murine response to dna encoding herpes simplex virus type-1 glycoprotein d targeted to the liver
TRANSCRIPT

Murine response to DNA encoding herpes simplex virus type-1glycoprotein D targeted to the liver
James V. Rogersa, Barbara E. Hullb, Pamela S. Finkc, Henry C. Chioud, NancyJ. Bigleyc,*
aBiomedical Sciences Ph.D. Program, Wright State University, Dayton, OH 45435, USAbDepartment of Biological Sciences, Wright State University, Dayton, OH 45435, USA
cDepartment of Microbiology and Immunology, Wright State University, Dayton, OH 45435, USAdThe Immune Response Corporation, Carlsbad, CA 92008, USA
Received 27 December 1998; accepted 17 September 1999
Abstract
Plasmid DNA encoding herpes simplex virus type-1 glycoprotein D (gD-1) was complexed with asialoorosomucoid conjugated
to poly-L-lysine. Following its intravenous injection into BALB/c mice, this complex was targeted to the liver. Liver cellsexpressing gD-1 were detected immunohistochemically through day 6 post-immunization, while gD-1 DNA was detectablethrough 14 days post-immunization. Decline of gD-1 expression and detectable gD-1 DNA in the liver correlated with in¯ux of
T cells, predominantly CD4+. The ASOR-poly-L-lysine DNA carrier system promotes hepatic expression of gD-1 and may beuseful in vaccination against herpes simplex virus type-1. # 2000 Elsevier Science Ltd. All rights reserved.
Keywords: Asialoorosomucoid; Liver; Vaccine
1. Introduction
Herpes simplex virus type-1 (HSV-1) causes recur-rent lesions in the oral mucosa of individuals world-wide. These lesions follow reactivation of the latentvirus from the trigeminal ganglion and serve as amajor site for HSV-1 transmission. HSV-1 infectionsstimulate cytotoxic T lymphocytes (CTLs), primarilyCD4+ T cells, leading to viral clearance [1±6]. A vac-cine against HSV-1 could limit the frequency andseverity of herpetic lesions.
Immunization with DNA encoding antigenic pro-teins can induce cellular and/or humoral responses byproviding prolonged antigen expression in vivo [7,8].Intracellular synthesis of DNA encoded antigens pos-sessing a conformation similar or identical to the wild-
type protein stimulate the immune system [8±10].Plasmid constructs of HSV DNA encoding either gly-coproteins B (gB) or D (gD) generate both antibodyand T cell responses. These two glycoproteins areexpressed during the early stages of HSV infection[11±14]. Immunization routes have included directinjection into footpad and skeletal muscle, intranasaladministration and injection of transfected dendriticcells [15±20]. Injection of animals with plasmids encod-ing gB and gD stimulated cell-mediated and humoralresponses, leading to protection against viral challengeof the vagina and skin [15±20].
A soluble, DNA carrier system consisting of asia-loorosomucoid (ASOR)-poly-L-lysine complexed witha plasmid DNA resists nuclease degradation and tar-gets foreign DNA to cells such as hepatocytes thatexpress receptors for ASOR [21±23]. Other cells withasialoglycoprotein receptors (ASGPr) include macro-phages and enterocytes [24±26]. Five minutes post-intravenous injection, ASOR/ASGPr complexes inter-nalize via receptor-mediated endocytosis, with 92±95%
Vaccine 18 (2000) 1522±1530
0264-410X/00/$ - see front matter # 2000 Elsevier Science Ltd. All rights reserved.
PII: S0264-410X(99 )00438 -7
www.elsevier.com/locate/vaccine
* Corresponding author. Tel.: +1-937-775-2824; fax: +1-937-775-
2012.
E-mail address: [email protected] (N.J. Bigley).

of the ASOR/ASGPr complex entering the endoso-mal±lysosomal pathway [27±29]. About 80±85% ofhepatocytes endocytose ASOR-poly-L-lysine DNAcomplexes [30], leading to the production of membranebound, intracellular and secreted proteins [22,31,32].These proteins are maximally expressed by 24±48 hand are detectable up to 6 days post-injection. Uptakeof ASOR-poly-L-lysine DNA complexes and sub-sequent transgene expression is inhibited in vivo and invitro by excess ASOR [30,33]. Partial hepatectomyprior to injection of ASOR-PL-DNA prolongs ex-pression for 11 weeks, since targeted DNA constructsremain in cytoplasmic vesicles which are not trans-ported to the nucleus [29,34].
Immunization with the ASOR-PL-DNA carrier sys-tem encoding gD-1 generated CTL responses inBALB/c (H-2d) and C3H (H-2k) mice [35]. Followingintravenous injection of mice with gD-ASOR, maximalgD-1 expression was detected in isolated liver cells at 3days post-immunization; these expression levelsdropped signi®cantly by day 7, persisting at low levelsthrough 14 days [35]. The purpose of this study was tomonitor expression of gD-1 by cells in the liver, heartand spleen and to characterize the in¯ux of T cells intothe liver following intravenous injection of nuclease-insensitive plasmid DNA encoding the gD-1 gene.
2. Materials and methods
2.1. Animals
Eight week-old female BALB/c (H-2d) mice werepurchased from Harlan Sprague-Dawley, Inc.(Indianapolis, IN) and housed in groups of 2±3 percage. Mice were housed at least one week prior to im-munization. Animal use was approved by theLaboratory Animal Care and Use Committee atWright State University.
2.2. Immunization of mice
As previously described [22,30], the pBK-CMV vec-tor expressing gD-1 [35] was complexed with asialoor-osomucoid (ASOR) conjugated to poly-L-lysine.Although the pBK-CMV vector contains the SV40eukaryotic origin of replication, this plasmid will onlyreplicate in cells infected with SV40 or expressingSV40 TAG (Stratagene, La Jolla, CA). Wilson et al.[34] characterized DNA targeted to the liver as non-replicating and remaining extrachromosomal with>95% of rescued plasmids maintaining restriction pat-terns indistinguishable from the input plasmid.Consequently, the pBK-CMV vector encoding gD-1 isnot likely to be integrated chromosomally. Mice wereimmunized intravenously via the tail vein with 1.0 ml
of phosphate-bu�ered saline (pH 7.3) containing 10 mgof DNA encoding gD-1 complexed with ASOR-poly-L-lysine (gD-ASOR). Control mice were injected withDNA encoding gD-1, but not complexed with ASOR(gD-control). Animals immunized with pBK-ASOR (aplasmid lacking the gD-1 gene, but complexed withASOR) or uninjected were used as negative controls.At 2, 4, 6, 8, 14, 21 and 28 days post-immunization,three mice were euthanized and samples of liver, spleenand heart were removed, snap frozen in liquid nitrogenand stored at ÿ708C for future DNA analysis.Additional samples of liver, spleen and heart werein®ltrated with Tissue-Tek O.C.T. embedding media(Miles Inc., Elkhart, IN) and frozen in isopentanecooled with liquid nitrogen or were ®xed in 2% buf-fered formalin (pH 7.4) and embedded in para�n.
2.3. Immunohistochemistry
Seven micron thick para�n sections of liver andheart tissues were incubated with polyclonal rabbitantibodies against HSV-1 gD, CD3 (Dako,Carpinteria, CA), or P-selectin (PharMingen, SanDiego, CA); gD-1 expression was also localized inspleen sections. The primary antibodies were detectedusing biotinylated goat anti-rabbit secondary anti-bodies from the ImmunoPure ABC AlkalinePhosphatase Staining kit, generating a red substrate(Pierce, Rockford, IL). Liver cryosections 7 mm thickwere cut at ÿ208C, air dried at room temperature and®xed for 10 min in acetone at 48C. These sections wereincubated with rat monoclonal antibodies against CD4or CD8 (Caltag Laboratories, Burlingame, CA), fol-lowed by biotinylated rabbit anti-rat secondary anti-bodies (Pierce, Rockford, IL) and localized asdescribed above. Control sections were incubated withnon-immune serum instead of primary antibody. Allslides were counterstained with 1% fast green.
2.4. DNA analysis
Frozen tissue samples were thawed and chromoso-mal DNA was isolated using TRI REAGENT1
(Molecular Research Center, Inc., Cincinnati, OH)according to the manufacturer's protocol. To detectthe presence of HSV-1 gD DNA in the tissues, thepolymerase chain reaction (PCR) was used to amplifya 309 base pair region of the HSV-1 gD gene with thefollowing primers: sense, 5 '-TTGGCGGATGCCTCTCTCAAG-3 '; antisense, 5 '-ACAGTTGCCTCCCATCCGAAAC-3 '. As an in-ternal control, a 195 base pair PCR product for glycer-aldehyde-3-phosphate-dehydrogenase (GAPDH) wasampli®ed with the following primers: sense, 5 '-CCATGGAGAAGGCTGGGG-3 '; antisense, 5 '-CAAAGTTGTCATGGATGACC-3 '. The PCR mix-
J.V. Rogers et al. / Vaccine 18 (2000) 1522±1530 1523
tures (Life Technologies, Gaithersburg, MD) contained1.0 mM MgCl2, 100 pmol of each primer and 20 ng ofchromosomal DNA. Reactions were carried out usinga Perkin Elmer GeneAmp System 2400 thermocycler.gD-1 was ampli®ed during 35 cycles with denaturation(958C), annealing (588C) and extension (728C) stepslasting 1 min, 30 s and 1 min, respectively. TheGAPDH annealing was carried out at 578C, with allsteps lasting 30 s through 25 cycles of ampli®cation.PCR products were analyzed by agarose gel (1.2%)electrophoresis followed by ethidium bromide staining.Fragments were transferred to nitrocellulose mem-branes and hybridized with an oligonucleotide probespeci®c for an internal region of the PCR product.The probe for gD-1 was 5 '-TTGCCGCGAAAGCGATTGGG-3 ' and 5 '-CTAAGCATGTGGTGGTGCA-3 ' for GAPDH.Probes were end-labeled with digoxigenin-11-dUTP/dATP using the 3 ' tailing reaction according to theGenius System User's Guide (Boehringer Mannheim,Indianapolis, IN). DNA was hybridized overnight at608C for the gD-1 probe and 428C for the GAPDHprobe. The Genius nonradioactive detection kit(Boehringer Mannheim, Indianapolis, IN) was usedaccording to the manufacturer's protocol to detectbound probes.
2.5. Quantitative data
The number of immunopositive cells for gD-1, CD3,CD4, CD8, or P-selectin was quanti®ed in 10 continu-ous ®elds from each of three tissue sections per animalper time point using an ocular grid that enclosed anarea of approximately 0.0625 mm2. P-selectin ex-pression was determined as a proportion of the 25quadrants within the ocular grid containing at leastone immunopositive cell or platelet aggregate.Quantitative data were expressed as mean2S.E.M.The Alternate Welch t test was used to comparemeans, using P < 0.05 as the level for signi®cance.
3. Results
3.1. Immunohistochemistry
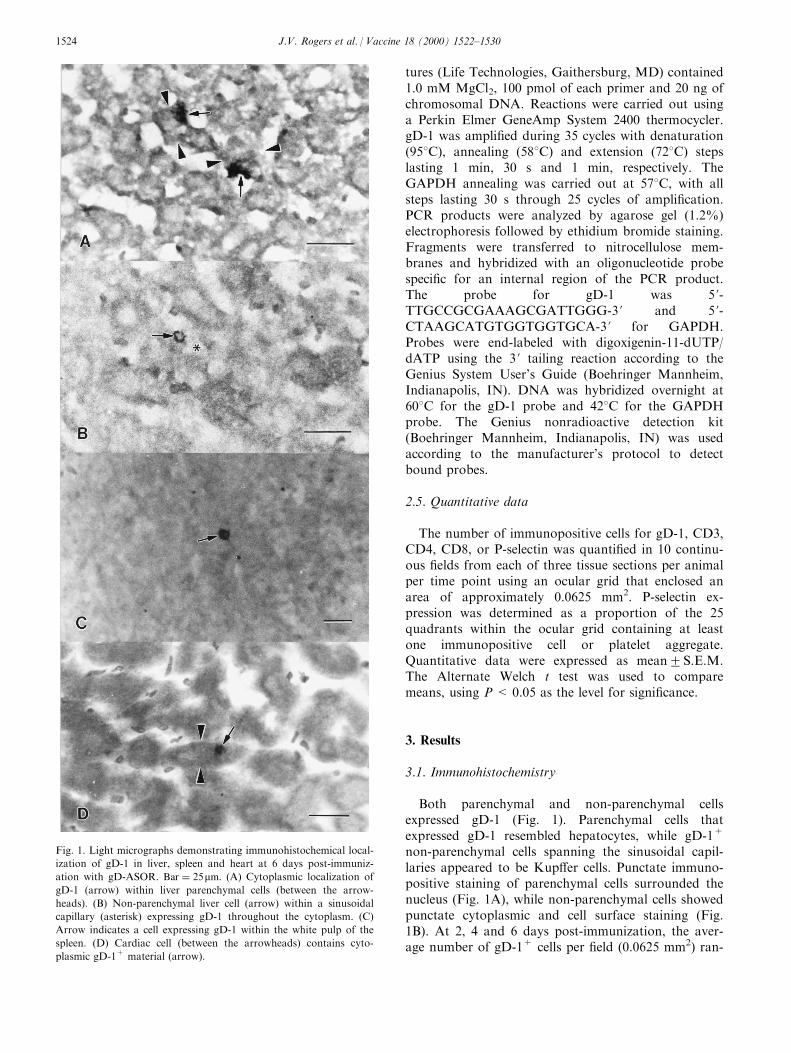
Both parenchymal and non-parenchymal cellsexpressed gD-1 (Fig. 1). Parenchymal cells thatexpressed gD-1 resembled hepatocytes, while gD-1+
non-parenchymal cells spanning the sinusoidal capil-laries appeared to be Kup�er cells. Punctate immuno-positive staining of parenchymal cells surrounded thenucleus (Fig. 1A), while non-parenchymal cells showedpunctate cytoplasmic and cell surface staining (Fig.1B). At 2, 4 and 6 days post-immunization, the aver-age number of gD-1+ cells per ®eld (0.0625 mm2) ran-
Fig. 1. Light micrographs demonstrating immunohistochemical local-
ization of gD-1 in liver, spleen and heart at 6 days post-immuniz-
ation with gD-ASOR. Bar � 25mm: (A) Cytoplasmic localization of
gD-1 (arrow) within liver parenchymal cells (between the arrow-
heads). (B) Non-parenchymal liver cell (arrow) within a sinusoidal
capillary (asterisk) expressing gD-1 throughout the cytoplasm. (C)
Arrow indicates a cell expressing gD-1 within the white pulp of the
spleen. (D) Cardiac cell (between the arrowheads) contains cyto-
plasmic gD-1+ material (arrow).
J.V. Rogers et al. / Vaccine 18 (2000) 1522±15301524
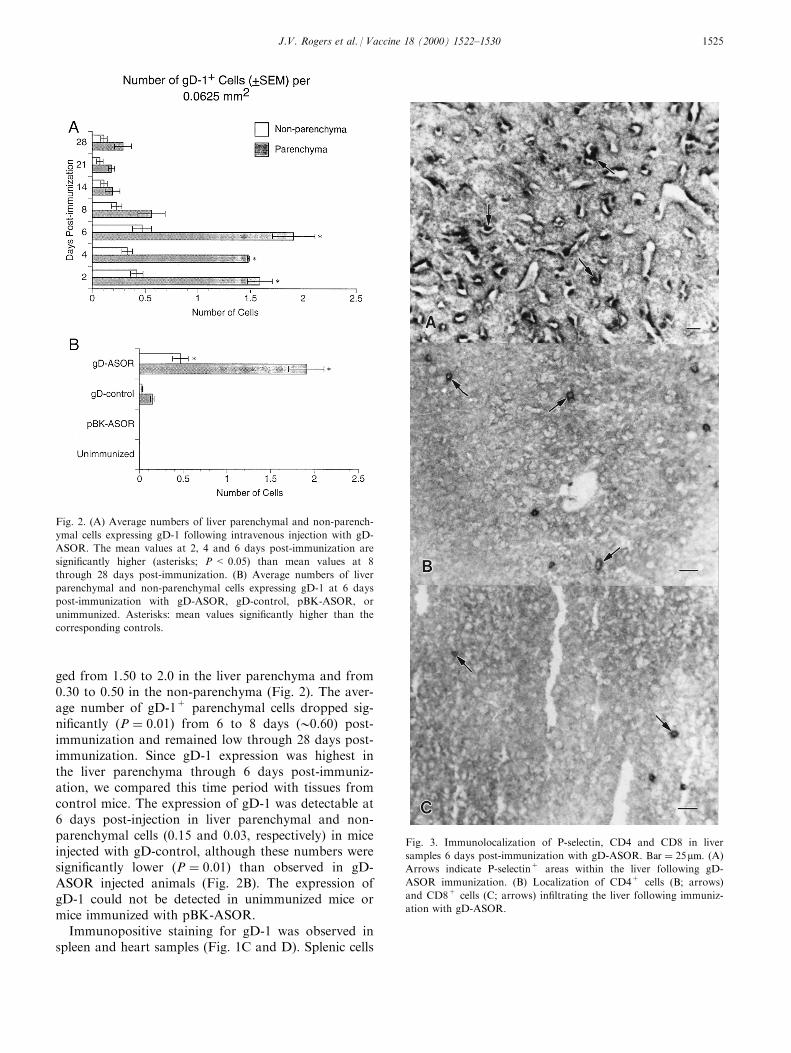
ged from 1.50 to 2.0 in the liver parenchyma and from0.30 to 0.50 in the non-parenchyma (Fig. 2). The aver-age number of gD-1+ parenchymal cells dropped sig-ni®cantly �P � 0:01� from 6 to 8 days (00.60) post-immunization and remained low through 28 days post-immunization. Since gD-1 expression was highest inthe liver parenchyma through 6 days post-immuniz-ation, we compared this time period with tissues fromcontrol mice. The expression of gD-1 was detectable at6 days post-injection in liver parenchymal and non-parenchymal cells (0.15 and 0.03, respectively) in miceinjected with gD-control, although these numbers weresigni®cantly lower �P � 0:01� than observed in gD-ASOR injected animals (Fig. 2B). The expression ofgD-1 could not be detected in unimmunized mice ormice immunized with pBK-ASOR.
Immunopositive staining for gD-1 was observed inspleen and heart samples (Fig. 1C and D). Splenic cells
Fig. 2. (A) Average numbers of liver parenchymal and non-parench-
ymal cells expressing gD-1 following intravenous injection with gD-
ASOR. The mean values at 2, 4 and 6 days post-immunization are
signi®cantly higher (asterisks; P < 0.05) than mean values at 8
through 28 days post-immunization. (B) Average numbers of liver
parenchymal and non-parenchymal cells expressing gD-1 at 6 days
post-immunization with gD-ASOR, gD-control, pBK-ASOR, or
unimmunized. Asterisks: mean values signi®cantly higher than the
corresponding controls.
Fig. 3. Immunolocalization of P-selectin, CD4 and CD8 in liver
samples 6 days post-immunization with gD-ASOR. Bar � 25mm: (A)
Arrows indicate P-selectin+ areas within the liver following gD-
ASOR immunization. (B) Localization of CD4+ cells (B; arrows)
and CD8+ cells (C; arrows) in®ltrating the liver following immuniz-
ation with gD-ASOR.
J.V. Rogers et al. / Vaccine 18 (2000) 1522±1530 1525
contained gD-1 positive material in both the cytoplasmand cell periphery, while cardiac cells expressed gD-1in the cytoplasm. The average number of gD-1+ cellsranged from 0.04 to 0.17 in the white pulp and from 0to 0.02 in the red pulp, while the average number ofgD-1+ cells in the heart ranged from 0.03 to 0.33. gD-1 expression in the spleen or heart could not be di�er-entiated from the background staining in tissue sec-tions from gD-control immunized animals.
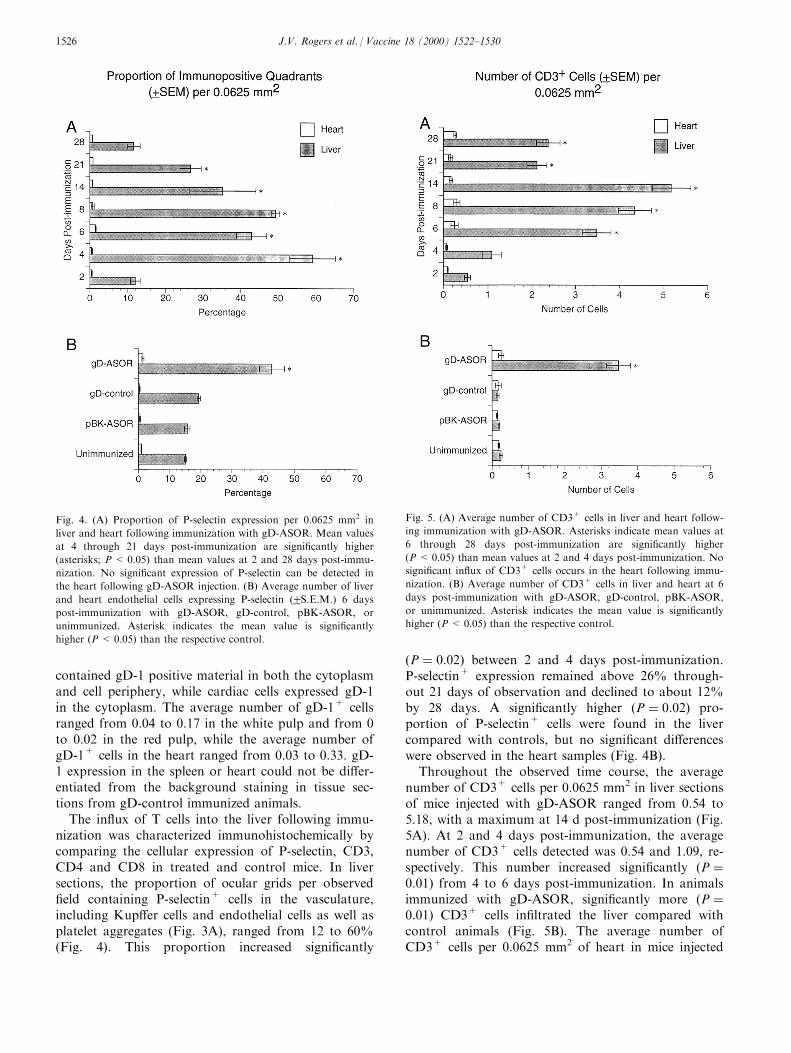
The in¯ux of T cells into the liver following immu-nization was characterized immunohistochemically bycomparing the cellular expression of P-selectin, CD3,CD4 and CD8 in treated and control mice. In liversections, the proportion of ocular grids per observed®eld containing P-selectin+ cells in the vasculature,including Kup�er cells and endothelial cells as well asplatelet aggregates (Fig. 3A), ranged from 12 to 60%(Fig. 4). This proportion increased signi®cantly
�P � 0:02� between 2 and 4 days post-immunization.P-selectin+ expression remained above 26% through-out 21 days of observation and declined to about 12%by 28 days. A signi®cantly higher �P � 0:02� pro-portion of P-selectin+ cells were found in the livercompared with controls, but no signi®cant di�erenceswere observed in the heart samples (Fig. 4B).
Throughout the observed time course, the averagenumber of CD3+ cells per 0.0625 mm2 in liver sectionsof mice injected with gD-ASOR ranged from 0.54 to5.18, with a maximum at 14 d post-immunization (Fig.5A). At 2 and 4 days post-immunization, the averagenumber of CD3+ cells detected was 0.54 and 1.09, re-spectively. This number increased signi®cantly �P �0:01� from 4 to 6 days post-immunization. In animalsimmunized with gD-ASOR, signi®cantly more �P �0:01� CD3+ cells in®ltrated the liver compared withcontrol animals (Fig. 5B). The average number ofCD3+ cells per 0.0625 mm2 of heart in mice injected
Fig. 4. (A) Proportion of P-selectin expression per 0.0625 mm2 in
liver and heart following immunization with gD-ASOR. Mean values
at 4 through 21 days post-immunization are signi®cantly higher
(asterisks; P < 0.05) than mean values at 2 and 28 days post-immu-
nization. No signi®cant expression of P-selectin can be detected in
the heart following gD-ASOR injection. (B) Average number of liver
and heart endothelial cells expressing P-selectin (2S.E.M.) 6 days
post-immunization with gD-ASOR, gD-control, pBK-ASOR, or
unimmunized. Asterisk indicates the mean value is signi®cantly
higher (P < 0.05) than the respective control.
Fig. 5. (A) Average number of CD3+ cells in liver and heart follow-
ing immunization with gD-ASOR. Asterisks indicate mean values at
6 through 28 days post-immunization are signi®cantly higher
(P < 0.05) than mean values at 2 and 4 days post-immunization. No
signi®cant in¯ux of CD3+ cells occurs in the heart following immu-
nization. (B) Average number of CD3+ cells in liver and heart at 6
days post-immunization with gD-ASOR, gD-control, pBK-ASOR,
or unimmunized. Asterisk indicates the mean value is signi®cantly
higher (P< 0.05) than the respective control.
J.V. Rogers et al. / Vaccine 18 (2000) 1522±15301526

with gD-ASOR ranged from 0.07 to 0.26; experimentaland control animals did not di�er signi®cantly (Fig.5B). During the peak periods of T cell in¯ux (6, 8 and14 days post-immunization), we observed three-foldhigher numbers of CD4+ cells than CD8+ cells (Figs.3 and 6).
3.2. DNA analysis
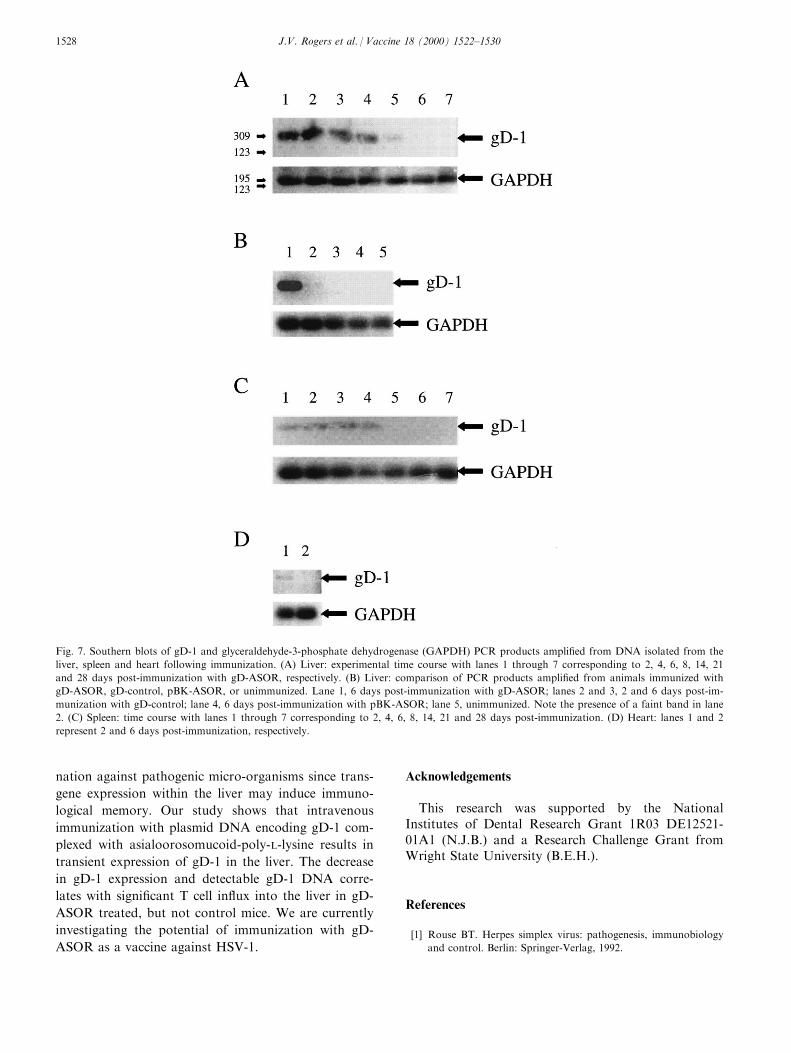
Southern analysis con®rmed the predicted sizes ofthe PCR products from liver, spleen and heart samplesof BALB/c mice as 309 bp for gD-1 and 195 bp forGAPDH. In liver samples of mice injected with gD-ASOR, DNA encoding gD-1 could be identi®ed up to14 days, but could not be detected at 21 and 28 dayspost-immunization (Fig. 7A). The GAPDH controlswere uniformly positive throughout the examined timecourse. Low, but detectable levels of DNA encodinggD-1 in liver samples occurred only at 2 days post-im-munization following gD-control immunization; gD-1DNA was not identi®ed in control liver samples ofunimmunized mice or animals injected with pBK-ASOR (Fig. 7B). The gD-1 gene was detectable inspleen and heart samples of mice injected with gD-ASOR through 8 days and 2 days post-immunization,respectively (Fig. 7C and D).
4. Discussion
In vitro and in vivo, ASGPr+ hepatocytes expresstransgenes following delivery of DNA complexed toasialoglycoprotein-poly-L-lysine [21,22,30±32,34,35].
Our results show that intravenous injection of the gD-ASOR complex leads to signi®cant expression of gD-1by parenchymal and non-parenchymal cells. Uptake ofthe gD-ASOR complex by hepatocytes presumablyre¯ects receptor-mediated endocytosis, as demon-strated with other ASOR-poly-L-lysine DNA com-plexes [21,22,30±35]. This complex could also be takenup by Kup�er cells via phagocytosis of larger aggre-gates of the ASOR-poly-L-lysine DNA complex [22].
Southern blots of PCR products ampli®ed fromliver DNA isolates demonstrated that the observedloss of gD-1 DNA leads to a decrease in the numberof hepatocytes expressing gD-1. We observed that Tcell in¯ux into the liver correlates with the decrease ingD-1 expression. In adenovirus-transduced livers ofimmunocompetent mice, transgene expression anddetectable viral DNA declines when lymphocyte in®l-tration and the number apoptotic cells increases [36±39]. These adenovirus-transduced cells are suggested tobe destroyed by the immune system and replaced byproliferating hepatocytes which do not express theforeign gene [36±39].
During T cell in¯ux, we observed signi®cantly higherexpression of P-selectin in gD-ASOR immunized ani-mals, consistent with the role P-selectin plays in leuko-cyte homing to the liver [40±44]. The T cells weobserved in®ltrating the liver are predominantlyCD4+, a subtype of T cells which respond to gD-1 inBALB/c mice [5,6,12,14,45]. We observed a 3:1 ratioof CD4+:CD8+ T cells during the ®rst two weekspost-immunization. Cruz et al. [35] also observed aCTL response in BALB/c mice immunized with gD-ASOR that was generated against cells expressingMHC class II, but not against cells expressing onlyMHC class I molecules. The signi®cant increase in Tcells in®ltrating the liver occurring as gD-1 expressiondeclines in gD-ASOR treated, but not control mice,suggests the observed T cell response may contributeto the decrease in gD-1 expression and detectable gD-1DNA.
In addition to the liver, we observed low levels ofuptake of gD-ASOR in the spleen and the heart. Inthe spleen, macrophages could phagocytize the gD-ASOR complex, resulting in low numbers of gD-1+
cells. gD-1 DNA could not be detected in heartsamples beyond 2 days post-immunization with gD-ASOR, presumably re¯ecting low levels of non-speci®cuptake of the DNA complex. Although in vivo ex-pression of DNA encoding a foreign gene by cardiaccells can occur following direct injection [46], onlyabout 0.4% of the total radiolabeled ASOR-poly-L-lysine DNA complex injected intravenously could bedetected in heart samples [30]. This low uptake of theDNA complex uptake presumably accounts for thelow number of cardiac cells expressing gD-1.
This targeted delivery system may be useful in vacci-
Fig. 6. Average number of CD4+ and CD8+ cells in liver at 6, 8
and 14 days post-immunization with gD-ASOR. Note the consis-
tantly higher proportion of CD4+ cells than CD8+ cells.
J.V. Rogers et al. / Vaccine 18 (2000) 1522±1530 1527
nation against pathogenic micro-organisms since trans-
gene expression within the liver may induce immuno-
logical memory. Our study shows that intravenous
immunization with plasmid DNA encoding gD-1 com-
plexed with asialoorosomucoid-poly-L-lysine results in
transient expression of gD-1 in the liver. The decrease
in gD-1 expression and detectable gD-1 DNA corre-
lates with signi®cant T cell in¯ux into the liver in gD-
ASOR treated, but not control mice. We are currently
investigating the potential of immunization with gD-
ASOR as a vaccine against HSV-1.
Acknowledgements
This research was supported by the NationalInstitutes of Dental Research Grant 1R03 DE12521-01A1 (N.J.B.) and a Research Challenge Grant fromWright State University (B.E.H.).
References
[1] Rouse BT. Herpes simplex virus: pathogenesis, immunobiology
and control. Berlin: Springer-Verlag, 1992.
Fig. 7. Southern blots of gD-1 and glyceraldehyde-3-phosphate dehydrogenase (GAPDH) PCR products ampli®ed from DNA isolated from the
liver, spleen and heart following immunization. (A) Liver: experimental time course with lanes 1 through 7 corresponding to 2, 4, 6, 8, 14, 21
and 28 days post-immunization with gD-ASOR, respectively. (B) Liver: comparison of PCR products ampli®ed from animals immunized with
gD-ASOR, gD-control, pBK-ASOR, or unimmunized. Lane 1, 6 days post-immunization with gD-ASOR; lanes 2 and 3, 2 and 6 days post-im-
munization with gD-control; lane 4, 6 days post-immunization with pBK-ASOR; lane 5, unimmunized. Note the presence of a faint band in lane
2. (C) Spleen: time course with lanes 1 through 7 corresponding to 2, 4, 6, 8, 14, 21 and 28 days post-immunization. (D) Heart: lanes 1 and 2
represent 2 and 6 days post-immunization, respectively.
J.V. Rogers et al. / Vaccine 18 (2000) 1522±15301528

[2] Koelle DM, Corey L, Burke RL, Eisenberg RJ, Cohen
GH, Pichyangkura R, Triezenberg SJ. Antigenic speci®cities
of human CD4+ T-cell clones recovered from recurrent
genital herpes simplex virus type 2 lesions. J Virol 1994;68:
2803±10.
[3] Smith PM, Wolcott RM, Chervenak R, Jennings SR. Control
of acute cutaneous herpes simplex virus infection: T cell-
mediated viral clearance is dependent upon interferon-g (IFN-
g). Virology 1994;202:76±88.
[4] Milligan GN, Bernstein DI. Analysis of herpes simplex virus-
speci®c T cells in the murine female genital tract following geni-
tal infection with herpes simplex virus type 2. Virology
1995;212:481±9.
[5] Ghiasi H, Cai S, Nesburn AB, Wechsler SL. MHC-II but not
MHC-I responses are required for vaccine-induced protection
against ocular challenge with HSV-1. Curr Eye Res
1997;16:1152±8.
[6] Ghiasi H, Roopenian DC, Slanina S, Cai S, Nesburn AB,
Wechsler SL. The importance of MHC-I and MHC-II responses
in vaccine e�cacy against lethal herpes simplex virus type 1
challenge. Immunology 1997;91:430±5.
[7] Pardoll DM, Beckerleg AM. Exposing the immunology of
naked DNA vaccines. Immunity 1995;3:165±9.
[8] Tighe H, Corr M, Roman M, Raz E. Gene vaccination: plasmid
DNA is more than just a blueprint. Immunol Today
1998;19:89±97.
[9] Ulmer JB, Sado� JC, Liu MA. DNA vaccines. Curr Opin
Immunol 1996;8:531±6.
[10] Donnelly JJ, Ulmer JB, Shiver JW, Liu MA. DNA vaccines.
Annu Rev Immunol 1997;15:617±48.
[11] Johnson RM, Lancki DW, Fitch FW, Spear PG. Herpes sim-
plex virus glycoprotein D is recognized as antigen by CD4+
and CD8+ T lymphocytes from infected mice. Characterization
of T cell clones. J Immunol 1990;145:702±10.
[12] Ghiasi H, Kaiwar R, Nesburn AB, Slanina S, Wechsler SL.
Expression of seven herpes simplex type 1 glycoproteins (gB,
gC, gD, gE, gG, gH and gI): comparative protection against
lethal challenge in mice. J Virol 1994;68:2118±26.
[13] Nugent CT, Wolcott RM, Chervenak R, Jennings SR. Analysis
of the cytolytic T-lymphocyte response to herpes simplex virus
type 1 glycoprotein B during primary and secondary infection. J
Virol 1994;68:7644±8.
[14] Ghiasi H, Cai S, Slanina S, Nesburn AB, Wechsler SL.
Vaccination of mice with herpes simplex virus type 1 glyco-
protein D DNA produces low levels of protection against lethal
HSV-1 challenge. Antiviral Res 1995;28:147±57.
[15] Rouse RJD, Nair SK, Lydy SL, Bowen JC, Rouse BT.
Induction in vitro of primary cytotoxic T-lymphocyte responses
with DNA encoding herpes simplex virus proteins. J Virol
1994;68:5685±9.
[16] Manickan E, Rouse RJD, Yu Z, Wire WS, Rouse BT. Genetic
immunization against herpes simplex virus. Protection is
mediated by CD4+ T lymphocytes. J Immunol 1995;155:259±
65.
[17] Bourne N, Stanberry LR, Bernstein DI, Lew D. DNA immuniz-
ation against experimental genital herpes simplex virus infection.
J Infect Dis 1996;173:800±7.
[18] Kriesel JD, Spruance SL, Daynes RA, Araneo BA. Nucleic acid
vaccine encoding gD2 protects mice from herpes simplex virus
type 2 disease. J Infect Dis 1996;173:536±41.
[19] Kuklin N, Daheshia M, Karem K, Manickan E, Rouse BT.
Induction of mucosal immunity against herpes simplex virus by
plasmid DNA immunization. J Virol 1997;71:3138±45.
[20] Manickan E, Kanangat S, Rouse RJD, Yu Z, Rouse BT.
Enhancement of immune response to naked DNA vaccines by
immunization with transfected dendritic cells. J Leukoc Biol
1997;61:125±32.
[21] Wu GY, Wu CH. Receptor-mediated in vitro gene transform-
ation by a soluble DNA carrier system. J Biol Chem
1987;262:4429±32.
[22] Wilson JM, Grossman M, Wei CH, Chowdhury NR, Wu GY,
Chowdhry JR. Hepatocyte-directed gene transfer in vivo leads
to transient improvement of hypercholesterolemia in low-density
lipoproteinreceptor-de®cient rabbits. J Biol Chem 1992;267:963±
7.
[23] Chiou HC, Tangco MV, Levine SM, Robertson D, Komis K,
Wu CH, Wu GY. Enhanced resistance to nuclease degradation
of nucleic acids complexed to asialoglycoprotein-polylysine car-
riers. Nucleic Acids Res 1994;22:5439±46.
[24] Hu CB, Lee EY, Hewitt JE, Baenziger JU, Mu JZ, DeSchryver-
Kecskemeti K, Alpers DH. The minor components of the rat
asialoglycoprotein receptor are apically located in neonatal
enterocytes. Gastroenterology 1991;101:1477±87.
[25] Mu JZ, Fallon RJ, Swanson PE, Carroll SB, Danaher M,
Alpers DH. Expression of an endogenous asialoglycoprotein
receptor in a human intestinal epithelial cell line, Caco-2.
Biochim Biophys Acta 1994;1222:83±91.
[26] Ozaki K, Lee RT, Lee YC, Kawasaki T. The di�erences in
structural speci®city for recognition and binding between asialo-
glycoprotein receptors of the liver and macrophages.
Glycoconjugate J 1995;12:268±74.
[27] Wall DA, Wilson G, Hubbard AL. The galactose-speci®c recog-
nition system of mammalian liver: the route of ligand internaliz-
ation in rat hepatocytes. Cell 1980;21:79±83.
[28] Bider MD, Cescato R, Jeno P, Spiess M. High-a�nity ligand
binding to subunit H1 of the asialoglycoprotein receptor in the
absence of subunit H2. Eur J Biochem 1995;230:207±12.
[29] Chowdhury NR, Wu CH, Wu GY, Yernei PL, Bummineni VR,
Chowdhury JR. Fate of DNA targeted to the liver by asialogly-
coprotein receptor-mediated endocytosis in vivo. Prolonged per-
sistence in cytoplasmic vesicles after partial hepatectomy. J Biol
Chem 1993;268:11265±71.
[30] Wu GY, Wu CH. Receptor-mediated gene delivery and ex-
pression in vivo. J Biol Chem 1988;263:14621±4.
[31] Wu GY, Wu CH. Evidence for targeted gene delivery to Hep
G2 hepatoma cells in vitro. Biochemistry 1988;27:887±92.
[32] Wu GY, Wilson JM, Shalaby F, Grossman M, Shafritz DA,
Wu CH. Receptor-mediated gene delivery in vivo. Partial cor-
rection of analbuminemia in nagase rats. J Biol Chem
1991;266:14338±42.
[33] Bunnell BA, Askari FK, Wilson JM. Targeted delivery of anti-
sense oligonucleotides by molecular conjugates. Somat Cell Mol
Genet 1992;18:559±69.
[34] Wilson JM, Grossman M, Cabrera JA, Wu CH, Wu GY. A
novel mechanism for achieving transgene persistance in vivo
after somatic gene transfer into hepatocytes. J Biol Chem
1992;267:11483±9.
[35] Cruz PE, Khalil PL, Dryden TD, Chiou HC, Fink PS,
Berberich SJ, Bigley NJ. A novel immunization method to
induce cytotoxic T lymphocyte responses (CTL) against plas-
mid-encoded herpes simplex virus type-1 glycoprotein D.
Vaccine 1999;17:1091±9.
[36] Engelhardt JE, Ye X, Doranz B, Wilson JM. Ablation of E2a
in recombinant adenoviruses improves transgene persistence and
decreases in¯ammatory response in mouse liver. Proc Natl Acad
Sci USA 1994;91:6196±200.
[37] Yang Y, Ertl HCJ, Wilson JM. MHC class I-restricted cytotoxic
T lymphocytes to viral antigens destroy hepatocytes in mice
infected with E1-deleted recombinant adenoviruses. Immunity
1994;1:433±42.
[38] Yang Y, Nunes FA, Berencsi K, Furth EE, Gonczol E, Wilson
JM. Cellular immunity to viral antigens limits E1-deletedadeno-
viruses for gene therapy. Proc Natl Acad Sci USA
1994;91:4407±11.
J.V. Rogers et al. / Vaccine 18 (2000) 1522±1530 1529

[39] Yang Y, Xiang Z, Ertl HC, Wilson JM. Upregulation of class I
major histocompatibility complex antigens by interferon gamma
is necessary for T-cell-mediated elimination of recombinant ade-
novirus-infected hepatocytes in vivo. Proc Natl Acad Sci USA
1995;92:7257±61.
[40] Rossiter H, Alon R, Kupper TS. Selectins, T cell rolling and in-
¯ammation. Mol Med Today 1997;3:214±22.
[41] Zibari JB, Brown MF, Burney DL, Granger N, McDonald JC.
Role of P-selectin in the recruitment of leukocytes in mouse
liver exposed to ischemia and reperfusion. Transplant Proc
1998;30:2327±30.
[42] Singh I, Zibari GB, Zizzi H, Granger DN, Cruz L, Gonsales E,
McDonald JC, Brown MF. Anti-P-selectin antibody protects
against hepatic ischemia-reperfusion injury. Transplant Proc
1998;30:2324±6.
[43] Essani NA, Fisher MA, Simmons CA, Hoover JL, Farhood A,
Jaeschke H. Increased P-selectin gene expression in the liver vas-
culature and its role in the pathophysiology of neutrophil-
induced injury in murine endotoxin shock. J Leukoc Biol
1998;63:288±96.
[44] Shi J, Kokubo Y, Wake K. Expression of P-selectin on hepatic
endothelia and platelets promoting neutrophil removal by liver
macrophages. Blood 1998;92:520±8.
[45] Martin S, Moss B, Berman PW, Laskey LA, Rouse BT.
Mechanisms of antiviral immunity induced by a vaccine virus
recombinant expressing herpes simplex virus type 1 glycoprotein
D: cytotoxic T cells. J Virol 1987;61:726±34.
[46] Lin H, Parmacek MS, Morle G, Bolling S, Leiden JM.
Expression of recombinant genes in myocardium in vivo after
direct injection of DNA. Circulation 1990;82:2217±21.
J.V. Rogers et al. / Vaccine 18 (2000) 1522±15301530