more than multiple introductions: multiple taxa contribute to the genesis of the invasive...
TRANSCRIPT

Journal of Systematics and Evolution 00 (00): 1–13 (2012) doi: 10.1111/j.1759-6831.2012.00222.x
Research Article
More than multiple introductions: Multiple taxa contribute to thegenesis of the invasive California’s wild artichoke thistle
Janet LEAK-GARCIA Jodie S. HOLT Seung-Chul KIM Lisa MU Jose A. MEJIASNorman C. ELLSTRAND
∗
(Department of Botany and Plant Sciences, University of California, Riverside, CA 92521, USA)
Abstract The history of some invasive species is so complex that their origins can be difficult to determine. Oneexample of such invasive species is the California invasive known as “wild artichoke thistle” (Cynara cardunculusvar. sylvestris), found in natural and disturbed ecosystems. Wild artichoke thistle is a Mediterranean native andthe progenitor of two domesticated horticultural taxa, artichoke and cardoon. Different hypotheses regarding theorigins of California plants have included introductions by 19th century Italian immigrants and the de-domestication(evolutionary reversion to wild-type morphology) of feral (escaped, free-living) cultivars. Using microsatellitemarkers, we compared the genetic constitutions of 12 artichoke thistle populations in California with possibleprogenitor populations: 17 Spanish and Italian wild populations and eight different artichoke and cardoon cultivars.Each California population was compared with its putative progenitors using STRUCTURE analysis. Our resultssuggest that California’s artichoke thistle populations are polyphyletic. Surprisingly, two-thirds of California’spopulations closely matched populations from the Iberian Peninsula. Three populations matched domesticatedartichoke. One population appears to have wild and cultivar hybrid ancestry. Alleles specific to Italian populationswere found at low frequencies in some California plants, suggesting that Italian wild plants may have been inCalifornia, but have left a trivial genetic legacy. Given that the de-domesticated plants in this study appear to be asinvasive as the wild taxon, we conclude with a discussion of the role that ferality and de-domestication may have inplant invasions.Key words artichoke thistle, Cynara cardunculus, de-domestication, ferality, invasive species, multiple origins.
Early studies that focused on determining the ori-gin of a biological invasion generally assumed a singlepoint of introduction stemming from a single source lo-cation (see Baker & Stebbins, 1965). More recently, theorigins of biological invaders have often been shownto be more complicated. In many cases, they are theculmination of a series of tenuous events and circum-stances that give rise to a unique evolutionary history(Ellstrand & Schierenbeck, 2000; Sakai et al., 2001;Kolbe et al., 2004; Hierro et al., 2005; Novak, 2007).In the last decade evidence has accumulated to suggestthat invasions are frequently the result of multiple intro-ductions of a species over time, and often from multiplegeographic locations (e.g. Kolbe et al., 2004; Durkaet al., 2005; Williams et al., 2005; Lavergne & Molof-sky, 2007; Novak, 2007; Cameron et al., 2008; Darlinget al., 2008; Dlugosch & Parker, 2008a; Ficetola et al.,2008; Rosenthal et al., 2008; Ghabooli et al., 2011).
Multiple introductions from diverse regions, co-introduction of sister taxa, and hybridization with in-
Received: 17 April 2012 Accepted: 28 June 2012∗ Author for correspondence. E-mail: [email protected]. Tel.: 1-951-827-
4194. Fax: 1-951-827-4437.
troduced or native relatives can result in a demograph-ically and genetically isolated set of immigrant popu-lations or in a loose network of populations (Bartsch& Ellstrand, 1999; Ellstrand & Schierenbeck, 2000;Gaskin & Schaal, 2002; Londo & Schaal, 2007; Novak,2007; Dlugosch & Parker, 2008a; Culley & Hardiman,2009; Gaskin & Kazmer, 2009). In evolutionarily com-plex cases, immigrant populations may be the productof a combination of factors: (i) colonizers from unre-lated native range sources; (ii) gene flow from cross-compatible domesticated relatives; (iii) gene flow fromcross-compatible wild relatives; and (iv) genetic back-crossing between, and subsequent introgression within,developing populations (Rieseberg & Wendel, 1993;Arnold, 1997; Rosenthal et al., 2008; Ellstrand, 2009;Ellstrand et al., 2010).
The elements of an introduction event define theinitial genetic conditions of diversity and variability thatmay be associated with evolutionary potential. The ini-tial diversity can be limited to what is carried withinone to a few individuals and represent a fraction of thegenetic diversity found in the source gene pool result-ing in a genetic bottleneck or “founder effect” (Durkaet al., 2005; Dlugosch & Parker, 2008b; Ficetola et al.,
C© 2012 Institute of Botany, Chinese Academy of Sciences
Journal of Systematics and Evolution 51 (3): 295–307 (2012)

296 Journal of Systematics and Evolution Vol. 51 No. 3 2012
2008; Rosenthal et al., 2008). Conversely, the diversityestablished during an introduction may closely reflectthe diversity of the source (Kang et al., 2007; Marrset al., 2008). For multiple introductions, overall ge-netic diversity of successful colonists from multiplesource gene pools may vary but will likely be greaterthan a single introduction (Durka et al., 2005; Williamset al., 2005; Dlugosch & Parker, 2008a; Marrs et al.,2008; Rosenthal et al., 2008). This “diversity of diver-sity” can be important to colonizing success if geneticvariation serves as a substrate for rapid adaptive re-sponses to novel conditions (Kolbe et al., 2004; Kanget al., 2007; Lavergne & Molofsky, 2007) or invents en-tirely novel genotypes (Ellstrand & Schierenbeck, 2000;Williams et al., 2005; Culley & Hardiman, 2009; Ell-strand, 2009; Gaskin & Kazmer, 2009; Schierenbeck &Ellstrand, 2009). Therefore, multiple introductions fromdifferent source populations may provide more evolu-tionary potential for colonizing populations in the intro-duced range than could be provided from a single sourcepopulation.
The origins of invasions resulting from multiple in-troductions and leading to different invasion outcomesare now well known (e.g. Gaskin & Schaal, 2002; Durkaet al., 2005; Williams et al., 2005; Kang et al., 2007; Dar-ling et al., 2008; Dlugosch & Parker, 2008a; Ficetola etal., 2008; Marrs et al., 2008; Ghabooli et al., 2011).Most of these studies focus primarily on the questionof genetic variability and its effect on evolutionary po-tential (Kolbe et al., 2004; Cameron et al., 2008; Faconet al., 2008; Rietzel et al., 2008; Rosenthal et al., 2008;Ward et al., 2008; Brown & Stepien, 2009; Culley &Hardiman, 2009). Relatively few studies, however, at-tempt to reconstruct the entire invasion history fromthe identification of specific multiple origins to presentstate (e.g. Dietz et al., 1999; Williams et al., 2005; Dlu-gosch & Parker, 2008b; Lozier et al., 2009). Here, wepresent such a study that involves multiple introductionsof multiple taxa.
The invasive wild artichoke thistle of California,Cynara cardunculus L. (Asteraceae), is our study sys-tem. Cynara cardunculus is a large thistle native tomuch of the Mediterranean basin (Rottenberg & Zo-hary, 1996; Sonnante et al., 2007). This species containsthree sub-taxa: the wild C. cardunculus var. sylvestris(Lam.) Fiori is the progenitor of the two domesti-cated cultivated varieties, C. cardunculus var. scoly-mus (L.) Fiori (artichoke), and C. cardunculus var.altilis DC (cardoon). The most apparent phenotypicdifference between wild and cultivated varieties ofC. cardunculus is the relative presence or absence ofspininess as part of the leaf, stem, and phyllary struc-tures. Cynara cardunculus is a diploid species (2n = 34).
Cynara cardunculus has a long history as an in-vasive in South America (Sturdevant, 1887; Barlow,1933) and since the late 19th century has also beena pernicious rangeland invader in both California andAustralia (Campbell, 1923; Thomsen & Barbe, 1986).Herbarium vouchers and the Calflora database indicatethat artichoke thistle and/or feral artichokes have beendocumented in at least 45 of California’s 58 counties(Calflora, 2012; CCH, 2012). Populations in Californiamay consist of scattered individuals or monospecificstands of varying sizes (Kelly & Pepper, 1996). Theprimary economic impact of this species is rangelanddegradation (Thomsen & Barbe, 1986). Ecological im-pacts can include physical exclusion from habitat dueto impassibility, changes in predator/prey habitat (e.g.rattlesnakes are attracted to large stands), replacementof native flora (White & Holt, 2005), and contributionsto shifts in the abiotic environment (Pepper & Kelly,1994; Potts et al., 2008).
The origin of California’s wild artichoke thistlehas been difficult to ascertain from historical notes andwritings that at times have made disparate claims. Allthree C. cardunculus taxa appear to have been intro-duced to California at about the same time. From thelatter part of the 19th century through the first half ofthe 20th century artichoke cultivation in California ex-panded both as a crop and as a popular garden veg-etable (Robbins et al., 1951; Jeffers, 2002). Cardoon,the other cultivar, was introduced during this period as agarden vegetable and flowering ornamental plant. Cal-ifornia wild artichoke thistle (sometimes referred to as“wild cardoon,” “wild artichoke,” or simply “artichokethistle”) reportedly first became noticeably invasive inthe late 1800s in Solano County, east of San FranciscoBay near the town of Benicia (Robbins et al., 1951).About this time newspaper articles reported that standsof escaped artichokes had “gone wild,” becoming spiny,while other newsletters and bulletins suggested that thewild variety might have been introduced inadvertentlyby Italian immigrants or was planted intentionally asliving fences and escaped (Robbins et al., 1951; Hill-yard, 1985; Thomsen et al., 1986). Cynara cardunculusachenes are large and smooth and are less likely thansmall, rough, or barbed seeds or fruits to be introducedinadvertently as hitchhikers on animals or in clothing,or as weed seed contaminants in feed or grain.
The introduction of artichoke thistle in South-ern California apparently happened at a similar timeas in Northern California. Samuel Parish charac-terized C. cardunculus as “abundant and well es-tablished” in San Diego County prior to sum-mer 1897 (Thomsen et al., 1986). There seems tohave been an anecdotal assumption of secondary
C© 2012 Institute of Botany, Chinese Academy of Sciences

LEAK-GARCIA et al.: California wild artichoke thistle’s complex origin 297
movement of plants from north to south in the state(Pepper & Kelly, 1994). However, although Californiapopulations are morphologically heterogeneous, themorphological variation appears geographically hap-hazard and exhibits no cline (Leak-Garcia, 2009), sug-gesting California populations have not establishedsequentially, and might not even share a commonpedigree.
This study examined the origins of C. cardunculusin California. Artichoke thistle has a generation time ofabout 2 years, long enough to permit any evolution thatmay have occurred since its first introduction to Califor-nia about 140 years ago, yet short enough to allow us tostudy historic traces of its progenitors. Using moleculartools and historical records, we explored its evolutionaryhistory leading to its current population structure. Not-ing the possibility of a complex origin for California’sartichoke thistle, we address several hypotheses:
(i) California artichoke thistle is the same as theMediterranean wild artichoke thistle and was in-troduced unintentionally by Italian immigrants.
(ii) California artichoke thistle is a feral form of cul-tivated artichoke that has evolved such that is hadreturned to its progenitor’s spiny morphology.
(iii) California artichoke thistle is a hybrid derivative,the result of gene flow between introduced wildplants and cultivar varieties.
(iv) California artichoke thistle is a composite set ofpopulations with diverse origins.
1 Material and methods
1.1 Sources of plant material and DNA extractionIn August 2005, we collected mature seed heads
from up to 20 individuals in each of 12 populations ofputative C. cardunculus var. sylvestris (artichoke thistle)in California (Fig. 1 provides four-letter code names).Seed families from individual plants were kept separatefrom each other. Because the plants occasionally re-produce vegetatively by offshoots or runners, care wastaken to avoid sampling different ramets of the samegenet. Specifically, we used a distance of 3 m betweensampled plants when possible. If the population wassufficiently large, 20 samples were collected at approx-imately equal distances from each other over the entirepopulation footprint. A few populations had been nearlyeradicated; in that case, all remaining individuals weresampled. In locations with significant herbivory, sam-ples represented the individuals in the population thatretained viable seeds. While collecting seed samples wesaw a number of populations (generally with very few
Fig. 1. Distribution of Cynara cardunculus collection sites used in thestudy. A, California sites (visited in August 2005). B, European sites(visited in August 2006). Population codes are consistent with thoseshown in Table 5.
plants) that appeared to be volunteer feral artichokes.For the most part, we avoided collecting from thesepopulations, but there were two exceptions. SACS andSACN, only a few kilometers from each other, appearedto have cultivar ancestry (they were nearly spineless withlarge fleshy leaves), but their flower heads were armedwith sharply rigid protruding phyllaries, indicating aprobable genetic difference from known cultivars. Allseeds were extracted from the flower heads in the lab andallowed a 3-month after-ripening period at room tem-perature before being planted. Previous work in the HoltLab at the University of California, Riverside (UCR) hasshown that this after-ripening period improves germina-tion (White V, 2007, personal communication).
C© 2012 Institute of Botany, Chinese Academy of Sciences
298 Journal of Systematics and Evolution Vol. 51 No. 3 2012
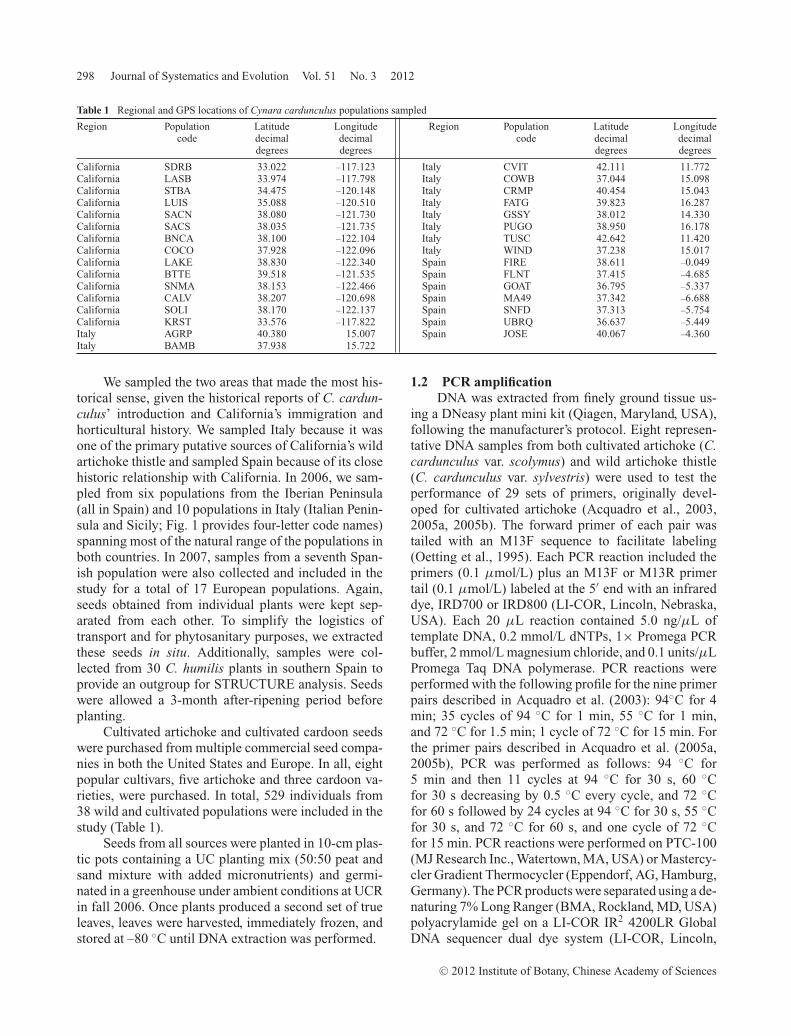
Table 1 Regional and GPS locations of Cynara cardunculus populations sampled
Region Population Latitude Longitude Region Population Latitude Longitudecode decimal decimal code decimal decimal
degrees degrees degrees degrees
California SDRB 33.022 –117.123 Italy CVIT 42.111 11.772California LASB 33.974 –117.798 Italy COWB 37.044 15.098California STBA 34.475 –120.148 Italy CRMP 40.454 15.043California LUIS 35.088 –120.510 Italy FATG 39.823 16.287California SACN 38.080 –121.730 Italy GSSY 38.012 14.330California SACS 38.035 –121.735 Italy PUGO 38.950 16.178California BNCA 38.100 –122.104 Italy TUSC 42.642 11.420California COCO 37.928 –122.096 Italy WIND 37.238 15.017California LAKE 38.830 –122.340 Spain FIRE 38.611 –0.049California BTTE 39.518 –121.535 Spain FLNT 37.415 –4.685California SNMA 38.153 –122.466 Spain GOAT 36.795 –5.337California CALV 38.207 –120.698 Spain MA49 37.342 –6.688California SOLI 38.170 –122.137 Spain SNFD 37.313 –5.754California KRST 33.576 –117.822 Spain UBRQ 36.637 –5.449Italy AGRP 40.380 15.007 Spain JOSE 40.067 –4.360Italy BAMB 37.938 15.722
We sampled the two areas that made the most his-torical sense, given the historical reports of C. cardun-culus’ introduction and California’s immigration andhorticultural history. We sampled Italy because it wasone of the primary putative sources of California’s wildartichoke thistle and sampled Spain because of its closehistoric relationship with California. In 2006, we sam-pled from six populations from the Iberian Peninsula(all in Spain) and 10 populations in Italy (Italian Penin-sula and Sicily; Fig. 1 provides four-letter code names)spanning most of the natural range of the populations inboth countries. In 2007, samples from a seventh Span-ish population were also collected and included in thestudy for a total of 17 European populations. Again,seeds obtained from individual plants were kept sep-arated from each other. To simplify the logistics oftransport and for phytosanitary purposes, we extractedthese seeds in situ. Additionally, samples were col-lected from 30 C. humilis plants in southern Spain toprovide an outgroup for STRUCTURE analysis. Seedswere allowed a 3-month after-ripening period beforeplanting.
Cultivated artichoke and cultivated cardoon seedswere purchased from multiple commercial seed compa-nies in both the United States and Europe. In all, eightpopular cultivars, five artichoke and three cardoon va-rieties, were purchased. In total, 529 individuals from38 wild and cultivated populations were included in thestudy (Table 1).
Seeds from all sources were planted in 10-cm plas-tic pots containing a UC planting mix (50:50 peat andsand mixture with added micronutrients) and germi-nated in a greenhouse under ambient conditions at UCRin fall 2006. Once plants produced a second set of trueleaves, leaves were harvested, immediately frozen, andstored at –80 ◦C until DNA extraction was performed.
1.2 PCR amplificationDNA was extracted from finely ground tissue us-
ing a DNeasy plant mini kit (Qiagen, Maryland, USA),following the manufacturer’s protocol. Eight represen-tative DNA samples from both cultivated artichoke (C.cardunculus var. scolymus) and wild artichoke thistle(C. cardunculus var. sylvestris) were used to test theperformance of 29 sets of primers, originally devel-oped for cultivated artichoke (Acquadro et al., 2003,2005a, 2005b). The forward primer of each pair wastailed with an M13F sequence to facilitate labeling(Oetting et al., 1995). Each PCR reaction included theprimers (0.1 μmol/L) plus an M13F or M13R primertail (0.1 μmol/L) labeled at the 5′ end with an infrareddye, IRD700 or IRD800 (LI-COR, Lincoln, Nebraska,USA). Each 20 μL reaction contained 5.0 ng/μL oftemplate DNA, 0.2 mmol/L dNTPs, 1× Promega PCRbuffer, 2 mmol/L magnesium chloride, and 0.1 units/μLPromega Taq DNA polymerase. PCR reactions wereperformed with the following profile for the nine primerpairs described in Acquadro et al. (2003): 94◦C for 4min; 35 cycles of 94 ◦C for 1 min, 55 ◦C for 1 min,and 72 ◦C for 1.5 min; 1 cycle of 72 ◦C for 15 min. Forthe primer pairs described in Acquadro et al. (2005a,2005b), PCR was performed as follows: 94 ◦C for5 min and then 11 cycles at 94 ◦C for 30 s, 60 ◦Cfor 30 s decreasing by 0.5 ◦C every cycle, and 72 ◦Cfor 60 s followed by 24 cycles at 94 ◦C for 30 s, 55 ◦Cfor 30 s, and 72 ◦C for 60 s, and one cycle of 72 ◦Cfor 15 min. PCR reactions were performed on PTC-100(MJ Research Inc., Watertown, MA, USA) or Mastercy-cler Gradient Thermocycler (Eppendorf, AG, Hamburg,Germany). The PCR products were separated using a de-naturing 7% Long Ranger (BMA, Rockland, MD, USA)polyacrylamide gel on a LI-COR IR2 4200LR GlobalDNA sequencer dual dye system (LI-COR, Lincoln,
C© 2012 Institute of Botany, Chinese Academy of Sciences

LEAK-GARCIA et al.: California wild artichoke thistle’s complex origin 299
Table 2 Microsatellite loci used in the study
Locus Alleles/ Size BP ReferenceBins range difference
CDAT-01 5 216–271 55 Acquadro et al. (2003)CDAT-02 6 162–242 80 Acquadro et al. (2003)CLIB-03 10 302–332 30 Acquadro et al. (2003)CLIB-12 8 212–241 29 Acquadro et al. (2003)CDAT-03 3 198–217 19 Acquadro et al. (2003)CMAL-24 8 197–217 20 Acquadro et al. (2005b)CMAL-06 35 134–178 44 Acquadro et al. (2005b)CMAL-07 3 192–201 9 Acquadro et al. (2005b)CMAL-25 22 136–169 30 Acquadro et al. (2005b)CMAL-144 3 194–201 7 Acquadro et al. (2005b)CLIB-02 5 201–225 24 Acquadro et al. (2003)CMAFLP-07 11 175–300 125 Acquadro et al. (2005a)CMAL-08 8 154–179 25 Acquadro et al. (2005b)Total 127
As a conservative approach, alleles were grouped into allele bins whendiscrete individuality could not be determined.
NE, USA). After addition of formamide loading dyeat a ratio of 1 μL dye per 10 μL PCR product, PCRproducts were denatured at 94 ◦C for 3 min and loadedon the gel. A 0.5 μL aliquot from each diluted and de-natured PCR amplification product was loaded in eachlane of the gel with three lanes (first, center, and lastlane) containing a 50–350 bp size standard (LI-COR).Primer performance on C. cardunculus var. sylvestrissamples was evaluated for each primer pair. Thirteensets of primers were considered to be informative basedon the PCR products analyzed on the LI-COR. Thesewere then utilized for all analyses.
Microsatellite loci were PCR amplified using the13 sets of primers (Table 2) for all samples; the re-actions were carried out with the same conditions de-scribed above. M13F primers labeled 5′ with FAM orNED and M13R primers labeled 5’ with VIC or PETwere included for visualization. The PCR products foreach individual plant were pooled in groups containingone primer set labeled with each of the four dyes andanalyzed on the ABI PRISM R© 3100 Genetic Analyzer(Applied Biosystems Inc., Foster City, CA, USA) at theInstitute for Integrative Genome Biology at UCR. Thedata were acquired by on-line fluorescence detectionand saved as a chromatography file. In some instances,it was unclear if a chromatography peak truly repre-sented a unique allele. If the allele in question differedfrom an established allele by one base pair (short of anexpected repeat sequence), the allele was binned withthe established allele to keep the analysis conservative.ABI GeneMapper 3.7 was used to analyze the data toobtain the number and size of alleles from each sam-ple. For economy, two primer pairs (CMAFLP07 andCMAL08) were analyzed using the LI-COR method in-stead of the ABI.
1.3 STRUCTURE analysisMicrosatellite data were analyzed to determine
genotypic grouping of California’s C. cardunculus pop-ulations using genetic cluster assignment and imple-menting an admixture model algorithm in STRUC-TURE v. 2.2 (Pritchard et al., 2000). Using a fixed-valuefrequency distribution parameter of λ = 1, a burn-inwas conducted over 100 000 iterations with subsequentsimulation runs of 500 000 iterations for K-parameterestimation. For determining proper K-parameters, threeruns each at K-values ranging from K = 2 to K = 9 wereperformed and compared for consistency. K-parameterestimation was guided by plotting the lnPr(X |K) valuesassociated with each run, or the log-transformed proba-bility of K clusters, which in graphical form shows thegreatest decrease in slope following the most correctlyestimated K-value (Pritchard et al., 2000).
An initial test run of the data in STRUCTURE wasperformed using the microsatellite data collected fromtissue of the seed-grown artichokes and cardoons ofknown genetic origin. These samples were expected toform two distinctly different genotypic groups or clus-ters, which they did (artichokes and cardoons), whenentered into STRUCTURE with a K = 2 parameter, pro-viding an initial quality check of the data. Subsequently,the congener C. humilis was included in the analysis toprovide an additional unique genotypic cluster. The C.humilis cluster was used as a control group in the largerdata runs and, as expected, remained completely discreetfrom the C. cardunculus clusters at all K-values testedand for all combinations of input data. After completingthe STRUCTURE analysis of our known taxa (the culti-vars and the congener), we made a number of runs fromK = 2 to K = 5 utilizing all of the data groups (congener,cultivars, and all Italian, Spanish, and California popu-lations). Because we were specifically interested in theorigins of individual California populations relative totheir potential progenitors and because the large numberof California populations we sampled could potentiallybias our STRUCTURE results, we chose a novel ap-proach.
To examine each population in relationship to itspotential progenitors, we chose instead to examine theCalifornia populations one by one using a group clus-ter analysis against which we could compare Californiapopulations. For the standard group cluster, we includedthe aforementioned five groups: the two domesticatedtypes, artichoke and cardoon (groups 1 and 2), the con-gener C. humilis (group 3), and the two native rangegeographic groups, Spain and Italy (groups 4 and 5).We then ran these standard group clusters and includedone California population at a time. Each Californiapopulation was individually assessed for its cluster
C© 2012 Institute of Botany, Chinese Academy of Sciences

300 Journal of Systematics and Evolution Vol. 51 No. 3 2012
assignment alongside the five group clusters using K= 5. To allow for the possibility that a California pop-ulation was not at all related to any of the groups, weperformed runs at higher K-values to test whether eachCalifornia population would form its own genetic clus-ter when released from the constraint of K = 5. Thisanalysis allowed for the determination of a Californiapopulation’s affiliation with a population from outsideof the regions sampled. These higher K-values were alsoused to graph the highest likelihood probability of thebest K-value for each individual California populationanalyzed to confirm the appropriateness of our chosenK-value (ln prob K = 2–7, respectively: –5352, –4964,–4654, –4444, –4238, –4145).
An example of this approach is given in Fig. 2.Figure 2-A shows the Iberian group assignment of Cali-fornia population LASB. Figure 2-B demonstrates whathappens when an additional cluster was introduced intothe analysis (K = 6); a split occurs among Italian pop-ulations but the California/Iberian cluster remains un-changed.
2 Results
Allelic diversity of microsatellites was high in Cy-nara cardunculus as a whole. Initially, 163 alleles over13 loci were detected. At several loci the independenceof two or more alleles was in question, and these alleleswere binned to ensure conservative analysis (see Section1.2). This combining of ambiguously assignable allelesreduced the overall allele count to 127 (Table 2). The av-erage number of alleles detected per population acrossall 13 loci differed slightly among population groups.California had the lowest average with 24.5 alleles perpopulation. Cultivated artichoke had the highest averageat 28.6 (Table 3).
Some private alleles were detected within popula-tion groups; three alleles were specific to cultivated arti-choke, one to cultivated cardoon, 12 to California, 21 tothe Italian Peninsula and Sicily, and eight to the IberianPeninsula. Seventeen alleles were shared solely betweenCalifornia and another population group (Table 4). Sixalleles representing four loci were shared only betweenCalifornia and Italy; four alleles were found in multi-ple populations in both California and Italy; the othertwo were unique to only a single Californian popula-tion and a single Italian population. Seven alleles at fiveloci were uniquely shared between California and theIberian Peninsula (Table 4). Of these alleles, five wereuniquely shared by just one population per region, andin total involved two California populations and threeIberian populations. With regard to the cultivated taxa,three alleles were shared solely by California and culti-
vated artichoke, involving multiple populations of each.One allele was shared uniquely between one Californiaand one cultivated cardoon population (Table 4). Allelefrequencies are available upon request.
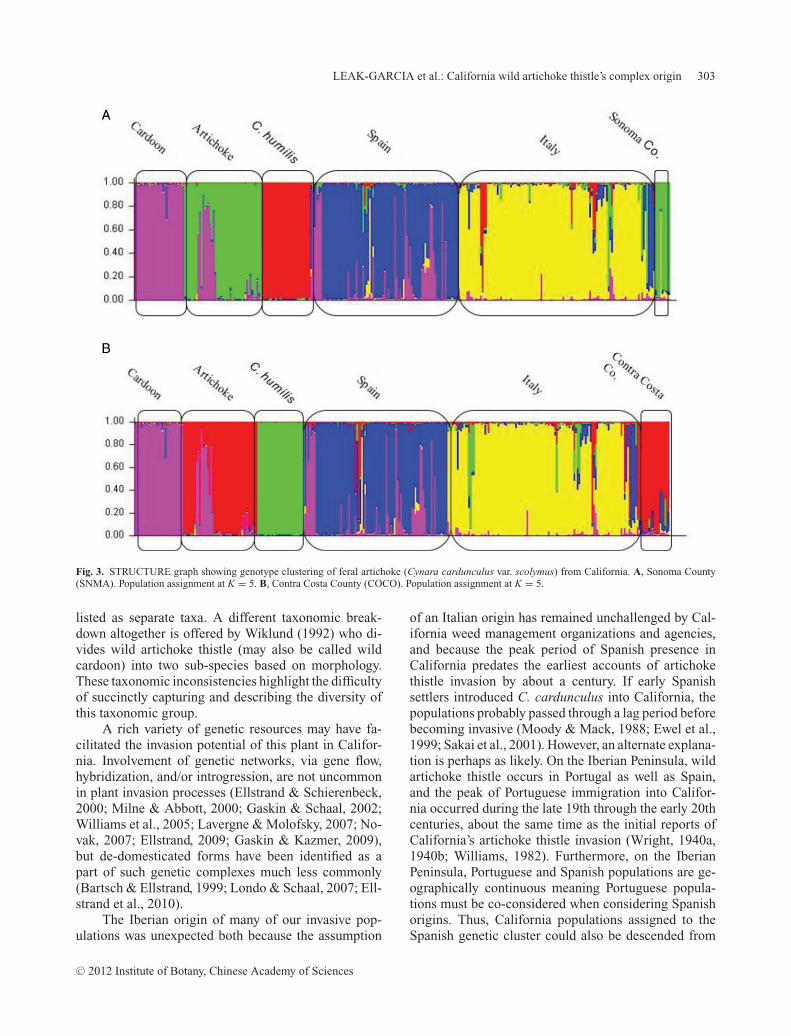
STRUCTURE analysis successfully assigned eachof the 12 California populations as affiliates of the pu-tative progenitor groups (Table 5) even when the higherK-value (K = 6) was used. The assignments were thesame whether just one or multiple California popula-tions were run concurrently. Three of the 12 Californiapopulations were identified as feral artichoke popula-tions (Fig. 3, Table 5). When given the opportunity toform a unique cluster using K = 6, K = 7, or K = 8,feral artichoke populations remained within their origi-nal artichoke assignment group. Seven populations wereprimarily assigned to the Iberian artichoke thistle clus-ter (Table 5). Three of these populations split off andformed a specific affiliation with an individual putativesource population when evaluated under K = 6. Figure2: C, D gives an example, showing one of these pop-ulations, BNCA, as closely affiliated with the Spanishpopulation GOAT. SOLI also affiliated with GOAT atK = 6; SDRB affiliated with Spanish FLNT.
One population (SACS) showed high levels of ad-mixture between artichoke and cardoon with a sec-ondary affiliation with both Italian and Iberian arti-choke thistle. Its close geographic neighbor, SACN (seeTable 1), depending on choice of K-value, is affiliatedwith either cardoon or the Iberian Peninsula. However,when the STRUCTURE analysis of SACN was per-formed using only the wild Spanish and cardoon pop-ulation data, SACN consistently affiliated with Spanishpopulations over cardoon varieties.
STRUCTURE analysis revealed the Spanish pop-ulation FIRE to be feral cardoon, which is substan-tiated by its field morphology (Leak-Garcia, 2009).One of the commercial seed-grown cultivated arti-choke populations was heavily affiliated with cardoons(Figs. 2, 3).
3 Discussion
We found that the evolutionary origins of Cynaracardunculus populations in California were diverse andpolyphyletic. Consequently, this complex group of pop-ulations consists of multiple sub-taxa and regionally dis-tinct genotypes. One-third of the populations sampled(four populations) were not wild artichoke thistle, as hadbeen assumed, but rather they were descendants of do-mesticated artichokes. Of these four populations, threehad evolved full spininess. These feral artichoke pop-ulations were morphologically cryptic artichokes that
C© 2012 Institute of Botany, Chinese Academy of Sciences

LEAK-GARCIA et al.: California wild artichoke thistle’s complex origin 301
Fig. 2. STRUCTURE graph showing genotype clustering of artichoke thistle (Cynara cardnuculus var. sylvestris) from California. A, B, Tres HermanosRanch, Los Angeles, and San Bernardino Counties (LASB). Population assignment at K = 5 (A) and K = 6 (B). C, D, Benicia (BNCA). Populationassignment at K = 5 (C) and K = 6 (D)
C© 2012 Institute of Botany, Chinese Academy of Sciences

302 Journal of Systematics and Evolution Vol. 51 No. 3 2012
Table 3 Allelic diversity of Cynara cardunculus sub-groups in the studyas detected by microsatellite analysis
Population Total Average Total Averagegroup individuals population alleles No. alleles/
sampled sample size population
California 211 15.07 319 24.5Italy 117 11.7 261 26.1Spain 88 12.57 191 27.3Cardoon 30 10.00 80 26.7Artichoke 46 9.20 143 28.6
evolutionarily converged with their wild progenitor.Free-living plants that have evolved from domesticatessuch that they resemble their progenitors are sometimesreferred to as “revertants.” We also demonstrated that
the remaining populations of California’s artichoke this-tle were most closely related to Iberian plants, not Italianplants as originally thought. Evidence for significant ge-netic admixture with artichoke and/or cardoon cultivarsand evidence for a low level of Italian ancestry werepresent for at least one (and possibly two) population inthis latter group. This co-occurrence of different geno-typic groups reveals the complex history of invasiveCalifornian C. cardunculus.
The taxonomic history of C. cardunculus has beencomplicated. Until recently, the taxon was split into twospecies, C. scolymus L. (artichokes) and C. cardunculusL. (artichoke thistle and cardoon); they are often still
Table 4 Alleles shared privately between groups
Alleles Locus-allele No. CA Other Population codeprivate to populations pops.
CA/IT CLIB-03–5 3 5 BNCA∗ SOLI∗ SNMA∗ FATG GSSY PUGO BAMB COWBCA/IT CMAL-06–34 3 1 BNCA∗ SOLI∗ COCO∗ WINDCA/IT CMAL-25–4 1 1 COCO∗ GSSYCA/IT CMAL-25–5 2 5 COCO∗ LAKE∗ FATG GSSY PUGO BAMB WINDCA/IT CMAL-24–6 1 1 LAKE∗ FATGCA/IT CMAL-06–20 4 1 BTTE∗ SACS∗ SOLI∗ SNMA∗ COWBTotal: 6CA/SP CMAL-07–3 1 1 LUIS∗ FLNTCA/SP CMAL-144–2 1 1 LUIS∗ FLNTCA/SP CMAL-25–16 1 1 SACN∗ JOSECA/SP CMAL-08–5 1 1 SACN∗ JOSECA/SP CMAL-25–12 1 1 SACN∗ MA49CA/SP CMAFLP -07–2 7 1 BNCA∗ COCO∗ SACS∗ SOLI∗ LUIS∗ LASB∗ SDRB∗ FLNTCA/SP CMAFLP -07–3 2 3 BTTE∗ CALV∗ GOAT MA49 UBRQTotal: 7CA/AR CLIB-12–2 4 3 BTTE∗ LAKE∗ LASB∗ SDRB∗ IMPS ROMA TSGGCA/AR CMAL-06–7 2 3 SACS∗ LAKE∗ ROMA TSGG VIOLCA/AR CMAL-24–3 2 2 CALV∗ LAKE∗ ROMA TSGGTotal: 3CA/CD CMAL-24–3 1 1 COCO∗ VCARTotal: 1∗ A California population. Other populations are listed by group in column one (CA/Other). “Other pops.” refers to number of populations from “other”group given in column one. Breakdown shows loci where alleles occurred, and all populations sharing those alleles.
Table 5 California Cynara cardunculus population progenitor assignments from STRUCTURE analysis
Genetic group assignment
Population Primary Secondary Comments
LAKE Artichoke Possibly cardoon Cardoon affiliation may come from cardoon-admixed artichoke (such as TSGG)with which it shares alleles representing a small proportion of the genotype.
SNMA Artichoke Italy Italy represents small proportion of the genotypeCOCO Artichoke Artichoke thistle Shares private alleles with Italy, Spain, and cardoonSACS Cardoon/
ArtichokeArtichoke thistle Highly admixed. Shares private alleles with artichoke, Italy, and Spain.
SACN Spain Spineless populationCALV Spain Artichoke, Artichoke
thistleShares private alleles with two artichoke cultivars. Assignment of artichoke thistle
origin uncertain— represents small proportion.BNCA Spain ItalySOLI Spain Artichoke Artichoke represents small proportionLUIS SpainLASB Spain Shares one private allele with multiple artichoke cultivars representing small
proportion of genotype.KRST SpainSDRB Spain; possibly feral
landrace cardoonArtichoke Shares one private allele with multiple artichoke cultivars representing small
proportion of genotype.
Population codes are consistent with those in Table 1.
C© 2012 Institute of Botany, Chinese Academy of Sciences
LEAK-GARCIA et al.: California wild artichoke thistle’s complex origin 303
Fig. 3. STRUCTURE graph showing genotype clustering of feral artichoke (Cynara cardunculus var. scolymus) from California. A, Sonoma County(SNMA). Population assignment at K = 5. B, Contra Costa County (COCO). Population assignment at K = 5.
listed as separate taxa. A different taxonomic break-down altogether is offered by Wiklund (1992) who di-vides wild artichoke thistle (may also be called wildcardoon) into two sub-species based on morphology.These taxonomic inconsistencies highlight the difficultyof succinctly capturing and describing the diversity ofthis taxonomic group.
A rich variety of genetic resources may have fa-cilitated the invasion potential of this plant in Califor-nia. Involvement of genetic networks, via gene flow,hybridization, and/or introgression, are not uncommonin plant invasion processes (Ellstrand & Schierenbeck,2000; Milne & Abbott, 2000; Gaskin & Schaal, 2002;Williams et al., 2005; Lavergne & Molofsky, 2007; No-vak, 2007; Ellstrand, 2009; Gaskin & Kazmer, 2009),but de-domesticated forms have been identified as apart of such genetic complexes much less commonly(Bartsch & Ellstrand, 1999; Londo & Schaal, 2007; Ell-strand et al., 2010).
The Iberian origin of many of our invasive pop-ulations was unexpected both because the assumption
of an Italian origin has remained unchallenged by Cal-ifornia weed management organizations and agencies,and because the peak period of Spanish presence inCalifornia predates the earliest accounts of artichokethistle invasion by about a century. If early Spanishsettlers introduced C. cardunculus into California, thepopulations probably passed through a lag period beforebecoming invasive (Moody & Mack, 1988; Ewel et al.,1999; Sakai et al., 2001). However, an alternate explana-tion is perhaps as likely. On the Iberian Peninsula, wildartichoke thistle occurs in Portugal as well as Spain,and the peak of Portuguese immigration into Califor-nia occurred during the late 19th through the early 20thcenturies, about the same time as the initial reports ofCalifornia’s artichoke thistle invasion (Wright, 1940a,1940b; Williams, 1982). Furthermore, on the IberianPeninsula, Portuguese and Spanish populations are ge-ographically continuous meaning Portuguese popula-tions must be co-considered when considering Spanishorigins. Thus, California populations assigned to theSpanish genetic cluster could also be descended from
C© 2012 Institute of Botany, Chinese Academy of Sciences

304 Journal of Systematics and Evolution Vol. 51 No. 3 2012
Portuguese populations, and consequently we have as-signed these populations to a more inclusive Iberianorigin. Although we did not detect any California popu-lations to be of direct Italian descent, a number of allelesdetected were specific to Italy and California. Given theethnobotanical history of California, these data suggestthat wild Italian plants that were once present in Califor-nia have left their genetic signature in current CaliforniaCynara populations.
Our genetic data uncovered genetic traces from apast introduction of wild Italian plants into California,but we have no evidence that such plants persist in Cal-ifornia. Despite evidence for low levels of introgressedItalian alleles in contemporary California C. cardun-culus, plants with a primary assignment to the Italianpopulations were not evident in our analysis. Not onlyis the Italian genotype missing from California, but theItalian phenotype is missing as well. This phenotypeis morphologically quite distinct from the Iberian phe-notype (Wiklund, 1992) and is conspicuously absent inCalifornia (Leak-Garcia, 2009). Typically Italian plantsare considerably shorter and bushier than Spanish orCalifornia plants and produce smaller, more numerousflower heads (Wiklund, 1992; Leak-Garcia, 2009). Ital-ian alleles seem to have been absorbed into the genepools of most of the extant wild Iberian and feral culti-var populations. A thorough review of historic herbar-ium specimens might help resolve the history of wildItalian artichoke thistle in California.
Our California sample had two nearly spinelesspopulations. One, SACS, was assigned hybrid ances-try involving cultivated artichoke and artichoke thistle.We assigned the other, SACN, to Iberian origin withsome affinity to cardoon as well (see Table 5). Spine-lessness occurs naturally in artichoke thistle popula-tions at a low frequency; however, spineless seedlingsare less likely to persist in arid environments due to her-bivory. Alternatively, the SACN population might havesome ancestry that includes admixture from an unsam-pled cardoon landrace, which would also explain itsspinelessness.
Hybridization has been thought to occasionallycontribute to the invasiveness of some of our most prob-lematic weeds and invasives (Ellstrand & Schierenbeck,2000; Gaskin & Schaal, 2002; Ellstrand, 2003; Williamset al., 2005; Lavergne & Molofsky, 2007; Culley &Hardiman, 2009; Gaskin & Kazmer, 2009). Despite themultiple origins of California’s wild artichoke thistle, itspresent invasiveness does not seem to result solely fromthe simple hybridization of two distinct taxa, as reportedin several other systems, where, for example, heterosisor transgressive traits may have evolved (Ellstrand &Schierenbeck, 2000; Gaskin & Schaal, 2002; Williams
et al., 2005; Lavergne & Molofsky, 2007; Culley &Hardiman, 2009; Gaskin & Kazmer, 2009). Rather, itappears that multiple distinct lineages—at least onehybrid-derived and some without evidence of hybridorigins—are involved in the evolution of the popula-tions.
Our study indicates that ferality leading to de-domestication can contribute to invasion processes.Studies of weedy or invasive de-domesticated crops arerelatively sparse, particularly those that provide infor-mation regarding the evolution of the problematic lin-eages (but see Gressel, 2005; Ellstrand et al., 2010). Ina study of California’s weedy beets, Bartsch & Ellstrand(1999) found they had complex polyphyletic origin in-volving some populations descended directly from morethan one wild taxon, some populations descended fromdomesticated taxa, and some populations descendedfrom hybrids. Burger et al. (2006) determined thatweedy rye in the far Western United States, previouslybelieved to be a crop × wild hybrid descendent, was in-stead directly descended from feral cultivated cereal rye.Recently, Cao et al. (2006) demonstrated that weedy ricethat recently appeared in northeastern China evolveddirectly from one or more local cultivars. These fewcases, along with the present results, indicate that rever-sion to free-living forms directly from cultivated geno-types without hybridization should be recognized as apotential evolutionary pathway to weediness or inva-siveness. They are of particular interest because theyhighlight an evolutionary potential in crops and horti-cultural plants that is not commonly recognized. It isimportant to elucidate what factors account for the evo-lution of weediness or invasiveness and require furtherstudy.
The feral artichoke populations we identified are ofspecial interest because they were misidentified as arti-choke thistle prior to this analysis. Leak-Garcia (2009)found that herbarium reports of volunteer feral artichokepopulations had decreased concurrently with increasingreports of artichoke thistle populations. It is possiblethat the evolution of individuals resembling artichokethistles is responsible for some of this shift in report-ing as they out-competed genotypes more similar to thecrop progenitor. Given that we have shown that feralartichokes can evolve morphological similarity to arti-choke thistle, and that they can evolve to become highlyinvasive, these seemingly innocuous volunteer or feralpopulations take on a new significance as foci for poten-tially nascent invasive populations. While we examinedthe evolutionary origin of these invasive lineages, thisinformation alone does not identify the precise evolu-tionary mechanism or specific traits that account for ashift to invasiveness. Consequently, our study has raised
C© 2012 Institute of Botany, Chinese Academy of Sciences

LEAK-GARCIA et al.: California wild artichoke thistle’s complex origin 305
several important questions. Does the invasion successof one genotype (Iberian artichoke thistle) and apparentfailure of another (Italian artichoke thistle) indicate vari-ation in potential weediness within this species? If so,elucidation of this point may contribute to understand-ing what characters are involved in invasiveness. Ques-tions remain about what triggers or favors feral artichokede-domestication of populations and how common thisevolutionary pathway may be. Given that we detectedprobable gene flow between horticultural artichokes andartichoke thistle populations in California, what are po-tential consequences of future gene flow between thesesub-taxa (Ellstrand, 2003)? Combining what we havelearned here about the origins of this invasion with fu-ture research on the detailed genetic structure of thecurrent populations will lay the necessary foundationfor understanding how these populations have evolvedand continue to evolve in California.
Acknowledgements We thank S. COZZOLINO forhis generosity in helping us locate and collect sam-ples in Italy. C. RIDLEY provided critical assistancecollecting in Europe. C. A. GARCIA, B. CASE, R.HILL, B. MACARTHUR, S. CASTILLE, J. SALZ, C.KOEHLER, E. PEARSON, P. CONGDON, V. WHITE,and D. JOHNSON, helped locate, escort, or otherwisefacilitated collecting Californian seed samples. Mem-bers of the Ellstrand and Holt Labs provided green-house assistance. Maps were created by L. SWEET. M.ROOSE and C. FEDERICI provided technical assis-tance and advice. J. HERATY and S. HEREDIA pro-vided helpful comments that improved the manuscript.Permit to import seeds collected in Europe (PPQ Form588) issued by United States Department of AgricultureAnimal and Plant Health Inspection Service (USDAAPHIS). This work was supported in part by Gradu-ate Assistance in Areas of National Needs (GAANN)Fellowship, United States National Science FoundationGrants (DEB 0409984 and DEB-1020799) and a JohnSimon Guggenheim Memorial Fellowship awarded toN. C. ELLSTRAND, a grant from the Elvenia J. SLOS-SON Endowment Fund awarded to J. S. HOLT, and aUC Dissertation Year Fellowship. A University of Cali-fornia Academic Senate Grant provided a portion of thetravel funds. Additional funding support was providedby J. S. HOLT and UCR’s Department of Botany andPlant Sciences.
References
Acquadro A, Portis E, Albertini E, Lanteri S. 2005a. M-AFLP-based protocol for microsatellite loci isolation in Cynara
cardunculus L. (Asteraceae). Molecular Ecology Notes 5:272–274.
Acquadro A, Portis E, Lanteri S. 2003. Isolation of microsatelliteloci in artichoke (Cynara cardunculus L. var. scolymus).Molecular Ecology Notes 3: 37–39.
Acquadro A, Portis E, Lee D, Donini P, Lanteri S. 2005b. De-velopment and characterization of microsatellite markers inCynara cardunculus L. Genome 48: 217–225.
Arnold ML. 1997. Natural hybridization and evolution. NewYork: Oxford University Press.
Baker HG, Stebbins GL. 1965. The genetics of colonizingspecies. London: Academic Press.
Barlow N. 1933. Charles Darwin’s Diary of the voyage of theH.M.S. Beagle. Edited from the MS. Cambridge: CambridgeUniversity Press.
Bartsch D, Ellstrand NC. 1999. Genetic evidence for the origin ofCalifornia wild beets (genus Beta). Theoretical and AppliedGenetics 99: 1120–1130.
Brown JE, Stepien CA. 2009. Invasion genetics of the Eurasianround goby in North America: Tracing sources and spreadpatterns. Molecular Ecology 18: 64–79.
Burger JC, Lee S, Ellstrand NC. 2006. Origin and genetic struc-ture of feral rye in the western United States. MolecularEcology 15: 2527–2539.
Calflora database. 2012. Available from http: //www.calflora.org/cgi-bin/library.cgi. [accessed June 2012].
Cameron EK, Bayne EM, Coltman DW. 2008. Genetic structureof invasive earthworms Dendrobaena octaedra in the borealforest of Alberta: Insights into introduction mechanisms.Molecular Ecology 17: 1189–1197.
Campbell DH. 1923. Australasian botanical notes II. Victoria,South Australia, and West Australia. American Journal ofBotany 10: 173–186.
Cao QJ, Lu BR, Xia H, Rong J, Sala F, Spada A, Grassi F. 2006.Genetic diversity and origin of weedy rice (Oryza sativaf. spontanea) populations found in North-eastern China re-vealed by simple sequence repeat (SSR) markers. Annals ofBotany 98: 1241–1252.
Consortium of California Herbaria (CCH). 2012. Availablefrom http: //ucjeps.berkeley.edu/consortium/ [accessed June2012].
Culley TM, Hardiman NA. 2009. The role of intraspecific hy-bridization in the evolution of invasiveness: A case studyof the ornamental pear tree Pyrus calleryana. BiologicalInvasions 11: 1107–1119.
Darling JA, Bagley MJ, Roman J, Tepolt CK, Geller JB. 2008.Genetic patterns across multiple introductions of the glob-ally invasive crab genus Carcinus. Molecular Ecology 17:4992–5007.
Dietz H, Fischer M, Schmid B. 1999. Demographic and ge-netic invasion history of a 9-year-old roadside populationof Bunias orientalis L. (Brassicaceae). Oecologia 120: 225–234.
Dlugosch KM, Parker IM. 2008a. Founding events in speciesinvasions: Genetic variation, adaptive evolution, and therole of multiple introductions. Molecular Ecology 17: 431–449.
Dlugosch KM, Parker IM. 2008b. Invading populations of anornamental shrub show rapid life history evolution despitegenetic bottlenecks. Ecology Letters 11: 701–709.
C© 2012 Institute of Botany, Chinese Academy of Sciences

306 Journal of Systematics and Evolution Vol. 51 No. 3 2012
Durka W, Bossdorf O, Prati D, Auge H. 2005. Molecular evi-dence for multiple introductions of garlic mustard (Alliariapetiolata, Brassicaceae) to North America. Molecular Ecol-ogy 14: 1697–1706.
Ellstrand NC. 2003. Dangerous liaisons: When cultivated plantsmate with their wild relatives. Baltimore: Johns HopkinsUniversity Press.
Ellstrand NC. 2009. Evolution of invasiveness in plants followinghybridization. Biological Invasions 11: 1089–1091.
Ellstrand NC, Schierenbeck KA. 2000. Hybridization as a stimu-lus for the evolution of invasiveness in plants? Proceedingsof the National Academy of Sciences USA 97: 7043–7050.
Ellstrand NC, Heredia SM, Leak-Garcia JA, Heraty JM, BurgerJC, Yao L, Nohzadeh-Malakshah S, Ridley CE. 2010. Cropsgone wild: Evolution of weeds and invasives from domesti-cated ancestors. Evolutionary Applications 3: 494–504.
Ewel JJ, O’Dowd DJ, Bergelson J, Daehler CC, D’Antonio CM,Gomez LD, Gordon DR, Hobb RJ, Holt A, Hopper KR,Hughes CE, LaHart M, Leakey RRB, Lee WG, Loope LL,Lorence DH, Louda SV, Lugo AE, McEvoy PB, Richard-son DM, Vitousek PM. 1999. Deliberate introductions ofspecies: Research needs. BioScience 49: 619–630.
Facon B, Pointier JP, Jarne P, Sarda V, David P. 2008. High ge-netic variance in life-history strategies within invasive pop-ulations by way of multiple introductions. Current Biology18: 363–367.
Ficetola GF, Bonin A, Miaud C. 2008. Population genetics re-veals origin and number of founders in a biological invasion.Molecular Ecology 17: 773–782.
Gaskin JF, Kazmer DJ. 2009. Introgression between invasivesaltcedars (Tamarix chinensis and T. ramosissima) in theUSA. Biological Invasions 11: 1121–1130.
Gaskin JF, Schaal BA. 2002. Hybrid tamarix widespread in U.S.invasion and undetected in native Asian range. Proceed-ings of the National Academy of Sciences USA 99: 11256–11259.
Ghabooli S, Shiganova TA, Zhan A, Cristescu ME, Eghtesadi-Araghi P, MacIsaac HJ. 2011. Multiple introductions andinvasion pathways for the invasive ctenophore Mnemiopsisleidyi in Eurasia. Biological Invasions 13: 679–690.
Gressel J. 2005. Crop ferality and volunteerism. Boca Raton:CRC Press. 1–7.
Hierro JL, Maron JL, Callaway RM. 2005. A biogeographicalapproach to plant invasions: The importance of studying ex-otics in their introduced and native range. Journal of Ecology93: 5–15.
Hillyard D. 1985. Artichoke thistle. Fremontia 12: 21–22.Jeffers HP. 2002. The Napoleon of New York: Mayor Fiorello La
Guardia. 2nd ed. New York: Wiley.Kang M, Buckley YM, Lowe AJ. 2007. Testing the role of genetic
factors across multiple independent invasions of the shrubScotch broom (Cytisus scoparius). Molecular Ecology 16:4662–4673.
Kelly M, Pepper A. 1996. Controlling Cynara Cardunculus (Ar-tichoke Thistle, Cardoon). In: Lovich J, Randall J, Kelly Meds. Proceedings of the California Exotic Pest Plant CouncilSymposium. San Diego: California Exotic Pest Plant Coun-cil. 97–100.
Kolbe JJ, Glor RE, Schettino LR, Lara AC, Larson A, Losos JB.2004. Genetic variation increases during biological invasionby a Cuban lizard. Nature 431: 177–191.
Lavergne S, Molofsky J. 2007. Increased genetic variation andevolutionary potential drive the success of an invasive grass.Proceedings of the National Academy of Sciences USA 104:3883–3888.
Leak-Garcia J. 2009. Genetic origins and the evolution of inva-siveness of Cynara cardunculus in California. Ph.D. Disser-tation. Riverside: University of California.
Londo JP, Schaal BA. 2007. Origins and population genetics ofweedy red rice in the USA. Molecular Ecology 16: 4523–4535.
Lozier JD, Roderick GK, Mills NJ. 2009. Tracing the invasionhistory of mealy plum aphid, Hyalopterus pruni (Hemiptera:Aphididae), in North America: A population genetics ap-proach. Biological Invasions 11: 299–314.
Marrs RA, Sforza R, Hufbauer RA. 2008. When invasion in-creases population genetic structure: A study with Centau-rea diffusa. Biological Invasions 10: 561–572.
Milne RI, Abbott RJ. 2000. Origin and evolution of invasivenaturalized material of Rhododendron ponticum L. in theBritish Isles. Molecular Ecology 9: 541–556.
Moody ME, Mack RN. 1988. Controlling the spread of plant in-vasions: The importance of nascent foci. Journal of AppliedEcology 20: 645–658.
Novak SJ. 2007. The role of evolution in the invasion process.Proceedings of the National Academy of Sciences USA 104:3671–3672.
Oetting WS, Lee HK, Flanders DJ, Wiesner GL, Sellers TA, KingRA. 1995. Linkage analysis with multiplexed short tandemrepeat polymorphisms using infrared fluorescence and M13tailed primers. Genomics 30: 450–458.
Pepper A, Kelly M. 1994. Portrait of an invader: The ecologyand management of the wild artichoke Cynara cardunculus.CalEPPC News. Winter 1994: 4–6.
Potts DL, Harpole WS, Goulden ML, Suding KN. 2008. The im-pact of invasion and subsequent removal of an exotic thistle,Cynara cardunculus, on CO2 and H2O vapor exchange in acoastal California grassland. Biological Invasions 10: 1073–1084.
Pritchard J, Stephens M, Donnelly P. 2000. Inference of popu-lation structure using multilocus genotypes data. Genetics155: 945–959.
Rieseberg LH, Wendel JF. 1993. Introgression and its conse-quences in plants. In: Harrison RG, Price J eds. Hybridzones and the evolutionary process. New York: Oxford Uni-versity Press. 70–109.
Rietzel AM, Darling JA, Sullivan JC, Finnerty JR. 2008. Globalpopulation genetic structure of the starlet anemone Ne-matostella vectensis: Multiple introductions and implica-tions for conservation policy. Biological Invasions 10: 1197–1213.
Robbins WW, Bellue MK, Ball WS. 1951. Weedsof California. Sacramento: California Department ofAgriculture.
Rosenthal DM, Ramakrishnan AP, Cruzan MB. 2008. Ev-idence for multiple sources of invasion and intraspe-cific hybridization in Brachypodium sylvaticum (Hudson)Beauv. in North America. Molecular Ecology 17: 4657–4669.
Rottenberg A, Zohary D. 1996. The wild ancestry of the cultivatedartichoke. Genetic Resources and Crop Evolution 45: 53–58.
C© 2012 Institute of Botany, Chinese Academy of Sciences

LEAK-GARCIA et al.: California wild artichoke thistle’s complex origin 307
Sakai AN, Allendorf FW, Holt JS, Lodge DM, Molofsky J, WithKA, Baughmann S, Cabin RJ, Cohen JE, Ellstrand NC, Mc-Cauley DE, O’Neil P, Parker IM, Thompson JN, Weller SC.2001. The population biology of invasive species. AnnualReview of Ecology and Systematics 32: 305–332.
Schierenbeck KA, Ellstrand NC. 2009. Hybridization and theevolution of invasiveness in plants and other organisms.Biological Invasions 11: 1093–1105.
Sonnante G, Pignone D, Hammer K. 2007. The domestication ofartichoke and cardoon: From Roman times to the genomicage. Annals of Botany 100: 1095–1100.
Sturdevant EL. 1887. History of garden vegetables (Continued).American Naturalist 21: 125–133.
Thomsen CD, Barbe GD. 1986. Artichoke thistle in California.California Agriculture 40: 7–10.
Thomsen CD, Barbe GD, Williams WA, George MR. 1986. Es-caped artichokes are troublesome pests. California Agricul-ture 40: 7–9.
Ward SM, Reid SD, Harrington J, Sutton J, Beck, KG. 2008.Genetic variation in invasive populations of yellow toad-
flax (Linaria vulgaris) in the Western United States. WeedScience 56: 394–399.
White VA, Holt JS. 2005. Competition of artichoke thistle (Cy-nara cardunculus) with native and exotic grassland species.Weed Science 53: 826–833.
Wiklund A. 1992. The genus Cynara L. (Asteraceae, Cardueae).Botanical Journal of the Linnean Society 109: 75–123.
Williams DA, Overholt WA, Cuda JP, Hughes CR. 2005. Chloro-plast and microsatellite DNA diversities reveal the intro-duction history of Brazilian peppertree (Schinus terebinthi-folius) in Florida. Molecular Ecology 14: 3643–3656.
Williams JR. 1982. And yet they come: Portuguese immigrationfrom the Azores to the United States. New York: Center forMigration Studies.
Wright DM. 1940a. The making of cosmopolitan California: Ananalysis of immigration 1848–1870. California HistoricalSociety Quarterly 19: 340.
Wright DM. 1940b. The making of cosmopolitan California:Part II an analysis of immigration 1848–1870. CaliforniaHistorical Society Quarterly 20: 72–73.
C© 2012 Institute of Botany, Chinese Academy of Sciences