mussel-farming effects on mediterranean benthic nematode communities
TRANSCRIPT

Nematology, 2008, Vol. 10(3), 323-333
Mussel-farming effects on Mediterranean benthicnematode communities
Ezzeddine MAHMOUDI 1,∗, Naceur ESSID 1, Hamouda BEYREM 1, Amor HEDFI 1,Fehmi BOUFAHJA 1, Patricia AÏSSA 1 and Pierre VITIELLO 2
1 Laboratoire de Biosurveillance de l’Environnement, Faculté des Sciences de Bizerte, 7021 Zarzouna, Bizerte, Tunisia2 Centre d’Océanologie de Marseille, Campus de Luminy, Case 901, 13009 Marseille Cedex 9, France
Received: 18 September 2007; revised: 22 October 2007Accepted for publication: 26 October 2007
Summary – Mussel aquaculture activities in coastal areas are growing rapidly throughout the world, inducing an increasing interestand concern for their potential impact on coastal marine environments. We have investigated the impact of organic loads due to thebiodeposition of a mussel farm in a lagoonar ecosystem of the Mediterranean Sea (Bizerta lagoon, northern Tunisia) on the benthicenvironment. The most evident changes in the benthic habitat under the farm were a strong reduction of oxygen penetration into thebottom sediments and a large accumulation of chlorophyll a (concentrations up to 16 µg g−1), phaeopigments (concentrations up to48 µg g−1) and total organic matter (concentrations up to 12%). Results from univariate analysis of the nematofaunal data show thatthe nematode abundance increased in all the stations located inside the mussel farm (I1, I2, I3) and the site I2, located in the centreof the mussel farm, was the most affected. At this site, Shannon-Wiener index H′, species richness (d), evenness (J′) and number ofspecies (S) decreased significantly. Results from multivariate analyses of the species abundance data demonstrated that responses ofnematode species to the organic matter enrichment were varied: Mesacanthion diplechma was eliminated at the most affected station(I2), whereas the abundances of Paracomesoma dubium, Terschellingia longicaudata and T. communis were significantly enhanced.Responses of free-living nematodes to mussel farm biodeposition (elimination of some species and increase or decrease of someothers) could lead to food limitation for their predators that, ultimately, could alter entire communities and ecosystems. Consequently,we suggest that site-specific hydrodynamic and biogeochemical conditions should be taken into account when planning new musselfarms, and meiobenthic communities should be monitored before and after farm development to prevent excessive modifications ofbenthic assemblage structure.
Keywords – biodeposition, free-living nematodes, lagoons, marine nematodes, mussel culture, Mytilus galloprovincialis, nematodecommunities.
Mussel production in the Mediterranean Sea alone isover 700 000 t yr−1 (FAO, 2000). The environmental ef-fects of mussel-farming activities have been extensivelystudied (e.g., Dahlbäck & Gunnarsson, 1981; Dinet et al.,1990; Miron et al., 2005) but many of these studies haveyielded different results, even in similar habitats and withthe use of the same taxonomic groups. For example, mus-sel farming is responsible for the intensive biodepositionof faeces and pseudo-faeces that causes strong changes inthe physical and chemical characteristics of the sedimentsbeneath the long-lines (Dahlbäck & Gunnarsson, 1981;Kaspar et al., 1985; Gilbert et al., 1997). The effects ofmussel farms on benthic environments are likely to be oflimited spatial extension and appear less relevant than theimpact of intensive fish farm activities employing exter-
* Corresponding author, e-mail: [email protected]
nal sources of organic matter (Mazzola et al., 1999, 2000).Mussel biodeposition affects benthic community structure(Castel et al., 1989; Dinet et al., 1990; Kröncke, 1996)and induces reducing conditions over a wide area of thesubstrate (Dahlbäck & Gunnarsson, 1981), which couldmarkedly alter the organic matter composition and cycling(Kaspar et al., 1985). Mussel farming is eco-sustainableas it does not alter the coastal marine ecosystem, either interms of functioning and trophic states, and it does notaffect the biota at the different levels of the biologicalorganisation (Danovaro et al., 2004). Many factors maycontribute to these seemingly conflicting results; the cul-turing method used, the density of the cultivated mussels,the water depth and the hydrographical conditions in thesystem being investigated.
© Koninklijke Brill NV, Leiden, 2008 323Also available online - www.brill.nl/nemy

E. Mahmoudi et al.
Among benthic organisms, the free-living nematodes,the most abundant taxa among the meiofauna (definedhere as microscopic metazoan invertebrates passingthrough 1 mm mesh size and retained on 40 µm meshsize sieves), have been used in impact studies in manyaquatic ecosystems (e.g., Teal & Wieser, 1966; Alke-made et al., 1993; Beyrem & Aïssa, 2000). Several au-thors demonstrated that these animals are ideal for pol-lution monitoring because of their potential for rapid re-sponses to environmental changes (e.g., Warwick, 1993;Beyrem & Aïssa, 2000; Burton et al., 2001; Mahmoudiet al., 2002). It is well known that meiofauna are relevantindicators of environmental perturbation (Warwick et al.,1988; Coull & Chandler, 1992; Somerfield et al., 1995).Indeed, Ansari et al. (1984) noted, in a sandy beach atGoa (India), a significant spatial and temporal variationof nematode density in response to organic discharge viasewage and the effect on prevailing environmental con-ditions. Schratzberger and Warwick (1998) revealed, ina microcosm experiment, that total abundance, diversityand species richness of the nematofauna decreased sig-nificantly with an increasing level of organic enrichment.Beyrem and Aissa (2000) reported that free-living nema-todes of the Bay of Bizerta (Tunisia) responded to hy-drocarbon pollution with a significant decrease of theirbiomass and diversity. At Ghar El Melh lagoon (Tunisia),Mahmoudi et al. (2002) found that salinity and dissolvedoxygen of the water, and ammonia content of the sedi-ment, affected density, biomass and diversity of the ne-matofauna. Recently, many authors (Mazzola et al., 1999,2000; Mirto et al., 2000; Danovaro et al., 2004) have pro-posed meiofaunal assemblages as tools for monitoring theimpact of organic enrichment following intensive aqua-culture activities.
In Tunisia, all mussel farms culturing Mytilus gal-loprovincialis are located in Bizerta lagoon (northernTunisia). Despite the growing interest in the critical ef-fects of coastal aquaculture on the environment, thereare no studies reporting the impacts on benthic lagoonarspecies. This study was designed to investigate the effectsof a mussel farm on the lagoonar benthos of a temperatearea of the western Mediterranean. Sediment characteris-tics and nematofaunal parameters were investigated in or-der to evaluate: i) changes in sediment characteristics, or-ganic matter composition and pigment content due to themussel-farm biodeposition; and ii) changes in nematofau-nal density and community structure.
Materials and methods
STUDY SITE AND SAMPLING
The study was carried out in December (the period ofhighest mussel-standing stocks) 2004 on a mussel farmlocated in the Bizerta lagoon (northern Tunisia) (Fig. 1).The Bizerta lagoon (37◦8′ 37◦14′N, 9◦48′ 9◦56′E) has asurface area of 150 km2 and a mean depth of 8 m. In thisaquatic ecosystem hydrodynamics are limited because thetides are negligible and most of the water circulationresults from wind-induced currents (Dellali et al., 2001;Sakka-Hlaili et al., 2006). The area covered by the farmwas 650 m2 and the average bottom depth was 5 m.The mussels were cultivated using a suspension-culturetechnique, in which the mussel larvae settle on ropeshanging down from long, horizontally anchored lines.Since the main objective of the study was to compare sitesunder the farm with sites outside the spatial extent of thefarm, we defined the position of two stations outside themussel farm (O1 and O2) and three stations within thearea of the long-lines of the mussel farm (I1, I2 and I3)(Fig. 1).
For meiofauna analysis, four replicate hand-cores (10cm2 sampling surface area) were taken at each station andfixed immediately in 4% buffered formaldehyde. Sam-pling depth was limited to the first 15 cm of the substrate.Twelve additional cores per station were subsampled forphotosynthetic pigments (chlorophyll a and phaeopig-ments), total organic matter and granulometric analyses.
SEDIMENT PARAMETERS
The redox potential discontinuity (RPD) layer wasvisually estimated as the depth at which the sedimentcolour turned from brown to grey-black. The distributionof particle grain size was determined by wet sievingof freshly collected sediment on a 63 µm sieve andthe fraction >63 µm was oven-dried at 80◦C for 24 h.The total organic matter (TOM) was determined as thedifference between the dry weight of the sediment (80◦C,24 h) and the residue left after combustion at 450◦C for 4h (Parker, 1983). Before the TOM analysis, the sedimentsamples were treated with an excess of 10% HCl toremove carbonates that may have interfered with the TOMdetermination. The chloroplastic pigment determinations(chlorophyll a and phaeopigments) of sediments werecarried out as described by Dell’Anno et al. (2002). Allanalyses were replicated three times.
324 Nematology

Mussel-farming effects on nematode communities
Fig. 1. Geographical location of the stations sampled in the Bizerta lagoon in December 2004. O1, O2: Stations outside the musselfarm; I1, I2, I3: Stations inside the mussel farm. Dashed arrows indicate the major direction of water circulation in the prospectedarea.
SAMPLE PROCESSING
Meiofauna samples were rinsed with a gentle jet offreshwater over a 1 mm sieve to exclude macrofauna,decanted over a 40 µm sieve, centrifuged three times withLudoxTM HS40 (specific density 1.18) and stained withRose Bengal. Meiofauna were counted and identified atthe higher taxon level (nematodes, copepods, polychaetes,turbellarians and ostracods) using a stereomicroscope.Nematodes, comprising ca two-thirds of total meiofaunaabundance, were identified to genus or species usingthe pictorial keys of Platt and Warwick (1983, 1988)and Warwick et al. (1998). From each sample 100nematodes were picked randomly (Ólafsson et al., 1995),impregnated with anhydrous glycerin (Platt & Warwick,1983), mounted on slides and used for microscopicspecies identification.
DATA PROCESSING
The majority of data analysis followed standard com-munity analysis methods described by Clarke (1993) andClarke and Warwick (2001) using the Plymouth Routinesin Multivariate Ecological Research (PRIMER) softwarepackage.
Since the main objective of the study was to assessthe response of nematofauna to mussel farming, statisticalanalyses focused on the faunal data. Univariate indices
were computed: total nematode abundance (I), number ofspecies (S), diversity (Shannon-Wiener index H′ = log e),species richness (Margalef’s d) and evenness (Pielou’sJ′) were calculated for each station to describe nematodeassemblage structure. To evaluate the effects of musselbiodeposition on the trophic relationships in nematodeassemblages, species were assigned to feeding typesaccording to the classification of Wieser (1953), basedon the structure of the buccal cavity: i) selective depositfeeders without buccal cavity, feeding on bacteria andsmall-sized particles; ii) non-selective deposit feederswith an unarmed cupshaped buccal cavity, feeding onlarger-sized particles; iii) epigrowth feeders with a buccalcavity armed with small teeth, scraping food off surfaces;and iv) predators with a large, strongly armed buccalcavity, feeding on other nematodes and small polychaetes.
The one-way ANOVA was used to test for overalldifferences between the computed indices in each stationand the Tukey HSD test was used in pairwise comparisonsof sites under the mussel farm and those outside the farmarea. A significant difference was assumed when P <
0.05.Multivariate data analysis was by non-parametric multi-
dimensional scaling (MDS) ordination with the Bray-Curtis similarity measure performed on square-root trans-formed species abundance data to determine whether thenematode assemblages responded to the mussel biodepo-
Vol. 10(3), 2008 325
E. Mahmoudi et al.
sition by changes in the relative abundance of species.Pairwise analysis of similarities (ANOSIM) was carriedout to determine if there were any significant differencesbetween nematode assemblages in the different stations(outside and inside the mussel farm). The nature of thecommunity groupings identified in the MDS ordinationwas explored further by applying the similarity percent-ages programme (SIMPER) to determine the contributionof individual species to the average dissimilarity betweenstations under the mussel farm and those outside the farmarea.
Results
SEDIMENT CHARACTERISTICS
The results of the environmental parameters investi-gated (RPD, TOM, grain size) are shown in Table 1. TheRPD depth was significantly higher (HSD test, P < 0.01)
at the stations outside the mussel farm (O1, O2) thanbeneath the stations within the area of the long-lines ofthe mussel farm (I1, I2, I3). The total organic matter(TOM) concentrations were significantly higher under themussel farm (HSD test, P < 0.01). The sediments inall prospected stations were characterised by medium tocoarse muddy sands, and sediment grain size results showno significant difference between the grain size distribu-tion patterns for most stations except for I2. Comparedwith stations outside the farm, the site I2 exhibited ahigher percentage of particles in the smallest (<63 µm)size fraction (HSD test, P < 0.01). Chlorophyll a con-centrations were significantly higher at the stations underthe farm (HSD test, P < 0.01). Similar patterns weredisplayed by phaeopigments and stations within the areaof the long-lines of the mussel farm maintained signifi-cantly higher values than those located outside the farm(Table 1).
MEIOFAUNAL ABUNDANCE AND COMMUNITY
STRUCTURE
Nematodes accounted on average for 68% of the totalmeiofaunal abundance at all prospected stations, followedby copepods (17 and 23% at stations outside and insidethe mussel farm, respectively), polychaetes (7% at allprospected stations), turbellarians (5 and 1% at stationsoutside and inside the mussel farm, respectively) andostracods (3 and 1% at stations outside and inside themussel farm, respectively).
A total of 34 nematode species were recorded fromthe areas around and in the mussel farms (Table 2).The stations located outside the mussel farm (O1, O2)were mainly dominated by Theristus sp., Mesacanthiondiplechma, Viscosia franzii, Metalinhomoeus setosus andParalinhomoeus deconincki. The stations I1 and I3 weredominated by Paracomesoma dubium, Viscosia franzii,Microlaimus cyatholaimoides, Theristus sp. and Terschel-lingia longicaudata. The station I2 was mainly dominatedby Paracomesoma dubium, Terschellingia communis andT. longicaudata.
At all stations nematode assemblages were dominatedby nonselective deposit feeders (49.0-61.5%), followed byepigrowth feeders (17.75-28.50%) and predators (12.25-19.75%), whereas the proportion of bacteria feeders wasgenerally low (<10%) except at I2 where we haverecorded a significant increase of selective deposit feeders(22.5%).
UNIVARIATE INDICES
The graphical summary of univariate indices for ne-matode assemblages from each station (Fig. 2) illustratessome variations between sites for most univariate com-munity attributes. The results of significance testing using
Table 1. Data on sediment parameters of the five stations investigated in Bizerte lagoon in December 2004.
Station RPD Chlorophyll a Phaeopigment TOM Grain size (%)(cm) (µg g−1) (µg g−1) (%)
Silt sand Clay sand
O1 2.3 ± 0.4 5.1 ± 1.2 13.8 ± 3.8 1.6 ± 0.8 8.66 91.34O2 1.7 ± 0.2 8.2 ± 1.5 15.5 ± 1.3 2.5 ± 0.2 8.74 91.26I1 0.6 ± 0.2 12.5 ± 3.9 36.6 ± 4.0 7.8 ± 0.4 23.24 76.76I2 0.4 ± 0.2 16.5 ± 4.5 48.8 ± 8.5 12.4 ± 3.3 30.39 69.61I3 0.7 ± 0.3 13.4 ± 4.0 38.5 ± 2.1 7.5 ± 0.3 12.51 87.49
O1, O2: stations outside the mussel farm; I1, I2, I3: stations inside the farm. RPD = redox potential discontinuity (visually estimated);TOM = total organic matter; 63 µm < sand < 1000 µm, Silt/Clay < 63 µm. The depth of the investigated station is 5 m.
326 Nematology
Mussel-farming effects on nematode communities
Table 2. Proportion (%) and standard deviation of nematode species identified in the investigated stations.
O1 O2 I1 I2 I3
Daptonema fallax 3.3 ± 2.2 1.0 ± 0.0 3.5 ± 0.6 0.5 ± 0.6 2.0 ± 1.2Daptonema hirsutum 3.5 ± 2.1 3.5 ± 1.0 0.8 ± 0.5 0.0 0.5 ± 0.6Daptonema normandicum 1.5 ± 1.3 0.8 ± 0.5 3.5 ± 0.6 0.0 2.0 ± 2.3Daptonema proprium 4.0 ± 1.4 2.5 ± 1.7 3.3 ± 0.5 4.0 ± 0.0 3.0 ± 0.0Dorylaimopsis mediterraneus 3.3 ± 1.7 3.3 ± 0.5 3.5 ± 0.6 0.0 3.0 ± 0.0Halichoanolaimus sp. 2.5 ± 2.4 2.0 ± 1.2 4.0 ± 0.8 0.0 3.0 ± 2.0Linhomoeus sp. 2.8 ± 1.3 1.8 ± 1.3 0.0 0.8 ± 0.5 0.0Marylynnia choanolaimoides 2.0 ± 0.8 2.8 ± 0.5 0.0 0.0 0.0Metalinhomoeus effilatus 3.0 ± 0.8 2.8 ± 0.5 1.3 ± 0.5 0.0 1.0 ± 0.0Metalinhomoeus setosus 9.5 ± 1.3 4.5 ± 2.4 5.0 ± 0.8 1.0 ± 0.0 5.5 ± 0.6Mesacanthion diplechma 9.0 ± 1.4 9.3 ± 2.2 3.5 ± 0.6 0.0 3.0 ± 1.2Microlaimus cyatholaimoides 1.3 ± 1.0 1.3 ± 0.5 8.8 ± 3.5 6.0 ± 0.0 10.0 ± 5.8Neotonchus chamberlaini 0.8 ± 0.5 0.3 ± 0.5 2.8 ± 0.5 0.0 3.0 ± 0.0Odontophora sp. 0.5 ± 0.6 1.0 ± 0.0 7.0 ± 0.0 0.0 7.0 ± 0.0Paracomesoma dubium 4.3 ± 3.8 4.0 ± 2.3 11.3 ± 1.0 46.3 ± 1.7 11.0 ± 1.2Paracomesoma sp. 0.0 0.0 1.0 ± 0.0 1.0 ± 0.0 0.5 ± 0.6Paralinhomoeus deconincki 6.8 ± 1.0 6.3 ± 1.0 3.5 ± 1.0 1.0 ± 0.0 4.0 ± 1.2Paralinhomoeus sp. 0.3 ± 0.5 0.0 0.3 ± 1.0 0.0 1.0 ± 1.2Paramesonchium angelae 1.8 ± 1.5 0.8 ± 1.5 0.0 0.0 0.3 ± 0.5Paramicrolaimus spirulifer 1.3 ± 0.5 0.3 ± 0.5 5.3 ± 0.5 0.0 5.5 ± 1.0Parodontophora quadristicha 0.8 ± 1.0 1.5 ± 1.0 1.0 ± 0.0 0.0 0.5 ± 0.6Prochromadorella longicaudata 1.0 ± 0.8 1.3 ± 0.5 1.0 ± 0.0 2.0 ± 0.0 0.8 ± 1.0Prochromadorella neapolitana 0.5 ± 1.0 1.3 ± 0.5 0.3 ± 0.5 0.0 0.5 ± 0.6Sabatieria lepida 4.3 ± 2.4 6.0 ± 2.9 0.0 0.0 0.0Setosabatieria hilarula 2.5 ± 1.4 2.5 ± 1.7 0.0 0.0Siphonolaimus sp. 1 0.8 ± 1.5 3.0 ± 3.4 1.0 ± 0.0 0.0 0.5 ± 0.6Siphonolaimus sp. 2 0.5 ± 0.6 1.3 ± 0.5 1.0 ± 0.0 0.0 0.3 ± 0.5Stylotheristus mutilus 1.5 ± 1.3 2.0 ± 0.8 0.3 ± 0.5 1.0 ± 1.4 0.5 ± 0.6Thalassironus sp. 1.0 ± 0.8 1.3 ± 0.5 0.0 0.0 0.0Theristus pertenuis 1.3 ± 1.0 1.5 ± 0.6 2.5 ± 1.0 0.0 2.3 ± 0.5Theristus sp. 15.0 ± 6.2 18.8 ± 2.1 8.3 ± 0.5 1.3 ± 1.7 10.0 ± 1.2Terschellingia longicaudata 0.3 ± 0.5 0.5 ± 0.6 7.0 ± 1.8 11.0 ± 0.0 9.0 ± 2.8Terschellengia communis 1.0 ± 1.2 0.8 ± 1.0 0.0 11.5 ± 1.0 0.0Viscosia franzii 8.8 ± 3.0 9.8 ± 2.2 8.3 ± 2.5 8.3 ± 1.5 10.5 ± 0.6
Total nematode abundance 123.5 ± 25.6 144.5 ± 25.7 426.0 ± 42.3 620.8 ± 59.9 245.3 ± 26.7
O1, O2: stations outside the mussel farm; I1, I2, I3: stations inside the mussel farm. Dominant species abundances (�10%) are indicatedin bold.
the one-way ANOVA for overall differences between theunivariate indices computed for each station indicate sig-nificant changes of univariate community attributes. Re-sults from multiple comparisons show that the nematodeabundance significantly increased in all stations under thefarm. At I2, located in the centre of the mussel farm,Shannon Wiener index H′, species richness (d), evenness(J′) and number of species (S) all decreased significantly(P < 0.05).
MULTIVARIATE INDICES
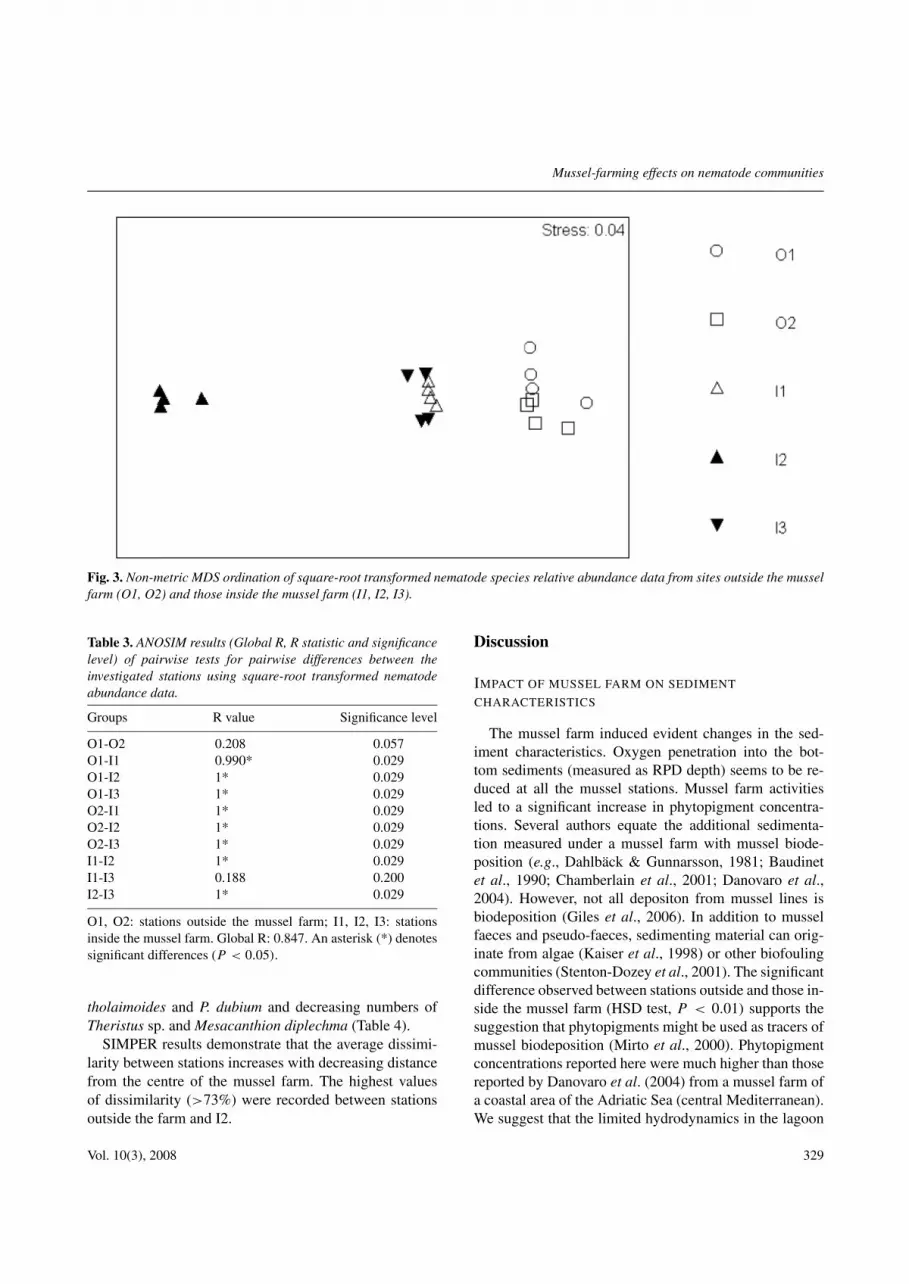
The MDS plot (Fig. 3) indicates the existence of agradient along an axis from the centre of the farm towardsthe edges and outside, but very near to the longlinesindependent of the direction on the axis. This is verifiedby the ANOSIM test (Table 3).
Significant differences between stations under the mus-sel farm and those outside the farm mainly resultedfrom changes in the abundances of the dominant species
Vol. 10(3), 2008 327

E. Mahmoudi et al.
Fig. 2. Graphical summary of means and 95% pooled confidence intervals of univariate indices for nematode assemblages from eachstation. H′ = Shannon-Wiener index; species richness = Margalef’s d; evenness = Pielou’s J; no. species = number of species (S);ns = no significant difference of the univariate index; * = significant difference of the univariate index.
(Table 4). Decreasing numbers of Theristus sp. andMesacanthion diplechma, and increasing abundance ofMicrolaimus cyatholaimoides, P. dubium and T. longi-caudata, were responsible for significant differences be-tween stations outside the mussel farm and I1. Elimina-tion of Mesacanthion diplechma, decreasing abundance
of Theristus sp., and increasing numbers of P. dubium,T. longicaudata and T. communis, were responsible forsignificant differences between stations outside the mus-sel farm and I2. Significant differences between stationsoutside the mussel farm and I3 were mainly due to in-creasing abundances of T. longicaudata, Microlaimus cya-
328 Nematology
Mussel-farming effects on nematode communities
Fig. 3. Non-metric MDS ordination of square-root transformed nematode species relative abundance data from sites outside the musselfarm (O1, O2) and those inside the mussel farm (I1, I2, I3).
Table 3. ANOSIM results (Global R, R statistic and significancelevel) of pairwise tests for pairwise differences between theinvestigated stations using square-root transformed nematodeabundance data.
Groups R value Significance level
O1-O2 0.208 0.057O1-I1 0.990* 0.029O1-I2 1* 0.029O1-I3 1* 0.029O2-I1 1* 0.029O2-I2 1* 0.029O2-I3 1* 0.029I1-I2 1* 0.029I1-I3 0.188 0.200I2-I3 1* 0.029
O1, O2: stations outside the mussel farm; I1, I2, I3: stationsinside the mussel farm. Global R: 0.847. An asterisk (*) denotessignificant differences (P < 0.05).
tholaimoides and P. dubium and decreasing numbers ofTheristus sp. and Mesacanthion diplechma (Table 4).
SIMPER results demonstrate that the average dissimi-larity between stations increases with decreasing distancefrom the centre of the mussel farm. The highest valuesof dissimilarity (>73%) were recorded between stationsoutside the farm and I2.
Discussion
IMPACT OF MUSSEL FARM ON SEDIMENT
CHARACTERISTICS
The mussel farm induced evident changes in the sed-iment characteristics. Oxygen penetration into the bot-tom sediments (measured as RPD depth) seems to be re-duced at all the mussel stations. Mussel farm activitiesled to a significant increase in phytopigment concentra-tions. Several authors equate the additional sedimenta-tion measured under a mussel farm with mussel biode-position (e.g., Dahlbäck & Gunnarsson, 1981; Baudinetet al., 1990; Chamberlain et al., 2001; Danovaro et al.,2004). However, not all depositon from mussel lines isbiodeposition (Giles et al., 2006). In addition to musselfaeces and pseudo-faeces, sedimenting material can orig-inate from algae (Kaiser et al., 1998) or other biofoulingcommunities (Stenton-Dozey et al., 2001). The significantdifference observed between stations outside and those in-side the mussel farm (HSD test, P < 0.01) supports thesuggestion that phytopigments might be used as tracers ofmussel biodeposition (Mirto et al., 2000). Phytopigmentconcentrations reported here were much higher than thosereported by Danovaro et al. (2004) from a mussel farm ofa coastal area of the Adriatic Sea (central Mediterranean).We suggest that the limited hydrodynamics in the lagoon
Vol. 10(3), 2008 329

E. Mahmoudi et al.
Table 4. The contribution percentage (%) of the SIMPER analysis for the species responsible for differences between stations insidethe mussel farm (I1, I2, I3) and those outside the mussel farm (O1, O2).
I1 I2 I3
Microlaimus cyatholaimoides (10.95) (+) Paracomesoma dubium (20.60) (+) Terschellingia longicaudata (12.13) (+)Theristus sp. (10.81) (−) Theristus sp. (9.02) (−) Microlaimus cyatholaimoides (11.65) (+)
Paracomesoma dubium (9.83) (+) Terschellingia longicaudata (7.32) (+) Paracomesoma dubium (10.05) (+)
Terschellingia longicaudata (9.42) (+) Terschellingia communis (7.15) (+) Theristus sp. (8.78) (−)
Mesacanthion diplechma (9.15) (−) Mesacanthion diplechma (6.13) (elim) Mesacanthion diplechma (7.50) (−)
(+) = more abundant; (−) = less abundant; (elim) = elimination. Species accounting for ca 50% of overall dissimilarity betweengroups are ranked in order of importance of their contribution to this dissimilarity.
(Dellali et al., 2001; Sakka-Hlaili et al., 2006) would limitthe dispersal of biodeposits and, therefore, result in highimpacts. Previous studies carried out on mussel farms sug-gested that high values of chlorophyll a are due to deposi-tion of pseudo-faeces, whilst high phaeopigment concen-trations are related to the deposition of faeces (Dahlbäck& Gunnarsson, 1981; Navarro & Thomposon, 1997). Inthe present study, we observed a phaeopigment content2-3 times higher than chlorophyll a and, on average, to-tal chloropigment concentrations (the sum of chlorophylla and phaeopigment concentrations) in the sediments be-neath the mussel farm were about three times higher thanin those outside the farm (Table 1).
Sedimentary organic carbon enrichment has been re-ported in most studies dealing with aquaculture impacts(Brown et al., 1987; Castel et al., 1989; Grenz et al., 1990;Weston, 1990; Gilbert et al., 1997; Karakassis et al., 2000;Mirto et al., 2000). Effects of mussel biodeposition wereevident in terms of total organic matter accumulation. Allstations inside the mussel farm showed TOM values sig-nificantly higher than those outside the farm area (HSDtest, P < 0.01). Previous investigations in mussel farmslocated in the Mediterranean (Gaeta Gulf, Tyrrhenian Sea)have reported that mussel biodeposition has a serious im-pact on the farm sediments. Indeed, such biodepositionhas been seen to be responsible for a significant increasein sediment concentration of organic material (Mirto etal., 2000).
IMPACT OF MUSSEL FARM ON NEMATODE
ASSEMBLAGES
The Bizerta lagoon nematofauna seemed to be affectedby mussel biodeposition. Univariate analysis showed thatat all stations under the farm nematode abundance wassignificantly higher than at sites outside the farm. Biode-position by bivalves generally provides a strong input of
organic matter of high quality and availability to benthicassemblages (Mirto et al., 2000), and previous studiesreported an increase of meiofaunal density in responseto this organic enrichment (Castel et al., 1989; Dinetet al., 1990; Guelorget et al., 1994). However, Mirto etal. (2000) demonstrated a clear decrease of turbellar-ian, ostracod and kinorhynch densities due to changesin the sedimentary characteristics of a mussel farm ina coastal area of the Tyrrhenian Sea (Western Mediter-ranean). Danovaro et al. (2004) reported that nematode,copepod, gastrotrich and polychaete densities were not af-fected by mussel farming in the Adriatic Sea. Many fac-tors can contribute to these seemingly conflicting results;the culturing method used, the density of the cultivatedmussels, the water depth, and the hydrographical condi-tions in the system under investigation (Danovaro et al.,2004).
Among stations inside the mussel farm, the site I2, lo-cated in the centre of the farm area and where we have re-corded the highest values of TOM (12.4 ± 3.3%), seemedto be the most affected. At this station all diversity in-dices decreased significantly. Mussel farm biodepositioncan lead to severe organic matter accumulation and re-ducing conditions in the sediments beneath mussel cul-tures (Kaspar et al., 1985; Kaiser et al., 1998; Mirto et al.,2000). This, in turn, affects benthic biodiversity. The sig-nificant decrease observed in all diversity indices may alsobe a consequence of changes in sediment characteristicsthat impact on the meiofauna. In fact, compared to stationsoutside the farm, the site I2 exhibited a higher percentageof silt-clay. Herman et al. (1985) reported that nematodeswere more sensitive to slight changes in sediment com-position than either macrofauna or harpacticoids. Wieser(1960) and Hopper and Meyers (1967) suggested thatmore species will be present in a habitat with a largernumber of niches, and presumed that marine sedimentsbecome more heterogeneous when the silt-clay content
330 Nematology

Mussel-farming effects on nematode communities
decreases and the grain size increases. Diversity is higherin sands, where an interstitial fauna exists, than in non-capillary sediments. Species numbers are also higher andsandy sediments contain more ‘specialist’ species thansilty ones (Heip & Decraemer, 1974). That the numberof niches is higher in sandy sediments is indicated by thecoexistence of closely related species in these sediments,indicating the existence of microhabitats. An extreme ex-ample of this is the presence of eight species of Micro-laimus on a sheltered beach found by McIntyre and Muri-son (1973).
The most abundant trophic groups in all stations werenonselective deposit feeders and epigrowth feeders. Non-selective deposit feeders were found dominant in manyTunisian lagoonar ecosystems and this was attributed tolow rates of organic matter decomposition (Beyrem et al.,2001; Hermi & Aïssa, 2001; Mahmoudi et al., 2001). Thedominance of epigrowth feeders may best be explained bythe nature of the sediment, being medium to coarse sandin most stations. In temperate habitats, epigrowth feedershave shown affinity for sandy sediments (Wieser, 1959;Heip et al., 1983) as they possess teeth capable of raspingfood particles off large sand grains or cracking open algalcells and sucking out cell fluids (Ólafsson et al., 1995).The increase of bacteria feeders observed at I1 (7%), I3(9%) and especially at I2 (22.5%) seems to be caused byan increase of microbial density commonly observed be-neath mussel cultures (Mirto et al., 2000).
Stimulation of selected nematode species has been ob-served in several studies of organic matter, and specif-ically in field experiments (Gyedu-Ababio et al., 1999;Mahmoudi et al., 2002, 2003) and microcosm experi-ments that have examined the influence of organic matteron nematofaunal abundance (Schratzberger & Warwick,1998; Gyedu-Ababio & Baird, 2006). In the present study,while M. diplechma was eliminated at the most affectedstation (I2), the abundances of P. dubium, T. longicaudataand T. communis were significantly enhanced. Differen-tial tolerance to organic matter may result in decreasedcompetition and a subsequent competitive release of moreresistant species to sediment organic enrichment. The twoTerschellingia species, T. longicaudata and T. communis,are both characterised by low oxygen consumption rates(Warwick & Price, 1979). This is of advantage in oxy-gen poor, organically enriched, sediment. At high levelsof organic enrichment, the environmental conditions werealtered to such an extent that only few species were ableto survive. Susceptible species were replaced by more tol-erant species (Schratzberger & Warwick, 1998).
Our field investigations show clearly that the lagoonarmussel farm accommodates different meiobenthic assem-blages to closely adjacent areas, and it exhibits poorer ne-matofauna in terms of numbers and altered assemblagestructure. Altered species composition could significantlyinfluence interactions between nematodes and interac-tions among major benthic taxa. In addition, many ju-venile fish and crustaceans have meiobenthos, and par-ticularly nematodes and copepods, as an obligate nutri-tional source (Gee, 1989; Coull, 1990). Response of free-living nematodes to mussel farm biodeposition (elimina-tion of some species, increase or decrease of some others)could lead to food limitation for their predators, which ul-timately could alter entire communities and ecosystems.Consequently, we suggest that site-specific hydrodynamicand biogeochemical conditions should be taken into ac-count when planning new mussel farms, and meiobenthiccommunities should be monitored before and after farmdevelopment to prevent excessive modifications of ben-thic assemblage structure.
References
ALKEMADE, R., WIELMAKER, A. & HEMMINGA, M.A.(1993). Correlation between nematode abundance and de-composition rate of Spartina anglica leaves. Marine EcologyProgress Series 99, 293-300.
ANSARI, Z.A., ANIL, C. & PARULEKAR, A.H. (1984). Effectof domestic sewage on sand beach meiofauna at Goa, India.Hydrobiologia 3, 229-233.
BAUDINET, D., ALLIOT, E., BERLAND, B., GRENZ, C.,PLANTE-CUNY, M.R., PLANTE, R. & SALEN-PICARD, C.(1990). Incidence of mussel culture on biogeochemical fluxesat the sediment-water interface. Hydrobiologia 207, 187-196.
BEYREM, H. & AÏSSA, P. (2000). Les nématodes libres,organismes-sentinelles de l’évolution des concentrationsd’hydrocarbures dans la Baie de Bizerte (Tunisie). Cahiersde Biologie Marine 41, 329-342.
BEYREM, H., MAHMOUDI, E. & AISSA, P. (2001). Évolutionspatiale de la structure trophique des peuplements de néma-todes libres de la lagune de Ghar El Melh pendant l’hiver2000. Revue de la Faculté des Sciences de Bizerte 1, 104-117.
BROWN, R., GOWEN, R.J. & MCLUSKY, D.M. (1987). Theeffects of salmon farming on the benthos of a Scottish sealoch. Journal of Experimental Marine Biology and Ecology109, 39-51.
BURTON, S.M., RUNDLE, S.D. & JONES, M.B. (2001). Therelationship between trace metal contamination and streammeiofauna. Environmental Pollution 111, 159-167.
CASTEL, J., LABOURG, P.J., ESCARAVAGE, V., AUBY, I. &GARCIA, M.E. (1989). Influence of seagrass beds and oyster
Vol. 10(3), 2008 331

E. Mahmoudi et al.
parks on the abundance and biomass patterns of meio- andmacrobenthos in tidal flats. Estuarine, Coastal and ShelfScience 28, 71-85.
CHAMBERLAIN, J., FERNANDES, T.F., READ, P., NICKELL,T.D. & DAVIES, I.M. (2001). Impacts of biodeposits fromsuspended mussel (Mytilus edulis L.) culture on the surround-ing surficial sediments. ICES Journal of Marine Science 58,411-416.
CLARKE, K.R. (1993). Non-parametric multivariate analysesof changes in community structure. Australian Journal ofEcology 18, 117-143.
CLARKE, K.R. & WARWICK, R.M. (2001). Changes inmarine communities: an approach to statistical analysis andinterpretation, 2nd edition. Plymouth, UK, Plymouth MarineLaboratory, PRIMER-E, 177 pp.
COULL, B.C. (1990). Are members of the meiofauna foodfor higher trophic levels? Transactions of the AmericanMicroscopical Society 109, 233-246.
COULL, B.C. & CHANDLER, G.T. (1992). Pollution and meio-fauna: field, laboratory and mesocosm studies. Oceanographyand Marine Biology Annual Review 30, 191-271.
DAHLBÄCK, B. & GUNNARSSON, L.A.H. (1981). Sedimen-tation and sulfate reduction under a mussel culture. MarineBiology 63, 269-275.
DANOVARO, R., GAMBI, C., LUNA, G.M. & MIRTO, S.(2004). Sustainable impact of mussel farming in the AdriaticSea (Mediterranean Sea): evidence from biochemical, micro-bial and meiofaunal indicators. Marine Pollution Bulletin 49,325-333.
DELLALI, M., GNASSIA-BARELLI, M., ROMEO, M. &AÏSSA, P. (2001). The use of acetylcholinesterase activityin Ruditapes decussatus and Mytilus galloprovincialis in thebiomonitoring of Bizerta lagoon. Comparative Biochemistryand Physiology C 130, 227-235.
DELL’ANNO, A., MEI, M.L., PUSCEDDU, A. & DANOVARO,R. (2002). Assessing the trophic state and eutrophicationof coastal marine systems: a new approach based on thebiochemical composition of sediment organic matter. MarinePollution Bulletin 44, 611-622.
DINET, A., SORNIN, J.M., SABLIÈRE, A., DELMAS, D. &FEUILLET-GIRARD, M. (1990). Influence de la biodéposi-tion de bivalves filtreurs sur les peuplements méiobenthiquesd’un marais maritime. Cahiers de Biologie Marine 31, 307-322.
FAO (2000). The state of world fisheries and aquaculture.Sofia, Bulgaria, The Editorial Group of the FAO InformationDivision, 142 pp.
GEE, J.M. (1989). An ecological and economical review ofmeiofauna as food for fish. Zoological Journal of the LinneanSociety 96, 243-261.
GILBERT, F., SOUCHU, P., BIANCHI, M. & BONIN, P. (1997).Influence of shellfish farming activities on nitrification, nitratereduction to ammonium and denitrification at the water-
sediment interface of the Thau lagoon, France. MarineEcology Progress Series 151, 143-153.
GILES, H., PILDITCH, C.A. & BELL, D.G. (2006). Sedimen-tation from mussel (Perna canaliculus) culture in the Firth ofThames, New Zealand: impacts on sediment oxygen and nu-trient fluxes. Aquaculture 261, 125-140.
GRENZ, C., HERMIN, M.N., BAUDINET, D. & DAUMAS,R. (1990). In situ biochemical and bacterial variation ofsediments enriched with mussel biodeposits. Hydrobiologia207, 153-160.
GUELORGET, O., PERTHUISOT, J.P., LAMY, N. & LEFEBRE,A. (1994). Structure et organisation de l’étang de Thaud’après la faune benthique (macrofaune, méiofaune): rela-tions avec le confinement. Oceanologica Acta 17, 105-114.
GYEDU-ABABIO, T.K. & BAIRD, D. (2006). Response ofmeiofauna and nematode communities to increased levelsof contaminants in a laboratory microcosm experiment.Ecotoxicology and Environmental Safety 63, 443-450.
GYEDU-ABABIO, T.K., FURSTENBERG, J.P., BAIRD, D. &VANREUSEL, A. (1999). Nematodes as indicators of pollu-tion: a case study from the Swartkops River estuary, SouthAfrica. Hydrobiologia 397, 155-169.
HEIP, C. & DECRAEMER, W. (1974). The diversity of nematodecommunities in the Southern North Sea. Journal of theMarine Biological Association of the United Kingdom 54, 1-3.
HEIP, C., HERMAN, R. & VINCX, M. (1983). Subtidal meio-fauna of the North Sea: a review. Biologisch Jaarboek Dodon-aea 51, 116-170.
HERMAN, R., VINCS, M. & HEIP, C. (1985). Meiofauna ofthe Belgian coastal waters: spatial and temporal variabilityand productivity. In: Heip, C. & Polk P. (Eds). Biologicalprocesses and translocations. Brussels, Belgium, Ministry ofScientific Policy, pp. 41-63.
HERMI, M. & AÏSSA, P. (2001). Structure printanière despeuplements nématologiques du lac Sud de Tunis (Tunisie).Marine Life 151, 143-153.
HOPPER, B.E. & MEYER, S.P. (1967). Foliicolous marinenematodes on turtle grass, Thalassia testudinum König, inBiscayne Bay, Florida. Bulletin of Marine Science 17, 471-517.
KAISER, M.J., LAING, I., UTTING, S.D. & BURNELL, G.M.(1998). Environmental impacts of bivalve mariculture. Jour-nal of Shellfish Research 17, 59-66.
KARAKASSIS, I., TSAPAKIS, M., HATZIYANNI, E., PA-PADOPOULOU, K. & PLAITI, W. (2000). Impact of cagefarming of fish on the seabed in three Mediterranean coastalareas. ICES Journal of Marine Science 57, 1462-1471.
KASPAR, H.F., GILLESPIE, P.A., BOYER, I.C. & MACKEN-ZIE, A.L. (1985). Effects of mussel aquaculture on the nitro-gen cycle and benthic communities in Kenepuru Sound, Mal-borough Sounds, New Zealand. Marine Biology 85, 127-136.
332 Nematology

Mussel-farming effects on nematode communities
KRÖNCKE, I. (1996). Impact of biodeposition on macrofaunalcommunities in intertidal sandflats. PSZN I: Marine Ecology17, 159-174.
MAHMOUDI, E., BEYREM, H. & AÏSSA, P. (2001). Structuretrophique des peuplements de nématodes libres d’un milieulagunaire anthropisé: la lagune de Ghar El Melh (Tunisie)en été 1999. Rapport de la Commission Internationale pourl’Exploration Scientifique de la Mer Méditerranée 36, 401.
MAHMOUDI, E., BEYREM, H. & AÏSSA, P. (2002). Réponsedes nématodes libres à la qualité des eaux et des sédiments dela lagune de Ghar El Melh (Tunisie) en été 1999. Cahiers deBiologie Marine 43, 83-93.
MAHMOUDI, E., BEYREM, H. & AÏSSA, P. (2003). Lespeuplements de nématodes libres, indicateurs du degréd’anthropisation des milieux lagunaires: cas de la lagune deBou Ghrara (Tunisie). Vie et Milieu 53, 47-59.
MAZZOLA, A., MIRTO, S. & DANOVARO, R. (1999). Initialfish-farm impact on meiofaunal assemblages in coastal sed-iments of the western Mediterranean. Marine Pollution Bul-letin 38, 1126-1133.
MAZZOLA, A., MIRTO, S., DANOVARO, R. & FABIANO, M.(2000). Fish-farming effects on benthic community structurein coastal sediments: analysis of the meiofaunal resilience.Journal of Marine Science 57, 1454-1461.
MCINTYRE, A.D. & MURISON, D.J. (1973). The meiofaunaof a flattish nursery ground. Journal of the Marine BiologicalAssociation of the United Kingdom 53, 93-118.
MIRON, G., LANDRY, T., ARCHAMBAULT, P. & FRENETTE,B. (2005). Effects of mussel-culture husbandry practices onvarious benthic characteristics. Aquaculture 43, 83-93.
MIRTO, S., LA ROZA, T., DANOVARO, R. & MAZZOLA,A. (2000). Microbial and meiofaunal response to intensivemussel-farm biodeposition in coastal sediments of the West-ern Mediterranean. Marine Pollution Bulletin 40, 244-252.
NAVARRO, J.M. & THOMPOSON, R.J. (1997). Biodepositionby the horse mussel Modiolus modiolus (Dillwyn) duringthe spring diatom bloom. Journal of Experimental MarineBiology and Ecology 209, 1-13.
ÓLAFSSON, E., JOHNSTONEA, R.W. & NDARO, S.G.M.(1995). Effects of intensive seaweed farming on the meioben-thos in a tropical lagoon. Journal of Experimental Marine Bi-ology and Ecology 191, 101-117.
PARKER, J.G. (1983). A comparison of method used forthe measurement of organic matter in marine sediments.Chemistry and Ecology 1, 201-210.
PLATT, H.M. & WARWICK, R.M. (1983). Free-living marinenematodes. Part I. British Enoplids. Synopses of the BritishFauna no. 28. Cambridge, UK, Cambridge University Press,314 pp.
PLATT, H.M. & WARWICK, R.M. (1988). Free-living marinenematodes. Part II. British Chromadorids. Synopses of the
British Fauna no 38. Leiden, The Netherlands, E.J. Brill,510 pp.
SAKKA HLAILI, A., CHIKHAOUI, M.A., EL GRAMI, B. &HADJ MABROUK, H. (2006). Effects of N and P supply onphytoplankton in Bizerte Lagoon (western Mediterranean).Journal of Experimental Marine Biology and Ecology 333,79-96.
SCHRATZBERGER, M. & WARWICK, R.M. (1998). Effectsof intensity and frequency of organic enrichment on twoestuarine nematode communities. Marine Ecology ProgressSeries 164, 83-94.
SOMERFIELD, P.J., REES, H.L. & WARWICK, R.M. (1995).Interrelationships in community structure between shallow-water marine meiofauna and macrofauna in relation to dredg-ings disposal. Marine Ecology Progress Series 127, 103-112.
STENTON-DOZEY, J., PROBYN, T. & BUSBY, A. (2001).Impact of mussel (Mytilus galloprovincialis) raft-culture onbenthic macrofauna, in situ oxygen uptake, and nutrient fluxesin Saldanha Bay, South Africa. Canadian Journal of Fisheriesand Aquatic Sciences 58, 1021-1031.
TEAL, J.M. & WIESER, W. (1966). The distribution andecology of nematodes in a Georgia salt marsh. Limnology andOceanography 11, 217-222.
WARWICK, R.M. (1993). Environmental impact studies onmarine communities: pragmatical considerations. AustralianJournal of Ecology 18, 63-80.
WARWICK, R.M. & PRICE, R. (1979). Ecological andmetabolic studies on free-living marine nematodes from anestuarine mud-flat. Estuarine, Coastal and Shelf Science 9,257-271.
WARWICK, R.M., CARR, M.R., CLARKE, K.R., GEE, J.M. &GREEN, R.H. (1988). A mesocosm experiment on the effectsof hydrocarbon and copper pollution on a sublittoral soft-sediment meiobenthic community. Marine Ecology ProgressSeries 46, 181-191.
WARWICK, R.M., PLATT, H.M. & SOMERFIELD, P.J. (1998).Free-living marine nematodes. Part III. British Monhysterida.Synopses of the British Fauna no. 53. Shrewsbury, UK, FieldStudies Council, 296 pp.
WESTON, D.P. (1990). Quantitative examination of macroben-thic community changes along an organic enrichment gradi-ent. Marine Ecology Progress Series 61, 233-244.
WIESER, W. (1953). Die Beziehung zwischen Mundhöh-lengestalt, Ernährungsweise und Vorkommen bei freileben-den marinen Nematoden. Arkiv für Zoologie 4, 439-484.
WIESER, W. (1959). The effect of grain size on the distributionof small invertebrates inhabiting the beaches of Puget Sound.Limnology and Oceanography 4, 181-194.
WIESER, W. (1960). Benthic studies in Buzzards Bay. II. Themeiofauna. Limnology and Oceanography 5, 121-137.
Vol. 10(3), 2008 333