morphometric analysis of neuronal and glial cell pathology in the caudate nucleus in late-life...
TRANSCRIPT

Morphometric Analysis of Neuronal andGlial Cell Pathology in the Caudate Nucleus
in Late-Life Depression
Ahmad Khundakar, Ph.D., Christopher Morris, Ph.D.,Arthur Oakley, C.Biol., M.I.Biol.,
Alan J. Thomas, Ph.D., M.R.C.Psych.
Objective: To assess glial and neuronal density and neuronal volume in two areasof the caudate nucleus in late-life major depression. Design: A postmortem studyusing the disector and nucleator methods to estimate neuronal density and volumeand glial density of cells from human brain tissue from the anterior portion (dor-solateral and ventromedial aspects) of the caudate nucleus. Setting: Brain tissueswere obtained from the Newcastle Brain Tissue Resource at Newcastle University, UK.Participants: The study group consisted of 13 subjects with late-life major depressionand nine comparison subjects of similar age. Results: Evidence of moderate reduc-tions in neuronal density was found in the depressed group in both the dorsolateraland ventromedial areas of the caudate nucleus. There were no significant changes inglial density or neuronal volume in either area nor was there any evidence ofdifferences in depression in early and late-onset subgroups. Conclusions: Neuroim-aging abnormalities in frontal and subcortical areas including ischemic hyperinten-sities and a reduction in volume and metabolism in the caudate nucleus have beenreported in late-life depression, and previous morphometric studies have reportedneuronal changes in prefrontal cortical areas. The findings in this study extend thesemorphometric investigations in late-life depression to the caudate nucleus, suggestingthat neuronal abnormalities are present in this subcortical nucleus as well as in theserelated prefrontal areas. (Am J Geriatr Psychiatry 2010; ●:000–000)
Key Words: Late-life depression, morphometric, disector, caudate nucleus
Major depression is common in older people,affecting at least 3% of the people older than 65
years,1 and is a severe, disabling illness with a veryhigh risk of relapse.2 Studies suggest that depression
in older people has a different etiology from depres-sion earlier in life.3 In particular, accumulating evi-dence showing that cerebrovascular disease (CVD)plays an important role in late-life depression (i.e., in
Received June 22, 2009; revised December 1, 2009; accepted December 21, 2009. From the Institute for Ageing and Health, Newcastle University,Newcastle upon Tyne, United Kingdom. Send correspondence and reprint requests to Alan J. Thomas, Ph.D., M.R.C.Psych., Wolfson ResearchCentre, Institute for Ageing and Health, Newcastle University, Campus for Ageing and Vitality, Newcastle upon Tyne NE4 5PL, United Kingdom.e-mail: [email protected]
© 2010 American Association for Geriatric Psychiatry
Am J Geriatr Psychiatry ●:●, ●●● 2010 1
AQ: 1
AQ: 7
rich3/jgp-ajgp/jgp-ajgp/jgp99910/jgp0690-10z xppws S�1 4/13/10 8:15 4/Color Figure(s): F1 Art: JGP200678 Input-rkp

older people generally regardless of age at depres-sion onset) has led to the “vascular depression” hy-pothesis.3,4 This proposes that CVD and associatedtissue ischemia in key brain areas (prefrontal andsubcortical gray matter) are an important cause ofdepression in older people. There is also evidencethat depression beginning in later life (late-onset de-pression [LOD]) has a larger contribution fromCVD,5 and therefore, it is important to explore therole of age at depression onset in late-life depression.
Several frontal-subcortical circuits have been iden-tified as abnormal in depression and one of theseprojects from the dorsolateral prefrontal cortex(DLPFC) to the dorsolateral aspect of the head of thecaudate nucleus and another from the orbitofrontalcortex (OFC) to the ventromedial aspect of the cau-date head.6–8 In both areas, these projection neu-rones originate in laminae 3 and 5, mainly layer 5,and in the striatum they synapse with �-aminobu-tyric acid (GABA)ergic neurones.6 Reductions in cau-date volume on magnetic resonance imaging havebeen reported in depressed patients of all ages,9–11
including one study looking only older depressedsubjects.12 Postmortem morphometric studies havealso reported volume reductions in the basal gangliain depression.13,14 Positron emission tomographystudies have shown hypometabolism in the basalganglia in general15 and in the caudate nucleus spe-cifically.16 Increases in hyperintensities on magneticresonance imaging have also been reported in late-life depression in the basal ganglia.17,18 Morphomet-ric studies of younger adults have reported abnor-malities in glia in the DLPFC in depression,19,20 andin late-life depression, this group has previouslyidentified a reduction in pyramidal neuronal volumein the DLPFC generally and in laminae 3 and 521 butno changes in glia. The only other previous morpho-metric study in late-life depression, in the OFC, alsoreported abnormalities in the pyramidal neuronesacross the cortex and in laminae 3 and 5, specificallya reduction in density22 but again no glial changes.Other studies have reported older subjects with de-pression to have no changes or increases in glialdensity.19,23 To the best of our knowledge, there hasbeen no previous morphometric assessment of thehead of the caudate nucleus in depression.
In this study, glial and neuronal density andneuronal volume in late-life major depression inboth the dorsolateral and ventromedial aspects of
the head of the caudate nucleus were examined. Itwas hypothesized that depressed subjects wouldhave reduced neuronal density and/or reducedneuronal volume but no changes in glial density inaccordance with the above reports.
METHODS
Subjects
Brain tissue was obtained from the NewcastleBrain Tissue Resource from elderly subjects whohad major depression during their life and elderlycomparison subjects. Permission for postmortemresearch was granted by the subjects, and ethicalapproval was granted for the use of the tissue inthis study. Most of the subjects analyzed have beenincluded in our previous reports,24,25 but new sub-jects have been added to the Newcastle Brain Tis-sue Resource. Case notes were reviewed for infor-mation on the depressive illness, including age atonset of depression, and information was obtainedfrom all subjects on vascular risk factors, cause ofdeath, and medication.
All subjects were older than 60 years and consistedof 13 subjects with Diagnostic and Statistical Manualof Mental Disorders, Fourth Edition, major depres-sion,26 and no history of any other psychiatric orneurological disorder, and nine psychiatricallyhealthy comparison subjects of similar age. All com-parison subjects were known to be capable of livingindependently and met the same criteria, except thatthey had never suffered a depressive episode. De-pressed subjects had all received clinical assessmentand diagnosis during life by senior psychiatrists, andthe Newcastle Clinicopathological committee (in-cluding senior research psychiatrists and neuropa-thologists) reviewed all their medical records afterdeath together with information from the postmor-tem and standardized neuropathological assess-ments. Diagnoses were confirmed, and any subjectswere excluded if they showed evidence of significantcognitive impairment during life or if they met neu-ropathological criteria for any known cause of de-mentia (such as Alzheimer disease, vascular demen-tia, or dementia with Lewy bodies) or had evidenceof any other neurological disorder.
Analysis of Neuronal and Glial Cell Pathology
Am J Geriatr Psychiatry ●:●, ●●● 20102
rich3/jgp-ajgp/jgp-ajgp/jgp99910/jgp0690-10z xppws S�1 4/13/10 8:15 4/Color Figure(s): F1 Art: JGP200678 Input-rkp

Tissue
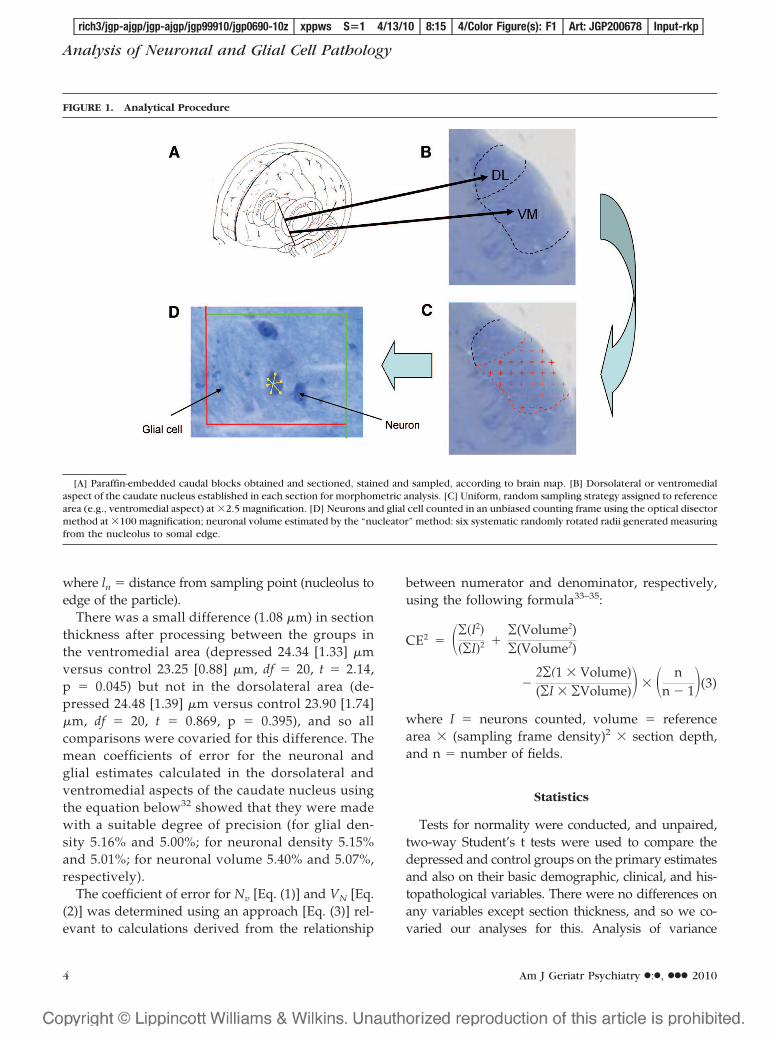
After death, the right hemisphere was fixed in 10%formalin. Tissue pH was recorded, and the delayfrom death to formalin fixation was noted. Tissueslices were obtained from predefined caudate blocks,taking the most rostral block available from the headof the caudate nucleus (i.e., tissue was from the an-terior aspect of the head of the caudate nucleus).These blocks were embedded in paraffin wax, andthe duration of fixation of these blocks was recorded.Sampling of the caudate began at the point coronallywhere the two reference areas were clearly formed,corresponding with the clear delineation of the cau-date nucleus from the putamen. The ventromedialand dorsolateral aspects of the caudate nucleus weredelineated in each section by the comparison ofNissl-stained sections against a detailed human brainatlas.27 A series of 18 30-�m sections was cut fromeach block and stained using cresyl fast violet. Thequality of the sections and the staining was carefullychecked for consistency, and all slides were coded sothat all analysis could be carried out blind to diag-nosis. For each subject, 3 of the 18 sections weresystematically randomly sampled from these sec-tions for morphometric analysis. Therefore, the firstsection was taken randomly from the first six sec-tions and then the proceeding section was sampledsix sections ahead (e.g., 3, 9, 15).
Neuronal and Glial Estimates
Neuronal density and volume and glial densitywere estimated using image analysis software(Histometrix Version 5, Kinetic Imaging Ltd., Liv-erpool, UK) to apply the optical disector28,29 andnucleator30 methods to obtain our density and par-ticle volume estimates, as in our previous study ofthe DLPFC.21 Neurones were identified using stan-dard criteria, i.e., the presence of a Nissl-stainedcytoplasm, pale nucleus, and single identifiablenucleolus in cells, which were not spherical likeglia. Glial cells were identified by their sphericalshape, absence of Nissl staining in the cytoplasm,and the heterogenous arrangement of chromatin inthe nucleus. The consistent application of thesecriteria were checked by the main rater (AK),working together with the senior investigatortrained in stereological methodology (AT) and the
experienced microscopist and research neuroscien-tist (AO). The sections were viewed in oil using aZeiss Axioplan Photomicroscope at �100 objectiveand numerical aperture of 1.25. The microscopewas attached to a JVC color video camera TK-C1360B (JVC U.K. Ltd., London, UK), a motorizedx and y axis stage accurate to 1 �m (OptiscanES110, Prior Scientific Instruments Ltd., Cam-bridge, UK), and a Heidenhain z axis depth gaugeaccurate to 0.5 �m (Heidenhain GB Ltd., London,UK) to ensure accurate measurement of disectordepth.
Based on our previous investigations,21 a uni-form, random sampling strategy (Fig. 1) was usedwithin the two regions of interest (the DLCN andVMCN), with a systematic random sampling pro-cedure applied to eliminate bias. The number ofdisectors for each patient was determined with theaim of achieving a low CE count. More than 100disectors were used per subject (mean 116 for boththe dorsolateral head and for the ventromedialhead). Each disector was 64.7-�m long, 54.4-�mwide, and 15-�m deep. A mean number of 892 gliaand 395 neurones were counted in each area foreach subject. Using the Heidenhain depth gauge,the local section thickness was assessed every 10readings. The readings showed consistent sectionthickness within individual sections, suggestinghomogeneous, uniform form of deformation fromthe section thickness (calibrated to 30 �m) beforetissue processing.31 The mean tissue thickness was24.25 and 23.89 �m, respectively, giving a meanguard zone of approximately 4.5 �m above and 4.5�m below the disector. The numerical density wascalculated using the following formula:
Nv ��p�
Q�
P � V (1)
where Nv � numerical density, p� � disector sam-ples, Q� � Q-weighted number of objects counted,P � total number of disectors, and V � disectorvolume.
The particle volume was calculated by the nucle-ator, which used the following formula30:
V�̂ N �4�
3 � ln3 �
4�
3 � n�i�1
n
ln,i3 (2)
Khundakar et al.
Am J Geriatr Psychiatry ●:●, ●●● 2010 3
F1
AQ: 2
rich3/jgp-ajgp/jgp-ajgp/jgp99910/jgp0690-10z xppws S�1 4/13/10 8:15 4/Color Figure(s): F1 Art: JGP200678 Input-rkp
where ln � distance from sampling point (nucleolus toedge of the particle).
There was a small difference (1.08 �m) in sectionthickness after processing between the groups inthe ventromedial area (depressed 24.34 [1.33] �mversus control 23.25 [0.88] �m, df � 20, t � 2.14,p � 0.045) but not in the dorsolateral area (de-pressed 24.48 [1.39] �m versus control 23.90 [1.74]�m, df � 20, t � 0.869, p � 0.395), and so allcomparisons were covaried for this difference. Themean coefficients of error for the neuronal andglial estimates calculated in the dorsolateral andventromedial aspects of the caudate nucleus usingthe equation below32 showed that they were madewith a suitable degree of precision (for glial den-sity 5.16% and 5.00%; for neuronal density 5.15%and 5.01%; for neuronal volume 5.40% and 5.07%,respectively).
The coefficient of error for Nv [Eq. (1)] and VN [Eq.(2)] was determined using an approach [Eq. (3)] rel-evant to calculations derived from the relationship
between numerator and denominator, respectively,using the following formula33–35:
CE2 � �¥�I2�
�¥I�2 �¥(Volume2)¥(Volume2)
�2¥(1 � Volume)(¥I � ¥Volume)� � � n
n � 1�(3)
where I � neurons counted, volume � referencearea � (sampling frame density)2 � section depth,and n � number of fields.
Statistics
Tests for normality were conducted, and unpaired,two-way Student’s t tests were used to compare thedepressed and control groups on the primary estimatesand also on their basic demographic, clinical, and his-topathological variables. There were no differences onany variables except section thickness, and so we co-varied our analyses for this. Analysis of variance
FIGURE 1. Analytical Procedure
[A] Paraffin-embedded caudal blocks obtained and sectioned, stained and sampled, according to brain map. [B] Dorsolateral or ventromedialaspect of the caudate nucleus established in each section for morphometric analysis. [C] Uniform, random sampling strategy assigned to referencearea (e.g., ventromedial aspect) at �2.5 magnification. [D] Neurons and glial cell counted in an unbiased counting frame using the optical disectormethod at �100 magnification; neuronal volume estimated by the “nucleator” method: six systematic randomly rotated radii generated measuringfrom the nucleolus to somal edge.
Analysis of Neuronal and Glial Cell Pathology
Am J Geriatr Psychiatry ●:●, ●●● 20104
rich3/jgp-ajgp/jgp-ajgp/jgp99910/jgp0690-10z xppws S�1 4/13/10 8:15 4/Color Figure(s): F1 Art: JGP200678 Input-rkp

(ANOVA) was used to compare the early onset, lateonset, and control groups in secondary analyses. Cor-relation coefficients were used to examine the effect ofage at depression onset. All comparisons were carriedout using SPSS software (version 15.0).
RESULTS
Demographic, clinical, and histopathological infor-mation on the study subjects is summarized in Table1. There were no significant differences between thegroups in age (df � 20, t � 4.00, p � 0.623), sex(Fisher’s exact test, p � 0.674), or any of the his-topathological variables. There were also no correla-tions between these variables and neuronal densityand volume and glial density. The causes of deathwere similar in the two groups, and the clinical fea-tures of the depressed subjects showed that in mostcases they had experienced severe depressive ill-nesses because all had required inpatient care fortheir depression and most had received at least onecourse of electroconvulsive therapy. They all had
received standard antidepressant treatment regimeswith selective serotonin reuptake inhibitors or tricy-clic antidepressants singly or often in combinationwith other agents. No comparison subject had takenany antidepressant or mood-stabilizing medication.Similar overall levels of key vascular risk factors inboth groups (myocardial infarction, angina pectoris,hypertension, diabetes mellitus, atrial fibrillation,cardiac failure, and smoking) were found from thecase note review, and the level of degenerative pa-thology was very low in all subjects, as in our earlierpublication.36
Figures 2 and 3 display the density of cells and thecell volumes, respectively, for the planned compari-sons from the estimations taken from the two sub-fields of the caudate nucleus. There were no signifi-cant differences in glial density (df � 1,19; F � 2.166,p � 0.142) or neuronal volume (df � 1,19; F � 1.765,p � 0.198) in the dorsolateral subfield of the caudatenucleus, but there was a significant reduction in neu-ronal density in the depressed group (df � 1,19; F �3.863, p � 0.039), which equates to a Cohen’s effectsize (difference between the two means divided bythe pooled SD) of 0.75. Similarly, there were also nosignificant differences in glial density (df � 1,19; F �1.164, p � 0.334) or neuronal volume (df � 1,19; F �1.831, p � 0.187) in the ventromedial subfield of thecaudate nucleus, but again there was a significantreduction in neuronal density in the depressed group(df � 1,19; F � 4.583, p � 0.024), equating to aCohen’s effect size of 0.63.
Because previous studies in late-life depression havesuggested that age at depression onset may be an im-portant etiological factor, it was analyzed, although thiswas merely exploratory because of the small numbersinvolved. Taking a cutoff of 60 to separate early onsetdepression (EOD) from LOD, there were four subjectswith EOD and nine with LOD in this investigationcompared with each other and with the nine healthycomparison subjects. Using ANOVA, in the dorsolat-eral area of the caudate nucleus, there was no signifi-cant difference in glial density (df � 2,18; F � 2.839, p �0.067). As the p value was �0.1, we conductedunpaired t tests, but these did not indicate any inter-group differences. Again, using ANOVA, there was nodifference across groups in neuronal density (df � 2,18;F � 2.703, p � 0.076), and again t tests indicated thatthere was just statistically significant reduction in neu-ronal density in the EOD group compared with the
TABLE 1. Demographic, Histopathological, and ClinicalInformation on Depressed and Control Subjectsa
Depressed(n � 13)
Control(n � 9)
Demographic informationAge (years) 73.46 (7.86) 75.11 (7.24)Sex (F:M) 7:6 6:3
Histopathological informationpH 6.29 (0.28) 6.04 (0.09)Fixation (months) 128.08 (57.40) 101.11 (58.10)PMI (hours) 24.23 (12.13) 32.11 (16.84)
Clinical featuresAge at onset of depression
(years)62.08 (16.52)
Depressed at death 12Required inpatient psychiatric
care13
Had received ECT 9Taking antidepressants at death 11
Cause of deathSuicide 2 0Pneumonia 2 0Carcinoma 1 3Ischaemic heart disease 2 1Cardiac failure 1 4Cardiac arrest 3 0Renal failure 1 0Hematemesis 0 1Aspiration pneumonia 1 0
aContinuous variables are expressed as mean (SD).
Khundakar et al.
Am J Geriatr Psychiatry ●:●, ●●● 2010 5
AQ: 3
T1,AQ:4
F2–3
AQ: 5
rich3/jgp-ajgp/jgp-ajgp/jgp99910/jgp0690-10z xppws S�1 4/13/10 8:15 4/Color Figure(s): F1 Art: JGP200678 Input-rkp
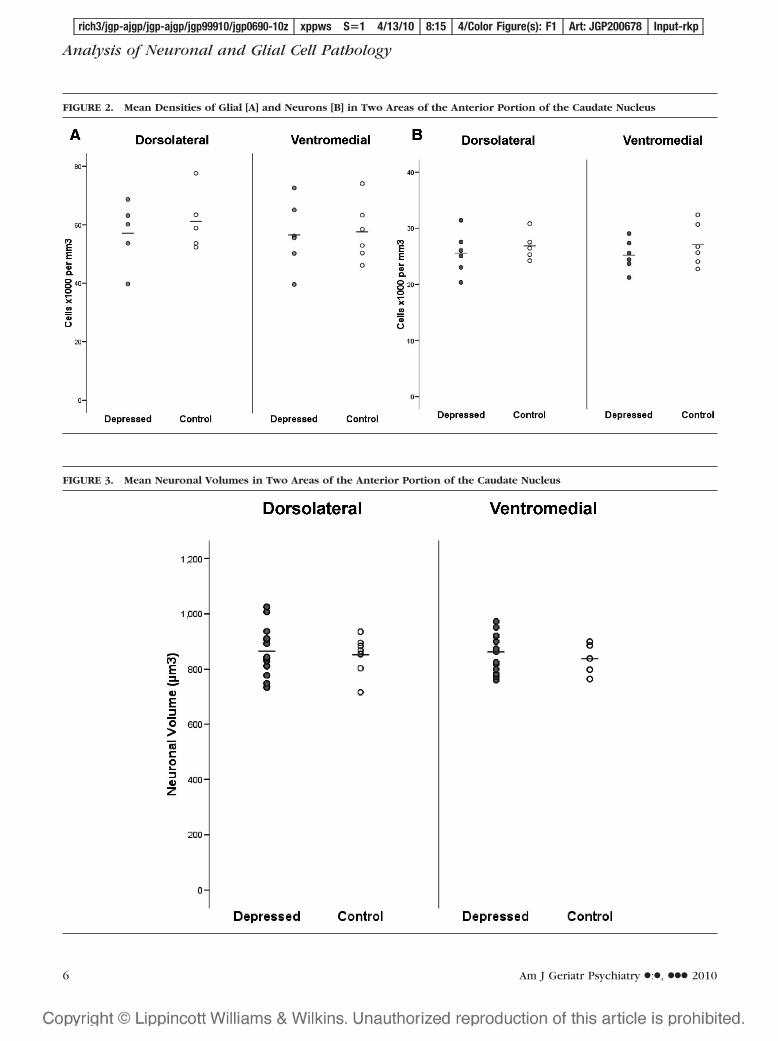
FIGURE 3. Mean Neuronal Volumes in Two Areas of the Anterior Portion of the Caudate Nucleus
FIGURE 2. Mean Densities of Glial [A] and Neurons [B] in Two Areas of the Anterior Portion of the Caudate Nucleus
Analysis of Neuronal and Glial Cell Pathology
Am J Geriatr Psychiatry ●:●, ●●● 20106
rich3/jgp-ajgp/jgp-ajgp/jgp99910/jgp0690-10z xppws S�1 4/13/10 8:15 4/Color Figure(s): F1 Art: JGP200678 Input-rkp

control group (df � 11, t � 2.273, p � 0.044). ANOVAsuggested no group differences in neuronal volume. Inthe ventromedial area, ANOVA indicated no differ-ences between these subgroups in glial density butsuggested differences in both neuronal density (df 2,18;F � 3.673, p � 0.032) and neuronal volume (df 2,18; F �3.240, p � 0.047). However, unpaired t tests did notindicate any intergroup differences in neuronal densityand also nonsignificant differences in neuronal volume.
DISCUSSION
In this first morphometric investigation of the cau-date nucleus in depression, evidence of a significantreduction in neuronal density was found in both thedorsolateral area and the ventromedial area, whichwere of moderate effect size. No evidence of changesin glial density or neuronal volume was found.
Interpretation and Limitations
A reduction in neuronal density in the caudate im-plies a reduction in neuronal number compared withage-matched comparison subjects, but in the absencean estimation of caudate volume (not possible in ourstudy), this remains an inference. Neuroimaging stud-ies in depression have reported caudate volume reduc-tion,9–12 which implies an increase in neuronal densityif the absolute numbers of neurones are the same. Thus,our finding of a decreased density, with no change inneuronal volume, suggests that neuronal numbers indepression may be even more reduced compared withhealthy subjects than imaging studies have suggested.There are two important caveats. First, it should benoted that “reduction” here is in comparison with thecontrol subjects and may or may not be due to a loss ofneurones or reduction in density over time, an impor-tant question but one a cross-sectional study cannotaddress. Second, although density reduction in suchmorphological studies is usually interpreted as indicat-ing a difference in absolute neuronal numbers, thiscannot be confirmed unless the caudate nucleus is sys-tematically randomly sampled throughout and its totalvolume estimated using the Cavalieri method. Onlysuch an approach could confirm this inference, but thisis not possible in any study of depression at the currenttime because of constraints on tissue availability. There
is the possibility of differential shrinkage between de-pressed and comparison groups during processing, al-though tissue from groups was processed randomly(rather than all subjects from a group being processedtogether), making such a systematic difference inshrinkage very unlikely. There are also considerableconstraints to working with predefined paraffin blocks,which prevented such a full stereological analysis ofthe whole caudate nucleus. However, tissue blockswere obtained from the anterior part of the caudatenucleus, which has been reported to have the mostdense glutamatergic projections from the OFC andDLPFC,7 and thus the most relevant parts of the cau-date were examined. The reduction in neuronal densityin this study is likely to be due to depressive illnessitself rather than any confounding factors because (asshown in Table 1) the two groups were well matchedfor important possible confounders, e.g., age and tissuepH, which would not therefore have biased the find-ings. Inevitably, in such a study, the groups were sys-tematically different in antidepressant medication use.However, previous studies have not suggested an ef-fect of such medication on neuronal morphometricvariables.21,22
Comparison With Previous MorphometricStudies
Our finding of a reduced neuronal density in thecaudate in late-life depression is consistent withthe previous reports of morphometric studies in theOFC22 and DLPFC,21 which also reported neuronalchanges but no glial changes in late-life depression.This study also explored whether subjects with anearly onset of their depressive illness (first episodebefore 60 years) differed from those with a late onsetand did not find consistent evidence for such differ-ences. The numbers involved in these comparisonswere very small, but the absence of clear differencesrelated to age at onset of depression is also consistentwith both the previous morphometric studies of late-life depression.21,22 This study found no change inglial cell density in either area of the caudate nucleus,which again is in accordance with these two previousstudies in cortical areas. In contrast, studies inyounger adults with depression have consistentlyreported a reduction in glial density.20,37,38 As a mor-phometric study, and similar to these earlier mor-phometric studies, we are unable to comment on the
Khundakar et al.
Am J Geriatr Psychiatry ●:●, ●●● 2010 7
AQ: 6
rich3/jgp-ajgp/jgp-ajgp/jgp99910/jgp0690-10z xppws S�1 4/13/10 8:15 4/Color Figure(s): F1 Art: JGP200678 Input-rkp

important issue of differences in glial subpopula-tions. However, we and others have reported anincrease or no difference in astroglial density in late-life depression19,23 but a reduction in younger adultswith depression.19 Although it is difficult to drawany definitive conclusions on cellular changes in dif-ferent age groups in depression because of the lim-ited number of design-based morphological studies,this study provides further evidence that glial den-sity is unchanged in late-life depression.
Potential Mechanisms Associated With NeuronalReduction
What mechanisms may be responsible for the ob-served differences in neuronal density? In our pre-vious report,21,22 it was suggested that the neuronalabnormalities may be due to the impact of ischemicbrain disease, which leads to damage to the axons ofpyramidal neurones traversing through the frontalwhite matter to the caudate nucleus from the DLPFCand OFC. An increase in WMH is well recognized inlate-life depression and such lesions have been foundto be ischemic in origin,39 especially in depressedsubjects and dominantly so in the frontal cortex. Alarge neuroimaging study in late-life depression hasreported a negative correlation between WMH vol-ume and caudate nucleus volume, especially forfrontal WMH, which supports this suggested mech-anism.17 However, as mentioned above, it is possiblethat reduced caudate volume and neuronal densityhave been present throughout life. Previous investi-gations in younger adults have reported a reductionin neuronal volume in the anterior cingulate cortexlayers 5b and 640 and in layer 3 of the OFC,41 and astudy in the OFC and DLPFC reported a trend to-ward smaller neurones in younger major depressionpatients.20 One study has reported an associationbetween reduced caudate volume in late-life depres-sion and the short allele of the serotonin transpor-ter,42 lending some support to this suggestion. Othermechanisms that may be relevant here include thepotential loss of trophic support for neurones and theeffects of hypothalamo-pituitary-adrenal axis dys-function. Decreased levels of neurotrophic factors,such as brain-derived neurotrophic factor (BDNF),have long been implicated in the pathophysiology ofdepression43 and could contribute to the atrophyfound in certain structures important in mood regu-
lation, including the caudate nucleus.17 BDNF ex-pression has been shown to be activity dependentand mediated by interplay between glutamate andGABA receptor activation.43,44 Reciprocally, chronicpretreatment with BDNF has been demonstrated topotentiate depolarization-induced GABA and gluta-mate release in cultured cortical neurones.45 Con-versely, chronic antidepressant drug treatment andelectroconvulsive shock have been shown to activatepostmonoamine receptor mechanisms governingneuronal plasticity in animal studies.46,47 Down-stream cAMP(cyclic adenosine monophosphate)/protein kinase targets such as cAMP-response ele-ment binding protein have been shown to beactivated by antidepressant treatment.48 cAMP-re-sponse element binding has been shown to regulatenumerous target genes associated with synapticfunction facilitation and cell survival, includingBDNF.46,49 The findings of this study and previousstudies21,22 are consistent with the view that dysfunc-tion within DLPFC and OFC circuitry as a result ofpathological changes to glutamatergic and GABAergicneurones may result in inadequate neurotrophic reg-ulation, further exacerbating preexisting damage. Ex-cessive hypothalamo-pituitary-adrenal axis activa-tion, which is present in major depressive disorder inyounger adults and continues into old age and tendsto become more severe,50 may also be a reason for theapparent neurone pathology. In this model, the ef-fects of elevated steroids may lead to neuronal atro-phy, which has been proposed to explain the hip-pocampal atrophy observed in depression.51
Finally, in such an elderly group of subjects, neu-rodegeneration needs to be considered as possiblyunderlying neuronal abnormalities. However, it isunlikely that neurodegenerative disease is a factor inthis sample because all subjects with any significantdegenerative disease were carefully excluded, al-though a subtle effect cannot be entirely eliminated.
Functional Importance of the Findings
A reduction in neuronal density in the caudate nu-cleus, even if combined with the effects previouslyreported of a reduction in neuronal number and/orvolume in the cortical pyramidal neurones,21,22 maynot be sufficient to produce an impact on function,because in other brain areas, e.g., the substantia nigra,much larger neuronal losses are needed to produce
Analysis of Neuronal and Glial Cell Pathology
Am J Geriatr Psychiatry ●:●, ●●● 20108
rich3/jgp-ajgp/jgp-ajgp/jgp99910/jgp0690-10z xppws S�1 4/13/10 8:15 4/Color Figure(s): F1 Art: JGP200678 Input-rkp

clinical effects. The prefrontal pyramidal projectionneurones synapse in the caudate nucleus withGABAergic neurones.6 These neurones in the caudatenucleus fall into two major functional categories: me-dium spiny projection cells and interneurones that con-nect local structures of the striatum.52 As both mediumspiny cells and interneurones in the caudate are inhib-itory and mediated by GABA,53 their loss could resultin the dysfunction in tonic regulation of the striatopal-lidal pathway and subsequent disinhibition of the glu-tamatergic thalamocortical pathway. This may result inexcitotoxic damage to the reciprocal regions of the OFCand DLPFC and further exacerbate underlying corticalpathology. Abnormalities in the GABAergic and gluta-matergic circuitry have long been implicated in thepathophysiology of depression, and reductions in neu-ronal density, as found in this study, could be associ-ated with a disruption in activity of the circuit in majordepression.
CONCLUSION
In summary, consistent with the previous studies inlate-life depression, this study reports further evi-dence of neuronal changes, which were not associ-ated with any changes in the overall glial population.These findings could be due to the influence of CVD,especially that affecting the white matter that dam-ages the axons of the pyramidal neurones as theyproject to the caudate nucleus.
The authors thank the NIHR Biomedical ResearchCentre for Ageing and Age-related Disease for their sup-port for this study. They also express gratitude to the staffat the Newcastle Brain Tissue Resource and the Neuropa-thology Department for their assistance in processing thetissue used in this study.
References
1. Beekman AT, Copeland JR, Prince MJ: Review of communityprevalence of depression in later life. Br J Psychiatry 1999; 174:307–311
2. Luijendijk HJ, van den Berg JF, Dekker MJ, et al: Incidence andrecurrence of late-life depression. Arch Gen Psychiatry 2008;65:1394–1401
3. Baldwin RC, O’Brien J: Vascular basis of late-onset depressivedisorder. Br J Psychiatry 2002; 180:157–160
4. Alexopoulos GS, Meyers BS, Young RC, et al: Clinically definedvascular depression. Am J Psychiatry 1997; 154:562–565
5. Salloway S, Malloy P, Kohn R, et al: MRI and neuropsychologicaldifferences in early- and late-life-onset geriatric depression. Neu-rology 1996; 46:1567–1574
6. Bonelli RM, Cummings JL: Frontal-subcortical circuitry and be-havior. Dialogues Clin Neurosci 2007; 9:141–151
7. Levy R, Friedman HR, Davachi L, et al: Differential activation ofthe caudate nucleus in primates performing spatial and non-spatial working memory tasks. J Neurosci 1997; 17:3870 –3882
8. Middleton FA, Strick PL: Basal ganglia output and cognition:evidence from anatomical, behavioral, and clinical studies. BrainCogn 2000; 42:183–200
9. Kim MJ, Hamilton JP, Gotlib IH: Reduced caudate gray mattervolume in women with major depressive disorder. Psychiatry Res2008; 164:114–122
10. Krishnan KR, McDonald WM, Escalona PR, et al: Magnetic reso-nance imaging of the caudate nuclei in depression. Preliminaryobservations. Arch Gen Psychiatry 1992; 49:553–557
11. Parashos IA, Tupler LA, Blitchington T, et al: Magnetic-resonancemorphometry in patients with major depression. Psychiatry Res1998; 84:7–15
12. Butters MA, Aizenstein HJ, Hayashi KM, et al: Three-dimensionalsurface mapping of the caudate nucleus in late-life depression.Am J Geriatr Psychiatry 2009; 17:4–12
13. Baumann B, Bielau H, Krell D, et al: Circumscribed numericaldeficit of dorsal raphe neurons in mood disorders. Psychol Med2002; 32:93–103
14. Bielau H, Trubner K, Krell D, et al: Volume deficits of subcorticalnuclei in mood disorders. A postmortem study. Eur Arch Psychi-atry Clin Neurosci 2005; 255:401–412
15. Buchsbaum MS, Wu J, DeLisi LE, et al: Frontal cortex and basalganglia metabolic rates assessed by positron emission tomogra-phy with [18F]2-deoxyglucose in affective illness. J Affect Disord1986; 10:137–152
16. Baxter LR Jr, Phelps ME, Mazziotta JC, et al: Cerebral metabolicrates for glucose in mood disorders. Studies with positron emis-sion tomography and fluorodeoxyglucose F 18. Arch Gen Psychi-atry 1985; 42:441–447
17. Hannestad J, Taylor WD, McQuoid DR, et al: White matter lesionvolumes and caudate volumes in late-life depression. Int J GeriatrPsychiatry 2006; 21:1193–1198
18. Steffens DC, Helms MJ, Krishnan KR, et al: Cerebrovascular dis-ease and depression symptoms in the cardiovascular health study.Stroke 1999; 30:2159–2166
19. Miguel-Hidalgo JJ, Baucom C, Dilley G, et al: Glial fibrillary acidicprotein immunoreactivity in the prefrontal cortex distinguishesyounger from older adults in major depressive disorder. BiolPsychiatry 2000; 48:861–873
20. Rajkowska G, Miguel-Hidalgo JJ, Wei J, et al: Morphometric evi-dence for neuronal and glial prefrontal cell pathology in majordepression. Biol Psychiatry 1999; 45:1085–1098
21. Khundakar A, Morris C, Oakley A, et al: Morphometric analysisof neuronal and glial cell pathology in the dorsolateral prefron-tal cortex in late-life depression. Br J Psychiatry 2009; 195:163–169
22. Rajkowska G, Miguel-Hidalgo JJ, Dubey P, et al: Prominentreduction in pyramidal neurons density in the orbitofrontalcortex of elderly depressed patients. Biol Psychiatry 2005;58:297–306
Khundakar et al.
Am J Geriatr Psychiatry ●:●, ●●● 2010 9
rich3/jgp-ajgp/jgp-ajgp/jgp99910/jgp0690-10z xppws S�1 4/13/10 8:15 4/Color Figure(s): F1 Art: JGP200678 Input-rkp

23. Davis S, Thomas A, Perry R, et al: Glial fibrillary acidic protein inlate life major depressive disorder: an immunocytochemicalstudy. J Neurol Neurosurg Psychiatry 2002; 73:556–560
24. Thomas AJ, Ferrier IN, Kalaria RN, et al: Cell adhesion moleculeexpression in the dorsolateral prefrontal cortex and anteriorcingulate cortex in major depression in the elderly. Br J Psychi-atry 2002; 181:129–134
25. Thomas AJ, Ferrier IN, Kalaria RN, et al: Elevation in late-lifedepression of intercellular adhesion molecule-1 expression in thedorsolateral prefrontal cortex. Am J Psychiatry 2000; 157:1682–1684
26. American Psychiatric Association: The Diagnostic and StatisticalManual of Mental Disorders.4th ed. Washington, DC, AmericanPsychiatric Association, 1994
27. Mai JK, Assheuer J, Paxinos G: Atlas of the Human Brain. SanDiego, Academic Press, 1997
28. Gundersen HJ, Bagger P, Bendtsen TF, et al: The new stereologi-cal tools: disector, fractionator, nucleator and point sampledintercepts and their use in pathological research and diagnosis.APMIS 1988; 96:857–881
29. Sterio DC: The unbiased estimation of number and sizes of arbi-trary particles using the disector. J Microsc 1984; 134:127–136
30. Gundersen HJ: The nucleator. J Microsc 1988; 151:3–2131. Dorph-Petersen KA, Nyengaard JR, Gundersen HJ: Tissue shrink-
age and unbiased stereological estimation of particle number andsize. J Microsc 2001; 204:232–246
32. Gundersen HJ, Jensen EB: The efficiency of systematic samplingin stereology and its prediction. J Microsc 1987; 147:229–263
33. Cruz-Orive L-M: Best linear unbiased estimators for stereology.Biometrics 1980; 36:595–605
34. Dorph-Petersen KA, Delevich KM, Marcsisin MJ, et al: Pyramidalneuron number in layer 3 of primary auditory cortex of subjectswith schizophrenia. Brain Res 2009; 1285:42–57
35. Howard CV, Reed MG: Unbiased Stereology: Three-Dimen-sional Measurement in Microscopy. Oxford, Bios ScientificPublishers, 1998
36. Thomas AJ, Ferrier IN, Kalaria RN, et al: A neuropathologicalstudy of vascular factors in late-life depression. J Neurol Neuro-surg Psychiatry 2001; 70:83–87
37. Cotter D, Landau S, Beasley C, et al: The density and spatialdistribution of GABAergic neurons, labelled using calcium bind-ing proteins, in the anterior cingulate cortex in major depressivedisorder, bipolar disorder, and schizophrenia. Biol Psychiatry2002; 51:377–386
38. Ongur D, Drevets WC, Price JL: Glial reduction in the subgenualprefrontal cortex in mood disorders. Proc Natl Acad Sci USA1998; 95:13290–13295
39. Thomas AJ, O’Brien JT, Davis S, et al: Ischemic basis for deepwhite matter hyperintensities in major depression: a neuropatho-logical study. Arch Gen Psychiatry 2002; 59:785–792
40. Cotter D, Mackay D, Landau S, et al: Reduced glial cell density andneuronal size in the anterior cingulate cortex in major depressivedisorder. Arch Gen Psychiatry 2001; 58:545–553
41. Cotter D, Hudson L, Landau S: Evidence for orbitofrontal pathol-ogy in bipolar disorder and major depression, but not in schizo-phrenia. Bipolar Disord 2005; 7:358–369
42. Hickie IB, Naismith SL, Ward PB, et al: Serotonin transporter genestatus predicts caudate nucleus but not amygdala or hippocampalvolumes in older persons with major depression. J Affect Disord2007; 98:137–142
43. Khundakar AA, Zetterstrom TS: Biphasic change in BDNF geneexpression following antidepressant drug treatment explained bydifferential transcript regulation. Brain Res 2006; 1106:12–20
44. Zafra F, Hengerer B, Leibrock J, et al: Activity dependent regulationof BDNF and NGF mRNAs in the rat hippocampus is mediated bynon-NMDA glutamate receptors. EMBO J 1990; 9:3545–3550
45. Matsumoto T, Numakawa T, Yokomaku D, et al: Brain-derivedneurotrophic factor-induced potentiation of glutamate and GABArelease: different dependency on signaling pathways and neuro-nal activity. Mol Cell Neurosci 2006; 31:70–84
46. D’Sa C, Duman RS: Antidepressants and neuroplasticity. BipolarDisord 2002; 4:183–194
47. Nair A, Vaidya VA: Cyclic AMP response element binding proteinand brain-derived neurotrophic factor: molecules that modulateour mood? J Biosci 2006; 31:423–434
48. Nibuya M, Nestler EJ, Duman RS: Chronic antidepressant admin-istration increases the expression of cAMP response elementbinding protein (CREB) in rat hippocampus. J Neurosci 1996;16:2365–2372
49. Chen B, Dowlatshahi D, MacQueen GM, et al: Increased hip-pocampal BDNF immunoreactivity in subjects treated with anti-depressant medication. Biol Psychiatry 2001; 50:260–265
50. O’Brien JT, Ames D, Schweitzer I, et al: Clinical and magneticresonance imaging correlates of hypothalamic-pituitary-adrenalaxis function in depression and Alzheimer’s disease. Br J Psychi-atry 1996; 168:679–687
51. Sapolsky RM, Romero LM, Munck AU: How do glucocorticoidsinfluence stress responses? Integrating permissive, suppressive,stimulatory, and preparative actions. Endocr Rev 2000; 21:55–89
52. Guehl D, Benazzouz A, Aouizerate B, et al: Neuronal correlates ofobsessions in the caudate nucleus. Biol Psychiatry 2008; 63:557–562
53. Tekin S, Cummings JL: Frontal-subcortical neuronal circuits andclinical neuropsychiatry: an update. J Psychosom Res 2002; 53:647–654
Analysis of Neuronal and Glial Cell Pathology
Am J Geriatr Psychiatry ●:●, ●●● 201010
rich3/jgp-ajgp/jgp-ajgp/jgp99910/jgp0690-10z xppws S�1 4/13/10 8:15 4/Color Figure(s): F1 Art: JGP200678 Input-rkp

JOBNAME: AUTHOR QUERIES PAGE: 1 SESS: 3 OUTPUT: Tue Apr 13 08:17:01 2010/rich3/jgp�ajgp/jgp�ajgp/jgp99910/jgp0690�10z
1—Please check whether the short title is OK as given.
2—Please spell out “DLCN,” “VMCN,” “CE,” and “WMH.”
3—Please provide manufacturer details for “SPSS software (version 15.0).”
4—Please spell out “PMI” in Table 1.
5—Kindly note that the citation of “Figures 1 and 2” has been changed to “Figures 2 and 3.”Please check.
6—Please note that the section title “conclusions” has been changed to “Discussion” as twosections were titled “Conclusion.”
7—Please provide department/division name (if any) for the affiliation.
AUTHOR QUERIES
AUTHOR PLEASE ANSWER ALL QUERIES 1