modulation of tumor necrosis factor-mediated cell death by fullerenes
TRANSCRIPT

Research Paper
Modulation of Tumor Necrosis Factor-mediated Cell Death by Fullerenes
Ljubica Harhaji,1 Aleksandra Isakovic,2 Ljubica Vucicevic,1 Kristina Janjetovic,1 Maja Misirkic,1
Zoran Markovic,3 Biljana Todorovic-Markovic,3 Nadezda Nikolic,3 Sanja Vranjes-Djuric,3 Zoran Nikolic,4
and Vladimir Trajkovic5,6
Received August 16, 2007; accepted October 19, 2007
Purpose. The fullerene (C60/C70 mixture—C60/70) nanocrystalline suspension prepared by solvent exchange
method using tetrahydrofyran (THF/nC60/70) and polyhydroxylated C60/70 [C60/70(OH)n] were compared
for their ability to modulate cytotoxicity of the proinflammatory cytokine tumor necrosis factor (TNF).
Materials and Methods. TNF-induced cytotoxicity was assessed in L929 fibrosarcoma cells by crystal
violet assay. The type of cell death (apoptosis/necrosis), production of reactive oxygen species, mitochondrial
depolarization and caspase activation were determined by flow cytometry using the appropriate reporter dyes.
Results. THF/nC60/70 augmented, while C60/70(OH)n reduced the cytotoxicity of TNF. The numbers of
cells undergoing apoptosis/necrosis, as well as of those displaying the activation of apoptosis-inducing
enzymes of caspase family, were respectively increased or reduced by THF/nC60/70 or C60/70(OH)n. The
antioxidant N-acetylcysteine and mitochondrial permeability transition inhibitor cyclosporin A each
partly blocked the cytotoxic action of TNF, indicating the involvement of oxidative stress and
mitochondrial dysfunction in the TNF cytotoxicity. Accordingly, THF/nC60/70 or C60/70(OH)n potenti-
ated or suppressed, respectively, TNF-triggered oxidative stress and mitochondrial depolarization.
Conclusion. The ability of different fullerene preparations to modulate TNF-induced oxidative stress
and subsequent cell death suggests their potential value in the TNF-based cancer therapy or prevention
of TNF-dependent tissue damage.
KEY WORDS: apoptosis; fullerene; necrosis; oxidative stress; tumor necrosis factor.
INTRODUCTION
Because of the unique chemical and physical propertiesthat enable interaction with living cells, fullerenes (C60) andtheir derivatives are promising candidates for many biomed-ical applications (1). The electrochemical features that providereactivity with cell-damaging reactive oxygen species (ROS)such as superoxide (O2
&-) and hydroxyl (&OH) radicals, makewater-soluble fullerene derivatives behave as Bfree radicalsponges.^ Accordingly, water-soluble fullerenes display a pro-tective effect in various experimental models of ROS-dependentcell death (2–6), and the polyhydroxylated fullerenes, known asfullerols or fullerenols, are particularly efficient in that respect(7,8). Somewhat surprisingly, it has recently been shown thatpure C60 brought into water by means of solvent extraction
forms water-stable crystalline aggregates (nano-C60 or nC60),which, in contrast to fullerol, generate reactive oxygen and killvarious cell types in a ROS-dependent manner (9–12). Whilethe mechanisms underlying this different behavior of watersoluble vs. nanocrystaline C60 are still to be revealed, it has beenproposed that derivatization of the fullerene core in order toprovide water solubility, might somehow decrease its prooxidantand increase the antioxidant capacity (9,10). On the other hand,as nC60 prepared by detergent solubilization or long-termmixing in water does not display ROS-dependent cytotoxicity(13,14), it has been suggested that the observed ROS generationby solvent exchange-prepared nC60 could actually result fromthe interaction of C60 with the relatively small amount (<10%)of otherwise nontoxic residual organic solvent (tetrahydrofuran)that remains trapped in nC60 crystalline lattice (12,15,16).
Tumor necrosis factor (TNF, previously known as TNF-a)is a pleiotropic proinflammatory cytokine produced by acti-vated macrophages, as well as by several other cell types,including lymphocytes, fibroblasts, and hepatocytes (17). Manycell types are targets for TNF action due to an extremelybroad expression of TNF receptors TNF-R1 and TNF-R2(18). One of the most prominent and most explored biologicalactions of TNF is its fairly selective cytotoxic activity ontumor cells (19), making it a plausible candidate for antican-cer treatment, particularly against surgically unresectable andchemotherapy- or radiation therapy-resistant tumors (20).However, the clinical use of TNF has been hampered by the
0724-8741/07/0000-0001/0 # 2007 Springer Science + Business Media, LLC
Pharmaceutical Research (# 2007)DOI: 10.1007/s11095-007-9486-y
1 Institute for Biological Research, Belgrade, Serbia.2 Institute of Biochemistry, School of Medicine, University of Belgrade,
Belgrade, Serbia.3 Vinca Institute of Nuclear Sciences, Belgrade, Serbia.4 Faculty of Physics, University of Belgrade, Belgrade, Serbia.5 Institute of Microbiology and Immunology, School of Medicine,
University of Belgrade, Dr. Subotica 1, 11000, Belgrade, Serbia.6 To whom correspondence should be addressed. (e-mail: vtrajkovic@
eunet.yu)
ABBREVIATIONS: DHR, dihydrorhodamine; FITC, fluorescein
isothiocyanate; MPT, mitochondrial permeability transition; PI,
propidium iodide; ROS, reactive oxygen species.

resistance of many tumors to TNF-mediated death (21), aswell as because of the severe side-effects of systemic TNFadministration, such as hypotension and hepatotoxicity (22,23).Due to systemic toxicity, TNF therapy is limited to sophisti-cated locoregional drug-delivery systems in patients with sometypes of organ-confined solid tumors (24). TNF is also involvedin liver damage during sepsis, viral hepatitis, alcoholic hepatitis,ischemia-reperfusion liver injury, fulminant hepatic failure andchemical-induced hepatotoxicity (25,26), which is consistentwith its ability to cause TNF-R1-dependent hepatocyte damagein vitro and in vivo (27). Therefore, exploring the mechanismsfor the enhancement of TNF-mediated tumor cell killing anddecreasing its toxicity against normal tissues is of paramountimportance for successful development of the TNF-relatedanticancer or anti-inflammatory therapeutic strategies.
As the cytotoxic action of TNF depends on its ability totrigger ROS production in target cells (28–30), ROS-modulatingagents such as fullerenes are potential candidates for theinterference with TNF-mediated cell death. Accordingly, ithas been demonstrated that malonic acid C60 derivatives(carboxyfullerenes) can protect human peripheral blood mono-nuclear cells from oxidative stress-dependent apoptosis inducedby combination of TNF and cycloheximide (31). However, theeffects of other fullerene derivatives on TNF-induced cell deathhave not been investigated to date. In the present study, weshow that nanocrystalline and polyhydroxylated fullerenesdifferently modulate the TNF toxicity towards L929 mousefibrosarcoma cell line, which has been widely used as a modelsystem for exploring TNF-induced cell death.
MATERIALS AND METHODS
Preparation of Fullerenes
For the preparation of fullerene colloid in water we usedC60/C70 extract of carbon soot (79% C60, 20% C70, 1% higher-order fullerenes) produced by arc discharge (32). C60/C70
colloid was produced by evaporating tetrahydrofuran (THF)from a mixture of water and molecularly dispersed C60/C70 inTHF (Sigma, St. Louis, MO), as previously described (33).The concentration of C60/C70 suspension in water, referred toas THF/nC60/70, was determined from the absorption spec-trum and adjusted to 10 mg/ml. Briefly, 1ml of fullerene watersuspension was mixed with equal amount of 2% NaCl solu-tion, and toluene (2 ml) was added. The mixture was sonicatedfor a few minutes and then set in the dark until the aqueousand toluene phases separated. The upper toluene phase, con-taining fullerene in molecular form, was withdrawn carefully,and the absorbance at 335 nm was measured and comparedwith referent spectrums. Polyhydroxylated fullerene, referredto as C60/70(OH)n (n = 2–24), was prepared as previouslydescribed (34) from the same C60/C70 extract of carbon sootused for the THF/nC60/70 preparation. Both THF/nC60/70 andC60/70(OH)n were characterized by dynamic light scattering,UV-Vis and Fourier transform infrared spectroscopy as pre-viously described (11). A schematic representation of thestructure of the two fullerene derivatives is given in Fig. 1.Both fullerene preparations were stored at 4-C until used forexperiments. Since we did not assess the ratio of C60 and C70
in THF/nC60/70 and C60/70(OH)n, only one batch of eachfullerene preparation was used in the experiments presented,
in order to avoid the possible influence of changes in C60/C70
ratio in different batches. Nevertheless, it should be notedthat similar results were obtained with different batches ofboth THF/nC60/70 and C60/70(OH)n.
Cells and Cell Cultures
The mouse fibrosarcoma cell line L929 was obtained fromthe European Collection of Animal Cell Cultures (Salisbury,UK), while the human glioma cell line U251 was kindlydonated by Dr. Pedro Tranque (Universidad de Castilla-LaMancha, Albacete, Spain). The primary macrophages wereobtained from peritoneal cavities of C57/BL6 mice (Institutefor Biological Research, Belgrade) as previously described(35), and in accordance with the Guide for the Care and Useof Laboratory Animals as adopted and promulgated by theUnited States National Institutes of Health. The tumor cellswere seeded in flat-bottom 96-well (1 � 104 cells/well) or 6-well (2 � 105 cells/well) cell culture plates (Sarstedt, Newton,NC) for the cell viability assessment or flow cytometry analysis,respectively. Peritoneal macrophages were seeded in 96-wellplates at 1 � 105 cells/well. After being rested for 24h, cellcultures were washed and incubated with TNF alone or incombination with THF/nC60/70 or C60/70(OH)n. U251 cellswere treated with cisplatin or exposed to hyperthermia (43-Cfor 1h), in the presence or absence of fullerene preparations.The cells were incubated at 37-C in a humidified atmospherewith 5% CO2, in a HEPES (20 mM)-buffered RPMI 1640 cellculture medium (Sigma, St. Louis, MO) supplemented with5% fetal calf serum, 2 mM L-glutamine, 50 mM 2-mercaptoethanol, 10 mM sodium pyruvate, and 100 IU/mlpenicillin and streptomycin (all from Sigma). Working
Fig. 1. Schematic representation of the structure of C60(OH)n (n = 2–
24) and THF/nC60/70. For clarity, only six C60 molecules are presented
per one side of THF/nC60/70 crystal, while C70 molecules are not
depicted. The size of THF/nC60/70 nanoparticles used in the present
study was approx. 100 nm.
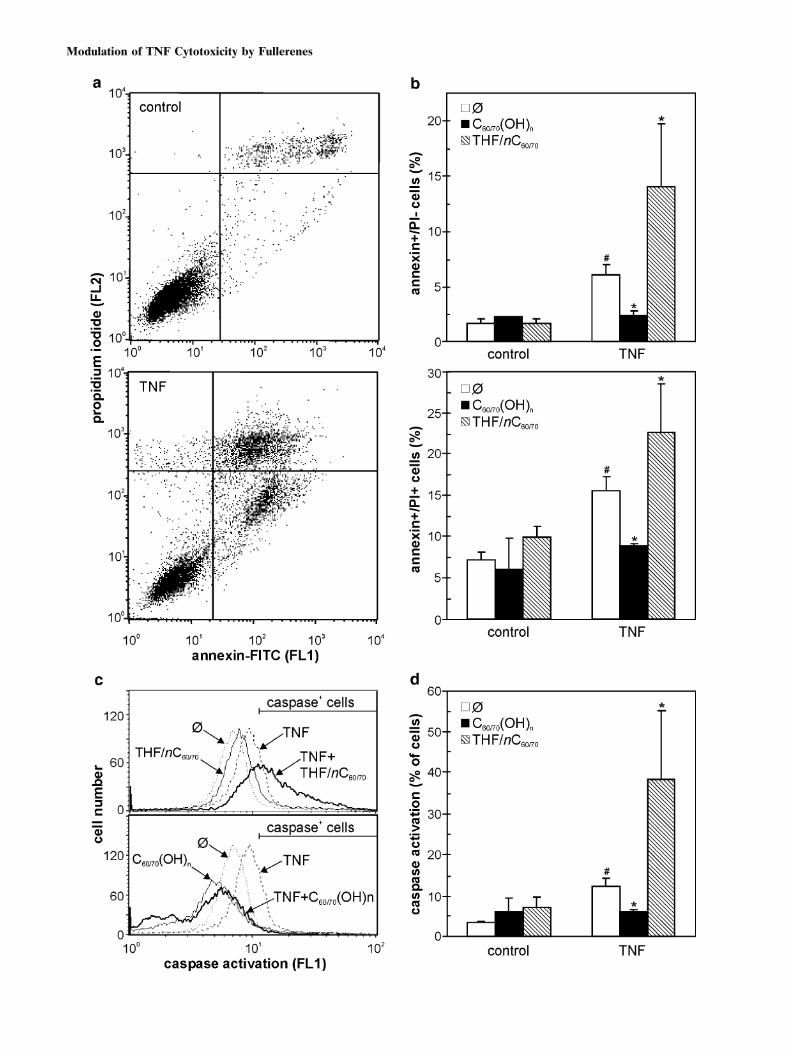
Fig. 2. Fullerene preparations differently modulate TNF-mediated
cytotoxicity. a, b L929 cells were incubated with different concen-
trations of TNF, THF/nC60/70 or C60/70(OH)n alone or in different
combinations, and the cell number was assessed after 24 h. The results
are mean values of triplicates from a representative of three experi-
ments. c The Chou–Talalay combination index was calculated and the
data are given as mean T SD values from three independent experi-
ments (*p < 0.05 denotes values significantly different from 1). d The
photomicrographs of the cells treated for 24 h with TNF (0.5 ng/ml), in
the presence or absence of THF/nC60/70 (125 ng/ml) or C60/70(OH)n
(250 mg/ml).
b
Harhaji et al.

Modulation of TNF Cytotoxicity by Fullerenes

solutions of THF/nC60/70 were prepared by addition of theappropriate amounts of 10-fold concentrated cell culturemedium and deionized water, while C60/70(OH)n wasdissolved directly in the cell culture medium.
Cell Viability Determination
For the assessment of cell viability we used a crystalviolet assay, which is based on the inability of dead cells toremain adherent to cell culture plastic (36). After incubation,cells were washed with PBS to remove dead, non-adherentcells. The remaining adherent, viable cells were fixed withmethanol and stained with 1% crystal violet solution at roomtemperature for 10min. The plates were thoroughly washedwith water, and crystal violet was dissolved in 33% aceticacid. The absorbance of dissolved dye, corresponding to thenumber of viable cells was measured in an automated micro-plate reader at 570nm. The results are presented as relative tothe control value (untreated cells).
Analysis of Apoptotic and Necrotic Cell Death
The type of cell death (apoptotic or necrotic) was analyzedby double staining with annexin V-FITC and propidium iodide(PI), in which annexin V bound to the early apoptotic cells withexposed phosphatidylserine, while PI labeled the late apoptoticand necrotic cells with the membrane damage. Staining wasperformed according to the instructions by the manufacturer(BD Pharmingen, San Diego, CA), and flow cytometricanalysis was conducted on a FACSCalibur flow cytometer(BD). The percentage of apoptotic (annexin+/PIj) and ne-crotic (annexin+/PI+) cells was determined using CellQuestPro software.
Detection of Caspase Activation
Activation of caspases was measured by flow cytometryafter labeling the cells with a cell-permeable, FITC-conjugatedpan-caspase inhibitor (ApoStat; R&D Systems, Minneapolis,MN) according to the manufacturer_s instructions. The in-crease in green fluorescence (FL1) is a measure of caspaseactivity within individual cells of the treated population. Theresults are expressed as % of cells containing active caspases.
Measurement of Intracellular ROS Production
Intracellular production of ROS was determined by mea-suring the intensity of green fluorescence emitted by redox-sensitive dye dihydrorhodamine 123 (DHR; Invitrogen, Paisley,UK), which was added to cell cultures (2 mM) at the beginningof treatment. At the end of incubation, cells were detached bytrypsinization, washed in PBS, and the green fluorescence (FL1)of DHR-stained cells was analyzed using a FACSCalibur flowcytometer.
Assessment of Mitochondrial Membrane Potential
The mitochondrial depolarization was assessed usingDePsipher (R&D Systems), a lipophilic cation susceptible tothe changes in mitochondrial membrane potential. It has the
property of aggregating upon membrane polarization form-ing an orange-red fluorescent compound. If the potential isdisturbed, the dye can not access the transmembrane spaceand remains or reverts to its green monomeric form. Thecells were stained with DePsipher as described by the man-ufacturer, and the green monomer and the red aggregateswere detected by flow cytometry. The results were presentedas a green/red fluorescence ratio (geomean FL1/FL2), theincrease of which reflects mitochondrial depolarization.
Mathematical Analysis of Synergism/Antagonism
To analyze the type (additive, synergistic or antagonistic)of fullerene interaction with TNF in inducing tumor cell death,cells were treated with each agent alone and their appropriatecombinations. The cell viability was assessed using a crystalviolet assay. The values of combination index, reflectingadditive (= 1), synergistic (<1) or antagonistic interactions(>1), were calculated according to the method based on themedian-effect principle of Chou and Talalay (37).
Statistical Analysis
The statistical significance of the differences was analyzedby t-test or ANOVA followed by the Student–Newman–Keuls test. The value of p < 0.05 was considered significant.
RESULTS
THF/nC60/70 and C60/70(OH)n Differently AffectTNF-mediated Cytotoxicity
To assess the influence of THF/nC60/70 and C60/70(OH)n
on TNF cytotoxicity, we treated L929 cells with differentdoses of fullerene preparations and TNF, separately or incombination. In accordance with our previous results (11),crystal violet assay showed that both fullerene preparationswere cytotoxic, but THF/nC60/70 was at least three orders ofmagnitude more potent than its hydroxylated counterpart(Fig. 2a,b). Interestingly, while combination of THF/nC60/70
and TNF was more toxic than each agent alone (Fig. 2a),addition of C60/70(OH)n clearly reduced the cytotoxic effectof TNF (Fig. 2b). Accordingly, the combination indexcalculated by Chou–Talalay approach was >1 throughout0.1–0.99 efficiency range for combination of C60/70(OH)n withTNF, and <1 for combination of THF/nC60/70 and TNF(efficiency range 0.4–0.99), thus confirming antagonistic andsynergistic interactions, respectively (Fig. 2c). For furtherinvestigation we used TNF at the concentration of 0.5 ng/ml,which displayed suboptimal cytotoxicity and thereforeallowed direct comparison of the opposite effects of nontoxicTHF/nC60/70 and C60/70(OH)n doses (125 and 250 mg/ml,
Fig. 3. The effects of fullerene preparations on TNF-induced
apoptosis, necrosis and caspase activation. a, b L929 cells were
incubated with TNF (0.5 ng/ml), in the absence or presence of THF/
nC60/70 (125 ng/ml) or C60/70(OH)n (250 mg/ml). Apoptosis, necrosis
(a, b) and caspase activation (c, d) were analyzed by flow cytometry
after 18 h. The representative dot plots and histograms are presented
in (a, c), while the data in (b, d) are mean T SD values from three
independent experiments (#p < 0.05 refers to untreated cells; *p < 0.05
refers to cells treated with TNF alone).
b
Harhaji et al.
Modulation of TNF Cytotoxicity by Fullerenes
Harhaji et al.

respectively) on TNF-mediated cell death. In accordancewith the cytotoxicity data, the TNF-treated cells lost theirpolygonal morphology and became smaller, with granularappearance and poorly defined margins (Fig. 2d). Whileneither THF/nC60/70 nor C60/70(OH)n alone were able tocause significant changes in L929 cell morphology, bothagents significantly affected morphological changes inducedby TNF treatment (Fig. 2d). Namely, the cells treated withTNF and THF/nC60/70 became completely round and de-tached from the surface of cell culture wells, which isconsistent with the synergistic induction of cell death. Onthe other hand, the cells treated with TNF and C60/70(OH)n
mainly retained normal polygonal morphology characteristicfor healthy, untreated cells, thus confirming the protectiveeffect of polyhydroxylated fullerenes.
The Effects of Fullerene Preparations on TNF-inducedApoptosis/Necrosis and Caspase Activation
The treatment with suboptimal dose of TNF led to asignificant increase in numbers of cells displaying early apopto-tic (annexin+/PIj) and late apoptotic/necrotic (annexin+/PI+)phenotype, as revealed by flow cytometric analysis (Fig. 3a,b).The TNF-mediated apoptosis/necrosis was associated by sig-nificant activation of apoptosis-executing enzymes caspases intarget cells (Fig. 3c,d). In accordance with the cytotoxicity dataobtained in a crystal violet assay (Fig. 2a,b), the addition ofTHF/nC60/70 markedly increased the numbers of both earlyapoptotic and late apoptotic/necrotic cells in TNF-treated L929cultures, while C60/70(OH)n clearly reduced TNF-triggeredapoptosis/necrosis (Fig. 3b). In addition, although unable tosignificantly activate caspases if applied alone, THF/nC60/70
caused synergistic potentiation of TNF-induced caspase activa-tion in L929 cells (Fig. 3c,d). On the other hand, TNF-mediatedcaspase activation was completely blocked by simultaneousaddition of C60/70(OH)n (Fig. 3c,d). Therefore, the oppositeeffects of THF/nC60/70 and C60/70(OH)n on TNF cytotoxicitywere associated with the enhancement and suppression, re-spectively, of TNF-induced caspase activation and apoptotic/necrotic cell death.
The Effects of Fullerene Preparations on TNF-induced ROSProduction and Mitochondrial Depolarization
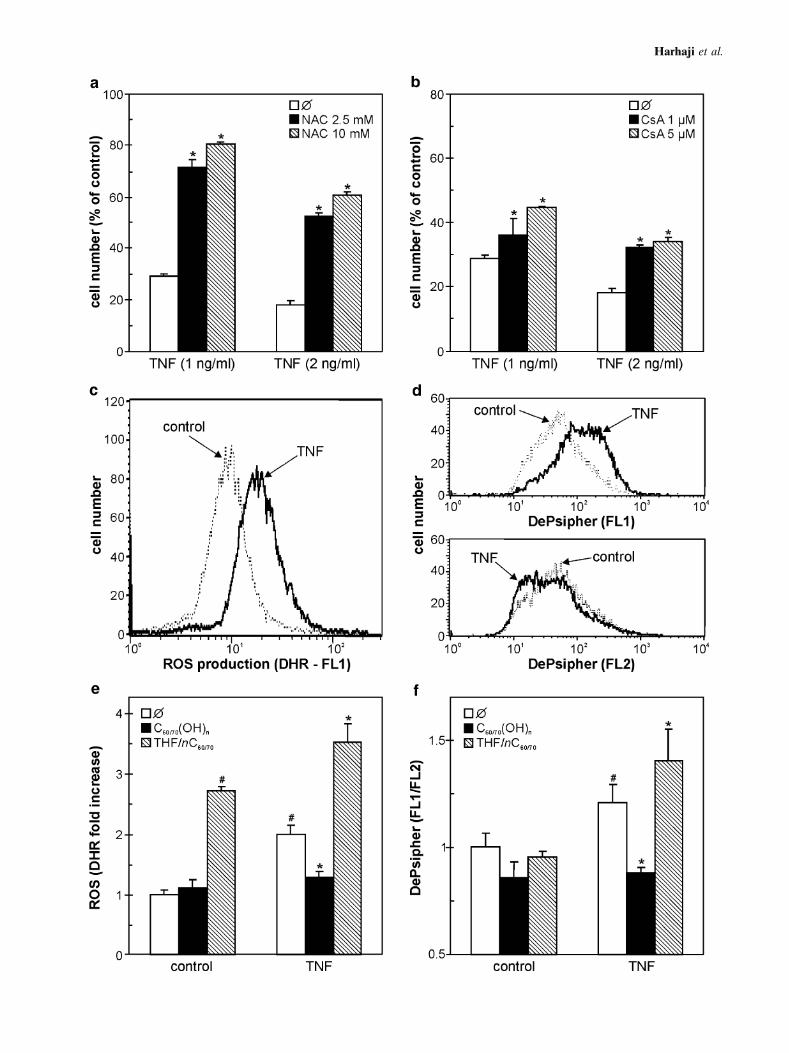
We next investigated the interference of THF/nC60/70
and C60/70(OH)n with the intracellular events responsible forTNF-induced cell death. A well-known antioxidant agent N-acetylcysteine and cyclosporine A, an inhibitor of mitochon-drial depolarization, both partially improved the viability of
TNF-treated L929 cells (Fig. 4a,b), thus indicating theinvolvement of oxidative stress and subsequent mitochondri-al failure in TNF cytotoxicity. Accordingly, treatment withTNF led to a significant production of ROS and loss ofmitochondrial membrane potential, as demonstrated by thefluorescence of redox-sensitive dye DHR (Fig. 4c) and in-crease in green vs. red fluorescence of mitochondrial-bindingdye DePsipher (Fig. 4d). THF/nC60/70 itself increased intra-cellular amount of ROS to a certain extent, while the cellstreated with both THF/nC60/70 and TNF produced more ROSthan in response to each agent alone (Fig. 4e). In contrast,C60/70(OH)n did not cause intracellular accumulation of ROSand completely prevented the TNF-triggered oxidative stress(Fig. 4e). Similar pattern of action of the two C60/70 prep-arations was observed when TNF-induced mitochondrialdepolarization was monitored in L929 cells. At the concen-trations used, fullerene preparations did not significantly affectmitochondrial membrane potential in L929 cells (Fig. 4f). TheTNF-mediated mitochondrial depolarization, however, wasmarkedly enhanced by THF/nC60/70 and efficiently preventedby C60/70(OH)n (Fig. 4f). Therefore, the opposite effects ofnC60 and C60(OH)n on TNF cytotoxicity could partly stemfrom their ability to augment and reduce, respectively, TNF-triggered ROS generation and subsequent mitochondrialdepolarization.
Synergistic Cytotoxicity of TNF and THF/nC60/70 ConfersSelectivity Towards Tumor Cells
It has previously been reported that THF-prepared nC60
at high doses is toxic to various types of transformed andprimary cells, including mouse peritoneal macrophages (12).Therefore, we wanted to examine whether combination ofsubtoxic doses of THF/nC60/70 and TNF could provide selec-tive cytotoxicity toward TNF-sensitive L929 cells, withoutdamaging TNF-resistant macrophages. Neither THF/nC60/70
nor TNF alone at subtoxic concentrations were able to mark-edly reduce viability of L929 cells or peritoneal macrophages(Fig. 5a,b). However, while combination of THF/nC60/70 andTNF exerted synergistic toxicity toward L929 cells (Fig. 5a),the viability of primary macrophages was not significantlyaffected (Fig. 5b). It therefore appears that cytotoxicity ofTHF/nC60/70 could be selectively targeted to TNF-sensitivetumor cells by combining non-cytotoxic doses of THF/nC60/70
with TNF.
Distinct Regulation of Cell Death by Fullerenesis not Specific for Cytotoxic Stimulus or Cell Type
Finally, we investigated whether the observed distincteffects of THF/nC60/70 and C60/70(OH)n are restricted to TNF-induced death of L929 cells, or they could be relevant forother cytotoxic stimuli and cell types as well. To that effect,we used human glioma cell line U251 treated with the classicanticancer drug cisplatin or hyperthermia (1h at 43-C), bothknown for their ability to induce oxidative stress-mediatedcell death (38,39). Accordingly, both treatments significantlyreduced the cell number in U251 glioma cultures (Fig. 6a–c).Consistent with the results obtained with TNF-treated L929cells, THF/nC60/70 or C60/70(OH)n potentiated or inhibited,respectively, both cisplatin- and hyperthermia-mediated kill-
R Fig. 4. The effects of fullerene preparations on TNF-induced ROS
production and mitochondrial depolarization. a, b L929 cells were
treated with TNF, in the absence or presence of N-acetylcysteine
(NAC) (a) or cyclosporin A (CsA) (b), and the cell number was
determined after 24 h. (c–f) L929 cells were incubated with TNF (0.5
ng/ml), in the absence (c, d) or presence (e, f) of THF/nC60/70 (125
ng/ml) or C60/70(OH)n (250 mg/ml). ROS production (DHR) (c, e)
and mitochondrial depolarization (DePsipher) (d, f) were analyzed
by flow cytometry after 8 h of incubation. The representative
histograms are presented in (c, d), while the data in (e, f) are mean T
SD values from three independent experiments (#p < 0.05 refers to
untreated cells; *p < 0.05 refers to cells treated with TNF alone).
Modulation of TNF Cytotoxicity by Fullerenes
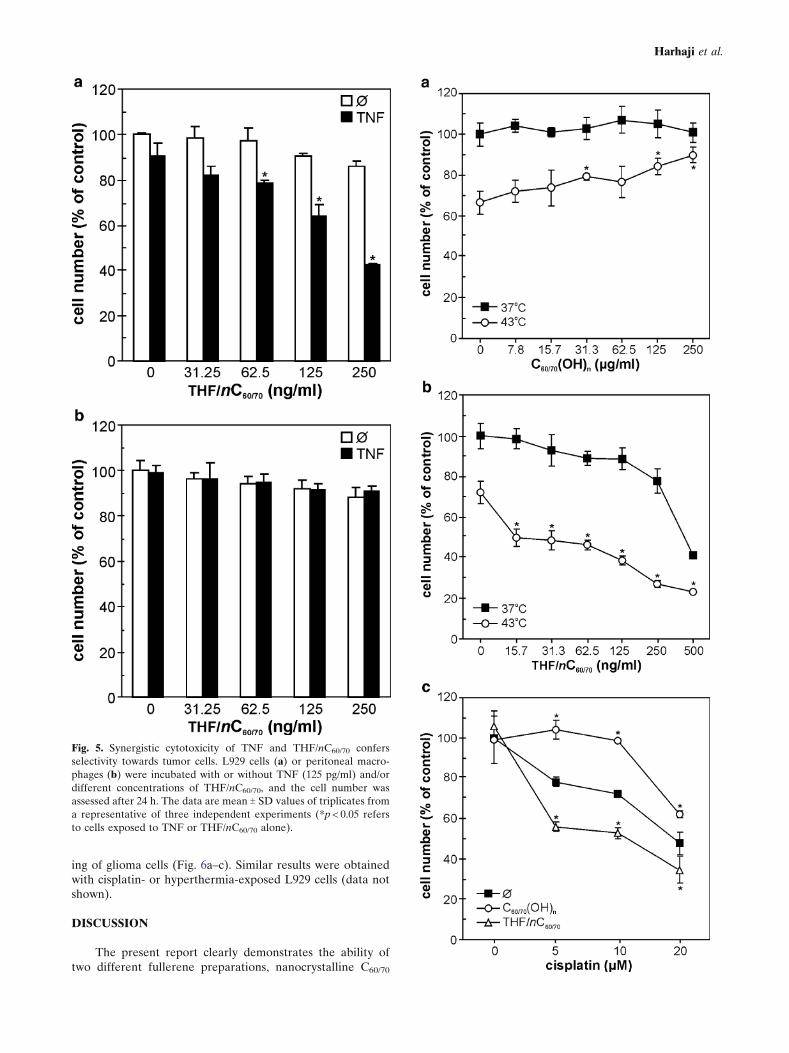
ing of glioma cells (Fig. 6a–c). Similar results were obtainedwith cisplatin- or hyperthermia-exposed L929 cells (data notshown).
DISCUSSION
The present report clearly demonstrates the ability oftwo different fullerene preparations, nanocrystalline C60/70
Fig. 5. Synergistic cytotoxicity of TNF and THF/nC60/70 confers
selectivity towards tumor cells. L929 cells (a) or peritoneal macro-
phages (b) were incubated with or without TNF (125 pg/ml) and/or
different concentrations of THF/nC60/70, and the cell number was
assessed after 24 h. The data are mean T SD values of triplicates from
a representative of three independent experiments (*p < 0.05 refers
to cells exposed to TNF or THF/nC60/70 alone).
Harhaji et al.

(THF/nC60/70) and polyhydroxylated C60/70 [C60/70(OH)n], toaugment and antagonize, respectively, the cytotoxicity of themain proinflammatory cytokine TNF. The mechanismsunderlying the observed effects possibly involved the inter-action of fullerene preparations with TNF-induced oxidativestress and subsequent mitochondrial depolarization in targetcells (Fig. 7). It should be noted that the UV absorbancespectra of the two fullerene preparations did not change inthe presence of TNF (Harhaji et al., unpublished), thusarguing against the possibility that the physico-chemicalinteraction between fullerene particles and TNF was involvedin the observed effects. This was further confirmed by the fail-ure of fullerenes to interfere with some other TNF-inducedresponses, such as expression of inducible nitric oxide synthasein the TNF-resistant C6 glioma cell line (Harhaji et al., un-published). While cytotoxic and cytoprotective properties ofTHF-prepared nanocrystalline and polyhydroxylated fuller-enes have previously been observed, this is the first reportdescribing their ability to modulate TNF-triggered cell death.It is important to note that the colloidal fullerene suspensionused in the present study was prepared from the mixture ofC60 (80%) and C70 (20%). Nevertheless, our previous studiesperformed with nanocrystals made from pure C60 and C60/C70
mixture (11,12), as well as preliminary results with pure C70
(Markovic et al., unpublished), suggest that THF/nC60 andTHF/nC70 might behave similarly, at least with regard totheir ability to exert ROS-dependent cytotoxicity.
Apoptosis and necrosis are two main cell death modeswith distinct morphological appearance and profoundly dif-ferent implications for the surrounding tissues (40, 41). Theability of TNF to induce apoptosis or necrosis has been a mat-ter of controversy, since different groups reported that TNF-mediated L929 cell killing proceeds preferentially throughapoptotic (42–44) or necrotic pathway (29, 30, 45), possiblydepending on the L929 subclone used. In some studies, how-ever, both apoptotic (internucleosomal DNA fragmentationand/or phosphatydilserine exposure) and necrotic changes(cell membrane damage) were observed in the same cell cul-ture following the TNF treatment (46–48), thus leading someresearchers to coin the terms such as Batypical apoptosis^ andBnecrapoptosis^ (49,50). This is consistent with the ap-pearance of two distinct cell populations in TNF-treatedL929 cells in our experiments, showing the characteristics ofeither apoptotic (phosphatydilserine exposure on the intactcell membrane) or necrotic cell death (the loss of cell mem-brane integrity). Interestingly, the modulation of TNF toxicityby THF/nC60/70 or C60/70(OH)n was associated with enhance-ment or suppression, respectively, of both apoptosis andnecrosis, indicating that the fullerenes might affect someintracellular event that is shared by both cell death pathways.It should be noted, however, that we could not exclude thepossibility that the cell population with membrane damage inour experiments actually consisted of late apoptotic, rather than
genuinely necrotic cells, in which case the fullerene interferencewith cell death would be limited to apoptosis induction.
Recent data indicate that process called mitochondrialpermeability transition (MPT), which precedes the loss ofmitochondrial membrane potential, could be a convergingpoint for various cell death signals, including those triggeredby TNF (50,51). The MPT pore opening and mitochondrialdepolarization are associated with the subsequent release ofsmall molecules such as cytochrome c, leading to activationof apoptosis-initiating cysteine proteases caspases. However,such perturbation of mitochondrial function may ultimatelyculminate not only in apoptosis, but also necrosis, or themixture of both, depending on cellular ATP level and othermodifying factors (50,51). The loss of mitochondrial mem-brane potential induced by TNF treatment in our study, aswell as cytoprotective effect of an MPT blocker cyclosporin A,indeed support the involvement of mitochondria in TNFtoxicity toward L929 cells. Moreover, our data indicate thatthe ability of THF/nC60/70 and C60/70(OH)n to modulate TNF-dependent apoptosis/necrosis might rely on the interferencewith TNF-induced mitochondrial depolarization, which isconsistent with the protective effect of carboxyfullerenes onTNF + cycloheximide-induced mitochondrial dysfunction inhuman peripheral blood mononuclear cells (31). This mayalso explain the observed fullerene-mediated modulation ofcaspase activation, having in mind the role of mitochondria-released factors in initiation of caspase cascades and subse-quent apoptosis. However, our results that fullerene-mediatedpotentiation (THF/nC60/70) and inhibition [C60/70(OH)n] ofcaspase activation were respectively associated with increased
Fig. 7. The mechanisms of TNF cytotoxicity modulation by fullerene
preparations. TNF induces caspase-dependent apoptosis and caspase-
independent necrosis, presumably through ROS-dependent mitochon-
drial permeability transition (MPT). THF/C60/70 and C60/70(OH)n could
increase or decrease, respectively, TNF-triggered apoptotic and necrotic
cell death by modulating ROS production and the subsequent onset of
MPT.
R Fig. 6. The effects of fullerene preparations on hyperthermia- or
cisplatin-induced death of U251 glioma cells. a, b U251 cells were
exposed to hyperthermia (1 h at 43-C) in the presence or absence of
C60/70(OH)n (a) or THF/nC60/70 (b). c U251 cells were incubated with
cisplatin, in the absence or presence of C60/70(OH)n (250 mg/ml) or
THF/nC60/70 (125 ng/ml). a–c The data are mean T SD values of
triplicates from a representative of three independent experiments
(*p < 0.05 refers to cells exposed to hyperthermia or TNF alone).
Modulation of TNF Cytotoxicity by Fullerenes

and reduced TNF cytotoxicity, might seem inconsistent withthe findings that caspase inhibition in TNF-treated L929 cellsactually enhances their sensitivity to necrosis and augmentscell death (29,30,45). A conceivable explanation for thisapparent discrepancy is that the caspase-modulating effectsof the two fullerene preparations, unlike those of the selectivecaspase inhibitors, are positioned upstream of caspase activa-tion, presumably at the mitochondrial level. In that way,fullerenes might affect a broad spectrum of cell death-associated events, including those related to necrotic celldeath pathway, and exert effects different from those ofselective caspase inhibition.
There is also a question of the mechanisms underlyingthe interference of the fullerenes with TNF-induced mito-chondrial dysfunction. It has been known that the excessiveROS generation could cause MPT pore opening, while, inturn, the collapse of the mitochondrial membrane potentialtriggers an increase in ROS generation by the electrontransfer chain, thus creating a vicious circle that leads tofurther mitochondrial and cellular injury (52). Indeed, theresults of several studies (28–30), including our experimentswith the antioxidant agent N-acetylcysteine, strongly supportthe involvement of oxidative stress in the cytotoxic action ofTNF. Moreover, the finding that ROS generation precedesTNF-triggered depolarization of mitochondrial membranepoints to an oxidative stress as a primary cause for mito-chondrial damage in TNF-treated cells (53). Accordingly, thecytoprotective effect of polyhydroxylated fullerenes wasassociated with the reduction of TNF-triggered ROS produc-tion. Having in mind the well-known ability of fullerols to actas Bfree radical sponges^ (1,7,8,11), it is conceivable toassume that this effect was due to a Bpassive^ scavenging ofROS, rather than to an Bactive^ interference with TNF-induced generation of reactive oxygen. On the other hand,nontoxic dose of THF/nC60/70 itself significantly increasedintracellular amount of ROS and enhanced TNF-induced ROSgeneration. However, the effects of TNF and THF/nC60/70 onROS generation were only additive, thus questioning the roleof oxidative stress in the synergistic cytotoxicity of the twoagents. Moreover, it should be noted that, despite the fairlylower ability for the oxidative stress induction in comparisonwith THF/nC60/70, TNF was much more efficient in causingROS-dependent mitochondrial depolarization resulting in celldemise. A plausible explanation for these discrepancies couldbe that TNF and THF/nC60/70 might induce reactive oxygenintermediates that are different and/or produced at distinctintracellular sites. Namely, while TNF treatment leads toproduction of superoxide by mitochondria and subsequentmitochondrial dysfunction (53,54), solvent exchange-preparedC60 nanocrystals apparently generate singlet oxygen (15,55)and accumulate mainly at the surface of cell membrane,causing its damage through lipid peroxidation (9,10).
CONCLUSION
The findings of the present study might be relevant forinvestigation of fullerenes as possible anticancer or cytopro-tective therapeutics. A synergistic enhancement of TNFcytotoxic action by THF/nC60/70 could be exploited in order
to reduce the toxicity of each agent alone, while still enablingefficient killing of TNF-sensitive tumor cells. As an exampleof such an approach, we have demonstrated how combiningthe noncytotoxic concentrations of TNF and THF/nC60/70 canconfer selectivity toward L929 fibrosarcoma cells withoutharming primary macrophages. On the other hand, fullerene-mediated protection from TNF cytotoxicity might be poten-tially useful in preventing TNF-mediated tissue damageassociated with TNF-based therapy or various inflammatory/autoimmune conditions. While polyhydroxylated fullerenes,because of the high retention in bone tissues (56), may not belikely drug candidates for the cytoprotective therapy, ourdata nevertheless support development of other eliminablefullerene derivatives, such as malonic acid derivatives, forsuch purpose. Finally, we have shown that the observedeffects of the two fullerene preparations are not restricted tocytotoxic action of TNF, but that they could also pertain todifferent oxidative stress-dependent cell death paradigms,such as cisplatin- or hyperthermia-mediated cytotoxicity. Ourstudy therefore warrants further investigation of potentialusefulness of fullerenes as either prooxidant/cytotoxic orantioxidant/cytoprotective therapeutics.
ACKNOWLEDGEMENTS
This work was supported by the Ministry of Science andEnvironmental Protection of the Republic of Serbia (GrantNo. 145073).
REFERENCES
1. S. Bosi, T. Da Ros, G. Spalluto, and M. Prato. Fullerenederivatives: an attractive tool for biological applications. Eur. J.Med. Chem 38:913–923 (2003).
2. L. L. Dugan, D. M. Turetsky, C. Du, D. Lobner, M. Wheeler,C. R. Almli, C. K. Shen, T. Y. Luh, D. W. Choi, and T. S. Lin.Carboxyfullerenes as neuroprotective agents. Proc. Natl. Acad.Sci. U S A 94:9434–9439 (1997).
3. L. L. Dugan, E. G. Lovett, K. L. Quick, J. Lotharius, T. T. Lin,K. L. O, and Malley. Fullerene-based antioxidants and neurode-generative disorders. Parkinsonism Relat. Disord 7:243–246 (2001).
4. C. Fumelli, A. Marconi, S. Salvioli, E. Straface, W. Malorni, A. M.Offidani, R. Pellicciari, G. Schettini, A. Giannetti, D. Monti, C.Franceschi, and C. Pincelli. Carboxyfullerenes protect humankeratinocytes from ultraviolet-B-induced apoptosis. J. Invest.Dermatol 115:835–841 (2000).
5. J. Lotharius, L. L. Dugan, and K. L. O_Malley. Distinctmechanisms underlie neurotoxin-mediated cell death in cultureddopaminergic neurons. J. Neurosci 19:1284–1293 (1999).
6. E. Straface, B. Natalini, D. Monti, C. Franceschi, G. Schettini,M. Bisaglia, C. Fumelli, C. Pincelli, R. Pellicciari, and W.Malorni. C3-fullero-tris-methanodicarboxylic acid protects epi-thelial cells from radiation-induced anoikia by influencing celladhesion ability. FEBS Lett 454:335–340 (1999).
7. L. L. Dugan, J. K. Gabrielsen, S. P. Yu, T. S. Lin, and D. W.Choi. Buckminsterfullerenol free radical scavengers reduceexcitotoxic and apoptotic death of cultured cortical neurons.Neurobiol. Dis 3:129–135 (1996).
8. H. Jin, W. Q. Chen, X. W. Tang, L. Y. Chiang, C. Y. Yang, J. V.Schloss, and J. Y. Wu. Polyhydroxylated C60, fullerenols, asglutamate receptor antagonists and neuroprotective agents. J.Neurosci. Res 62:600–607 (2000).
9. C. M. Sayes, J. D. Fortner, W. Guo, D. Lyon, A. M. Boyd, K. D.Ausman, Y. J. Tao, B. Sitharaman, L. J. Wilson, J. L. West, and
Harhaji et al.

V. L. Colvin. The differential cytotoxicity of water-solublefullerenes. Nano. Lett 4:1881–1887 (2004).
10. C. M. Sayes, A. M. Gobin, K. D. Ausman, J. Mendez, J. L. West,and V. L. Colvin. Nano-C60 cytotoxicity is due to lipidperoxidation. Biomaterials 26:7587–7595 (2005).
11. A. Isakovic, Z. Markovic, B. Todorovic-Markovic, N. Nikolic, S.Vranjes-Djuric, M. Mirkovic, M. Dramicanin, L. Harhaji, N.Raicevic, Z. Nikolic, and V. Trajkovic. Distinct cytotoxicmechanisms of pristine versus hydroxylated fullerene. Toxicol.Sci 91:173–183 (2006).
12. A. Isakovic, Z. Markovic, N. Nikolic, B. Todorovic-Markovic, S.Vranjes-Djuric, L. Harhaji, N. Raicevic, N. Romcevic, D.Vasiljevic-Radovic, M. Dramicanin, and V. Trajkovic. Inactiva-tion of nanocrystalline C60 cytotoxicity by gamma-irradiation.Biomaterials 27:5049–5058 (2006).
13. N. Gharbi, M. Pressac, M. Hadchouel, H. Szwarc, S. R. Wilson,and F. Moussa. [60]fullerene is a powerful antioxidant in vivo withno acute or subacute toxicity. Nano. Lett 5:2578–2585 (2005).
14. E. Oberdorster, S. Zhu, T. M. Blickley, P. McClellan-Green, andM. L. Haasch. Ecotoxicology of carbon-based engineered nano-particles: Effects of fullerene (C60) on aquatic organisms. Carbon44:1112–1120 (2006).
15. Z. Markovic, B. Todorovic-Markovic, D. Kleut, N. Nikolic, S.Vranjes-Djuric, M. Misirkic, L. Vucicevic, K. Janjetovic, A.Isakovic, L. Harhaji, B. Babic-Stojic, M. Dramicanin, and V.Trajkovic. The mechanism of cell-damaging reactive oxygengeneration by colloidal fullerenes. Biomaterials 28:5437–5448(2007).
16. G. Andrievsky, V. Klochkov, and L. Derevyanchenko. Is the C60
fullerene molecule toxic?!. Fullerenes, Nanotubes and CarbonNanostructures 13:363–376 (2005).
17. P. Vassalli. The pathophysiology of tumor necrosis factor. Annu.Rev. Immunol 10:411–452 (1992).
18. P. Vandenabeele, W. Declerq, R. Beyaert, and W. Fiers. Twotumor necrosis factor receptors: structure and function. TrendsCell. Biol 5:392–399 (1995).
19. R. Beyaert and W. Fiers. Molecular mechanisms of tumornecrosis factor-induced cytotoxicity. What we do understandand what we do not. FEBS Lett 340:9–16 (1994).
20. T. Nakamoto, H. Inagawa, K. Takagi, and G. Soma. A newmethod of antitumor therapy with a high dose of TNF perfusionfor unresectable liver tumors. Anticancer Res 20:4087–4096 (2000).
21. H. M. Shepard and G. D. Lewis. Resistance of tumor cells totumor necrosis factor. J. Clin. Immunol 8:333–341 (1988).
22. R. Lucas, M. Kresse, M. Latta, and A. Wendel. Tumor necrosisfactor: how to make a killer molecule tumor-specific?. CurrentCancer Drug Targets 5:381–392 (2005).
23. D. R. Spriggs, M. L. Sherman, H. Michie, K. A. Arthur, K.Imamura, D. Wilmore, E. Frei 3rd, and D. W. Kufe. Recombi-nant human tumor necrosis factor administered as a 24-hourintravenous infusion. A phase I and pharmacologic study. J.Natl. Cancer Inst 80:1039–1044 (1988).
24. S. Mocellin, P. Pilati, and D. Nitti. Towards the development oftumor necrosis factor (TNF) sensitizers: making TNF workagainst cancer. Curr. Pharm. Des 13:537–551 (2007).
25. S. Ghavami, M. Hashemi, K. Kadkhoda, S. M. Alavian, G. H.Bay, and M. Los. Apoptosis in liver diseases—detection andtherapeutic applications. Med. Sci. Monit 11:RA337–RA345 (2005).
26. M. I. Luster, P. P. Simeonova, R. M. Gallucci, A. Bruccoleri, M. E.Blazka, B. Yucesoy, and J. M. Matheson. The role of tumornecrosis factor-a in chemical-induced hepatotoxicity. Ann. N.Y.Acad. Sci 919:214–220 (2000).
27. M. Leist, F. Gantner, I. Bohlinger, P. G. Germann, G. Tiegs, andA. Wendel. Murine hepatocyte apoptosis induced in vitro and invivo by TNF-a requires transcriptional arrest. J. Immunol153:1778–1788 (1994).
28. V. Goossens, K. De Vos, D. Vercammen, M. Steemans, K.Vancompernolle, W. Fiers, P. Vandenabeele, and J. Grooten.Redox regulation of TNF signaling. BioFactors 10:145–156(1999).
29. Y. Liu, V. Tergaonkar, S. Krishna, and E. J. Androphy. Humanpapillomavirus type 16 E6-enhanced susceptibility of L929 cellsto tumor necrosis factor-alpha correlates with increased accu-
mulation of reactive oxygen species. J. Biol. Chem 274:24819–24827 (1999).
30. D. Vercammen, R. Beyaert, G. Denecker, V. Goossens, G. VanLoo, W. Declercq, J. Grooten, W. Fiers, and P. Vandenabeele.Inhibition of caspases increases the sensitivity of L929 cells tonecrosis mediated by tumor necrosis factor. J. Exp. Med187:1477–1485 (1998).
31. D. Monti, L. Moretti, S. Salvioli, E. Straface, W. Malorni, R.Pellicciari, G. Schettini, M. Bisaglia, C. Pincelli, C. Fumelli, M.Bonafe, and C. Franceschi. C60 carboxyfullerene exerts aprotective activity against oxidative stress-induced apoptosis inhuman peripheral blood mononuclear cells. Biochem. Biophys.Res. Commun 277:711–717 (2000).
32. Z. Markovic, B. Todorovic-Markovic, M. Marinkovic, and T.Nenadovic. Temperature measurement of carbon arc plasma inhelium. Carbon 41:369–371 (2003).
33. J. D. Fortner, D. Y. Lyon, C. M. Sayes, A. M. Boyd, J. C.Falkner, E. M. Hotze, L. B. Alemany, Y. J. Tao, W. Guo, K. D.Ausman, V. L. Colvin, and J. B. Hughes. C60 in water:nanocrystal formation and microbial response. Environ. Sci.Technol 39:4307–4316 (2005).
34. G. C. Zhao, P. Zhang, X. W. Wei, and Z. S. Yang. Determina-tion of proteins with fullerol by a resonance light scatteringtechnique. Anal. Biochem 334:297–302 (2004).
35. L. Harhaji, D. Popadic, D. Miljkovic, I. Cvetkovic, A. Isakovic,and V. Trajkovic. Acidosis affects tumor cell survival throughmodulation of nitric oxide release. Free Radic. Biol. Med 40:226–235 (2006).
36. D. A. Flick, and G. E. Gifford. Comparison of in vitro cellcytotoxic assays for tumor necrosis factor. J. Immunol. Methods68:167–175 (1984).
37. T. C. Chou and P. Talalay. Quantitative analysis of dose-effectrelationships: the combined effects of multiple drugs or enzymeinhibitors. Adv. Enzyme Regul 22:27–55 (1984).
38. M. Berndtsson, M. Hagg, T. Panaretakis, A. M. Havelka, M. C.Shoshan, and S. Linder. Acute apoptosis by cisplatin requiresinduction of reactive oxygen species but is not associated withdamage to nuclear DNA. Int. J. Cancer 120:175–180 (2007).
39. D. M. Katschinski, K. Boos, S. G. Schindler, and J. Fandrey.Pivotal role of reactive oxygen species as intracellular mediatorsof hyperthermia-induced apoptosis. J. Biol. Chem 275:21094–21098 (2000).
40. A. L. Edinger and C. B. Thompson. Death by design: apoptosis,necrosis and autophagy. Curr. Opin. Cell Biol 16:663–669 (2004).
41. J. L. Au, N. Panchal, D. Li, and Y. Gan. Apoptosis: a newpharmacodynamic endpoint. Pharm. Res 14:1659–1671 (1997).
42. S. Bulfone-Paus, E. Bulanova, T. Pohl, V. Budagian, H. Durkop,R. Ruckert, U. Kunzendorf, R. Paus, and H. Krause. Deathdeflected: IL-15 inhibits TNF-a-mediated apoptosis in fibroblastsby TRAF2 recruitment to the IL-15Ra chain. FASEB J 13:1575–1585 (1999).
43. J. P. Piret, T. Arnould, B. Fuks, P. Chatelain, J. Remacle, and C.Michiels. Caspase activation precedes PTP opening in TNF-a-induced apoptosis in L929 cells. Mitochondrion 3:261–278(2004).
44. J. C. Trent, D. J. McConkey, S. M. Loughlin, M. T. Harbison, A.Fernandez, and H. N. Ananthaswamy. Ras signaling in tumornecrosis factor-induced apoptosis. EMBO J 15:4497–4505(1996).
45. M. Los, M. Mozoluk, D. Ferrari, A. Stepczynska, C. Stroh, A.Renz, Z. Herceg, Z. Q. Wang, and K. Schulze-Osthoff.Activation and caspase-mediated inhibition of PARP: a molec-ular switch between fibroblast necrosis and apoptosis in deathreceptor signaling. Mol. Biol. Cell 13:978–988 (2002).
46. C. Fady, A. Gardner, F. Jacoby, K. Briskin, Y. Tu, I. Schmid,and A. Lichtenstein. Atypical apoptotic cell death induced inL929 targets by exposure to tumor necrosis factor. J. InterferonCytokine Res 15:71–80 (1995).
47. D. T. Humphreys and M. R. Wilson. Modes of L929 cell deathinduced by TNF-a and other cytotoxic agents. Cytokine 11:773–782 (1999).
48. X. Wang, N. Li, B. Liu, H. Sun, T. Chen, H. Li, J. Qiu, L. Zhang,T. Wan, and X. Cao. A novel human phosphatidylethanolamine-
Modulation of TNF Cytotoxicity by Fullerenes

binding protein resists tumor necrosis factor-a-induced apoptosisby inhibiting mitogen-activated protein kinase pathway activa-tion and phosphatidylethanolamine externalization. J. Biol.Chem 279:45855–45864 (2004).
49. A. Strelow, K. Bernardo, S. Adam-Klages, T. Linke, K. Sandhoff,M. Kronke, and D. Adam. Overexpression of acid ceramidaseprotects from tumor necrosis factor-induced cell death. J. Exp.Med 192:601–612 (2000).
50. J. J. Lemasters. Necrapoptosis and the mitochondrial perme-ability transition: shared pathways to necrosis and apoptosis.Am. J. Physiol 276:G1–G6 (1999).
51. G. Denecker, D. Vercammen, W. Declercq, and P. Vandenabeele.Apoptotic and necrotic cell death induced by death domainreceptors. Cell. Mol. Life Sci 58:356–370 (2001).
52. D. B. Zorov, M. Juhaszova, and S. J. Sollott. MitochondrialROS-induced ROS release: an update and review. Biochim.Biophys. Acta 1757:509–517 (2006).
53. S. Ko, T. T. Kwok, K. P. Fung, Y. M. Choy, C. Y. Lee, and S. K.Kong. Tumour necrosis factor induced an early release ofsuperoxide and a late mitochondrial membrane depolarizationin L929 cells. Increase in the production of superoxide is notsufficient to mimic the action of TNF. Biol. Signals Recept10:326–335 (2001).
54. T. Hennet, C. Richter, and E. Peterhans. Tumour necrosisfactor-a induces superoxide anion generation in mitochondria ofL929 cells. Biochem. J 289:587–592 (1993).
55. M. Fujitsuka, H. Kasai, A. Masuhara, S. Okada, H. Oikawa, H.Nakanishi, O. Ito, and K. Yase. Laser flash photolysis study onphotophysical and photochemical properties of C60 fine par-ticles. J. Photochem. Photobiol., A Chem 133:45–50 (2000).
56. L. Qingnuan, X. Yan, Z. Xiaodong, L. Ruili, D. Qieqie, S.Xiaoguang, C. Shaoliang, and L. Wenxin. Preparation of 99mTc-C60(OH)x and its biodistribution studies. Nucl. Med. Biol29:707–710 (2002).
Harhaji et al.