Молекулярно-генетический анализ пяти сохранившихся...
TRANSCRIPT

ГЕНЕТИКА, 2016, том 52, № 8, с. 931–942
931
МОЛЕКУЛЯРНО-ГЕНЕТИЧЕСКИЙ АНАЛИЗПЯТИ СОХРАНИВШИХСЯ РЕЗЕРВАТОВ ТЕМНОЙ ЛЕСНОЙ
ПЧЕЛЫ Apis mellifera mellifera УРАЛА И ПОВОЛЖЬЯ© 2016 г. Р. А. Ильясов1, А. В. Поскряков1, А. В. Петухов2, А. Г. Николенко1
1Институт биохимии и генетики Уфимского научного центра Российской академии наук, Уфа 4500542Пермский государственный гуманитарно-педагогический университет, кафедра зоологии, Пермь 614990
е-mail: [email protected]е-mail: [email protected]
Поступила в редакцию 27.10.2015 г.
Изучены локальные популяции темной лесной пчелы Apis mellifera mellifera Урала и Поволжья всравнении с локальными популяциями пчел южных подвидов A. m. caucasica и A. m. carpatica Кавка-за и Карпат. Генетический анализ проводился на основе полиморфизма девяти микросателлитныхлокусов ядерной ДНК и локуса COI-COII мтДНК. На территории Урала и Поволжья нами были вы-делены пять сохранившихся популяций (резерватов) темной лесной пчелы A. m. mellifera: бурзян-ская, татышлинская, южно-прикамская, вишерская и камбарская. Эти пять популяций составляютоснову современного генофонда темной лесной пчелы A. m. mellifera Урала и Поволжья. Основноймассив сохранившегося аборигенного генофонда A. m. mellifera (ядро генофонда популяции A. m. mel-lifera) располагается на территории всего Пермского края и Севера Республики Башкортостан. Для по-пуляции пчел A. m. mellifera Урала и Поволжья были рассчитаны генетические стандарты, которые бу-дут полезны для последующих популяционных исследований медоносной пчелы.
Ключевые слова: темная лесная пчела, Apis mellifera mellifera, сохранение аборигенного генофонда,COI-COII мтДНК, микросателлитные локусы, сохранившиеся резерваты, интрогрессия, генетиче-ский стандарт, гетерозиготностьDOI: 10.7868/S0016675816060059
Из 30 известных европейских подвидов медо-носной пчелы Apis mellifera [1–3] только одинподвид A. m. mellifera приспособлен к жизни вусловиях с экстремально холодными и длительны-ми зимовками продолжительностью до 6–7 мес. икритически короткими периодами летнего медо-сбора [1–3]. Аборигенный генофонд темной лес-ной пчелы A. m. mellifera является источником ло-кальных адаптаций и уникальной комбинацииценных свойств, сформировавшейся в ходе дли-тельного естественного отбора [4].
Темная лесная пчела A. m. mellifera, представи-тель эволюционной ветви М [5, 6], на сегодняш-ний день признается подвидом, находящимсяпод угрозой вымирания в результате массовойинтрогрессии генофонда южных подвидов пчелэволюционной ветви С [5–9]. Межрегиональныеперемещения пчел разных подвидов приводят кпотере чистоты аборигенного генофонда в ре-зультате гибридизации [2].
Генофонд аборигенных темных лесных пчелA. m. mellifera считают утраченным во многихстранах Европы [10]. Известна полная заменааборигенной темной лесной пчелы A. m. mellifera
краинской пчелой A. m. carnica в Германии [10,11]. Предпочтение пчеловодов Западной и Север-ной Европы в разведении пчел эволюционнойветви С (A. m. carnica, A. m. ligustica и гибриднаяпчела Бэкфаст) по причине их дешевизны, до-ступности и раннего созревания маток по сравне-нию с темной лесной пчелой способствовало по-тере целостности ареала A. m. mellifera и интро-грессии генофонда южных подвидов [5].В Скандинавских странах и на Британских остро-вах большинство пчеловодов на данный моментпредпочитают разводить A. m. ligustica, A. m. cecro-pia, A. m. carnica или искусственно выведеннуюпороду бэкфаст [10]. В России подвид A. m. mel-lifera был практически повсеместно подверженгибридизации с подвидами A. m. caucasica иA. m. carpatica [3, 12–15].
Однако морфологические исследования и ис-следования митохондриальной ДНК предполага-ют сохранение темной лесной пчелы A. m. mellif-era на территории Урала и Поволжья [3, 12–21].Широкомасштабные исследования популяцийпчел на основе локусов ядерной ДНК в России непроводились. Отсутствие полноценной адекват-
УДК 575.17:638.123
ГЕНЕТИКА ЖИВОТНЫХ
4*

932
ГЕНЕТИКА том 52 № 8 2016
ИЛЬЯСОВ и др.
Таблица 1. Объем выборки семей медоносных пчел по районамРегион Район Число семей Число пасек
Республика Башкортостан Абзелиловский 90 11Альшеевский 35 6Баймакский 70 13Балтачевский 36 7Белебеевский 16 2Белорецкий 114 29Бирский 91 18Бурзянский 326 90Гафурийский 62 9Зилаирский 141 33Иглинский 197 12Ишимбайский 226 42Караидельский 132 19Кушнаренковский 37 8Куюргазинский 61 7Мелеузовский 73 14Мишкинский 55 12Татышлинский 200 17Уфимский 30 4Учалинский 10 2Хайбуллинский 130 19Чекмагушевский 62 12Чишминский 15 2Янаульский 100 10
Всего по Республики Башкортостан 2309 398Пермский край Добрянский 20 2
Красновишерский 41 9Нытвенский 18 2Ординский 25 3Осинский 38 2Пермский 76 4Уинский 59 7Усольский 20 2Частинский 28 2Юсьвенский 37 8
Всего по Пермскому краю 362 41Республика Удмуртия Завьяловский 39 3
Камбарский 46 2Малопургинский 26 5Можгинский 22 2Шарканский 34 2Якшур-Бодьинский 33 3
Всего по Республики Удмуртия 200 17Республика Татарстан Кукморский 24 4
Мамадышский 16 2Нижнекамский 12 2

ГЕНЕТИКА том 52 № 8 2016
МОЛЕКУЛЯРНО-ГЕНЕТИЧЕСКИЙ АНАЛИЗ 933
Всего по Республики Татарстан 52 8Кировская область Кирово-Чепецкий 20 2
Даровской 20 2Орловский 14 2Кильмезский 10 2
Всего по Кировской области 64 8Свердловская область Красноуфимский 58 8Республика Чувашия (РЧ) Чебоксарский 14 2Краснодарский край (КК) Сочинский 32 3Республика Адыгея (РА) Майкопский 15 2Закарпатская область (ЗО) Мукачевский 17 6
Всего 3123 493
Регион Район Число семей Число пасек
Таблица 1. Окончание
ной информации о состоянии и структуре медо-носной пчелы в России не позволяет эффективновыполнять мероприятия по сохранению и восста-новлению аборигенного генофонда темной лес-ной пчелы в локальных популяциях, подвержен-ных угрозе интенсивной внутривидовой гибри-дизации и интрогрессии “южных” генов.
Целью нашей работы является изучение ло-кальных популяций темной лесной пчелы Уралаи Поволжья, оценка их основных генетическиххарактеристик, анализ уровня интрогрессии и ло-кализация географических границ сохранивших-ся резерватов темной лесной пчелы A. m. mellifera наоснове анализа полиморфизма девяти микросател-литных локусов ядерной ДНК и локуса COI-COIIмтДНК.
МАТЕРИАЛЫ И МЕТОДЫБыли проанализированы образцы рабочих
особей местных пчел подвида A. m. mellifera из2729 семей с 447 пасек Урала, из 330 семей с 35 па-сек Поволжья. Для сравнительного анализа былииспользованы образцы южных пчел подвидов
A. m. caucasica, A. m. carpatica из 64 семей с 11 па-сек Кавказа и Карпат (табл. 1).
Рабочие особи пчел до выделения ДНК былизафиксированы в 96%-ном этаноле и хранилисьпри температуре –20°C. Выделение ДНК из мышцторакса проводили набором ДНК-ЭКСТРАН-2 попротоколу “Синтол” (Москва) (www.syntol.ru). Ка-чество и количество выделенной ДНК анализи-ровали на спектрофотометре NanoDrop 1000(Thermo, США).
Амплификация девяти микросателлитных ло-кусов (Ap243, 4a110, A24, A8, A43, A113, A88, Ap049и A28) ядерной ДНК и локуса COI-COII митохон-дриальной ДНК (табл. 2) [22–25] была выполненав термоциклере BIO-RAD T100 (США) по прото-колу СИЛЕКС (Москва) (www.sileks.com/ru). Фраг-ментарный анализ продуктов ПЦР был выполненна автоматическом секвенаторе Applied Biosystemsequencer (США).
Продукты амплификации разделяли в 8%ПААГ при силе тока 40 мА, окрашивали броми-стым этидием и фотографировались в гель-до-
Таблица 2. ДНК-маркеры, использованные в генетическом анализе популяций пчел
Примечание. В скобках (Q), (PQQ), (PQQQ) – обозначения фрагментов ДНК.
№ Локус Расположение на хромосоме Аллели, пн
1 Ap243 LG1 254, 257, 2602 4a110 LG4 160, 163, 1683 A24 LG7 98, 106, 1084 A8 LG2 154, 156, 158, 164, 1735 A43 LG3 128, 134, 140, 1426 A113 LG6 216, 218, 220, 222, 228, 2347 A88 LG8 143, 146, 148, 152, 1558 Ap049 LG1 123, 129, 130, 1429 A28 LG14 134, 140, 144
10 COI-COII Митохондрия 563 (Q)*, 823 (PQQ), 1023 (PQQQ)

934
ГЕНЕТИКА том 52 № 8 2016
ИЛЬЯСОВ и др.
Таблица 3. Значения гетерозиготности, коэффициентов инбридинга и родства в локальных популяциях пчел Ура-ла, Поволжья, Кавказа и Карпат, рассчитанные на основе анализа полиморфизма микросателлитных локусов
Локальная популяция N Ho ± σ Hs ± σ Ht ± σ Fis ± σ Fit ± σ Fst ± σ R ± σ
Абзелиловская 90 0.32 ± 0.08 0.35 ± 0.06 0.41 ± 0.07 0.07 ± 0.07 0.21 ± 0.08 0.15 ± 0.03 0.25 ± 0.03Альшеевская 35 0.42 ± 0.08 0.45 ± 0.06 0.48 ± 0.06 0.09 ± 0.08 0.16 ± 0.10 0.08 ± 0.03 0.14 ± 0.04Баймакская 70 0.33 ± 0.11 0.32 ± 0.11 0.35 ± 0.12 0.03 ± 0.03 0.12 ± 0.08 0.09 ± 0.02 0.16 ± 0.03Балтачевская 36 0.29 ± 0.10 0.33 ± 0.09 0.33 ± 0.08 0.10 ± 0.09 0.12 ± 0.11 0.02 ± 0.02 0.03 ± 0.03Белебеевская 16 0.37 ± 0.07 0.43 ± 0.06 0.48 ± 0.05 0.15 ± 0.08 0.31 ± 0.10 0.19 ± 0.07 0.29 ± 0.10Бирская 91 0.37 ± 0.10 0.38 ± 0.07 0.41 ± 0.07 0.04 ± 0.03 0.11 ± 0.10 0.07 ± 0.02 0.12 ± 0.03Бурзянская 326 0.28 ± 0.04 0.25 ± 0.04 0.34 ± 0.06 0.01 ± 0.01 0.179 ± 0.06 0.18 ± 0.02 0.30 ± 0.02Белорецкая 114 0.30 ± 0.06 0.32 ± 0.04 0.34 ± 0.05 0.08 ± 0.06 0.14 ± 0.05 0.06 ± 0.02 0.11 ± 0.04Гафурийская 62 0.37 ± 0.07 0.40 ± 0.04 0.45 ± 0.05 0.09 ± 0.08 0.16 ± 0.08 0.07 ± 0.02 0.13 ± 0.03Зилаирская 141 0.37 ± 0.09 0.33 ± 0.07 0.39 ± 0.09 –0.09 ± 0.08 0.06 ± 0.01 0.05 ± 0.04 0.10 ± 0.08Иглинская 197 0.43 ± 0.09 0.40 ± 0.05 0.42 ± 0.05 –0.05 ± 0.04 –0.01 ± 0.07 0.04 ± 0.02 0.08 ± 0.03Ишимбайская 226 0.42 ± 0.04 0.43 ± 0.03 0.48 ± 0.04 0.03 ± 0.02 0.12 ± 0.04 0.10 ± 0.01 0.17 ± 0.02Караидельская 132 0.35 ± 0.08 0.37 ± 0.07 0.38 ± 0.08 0.08 ± 0.06 0.11 ± 0.05 0.03 ± 0.02 0.06 ± 0.03Кушнаренковская 37 0.50 ± 0.12 0.41 ± 0.06 0.43 ± 0.06 –0.17 ± 0.12 –0.11 ± 0.10 0.06 ± 0.04 0.13 ± 0.08Куюргазинская 61 0.29 ± 0.10 0.32 ± 0.09 0.32 ± 0.09 0.09 ± 0.16 0.11 ± 0.10 0.03 ± 0.06 0.05 ± 0.10Мелеузовская 73 0.34 ± 0.07 0.35 ± 0.06 0.39 ± 0.05 0.06 ± 0.05 0.14 ± 0.09 0.09 ± 0.04 0.15 ± 0.06Мишкинская 55 0.28 ± 0.10 0.31 ± 0.10 0.32 ± 0.10 0.09 ± 0.07 0.13 ± 0.06 0.05 ± 0.04 0.08 ± 0.07Татышлинская 200 0.31 ± 0.10 0.30 ± 0.10 0.32 ± 0.10 0.01 ± 0.01 0.05 ± 0.04 0.14 ± 0.02 0.26 ± 0.04Янаульская 100 0.21 ± 0.08 0.25 ± 0.10 0.26 ± 0.11 0.16 ± 0.04 0.21 ± 0.05 0.18 ± 0.03 0.27 ± 0.05Уфимская 30 0.30 ± 0.07 0.38 ± 0.06 0.43 ± 0.06 0.15 ± 0.07 0.30 ± 0.09 0.06 ± 0.05 0.10 ± 0.06Учалинская 10 0.53 ± 0.17 0.41 ± 0.09 0.41 ± 0.09 –0.32 ± 0.18 –0.28 ± 0.20 0.03 ± 0.02 0.08 ± 0.06Чишминская 15 0.31 ± 0.09 0.41 ± 0.07 0.40 ± 0.07 0.20 ± 0.13 0.17 ± 0.12 –0.04 ± 0.02 –0.07 ± 0.03Чекмагушевская 62 0.58 ± 0.18 0.38 ± 0.10 0.38 ± 0.10 –0.56 ± 0.14 –0.55 ± 0.01 0.01 ± 0.14 0.03 ± 0.05Хайбуллинская 130 0.27 ± 0.09 0.29 ± 0.10 0.33 ± 0.13 0.09 ± 0.06 0.21 ± 0.08 0.13 ± 0.04 0.22 ± 0.05Уинская 59 0.32 ± 0.12 0.32 ± 0.11 0.32 ± 0.11 –0.01 ± 0.01 –0.01 ± 0.01 0.01 ± 0.01 0.02 ± 0.01Ординская 25 0.34 ± 0.13 0.33 ± 0.13 0.33 ± 0.13 –0.01 ± 0.01 –0.01 ± 0.01 –0.01 ± 0.01 –0.01 ± 0.01Частинская 28 0.26 ± 0.14 0.25 ± 0.13 0.24 ± 0.13 –0.06 ± 0.06 –0.10 ± 0.06 –0.04 ± 0.01 –0.08 ± 0.01Нытвенская 18 0.27 ± 0.13 0.25 ± 0.11 0.25 ± 0.11 –0.07 ± 0.05 –0.13 ± 0.05 –0.05 ± 0.01 –0.13 ± 0.01Осинская 38 0.33 ± 0.17 0.25 ± 0.11 0.24 ± 0.11 –0.33 ± 0.13 –0.35 ± 0.13 –0.02 ± 0.01 –0.06 ± 0.01Пермская 76 0.29 ± 0.11 0.31 ± 0.09 0.33 ± 0.11 0.04 ± 0.10 0.15 ± 0.12 0.11 ± 0.04 0.19 ± 0.05Юсьвенская 37 0.24 ± 0.11 0.26 ± 0.11 0.27 ± 0.11 0.07 ± 0.05 0.09 ± 0.05 0.02 ± 0.02 0.04 ± 0.05Красновишерская 41 0.18 ± 0.10 0.15 ± 0.08 0.24 ± 0.12 –0.04 ± 0.03 0.12 ± 0.08 0.16 ± 0.07 0.28 ± 0.04Усольская 20 0.26 ± 0.14 0.29 ± 0.09 0.29 ± 0.09 0.13 ± 0.13 0.08 ± 0.06 –0.06 ± 0.01 –0.11 ± 0.01Добрянская 20 0.30 ± 0.12 0.27 ± 0.10 0.27 ± 0.10 –0.10 ± 0.07 –0.15 ± 0.07 –0.05 ± 0.01 –0.11 ± 0.01Красноуфимская 58 0.38 ± 0.12 0.34 ± 0.08 0.39 ± 0.08 –0.09 ± 0.09 0.05 ± 0.05 0.12 ± 0.11 0.24 ± 0.09Кукморская 24 0.33 ± 0.14 0.28 ± 0.09 0.33 ± 0.11 –0.12 ± 0.10 0.13 ± 0.08 0.22 ± 0.15 0.39 ± 0.10Нижнекамская 12 0.19 ± 0.13 0.29 ± 0.05 0.41 ± 0.07 0.35 ± 0.18 0.65 ± 0.10 0.46 ± 0.05 0.55 ± 0.06Мамадышская 16 0.39 ± 0.11 0.37 ± 0.11 0.38 ± 0.11 –0.04 ± 0.04 –0.03 ± 0.02 0.01 ± 0.01 0.03 ± 0.02Малопургинская 26 0.23 ± 0.09 0.27 ± 0.08 0.29 ± 0.10 0.13 ± 0.10 0.23 ± 0.08 0.11 ± 0.04 0.18 ± 0.06Шарканская 34 0.34 ± 0.09 0.38 ± 0.08 0.38 ± 0.07 0.08 ± 0.06 0.10 ± 0.05 0.02 ± 0.01 0.04 ± 0.03
Камбарская 46 0.24 ± 0.12 0.29 ± 0.12 0.29 ± 0.12 0.17 ± 0.05 0.17 ± 0.05 0.01 ± 0.01 0.01 ± 0.01
Можгинская 22 0.26 ± 0.11 0.27 ± 0.11 0.27 ± 0.11 0.03 ± 0.02 –0.01 ± 0.01 –0.03 ± 0.02 –0.06 ± 0.01
Завьяловская 39 0.33 ± 0.10 0.32 ± 0.08 0.34 ± 0.09 0.01 ± 0.01 0.09 ± 0.04 0.08 ± 0.02 0.15 ± 0.03

ГЕНЕТИКА том 52 № 8 2016
МОЛЕКУЛЯРНО-ГЕНЕТИЧЕСКИЙ АНАЛИЗ 935
Примечание. Ho – усредненная наблюдаемая гетерозиготность субпопуляций; Hs – усредненная ожидаемая гетерозигот-ность субпопуляций; Ht – общее генное разнообразие всей популяции в целом; Fis– коэффициент инбридинга особи отно-сительно субпопуляции; Fit – коэффициент инбридинга особи относительно всей популяции в целом; Fst – коэффициент ин-бридинга выборки относительно всей популяции в целом (межпопуляционый компонент изменчивости, подразделенностьпопуляции); R – коэффициент родства; σ – стандартная ошибка.
Якшур-Бодьинская
33 0.27 ± 0.10 0.31 ± 0.12 0.31 ± 0.12 0.13 ± 0.03 0.15 ± 0.03 0.02 ± 0.01 0.04 ± 0.01
Чебоксарская 14 0.36 ± 0.14 0.30 ± 0.11 0.30 ± 0.10 –0.17 ± 0.03 –0.25 ± 0.02 –0.06 ± 0.01 –0.16 ± 0.01Кирово-Чепецкая 20 0.39 ± 0.12 0.44 ± 0.04 0.43 ± 0.04 0.12 ± 0.10 0.07 ± 0.07 –0.05 ± 0.01 –0.10 ± 0.01Даровская 20 0.63 ± 0.07 0.53 ± 0.05 0.52 ± 0.05 –0.19 ± 0.07 –0.23 ± 0.07 –0.04 ± 0.01 –0.10 ± 0.01Орловская 14 0.50 ± 0.17 0.39 ± 0.11 0.38 ± 0.11 –0.27 ± 0.09 –0.33 ± 0.08 –0.05 ± 0.01 –0.14 ± 0.01Кильмезская 10 0.48 ± 0.18 0.36 ± 0.11 0.35 ± 0.10 –0.34 ± 0.10 –0.40 ± 0.09 –0.04 ± 0.01 –0.13 ± 0.02Майкопская 15 0.19 ± 0.10 0.24 ± 0.12 0.24 ± 0.12 0.25 ± 0.04 0.24 ± 0.04 –0.02 ± 0.02 –0.03 ± 0.02Сочинская 32 0.18 ± 0.10 0.17 ± 0.08 0.24 ± 0.10 0.05 ± 0.04 0.48 ± 0.15 0.46 ± 0.15 0.61 ± 0.17Мукачевская 17 0.26 ± 0.07 0.24 ± 0.08 0.30 ± 0.10 –0.09 ± 0.07 0.16 ± 0.07 0.22 ± 0.03 0.39 ± 0.05
Локальная популяция N Ho ± σ Hs ± σ Ht ± σ Fis ± σ Fit ± σ Fst ± σ R ± σ
Таблица 3. Окончание
кументирующей системе DocPrint Vilber Lour-mat (Франция).
Оценка доли пчелиных семей южного проис-хождения по митохондриальному геному в ло-кальных популяциях A. m. mellifera Урала и По-волжья была выполнена на основе данных по по-лиморфизму локуса COI-COII мтДНК, которыйобладает значительной вариабельностью длины,где наименьший фрагмент (Q) характеризуетпроисхождение по материнской линии от пчелюжных подвидов A. m. caucasica, A. m. carpatica, авсе остальные фрагменты (PQQ и PQQQ) боль-шего размера – от темной лесной пчелы подвидаA. m. mellifera [25].
Оценка уровня интрогрессии южных генов вядерном геноме локальных популяций пчелA. m. mellifera Урала и Поволжья была выполненана основе данных полиморфизма микросател-литных локусов с использованием программыSTRUCTURE 2.3.4 на основе байесовского ана-лиза (Bayesian analysis) с применением методакластеризации Монте-Карло с цепями Маркова(Monte Carlo Markov Chain) (MCMC) [26] при за-данном числе кластеров K=2 с использованиеммодели смешивания (Admixture model) и повтор-ности MCMC (iteration) 5000.
Статистический анализ полученных резуль-татов проводили с использованием программFSTAT 2.9.3.2, GENEPOP 4.2.2, POPULA-TIONS 1.2.28, STRUCTURE 2.3.4, STATISTICA8.0, MICROSOFT EXCEL 2010.
РЕЗУЛЬТАТЫДля локальных популяций пчел Урала, Повол-
жья, Кавказа и Карпат были рассчитаны значе-
ния гетерозиготности, коэффициентов инбри-динга и родства на основе анализа полиморфизмамикросателлитных локусов (табл. 3).
Гетерозиготность – это доля особей в популя-ции, гетерозиготных по изучаемым локусам. Дляпопуляции каждого вида существует оптималь-ный уровень гетерозиготности. Понижение и по-вышение значения гетерозиготности в популя-ции относительно оптимального значения можетпривести к неблагоприятным последствиям.Значения гетерозиготности в локальных попу-ляциях местных пчел Урала и Поволжья былисходны с европейскими популяциями пчел под-вида A. m. mellifera эволюционной ветви М [6, 27–29] и выше по сравнению с локальными популя-циями южных пчел с Кавказа и Карпат (подвидыA. m. caucasica, A. m. carpatica эволюционной ветвиС). Значения гетерозиготности в локальных по-пуляциях пчел Италии (A. m. Ligustica) [30], (Сер-бии (A. m. carnica эволюционной ветви С) [31],Китая (A. m. ligustica эволюционной ветви С) [32],Турции (A. m. anatoliaca эволюционной ветви О)[33], Пуэрто-Рико (африканизированные пчелыэволюционной ветви А) [34] оказались немноговыше, по сравнению с локальными популяциямиместных пчел Урала и Поволжья.
Коэффициент инбридинга – вероятность то-го, что аллели изучаемых локусов идентичны попроисхождению. Положительные значения ко-эффициента инбридинга характеризуют преобла-дание близкородственного скрещивания в попу-ляции, а отрицательные значения – отдаленного.Коэффициенты инбридинга в локальных популя-циях местных пчел Урала и Поволжья были сход-ны с европейскими популяциями пчел подвидаA. m. mellifera эволюционной ветви М [6, 27–29] и

936
ГЕНЕТИКА том 52 № 8 2016
ИЛЬЯСОВ и др.
Таблица 4. Доля интрогрессии по ядерному и митохондриальному геномам в локальных популяциях пчел Урала,Поволжья, Кавказа и Карпат, рассчитанная на основе анализа полиморфизма микросателлитных локусов и ло-куса COI-COII мтДНК
Локальная популяция NЯдерный геном Митохондриальный геном
M* C* M C
Абзелиловская 90 0.23 0.77 0.37 0.63Альшеевская 35 0.21 0.79 0.31 0.69Баймакская 70 0.34 0.66 0.57 0.43Балтачевская 36 0.98 0.02 1.00 0.00Белебеевская 16 0.36 0.64 0.44 0.56Бирская 91 0.75 0.25 0.79 0.21Бурзянская 326 0.96 0.04 1.00 0.00Белорецкий 114 0.62 0.38 0.63 0.37Гафурийская 62 0.35 0.66 0.40 0.60Зилаирская 141 0.30 0.70 0.54 0.46Иглинская 197 0.62 0.38 0.75 0.25Ишимбайская 226 0.35 0.65 0.50 0.50Караидельская 132 0.80 0.20 0.86 0.14Кушнаренковская 37 0.86 0.14 0.92 0.08Куюргазинская 61 0.08 0.92 0.15 0.85Мелеузовская 73 0.12 0.88 0.15 0.85Мишкинская 55 0.93 0.07 0.95 0.06Татышлинская 200 0.97 0.03 0.99 0.02Янаульская 100 0.98 0.02 1.00 0.00Уфимская 30 0.36 0.64 0.47 0.53Учалинская 10 0.19 0.81 0.20 0.80Чишминская 15 0.17 0.83 0.20 0.80Чекмагушевская 62 0.61 0.39 0.61 0.39Хайбуллинская 130 0.06 0.94 0.13 0.87Уинская 59 0.85 0.15 0.85 0.15Ординская 25 0.95 0.05 0.96 0.04Частинская 28 0.99 0.01 1.00 0.00Нытвенская 18 0.99 0.01 1.00 0.00Осинская 38 0.99 0.01 1.00 0.00Пермская 76 0.93 0.07 0.95 0.05Юсьвенская 37 0.99 0.01 1.00 0.00Красновишерская 41 0.99 0.01 1.00 0.00Усольская 20 0.97 0.03 1.00 0.00Добрянская 20 0.96 0.04 1.00 0.00Красноуфимская 58 0.31 0.70 0.26 0.74Кукморская 24 0.60 0.41 0.58 0.42Нижнекамская 12 0.16 0.85 0.67 0.33Мамадышская 16 0.97 0.03 1.00 0.00Малопургинская 26 0.92 0.08 0.92 0.08Шарканская 34 0.79 0.21 0.79 0.21Камбарская 46 0.98 0.02 1.00 0.00Можгинская 22 0.99 0.01 1.00 0.00

ГЕНЕТИКА том 52 № 8 2016
МОЛЕКУЛЯРНО-ГЕНЕТИЧЕСКИЙ АНАЛИЗ 937
* М и С – эволюционные ветви.
Завьяловская 39 0.64 0.36 0.62 0.39Якшур-Бодьинская 33 0.90 0.10 0.91 0.09Чебоксарская 14 0.98 0.02 1.00 0.00Кирово-Чепецкая 20 0.23 0.77 0.70 0.30Даровская 20 0.37 0.63 0.50 0.50Орловская 14 0.53 0.48 1.00 0.00Кильмезская 10 0.94 0.06 1.00 0.00Майкопская 15 0.01 0.99 0.00 1.00Сочинская 32 0.01 0.99 0.00 1.00Мукачевская 17 0.01 0.99 0.00 1.00
Локальная популяция NЯдерный геном Митохондриальный геном
M* C* M C
Таблица 4. Окончание
ниже по сравнению с локальными популяциямиюжных пчел с Кавказа и Карпат. Значения коэф-фициентов инбридинга в локальных популяцияхпчел Италии [30], Сербии [31], Китая [32], Тур-ции [33], Пуэрто-Рико [34] оказались немногониже по сравнению с локальными популяциямиместных пчел Урала и Поволжья.
Коэффициент родства – показатель родства впопуляции, характеризует долю генов, идентич-ных по происхождению, среди пчелиных семей.Локальные популяции с неродственными пчели-ными семьями будут характеризоваться коэффи-циентом родства менее 0.25, а с родственными –более 0.25. Коэффициенты родства в локальныхпопуляциях местных пчел Урала и Поволжья бы-ли сходны с европейскими популяциями пчелподвида A. m. mellifera эволюционной ветви М[35] и ниже по сравнению с локальными популя-циями южных пчел с Кавказа и Карпат. Значениякоэффициентов родства в локальных популяцияхпчел Германии (A. m. carnica), Италии (A. m. ligus-tica) оказались сходными с локальными популя-циями местных пчел Урала и Поволжья.
Сходство генетических показателей локаль-ных популяциий местных пчел Урала и Поволжьяс европейскими популяциями A. m. mellifera эво-люционной ветви М подтверждает гипотезу оединстве происхождения всех разрозненных наданный момент популяций темной лесной пчелыв Европе. Небольшие различия генетических по-казателей между разными локальными популя-циями связаны с климатическими факторами иособенностями разведения пчел. Сильные откло-нения генетических показателей от оптимальныхзначений, несомненно, вызваны процессами ги-бридизации с южными подвидами пчел эволюци-онной ветви С.
ОБСУЖДЕНИЕДля локальных популяций пчел Урала, Повол-
жья были рассчитаны доли генов южных подви-дов по ядерному и митохондриальному геномам(табл. 4).
Гистограмма (plot), построенная по даннымполиморфизма микросателлитных локусов, на-глядно показывает сохранение ядерного геноматемной лесной пчелы A. m.mellifera в локальныхпопуляциях Урала и Поволжья (рис. 1).
Минимальным уровнем интрогрессии южныхгенов по ядерному геному характеризовались ло-кальные популяции темной лесной пчелы Рес-публики Башкортостан (бурзянская, татышлин-ская, янаульская, балтачевская, караидельская,мишкинская, кушнаренковская), Пермского края(ординская, осинская, частинская, добрянская,красновишерская, юсьвенская, нытвенская,усольская, уинская, пермская), Республики Уд-муртия (камбарская, можгинская, якшур-бо-дьинская, малопургинская), Республики Татар-стан (мамадышская), Республики Чувашия (че-боксарская), Кировской области (кильмезская).
На основе попарных значений Fst между ло-кальными популяциями темной лесной пчелыA. m. mellifera Урала и Поволжья, полученных порезультатам анализа полиморфизма микросател-литных локусов, была построена дендрограммаметодом кластеризации ближайшего соседа, ко-торая подтверждает данные уровня интрогрессиипо ядерному геному. Локальные популяции пчелюжных подвидов A. m. caucasica и A. m. carpaticaКавказа и Карпат были использованы для сравне-ния в качестве внешней группы (рис. 2).
На дендрограмме локальные популяции юж-ных пчел Кавказа и Карпат четко дифференциру-ются и располагаются отдельно (группа I). Ло-кальные популяции местных пчел Урала и По-

938
ГЕНЕТИКА том 52 № 8 2016
ИЛЬЯСОВ и др.
Рис
. 1. Г
исто
грам
ма
уров
ня и
нтро
грес
сии
гено
в ю
жны
х по
двид
ов в
лок
альн
ых
попу
ляци
ях т
емно
й ле
сной
пче
лы У
рала
и П
овол
жья
, пос
трое
нная
на
осно
ве а
на-
лиза
пол
имор
физ
ма
мик
роса
телл
итны
х ло
кусо
в. Ч
ерны
й цв
ет х
арак
тери
зует
сод
ерж
ание
ген
ов м
естн
ых
пчел
под
вида
A. m
. mel
lifer
a, а
сер
ый
цвет
– ю
жны
х пч
елпо
двид
ов A
. m. c
auca
sica
и A
. m. c
arpa
tica.
100
Абз
елил
овск
ая%
N =
90
80 60 40 20 0
Бай
мак
ская
N =
70
Аль
шее
вска
яN
= 3
5Б
ирск
аяN
= 9
1
Бал
таче
вска
яN
= 3
6 Бел
ебее
вска
яN
= 1
6
Игл
инск
аяN
= 1
97Га
фур
ийск
аяN
= 6
2Зи
лаир
ская
N =
141
Куш
наре
нков
ская
N =
37
Тат
ыш
линс
кая
N =
200
Миш
кинс
кая
N =
55
Мел
еузо
вска
яN
= 7
3
Кую
ргаз
инск
аяN
= 6
1
Уин
ская
N =
59
Орд
инск
аяN
= 2
5Час
тинс
кая
N =
28
Ны
твен
ская
N =
18 О
синс
кая
N =
38
Кра
снов
ишер
ская
N =
41
Пер
мск
аяN
= 7
6Ю
сьве
нска
яN
= 3
7
Дар
овск
аяN
= 2
0У
соль
ская
N =
20
Доб
рянс
кая
N =
20
Кра
сноу
фим
ская
N =
58К
укм
орск
аяN
= 2
4
Ниж
нека
мск
аяN
= 1
2
Мам
ады
шск
аяN
= 1
6
Мал
опур
гинс
кая
N =
26Ш
арка
нска
яN
= 3
4 Кам
барс
кая
N =
46
Хай
булл
инск
аяN
= 1
30Я
наул
ьска
яN
= 1
00У
фим
ская
N =
30Уча
линс
кая
N =
10
Чек
маг
ушев
ская
N =
62
Чиш
мин
ская
N =
15
Мож
гинс
кая
N =
22 За
вьял
овск
аяN
= 3
9
N =
33
Чеб
окса
рска
яN
= 1
4
N =
20
Орл
овск
аяN
= 1
4
Кил
ьмез
ская
N =
10
Соч
инск
аяN
= 3
2
Май
копс
кая
N =
15
Мук
ачев
ская
N =
17
Эво
люци
онна
я ве
твь
CЭ
волю
цион
ная
ветв
ь М
Бур
зянс
кая
N =
326
Бел
орец
кая
N =
114
Иш
имба
йска
яN
= 2
26К
араи
дель
ская
N =
226

ГЕНЕТИКА том 52 № 8 2016
МОЛЕКУЛЯРНО-ГЕНЕТИЧЕСКИЙ АНАЛИЗ 939
волжья подразделяются на две группы (II и III).В группе III преобладают локальные популяциипчел Республики Башкортостан, имеющие зна-чительный уровень интрогрессии генов южныхподвидов эволюционной ветви С. В группе II ло-кальные популяции пчел Урала и Поволжья под-разделяются на две подгруппы – II-1 и II-2.В подгруппе II-1 представлены примерно поров-ну локальные популяции пчел Урала и Поволжья,имеющие незначительный уровень интрогрессиигенов южных подвидов. В подгруппе II-2 преоб-ладают локальные популяции темной леснойпчелы Урала, характеризующиеся минимальнымуровнем интрогрессии генов южных подвидов.
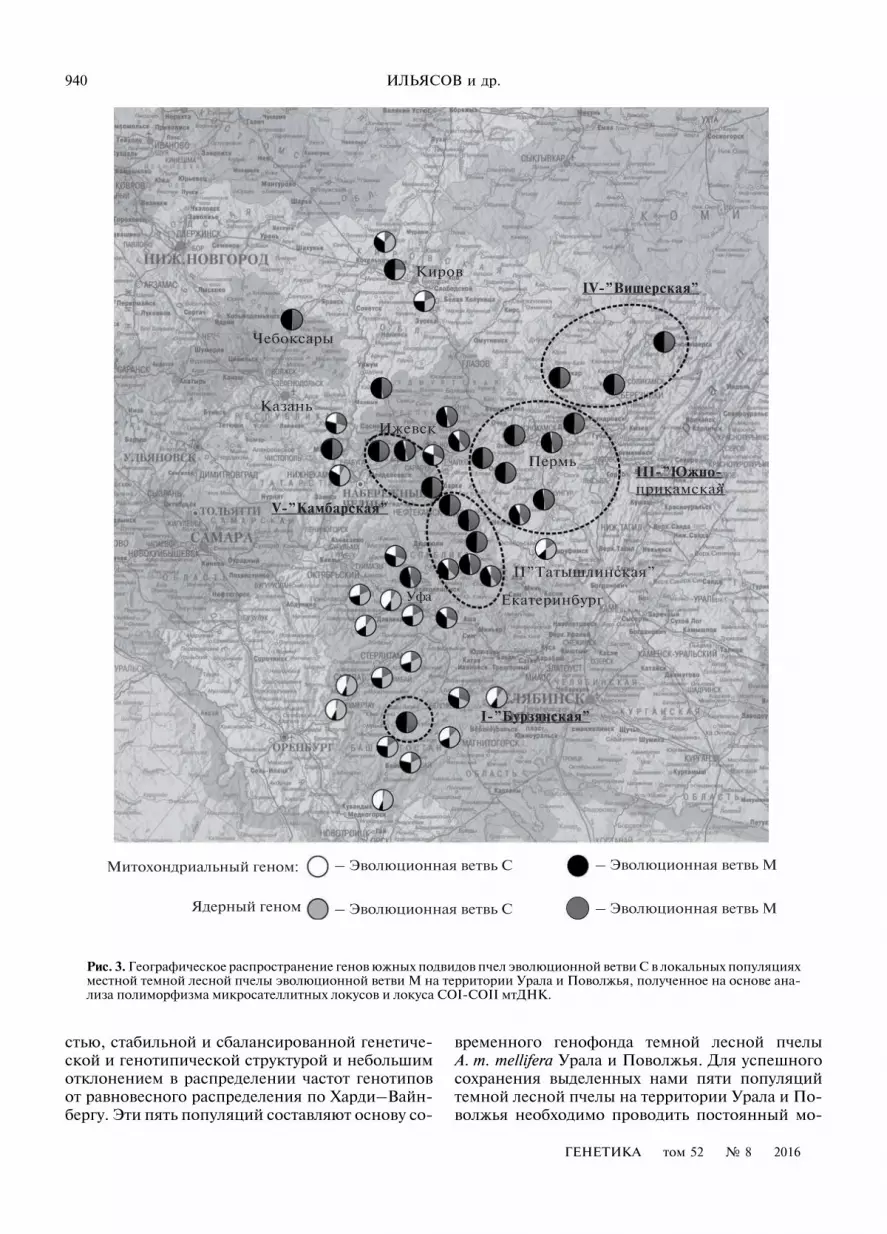
Для проведения геногеографического анализасекторные диаграммы, отражающие доли интро-грессии генов южных подвидов пчел эволюцион-ной ветви С в локальных популяциях темной лес-ной пчелы Урала и Поволжья эволюционной ветвиМ, были расположены по местам их локализациина географической карте (рис. 3).
На территории Урала и Поволжья гены темнойлесной пчелы A. m. mellifera сохранились неравно-мерно – наблюдается тенденция возрастания до-ли генов эволюционной ветви М по ядерному имитохондриальному геномам с юга на север.В южной части Урала и Поволжья наблюдаетсяединственная сохранившаяся в чистоте локаль-ная популяция темной лесной пчелы – бурзян-ская, которая находится под охраной заказника“Алтын Солок”, заповедника “Шульган-Таш” инационального парка “Башкирия”.
Исходя из пространственного распределениялокальных популяций, характеризующихся ми-нимальной интрогрессией генов южных подви-дов по ядерному и митохондриальному геномам,нам удалось выделить на территории Урала и По-волжья пять сохранившихся популяций (резерва-тов) темной лесной пчелы A. m. mellifera: бурзян-ская, татышлинская, южно-прикамская, вишер-ская и камбарская. Эти популяции на данныймомент характеризуются достаточной численно-
Рис. 2. Дендрограмма генетических отношений локальных популяций пчел Урала и Поволжья, построенная на основеанализа полиморфизма микросателлитных локусов.
Сочинская (КК)Мукачевская (ЗО)Майкопская (РА)
I
II
III
Орловская (КО)
Можгинская (РУ)Белорецкая (РБ)Кукморская (РТ)
Иглинская (РБ)Усольская (ПК)
Добрянская (ПК)Чебоксарская (РЧ)
Нижнекамская (РТ)Кильмезская (КО)
Мамадышская (РТ)
Кушнаренковская (РБ)Караидельская (РБ)
Бирская (РБ)Завьяловская (РУ)
Камбарская (РУ)Янаульская (РБ)
Частинская (ПК)Нытвенская (ПК)Юсьвенская (ПК)
Осинская (ПК)Красновишерская (ПК)
Уинская (ПК)Бурзянская (РБ)
Шарканская (РУ)
Пермская (ПК)Ординская (ПК)
Малопургинская (РУ)
Чишминская (РБ)Уфимская (РБ)
Хайбуллинская (РБ)Куюргазинская (РБ)
Чекмагушевская (РБ)Балтачевская (РБ)
Татышлинская (РБ)Якшур-Бодьинская (РУ)
Мишкинская (РБ)
Учалинская (РБ)Мелеузовская (РБ)
Красноуфимская (СО)
Белебеевская (РБ)Гафурийская (РБ)
Ишимбайская (РБ)Зилаирская (РБ)Даровская (КО)
()
()
(
Баймакская (РБ)
Абзелиловская (РБ)Альшеевская (РБ)
)
1.41.21.00.80.60.40.20
940
ГЕНЕТИКА том 52 № 8 2016
ИЛЬЯСОВ и др.
стью, стабильной и сбалансированной генетиче-ской и генотипической структурой и небольшимотклонением в распределении частот генотиповот равновесного распределения по Харди–Вайн-бергу. Эти пять популяций составляют основу со-
временного генофонда темной лесной пчелыA. m. mellifera Урала и Поволжья. Для успешногосохранения выделенных нами пяти популяцийтемной лесной пчелы на территории Урала и По-волжья необходимо проводить постоянный мо-
Рис. 3. Географическое распространение генов южных подвидов пчел эволюционной ветви С в локальных популяцияхместной темной лесной пчелы эволюционной ветви М на территории Урала и Поволжья, полученное на основе ана-лиза полиморфизма микросателлитных локусов и локуса COI-COII мтДНК.
Чебоксары
Казань
Киров
Ижевск
Пермь
Уфа
II”Татышлинская”
Екатеринбург
Митохондриальный геном:
Ядерный геном:
– Эволюционная ветвь C – Эволюционная ветвь М
– Эволюционная ветвь C – Эволюционная ветвь М
прикамская”

ГЕНЕТИКА том 52 № 8 2016
МОЛЕКУЛЯРНО-ГЕНЕТИЧЕСКИЙ АНАЛИЗ 941
ниторинг и управлять их генофондом в соответ-ствии с принципами популяционной генетики.
Работа выполнена при финансовой поддержкегранта РФФИ 14-04-97084 р_поволжье_а на обо-рудовании ЦКП “Биомика” отделения биохими-ческих методов исследований и нанобиотехноло-гии РЦКП “Агидель”.
СПИСОК ЛИТЕРАТУРЫ1. Sheppard W.S., Meixner M.D. Apis mellifera pomonella,
a new honey bee subspecies from Central Asia // Api-dologie. 2003. V. 34. P. 367–375.
2. Meixner M.D., Leta M.A., Koeniger N., Fuchs S. Thehoney bees of Ethiopia represent a new subspecies ofApis mellifera - Apis mellifera simensis n. ssp. // Apidol-ogie. 2011. V. 42. P. 425–437.
3. Николенко А.Г., Поскряков А.В. Полиморфизм ло-куса COI-COII митохондриальной ДНК Apis mellif-era L. на Южном Урале // Генетика. 2002. Т. 38.№ 4. С. 458–462.
4. Büchler R., Costa C., Hatjina F. et al. The influence ofgenetic origin and its interaction with environmental ef-fects on the survival of Apis mellifera L. colonies in Eu-rope // J. Apic. Res. 2014. V. 53. P. 205–214.
5. Jensen A.B., Palmer K.A., Boomsma J.J., Pedersen B.V.Varying degrees of Apis mellifera ligustica introgressionin protected populations of the black honeybee, Apismellifera mellifera, in northwest Europe // Mol. Ecol.2005. V. 14. P. 93–106.
6. Soland-Reckeweg G., Heckel G., Neumann P. et al. Geneflow in admixed populations and implications for theconservation of the Western honeybee, Apis mellifera //J. Insect Conserv. 2009. V. 13. P. 317–328.
7. Muñoz I., Dall’Olio R., Lodesani M., De La Rúa P. Pop-ulation genetic structure of coastal Croatian honeybees(Apis mellifera carnica) // Apidologie. 2009. V. 40.P. 617–626.
8. Nedić N., Francis R.M., Stanisavljević L. et al. Detectingpopulation admixture in the honey bees of Serbia //J. Apic. Res. 2014. V. 53. P. 303–313.
9. Uzunov A., Meixner M.D., Kiprijanovska H. et al. Ge-netic structure of Apis mellifera macedonica populationbased on microsatellite DNA polymorphism // J. Apic.Res. 2014. V. 53. № 2. P. 288–295.
10. Jensen A.B., Pedersen B.V. Honeybee Conservation: acase story from Læsø island, Denmark // Beekeepingand conserving biodiversity of honeybee. Sustainablebee breeding / Eds Lodesani M., Costa C. HebdenBridge: Northern Bee Books, 2005. P. 142–164.
11. Maul V., Hähnle A. Morphometric studies with purebred stock of Apis mellifera carnica Pollmann from Hes-sen // Apidologie. 1994. V. 25. P. 119–132.
12. Ильясов Р.А., Петухов А.В., Поскряков А.В., Нико-ленко А.Г. Локальные популяции Apis mellifera mel-lifera L. на Урале // Генетика. 2007. Т. 43. № 6.С. 855–858.
13. Ильясов Р.А., Косарев М.Н., Юмагужин Ф.Г. Бур-зянская бортевая пчела и бортевое пчеловодствона Южном Урале // Пчеловодство. 2015. № 7.С. 12–15.
14. Петухов А.В., Шураков А.И., Еськов Е.К. и др. Мор-фологическая характеристика среднерусских пчелверхнекамской популяции // Пчеловодство. 1996.№ 5. С. 8–10.
15. Никоноров Ю.М., Беньковская Г.В., Поскряков А.В.и др. Использование метода ПЦР для контроля чи-стопородности пчелосемей Apis mellifera mellifera L.в условиях Южного Урала // Генетика. 1998. Т. 34.№ 11. С. 1574–1577.
16. Ильясов Р.А., Поскряков А.В. Филогенетика подвидовApis mellifera // Пчеловодство. 2006. № 7. С. 18–19.
17. Ильясов Р.А., Поскряков А.В., Колбина Л.М., Нико-ленко А.Г. Сохранение Apis mellifera mellifera L. в Уд-муртской республике // Пчеловодство. 2007. № 6.С. 13–14.
18. Колбина Л.М., Непейвода С.Н., Воробьева С.Л. и др.Генетическая дифференциация популяций Apismellifera L. в Удмуртской Республике // Аграрнаянаука Евро-Северо-Востока. 2011. Т. 6. № 25.С. 46–50.
19. Брандорф А.З., Ивойлова М.М., Ильясов Р.А. и др.Популяционно-генетическая дифференциациямедоносных пчел Кировской области // Пчело-водство. 2012. № 7. С. 14–16.
20. Ильясов Р.А., Поскряков А.В., Петухов А.В., Нико-ленко А.Г. Генетическая дифференциация локаль-ных популяций темной лесной пчелы Apis melliferamellifera L. на Урале // Генетика. 2015б. Т. 51. № 7.С. 792–798.
21. Ильясов Р.А., Поскряков А.В., Николенко А.Г. НовыеSNP маркеры в гене вителлогенина Vg медоноснойпчелы для идентификации Аpis mellifera mellifera L. //Генетика. 2015, Т. 51. № 2. С. 194–199.
22. Estoup A., Garnery L., Solignac M., Cornuet J.- M. Mi-crosatellite variation in honey bee (Apis mellifera L.)populations: hierarchical genetic structure and test ofthe infinite allele and stepwise mutation models //Genetics. 1995. V. 140. P. 679–695.
23. Haberl M., Tautz D. Tri- and tetranucleotide microsat-ellite loci in honey bees (Apis mellifera) – a stap towardsquantitative genotyping // Mol. Ecol. 1999. V. 8.P. 1351–1362.
24. Solignac M., Vautrin D., Loiseau A. et al. Five hundredand fifty microsatellite markers for the study of thehoneybee (Apis mellifera L.) genome // Mol. Ecol.Notes. 2003. V. 3. P. 307-311.
25. Garnery L., Solignac M., Celebrano G., Cornuet J.-M.A simple test using restricted PCR-amplified mito-chondrial DNA to study the genetic structure of Apismellifera L. // Experientia. 1993. V. 49. P. 1016–1021.
26. Pritchard J.K., Stephens M., Donnelly P. Inference ofpopulation structure using multilocus genotype data //Genetics. 2000. V. 155. P. 945–959.
27. Franck P., Garnery L., Solignac M., Cornuet J. M. Theorigin of West European subspecies of honeybees (Apismellifera): new insights from mitodhondrial DNA andmicrosatellite data // Evolution. 1998. V. 52. № 4. P.1119–1134.
28. Garnery L., Franck P., Baudry E. et al. Genetic biodi-versity of the West European honeybee (Apis melliferamellifera and A. m. iberica). I. Mitochondrial DNA //Genet. Sel. Evol. 1998. V. 30. P. 31–47.

942
ГЕНЕТИКА том 52 № 8 2016
ИЛЬЯСОВ и др.
29. Franck P., Garnery L., Loiseau A. et al. Genetic diversi-ty of the honeybee in Africa: microsatellite and mito-chondrial data // Heredity. 2001. V. 86. P. 420–430.
30. Franck P., Garnery L., Celebrano G. et al. Hybrid originsof the Italian honeybees, Apis mellifera ligustica and A.m. sicula // Mol. Ecol. 2000. V. 9. P. 907–923.
31. Pihler I., Kiprijanovska H., Plavsa N. et al. Population-genetical characteristics of the bee population of Vojvo-dina // Genetika. 2014. V. 64. № 1. P. 219–226.
32. Yin L., Ji T., Chen G., Peng W. Genetic characterizationof three breeds of high royal jelly producing honeybee(Apis mellifera ligustica) in China // African J. Agricul-tur. Res. 2011. V. 6. № 2. P. 331–337.
33. Bodur Ç. Genetic structure analysis of honeybee popu-lations based on microsatellites: A thesis for the degreeof doctor of philosophy in biology. The graduate schoolof natural and applied sciences of middle east technicaluniversity. A 2005. 116 p.
34. Galindo-Cardona A., Acevedo-Gonzales J.P., Rivera-Marchand B., Giray T. Genetic structure of the gentleAfricanized honey bee population (gAHB) in PuertoRico // BMC Genetics. 2013. V. 14. P. 1–12.
35. Estoup A., Solignac M., Cornuet J.-M. Precise assess-ment of the number of patrilines and of genetic related-ness in honeybee colonies // Proc. R. Soc. London. B.Biolo. Scie. 1994. V. 258. P. 1–7.
Molecular Genetic Analysis of Five Extant Reserves of Black HoneybeeApis melifera melifera in the Urals and the Volga Region
R. A. Ilyasova, A. V. Poskryakova, A. V. Petukhovb, and A. G. Nikolenkoa
aInstitute of Biochemistry and Genetics, Ufa Scientific Center, Russian Academy of Sciences,Ufa, 450054, Russia
e-mail: [email protected] of Zoology, Perm State Humanitarian Pedagogical University, Perm, 614990 Russia
Abstract—Local populations of the black honeybee Apis melifera melifera from the Urals and the Volga regionwere examined in comparison with local populations of southern honeybee subspecies A. m. caucasica andA. m. carpatica from the Caucasus and the Carpathians. Genetic analysis was performed on the basis of thepolymorphism of nine microsatellite loci of nuclear DNA and the mtDNA COI–COII locus. On the territoryof the Urals and the Volga region, five extant populations (reserves) of the black honeybee A. m. mellifera wereidentified, including the Burzyanskaya, Tatyshlinskaya, Yuzhno-Prikamskaya, Visherskaya, and Kam-barskaya populations. These five populations are the basis of the modern gene pool of the black honeybeeA. m. mellifera from the Urals and the Volga region. The greatest proportion of the remaining indigenous genepool of A. m. mellifera (the core of the gene pool of the population of A. m. mellifera) is distributed over theentire territory of Perm krai and the north of the Republic of Bashkortostan. For the population of A. m. mel-lifera from the Urals and the Volga region, the genetic standards were calculated, which will be useful for fu-ture population studies of honeybees. English translation of the paper published in Russian Journal of Genet-ics, 2016, Vol. 52, No. 8, is available ONLINE by subscription from: http://www.springer.com/, http://link.springer.com.
Keywords: black honeybee Apis melifera melifera, conservation of indigenous gene pool, COI–COIImtDNA, microsatellite loci, microsatellite loci, extant reserves, genetic standard, heterozygosity.