, Inamori Y. Purification and propertiesof three types of xylanases produced by an alkalophilic actinomycete. J ApplBacteriol 1990b;69:398–405.
Tul’skaya EM, Shashkov AS, Streshinskaya GM, Potekhina NV, Evtushenko LI.New structures and composition of cell wall teichoic acids from Nocardiop-sis synnemataformans, Nocardiopsis halotolerans, Nocardiopsis composta andNocardiopsis metallicus: a chemotaxonomic value. Antonie Van Leeuwenhoek2014;106:1105–17.
Vasanthi V, Hoti SL. Microbial flora in gut of Culex quinquefasciatus breeding in cesspits. Southeast Asian J Trop Med Public Health 1992;23:312–7.
Wahl M, Goecke F, Labes A, Dobretsov S, Weinberger F. The second skin: ecologicalrole of epibiotic biofilms on marine organisms. Front Microbiol 2012;3:292.
Waksman SA, Schatz A, Reynolds DM. Production of antibiotic substances by Acti-nomycetes. Ann N Y Acad Sci 2010;1213:112–24.
Walker D, Ledesma P, Delgado OD, Breccia JD. High endo-�-1,4-d-glucanase activityin a broad pH range from the alkali-tolerant Nocardiopsis sp. SES28. World JMicrobiol Biotechnol 2006;22:761–4.
Wang Y, Kern SE, Newman DK. Endogenous phenazine “antibiotics” promote anaer-obic survival of Pseudomonas aeruginosa via extracellular electron transfer. JBacteriol 2009;192:365–9.
Wang Z, Fu P, Liu P, Wang P, Hou J, Li W, et al. New pyran-2-ones from alkalophilicactinomycete Nocardiopsis alkaliphila sp. Nov. YIM-80379. Chem Biodivers2013;10:281–7.
Welsh DT. Ecological significance of compatible solute accumulation by microor-ganisms: from single cells to global climate. FEMS Microbiol Rev 2000;24:263–90.
Wildermuth H. Development and organization of the aerial mycelium in Strepto-myces coelicolor. J Gen Microbiol 1970;60:43–50.
Wu Z, Xie L, Xia G, Zhang J, Nie Y, Hu J, et al. A new tetrodotoxin-producing acti-nomycete Nocardiopsis dassonvillei, isolated from the ovaries of puffer fish Fugurubripes. Toxicon 2005a;45:851–9.
Wu Z, Yang Y, Xie L, Xia G, Hu J, Wang S, et al. Toxicity and distribution oftetrodotoxin-producing bacteria in puffer fish Fugu rubripes collected from theBohai Sea of China. Toxicon 2005b;46:471–6.
Wu ZC, Li S, Nam SJ, Liu Z, Zhang C. Nocardiamides A and B, two cyclohexapep-tides from the marine-derived actinomycete Nocardiopsis sp. CNX037. J Nat Prod2013;76:694–701.
Xu S, Yan L, Zhang X, Wang C, Feng G, Li J. Nocardiopsis fildesensis sp. nov., an actino-mycete isolated from soil. Int J Syst Evol Microbiol 2014;64:174–9.
Yamamura H, Ohkubo SY, Ishida Y, Otoguro M, Tamura T, Hayakawa M. Nocardiopsisnikkonensis sp. nov., isolated from a compost sample. Int J Syst Evol Microbiol2010;60:2967–71.
Yan X, Yan H, Liu Z, Liu X, Mo H, Zhang L. Nocardiopsis yanglingensis sp. nov., athermophilic strain isolated from a compost of button mushrooms. Antonie VanLeeuwenhoek 2011;100:415–9.
Yang R, Zhang LP, Guo LG, Shi N, Lu Z, Zhang X. Nocardiopsis valliformis sp. nov., analkaliphilic actinomycete isolated from alkali lake soil in China. Int J Syst EvolMicrobiol 2008;58:1542–6.
Yassin AF, Galinski EA, Wohlfarth A, Jahnke KD, Schaal KP, Trüper HG. A new acti-nomycete species Nocardiopsis lucentensis sp. nov. Int J Syst Evol Microbiol1993;43:266–71.
Yassin AF, Rainey FA, Burghardt J, Gierth D, Ungerechts J, Lux I, et al. Descrip-tion of Nocardiopsis synnemataformans sp. nov., elevation of Nocardiopsis albasubsp. prasina to Nocardiopsis prasina comb. nov., and designation of Nocar-diopsis antarctica and Nocardiopsis alborubida as later subjective synonyms ofNocardiopsis dassonvillei. Int J Syst Bacteriol 1997;47:983–8.
Yassin AF, Spröer C, Hupfer H, Siering C, Klenk HP. Nocardiopsis potens sp.nov., isolated from household waste. Int J Syst Evol Microbiol 2009;59:2729–33.
Yu CF, Yu PH, Chan PL, Yan Q, Wong PK. Two novel species of tetrodotoxin-producingbacteria isolated from toxic marine puffer fishes. Toxicon 2004;44:641–7.
Zhang H, Lee YK, Zhang W, Lee HK. Culturable actinobacteria from the marine spongeHymeniacidon perleve: isolation and phylogenetic diversity by 16S rRNA gene-RFLP analysis. Antonie Van Leeuwenhoek 2006;90:159–69.
Zhang JW, Zeng RY. Purification and characterization of a cold-adapted alpha-amylase produced by Nocardiopsis sp. 7326 isolated from Prydz Bay Antarctic.Mar Biotechnol (NY) 2008;10:75–82.
Zhang X, Zhang LP, Yang R, Shi N, Lu Z, Chen WX, et al. Nocardiopsis ganjiahuen-sis sp. nov., isolated from a soil from Ganjiahu China. Int J Syst Evol Microbiol2008;58:195–9.
Zhang XY, He F, Wang GH, Bao J, Xu XY, Qi SH. Diversity and antibacterial activityof culturable actinobacteria isolated from five species of the South China Sea
gorgonian corals. World J Microbiol Biotechnol 2013;29:1107–16.Zitouni A, Boudjella H, Lamari L, Badji B, Mathieu F, Lebrihi A, et al. Nocardiop-sis and Saccharothrix genera in Saharan soils in Algeria: isolation, biologicalactivities and partial characterization of antibiotics. Res Microbiol 2005;156:984–93.