morphology, molecular characterisation and systematic position of hemiplectus muscorum zell, 1991...
TRANSCRIPT

Nematology, 2009, Vol. 11(5), 719-737
Morphology, molecular characterisation and systematic positionof Hemiplectus muscorum Zell, 1991 (Nematoda: Plectida)
Oleksandr HOLOVACHOV 1,!, Sven BOSTRÖM 2, Manuel MUNDO-OCAMPO 1,Irma TANDINGAN DE LEY 1, Melissa YODER 1, A. H. Jay BURR 3 and Paul DE LEY 1
1 Department of Nematology, University of California–Riverside, Riverside, CA 92521, USA2 Department of Invertebrate Zoology, Swedish Museum of Natural History, Box 50007, SE-104 05 Stockholm, Sweden
3 Department of Biological Sciences, Simon Fraser University, Burnaby, BC V5A 1S6, Canada
Received: 18 September 2008; revised: 11 December 2008Accepted for publication: 12 December 2008
Summary – Hemiplectus muscorum, the type and single representative of its genus, is redescribed on the basis of abundant new materialcollected in the UK, Canada and the USA using both light and scanning electron microscopy. The phylogenetic relationships of thespecies are inferred from morphological as well as molecular data. Maximum parsimony, neighbour joining and maximum likelihoodanalyses of small subunit (SSU) rRNA sequences support a position nested among the Plectidae. This conflicts with our morphologicalassumptions of character polarity, as it implies that the absence of a valvate bulb in Hemiplectus is a reversal rather than a plesiomorphy.The excretory system of Hemiplectus is described more precisely. Its structure is highly reminiscent of the system in Plectus but differsin the presence of an anterior and posterior pair of pseudocoelomocytes flanking the renette cell. A pair of lateral somatic setae isidentified as possible homologues of the ‘deirids’ in Plectus and Rhabditida. Measurements and descriptions are given of all fourjuvenile stages.
Keywords – Canada, cladistic analysis, morphology, morphometrics, phylogeny, rDNA, SEM, UK, USA.
The genus Hemiplectus Zell, 1991, with its only speciesHemiplectus muscorum Zell, 1991, was originally de-scribed on the basis of one female, three males and asingle juvenile. These specimens were collected in mossfrom the USA (Olympic National Park in the state ofWashington) and Europe (Orkney and the Azores islands).Subsequently, male paratypes were re-examined and de-scribed within the framework of a phylogenetic analy-sis of the order Plectida Malakhov, 1982 (Holovachov &Boström, 2004; Holovachov, 2006). In this paper we re-port on numerous specimens of H. muscorum collectedin the UK, USA and Canada. Our observations extendthe morphometric ranges and provide new information onthe morphology of the species. A molecular analysis wasmade by sequencing of the small subunit (SSU) rRNAgene. The resulting phylogenies conflict with our morpho-logical assumptions and form the basis for a re-evaluationof the phylogenetic relationships of the species.
! Corresponding author, e-mail: [email protected]
Materials and methods
COLLECTION AND PREPARATION FOR MICROSCOPY
Specimens from Olympic National Park collected in2005 were extracted by a modified wet funnel methodand fixed in DESS (Yoder et al., 2006). Material fromVancouver was extracted in the same way but fixed with4% formaldehyde solution. All other material was alreadyavailable mounted in glycerin on permanent glass orCobb’s slides. All curved structures were measured alongthe curved median line. Stoma length was measured to theposterior end of the visible lumen only since the positionof the radial tubes cannot be identified with confidence inmany specimens. The length of the supplements and theirposition (as distance from cloacal opening or anterior end)were measured as proposed by Zell (1993) for plectids.On the figures, setae in black are located on the side of
© Koninklijke Brill NV, Leiden, 2009 DOI:10.1163/156854109X404580Also available online - www.brill.nl/nemy 719

O. Holovachov et al.
the body facing the viewer and setae in white are locatedon the opposite side. For SEM studies, several adultsfrom Vancouver and one juvenile from Brecon, Wales,UK, were postfixed in 1% osmium tetroxide (OsO4) andtransferred to pure acetone through an acetone/distilledwater series. The specimens were critical point dried inliquid CO2, mounted on a stub and gold-plated undervacuum to a thickness of 200 Å in a sputter coater. Theywere examined in a XL30 FEG (adults) or Hitachi S-4300SEM at an accelerating voltage of 10 kV.
MOLECULAR ANALYSIS
Individual nematodes were removed from a fluores-cence slide and cut in a drop of worm lysis buffer(WLB: 50 mM KCl, 10 mM Tris-Cl pH 8.3, 2.5 mMMgCl2, 0.45% NP40 and 0.45% Tween 20, as describedin Williams et al., 1992), transferred to a microcentrifugetube with Proteinase K (60 µg ml"1) and frozen at "80#C.Genomic DNA was prepared from the lysate by incubat-ing the tubes at 64#C for 1 h followed by enzyme deactiva-tion at 90#C for 10 min and centrifugation at 13 226 g for3 min. Near full length small subunit (SSU) rDNA genewas amplified, sequenced and assembled as in TandinganDe Ley et al. (2007).
PHYLOGENETIC ANALYSIS OF RDNA SEQUENCES
The newly determined SSU rDNA sequences of H. mus-corum, Pakira orae Yeates, 1967 and Ceramonema reti-culatum Chitwood, 1936 were analysed against a back-ground of selected sequence data available in public data-bases for Plectida as well as representatives from otherorders within the class Chromadorea sensu De Ley andBlaxter (2004). We aligned the resulting set of 39 se-quences with a suitably broad range of automated align-ment algorithms including Clustal X, DIALIGN andMAFFT v6 (Morgenstern, 2004; Larkin et al., 2007; Ka-toh & Toh, 2008) as well as a manually constructedsecondary structure alignment. The secondary structurealignment was created by downloading 12 sequences withsecondary structure annotation from the European ribo-somal RNA Database (Wuyts et al., 2004), convertingthe annotation from the format of the secondary structureeditor DCSE (De Rijk & De Wachter, 1993; no longersupported) to a contemporary format compatible with theJAVA-based editor 4SALE (Seibel et al., 2006), import-ing the remaining 27 sequences into 4SALE, manually
adding secondary structure annotation for these patternedafter the DCSE model, and aligning the result by eye tomaximise apparent homology and minimise gaps.
All resulting alignments were analysed with likeli-hood algorithms in RAxML and PAUP* as well as max-imum parsimony (MP) and neighbour joining (NJ) al-gorithms in PAUP* (Ronquist & Huelsenbeck, 2003;Swofford, 2003; Stamatakis et al., 2008). RAxML wasaccessed through the RAxML Black Box online inter-face (http://phylobench.vital-it.ch/raxml-bb/) and ran atdefault settings. Before analysis in PAUP*, alignmentswere processed in ModelTest (Posada & Crandall, 1998)to determine the appropriate models of DNA substitu-tion and to estimate substitution rates, relative base fre-quencies, gamma distribution shape parameter and pro-portions of invariable sites were estimated. NJ distancecalculations were performed with 3000 bootstrap repli-cations. Minimum evolution analyses included correc-tions for multiple substitutions using the Log Determi-nant distance measure (Lockhart et al., 1994) and the gen-eral time reversible model (GTR, Rodriguez et al., 1990)based on ModelTest parameter estimates. For MP analy-ses, characters were independent, unordered and equallyweighted with gaps treated as missing data. To discoverthe most parsimonious trees, an analysis was performedusing heuristic search options with 100 replicates of ran-dom branch addition, branch swapping algorithm, andtree-bisection-reconnection. Clade support was evaluatedby bootstrap analysis from 1000 replicates. In this study,we regarded bootstrap values <65% to be weakly sup-ported and >85% as highly supported (see analyses of sig-nificance vs bootstrap values in Hillis and Bull, 1993). TheModelTest parameter estimates were also used in PAUP*for a non-bootstrap heuristic maximum likelihood (ML)search with 100 replicates of random branch addition.The resulting trees from all the analyses were then com-pared statistically using Kishino and Hasegawa (1989)pairwise tests with ML and MP optimality criteria, andthe Templeton test with MP optimality criterion. Clado-grams were examined with TREEVIEW X (Page, un-publ.; http://darwin.zoology.gla.ac.uk/$rpage/treeviewx/)and converted into graphic files for Adobe Illustrator™CS2.
In addition to tree-construction algorithms requiringprior construction of alignments, we also subjected ourset of 39 sequences to Dynamic Homology analysis withPOY 4.0 (Varón et al., 2008), resulting in simultaneousproduction of the phylogenetic tree and alignment. POYwas set to build 200 trees with swap threshold of 5 and
720 Nematology
Morphology, molecular characterisation and systematic position of Hemiplectus muscorum
Table 1. List of nematode SSU ribosomal RNA gene sequences (partial) used in this study.
Accession Genus/species Reference
AF034391 Acrobeloides sp. Goldstein et al. (1998)AY284697 Anaplectus grandepapillatus Holterman et al. (2006)AY593932 Aphanolaimus aquaticus Holterman et al. (2006)U94366 Ascaris lumbricoides Nadler and Hudspeth (1998)EF591330 Ascolaimus cf. elongatus Holterman et al. (2008)EF591331 Axonolaimus sp. Holterman et al. (2008)EF591325 Camacolaimus sp. Holterman et al. (2008)FJ474095 Ceramonema reticulatum NewFJ474096 Ceratoplectus armatus NewFJ474097 C. assimilis NewAY593931 Chronogaster boettgeri Holterman et al. (2006)AY593939 Cylindrolaimus communis Holterman et al. (2008)AF047889 Daptonema procerus Aleshin et al. (1998)EF591322 Deontolaimus papillatus Holterman et al. (2008)Y16913 Desmodora ovigera Kampfer et al. (1998)AJ966482 Diplolaimella dievengatensis Meldal et al. (2007)EF591329 Diplopeltula sp. Holterman et al. (2008)AY284684 Euteratocephalus palustris Holterman et al. (2006)AJ966485 Halomonhystera disjuncta Meldal et al. (2007)FJ474098 Hemiplectus muscorum NewEF591324 Leptolaimus sp. Holterman et al. (2008)AJ966491 Metadesmolaimus sp. Meldal et al. (2007)AY284686 Metateratocephalus crassidens Holterman et al. (2006)AY593938 Monhystera riemanni Holterman et al. (2006)EF591328 Onchium sp. Holterman et al. (2008)FJ474099 Pakira orae NewAF047888 Paracanthonchus caecus Aleshin et al. (1998)EF591320 Paraplectonema pedunculatum Holterman et al. (2008)AF037628 Plectus acuminatus Frisse et al. (unpubl.)AF036602 P. aquatilis Blaxter et al. (1998)AY284702 P . cf. parietinus Holterman et al. (2006)EF591326 Procamacolaimus n. sp. Holterman et al. (2008)EF591335 Sabatieria pulchra Holterman et al. (2008)EF591321 Setostephanolaimus spartinae Holterman et al. (2008)Y16922 Stilbonema majum Kampfer et al. (1998)AF083015 Teratorhabditis synpapillata Fitch et al. (1995)AF036607 Teratocephalus lirellus Blaxter et al. (1998)AY284695 Theristus agilis Holterman et al. (2006)AF202155 Tylocephalus auriculatus Félix et al. (2000)
static approximation of perturbance transformation in 15iterations, with ratchet up-weighting by a factor of 3for 20% of characters. Paracanthonchus caecus (Bastian,1865) Micoletzky, 1924 was designated as outgroup forall the above analyses. POY ran in Mac OS X 10.4.11 ona dual Intel processor Apple iMac 2.33 Ghz with 1.5 GbRAM, PAUP* ran in Mac OS 9.2 on a single G5 processorApple iMac 2 Ghz with 2 Gb RAM.
Hemiplectus muscorum Zell, 1991(Figs 1-6)
MEASUREMENTS
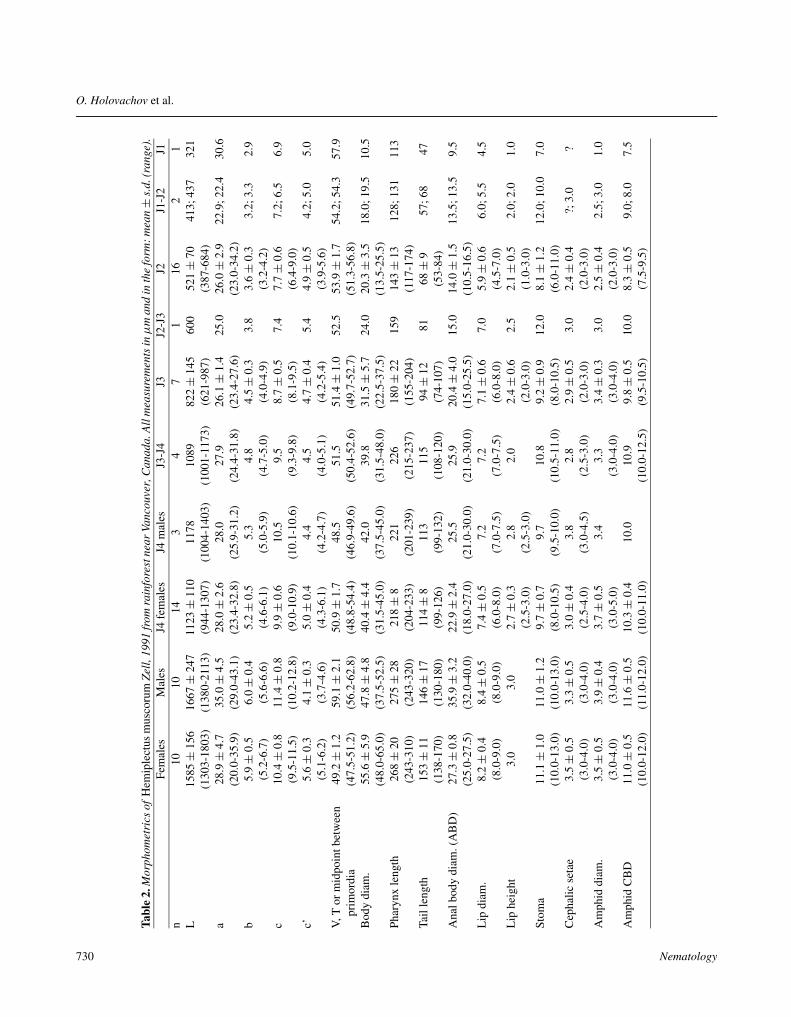
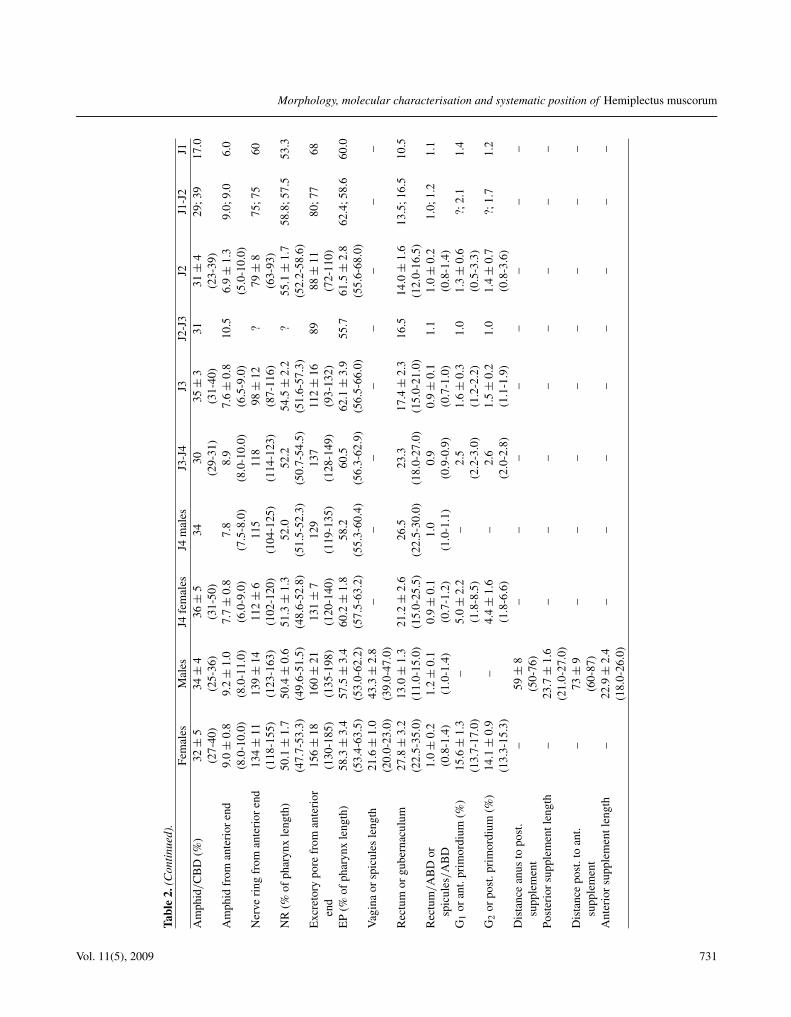
See Tables 2, 3.
Vol. 11(5), 2009 721
O. Holovachov et al.
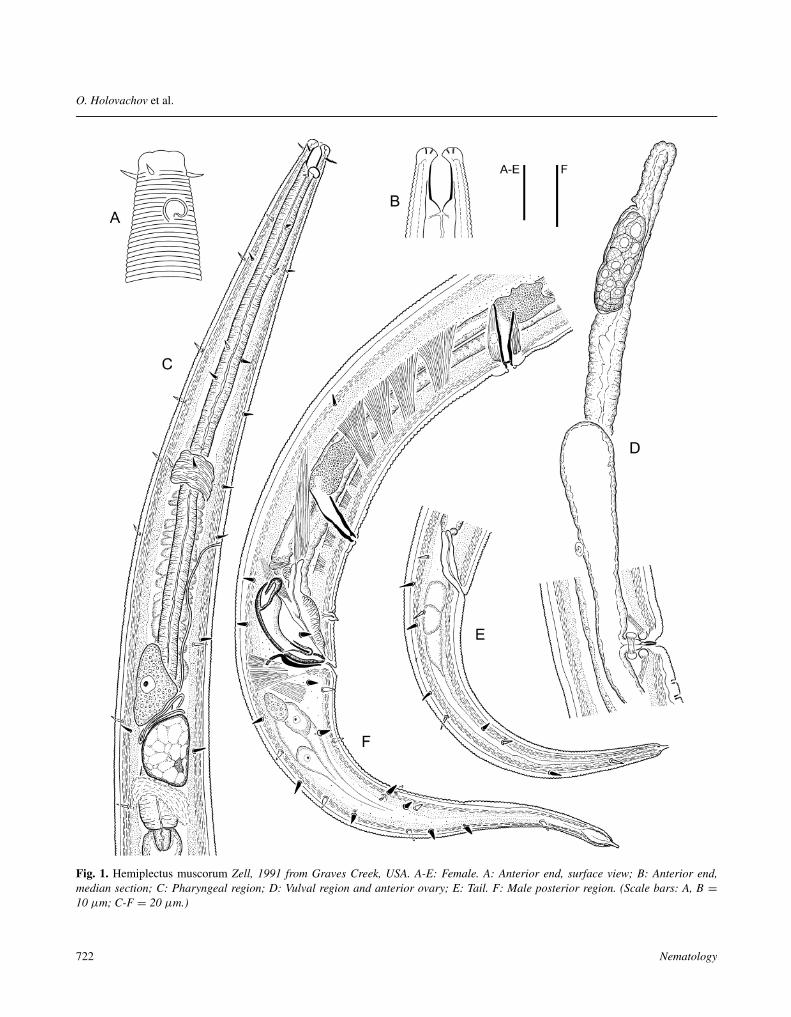
Fig. 1. Hemiplectus muscorum Zell, 1991 from Graves Creek, USA. A-E: Female. A: Anterior end, surface view; B: Anterior end,median section; C: Pharyngeal region; D: Vulval region and anterior ovary; E: Tail. F: Male posterior region. (Scale bars: A, B =10 µm; C-F = 20 µm.)
722 Nematology

Morphology, molecular characterisation and systematic position of Hemiplectus muscorum
Fig. 2. Hemiplectus muscorum Zell, 1991 from Windermere, UK. A-E: Female. A: Anterior end, combined view; B: Tail; C: Vulvalregion; D: Anterior ovary; E: Posterior part of pharynx. F, G: Male. F: Anterior end, combined view; G: Posterior region. (Scale bar:A-G = 20 µm.)
DESCRIPTION
Adult
Body more or less spindle-shaped, markedly taper-ing anteriorly in pharyngeal region and posteriorly ontail; ventrally or sigmoidally curved upon fixation, morestrongly curved ventrad in posterior part. Cuticle annu-lated; annuli 1.0-1.3 µm wide at mid-body, without orna-mentation. Lateral alae absent. Epidermal glands present,
two or three on each body side, connected with smallpapillae on cuticle surface, distinct body pores absent. So-matic setae present, not connected to epidermal glands,evenly distributed throughout body. Pair of lateral setaejust posterior to or level with nerve ring (one on eachbody side) shorter and plumper than somatic setae else-where and possibly homologous to deirids. Lip regiontruncate, slightly offset from rest of body as seen in LM,lips low and basally fused. First annulus appearing at level
Vol. 11(5), 2009 723
O. Holovachov et al.
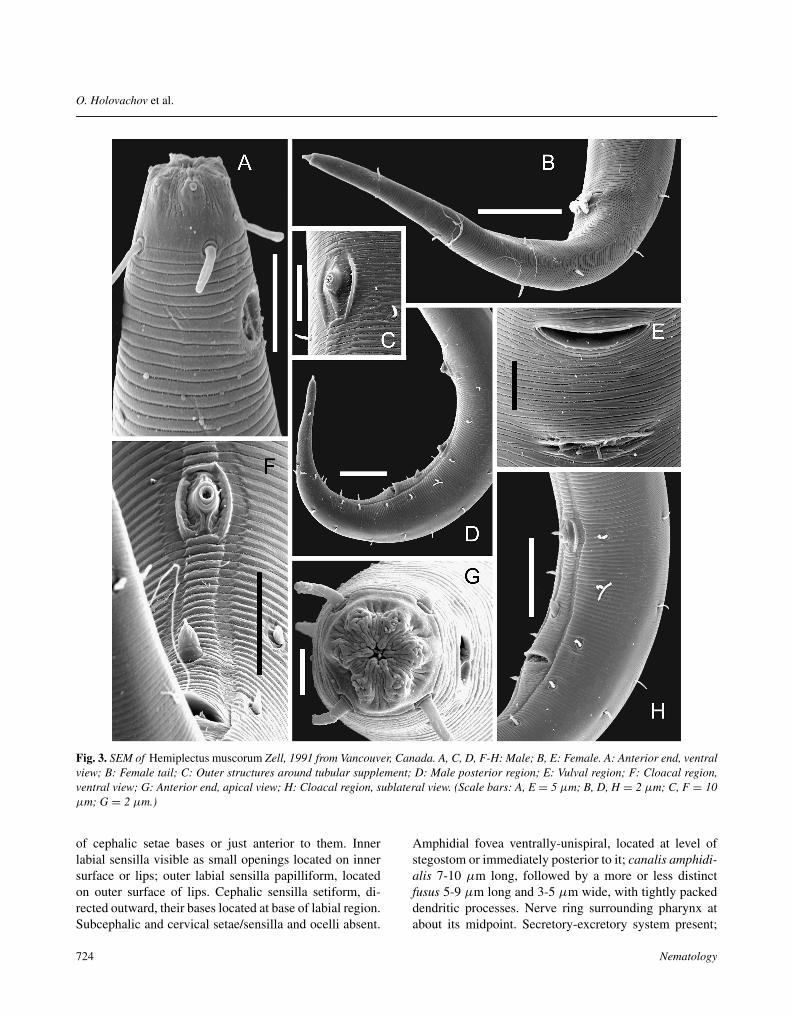
Fig. 3. SEM of Hemiplectus muscorum Zell, 1991 from Vancouver, Canada. A, C, D, F-H: Male; B, E: Female. A: Anterior end, ventralview; B: Female tail; C: Outer structures around tubular supplement; D: Male posterior region; E: Vulval region; F: Cloacal region,ventral view; G: Anterior end, apical view; H: Cloacal region, sublateral view. (Scale bars: A, E = 5 µm; B, D, H = 2 µm; C, F = 10µm; G = 2 µm.)
of cephalic setae bases or just anterior to them. Innerlabial sensilla visible as small openings located on innersurface or lips; outer labial sensilla papilliform, locatedon outer surface of lips. Cephalic sensilla setiform, di-rected outward, their bases located at base of labial region.Subcephalic and cervical setae/sensilla and ocelli absent.
Amphidial fovea ventrally-unispiral, located at level ofstegostom or immediately posterior to it; canalis amphidi-alis 7-10 µm long, followed by a more or less distinctfusus 5-9 µm long and 3-5 µm wide, with tightly packeddendritic processes. Nerve ring surrounding pharynx atabout its midpoint. Secretory-excretory system present;
724 Nematology

Morphology, molecular characterisation and systematic position of Hemiplectus muscorum
renette cell irregularly ovoid, enveloping distal part ofpharynx from ventral and lateral sides. Excretory pore lo-cated somewhat posterior to nerve ring; excretory canalstrongly cuticularised, extending from pore along ventralregion of pharynx towards renette cell, making two loops(one on each side of pharynx) before entering renette cell(point of entry not clear). Renette cell usually with verylarge nucleus, cytoplasm appearing coarsely reticulate.Two large triangular or elongate ovoid pseudocoelomo-cytes with large nuclei and more finely granular cytoplasmlocated anterior to renette cell, in very close proximity toloops of excretory canal on left/right dorsosublateral sec-tors of body. Base of pharynx enveloped just posterior torenette and ventrosublaterally by two additional cells ofvery similar appearance to dorsosublateral pair; these twopairs of pseudocoelomocytes possibly part of secretory-excretory system, but not clearly visible in some spec-imens, especially so for posterior pair. More distinct informalin-fixed specimens from Vancouver and in paratypematerial described by Holovachov and Boström (2004,Fig. 12A). Buccal cavity barrel-shaped, 8-10 µm longand 4-5 µm broad. Cheilostom usually constricted, lack-ing prominent sclerotisations. Gymnostom barrel-shaped,with parallel, sclerotised, short walls. Stegostom closed,linear, its slender muscular lining continuous with thatof corpus. An apparent cell boundary sometimes visibleat 2-3 µm posterior to barrel-shaped cavity, with deli-cate, rarely visible, radial tubes emerging at a shallow an-gle 5-12 µm posterior to barrel-shaped cavity. Pharynxcylindrical anteriorly, forming a slight expansion anteriorto nerve ring (see below), gradually widening again pos-teriorly; heavily muscularised with uniformly thickenedlumen throughout, lacking valves and bulbs. Additionaldiscontinuities corresponding to probable cell boundariessometimes visible at 20-28 µm (n = 19) and 35-48 µm(n = 12) from first discontinuity in lining of stegostom.Pharyngeal lumen penetrated by two subventral glandorifices just anterior to nerve ring level, at 85-107 µm(n = 19) from first discontinuity in lining of stegostom.Contour of pharynx somewhat expanded at this point, ap-pearing like corpus-isthmus junction in Plectus. Nucleusof dorsal pharyngeal gland distinct in some specimens, lo-cated at 10-15 µm (n = 6) from base of pharynx; onepair of nuclei of subventral pharyngeal glands occasion-ally seen at 21-40 µm (n = 6) from base of pharynx(see also Figure 12C in Holovachov and Boström, 2004).Cardia cylindrical, conical or hemispherical, embedded inintestine. Two irregularly spaced subventral pseudocoelo-mocytes with granular contents are usually visible at 46-
148 µm (n = 14) from pharynx base. Tail conoid, arcuateventrad. Three caudal glands present, their nuclei incau-dal. Single pseudocoelomocyte with granular cytoplasmlocated anterior to caudal glands. Spinneret functional.
Female
Reproductive system didelphic, amphidelphic; ovarybranches reflexed antidromously. Anterior ovary 40-128µm long, located on right-hand side of intestine, pos-terior ovary 45-118 µm long, located on left-hand side(n = 5). Oviduct a narrow U-shaped tube. Spermath-eca oval, empty in specimens from Olympic NationalPark (apparently young females), but filled with spermin specimens from Vancouver. Intrauterine spermatozoarounded-polygonal to oval, 5-10 µm long and 3-7 µmdiam. (n = 15), with comparatively large nucleolus-likeaggregations of chromatin. Vagina straight, 0.2-0.5 timesvulval body diam. long; pars proximalis vaginae encircledby single sphincter muscle; middle part of vagina con-nected to two pairs of oblique muscles extending towardslateral body sides (midlateral sectors), two muscles pass-ing from anterior wall of vagina anteriorly towards leftand right body sides each, two more muscles passing fromposterior wall of vagina posteriorly towards left and rightbody sides each. Epiptygmata present but delicate, weaklyrefringent. Intrauterine egg(s) not seen. Transverse cuticu-lar grooves or folds present in vulva region in specimensfrom Olympic National Park and Vancouver: 0-1 anteriorand 2-4 posterior to vulva; females from Mendocino andWindermere without cuticular grooves, but cuticular an-nulation along their ventral surface around vulva coarserand more irregular. Rectum 0.7-1.3 anal body diam. long,clearly flattened transversely; surrounded by three gland-like cells at junction with intestine: one flattened dorsalcell and two more rounded ventrosublateral cells. Bandsof muscles passing from dorsal wall of rectum (its prox-imal part) towards dorsal body wall. Six to eight caudalsetae arranged in one subventral and three subdorsal pairs.
Male
Reproductive system diorchic; anterior testis outstret-ched, 173-202 µm long; posterior one reflexed, 122-152µm long. Spermatids rounded-polygonal to oval, 6-11 µmlong and 4-8 µm diam. (n = 15), with comparatively largenucleolus-like aggregations of chromatin. Spicules paired,slightly asymmetrical, arcuate, with ovoid manubrium,narrowing shaft and thin velum. Spicular fenestra oval,anterio-laterally located on manubrium. Gubernaculumarcuate, plate-like, with dorsal apophysis less promi-nent in European males. Two similar midventral tubu-
Vol. 11(5), 2009 725
O. Holovachov et al.
726 Nematology

Morphology, molecular characterisation and systematic position of Hemiplectus muscorum
lar supplements and single midventral precloacal sensil-lum present, postcloacal sensilla absent. Each tubule con-nected to one large rounded anteriorly directed and finelygranular uninucleate gland-like cell, plus a smaller elon-gate cell with a flattened-triangular cell body positionedanteriodorsally of this large cell and connected to poste-rior rim of supplement by a slender process covering dor-sal surface of the large cell. All these cells lying exclu-sively on right-hand side of intestine. Distal part of eachtubule connected to ventral body wall via muscular cells,which are more distinct around anterior tubule (clearlyseen around posterior tubule only in one male). Midven-tral precloacal sensillum papilliform with a distinct nerveending, located 17.5-24 µm anterior to cloacal opening,i.e. usually at level of spicule manubrium. Caudal setaenumbering 11-20, arranged along subventral and subdor-sal body sectors.
J4 female
Similar to adult in most respects except for measure-ments and following features. Epidermal glands indis-tinct. Two pseudocoelomocytes along anterior part of in-testine and three caudal glands distinct. Somatic setaepresent, not connected to epidermal glands; four or fivepairs arranged along pharyngeal corpus, one lateral pair(‘deirids’) somewhat posterior to nerve ring, three pairs atlevel of secretory-excretory complex and a few scatteredalong remainder of body. Secretory-excretory system asin adults. Gonad primordia small, variable in size, fusedand multicellular, in older specimens vagina precursor aredistinct. Tail with four to eight caudal setae: one left sub-lateral seta along distal fifth of tail, one right sublateralseta along distal fourth of tail, one pair lying ventrosub-laterally at about middle of tail length and sometimes (notin all specimens) one to four subventral setae at level ofrectum.
J4 male
Similar to adult in most respects except for measure-ments and following features. Epidermal glands indis-tinct. Two pseudocoelomocytes along anterior part ofintestine and three caudal glands distinct. Somatic se-tae present, not connected to epidermal glands; fourpairs arranged along pharyngeal corpus, one lateral pair(‘deirids’) somewhat posterior to nerve ring, three pairsat level of secretory-excretory complex and a few scat-tered along remainder of body. Secretory-excretory sys-tem as in adults. Gonad primordia large, multicellular, de-veloped into testes and gonoduct precursors. Spicule pri-mordia visible on both sides of rectum. Tail with five orsix caudal setae: one left sublateral seta along distal fifthof tail, one right sublateral seta along distal fourth of tail,one pair lying ventrosublaterally at about mid-tail and oneto two (a pair) subventral setae at level of rectum.
J3
Similar to adult in most respects except for measure-ments and following features. Epidermal glands indis-tinct. Two pseudocoelomocytes along anterior part of in-testine and three caudal glands distinct. Somatic setaepresent, not connected to epidermal glands; three pairsarranged along corpus, one lateral pair (‘deirids’) some-what posterior to nerve ring, one pair at level of secretory-excretory complex and a few scattered along remainder ofbody. Secretory-excretory system as in adults. Gonad pri-mordia small, oval, usually multicellular, in some spec-imens fused. Tail with four to six caudal setae: one leftsublateral seta along distal fifth of tail, one right sublat-eral seta along distal fourth of tail, one pair lying ventro-sublaterally at about mid-tail and sometimes (not in allspecimens) one or two (a pair) subventral setae at level ofrectum.
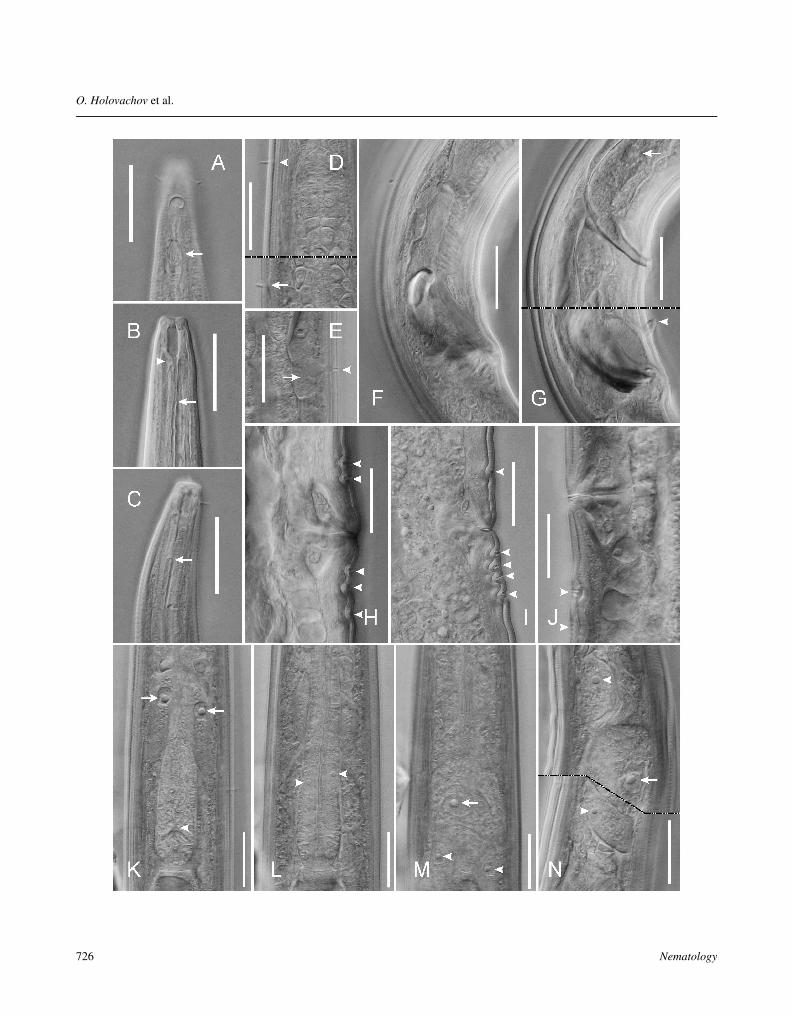
Fig. 4. Hemiplectus muscorum Zell, 1991 from Vancouver, Canada. A: Anterior end, surface view, arrow points at fusus amphidialis; B,C: Anterior end, longitudinal optical sections, arrowhead points at dorsal gland ampulla, arrows point at pharyngeal radial tubes; D:Nerve ring region, dorsal view, arrowhead points at somatic setae, arrow points at ‘deirid’; E: Epidermal gland (arrow) and somaticsensilla (arrowhead) along intestinal region; F, G: Male cloacal region at different focal planes, arrow points at nucleus of gland-likecell, arrowhead points at midventral precloacal sensillum; H-J: Vulval region, arrowheads point at transverse cuticular groves; K-M: Posterior part of pharyngeal region of same specimen at different focal planes in dorso-ventral view, showing dorsal pharyngealgland nucleus (arrowhead on K), nuclei of anterior dorsosublateral pair of pseudocoelomocytes (arrows on K), subventral pharyngealgland nuclei (arrowheads on L), renette nucleus (arrow on M) and nuclei of posterior ventrosublateral pair of pseudocoelomocytes(arrowheads on M); N: Posterior part of pharyngeal region in lateral view, arrowheads point at nuclei of anterior dorsosublateraland posterior ventrosublateral pair of pseudocoelomocyte, arrow points at renette nucleus. Figures D, G and N are composed of twooverlapping images (broken line = boundary) taken at somewhat different focal planes to superimpose/combine different features onthe same image. (Scale bars: A-N = 20 µm.)
Vol. 11(5), 2009 727
O. Holovachov et al.
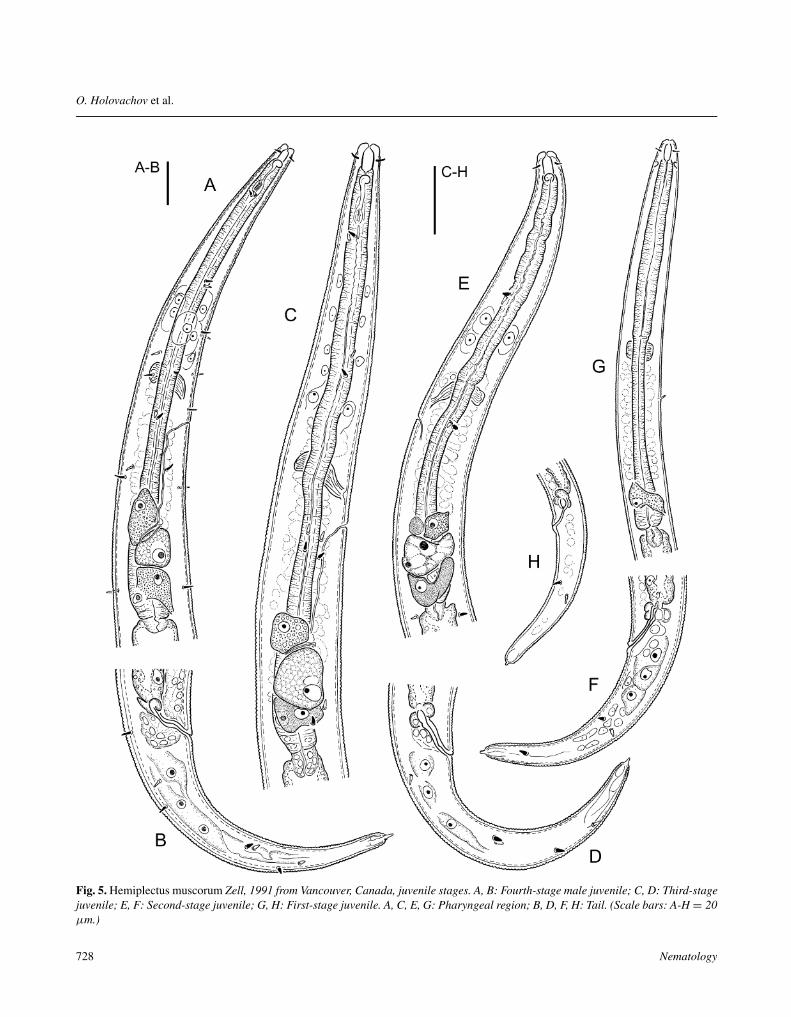
Fig. 5. Hemiplectus muscorum Zell, 1991 from Vancouver, Canada, juvenile stages. A, B: Fourth-stage male juvenile; C, D: Third-stagejuvenile; E, F: Second-stage juvenile; G, H: First-stage juvenile. A, C, E, G: Pharyngeal region; B, D, F, H: Tail. (Scale bars: A-H = 20µm.)
728 Nematology
Morphology, molecular characterisation and systematic position of Hemiplectus muscorum
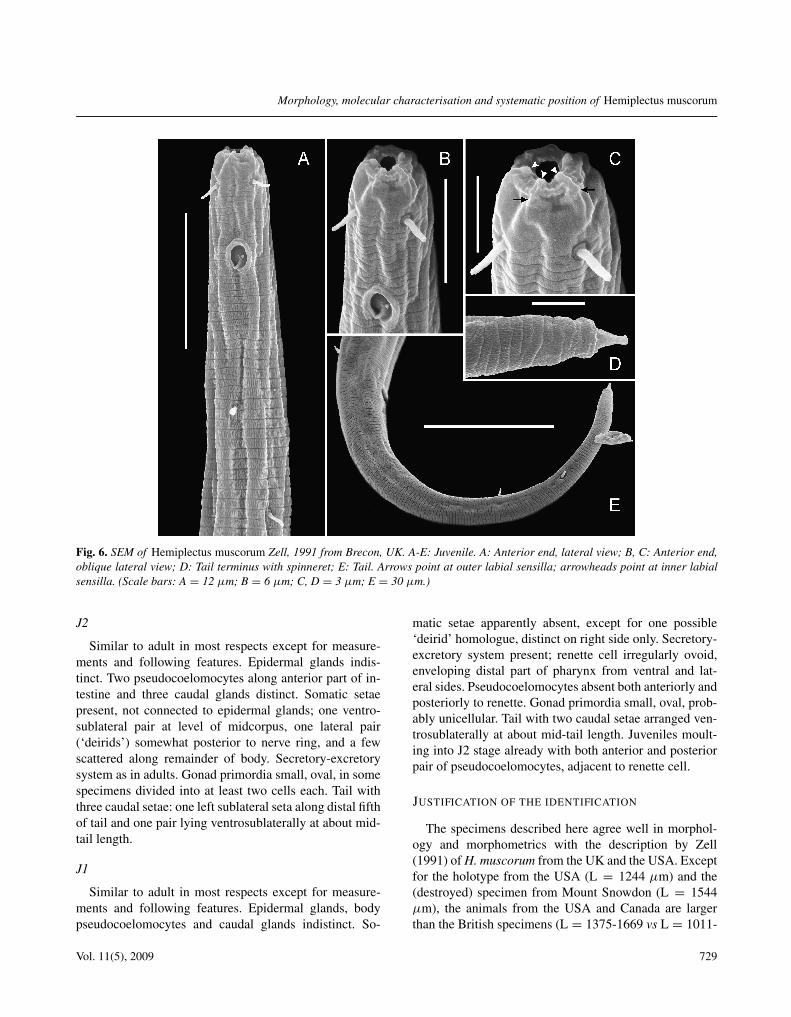
Fig. 6. SEM of Hemiplectus muscorum Zell, 1991 from Brecon, UK. A-E: Juvenile. A: Anterior end, lateral view; B, C: Anterior end,oblique lateral view; D: Tail terminus with spinneret; E: Tail. Arrows point at outer labial sensilla; arrowheads point at inner labialsensilla. (Scale bars: A = 12 µm; B = 6 µm; C, D = 3 µm; E = 30 µm.)
J2
Similar to adult in most respects except for measure-ments and following features. Epidermal glands indis-tinct. Two pseudocoelomocytes along anterior part of in-testine and three caudal glands distinct. Somatic setaepresent, not connected to epidermal glands; one ventro-sublateral pair at level of midcorpus, one lateral pair(‘deirids’) somewhat posterior to nerve ring, and a fewscattered along remainder of body. Secretory-excretorysystem as in adults. Gonad primordia small, oval, in somespecimens divided into at least two cells each. Tail withthree caudal setae: one left sublateral seta along distal fifthof tail and one pair lying ventrosublaterally at about mid-tail length.
J1
Similar to adult in most respects except for measure-ments and following features. Epidermal glands, bodypseudocoelomocytes and caudal glands indistinct. So-
matic setae apparently absent, except for one possible‘deirid’ homologue, distinct on right side only. Secretory-excretory system present; renette cell irregularly ovoid,enveloping distal part of pharynx from ventral and lat-eral sides. Pseudocoelomocytes absent both anteriorly andposteriorly to renette. Gonad primordia small, oval, prob-ably unicellular. Tail with two caudal setae arranged ven-trosublaterally at about mid-tail length. Juveniles moult-ing into J2 stage already with both anterior and posteriorpair of pseudocoelomocytes, adjacent to renette cell.
JUSTIFICATION OF THE IDENTIFICATION
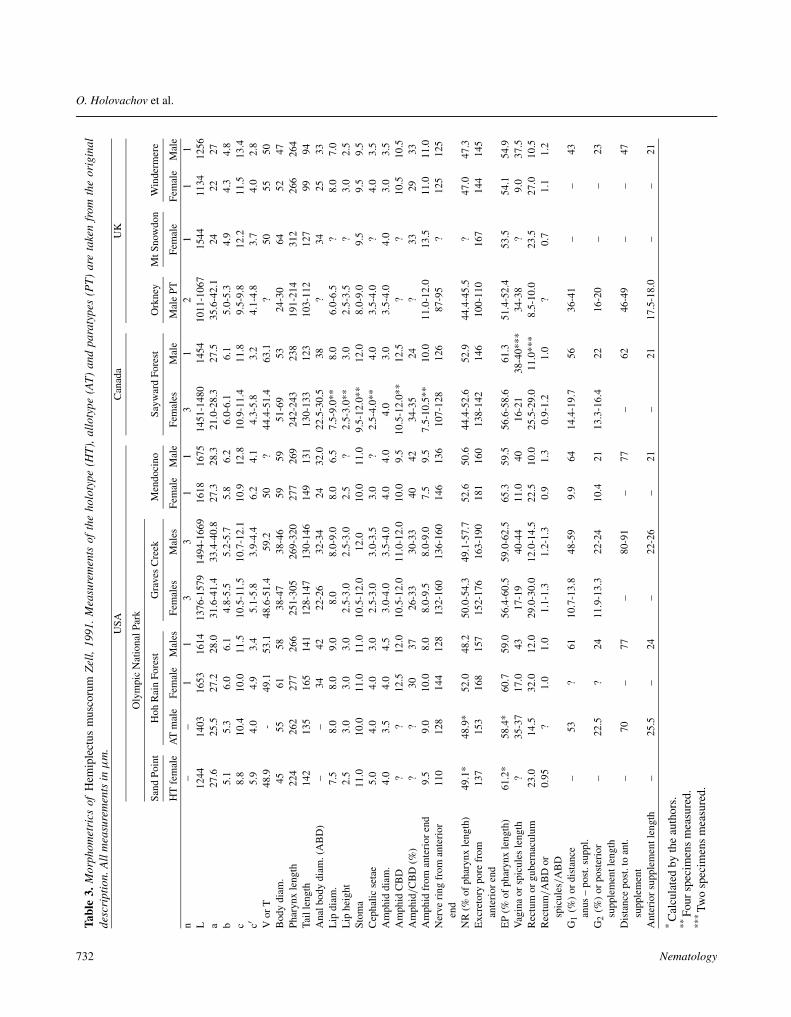
The specimens described here agree well in morphol-ogy and morphometrics with the description by Zell(1991) of H. muscorum from the UK and the USA. Exceptfor the holotype from the USA (L = 1244 µm) and the(destroyed) specimen from Mount Snowdon (L = 1544µm), the animals from the USA and Canada are largerthan the British specimens (L = 1375-1669 vs L = 1011-
Vol. 11(5), 2009 729
O. Holovachov et al.
Tabl
e2.
Mor
phom
etri
csof
Hem
iple
ctus
mus
coru
mZe
ll,19
91fr
omra
info
rest
near
Vanc
ouve
r,C
anad
a.A
llm
easu
rem
ents
inµ
man
din
the
form
:mea
n±
s.d.
(ran
ge).
Fem
ales
Mal
esJ4
fem
ales
J4m
ales
J3-J
4J3
J2-J
3J2
J1-J
2J1
n10
1014
34
71
162
1L
1585
±15
616
67±
247
1123
±11
011
7810
8982
2±
145
600
521
±70
413;
437
321
(130
3-18
03)
(138
0-21
13)
(944
-130
7)(1
004-
1403
)(1
001-
1173
)(6
21-9
87)
(387
-684
)a
28.9
±4.
735
.0±
4.5
28.0
±2.
628
.027
.926
.1±
1.4
25.0
26.0
±2.
922
.9;2
2.4
30.6
(20.
0-35
.9)
(29.
0-43
.1)
(23.
4-32
.8)
(25.
9-31
.2)
(24.
4-31
.8)
(23.
4-27
.6)
(23.
0-34
.2)
b5.
9±
0.5
6.0
±0.
45.
2±
0.5
5.3
4.8
4.5
±0.
33.
83.
6±
0.3
3.2;
3.3
2.9
(5.2
-6.7
)(5
.6-6
.6)
(4.6
-6.1
)(5
.0-5
.9)
(4.7
-5.0
)(4
.0-4
.9)
(3.2
-4.2
)c
10.4
±0.
811
.4±
0.8
9.9
±0.
610
.59.
58.
7±
0.5
7.4
7.7
±0.
67.
2;6.
56.
9(9
.5-1
1.5)
(10.
2-12
.8)
(9.0
-10.
9)(1
0.1-
10.6
)(9
.3-9
.8)
(8.1
-9.5
)(6
.4-9
.0)
c’5.
6±
0.3
4.1
±0.
35.
0±
0.4
4.4
4.5
4.7
±0.
45.
44.
9±
0.5
4.2;
5.0
5.0
(5.1
-6.2
)(3
.7-4
.6)
(4.3
-6.1
)(4
.2-4
.7)
(4.0
-5.1
)(4
.2-5
.4)
(3.9
-5.6
)V
,Tor
mid
poin
tbet
wee
n49
.2±
1.2
59.1
±2.
150
.9±
1.7
48.5
51.5
51.4
±1.
052
.553
.9±
1.7
54.2
;54.
357
.9pr
imor
dia
(47.
5-51
.2)
(56.
2-62
.8)
(48.
8-54
.4)
(46.
9-49
.6)
(50.
4-52
.6)
(49.
7-52
.7)
(51.
3-56
.8)
Bod
ydi
am.
55.6
±5.
947
.8±
4.8
40.4
±4.
442
.039
.831
.5±
5.7
24.0
20.3
±3.
518
.0;1
9.5
10.5
(48.
0-65
.0)
(37.
5-52
.5)
(31.
5-45
.0)
(37.
5-45
.0)
(31.
5-48
.0)
(22.
5-37
.5)
(13.
5-25
.5)
Phar
ynx
leng
th26
8±
2027
5±
2821
8±
822
122
618
0±
2215
914
3±
1312
8;13
111
3(2
43-3
10)
(243
-320
)(2
04-2
33)
(201
-239
)(2
15-2
37)
(155
-204
)(1
17-1
74)
Tail
leng
th15
3±
1114
6±
1711
4±
811
311
594
±12
8168
±9
57;6
847
(138
-170
)(1
30-1
80)
(99-
126)
(99-
132)
(108
-120
)(7
4-10
7)(5
3-84
)A
nalb
ody
diam
.(A
BD
)27
.3±
0.8
35.9
±3.
222
.9±
2.4
25.5
25.9
20.4
±4.
015
.014
.0±
1.5
13.5
;13.
59.
5(2
5.0-
27.5
)(3
2.0-
40.0
)(1
8.0-
27.0
)(2
1.0-
30.0
)(2
1.0-
30.0
)(1
5.0-
25.5
)(1
0.5-
16.5
)L
ipdi
am.
8.2
±0.
48.
4±
0.5
7.4
±0.
57.
27.
27.
1±
0.6
7.0
5.9
±0.
66.
0;5.
54.
5(8
.0-9
.0)
(8.0
-9.0
)(6
.0-8
.0)
(7.0
-7.5
)(7
.0-7
.5)
(6.0
-8.0
)(4
.5-7
.0)
Lip
heig
ht3.
03.
02.
7±
0.3
2.8
2.0
2.4
±0.
62.
52.
1±
0.5
2.0;
2.0
1.0
(2.5
-3.0
)(2
.5-3
.0)
(2.0
-3.0
)(1
.0-3
.0)
Stom
a11
.1±
1.0
11.0
±1.
29.
7±
0.7
9.7
10.8
9.2
±0.
912
.08.
1±
1.2
12.0
;10.
07.
0(1
0.0-
13.0
)(1
0.0-
13.0
)(8
.0-1
0.5)
(9.5
-10.
0)(1
0.5-
11.0
)(8
.0-1
0.5)
(6.0
-11.
0)C
epha
licse
tae
3.5
±0.
53.
3±
0.5
3.0
±0.
43.
82.
82.
9±
0.5
3.0
2.4
±0.
4?;
3.0
?(3
.0-4
.0)
(3.0
-4.0
)(2
.5-4
.0)
(3.0
-4.5
)(2
.5-3
.0)
(2.0
-3.0
)(2
.0-3
.0)
Am
phid
diam
.3.
5±
0.5
3.9
±0.
43.
7±
0.5
3.4
3.3
3.4
±0.
33.
02.
5±
0.4
2.5;
3.0
1.0
(3.0
-4.0
)(3
.0-4
.0)
(3.0
-5.0
)(3
.0-4
.0)
(3.0
-4.0
)(2
.0-3
.0)
Am
phid
CB
D11
.0±
0.5
11.6
±0.
510
.3±
0.4
10.0
10.9
9.8
±0.
510
.08.
3±
0.5
9.0;
8.0
7.5
(10.
0-12
.0)
(11.
0-12
.0)
(10.
0-11
.0)
(10.
0-12
.5)
(9.5
-10.
5)(7
.5-9
.5)
730 Nematology
Morphology, molecular characterisation and systematic position of Hemiplectus muscorum
Tabl
e2.
(Con
tinue
d).
Fem
ales
Mal
esJ4
fem
ales
J4m
ales
J3-J
4J3
J2-J
3J2
J1-J
2J1
Am
phid
/CB
D(%
)32
±5
34±
436
±5
3430
35±
331
31±
429
;39
17.0
(27-
40)
(25-
36)
(31-
50)
(29-
31)
(31-
40)
(23-
39)
Am
phid
from
ante
rior
end
9.0
±0.
89.
2±
1.0
7.7
±0.
87.
88.
97.
6±
0.8
10.5
6.9
±1.
39.
0;9.
06.
0(8
.0-1
0.0)
(8.0
-11.
0)(6
.0-9
.0)
(7.5
-8.0
)(8
.0-1
0.0)
(6.5
-9.0
)(5
.0-1
0.0)
Ner
veri
ngfr
oman
teri
oren
d13
4±
1113
9±
1411
2±
611
511
898
±12
?79
±8
75;7
560
(118
-155
)(1
23-1
63)
(102
-120
)(1
04-1
25)
(114
-123
)(8
7-11
6)(6
3-93
)N
R(%
ofph
aryn
xle
ngth
)50
.1±
1.7
50.4
±0.
651
.3±
1.3
52.0
52.2
54.5
±2.
2?
55.1
±1.
758
.8;5
7.5
53.3
(47.
7-53
.3)
(49.
6-51
.5)
(48.
6-52
.8)
(51.
5-52
.3)
(50.
7-54
.5)
(51.
6-57
.3)
(52.
2-58
.6)
Exc
reto
rypo
refr
oman
teri
or15
6±
1816
0±
2113
1±
712
913
711
2±
1689
88±
1180
;77
68en
d(1
30-1
85)
(135
-198
)(1
20-1
40)
(119
-135
)(1
28-1
49)
(93-
132)
(72-
110)
EP
(%of
phar
ynx
leng
th)
58.3
±3.
457
.5±
3.4
60.2
±1.
858
.260
.562
.1±
3.9
55.7
61.5
±2.
862
.4;5
8.6
60.0
(53.
4-63
.5)
(53.
0-62
.2)
(57.
5-63
.2)
(55.
3-60
.4)
(56.
3-62
.9)
(56.
5-66
.0)
(55.
6-68
.0)
Vag
ina
orsp
icul
esle
ngth
21.6
±1.
043
.3±
2.8
––
––
––
––
(20.
0-23
.0)
(39.
0-47
.0)
Rec
tum
orgu
bern
acul
um27
.8±
3.2
13.0
±1.
321
.2±
2.6
26.5
23.3
17.4
±2.
316
.514
.0±
1.6
13.5
;16.
510
.5(2
2.5-
35.0
)(1
1.0-
15.0
)(1
5.0-
25.5
)(2
2.5-
30.0
)(1
8.0-
27.0
)(1
5.0-
21.0
)(1
2.0-
16.5
)R
ectu
m/A
BD
or1.
0±
0.2
1.2
±0.
10.
9±
0.1
1.0
0.9
0.9
±0.
11.
11.
0±
0.2
1.0;
1.2
1.1
spic
ules
/AB
D(0
.8-1
.4)
(1.0
-1.4
)(0
.7-1
.2)
(1.0
-1.1
)(0
.9-0
.9)
(0.7
-1.0
)(0
.8-1
.4)
G1
oran
t.pr
imor
dium
(%)
15.6
±1.
3–
5.0
±2.
2–
2.5
1.6
±0.
31.
01.
3±
0.6
?;2.
11.
4(1
3.7-
17.0
)(1
.8-8
.5)
(2.2
-3.0
)(1
.2-2
.2)
(0.5
-3.3
)G
2or
post
.pri
mor
dium
(%)
14.1
±0.
9–
4.4
±1.
6–
2.6
1.5
±0.
21.
01.
4±
0.7
?;1.
71.
2(1
3.3-
15.3
)(1
.8-6
.6)
(2.0
-2.8
)(1
.1-1
.9)
(0.8
-3.6
)D
ista
nce
anus
topo
st.
–59
±8
––
––
––
––
supp
lem
ent
(50-
76)
Post
erio
rsup
plem
entl
engt
h–
23.7
±1.
6–
––
––
––
–(2
1.0-
27.0
)D
ista
nce
post
.to
ant.
–73
±9
––
––
––
––
supp
lem
ent
(60-
87)
Ant
erio
rsup
plem
entl
engt
h–
22.9
±2.
4–
––
––
––
–(1
8.0-
26.0
)
Vol. 11(5), 2009 731
O. Holovachov et al.Ta
ble
3.M
orph
omet
rics
ofH
emip
lect
usm
usco
rum
Zell,
1991
.Mea
sure
men
tsof
the
holo
type
(HT)
,allo
type
(AT)
and
para
type
s(P
T)ar
eta
ken
from
the
orig
inal
desc
ript
ion.
All
mea
sure
men
tsin
µm
.U
SAC
anad
aU
K
Oly
mpi
cN
atio
nalP
ark
Sand
Poin
tH
ohR
ain
Fore
stG
rave
sC
reek
Men
doci
noSa
ywar
dFo
rest
Ork
ney
MtS
now
don
Win
derm
ere
HT
fem
ale
AT
mal
eFe
mal
eM
ales
Fem
ales
Mal
esFe
mal
eM
ale
Fem
ales
Mal
eM
ale
PTFe
mal
eFe
mal
eM
ale
n–
–1
13
31
13
12
11
1L
1244
1403
1653
1614
1376
-157
914
94-1
669
1618
1675
1451
-148
014
5410
11-1
067
1544
1134
1256
a27
.625
.527
.228
.031
.6-4
1.4
33.4
-40.
827
.328
.321
.0-2
8.3
27.5
35.6
-42.
124
2227
b5.
15.
36.
06.
14.
8-5.
55.
2-5.
75.
86.
26.
0-6.
16.
15.
0-5.
34.
94.
34.
8c
8.8
10.4
10.0
11.5
10.5
-11.
510
.7-1
2.1
10.9
12.8
10.9
-11.
411
.89.
5-9.
812
.211
.513
.4c%
5.9
4.0
4.9
3.4
5.1-
5.8
3.9-
4.4
6.2
4.1
4.3-
5.8
3.2
4.1-
4.8
3.7
4.0
2.8
Vor
T48
.9-
49.1
53.1
48.6
-51.
459
.250
?44
.4-5
1.4
63.1
?50
5550
Bod
ydi
am.
4555
6158
38-4
738
-46
5959
51-6
953
24-3
064
5247
Phar
ynx
leng
th22
426
227
726
625
1-30
526
9-32
027
726
924
2-24
323
819
1-21
431
226
626
4Ta
ille
ngth
142
135
165
141
128-
147
130-
146
149
131
130-
133
123
103-
112
127
9994
Ana
lbod
ydi
am.(
AB
D)
––
3442
22-2
632
-34
2432
.022
.5-3
0.5
38?
3425
33L
ipdi
am.
7.5
8.0
8.0
9.0
8.0
8.0-
9.0
8.0
6.5
7.5-
9.0*
*8.
06.
0-6.
5?
8.0
7.0
Lip
heig
ht2.
53.
03.
03.
02.
5-3.
02.
5-3.
02.
5?
2.5-
3.0*
*3.
02.
5-3.
5?
3.0
2.5
Stom
a11
.010
.011
.011
.010
.5-1
2.0
12.0
10.0
11.0
9.5-
12.0
**12
.08.
0-9.
09.
59.
59.
5C
epha
licse
tae
5.0
4.0
4.0
3.0
2.5-
3.0
3.0-
3.5
3.0
?2.
5-4.
0**
4.0
3.5-
4.0
?4.
03.
5A
mph
iddi
am.
4.0
3.5
4.0
4.5
3.0-
4.0
3.5-
4.0
4.0
4.0
4.0
3.0
3.5-
4.0
4.0
3.0
3.5
Am
phid
CB
D?
?12
.512
.010
.5-1
2.0
11.0
-12.
010
.09.
510
.5-1
2.0*
*12
.5?
?10
.510
.5A
mph
id/C
BD
(%)
??
3037
26-3
330
-33
4042
34-3
524
?33
2933
Am
phid
from
ante
rior
end
9.5
9.0
10.0
8.0
8.0-
9.5
8.0-
9.0
7.5
9.5
7.5-
10.5
**10
.011
.0-1
2.0
13.5
11.0
11.0
Ner
veri
ngfr
oman
teri
or11
012
814
412
813
2-16
013
6-16
014
613
610
7-12
812
687
-95
?12
512
5en
dN
R(%
ofph
aryn
xle
ngth
)49
.1*
48.9
*52
.048
.250
.0-5
4.3
49.1
-57.
752
.650
.644
.4-5
2.6
52.9
44.4
-45.
5?
47.0
47.3
Exc
reto
rypo
refr
om13
715
316
815
715
2-17
616
3-19
018
116
013
8-14
214
610
0-11
016
714
414
5an
teri
oren
dE
P(%
ofph
aryn
xle
ngth
)61
.2*
58.4
*60
.759
.056
.4-6
0.5
59.0
-62.
565
.359
.556
.6-5
8.6
61.3
51.4
-52.
453
.554
.154
.9V
agin
aor
spic
ules
leng
th?
35-3
717
.043
17-1
940
-44
11.0
4016
-21
38-4
0***
34-3
8?
9.0
37.5
Rec
tum
orgu
bern
acul
um23
.014
.532
.012
.029
.0-3
0.0
12.0
-14.
522
.510
.025
.5-2
9.0
11.0
***
8.5-
10.0
23.5
27.0
10.5
Rec
tum
/AB
Dor
0.95
?1.
01.
01.
1-1.
31.
2-1.
30.
91.
30.
9-1.
21.
0?
0.7
1.1
1.2
spic
ules
/AB
DG
1(%
)or
dist
ance
–53
?61
10.7
-13.
848
-59
9.9
6414
.4-1
9.7
5636
-41
––
43an
us–
post
.sup
pl.
G2
(%)
orpo
ster
ior
–22
.5?
2411
.9-1
3.3
22-2
410
.421
13.3
-16.
422
16-2
0–
–23
supp
lem
entl
engt
hD
ista
nce
post
.to
ant.
–70
–77
–80
-91
–77
–62
46-4
9–
–47
supp
lem
ent
Ant
erio
rsup
plem
entl
engt
h–
25.5
–24
–22
-26
–21
–21
17.5
-18.
0–
–21
*C
alcu
late
dby
the
auth
ors.
**Fo
ursp
ecim
ens
mea
sure
d.**
*Tw
osp
ecim
ens
mea
sure
d.
732 Nematology

Morphology, molecular characterisation and systematic position of Hemiplectus muscorum
1256 µm), resulting in small differences in a few mea-surements that are not considered significant for speciesdifferentiation.
MATERIAL EXAMINED
Canada: British Columbia, Vancouver, local rainforest,seven moss species collected from tree trunks and rocks,namely Plagiothecium undulatum (Hedw.) Schimp.,Isothecium myosuroides Brid., Rhytidiadelphus loreus(Hedw.) Warnst., Hylocomium splendens (Hedw.)Schimp., Kindbergia oregana (Sull.) Ochyra, Dicranumfuscescens Turn. and Rhizomnium glabrescens (Kind.)Kop., February 2007, legit J. Burr; Vancouver Island,Campbell River Forest District, Sayward Forest dom-inated by second-growth, even-aged, 40-60 year Dou-glas fir trees, soil, October-November 2005 and October-November 2006, legit J. Addison. USA: Washington,Olympic National Park, Hoh Rain Forest Visitor Centerand Graves Creek, different species of mosses growing ontree trunks, 8 July 2006, legit J. Burr; California, Men-docino County, Cupressus pygmaea (Lemmon) Sarg., 6February 1980, legit A. Maggenti. UK: England, LakeDistrict, Windermere, Rydal Tell, Ambleside, mosses on astone, 12 June 2000 and Wales, Mount Snowdon, mosseson a stone, 16 June 2000, legit B. Sohlenius.
Results of alignment and phylogenetic analysis
The different alignment methods employed producedcharacter matrices that included between 1729 and 1809positions, including 632-652 parsimony-informative char-acters and 833-925 constant characters. Overall structureof the obtained trees is generally consistent in placing bothH. muscorum and Pakira orae inside the family Plecti-dae (Fig. 7). Resolution of relationships between majorclades of Plectida is poor, but there is strong support fortwo monophyletic clades corresponding respectively tothe families Camacolaimidae and Metateratocephalidae.Another strongly supported clade includes both Lepto-laimidae and Aphanolaimidae, but implies that the formerfamily is paraphyletic. Relationships between Plectidaeand Chronogastridae are not well resolved – they are usu-ally placed in a monophyletic clade but without significantsupport or clear resolution of branch order. Monophyly forPlectida is strongly supported, to the exclusion of C. reti-culatum which appears to be closer to Chromadorida andDesmodorida.
Relationships and systematic position
Zell (1991) placed Hemiplectus in the family Lepto-laimidae based on the shape of the excretory canal, thebasal expansion of pharynx, and the absence of deirids.Holovachov and Boström (2004) grouped Hemiplectusand Plectidae in one clade on the basis of three sharedderived characters: a long excretory canal, a reducedexcretory ampulla and a complex multicellular renettecell. Holovachov (2006) placed Hemiplectus together withStephanolaimus Ditlevsen, 1918 and SetostephanolaimusTchesunov, 1994 incertae sedis in Camacolaimoidea, al-though some features suggested a closer relationship withPlectoidea (both superfamilies in the order Plectida). Ourphylogenetic analyses of the SSU rDNA sequences of H.muscorum and other Plectida now lead to some surpris-ingly different hypotheses. Most importantly, Hemiplec-tus is always placed robustly within Plectidae and in manyanalyses even within the genus Plectus (Fig. 8). This po-sition suggests that the absence of a well defined basalbulb in Hemiplectus is not necessarily a plesiomorphiccondition, but could very well be a reversal from a Plec-tus- or Chronogaster-like ancestor with a well developedbulb. Also surprising is the position of P. orae with ro-bust support as sister taxon to Anaplectus grandepapilla-tus (Ditlevsen, 1928) Andrássy, 1973, since Pakira alsolacks a basal bulb and the genus was previously consid-ered to belong in its own family rather than inside Plecti-dae. However, in this case our phylogenetic analyses arenot sufficiently well resolved to imply unequivocally thatthe shape of the posterior pharynx in Pakira represents acharacter state reversal and not a plesiomorphy. Contraryto the system of De Ley and Blaxter (2004) and consistentwith the hypothesis of Holovachov and De Ley (2006), C.reticulatum is placed well outside Plectida.
Overall, our sequence analyses clearly indicate thatplectid classification needs substantial changes. Unfortu-nately, SSU rDNA sequence alone appears to lack the res-olution needed to pry apart relationships among Plectusspecies with sufficient clarity to pinpoint the exact posi-tion of Hemiplectus and to propose taxonomic changes atthis point. This makes it as yet impossible to revise genusdiagnoses – more taxa, as well as more genes, are neededto support any well-defined new system for Plectida.
Acknowledgements
The authors are grateful to Drs T. Panesar (Royal RoadsUniversity, Victoria, Canada) and B. Sohlenius (Swedish
Vol. 11(5), 2009 733

O. Holovachov et al.
Fig. 7. Phylogenetic relationships of the family Plectidae inferred by the RaxML using alignment of 18S rDNA of 39 nematode specieswith Paracanthonchus caecus (Bastian, 1865) Micoletzky, 1924 as an outgroup. Tree topology combines data obtained using secondarystructure-based, Clustal-based, Dialign-based and Mafft-based alignments and is consistent between all four alignments. Branchlength is given for the tree inferred from the secondary structure-based alignment. Support values for each clade are given for allfour alignments: secondary structure-based, Clustal, Dialign and Mafft, respectively. While other algorithms (maximum parsimony,neighbour-joining and dynamic homology) produce the same clades, greater variation in topology occurs among the different familieswithin Plectida and with respect to the placement of members of the Araeolaimida.
Museum of Natural History, Stockholm, Sweden) for-putting material for this study at their disposal and to DrR. C. Brooke (Simon Fraser University, Burnaby, Canada)
for identification of moss samples from Vancouver rain-forest.
734 Nematology

Morphology, molecular characterisation and systematic position of Hemiplectus muscorum
Fig. 8. Phylogenetic placement of Hemiplectus muscorum Zell, 1991 within the family Plectidae as inferred by the RaxML usingalignment of 18S rDNA of 39 nematode species with Paracanthonchus caecus (Bastian, 1865) Micoletzky, 1924 as an outgroup –terminal part of the phylogenetic tree given on Figure 7 for each type of alignment separately. Other algorithms (maximum parsimony,neighbour joining and dynamic homology) produce similar variation in topology; Hemiplectus and Pakira are never placed outsidethis clade.
References
ALESHIN, V.V., KEDROVA, O.S., MILYUTINA, I.A., VLADY-CHENSKAYA, N.S. & PETROV, N.B. (1998). Relationshipsamong nematodes based on the analysis of 18S rRNA genesequences: molecular evidence for monophyly of chromado-rian and secernentian nematodes. Russian Journal of Nema-tology 6, 175-184.
BLAXTER, M.L., De LEY, P., GAREY, J.R., LIU, L.X.,SCHELDEMAN, P., VIERSTRAETE, A., VANFLETEREN, J.R.,MACKEY, L.Y., DORRIS, M., FRISSE, L.M., VIDA, J.T. &THOMAS, W.K. (1998). A molecular evolutionary frameworkfor the phylum Nematoda. Nature 392, 71-75.
DE LEY, P. & BLAXTER, M.L. (2004). A new system for Ne-matoda: combining morphological characters with moleculartrees, and translating clades into ranks and taxa. In: Cook, R.
& Hunt, D.J. (Eds). Proceedings of the fourth internationalcongress of nematology, Tenerife, Spain, 8-13 June 2002, Ne-matology Monographs and Perspectives, Volume 2. Leiden,The Netherlands, Brill, pp. 633-653.
DE RIJK, P. & DE WACHTER, R. (1993). DCSE, an interactivetool for sequence alignment and secondary structure research.Computer Applications in the Biosciences 9, 735-740.
FÉLIX, M.-A., DE LEY, P., SOMMER, R.J., FRISSE, L.,NADLER, S.A., THOMAS, W.K., VANFLETEREN, J. &STERNBERG, P.W. (2000). Evolution of vulva developmentin the Cephalobina (Nematoda). Developmental Biology 221,68-86.
FITCH, D.H.A., BUGAJ-GAWEDA, B. & EMMONS, S.W.(1995). 18S ribosomal RNA gene phylogeny for some Rhab-ditidae related to Caenorhabditis. Molecular Biology andEvolution 12, 346-358.
Vol. 11(5), 2009 735

O. Holovachov et al.
GOLDSTEIN, B., FRISSE, L.M. & THOMAS, W.K. (1998).Embryonic axis specification in nematodes: evolution of thefirst step in development. Current Biology 8, 157-160.
HILLIS, D.M. & BULL, J.J. (1993). An empirical test of boot-strapping as a method for assessing confidence in phyloge-netic analysis. Systematic Biology 42, 182-192.
HOLOVACHOV, O. (2006). Morphology and systematics of theorder Plectida Malakhov, 1982 (Nematoda). Ph.D. Thesis,Wageningen University, Wageningen, The Netherlands, 246pp.
HOLOVACHOV, O. & BOSTRÖM, S. (2004). Morphology andsystematics of the superfamilies Leptolaimoidea Örley, 1880and Camacolaimoidea Micoletzky, 1924 (Nematoda: Plec-tida). Journal of Nematode Morphology and Systematics 7,1-49.
HOLOVACHOV, O. & DE LEY, P. (2006). Order Plectida. In:Abebe, E., Traunspurger, W. & Andrássy, I. (Eds). Freshwaternematodes: ecology and taxonomy. Wallingford, UK, CABIPublishing, pp. 611-647.
HOLTERMAN, M., VAN DER WURFF, A., VAN DEN ELSEN,S., VAN MEGEN, H., BONGERS, T., HOLOVACHOV, O.,BAKKER, J. & HELDER, J. (2006). Phylum-wide analysis ofSSU rDNA reveals deep phylogenetic relationships amongnematodes and accelerated evolution towards crown clades.Molecular Biology and Evolution 23, 1792-1800.
HOLTERMAN, M., HOLOVACHOV, O., VAN DEN ELSEN, S.,VAN MEGEN, H., BONGERS, T., BAKKER, J. & HELDER,J. (2008). Small subunit ribosomal DNA-based phylogenyof basal Chromadoria (Nematoda) suggests that transitionsfrom marine to terrestrial habitats (and vice versa) requirerelatively simple adaptations. Molecular Phylogenetics andEvolution 48, 758-763.
KAMPFER, S., STURMBAUER, C. & OTT, J. (1998). Phyloge-netic analysis of rDNA sequences from adenophorean nema-todes and implications for the Adenophorea-Secernentea con-troversy. Invertebrate Biology 117, 29-36.
KATOH, K. & TOH, H. (2008). Recent developments in theMAFFT multiple sequence alignment program. Briefings inBioinformatics 9, 286-298.
KISHINO, H. & HASEGAWA, M. (1989). Evaluation of the max-imum likelihood estimate of the evolutionary tree topologiesfrom DNA sequence data, and the branching order of theHominoidea. Journal of Molecular Evolution 29, 170-179.
LARKIN, M.A., BLACKSHIELDS, G., BROWN, N.P., CHENNA,R., MCGETTIGAN, P.A., McWILLIAM, H., VALENTIN, F.,WALLACE, I.M., WILM, A., LOPEZ, R., THOMPSON, J.D.,GIBSON, T.J. & HIGGINS, D.G. (2007). Clustal W andClustal X version 2.0. Bioinformatics 23, 2947-2948.
LOCKHART, P.J., STEEL, M.A., HENDY, M.D. & PENNY,D. (1994). Recovering evolutionary trees under a morerealistic model of sequence evolution. Molecular Biology andEvolution 11, 605-612.
MELDAL, B.H.M., DEBENHAM, N.J., De LEY, P., TANDIN-GAN DE LEY, I., VANFLETEREN, J.R., VIERSTRAETE,
A.R., BERT, W., BORGONIE, G., MOENS, T., TYLER, P.A.,AUSTEN, M.C., BLAXTER, M.L., ROGERS, A.D. & LAMB-SHEAD, P.J.D. (2007). An improved molecular phylogeny ofthe Nematoda with special emphasis on marine taxa. Molec-ular Phylogenetics and Evolution 42, 622-636.
MORGENSTERN, B. (2004). DIALIGN: multiple DNA and pro-tein sequence alignment at BiBiServ. Nucleic Acids Research32, W33-W36.
NADLER, S.A. & HUDSPETH, D.S.S. (1998). Ribosomal DNAand phylogeny of the Ascaridoidea (Nemata: Secernentea):implications for morphological evolution and classification.Molecular Phylogenetics and Evolution 10, 221-236.
POSADA, D. & CRANDALL, K.A. (1998). MODELTEST: test-ing the model of DNA substitution. Bioinformatics 14, 817-818.
RODRIGUEZ, F., OLIVER, J.L., MARIN, A. & MEDINA, J.R.(1990). The general stochastic model of nucleotide substitu-tion. Journal of Theoretical Biology 142, 485-501.
RONQUIST, F. & HUELSENBECK, J.P. (2003). MRBAYES3: Bayesian phylogenetic inference under mixed models.Bioinformatics 19, 1572-1574.
SEIBEL, P.N., MÜLLER, T., DANDEKAR, T., SCHULTZ, J. &WOLF, M. (2006). 4SALE – a tool for synchronous RNAsequence and secondary structure alignment and editing.BMC Bioinformatics 7, 498.
STAMATAKIS, A., HOOVER, P. & ROUGEMONT, J. (2008).A rapid bootstrap algorithm for the RAxML Web-Servers.Systematic Biology 57, 758-771.
SWOFFORD, D.L. (2003). PAUP*. Phylogenetic analysis usingparsimony (*and other methods). Version 4b10. Sunderland,MA, USA, Sinauer Associates.
TANDINGAN DE LEY, I., MUNDO-OCAMPO, M., YODER, M.& DE LEY, P. (2007). Nematodes from vernal pools inthe Santa Rosa Plateau Ecological Reserve, California I.Hirschmanniella santarosae sp. n. (Nematoda: Pratylenchi-dae), a cryptic sibling species of H. pomponiensis Abdel-Rahman & Maggenti, 1987. Nematology 9, 405-429.
VARÓN, A., VINH, L.S., BOMASH, I. & WHEELER, W.C.(2008). POY 4.0.2870. American Museum of Natural His-tory. Available online at http://research.amnh.org/scicomp/projects/poy.php
WILLIAMS, B.D., SCHRANK, B., HUYNH, C., SHOWNKEEN,R. & WATERSON, R.H. (1992). A genetic mapping systemin Caenorhabditis elegans based on polymorphic sequencetagged sites. Genetics 131, 609-624.
WUYTS, J., PERRIERE, G. & VAN DE PEER, Y. (2004). TheEuropean ribosomal RNA database. Nucleic Acids Research32, D101-D103.
YODER, M., TANDINGAN DE LEY, I., KING, I.W., MUNDO-OCAMPO, M., MANN, J., BLAXTER, M., POIRAS, L. & DE
LEY, P. (2006). DESS: a versatile solution for preservingmorphology and extractable DNA of nematodes. Nematology8, 367-376.
736 Nematology

Morphology, molecular characterisation and systematic position of Hemiplectus muscorum
ZELL, H. (1991). Hemiplectus muscorum n. gen., n. spec.(Nematoda, Leptolaimidae). Zoologischer Anzeiger 226, 298-306.
ZELL, H. (1993). Die Gattung Plectus Bastian, 1865 sensulato (Nematoda: Plectidae) – Ein Beitrag zur Ökologie,
Biogeographie, Phylogenie und Taxonomie der Plectidae.Andrias 11, 1-171.
Vol. 11(5), 2009 737