minireview microbial degradation of phthalates: biochemistry
TRANSCRIPT

Minireview
Microbial degradation of phthalates: biochemistryand environmental implications
Matthias Boll, 1* Robin Geiger,1 Madan Junghare2
and Bernhard Schink21Faculty of Biology, Microbiology, University of Freiburg,Freiburg, Germany.2Department of Biology and Microbial Ecology,University of Konstanz, Constance, Germany.
Summary
The environmentally relevant xenobiotic esters ofphthalic acid (PA), isophthalic acid (IPA) and ter-ephthalic acid (TPA) are produced on a million ton scaleannually and are predominantly used as plastic poly-mers or plasticizers. Degradation by microorganisms isconsidered as the most effective means of their elimina-tion from the environment and proceeds via hydrolysisto the corresponding PA isomers and alcohols underoxic and anoxic conditions. Further degradation of PA,IPA and TPA differs fundamentally between anaerobicand aerobic microorganisms. The latter introducehydroxyl functionalities by dioxygenases to facilitatesubsequent decarboxylation by either aromatizingdehydrogenases or cofactor-free decarboxylases. Incontrast, anaerobic bacteria activate the PA isomers tothe respective thioesters using CoA ligases or CoAtransferases followed by decarboxylation to the centralintermediate benzoyl-CoA. Decarboxylases acting onthe three PA CoA thioesters belong to the UbiD enzymefamily that harbour a prenylated flavin mononucleotide(FMN) cofactor to achieve the mechanistically challeng-ing decarboxylation. Capture of the extremely instablePA-CoA intermediate is accomplished by a massiveoverproduction of phthaloyl-CoA decarboxylase and abalanced production of PA-CoA forming/decarboxylatingenzymes. The strategy of anaerobic phthalate degrada-tion probably represents a snapshot of an ongoingevolution of a xenobiotic degradation pathway via ashort-lived reaction intermediate.
Introduction
Phthalic acids (benzene dicarboxylic acids; phthalates)consist of a benzene ring to which two carboxylic groupsare attached. Depending on the position of the carboxylicgroups to each other, three isomers are possible, namelyphthalic acid (PA; ortho-positioned), isophthalic acid (IPA;meta-positioned) and terephthalic acid (TPA; para-posi-tioned). In colloquial usage, the term ‘phthalate’ refersmost often to the ortho isomer and is also used for itsesters. Phthalic acids are essential constituents of syntheticorganic polymers that are commonly called plastics. Inthese materials, phthalates are either part of the polymericstructure as in poly(ethylene terephthalate) (PET), orthey are non-covalently dissolved as low-molecular-weightdi-esters within the polymeric structure to act as softenersor plasticizers.
Plastic materials are used in almost every aspect of mod-ern life and offer solutions and features that other materialswere not able to provide before (Andrady and Neal, 2009).Due to their low production costs and lack of cheap alterna-tives, there is an increasing demand of synthetic organicpolymers worldwide (Thompson et al., 2009). For instance,the packaging industry alone accounts for approximately42% of the global use of all non-fibre plastics, followed bythe building and construction sector accountable for about19% of non-fibre plastics (Geyer et al., 2017). It is estimatedthat the worldwide annual production of plastics will doubleto 600 million tons within 20 years (Kumar, 2018). Despitetheir numerous advantages, plastics cause increasing envi-ronmental problems and health concerns worldwide(Thompson et al., 2009). Some of these problems start onlyat the end of their designated purpose when plasticsbecome waste. Europe disposes approximately 50% of itsplastic waste in engineered landfills, while in other parts ofthe world it is still incinerated openly, emitting large amountof toxic or greenhouse gases. Huge freights of plastic mate-rials end up in the oceans where they persist for long timesand endanger birds and other marine animals. Althoughrecycling might be an option to limit production of new plas-tics, this material primarily ends up in secondary productswith less demanding material properties and unclear fate,and is hence not a permanent solution. Furthermore,
*For correspondence. E-mail [email protected];Tel. (+49) 761 2032649; Fax (+49) 761 2032626.
© 2019 The Authors. Environmental Microbiology Reports published by Society for Applied Microbiology and John Wiley & Sons Ltd.This is an open access article under the terms of the Creative Commons Attribution License, which permits use, distribution andreproduction in any medium, provided the original work is properly cited.
Environmental Microbiology Reports (2020) 12(1), 3–15 doi:10.1111/1758-2229.12787

recycling will not prevent the rise of microplastics in marineenvironments which is a growing problem that is difficult tosolve (Hahladakis et al., 2018). Therefore, it is important todevelop new strategies of waste management and to betterunderstand plastic biodegradation as a sustainable per-spective for removal of plastics in nature.
Phthalic acid ester use and biodegradation
Phthalic acid polyesters. Phthalates can form essential build-ing blocks inside plastic polymers (1). In these cases,phthalates (most often TPA) are bound via ester linkages todivalent alcohols such as ethylene glycol, 1,4-butanediol,1,6-hexanediol, and so on. Needless to say, these covalentlylinked TPA residues are rather difficult to release from theirpolymeric mother backbones, as this is true for synthetic poly-mers in general. Although ester bonds are comparably easy tocleave by hydrolysis and require only water as a co-substrate,degradation of such polymers is difficult because a hydrolyticenzyme first has to get access to such polymer fibres, and thiswould require swelling of the plastic material by excessivewater uptake. Plastics are typically produced as water-resis-tant, rather hydrophobic structures that are hardly accessibleto hydrolytic enzymes. Moreover, hydrolytic enzymes pro-duced by microbes are typically synthesized rather specificallyfor attack on specific ester structures and do not interact freelywith novel ester linkages. Last but not least, synthesis of extra-cellular enzymes for polymer degradation is an expensiveeffort for a microbial cell and has to guarantee a sufficient pay-back by easily degradable breakdown products that can coverthe energy expenditure for enzyme synthesis on the side ofthe producer. Thus, the breakdown products should be ‘palat-able’ and should easily provide metabolic energy for a broadcommunity of microbes of which then only few would dare toinvest into the development of suited enzymes for polymerhydrolysis. As we will see later, the biochemistry of phthalatedegradation is a rather demanding task, and the aromaticderivatives formed in its breakdown are energetically not toorewarding to justify every effort into such an enterprise. Poly-saccharides, proteins, nucleic acids and so on are much moreappealing in this respect. This is true also for the natural poly-esters, e.g., poly-3-hydroxybutyric acid, a bacterial storagepolymer which is easily degradable by a broad community ofaerobic and anaerobic bacteria (Jendrossek and Handrick,2002; Hazer and Steinbüchel, 2007).
PET (Fig. 1A) is produced at high rate (56 million tons in2013; Kawai et al., 2019), mainly as containers for soft drinksand so on. Its microbial degradation is very slow and incom-plete. It appears that only surface-exposed residues aredegraded by microbes to a limited extent through the action ofcutinases, lipases and other carboxylesterases (Kawai et al.,2019). As this plastic material is applied mainly as drink con-tainers it has little tendency to take up water, and this limitsseverely a sufficient interaction of microbial esterases with themicrofibers of this plastic material (Tokiwa et al., 2009; Wei andZimmermann, 2017). Slow degradation of low-crystallinity PETto TPA and ethylene glycol was reported for a new aerobic bac-terial isolate, Ideonella sakaiensis (Yoshida et al., 2016).
Also ‘biodegradable’ synthetic phthalate polyesters arecommercially available, especially for use as protecting
covers in agriculture, or for plastic shopping bags. The BASFcompany produces Ecoflex®, a copolymer of 1,4-butanediolwith alternating terephthalate and adipate (hexane dioic acid)residues (poly butylene-adipate-terephthalate; PBAT). Thematerial decomposes in compost and agricultural soil withinseveral months and thus fulfils its task to survive one vegeta-tion period and to disappear before the next one starts. Inour lab, we could confirm that this material decomposesunder oxic conditions and also anaerobically in the presenceof nitrate, although nitrate-dependent degradation turned outto be far slower with terephthalate-containing polymers thanwith adipate-containing ones. There was no significant deg-radation of either type of polymer in incubations under strictlyanoxic conditions with either sulfate or CO2 as the only elec-tron acceptor (Schink, unpublished). This is surprising ashydrolytic depolymerization of such synthetic material shouldbe possible under any condition, independent of the avail-able electron acceptor. We assume that the probability ofdevelopment of efficient polymer-degrading hydrolases maybe higher in the aerobic and nitrate-dependent microbialworld than among the strict anaerobes. Moreover, productionof extracellular hydrolases for unusual polymeric substratesmay be too ‘expensive’ for strict anaerobes with their limitedenergy budgets. We have to conclude that phthalate resi-dues that are covalently bound inside polymeric plasticmaterials are released only slowly, and their releasedepends largely on the physicochemical properties of theplastic material and the incubation conditions.
Occurrence and use of isophthalic acid. IPA is an importantisomer produced on the million ton per year scale by oxida-tion of m-xylene (Sheehan, 2005). It is mainly used as a
Fig. 1. Examples of industrially produced esters of the three phthalicacid isomers. A. Polyethylenglycol terephthalate ester polymer (PET).B. Polyethylenglycol IPA esters used as co-polymer of PET. C. PA estersused as plasticizers, for typical alcohols residues (=R) see Table 1.
© 2019 The Authors. Environmental Microbiology Reports published by Society for Applied Microbiology and John Wiley & Sons Ltd.,Environmental Microbiology Reports, 12, 3–15
4 M. Boll, R. Geiger, M. Junghare and B. Schink

feedstock for the production of unsaturated polyesters, anal-ogous to terephthalate, and is also a co-monomer in somecommon plastics such PET (Fig. 1B). Furthermore, iso-phthalate is an important constituent of the extremely heat-stable polymer polybenzimidazole (Sheehan, 2005).
Phthalic acid esters as synthetic plasticizers. Low-molecu-lar-weight PA isomer esters–nearly all of them PA esters(PAE) (Fig. 1C)–are essential constituents of synthetic plas-tics where they act as plasticizers. These are additives thatare freely dissolved within the polymeric plastic resins andincrease the plasticity or elasticity of a material (Giam et al.,1984; Nilsson, 1994). PAEs were introduced in the 1920s toreplace camphor-based plasticizers. In order to provide therequired elasticity to plastic products, they are not boundcovalently to the plastic material and can easily leach out(Nilsson, 1994). Today, more than 60 kinds of PAEs are pro-duced for different commercial applications such as plasticmanufacturing, additives in paints, lubricants, adhesives,insecticides, material packaging, and cosmetics (Vats et al.,2013; Benjamin et al., 2015). Their estimated global produc-tion is growing rapidly with 1.8 million tons in 1975, reaching6.2 million tons in 2009 and more than 8 million tons in 2011(Peijenburg and Struijs, 2006; Meng et al., 2014; Net et al.,2015). The most frequently utilized PAEs for different appli-cations are listed in Table 1.
Due to the high global utilization of plasticizers, they arereleased during production and distribution of plastics in nearlyevery environment (Giam et al., 1984) such as air, freshwater,oceans, sediments, soil, wastewater, household dust and foodproducts (Gao and Wen, 2016), independent of the degrada-tion of the mother polymer itself. Their continuous release fromplastics leads to environmental contamination. PAEs aredescribed as man-made priority pollutants due to their highexposure to humans and adverse effects on their health. Cer-tain PAEs have adverse effects on the development of themale reproductive system due to their anti-androgenic proper-ties (Latini et al., 2006; Lambrot et al., 2009). They are alsoconsidered as potential carcinogens, teratogens, mutagens,
and endocrine disruptors (Fushiwaki et al., 2003). Thus, fromthe beginning there has been great concern about the releaseof PAEs into the environment (Mayer et al., 1972; Giam et al.,1978). Furthermore, many PAEs have been blacklisted by theU.S. Environmental Protection Agency and the Chinese Envi-ronment Monitoring Center and banned, especially for theiruse in children’s toys (Net et al., 2015).
Abiotic and biotic degradation of PA esters. PAEs can bedegraded in the environment by abiotic or biotic reactions suchas hydrolysis (Jonsson et al., 2006), UV light photolysis (Lauet al., 2005), TiO2-enhanced photocatalysis (Sin et al., 2012)and microbial degradation (Chao et al., 2006; Li et al., 2006).However, abiotic (non-biological) degradation of PAEs is veryslow, e.g., butyl-benzyl phthalate has an aqueous photolysishalf-life of >100 days, for dimethyl phthalate it is about 3 years,and for ethyl hexylphthalate 2000 years (Gledhill et al., 1980;Staples et al., 1997). Compared to non-biological destruction,microbial degradation is rather fast and a promising, cost-effective and environmentally friendly method for removal ofphthalates from contaminated sites. For instance, removal ofPAEs has been reported from various environments, includingnatural water, soils (Carrara et al., 2011), sediments (Changet al., 2005), wastewater (Camacho-Munoz et al., 2012), andlandfills (Boonyaroj et al., 2012).
PAE-degrading microbes are either aerobic, facultativelyanaerobic or strictly anaerobic microorganisms. In most cases,the primary step is the initial hydrolysis of PAEs by esterasesthat release the free phthalate moiety and side chain alcohols.In addition, transesterification or the conversion of longer alkylresidues to shorter ones by unknown enzymes have beenreported occasionally (Liang et al., 2008). Esterases acting onPAEs have been characterized from several bacteria:e.g., esterases purified from Micrococcus sp. strain YGJ1, andfrom Bacillus sp. hydrolyze medium-chain (C3-C5) mono/di-alkyl phthalate esters (Niazi et al., 2001; Maruyama et al.,2005). Most often the aliphatic or aromatic side chain alcoholsare easily utilized by bacteria, and phthalate accumulates dueto a lack of essential enzymes for its degradation. At leastunder anoxic conditions, degradation of phthalate is consideredthe rate-limiting step in degradation of PAEs (Gao and Wen,2016). The complete mineralization of PAEs requires a combi-nation of the metabolic capacities of diverse microbes thatcould accomplish the degradation of side chain alcohols aswell as of the phthalate residues. For instance, degradation ofdi-n-octyl phthalate is carried out by a metabolic cooperationbetween two bacteria: e.g., Gordonia sp. strain JDC-2 andArthrobacter sp. strain JDC-32 (Wu et al., 2010). In this co-cul-ture, Gordonia sp. converts di-n-octyl phthalate (DOP) into PA,which is degraded by Arthrobacter sp. strain JDC-32 that can-not degrade DOP alone.
Aerobic biodegradation of PA isomers
The aerobic biodegradation of PA, IPA and TPA hasbeen studied since almost 60 years (Ribbons andEvans, 1960; Keyser et al., 1976), and has meanwhilebeen demonstrated in a large number of gram-positiveand -negative bacteria, as well as in some fungi. Fordetailed lists of species that use the three PA isomers as
Table 1. Applications of selected phthalate esters that are commonlyused in industry.
Phthalate ester Application
Dimethyl phthalate Insect repellent/ectoparasiticide, e.g.,against mosquitoes and flies
Diethyl phthalate Often used in cosmetics, fragrancesand other industrial uses includingplasticizers, aerosol sprays etc.
Butyl-benzyl phthalate Mostly used as a plasticizer for PVC.Other uses include traffic cones, foodconveyor belts, artificial leather etc.
Diisononyl phthalate Mostly used as a plasticizerDi(2-ethylhexyl)
phthalateUsed as plasticizer in medical devices,
e.g., intravenous tubing, IV catheters,nasogastric tubes, dialysis bags andtubing, blood bags and transfusiontubing, air tubes
Diisodecyl phthalate Mostly used as a plasticizerDibutyl phthalate Used as a plasticizer or as an additive
in adhesives or printing inks
© 2019 The Authors. Environmental Microbiology Reports published by Society for Applied Microbiology and John Wiley & Sons Ltd.,Environmental Microbiology Reports, 12, 3–15
Anaerobic degradation of phthalates 5

sole source of carbon and energy we refer to previousreviews (Vamsee-Krishna and Phale, 2008; Gao andWen, 2016). Here, we summarize the common enzymaticstrategies of aerobic PA, IPA and TPA degradation inorder to compare them with the alternative ones employedby anaerobic bacteria.Uptake of PA has been studied only in a few cases on
the biochemical level. In Burkholderia multivorans ATCC17616, there are two transport systems for PA uptake: theOphD permease and an ABC transporter-type system(Chang et al., 2009). Here, the specific porin OphP medi-ates PA transport across the outer membrane. An ABCtransporter-type system has also been reported to beinvolved in PA uptake during PAE degradation inRhodococcus jostii RHA1 (Hara et al., 2010). In contrast, atripartite tricarboxylate transporter-type is involved in TPAuptake in Comamonas sp. Strain E6 (Hosaka et al., 2013).In almost all aerobic microorganisms the three phthalate
isomers are converted to protocatechuate (3,4-dihydroxy-benzoate), a central intermediate of aerobic degradation ofaromatic compounds (Fig. 2). The conversion of PA to pro-tocatechuate in aerobicmicroorganisms involves three steps:(i) dioxygenation to cis-dihydroxy-dihydro-PA, (ii) dehydroge-nation to dihydroxy-PA and (iii) decarboxylation to pro-tocatechuate. In contrast, formation of protocatechuate fromTPA and most possibly also IPA affords only two steps asdecarboxylation is achieved directly in step (ii). Thus, themandatory decarboxylation of the three phthalic acid isomersis accomplished either by special decarboxylases (PA), orby cis-dihydrodiol dehydrogenases that abstract CO2
instead of a proton during hydride transfer (IPA, TPA). Theprotocatechuate formed is finally cleaved by dioxygenasesand converted to central intermediates either via the ortho-or meta-cleavage pathways (Dagley, 1971; Stanier andOrnston, 1973). A variant of these canonical PA isomer degra-dation pathways has been described recently in Pseudomo-nas sp. strain PTH10 during growth with PA involving aphthalate-2,3-dioxygenase and a decarboxylating 2,3-dihydro-cis-2,3-dihydroxy-PA dehydrogenase (Kasai et al.,2019). The ring system of the cis-2,3-dihydroxybenzoate inter-mediate formed is then directly cleaved by a 3,4-dioxygenase.The enzymes involved in the degradation of the three PA iso-mers are briefly described in the following.
Two-component dioxygenases. The initial dihydroxylation ofPA isomers has been studied most intensively with PA as sub-strate. Its dihydroxylation to cis-dihydrodiol-PA is catalysed bytwo-component Rieske non-heme iron oxygenase systemscomprising (i) a NAD(P)H-dependent dioxygenase reductasecomponent that harbours FMN and a plant-type [2Fe-2S] ferre-doxin, and (ii) a non-heme dioxygenase component binding aRieske-type [2Fe-2S] cluster and an active site Fe atom [someselected references: (Batie et al., 1987; Correll et al., 1992;Tarasev and Ballou, 2005; Karandikar et al., 2015)]. There aretwo types of PA dioxygenases yielding either 4,5-dihydro-cis-
4,5-dihydroxy-PA (mostly in gram-negative bacteria) or3,4-dihydro-cis-3,4-dihydroxy-PA (mostly in gram-positive bac-teria) (Fig. 2). In both products a hydroxyl group is positioned inortho or para to one of the two carboxyl functionalities, which isessential for the down-stream decarboxylation reaction.
TPA is initially attacked by a 1,2-dioxygenase yielding2-hydro-cis-1,2-dihydroxy-TPA (Schläfli et al., 1994; Choiet al., 2005; Fukuhara et al., 2008), which can be directlydecarboxylated by a cis-1,2-dihydroxy dehydrogenase. Lessis known about the ring-hydroxylating oxygenase involved inIPA degradation. It has been shown that it specifically formsthe 4-hydro-cis-3,4-dihydroxy-IPA product (Vamsee-Krishnaet al., 2006).
Aromatizing cis-diol-dehydrogenases. Decarboxylating,NAD+-dependent 2-hydro-cis-1,2-dihydroxy-TPA dehydroge-nases have been purified and structurally characterized fromdifferent bacteria (Saller et al., 1995; Bains et al., 2012). Here,dehydrogenation proceeds via elimination of a hydride from C2accompanied by decarboxylation at C1 (Fig. 2B). A number ofdecarboxylating 2-hydro-cis-dihydro-dihydroxy-PA dehydroge-nases have been identified based on the correspondingphthalate-induced genes (Chang and Zylstra, 1998; Stingleyet al., 2004; Choi et al., 2005; Carbone et al., 2008).
Dihydroxyphthalate decarboxylases. So far, two 4,5-dihydroxy-PA decarboxylases have been purified and initiallycharacterized that both form the common protocatechuate inter-mediate. Here, decarboxylation of the carboxyl functionality inpara-position to a phenolic group is largely facilitated, and conse-quently these decarboxylases do not require a special cofactorfor catalysis (Fig. 2C), (Nakazawa and Hayashi, 1978; Pujar andRibbons, 1985).
Environmental importance of anaerobic biodegradation
Being surrounded by air containing 21%ofmolecular oxygen,we consider aerobic processes to be most important for deg-radation of organic compounds. For this reason microbialdegradation of novel synthetic compounds has always beenstudied under oxic conditions in the first place. However, oxy-gen can quickly become limiting wherever its supply dependson diffusive rather than convective transport. In the late1970s, Julian Wimpenny and coworkers showed that evencolonies of aerobic bacteria on a Petri dish may be entirelyanoxic inside, i.e., that a film of about 50 μm thickness (about50 layers of bacterial cells) is sufficient to consume the atmo-spheric oxygen entirely (Wimpenny and Coombs, 1983). Insediments of freshwater lakes or marine water bodies, oxy-gen contents decrease from air saturation to zero within fewmillimetres, depending on their trophic status and on theirsupply with easily degradable organic matter. In soil, oxygenmay reach down into the ground by few centimetres, strictlydepending on thewater content.Water-logged soils, e.g., ricepaddies, are largely oxygen-free, similar to lake sediments.Waste dump sites such as landfills contain a broad variety oforganic materials including various types of plastics. Their ini-tial aerobic biodegradationwill consume the available oxygen
© 2019 The Authors. Environmental Microbiology Reports published by Society for Applied Microbiology and John Wiley & Sons Ltd.,Environmental Microbiology Reports, 12, 3–15
6 M. Boll, R. Geiger, M. Junghare and B. Schink
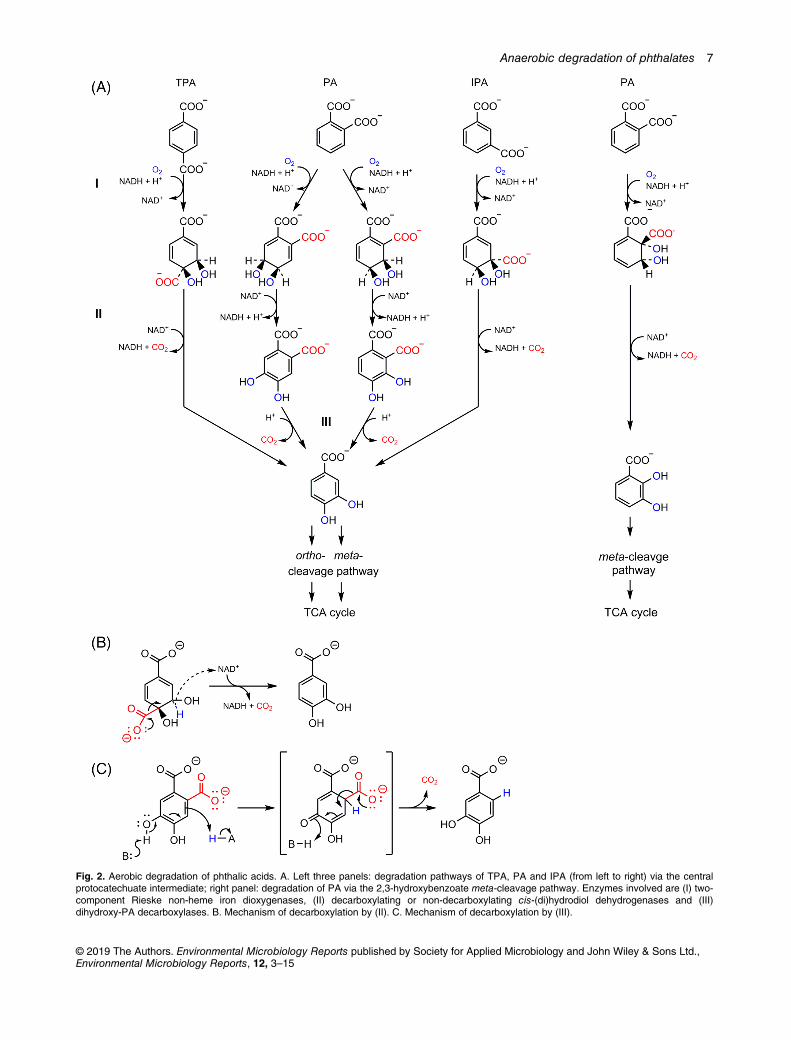
Fig. 2. Aerobic degradation of phthalic acids. A. Left three panels: degradation pathways of TPA, PA and IPA (from left to right) via the centralprotocatechuate intermediate; right panel: degradation of PA via the 2,3-hydroxybenzoate meta-cleavage pathway. Enzymes involved are (I) two-component Rieske non-heme iron dioxygenases, (II) decarboxylating or non-decarboxylating cis-(di)hydrodiol dehydrogenases and (III)dihydroxy-PA decarboxylases. B. Mechanism of decarboxylation by (II). C. Mechanism of decarboxylation by (III).
© 2019 The Authors. Environmental Microbiology Reports published by Society for Applied Microbiology and John Wiley & Sons Ltd.,Environmental Microbiology Reports, 12, 3–15
Anaerobic degradation of phthalates 7

quickly, resulting in an entirely anoxic zone below a thin coverlayer. It is obvious from these few examples that microbialdegradation of plastic wastes often has to face oxygen limita-tion or even entirely anoxic conditions, be it in soils (protectivefoils in agriculture), sediments (accidental spills and losses)or landfills. Thus, considerations on the (bio)degradability ofplastic constituents under entirely anoxic (reducing) condi-tions are of global relevance. Even if the polymeric backbonestructures may be hardly degraded under anoxic conditions,the slow but steady release of low-molecular-mass plasti-cizers such as phthalate di-esters continues in any case.
Anaerobic biodegradation of PA isomers
The use of dioxygen as substrate for ring-hydroxylating andring-cleaving dioxygenases is not an option for anaerobicphthalate degradation. In anaerobic bacteria most monocy-clic aromatic compounds, including the three PA isomers,are converted to the central intermediate benzoyl-CoA(Carmona et al., 2009; Fuchs et al., 2011; Rabus et al.,2016). Exceptions are only aromatics that contain at leasttwo meta-positioned hydroxyl functionalities which substan-tially weaken the aromatic character (Philipp and Schink,2012). Benzoyl-CoA is dearomatized to cyclohexa-1,-5-diene-1-carboxyl-CoA either by ATP-dependent classI benzoyl-CoA reductases (BCRs) present in facultativeanaerobes (Boll et al., 2014; Tiedt et al., 2018), or by ATP-independent class II BCRs that occur in strict anaerobes(Weinert et al., 2015; Huwiler et al., 2019). The further degra-dation to acetyl-CoA units and CO2 proceeds in the so-called benzoyl-CoA degradation pathway by a series ofmodified β-oxidation like reactions including a hydrolytic ringcleavage step (Carmona et al., 2009; Fuchs et al., 2011; Bollet al., 2014). Thus, the basic question of anaerobic PA, IPAand TPA degradation is how they are converted to the cen-tral intermediate benzoyl-CoA.
Anaerobic degradation of phthalate
Degradation of phthalate in denitrifying bacteria.Biodegradability of PA in denitrifying bacteria was reportedalready in the early eighties (Aftring et al., 1981). It was soonpostulated that in anaerobic bacteria PA is decarboxylated tobenzoate followed by thioesterification, e.g. by an ATP-dependent benzoate CoA ligase (Eaton and Ribbons, 1982;Taylor and Ribbons, 1983). However, decarboxylation of anyof the three phthalate isomers to benzoate would afford anextremely unfavourable dianionic intermediate. As an alter-native, initial activation to phthaloyl-CoA was suggested,followed by decarboxylation to benzoyl-CoA (Nozawa andMaruyama, 1988). Experimental evidence for such a sce-nario was lacking for almost another 20 years. In two recent stud-ies, complete PA degradation to CO2 coupled to denitrificationwas reported for the newly isolated Azoarcus sp. strain PA01(Junghare et al., 2015; Junghare et al., 2016) and for a number ofknown aromatic compound degrading denitrifyers such as
Thauera chlorobenzoica 3CB-1, Aromatoleum aromaticumEbN1 and Azoarcus evansii KB740 [now reclassified asAromatoleum evansii KB740, (Rabus et al., 2019)], (Ebenau-Jehle et al., 2017). All denitrifying phthalate degraders grow withphthalate and nitrate at doubling times in the range of 10 to 15 hwhich opened the door for obtaining sufficient biomass to studythe genes, enzymes and metabolites involved in anaerobic PAdegradation.
In vitro assays with extracts of these phthalate-degrading,denitrifying pure cultures provided evidence for the succinyl-CoA-dependent conversion of phthalate to the central intermedi-ate benzoyl-CoA indicating the following reaction sequence:(i) activation of phthalate to phthaloyl-CoA by a succinyl-CoAdependent CoA transferase and (ii) decarboxylation of theformed phthaloyl-CoA to benzoyl-CoA by a decarboxylase(Junghare et al., 2016; Ebenau-Jehle et al., 2017). Differentialproteome analyses of Aromatoleum EbN1 (Ebenau-Jehle et al.,2017) and Azoarcus sp. strain PA01 (Junghare et al., 2016) ofcells grown with phthalate versus benzoate revealed phthalate-induced gene clusters comprising genes encoding transportersand enzymes involved in uptake and degradation of PA. InA. aromaticum EbN1 the genes involved in anaerobic phthalatedegradation are located on the conjugative p2A plasmid(0.22 Mb), whereas in other denitrifying bacteria they are inte-grated into the chromosome. The genes, enzymes and reac-tions involved in phthalate degradation in selected anaerobicbacteria are depicted in Fig. 3, and will be discussed in thefollowing.
Uptake of PA All phthalate-induced gene clusters of den-itrifying phthalate degraders contain genes putatively encodinga tripartite ATP-independent periplasmic (TRAP) transporter(Junghare et al., 2016; Ebenau-Jehle et al., 2017), a kind oftransport system typically involved in the uptake of dicarboxylicacids (Forward et al., 1997; Mulligan et al., 2011). These sec-ondary transporters involved in phthalate uptake comprise aperiplasmic binding protein and a permease subunit with twofused domains. In A. aromaticum EbN1 and A. toluclasticus,genes for an additional three-component ABC transporter sys-tem were identified including a periplasmic binding protein, apermease and an ATPase domain. It is unknown whether theTRAP- and ABC-type components both transport phthalateunder different conditions or whether they transport differentsubstrates.
Succinyl-CoA:phthalate CoA transferase (SPT) and insta-bility of phthaloyl-CoA. In denitrifying bacteria PA degrada-tion is initiated by a class III CoA transferase that specificallyuses succinyl-CoA as CoA donor, and PA as CoA acceptor(Junghare et al., 2016; Ebenau-Jehle et al., 2017). Theheterodimeric enzyme from A. aromaticum EbN1 was heter-ologously produced in Escherichia coli by expression of theencoding sptAB genes. The isolated enzyme predominantlycatalysed the phthalate-dependent hydrolysis of succinyl-CoA, whereas phthaloyl-CoA was formed only at sub-micromolar concentrations (Mergelsberg et al., 2018). Thisfinding is rationalized by the extreme instability of phthaloyl-CoA (half-life <7 min) that is prone to intracellular substitu-tion yielding CoA and phthalic anhydride, the latter is thenimmediately hydrolyzed to phthalate (Fig. 4). The extremeinstability is also reflected by the low cellular phthaloyl-CoAconcentration which is more than three orders of magnitudebelow that of benzoyl-CoA (Mergelsberg et al., 2018).
© 2019 The Authors. Environmental Microbiology Reports published by Society for Applied Microbiology and John Wiley & Sons Ltd.,Environmental Microbiology Reports, 12, 3–15
8 M. Boll, R. Geiger, M. Junghare and B. Schink
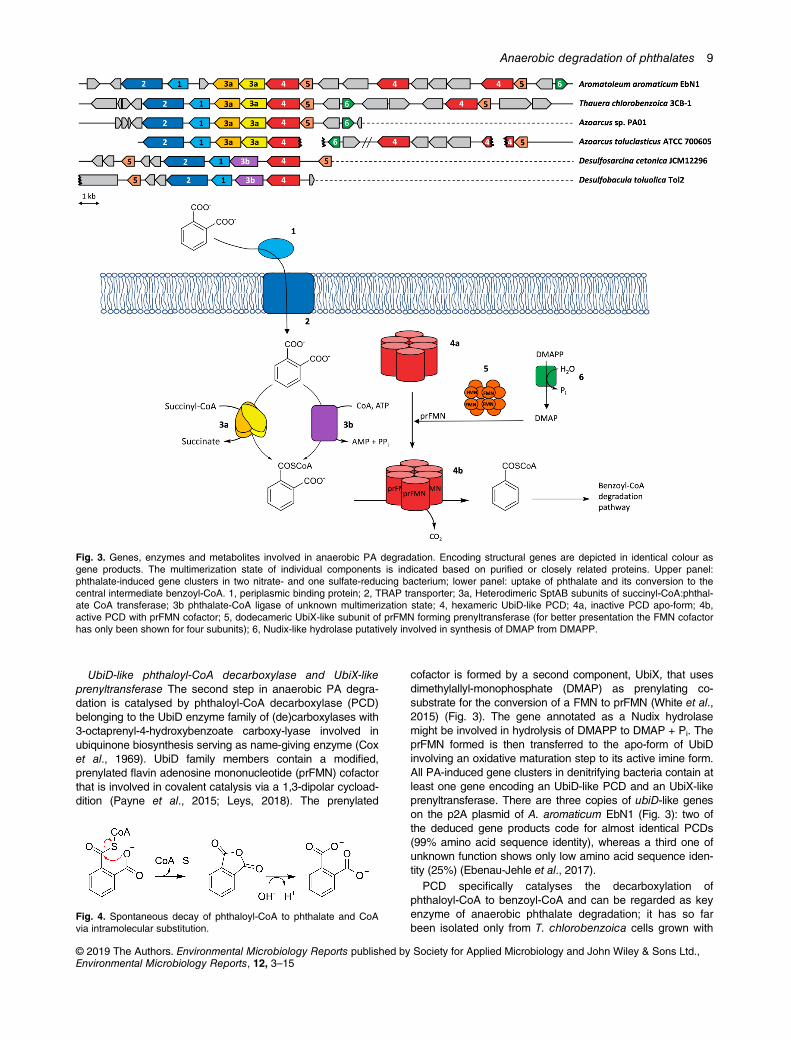
UbiD-like phthaloyl-CoA decarboxylase and UbiX-likeprenyltransferase The second step in anaerobic PA degra-dation is catalysed by phthaloyl-CoA decarboxylase (PCD)belonging to the UbiD enzyme family of (de)carboxylases with3-octaprenyl-4-hydroxybenzoate carboxy-lyase involved inubiquinone biosynthesis serving as name-giving enzyme (Coxet al., 1969). UbiD family members contain a modified,prenylated flavin adenosine mononucleotide (prFMN) cofactorthat is involved in covalent catalysis via a 1,3-dipolar cycload-dition (Payne et al., 2015; Leys, 2018). The prenylated
cofactor is formed by a second component, UbiX, that usesdimethylallyl-monophosphate (DMAP) as prenylating co-substrate for the conversion of a FMN to prFMN (White et al.,2015) (Fig. 3). The gene annotated as a Nudix hydrolasemight be involved in hydrolysis of DMAPP to DMAP + Pi. TheprFMN formed is then transferred to the apo-form of UbiDinvolving an oxidative maturation step to its active imine form.All PA-induced gene clusters in denitrifying bacteria contain atleast one gene encoding an UbiD-like PCD and an UbiX-likeprenyltransferase. There are three copies of ubiD-like geneson the p2A plasmid of A. aromaticum EbN1 (Fig. 3): two ofthe deduced gene products code for almost identical PCDs(99% amino acid sequence identity), whereas a third one ofunknown function shows only low amino acid sequence iden-tity (25%) (Ebenau-Jehle et al., 2017).
PCD specifically catalyses the decarboxylation ofphthaloyl-CoA to benzoyl-CoA and can be regarded as keyenzyme of anaerobic phthalate degradation; it has so farbeen isolated only from T. chlorobenzoica cells grown with
Fig. 4. Spontaneous decay of phthaloyl-CoA to phthalate and CoAvia intramolecular substitution.
Fig. 3. Genes, enzymes and metabolites involved in anaerobic PA degradation. Encoding structural genes are depicted in identical colour asgene products. The multimerization state of individual components is indicated based on purified or closely related proteins. Upper panel:phthalate-induced gene clusters in two nitrate- and one sulfate-reducing bacterium; lower panel: uptake of phthalate and its conversion to thecentral intermediate benzoyl-CoA. 1, periplasmic binding protein; 2, TRAP transporter; 3a, Heterodimeric SptAB subunits of succinyl-CoA:phthal-ate CoA transferase; 3b phthalate-CoA ligase of unknown multimerization state; 4, hexameric UbiD-like PCD; 4a, inactive PCD apo-form; 4b,active PCD with prFMN cofactor; 5, dodecameric UbiX-like subunit of prFMN forming prenyltransferase (for better presentation the FMN cofactorhas only been shown for four subunits); 6, Nudix-like hydrolase putatively involved in synthesis of DMAP from DMAPP.
© 2019 The Authors. Environmental Microbiology Reports published by Society for Applied Microbiology and John Wiley & Sons Ltd.,Environmental Microbiology Reports, 12, 3–15
Anaerobic degradation of phthalates 9

phthalate (Mergelsberg et al., 2017). Due to the instability ofthe phthaloyl-CoA substrate, PCD is routinely measured in acoupled assay together with SPT and the formation ofbenzoyl-CoA from phthalate and succinyl-CoA is followed.Hexameric PCD contains prFMN, K+ and Fe2+ as cofactors.In the presence of oxygen, the Fe2+ is oxidized to Fe3+
resulting in a loss of the prFMN cofactor and activity. It isproposed that K+ and Fe2+ (evidenced by Fe-determinationand EPR spectroscopy) are both involved in prFMN binding(Mergelsberg et al., 2017), with the Fe2+ found in PCDsubstituting for the Mn2+ found in UbiD-like enzymes fromaerobic organisms (Payne et al., 2015). It is unknownwhether all UbiD-like enzymes from anaerobes contain Fe2+
instead of Mn2+, and whether the metal is involved in a redoxprocess during maturation of the prFMN cofactor.
PCD is the only member of the UbiD-like family of (de)car-boxylases that use a CoA-ester substrate. The CoA-ester func-tionality may contribute to the stabilization of a putative anionicintermediate in the course of catalysis. However, it is unknownhow the prFMN cofactor promotes phthaloyl-CoA decarboxyl-ation. The 1,3-dipolar cycloaddition mechanism proposed forthe decarboxylation of cinnamic acid and related derivativescannot be applied for PCD as the direct cleavage of a carboxylfunctionality from the aromatic ring would necessitate anunfavourable dearomatization step (Mergelsberg et al., 2017).
Capture of labile phthaloyl-CoA The formation of theextremely labile phthaloyl-CoA intermediate is a major chal-lenge of anaerobic PA degradation. The relatively high appar-ent Km of PCD for phthaloyl-CoA of around 120 μM, 200-foldhigher than the cellular phthaloyl-CoA concentration, togetherwith the low specific activity (Vmax ≈ 35 nmol min−1 mg−1) arenot indicative for an efficient phthaloyl-CoA turnover by PCD(Mergelsberg et al., 2017). For this reason, it had beensuggested initially that SPT and PCD may form a complexwithout releasing the phthaloyl-CoA intermediate. However, aseries of assays with SPT and PCD, separated by dialysismembranes, ruled out a necessary direct interaction of bothenzymes to efficiently form benzoyl-CoA, CO2 and CoA fromphthalate and succinyl-CoA (Mergelsberg et al., 2018). Instead,PCD was found as the most abundant enzyme in phthalate-grown cells of denitrifiers with an estimated cellular concentra-tion of around 140 μM per active site-containing PCD monomer(10%–15% of soluble protein). Thus, the concentration of PCDis around 250-fold higher than that of its substrate phthaloyl-CoA which guarantees an efficient capture and decarboxylationof the substrate (Mergelsberg et al., 2018).
To avoid phthaloyl-CoA accumulation and its subsequenthydrolysis balanced SPT and PCD activities are mandatory.In vitro titration experiments with purified SPT and PCD indi-cated that the optimal PCD:SPT ratio for the conversion ofphthalate + succinyl-CoA to benzoyl-CoA + CO2 + succinatewas 4:1 (normalized to active site subunits). Remarkably,the relative cellular abundance of both enzymes was almostin the same ratio (Mergelsberg et al., 2018). Thus, highly bal-anced SPT and PCD production levels are essential to mini-mize phthaloyl-CoA hydrolysis. Even under these optimizedconditions found in T. chlorobenzoica cells, still around 20%of the succinyl-CoA added to the coupled SPT/PCD assayswas hydrolyzed rather than converted to benzoyl-CoA, indi-cating that phthalate degradation via phthaloyl-CoA repre-sents a poorly optimized xenobiotic degradation pathway.
Degradation of phthalate in sulfate-reducing bacteria. Untilrecently, knowledge of anaerobic degradation of PA in obli-gately anaerobic bacteria was restricted to a few studies withsediment slurries and enrichment cultures (Kleerebezem et al.,1999a; Kleerebezem et al., 1999b; Liu and Chi, 2003; Liuet al., 2005a; Liu et al., 2005b). Further, a Pelotomaculum iso-pthalicum species has been described that grows with all threephthalate isomers in syntrophic association with a methanogen(Qiu et al., 2006). In a recent study, the previously overseencapacity of the sulfate-reducing, benzoate-degrading Des-ulfosarcina cetonica to use phthalate as carbon and togetherwith sulfate as energy source was reported (Geiger et al., 2019).A differential proteome analysis identified a gene cluster that wasinduced in D. cetonica cells grown with phthalate versus benzo-ate. In this cluster a number of genes putatively encoding aTRAP transporter, a periplasmic binding protein, an UbiD-likePCD, and the corresponding UbiX-like prenyltransferase werefound, indicating significant similarities to the gene productsinvolved in phthalate degradation in denitrifying bacteria (Fig. 3).However, a gene encoding a CoA transferase was missing,instead a gene encoding a putative ATP-dependent CoA ligasewas identified. In vitro enzyme activity assays confirmed thatindeed phthaloyl-CoA is formed by an ATP-dependent phthalateCoA ligase. Similar as in denitrifying bacteria, phthaloyl-CoAaccumulated only to submicromolar concentrations as a result ofits rapid decay by intramolecular hydrolysis. In the coupledassays with enriched PCD, the ready formation of benzoyl-CoAfromATP, CoA andPAwas demonstrated (Geiger et al., 2019).
The rationale for using a phthalate CoA ligase instead of aless energy consuming SPT in sulfate-reducing bacteriacould be that they oxidize fatty acids via the Wood-Ljungdahlpathway rather than via the Krebs cycle (Schauder et al.,1986; Janssen and Schink, 1995). As a consequence,succinyl-CoA is not available to significant amounts to serveas CoA donor for phthalate activation. Indeed, the cellularsuccinyl-CoA concentrations in D. cetonica grown withphthalate are several orders of magnitude lower than in den-itrifying bacteria (Geiger et al., 2019).
A phthalate degradation gene cluster highly similar to thatof D. cetonica was identified in Desulfobacula toluolica, asulfate-reducing δ-proteobacterium known to degrade sev-eral monocyclic aromatic compounds (Wöhlbrand et al.,2013), leaving little doubt that this organism has the capacityto growth with phthalate and sulfate. However, no completephthalate catabolic gene cluster was found in other knownaromatic compound degrading strictly anaerobic bacteria(Geiger et al., 2019). A number of putative δ-proteobacterialPCD encoding genes (amino acid sequence identity ≥66%)are present in the metagenomes of a shallow sediment-hosted perennially suboxic/oxic aquifer (Anantharamanet al., 2016), suggesting that phthalate degradation capacityis present in many so far unknown δ-proteobacteria.
Anaerobic degradation of isophthalate
Anaerobic degradation of isophthalate has been studiedonly with very few bacterial cultures. Syntrophorhabdusaromaticivorans degrades isophthalate and other aro-matic compounds in syntrophic association with a H2-consuming partner organism (Qiu et al., 2004; Qiu et al.,
© 2019 The Authors. Environmental Microbiology Reports published by Society for Applied Microbiology and John Wiley & Sons Ltd.,Environmental Microbiology Reports, 12, 3–15
10 M. Boll, R. Geiger, M. Junghare and B. Schink

2008). Further, a defined co-culture of strictly anaerobic,syntrophically isophthalate fermenting bacteria belongingto the genus Pelotomaculum (family Peptococcaceae),i.e. P. isophthalicum has been described (Qiu et al., 2006).A recent study of anaerobic isophthalate degradation inS. aromaticivorans provided experimental evidence of thegenes, key enzymes and intermediates involved in anaero-bic degradation of IPA (Junghare et al., 2019). Similar to PAdegradation by sulfate-reducing bacteria (Junghare et al.,2016; Ebenau-Jehle et al., 2017), S. aromaticivorans acti-vates isophthalate by an ATP-dependent isophthalate:CoAligase enzyme to isophthaloyl-CoA which is subsequentlydecarboxylated to benzoyl-CoA (Junghare et al., 2019).Decarboxylation of isophthaloyl-CoA to benzoyl-CoA iscatalysed by the UbiD enzyme family member isophthaloyl-CoA decarboxylase that requires an UbiX-like flavinprenyltransferase to generate the active site prFMN cofactor(Junghare et al., 2019). Thus, anaerobic isophthalate degra-dation appears to employ a highly similar enzyme inventoryas anaerobic phthalate degradation (Junghare et al., 2016;Ebenau-Jehle et al., 2017). A major difference is thatisophthaloyl-CoA is not prone to intramolecular hydrolysissuch as phthaloyl-CoA, and has a half-life time of severalhours (Junghare et al., 2019).
Anaerobic degradation of terephthalate
In comparison to the anaerobic degradation of PA andIPA, much less is known about the genes and enzymesinvolved in anaerobic TPA degradation. It has beenreported to occur in syntrophic methanogenic consortiain bioreactors (Qiu et al., 2004; Lykidis et al., 2011), andPelotomaculum species have been described that werecapable of degrading TPA in syntrophic co-culture with amethanogen (Qiu et al., 2006). Further, ubiD-like geneshave been identified as potentially encoding a decarbox-ylase involved in TPA degradation, either directly or afteractivation to terephthaloyl-CoA (Lykidis et al., 2011,Nobu et al., 2015). However, any experimental evidencefor the involvement of the proposed gene products inanaerobic TPA is missing. It is generally difficult, if notimpossible to make any prediction to deduce a specificPA, IPA or TPA degradation potential purely based onthe presence of ubiD-like genes. As the reactivity of theortho- or para-positioned carboxyl-functionalities in PA andTPA should be equivalent, there remains little doubt thatanaerobic TPA degradation proceeds via activation toterephthaloyl-CoA followed by decarboxylation. As thepredicted terephthaloyl-CoA intermediate is muchmore sta-ble than phthaloyl-CoA, the necessity of highly balancedterephthaloyl-CoA formation/decarboxylation activitiesappears to be lessmandatory.
Concluding remarks
The predominantly xenobiotic esters of the three PA iso-mers play a globally important role as plasticizers andpolymer constituents. Though massively produced onlysince around 60 years, a remarkable variety of degrada-tion strategies have evolved in a relatively short time span.Degradation pathways are usually initiated by hydrolysis tothe individual PA isomers plus the corresponding alcohols,followed by decarboxylation of one of the two carboxylicacid functionalities. The decarboxylation of non-activatedphthalic acids via negatively charged intermediates ismechanistically difficult. For this reason, two aerobic andone anaerobic strategy have evolved to accomplish decar-boxylation: (i) during aerobic degradation of IPA and TPA, adioxygenase introduces hydroxyl groups directly at a car-boxyl functionality yielding cyclic cis-diol-dienes that candirectly be decarboxylated during the rearomatizing dehy-drogenation reaction; (ii) aerobic PA degradation proceedsvia dihydroxyphthalates with phenolic functionalities inortho- or para-position to a carboxylic group which greatlyfacilitates their decarboxylation; (iii) anaerobic degradationof most likely all three phthalic acid isomers proceeds viaCoA thioesterification followed by decarboxylation tobenzoyl-CoA by UbiD-like decarboxylases. The mechanismof how this activation promotes the subsequent decarboxyl-ation is unknown, and there is a high research demand toelucidate the structure and function of UbiD-like decar-boxylases involved in the degradation of all three PA iso-mers. The mechanism of phthaloyl-CoA and terephthaloyl-CoA decarboxylase should be similar but markedly differfrom that of isophthaloyl-CoA decarboxylase due to the dif-fering reactivities of the carboxyl functionalities.
The phthaloyl-CoA intermediate during anaerobic PAdegradation is possibly the most unstable CoA thioesterin biology, which demands additional adaptations to cir-cumvent its immediate futile hydrolysis. The solution tooverproduce PCD up to 140 μM is accompanied by ahigh energetic burden for its synthesis. Taken intoaccount that even under optimized conditions a signifi-cant phthaloyl-CoA hydrolysis of up to 20% occurred,anaerobic phthalate degradation via phthaloyl-CoA is anon-optimized, rather wasteful strategy. PA representsan intermediate during the aerobic degradation of natu-rally occurring polycyclic aromatic compounds. Thus, itscomplete aerobic degradation pathways cannot beassigned to recent evolutionary inventions. However, PAis not known as an intermediate of an anaerobic degra-dation pathway. In conclusion, the current anaerobicsolution may indeed represent an early stage of theongoing evolution of a degradation pathway for an essen-tially xenobiotic compound. Future evolution of the anaer-obic pathway may result in complex formation betweenSPT and PCD that would allow a direct transfer of
© 2019 The Authors. Environmental Microbiology Reports published by Society for Applied Microbiology and John Wiley & Sons Ltd.,Environmental Microbiology Reports, 12, 3–15
Anaerobic degradation of phthalates 11

phthaloyl-CoA between the individual active sites of itssynthesizing and decarboxylating enzymes; such a strat-egy is observed for highly reactive and/or toxic intermedi-ates in nature (Bernhardsgrütter et al., 2018).
Acknowledgements
The Konstanz research group is grateful to Dr. Dieter Spitellerfor expert help in analytical chemistry and chemical synthesisof phthtaloyl-CoA isomers. This work was financially supportedby the German research council (BO 1565/16-1), the DeutscheAkademische Austauschdienst (DAAD) and funds by the Uni-versity of Konstanz.
References
Aftring, R.P., Chalker, B.E., and Taylor, B.F. (1981) Degra-dation of phthalic acids by denitrifying, mixed cultures ofbacteria. Appl Environ Microbiol 41: 1177–1183.
Anantharaman, K., Brown, C.T., Hug, L.A., Sharon, I.,Castelle, C.J., Probst, A.J., et al. (2016) Thousands ofmicrobial genomes shed light on interconnected biogeo-chemical processes in an aquifer system. Nat Commun 7:13219.
Andrady, A.L., and Neal, M.A. (2009) Applications and socie-tal benefits of plastics. Philos Trans R Soc Lond B BiolSci 364: 1977–1984.
Bains, J., Wulff, J.E., and Boulanger, M.J. (2012) Investigat-ing terephthalate biodegradation: structural characteriza-tion of a putative decarboxylating cis-dihydrodioldehydrogenase. J Mol Biol 423: 284–293.
Batie, C.J., LaHaie, E., and Ballou, D.P. (1987) Purificationand characterization of phthalate oxygenase and phthal-ate oxygenase reductase from Pseudomonas cepacia.J Biol Chem 262: 1510–1518.
Benjamin, S., Pradeep, S., Josh, M.S., Kumar, S., andMasai, E. (2015) A monograph on the remediation of haz-ardous phthalates. J Hazard Mater 298: 58–72.
Bernhardsgrütter, I., Vögeli, B., Wagner, T., Peter, D.M.,Cortina, N.C., Kahnt, J., et al. (2018) The multicatalyticcompartment of propionyl-CoA synthase sequesters atoxic metabolite. Nat Chem Biol 14: 1127–1132.
Boll, M., Löffler, C., Morris, B.E.L., and Kung, J.W. (2014)Anaerobic degradation of homocyclic aromatic compoundsvia arylcarboxyl-coenzyme A esters: organisms, strategiesand key enzymes. Environ Microbiol 16: 612–627.
Boonyaroj, V., Chiemchaisri, C., Chiemchaisri, W., andYamamoto, K. (2012) Removal of organic micro-pollutantsfrom solid waste landfill leachate in membrane bioreactoroperated without excess sludge discharge. Water SciTechnol 66: 1774–1780.
Camacho-Munoz, G.A., Llanos, C.H., Berger, P.A., Miosso, C.J., and da Rocha, A.F. (2012) Occurrence of pharmaceuticalcompounds in wastewater and sludge from wastewatertreatment plants: removal and ecotoxicological impact ofwastewater discharges and sludge disposal. Conf Proc IEEEEng Med Biol Soc 2012: 6508–6513.
Carbone, V., Hara, A., and El-Kabbani, O. (2008) Structuraland functional features of dimeric dihydrodiol dehydroge-nase. Cell Mol Life Sci 65: 1464–1474.
Carmona, M., Zamarro, M.T., Blázquez, B., Durante-Rodríguez, G., Juárez, J.F., Valderrama, J.A., et al. (2009)Anaerobic catabolism of aromatic compounds: a geneticand genomic view. Microbiol Mol Biol Rev 73: 71–133.
Carrara, S.M., Morita, D.M., and Boscov, M.E. (2011) Bio-degradation of di(2-ethylhexyl)phthalate in a tropical soil.J Hazard Mater 197: 40–48.
Chang, H.K., and Zylstra, G.J. (1998) Novel organization ofthe genes for phthalate degradation from Burkholderiacepacia DBO1. J Bacteriol 180: 6529–6537.
Chang, B.V., Liao, C.S., and Yuan, S.Y. (2005) Anaerobicdegradation of diethyl phthalate, di-n-butyl phthalate, anddi-(2-ethylhexyl) phthalate from river sediment in Taiwan.Chemosphere 58: 1601–1607.
Chang, H.K., Dennis, J.J., and Zylstra, G.J. (2009) Involve-ment of two transport systems and a specific porin in theuptake of phthalate by Burkholderia spp. J Bacteriol 191:4671–4673.
Chao, W.L., Lin, C.M., Shiung, I.I., and Kuo, Y.L. (2006)Degradation of di-butyl-phthalate by soil bacteria. Chemo-sphere 63: 1377–1383.
Choi, K.Y., Kim, D., Sul, W.J., Chae, J.C., Zylstra, G.J., Kim, Y.M., and Kim, E. (2005) Molecular and biochemical analysisof phthalate and terephthalate degradation by Rhodococcussp. strain DK17. FEMS Microbiol Lett 252: 207–213.
Correll, C.C., Batie, C.J., Ballou, D.P., and Ludwig, M.L.(1992) Phthalate dioxygenase reductase: a modular struc-ture for electron transfer from pyridine nucleotides to [2Fe-2S]. Science 258: 1604–1610.
Cox, G.B., Young, I.G., McCann, L.M., and Gibson, F. (1969)Biosynthesis of ubiquinone in Escherichia coli K-12: loca-tion of genes affecting the metabolism of 3-octaprenyl-4-hydroxybenzoic acid and 2-octaprenylphenol. J Bacteriol99: 450–458.
Dagley, S. (1971) Catabolism of aromatic compounds bymicroorganisms. Adv Microb Physiol 6: 1–46.
Eaton, R.W., and Ribbons, D.W. (1982) Metabolism of dibu-tylphthalate and phthalate by Micrococcus sp. strain 12B.J Bacteriol 151: 48–57.
Ebenau-Jehle, C., Mergelsberg, M., Fischer, S., Bruls, T.,Jehmlich, N., von Bergen, M., and Boll, M. (2017) Anunusual strategy for the anoxic biodegradation of phthal-ate. ISME J 11: 224–236.
Forward, J.A., Behrendt, M.C., Wyborn, N.R., Cross, R., andKelly, D.J. (1997) TRAP transporters: a new family of peri-plasmic solute transport systems encoded by the dctPQMgenes of Rhodobacter capsulatus and by homologs indiverse gram-negative bacteria. J Bacteriol 179: 5482–5493.
Fuchs, G., Boll, M., and Heider, J. (2011) Microbial degrada-tion of aromatic compounds - from one strategy to four.Nat Rev Microbiol 9: 803–816.
Fukuhara, Y., Kasai, D., Katayama, Y., Fukuda, M., andMasai, E. (2008) Enzymatic properties of terephthalate1,2-dioxygenase of Comamonas sp. strain E6. Biosci Bio-technol Biochem 72: 2335–2341.
Fushiwaki, Y., Niino, T., Ishibashi, T., Takeda, K., andOnodera, S. (2003) Tumor-promoting activity of phthalateesters estimated by in vitro transformation using Bhascells. J Health Sci 49: 82–87.
Gao, D.-W., and Wen, Z.-D. (2016) Phthalate esters in theenvironment: a critical review of their occurrence,
© 2019 The Authors. Environmental Microbiology Reports published by Society for Applied Microbiology and John Wiley & Sons Ltd.,Environmental Microbiology Reports, 12, 3–15
12 M. Boll, R. Geiger, M. Junghare and B. Schink

biodegradation, and removal during wastewater treatmentprocesses. Sci Total Environ 541: 986–1001.
Geiger, R.A., Junghare, M., Mergelsberg, M., Ebenau-Jehle, C., Jesenofsky, V.J., Jehmlich, N., et al. (2019)Enzymes involved in phthalate degradation in sulphate-reducing bacteria. Environ Microbiol.. https://doi.org/10.1111/1462-2920.14681.
Geyer, R., Jambeck, J.R., and Law, K.L. (2017) Production,use, and fate of all plastics ever made. Sci Adv 3:e1700782.
Giam, C.S., Chan, H.S., Nett, G.S., and Atlas, E.L. (1978)Phthalate ester plasticizers: a new class of marine pollut-ant. Science 199: 419–421.
Giam, C.S., Atlas, E., Powers, M.A., and Leonard, J.E.(1984) Phthalic acid esters. In The Handbook of Environ-mental Chemistry – Anthropogenic Compounds, Vol.3 (C). Hutzinger, b.O. (ed). Berlin: Springer, pp. 67–142.
Gledhill, W.E., Kaley, R.G., Adams, W.J., Hicks, O.,Michael, P.R., Saeger, V.W., and Leblanc, G.A. (1980) Anenvironmental safety assessment of butyl benzyl phthal-ate. Environ Sci Technol 14: 301–305.
Hahladakis, J.N., Velis, C.A., Weber, R., Iacovidou, E., andPurnell, P. (2018) An overview of chemical additives pre-sent in plastics: migration, release, fate and environmentalimpact during their use, disposal and recycling. J HazardMater 344: 179–199.
Hara, H., Stewart, G.R., and Mohn, W.W. (2010) Involve-ment of a novel ABC transporter and monoalkyl phthalateester hydrolase in phthalate ester catabolism byRhodococcus jostii RHA1. Appl Environ Microbiol 76:1516–1523.
Hazer, B., and Steinbüchel, A. (2007) Increased diversifica-tion of polyhydroxyalkanoates by modification reactionsfor industrial and medical applications. Appl Microbiol Bio-technol 74: 1–12.
Hosaka, M., Kamimura, N., Toribami, S., Mori, K., Kasai, D.,Fukuda,M., andMasai, E. (2013)Novel tripartite aromatic acidtransporter essential for terephthalate uptake in Comamonassp. strain E6.Appl EnvironMicrobiol 79: 6148–6155.
Huwiler, S.G., Löffler, C., Anselmann, S.E.L., Stark, H.J.,von Bergen, M., Flechsler, J., et al. (2019) One-megadalton metalloenzyme complex in Geobacter meta-llireducens involved in benzene ring reduction beyond thebiological redox window. Proc Natl Acad Sci 116:2259–2264.
Janssen, P.H., and Schink, B. (1995) Pathway of butyratecatabolism by Desulfobacterium cetonicum. J Bacteriol177: 387–3872.
Jendrossek, D., and Handrick, R. (2002) Microbial degrada-tion of polyhydroxyalkanoates. Annu Rev Microbiol 56:403–432.
Jonsson, S., Vavilin, V.A., and Svensson, B.H. (2006)Phthalate hydrolysis under landfill conditions. Water SciTechnol 53: 119–127.
Junghare, M., Patil, Y., and Schink, B. (2015) Draft genomesequence of a nitrate-reducing, o-phthalate degradingbacterium, Azoarcus sp. strain PA01(T). Standards ingenomic sciences 10: 90.
Junghare, M., Spiteller, D., and Schink, B. (2016) Enzymesinvolved in the anaerobic degradation of ortho-phthalate
by the nitrate-reducing bacterium Azoarcus sp. strainPA01. Environ Microbiol 18: 3175–3188.
Junghare, M., Spiteller, D., and Schink, B. (2019) Anaerobicdegradation of xenobiotic isophthalate by the fermentingSyntrophorhabdus aromaticivorans. ISME J 13: 1252–1268.
Karandikar, R., Badri, A., and Phale, P.S. (2015) Biochemi-cal characterization of inducible ’Reductase’ componentof benzoate dioxygenase and phthalate isomerdioxygenases from Pseudomonas aeruginosa strain PP4.Appl Biochem Biotechnol 177: 318–333.
Kasai, D., Iwasaki, T., Nagai, K., Araki, N., Nishi, T., andFukuda, M. (2019) 2,3-Dihydroxybenzoate meta-cleavagepathway is involved in o-phthalate utilization in Pseudo-monas sp. strain PTH10. Sci Rep 9: 1253.
Kawai, F., Kawabata, T., and Oda, M. (2019) Current knowl-edge on enzymatic PET degradation andits possible appli-cation to waste stream management and other fields.Appl Microbiol Biotechnol 103: 4252–4268.
Keyser, P., Pujar, B.G., Eaton, R.W., and Ribbons, D.W.(1976) Biodegradation of the phthalates and their estersby bacteria. Environ Health Perspect 18: 159–166.
Kleerebezem, R., Pol, L.W., and Lettinga, G. (1999a) Anaer-obic biodegradability of phthalic acid isomers and relatedcompounds. Biodegradation 10: 63–73.
Kleerebezem, R., Hulshoff Pol, L.W., and Lettinga, G.(1999b) Anaerobic degradation of phthalate isomers bymethanogenic consortia. Appl Environ Microbiol 65:1152–1160.
Kumar, P. (2018) Role of plastics on human health. Indian JPediatr 85: 384–389.
Lambrot, R., Muczynski, V., Lécureuil, C., Angenard, G.,Coffigny, H., Pairault, C., et al. (2009) Phthalates impairgerm cell development in the human fetal testis in vitrowithout change in testosterone production. Environ HealthPerspect 117: 32–37.
Latini, G., Del Vecchio, A., Massaro, M., Verrotti, A., and DeFelice, C. (2006) Phthalate exposure and male infertility.Toxicology 226: 90–98.
Lau, T.K., Chu, W., and Graham, N. (2005) The degradation ofendocrine disruptor di-n-butyl phthalate by UV radiation: aphotolysis and product study. Chemosphere 60: 1045–1053.
Leys, D. (2018) Flavin metamorphosis: cofactor transforma-tion through prenylation. Curr Opin Chem Biol 47:117–125.
Li, J., Chen, J.A., Zhao, Q., Li, X., and Shu, W. (2006) Biore-mediation of environmental endocrine disruptor di-n-butylphthalate ester by Rhodococcus ruber. Chemosphere 65:1627–1633.
Liang, D.W., Zhang, T., Fang, H.H., and He, J. (2008)Phthalates biodegradation in the environment. ApplMicrobiol Biotechnol 80: 183–198.
Liu, S.-M., and Chi, W.-C. (2003) CO2–H2-dependent anaer-obic biotransformation of phthalic acid isomers in sedi-ment slurries. Chemosphere 52: 951–958.
Liu, S.M., Lin, Y.L., and Chi, W.C. (2005a) The effects ofbenzoate, cyclohex-1-ene carboxylate, and cyclohexanecarboxylate on biotransformation of o-phthalate in sedi-ment slurries under sulfate-reducing conditions. Chemo-sphere 59: 41–48.
© 2019 The Authors. Environmental Microbiology Reports published by Society for Applied Microbiology and John Wiley & Sons Ltd.,Environmental Microbiology Reports, 12, 3–15
Anaerobic degradation of phthalates 13

Liu, S.M., Lin, Y.L., and Tsai, T.L. (2005b) Growth dynamicsof major microbial populations during biodegradation of o-phthalate in anaerobic sediment slurries under a CO2/H2
atmosphere. Chemosphere 59: 91–98.Lykidis, A., Chen, C.L., Tringe, S.G., McHardy, A.C.,
Copeland, A., Kyrpides, N.C., et al. (2011) Multiplesyntrophic interactions in a terephthalate-degradingmethanogenic consortium. ISME J. 5: 122–130.
Maruyama, K., Akita, K., Naitou, C., Yoshida, M., andKitamura, T. (2005) Purification and characterization of anesterase hydrolyzing monoalkyl phthalates from Micro-coccus sp. YGJ1. J Biochem 137: 27–32.
Mayer, F.L., Stalling, D.L., and Johnson, J.L. (1972) Phthalateesters as environmental contaminants. Nature 238: 411–413.
Meng, X.Z., Wang, Y., Xiang, N., Chen, L., Liu, Z., Wu, B.,et al. (2014) Flow of sewage sludge-borne phthalateesters (PAEs) from human release to human intake: impli-cation for risk assessment of sludge applied to soil. SciTotal Environ 476–477: 242–249.
Mergelsberg, M., Willistein, M., Meyer, H., Stark, H.J.,Bechtel, D.F., Pierik, A.J., and Boll, M. (2017) Phthaloyl-coenzyme a decarboxylase from Thauera chlorobenzoica:the prenylated flavin-, K(+) - and Fe(2+) -dependent keyenzyme of anaerobic phthalate degradation. EnvironMicrobiol 19: 3734–3744.
Mergelsberg, M., Egle, V., and Boll, M. (2018) Evolution of axenobiotic degradation pathway: formation and capture ofthe labile phthaloyl-CoA intermediate during anaerobicphthalate degradation. Mol Microbiol 108: 614–626.
Mulligan, C., Fischer, M., and Thomas, G.H. (2011) TripartiteATP-independent periplasmic (TRAP) transporters in bac-teria and archaea. FEMS Microbiol Rev 35: 68–86.
Nakazawa, T., and Hayashi, E. (1978) Phthalate and4-hydroxyphthalatemetabolism inPseudomonas testosteroni:purification and properties of 4,5-dihydroxyphthalate decar-boxylase.Appl EnvironMicrobiol 36: 264–269.
Net, S., Delmont, A., Sempéré, R., Paluselli, A., andOuddane, B. (2015) Occurrence, fate, behavior and eco-toxicological state of phthalates in different environmentalmatrices. Environ Sci Technol 49: 4019–4035.
Niazi, J.H., Prasad, D.T., and Karegoudar, T.B. (2001) Initialdegradation of dimethylphthalate by esterases from Bacil-lus species. FEMS Micobiol Lett 196: 201–205.
Nilsson, C. (1994) Phthalate Esters Used as Plastic Addi-tives – Comparisons of Toxicological Effects. SwedishNational Chemicals Inspectorate: Brussels.
Nobu, M.K., Narihiro, T., Hideyuki, T., Qiu, Y.L., Sekiguchi,Y., Woyke, T., et al. (2015) The genome of Syn-trophorhabdus aromaticivorans strain UI provides newinsights for syntrophic aromatic compound metabolismand electron flow. Environ Microbiol 17: 4861–4872.
Nozawa, T., and Maruyama, Y. (1988) Anaerobic metabo-lism of phthalate and other aromatic compounds by a den-itrifying bacterium. J Bacteriol 170: 5778–5784.
Payne, K.A.P., White, M.D., Fisher, K., Khara, B., Bailey, S.S., Parker, D., et al. (2015) New cofactor supportsα,β-unsaturated acid decarboxylation via 1,3-dipolar cyclo-addition. Nature 522: 497–501.
Peijenburg, W.J., and Struijs, J. (2006) Occurrence of phthal-ate esters in the environment of the Netherlands.Exotoxicol Environ Saf 63: 204–215.
Philipp, B., and Schink, B. (2012) Different strategies inanaerobic biodegradation of aromatic compounds: nitratereducers versus strict anaerobes. Environ Microbiol Rep4: 469–478.
Pujar, B.G., and Ribbons, D.W. (1985) Phthalate metabolismin Pseudomonas fluorescens PHK: purification and prop-erties of 4,5-dihydroxyphthalate decarboxylase. Appl Envi-ron Microbiol 49: 374–376.
Qiu, Y.-L., Sekiguchi, Y., Imachi, H., Kamagata, Y., Tseng, I.C., Cheng, S.S., et al. (2004) Identification and isolation ofanaerobic, syntrophic phthalate isomer-degradingmicrobes from methanogenic sludges treating wastewaterfrom terephthalate manufacturing. Appl Environ Microbiol70: 1617–1626.
Qiu, Y.-L., Sekiguchi, Y., Hanada, S., Imachi, H., Tseng, I.-C.,Cheng, S.-S., et al. (2006) Pelotomaculum terephthalicumsp. nov. and Pelotomaculum isophthalicum sp. nov.: twoanaerobic bacteria that degrade phthalate isomers insyntrophic association with hydrogenotrophic methanogens.Arch Microbiol 185: 172–182.
Qiu, Y.-L., Hanada, S., Ohashi, A., Harada, H., Kamagata, Y.,and Sekiguchi, Y. (2008) Syntrophorhabdus aromaticivoransgen. nov., sp. nov., the first cultured anaerobe capable ofdegrading phenol to acetate in obligate syntrophic associa-tions with a hydrogenotrophic methanogen. Appl EnvironMicrobiol 74: 2051–2058.
Rabus, R., Boll, M., Heider, J., Meckenstock, R.U.,Buckel, W., Einsle, O., et al. (2016) Anaerobic microbialdegradation of hydrocarbons: from enzymatic reactions tothe environment. J Mol Microbiol Biotechnol 26: 5–28.
Rabus, R., Wöhlbrand, L., Thies, D., Meyer, M., Reinhold-Hurek, B., and Kämpfer, P. (2019) Aromatoleum gen. nov.,a novel genus accommodating the phylogenetic lineageincluding Azoarcus evansii and related species, and pro-posal of Aromatoleum aromaticum sp. nov., Aromatoleumpetrolei sp. nov., Aromatoleum bremense sp. nov.,Aromatoleum toluolicum sp. nov. and Aromatoleum diolicumsp. nov. Int J Syst Evol Microbiol 69: 982–997.
Ribbons, D.W., and Evans, W.C. (1960) Oxidative metabo-lism of phthalic acid by soil pseudomonads. Biochem J76: 310–318.
Saller, E., Laue, H., Schläfli-Oppenberg, H.R., and Cook, A.M. (1995) Purification and some properties of (1R,2S)-1,2-dihydroxy-3,5-cyclohexadiene-1,4-dicarboxylate dehy-drogenase rom Comamonas testosteroni T-2. FEMSMicrobiol Lett 130: 97–102.
Schauder, R., Eikmanns, B., Thauer, R.K., Widdel, F., andFuchs, G. (1986) Acetate oxidation to CO2 in anaerobicbacteria via a novel pathway not involving reactions of thecitric acid cycle. Arch Microbiol 145: 162–172.
Schläfli, H.R., Weiss, M.A., Leisinger, T., and Cook, A.M.(1994) Terephthalate 1,2-dioxygenase system from Com-amonas testosteroni T-2: purification and some properties ofthe oxygenase component. J Bacteriol 176: 6644–6652.
Sheehan, R.J. (2005) Terephthalic acid, dimethyl ter-ephthalic acid, and isophthalic acid. In Ullmann’s Encyclo-pedia of Industrial Chemistry. Weinheim: Wiley, pp. 1–13.
Sin, J.-C., Lam, S.-M., Mohamed, A.R., and Lee, K.-T. (2012)Degrading endocrine disrupting chemicals from wastewaterby TiO2 photocatalysis: a review. Int J Photoenerg 2012:23. https://doi.org/10.1155/2012/185159.
© 2019 The Authors. Environmental Microbiology Reports published by Society for Applied Microbiology and John Wiley & Sons Ltd.,Environmental Microbiology Reports, 12, 3–15
14 M. Boll, R. Geiger, M. Junghare and B. Schink

Stanier, R.Y., and Ornston, L.N. (1973) The beta-ketoadipatepathway. Adv Microb Physiol 9: 89–151.
Staples, C.A., Peterson, D.R., Parkerton, T.F., andAdams, W.J. (1997) The environmental fate of phthalatesesters: a literature review. Chemosphere 35: 667–749.
Stingley, R.L., Brezna, B., Khan, A.A., and Cerniglia, C.E.(2004) Novel organization of genes in a phthalate degra-dation operon of Mycobacterium vanbaalenii PYR-1.Microbiology 150: 3749–3761.
Tarasev, M., and Ballou, D.P. (2005) Chemistry of the cata-lytic conversion of phthalate into its cis-dihydrodiol duringthe reaction of oxygen with the reduced form of phthalatedioxygenase. Biochemistry 44: 6197–6207.
Taylor, B.F., and Ribbons, D.W. (1983) Bacterial decarboxyl-ation of o-phthalic acids. Appl Environ Microbiol 46:1276–1281.
Thompson, R.C., Moore, C.J., vom Saal, F.S., and Swan, S.H. (2009) Plastics, the environment and human health:current consensus and future trends. Philos Trans R SocLond B Biol Sci 364: 2153–2166.
Tiedt, O., Fuchs, J., Eisenreich, W., and Boll, M. (2018) Acatalytically versatile benzoyl-CoA reductase, key enzymein the degradation of methyl- and halobenzoates in den-itrifying bacteria. J Biol Chem 293: 10264–10274.
Tokiwa, Y., Calabia, B.P., Ugwu, C.U., and Aiba, S. (2009)Biodegradability of plastics. Int J Mol Sci 10: 3722–3742.
Vamsee-Krishna, C., and Phale, P.S. (2008) Bacterial degra-dation of phthalate isomers and their esters. Indian JMicrobiol 48: 19–34.
Vamsee-Krishna, C., Mohan, Y., and Phale, P.S. (2006) Bio-degradation of phthalate isomers by Pseudomonasaeruginosa PP4, Pseudomonas sp. PPD and Acinetobacterlwoffii ISP4. Appl Microbiol Biotechnol 72: 1263–1269.
Vats, S., Singh, R.K., and Tyagi, P. (2013) Phthalates – apriority pollutant. Int J Adv Biol Res 3: 1–8.
Wei, R., and Zimmermann, W. (2017) Microbial enzymes forthe recycling of recalcitrant petroleum-based plastics: howfar are we? Microb Biotechnol 10: 1308–1322.
Weinert, T., Huwiler, S.G., Kung, J.W., Weidenweber, S.,Hellwig, P., Stärk, H.-J., et al. (2015) Structural basis ofenzymatic benzene ring reduction. Nat Chem Biol 11:586–591.
White, M.D., Payne, K.A.P., Fisher, K., Marshall, S.A.,Parker, D., Rattray, N.J.W., et al. (2015) UbiX is a flavinprenyltransferase required for bacterial ubiquinone biosyn-thesis. Nature 522: 502–506.
Wimpenny, J.W., and Coombs, J.P. (1983) Penetration ofoxygen into bacterial colonies. J Gen Microbiol 129:1239–1242.
Wöhlbrand, L., Jacob, J.H., Kube, M., Mussmann, M.,Jarling, R., Beck, A., et al. (2013) Complete genome,catabolic sub-proteomes and key-metabolites ofDesulfobacula toluolica Tol2, a marine, aromaticcompound-degrading, sulfate-reducing bacterium. Envi-ron Microbiol 15: 1334–1355.
Wu, X., Liang, R., Dai, Q., Jin, D., Wang, Y., and Chao, W.(2010) Complete degradation of di-n-octyl phthalate bybiochemical cooperation between Gordonia sp. strainJDC-2 and Arthrobacter sp. strain JDC-32 isolated fromactivated sludge. J Hazard Mater 176: 262–268.
Yoshida, S., Hiraga, K., Takehana, T., Taniguchi, I.,Yamaji, H., Maeda, Y., et al. (2016) A bacterium thatdegrades and assimilates poly(ethyleneterephthalate).Science 35: 1196–1199.
© 2019 The Authors. Environmental Microbiology Reports published by Society for Applied Microbiology and John Wiley & Sons Ltd.,Environmental Microbiology Reports, 12, 3–15
Anaerobic degradation of phthalates 15