molecular engineering: applicationsto the clinical laboratory
TRANSCRIPT

GUN. CHEM. 39/9, 1988-1997 (1993)
1988 CLINICAL CHEMISTRY, Vol. 39, No. 9, 1993
Molecular Engineering: Applications to the Clinical LaboratoryRobert G. Hamilton
Advances in cellular and molecular biology methods haveled to the molecular engineering of novel human biomol-ecules, some of which have been successfully applied tothe documentation of clinical laboratory assays. Here Idescribe the use of engineered chimeric antibodies in theclinical immunology laboratory in three principal applica-tions: (a) as reference proteinsto document the specificityof clinical assay reagents, be used as reagent-grade pu-rifled antigens, and facilitate the epitope mapping of an-tibody reagents; (b) as calibration proteins to assignmass/volume estimates to proposed antibody standards;and (c) as interference proteins to study the effects ofnaturally occurring autoantibodies on the accuracy andsensitivity of current clinical assays. The model recombi-nant proteins used for these illustrations are chimericantibodies with a defined V-region specfficity for one oftwo haptens (nitrophenylor dansyl) and C-region domainscovering a spectrum of human isotypes. I also describe apanel of mutant human lgGl-4 anti-dansyl chimeric an-tibodies that have been genetically engineered withswapped, deleted, or point-mutated wild-type C-regionexons and used as specialized reagents for mapping theepitopes to which clinically used human lgG-specificmonoclonal antibodies bind. Finally, the use of a recom-binant human IgGi anti-human IgE Fc chimeric antibodyto simulate human lgG anti-IgE autoantibody interferencein assays of total serum IgE is investigated.
IndexIng Terms: monoclonal antibodies . chimeric (human-mouse) antibodies immunoglobulins . autoantibodiesrecombinant proteins . reference materials . calibration
Molecular engineering can be defined within the con-text of biomolecules as the science and mathematics bywhich the elemental organization of proteins, carbohy-drates, lipids, and nucleic acids is modified to makethem more useful to humans in structures, processes,and products (1, 2). Scientists have combined tech-niques from protein chemistry and cellular and molec-ular biology with mathematics and computer modelingto design and generate biomolecules for use as replace-ment hormones and therapeutic drugs. Investigators in
Clinical Immunology Division, Johns Hopkins Asthma and Al-tergy Center, Johns Hopkins University School of Medicine, Bal-timore, Ml) 21224.
Address for correspondence:Johns Hopkins Asthma and AllergyCenter, Johns Hopkins Dermatology, Allergy and Clinical Immu-nology(DACI) Reference Laboratory, 5501 HopkinsBayview Cir-cle, Baltimore, MD 21224. Fax 410-550-2030.
Received April 27, 1993; acceptedJune 14, 1993.
clinical laboratories have benefited from this work be-cause it has generated some novel recombinant biomol-ecules that are useful as ligands (standards, controls)and receptors in clinical assays.
Overviewof ClinicalLaboratoryApplications
Engineered biomolecules may have their greatest ap-plication as therapeutic agents, e.g., as hormones anddrugs (3), especially because some human proteins are
difficult to obtain in large quantities and in a purified,virus-free form. Moreover, molecular biology techniquesallow individual regions of the biomolecule (e.g., singleamino acids of a protein) to be modified by point-muta-tion of DNA, allowing the production of structural vari-ations that provide insight into molecular structure-function relationships. Investigators in the clinicallaboratory have not generally had a need for these so-phisticated and often expensive engineered biomole-cules. It has been relatively easy to obtain from humantissue and serum sources small quantities of purifiedhuman proteins for use as assay standards and controls.Moreover, monospecific antiserum for use in clinicalimmunoassays can be readily prepared in animals(mice, rabbits, goats, horses) and function well after
absorption and affinity purification. Thus, the primaryconcern of immunogenicity ofheterologous proteins thathas driven the production of engineered biomoleculesfor use in vivo has not been an issue with reagents thatare used exclusively in vitro in a laboratory setting. Inat least three areas, however, engineered biomoleculesare of particular value in the clinical laboratory: refer-ence proteins (e.g., purified proteins for specificity anal-ysis, reagent-grade purified antigens, and antigens forepitope mapping of antibody reagents), calibration pro-teins (e.g., heterologous antibody calibration reagents),and interference proteins (e.g., autoantibodies to add tobiological specimens to simulate natural autoantibodyinterference in clinical immunoassays). In this paper Ifocus on engineered human proteins as models for ifius-trating these applications of engineered biomolecules inthe clinical immunology laboratory.
Reference proteins. Engineered proteins can be usefulas reference proteins to test the specificity of clinicallaboratory reagents (e.g., antisera) when purified hu-man proteins are either technically difficult to obtain orexpensive to purify from serum (e.g., human 1gB) ortissue (e.g., human insulin). Additionally, identifyingsource sara with exceptionally high concentrations ofselect proteins for subsequent use in isolation can be

CLINICALCHEMISTRY, Vol. 39, No. 9, 1993 1989
difficult. For example, in a clinical immunology labora-tory that measures total concentrations of IgGi, IgG2,IgG3, and IgG4 in serum, it can be difficult both toidentify quantities of paraprotein-containing sara andto isolate the minor IgG subclasses (IgG3 or IgG4) fromother serum components to the required >99% purity.Later I describe a panel of human IgG1, IgG2, IgG3,IgG4, and IgE chimeric proteinswith defined isotypes
and allotypes. They are useful as reference materials inthe quality control of murine monoclonal antibodies(mAbs) used in clinical assays to quantify human 1gBand individual IgG subclasses in serum.’
Calibration proteins. Engineered human proteins canbe used as calibration proteins. Characterized humanchimeric antibodies with a defined isotype (IgGl-4,IgA2, IgE, 1gM) can be used to construct heterologouscalibration curves for a variety of human antibody im-munoassays. By using such a calibration curve, one canconvert the response signal obtained in the assay thatmeasures a particular antibody in a patient’s serum intoan estimate of the concentration of that antibody in thespecimen. For example, in a human IgG anti-Haemo-philus influenzae enzyme immunoassay, one can esti-mate the mass quantity per unit volume of IgGi, IgG2,IgG3, and IgG4 antibodies that react with insolubilizedantigen by cross-interpolation of response data from acalibration curve for the heterologous proteins.
Interference proteins. Engineered biomolecules maybe used as interference proteins to study the effects ofnaturally occurring autoantibodies on the performanceof clinical immunoassays for both ligands (antigens) andreceptors (antibodies). Engineered human antibodiesthat mimic autoantibody reactivities can be added tohuman sara containing a defined quantity of the ligandbeing measured. By varying the concentrations of theligand and the interfering autoantibody, one can exam-ine the extent to which the results of a clinical inunu-noassay are altered. This application is ifiustrated be-low in a study of naturally occurring human IgG auto-antibodies to human IgE that may interfere in assays oftotal serum IgE.
Reagents
Murine monoclonal antibodies. A panel of 29 mousemAbs with internationally documented specificity forhuman IgG, IgA, 1gM, 1gB, K and A light chains, orsecretory component was used to examine the use of thegenetically engineered chimeric antibodies as specificityand calibration reagents (4-7). The mouse isotype, hu-man Ig specificity, and clone number of the mAbs arelisted in Table 1. Hybridoma product (HP) 6014, forexample, is a mouse IgGi mAb to human IgG2 Fabproduced by clone 6014. All the mAbs were isolatedfrom ascites by the Hybridoma Reagent Lab., Balti-more, MD, by sequential chromatography on an anion-exchange resin (Whatman, Maidstone, UK) and hydrox-
‘Nonstandard abbreviations: mAb, monoclonal antibody; BSA,bovine serum albumin; HP, hybridoma product; V, variable; C,constant; NP, nitrophenyl; and H1B, H. influenzae type B.
Table 1. Human Ig-Specific Murine mAbsMouss
Clan. Isotyp. HumanIg sp.cflIcfty spscfflclty
HP60005 IgG2b lgG-PAN Fc-CH2HP6017 IgG2a IgG-PAN Fc-CH2HP6043 IgG2b lgG-PAN Fc-CH2HP8045 IgGi IgG-PAN Fd-CH1HP6046b IgGi IgG-PAN Fd-CH1HP6069b IgGi IgG1 Fc-CH2HP6070 IgGi IgGi Fc-CH2HP6091 IgG1 IgG1 Fc-CH2HP6002b IgG2b lgG2 Fc-CH2HP60141’ IgG1 lgG2 Fd-CH1HP60475 IgGi lgG3 Fd-hlngeHP6050 IgGi lgG3 Fd-hlngeHP6023 lgG3 IgG4 Fc-CH3HP6025b IgGi IgG4 Fc-CH3HP6019’ IgGi IgGl,3,4 Fc-CH2HP6O305 IgGi IgG2,3,4 Fc-CH2HP6058b IgGi IgGl,2,3 Fc-CH2HP6029 IgGi IgE FcHP6061 1gM IgE FcHP6081 IgG1 1gM FcHP6083 IgGi 1gM FcHP6084 IgGi 1gM FcHP6086 IgGi 1gM FcHP6111 IgGi IgA-PAN FcHP6123 IgG1 IgA-PAN FdHP6054 lgG2a A lightchain -
HP6062 lgG3 K lightchain -
HP6130 IgGi Secretorycomponent -
HP6141 IgGi Secretorycomponent -
a Designation of thedomain towhich theIgG-specfflc mAbs bind has been
obtained from ret. 11. Fd, heavy chain of F(ab’)2.b mAbs specific for humanI9Gthatwere selected as components for the
poly/monoclonalanti-human 190 reagent (see text).
ylapatite (Bio-GelHT; Bio-Rad, Richmond, CA) (8). Al-ter being concentrated to 2 gIL (Amicon, Beverly,MA;YM-lO membrane), the mAbs were either frozen at-20 #{176}Cfor future use or immediately biotinylatedwithbiotin-X-hydroxysuccinimide ester (biotin-X-HSE; Cal-biochem, La Jolla,CA) in diniethylformamide (30:1 bio-tin-HSE IgG molar ratio). Isoelectric focusing ixnmuno-blot spectrotype and the degree of inimunoreactivity tocharacterized human paraproteins were used to confirmthe isoelectric focusing spectrotype and specificity of themAbs (9, 10).
Chimeric human antibodies. The chimeric antibodieswere produced by manipulation of the DNA exons cod-ing for the Ig variable (V)- and constant (C)- regiondomains (Figure 1). Each chimeric protein discussedhere has a mouse V region that reacts with one of threeantigen speciflcitie8:mtrophenyl (NP), dansyl, or, in thecase of ChlO-10-3, the Fc region of human IgE. DNAcoding for the amino acids that form one of these mouseV regions is genetically linked to DNA that codes forone of the following “wild-type” human C-region do-mains: IgGi, IgG2, IgG3, IgG4, IgA2, 1gM, or IgE. Forthe descriptor code, antigen specificity, and isotype des-ignation of the antibody’s C-region domains (CH1,

Proteb,,’vi-ci-’
“1Mm,.. L C3iai,
Protein
Itin., i- au
d,im.nci-chuG...
Moias. H chU,
NVt/n.o suction
Iexpansion
tTWSfICtonS call tio.
VMous.-4tius cMsrlc astaod,
GEEGEE GEE
1990 CUNICALCHEMISTRY,Vol.39, No. 9, 1993
Protein/VH
CH2’
Cotranstection intononproiicing myelcma cal un.
\Prot.i
)Un. H ch.1
H Oski Gas.
Fig. 1. Transfectoma productionmethodusedto generatechimericantibodiesMouse heavy-chain (H-chain)V-regionVDJgenes(presumably froman existing mousehybridomacell line such as the one secreting NP-, dansyl-, orhuman lgE Fc-speclflc mouse antibody) are in-serted into a plasmld vector containing humanH-chain C-region genes. SlmIIa,ly, mouse fight-chain (L-chain)V-region genes are inserted into aseparate plasmid containing human L-chaln C-re-gion genes. Both engineered mouse-human Iggene vectors are then cotransfected into non-Ig-producing myeloma cells. Transfected myelomacellssynthesizing,assemblIng,andsecretingcom-pletechimeric (human-mouse) antibodiesthatareimmunoreactivewith NP or dansl onBSAor hu-manIgEare thenselectedandexpanded incultureand purifiedfrom culture fluidfor use as laboratoryreagents. Reproduced with peonisslon from ret. 16
hinge, CH2, and CH3), see Table 2. As examples, thewild-type IgG2 anti-dansyl antibody is denotedIgG(2,2,2,2); TNO, a domain-switched IgG anti-dansyl(2,2,2,3) antibody, is an IgG2 chimeric antibody with anIgG3 CH3 domain; and the TUH point-mutated IgG2Phe(296) -* Tyr is an IgG2 with a singleamino acidsubstitution from phenylalanine to tyrosine at position296. The C-region domains of the dansyl-specific IgGantibodies are schematically depicted in Figure 2(11).Details of the preparation of these chinieric antibodiesare presented elsewhere (12, 13). Chinieric antibodyChlO-10-3, which was prepared by L. Sun (Tanox Corp.,Houston, TX), is a human IgGi with V-region specificityfor the Fc region of human IgE (14).
Human immunoglobulins. Reactivity of the humanIg-specific mAbs to the chitneric antibodies was com-pared with their reactivity to chromatographically pu-rified polyclonal Igs and human paraproteins from pa-tients with multiple myeloma. The paraproteins withdefined light-chain specificity were kindly provided byH. Spiegelberg (IgGi, IgG2, IgG3, IgG4) while he was atScripps Clinic, La Jolla, CA; by K. Ishizaka (point-mu-tated myeloma IgE), while he was at Johns HopkinsUniversity, Baltimore, MD; and by J. Mestecky (poly-clonal secretory IgA), University of Alabama, Binning-
ham, AL. Polyclonal human IgG, IgA, and 1gM werefrom Jackson Immunoresearch, Malvern, PA.
Solid-phase antigens. The haptens NP and dansylwere separately coupled to bovine serum albumin (BSA)and human serum albumin and chromatographicallypurified for use as a solid-phase antigen (12-13). Foroptimal insolubilization on microtiter plates they wereused at 10 mgfL in phosphate-buffered saline (0.1 mLper well). The NP- or dansyl-BSA conjugates were cou-pled to CNBr-activated Sepharose CL-4B as describedpreviously (15) and subsequently used to aflunity-purilyeach chimeric antibody from culture supernate.
Molecular-Engineered ReferenceProteinsSpecificity reagents. In one of their earliest applica-
tions in the clinical laboratory, engineered antibodieswere used as human immunological quality-control orreference proteins with defined specificity (17). Chi-meric antibodies are particularly useful for validatingthe specificity and immunoreactivity of capture and de-tection polyclonal and mAb reagents used in clinicalassays of human antibodies with minor isotypes, e.g.,IgE, IgG3, IgG4. In these examples, the purified para-proteins are rare and expensive and often their sIlo-types are unknown. Moreover, because of conforma-

Table 2. Human Igand Chimeric Antibodies
CUNICAL CHEMISTRY,Vol.39, No. 9, 1993 1991
V-regionProteincod. sp.ciflclty C-regIon domaIn. Allotyp./deacvlptlon
G1-myeloma Unknown I9G(1-1-1-1) NRGla-NP-wIId NP IgG(1-1-1-1) NRGla-dansyl-wild Dansyl IgG(1-1-1-1) Glm(a, g, z)PM-TUS Dansyl IgGi Asn(276) -. Lys IgGi withmutated aa276DS-TLL Dansl IgG(1-1-1/4-4) IgGi with CH2 from aa292 and CH3 from lgG4DD-rrJ Dansyl IgG(X-1-1-1) IgGi with missingCH1G2-myeloma Unknown IgG(2-2-2-2) NRG2a-NP-wild NP IgG(2-2-2-2) NRG2a-dansyl-wlld Dansyl IgG(2-2-2-2) G2m(n)DS-TNO Dansyl IgG(2-2-2-3) IgG2 withIgG3CH3DS-TSR Dansyl IgG(2-2-3-2) IgG2 with IgG3 CH2PM-TUH Dansyl IgG2Phe(296)-, Tyr lgG2 withmutated aa296PM-TUV Dansyl lgG2 Asn(276) -* Lys lgG2 with mutated aa276G3-myeloma Unknown lgG(3-3abcd-3-3) NRG3a-NP-wIId NP IgG(3-3abcd-3-3) G3m(b)G3a-dansyl-wlId Dansyl IgG(3-3abcd-3-3) G2m(g,t), non-Gm3(bo),non-Gml(z)DS-TNL Dansyl IgG(3-3abcd-2-2) IgG3withIgG2CH2-CH3DS-TSV Dansyl IgG(3-3abcd-2-3) IgG3 with lgG2 CH2DS-TON Dansyl lgG(3-4-3-3) lgG3withIgG4 hingeDD-TJH Dansyl IgG(3-3abcX-3-3) IgG3 with hinge exons 1,2,3DD-THK Dansyl IgG(3-3aXXX-3-3) IgG3 onlywith hingeexon1
G4-myeloma Unknown IgG(4 4 4 4) NRG4a-NP-wIId NP lgG(4-4-4-4) NRG4a-dansyl-wild Dansyl IgG(4-4-4-4) G4m(a), non-Gim(z), G3m(g)DS-1TK Dansyl IgG(4-4-411-1) IgG4 with IgGi CH2 from aa292 and CH3PM-NC Dans,l IgG4 Ser(331) - Pro IgG4 with mutated aa331PM-TZB Dansyl IgG4 Ser(330) -. Pro lgG4 with mutated aa330DS-TJJ Dansyl I9G(4-3abcd-4-4) lgG4 with 4 IgG3hingeexonsE-myeloma(PS) Unknown lgE(E-E-E-E) NAEa-NP-wlld NP lgE(E-E-E-E) NAM-polyclonal Unknown 1gM NAMa-NP-wild NP IgM(M-M-M-M) NAA-polyclonal Unknown IgAl/A2 NAA2a-NP-wild NP IgA(A2-A2-A2-A2) NASec-lgA Unknown Secretory lgAl/2 NA
as, amino acid; domaIn-switched1g.DS-(3-lettercode); domaIn-deleted1g.DD-(3.Iettercode); poInt-mutated1g.PM-(3.Iettercode); NA, notavailable;NR, notreported; “abed’ refers to the four exons that code for the human IgG3 hingeregion.
tional alterations, insolubilizationof a paraprotein byadsorption onto a solid-phase surface may not mimic thebinding of the test human antibody to its insolubilizedantigen (18). The binding of chimeric antibody, e.g.,from the NP or dansyl series, onto a capture antibody orits appropriately insolubilized antigen appears to ad-dress these concerns.
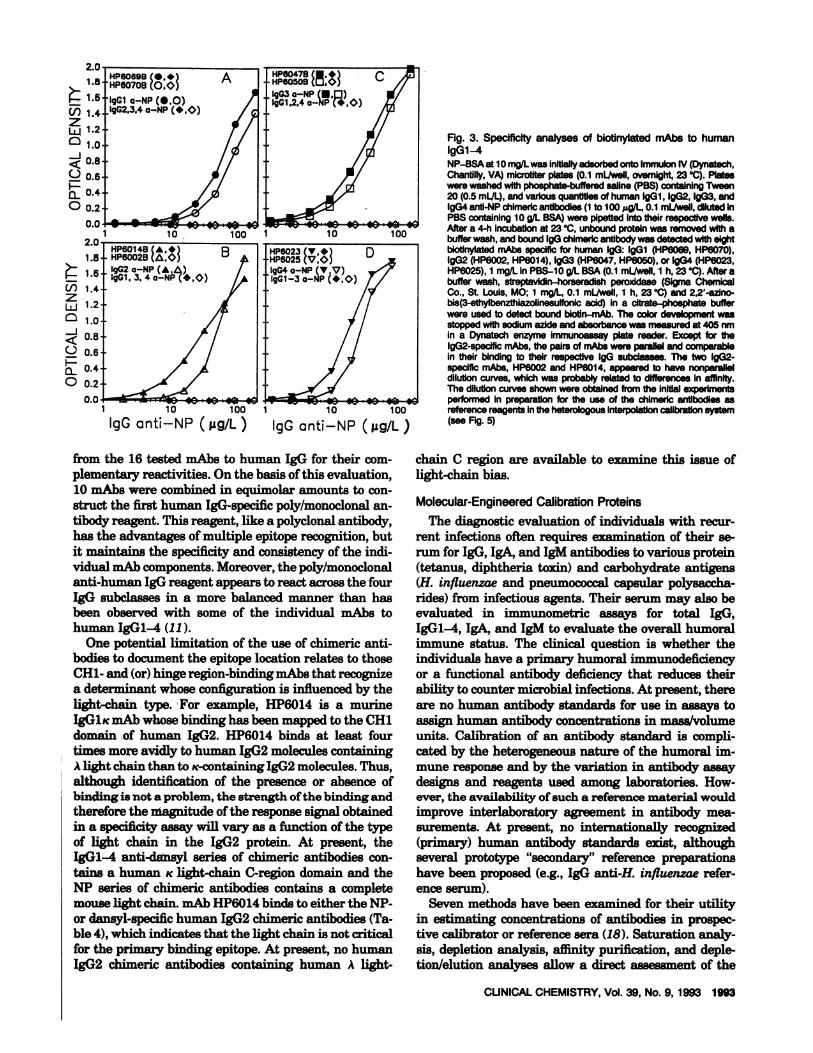
The subclass-restricted immunoreactivity of biotiny-lated mouse mAbs to human IgGi (HP6069, HP6070),IgG2 (HP6002, HP6014), IgG3 (HP6047, HP6050), andIgG4 (HP6023, HP6025) was demonstrated by usingIgGl-4 anti-NP chimeric antibodies bound to solid-phase NP-BSA antigen (Figure 3). The value of the“clean” or single IgG subclass nature of the chimericantibodies was demonstrated by comparing the resultsof similar analyses with human IgGl-4 paraproteinsand showing the presence of multiple human IgG sub-class proteins in IgG paraprotein preparations reportedto contain a single subclass. This observation demon-strates the difficulty of preparing single human IgG
subclass-restricted specificity reagents by chromato-graphically separating one human IgG subclass from itsthree structurally imi1ar counterparts. These chiniericproteins have also been used to confirm the specificity ofthe capture and detection antibody reagents in immu-noenzymometric assays of total serum IgE and IgGl-4(15, 19). Results of these specificity analyses are re-ported elsewhere (17).
Reagent-grade antigens. Molecular-engineered bio-molecules can be used as reagent-grade antigens to re-place proteins that need to be extracted from humanserum or tissue. Their application is predicated on theassumptions that the recombinant protein has foldedinto its native configuration after secretion from theexpression system (e.g., Escherichia coli, transfectedmyeloma cells) and that it has the relevant biologicalactivity. One nonantibody recombinant protein that hasbeen used as a solid-phase antigen is human insulin,which has been valuable in the study of human IgE andIgG antibodies that react with human insulin in vivo,

HumanCk
IgO(1.11/4/4)
-1---fl---ED-i-
-E---fl--cJ--
-E1---U1---Efl--
IgG(3.3abcd,2,3)
nhI33COC3)
-------
OS-Tn-
Do-Tru
PM-TUS
OS-mO
OS-TSR
PM-TUH
PM-TUV
DS.TNL
DS-TSV
DS-TON
DO-TiN Fig. 2. C-region exons of the lgG wild-type and mutant IgG
anti-dansyl chlmeric antibodiesOD-mK Eachdansyl-specificchimenicIg contains identicaldomainsfor munne
VK- and V-heavychain(VH)and humanCc The heavy-chainC-regionDS-TrK (CH) domainsdlfler among the chimenlc proteins. DS, domain-
switched Ig; OD, domain-deletedIg; PM, poInt-mutated1g.On the left,PM-TVC the human IgG subclass of each CH1, hInge,CH2, and CH3 exon is
descilbed.OS-TON: IgG(3,4,3,3) = IgG3-CH1, lgG4-hlnge, lgG3-CH2, and IgG3-CH3; DD-TIJ: lgG(X,1.1,1) missing-CH1,IgGi-
PM-TZB hinge, IgGl-CH2,and IgGl-CH3; DS-TTL: IgG(1,1,1/4,4) = IgGi.CH1, lgGl-hlnge, CH2 with the amino terminusfrom IgG1 and thecazboxyl terminus from lgG4, and IgG4-CH3. Reproducedwith per-
DS-fJJ missionfrom net. II--HI-----
1992 CUNICALCHEMISTRY,Vol.39, No.9, 1993
Mouse
aVH /
IgG(I.1.1.1) Ifl.1 J-f v.....Wild-Typci1IgG(2.22.2) Wild-Typel2
IJG(3.3abcd.3.3) - Wild-Type 13
IgG(4.44,4) Wdd.Typci4
IgG(X.I.I,1)
IgGI Asn(276)- Lys
IgG(2.2.2J)
IgG2.13.)
IgG2 Phe(296)-‘Tyr
LgG2 Asn(276)
- Lyo
IgG(3.3abcd,2.2)
IgG(3.4.3.3)
IG(33bX.3)
IgG(4.4.4/I.1)
IgG4 Scr(331)- Pro
lgG-l Ser(330)
IgG4.3.4.4)
leading in some cases to insulin resistance (20). Chi-meric antibodies can be used as reagent-grade purifiedhuman IgG1-4 proteins that can be labeled and used incompetitive binding immunoassays to quantify the hu-man IgG subclasses.
Epitope-mapping reagents. Morrison et al. (12) ini-tially constructed a panel of IgG anti-dansyl chimericantibodies for studying IgG antibody structure-functionrelationships. The wild-type IgG1-4 dansyl antibodieswere then modified at the DNA level by switching, de-leting, and point-mutating amino acids in the exonscoding for the CH1, hinge, CH2, and (or) CH3 domains.An exotic set of mutant IgG antibodies was thus createdthat can bind to solid-phase dansyl-B8A and has uniqueatypical C-region domains with ideal properties for
mapping epitopes that bind to murine mAbs to humanIgG reagents. Domain switching, for instance, was usedto switch an exon coding for the 12-amino acid hinge ofhuman IgG4 with an exon coding for the extended 62-amino acid hinge in human IgG3 molecules [DS-TJJIgG(4,3,4,4); see Figure 2 and Table 2]. In another in-stance, the Asn at position 276 in the IgG2 was con-verted to a Lye by point mutation. This change produceda loss of reactivity of the IgG Fc PAN mAbs (HP6017,HP6000, and HP6043) binding to this point-mutatedIgG2. Tables 3 and 4 summarize other reactivities thathave led reagent manufacturers to a better understand-ing of where these mAbs bind the IgG molecule. Themap of reactivities shown in Figure 4 provides an objec-tive method by which individual mAbs can be selected
HP6O69B C...HP6O7OB(0,0igGi a-NP (#{149}.O)IgG2,3,4 a-NP (#{149},0)
100 10 100
IgG anti-NP (j.ig/L) gG anti-NP (iig/L)
CUNICAL CHEMISTRY, Vol. 39, No. 9, 1993 1993
U,zLU0-J
0
0
0
Cl)zLU0-J
0
00
Fig. 3. Specificity analyses of biotinytatedmAbs to humanIgGl-4NP-BSA at 10 maJLwas inltiailyadsorbed onto Immulon IV (Dynatech,Chantilly,VA) microtiter plates (0.1 mI/well, overnight, 23#{176}C).Plateswerewashedwithphosphate-butteredsame (PBS)containingTween20(0.5 mL/L),andvarious quantities of human IgGi, IgG2,lgG3, andlgG4 anti-NPchimericantibodies(ito 100 g/L, 0.1 mI/well, diluted inPBS containing 10 g/L BSA)were pipetted into theirrespectivewells.Aftera 4-h incubationat 23 #{176}C,unboundproteinwas removed with abuffer wash, and bound lgG chimeric antibodywas detectedwith eightbiotinylated mAbs specific for human IgG: lgGl (HP6069, HP6070),IgG2 (HP6002,HP6014),IgG3 (HP6047, HP6050),or IgG4 (HP6023,HP6025),1 mg/L in PBS-b g/L BSA(0.1 mI/well, 1 h, 23#{176}C).After abuffer wash, streptavidin-horseradish pero,ddase (Sigma ChernicaiCo.,St. Louis,MO; 1 mg/L 0.1 mI/well, 1 h, 23#{176}C)and 2,2’-azlno-bis(3-ethylbenzthiazolinesuifonicacid) in a citrate-phosphate bufferwereused to detect boundbiotin-mAb. Thecolordevelopment wasstoppedwith sodium azide and absorbance wasmeasuredat405 nmin a Dynatech enzymeimmunoassayplate reader. Except for thelgG2-speciflcmAbs,the pairsof mAbswere parallelandcomparablein their binding to their respective IgG subclasses. The two lgG2-specific mAbs, HP6002 andHP6014, appeared to have nonparalleldilutioncurves, which was probably related todifferencesinaffinity.The dilution curves shown were obtained from the initial experimentsperformed in preparation for the use of the chimeric antibodies asreferencereagentsin the heterologousinterpolationcalibrationsystem(see Fig. 5)
from the 16 tested mAbs to human IgO for their com-plementary reactivities. On the basis ofthis evaluation,10 mAbs were combined in equimolar amounts to con-struct the first human IgG-specific poly/monoclonal an-tibody reagent. This reagent, like a polyclonal antibody,has the advantages of multiple epitope recognition, butit maintains the specificity and consistency of the indi-vidual rnAb components.Moreover, the poly/monoclonalanti-human IgG reagent appears to react across the fourIgG subclasses in a more balanced manner than hasbeen observed with some of the individual mAbs tohuman IgGl-4 (11).
One potential limitation of the use of chimeric anti-bodies to document the epitope location relates to thoseCH1- and (or) hinge region-binding mAbs that recognizea determinant whose configuration is influenced by thelight-chain type. For example, HP6014 is a murineIgGlic mAb whose binding has been mapped to the CH1domain of human IgG2. HP6014 binds at least fourtimes more avidlytohuman IgG2 molecules containingA light chain than to ic-containing IgG2 molecules. Thus,although identification of the presence or absence ofbinding is not a problem, the strength of the binding and
therefore the magnitude of the responsesignal obtainedin a specificity assay will vary as a function of the typeof light chain in the IgG2 protein. At present, theIgGl-4 anti-dansyl series of chimeric antibodies con-thins a human ic light-chain C-region domain and theNP series of chimeric antibodies contains a completemouse light chain. inAb HP6014 binds to either the NP-or dansyl-specific human IgG2 chimeric antibodies (Ta-ble 4), which indicates that the light chain is not criticalfor the primary binding epitope. At present, no humanIgG2 chimeric antibodies containing human A light-
chain C region are available to examine this issue oflight-chain bias.
Molecular-Engineered Calibration Proteins
The diagnostic evaluation of individuals with recur-rent infections often requires examination of their so-rum for IgG, IgA, and 1gM antibodies to various protein(tetanus, diphtheria to,dn) and carbohydrate antigens(H. influenzae and pneumococcal capsular polysaccha-rides) from infectious agents. Their serum may also beevaluated in immunometric assays for total IgG,IgG1-4, IgA, and 1gM to evaluate the overall humoralimmune status. The clinical question is whether theindividuals have a primary humoral immunodeficiencyor a functional antibody deficiency that reduces theirability to counter microbial infections. At present, thereare no human antibody standards for use in assays toassign human antibody concentrations in mass/volumeunits. Calibration of an antibody standard is compli-cated by the heterogeneous nature of the hunioral mm-mime response and by the variation in antibody assay
designs and reagents used among laboratories. How-ever, the availability of such a reference material wouldimprove interlaboratory agreement in antibody mea-surements. At present, no internationally recognized(primary) human antibody standards exist, althoughseveral prototype “secondary” reference preparationshave been proposed (e.g., IgG anti-H. in/luenzae refer-ence serum).
Seven methods have been examined for their utilityin estimating concentrations of antibodies in prospec-tive calibrator or reference sera (18). Saturation analy-sis, depletion analysis, affinity purification, and deple-tion/elution analyses allow a direct assessment of the

6 6 61 1 12 3 43 0 1
p - -
P - -
P P P
6 6 60 0 08 8 81 3 4
P P PP P P
P, positive reactivityof mAbfor thelistedhuman Igorchimericantibodythat has beenattachedto thesolidphase;-, no detectablemAb immunoreactlvltywithinsolubilized Ig or chime,lc protein,comparedwithbindingto BSA sham control.
1994 CUNICAL CHEMISTRY, Vol. 39, No. 9, 1993
Table 3. lmmunoreactlvtty Patterns of Human Pg-Specific mAbs for Chimeric Antibodies
Human Ig or chlmsrlcanUbody
IgG(1-1-1-1 )-?c-myelomalgG(1-1-1-1)-a-NPIgG(1-1-1-1 )-a-dansylTUS IgGi Asn(276) LysTLL lgG(1-1-1/4-4)TTJ IgG(X-1-1-1)lgG(2-2-2-2)-A-myelomaIgG(2-2-2-2)-a-NPlgG(2-2-2-2)-a-dansylTNO-IgG(2-2-2-3)TSR-lgG(2-2-3-2)TUH-IgG2 Phe(296) - TyrTUV-IgG2 Asn(276) - LyelgG(3-3-3-3)-c-myelomalgG(3-3abcd-3-3)-a-NPI9G(3-3abcd-3-3)-a-dansylTNL-IgG(3-3abcd-2-2)TSV-IgG(3-3abcd-2-3)TON-IgG(3-4-3-3)TJH-lgG(3-3abcX-3-3)THK-lgG(3-3aXXX-3-3)IgG(4-4-4-4)-A-myelomaIgG(4-4-4-4)-a-NPIgG(4-4-4-4)-a-dansylTTK-lgG(4-4-4/1-1)TVC-IgG4 Ser(331) - ProTZB-IgG4Ser(330) - ProTJJ-IgG(4-3abcd-4-4)IgE(E-E-E-E-E)-A-myelomalgE(E-E-E-E-E)-a-NPIgM-polyclonallgM(M-M-M-M)-a-NPIgAl /A2-polyclonaiIgA(A2-A2-A2-A2)-a-NPSecretoryIgAl /2
6 6 6 6 6 6o o 0 0 0 11 0 4 4 4 17 0 3 6 5 1
PP P P P -
PP P P P -
PP P P P -
PP P P P -
PP P P P -
PP P - - -
PP P P P -
PP P P P -
PP P P P -
PP P P P -
PP P P P -
PP P P P -
- - - P P -
PP P P P -
PP P P P -
PP P P P -
PP P P P -
PP P P P -
P - - - P -
P - - - P -
P - PP P -
PP P P P -
PP P P P -
PP P P P -
pp p p p -
PP P P P -
PP P P P -
P - P - P -
ppp
6 6 6 6 6 6 6 6o o 0 0 0 0 0 08 2 6 1 3 5 6 56 9 1 9 0 8 2 4
- - P - PP -
- - P - P - -
- - - P - PP -
- - - P - PP -
- - - P - P P -
- P - Pp -
- - - - P P - P- - - - p p - -
- - - - PP P -
- - - - PP P -
PP P P -
- pp p -
P -
PP P P -
p p p - -
P P P P -
- - - - pp p -
- - - - P P p -
- P P P P -
- - - PP P P -
- - - PP P P -
- - - pp - - p- - - PP - - -
- - - PP - P -
- - - pp p p -
- - - pp - p -
- - - PP - P -
- - - - P - P -
- p p - - - - P
p p
p P Pp
p p
P P
amount of antibody in the serum specimen by competi-tive inhibition or extraction with and without elution.For the comparative adsorbed-Ig method, the compara-tive antibody-Ig capture assay, and heterologous inter-polation with the use of a chimeric antibody standard, aheterologous reference curve is constructed for estimnat-ing the amount of reference antibody bound to its re-spectiveantigen in the actual assay. Estimates of themass per volume amounts of antibody in the serum are
then made from determinations of the amount of anti-body bound. The chimeric antibody-based method is po-tentially the most attractive because it mimics the ori-entation in which the test antibody binds to its antigen
In the chimeric antibody-based heterologous interpo-lation method, a heterologous (different antigen system)dose-response curve is constructed by binding knownquantities of a chimeric antibody having a single iso-
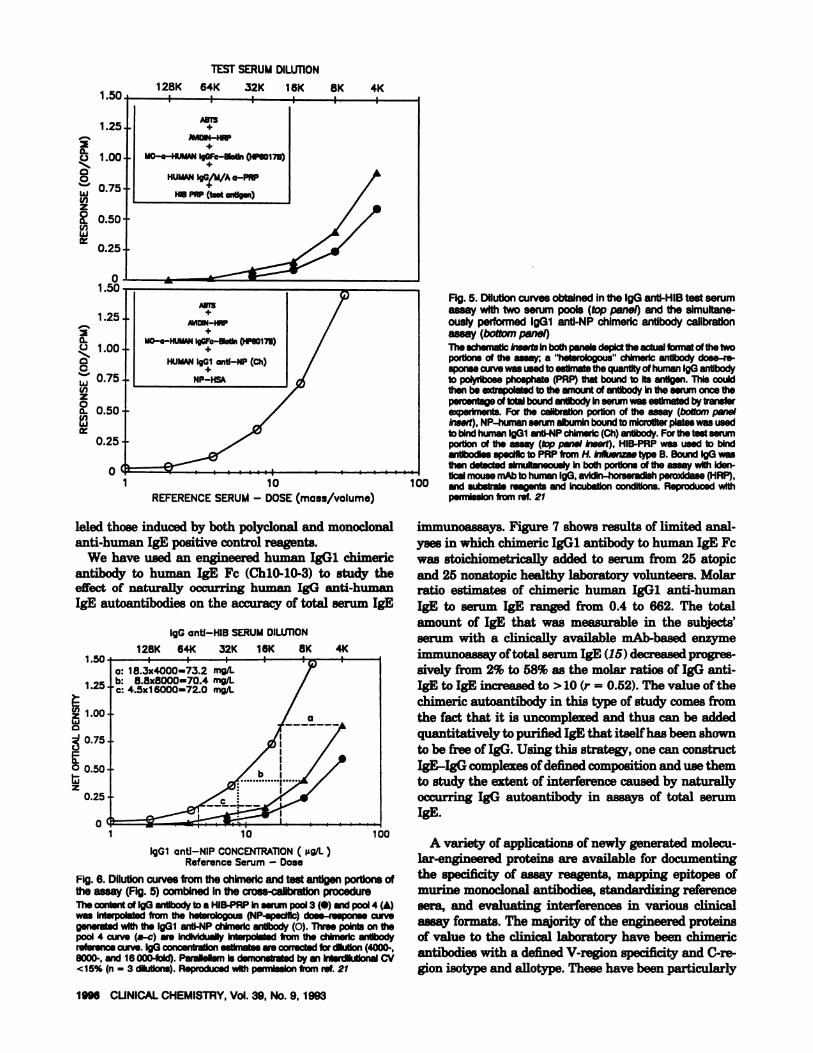
type and specificity to its insolubilized antigen. As illus-trated in Figure 5 (insert in bottom panel), chimerichuman IgGi anti-NP is bound to NP-human serumalbumin that has been attached to a well in a microtiterplate. In a simultaneous reaction performed with a testserum in a different portion of the plate (top panel in-sort), human IgG, IgA, and 1gM antibody to H. in/luen-zae type B (HIB) polyribose phosphate from a prospec-tive reference serum bind the insolubilized antigen(HIB-polyribose phosphate). Once these two simultane-ous reactions have been carried to completion (typically16-18 h at room temperature), unbound human serumproteins are removed with buffer washes, and theamount of IgG that is bound in both portions of theassay is detected by adding the same reagents to each:mouse anti-human IgG Fc-biotin (HP6O17B), avidin-horseradish peroxidase, and substrate [ABTS”; 2,2’-azinobis(3-ethylbenzthiazolinesulfonic acid)1. Figure 5

6014-G26045-P6046-P
6023-G46025-G4
6047-03 6069-G16050-G3
6091-GI6002-G26017-P6000-P6043-P6019-G 1346030-G2346058-G123
6666666660000000006790 1 4522901247035
P P P
P P P
P P P
P P P - - - - P P
P P P
- - - P P - - - -
- - - P P - - - -
- - - P P - - - -
- - - P P - - - -
- - - - P - - - -
- - - P P - - - -
- - - - P - - - -
P P - -
P P - -
P P - -
P- PP--
P- PP--
P P - -
P P
P P
P P
P P
P P
P P P P
Molecular-EngineeredInterferenceProteins- - Numerous reports indicate that human proteins, es-
pecially autoantibodies such as rheumatoid factor (hu-man 1gM anti-human IgG), artifactually increase ordecrease analyte estimates produced by clinical immu-noassays. The extent and nature of the interferencedepend on the assay design and the species of immuno-chemical reagents (22, 23). In the field of allergy, simi-lar problems have been suggested as a result of theobservation that naturally occurring human IgG anti-bodies specific for human IgE circulate in the blood ofmost individuals (24). We have recently extended theseearlier findings by showing that, in most individuals,human IgG autoantibothes are bound to human IgE oncirculating basophils (25). niAbs specific for human IgGPAN and IgGl-4 (e.g., Table 1) successfully cross-linkedIgG anti-IgE autoantibodies that were attached to IgE,which was itself bound to high-affinity IgE receptors onthe basophil surface. The incubation of peripheral bloodbasophils with these human IgG-specific mAbs inducedor triggered the release of vasoactive mediators such ashistamine, presumably by means of cross-linking IgEcell-surface receptors. These patterns of release paral-
VH CHI Hinge CH2 CH3
CLINICAL CHEMISTRY, Vol. 39, No. 9, 1993 1995
Table 4. ImmunoreactIvity Patterns of IgGSubclass-Specific mAbs for Chimeric Antibodies
Human Ig or chimeric
antibodyIgG(1-1-1-1)-K-myelomaIgG(1-1-1-1)-a-NPIgG(1-1-1-1 )-a-dansyl
TUS IgGi Asn(276) - LysTLL IgG(1-1-1/4-4)TTJ IgG(X-1-1-1)IgG(2-2-2-2)-A-myelomaIgG(2-2-2-2)-a-NPIgG(2-2-2-2)-a-dansylTNO-IgG(2-2-2-3)TSR-IgG(2-2-3-2)TUH-IgG2 Phe(296) -+ TyrTUV-IgG2 Asn(276)-* LysIgG(3-3-3-3)-K-myelomaIgG(3-3abcd-3-3)-a-NPIgG(3-3abcd-3-3)-a-dansylTNL-IgG(3-3abcd-2-2)TSV-lgG(3-3abcd-2-3)TON-IgG(3-4-3-3)TJH-IgG(3-3abcX-3-3)THK-IgG(3-3aXXX-3-3)IgG(4-4-4-4)-A-myeloma
IgG(4-4-4-4)-a-NPIgG(4-4-4-4)-a-dansylTTK-IgG(4-4-4/1 -1)TVC-IgG4 Ser(331) - ProTZB-IgG4 Ser(330) - ProTJJ-IgG(4-3abcd-4-4)IgE(E-E-E-E-E)-A-myelomaIgE(E-E-E-E-E)-a-NPIgM-polyclonalIgM(M-M-M-M)-a-NPIgAl/A2-polyclonalIgA(A2-A2-A2-A2)-a-NPSecretoryIgAl/2
P, positive reactivityofmAb for the listed humanIgor chimencantibody thathas been attachedto the solid phase; - no detectableimmunoreactivityof themAbwith insolubilized Ig or chimencproteinwhencomparedwith binding tothe BSAshamcontrol.
displays the response (absorbance) detected in each ofthe chimeric antibody (bottom curve) and test serum(top curve) segments of this assay. Finally, the dose-response curves from these two distinct but simultane-
ous reactions are plotted on the same graph (Figure 6).By interpolating multiple points of the IgG anti-HIBcurve from the heterologous chimeric antibody curve, anestimate is made of the amount of IgG anti-HIB bound.The curve for the chimeric antibody simply allows one torelate the response signal generated in an IgG anti-HIBassay to a particular amount of IgG bound. Translatingthe quantity of IgG bound into the quantity of IgGantibody in the serum requires a knowledge of the per-cent of total antibody in serum that has bound underassay conditions. This is determined by multiple Se-
llf lI I I II I ______
Fig. 4. C-region domains of the H chain of the human IgG molecule,showingthe bindinglocationof each of the 17 HP mAbs (Table 1)‘, the 10 mAbs selected for combination into the poly/monoclonal antibodyreagent as a result of the complementarityof their binding sites (reproducedwith permissionfrom ref. 11)
quential transfer experiments into new antigen-coatedplates until >90% of the total IgG antibody has beenbound (21). Figure 6 shows that the IgGi anti-NP andIgG anti-HIB dose-response curves are parallel; inter-dilutional CVs based on estimates of interpolated anti-body from three portions of the dose-response curve are
<15%. The accuracy of the chimeric interpolationmethod can be influenced by differences in the affinitiesof the calibration (e.g., anti-NP chimeric antibodies) andthe test antibodies (e.g., IgG anti-HIB). Both parallel-ism and antibody-transfer studies can be used to addressthis potential limitation. Therefore, the chimeric inter-polation method has the potential to provide an esti-mate of antibody content for any isotype of antibody andantigen system, when the chimeric antibodies are avail-able in a purified and calibrated form for the desiredisotype (18, 21).
I fl
TESTSERUMDILUTION
128K 64K 32K 16K 8K 4K
+
MDN-HRP
MO-a-HUMAN Ig -SIaSn (HPSO17S)
HUMAN lgG/M/A a-PRP+
HIDPftP (taut aMlg.n)
a-
C0
LU
z00U,LU
00
CCLUU,z0a-
1.25
1.00
0.75
0.50
0.25
01.50
1.25
1.00
0.75
0.50
0.25
+
A’.lOlN-HP+
MO-a-HUMAN I9GFc-aotln peeOl7D)
HUMAN lgGl anti-NP (Ch)
NP-lISA
1 10REFERENCE SERUM- DOSE(mass/volume)
leled those induced by both polyclonal and monoclonalanti-human IgE positive control reagents.
We have used an engineered humsn IgGi chimeric
antibody to human IgE Fc (ChlO-10-3) to study theeffect of naturally occurring human IgG anti-humanIgE autoantibodies on the accuracy of total serum IgE
IgG anti-HIS SERUMDILUTION
128K 64K 32K 16K 8K 4K1.50
1.25I 8.3x4000-73.2 mg/I..8.8*8000=70.4 mg/I-4.5x1 6000=72.0 mg/I-
1 10
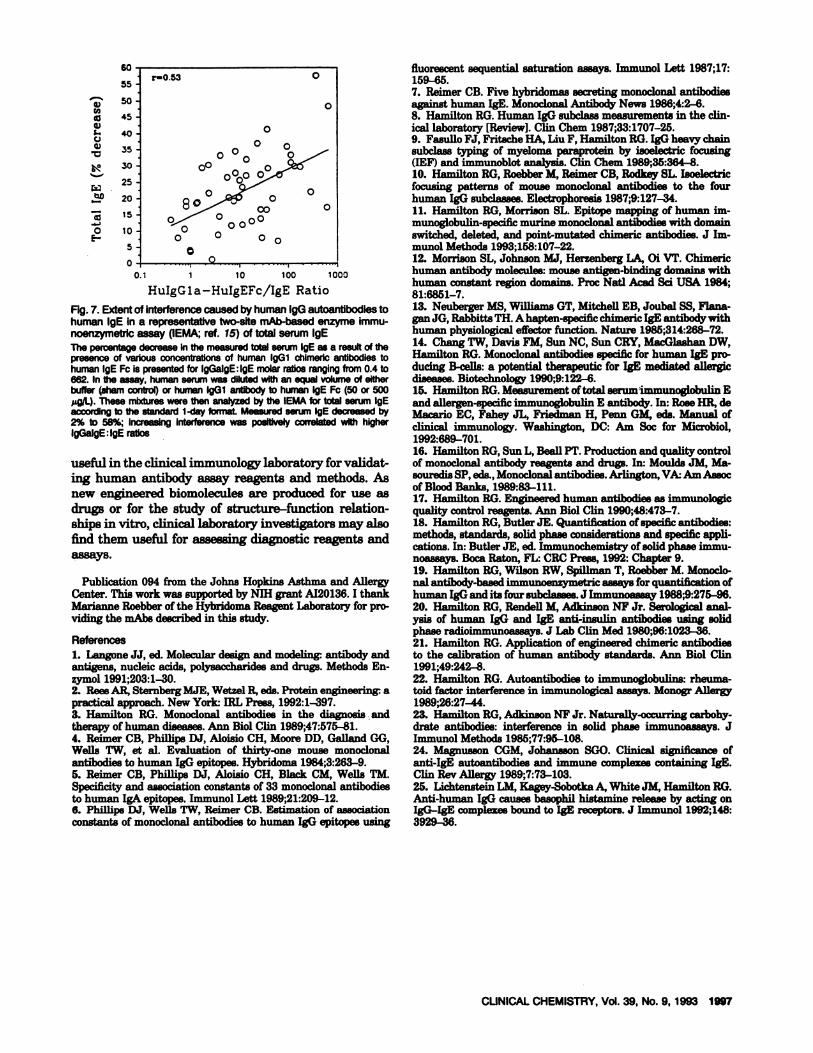
immunoassays. Figure 7 shows results of limited anal-yses in which chimeric IgGi antibody to human IgE Fcwas 8toichiometrically added to serum from 25 atopicand 25 nonatopic healthy laboratory volunteers. Molarratio estimates of chimeric human IgGi anti-human1gB to serum IgE ranged from 0.4 to 662. The total
amount of 1gB that was measurable in the subjects’serum with a clinically available mAb-based enzymeimmunoassay oftotal serum IgE (15) decreased progres-sively from 2% to 58% as the molar ratios of IgG anti-1gB to IgE increased to > 10 (r = 0.52). The value of thechimeric autoantibody in this type of study comes fromthe fact that it is uncomplexed and thus can be addedquantitatively to purified IgE that itselfhas been shown
to be free of IgG. Using this strategy, one can constructIgE-IgG complexesofdefined composition and use themto study the extent of interference caused by naturallyoccurring IgG autoantibody in assays of total serum1gB.
A variety of applications of newly generated molecu-lar-engineered proteins are available for documentingthe specificity of assay reagents, mapping epitopes ofmurine monoclonal antibodies, standardizing referencesara, and evaluating interferences in various clinicalassay formats. The majority of the engineered proteinsof value to the clinical laboratoty have been chimericantibodies with a defined V-region specificity and C-re-gion isotype and allotype. These have been particularly
1996 CUNICAL CHEMISTRY, Vol. 39, No.9, 1993
1.00w
0.75
0 0.50Iiiz
0.25
0
IgGi anti-NIP CONCENTRATION (g/L)Reference Serum - Dose
Fig. 6. Dilutioncurvesfromthe chimencand test antigen portions ofthe assay (Fig.5) combined in the cross-calibrationprocedureThe contentof IgG antibody to a HIB-PRPin serum pool 3(S) and pool 4(4)was Interpolated from the heterologous (NP-specific) dose-response curvegenerated with the lgGl anti-NP chlmeric antibody (0). Three points on thepool 4 curve (a-c) are Individually Interpolated from the chimeric antibodyreferencecurve. IgGconcentrationestimatesare correctedfor dilutIon(4000-,8000-, and 16000-fold). ParallelismIs demonstratedby an lnterdllutlon& CV<15% (n - 3 dIlutions).Reproducedwith permissionfrom ref. 21
Fig. 5. Dilutioncurvesobtained inthe IgGanti-HIB test serumassay with two serum pools (top panel) andthe simultane-ouslyperformed IgGi anti-NP chimeric antibody calibrationassay (bottom panel)The schematic Inserts In both panels depict the actual format of the twoportions of the assay; a ‘heterologous chlmedc antibody dose-re-sponsecurvewas usedtoestimatethequantityof human 190 antibodyto polyribose phosphate (PRP)that bound to Its antigen. This couldthen be extrapolatedto the amount of antibody In the serum once thepercentageof total boundantibody inserumwas estimatedby transferexperiments. For the calibratIon portion of the assay (boltom panelInsert), NP-human serumalbuminboundto mlcroliter plateswas usedto bIndhumanlgGl anti-NPchimerlc(Ch)antibody.For the test serumportion of the assay (tep panel Insert), HIB-PRP was used to bindantibodIesspecific to PAP from H. Influenzaetype B. Bound IgG wasthen detected simultaneously In both portIonsof the assay with Iden-
‘ tical mousemAb to humanlgG, avidln-horseradlsh peroxldase(HAP),100 and substrate reagents and Incubation conditions. Reproduced with
permissionfrom ret. 21
U)
5,I..C)5)
a0
5-
6055.
5045
40
35
30
25
2015
10
5
0
r-O.53 0
0
00 0
0 0000
800 0 0O o
o0 0
0C)
CUNICAL CHEMISTRY, Vol. 39, No. 9, 1993 1997
0.1 1 10 100 1000
HuIgGla-HuIgEFc/IgE Ratio
Fig.7. Extentof interferencecaused byhumanlgGautoantibodiestohuman IgE in a representativetwo-site mAb-basedenzyme immu-noenzymetricassay (IEMA;ref. 15) of total serum IgEThepercentagedecrease in the measuredtotalserum IgE as a result of thepresenceof variousconcentrationsof human lgGl chlmericantibodies tohumanIgEFcispresentedfor IgGalgEIgEmolarratiosrangingfrom0.4to662. In the assay, human serum was dilutedwith an equal volume of eitherbuffer (sham control) or human IgGi antibody to human IgE Fc (50 or 500pg/I). These mixtures were thenanalyzedby the IEMA for total serum IgEaccording to the standard 1-day format Measuredserum IgE decreased by2% to 58%; Increasing interference was positively correlated with higherIgGalgE:IgE ratios
useful in the clinical immunology laboratory for validat-ing human antibody assay reagents and methods. Asnew engineered biomolecules are produced for use asdrugs or for the study of structure-function relation-ships in vitro, clinical laboratory investigators may alsofind them useful for assessingdiagnostic reagents andassays.
Publication 094 from the Johns Hopkins Asthma and AllergyCenter. This work was supported by NIH grant A120136. I thankMarianne Roebber of the Hybridoma Reagent Laboratory for pro-viding the mAbs described in this study.
References1. Langone JJ, ed. Molecular design and modeIing antibody andantigens, nucleic acids,polysaccharides and drugs. Methods En-zymol 1991;203:1-30.2. Rees AR, Sternberg MJE, Wetzel R, eds. Protein engineering apractical approach.NewYork IRL Press, 1992:1-397.3. Hamilton RG. Monoclonal antibodies in the diagnosis andtherapy of human diseases. Ann Biol Clin 1989;47:575-81.4. Rainier CB, Phillips DJ, Aloisio CH, Moore DD, Galland (Xl,Wells TW, et al. Evaluation of thirty-one mouse monoclonalantibodies to human IgG epitopes. Hybridoma 1984;3:263-9.5. Reimer CB, Phillips DJ, Aloisio CH, Black CM, Wells TM.Specificity and association constants of 33 monoclonal antibodiesto human IgA epitopes. Immunol Lett 1989;21:209-12.6. Phillips DJ, Wells TW, Reimer CB. Estimation of associationconstants of monoclonal antibodies to human IgG epitopes using
fluorescent sequential saturation assays.Immunol Lett 1987;17:159-65.7. Reimer CB. Five hybridomas secreting monoclonalantibodiesagainst human IgE. MonoclonalAntibOdy News 1986;4:2-6.8. Hamilton RG.HumanIgG subclassmeasurementsin the olin-ical laboratory [Review]. Clin Chem 1987;33:1707-25.9. FasulloFJ, Fritache HA, Liu F, Hamilton RG. IgO heavy chainsubclass typing of myeloma paraprotein by isoelectric focusing(IEF) and immunoblotanalysis. Clin Chem 1989;35:364-8.10. Hamilton RG, Roebber M, Reimer CE, Rodkey SL. Isoelectricfocusing patterns of mouse monoclonal antibodies to the fourhuman IgG subclasses.Electrophoresis1987;9:127-34.11. Hamilton RG,Morrison SL. Epitope mapping of human fin-munoglobulin-specilicmurine monoclonal antibodies with domainswitched, deleted, and point-mutated chimeric antibodies. J Im-munol Methods 1993;158:107-22.12. Morrison SL, Johnson MJ, Herzenberg LA, Oi VT. Chimerichuman antibody molecules:mouse antigen-binding domains withhuman constant region domains. Proc Nati Aced Sd USA 1984;81:6851-7.13. NeubergerMS. Willifims GT, Mitchell EB, Joubal 58, Flana-gan JG, Rabbitta PH. A hapten-specific chimeric IgE antibody withhuman physiological effector function. Nature 1985;314:268-72.14. Chang TW, DavisFM, Sun NC, Sun CRY,MaCGlaShan DW,Hamilton RG. Monoclonal antibodies specific for human IgE pro-ducing B-cells: a potential therapeutic for IgE mediated allergicdiseases.Biotechnology 1990;9:122-6.15. Hamilton RG.Measurementoftotal serum immunoglobulin Eand allergen-specific immunoglobulin E antibody. In: RoseHR, deMacario EC, Fahey JL, Friedman H, Penn GM, ads. Manual ofclinical immunology. Washington, DC: Am Soc for Microbiol,1992:689-701.16. Hamilton RG,Sun L, BeaU PT. Production and quality controlof monoclonal antibody reagents and drugs. In: Moulds JM, Ma-souredis SP,ads.,Monoclonal antibodies. Arlington, VA Am Assocof Blood Banks, 1989:83-111.17. Hamilton RG.Engineered human antibodies as immunologicquality control reagents. Ann BiolCliii 1990;48:473-7.18. Hamilton RG,Butler JE. Quantification of specific antibodies:methods, standards, solid phase considerations and specific appli-cations.In: Butler JE, ad. Immunochemistry of solid phase immu-noassays. Boca Raton, FL: CRC Press, 1992: Chapter 9.19. Hamilton RG, Wilson RW, Spilimfin T, Roebber M. Monoclo-nal antibody-based immunoenzymetric assays for quantification ofhuman IgG and its foursubclasses.J Immunoassay1988;9:275-96.20. Hamilton RG, Rendell M, Mkinson NF Jr. Serological anal-ysis of human IgG and 1gB anti-insulin antibodies using solidphase radioimmunoassays. J Lab Clin Med 1980;96:1023-36.21. Hamilton RG. Application of engineered chimeric antibodiesto the calibration of human antibody standards. Ann Biol Cliii1991;49:242-8.22. Hamilton RG. Autoantibodies to immunoglobulins: rheuma-toid factor interference in immunological assays. Monogr Allergy1989;26:27-44.23. Hamilton RG,Mkinson NF Jr. Naturally-occurring carbohy-drate antibodies: interference in solid phase immunoassays. JImmunol Methods 1985;77:95-108.24. Magnuason CGM, Johansson SGO. Clinical significance ofanti-IgE autoantibodies and immune complexescontaining 1gB.Clin Rev Allergy 1989;7:73-103.25. Lichtenstein LM, Kagey-Sobotka A, White JM, Hamilton RG.Anti-human IgG causes basophil histamine release by acting onIgG-IgE complexes bound to IgE receptors. J Immunol 1992;148:3929-36.