methylphenidate to adolescent rats drives enduring changes of accumbal htr7 expression: implications...
TRANSCRIPT

Genes, Brain and Behavior (2009) 8: 356–368 # 2009 The AuthorsJournal compilation # 2009 Blackwell Publishing Ltd/International Behavioural and Neural Genetics Society
Methylphenidate to adolescent rats drives enduringchanges of accumbal Htr7 expression: implications forimpulsive behavior and neuronal morphology
D. Leo†,1, W. Adriani*,‡,1, C. Cavaliere§, G. Cirillo§,
E. M. Marco‡, E. Romano‡, U. di Porzio†,
M. Papa§, C. Perrone-Capano†,¶ and G. Laviola‡
†Institute of Genetics and Biophysics ‘A. Buzzati Traverso’, CNR,
Naples, ‡Department of Cell Biology & Neurosciences, Behavioral
Neuroscience Section, Istituto Superiore di Sanita, Rome, and§Faculty of Medicine, Second University of Naples and¶Department of Biological Sciences, ‘Federico II’ University of
Naples, Naples, Italy
*Corresponding author: W. Adriani, Department of Cell Biology &
Neurosciences, Behavioral Neuroscience Section, Istituto Supe-
riore di Sanita’, viale Regina Elena 299 – I-00161 Roma, Italy.
E-mail: [email protected]
Methylphenidate (MPH) administration to adolescent
rodents produces persistent region-specific changes in
brain reward circuits and alterations of reward-based
behavior. We show that these modifications include
a marked increment of serotonin (5-hydroxy-tryptamine)
receptor type 7 (Htr7) expression and synaptic contacts,
mainly in the nucleus accumbens, and a reduction of basal
behavioral impulsivity. We show that neural and behav-
ioral consequences are functionally related: administration
of a selective Htr7 antagonist fully counteracts the MPH-
reduced impulsive behavior and enhances impulsivity
when administered alone in naive rats. Agonist-induced
activation of endogenous Htr7 significantly increases
neurite length in striatal neuron primary cultures, thus
suggesting plastic remodeling of neuronal morphology.
The mixed Htr (1a/7) agonist, 8-OH-DPAT, reduces impul-
sive behavior in adolescent rats and in naive adults, whose
impulsivity is enhanced by the Htr7 antagonist. In sum-
mary, behavioral pharmacology experiments show that
Htr7 mediates self-control behavior, and brain primary
cultures experiments indicate that this receptor may be
involved in the underlying neural plasticity, through
changes in neuronal cytoarchitecture.
Keywords: Animal model, attention deficit/hyperactivitydisorder,delay intolerance, neuralplasticity, nucleus accumbens
Received 3 September 2008, revised 13 January 2009,accepted form publication 19 January 2009
Repeated exposure to psychostimulant drugs causes com-
plex molecular and structural modifications in discrete brain
regions of the mesocorticolimbic dopamine (DA) system, the
main pathway of the brain reward circuitry (Nestler 2005;
Robinson & Kolb 2004; Volkow & Li 2005). This structural
remodeling is similar to other forms of experience-dependent
plasticity, and it is thought to underlie neurobehavioral adap-
tive responses (Hyman et al. 2006; Kelley 2004). The impact
of pharmacological manipulation over DA systems is
enhanced during critical periods of postnatal development,
when this, and other systems, undergo key maturational
changes (Cao et al. 2007, Frantz et al. 2007).
Recently, we have investigated, in rats, the enduringconsequences of adolescent exposure to methylphenidate
(MPH), a psychostimulant drug commonly used for attention
deficit/hyperactivity disorder (ADHD). Such exposure elicited
a long-lasting striatal upregulation (Adriani et al. 2006) of the
kainate 2 subunit of the ionotropic glutamate receptor (Grik2,
also known as GluR6) and of the serotonin (5-hydroxy-
tryptamine, 5-HT) receptor type 7 (Htr7). The striatal complex
is crucial for regulating innate and acquired behavioral habits
and natural reinforcement (Robbins & Everitt 1996) through
its dorsal (i.e. caudate putamen, CPu) and ventral (i.e. nucleus
accumbens, Acb) components, respectively.
The Htr7 is a seven-transmembrane-domain receptor thatis positively coupled to adenylate cyclase (Hedlund & Sutcliffe
2004; Shen et al. 1993) and displays low homology (40%) to
other Gs-coupled 5-HT receptors (Raymond et al. 2001). The
Htr7, localized within the spinal cord (Dogrul & Seyrek 2006),
hypothalamus, hippocampus and (pre)frontal cortex (PFC)
(Neumaier et al. 2001; Ruat et al. 1993; Russo et al. 2005;
Sleight et al. 1995; Venero et al. 1997), has been associated
with several central processes, including learning and mem-
ory (Ballatz et al. 2007; Gasbarri et al. 2008; Hedlund &
Sutcliffe 2004; Meneses 2004; Perez-Garcia & Meneses
2005). Htr7 also modulates DA firing activity in the ventral
tegmental area and plays a prominent role in shaping neuronal
cytoarchitecture (van Kesteren & Spencer 2003; Kvachnina
et al. 2005; Mnie-Filiali et al. 2007). Its central nervous system
distribution together with a high affinity for psychoactive
drugs suggest that Htr7 could arise as a potential therapeutic
target in depression and schizophrenia (Dean et al. 2006; East
et al. 2002; Guscott et al. 2005; Hedlund & Sutcliffe 2007).
The involvement of this receptor in mood regulation and
stress reactivity has been confirmed extensively (Hedlund
et al. 2005; Le Corre et al. 1997; Mullins et al. 1999;
Wesolowska et al. 2006).
We have now explored the neural architecture of the brainreward circuitry in adult rats, following exposure to MPH1Both authors have contributed equally to this work.
356 doi: 10.1111/j.1601-183X.2009.00486.x

during adolescence. We observed a marked increase of Htr7,mainly in the Acb, associated with an increased number of
synaptic contacts. As subchronic MPH exposure in adoles-cent rats also reduces behavioral impulsivity (Adriani & Laviola
2004), an effect that persists into adulthood (Adriani et al.2007; Bolanos et al. 2003), we hypothesized that a functional
link might exist between accumbal Htr7 expression and self-control ability. Therefore, we assessed whether a selective
Htr7 antagonist was able to block the enduring reduction inimpulsivity, resulting from adolescent MPH exposure. We
decided to focus on ‘cognitive’ rather than ‘motor’ impulsivity(Evenden 1999), and therefore we adopted a previously
validated adaptation of delay-discounting task (Adriani &Laviola 2004; Evenden & Ryan 1999) instead of other
protocols, more based on premature and/or non-reinforcedresponding. We hypothesized that the Htr7 receptor could play
its key role in behavioral modulation by means of persistenteffects on neuronal architecture. Thus, we looked for morpho-
logical changes in cultured striatal neurons following adminis-tration of Htr7 agonist and antagonist drugs. Finally, we
investigated the putative role played by Htr1a and Htr7 in basaland pharmacologically modulated impulsive behavior.
Materials and methods
Animal subjects and treatment design
Animal experimental protocols were approved by institutional author-ities, on behalf of Ministry of Health, in close agreement withEuropean Community Directives and Italian Law. All efforts weremade to minimize animal suffering, to reduce the number of animalsused and to use alternatives to in vivo testing. For the first twoexperiments, pregnant Wistar female rats (Harlan, Italy) were housedin an air-conditioned room (temperature 21 � 18C, relative humidity60 � 10%), with a 12-h light/dark cycle (lights on at 0800 h). Waterand food (Enriched Standard Diet; Mucedola, Settimo Milanese, Italy)were available ad libitum. The day of delivery was considered aspostnatal day (PDN) zero, pups being culled to six males and twofemales. Even if only male subjects were used in the presentexperiments, culling included the female gender to avoid carryovereffects of rearing animals in sexual segregation. Pups were thenweaned on PND 21 and housed in groups of two siblings, according tosex. Within litters, each of the siblings was assigned to a different finalgroup. Two separate batches (of eight litters each) were used.
In a first experiment, the consequences of adolescent MPH (Ritalin,CIBA-GEIGY SpA, Milan, Italy) exposure were investigated. Withineach of the eight litters, three pups were assigned to administrationwith MPH (2 mg/kg/day), the other three siblings were assigned toinjection with saline (SAL) during adolescence (PND 30–44). Giventhat learning and memory have significant effects on neuroplasticity,we decided animals for the ex vivo analyses to have no experience atall between last treatment day and the killing. Thus, one SAL and oneMPH sibling per litter (n ¼ 8 rats per group) were killed, at least6 weeks after treatment (PND > 90), for ex vivo analyses. The otherfour animals (two SAL and two MPH sibling per litter, n ¼ 8 rats perfinal group) were tested, at least 3 weeks after treatment(PND > 70), for impulsive behavior.
The latter was measured using the intolerance-to-delay (ID) task. Atthe three highest delays, rats underwent the operant sessionsfollowing challenge with saline vehicle (SAL) or with the Htr7antagonist (R)-3-(2-(2-(4-methyl-piperidin-1-yl)ethyl)-pyrrolidine-1-sulfonyl)-phenol (SB269970, 3 mg/kg/day for 3 days, 15 min beforethe session; Tocris, Bristol, UK). This was to assess whether long-termMPH consequences could be blocked by a selective Htr7 antagonist.
A second experiment used a second batch of eight litters. Fourmale pups per litter were assigned to another experiment, the othertwo siblings per litter aimed to study the effects of ongoing treatment
witha mixed Htr (1a/7)agonist during theadolescent period (PND 30–44)on impulsivity, as assessed in the intolerance-to-delay task. At thethree highest delays, rats underwent the operant sessions followingchallenge with SAL or with the Htr (1a/7) agonist 8-hydroxy-2(di-n-propylamino)tetralin (8-OH-DPAT, 0.060 mg/kg for 3 days, 30 minbefore the session; Sigma-Aldrich, Milan, Italy). This was to assessif the ongoing effects of MPH, observed in adolescent rats (Adriani &Laviola 2004), were mimicked by a mixed agonist also targeting Htr7.
For the third experiment, Wistar male rats (Harlan) were purchasedwhen adult. At arrival (PND > 60), they were housed in pairs underthe same environmental conditions previously described. After atleast 1 week of acclimation (PND > 70), they underwent the intoler-ance-to-delay task following challenge with either SB269970, the Htr7selective antagonist (or SAL, 45 min before the session), or 8-OH-DPAT, the mixed Htr (1a/7) agonist (or SAL, 30 min before thesession), both at the three highest delays. These naive adult animalswere randomly assigned to one of the four pharmacological treatmentgroups. This experiment aimed to further investigate the role playedby Htr7 in the modulation of baseline impulsive behavior and toinvestigate the participation of Htr7 in the well-known modulatoryeffects of 8-OH-DPAT.
Behavioral impulsivity task
Apparatus and procedureAnimals were tested in an intolerance-to-delay protocol, involvinga choice between a smaller, immediate or a larger, delayed reward(Adriani & Laviola 2004; Evenden & Ryan 1999). Few days before theschedule started, animals were food-restricted to keep them at80–85% of their free-feeding weight, in order to increase theirmotivation to work for food delivery. Each animal was then placeddaily in a computer-controlled operant chamber (Coulbourn Instru-ments, Allentown, PA, USA), provided with two nose-poking holes,a chamber light, a feeder device, a magazine where pellets (45 mg,BioServ, Frenchtown, NJ, USA) were dropped and a magazine light.The nose poking in either hole was detected by a photocell and wasrecorded by a computer, which also controlled food delivery. After the25-min session, animals were returned to their home cage, wherethey were given standard chow (approximately 10 g/each).
During the training phase (1 week), nose poking in one of the twoholes resulted in the delivery of five pellets of food, whereas nosepoking in the other hole resulted in the delivery of one pellet of food.After nose poking and before food delivery, the chamber light wasturned on for 1 second. Following food delivery, the magazine lightwas turned on for 20 seconds, during which nose poking wasrecorded, but was without scheduled consequences (time-out).During the testing phase (1 week), a signaled delay was insertedbetween nose-poking and large-reward delivery. The chamber lightwas kept on during the entire length of this delay. The small rewarddelivery was unchanged. Hence, animals had choice betweena ‘LARGE & LATE’ (LL) or a ‘SMALL & SOON’ (SS) reward. The delaylength was kept fixed for each daily session and was changedprogressively over days. Animals received the corresponding drugchallenge(s) at the three longest delays of this task.
Design and data analysisData were analyzed by split-plot analysis of variance (ANOVA). Thegeneral design of the experiment had two-level drug factors (denotingpretreatment and/or acute challenge with agonist and/or antagonistdrug vs. vehicle) � 4-level delay (set for each session). For Exp. 1 and 2,the litter was always the blocking unit, in that the design comprised allwithin-litter factors. This approach is often used and even recom-mended in studies involving developmental treatments (Zorrilla 1997).In the third experiment, the blocking unit was the subject, and allfactors were between-subject, as adults were purchased as indepen-dent individuals. Multiple comparisons were then performed withTukey HSD when allowed.
The test phase consisted indeed of two halves: (1) the first foursessions, when delays were introduced but animals were allowed toface them without acute drug challenge and (2) the last four sessions,at the highest delays, including a baseline reference point plus the lastthree sessions, when the acute challenge was indeed administered.
Genes, Brain and Behavior (2009) 8: 356–368 357
Long-term consequences of methylphenidate during adolescence

ANOVAs to evaluate drugs’ effects were conducted on the second half.Separate analyses were performed within levels of a given factorwhen allowed by a main effect of that factor.
The first dependent variable considered was the choice (%) for thelarge reinforcer, namely percentage of LL over total LL þ SS choices,during the test phase. The second variable considered was the slopeof the preference–delay curve, calculated using MICROSOFT EXCEL
‘slope’ function, with the large-reward preference as y-axis data andlog (delay þ 1) as x-axis data (Evenden & Ryan 1999). Criteria forexclusion were: large-reward preference lower than 50% at the endof the training phase, or large-reward preference at highest delayhigher than that shown at delay ¼ 0. Accordingly, in Exp. 3, a total ofthree rats were excluded from data analyses.
Brain markers and morphology
The animals assigned to ex vivo analyses were killed at a point whenthe corresponding siblings had completed the behavioral testing.Three rats per group were randomly assigned to the brain dissectionand five rats per group were assigned to the brain perfusion.
Western blot analysisAfter killing, the brain was quickly dissected on dry þ wet ice. Thewhole-striatal complexes were collected bilaterally and used forWestern blot analysis. Brain striatal hemispheres were weighed,homogenized, split in two halves and pooled. To the purpose of usingsix final samples per group, we prepared six independent poolscoming from two contra-lateral hemispheres taken from two differentanimals out of the three. Striatal proteins were obtained using theTri-Reagent isolation system, following manufacturer’s instructions.Total protein concentration was determined by Bradford method(Bradford 1976). To generate Western blots, 25 mg of protein/lanewas loaded onto 10% SDS-polyacrylamide gels and resolved usinga mini-Protean II (Bio-Rad Laboratories) gel apparatus for 1 h at 100 V.A molecular weight standard was also included on the gel (Rainbowstandards; Bio-Rad Laboratories). Proteins were electrophoreticallytransferred onto polyvinylidene difluoride membranes (Immobilon-P,Millipore, Watford, UK) with a semi-dry transblotting apparatus. Themembranes were blocked in 5% (w/v) non-fat milk in Tris-bufferedsaline Tween-20 (TBST) (0.1% Tween, 150 mM NaCl, 10mM Tris-HCI,pH 7.5) and incubated with primary antibodies for 1 h at roomtemperature. After washing in TBST, membranes were incubatedfor 1 h at room temperature with HRP-conjugated secondary anti-bodies (dilution 1:1000; Bio-Rad) in TBST buffer containing 5% non-fatmilk. The blots were then washed three times for 15 min in Tris-buffered saline, and exposed to Kodak film using ECL chemilumines-cence (GE Health Care, Milan, Italy). Finally, the blots were strippedand probed with a b-actin antibody. The bands from the Western blotswere densitometrically quantified on X-ray films using a software(IMAGEQUANT) to determine the levels of immunoreactivity. Monoclonalmouse antibodies to tyrosine hydroxylase (TH) (#MAB318, 1:500;Chemicon, Milan, Italy), polyclonal rabbit antibodies to Grik2 (#06-315;1:200; Upstate Biotechnology Inc., Lake Placid, NY, USA), polyclonalrabbit antibodies to Htr7 (#PC249L, 1:500; Calbiochem, Milan, Italy),and monoclonal mouse b-actin antibodies (#A2668, 1:5000; Sigma-Aldrich, Milan, Italy) were used. Data from bands corresponding to themolecular weight of the detected protein were normalized to b-actin. Thedensity in the MPH group was then expressed as fold change over theSAL group, set as reference level at 100%.
Tissue preparationAfter killing, the brains of perfused animals were removed (fiveanimals per group). Rats were deeply anesthetized with a chloralhydrate (300 mg/kg i.p.) and perfused transcardially with a solution(Tris–HCl 0.1 M, ethylenediaminetetraacetic acid (EDTA) 10 mM) fol-lowed by 4% paraformaldehyde plus 0.1% glutaraldehyde in 0.01 M
phosphate-buffered saline (PBS), pH 7.4 at 48C. For light microscopy,the brain was removed and post-fixed 2 h in the same fixative, thensoaked in 30% sucrose PBS and frozen in chilled isopentane on dryice. Serial sections were cut on a cooled sliding microtome ata thickness of 25 mm and collected in cold PBS for immunocytochem-
istry. For electron microscopy, the brain was removed and placedovernight in 4% paraformaldehyde added to 1% glutaraldehyde in0.01 M PBS, pH 7.4 at 48C. Then, the tissue was rinsed in phosphatebuffer and post-fixed in 2% osmium tetroxide in phosphate buffer for20 min, dehydrated in ethanol and embedded in araldite resinembedding kit (TAAB Laboratories Equipment Ltd, AldermastonBerkshire, UK). Ultra-thin sections, then, were stained with uranylacetate and lead citrate and examined with an EFTEM Lybra 120electron microscope (Zeiss, Oberchoken, Germany).
ImmunocytochemistryBrain sections were blocked in 10% normal serum in 0.01 M PBS,0.25% Triton for 1 h at room temperature. Each primary antibody wasdiluted in 0.01 M PBS containing 10% normal serum 0.25% Triton. Free-floating slices were incubated for 48 h at 48C on an oscillating plate.Sections were washed several times in PBS and incubated with theappropriate biotinylated secondary antibody (Vector Labs Inc., Burlin-game, CA, USA; 1:200) for 90 min at room temperature, washed in PBSand processed using the Vectastain avidin–biotin peroxidase kit (VectorLabs Inc.) for 90 min, also at room temperature. The sections werewashed in 0.05 M Tris–HCl and reacted with 3,3-diaminobenzidinetetrahydrochloride (DAB; Sigma, Milan, Italy) in 0.5 mg/ml Tris–HCland 0.01% hydrogen peroxide. Sections were mounted on chrome-alume–gelatine-coated slides, dehydrated and coverslipped. Adjacentsections were Nissl-stained. Slides were imaged with a Zeiss Axio-skope 2 light microscope (Zeiss) equipped with high-resolution digitalcamera (C4742-95, Hamamatsu Photonics, Milan, Italy).
A densitometry of the several markers and morphometric featuresof cells in CPu, Acb and PFC was accomplished using computer-assisted image analysis system (MCID 7.0; Imaging Res. Inc.,Ontario, Canada). Sections of the CPu were analyzed within theinterval þ2.20 to �1.30 from bregma (Paxinos & Watson 2001). TheCPu was divided in two halves a dorsolateral (Dl-) and ventro-medial(Vm-) one. The Acb was outlined for each section and analyzedaccording to Nissl-stained adjacent sections and the calbindin-treatedsections of Zahm and Brog (1992). The core was divided from theshell throughout the posterior horn of the anterior commissure.Sections of the PFC were analyzed within the interval þ5.20 toþ3.20 from bregma (Paxinos & Watson 2001).
For Htr7 and TH expression, where labeling gave a diffuse appear-ance, we measured the mean density (density � area) within theselected regions. For Substantia P SubP and Grik2 expression, wherelabeling allowed discrete imaging of structures, we counted thenumber of positive cells elements, and therefore these two markerswere expressed as proportion of the total target area relative to thescanned area. Data were obtained from 10 randomly selectedsections for each region and then averaged for treated (MPH) andfor saline control (SAL) group.
Electron microscopyFor electron microscopy analysis, to estimate the synaptic density andactive zone length in the nucleus accumbens (Schikorski & Stevens1997), we choose a previously reported systematic sampling methodto reduce sampling bias (French & Trotterdel 2004). This wasachieved by collecting five sections per grid on three grids and then4 mm of the block was removed and the collection repeated; thisprocedure was repeated eight times. This technique allows to samplethroughout the depth of the sample. The middle section of the middlegrid in each collection of three grids was photographed at 42 000�magnification. In an alternative pattern, on one section we acquiredfour pictures proceeding from the top left corner to the bottom rightcorner; on the following section, we inverted direction of acquisitionfrom the top right corner to the bottom left one. Then synapticcomplexes were quantified. Synaptic complexes were defined asstructures that met two criteria: (1) restricted zone of two membranesin apposition, one or both exhibiting increased thickness or electroncontrast and (2) presence of vesicles, 30–50 nm in size, in one or bothof the two cytoplasmic domains separated by the opposing mem-branes. Digital images were analyzed using computer-assisted imageanalysis system (MCID 7.0; Imaging Res. Inc.).
All data collection was performed blind, with the observer being notaware of the treatment groups. These data were exported and
358 Genes, Brain and Behavior (2009) 8: 356–368
Leo et al.

converted to a frequency distribution histogram using the SIGMA-PLOT
v8.0 program (SPSS, Erkrath, Germany). Individual images of controland treated rats were assembled, and then the same adjustmentswere made for brightness, contrast and sharpness using ADOBE
PHOTOSHOP (Adobe Systems, San Jose, CA, USA).
Neuronal primary cultures
PreparationTimed pregnant Sprague–Dawley rats (Charles River, Italy) were killedin accordance with Society for Neuroscience guidelines. The embry-onic age (E) was determined by day of insemination (i.e. the ap-pearance of vaginal plug was considered as day E0). About 25–30embryos from different dams were pooled at E16 and were used forpreparation of four culture dishes.
The striatum (STR) was quickly dissected from embryos, undera stereoscope in sterile conditions, and placed in PBS withoutcalcium and magnesium and supplemented with 33 mM glucose.Cells were dissociated from embryonic STR and cultured as pre-viously described (di Porzio et al. 1980). Briefly, the dissectedE16 striata were enzymatically dissociated by incubation for30 min at 378C in a papain solution (Warthington, 20 U/mL) in Earle’sbalanced salts containing 1 mM EDTA, 1 mM cysteine and 0.01%pancreatic DNAse. After addition of 1 mg/mL of bovine serumalbumin (fraction V, Sigma-Aldrich) and 1 mg/mL ovomucoid(Sigma-Aldrich) the cell suspension was centrifuged 10 min at800 g, suspended in plating medium with a fire-polished Pasteurpipette and counted (Fiszman 1991).
Cells were plated at a density of 300 000/cm2 (Lab-Tek� ChamberSlides�, Nunc, Milan, Italy), in four dishes coated with 15 mg/mL ofpoly-D-lysine (Sigma). Cells were grown in neurobasal medium(Invitrogen, Milan, Italy), supplemented with B27 (Invitrogen). Cul-tures were maintained at 378C in a humidified incubator for 3 days.The cell cultures were treated with 100 nM of the Htr (1A/7) receptoragonist (8-OH-DPAT, Sigma-Aldrich, Milan, Italy), or with 100 nM ofthe Htr7 antagonist (SB269970, Tocris, Bristol, UK), or with a combi-nation of these two drugs. Agonists of D1 or D2 DA receptors (i.e.either deoxyepinephrine HCl or Quinpirole, respectively; Sigma) werealso administered. Drugs were added to cultures 48 h after cell platingand the drugs were left in culture medium for 24 h.
Measurement of neurite outgrowth and numberCell cultures were fixed for 30 min at room temperature, in 4%paraformaldehyde in PBS, followed by three washes in PBS, per-meabilized for 15 min in PBS containing 0.1% Triton-X-100 and 10%normal goat serum (NGS) and incubated for 2 h at room temperatureor overnight at 48C in the primary antibodies diluted in PBS containinga 10% of NGS. We used the monoclonal antibody (mAb) againstneuronal class III-tubulin (anti-Tuj1; 1:750, Chemicon). After incuba-tion with the primary antibody, cell cultures were rinsed in PBS andwere either stained according to standard avidin–biotin immunocyto-chemistry procedures (Vectastain Elite, Vector Labs Inc.) usingperoxidase substrate kit (dia-aminobenzidine) or incubated withfluorescent-labeled secondary antibodies (Texas-red goat anti-rabbit,Molecular Probes Inc., Eugene, OR, USA; goat anti-mouse fluores-cein-conjugated, Chemicon) in PBS containing a 5% solution of NGS.Control cells were incubated in the same solutions without primaryantibodies and subsequently processed as above.
After Tuj1 staining, cell-culture slides were analyzed by a ZeissAxioskop 20 XYZ motorized stage (Prior Proscan II) microscope usingthe software MCID Elite High Definition analog Camera HamamatsuC5405. The MCID Elite imaging system (Imaging Research Inc.,Linton, UK) was used for imagine acquisition and operator-controlledtracing of neuritis. A total of 12 fields for each cell-culture conditionwere selected from four independently treated culture wells, eachcontaining from 4–10 cells. The number, total and mean lengths ofbranches per neuron were measured.
Statistical analysesData were analyzed using the Student’s t-test for both immunohisto-chemical (Fig. 1a–d) and electron microscopy (Fig. 2) analyses, for
quantification of proteins in Western blot analyses (Fig. 1e) and formeasurement of neurite outgrowth (Fig. 6b,c). A difference wasaccepted as significant if the probability level was less than 5%(P < 0.05).
Results
Neuronal markers and cytoarchitecture in brain
reward pathways are altered by MPH
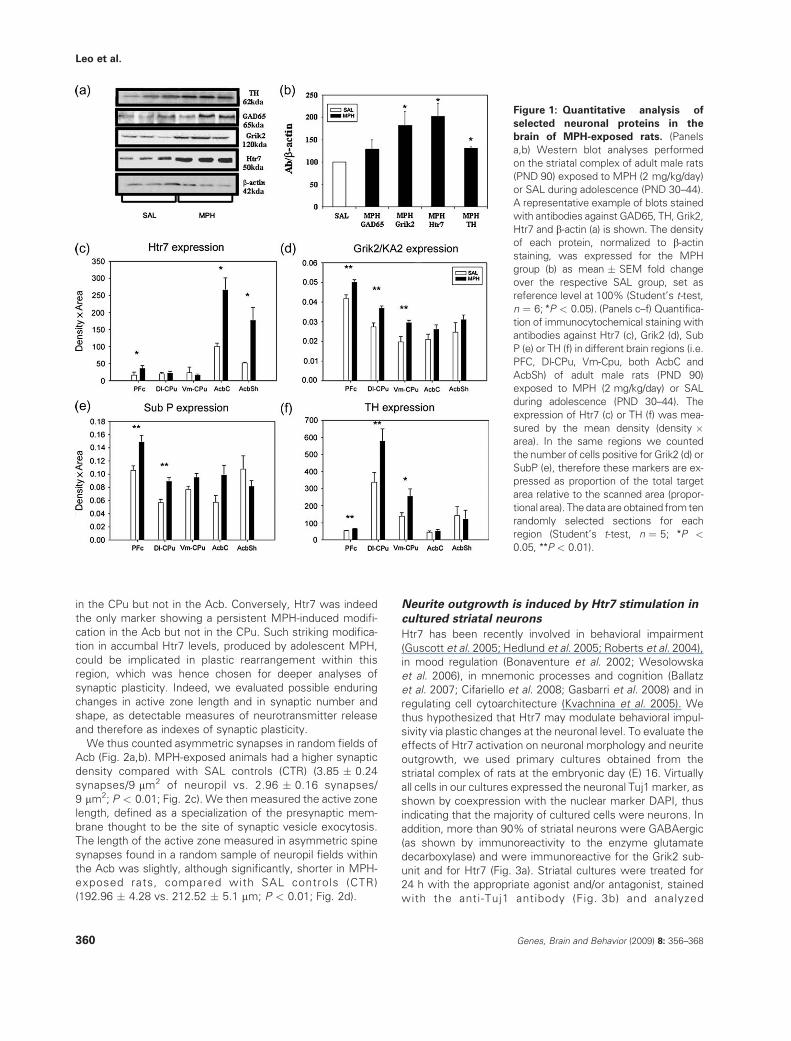
Rats, exposed to MPH (2 mg/kg/day) or saline (SAL) duringadolescence, at postnatal days (PND) 30–44, were used at
adulthood either for the behavioral studies (PND 70) or for theneurochemical and morphological analyses (PND 90). We first
examined, in samples coming from the whole-striatal com-plexes, protein levels of two neurotransmitter receptors,
Grik2 and Htr7, whose mRNA transcripts were found to bepersistently upregulated following adolescent MPH exposure
(Adriani et al. 2006). Western blot analyses showed that the
levels of both proteins were significantly higher in the whole-striatal complex of adult rats as a consequence of MPH
exposure during adolescence, when compared with SALcontrols (P < 0.05; Fig. 1a,b).
We also analyzed the protein levels of the glutamic aciddecarboxylase (GAD65) and the tyrosine hydoxylase (TH),
two key neuronal enzymes. GAD65 was investigated asa marker for g-aminobutyric acid (GABA)ergic neurons that
constitute the majority of striatal neurons. TH was studied asit labels catecholaminergic fibers, most of which are dopami-
nergic terminals of the mesocorticolimbic pathway (Graybiel1990). The protein levels of both enzymes were significantly
higher in the adult striatal complex as a consequence of MPHexposure during adolescence, when compared with SAL
controls (P < 0.05; Fig. 1a,b).To map the anatomical localization of the Grik2 and Htr7
increase in target forebrain areas of the mesocorticolimbicpathway, we performed immunohistochemical analyses.
Grik2 showed more intense staining in the dorsolateral (Dl-)and ventromedial (Vm-) Cpu and in the PFC of the MPH group,
compared with the SAL group. In both core and shell ofthe Acb (AcbC and AcbSh), the densitometric analysis for
Grik2 expression did not show differences between the twogroups (Fig. 1d). As for the Htr7 immunoblotting data, the
densitometric value in the Acb and in the PFC was signifi-cantly higher for MPH-exposed animals, compared with the
SAL group. In the CPu, however, the Htr7 staining wascomparable in both groups (Fig. 1c). We also analyzed by
immunohistochemistry the expression of Substantia P (SubP)and TH. SubP was investigated as it is expressed by a subset
of GABAergic neurons that constitute the direct striatal out-put pathway (Gerfen 1988). The SubP and the TH staining
were both significantly more intense in the Dl-CPu and in the
PFC of MPH-exposed rats than in the SAL group. In theVm-CPu and in both core and shell of Acb, SubP and TH
staining did not show any difference between the two groups(Fig. 1e,f).
In summary, we observed region-specific long-term con-sequences of an adolescent MPH exposure in selected
forebrain areas. All of the markers were increased in thePFC. Among these markers SubP, TH and Grik2 were altered
Genes, Brain and Behavior (2009) 8: 356–368 359
Long-term consequences of methylphenidate during adolescence
in the CPu but not in the Acb. Conversely, Htr7 was indeedthe only marker showing a persistent MPH-induced modifi-
cation in the Acb but not in the CPu. Such striking modifica-tion in accumbal Htr7 levels, produced by adolescent MPH,
could be implicated in plastic rearrangement within thisregion, which was hence chosen for deeper analyses of
synaptic plasticity. Indeed, we evaluated possible enduringchanges in active zone length and in synaptic number and
shape, as detectable measures of neurotransmitter releaseand therefore as indexes of synaptic plasticity.
We thus counted asymmetric synapses in random fields ofAcb (Fig. 2a,b). MPH-exposed animals had a higher synaptic
density compared with SAL controls (CTR) (3.85 � 0.24synapses/9 mm2 of neuropil vs. 2.96 � 0.16 synapses/
9 mm2; P < 0.01; Fig. 2c). We then measured the active zonelength, defined as a specialization of the presynaptic mem-
brane thought to be the site of synaptic vesicle exocytosis.The length of the active zone measured in asymmetric spine
synapses found in a random sample of neuropil fields withinthe Acb was slightly, although significantly, shorter in MPH-
exposed rats, compared with SAL controls (CTR)(192.96 � 4.28 vs. 212.52 � 5.1 mm; P < 0.01; Fig. 2d).
Neurite outgrowth is induced by Htr7 stimulation in
cultured striatal neurons
Htr7 has been recently involved in behavioral impairment
(Guscott et al. 2005; Hedlund et al. 2005; Roberts et al. 2004),in mood regulation (Bonaventure et al. 2002; Wesolowska
et al. 2006), in mnemonic processes and cognition (Ballatzet al. 2007; Cifariello et al. 2008; Gasbarri et al. 2008) and in
regulating cell cytoarchitecture (Kvachnina et al. 2005). Wethus hypothesized that Htr7 may modulate behavioral impul-
sivity via plastic changes at the neuronal level. To evaluate theeffects of Htr7 activation on neuronal morphology and neurite
outgrowth, we used primary cultures obtained from thestriatal complex of rats at the embryonic day (E) 16. Virtually
all cells in our cultures expressed the neuronal Tuj1 marker, asshown by coexpression with the nuclear marker DAPI, thus
indicating that the majority of cultured cells were neurons. Inaddition, more than 90% of striatal neurons were GABAergic
(as shown by immunoreactivity to the enzyme glutamatedecarboxylase) and were immunoreactive for the Grik2 sub-
unit and for Htr7 (Fig. 3a). Striatal cultures were treated for24 h with the appropriate agonist and/or antagonist, stained
with the anti-Tuj1 antibody (Fig. 3b) and analyzed
Figure 1: Quantitative analysis of
selected neuronal proteins in the
brain of MPH-exposed rats. (Panels
a,b) Western blot analyses performed
on the striatal complex of adult male rats
(PND 90) exposed to MPH (2 mg/kg/day)
or SAL during adolescence (PND 30–44).
A representative example of blots stained
with antibodies against GAD65, TH, Grik2,
Htr7 and b-actin (a) is shown. The density
of each protein, normalized to b-actin
staining, was expressed for the MPH
group (b) as mean � SEM fold change
over the respective SAL group, set as
reference level at 100% (Student’s t-test,
n ¼ 6; *P < 0.05). (Panels c–f) Quantifica-
tion of immunocytochemical staining with
antibodies against Htr7 (c), Grik2 (d), Sub
P (e) or TH (f) in different brain regions (i.e.
PFC, Dl-CPu, Vm-Cpu, both AcbC and
AcbSh) of adult male rats (PND 90)
exposed to MPH (2 mg/kg/day) or SAL
during adolescence (PND 30–44). The
expression of Htr7 (c) or TH (f) was mea-
sured by the mean density (density �area). In the same regions we counted
the number of cells positive for Grik2 (d) or
SubP (e), therefore these markers are ex-
pressed as proportion of the total target
area relative to the scanned area (propor-
tional area). The data are obtained from ten
randomly selected sections for each
region (Student’s t-test, n ¼ 5; *P <
0.05, **P < 0.01).
360 Genes, Brain and Behavior (2009) 8: 356–368
Leo et al.

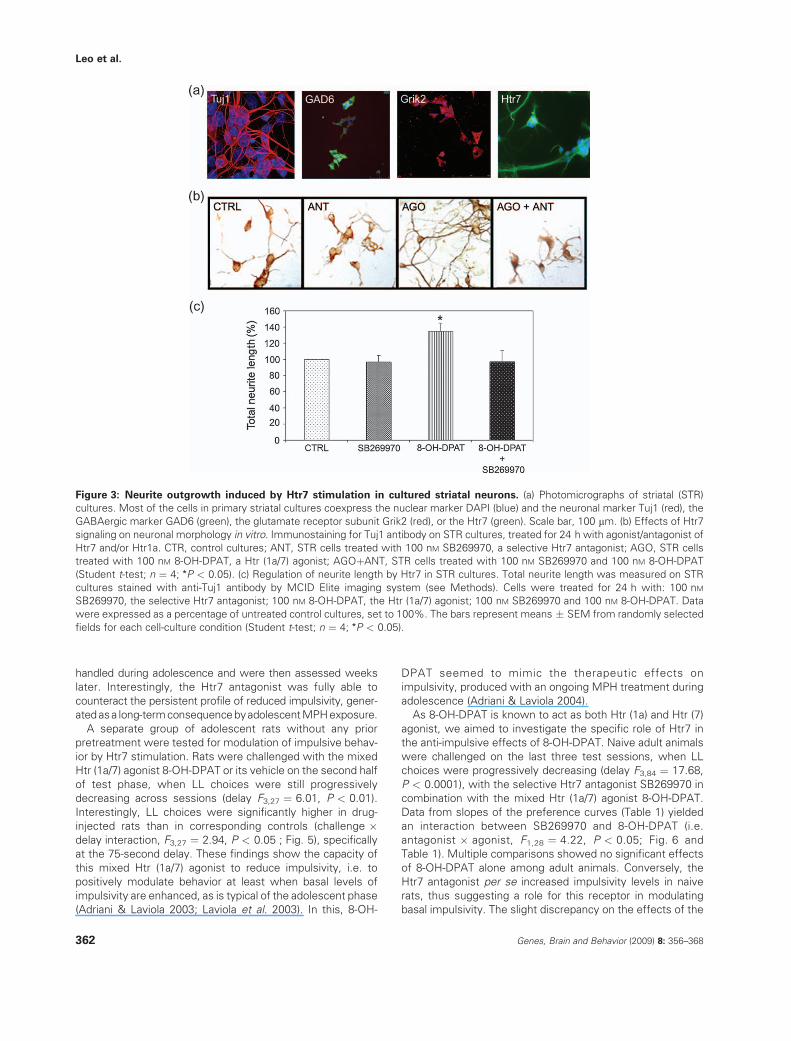
morphometrically. Exposure of cultures to 8-OH-DPAT, the
mixed Htr (1a/7) agonist, resulted in a significant increase inthe total length (Fig. 3c) and mean length (data not shown) of
neurites, when compared with untreated control cultures(CTR, set to 100%). These effects were at least partially
induced by selective stimulation of the Htr7 because theywere completely blocked by the simultaneous treatment with
the highly selective Htr7 receptor antagonist SB269970.Treatment with the Htr7 receptor antagonist alone had no
effect on the total length of neurites. Other morphological
parameters, such as the number of neurites, the end nodes,the branch nodes, the soma perimeter and the total area of
each neuron were not affected by stimulation of the cultureswith 8-OH-DPAT (data not shown).
Treatment with agonists of D1 or D2 DA receptors did notaffect any parameter in the morphology of striatal cultures
(data not shown). These results further emphasize theselective role of Htr7 activation in the outgrowth of striatal
neurites, as stimulation of other abundantly expressedG-protein-coupled receptors was ineffective.
Modulation of behavioral impulsivity by
Htr7-targeting drugs
Behavioral impulsivity can be studied in laboratory settings,
based on the assumption that impulsive subjects are lesstolerant to (and feel aversion for) situations of delayed reward
(Adriani & Laviola 2004; Evenden 1999; Evenden & Ryan1999). Adult animals, exposed to either MPH or SAL during
adolescence, were given choice between the immediatedelivery of one pellet of food (small and soon, SS), or the
delivery of five pellets of food after a delay (large and late, LL)
that was increased progressively over subsequent dailysessions (see Materials and methods). As expected, all rats
expressed significant preference for LL at the end of thetraining phase, when LL had no delay, then started to shift
toward SS choices (delay F3,33 ¼ 5.57, P < 0.01), as delayprogressively increased during the first half of the test phase.
To evaluate if a selective Htr7 blockade could counteract theMPH-reduced impulsivity, animals were assessed with the
selective Htr7 antagonist, SB269970, or its saline vehicle (SAL)
on the second half of the test phase, when LL choices were stillprogressively decreasing (delay F3,33 ¼ 9.02, P < 0.001). Con-
sistent with previous work (Adriani et al. 2007), a reduced levelof impulsivity appeared as a long-term consequence in MPH-
exposed subjects, compared with SAL-exposed controls (pre-treatment, F1,11 ¼ 8.56, P < 0.05), especially at the 45- and
60-second delays (Fig. 4). As the SAL and MPH pretreated ratsdo differ considerably, as we expected based on our previous
work (Adriani et al. 2007), we had no possibility of formulatinga null hypothesis for a general ANOVA, i.e. to assume that these
two pretreatment groups do not differ. The two pretreatmentgroups were hence analyzed separately.
In response to the Htr7 antagonist SB269970, no significanteffects were evident within the SAL-exposed group (challenge,
F1,6 ¼ 0.05, not significant). Conversely, within the MPH-exposed group, rats challenged with the Htr7 antagonist
exhibited significantly lower LL choices, compared with theircorresponding SAL controls (challenge, F1,5 ¼ 7.79, P < 0.05),
at the 45- and 60-second delays (Fig. 4b). In summary, Htr7antagonist administration per se failed to exert consistent
effects, at least in rats that were born in our facility, were
Figure 2: Synaptic organization in the
Acb of MPH-exposed rats. Changes in
the number and structure of synaptic
contacts in adult male rats (PND 90)
exposed to MPH (0 or 2 mg/kg/day) during
adolescence (PND 30–44). The synaptic
complexes are shown (arrow) for SAL (a)
and MPH (b) exposed groups (individual
sampling area, 9 mm2; total sampling area,
270 mm2). (c) The number of Acb synapses
shows a significant increase (Student‘s
t-test, n ¼ 5; **P < 0.01) in MPH-exposed
rats (filled column) (3.85 � 0.24 synapses/
9 mm2) compared with controls (CTR, open
column) (2.96 � 0.16 synapses/9 mm2). (d)
The active zone (AZ) length (mm) of the
accumbal synapses (post-synaptic density)
shows a significant decrease (Student’s
t-test, n ¼ 5; **P < 0.01) in MPH-exposed
rats (filled column) (192.96 � 4.28 mm)
compared with controls (CTR, open col-
umn) (212.52 � 5.1 mm). Scale bar:
0.5 mm.
Genes, Brain and Behavior (2009) 8: 356–368 361
Long-term consequences of methylphenidate during adolescence
handled during adolescence and were then assessed weekslater. Interestingly, the Htr7 antagonist was fully able to
counteract the persistent profile of reduced impulsivity, gener-ated asa long-term consequence byadolescent MPH exposure.
A separate group of adolescent rats without any priorpretreatment were tested for modulation of impulsive behav-
ior by Htr7 stimulation. Rats were challenged with the mixedHtr (1a/7) agonist 8-OH-DPAT or its vehicle on the second half
of test phase, when LL choices were still progressivelydecreasing across sessions (delay F3,27 ¼ 6.01, P < 0.01).
Interestingly, LL choices were significantly higher in drug-injected rats than in corresponding controls (challenge �delay interaction, F3,27 ¼ 2.94, P < 0.05 ; Fig. 5), specificallyat the 75-second delay. These findings show the capacity of
this mixed Htr (1a/7) agonist to reduce impulsivity, i.e. topositively modulate behavior at least when basal levels of
impulsivity are enhanced, as is typical of the adolescent phase(Adriani & Laviola 2003; Laviola et al. 2003). In this, 8-OH-
DPAT seemed to mimic the therapeutic effects onimpulsivity, produced with an ongoing MPH treatment during
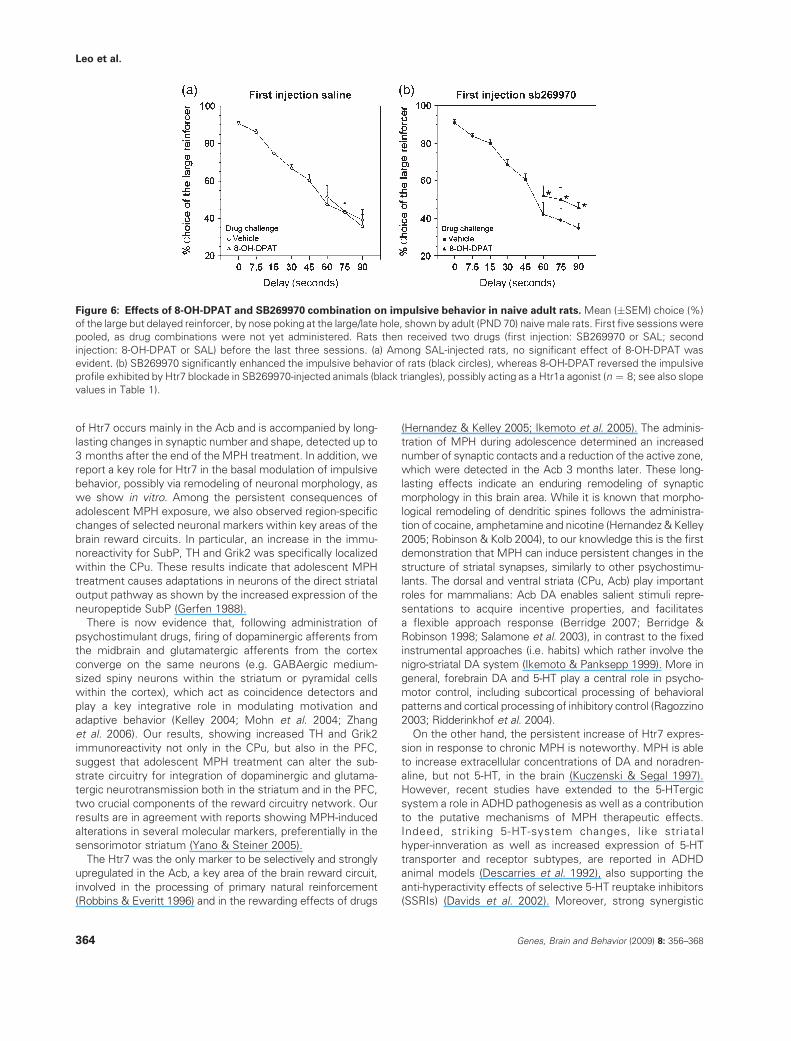
adolescence (Adriani & Laviola 2004).As 8-OH-DPAT is known to act as both Htr (1a) and Htr (7)
agonist, we aimed to investigate the specific role of Htr7 inthe anti-impulsive effects of 8-OH-DPAT. Naive adult animals
were challenged on the last three test sessions, when LLchoices were progressively decreasing (delay F3,84 ¼ 17.68,
P < 0.0001), with the selective Htr7 antagonist SB269970 incombination with the mixed Htr (1a/7) agonist 8-OH-DPAT.
Data from slopes of the preference curves (Table 1) yieldedan interaction between SB269970 and 8-OH-DPAT (i.e.
antagonist � agonist, F1,28 ¼ 4.22, P < 0.05; Fig. 6 andTable 1). Multiple comparisons showed no significant effects
of 8-OH-DPAT alone among adult animals. Conversely, theHtr7 antagonist per se increased impulsivity levels in naive
rats, thus suggesting a role for this receptor in modulatingbasal impulsivity. The slight discrepancy on the effects of the
Figure 3: Neurite outgrowth induced by Htr7 stimulation in cultured striatal neurons. (a) Photomicrographs of striatal (STR)
cultures. Most of the cells in primary striatal cultures coexpress the nuclear marker DAPI (blue) and the neuronal marker Tuj1 (red), the
GABAergic marker GAD6 (green), the glutamate receptor subunit Grik2 (red), or the Htr7 (green). Scale bar, 100 mm. (b) Effects of Htr7
signaling on neuronal morphology in vitro. Immunostaining for Tuj1 antibody on STR cultures, treated for 24 h with agonist/antagonist of
Htr7 and/or Htr1a. CTR, control cultures; ANT, STR cells treated with 100 nM SB269970, a selective Htr7 antagonist; AGO, STR cells
treated with 100 nM 8-OH-DPAT, a Htr (1a/7) agonist; AGOþANT, STR cells treated with 100 nM SB269970 and 100 nM 8-OH-DPAT
(Student t-test; n ¼ 4; *P < 0.05). (c) Regulation of neurite length by Htr7 in STR cultures. Total neurite length was measured on STR
cultures stained with anti-Tuj1 antibody by MCID Elite imaging system (see Methods). Cells were treated for 24 h with: 100 nM
SB269970, the selective Htr7 antagonist; 100 nM 8-OH-DPAT, the Htr (1a/7) agonist; 100 nM SB269970 and 100 nM 8-OH-DPAT. Data
were expressed as a percentage of untreated control cultures, set to 100%. The bars represent means � SEM from randomly selected
fields for each cell-culture condition (Student t-test; n ¼ 4; *P < 0.05).
362 Genes, Brain and Behavior (2009) 8: 356–368
Leo et al.

antagonist per se may be ascribed to uncontrolled effects ofprevious handling for drug exposure during adolescence.
Our findings clearly show the capacity of a mixed Htr (1a/7)agonist to positively modulate behavior in the presence of
enhanced basal levels of impulsivity, which were found innaive adult animals treated with the Htr7 antagonist. In this
group, indeed, the coadministration of 8-OH-DPAT reducedimpulsivity levels, by counteracting the pro-impulsive effects
of the Htr7 antagonist alone. Present results confirm the Htr7as an important player in the modulation of basal levels of
impulsive behavior, suggesting a potential benefit from this
receptor as a target for novel therapy in self-control disorders.
Discussion
We have recently reported that MPH produces differential
brain effects during adolescence, compared with adult rats(Canese et al. 2009). Adolescent MPH exposure modulates
gene expression in the striatal complex, suggesting itslong-lasting impact on many neuronal functions: Notably, a
persistent upregulation of striatal Htr7 mRNA was associatedwith a low basal level of behavioral impulsivity (Adriani et al.
2006 2007). We are now reporting that increased expression
Figure 4: Blockade of adolescentMPH consequences on impulsivity by the Htr7 antagonist, SB269970. Mean (�SEM) choice (%)
of the large but delayed reinforcer, by nose poking at the large/late hole, shown in drug-free state by adult male rats (PND 70) exposed to
MPH (0 or 2 mg/kg/day) during adolescence (PND 30–44). First five sessions were pooled, as the challenge (SB269970 or SAL) was not
yet administered to rats. Impulsivity was reduced as a long-term consequence of adolescent MPH (Tukey post hoc tests between
adolescent exposure groups, n ¼ 8; *P < 0.05), as shown on saline vehicle (SAL) challenge (white circles). Impulsivity levels, shown by
MPH-exposed rats (b) on SB269970 (3 mg/kg once/day) challenge (black circles) were drawn to those expressed by the SAL-exposed (a)
control groups (Tukey post hocs between challenge groups, n ¼ 8; *P < 0.05).
Figure 5: Acute effects of the Htr (1a/7) agonist, 8-OH-DPAT,
on impulsivity in adolescent rats. Mean (�SEM) choice (%) of
the large but delayed reinforcer, by nose poking at the large/late
hole, shown by adolescent (PND 30–44) male rats born in our
colony. The first five sessions were pooled, as no challenge was
yet administered. Rats were challenged with 8-OH-DPAT (0 or
0.06 mg/kg once/day) during the last three sessions at the
longest delay (Tukey post hoc tests; n ¼ 8; *P < 0.05).
Table 1: Effects of 8-OH-DPAT and SB269970 combination on
impulsive behavior in naive adult rats
Group Slope value
Vehicle Vehicle �23.97 � 2.48
Vehicle 8-OH-DPAT �25.22 � 2.08
SB269970 Vehicle �32.36 � 2.31*
SB269970 8-OH-DPAT �22.97 � 3.33†
Slope (mean � SEM) of the preference–delay curve from naive adult
(PND 70) male rats receiving two drugs (first injection: SB269970 or
SAL; drug challenge: 8-OH-DPAT or SAL) on the last 3 days of the
intolerance-to-delay task (Tukey post hoc tests, n ¼ 8). *P < 0.05 vs.
vehicle-vehicle animals; †P < 0.05 vs. SB269970-vehicle animals.
Genes, Brain and Behavior (2009) 8: 356–368 363
Long-term consequences of methylphenidate during adolescence
of Htr7 occurs mainly in the Acb and is accompanied by long-lasting changes in synaptic number and shape, detected up to
3 months after the end of the MPH treatment. In addition, wereport a key role for Htr7 in the basal modulation of impulsive
behavior, possibly via remodeling of neuronal morphology, aswe show in vitro. Among the persistent consequences of
adolescent MPH exposure, we also observed region-specificchanges of selected neuronal markers within key areas of the
brain reward circuits. In particular, an increase in the immu-noreactivity for SubP, TH and Grik2 was specifically localized
within the CPu. These results indicate that adolescent MPH
treatment causes adaptations in neurons of the direct striataloutput pathway as shown by the increased expression of the
neuropeptide SubP (Gerfen 1988).There is now evidence that, following administration of
psychostimulant drugs, firing of dopaminergic afferents fromthe midbrain and glutamatergic afferents from the cortex
converge on the same neurons (e.g. GABAergic medium-sized spiny neurons within the striatum or pyramidal cells
within the cortex), which act as coincidence detectors andplay a key integrative role in modulating motivation and
adaptive behavior (Kelley 2004; Mohn et al. 2004; Zhanget al. 2006). Our results, showing increased TH and Grik2
immunoreactivity not only in the CPu, but also in the PFC,suggest that adolescent MPH treatment can alter the sub-
strate circuitry for integration of dopaminergic and glutama-tergic neurotransmission both in the striatum and in the PFC,
two crucial components of the reward circuitry network. Ourresults are in agreement with reports showing MPH-induced
alterations in several molecular markers, preferentially in thesensorimotor striatum (Yano & Steiner 2005).
The Htr7 was the only marker to be selectively and stronglyupregulated in the Acb, a key area of the brain reward circuit,
involved in the processing of primary natural reinforcement(Robbins & Everitt 1996) and in the rewarding effects of drugs
(Hernandez & Kelley 2005; Ikemoto et al. 2005). The adminis-tration of MPH during adolescence determined an increased
number of synaptic contacts and a reduction of the active zone,which were detected in the Acb 3 months later. These long-
lasting effects indicate an enduring remodeling of synapticmorphology in this brain area. While it is known that morpho-
logical remodeling of dendritic spines follows the administra-tion of cocaine, amphetamine and nicotine (Hernandez & Kelley
2005; Robinson & Kolb 2004), to our knowledge this is the firstdemonstration that MPH can induce persistent changes in the
structure of striatal synapses, similarly to other psychostimu-
lants. The dorsal and ventral striata (CPu, Acb) play importantroles for mammalians: Acb DA enables salient stimuli repre-
sentations to acquire incentive properties, and facilitatesa flexible approach response (Berridge 2007; Berridge &
Robinson 1998; Salamone et al. 2003), in contrast to the fixedinstrumental approaches (i.e. habits) which rather involve the
nigro-striatal DA system (Ikemoto & Panksepp 1999). More ingeneral, forebrain DA and 5-HT play a central role in psycho-
motor control, including subcortical processing of behavioralpatterns and cortical processing of inhibitory control (Ragozzino
2003; Ridderinkhof et al. 2004).On the other hand, the persistent increase of Htr7 expres-
sion in response to chronic MPH is noteworthy. MPH is ableto increase extracellular concentrations of DA and noradren-
aline, but not 5-HT, in the brain (Kuczenski & Segal 1997).However, recent studies have extended to the 5-HTergic
system a role in ADHD pathogenesis as well as a contributionto the putative mechanisms of MPH therapeutic effects.
Indeed, striking 5-HT-system changes, like striatalhyper-innveration as well as increased expression of 5-HT
transporter and receptor subtypes, are reported in ADHDanimal models (Descarries et al. 1992), also supporting the
anti-hyperactivity effects of selective 5-HT reuptake inhibitors(SSRIs) (Davids et al. 2002). Moreover, strong synergistic
Figure 6: Effects of 8-OH-DPAT and SB269970 combination on impulsive behavior in naive adult rats. Mean (�SEM) choice (%)
of the large but delayed reinforcer, by nose poking at the large/late hole, shown by adult (PND 70) naive male rats. First five sessions were
pooled, as drug combinations were not yet administered. Rats then received two drugs (first injection: SB269970 or SAL; second
injection: 8-OH-DPAT or SAL) before the last three sessions. (a) Among SAL-injected rats, no significant effect of 8-OH-DPAT was
evident. (b) SB269970 significantly enhanced the impulsive behavior of rats (black circles), whereas 8-OH-DPAT reversed the impulsive
profile exhibited by Htr7 blockade in SB269970-injected animals (black triangles), possibly acting as a Htr1a agonist (n ¼ 8; see also slope
values in Table 1).
364 Genes, Brain and Behavior (2009) 8: 356–368
Leo et al.

interactions exist between MPH and 5-HT (1B) receptoragonists (Borycz et al. 2008), and treatment with fluoxetine,
a classical SSRI, can normalize adult behavioral deficitsinduced by early-life exposure of rats to MPH (Bolanos et al.
2008). These findings suggest a functional link betweenADHD, the effects of MPH, and the 5-HTergic system. Here,
we extend these putative interactions, highlighting thata strong and enduring Htr7 upregulation (following adolescent
MPH administration) has functional implications for impulsivebehavior, and is consistent with the known interactions
between Htr7 and neural effects induced by psychostimulantdrugs of abuse (Mnie-Filali et al. 2007).
The neurophysiological mechanism responsible for theseeffects is still unknown. As for MPH-induced Htr7 upregulation,
it can be speculated that MPH-increased catecholamine levelscould trigger a cascade of adaptive events, leading to a long-term
Htr7 increase. As for the role of Htr7 in behavioral modulation,we sought for neurobiological substrates by probing its possible
plastic effects on the neuronal cytoarchitecture (Kvachnina et al.2005). Indeed, the stimulation of cultured striatal neurons with
the mixed Htr (1a/7) agonist, 8-OH-DPAT, induced a markedneurite outgrowth. These effects were specifically triggered by
Htr7 activation, because they were blocked by application of theHtr7 selective antagonist, SB-269970. Noteworthy, the applica-
tion of DA receptor agonists, well known to trigger plastic CPu/Acb changes (Geldwert et al. 2006; Gerfen 2000), did not have
any effect on the neurite length nor on other morphological
parameters in our culture conditions, thus strengthening thespecificity of the 8-OH-DPAT-mediated effects.
Our data are consistent with the increased neurite elonga-tion previously reported in mouse hippocampal neurons
following Htr7 activation (Kvachnina et al. 2005) and furthersupport the crucial role of this receptor in the modulation of
neuronal morphology. The effects observed following Htr7stimulation in vitro could be exploited in vivo to exert
beneficial effects on behavioral output. Similarly to adoles-cent MPH, a direct Htr7-targeting drug might act through
a plastic morphofunctional remodeling of the Htr7-expressingforebrain neurons, possibly in the accumbens. Further studies
are required to confirm this hypothesis, and development ofnovel Htr7 agonist drugs would certainly help.
Htr7 function and cognitive impulsivity
Present data provide evidence for a direct relationshipbetween tonic Htr7 function and the modulation of impulsive
behavior and self-control capacity. A persistent overexpres-sion of the Htr7 gene in the striatal complex, generated by
adolescent MPH administration, was indeed accompanied inthe behavioral domain by a reduced basal level of cognitive
impulsivity (see also Adriani et al. 2007). Accordingly, in thepresent study, an acute drug-induced blockade of such over-
expressed Htr7 function effectively reversed the reduction ofbasal impulsivity produced by MPH.
Self-control, a component of decision-making processes,requires: (1) to withhold the rapid ‘subcortical’ processing of
contingencies and (2) to wait for the slower elaboration ofa ‘planned’ decision, by foresight on the future consequences
of a given action or choice strategy (McClure et al. 2004;
Ridderinkhof et al. 2004). The positive modulatory effects ofMPH in decreasing impulsivity levels suggest a more efficient
(cortical) decision-making process, possibly through enhancedcorticolimbic cross-talk (Cardinal et al. 2001; Christakou et al.
2001). The observed Acb overexpression of Htr7 might beinvolved in these respects. It is worth underlining that Htr7 is
involved in ultra- and circadian rhythms, via the hypothalamicinternal clock (Hedlund & Sutcliffe 2004). Any effect in the
delay-discounting task, found in rats treated with Htr7 agonistsand/or antagonist, could reflect a difference in the internal
perception of time, rather than effects on cognitive impulsivity.Some results in behavioural tasks show that 8-OH-DPAT may
disrupt timing abilities (Chiang et al. 2000). In the present study,we decided to probe the drug challenge only at the highest
delays, so that rats were able to face the introduction andprogression of delays under fully untouched ability to evaluate
the elapsing time. The first half of all ID-task testing phaseswas thus unaffected by altered perception of the delay
intervals. As for the three drug challenge sessions, as a matterof fact, we cannot exclude that rats were detecting delay
intervals as lasting less (or more) and thus bearing them more(or less) easily. Anyway, time estimation is one of the factors,
which does account for decision making in hyperbolic delay-discounting models (Ho et al. 1999; Kheramin et al. 2003), and
the role of Htr7 in cognitive impulsivity may well tap onto delay-interval affective impact, rather than onto corticostriatal cross-
talk and top-down inhibitory control.
In addition, the selective blockade of Htr7 per se mayenhance impulsive behavior in adult rats, depending on intrinsic
and extrinsic factors such as age and/or previous handlingexperience. Recently, in vitro experiments have reported
inverse agonist properties for the SB269970 (Rauly-Lestienneet al. 2007), which may therefore explain the pro-impulsive
actions of this compound. Alternatively, serotonin is knownto exert a tonic control over intolerance-to-delay processes
(Talpos et al. 2006; van den Bergh et al. 2006; Winstanley et al.2004), with Htr7 possibly playing an essential role. However,
participation of Htr1a in the general control of impulsivebehavior cannot be ruled out. Accordingly, a combined involve-
ment of Htr7 and Htr1a has been described for regulatorymechanisms, with both receptor subtypes acting in the same
direction (Faure et al. 2006; Guscott et al. 2005; Heisler et al.1998; Landry et al. 2006). Hence, anti-impulsive properties
might be proposed for drugs acting at either receptor, althoughcomplex interactions between these receptors deserve further
investigations. Fully validated, selective drugs for Htr7 overHtr1a are not available yet, unfortunately: the most useful
compound to target Htr7 is 8-OH-DPAT, which however hasbeen used for decades as a classical Htr1a agonist. Available
literature depicts 8-OH-DPAT‘s abilities to decrease prematureresponding in reaction-time tasks, following intra-PFC infusion
(Winstanley et al. 2003) and systemic administration (Bloklandet al. 2005), but not subcutaneously (Carli & Samanin 2000). We
thus decided to more deeply investigate the possible contribu-tion of Htr7 stimulation in 8-OH-DPAT’s effects on impulsivity.
In this respect, we found a consistent ‘therapeutic’ effectfor 8-OH-DPAT, similarly to what reported by some authors
(Bizot et al. 1999; Evenden & Ryan 1999; Poulos et al. 1996)but not by others (Winstanley et al. 2005). In our hands, the
8-OH-DPAT counteracted indeed any profile of altered
Genes, Brain and Behavior (2009) 8: 356–368 365
Long-term consequences of methylphenidate during adolescence

self-control baseline (as observed during adolescence, or afternaive adults were treated with Htr7 antagonist). Interestingly,
the 8-OH-DPAT did not influence baseline impulsivity in non-naive adult rats (which indeed faced previous experimental
manipulation during adolescence). We suggest that a baselineof increased impulsivity was necessary to observe the ‘ther-
apeutic’ action of this compound. In other words, it onlyreduced an already elevated impulsivity, like in adolescent rats
or when Htr7 was blocked in naive adult rats, presently. Asa matter of fact, however, Htr7 was not the primary site of
action for 8-OH-DPAT, which acted possibly through Htr1A.Altogether, we provide clear evidence that basal levels of
impulsivity may depend on the physiological tone at Htr7, andmay also be influenced by previous experimental manipula-
tion, thus tapping onto the delicate psychobiological equilib-rium underlying self-control parameters. The possibility that
selective Htr7 agonists may be useful for therapy of impulse-control disorders still remains open. Indeed, the effect of
8-OH-DPAT should be attributed to drug‘s action on a targetother than Htr7, as was evident at least from the third of our
present experiments.
Conclusion
In summary, our data show that chronic MPH exposure duringrat adolescence produces enduring structural rearrangement of
neural pathways involved in reward-based behavior, in terms of
synaptic contacts and accumbal Htr7 upregulation. We alsoshow that Htr7 stimulation plays a key role in the regulation of
neurite outgrowth in striatal neurons, at least in vitro. Finally,we show that a selective Htr7 antagonist effectively reverses
the persistent increase in the basal self-control profile, pro-duced by adolescent MPH, and may also enhance the basal
levels of behavioral impulsivity, at least in naive adult rats.Taken together, these results suggest that Htr7 may sub-
serve the persistent changes in neuronal morphologyobserved following adolescent MPH exposure, and may play
a major role in the modulation of self-control behavior (seealso Ballatz et al. 2007). Investigation on self-control capacity
may provide insights on the efficiency of corticolimbic cross-talk, possibly via accumbal Htr7 expression, thus improving
our understanding of the molecular and structural determi-nants of impulsive behavior. New avenues can be explored to
identify more selective tools for alternative therapeutic tar-gets to treat this symptom, which is common to many
neuropsychiatric disorders (Peluso et al. 2006; Swann et al.2003 2004; Winstanley et al. 2006).
Author contributions
D.L. contributed to the experimental design and carried out
the experiments on neuronal primary cultures and the Westernblots. W.A. contributed to the experimental design, treated
the animals, carried out the behavioral experiments andperformed the statistical analyses. C.C. realized the experi-
ments on neuronal markers and morphology. G.C. realizedthe experiments on neuronal markers and morphology. E.R.
realized the behavioral experiments. E.M.M. realized thebehavioral experiments and performed the statistical analy-
ses. U.P. devised the experiments on neuronal primary
cultures. M.P. devised the experiments on neuronal markersand morphology. C.P.-C. devised the experiments and coor-
dinated the experimental work. G.L. devised the experimentsand coordinated the experimental work.
Disclosure/Conflict of interest
The author(s) declare that except for income received from
their primary employer no financial support or compensationhas been received from any individual or corporate entity over
the past 3 years for research or professional service and thereare no personal financial holdings that could be perceived as
constituting a potential conflict of interest.
References
Adriani, W., Leo, D., Greco, D., Rea, M., Di Porzio, U., Laviola, G. &Perrone-Capano, C. (2006) Methylphenidate administration to ado-lescent rats determines plastic changes on reward-related behaviorand striatal gene expression. Neuropsychopharmacology 31,1946–1956.
Adriani, W., Canese, R., Podo, F. & Laviola, G. (2007) Reducedimpulsive behavior and 1H MRS detectable metabolic brainchanges in adult rats exposed to methylphenidate during adoles-cence. Neurotoxicol Teratol 29, 116–125.
Adriani, W. & Laviola, G. (2003) Elevated levels of impulsivity andreduced place conditioning with d-amphetamine: Two behavioralfeatures of adolescence in mice. Behav Neurosci 117, 695–703.
Adriani, W. & Laviola, G. (2004) Windows of vulnerability to psycho-pathology and for therapeutic strategy in the adolescent rodentmodel. Behav Pharmacol 15, 341–352.
Ballatz, S.J., Akil, H. & Watson, S.J. (2007) The 5-HT(7) receptor: rolein novel object discrimination and relation to novelty-seekingbehaviour. Neuroscience 149, 192–202.
van den Bergh, F.S., Bloemarts, E., Groenink, L., Olivier, B. & Oosting,R.S. (2006) Delay aversion: effects of 7-OH-DPAT. 5-HT(1A/1B)-receptor stimulation and D-cycloserine. Pharmacol Biochem Behav85, 736–743.
Berridge, K.C. (2007) The debate over dopamine’s role in reward: thecase for incentive salience. Psychopharmacology 191, 391–431.
Berridge, K.C. & Robinson, T.E.(1998) What is the role of dopamine inreward: hedonic impact, reward learning, or incentive salience?Brain Res Rev 28, 309–369.
Bizot, J., Le Bihan, C., Puech, A.J., Hamon, M. & Thiebot, M. (1999)Serotoninand tolerance todelayof reward in rats. Psychopharmacology146, 400–412.
Blokland, A., Sik, A. & Lieben, C. (2005) Evaluation of DOI, 8-OH-DPAT,eticlopride and amphetamine on impulsive responding in a reactiontime task in rats. Behav Pharmacol 16, 93–100.
Bolanos, C.A., Barrot, M., Berton, O., Wallace-Black, D. & Nestler,E.J. (2003) Methylphenidate treatment during pre- and periadoles-cence alters behavioral responses to emotional stimuli at adult-hood. Biol Psychiatry 54, 1317–1329.
Bolanos, C.A., Willey, M.D., Maffeo, M.L., Powers, K.D., Kinka, D.W.,Grausam, K.B. & Henderson, R.P. (2008) Anti-depressant treat-ment can normalize adult behavioral deficits induced by early-lifeexposure to methylphenidate. Biol Psychiatry 63, 309–316.
Bonaventure, P., Nepomuceno, D., Kwok, A., Chai, W., Langlois, X.,Hen, R., Stark, K., Carruthers, N., Lovenberg, TW. (2002) Recon-sideration of 5-hydroxy-tryptamine (5-HT) (7) receptor distributionusing [(3)H]-5-carboxamido-tryptamine and [(3)H]-8-hydroxy-2-(-di-n-proplamino)-tetraline: analysis in brain of 5-HT(1A) knockoutand 5-HT(1A/1B) double knockout mice. J Pharmacol Exp Ther 302,240–248.
Borycz, J., Zapata, A., Quiroz, C., Volkow, N.D. & Ferre, S. (2008)A 5-HT(1B) receptor-mediated serotonergic modulation of themethylphenidate-induced locomotor activation in rats. Neuro-psychopharmacology 33, 619–626.
366 Genes, Brain and Behavior (2009) 8: 356–368
Leo et al.

Bradford, M.M. (1976) A rapid and sensitive method for the quanti-tation of microgram quantities of protein utilizing the principle ofprotein-dye binding. Anal Biochem 72, 248–254.
Canese, R., Adriani, W., Marco, E.M., De Pasquale, F., Lorenzini, P.,De Luca, N., Fabi, F., Podo, F. & Laviola, G. (2009) Peculiarresponse to methylphenidate in adolescent compared to adult rats:a ph-MRI study. Psychopharmacology 203: 143–153.
Cao, J., Lotfipour, S., Loughlin, S.E. & Leslie, F.M. (2007) adolescentmaturation of cocaine-sensitive neural mechanisms. Neuro-psychopharmacology 32, 2279–2289.
Cardinal, R.N., Pennicott, D.R., Sugathapala, C.L., Robbins, T.W. &Everitt, B.J. (2001) Choice induced in rats by lesions of the nucleusaccumbens core. Science 292, 2499–2501.
Carli, M. & Samanin, R. (2000) The 5-HT(1A) receptor agonist8-OH-DPAT reduces rats‘ accuracy of attentional performance andenhances impulsive responding in a five-choice serial reaction timetask: role of presynaptic 5-HT(1A) receptors. Psychopharmacology149, 259–268.
Chiang, T.J., Al-Ruwaitea, A.S., Mobini, S., Ho, M.Y., Bradshaw, C.M. &Szabadi, E. (2000) Effects of 8-hydroxy-2-(di-n-propylamino)tetralin(8-OH-DPAT) on performance on two operant timing schedules.Psychopharmacology 151, 379–391.
Christakou, A., Robbins, T.W. & Everitt, B.J. (2001) Functionaldisconnection of a prefrontal cortical–dorsal striatal system disruptschoice reaction time performance: implications for attentionalfunction. Behav Neurosci 115, 812–825.
Cifariello, A., Pompili, A. & Gasbarri, A. (2008) 5-HT(7) receptors inthe modulation of cognitive processes. Behav Brain Res 195,171–179. [Epub 2007 Dec 23].
Davids, E., Zhang, K., Kula, N.S., Tarazi, F.I. & Baldessarini, R.J. (2002)Effects of norepinephrine and serotonin transporter inhibitors onhyperactivity induced by neonatal 6-hydroxy-dopamine lesioning inrats. J Pharm Exp Ther 301, 1097–1102.
Dean, B., Pavey, G., Thomas, D. & Scarr, E. (2006) Cortical serotonin(7, 1D) and (1F) receptors: Effects of schizophrenia, suicide andantipsychotic drug treatment. Schizophr Res 88, 265–274.
Descarries, L., Soghomonian, J.J., Garcia, S., Doucet, G. & Bruno, J.P.(1992) Ultrastructural analysis of the serotonin hyperinnervation inadult rat neostriatum following neonatal dopamine denervationwith 6-hydroxy-dopamine. Brain Res 569, 1–13.
Dogrul, A. & Seyrek, M. (2006) Systemic morphine produce anti-nociception mediated by spinal 5-HT(7), but not 5-HT(1A) and5-HT(2) receptors in the spinal cord. Br J Pharmacol 149, 498–505.
East, S.Z., Burnet, P.W., Kerwin, R.W. & Harrison, P.J. (2002) AnRT-PCR study of 5-HT(6) and 5-HT(7) receptor mRNAs in thehippocampal formation and the prefrontal cortex in schizophrenia.Schizophr Res 57, 15–26.
Evenden, J.L. (1999) Varieties of impulsivity. Psychopharmacology146, 348–361.
Evenden, J.L. & Ryan, C.N. (1999) The pharmacology of impulsivebehaviour in rats VI: the effects of ethanol and selective serotoner-gic drugs on response choice with varying delays of reinforcement.Psychopharmacology 146, 413–421.
Faure, C., Mnie-Filali, O., Scarna, H., Debonnel, G. & Haddjeri, N.(2006) Effects of the 5-HT(7) receptor antagonist SB-269970 on therat hormonal and temperature responses to the 5-HT(1A/7) recep-tor agonist 8-OH-DPAT. Neurosci Lett 404, 122–126.
Fiszman, M.L. (1991) Dopamine synthesis precedes dopamineuptake in embryonic rat mesencephalic neurons. J Neurochem 56,392–399.
Frantz, K.J., O’dell, L.E. & Parsons, L.H. (2007) Behavioral andneurochemical responses to cocaine in periadolescent and adultrats. Neuro-psychopharmacology 32, 625–637.
French, S.J., Totterdell, S. (2004) Quantification of morphologicaldifferences in boutons from different afferent populations to thenucleus accumbens. Brain Res 1007, 167–177.
Gasbarri, A., Cifariello, A., Pompili, A. & Meneses, A. (2008) Effect of5-HT(7) antagonist SB-269970 in the modulation of working andreference memory in the rat. Behav Brain Res 195, 164–170. [Epub2008 Jan 9].
Gerfen, C.R. (1988) Synaptic organization of the striatum. J ElectronMicrosc Tech 10, 265–281.
Gerfen, C.R. (2000) Molecular effects of dopamine on striatal-projectionpathways. Trends Neurosci 23, S64–S70.
Geldwert, D., Norris, J.M., Feldman, I.G., Schulman, J.J., Joyce, M.P. &Rayport, S. (2006) Dopamine presynaptically and heterogeneouslymodulates nucleus accumbens medium-spiny neuron GABA syn-apses in vitro. BMC Neurosci 7, 53.
Graybiel, A.M. (1990) Neurotransmitters and neuromodulators in thebasal ganglia. Trends Neurosci 13, 244–254.
Guscott, M.R., Bristow, L.J., Hadingham, K., Rosahl, T.W., Beer,M.S., Stanton, J.A., Bromidge, F., Owens, A.P., Huscroft, I.,Myers, J., Rupniak, N.M., Patel, S., Whiting, P.J., Hutson, P.H.,Fone, K.C., Biello, S.M., Kulagowski, J.J. & McAllister, G. (2005)Genetic knockout and pharmacological blockade studies of the5-HT(7) receptor suggest therapeutic potential in depression.Neuro-pharmacology 48, 492–502.
Hedlund, P.B., Huitron-Resendiz, S., Henriksen, S.J. & Sutcliffe, J.G.(2005) 5-HT(7) receptor inhibition and activation induce antidepressant-like behavior and sleep pattern. Biol Psychiatry 58, 831–837.
Hedlund, P.B. & Sutcliffe, J.G. (2004) Functional, molecular andpharmacological advances in 5-HT(7) receptor research. TrendsPharmacol Sci 25, 481–486.
Hedlund, P.B. & Sutcliffe, J.G. (2007) The 5-HT(7) receptor influencesstereotypic behaviour in a model of obsessive-compulsive disorder.Neurosci Lett 414, 247–251.
Heisler, L.K., Chu, H.M., Brennan, T.J., Danao, J.A., Bajwa, P.,Parsons, L.H. & Tecott, L.H. (1998) Elevated anxiety and antide-pressant-like responses in serotonin 5-HT(1A) receptor mutantmice. Proc Natl Acad Sci USA 95, 15049–15054.
Hernandez, P.J. & Kelley, A.E. (2005) Cracking addiction the secondtime around: reconsolidation of drug-related memories. Neuron47, 772–775.
Ho, M.Y., Mobini, S., Chiang, T.J., Bradshaw, C.M. & Szabadi, E.(1999) Theory and method in the quantitative analysis of ‘impulsivechoice’ behavior: the implications for psychopharmacology.Psychopharmacology 146, 362–372.
Hyman, S.E., Malenka, R.C. & Nestler, E.J. (2006) Neural mecha-nisms of addiction: the role of reward-related learning and memory.Annu Rev Neurosci 29, 565–598.
Ikemoto, S. & Panksepp, J. (1999) The role of nucle us accumbensdopamine in motivated behavior: a unifying interpretation withspecial reference to reward-seeking. Brain Res Rev 31, 6–41.
Ikemoto, S., Qin, M. & Liu, Z.H. (2005) The functional divide forprimary reinforcement of D-amphetamine lies between the medialand lateral ventral striatum: is the division of the accumbens core,shell, and olfactory tubercle valid? J Neurosci 25, 5061–5065.
Kelley, A.E. (2004) Memory and addiction: shared neural circuitry andmolecular mechanisms. Neuron 30, 161–179.
van Kesteren, R.E. & Spencer, G.E. (2003) The role of neurotransmit-ters in neurite outgrowth and synapse formation. Nat Rev Neurosci14, 217–231.
Kheramin, S., Body, S., Ho, M., Velazquez-Martinez, D.N., Bradshaw, C.M.,Szabadi, E., Deakin, J.F. & Anderson, I.M. (2003) Role of the orbitalprefrontal cortex in choice between delayed and uncertainreinforcers: a quantitative analysis. Behav Processes 64, 239–250.
Kuczenski, R. & Segal, D.S. (1997) Effects of methylphenidate onextracellular dopamine, serotonin, and norepinephrine: comparisonwith amphetamine. J Neurochem 68, 2032–2037. Erratum in:J Neurochem 69: 1332.
Kvachnina, E., Liu, G., Dityatev, A., Renner, U., Dumuis, A., Richter,D.W., Dityateva, G., Schachner, M., Voyno-Yasenetskaya, T.A. &Ponimaskin, E.G. (2005) 5-HT(7) receptor is coupled to G alphasubunits of heterotrimeric G12-protein to regulate gene transcrip-tion and neuronal morphology. J Neurosci 25, 7821–7830.
Landry, E.S., Lapointe, N.P., Rouillard, C., Levesque, D., Hedlund, P.B.& Guertin, P.A. (2006) Contribution of spinal 5-HT(1A) and 5-HT(7)receptors to locomotor-like movement induced by 8-OH-DPAT inspinal cord-transected mice. Eur J Neurosci 24, 535–546.
Laviola, G., Macrı, S., Morley-Fletcher, S. & Adriani, W. (2003) Risk-taking behavior in adolescent mice: Psychobiological determinantsand early epigenetic influence. Neurosci Biobehav Rev 27, 19–31.
Le Corre, S., Sharp, T., Young, A.H. & Harrison, P.J. (1997) Increaseof 5-HT(7) and 5-HT(1A) mRNA expression in rat hippocampusafter adrenalectomy. Psychopharmacology 130, 368–374.
McClure, S.M., Laibson, D.I., Loewenstein, G. & Cohen, J.D. (2004)Separate neural systems value immediate and delayed monetaryrewards. Science 306, 503–507.
Genes, Brain and Behavior (2009) 8: 356–368 367
Long-term consequences of methylphenidate during adolescence

Meneses, A. (2004) Effects of the 5-HT(7) receptor antagonistsSB-269970 and DR 4004 in auto-shaping Pavlovian / instrumentallearning task. Behav Brain Res 155, 275–282.
Mnie-Filali, O., Dahan, L., Zimmer, L. & Haddjeri, N. (2007) Effects ofthe serotonin 5-HT(7) receptor antagonist SB-269970 on inhibitionof dopamine neuronal firing induced by amphetamine. Eur JPharmacol 570, 72–76.
Mohn, A.R., Yao, W.D. & Caron, M.G. (2004) Genetic and genomicapproaches to reward and addiction. Neuro-pharmacology 47,S101–S110.
Mullins, U.L., Gianutsos, G. & Eison, A.S. (1999) Effects of antide-pressants on 5-HT(7) receptor regulation in the rat hypothalamus.Neuro-psychopharmacology 21, 352–367.
Nestler, E.J. (2005) Is there a common molecular pathway foraddiction? Nat Neurosci 8, 1445–1449.
Neumaier, J.F., Sexton, T.J., Yracheta, J., Diaz, A.M. & Brownfield,M. (2001) Localization of 5-HT(7) receptors in rat brain by immu-no-cytochemistry, in situ hybridization and agonist stimulated cFosexpression. J Chem Neuroanat 21, 63–73.
Paxinos, G. & Watson C. (2001) The Rat Brain in StereotaxicCoordinates [Atlas]. Academic Press, London.
Peluso, M.A., Hatch, J.P., Glahn D.C., Monkul E.S., Sanches M., NajtP., Bowden C.L., Barratt E.S. & Soares J.C. (2007) Trait impulsivityin patients with mood disorders. J Affect Disord 100, 227–231.
Perez-Garcia, G.S. & Meneses, A. (2005) Effects of the potential5-HT(7) receptor agonist AS 19 in an autoshaping learning task.Behav Brain Res 163, 136–140.
di Porzio, U., Daquet, M.C., Glowinski, J. & Prochiantz, A. (1980)Effect of striatal cells on in vitro maturation of mesencephalicdopaminergic neurones grown in serum-free conditions. Nature288, 370–373.
Poulos, C.X., Parker, J.L. & Le, A.D. (1996) Dexfenfluramine and8-OH-DPAT modulate impulsivity in a delay-of-reward paradigm:Implications for a correspondence with alcohol consumption.Behav Pharmacol 7, 395–399.
Rauly-Lestienne, I.,Boutet-Robinet,E.,Ailhaud,M.C.,Newman-Tancredi,A.&Cussac,D. (2007) Differential profile of typical, atypical and third generationantipsychotics at human 5-HT(7a) receptors coupled to adenylyl cyclase:detection of agonist and inverse agonist properties Naunyn Schmiede-bergs. Arch Pharmacol 376, 93–105.
Raymond, J.R., Mukhin, Y.V., Gelasco, A., Turner, J., Collinsworth, G.,Gettys, T.W., Grewal, J.S. & Garnovskaya, M.N. (2001) Multiplicityof mechanisms of serotonin receptor signal transduction. Pharma-col Ther 92, 179–212.
Ragozzino, M.E. (2003) Acetylcholine actions in the dorsomedialstriatum support the flexible shifting of response patterns. Neuro-biol Learn Mem 80, 257–267.
Ridderinkhof, K.R., van den Wildenberg, W.P.M., Segalowitz, S.J. &Carter, C.S. (2004) Neurocognitive mechanisms of cognitive con-trol: The role of prefrontal cortex in action selection, responseinhibition, performance monitoring and reward-based learning.Brain and Cognition 56, 129–140.
Robbins, T.W. & Everitt, B.J. (1996) Neuro-behavioural mechanismsof reward and motivation. Curr Opin Neurobiol 6, 228–236.
Roberts, A.J., Krucker, T., Levy, C.L., Slanina, K.A., Sutcliffe, J.G.,Hedlund, PB (2004) Mice lacking 5-HT receptors show specificimpairments in contextual learning. Eur J Neurosci 19, 1913–1922.
Robinson, T.E. & Kolb, B. (2004) Structural plasticity associated withexposure to drugs of abuse. Neuro-pharmacology 47, 33–46.
Ruat, M., Traiffort, E., Leurs, R., Tardivel-Lacombe, J., Diaz, J.,Arrang, J.M. & Schwartz, J.C. (1993) Molecular cloning, character-ization and localization of a high-affinity serotonin receptor 5-HT(7)activating cAMP formation. Proc Natl Acad Sci USA 90, 8547–8551.
Russo, A., Pellitteri, R., Monaco, S., Romeo, R. & Stanzani, S. (2005)In vitro postnatal expression of 5-HT(7) receptors in the rathypothalamus: an immunohistochemical analysis. Brain Res DevBrain Res 154, 211–216.
Salamone, J.D., Correa, M., Mingote, S. & Weber, S.M. (2003)Nucleus accumbens dopamine and the regulation of effort infood-seeking behavior: implications for studies of natural motiva-tion, psychiatry, and drug abuse. J Pharmacol Exp Ther 305, 1–8.
Schikorski, T., Stevens, C.F. (1997) Quantitative ultrastructural anal-ysis of hippocampal excitatory synapses. J Nerosci 17, 5858–5867.
Shen, Y., Monsma, F.J. Jr, Metcalf, M.A., Jose, P.A., Hamblin, M.W. &Sibley, D.R. (1993) Molecular cloning and expression of a5-hydroxy-tryptamine7 serotonin receptor subtype. J Biol Chem268, 18200–18204.
Sleight, A.J., Carolo, C., Petit, N., Zwingelstein, C. & Bourson, A.(1995) Identification of 5-hydroxy-tryptamine-7 receptor bindingsites in the rat hypothalamus: sensitivity to chronic antidepressanttreatment. Mol Pharmacol 47, 99–103.
Swann, A.C., Pazzaglia, P., Nicholls, A., Dougherty, D.M. & Moeller,F.G. (2003) Impulsivity and phase of illness in bipolar disorder.J Affect Disord 73, 105–111.
Swann, A.C., Dougherty, D.M., Pazzaglia, P.J., Pham, M. & Moeller,F.G. (2004) Impulsivity: a link between bipolar disorder and sub-stance abuse. Bipolar Disord 6, 204–212.
Talpos, J.C., Wilkinson, L.S. & Robbins, T.W. (2006) A comparison ofmultiple 5-HT receptors in two tasks measuring impulsivity. JPsychopharmacol 20, 47–58.
Venero, J.L., Vizuete, M.L., Machado, A. & Cano, J. (1997) Develop-mental expression of 5-HT(7) receptor mRNA in rat brain visualstructures after neonatal enucleation. Neuroreport 8, 1531–1535.
Volkow, N. & Li, T.K. (2005) The neuroscience of addiction. NatNeurosci 8, 1429–1430.
Wesolowska, A., Nikiforuk, A., Stachowicz, K. & Tatarczynska, E.(2006) Effect of the selective 5-HT(7) receptor antagonist SB269970 in animal models of anxiety and depression. Neuro-pharmacology 51, 578–586.
Winstanley, C.A., Chudasama, Y., Dalley, J.W., Theobald, D.E.,Glennon, J.C. & Robbins, T.W. (2003) Intra-prefrontal 8-OH-DPATand M100907 improve visuospatial attention and decreaseimpulsivity on the five-choice serial reaction time task in rats.Psychopharmacology 167, 304–314.
Winstanley, C.A., Dalley, J.W., Theobald, D.E. & Robbins, T.W. (2004)Fractionating impulsivity: contrasting effects of central 5-HT deple-tion on different measures of impulsive behavior. Neuro-psychopharmacology 29, 1331–1343.
Winstanley, C.A., Theobald, D.E., Dalley, J.W. & Robbins, T.W. (2005)Interactions between serotonin and dopamine in the control ofimpulsive choice in rats: therapeutic implications for impulsecontrol disorders. Neuro-psychopharmacology 30, 669–682.
Winstanley, C.A., Eagle, D.M. & Robbins, T.W. (2006) Behavioralmodels of impulsivity in relation to ADHD: translation betweenclinical and preclinical studies. Clin Psychol Rev 26, 379–395.
Yano, M. & Steiner, H. (2005) Topography of methylphenidate(Ritalin)-induced gene regulation in the striatum complex: Differen-tial effects on c-Fos, substance P and opioid peptides. Neuro-psychopharmacology 30, 901–915.
Zahm, D.S. & Brog, J.S. (1992) On the significance of sub-territories inthe ‘accumbens’ part of the rat ventral striatum. Neuroscience 50,751–767.
Zhang, J., Zhang, L., Jiao, H., Zhang, Q., Zhang, D., Lou, D., Katz, J.L. &Xu, M. (2006) c-Fos facilitates the acquisition and extinction ofcocaine-induced persistent changes. J Neurosci 26, 13287–13296.
Zorrilla, E.P. (1997) Multiparous species present problems(and possibilities) to developmentalists. Devl Psychobiol 30,141–150.
Acknowledgments
Research performed along the lines of the ‘‘ADHD-sythe’’ young-investigator projects (to WA): ‘‘under-40 call’’ from the ItalianMinistry of Health (assigned); "ERC-StG" from EU-FP7 ideas(pending). Supported by the bilateral Italy–USA Program on RareDiseases ‘X-linked or autosomal rare mental retardation syn-dromes’ (7NR1), the ERARE-EuroRETT Network (ERAR/6) (toGL); by PRIN/MIUR 2007_STRNHK_002 (to CPC); by PRIN/MIUR2007_F7AJYJ_004 (to U.dP.); by the Federation of EuropeanBiochemical Societies (FEBS) fellowship (to D.L.) and theEuropean Mind and Metabolism Association (EMMA) fellowship(to E.M.M. and E.R.). Authors wish to thank B. Duvillard (Universityof Lyon, France), L. Leone (IGB, CNR, Naples, Italy), A. De Simoneand M. R. Bianco (SUN, Naples, Italy) for their invaluable help.
368 Genes, Brain and Behavior (2009) 8: 356–368
Leo et al.