morphological differences among radopholus simile (cobb, 1893) thorne, 1949 populations
TRANSCRIPT

7 Rus~ Joumal of Nematology, 1999, 7 (2), 139 - 153
Morphological differences among Radopholus similis (Cobb, 1893) Thorne, 1949 populations
Gamal A.A. Elbadri*, Etienne Geraert** and Maurice Moens***
"'Agrlcu1tural Research Corporation, Wad Medoni, P.O. BOX 126, Sudan, **Rijksuniversit.ell Gent, Instituut voor Dierkunde, K.L. Ledegonckstroal 35, B-9000 Gent, Belgium,
***CLO-Departement Gewasbescherming, Burg. Van Gansbcrghelaan 96, B-9820 Merelbeke, Belgium.
Accepted for publication 31 August 1999
Summary. Eight Radopholus similis populations from tmee eontinenls and different hosts were compared morphologicaUy and morphometrically. ReQection of the posterior or anterior ovary or testis was common in aU the populati os. Vari bilily of most taxonomie eharacters was frequent. Many of the individuals from the lndonesian population had a forked tail end. Scanning microscopy showed that males and females [rom different populations had scveral morphological difTerences, especially in the number of anterior hypoptygmata in the males and annllles tcrminating the vulva of the females. Reslilts of th is study confinu that R. citrophilus is a ynonym of R. similis. Key words: banana, black pepper, morphology, morphometrics, ornamentals, Radopholus similis, SEM, taxonomy.
Scanning electron JllÎcroscapy (SEM) tudies on cauntries and hosts (Tabie 1). Roots ofinfected host Radopholus citrophilus and R. similis populations plants were cut into sma.IJ pieces, rinsed with water isolated from Anthurium andreaenum L. in Hawaii and placed on a sieve in a funnel in a mistifier and from Citrus aurantium L. ijl Flarida, revealed (Seinhorst, 1950). After 3-4 days Radopholus nemaseverai morphological differences between the two todes were collected and hand-picked, sUlface sterispeciec (e.g., the number of genital papWae at the lised with streptomycin sulphat 4 ppm, and culcloaca! spicular orifice and number of annul s at tmed on canot (Daucas carata) di ks (Moody et al., the vulva terminus Ul females (Huettel & Yaegashi, 1973) at 22°C for everaJ generations. Nematodes 1988). A simiJar SEM study of Radoplzolus popula were extracted from carrot disks placed ln a mistifier tions from banana growing in Africa, revealed ,ub as described. Aftel' 2-3 day mixed life stages were stantiaJ variation and overlapping of morphological collected, specimens were heat killed and fixed by characters previously considered useful for distin adding 4% hot fonnaldehyde, and transferred to gui hing R. citrophilus [rom R. similis, and it was anhydrous g1y erine according to Seinhorst's rapid concluded that R. citrophilus was a j nior synonym method (Seinharst, 1959 as modified by De Grisse, of R. similis (Valette et af., 1998; Elbadri et al., 1999). (969). Females and males were separated and moun
As both species are quarantule organisms in the ted on permanent slide (Cobb, 1918). Thirty adult European Vnion accurate identification is required. females and males were measured far each populaTo obtain more infonnation on the taxonomie status tion and all measurements and drawings were made of R. citrophilus, tJle morphology of an additional usiug a camera lueida attached to a light microeight Radopholus populations was studied. These scope. For study by scanning electron microscopy population originated from Australia Belgium, the (SEM), nematodes embedded in glycerine were Caribbean, Gennany, and Indonesia, and tJle results transfeued into a drop of glycerin to which disof the study are described here. tilled water was added gradually untü the nematodes
were in almost pme water. Ultrasonlc treatment was MATERIALS AND METHODS used for about 8-10 minutes to remove paIticles
adherÎJlg to tJle surface afthc nematodes. 1l1e nemaSpecinlcns of Radopholus (Tylenchida: Praty lodes wcre dehydrated by passing them througll
lenchldae) were obtauled from estabLished carrot disk ethanol coneentrations of25, 50, 75, 95, and 100% culture ofpopulatians originally isolated in different at two hour1y intervals, foUowed by all overnight
139

G.A.A. Elbadri et al.
Table 1. Origin of Radopholus similis populations lIsed in this stlldy,
Country Locality Host plant
J\ustrali CairrIS Banana Belgium Aalst Calathen makoyana
Belgium Leuven Anthun'um andreanum
CostaRica La Estrella Banana Cuba VillaClara Banana
Germany Hamburg Philodendron sp,
lndonesia Bangka Black pepper Panama Changuinola Banana
dehydmtion in 100% ethanol. Aftel' critical-point drying with carbon djox.idc, dried pecimens were sputt I' coated with a layer of gold and examined with a JEOL JSM-840 SEM at 15 kV.
RESULTS
Light microscopy Females (TabIe 2). The largest females were from
an Jndonesian population (average body lengtIl = 686 ~1l1) nnd th smallest from a population from Co taRica (average body length = 614 ~m). Head measurements were almost identicaJ in all populations, and ranged from 10.5 x4.9 ~m (Indonesia) to 9.3 x 4.2 ~m (C sta Rica). The number of head
11l1UIeS varied btween 3 and 4 in the Co ta Rican Panamanian, and Austrahan populations, 2-4 in th~ Cuban and lndon ian populations, and populatiol from ornamental plant hosts (Calatlzea makoyana, Anthurium andreanllm nnd Philodendron . p.) had only 3. The tylet length varied between 19.2 ~n
(Austral ia) to 17.8 ~m (Cuba), and tl1e DGO was situated near the, tylet base in most populations. The excretolY pore opened ventrally 1-2 annuJes behind the hemizorud. In most specimens the oesophageal glan ha tJuce nuclei, mostly in one extended gland lobe, but in some specimens of populatiol from C. makoyana and Cuba two separate lobes were pres nt. LateraJ fi Ids had 4 lines, trongly areolated in the ante.rior and posterior parts of the body and fajntly, partlally nreolated in the mid-body regi n. In all populations the vulvaJ opening was at 57-58% oftl1e body length. Ovaries were extended anteriody nnd posteriorly witl1 asingle row of oocytes , with reflection of the anterior and/or po terior ovary occurring in all populations. llle pelmatlleca was rounded t oval. Tail shape and length varied bOtll i~in al~d between populations, with th tail shape
bemg pomted to tapering with a smooth temUnus and coarse annulation (Figs. 1 & 2). Howev I 47% of the individuaJ in the Indone ia.n population had a tail h pe with forked end . The longest tail (8 1 ~lm) was present in tbe population from Cala/hea
makoyana, and the shortest (63.8 ~m) in the Costa Rican population.
Males (Tabie 3). Males were more slender and shorter than femaJes. 1l1e longest males (625.7 ~m)
were present in the population from Philodendron sp., and the shortest (552.7 ~m) in the population from A. andreanum. Heads w re rounded set ofT with weak cephalic sel rotisation. Tbe s~Ylet wa~ reduced and stylet kJlobs wem ab ent. Head measuremen showed little difference within and between populatioll . Lateral fjelds had 4lines, aS iJl females. The spicule length varied between populations being 16 to 21 ~m in the populations from Indonesia and C sta Rica, resp tively. llle gubemaculum was stout, slightly protruding from the cloaca, and almo t half of the picule lengtil. al gon ds were g n rally outstr t hed with < riabIe length and the g nital tract c n ined rod-like . perm. However, some specimens with flected gonads were present in all of the populations.
Male tail ends were thinner than those in the females. Tail length and shape vali d within alld betwe n population (Figs. I & 2). Male taü shape was g neralJy poi.nted to tap ring ith a smooth terminus, but ill th lndonesi n popul tioJl 24% of the individuaJs had forke rails (Fig. I). llle IOllgest male tails (82.3 ).lrn) werc present in the population from C. makoyana and the sJ ortc t ( 4.6 ~m) in the population (rom A. andreanum.
Scanning electron micro copy Females. Morphological differences in the head
regjon and the labiaJ disc werc bserved in ti males from aU populations tudied. The labial di.c was hexagonal (Fig. 3A) in the population from Co ta Rica, slightly hexagonal in the populations (rom A. andreanum, Philodendron sp., nnd C. makoyana (Figs. 6A, 7A & 8A), to rounded-elongate in the population from Panama and Ind ne. ia (Figs. 4A & 5A). 1l1e lateral lips ternUnated difTel ntly, depcnding upon the population. ln the Co ta Rican and Panamani.an populations, tl1e lat "ral li ps appeared to tcmullate bcfore tl1e end of th third head
140 =-=~ -l

Variation of Radopholus similis populations
.1 dl
f \
ij 1
A B C D E F G 85% 15% 76% 24% 44% 56% 47%
. \
H I J K L M N 53% 23% 77% 58% 42% 89% 11%
lJ
«
o P o R S T 97% 3% 76% 24% 88% 12%
Fig. 1. Tail ends of Radopholussimilis populations. A-J: Females; K-T: Males. A-Band K-L: Panama; C-D and M-N: Cuba; E-Fand O-P: CostaRica; G-H and Q-R: Indonesia; I-J and S-T: Australia. Figures indicate the % presence of the tail shape in the population.
141

..,. N
o » ~
Table 2. Morphometrics (in J,.t.m) of Radopholus similis females from eight populations. m 0= Pol
Country Costa Rica Cuba Panama Australia Indonesia Germany Belgium Belgium 0::J.
Area
Host
La Estrella
Banana
Villa Clara
Banana
Changuinola
Banana
Cairns
Banana
Bangka
Black pepper
Hamburg
Philodendron
Leuven
Anthurium
Aalst
Calathea
~
l:l:
L* 614±30.3 631±605 6818±595 647±44.6 686±35.7 6787±39 640±44.2 664±52.4 (550-680) (510-756) (542-819) (552-750) (612-752) (601-752) (572-727) (576-765)
Stylet 1ength 18.4±0.8 (17-20) 17.8±0.7 (17-19) 191±1.5 (17-23) 19.2±0.8 (18-20) 18.3±0.8 (16-20) 18.4±1.2 (20-16) 18.5±1.0 (16-20) 19±1.3 (17-22) a 25.7±2.6 265±19 27.3±3.1 267±3 265±2.1 27.4±2.3 28.8±2.1 28.1±2.3
(20.2-30) (21.9-29.5) (20.6-32.9) (20.4-32) (22.2~33) (22.4- 31.5) (24-32.4) (23-34) b 79±06 (6.5-9.1) 7.8±0.6 (6.8-9.4) 7.7±0.8 (6.2-10) 7.6±06 (6.7-9.3) 8.5±0.42 (7.6-9.0) 7.9±0.4 (7.2-9.1) 81±05 (7.3-8.9) 7.4±0.7 (6.3-9.3) b' 4.4±0.3 (3.8-5.2) 4.0±0.46 (3.3-4.8) 4.5±0.4 (3.6-5.3) 4.3±0.4 (3.8-5.2) 4.6±0.2 (4.2-5.1) 4.4±0.3 (3.9-5) 4.4±0.2 (3.9-4.9) 4.4±0.5 (3.7-5.7) c 9.7±1.0 (6.8-116) 9.6±0.7 (8.5-12.2) 95±0.8 (7.6-12.1) 9.8±0.9 (8.2-11.6) 8.9±0.5 (7.9-9.9) 9.6±0.4 (86-10.6) 9.8±0.8 (8.9-10.8) 8.3±0.7 (7.3-10.5) c 3.8±05 (28-5) 4.0±0.35 (3.0-46) 41±0.6 (3.1-6.2) 3.8±0.4 (3.1-4.7) 4.0±0.4 (3.2-4.8) 3.7± 03 (3.3-4.5) 4.0±0.4 (3.4-4.8) 4.6±0.5 (3.8-5.7) Head height 4.2±0.4 (4.0-5.0) 48±05 (4.0-6.0) 4.0±0.6 (2.5-5.0) 4.5±0.6 (40-6.0) 4.9±0.3 (4.0-5.0) 4.4±0.6 (3.0-5.0) 4.3± 0.6 (3.0-5.0) 4.2±0.5 (2.5-6.0) Head width 93±0.6 (8.0-10) 10±064 (9.0-11) 98±0.6 (9.0-11) 10.l±0.6 (9.0-11.0) 10.5±0.6 (9.0-11) 97± 0.7 (8.0-110) 9.4±06 (8.0-10.0) 10.l±0.6 (9.0-11.0) DGO 4.9±0.8 (4.0-6.0) 4.8±0.6 (4.0-6.0) 5.4±0.8 (4.5-7.5) 4.8±0.7 (4.0-6.0) 4.1±0.7 (3.0-5.0) 4.8±0.7 (4-6) 4.0±0.8 (2.5-6) 5.3±0.8 (3.5-7) Oeso length 785±69 (67-99) 807±58 (69-99) 88.6±10 (72-120) 85.4±6.4 (69-96) 81.4±3.8 (70-88) 85.6±5 (72-94) 79.7±42 (66-86) 898±6.1 (74-102) Oeso gL length 140.5±10.6 153.8±15.6 1536± 18.1 149.8±123 148.1±93 154.8±11 145.1±9.5 152.7±12.1
(116-161) (115-207) (119-204) (122-171) (128-165) (131-183) (122-166) (132-177)
Ex. pore 88.3±7.5 83.5±7.9 94.1± 11.3 911±6.8 897±5.1 87.3±6.9 88.1±5.6 82.8±5.4 (72-103) (59-65) (75-115) (69-101) (79-101) (74-104) (75- 100) (75-97)
m.b.w. 24.2±3.0 (19-33) 24±21 (21-29) 25.2±2.8 (20-32) 245±3.2 (20-33) 26±2.4 (21-30) 24.9±19 (22-29) 22.4±18 (19-26) 23.8±2.3 (20-29) b.w.a. 17.1±1.7 (14-21) 16.7±14 (15-19) 17.9±2.0 (13-22) 175±1.7 (15-21) 19.5±2.1 (16-24) 19.1±1.7 (16-22) 16.4±L5 (14-19) 17. 6±2.2 (15-24) Head-anus 548.3±31.6 565±55.9 610.8±55.3 580.8±42.6 608.3±327 608±36 572±39 583± 46.6
(491-612) (456-680) (485-728) (485 - 676) (546-668) (536-676) (509-653) (498-675) Head-vLl.1va 348.1±21.1 357.3±32.7 3933±375 371.8±31.3 388±23 380±24 361±26.3 375.5±30
(305-399) (293-405) (315-467) (300-442) (344-450) (336-420) (320-420) (324-428) V% 56.6± 1.5 567±18 57.7±18 57.5±2.4 56.8±2.5 55.9±1.0 56± 1.6 56.6±2.9
(54.1-60.7) (50.7 -59) (532-61) (51.1-61) (51-62) (53.3-58) (50.6-59) (53.3-67) V'% 63.5± 1.7 63.4±20 64.4±19 64±26 64±2.8 62.5± l.l 63±1.4 64.5±3.1
(59.9-67.7) (56.3-66) (59.7-67.7) (57 -68.1) (57.3-69) (60-64.5) (60.1-66.3) (60.4-76.8) Taillength 63.8±7.1 (52-85) 66.4±6.5 (52-76) 72.2±85 (53-91) 66.7±6.9 (55-84) 77. 7±5.7 (66-90) 71.1±3.5 (65-78) 65±5 (58-81) 80.9±8.9 (57-100) Gl 163±27.8 171.2±26.5 191±35.3 171±34 190±21 181.3±39 151±34.2 170.3±44
(130- 238) (131-225) (131-263) (135-266) (145-228) (120 -249) (115-226) ( 112-297) G2 150.6±25.7 1625±31.5 187±35.3 160.5±31 185±295 169.4±34 147.8±34 156.8±35
(120-241) (117-235) (122-269) (121-228) (120-244) (120-224) (105-230) (103-226)

Table 3. Morphometrics (in ~m) of Radopholus similis males from eight populations.
Country Costa Rica Cuba Panama Australia lndonesia Gennany Bclgium Belgium
Area La Estrella VLlla Clara Changuinola Caims Bangka Hamburg Leuven Aalst
Host Banana Banana Banana Banana Black pepper Philodendron Anthurium Calathea
L* 576.1±30.9 556.&±42.6 610±26.9 592.2±25 600.1±37.7 6257±26.7 552.7±32 603±39.4 (538-660) (450-648) (538-660) (543-681) (531-675) (575-670) (483-588) (546-723)
Stylet length 12.6±0.9 (11-14) 14±0.9 (12-16) 13.7±1.1 (11-16) 13.3±0.7 (11-14) 13.3±1.0 (12-16) 13±09 (12-15) 13±1.0 (l2-15) 13.2±0.9 (10-15)
a 32.1±2.5 31.7±4 32.6±3.2 31.3±3.4 32±2.4 34.4±3.6 33.8±3.3 36.7±3.0 (27-35.5) (24.5-40.5) (25.6-38.1) (25.1-40) (24.5-35) (26.4-41.5) (26.7-39.1) (31.8-42.5)
b 7.4±0.5 7.5±0.42 7.6± 1.1 7.5±0.3 8.3±0.5 7.8±0.4 7.8±OA 7.3±0.7
(6.1-8.9) (6.6-8.5) (5.0-9.6) (6.8·8.5) (7.4-9.3) (7.3-8.5) (7.0-9.0) (5.4-8.5)
b' 5.5±0.5 (4.8-6.9) 5.2±0.4 (3.7-5.7) 56±0.6 (4.0-6.6) 5.2±0.3 (4.6-5.9) 5.6±0.5 (4.8-6.8) 5.7±OA (4.9-6.5) 5.4±0.5 (4.6·6.7) 5.7±0.5 (4.3-6.6)
c 8.6±0.5 (7.8-9.6) 8.5±0.45 (7.8-9.4) 8.3±0.7 (6.8-10.2) 8.4±0.6 (7.5-9.8) 8.2±0.5 (7.0-9.5) 8.5±0.5 (7.6-9.4) 8.6±0.5 (7.6-9.6) 7.4±0.6 (6.3-87) c 4.7±OA (3.7-5.6) 4.9±0.44 (4-5.6) 52±0.6 (3.9-6.4) 5.1±0.4 (4.3-5.7) 5.2±0.5 (4.2-6.5) 5.2±0.4 (3.9-5.9) 51±0.4 (4.0-6.0) 6.0 ±0.5 (4.9-6.9)
Head hcight 6.1±0.7 (5.0-7.0) 62±0.6 (5-7) 56±0.6 (50-7.0) 5.6±0.5 (5.0-6.0) 5.6±0.6 (4.5-7.0) 5.8±0.6 (5-7) 5.4±OA (5.0-6.0) 5.7±0.7 (5-75) Head width 8.0±0.9 (70-90) 85±06 (7-9) 8.2±0.8 (7.0-9.5) 78±0.8 (6.0-9.0) 82±0.6 (7.0-9.0) 72±0.8 (7.0-8.0) 7.1±0.8 (6.0-9.0) 7.3±0.6 (6.0-9.0) Oeso length 78.2±7.9 74±4 82.U 13.4 79.5±3.3 72±4.0 80.2±3.6 71.3±5.2 83.2±6.4
(62-103) (66-85) (62-123) (72-88) (66-80) (73-90) (58-78) (73-101) Oeso gl. length 104.7±10.7 108±6 109±12.7 I 14.9±8.6 10S.3± 10 IIO±6.4 104. I± 11.7 107±7.2
(80-125) (97-123) (90-123) (9S-138) (S7-128) (97-130) (75-120) (83-126)
Ex. pore 882±6.7 (66-100) 833±5 (74-93) 867±8.8 (70-104) 90±3.7 (81-102) 85.3±5.0 (70-95) 88.1±4.3 (79-96) 83±6.8 (65-95) 86.7±5.7 (70-103)
m.b.w. 18.1±1.8 (16-22) 178±2.3 (14-23) 18.8±1.7 (16-23) 19.1±1.& (16-22) 18.8± 1.4 (16-22) IS.3±1.8 (15-22) 16.4±1.8 (14-22) 16.5±1.4 (14-20) -<b.w.a. 14.2±1.3 (11-18) 13.5±0.9 (12-15) 14.4± 1.6 (12-20) 14±1.0 (12-16) 14.2±0.9 (l3-16) 14.3±1.0 (12-17) 12.S±1.1 (11-16) 13.9±1.0 (12-19) t>:> ;:1. t>:>Taillength 66.8±43 (60-75) 65.5±59 (4S-76) 73.7±6.4 (60-90) 70.6±5.0 (60-SI) 73.4±4.9 (64-87) 74±5.0 (67 -85) 64.6±4.4 (55-74) 82.3±5.8 (70-96) ~
ö'Testis length 184±23.4 178±20 214.3±23.5 201.2±22.1 194±29.7 216.3±26 185.7±22 195±26.8 ::J
( .136-236) (131-215) (152-250) (165-246) (128-233) (128-245) (140-223) (104-236) o...., Spicule length 21±1.3 (18-24) 19.5±1.6 (17-24) 20.3±1.9 (17-23) 19.9±1.3 (18-23) 16± 1.4 (13-18) 20±0.8 (18-22) 19±1.4 (16-21) 18.5±1.9 (15-24) S' GubemacuJum 11.9±0.6 (10-13) 10.2±0.9 (9-12) 107±1.5 (8-14) 11.0±0.6 (10-12) 8.8±0.& (7.0- 10) I l± 0.7 (10-12) 10.4±0.8 (9-12) 10.4±0.8 (9-12) 2
~
c ~
List of abbreviations used. L, total body length; a, total body lengthjmaximum body width; m.b.w., maximum body width; Oeso length, ~
oesophagus length; b, total body lengthjdistance from anterior end to junction of oesophagus and intestine; Oeso gl. length, oesophagus gland '" length; b', total body lengthjdistance from anterior end to posterior end of oesophageal glands; c, total body lengthjtail length; c', tail lengthjbody ~:
width at anus; b.w.a., body width at anus; V%, distance from head end to vulvajbody length x 100; V'%, distance from head end to vulvajdistance ~
from head end to anus x 100; DGO, dorsal oesophageal gland arifice; Ex. pare, excretory pore; Gl, anteriar ovary length; 02, posterior ovary '0o
Iength. '0t: Ei' ~
ö' .j:o w
r;;

G.A.A. Elbadri et al.
)
!
A B c o E F
72% 24% 4% 51% 26% 23%
(
\\ \\
H J K L
95% 4% 1% 96% 2% 2%
RM 17% 3%68% 33% 1% 80%
Fig. 2. Tail end,> of femalcs and males of Radopho/us similis population. A- I: Females; J-R: Males. A-C and J- L: Phi/odendron; 0- F and M-0: Anthurium; G- land P-R: Calathea. Figurcs indicalc the % presence of thc tail shape in thc population.
J
144

VaIiation of Radopholus similis populations
Fig. 3. Scanning electron microscopy of a population of Radopholus similis from Cosla Rica. A: Female en-face view; B: Female hcad lalcral view; C-O: Vulva, entral view; E-H: Male cloacal oJifice, antelior view. Anows indicate head direction. Scale bar - 1 flm.
145

GAA. Elbadri et al.
Fig. 4. Scanning electron mieroscopy of a population of Radopholus similis from Panama. A: Female en-face view; 8: Female llead lateral view' C-O: Vulva, ventral view. E-H: Male cloacal orifice, anterior view. Arrows indicatc head direction. Scale bar - 1 ~m.
146
Variation of Radopholus similis populatiüns
,.
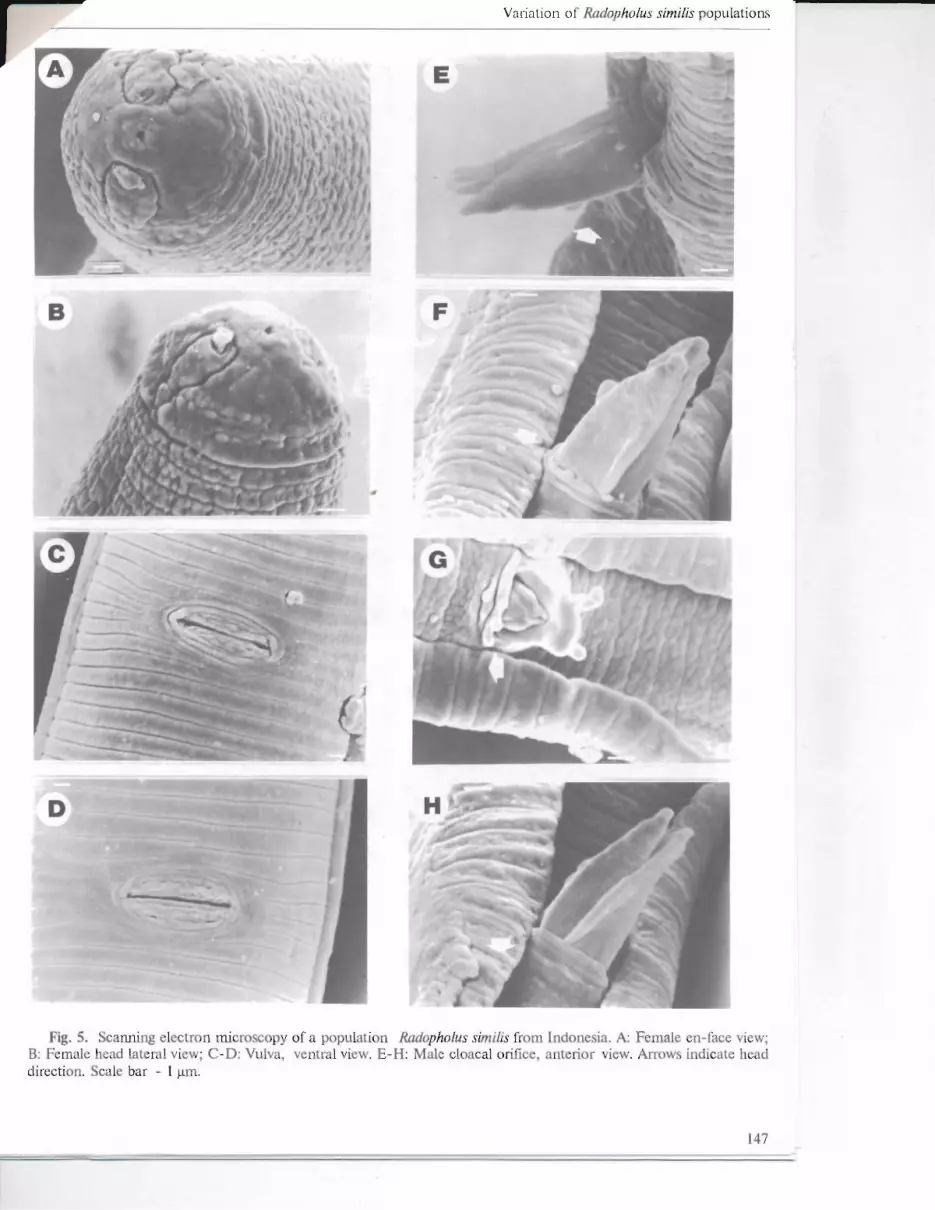
Fig. 5. ScatUling electron mi roscopy of a population Rcuiopholus similis fIom Indon sia. A Female en-face view; B: Fernale head lateral view; C- D: Vulva, ventral view. E- H: Male cloacal ürifice, anterior view. Arrows indicate head direction. Scale bar - 1 ~m.
147

G.A.A. Elbadri et al.
B
Fig. 6. Scanning electron mieroscopy of a population of Radopholus similis [rom A1lthurium andreanum. A: Female en-face view; B: Female llead lat ral view; C: Vulva, ventral iew. D-F: Male cloaca! orifice; D-E: Antelior view; F: Posterior view. Arrows indicate head direction. Scale bar - I Ilm.
148

Variation of Radopholus similis populations
Fig. 7. Scanning eleclron microscopy of a p pulation of Radopholus similis fr m Philodendron p. A: Female en-face view; B: Female hcad lateral view; C-O: Vulva, ventral view; E-H: Male cloacal orifice; E-G: Anterior view; H: Posterior view. Anows indicate head directi n. Scale bar - I llm.
149

G.AA Elbadri et al.
Fig. 8. Scanning electron microscopy of a population of Radopholus similis from Calathea makoyana. A: Fcmale en-face view; B: Head lateral view; C: Vulva ventral view. D-f: Male cloacal orifice; D-E: Anterior view; F: Posterior view. Anows indicate head direction. Scale bar - J Ilm.
150

Variation of Radopholus similis populations
annule (Figs. 3B & 4B); in the populations from A. andreanum and Philodendron sp. the lateraJ lips tenninated within the third annule (Figs. 6B & 7B); and in the populations from Indonesia and C. makoyana the lateral Ii ps telminated within the second annule (Figs. 5B & 8B).
SEM observations of the vulval area showed inter-population variation. In the population from Costa Rica (Figs. 3C-0) and Indonesia (Figs. 5C-0) three and two annules terminated at the vulva; in that [rom A. andreanum (Fig. 6C) and C. makoyana (Fig. 8C) two annules terrninated at the vulva; in the population from Philodendron sp. the number of annules teIminating at the vulva were one and two on each side, respectively, in the same specimen (Figs. 7C), or in some specimens only one annule was present on each side (Fig. 70). In another population from Panama one or two annules were present on each side (Figs. 4C-0).
Males. Most maIes in all populations had genital papilJac or hypoptygmata on the anterior cloacal aperturcs. However, these hypoptygmata varied in number both within and between populations, e.g. in the Costa Rican popuJation they varied between 5, 3, and 0 (Figs. 3E, F & H); in the Panamanian population between 1, 5, 7 and 4 (Figs. 4E-H); in the population from A. andreanum there were 6 and 2 hypoptygmata (Figs. 60-E); in the population from Philodendron sp. there were 7 and 6 (Figs. 7E-G); and in the popuJation from C. makoyana there were 2 and 3 (Figs. 8D-E).
DISCUSSION
Our study revealed the presence of substantiaJ variation in cuticular structures and in morphomctlics within and bctwccn eight Radopholus populations. The morphometrical data are similar to those obtained by Huettel et al. (1986) and Koshy et al. (1991), although in our study the spicule Jength in an Indonesian population was somewhat sho/ter, being 16±1.4 ~m. However, Van Weerdt (1958) reported a spicule length of 17.3±1.06 ~m
from a citlus popuJation and 15±1.3 ~m from a banana population . Sher (1968) repolted a spicuJe length of R. similis from a sugarcane population from Hawaii of 21 ~m.
The number of head allllUIes varied between 3, 2-4 and 3-4 in the various populations studied. In previous studies on R. similis from Africa the number varied between 3, 2-5, and 5 (Elbadri et al., 1999), and Valette et al. (1998) found a range of 2-5 in two African R. similis populations, whereas Williams & Siddiqi (1973) and Sher (1968) repolted 3-4 head annules for the species. Colbran (1970), when
describing 11 new Radopholus species reported the number of annuJes for R. rectus as being 3-4, whereas R. intermedius, R. inanis, and R. brevicaudatus had 3, R. ferax and R. dams had 4, R. vacuus and R. capitatus had only 2, and R. crenatus and R. megadoms did not have distinct annuJes. More recently Ryss and Wouts (1997) reported two new species of thc genus Radopholus, R. nelsonensis and R. kahikateae, as having 4-6 and 4-5 annules, respectivcly. From the present study, and our previous studies of R. similis popuJations we conclude that variation in the number of hcad annules exists both within and between the species of Radopholus. This is in contrast to the closely related genus Pratylenchus, where the number of head annules is considered a good taxonomie character (Sher & Allen, 1953; Corbett & CJark, 1983).
The oesophageal glands were present as an extended gland lobe, except in the populations from Cuba and from C. makoyana, each of which have two separate gland lobes in some specimens. The length of the postelior ovary is slightly shorter than the anterior ovalY and this confums previously repolted data obtained from African populations (Elbadri et al., 1999).
The morphology ofthe tail end varied within and between popuJations. In the Indonesian popuJation 47% of female and 27% of male tail ends had forked ends and this is the first report of this type of tail end for R. similis. In generaJ, the male tail tapered more than that ofthe female and this observation confiIms the findings of Siddiqi (1986).
Huettel & Yaegashi (1988) separated R. similis from R. citrophilus by severaJ minor morphological characteristics, which were used in combination with cytogeneticaJ differences (n=4 vs n = 5; Huettel & Oickson, 1981) and differences in isozyme patterns (Huettel et al., 1983a, J983b). Hahn et al. (1996), however, demonstrated that the chromosome number cannot be used as a diagnostic feature in R. similis. The morphological differences included the number of hypoptygmata on the cloaca-spicular orifice of males, the number of annules terminating the vulva, the shape of the head reg ion, and the position of the JateraJ lips. In our previous study (Elbadri et al., 1999), and that by Valette et al. (1998), and confinned in the present study, the values of these characteristics showed considerabIe overlap. The eight popuJations studied here could not be differentiated by ligl1t microscopy or SEM. As the populations studied originated from a large host range and have a very wide geographical distribution, our results confirm that R. citrophilus is a synonym of R. similis and thus the eight populations studied here are each R. similis. This synonimization has recently been supported by independent molecular
151

G.A.A. Elbadri et al.
and biochemical studies (Kaplan, 1999; Kaplan et al., 1999).
ACKNOWLEDGEMENTS
11le enior author thanks the Islamic D veIopment Bank for financial as i tance for this tudy. 11le authors thankDr. J. Pinoch t for making avaiJable ome of tl1e populations; Mr. Herdradjat for sending the Indone ian population from Bangka and Rita Van den Dries che and M. Bruyneel for their help with thc scanning electron microscopy.
REFERENCES
Cobb, N.A. 1918. Estimating the nemapopulation of soil, with special reil rence to the sugar beet and root gall
n mas, Heterodera schachtii Schmidt and H. radicicola
(Gr cf) Müller, and with de cri ption of Tylencholaimus aequalis n. sp. Agricultural Technical Circular I. 48 pp. US Departm nt f Agriculture.
Colbran, R.C. 1970. Study of plant and soU nematodes. 15. Eleven new specie. of Radopholus Th me and a new species of Radopholoides de Guiran (N m toda: Tylenehoidea) from Australia. Queensland Journalof Agricu/tural and Allimal Science.l 27: 437-46 .
Corbett, D.C.M. & Clark, S.A. 1983. SUlfaee features in the taxon my f Pratylenchu.l . pecie.. Revue de Nématologie. 6: 85-98.
De Grisse, A.T. 1969. Rédescription ollmodifications de quelque teclmiqlle utilisées dans I étude de nématode phytoparasitaires. Mededelingen van de Rijksfaculteit vall de Landbouwwetenschappen Gent 34: 315359.
Elbadri, GAA., Geraert, E. & Moens, M. 1999. MorphoI gical difference. among Radopholus populations from banana from Africa (N matoda: Tyl nch.ida). Joumal of Nematode Morphology and Systematic. 2: 1-16.
Halm, M.L. Wright, D.J. & Burrows, P.R. 1996. TIle ch.romo ome number in Radophollis similis a diagno. tic [I ature? Nematologica 42: 382-386.
Huettel, R.N. & Dickson, D.W. 1981. Karyology aud oogene is f Radopholus similis (Cobb) Thome. Journal of Nematology 13: 16-20.
Huettel, R. . & Yaegasld, T. J988. MorphoLogical difference b tw n Radopholus citrophilus and R. similis. Journalof Nematology 20: 150- 157.
Huettel. R. ., Dickson, D.W. & KAplan, D.T. 1983a. Biochemical identifieation f the two races of RadoplIolus similis by starclt gel electr phoresis. Journalof Nematology 15: 338-344.
Huettel, R.N., Dickson,� O.W. & Kaplan, D.T. 1983b. Bi chemical identificati n of tlte two raees of Radopholus similis by polyacrylamid gel eleetroph resis. JOllmal of Nematology 15: 345-348.
Huettel, RN., KapLan, D.T. & Dickson, D.W. 1986. Characterisation of a new burr wing nematode population, Radopholus citrophilus, from Hawail. Joumal ofNema
t%gy 18: 50-54. Kaplan, D.T. 1999. App!ying evolutionary and phyloge
netie speeies c ncepts to cLarify the taxon mie status of Radopholus similis. Proceedings of the American Society of Parasitologists and Society of Nematologists Meeting. 6-9 July ]999 Monterey, USA; 130.
Kaplan, D.T., Thomas W.K., Fri e L.M., Morris K. Sarah, J.-L. StantonJ.M. Speijer, P.R., Maria, D.H. & Opperman, C.H. 1999. Towards resolution of the Radopholus conllndrum. Proceedings of the American Society of Parasitologi and Nematologists Meeting, 6-9 July 1999, Monterey, USA: 163.
Koshy, P.K., Jasy, T., Mathew, J. & Nampoothiri, C.K. 1991. Morpltometric variabiLity in Radopho/us similis. Indian Journalof Nematology 21: 24-38.
Moody, E.H., Lownsbery, B.F. & Ahmed, J.M. 1973. Culture of th root-l ion nematode Praty/enchus vulnus on carr t disks. Joumal of Nematology 19: 125-134.
Ryss, A.Y. & Wouts, W.M. 1997. 1l1e genus Radopholus (Nematoda: Pratylenehidae) from native vegetation in New Zealand, with de clipti n of two new species. International Joumal of Nematology 7: 1-17.
Seïnhorst, J.W. 1959. A rapid method ror thc transfer of nematodes from Clx tive to aohydrous gLycerine. Nematologica 4: 67-69.
Seinhorst, J.W. 1950. De betekenis van de toestand v n de gr nd voor het optreden van aantasting door het stengelaaltje (Ditylenchu dipsaci Külm Filipjev). Tijdschrift voor Plantenziekten 52: 103-108.
Sher, S.A. 1968. Revü;ion ofthegenus Radopholus home, 1949 (Nematoda: Tylenchidae). Proceedings of the Helminthological Society of Washington 35: 219- 37.
Sher, S.A & Allen, M.W. 1953. Revision of the genus Pratylenchus (Nematoda: Tylenchidae). University of Califomia Publications in Zoology 57: 441-470.
Siddiqi, M.R. 1986. Tylenchida. Parasites of Plant and {nsec/s. Wallingford, UK., CAB International. 645 pp.
Valette, C. Mounport, 0., Nicole, M., arah J.-L. & Baujard, P. 1998. Scanning electron mieroscopy study of two African populalions f Radopholus simili.'> (Nematoda: Pratylenchidae) and proposal of R. citrophilus as junior synonym of R. similis. Fundamental and Applied Nematology 21: 139-146.
Van Weerdt, L.G. 1958. Studies on the bioLogy of Radopholus imilis ( Cobb, 189 ) Thome, 1949. Par1 Il. Morphological variation within and bctween progerues of single females. Nematologica 3: 184-196.
Williams, O.K.J. & Siddiqi, M.R. 1973. Radopholus similis. CJH Descriptiofl of Plant-Parasitic Nematodes, Set 2 No. 27, 4 pp.
152

r-:z~/~-----
Variation of Radopho/us simi/is populations
Elbadri G.A.A., Geraert E., Moens M. MOP<P0JIO[I1l{CCKMC pa3JHf'{]{51 MC)I(,Dy nonYJI51UvUIMJil
Radopho/us simi/is (Cobb, 1893) Thomc, 1949. Pe31OMe. TIpoBC.uCHO MOP<P0J[OfJil"lCCKOC Jil MOP<P0MCTPJil'{CCKOC cpaBIICHJilC 8-MJil nonYJI51UJilH
Radopho/us similis C Tpex KOHTYlHCHTOB Yl OT pa3JIJil'IHbIX X0351CB. 3afJil6 3a,1lHcro H IIcpC,D,HcrO
5lH"lIIYlKOB HJIH CCMCHHHKa 6blJT OTMe"lCH Y BCCX nonYJI51UYlH. lJacTo OTMe'laJlH Yl3MCH"lYlBOCTb no
OCIIOBHblM TaKCOHOMI1"lCCKHM I1pYl3HaKaM. MHorl1C oc06Yl 113 Yl]-[.IJ:OI [C3JilHCKOH nonYJr5II(YlYl H:\1eJIJil
npYlOCTpmOIllI1HC5I XBOCTOBOH KOHeu. 113Y'leHl1e B C3M 110Ka3aJlO, "lTO caMUbl H caMKH H3
paJJIH"lHblX 110rrYJI5IUYlH OTJlWlalOTC5I HeKOTopblMH MOp<pOJIOm"leCKI1MI1 OC06CHHOCTlIMJil, oc06eHHo
B '{I1CJIe nepe.LO-Il1X fl1nOrrmrM y caMUOB Jil llYlCJIY KOJIeu KYTYl K)'JI bI OKp)')KaIOlllYlX BYJlbBY Y caMOK.
Pe3YJIbTaTbI I1CCJIe.uOBaHI151 nO.IJ:TBep)l(Jla lOT, '{TO R. citrophi/us lIBJIlIeTC51 CI1HOHJ1MOM R. similis.
153