metal-atp complexes as substrates and free metal ions as activators of the red cell calcium pump
TRANSCRIPT

Cell Calcium 2: 449-458, 1981
METAL-ATP COMPLEXES AS SUBSTRATES AND FREE METAL IONS AS ACTIVATORS OF THE RED CELL CALCIUM PUMP
B. Sarkadi, Agnes Enyedi, and G. Gdrdos
National Institute of Haematology and Blood Transfusion, 1113 Budapest, Hungary
ABSTRACT
The plasma membrane calcium pump in most mammalian cells is the basic mechanism for assuring a low cyto- plasmic calcium concentration. In inside-out human red cell membrane vesicles /IOVs/ the substrate and metal specificity as well as the intracellular protein /Cal- modulin/ regulation of the ATP-dependent active calcium
;~;~;~=;~t~a:h;; ~~qpstft~~~e~i~t~~l:~w:~~sMX~~~; we .
complexes, including MgATP, MnATP, CoATP, FeATP, and NiATP, can serve as substrates for the calcium pump in IOVS. Calcium pumping is activated by the above metals, while Sr, Ba, Cu, Cd ions or the trivalent cations are ineffective in this respect. Calmodulin-stimulation of the calcium transport is present independent of the metal ions used for the activation of the pump. Based on kinetic studies we suggest that divalent metal ions interact with the red cell calcium pump at four different sites: 1.1 MeATP complex is the true substrate of the pump; 2.1 Ca or Sr ions activate the system by binding to the transport site/s/ and other metal ions competitively inhibit this binding; 3.1 the presence of free divalent metal ions /Mg, Mn, Co, Fe, or Ni, but not Ca, Sr, Ba I is required for activating calcium translocation; 4.1 interaction with a Ca - calmodulin complex specifically stimulates calcium pumping.
INTRODUCTION
The ATP-dependent calcium pump in the human erythrocyte provides a model-system for the investigation
449

of this transport nrocess with general prevalence and vital importance in mammalian cells. In inside-out red cell membraq$ vesicle an ATP + Mg
/IOVl prenarations the pump p:;zuces -dependent uphill calcium uptake and
calcium + magnesium + ATP-binding sites are at the external surface of these vesicles. In IOVs, containing the pump in its natural environment, calcium transport, ATPase activity, membrane phosphorylation, as well as the stimulation by the intracellular regulatory protein,cal- modulin, can be studied simultaneously /see ref.11.
In the past few years there has been a controversy on the substrate specificity of the red cell calcjium pyr~p. Wolf /2/ suggested that the substrate of the /Ca +Mg i- -ATPase is MgATP, while the experiments of Schatzmann 131 and Sarkadi et al /4/ indicated that the substrate is either uncomplexed ATP,or that the pump cannot discrimin- ate between MgATP and free ATP. The latter conclusion is supported by the results of Muallem and Karlish 151, measuring calcium-dependent ATPase activity and membrane phosphorylation in red cell membranes. Experiments on the
'urn-induced phosphorylation of the pump protein by [CyatSt P)ATP clearly show that CaATP is accepted as a substrate and magnesium is not necessary for this 2+ phosphorylation 15-91. The possible role of free Mg is to stimulate dephosphorylation of the pump protein and thus to complete the calcium transport cycle /b-10/. Recent work of Penniston et al Ill/, and Graf and Penniston 1121, using isolated preparations of the red cell calcium pump protein,gave rise to the conclusion that the true, physiological substrate of the pump is CaATP. The key-experiments supporting this conclusion were those demonstrating that at low ATP I1 pM/ and calcium IO.6 - 1.0 FM/ concentrations the activity of the isolated and vrrified calcium pump ATPase was strongly reduced if Mg concentration was increased from 1 PM up to 1 mM . In these experiments the reduction in ATPase activity was parallel to the decrease in the calculated concentration of CaATP /11,12/. In the present paper we report experiments which indicate that CaATP is unlikely to be the physiological substrate of the in situ red cell calcium pump and demonstrate the effects z - various divalent cations on this transport system.
NATERIALS APUD METHODS
All the chemicals used were of reagent grade- IOVs were prepared and assayed for their sidedness as Zlescribed in ref. 13. Active calcium uptake was assessed by 'I);, rapid filtration of the vesicles and by measuring radioactivity0 in the IOVs, as in refs. 4 and 13. Each data point represents triplicate experiments by measuging IOV calcium uptake in 3 min incubation periods at 37 C. The
450

calcium uptake was linear at any ATP or metal concentrations examined at least up to 5 min. Red cell calmodulin was prepared as described in ref. 13.
RESULTS AND DISCUSSION
Fig. 1. Effects of changes in magnesium concentration on the rate of active calcium uptake by IOVs.
Incubation media: 120 mM KCl, 17 mM imide3ole- -HCl, /pH 7.01, 50 pM CaC12 / including Ca tracer/, 70 ).IM EGTA /free calcium = 1 pM/, 1 /_IM ATP, and l-1000 p" MgC12. IOV concentra- tion = 20 pg/ ml medium. The dotted lines represent the concentrations of free ATP, CaATP, and MgATP, respectively, as calculated by Penniston et al Ill/. Calmodulin = 0.5 pg/ml medium.
451

The data presented in Fig. 1 were obtained by repeating the experiments of Penniston and co-workers /11,12/, using similar incubation conditions but measuring active calcium uptake by the in situ trans- port enzyme in IOVs. In this system the= iso measurable calcium uptake in the absence of magnesium, while at increasing magnesium concentrations the transport rate increaEas and reaches its maximum value between 200-500 PM Mg . At magnesium levels higher than 500 PM the concentration of CaATP complex is practically zero, thus it is improbable that CaATP would be the only acceptable substrate for the red cell calcium pump. Active calcium uptake in IOVs is stimulated by added calmodulin at all the magnesium concentrations examined, although this stimulation is smaller at higher magnesium levels. This is most probably the result of a competition between calcium and magnesium ions in binding to calmodulin, which is active only in its calcium- -1iganded form 1141. Based on these results we suggest that in the isolated pump protein preparation /11,12/ magnesium may directly inhibit ATPase activity by binding to sites normally hidden in the hydrophobic areas of the red cell membrane. This inhibition might have led Penniston and his co-workers to an inappropriate conclusion. This view is further supported by the fact that while the activity of the isolated pump ATPase under the conditions presented in Fig.1 was only 0.1 % of the maximum rate /11,12/, IOVs transported calcium with about 10 % of the maximum rate /see Table 1.1.
Table I. Effects of changes in calcium and ATP concentrations on the value of K
Mg and the maximum transport rate in IOVs.
pM ATP
2; 1.5
10 150 14 100 15 250 18 500 18 500 22
6 30 60
200
r 1 PM Ca 2+ T
K~glw/
100 p Ca 2+
‘MgIPMl ( 'max V max
K = total magnesium concentration, producing half- -!&ximum c alcium_transpor$ rate: V = nanomol calcium x x mg IOV protein x min max
Incubation media: 120 mM K&l, 17 mM imidazole-HCl IpH 7.01, 100 w EGTA, 70 PM CaCl
8 /free calcium = 1 pMI, or 190 )iM
CaC12 /free calcium = 1 0 ~.&!/,l-500 q ATP. IOV = 20 )Iglml medium. Means of three different experiments.
452

When studying active calcium uptake by IOVs at two different calcium /l FM and 100 )IM/ and at variable ATP concentrations, the values for magnesium concentrations producing half-maximum transport rates /K / varied significantly /Table 1.1. The observed in%ease in K higher ATP concentrations, in accordance with2$he
Mg at
experiments demonstrating the role of free Mg in the dephosphorylation of the calcium pump protein 16-101, suggest the need for free magnesium as an activator of the calcium pump at the internal membrane surface. The data also indicate a competition between magnesium and calcium ions at this site /see also ref.51.
In the experiments presented in Fig.2, we examined the effects of various metal ions on the active calcium uptake by IOVs. In these experiments we did not use EGTA buffers and added 20 w CaCl and 20 PM ATP to the media. As shown in the figure,
such as Fe The explanation for the differences in the maximum transport values and the shapes of the curves in Fig.2 requires further investigation, but the calculations show that - since Mn, Co, Fe, and Ni form 10 - 50 times more stable complexes with ATP than Ca - at 1 m?4 concentrations of the above metals there is practically no CaATP in the medium. Therefore, the suggestion that the only acceptable substrate for the red cell calcium pump is CaATP 11.1,12/, proves untenable. Another important conclusion from these experiments is that free magnesium canbe substituted in activating the calcium pump by other divalent metal ions. The relative efficacy of various MeATP complexes as suhstra?es, and that of the free metals as activators of the pump should be further investigated by studying the partial reactions, that is the phosphorylation and dephosphorylation of the transport protein. Our preliminary measurements of the dephosphoryl- ation rate of the calclym ~~@,e;znq_~; ;;dI;;~,/~;~ublished data/ indicate th?$ Mn substitute for Mg in this reaction in a decreasing sequence of effectivity. In experiments to be reported in detail elsewhere we also examined the effects of changes in ATP and calcium concentrations on the rate of calcium uptake in the presence of various activating metals. The results are essentially similar to those obtained in the presence of magnesium, that is Kple is increased at higher ATP and/or calcium concens$ag;o:;e T';;ilha;;z; ;;d;;~t;,, again the role of free Me and a competition between Me and Ca at least at two types of sites.
453

i lb 100 lbOO~~h4 Me’+
Fig. 2. Effects of divalent metal ions on the rate of active calcium uptake by IOVs.
The incubation media contained 120 mM KCl, 17 mN @idasole-HCl /pH 7.01, 20 PM CaC12 / including
Ca tracer/, 20 I_'" ATP, and the divalent metals in the total concentratioy? indica$$d. When studying the effect of Co and Fe , the media were completed with 1 mM ascorbic acid to keep these ions in their reduced forms. Ascorbic acid had no effect on active calcium uptake in the presence of magnesium. All the calcium transport values represent uphill uptake of calcium into IOVS, as tested by the A23187 ionophore releasing effect /ref. 131. The dashed line indicates the rate of calcium uptake in the absence of added metals I magnesium concentration in the media was 2-3 PM/.
454
: : ‘\
: ’ 6
i\M"=+ : +CALMOnULfN
3 *
IO too lOOO& M Me 2+
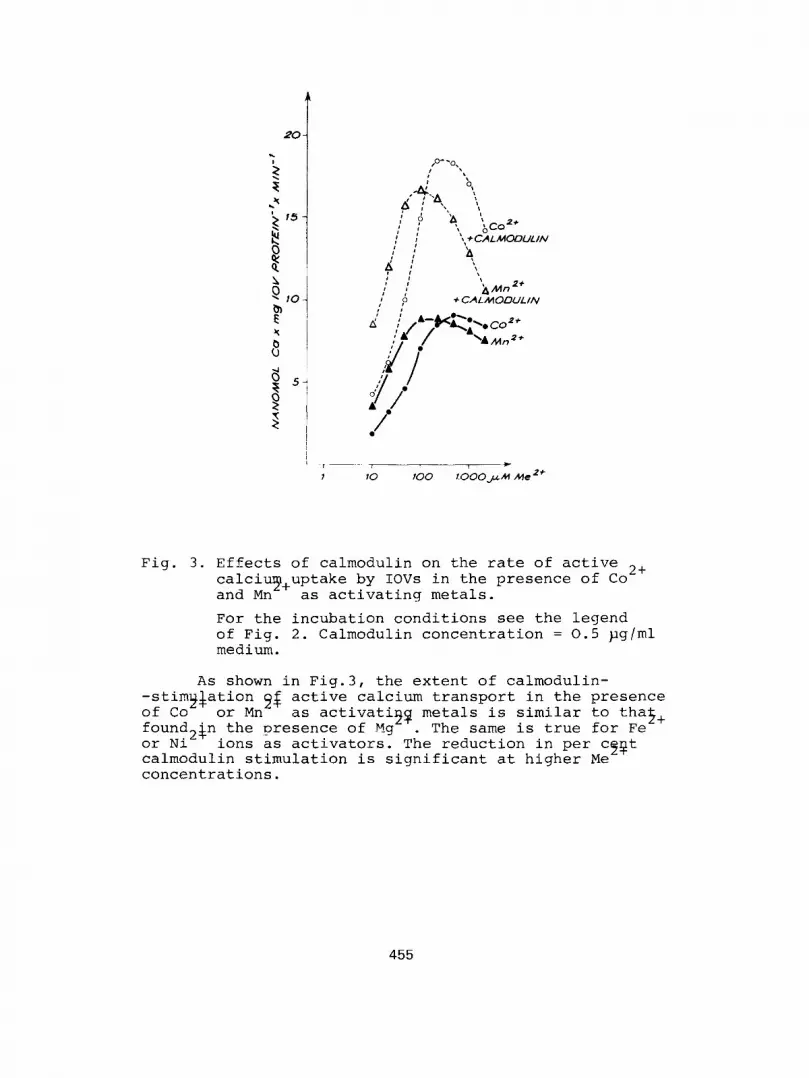
Fig. 3. Effects of calmodulin on the rate of active calciy+uptake by IOVs in the presence of Co
2+
and Mn as activating metals.
For the incubation conditions see the leqend of Fig. 2. Calmodulin concentration = 0.5 pqlml medium.
As shown in Fiq.3, the extent of calmodulin- active calcium transport in the presence as activatiqq metals is similar to that,
found2$n the presence of Mq . The same is true for Fe or Ni ions as activators. The reduction in per cfiqt calmodulin stimulation is significant at higher Me concentrations.
455

In conclusion, the present experiments suggest that the red cell calcium pump accepts various metal-liganded forms of ATP / and probably also free ATP/ as substrates. The true, physiological substrate of the in situ pump is most probably MgATP. The pump is specifically activated by2$alciyv lor Mg , Mn
strontiyT/ 115,161, co
bind@g to the transport site/s/. or Fe ISarkadi, unpublished data/
are not transported by the calcium pump but these metal ions can interact with the calcium-site and inhibit the enzyme activity /1,3,35-184+ A partial stimulation of the red cell membrane ICa +Mg /-ATPase activity by various metal ions has been shown by Pfleger and Wolf 1171.
Asgive calcium transport requires the presence of free Mg at the internal membrane surface. Magnesium in this role can be substituted by metals which have, similarly to magn?gium, smaller ic@c radii thZP calcium
'% radius of Fe = 0.074+ Ni itself, Sr
%= ;.;;;"'C",%=M; o~90~~~~~w~ple=C~207"~
IO.112 nm io; radius/; or Ba IO.134 nm ion radiusd+are not+ accepted at this site. Some metals, such as Cu or Cd directly interfere with protein actions and block calcium pumping.
lie have to note that the sarcoplasmic reticulum /SRI calcium pump has most probably a similar metal- -dependence as the shown 1191, that ?4n
sr;d cell2$alcium pump. It has be?? or Co- can substitute for Mg in
supporting the SR calcium pump, and a recent paper by Yamada and Ikemoto 1201 indicates that CoATP and MnATP can be utilized as substrates in the SR. Moreover, the experiments of the latter aut$o;; ;i",iz&e,,;;;t,;~e;~_ metals can substitute for Mg phosphorylation of the pump protein.
The broad range of metals accepted by the calcium pump as ATP complexes and activators is an intriguing phenomenon and may provide an experimental tool for the investigation of the molecular mechanism of this transport system. The characteristics of paramagnetic ions in their interactions with ATP and the pump protein may certainly be used for spectroscopic and other physical studies in this research. Moreover, as the stimulation of membrane phosphorylation and dephosphoryl- ation is different by different metal ions I Enyedi, unpublished data/, the metal-dependence of the pump may help in the separation of2$alcium traqgport linked. ATPase activity from other Ca and/or P4g -dependent ATP splitting processes in the plasma membranes.
456

ACKNOWLEDGEMENTS
This work was supported by the Scientific Research Council, Ministry of Health, Hungary /6-03-0306-01-1/G&/. The authors wish to thank Mrs. M. Sarkadi and Mrs. M, Berenyi for the skilful technical assistance.
1.
2.
3.
4.
5.
6.
7.
8.
9.
10.
11.
REFERENCES
Sarkadi, B. /198O/.Active calcium transport in human red cells. Biochim. Biophys. Acta2$04, 159-190. Wolf, H.U. /1972/.Studies on a Ca of human erythrocyte membranes.
Eff~~~e~~e;~24T;=$e
H. Biochim. Biophys. Acta 266, 361-375. Sch?$zmang+ H.J. /1977/.Role of magnesium in the
s A:: b:o:: c&l:
imulated membrane ATPase of human . J. Membrane Biol. 35, 149-158.
Sarkadi, B., .Macintyre, J.D., and Gbrdos, G. 119781. Kinetics of active calcium transport in inside-out red cell membrane vesicles. FEBS Letters 89, 78-82. Muallem, S., and Karlish, S.J.D. 119811. On the mechanism of regulation of the red cell calcium pump by calmodulin and ATP. Biochim. Biophys. Acta in press. Rega, A.F., and Garrahan, P.J. 119751. Calcium ion- -dependent phosphorylation of human erythrocyte membranes. Katz, s.,
J. Membrane Biol. 22, 313-237. and Blostein, R. 119751. Ca -stimulated
membrane phosphorylation and ATPase activity of the human erythrocyte. Biochim. Biophys. Acta 389, 314-324. Schatzmann, H-J., and Biirgin, H. 119781. Calcium in human red blood cells. Ann.N.Y.Acad.Sci. 307, 125-147. Szasz, I., Hasitz, M., Sarkadi, B2+ and Gbrdos,G. 119781. Phosphorylation of the Ca pump intermediate in intact red cells, isolated membranes and inside- -out vesicles. Mol. Cell. Biochem. 22, 147-152. Garrahan, P.J., and Rega, A2$'. 119781. Activation of partial rea?$ions of the Ca -ATPase from human red cells by Mg and ATP. Biochim. Biophys. Acta 513, 59-65. Penniston, J.T., Graf, E., Niggli, V., Verma, A.K., and Carafoli, E. /1980/. The plasma membrane calcium ATPase. In: Calcium Binding Proteins: Structure and Function leds. Siegel, F.L., Carafoli, E., Kretsinger, R.H., MacLennan, D.H., and Wasserman, R-H./, pp 23-30, Elsevier-North Holland, Amsterdam.
457

12. Graf, E., and Penniston, J.T. 1198ll. substrate,
CaAT$ - The at low ATP concentration of Ca ATPase
from human erythrocyte membranes. J. Biol. Chem. 256, 1587-1592.
13. Sarkadi, B., Szasz, I., and Gardos, G. /1900/. Characteristics and regulation of active calcium transport in inside-out red cell membrane vesicles. Biochim. Biophys. Acta 598, 326-338.
14. Scharff, 0. /1981/. Calmodulin - and its role in cellular activation. Cell Calcium 2, l-27.
15. Schatzmann, H.J., and Vincenzi, F.F. 119691. Calcium movements across the membrane of human red cells. J. Physiol. 201, 369-395.
16. Olson, E:J. 119791. Inhibition of active strontium transport from erythrocyte ghosts by internal calcium: evidence for a specific controlling site. J. Membrane Biol. 48, 265-284.
17. Pfleger, H., and Wolf, H.U. 119751. Activation of membrane bound high-affinity calcium ion sensitive adenosine triphosphatase of human erythrocytes by bivalent metal ions. Biochem. J. 147, 359-361.
18. Schatzmann, H.J., and Roelofsen, B. 119771. Some aspects of the Ca-pump in human red blood cells. In: Biochemistry of Membrane Transport leds. Semenza, G., and Carafoli, E./ pp 389-400, Springer-Verlag, Berlin.
19. Martonosi, A., and Feretos, R.+i1964/. Sarcoplasmic reticulum: I. The uptake of Ca by sarcoplasmic reticulum fragments. J. Biol. Chem. 299, 648-658.
20. Yamada, S., and Ikemoto, N. 119801. Reaction Mechanism of calcium-ATPase of sarcoplasmic reticulum. Substrates for phosphorylation reaction and back reaction, and further resolution of phosphorylated intermediates. J. Biol. Chem. 255, 3108-3119.
Received: 29.5.81
Revised version received: 27.7.81 accepted: 27.7.81
458