mechanisms of herbicide resistance in wild oats (avena spp.)
TRANSCRIPT

_*"Íot[il*uu **:{P" -."_
Mechanisms of He
in Witd Oats
Chanya Maneechote
Thesis Submitted for the Degree of
Doctor of Philosophy
in Faculty of Agricultural and Natural Resource Sciences
The University of Adelaide
Department of Crop Protection
Waite Agricultural Research Institute
July 1995

u
(;'ì,'i i "Table of Contents
Title page
Table of Contents
AbsEact
Declaration
Acknowledgements
Abbreviations
Chapter I Introduction
I .1 History of Herbicide Resistance
1.2 Herbicide Resistance in Ausualia
1.3 Mechanisms of Herbicide Resistance
1.3.1 Reduced Herbicide Uptake and Translocation
I.3.2 Enhanced Herbicide Metabolism
1.3.3 Modified Site of Action
I.3.4 Sequesfration of Herbicide within Plant Cells
1.4 Inheritance of Herbicide Resistance
1.5 Aryloxyphenoxypropionate and Cyclohexanedione Herbicides
1.5.1 Antagonism of APP and CHD Herbicides
L.5.2 Uptake and Translocation
1.5.3 Metabolism
I .6 Mode of Action of APP and CHD Herbicides
I .6.1 Inhibition of Fatty Acid Biosynthesis
1.6.2 Characteristics of ACCase Enzyme
1.6.3 Kinetics of Inhibition by APP and CHD Herbicides
I.6.4 Inheritance of Herbicide Resistant ACCase in Formerly
Susceptible Weed Species
I
ü
vtl
x
xi
xü
1
I
2
4
4
5
6
8
8
10
L2
12
t4
20
2t
23
25
25

ru
1.7 Disruption of Membrane Function by APP and CHD Herbicides
t.7 .l Diclofop as a ProtonoPhore
1.7 .2 Recovery from Diclofop Acid Induced Depolarization of the
PlasmaMembrane Electrogenic Potential in Some Resistant
V/eeds
1.8 Resistance to Acetyl-CoA Carboxylase-krhibiting Herbicides
1.9 V/ild Oat(Avena spp.)
1.9.1 Seed Dormancy and Germination
I.9.2 Seed Bank of lVild Oat
1.9.3 ControlofWildOat
1.10 Herbicide Resistance in Wild Oat
l.l 1 Objectives of this Project
Chapter 2 Materials and Methods
2.1 PlantMaærial
2.2 PlantCulture
2.2.I Germination
2.2.2 Pot culnue
2.2.3 Hydroponic Culnue
2.3 Dose Response Experiments
2.4 ACCase Extraction and Assay
2.5 Uptake and Translocation of ¡l4qDiclofop-Methyl
2.6 Metabolism of Herbicides in Plants
2'6.1 ¡l4clDiclofoP-MethYl
2.6.2 ¡4-t+ClSethoxydim
2.7 Measr¡rement of Plasma Membrane Potentials
26
27
27
28
30
3l
32
33
33
34
35
35
37
37
37
37
39
40
43
42
42
43
44

lv
Chapter 3 Spectrum of Resistance to Arylox¡phenoxy¡lropionate and
Cyclohexanedione Herbicides in lVild Oat Biot¡pes:
\ilhole Plant and Target Site Inhibition Studies
3.1 Introduction
3.2 Whole Plant Assessment of Resistance
3.3 Inhibition of ACCase Activity by Herbicides
3.4 Discussion
Chapter 4 Herbicide Resistance Endowed by an APP Insensitive
ACCase in BiotYPe SAS 1
4.t Introduction
4.2 lau'flaærials andMethods
4.2.1 Variations in ACCase During Development
4.3 Results
4.3.1 ACCase Activity During Growth Development
4.3.2 Uptake and Translocation of [14C]D'clofop-Methyl
4.3.3 Meøbolism of l14c]D'clofop-Methyl
4.3 .4 Effect of Diclofop Acid on Plasma Membrane Potential
4.3.5 Inhibition of ACCase Activity InVivo
4.4 Discussion
Chapter 5 Herbicide Resistance Endowed by an APP- and CHD-
Insensitive ACCase in Biotype NAF 6
5.1 Intoduction
5.2 Results
5.2.1 Uptake, Translocation and Metabolism of ¡llgtD'tlofop-Methyl
5.2.2 Uptake and Metabolism of ¡4-l4clsethoxydim
46
46
47
58
69
74
74
74
74
75
75
77
80
83
83
86
88
88
89
89
93

v
5.2.3 Effect of Diclofop Acid on Plasma Membrane Potential
5.3 Discussion
Chapter 6 Resistance to APP Herbicides Endowed by Two
Mechanisms in BiotYPe NAS 4
6.1 Introduction
6.2 Maænals andmethods
6.2.1 Effects of the Cytochrome P450Inhibitor Tetcyclasis on Diclofop
Metabolism
6.2.2 Effects of the Cytochrome P450 Inhibitor Tetcyclasis on Plant
Growth
6.2.3 Inhibition of ACCase by Diclofop in Single Lines of NAS 4
6.3 Results
6.3.1 Uptake and Translocation of ¡l4C¡Diclofop-Methyl
6.3.2 Metabolism of l14c]D'ctofop-Methyl
6.3.3 Effect of Tetcyclasis on Diclofop Metabolism
6.3.4 Effects of Tetcyclasis on Plant Growth
6.3.5 Inhibition of ACCase Activity by Diclofop Acid in Single Lines
of Biotype NAS 4
6.3.6 Effect of Diclofop Acid on Plasma Membrane Potential
6.4 Discussion
Chapter 7 Non-Target Site Mechanism Conferring Resistance to
ACCase-Inhibiting Herbicides in Biotype NAS 14
7.1 Innoduction
7 .2 Matenals and Methods
7 .2.1 Uptake, Translocation and Metabolism of ¡l+grD'.lofop-Methyl
7 .2.2 Absorption of ll4c]D'clofop-Methyl to Leaf Cuticle
97
97
100
100
t0l
l0l
101
t02
t02
t02
105
108
It2tt7
119
il9
t23
r23
t24
t24
t24

7.3 Results
7.3.t
7.3.2
7.3.3
7.3.4
7.3.5
Uptake and Metabolism of I l4C]Diclofop-Methyl
Translocation of [14c]Diclofop-Methyl
Absorption of Herbicide to I-eaf Cuticle
ACCase Activity inVivo
Effect of Diclofop on Plasma Membrane Potential
vl
t24
124
t27
t27
t29
t32
132
135
135
136
136
136
t37
137
138
142
t45
152
185
7.4 Discussion
Chapter 8 Investigations of Alterations of Membrane Responses to
Diclofop as Mechanism of Resistance in Wild Oat
8.1 InEoduction
8.2 Materials and Methods
8.2.I Effect of pH on Membrane Response to Diclofop Acid
8.2.2 Effect of Diclofop Acid on Acidification of External Solution
8.3 Results
8 .3 . 1 Effect of Diclofop Acid on Plasma Membrane Potential
8.3.2 Effects of pH on PlasmaMembrane Repolarization
8 .3 . 3 Acidification of the External Solution by Roots of Intact Plants
8.4 Discussion
Chapter 9 General Conclusions
Literature Cited
Appendix
159

vll
Abstract
Herbicide resistance, specifically to the acetyl coenzyme A carboxylase (ACCase)-inhibiting
herbicides aryloxyphenoxypropionate (APP) and cyclohexanedione (CIID) herbicides, in
wild oat has become a serious problem in cropping regions of Australia. This study was
conducted to elucidate the mechanisms endowing resistance to herbicides in wild oat. Ten
biotypes of wild oat (Avena fatua and A. sterilis ssp. ludovicíana) were examined for
resistance to ACCase-inhibiting herbicides. At the whole plant level, all biotypes were
resistant to at least one APP (diclofop-methyl, fluazifop-butyl, and haloxyfop-ethoxyethyl),
or CHD (sethoxydim and tralkoxydim) herbicide. The response of the target site, ACCase,
to the APP and CHD herbicides was examined in vitro. All resistant biotypes excePt one
contained a target site resistant to at least one APP and CHD herbicide. The response of the
target site of different biotypes to APP and CHD herbicides varied widely suggesting that
several different mutations within ACCase can endow resistance to these herbicides. Four
resistant biotypes (SAS 1, NAF 6, NAS 4 and NAS 14) of wild oat with contrasting
patterns of resistance at the whole plant and ACCase levels were studied further to fully
elucidate mechanisms endowing resistance.
Biotype SAS I is highly-resistant to APP herbicides but is not resistant to CHD herbicides.
The ACCase from this biotype displays high-level resistance to APP herbicides and low-
level resistance to CHD herbicides. Absorption, translocation and metabolism of
¡l4Cldiclofop-methyl were simila¡ in both resistant biotype SAS I and susceptible biotype
SAS 2. Hence, the target-site based mechanism is the only mechanism responsible for
resistance in biotype SAS 1.
Biotype NAF 6 is moderately resistant to both APP and CHD herbicides at the whole plant
level and contains a target enzyme ACCase with moderate resistance to both groups of
herbicides. There were no differences in uptake, translocation and metabolism of
llaC]diclofop-methyl between susceptible SAS 2 and resistant NAF 6 wild oat biotypes. In

vlu
addition, absorption and meøbolism of ¡+laClsethoxydim in both biotypes were simila¡. A
modified target site is solely responsible for resistance to both APP and CHD herbicides in
biotype NAF 6.
Biotype NAS 4 is highly resistant to diclofop-methyl in the field yet contains an ACCase
which displays only moderate resistance to this herbicide. No differences in uptake and
translocation of diclofop-methyl were observed in this biotype compared to the susceptible.
However, this biotype showed a 1.S-fold enhanced rate of metabolism of diclofop-methyl.
In the prcsence of the cytochrome P450 inhibitor tetcyclasis, the rate of diclofop metabolism
in this biotype was decreased to the same level as that of the susceptible biotype SAF 19
suggesting the involvement of cytochrome P450 monooxygenases in resistance. In
addition, tetcyclasis increased the phytotoxicity of diclofop-methyl against the resistant
biotype. Studies with ten individual families of biotype NAS 4 indicate that two
mechanisms of resistance, an altered target site and enhanced metabolism, are present in each
individual of the NAS 4 population.
Biotype NAS 14 shows low-level resistance to APP and CHD herbicides but contains á
sensitive ACCase. Reduced uptake or enhanced metabolism of diclofop-metþl do not
account for resistance in this biotype. A small difference between susceptible and resistant
biotypes was observed in translocation of diclofop to the meristematic region following
application of ll4c]d'clofop-methyl to the leaf axil of twoleaf stage plants. This reduced
translocation of diclofop was correlated with a reduction in the inhibition of ACCase in vivo
following application of diclofop-methyl to intact plants. This non-target site-based
mechanism may be responsible for the low level of resistance to diclofop-methyl in this
biotype.
The relationship between resistance to graminicides and the ability to repolarize the plasma
membrane potential following diclofop acid-induced depolarization was studied in all
biotypes of wild oat. No biotype demonstrated recovery of the plasma membrane potential
at pH 5.8 following the removal of diclofop acid. pH was found to be an important
determinant of the ability of plasma membranes to repolarize following removal of diclofop

D(
acid. An initial dictofop acid-induced depolarization of the plasma membrane potential was
reversible in herbicide-free solution buffered at pH 6.5, but not when buffered at pH 5.8. It
is clear from this study that resistance to graminicides is not correlated with differences in the
ability of wild oat biotypes to recover from diclofop acid-induced depolarization of the
plasma membrane potential.
In conclusion, at least three mechanisms of resistance to ACCase-inhibiting herbicides, a
modified target-site, enhanced metabolism and reduced translocation are found in Avena
populations. The target-site based mechanism, which is responsible for the moderate and
high resistance to herbicides at the whole plant level, is the most cornmon mechanism. The
non-target site based mechanisms, enhanced herbicide metabolism and reduced translocation
of herbicide to the target site, were observed in one resistant biotype each. The variety of
resistance mechanisms which have appea¡ed in wild oat biotypes may lead to cross-
resistance to other herbicide classes and make control of resistant wild oat with alternative
herbicides difficult.

x
Declaration
This work contains no material which has been accepted for the awa¡d of any other degree in
any university or other tertiary institution and to the best of my knowledge and belief
contains no material previously published or written by another person, except where due
reference has been made in the text.
I give consent to this copy of my thesis, when deposited in the University Library, being
available for loan and photocopying.
Chanya Maneechote
July 1995

xr
Acknowledgements
I wish to thank my supervisors, Dr. S.B. Powles and Dr. C. Preston for their valuable
supervision, advice and constructive criticism during the course of this study. I express my
sincere thanks to Dr. J.A.M. Holtum for his expert advice and also special thanks to Dr.
F.J. Tardif and Dr. J. Karotam for critical reading of this manuscript.
Sincere thanks is extended to Dr. S.D. Tyerman from School of Biological Sciences,
Flinders University, Bedford Parh South Australia for providing facilities for the membrane
potential work.
I would like to thank Dr. A.M. Mansooji for providing the dose response data of wild oat
biotypes SAS I and SAS 2.
I would also like to thank the members of herbicide resistance group at the V/aite
Agricultural Research Institute: Dr. J.C. Christopher, Dr. E. Purba, J.M. Matthews, P.
Boutsalis, S. Hole, B. Nietschke, H. Alizadeh, N. Farida and I. Hidayat for their help,
encouragement and companionship.
Special thanks a¡e extended to my dear friends, S. Jamjod, C. Koshawatana, S. Agung and
K. Beaton for their help and freindship.
I thank the Australian Agency for International Development (AusAID) for the financial
support and also thank Thai Goverment for study leave permission.
Lastly, my deepest gratitude to my parents and family for their support and understanding
throughout the study.

xlt
Abbreviations
ai
APP
ACCase acetyl co€nzyme A carboxylase (EC 6.4.1.2)
active ingredient
Aryloxyphenoxypropionate
CHD Cyclohexanedione
Clethodim (E,E)-(!)-2-[ I -[[(3-chloro-2-propenyl) oxy] iminol propyll-S-
[(ethylthio) propyl] -3-hydroxy-2-cyclohexene- I -one
Cycloxydim (t)-2-[ 1 -(ethoxyimino) butyl] -3-hydroxy-5-thian-3-ylcyclohex-
2-enone
Diclofopmethyl methyl(t)-2-Í4-(2,4-dtchlorophenoxy) phenoxyl propanoate
EDTA
dithiothreitol
ethylenediamineteEaacetic acid
ethyl- I -(2,4-dicilorphenyl)-5-trichloromethyl- lH-t,2,4-
tiazole-3-carboxylic acid
ethyl(t)2-[4-[(6-chloro -2-betuoxazolyl) oxy] phenoxyl
propanoate
buty(t)-2-[a-[[5-(trifluoromethyl)-2-pyridinyl] phenoxyl
propanoate
fresh weight
fenchloraz ole-ethyl
Fenoxaprop-ethyl
Fluazifop-butyl
DTT
FW

ruu
Haloxyfoperhoxyethyl ethoxyerhyt(t)-2-[4-[[3-chloro-5-(trifluoromethyl)-2-pyndinyu
oxyl phenoxYlProPanoate
HPLC High Pressure Liquid Chromatography
herbicide concenEation required to reduce invitro ACCase
activity 507o withrespect to control assay without herbicide
Iso
LDso
LSS
MES
PMSF
PPO
POPOP
PVP
Quizalofopethyl
Sethoxydim
Tralkoxydim
Tricine
dose of herbicide required to kill 507o of plants
Liquid Scintillation SpecEometrY
2-(N-morpholino)ethanesulfonic acid
phenylmethylsulfonyl fluoride
2,5-diphenyloxazole
1,4-bis (5 -phenyloxazoyl)benzene
polyvinylpyrrolidone
ethyl(+)-2- [4- [(6-chloro-2-quinoxdinyl) oxy]phenoxyl
propanoate
2- [ I -ethoxyimino)butyl] -5 - l2-(ethylthio)propyl] -3-hydroxy-2-
cyclohexen-1-one
2- [ 1 (ethoxyimino)propyl] -3-hydroxy -5-(2,4,6'
trimethylphenyl)cyclohex-2-enone
N-[2-hydroxy- 1 - 1 -bis (hydroxymethyl) ethyl] glycine
Tris (hydroxymethyl) aminomethaneTris

xivErrata
Page 6,
Page 13,
Page 2I,
Page 24,
Page 25,
Page 26,
Page 27,
Page 29,
Page 34,
Page 40,
Page 44,
Page 72,
Page 77 ,
Page 80,
line 4. Change to: Reduced herbicide sensitivity ...
line 14. Change to: The predominant translocated form is diclofop acid ...
line 7-8. Change to: Initially, the ATP-dependent carboxylation of biotin is
catalysed in the biotin carboxylase domain.
line 26. Change "containg" to containing.
lirc 21. Change "diclots" to dicots.
lines 5-6. Remove last sentence from paragraph.
lines 19-20. Remove reference to Stoltenberg et al. 1989.
line 10. Change to: A second mode of action of aryloxyphenoxypropionic acids
and cyclohexanediones is hypothesised to ...
line 11. Change to: ...proton-translocating plasmalemma ATPase.
Table 1.9. Remove line containing Festuca rubra.
line 2I. Change to: Ten putative resistant biotypes with varying levels of resistance
were examined.
line 11. Change to: The supernatant was slowly...
line 22. Add to the end of the sentence , giving a final concentration of | 7o
acetone in the assay.
line 4. Add to the end of paragraph. Recovery of radiolabel was in the
range of 90 to L00 7o of. that applied in all cases.
Iine 24. Change to: To test the assumption...
line 12. Change "as" to \ilas.
line2l. Change to: Metabolism occurred in both the meristematic regions (Figure
4.4) and in the leaves (data not shown).
figure legend. Change to: ...extracted from the meristematic region, 0 to 180 h
after treatment.
Page 81,

Page 82, figure legend. Change to: HPLC elution profiles of extract, frorr, 1I"
meristematic region of susceptible ...
Page 92, figure legend. Change to: Percentage of radioactivity extracted from the
meristematic region as diclofop-methyl ...
Page 109, table title. Change to: The percentage of radioactivity extracted from the
meristematic region as ¡14çrd'"lofop acid ...
Page 1 lT,line 22. Change to: ...possessing an ACCase that is 3- to 8-fold less resistant.
Page l22,line 7. Change to: In L. rigidum, biotype SLR 31 has at least two mechanisms of
resistance, enhanced metabolism to detoxify diclofop-metþl and resistant ACCase,
as well as an alteration of the plasma membrane that has been correlated with
resistance (Haüsler et al. 1991, Holtum et a1.,1991, Tardif and Powles 1994).
Page t24,line 5. Change "absorption" to adsorption.
Page 127,line 21. Change "absorption" to adsorption.
Page l29,line 17. Change "diclofop-methyl" to diclofop.
Page 130, Figure legend. Change "[14C]diclofop" to radiolabel.
Page 132,line22. Change "diclofop-methyl" to diclofop.
line 25. Change "diclofop-methyl" to diclofop.
Page 135, line 7 . Change "resistance" to action.
Page [49,line23. Change "have" to has.
Page 153, line 16. Change to: ...where three mutant alleles of the ACCase structural
gene...
Page 156, line 25. Change to: The recovery of membrane polarity is dependent on the pH...
Page 157 ,line l. Change "documented as a mechanism of resistance" to correlated with
resistance.
Iine 2. Change "appeared" to apparent.
line 27. Change "has" to have.
Page 158,line 3. Change "could" to can.
Page 159, line 2. Change "Physiological" to physiological.
Page 160,line 15. Change "herbicide" to Herbicide.
Page l7O,line 7. Change "ishikawa" to Ishikawa.

..""' - ,,-*"*.*"'*l
' Lr+.qliË {},¿,i,,{Pli$ L!ffi{p,nY :
';'iiã tl|,åTF:[1¡ii]]l tf: li,'[L/,i¡i ,Chapter 1
Introduction
1.1 History of Herbicide Resistance
The exposure of a weed population to persistent herbicide application can lead to the
occrurence of herbicide resistance in that population. The first report of herbicide resistance
was in a biotype of. Senecio vulgaris L. from Washington State, USA, in 1968 which was
no longer connolled by the photosystemtr-inhibiting herbicide simazine (Ryan 1970). This
biotype appeared in a conifer plantation where simazine had been sprayed repeatedly for
several years. By t914,triazine-resistant weeds were documented as a widespread problem
in maize fields of north-west'Washington and throughout the Pacific North West, from
California to British Columbia. Later, triazine resistant weeds became cortmon in eastern
Europe, Israel and Australia (Ritter 1989). Now more than 57 weed species (40 dicots and
17 monocots) have been identified as triazine resistant (Holt et at. 1993).
Resistance to some other early and widely-used herbicide groups has developed more
slowly. For example, resistance to the bipyridiliums (paraquat and diquat), released in
1960, has been identified in 18 weed species (Preston L994). Purba (1993) noted that
resistance to paraquat and diquat has evolved slowly compared to the other groups of
herbicides. Resistance to dinitroanilines has been identified in 4 weed species following
annual applications of herbicides for at least l0 years (Smeda and Vaughn 1994).
Resistance to auxin analog herbicides, released in 1940s, has been documented in 8 species
following repeated herbicide application (Coupland 1994).
Recently, resistance to herbicides inhibiting acetyl coenzyme A carboxylase (ACCase) and
acetolactate synthase (ALS) has become widespread. Resistance to ACCase-inhibiting
herbicides (aryloxyphenoxypropionate and cyclohexanedione) has been reported in nine
grass weeds (Devine and Shimabukuro L994). Resistance to AlS-inhibiting herbicides

2
(sulfonylureas, imidazolinones, triazolopyrimidines and pyrimidinyl thiobenzoates) has
evolved in 13 weed species in North America and Australia (Saari et al. 1994). These
herbicides have only been in use for less than two decades yet resistance to these herbicides
has been evolved more rapidly than to other herbicide groups. Thus, increased use of these
herbicides will probably increase the number of resistant weeds developing in the future.
1.2 Herbicide Resistance in Australia
The fust report of herbicide resistance in Australia was a diclofop-methyl resistant biotype of
Lolium rigidum in 1982 following the use of this herbicide in four consecutive years (Heap
and Knight 1982). Subsequently, hundreds of populations of L. rigidum in Australia have
developed resistance to diclofop-methyl and some are cross-resistant to different classes of
herbicides which have a different mode of action (Heap and Knight 1990). To date, eleven
weed species (seven monocots and four dicots) have developed resistance to ten classes of
herbicides with six different modes of action (Table 1.1).

3
Table 1.1 Herbicide-resistant weed species in Australia
Species Class of herbicide (site of action) Reference
Arcøtlvca calenùtla
Avenafatrn
Avena sterilß
Brassica touneþrtü
Cyperus diformis
Hordeum gbucum
Hordeum leporinum
Lolium rigidum
Sisymbrium orientale
Sonchus oleraceus
Bipyridilium (photosystem I)
Aryloxyphenoxypropionates (ACCase)
Aryloxyphenoxypropionates (ACCase)
Sulfonylurea (ALS)
Sulfonylurea (ALS)
Bipyridilium (photosystem I)
Bipyridilium (photosystem I)
Aryloxyphenoxypropionate (ACCase)
Cyclohexanedione (ACCase)
Dinitroaniline (microtubule forrration)
Phenylurea (photosystem II)
Sulfonylurea (ALS)
Triazine (photosystem tr)
Triazinone (photosystem tr)
Triazole (pigment biosynthesis)
Imidazoünone (ALS)
Sulfonylurea (ALS)
Imidazolinone (ALS)
Sulfonylurea t.qlSl
Triazolopyrimidine (ALS)
Bipyridilium (photosystem I)
Powles et al. 1989
Mansooji e¡ al. 1992
Mansooji et al. 1992
Boutsalis and Powles 1995a
Gratram et al. unpublised
Powles 1986
Tucker and Powles 1991
Heap and Knight 1982
Heap and lftright 1990
McAlister et al. 1995
Burnetetal. l99l
Christopher etal. t992
Burnetetal. l99l
Burnet et al. l99l
Burnet et al. l99l
Boutsalis and Powles 1995a
Boutsalis and Powles 1995a
Vulpiabromoides Purba et al. 1993b

4
1.3 Mechanisms of Herbicide Resistance
To be effective, a herbicide must enter the plant, reach the target site at a sufficient
concentration and be maintained there for a sufficient period to kill the plant. Changes in
morphological, physiological and/or biochemicat processes in plants that exclude herbicide
from the tatget site can lead to herbicide resistance. Possible mechanisms of resistance to
herbicides are discussed below.
1.3.1 Reduced Herbicide Uptake and Translocation
Herbicide can be prevented from reaching the target tissue by reduced uptake and
translocation. To date, there have been no reports of resistance due to reduced absorption of
herbicide into resistant weed biotypes. Examples of reduced translocation of herbicide
conferring resistance are given in Table 1.2.
Table 1.2 Reduced franslocation as a mechanism of resistance in weeds
Species Herbicide Reference
Arctothcca calendula diquat
Conyza bonariensis paraquat
Erigeron canadensis pa¡aquat
Erigeron philadelphicus pa¡aquat
Hordeum glaucum paraquat
Preston etal.L994
Fuerstet al. 1985
Tanaka et al. 1986
Tanaka et al. 1986
Bishop et al. 1987
Preston etal.t992
Preston etaI. t992Hordeum leporinum paraquat

5
1.3.2 Enhanced Herbicide Metabolism
After entering the plant, the herbicide may be metabolized within the cell to an inactive
product. Changes that enhance metabolism of the herbicide within the plant, may result in a
reduction in the amount of the active form of herbicide. Several weed species have evolved
resistance to herbicides due to an increased capacity to detoxify the herbicide into less toxic
compounds as shown in Table 1.3.
Table 1.3 Enhanced metabolism of herbicide as a mechanism of resistance in weeds
Species Herbicide Reference
Abutilon theophrasti atazine Gronwald et al. 1989
Alopecurus myosuroides chlorotoluron Moss 1990
Alopecurus myosuroides diclofop, fenoxaprop Hall et al. 1993
Lolium rigid.um diclofop-methyl Holtum et al. 1991
Loliurn rigi.dum PS tr inhibitors Burnet et al. 1993a and 1993b
Inlium rigidum chlorsulfuron
Loliutn rigidum
Stellariamedia
Ealkoxydim
mecoprop
Christopher et al. 1991
Cotterman and Saari 1992
Preston et al. unpublished
Coupland et al. 1990

6
1.3.3 Modified Site of Action
When the herbicide reaches the target site in a sufficient concentration to inhibit the
biochemical target site, a modification of the target site may reduce the herbicidal effect and
provide resistance. Reduced sensitivity of photosystem II, ACCase and ALS are
documented as target site mechanisms of resistance. Most of the 57 species with resistance
to PS tr-inhibiting herbicides have a target site change which renders them resistant to the
triazine herbicides. A single amino acid change (ser 264 to gly) in the Qs-binding site
within the D1 protein of PS II is responsible for resistance in these plants (Gronwald 1994).
Examples of resistant species exhibiting reduced sensitivity of herbicides at the target sites
ACCase and ALS are shown in Tables 1.4 and 1.5, respectively.
Table 1.4 Reduced sensitivity of ACCase enzyme as a mechanism of resistance in plants
Species Herbicide Reference
Festucarubra APP, CHD Stoltenberg et al. 1989
Festtrca ovina
Festuca amethystina
APP, CHD Catanzaro et al. 1993
Lolium multiforum diclofop-methyl Gronwald et aL. 1992
I-oliutn rigidum APP, CHD Tardif et al. 1993
Tardif and Powles 1994
Setarinviridis APP, CHD Ma¡les et al. 1993
Tzamays haloxyfop,sethoxydim Parkeretal. 1990a

7
T¡ble 15 Insensitive ALS enzyme as a mechanism of resistance in weeds
Species Refercnce
Brassicatourneþrtü BoutsalisandPowlesunpublished
Kochia scoparia Saarietal.1990
Loliwn perenne Saa¡i et al.1992
Lolium rigidum Christopher etal.1992
Salsola iberica Saari et aL.1992
Sisynbrium orientale Boutsalis and Powles unpublished
Sonchus oleraceus Boutsalis and Powles 1995b
Stellariamedia Devine et al. 1991
Xanthium strumarium Schmiøer et al. 1993
In addition, some resistant weed species containing a sensitive target enzyme ACCase
exhibit altered membrane function. This is manifest as a recovery of the plasma membrane
electrogenic potential following diclofop acid-induced depolarization (Holtum et al. 1991,
DiTomaso 1993, Devine et al. 1993). Membrane depolarization has been proposed as a
second site of action of ACCase-inhibiting herbicides (more detail in section 1.7).
Therefore, repolarization of the plasma membrane potential following removal of herbicide
has been proposed to be a mechanism of resistance to these herbicides in some weed species
(Table 1.6).

8
Table 1.6 Alæred plasma membrane response as a mechanism of resistance in weeds
Species Herbicide Reference
Avenafatua
I¡Iiutn rigidum
Loliumrigidum
APP Devine et al. 1993
APP, CHD Häusler et al. 1991
Holtum et aI. 1991
DiTomaso 1993
APP Shimabuhuo and Hoffer 1992
1.3.4 Sequestration of Herbicide within Plant Cells
Sequestration is another possible mechanism of herbicide resistance. Holtum et al. (1991)
proposed that resistant L. rigidurø plants which were able to grow in the presence of
substantial pools of diclofop acid in the tissue could do so because the toxic form is
sequestered in the vacuole or in the apoplastic space. Under this scenario, the concentration
of diclofop acid at the active site is too low to substantially inhibit ACCase activity or
depolarize the plasma membrane potential. However, this phenomenon has yet to be
proved.
1.4 Inheritance of Herbicide Resistance
In most cases of resistance, inheritance of resistance is conferred by a single gene. In the
case of triazine resistance, inheritance is usually maternal, whereas in other cases of
resistance it is often a partially-dominant, nuclear gene as shown in Table 1.7. Resistance to
dinitroaniline herbicides in Setaria viridis is an exception and is due to a single recessive

9
gene (Jasieniuk et aI. 1994). However, resistance to ALS inhibitors in some biotypes of
Loliutn rigidwt in Australia is controlled by at least ttto genes (Christopher 1993).
Table 1.7 Inheritance of herbicide resistance in weed biotypes
Herbicide Species Number of gene Reference
a¡razme Abutilon theophrasti I partially-dominant Anderson and Gronwald 1987
cblorsulfuron Sonchus oleraceus I partially-dominant Boutsalis and Powles 1995b
chlortoluron Alopecurus myosuroides 2 addiúve Chauvel l99l
diclofop Lolium multiþrum l partially-dominant Betts et al.1992
fenoxaprop Avena sterilis 1 partially-dominant Barr et al. 1992
fluazifop Avena sterilis 1 partially-dominant Barr et al.1992
haloxyfop Lolium rigidum 1 partially-dominant Ta¡dif et al. submined
metsulfuron l-actuca seniala 1 partially-dominant Mallory-Snith et al. 1990
paraquat Erigeroncanadensß l dominant Itoh and Miyahara 1984
paraquaf Conyzt bonariensis l dominant Shaaltiel et al. 1988
pa¡aquat Hordeum glaucum I partially-dominant Islam and Powles 1988
praquat/diquat Arcthotheca calendulo I partially-dominant Purba et al. 1993a
paraqur Hordeum leporinum I partially-dominant Pu¡ba et al. 1993a
siduron Hordeum jubatum 3 complementary Shooler et al. 1972
trifluralin Setariaviridis I recessive Jasieniuk er al. 1994

l0
1.5 Aryloxyphenoxypropionate and Cyclohexanedione Herbicides
ACCase-inhibiting herbicides are represented by two classes of herbicides: the
aryloxyphenoxypropionate (APP) and substituted l,3-cyclohexanedione (CID) herbicides.
These are stn¡cturally different but exhibit simila¡ ph¡otoxicity and symPtomology. The
chemical structure and common name of these herbicides are given in Table 1.8. APP and
CHD herbicides are sometimes called grarninicides because they selectively control grass
weeds in dicot crops (Gronwald 1991). Diclofop-methyl, fenoxaprop-ethyl and
tralkoxydim are used for selective post-emergent wild oat control in wheat, barley, rye and
dicot crops. Fluazifop-butyl, haloxyfop-ethoxyethyl and sethoxydim are widely used as
selective post-emergent herbicides in dicot crops. Better confrol of grass weeds is obtained
when these herbicides a¡e applied at the two to th¡ee leaf stage (Ashton and Monaco t99I,
Harker and Blackshaw 1991). The site of action of APP and CHD herbicides is mainly in
the meristematic regions of the susceptible plants (Hoerauf and Shimabukuro 1979, Hosaka
et al. 1984, Hosaka et at. 1987). Injury symptoms produced by these graminicides on
susceptible plants are necrosis in the meristematic tissue, necrosis or chlorosis in developing
leaf tissue and appearance of purple leaf colour due to anthocyanin accumulation (Hoerauf
and Shimabukuro Ig7g,Ishigawa et al. 1985, Köcher et aI. 1982, Swisher and Corbin
1982, Asare-Boamah and Fletcher 1983). General usage, mode of action, uptake,
translocation and metabolism of APP (diclofop, fluazifop and haloxyfop) and CHD
(sethoxydim and tralkoxydim) herbicides will be discussed in the following sections.
The APP herbicides are chiral due to an asymetrical carbon atom in the propionic side chain.
These herbicides are usually a racemic mixture of two isomers, R(+) and S(-) (Duke and
Kenyon 1988). The R-enantiomer is the herbicidally-active form (Dicks et al. 1985,
Uchiyama et al. 1986, Harwood et al. 1987, Gerwick et al. 1988, Secor et al. 1989). The
APP herbicides a¡e formulated as esters of the parent acid to facilitate penetration into the
leaf cuticle. After entering the plant, the ester form is hydrolysed rapidly to the phytotoxic

1t
Table 1.8 Structure of graminicides
(a) Aryloxlphenoxlpropionate (APP) herbicides
F'o&R1-
R1 Rz Common tame
CH¡ diclofop-methyl
\)- CzHs fenoxaprop-ethyl
c¿Hs fluazifopbutyl
cH¡ haloxyfop-methyl
czHs quizalofopethyl
(b) Cyclohexanedione (CtD) herbicides
o-R¿R1
R1 R2 R3 R4 Commonname
l"' CzHs CH2CIFCH clethodimC2H5S-C-CH2- H2
H2
H2
H2
H
CtHt CzHs cycloxydim
CtHt CzHs sethoxydim
cHr
c¡I
S-r(}
1",qH5rc-cH2-H
cHs
CH¡
CzHs CzHs Ealkoxydim

12
carboxylic acid (Shimabukuro et al. 1979, Stonebridge 1981, Duke and Kenyon 1988,
Harwood et al. 1989) which is then translocaæd to meristematic regions (Buhler et al. 1985,
Hendley et al. 1985, Chandrasena and Sagar 1987 and 1989).
1.5.1 Antagonism of APP and CIID Herbicides
The general lack of activity of APP and CHD herbicides on dicot weeds makes mixtures
with other herbicides important in order to broaden the spectrum of weed control.
However, auxinic compounds and some other herbicides can antagonise the efficacy of APP
and CHD herbicides when applied as a tank-mixture. For example, addition of 2,4-D
dimethylamine reduced the efficiency of diclofop-methyl on wild oat (Todd and Stobbe
1980), fenoxaprop and haloxyfop on Sorghum halepense (Mueller et al. 1989 and 1990),
and tralkoxydim on wild oat (Jensen and Caseley 1990). In most cases, 2,4-D reduced
uptake and/or translocation of those herbicide into plants. Dicamba can also antagonise the
effect of haloxyfop-methyl onFestuca arund.inaceø (Aguero-Alva¡ado et al. 1991).
Bentazon is another compound that can antagonize the effects of APP and CHD herbicides.
Penetration of haloxyfop into Setaria lutescens leaves was reduced3TTo in the presence of
bentazon due to crystallization of bentazon on the leaf surface, thus forming a physical
barrier that impaired the penetration of other xenobiotics through the cuticle (Gerwick 1988).
Aguero-Alvarado et al. (1991) found that bentazon alone or mixed with haloxyfop
significantly inhibited ACCase activity only at high concentrations. For example, l0 mM
bentazon was required to reduce ACCase activity of a susceptible grass (F. arundinøcea)by
40Vo. They suggested that antagonism may be due to mechanisms that reduce the amount of
haloxyfop reaching the target site. In addition, bentazon reduced the uptake and
translocation of tralkoxydim resulting in a reduction of wild oat control (Jensen and Caseley
1990).
1.5.2 Uptake and Translocation
The ester forms of APP herbicides are readily absorbed by leaves and roots. Tritter et al.
(1987) found that uptake of diclofop-methyl was ten times greater than diclofop acid into oat

13
protoplasts over the same period. They suggested that this might be due to the acid group of
diclofop being dissociated at physiological pH giving a charged molecule which would be
less penneable to plant membranes and result in reduced uptake compared to diclofop-
methyl which is less polar. Uptake of [lag¡Ooazifop-butyl was more rapid in young leaves
compared to older leaves (Ca¡r 1985). Absorption of fluazifop was greater at 30'C to 35'C
than at 18'C to 20'C (Ca¡r 1985). Cycloxydim absorption by suspension-cultured cells of
Abutilon theophrasti was significantly greater at2O'C compared with 4 and 12"C, but no
difference was observed between the two lower temperatures @gyneberg et al. 1994).
Translocation of APP and CHD herbicides varies between individual herbicides.
Translocation of diclofop-methyl in plants is limited. A number of studies have established
that less than 5Vo of absorbed [14C]diclofop-methyl is translocated out of the treated zone in
a range of species (Brezeanu et al. 1976, Boldt and Putman 1980, Hall et al. t982,
Jacobson and Shimabukuro t982, Dahroug and Muller 1990, Baker and Chamel 1990,
Devine etal.1992). The predominant tanslocated forms are diclofop-methyl and diclofop
acid (Jacobson and Shimabukuro 1982). Diclofop was found to move mainly in an
acropetal direction when applied to different parts of wild oat plants (Friesen et al. 1976).
Beckett et al. (1992) found that translocation of ¡l+grn rizalofop out of the treated area was
limited and translocation occurred both acropetally and basipetatly in similar arnounts.
Fluazifop acid or its glucose conjugated products a¡e the mobile forms of fluazifop (Hendley
et al. 1985, Carr et al. 1985). Fluazifop is readily translocated from treated leaves to
rhizomes and stolon of perennial grasses and accumulates in meristematic tissue
(Stonebridge 1981, Ca¡r et al. 1985, Chandrasena and Sagar 1987). Fluazifop moves in the
phloem with the assimilate stream along 'source-sink' relationships of the plants (Kell et al.
1984, Hendley et al. 1985, Chandrasena and Sagar 1987). As a result, applications of
[l4C]fluazifop to mature leaves resulted in large quantities of radioactivity accumulating in
lower stem regions, roots and rhizomes of Agropyron repens, compared with treatments to
younger leaves (Chandrasena and Sagar l9S7). In addition, translocation of [l4C]fluazifop

t4
was greater from lamina base applications than from treatments to the lamina apex and
middle (Chandrasena and Sagar 1987).
Little translocation of l4C out of the treated leaf occurred following [l4C]traloxyfopmethyl
application to the youngest leaf of susceptible and tolerant species @uhler et al. 1985).
These authors also found that highly susceptible Sorghum bicolor translocated more 14C
than did the susceptible Setaria glauca. Following llaC]haloxyfop-methyl application to
leaves of a stolon node of Cynodon dactylon less than 37o of the radiolabel translocated to
the apex and base (Marorder et al. 1987).
CHD herbicides a¡e absorbed rapidly into plants and translocated to the target site. Rapid
uptake of sethoxydim occurred in the first 3 h and by 6 h more than half of heöicide applied
had entered wild oat leaves (Smith and Vanden Born 1992). Absorption of
[laC]cycloxydim by suspension-cultured velvetleaf (Abutilon theophrasti) cells was
dependent on concentration, temperature and pH (Rgyneberg et al. 1994). Absorption of
cycloxydim was linea¡ly related to the extracellular herbicide concentration. Cycloxydim
absorption was significantly greater at2O'C compared to at 4 or l2'C. Cycloxydim was
absorbed >5.5 fold at pH 3.7 compared to pH 5.7 (Røyneberg et al. L994). Ammonium
sulfate can increase the efficiency of sethoxydim by increased absorption and translocation
in Digitaria sanguinalis, Hordeum vulgare and,Avenafatua (Jordanet al. 1989, Smith and
Vanden Born 1992).
1.5.3 Metabolism
APP herbicides are absorbed into plants as an ester form and then are hydrolyzed to the
ph¡otoxic acid form. Grass crops such as wheat rapidly metabolize some APP herbicides
to inactive products (Shimabukuro et al. 1979 and 1987). However, wheat is susceptible to
fluazifop, haloxyfop and some other APP herbicides because it is unable to detoxify these
herbicides. The metabolic pathway for diclofop-methyl in resistant (e.g. wheat) and
susceptible plants (e.9. wild oat, muze and other susceptible grasses) is shown in Figure
1.1. In susceptible species, the phytotoxic diclofop acid is mostly converted to the neutral

15
glycoester which can be hydrolyzed back to the toxic form (Shimabuku¡o 1990). In wheat
diclofop acid undergoes aryl-hydroxylation by a cytochrome P450 monooxygenase
(Zimmerlin and Durst 1992). Subsequently, most of the hydroxylated diclofop is
conjugated rapidly to glucose to form the aryl-O-glucoside metabolite (Shimabukuro et al.
1987).
Metabolism of fenoxaprop-ethyl was studied in soybean (Wink et al. 1984) and grass
species (Lefsrud and Hall 1989, Tal et al. 1993). In soybean, fenoxaprop-ethyl is rapidly
hydrolyzed to the ph¡otoxic form, fenoxaprop acid, which is subsequently metabolized to
polar conjugates $fink et al. 1984). Tal et al. (1993) proposed that fenoxaprop-ethyl is
detoxified by glutathione, cysteine and glucose conjugations in some grass species. In
wheat (Triticum aestivum), barley (Hordeum vulgare), crabgrass (Digitaria ischaemum) and
oat (Avena sativa), fenoxaprop-ethyl is rapidly hydrolyzed to fenoxaprop acid (Figure 1.2).
After 8 h of incubation, in susceptible crabgrass and oat, most of the radioactivity was
recovered as the ph¡otoxic form of fenoxaprop acid and only small amounts were convefed
to water-soluble metabolites. In contrast, in wheat and barley, fenoxaprop acid underwent
rapid displacement of the phenyl group by glutathione (GSH) and/or cysteine, resulting in
production of S-(6-chlorobenzoxazole-2-yl)-glutathione (GSH conjugate), 5-(6-
chlorobenzoxazole-2-yl)-cysteine (cysteine conjugate), and 4-hydroxyphenoxy-propanoic
acid. The GSH conjugate may be catabolized to form the cysteine conjugate which was
subsequently metabolized to an unidentified metabolite. The 4-hydroxyphenoxypropionic
acid was further metabolized to form a glucoside conjugate.

l6
o
o
F.,IH
diclofopmethyl
diclofop acid
H
susceptible
COO-Glucose
H
neutral glucose ester of diclofop
CH¡t-
oresistant
ct
o cooHIH
OH
aryl-hydroxylated diclofop
oCH.t-IH
cooH
acidic aryl-O-glucoside of diclofop
Figure 1.1 Metabolism of diclofop-methyl in plants (redrawn from Shimabukuro 1990)

t7
1",O- CH- COOC2H5
CH¡l-
o- cH- cooH
1",o- cH- cooH
4-hydroxylated-propanoic acidS.
cl o co-NH-cH2-cooH
S-(6-chlorobenzoxazole-2-yl)-glutathione
S-
N
cl o
fenoxaproprthyl
o
fenoxaprop acid
,l
cooH
l"'o- cH- cooH
N
cl
NHnl'
N NH-CO-CHr-CHI-CH-COOHl-
CHz-1
H
pO-glucoside conjugate
NHr
cHz-lN
ocl
S-(6-ctrlorobenzoxazole-2-yl)-cysteine N-glucoside
unidentified metabolite
Figure 1.2 Metabolism of fenoxaprop-etþl in wheat, barley, crabgrass and oat (redrawn
from Tal et al. 1993)

l8
Less information is available on the metabolism of CHD herbicides in plants.
Cyclohexanediones are unstable compounds in both biotic and abiotic systems (Swisher and
Corbin 1982, Røyneberg et al. 1994). Decomposition of sethoxydim is enhanced by
alkaline conditions (Shoaf and Carlson 1986) and ultraviolet light (Shoaf and Ca¡lson
1992). Sethoxydim decomposes in water, especially at basic pH, leading to generation of at
least 14 possible degradation products (Shoaf and Carlson 1986). The metabolism of
sethoxydim in plants is rapid and most of the parent herbicide is degraded within 6 to 72 h
after application (Campbell and Penner 1985, Hosaka et al. 1987, Ishihara et al. 1988,
Tardif et al. 1993). Campbell and Penner (1985) found six major breakdown products of
sethoxydim and two of them were speculated to be toxic forms. However, it is unclear
whether sethoxydim or its breakdown products are the actual herbicidal agent. The
metabolism of tralkoxydim was studied in wheat gfolvn under field condition and sprayed at
a rate of 345 g ai ha-l (Hadfield et al. 1994). In wheat, the metabolism of tralkoxydim
involved hydroxylation of the p-methyl function group of the a¡omatic ring (Figure 1.3).
This oxidation may occur in conjunction with either transformation of the
ethoxy(imino)propyl function or with oxidative cleavage of the cyclohexanedione ring. All
the initial products were then conjugated to a range of endogeneous sugars. Although there
is no conclusive evidence, it is likely that the selectivity of tralkoxydim between susceptible
and resistant grass species is based on different rates of herbicide metabolism.

/CH
cH2 - co2H
CHO -COOHcll3
3-(2,4,Gtimethylphenyl)pentanedioic acid
l9
QHs
NOC2H5
CzHs
cH¡ o
2-( I -(ethoxyimino)propyl)-3-hydroxy-S-(zl-hydroxymethyl-2,Gdimethylphenyl)cyclohex-2-enone
cH3 o
Tralkoxydim
CuHs
N
cH3
2- ethyl-G (4-hydroxymethyl-2, 6-dimethylphenyl)-4,5,6,7-tetrahydrozoxazol-4-one
HO HorcH2 cH2
OH
NI{
czHs
cH¡ o
3-hydroxy-2-( 1 -iminopropyl)-5-(a-hydroxymethyl-2,6-dimethylphenyl)cyclohex-2-enone
cH, - co,H/
CH
CH¡cH2 - co2H
3-(zt-hydroxymethyl-2,Gdinethylphenyl)pentanedioic acid
Figure 1.3 Metabolic pathway of tralkoxydim in wheat foliage and straw (redrawn from
Hadfield et al. 1994)
"ol
Hol
cH.
cHz

20
1.6 Mode of Action of APP and CHD Herbicides
Two basic mechanisms of action have proposed for both APP and CHD herbicides. One is
a biochemically-based mechanism involving the inhibition of acetyl-CoA carboxylase
(ACCase) and the second is a biophysically-based mechanism involving a perturbation of
the transmembrane proton gradient across plasma membrane (Figure 1.4).
PlasmalemmaInside
ATP
Outside
IT+.ATP
ADP+Pi
ÂpH
PlastidsH+ channel (?)
¿tH'1
¿H.
I
tH'1
diclofop-methyl
diclofop
and/or diclofop-methyl
Em
G120mV) (0mV)
Figure 1.4 A diagram of biophysical and biochemical mechanisms of action of APP
herbicides (redrawn from Devine and Shimabukuro 1994)
diclofopI +
+
+
+
+
+
IAA,2,4-D
TTITITTTTTTTI
acetyl-CoA
malonyl-CoA
()ØCÚ
UU
+

2l
1.6.1 Inhibition of Fatty Acid Biosynthesis
Fatty acid biosynthesis in plants occrus exclusively in plastids (Harwood 1988). ACCase
(acetyl-coerizyme A:bicarbonate ligase [ATP], E.C. 6.4.1.2), a biotin-containing ewzymø
catalyzes the first step in fatty acid biosynthesis via ATP-dependent carboxylation of acetyl-
CoA to fonn malonyl-CoA (Harwood 1989). Plant ACCase is a high molecular weight,
multifunctional enzyme comprised of three catalytic domains, biotin carboxylase, biotin
carboxyl carrier protein and carboxyltansferase domains (Figure 1.5). Initially, a carboxyl
group is donated from a bica¡bonate anion, and ATP hydrolysis is used to a ca¡boxybiotin
intermediate by biotin carboxylase. Carboxybiotin is attached to an e-amino group of a
lysine residue on the biotin carboxyl ca¡rier protein (BCCP). Carboxybiotin then functions
as a CO2 donor in malonyl-CoA formation (Harwood 1989). Malonyl-CoA is an
intermediate in a number of metabolic pathways, such as the biosynthesis of cuticular waxes
and flavonoids (Harwood 1989). The condensation of acetyl-CoA and malonyl-ACP
catalyzedby 3-ketoacyl-ACP synthase is the fust step in a series of reactions in the fatty acid
synthase complex that results in 16:0-ACP and l8:I-ACP as major products of fatty acid
synthesis in plastids (Browse and Sommerville 1991). Phospholipids are synthesized at the
endoplasmic reticulum through initial acylation of glycerol phosphate by acyl-CoA
transferase.
ACCase is the target site of the aryloxyphenoxypropionic acid and the cyclohexanedione
herbicides in susceptible grass species (Burton et al. 1987 and 1989, Rendina et al. 1988
and 1989, Secor and Cséke 1988, Secor et al. 1989, Walker et al. 1989, Howard and
Ridley 1990). Generally, ACCase from monocots is sensitive to both classes of herbicides
and that of dicots is not (Cho et al. 1986, Burton et al. 1987, Litchtenthaler and Kobek
1987, Kobek et al. 1988, Rendina and Felts 1988, Rendina et al. 1988, Secor and Cséke
1988, Walker et al. 1989, Boldt and Barett 1991). However, some monocot crops such as
wheat are tolerant to diclofop and fenoxaprop despite containg a herbicide-sensitive
ACCase, as a result of rapid metabolism of these herbicides to nontoxic products
(Shimabukuro et al. 1979, Lefsrud and Hall 1989).

22
acetyl-CoA carboxylase
biotin biotin carboxyl BCCP:acetyl-CoAca¡rier protein fianscarboxylase
(BCCP)carboxylase
aryloxyphenoxypropionic acids
cyclohexanediones
MSATP
biotin \ carboxylbiotinbiotin-CO,
malonyl-CoA
"4 biotin
/pyruvate acetate
malonyl-ACPoCË
ÊøEo6!
àd
J3-ketoacyl-ACP
+l6:GACP
+18:O-ACP
Plastid vl8:1-ACP
vacyl-CoA
phospholipidsacyl-CoA transferase
Figure 1.5 Schematic of fatty acid biosynthesis in plants
glycerol-3-
reticulum

23
1.6.2 Characteristics of ACCase Enzyme
ACCase is found in plastids and is active over a wide pH range with an optimum at pH 8.0
in purifred plastids of developing endosperm of castor seeds (Finlayson and Dennis 1983)
and with optimum at pH 8.2 in soybean seeds(G/ycine max L. Men.) (Charles et al. 1986).
A divalent cation, Vtn2+ or Mg2+, is necessary for its activity @nlayson and Dennis 1983),
but Mn2+ could not replace Mg2+ as an essential activator of ACCase in soybean (Charles et
al. 1986). Based on experiments on the utilization of acetate by chloroplasts in the light and
dark, the biosynthesis of fatty acids from acetate in chloroplasts was induced by light
(Nakamura and Yamada 1975) with little or no accumulation of fatty acids in leaves in the
dark (Browse et al. 1981). Nakamura and Yamada (1979) proposed that the carbon flow
for fatty acid synthesis is limited at the step of ACCase in a light-dependent reaction.
Moreover, the kinetic properties of this enzyme in maize chloroplasts demonstrated that its
activity may be modulated by light-dependent changes of metabolite levels, particularly by
alterations in the stromal concentration of ATP, ADP, Mg2+ and by pH (Nikolau and
Hawke 1984). During seed maturation, the level of ACCase markedly increased just before
the onset of major lipid accumulation (Turnham and Northcote 1983).
There are two types of ACCase: prokaryotic ACCase, in which the three functional
domains-biotin carboxylase (BC), biotin carboxyl carrier protein (BCCP) and
carboxyltransferase (CT) a¡e located in separable proteins (as found in Escherichia coli and
probably pea chloroplast) (Li and Cronan l992aa¡d 1992b, Alban et al. 1994, Konishi and
Sasaki 1994), and the eukaryotic ACCase, in which all the domains a¡e located on one large
polypeptide (as found in rat, chicken, yeast, diatom and wheat) (I-ópez-Casillas et al. 1988,
Takai et al. 1988, Al-Feel et al. 1992). In plants, both eukaryotic and prokaryotic forms
have been found (Kannangara and Stumpf 1972, Nikolau and Hawke 1984, Egli et al.
1993, Sasaki et al. 1993, Alban et al. 1994, Konishi and Sasaki 1994).
Plants contain two isoforms of ACCase that appear to have different subcellular localization
and metabolic roles. A major isoform, which is responsible for the bulk of the cell ACCase
activity, is located in the chloroplasts and other plastids, whereas a minor isoform is

24
presumably found in only the cytosol @gli et al. 1993, Alban et al. 1994, Ashton et al.
1994, Konishi and Sasaki 1994). The major isoform plays a key role in fatty acid
metabolism and the minor isoform is involved in plant defence against pathogens and
elongation of very long chain fatty acids (Shorrosh et al. 1994).
Recent evidence indicates that the types of ACCase present in dicotyledonous species are
fundamentally different to the types present in monocot species. Pea contains a eukaryotic
ACCase in the cytoplasm but a prokaryotic multi-subunit ACCase in the chloroplasts (Alban
et aL. t994, Konishi and Sasaki 1994, Sasaki et al. 1994). In contrast, grasses contain
eukaryotic ACCase in both the plastids and cytoplasm (Egli et al. 1993, Ashton et al. 1994).
In young pea leaves, a minor eukaryotic ACCase (-20Vo of the total ACCase activity in the
whole leaf), located in the epidermis, \r,as strongly inhibited by diclofop (Alban et al. 1994).
In contrast, a major prokaryotic ACCase, present in both epidermis and mesophyll, was
highly resistant to herbicide (Alban et al. 1994). Konishi and Sasaki (1994), working with
pea, wheat and rice, found that prokaryotic ACCase \ryas located in the plastids and the
eukaryotic ACCase was presumably in the cytosol. The prokaryotic ACCase was
insensitive to ACCase-inhibitng herbicides whereas the eukaryotic ACCase was sensitive to
herbicide inhibition. Pea is resistant to ACCase-inhibiting herbicides since it contains the
prokaryotic, herbicide-insensitive form of ACCase. In contrast, wheat and rice are
susceptible to ACCase-inhibiting herbicides because they lack the prokaryotic ACCase
(Konishi and Sasaki 1994). This probably explains why grasses a¡e sensitive to APP and
CHD herbicides, whereas diclots are susceptible.
In grass species like maize, a herbicide-insensitive form of ACCase firstly repofed by
Howa¡d and Ridley (1990) has been observed (Egli et al. 1993, Ashton etal.1994} Egli et
al. (1993) found that a major isoform of ACCase (ACCase I), located in most tissues, was
strongly inhibited by haloxyfop and sethoxydim whereas a minor isoform (ACCase II), not
found in mesophyll plastids, was insensitive to those herbicides. Ashton et aI. (1994) also
found that a minor form of ACCase from maize leaves was insensitive to fluazifop and
talkoxydim while the major ACCase form was strongly inhibited.

25
ACCase from susceptible and APP herbicide resistant biotypes of Lolium multiþrumwas
purified and characteizedby Evenson et al. (L994). They found that both susceptible and
resistant forms of ACCase were simila¡ in subunit molecular mass, pH optimum and
substrate kinetics. However, the resistant form of ACCase exhibited altered kinetics of
diclofop binding and a different isoelectric point. They speculated that the ACCase fraction
from the resistant biotype contains both a diclofop-susceptible and -resistant forms.
1.6.3 Kinetics of ACCase Inhibition by APP and CHD Herbicides
The kinetics of inhibition of ACCase by APP (diclofop, fluazifop, haloxyfop) and CHD
(clethodim and sethoxydim) herbicides are linear, reversible and non-competitive with
respect to all three substrates, acetyl-CoA, MgATP and bicarbonate (Rendina and Felts
1988, Rendina et al. 1989, Howa¡d and Ridley 1990, Rendina et al. 1990, Burton et al.
1989 and l99l). Kinetic analyses suggested that APP and CHD herbicides are mutually
exclusive inhibitors of ACCase erzyme and both classes of herbicides compete for the same
binding site (Rendina et al. 1989, Rendina et al. 1990, Burton et al. 1991). In addition,
APP and CHD herbicides inhibit the transcarboxylation rather than the biotin carboxylation
reaction (Rendina et al. 1990, Burton et at. 1991).
1.6.4 Inheritance of Herbicide Resistant ACCase in Formerly Susceptible
Weed Species
Recently, a resistant form of ACCase has been reported in several grass species previously
controlled by ACCase-inhibiting herbicides (Stoltenberg et al. 1989, Betts et aI. t992,
Holtum and Powles 1991, Mansooji et al. 1992. Tardif and Powles 1993) and in maize
(Parker et al. 1990a). In each species, the degree of resistance at the enzyme level is
correlated with resistance at the whole plant level. Inheritance of resistance in wild oat (r4,.
sterilis ssp. ludoviciana) and Italian ryegrass (L. multiþrum) is due to a single, partially-
dominant, nuclear gene (Barr et al. 1992, Betts et al. 1992). Inheritance of resistance to
ACCase-inhibiting herbicides n L. rigidun biotype rWLR 96 is also controlled by a single,
partially-dominant, nuclear gene (Tardif and Powles 1993). Inheritance studies conducted

26
with regenerated plants from graminicide-resistant maize lines selected in tissue culture,
showed that resistance is due to a single nuclea¡ gene er.hibiting partial dominance (Parker
et al. 1990a, Marshall et al. 1992). Among those graminicide-resistant maize lines, th¡ee
mutant ACCase alleles (Accl-S,AccI-HI,Accl-H2) were identified (Ma¡shall et al.
1992). The Accl-S and Accl-H2 alleles conferred target-site cross-resistance to both APP
and CHD herbicides, whereas the AccI-Hl allele conferred selective resistance to the APP
herbicides.
1.7 Disruption of Membrane Function by APP and CHD
Herbicides
The second mode of action of aryloxyphenoxypropionic acids and cyclohexanediones is to
depolarize the plasma membrane potential of susceptible species. Plant cells establish and
maintain a transmembrane proton gradient which is vital to their growth and development.
Any chemical such as diclofop-methyl that disrupts proton electrochemical gradients may
have significant effects on many aspects of the physiology of the plant (Shimabukuro
1ee0).
The free acids of APP herbicides (such as diclofop) depolarize membrane potentials in
Chara spp. (Lucas et al. 1984), parenchyma cells of oat and wheat (Wright and
Shimabukuro 1987), coleoptiles and root tips of A. fatua and Z. rigidum (Häusler et al.
L99t, Holtum et al. 1991, Shimabukuro and Hoffer 1992, Devine et al. 1993). Similar
effects have been found in isolated vesicles of plasma membrane and tonoplast of oat root
(Ratterman and Balke 1988, 1989). Similarly, CHD herbicides such as sethoxydim
depolarize membrane potentials of leaf cells from sensitive grasses (Weber and Lünge 1988,
Weber et al. 1988), and coleoptiles of susceptible and resistant L. rigidum biotypes (Häusler
et al. 1991). The mechanism by which diclofop affects the electrogenic properties of the cell
membrane is poorly understood.

t1
1.7.1 Diclofop as a ProtonoPhore
Diclofop was proposed to act as a specific protonophore that moves protons across the
plasmalemma of plant cells resulting in an influx of protons into the protoplast (Lucas et al.
1984,V/right and Shimabukuro 1987). According to this model, the methyl ester form of
diclofop crosses the plasmalemma and is rapidly hydrolyzed to the free acid form. This
undissociated acid then releases a proton on the c¡oplasmic side of the membrane (Figure
1.4). The deprotonated anionic species then moves back across the plasmalemma to the
apoplasmic side in response to the root cell membrane potential (inside negative) and is
again protonated. This cycle is repeated until the proton gradient is dissipated and
eventually leads to a reduction in cellular ATP levels by short circuiting the proton-
translocating plasmalerlma. Later, Shimabukuro and Hoffer (1992) found that diclofop did
not act like a classic proton ionophore such as z-chlorophenylhydrüzone (CCCP) and
membrane depolarization by diclofop could be prevented by p-
chloromercuribenzenesulphonic acid (PCMBS), a non-permeant molecule that binds -SH
groups on proteins. This suggests that diclofop may interact with a specific plasma
membrane-associated protein to increase proton influx into sensitive cells.
rWhether the membrane depolarization caused by this herbicide leads to phytotoxicity in
plants is the subject of some debate. DiTomaso et al. (1991) found that diclofop acid was
capable of depolarizing the plasma membrane potential of susceptible maize and oat roots, as
well as resistant pea roots. The similar membrane response of resistant pea and susceptible
muze to diclofop acid suggested that the membrane depolarization is not phytotoxic. In
addition, after removal of diclofop acid from the treatment solution, complete repolarization
was observed in both maize and pea root cells (DiTomaso et al. 1991).
1.7.2 Recovery from Diclofop Acid Induced Depolarization of the Plasma
Membrane Electrogenic Potential in Some Resistant Weeds
Evidence that differences in the ability of plasma membranes to recover from depolarization
may be correlated with the resistance responses of some resistant weeds has been found in

28
diclofop-resistant L. rigidum and.á. fatua biotypes (Holn¡m et al. 1991, Häusler et al.
1991, Shimabukuro and Hoffer 1992, Devine et al. 1993). Etiolated coleoptiles and root
tips of foru diclofop-methyl-resistant annual ryegrass biotlpes recovered membra¡e polarity
following exposure to diclofop acid, whereas the susceptible plants did not (Häusler et al.
199L, Holtum et al. 1991, Shimabukuro and Hoffer t992). Although there is a good
correlation between the recovery and resistance, the mechanism producing the difference in
membrane response between the susceptible and resistant biotypes is still unclear.
DiTomaso (1993) suggested that the differential membrane response between susceptible
and resistant L. rigidurn biotypes may be the result of differences in cell wall pH. He also
demonsEated that the repolarization of the membrane potential was pH dependent and use of
a buffered solution could eliminate the differential membrane responses. In solution
buffered at pH 5 or pH 6 (2 mM MES-Tris), both susceptible and resistant L. rigidum
biotypes behaved similarly following diclofop acid-induced depolarization (DiTomaso
1993).
1.8 Resistance to Acetyl-CoA Carboxylase-Inhibiting Herbicides
The first reported case of resistance to ACCase-inhibiting herbicides was a diclofop-methyl
resistant biotype of L. rigi.dum in Ausfralia reported by Heap and Knight (1982). To date, at
least ten species of weeds have become resistant to ACCase-inhibiting herbicides (Table
1.9). Three of these resistant species, L. rigidum,Avenafatua andA. sterilis, a¡e found in
Australia.

29
Table 1.9 Developed resistance to ACCase-inhibiting herbicides in grass species
Species Herbicide Origin Reference
Avenalattn APP, CHD Canada Heap et al. 1993
Avenafatrn APP Austalia Mansooji et al. 1992
Avena sterilis APP Australia Mansooji et al. 1992
Eleusine itúica fluazifop Malaysia Leach et al.t993
Fesucarubm APP, CHD USA Stoltenberg et al. 1989
I-olium multþrum diclofop USA Stanger and Appleby 1989
I-olium rigidum APP, CHD Australia Heap and Knight 1982 and 1986
Alopecurus myosuroides APP, CHD TJK Hall et al.1993
Setariafaberi APP, CHD USA Stoltenberg and V/iederholt 1993
Senriaviridis APP, CHD Anada Ma¡les et al. 1993
Sorghum lulepense APP, CHD USA Barrentine etal. 1992
Smeda et al. 1993

30
1.9 \üild Oat (Avena spp.)
lVild oat is an important weed of cereal and a¡able crops throughout the world, and is of
major importance in Australia. It is estirnaæd that wild oat infestations cost Austalian wheat
growers at least M2 million per year (Medd and Pandey 1990) due to the reduction of yields
through wild oat competition and the cost of herbicides applied to control wild oat. There
are three species of wild oat, Avena latuaL., A. sterilis ssp. ludoviciana Dur. G&M and A.
barbata Pott. ex Link, occur in Australia. Only the first two species are documented as a
major weed of cereal crops while /,. barbata is found mainly in vacant, undisturbed land and
in pastures (V/halley and Burfit t972, Paterson t976, Ma¡tin et al. 1988). The detailed
descriptions for those two species of wild oat are given in Table 1 .10.
Table 1.10 Diagnostic cha¡acters of the two main wild oat weeds (Thomas and Jones 1976)
Character A. fatuaL. A. sterilis ssp. ludoviciana Dw. G&M
Chromosomenumber Hexaploid(2n=6x42) Hexaploid(2n=6x=42)
Season of germination
Awn on third seed
Abscission sca¡
Shedding of ripe seed
Spring
Present
Present at the base of
every seed
All seeds of spikelet fall
separately
V/inter
Absent
At the base of first seed only (second
and third seeds end in stalk)
Seeds of spikelet fall together as a
unit and force necessary to separate
the grains benveen a spikelet

31
1.9.1 Seed Dormancy and Germination
Dormancy in wild oat seeds is a major problem in controlling this weed throughout the
world. Seeds of. A. fatua and A. sterilis are dormant for at least two months after harvest
(Thurston 1959). About 957o of A. faru¿ seeds are dormant at the time of shedding whereas
orty 50Vo of A. sterilis seeds are dormant at this time (Chancellor 1976). ln A. steríI¡s, the
fust seed in the spikelet is usually non-dormant and germinates within a few weeks of being
shed (Thurston 1951, Gwynne and Murray 1985). Dorrrancy of wild oats is due to the
impermeability of seed coat to oxygen (Naylor and Simpson 1961) and is more prolonged
under cool, moist conditions (Gwynne and Murray 1985).
There are several treatments for breaking dormancy in wild oat seeds:
(a) Scarification. The hull (lemma and palea) of wild oat seed retards water uptake and
contains germination inhibitors. Thus, the germination of dormant seeds can be brought
about by removing their hulls @lack 1959). It was reported that over 9O7o germination of
wild oat seeds can be obtained by pricking the seeds (Whaltey and Burfitt 1972).
(b) Fertilizers. Watkins (1971) found that the number of germinating wild oats increased as
the rate of the applied nitrogen increased. Adkins et at. (1984a and 1984b) demonstrated
that sodium nitrate and nitrite (50-100 mM) induced germination in a partly after-rþened
dormanthneof A.fatuaby promotingoxygenuptakeasmuch asTOVo priortothefustsigns
of germination. Nitrogen fertilizers significantly enhanced seedling emergence of partially-
dormant seeds of wild oat (Agenbag and DeVillier 1989).
(c) Chemical treatments. Ethanol at 50-200 mM overcame dormancy in freshly han¡ested
and partially after-ripened caryopses of A. fatua after five days of incubation (Adkins et al.
1984c). Cairns and DeVilliers (1986) broke A.fatua seed dormancy by treating the air-dry
seeds with ammonia for 48 h. Ammonia caused an increase in leakage of electrolytes from
treated seeds, indicating an increase in permeability of their seed coats. Giberellic acid (GA)

32
at > 0.1 ppm was sufficient to induce germination of both newly matured and old embryos
of A. fatua (Naylor and Simpson 1961).
Wild oat seed must pass through a wa¡m imbibed condition in the soil before chilling can
stimulate its germination. Quail and Carter (1968) proposed that the optimum temperature
for germination of .4. sterilis seeds was between 0-10'C whereas A. fatua required 20-30'C.
However, Peters (1986) found that germination of dormant A. fatw seeds occurred when
removed from the soil and placed at 15'C. Paterson et al. (1976) found that maximum
germination of A. fatua occurred in 11-23'C range and temperatures above 23'C reduced
germination.
Wild oat seeds do not germinate under water-logged conditions. No germination was
observed for seeds following submersion for 6, 8 or 42 months in fresh water (Brun 1965).
In addition, seeds being in water-logged soil for only t to 2 months could not survive
(Lewis 196l).
1.9.2 Seed Bank of Wild Oat
The seed bank consists of seeds of different ages, some of which a¡e dormant with some
being exposed to favourable and some to unfavourable conditions for germination (Wilson
1988). Seed bank life of wild oat varies depending on soil type, soil depth and cultivation.
For example, under grassland or in undisturbed a¡able soil, the survival period of A. fatua
seeds is up to 8-10 years and about half of that for A. sterilis (Gwynne and Murray 19S5).
Longevity of buried wild oat seeds can range from 2 to 6 years (Thurston 1966) or 7 to 9
years (Chancellor 1976, Miller and Nalewaja 1990) depending on the local environmental
and cultural practices. Injection of wild oat seed into the seed bank can be reduced by straw
burning (Wilson and Cussans 1975) or lost by natural deterioration or by predation
(Marshall and Jain 1970, V/ilson 1972).

33
1.9.3 Control of Wild Oat
Wild oats are highly adaptable and produce large number of seeds which ænd to persist from
year to year. The control of wild oat not only reduces the yield loss in the current year but
also affects future infestation by reducing the numbers of seeds in the soil (seed bank)
carried over to the next year. The earliest herbicides used for wild oat control were non-
selective graminicides such as dalapon, propham, chloropham (Hutson and Roberts 1987).
In the late 1950s, diallate and ba¡ban were inEoduced and these herbicides were selective for
the control of wild oat in cereals. However, all varieties of oats were susceptible whereas
wheat and barley were relatively resistant. Triallate proved to be better for wheat and barley
than diallate (both species being more tolerant to triallate) and the use of diallate for wild oat
control was eventually discontinued (Hutson and Roberts 1987). In the late 1970s,
diclofop-methyl was fustly used as a post-emergence herbicide for the control of wild oat in
wheat and barley (Friesen et al. 1976). Since then, many more APP and CHD herbicides
have become available for the control of wild oat and other grass weeds. Of these, diclofop-
methyl, fenoxaprop-ethyl and tralkoxydim give selective control in grass crops such as
wheat and barley. The other APPs (such as fluazifop-butyl, haloxyfop-ethoxyethyl,
quizalofop-ethyl) and CHDs (such as sethoxydim, cycloxydim and alloxydim) are widely
used to control wild oat in dicot crops (Worthing and Hance 1991). In addition,
fenoxaprop-etþI, flamaprop-methyl or tralkoxydim can be used during stem elongation and
booting stage of wild oat to reduce the seed production (Medd et al. 1992).
1.10 Herbicide Resistance in Wild Oat
The occurrence of resistance to ACCase-inhibiting herbicides in wild oat is of considerable
importance as wild oat is one of the most serious weeds of cereal cropping in many
countries. Also there are few altemative selective chemistries for wild oat control. Resistant
wild oat has now appeared in N. America, Europe and Australia. In western Canada, there
are 15 populations of Avena fatua that have developed high-level resistance to triallate
(O'Donavan et al. 1994). Most triallate-resistant populations are also resistant to

34
difenzoquat. However, the mechanisms conferring resistance to triallaæ and difenzoquat in
those populations have not yet been elucidated. In addition, wild oat resistant to APP and
CHD herbicides in N. America developed following 6 to l0 herbicide applications (Heap et
al. 1993). Among these populations, A. fatua biotype UM-l has been examined for
possible mechanisms conferring resistance to APP herbicides. This study showed that this
biotype was able to repolarize the plasma membrane potential following depolarization by
diclofop acid and fenoxaprop acid (Devine et al. 1993) and has a resistant ACCase (Marles
and Devine, unpublished).
In Australia, resistance to diclofop-methyl tnAvenafatwtwas first evident in 1990 (T. Pþr
unpublished). This biotype was subsequently shown low-level resistance to diclofop-
methyl but was not resistant to other APP and CHD herbicides (Mansooji et al. 1992). The
mechanism of resistance in this biotype has not been elucidated but this biotype does contain
a sensitive target ACCase (Boutsalis et al. 1990). At present, resistance to APP and CHD
herbicides has been confirmed in populations of A. fatua and A. sterilis from New South
Wales, South Australia, Victoria and S/estern Australia (Mansooji et aI. 1992). At least 20
populations of Avena spp. have developed resistance to APP and/or CHD herbicides
(Mansooji 1994).
1.11 Objectives of this Project
The aim of this project is to elucidate the mechanism or mechanisms by which biotypes of
wild oat (Avena fatua and Avena sterilis) in Australia have become resistant to ACCase-
inhibiting herbicides. Ten putative resistant biotypes were examined with varying levels of
resistance. Four biotypes which showed different patterns of resistance were examined in
more detail.

35
Chapter 2
Materials and Methods
In this chapter, general methods will be described. Some modifications to these methods
and specific techniques are detailed in the relevant chapters.
2.1 Plant Material
Wild oat (4. fatw L. andÁ. sterilis L.) biotypes were collected from the following sites:
SAS I Collected in 1989 from a barley field near Bordertown, South Australia.
NAS 4 Collected in 1990 from a wheat field near Greenthorpe, New South V/ales.
NAF 6 Collected in 1991 from a wheat field near Orange, New South Wales.
NAS 14 Collected in 1989 from a wheat field nea¡ Henty, New South V/ales.
NAF 8 Collected in 1991 from a wheat field near Junee, New South rWales.
NAS 2 Collected in 1990 from a wheat field nea¡ Greenthorpe, New South rùfales.
NAS 6 Collected in 1991 from a wheat trial site nea¡ Tamworttr, New South Wales.
SAF 14 Collected in 1990 from a wheat field near Bordertown, South Australia.
SAF 34 Collected in 1991 from a wheat field near Mintaro, South Australia.
V/AF 12 Collected in 1991 from a wheat field nea¡ Coomberdale,'Western Austalia.
SAS 2 Collected from unsprayed area adjacent to biotype SAS 1.
SAF 19 Collected from unsprayed pasture area near Bordertown, South AusEalia

36
A coding system was used to classify the wild oat seed samples. The fust letter is taken
from the state of Australia in which the biotlpe was collected. The following letters are the
genus and species of wild oat. The number is the order of collection from each state. For
exarrple, bioqpe SAS I reprcsents wild oat from South Australia, Avena sterílis, collection
No. 1. Biotype \MAF 12 represents wild oat from'Western Australia, Avena fatua,
collection No. 12. The full details of histories of herbicide applications of all resistant
biotypes are shown in Appendix l. The number of herbicide applications in four
representative resistant biotypes of wild oat are shown in Table 2.1. Biotypes SAS 2 and
SAF 19 a¡e known to be susceptible to all ACCase-inhibiting herbicides.
Other species used:-
Oat (Avena sativa L.) cultivar Echidna
Wheat (Triticum aestivum L.) cultivar Halberd
Table 2.1 Number of herbicide applications prior to the development of herbicide
resistance in four resistant biotypes of wild oats
Biotype APP herbicides CHD herbicides Other herbicides
SAS 1 3 diclofop, 2fluaztfop
NAF 6 2 diclofop
NAS 4 5 diclofop, 1 fluazifop, 2 tralkoxydim
I haloxyfop, 1 fenoxaprop
0
0
0
0
0
3 trifluralin
NAS 14 4 diclofop, 2f\razifop 2 simazine

37
2.2 Plant Culture
2.2.1 Germination
Seeds of wild oats were dehusked and gerurinated in containers containing O.6Vo agar and
kept at 5"C for 7 d then transferred to a seed germinator at 15'C 12 h light/dark cycle.
Seedlings were transplanted at one-leaf stage approximately 5 d after conrmencement of
germination. Plants of even size and vigour were selected. V/heat and oat seeds were
incubated for 48 h at 5'C prior to transfer to l5'C. These seedlings were transplanted one
day later than wild oat seedlings so that plants reached the two-leaf stage at the same time.
2.2.2 Pot Culture
Germinated seedlings were transplanted into sterile potting mix based on sand and peat (1:1)
in 15-cm pots. For dose response experiments, plants were grown outdoors during autumn
and winter. For all other experiments, plants were grown in the growth chamber with 20"C,
14 h, 330 p mol photons m-2 s-1 light period/16'C, 10 h dark period.
2.2.3 Hydroponic Culture
Seedlings germinated as in 2.2.1 were transferred to I L rectangular plastic containers
containing 500 mL of nutrient solution (Table 2.2). Containers were $rrapped with
aluminium foil to prevent light reaching the roots. Seedlings were supported by black poly
propylene beads (400 ml/container) then transferred to the growth room with the same
conditions as in section2.2.2. The containers were topped up with 100 mL water every day
to replace evapo-transpirational losses in the first week. Afterwards, half strength nutrient
solution was added until the end of the experiments.

38
Table2.2 Hydroponic nutrient solution (Hoagland and Arnon 1938)
Source of nutrient Final concenEation (¡tM)
Ca(NOg).4HzO
CaSO¿.2HzO
KHzPO¿
K2SOa
KNO¡
MgSO+.7HzO
FeSOa.TH2O
EDTA Na2.2H2O
H3B03
CuSOa.5H2O
NazMoO¿.2HzO 0.25
MnCl2.4H2O 4.60
7-nSO+.7HzO 0.38
1670
800
500
400
t670
1000
72
&
23
0.16

39
2.3 l)ose Response Experiments
Plants were sprayed at the two to three-leaf stage, the most susceptible stage under
agricultural conditions. Herbicides assigned as treatments (Table 2.3) werc applied as
commercial fonnulations with O.2Vo (vlv) non-ionic surfactant (Agral600@) in a laboratory
spray cabinet and delivered via two 110' flat fan hydraulic nozzles suspended 40 cm above
the plant axils. At a pressure of 25O kPa and a boom speed of 1 m s-1, the sprayer output
was equivalent to 113 L ha-l.
Six plants of each biotype were grown in each pot. rü/ithin each herbicide treatment, pots
\ilere aranged as a completely randomized block design with six replicates. Plants were
harvested 4 weeks after spraying which allowed surviving plants to make noticeable growth.
Plants which were green, had live meristems and had increased in size beyond the two leaf-
stage were classified as live plants and were harvested at the ground level. Plant shoots
were dried at 80"C for 48 h prior to dry weight measurement.
Table 2.3 Commercial formulations of aryloxyphenoxypropionates and cyclohexanedione
herbicides registered for use in Australia were used as treatments for dose response
experiments.
Common name Trade name Recommendedrate Manufacturer
(g ai ha-l)
diclofopmethyl Hoegrass@
fluazifopbutyl Fusilade@
haloxyfop-ethoxyethyl Verdict@
cycloxydim Focus@
sethoxydim Sertin@
Hoechst AustraliaLtd.
104 ICI AustraliaLtd.
52 Dow Elancoltd.
40 BASF
100 Schering Australia Ltd.
563
Grasp@talkoxydim 150 Hoechst Australia Ltd.

40
2.4 ACCase Extraction and Assay
The shoot meristematic region of wo to th¡ee-leaf plants was harvested and 3 g of this tissue
was ground in a chilled mortar with 5 rnl- of Buffer A (Table 2.4). All subsequent
operations were performed at 4'C. The brei was centrifuged for 10 min at 27,0009. The
supernatant (2.7 mL) was centrifuged for a few minutes at 10,0009 to remove non-protein
solid material and 2.5 rnL was loaded onto a column containing Sephadex G-25 (PD-10
column, Phannacia, Uppsala, Sweden) previously equilibrated with Buffer B (Table 2.4).
The void volume was eluted with 3.5 mT of Buffer B. The eluate was stored on ice and
assayed immediately for ACCase activity.
To obtain partially purified ACCase enzyme, plant tissue was ground in Buffer A and
centrifuged as mentioned above. The supernatant slowly brought to 1.0Vo (NH4)2SO4
saturation, kept on ice for l0 min and then centrifuged for 20 min at 27,OC[lg at 4'C. The
precipitate was discarded, the supernatant was broughtto 4O7o saturation with (NHa)zSO¿
and centrifuged as above. The supernatant was disca¡ded and the pellet resuspended in
2.7 nL Buffer B and 2.5 nL applied to a column of Sephadex G-25 previously equilibrated
with Buffer B. The eluate (3.5 mL) was stored on ice and immediately assayed for ACCase
activity.
ACCase activity was assayed by following the incorporation of l4C from NaHl4CO, into
acid-stable products at 30'C. Assays, final volume 200ltL, contained Buffer C (Table 2.4),
30 pL protein extract and herbicides or HrO as required. Herbicide stock solutions (1 mM
diclofop acid, fluazifop acid, haloxyfop acid, sethoxydim and tralkoxydim) were dissolved
in 50Vo acetone and made up as serial dilutions in tÙVo acetone. Assay duration was 3 min
and the reaction was terminated by the addition of 30 pL glacial acetic acid. A piece of
glass-micro fibre filter (GF-A; Whatman, Maidstone, Kent, UK) was added to each reaction
tube and the liquid evaporated for 5 h under a stream of air at 70-80'C. Scintillation fluid
(3 mL of toluene:Triton X-100 f2:L, vlvf containing O.4Vo wlv PPO, and O.OlVo (w/v)
POPOP was added. Radioactivity was determined by LSS.

4l
Table 2.4 ExEaction and assay buffers in ACCase assay
BufferA BufferB Buffer C
100 mI\d Tris (pH 8.0)
I nIVIEDTA
2 mM isoascorbic acid
I mMPMSF
20mMDTT
0.57o (wlv) PVP-40
0.5Vo (wlv) insoluble PVP
lOVo (vlv) glycerol
50 mN,f Tricine (pH 8.0)
2.5 mM MgClr.6HrO
50 mM KCI
I mMDTT
50 mM Tricine (pH 8.0)
2.5 mM MgClr.6HrO
50 mM KCI
I TnIvIDTT
I mMATP
0.1 mM acetyl-CoA
10 mM NaH14CO3
containing 0.5 pCi of 14C
2.5 Uptake and Translocation of llaC]Diclofop-Methyl
Resistant (NAF 6, NAS 4 and NAS 14) and susceptible (SAF 19 and SAS 2) plants were
grown hydroponically as in section 2.2.3. At the two leaf-stage, plants were treated with I
pL of 5 mM [l4C] ¿ictofop-methyl (4.5 mM llaC]diclofop-methyl with specific activity of
592 Mbq ml--l dissolved in 0.5 mM commercial product, Hoegrass) onto the leaf æril. Ten
plants of NAS 14 (one plant/replicate) were harvested L,3, 6, 12,24,48 and 72 h after
treatment. For NAF 6 and NAS 4 biotypes, five plants were used in each replicate.
Unabsorbed radioactivity was removed by washing the plant in 5 mL of ZOVo (v/v) methanol
plus 0.17o (v/v) Triton X-100 (contained in 20 ml-scintillation vials). Radioactivity in the
washes was measured by LSS. The plant was sectioned into four fractions:- shoot

42
meristematic region (l cm above the initiation root zone), stem (from meristematic zone up to
2 cm above the leaf axil), leaves and roots (Figure 2.1) except for biotype NAF 6 where
plants were sectioned into the shoot meristematic region and the rest.
second leaffirst leaf
stem
shoot meristematic region
root
Figure 2.1 Plant sections in translocation experiments when harvested
Plant parts were combusted in a Biological Sample Oxidizer OX-600 (R.J. Harvey
Corporation, Milldale, N.Y.) and the l4COzreleased was trapped in 14 mL of scintillation
fluid lCarbosorb@:Permafluor@E+, l:1 (v/v), Canberra-Packard, Tampa, FL, USA] and
laC quantified by LSS.
2.6 Metabolism of Herbicides in Plants
2.6.1 [14C]nictofop-Methyl
Plants \ilere grown in the soil and treated as for the uptake and translocation experiments.
Treated plants were rinsed, blotted dry and frozen in liquid nitrogen. Frozen tissue was
pulverised in liquid nitrogen using a mortar and pestle. Tissue powder was extacted with 5

43
mI . of 807o methanol at -20'C. The brei was centrifrrged at 15,0009 for 20 min at 2'C. The
pellet was washed twice in 2 mI of 807o methanol and recentriñ¡ged at 15,0009 for 10 min
at 4'C. The supenratants were pooled and stored at -20'C for 6 h, then centrifuged for 3
min to remove any precipitate before evaporation. The pooled supernatant was reduced in
volume to near dryness under vacuum and resuspended in enough 507o methanol to provide
radioactivity levels suitable for the radioactivity detector. Samples were filtered through a
O.22 ¡tmNylon filter prior to injection onto the HPLC column.
Radiolabelled metabolites were separated by HPLC equipped with a reverse-phase C18
column (250 mm long, 4.6 mm i.d.; Brownlee Labs ODS-5 Spherisorb; Applied
Biosystems, Lincoln City, California, USA). Radioactivity was detected with an inline
radioactivity detector (Flo-one 1 Beta Model A-140, Radiomatic Instmments and Chemical,
Canberra-Packard, Tampa, FL, USA). Solvents used were 89:10:1 (v/v)
H2O:acetonitile:acetic acid (solvent A) and 90:9:l (v/v) acetoninile:H2O:acetic acid (solvent
B). Elution conditions were a lO-rnin linear gradient from 30 to 35Vo solvent B, followed
by a 12-min linear gradient from 35 to 50Vo solvent B, followed by a 3-min linea¡ gradient
from5OVo to lOÙVo solvent B. The column was then eluted with 1007o solvent B for l0 min
and re-equilibrated with307o solvent B for l0 min prior to injection of next sample. At all
times, the combined flow rate of solvents A and B was 1.5 mL min-l.
2.6.2 ¡4-l4ClSethoxydim
A single droplet of I pL of I mM ¡4-laClsethoxydim (a33 Mbq mmol-l) dissolved in
water:methanol:Triton X-l0O:toluene (96:16.4:L.7:I, by vol.) was placed on the leaf axil of
the resistant NAF 6 and susceptible SAS 2 plants at the two-leaf stage. Plants were
harvested after varying periods of exposure (1,3, 6, t2, 24 and 48 h). Tissue was rinsed
and exfiacted with the same methods as for diclofop metabolism. Six plants were pooled at
each time point. Extracted [laC]sethoxydim and metabolites were separated by HPLC.
Solvents A and B as described in section 2.6.1were used. The elution conditions were a 5-
min linear gradient from 19 to 607o solvent B, followed by a l0-min linear gradient from 60
to 85Vo solvent B, followed by a l0-min linear gradient from 85 to LÙOVo solvent B. Then

4
the column was eluted with IOOEI solvent B for 10 min and re-equilibrated with l9%o
solvent B for l0 min before injection of the next sample. Flow rate was 1.2 mL min-I.
Recovery of radiolabel was deterurined by adding the radioactivity in the wash to the
radioactivity in the extract and dividing by the radioactivity applied.
2.7 Measurement of Plasma Membrane Potentials
Etiolated coleoptiles were collected 3 to 5 d after gerrrination on0.67o agar. The top 3 mm
of the coleoptiles were decapitated and the leaves within the coleoptiles were removed. The
epidermis was peeled using fine forceps under Higinbotham's solution (Table 2.5). A
peeled coleoptile was placed in a plexiglass chamber attached to the stage of a microscope.
During the impalement of a cell, the coleoptile was continuously inigated with
Higinbotharn's solution, pH adjusted to 5.8, at a flow rate of 2.5 mL min-I.
Table 2.5 Higinbotham's solution (Higinbotham et al. 1970).
Source of nutrient Concentation (¡tM)
KCI
NaII2POa
Na2HPOa
Ca(NO¡)z.4HzO
MgSOa.TH2O
Micropipettes \ilere pulled from borosilicate glass capillaries, 0.86 mm i.d. with an intcrnal
filament (GCl50F-10, Clark Electrochemical Instn¡ments, UK) to a tip diameter of about I
pm. The electrodes, with resistances of between 2 to L5 Mll, were mounted on a head-
1000
9U
48
1000
250

45
stage and holder with a Ag-AgCl electrode. The reference electrode, filled with lM KCI in
2Vo (wlv) agar, was placed close to the coleoptile in the bathing solution, but downstreart
from the measuring electrode. Electrode offsets were nulled before each impalement. The
membrane potential was measured by inserting a glass microelectrode into a single cell by a
micromanipulator. Membrane potentials were monitored using a Neuroprobe Amplifier
model 1600 (A-M Systems, USA) connected to a recorder. The initial membrane potential
was allowed to stabilize for 5 min before herbicide application. The basal solution was
replaced with 50 pM diclofop acid in Higinbotham's solution with the final concentration of
l%o (vlv) acetone. After 15 min exposure, the Eeafinent solution was replaced with the basal
solution. Ten coleoptiles from resistant and susceptible seedlings were used and the data
presented a typical run of each biotype.

46
Chapter 3
Spectrum of Resistance to Aryloxyphenoxypropionate and
Cyclohexanedione Herbicides in WiId Oat Biotypes:
Whole Ptant and Target Site Inhibition Studies
3.1 Introduction
The aryloxyphenoxypropionate (APP) and cyclohexanedione (CHD) herbicides are post-
emergent herbicides used to control annual and perennial grass weeds in dicot and some
monocot crops. These are structurally different chemical groups with the same mode of
action, inhibition of ACCase. Weeds can develop resistance to these herbicides after
exposure to these compounds for as few as three consecutive years (Tardif et al. 1993). In
the past 15 years diclofop-methyl and subsequent herbicides which target the plastidic
ACCase enzyme have become widely used for wild oat control in world agriculture.
V/idespread and persistent use of ACCase-inhibiting herbicides has resulted in the recent
appearance of resistant populations of wild oats in AusEalia and North America (Mansooji et
al. L992, Devine et al. 1992, Heap et al. 1993). In Australia, there a¡e increasing numbers
of biotypes of two species of wild oats (Avena fatua L. and A. sterilis L.) exhibiting
resistance to ACCase-inhibiting herbicides. In addition, biotypes of several other grass
weeds are resistant to APP and CHD herbicides such as Alopecurus rnyosuroides (Moss
1990), Lolium multiþrum (Gronwald e¡ aL. 1992), L. rigidum (Heap and Knight 1982,
Powles and Howat 1990) and Setaria viridis (Marles et al. 1993).
Putative resistant biotypes of wild oat were investigated for their responses to ACCase-
inhibiting herbicides. It is suspected that there would be some va¡iations in response to
ACCase-inhibiting herbicides as a result of different herbicide treaünents in the field. The
response of ten putative biotypes of wild oat were tested with the two chemical groups, APP
and CHD herbicides. Herbicides examined were diclofop-methyl, fluazifop-butyl,

47
haloxyfop-ethoxyethyl, sethoxydim and tralkoxydim. The ability of these herbicides to
inhibit ACCase activity invitro was also determined.
3.2 Whole Plant Assessment of Resistance
(a) Response to APP Herbicides
Ten putative resistant wild oat biotypes with varying histories of exposure to APP and CHD
herbicides (Appendix l) were exaurined for resistance to APP and CHD herbicides. These
biotypes were found to express very different patterns of resistance to APP and CHD
herbicides. Detailed dose response experiments were conducted to the ACCase-inhibiting
herbicides as shown in Figure 3.1. At the recoûrmended rate for wild oat control in
Australia, 563 g diclofop-methyl ha-1, the susceptible biotypes exhibited no survival
whereas three resistant biotypes, NAS 14, NAF 6 and NAS 4 exhibited 60, 100 and l00%o
survival, respectively (Figure 3.lA). Biotype NAS 14 and the susceptible biotype were
easily controlled by the recommended rate of fluazifop-butyl (104 g ha:l) whereas biotypes
NAS 4 and NAF 6 were resistant at this rate exhibiting90%o andTOVo survival, respectively
(Figure 3.lB). The three resistant biotypes were also resistant to haloxyfop-ethoxyethyl
with3OVo,TOVo and 907o survival at the recommended rate of 52 gha-l for biotypes
NAS 14, NAF 6 and NAS 4, respectively whereas the susceptible biotype was controlled
by this rate (Figure 3.lC).

48
1finO 2finO 3finO
100
00 200 400 600 800
100
Herbicide rate (g ai h"-1)
Figure 3.14.C Survival of susceptible SAF 19 (O) and putative resistant NAS 14 (A),
NAF 6 (O) and NAS 4 (I) wild oat biotypes 30 d after exposure to diclofop-methyl (A),
fluazifop-butyl (B) and haloxyfop-ethoxyethyl (C). A¡rows indicate rates recommended for
wild oat control. Each point is the mean + standard error of six replicates.
100
s80ã60a-bã40u)
20
s80ã60.-340
ct)20
s80ã60'E
40(n
20
00
04003002001000
Adiclofop-methyl
Bfluazifop-butyl
ü
chaloxyfop-ethoxyethyl

49
The susceptible wild oat biotypes SAF 19 and SAS 2 werc easily controlled by the
reconrmended rates of all APP and CHD herbicides tested. The rates of APP herbicides
required to kill 50Vo of. the individuals of the wild oat populations are given in Tables 3.1 to
3.3. The level of resistance to the herbicides among the resistant populations was classified
into three $oups by using the resistanUsusceptible ratio of LD5g values as follows:
Level of resistance APP herbicides CHD herbicides
>50
Moderate >10
Low
Sensitive <2
Diclofop-methyl is widely used to selectively control grass weeds in wheat. rrl/heat is
resistant to diclofop-methyl due to the ability to rapidly detoxify the herbicide (Shimabukr¡ro
et al. 1979). However, resistant biotypes SAS I and NAS 4 were less sensitive to diclofop-
methyl than wheat cv. Halberd. At the rate of 16 kg diclofop-methyl ha-l they had l00Vo
survival while in this wheat cultivar some mortality was observed. The biotypes SAS l,
NAS 4, NAF 8, SAF 14 and NAS 2 were highly resistant to diclofop-methyl with LD59
ratios of. >2L2, >192, 190, 159, and 113, respectively (Table 3.1). Biotype NAF 6 was
moderately resistant to diclofop-methyl with an LDso ratio of 45. The other four resistant
biotypes, NAS 14, ÌWAF 12, NAS 6 and SAF 34, expressed either a low level resistance
to diclofop-methyl; 3.2 and 2.3 times, respectively for biotypes NAS L4 and WAF 12, or
showed no resistance <2 fold (Table 3.1).
>20
>4
>2
<2
>2
High

50
Table 3.1 Amounts of diclofop-methyl in g ai ha-I, required to kill 50Vo ofpopulations
of twelve biotypes of wild oat and wheat cv. Halberd. The recommended rate of
diclofo¡methyl for wild oat control in Austalia is 563 g ai hrl.
Biotype LDso G ai ha:t) Resistant/Susceptible
wheat cv. Halberd
Resistant biotvoes
SAS I*
NAS 4
NAF 8
SAF 14
NAS 2
NAF 6
NAS 14
rwAF 12
NAS 6
SAF 34
Susceotible biowoes
SAS 2*
SAF 19
>16,000
>30,000
>32,000
31,660
26,550
18,890
7,490
392
227
t4t
167
>2t2
>192
190
159
113
45
534
286
3.2
2.3
t.7
1.4
* Data from Mansooji et al. 1992

51
Growth response of biotypes having moderate and high levels of resistance to APP
herbicides were not the same for all the herbicide rates. When exposed to the rccommended
rate, plants looked unaffected. At very high rates, from 8,000 to 32,0O0 g ha-l diclofop-
methyl, growth was obviously inhibited and transient yellowing or leaf burn appeared on the
older leaves. However, emerging new leaves appeared normal. In contrast, biotypes with a
low level of resistance like biotype NAS 14 were badly affected by application rates as low
as 2,000 g ha-l diclofop-methyl. Newly emerging leaves of these biotypes showed severe
chlorosis.
The biotypes SAS l, NAS 2, SAF 14 and NAF 8 were highly resistant to fluazifop-butyl
and had LDso ratios of >1,000, >67,>67 and 53 times that of the susceptible biotypes,
respectively (Table 3.2). Moderate resistance to fluazifop-butyl was observed in the
biotypes NAS 4, NAF 6 and NAS 6 with LDso ratios ranging from 7.7 to 37. The
remaining biotypes 1WAF 12, SAF 34 and NAS 14, showed low level resistance to
fluazifop-butyl with LD59 ratios of 4.3,3.3 and 2.7 times that of the susceptible,
respectively (Table 3.2). These last three biotypes would be controlled by the recommended
rate of fluazifop-butyl.
Biotype SAS 1 was the only biotype to show high level resistance to haloxyfop-ethoxyethyl
(Table 3.3). Biotypes NAS 2, NAF 6 and NAS 4 were moderately resistant to haloxyfop-
ethoxyethyl with LDso ratios of 25,10 and 10 times that of the susceptible, respectively.
The remaining biotypes exhibited low-level resistance to haloxyfop-ethoxyethyl with LDso,
ranging from 8.5 to 4 times that of the susceptible.

52
Table 3.2 Amount of fluazifopbutyl in g ai ha-I, requi¡ed to kill SOVo of populations
of twelve biotypes of wild oat. The recoûlmended rate of fluazifop-butyl for wild oat
conEol in Australia is 106 g ai ha-I.
Biotype LDso G ai ha-t) Resistant/Susceptible
Resistant biotvoes
SAS I*
NAS 2
SAF 14
NAF 8
NAS 4
NAF 6
NAS 6
IüVAF 12
SAF 34
NAS 14
Susceotible biowoes
SAS 2*
SAF 19
>16960
>800
>800
uo
48
t28
92
52
40
25
>1000
>66.7
>66.7
53.3
37.3
t0.7
7.7
4.3
3.3
2.t
<16
t2
* Data fromMansooji et al.1992

53
Table 3.3 Amount of haloxyfop-ethoxyethyl in g ai ha-I, required to kilt SOVo of
populations of twelve biotypes of wild oat. The recommended rate of haloxyfop-
ethoxyethyl for wild oat control is 52 g ai ha-I.
Biotype LD59 G ai tra-t¡ ResistanUsusceptible
Resistant biotvoes
SAS 1* 177
NAS 2
NAF 6 10
NAS 4 t0
NAF 8 8.5
SAF 14
NAS ó 5.5
NAS 14
WAF 12 4.8
SAF 34
SAS 2* l0
SAF 19
* Data from Mansooji et al. 1992
5
1768
200
80
80
68
56
4
40
38
32
25
7
4
8

54
O) Response to CHD Herbicides
None of the putative resistant biotypes tested were found to be highly resistant to
tralkoxyrlim (Table 3.4). Biotype NAF 6 exhibited 507o survival at the recommended rate of
100 g ha-l tralkoxydim whereas biotypes NAS 4 and NAS 14 and the susceptible biotype
SAF 19 were controlled by this rate of herbicide (Figure 3.2). Biotypes NAF 6 and
V/AF 12 were moderately resistant to tralkoxydim with LDso. 5 and 3.6 times that of the
susceptible, respectively (Table 3.4). Biotypes NAS 14, NAF 8, NAS 2 and NAS 6 were
slightly resistant to tralkoxydim but were completely contolled by the recommended rate of
100 g ¡¿-l (Table 3.4). The other biotypes were not resistant with LD5g ratios of less
than 2.
Only a single biotype of wild oat, NAF 6, showed substantial survival at the recommended
rate of 100 g ha-l tralkoxydim. This biotype was also resistant to the other CHD herbicides,
cycloxydim and sethoxydim with 90 and lÙ07o survival, respectively, at the recommended
rates (Figure 3.34-B). Rates of CHD herbicides, tralkoxydim, cycloxydim, and
sethoxydim required to kill 5OVo of biotype NAF 6 were 5 to 7 times those required to kill
SOVI of the susceptible population (Table 3.4 and Figure 3.34-8).

55
Table 3.4 Amount of tralkoxydim, in g ai ha:I, required to kill SOVo of populations of
twelve biotypes of wild oat. The recommended rate of tralkoxydim for wild oat contol in
Australia is 100 g ai ha-l.
Biotype LD5s (g ai ha-t) ResistanUsusceptible
Resistant biotvoes
NAF 6
rùrAF 12
NAS 14
NAF 8
NAS 2
NAS 6
NAS 4
SAS 1*
SAF 14
SAF 34
Susceotible biowoes
SAS 2*
SAF 19
5100
72
52
50
50
50
36
34
28
28
3.6
2.6
2.5
2.5
2.5
1.8
2.6
1.4
1.4
l3
20
* Data from Mansooji et aL.1992

56
100
80
èa
ã60.-¡i
ã40
20
00 100 300 400
Tralkoxydim (g ai ha'1)
Figure 3.2 Survival of susceptible SAF 19 (O) and resistant NAS 14 (A), NAF 6 (O)
and NAS 4 (f) wild oat biotypes 30 d after exposure to tralkoxydim, Arrow indicates the
rate recommended for wild oat control. Point symbols a¡e the mean t standa¡d error of the
six replicate experiments.
240

57
100 A I Bsethoxydim
20
0 40 80 t20 160 0 100 2t0 300 400
Herbicide rate (g ai h"-1)
Figure 3.34-B Survival of susceptible SAS 2 (O) and resistant NAF 6 (O) wild oat
biotypes following treatment with cycloxydim (A) and sethoxydim (B). Point symbols are
the mean * standard error of six replicate experiments. Arrows indicate the recommended
rate of herbicides for use in AusEalia.
EO
èa
:60G
.l
i40ct)
0

58
3.3 Inhibition of ACCase Activity by Herbicides
Partially purified ACCase enzyme extracted from resistant and susceptible plants was
assayed (as described in Chapter 2) to determine the effect of ACCase-inhiþi¡i¡g heùicides.
The level of resistance to the herbicides among the resistant populations w¿ts classified as in
the whole plant level (section 3.2)by using the resistant and susceptible ratio of I5g values.
Inhibition of ACCase by diclofop acid and tralkoxydim a¡e shown in Figures 3.4 to 3.7 for
four representative resistant biotypes compared to susceptible biotypes. These figures show
that ACCase extracted from the susceptible biotypes SAS 2 and SAF 19 was sensitive to
inhibition by diclofop acid with an Iso of less than 1.1 UM. Similarly, the enzyme from the
susceptible biotypes was sensitive to tralkoxydim with an I5g of about I to 2 ¡tM. ACCase
from four resistant biotypes displayed varying responses to these two herbicides. ACCase
from biotype SAS I was highly resistant to diclofop acid with an Iso >10 pM (Figure 3.4A)
but was only moderately resistant to tralkoxydim (Figure 3.48). ACCase from biotype
NAS 4 showed moderate resistance to diclofop acid, I5g -6 ttM (Figure 3.54), and only
slight resistance to tralkoxydim (Figure 3.58). ACCase from biotype NAF 6 showed
moderate resistance to both diclofop acid and tralkoxydim with 15¡ of -4 ¡rM for diclofop
acid and 4 lrM for tralkoxydim (Figure 3.6A-8). ACCase from biotype NAS 14 showed
identical inhibition kinetics to the susceptible biotype for both diclofop acid and tralkoxydim
(Figure 3.74-B).
ACCase was extracted from all biotypes and herbicide inhibition curves were generated for
diclofop acid, fluazifop acid, haloxyfop acid, sethoxydim and tralkoxydim. The I5gs and
R/S ratios for these herbicides for each biotype are shown in Tables 3.5 to 3.9. The
ACCase from biotype SAS I was the most resistant to diclofop acid. ACCase from biotypes
NAS 4, NAF 8 and SAF 14 was also strongly resistant to this herbicide. ACCase from
biotypes NAF 6 and NAS 2 was moderately resistant, whereas that from biotypes NAS 6,
NAS 14, WAF 12 and SAF 34 was not resistant (Table 3.5).

59
The ACCase from biotype NAS 2 was most resistant to fluazifop acid (Table 3.6). ACCase
from biotypes SAF 14, NAF 8, NAS 4, NAS 6 and SAS I was also strongly resistant,
whereas the ACCase from biotypes NAF 6, WAF 12 and SAF 34 showed low-level
resistance to fluazifop acid. ACCase from biorype NAS 14 was susceptible to fluazifop
acid.
None of the biotypes examined had ACCase with high-level resistance to haloxyfop acid
(Table 3.7), such as was seen for diclofop acid and fluazifop acid. ACCase from biotypes
SAS I and NAF 8 were moderately resistant to this herbicide, whereas that from biotypes
SAF 14, NAS 4, NAS 2, NAF 6, NAS 6, WAF 12 and SAF 34 exhibited low-level
resistance to this herbicide. ACCase from biotype NAS 14 was susceptible to haloxyfop
acid.
ACCase from biotypes SAS 1, NAF 6, WAF 12 and NAS 2 was moderately resistant to
sethoxydim (Table 3.8). The ACCase from biotypes NAF 8, NAS 4, NAS 6, SAF 34 and
SAF 14 showed low-level resistance to sethoxydim whereas that from biotype NAS 14 was
as sensitive as the susceptible enzyme.
For tralkoxydim inhibition, ACCase from biotypes SAS l, NAF 6 and IWAF 12 was
moderately resistant to this herbicide (Table 3.9). The enzyme from biotypes NAS 14 and
SAF 34 was sensitive to tralkoxydim whereas that from biotypes NAF 8, NAS 2, NAS 6,
NAS 4 and SAF 14 showed lowJevel resistance to tralkoxydim.

60
-oLۃoI
CHoñ
-a-
a-ìIcÉ(ÐU2GI(JU
100
80
60
40
20
100
80
60
40
20
0
00.1 1 10 100
Concentration (pM)
Figure 3.44-8 Inhibition of ACCase activity by diclofop acid (A) and tralkoxydim (B)
in susceptible SAS 2 (O) and resistant SAS 1 (O) wild oat biotypes. Data are the means of
seven replicate experiments. Vertical bars represent the standard errors.
A
B
a

61
0.1 I 10 100
Concentration (ttM)
Figure 3.54-8 Inhibition of ACCase activity by diclofop acid (A) and tralkoxydim (B)
in susceptible SAF 19 (O) and resistant NAS 4 (O) wild oat biotypes. Data are the means of
four replicate experiments. Vertical bars represent the standard enors.
100
80
60
40
20
0
100
80
60
40
20
-oLrÅIocJ
CHo
Ba
È¡.-.a€I
cÉ
c)at2c!QL)
0
e
B

62
0.1 1 10
Concentration (pM)
Figure 3.64-B Inhibition of ACCase activity by diclofop acid (A) and tralkoxydim (B)
in susceptible SAF 19 (O) and resistant NAF 6 (O) wild oat biotypes. Data are the means of
four replicate experiments. Vertical bars represent the standard errors.
-oLÈIoI
CHoBa
.aJ
-IcÉ(¡)u)6lU(J
100
80
60
40
20
00
80
60
40
20
I0
0100
A
B
0

63
100
80
60
-oËã40c.)c:o
Ð20*¡'F0.lË 1oocË
é)(t)
880t)
40
20
A
B
0.1 I 100
100
Concentration (pM)
Figure 3.74-B Inhibition of ACCase activity by diclofop acid (A) and tralkoxydim (B)
in susceptible SAF 19 (O) resistant NAS 14 (O) wild oat biotypes. Data are the means of
four replicate experiments. Venical bars represent the standa¡d enors.

&
Table 3.5 Concentrations of diclofop acid giving 507o inhibition of ACCase from
susceptible and resistant wild oat biotypes. Ratios are the value for the resistant divided
by the value for the susceptible biotype. ACCase activity \ilas measured in partially
purifred extracts of the shoot meristematic regions. Values are the mean È standard error
of four replicate experiments.
Biotype Iso (tM) ResisanUSusceptible
Resistant biotvoes
SAS 1*
NAS 4
NAF 8
SAF 14
NAF 6
NAS 2
NAS 6
NAS 14
SAF 34
WAF 12
Susceptible biotypes
SAS 2*
SAF 19
11.0 r 3.0
6.30 + 0.8
6.95 t 0.8
5.85 + 0.6
3.80 + 0.7
2.90 + 0.8
o.93 to.2
0.93 + 0.4
1.51 + 0.3
r.22 t 0.3
0.2 r 0.1
1.1 + 0.9
52
6
6
5
3
3
1
I
I
I
* Values a¡e the mean t standard error of seven replicate experiments

65
Table 3.6 Concentrations of fluazifop acid giving 507o inhibition of ACCase from
susceptible and resistant wild oat biotypes. Ratios are the value for the resistant divided
by the value for the susceptible biotype. ACCase activity was measured in partially
purified extracts of the shoot meristematic region. Values are the mean t standa¡d error
of foru replicate experiments.
Biot¡pe Iso (tM) Resistant/Susceptible
Resistant biowoes
NAS 2
SAF 14
NAF 8
NAS 4
SAS 1*
NAF 6
NAS 6
I9VAF 12
SAF 34
NAS 14
Susceptible biotypes
sAs 2*
SAF 19
157 + t5
49+tt
97 t7.0
78 + 6.0
23 + 6.0
14 + 6.0
3l + 3.0
r0 t 2.5
16 t 4.0
6t2.0
2.5 + 0.8
3.2 + r.0
50
15
30
25
9
4
10
3
5
1.9
* Values are the mean + standard error of seven replicate experiments

66
Table 3.7 Concentrations of haloxyfop acid giving 507o inhibition of ACCase from
susceptible and resistant wild oat biotypes. Ratios are the value for the resistant divided
by the value for the susceptible biotype. ACCase activity was measured in partially
purifred extracts of the shoot meristematic region. Values are the mean * standard error
of four replicate experiments.
Biotype Iso (tM) ResistanlSusceptible
Resistant biotvoes
SAS I*
NAS 2
NAF 6
NAS 4
NAF 8
SAF 14
NAS 6
NAS 14
WAF 12
SAF 34
Susceotible biotvoes
SAS 2*
SAF 19
17 r 1.0
1l + 1.0
5r1.6
16 r 3.0
24 t3.5
18 r 0.4
5.6 r 0.1
2.0 + 0.6
4.7 + 1.2
3.4 r 0.3
0.7 r 0.1
1.8 + 0.6
25
6
3
9
13
10
3
1
3
2
* Values a¡e the mean t standa¡d error of seven replicate experiments

67
Table 3.8 Concentrations of sethoxydim giving 507o inhibition of ACCase from
susceptible and resistant wild oat biotypes. Ratios a¡e the value for the resistant divided
by the value for the susceptible biotype. ACCase activity was measured in partially
purified extracts of the shoot meristematic region. Values a¡e the mean t standard error
of four replicate experiments.
Biotype Iso (pM) ResistanlSusceptible
Resistant biotvoes
NAF 6
V/AF 12
NAS 2
sAs 1*
NAF 8
NAS 4
NAS 6
SAF 34
SAF 14
NAS 14
Susceotible biowoes
SAS 2*
SAF 19
38 r 6.8
28 t2.8
25 + 5.t
22 t 5.0
22t 4.3
t4+ 2.5
t5 + 2.6
16 + 5.0
18 r 3.6
410.8
2.7 X0.7
5.4 + 0.9
7
5
5
8
4
3
3
3
3
I
* Values a¡e the mean t standa¡d error of seven replicate experiments

68
Table 3.9 Concentrations of tralkoxydim giving 507o nhtbition of ACCase from
susceptible and resistant wild oat biotypes. Ratios are the value for the resistant divided
by the value for the susceptible biotype. ACCase activity $,as measured in partially
purified extracts of the shoot meristematic region. Values a¡e the mean t standard error
of four replicate experiments.
Biotype Iso (pM) Resistant/Susceptible
Resistant biotvoes
SAS I*
NAF 6
WAF 12
NAS 14
NAF 8
NAS 2
NAS 6
NAS 4
SAF 14
SAF 34
Susceotible biowoes
sAs 2*
SAF 19
6.s + 5.0
3.7 + 0.9
5.5 + 1.4
0.9 r 0.1
2.t + o.3
1.8 t 0.4
t.9 t 0.2
1.7 r 0.3
2.6 + t.r
1.0 + 0.3
2.1t 0.6
0.8 r 0.r
3
4
6
I
2
2
2
2
3
I
* Values are the mean t standard error of seven replicate experiments

69
3.4 l)iscussion
Of the ten putative resistant populations of wild oat tested for resistance, all proved to be
resistant to at least one APP or CHD herbicide (Tables 3.1 to 3.4). Not all biotypes were
resistant to diclofop-methyl (Table 3.1), and only one biotype NAF 6 displayed
substantial resistance to the CHD herbicides (Table 3.4). The pattern of resistance at the
whole plant level could be generally classified into four groups (Table 3.10). A group
with high resistance to diclofop-methyl but sensitive to tralkoxydim consisted of biotypes
SAS l, NAS 4 and SAF 14. High resistance to diclofop-methyl and low resistance to
tralkoxydim was found in biotypes NAF 8 and NAS 2. Moderate resistance to both
diclofop-methyl and ualkoxydim was found in biotype NAF 6. Low resistance to both
herbicides was observed in biotypes NAS 14 and }VAF 12. Apart from those groups,
biotypes NAS 6 and SAF 34 were rather sensitive to both herbicides.
Most of these populations also contained an ACCase that was resistant to one or more of
the APP herbicides. In parallel with whole plant experiments, there was much variation
in the level of resistance exhibited at the enzyme level. Differences in the spectra of
resistance to APP and CHD herbicides at the whole plant and enzyme level in Table 3.10
for the populations of A. fatua and A. sterilis suggest that some of the va¡iation in
patterns of resistance in wild oat may be due to the presence of different physiological and
biochemical mechanisms of resistance.
Resistance to ACCase-inhibiting herbicides has been documented in several weed species
such as Lolium rigidum (Heap and Knight L982), L. tnultiþrum (Gronwald et al.1992),
Avena spp. (Joseph et al. 1990, Heap et al. 1993, Mansooji et aL L992), Setaria viridis
(Marles et al. 1993), Eleusine indica, Digitaria sanguinalis, Sorghum halepense and,
S. faberii (reviewed in Devine and Shimabukuro 1994). In most cases, resistance has
become evident after repeated applications of either, or both, APP and CHD herbicides for
several years. The patterns of resistance to these herbicides in resistant biotypes vary.
There may be resistance to diclofop only, to all APP but not CHD herbicides, or to both
APP and CHD herbicides.

70
Table 3.10 Degree of resistance at the whole plant level and at the target enzyme,
ACCase, of ten resistant biotypes of wild oat.
Biotype Whole plant response to: Enzyme rcsponse to:
Diclofop-methyl Tralkoxydim Diclofop acid Tralkoxydim
SAS I
NAS 4
NAF 8
NAS 2
SAF 14
NAF 6
NAS 14
V/AF 12
NAS 6
SAF 34 sensitive
sensitive
sensitive
moderate
loilmoderate
sensitive
high
high
high
high
high
low
low
low
low
low
high
low
low
sensitive
moderate
moderate
moderate
moderate
moderate
sensitive
sensitive
sensitive
sensitive
sensitive
moderate
sensitive
moderate moderate
lowlowsensitive
lowlow
low

7t
As for the whole plant response, the pattern of resistance of ACCase to herbicides also
varies. Most ACCase from resistant weed biotypes demonstrated a greater level of resistance
to APP herbicides than to CHD herbicides (Table 3.10) e.g. biotypes SAS I, NAS 4,
NAF 8, NAS 2 and SAF 14. Similar results were obtained from some resistant biotypes of
L. rigidum which were 30 to 85 fold resistant to diclofop but I to 8 fold resistant to
sethoxydim (Tardif et al. 1993; Tardif, Preston, Holtum and Powles unpublished; Preston,
Tardif, Christopher and Powles unpublished). ACCase from biotype NAF 6 was
moderately resistant to both APP and CHD herbicides. Likewise, two biotypes of .á..
myosuroider were moderately resistant to both diclofop and sethoxydim but they were more
resistant to fenoxaprop (Hall, Moss and Powles unpublished). In contrast to the biotypes
above, a biotype of S. viridir was highty resistant to both diclofop and sethoxydim (Marles
et al. 1993) and ACCase from a l27o subset of L. rigidum biotype SLR 3l showed low level
resistance to diclofop but was highly resistant to sethoxydim (Tardif and Powles 1994).
ACCase from biotypes NAS 14, ÌWAF 12, NAS 6 and SAF 34 was sensitive to diclofop and
only ACCase from biotypes NAS 6 and ÌWAF 12 showed a moderate level of resistance to
tralkoxydim (Table 3. 10).
Target site cross-resistance across APP and CHD herbicides in L. rigidum can result from
selection either with an APP herbicide (Holtum and Powles 1991) or a CHD herbicide
(Tardif et al. 1993). Target site cross-resistance has occurred in biotype NAF 6 which has
been selected with two applications of diclofop-methyl and has a moderate cross-resistance to
CHD herbicides. In contrast, wild oat biotypes SAS I and NAS 4, which had been exposed
to APP herbicides for at least five years, are highly resistant to APP herbicides but remain
sensitive to a CHD herbicide, tralkoxydim. A similar result was observed in a biotype of Z.
multiþrum which did not exhibit cross-resistance to CHD herbicides following selection
with APP herbicides (Gronwald et ^1.
1992). These results suggest that, although the
herbicide is a strong selection pressure for weeds carrying a herbicide-resistant ACCase,
cross-resistance across the APP and CHD classes cannot be predicted by herbicide exposure.
It is likely that there are a number of possible mutations of ACCase which can confer

72
resistance to the APP and CHD herbicides and those different mutations of ACCase affect the
action of different herbicides leading to resistance either within or across herbicide classes.
The data presented in this chapter establishes that there are clear differences in the spectra of
herbicide resistance among resistant wild oat biotypes. Thus, in the following chapters, four
resistant biotypes SAS 1, NAS 4, NAF 6 and NAS 14 were used to further investigate the
mechanisms of herbicide resistance. These biotypes are representative of the different
responses of the wild oat biotypes to both application of herbicides to plants and inhibition of
ACCase by herbicides (Table 3.10). Resistant biotypes SAS I and NAS 4 both have a high
level of resistance to APP herbicides, however, there are differences in the responses of the
ACCase from these two biotypes to the APP and CHD herbicides. ACCase from biotype
SAS I is highly resistant to diclofop and slightly resistant to tralkoxydim whereas ACCase
from biotype NAS 4 is moderately resistant to diclofop and slightly resistant to tralkoxydim.
Resistant biotype NAF 6 has moderate resistance to both APP and CHD herbicides at the
whole plant and ACCase levels, whereas biotype NAS 14 has a low level of resistance to
APP and CHD herbicides but contains a sensitive form of the ACCase erizyme.
ACCase from the susceptible biotype SAS 2 was strongly inhibited by a range of herbicides
whereas that of the resistant biotype SAS 1 was much less inhibited (Figure 3.4 and Tables
3.5 to 3.9). ACCase from the resistant biotype was markedly less sensitive to the APP
herbicides (diclofop acid, fluazifop acid and haloxyfop acid) than was ACCase from the
susceptible biotype by 52,25 and 9 fold, respectively. Likewise, the enzyme extracted from
resistant plants was less sensitive to CHD herbicides (sethoxydim and tralkoxydim) by 8 and
6 fold, respectively. Hence, a mechanism for resistance to ACCase inhibiting herbicides in
SAS 1 biotype is the possession of a less sensitive, mutant, target enzyme which has a good
correlation to the resistance at the whole plant level (Mansooji et al. 1992). To support the
assumption that an APP insensitive ACCase is a single mechanism in this biotype, other
possible mechanisms conferring resistance were investigated in Chapter 4.
Biotype NAF 6 shows moderate resistance to both diclofop-methyl and tralkoxydim and
contains an ACCase less sensitive to the APP herbicides diclofop acid, fluazifop acid and

73
haloxyfop acid by 3,4 and 3 times, respectively (Tables 3.5 to 3.7). This ACCase is also
less sensitive to the CHD herbicides sethoxydim and tralkoxydim by 7 and 4 fold,
respectively (Tables 3.8 to 3.9). The effects of APP and CHD herbicides on the target site
correlates well with the results of response to the herbicides at the whole plant level.
A modified target site, ACCase, which is moderately resistant to both APP and CHD
herbicides is probably the mechanism of resistance in this resistant biotype. Other possible
¡¡scþanism of resistance in this biotlpe will be examined in Chapter 5.
Resistant biotype NAS 4 exhibited a greater level of resistance in the field than biotype
SAS 1, whereas the ACCase enzyme from biotype NAS 4 is less sensitive to diclofop acid,
fluazifop acid and haloxyfop acid than ACCase from susceptible SAF 19 by 6,25 and 9 fold,
respectively (Figure 3.54 and Tables 3.5 to 3.7). Similarly, ACCase from resistant NAS 4
plants is slightly less sensitive to CHD herbicides sethoxydim and tralkoxydim by 3 and?
fold, respectively (Tables 3.8 to 3.9). Despite being more resistant to diclofop-methyl than
biotype SAS l, the ACCase of this biotype is considerably more sensitive to diclofop acid
than the ACCase of biotype SAS l. This suggests that other mechanisms may contribute to
resistance in this biotype, which will be examined in Chapter 6.
Resistant biotype NAS 14 is three times less sensitive than the susceptible biotype to
diclofop-methyl and tralkoxydim in the field (Tables 3.1 and 3.4), however, this biotype
contains an ACCase enzyme that is as sensitive to diclofop acid and tralkoxydim as that of the
susceptible (Figure 3.74-8). In addition, the ACCase from biotype NAS 14 exhibits the
same sensitivity to inhibition by haloxyfop acid and sethoxydim as that of the susceptible
plants (Tables 3.5 to 3.9). These results suggest that a modified target enzyme ACCase is
not the mechanism of resistance in this biotype. Possible mechanisms of resistance in this
biotype will be investigated in Chapter 7.

74
Chapter 4
Herbicide Resistance Endowed by an APP Insensitive ACCase
in Biotype SAS 1
4.1 Introduction
The response of wild oat biotype SAS I at the whole plant and enzyme level suggested that
this biotype contains a target site ACCase that is insensitive to these herbicides (Chapter 3).
Reduced sensitivity of the target site is the most commonly encountered mechanism of
herbicide resistance (Holt et al. 1993). This is also true of ACCase-inhibiting herbicides
where, for example, herbicide-resistant ACCase has been found in resistant biotypes of
Lolium multiþrum @etts et al. 1992, Gronwald et al. 1992); L. rigidum (Tardif et al. 1993,
Tardif and Powles 1994) and Setaria viridis (Marles et al. 1993). In wild oat, however, no
cases of resistance due to ACCase insensitivity to APP and CHD herbicides have been
reported (Boutsalis et al. 1990, Devine et al. 1992)
Herbicide resistance may occur as a result of one or more mechanisms. Therefore,
investigations of herbicide uptake, translocation, metabolism and herbicide-induced
membrane depolarization were conducted to establish if any other mechanisms were
contributing to herbicide resistance in this biotype.
4.2 Materials and Methods
Most materials and methods are as described in Chapter 2. Additional methods are described
below.
4.2.1 Variations in ACCase During Development
To verify whether there were differences between the two biotypes, susceptible SAS 2 and
resistant SAS 1, in the amount of ACCase enzyme during development, meristematic tissue
was harvested 5, 10, 20, 30, 40 and 45 days after planting. Crude extracts from both

75
biotypes were assayed with 0.2 ml\d acetyl-CoA. To measure ACCase activity at day 0,
seeds were imbibed with water fot 24 h and I g of seed was extracted and assayed as
described in Chapter 2.
4.3 Results
4.3.1 ACCase Activity During Growth Development
There were no large differences in the level of ACCase activity in crude extracts from
susceptible and resistant plants in any of the plant fractions examined (Table 4.1). The
highest relative ACCase activity was found in the meristematic regions in both biotypes,
while the leaf had the lowest relative activity. As relative activity was highest in meristematic
regions this tissue was used in all subsequent assays.
Table 4.1 ACCase activity (nmoles-rlCOz fixed g-1 FW min-l) in crude extract from
different parts of susceptible (SAS 2) and resistant (SAS 1) A. sterilis plants at the two leaf
stage. Values are the mean + standard error of six replicates.
Plant part Susceptible Resistant
Meristematic region 205.6 + 7.0 224.5 + 6.3
Stem 174.2 + 7.6 180.1 + 6.3
I-eaf lamina 160.7 + 3.2 t&.3 i.3.7
In addition, the amount of ACCase activity in the shoot meristematic regions of both
resistant and susceptible biotypes was found to be similar during the course of plant
development @gure 4.1). ACCase activity in the shoot meristem of both biotypes was low

76
in the imbibed seed at planting. Activity increased in a linea¡ fashion reaching a peak in bottt
biotypes 30 d after planting. ACCase activity in the meristematic regions subsequently
rapidly decreased such that 45 d after planting when plants \ilere at the booting stage
ACCase activity was very low. ACCase activity in the susceptible shoot meristem was
forurd to be slightly higher than in the resistant shoot meristem at most time points.
100
0 10 20 30 40 50
Days after planting
Figure 4.1 ACCase activity in the shoot meristem of the susceptible SAS 2 (O) and
resistant SAS 1 (a) A. sterilis from zero to 45 d after planting. ACCase activity was
measured in crude extracts and each point is the average of three replicates. Vertical bars
represent standard errors and are sometimes obscured by point symbols. The arrow
indicates that soil emergence occurred 5 d after planting the seeds.
î80-I
.l--'tB60.-i
Ë l¡i
Ë ä40z\ (9xãv-
920
0

77
4.9.2 Uptake and Translocation of [lag¡Oictofop-Methyl
To quantify the rates of uptake and Eanslocation of diclofopmethyl, radiolabelled herbicide
was applied to the leaf axil of seedlings at the two-leaf stage. The amount of [tag¡6¡clofop-
methyl absorbed into treated plants was mærimal after ?.4hin both biotypes (Figure 4.2).
The pattems of [l4C1¿¡clofop-methyl uptake in the resistant and susceptibte biotypes rvere
similar with SOVo of uptake evident 6 h following treatment. The amount of radioactivity
that could be washed from the leaves declined rapidly and reached 0 at 180 h after
application. Not all of the label appeared in the extract with significant losses occurring due
either to volatilization, or binding of the label to insoluble material.
For the fi¡st 12 h after treatment, more than SOVo of the radioactivity absorbed was found in
the stem (Figure 4.34). After this time, increasing amounts of label were observed in the
second leaf until about 50Vo of the radioactivity as in the second leaf from 48 h (Figure
4.38). Little radiolabel was detected in the first or third leaves at any time @gure 4.3D-E).
After 6 h exposure, l%o and 3Vo of the radiolabel absorbed was located in the shoot
meristematic regions of the susceptible and resistant biotypes respectively and then rapidly
declined (Figure 4.3C). Although the radiolabel in the shoot meristematic region of the
susceptible biotype was higher than that in the resistant, they were not significantly different.
At no time were there clear differences between the resistant and susceptible biotypes.
Although the total amount of radiolabel recovered in the susceptible and resistant biotypes
during the period of the experiment did not differ, the susceptible plants showed necrosis on
the first, second and third leaves whereas the resistant plants showed some chlorosis at 24 h
at the point of application but newly emerged leaves were unaffected. Very little
radioactivity lvas found in the roots of either biotype (data not shown).

78
100
80
60
40
20
0
E(¡).--ÈÊG-ctto€tHo
Ba
J.l.lIIGo.-EÉË
ú
061218z-4896144192Time after application (h)
Figure 4.2 The absorbed (O, O) and unabsorbed (1, tr) radioactivity in the seedlings of
susceptible SAS 2 (open symbols) and resistant SAS I (closed symbols) A. sterilis. Each
point is the mean of two experiments with two replicates.

79
100
50
100
50
0
20
10
0
10
0
0q)¡cl-È=-cl€o.aJ
tHo
èa
tal
.¡tIcúo.tE6É
ú
10
006121824 48 96
Time after application (h)
Figure 4.34-E Distribution of radioactivity following application of [l4ç¡*.lofop to the
leaf ædls of the susceptible SAS 2(O) and resistant SAS I (O) A. sterilis; stem (A), second
leaf (B), shoot meristematic region (C), fust leaf (D) and third leaf (E). Each point is the
mean of two experiments with two replicates. The thi¡d leaf of resistant plants emerged after
48 h but the thi¡d leaf of susceptible plants did not emerge until 140 h. Vertical bars represent
standa¡d enors and are sometimes obscured by symbols.
192
A
B
c
D
E

80
4.3.3 Metabolism of [14C]n¡ctofop-Methyl
[laC]Diclofop-methyl was converted to diclofop acid at a similar rate in both susceptible and
resistant biotypes. Conversion of diclofop-methyl to diclofop acid was rapid with less than
lÙVo of the applied radioactivity recovered as diclofop-methyl in both biotypes 12 h after
treatment @igure 4.4A). The amount of radioactivity recovered as diclofop acid, the
phytotoxic form of the herbicide, reached a peak 6 h after treatment. The level of
[l4C]diclofop acid rapidly decreased to 20Vo after 48 h. Little label was recovered as
ll4Cldictofop acid in either biotype 180 h after treatment (Figure 4.48). HPLC elution
profiles suggest that diclofop acid was metabolized to similar polar compounds in both
biotypes (Figure 4.5). At 180 h after treatment, more than 95Vo of the radioactivity
recovered from both biotypes was in the form of metabolites of diclofop acid with no
differences between the two biotypes (Figure 4.4C). Therefore, in both resistant and
susceptible biotypes, diclofop-methyl was rapidly converted to diclofop acid, which was in
turn converted to more polar metabolites. Although most of the herbicide in both biotypes
was metabolized, only the susceptible plants were killed by herbicide application.
The HPLC elution profiles of the parent herbicide a¡rd its metabolites at24h after treatment
were similar for both susceptible and resistant biotypes (Figure 4.54-8). Although the
elution profile of diclofop acid and metabolites was simila¡ between susceptible and resistant
wild oat (Figure 4.54-8), cultivated oat (4. sativa cv. Echidna) showed a different pattern
with more polar metabolites eluting between 2 and l0 min than either wild oat biotype
(Figure 4.5C). Metabolism occurred in both the shoot meristematic regions and in the
leaves. In translocation experiments, about 25Vo of radiolabel in the leaves was diclofop
acid in both biotypes 48 h after application and the HPLC elution profiles were similar (data
not shown). Therefore, differences in the amount of unmetabolized herbicide at the
meristem cannot account for resistance.

81
A
0
0612182A-4896144192
Time after application (h)
Figure 4.4L-C Percentage of [laqdt.lofop-methyl (A), diclofop acid (B) and turther
metabolites (C) in the susceptible SAS 2 (O) and resistant SAS I (O),A. stertHs extracted
from 0 to 180 h after treatment. Each point is the mean of two experiments with two
replicates. Venical bars represent standard enors and are sometimes obscr¡red by symbols.
100
t0
60vt-Ë40€Xo20-I
äos 100CH
:BoB\
ã60.-
Ë40GIoË206l¡rts0Ë 100o.i
=80Êa-L.Ë60ä
40
20
B
c

82
:rA
susceptible wild oat
:l€B
resistant wild oat
{(
c
oat cv. Echidna
Ð.--taLGIL-.l¡¡ictl
Ì.l
.l
-9cËo.-E6lú
0 5 10 15 20
Elution time (min)25 30
Figure 4.54-C HPLC elution profiles of extracts from susceptible SAS 2 (A), resistant
SAS 1 (B) ,4. sterilis, and cultivated oat .4. sativa cv. Echidna (C) 24 h after trearnent with
l14C]diclofop-methyl. ll4C]piclofop acid eluted at the retention time of 26 min (*).

83
4.3.4 Effect of Diclofop Acid on Plasma Membrane Potential
In the presence of 50 ¡rM diclofop acid, the plasma membrane potential of etiolaæd coleoptile
cells from both susceptible and resistant wild oat depolarized within 15 min @igure 4.6).
After removal of diclofop acid, the plasma membrane potential remained depolarized for
more than 20 min for both biotypes (Figure 4.64-8). Therefore in this biotype of wild oat,
resistance to diclofop does not correlate with recovery of the plasma membrane electrogenic
potential following removal of diclofop acid.
4.3.5 Inhibition of ACCase Activity Iz Vivo
Translocation experiments revealed that the herbicide concentration reached a mærimum in
the meristem of both resistant and susceptible biotypes 6 h after treatment (Figure 4.3C).
Hence, following herbicide application to intact seedlings, invivo ACCase activity from the
susceptible biotype should become inhibited due to the susceptibility of this enzyme to
ACCase-inhibiting herbicides. However, as the ACCase in the resistant biotype is much less
sensitive to these herbicides (Table 3.5), in vivo ACCase activity should be less inhibited.
To measure this directly, plants were Eeated with herbicide and then the activity of ACCase
from crude extract of the meristems was measured at l, 3, 6,48, 140 and 180 h after
herbicide application. These experiments revealed that extractable ACCase activity was
reduced by SOVo for the susceptible biotype 6 h after treatment whereas ACCase activity
extracted from the treated resistant biotype was reduced by only 3OVo (Figure 4.7). The
extractable ACCase activity in the susceptible biotype remained inhibited up to 180 h after
treatment. However, in the resistant biotype, extractable ACCase activity was only reduced
in the ftrst?4 h after treatment and completely recovered within 180 h. This in vivo result
confirmed the in vitro measurements demonstrating strong inhibition of ACCase activity
from the susceptible but not resistant plants (Table 3.5) and allows the conclusion that
resistance to ACCase-inhibiting herbicides in the resistant biotype is endowed by a mutant
form of ACCase which is markedly less sensitive to inhibition by herbicides.

84
20 30
mtn
Figure 4.64-B The effect of 50 pM diclofop acid on plasma membrane potential in
coleoptile cells of the susceptible SAS 2 (A) and resistant SAS I (B) A. sterilis. Arrows,
addition (+) and removal (-) of diclofop acid (DA). The data shown is from a single
coleoptile and is representative of ten coleoptiles each of the susceptible and resistant
biotypes.
-120
-90
-60
-30
0
-tI
-CE.-Ð-:()*aoÈc)IGL¡-tI()Hà
-120
-90
-60
-30
0100
+ A+DA
.DA
¡
- +DA B
DA
I
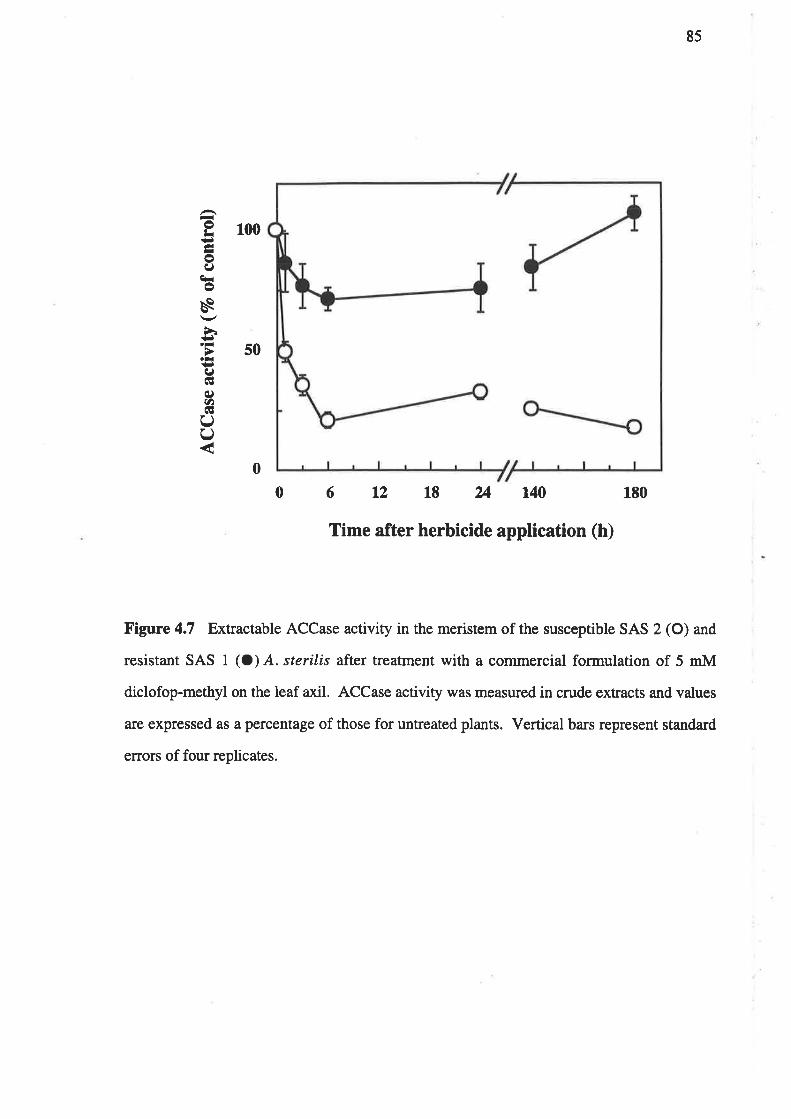
85
100
50
061218z-140 180
Time after herbicide application (h)
Figure 4.7 Extractable ACCase activity in the meristem of the susceptible SAS 2 (O) and
resistant SAS I (O) A. sterilis after treatment with a cornmercial formulation of 5 mM
diclofop-methyl on the leaf axil. ACCase activity was measured in crude extracts and values
are expressed as a percentage of those for untreated plants. Vertical bars represent standard
errors of four replicates.
-oLr-toICIoèa
Ða-
a-IIcË(¡)trhG()L)
0

86
4.4 Discussion
There were no differences in diclofop absorption, translocation or metabolism between the
resistant A. sterìlis biotype SAS 1 and susceptible SAS 2 (Figures 4.2, 4.3A-E and 4.4).
Therefore, these potential resistance mechanisms do not account for diclofop-methyl
resistance in this biotype. To date, there have been no reports of APP or CHD resistance
due to altered absorption or translocation of these herbicides in the related species A. fatua
(Devine et al. 1992). Similarly, absorption and translocation of diclofop-methyl a¡e simila¡
for susceptible and resistant biotypes of L. rigidum (Holtum et al. 1991), L. multiþrum
(Gronwald et ^1.
L992) and S. viridis (Marles et al. 1993).
Häusler et al. (1991) reported that in a number of biotypes of L. rigidurn resistance to APP
and CHD herbicides was correlated with the ability of the plasma membrane electrogenic
potential to recover from diclofop acid-induced depolarization following removal of the
herbicide. A similar situation has been observed for a resistant biotype of A. fatua (Devine
et al. 1992). In this study, neither the resistant nor susceptible A. sterilis biotypes exhibited
recovery of the plasma membrane potential following removal of diclofop acid (Figure 4.6).
Therefore, resistance to APP herbicides in biotype SAS I differs from resistance in the
resistant biotype of A. fatw (Devine et al. 1992) in that it does not correlate with recovery of
electrogenic membrane potential. This phenomenon will be considered at greater length in
Chapter 8.
In certain tolerant maize cell lines selected in vitro, resistance to APP and CHD herbicides is
due to the overproduction of the normal, sensitive ACCase (Parker et al. 1990b). The 58-
and 9O-fold resistance to sethoxydim and haloxyfop, respectively, in the selected line B50S
was conferred by a 2.6-fold increase in herbicide-sensitive ACCase activity. However, in
biotype SAS 1, differences in the activity of the target enzyme, ACCase, cannot explain
resistance as the level of activity in the susceptible and resistant biotypes were similar at all
stages of development (Figure 4.1). The distribution of ACCase activity in shoot meristem,
stem and leaf lamina in the resistant and susceptible biotypes was also simila¡ (Table 4.1).

87
However, ACCase activity in panially-purifred exEacts from the resistant biotype was much
less susceptible to inhibition by APP herbicides than that of the susceptible (Tables 3.5 to
3.e).
At the whole plant level, the resistant biotype exhibits a high level of resistance to APP
herbicides and slight resistance to CHD herbicides (Mansooji et al. 1992). This difference
between herbicide groups is reflected at the ACCase level with the level of resistance of the
enzyme being greater for APP herbicides (9- to 52-fold) than CHD herbicides (6- to 8-fold)
(Tables 3.5 to 3.9). Furthennore, when ACCase was extracted from tissue of susceptible
and resistant plants at various periods after herbicide treatment, the enzyme extracted from
susceptible plants was found to be inhibited to a gteater extent and for a longer period than
that of the resistant (Figure 4.6). Therefore, there is a good correlation between the
inhibition of ACCase activity in vivo and in vitro (Table 3.5 and Figure 4.7). Clearly, the
effects of herbicides on ACCase activity observed both in vivo and in vitro provide direct
evidence that the resistance mechanism in this resistant biotype involves a modified form of
ACCase. Preliminary genetic studies with this resistant A. sterilis biotype suggest that
resistance is endowed by a single, major nuclear gene (Barr etal.1992} It is likely that this
single gene is that encoding for the resistant ACCase enzyme in the biotype SAS l.

88
Chapter 5
Herbicide Resistance Endowed by an APP- and CHD-Inseruitive
ACCase in Biotype NAF 6
5.1 Introduction
Many of the wild oat biotypes examined in Chapter 3 showed resistance to APP herbicides
due to an APP-insensitive ACCase. A few of these biotypes show slight resistance to the
CHD herbicides, however, biotype NAF 6 is moderately resistant to both APP and CHD
herbicides. The mechanism of resistance to herbicides in this resistant biotype NAF 6
appears due to a modified target enzyme, ACCase, which is moderately resistant to both
APP herbicides and CHD herbicides (Chapter 3). This is the only wild oat biotype to show
substantial resistance to CHD herbicides.
The mechanisms providing resistance to CHD herbicides have been elucidated in a number
of studies. Non-regenerable muze cell lines selected with sethoxydim were resistant due to
an overproduction of the target enzyme, ACCase (Parker et al. 1990b). Resistance to
sethoxydim n Inlium rigidum biotype SLR 3 is due to a modified form of ACCase (Tardif
et al. 1993), as is resistance ín Setaria viridis (Marles et al. 1993). A L2Vo subset of L.
rigidum biotype SLR 3l is highly resistant to sethoxydim and other ACCase-inhibiting
herbicides and contains a resistant ACCase target enzyme (Tardif and Powles 1994),
however, the remainder of the population is susceptible to this herbicide. It seems therefore
that most cases of resistance to CHD herbicides, and especially sethoxydim, is conferred by
an insensitive ACCase. Although target site modification appeÍìrs to confer resistance to
CHD herbicides in some species, there may be more than one mechanism of resistance to
CHD herbicides in biotype NAF 6.
To verify whether other mechanisms were contributing to resistance in biotype NAF 6,
uptake, translocation and metabolism of diclofop were examined in both susceptible SAS 2

89
and resistant NAF 6 biotypes. Uptake and metabolism of sethoxydim in this biotype were
also investigated.
5.2 Results
5.2.1 Uptake, Translocation and Metabolism of [14C]O¡ctofop-Methyl
Foliar absorption of diclofop-methyl occurred readily in both susceptible and resistant wild
oat biotypes (Figure 5.1). At I and 6 h after treatment, 25 and427o of the radioactivity was
absorbed in the resistant biotype NAF 6 compared with 27 and 45Vo for the susceptible
biotype SAS 2. Absorption of radioactivity reached a mærimum at 80Vo by 24 h after
application in both biotypes.
The distribution of radioactivity between the meristematic region and the rest of the plant was
found to be identical in both susceptible and resistant biotypes. Translocation of
[lac]diclofop was limited with approximately lOVo reaching the meristematic tissue in both
biotypes by 6 h after application (Figure 5.2A). Most of the radioactivrty absorbed (-907o)
was found in the rest of the foliage (Figure 5.28).
The rate of metabolism of llaC]¿ictofop-methyl was identical in both susceptible and
resistant biotypes (Figure 5.3). Metabolism of diclofop-methyl by susceptible and resistant
plants was nearly complete within 48 h (Figure 5.34). Diclofop-methyl was rapidly
converted to the phytotoxic form, diclofop acid in the fust 6 h following treatment (Figure
5.3B). At72 h after treatment, more than SOVo of the label recovered from both biotypes
was in the form of metabolites of diclofop acid with no differences between two biotypes
(Figure 5.3C). The major pathways for metabolism of diclofop-methyl appear to be similar
in the susceptible SAS 2 and resistant NAF 6 biotypes.

90
Time after application (h)
Figure 5.1 The absorption of radioactivity following application of [lag¡diclofop-methyl
to the axil of two-leaf stage seedlings of susceptible SAS 2 (O) and resistant NAF 6 (O)
wild oat biotypes. Points are the mean t standard error of three replicate experiments.
100
80
60
40
20
0
Eq).I
-ÈÈ6l
-6tto¡fHo
èa
-.-.--IGIo.-EcÉ
ú
A0 7248

9l
20
10
0
100
80
60
40
20
(¡),¡lcÉÐÈ!a-
-acË
-ÌCHoèa
-.ã-CJ6úoEc!ú
0
u18t260
Time after application (h)
Figure 5.24-B Radioactivity found in the meristematic region (A) and the rest of the
foliage (B) of susceptible SAS 2 (O) and resistant NAF 6 (O) wild oat biotypes following
application of [l4c]diclofop-methyl to the axil of nvo-leaf seedlings. Point symbols are the
mean of five replicate experiments. Vertical bars represent the standard errors of the mean
and a¡e sometimes obscured by point symbols.
A
I I
B
I

92
100
80
60
40
20
0100
80
60
40
20
0100
80
60
40
20
0u 48 72
Time after application (h)
Figure 5.34-C Percentage of extracted radioactivity as diclofop-methyl (A), diclofop
acid (B) and further metabolites (C) following application of ¡l4cldiclofop-methyl to the
susceptible SAS 2 (O) and resistant NAF 6 (O) biotypes of wild oat up to 72 h after
treatnent. Each point is the mean of th¡ee replicates. Vertical bars represent standard errors
and are sometimes obscured by symbols.
et)
-IG'LÈX(¡)
-L.l
-ÉË
-o-CHo
èa
5.I
.--IeÉo.lËc!¡rr:o-I
.tÐI-¡
alL-(t2â
0
A
B
c

93
5.2.2 Upteke and Metabolism of [4-149]Sethoxydim
Absorption of [4-laC¡sethoxydim was identical in both susceptible SAS 2 and resistant
NAF 6 biotypes. Absorption of [4-1aC¡sethoxydim, when applied onto the leaf axil of two-
leaf stage seedlings, increased rapidly from I to 6 h after treatment and reached a maximum
of about 9OVo by 72 h after herbicide application (Figure 5.4). The susceptible biotype
SAS 2 absorbed slightly more radioactivity than did the resistant biotype NAF 6, however,
this difference was not significant.
The parent herbicide, ¡4-laClsethoxydim, was metabolized by both biotypes at the same rate
(Figure 5.54). By t h after treatment, only 30Vo of the radioactivity was recovered as
sethoxydim and by 24 h very little sethoxydim was detected. Sethoxydim was rapidly
converted to a metabolite (Ml) which was more polar than sethoxydim (Figure 5.6). This
metabolite reached a maximum concentration 6 h following application (Figure 5.5B) and
was subsequently converted to other products. Radioactivity in other peaks was summed as
polar metabolites (Figure 5.5C). These metabolites increased very rapidly from I to 36 h
after treatment and represented 85Vo of the radioactivity in the extract at the end of the
sampling period @igure 5.5C). There was no difference between susceptible SAS 2 and
resistant NAF 6 in the amount of either the major metabolite or the other polar metabolites
produced. HPLC chromatograms of sethoxydim and metabolites were identical in the
susceptible and resistant biotypes as shown in Figure 5.64-8.

94
00
80
60
40
20
0
1Eq)al
-ÈÈeÉ
-ÉÉ€oICEoñÐ.-.l
-Ic!o
.lEc!ú
72
Time after application (h)
Figure 5.4 Absorption of [4-14c]sethoxydim by susceptible SAS 2 (O) and resistant
NAF 6 (O) wild oat biotypes following application to the axil of two-leaf stage seedlings.
Point symbols are the mean * standard error of three replicate experiments.
5436180

95
100
80
60
40
20
0100
80
60
40
20
0100
80
60
40
20
0
E)-IqB¡r-Xq)
-I.--6!-otÞ
cHo
Ba
>¡-.-a--c.)clo.l€ÉtlL
CHo-Io.----I
li€ct2.-â
54
Time after application (h)
Figure 5.54-C Metabolism of [4-l4c¡sethoxydim in the susceptible SAS 2 (O) and
resistant NAF 6 (O) wild oat biotypes when applied to the leaf axil at the twoleaf stage.
Percentages of [4-l4g]sethoxydim (A), metabolite Ml (B) and other metabolites (C) in the
extracts. Points are the mean t standard efrors of the experiment with four replicates. Error
ba¡s a¡e sometimes obscured by point symbols.
720 18 36
A
B
c

96
*
*
^^^^
A
susceptible wild oat
:lc
*
B
resistant wild oat
Ð.l-tãliÉËL-.Fl¡L(!I.-.-ùI6!o.-EcË
ú
051015 30
Elution time (min)
Figure 5.64-8 HPLC elution profiles of the extracts from the susceptible SAS 2 (A) and
resistant NAF 6 (B) t h after treatment with ¡4-laClsethoxydim. Metabolite (Ml) and
sethoxydim eluted at a retention time of 13 (tt) and 20 min (*), respectively.

97
5.2.3 Effect of Diclofop Acid on Plasma Membrane Potential
To determine whether an altered plasma membrane response to diclofop acid is a mechanism
conferring resistance in this biotype, the plasma membrane potential of etiolated coleoptile
cells of both susceptible SAS 2 and resistant NAF 6 were measured. In the presence of
50 pM diclofop acid, the plasma membrane potential from both susceptible and resistant
biotypes was depolarized within 15 min (Figure 6.6,{-8). After removal of diclofop acid,
the plasma membrane potential from both biotypes remained depolarized for more than
20 min. Hence, in this biotype resistance is not correlated with a recovery of the plasma
membrane electrogenic potential following removal of diclofop acid.
5.3 l)iscussion
The mechanism of resistance to APP herbicides in biotype NAF 6 does not involve changes
in uptake, translocation or metabolism of diclofop-methyl (Figures 5.1 to 5.3) or an
alteration of the plasma membrane response to diclofop acid (Figure 5.6). In addition,
resistance to CHD herbicides in this biotype is not due to reduced uptake or enhanced
metabolism of sethoxydim (Figures 5.4 and 5.5). A modified target site, ACCase enzyme,
appe¿us to be the only mechanism responsible for resistance for both APP and CHD
herbicides in this biotype (Chapter 3). The ACCase enzyme from this resistant biotype is 3,
4 and 3 times more resistant to diclofop acid, fluazifop acid and haloxyfop acid, respectively
(Tables 3.5 to 3.7) and is 7 and 4 times more resistant to sethoxydim and tralkoxydim
(Tables 3.8 and 3.9) than that of the susceptible biotype.
The level of resistance to diclofop-methyl may be correlated with the history of herbicide
application. Herbicide resistance to ACCase-inhibiting herbicides has been shown to occur
after three years or more of herbicide application (Tardif et al. t993, Devine and
Shimabukuro 1994). Exposure to APP herbicides for five years in biotype SAS I and nine
years in biotype NAS 4 resulted in a high level of resistance of both biotypes to diclofop-
methyl (Table 2.l,Table 3.10). In contrast, biotype NAF 6 had been exposed to only two
applications of APP herbicide before developing a moderate level of resistance to diclofop-

98
-LI-Grt-I()i¡cÈé)I6lL¡-Iq)
FTÀ
-100
-80
-60
-120
-120
0
-100
-80
-60
00 10 20
Time (min)
30
Figure 5.64-B The effect of 50 pM diclofop acid on plasma membrane potentials in
coleoptile cells of the susceptible SAS 2 (A) and resistant NAF 6 (B) biotypes. Arrows,
addition (+) and removal (-) of diclofop acid (DA). The data shown is from a single
coleoptile and is representative of ten coleoptiles each of the susceptible and resistant
biotypes.
+DA
ü A
.DA
,
+DA
ü B
.DA
+
,I I

99
methyl. It is difficult to correlate resistance to CHD herbicides with herbicide use histories.
Biotype NAF 6 had not been exposed to CHD herbicides yet it exhibited a moderate
resistance to CHD herbicides (cycloxydim, sethoxydim and tralkoxydim). In contrast,
biotype NAS 4 had been exposed to two applications of CHD herbicides but this biotype is
still sensitive to tralkoxydim (Table 3.10). In the case of biotype NAF 6, it is clea¡ that
selection by diclofop-methyl has concomitantly provided target site cross-resistance to CHD
herbicides. This does not always occur and many wild oat biotypes only have resistance to
APP herbicides.
Some resistant biotypes possessing resistant ACCase are highly resistant to both APP and
CHD herbicides. For example, a high level of resistance to APP and CHD herbicides has
been described in Setaria viridis (Marles et ai. 1993) and Z. rigidum (Tardif et al. 1993,
Tardif and Powles 1994) based on a modified form of ACCase. Yet, wild oat biotypes
SAS I and NAS 4 that possess APP-resistant forms of ACCase are highly resistant to APP
herbicides only. A similar pattern is also evident in a resistant biotype of. L. multiþrum
(Gronwald et al. L992). Biotype NAF 6 also contains an APP-resistant ACCase but it is
moderately resistant to both groups of herbicides.
The ACCase from biotype NAF 6 appears to be different to that of biotypes SAS I and NAS
4 since it exhibits moderate resistance to both APP and CHD herbicides. Probably, the
mutations of the herbicide binding sites within ACCase are not the same among the three
resistant wild oat biotypes. Modified forms of ACCase from mutantmuze cell cultures,
selected by sethoxydim or haloxyfop are encoded by different allelic mutations at the same
locus (Marshall et al.1992). These data indicate that there a¡e several potential modifications
of ACCase which can confer resistance to APP and CHD herbicides. The differences in
sensitivity of resistant forms of ACCase to CHD herbicides indicates that the CHD and APP
herbicides probably bind to overlapping but non-identical sites on the enzyme. In the ten
resistant biotypes examined in this thesis, modifications of ACCase which confer resistance
to APP herbicides only are more common than those which confer target site cross-
resistance to CHD herbicides (Chapter 3).

100
Chapter 6
Resistance to APP Herbicides Endowed by Two Mechanisms in
Biotype NAS 4
6.1 Introduction
At the whole plant level, wild oat biotypes NAS 4 and SAS I could not be controlled by the
very high rate of 30,000 g diclofop-methyl ¡¿-l (Table 3.1). However, at the enzyme level,
biotype NAS 4 exhibited an ACCase that was six-fold less sensitive to diclofop (Table 3.6),
whereas the ACCase from biotype SAS I showed 52-fold less sensitivity to diclofop than
that of susceptible plants (Table 3.5). Thus, the modified target site, ACCase enzyme, in
biotype NAS 4 may not be sufficient to confer the high level of resistance observed in the
whole plants. Therefore, studies were undertaken with biotype NAS 4 to establish whether
herbicide resistance in this biotype may involve more than one mechanism.
That different mechanisms of resistance to ACCase-inhibiting herbicides can occur in
populationsof Avena isshownbyacomparisonof Avenasterilis biotypeSAS l whichhas
a mutant ACCase enzyme (Chapter 4) andA.fatuabiotype UM-l which possess alterations
in membrane properties (Devine et al. 1993). Other mechanisms, such as enhanced
metabolism of diclofop a¡e also possible as, has been documented in some Inlium rigidum
biotypes. For example, L. rigidumbiotype SLR31 was able to detoxify diclofop acid at
about 1.5 times of the rate of the susceptible biotype (Holtum et al. l99l). However, to
date, enhanced metabolism of diclofop has not been observed in biotypes of wild oat.
To determine whether non-target site mechanisms co-exist with resistant ACCase in biotype
NAS 4, uptake, tanslocation, and metabolism of ll4c]diclofop-methyl as well as the ability
of plasma membrane to recover from diclofop-induced membrane depolarization by diclofop
acid were investigated.

101
6.2 Materials and Methods
I;.:l¡ I[ tfrilf F,i.jf.å Í-li:iii¡,nY; ;: ühi!'il:fi;ìi'i T fii: fiüf; #li]_.
Most materials and methods are as described in Chapter 2. Additional methods are
described below.
6.2.1 Effects of the Cytochrome P450 Inhibitor Tetcyclasis on Diclofop
Metabolism
To demonstrate the effects of tetcyclasis on the metabolism of diclofop-methyl, wild oat
biotypes NAS 4, SAF 19 and wheat plants were grown in hydroponic solution as described
in section 2.2.3. Tetcyclasis (20 ttM) was added to the nutrient solution 24 h prior to
treatment with I pL of 5 mM ll4C]diclofop-methyl applied to the leaf axil. Plants were
harvested 24 h following herbicide treatment, ground, radioactivity extracted and the
metabolites examined by HPLC. The experiments were a completely randomized block
design with four replicates.
6.2.2 Effects of the Cytochrome P450 Inhibitor Tetcyclasis on Plant
Growth
To determine the effects of tetcyclasis on the growth response of wild oat biotypes, resistant
(NAS 4 and SAS 1), susceptible (SAF 19) wild oat biotypes, and wheat cv. Halberd were
cultured using hydroponics in I L rectangular plastic trays. Each tray contained a single
row of each biotype (six plants/row). The nutrient solution was replaced with solution
containing 20 pM tetcyclasis when plants were at the two-leaf stage. After t¡eatment with
20 pM tetcyclasis for 24 h, a I pL droplet of l0 mM diclofop-methyl (commercial
formulation) was placed on the leaf axil of the two-leaf stage plants. Seven days following
herbicide application, shoot and root length of individual plants tvere measured. Dry weight
of shoot and root were also determined. The experiments were a completely randomized
block design with four replicates.
To determine the growth response of wild oat biotypes to various rates of diclofop-methyl in
the presence and absence of tetcyclasis, susceptible SAF 19 and resistant NAS 4 were

t02
grown in hydroponic culnue and treated with 20 pM tetcyclasis as described above. After
treatment with 20 ¡rM tetcyclasis for 24h, a I ¡tL droplet of 0, 2.5, 5 and l0 mM diclofop-
methyl (commercial formulation) were placed on the leaf æril of the two-leaf stage plants.
Seven days following herbicide application, shoot and root length of individual plants were
measured. Dry weight of shoot and root were also determined. The experiments were a
completely randomized block design with four rreplicates.
6.2.3 Inhibition of ACCase by Diclofop in Single Lines of NAS 4
To test the hypothesis that two mechanisms of resistance to diclofop are present in each
individual of the NAS 4 population, seeds from ten different plants of biotype NAS 4 which
had survived 52 g haloxyfop-ethoxyethyl h¿-l were separately collected and used for
ACCase measurements. Two lines of NAS 4, resistant SAS I and susceptible SAF 19
biotypes were arranged in each assay with four replicates.
6.3 Results
6.3.1 Uptake and Translocation of ¡ugfDiclofop-Methyl
Labelled diclofop-methyl was absorbed rapidly when applied to the leaf axil of two-leaf
stage wild oat seedlings (Figure 6.1) with no differences between the two biotypes. More
than 9OVo of the applied herbicide was observed in the stem following application of
herbicide onto the leaf axil of two-leaf seedlings of both biotypes. The treated area (stem)
contained the major fraction of the ¡l4C1diclofop at atl harvest times (Figure 6.2B). Very
little radioactivity was translocated to the leaf tips and roots of either biotype (Figure 6.2C-
D). Translocation of radiolabel from the treated area to the meristematic region was similar
and reached a maximum of l2%o and lïVo respectively in the susceptible SAF 19 and
resistant NAS 4 biotypes (Figure 6.24). Therefore, uptake and translocation do not
contribute to resistance to diclofop in this biotype. This result is simila¡ to that observed for
biotype SAS I in Chapter 4, however, more radioactivity appeared in the leaves of both
biotypes in that experiment. This may be because the plants in the experiment described
here were growing more slowly.

103
e 1oo.l
-ÈÈcÉ Bo-cl-o-860èa
b40.¡.t-I820.-€GI
ú0
0 44872Tme after application (h)
96
Figure 6.1 Absorption of [l4C]d'clofop-methyl by susceptible SAF 19 (O) and resistant
NAS 4 (O) wild oat biotypes. ll4c]diclofop-methyl was applied to the leaf axil of the two-
leaf stage seedlings. Points are the mean * standard error of four replicate experiments.

104
0612182/4896
Time after application (h)
Figure 6.24-D Distribution of radioactivity in the meristem (A), stem (B), leaf (C) and
root (D) from ¡lag¡diclofop-methyl deposited on the leaf axil of twoJeaf stage seedlings of
the susceptible SAF 19 (O) and resistant NAS 4 (O) wild oat biotypes. Points a¡e the mean
* standard error of five replicate experiments.
20
10
6.04 100
-3Bo-dË60€¡a
:40B\
;20i.l.r
Ëoclo.i
H10ú
0
10
0
A
Hù-o
B
c
D

105
6.3.2 Metabolism of [14C]nictofop-Methyl
In susceptible (SAF 19) and resistant (NAS 4) biotypes, ¡l4c¡diclofop-methyl was
degraded to diclofop acid at an equal rate @igure 6.3A). Radioactivity rapidly accumulated
in the ph¡otoxic form, diclofop acid, with 6OVo of the label in this form t h after treatment
(Figure 6.38). Conversion of diclofop-methyl to diclofop acid was similar in both biotypes
(Figure 6.38), however, the conversion of diclofop acid to the other metabolites was more
rapid in the resistant biotype compared to the susceptible. From 6 to 48 h after treatment,
the susceptible tissue contained between 5 to lsEo more of the radiolabel as ¡laç¡diclofop
acid than did the resistant tissue. The decline in diclofop acid was accompanied by an
increase in more polar metabolites in both biotypes. These metabolites were evident at 6 h
after the treatment in the resistant biotype and were produced more rapidly by this biotype
(Figure 6.3C). By 72 h after treatment, the content of ll4C]diclofop acid had decreased to
11 and l4%o in biotypes NAS 4 and SAF 19, respectively (Figure 5.38) with no differences
between the two biotypes. However, the initial rate of production of polar metabolites wÍrs
1.5 times faster in the resistant biotype compared to the susceptible biotype (Figure 6.3C).
The HPLC elution profiles for the two biotypes at24 h after treatment were similar except
the resistant biotype, NAS 4, had more polar metabolites eluting between 4 to 7 min than
did the susceptible (Figure 6.44-8).
It is clear that the resistant biotype is able to convert diclofop acid to more polar compounds
at a faster rate compared to the susceptible biotype. There are two reactions which may be
involved in this conversion. In wheat, diclofop is predominantly metabolized via an aryl-
hydroxylation reaction catalyzed by a cytochrome P450 (McFadden et aI. 1989, Zimmerlin
and Durst 1992). The arylhydroxylated diclofop is then conjugated to sugars and other
compounds (Shimabukuro et aI. 1979). In some susceptible grass species, diclofop is
predominately metabolized via an ester linkage to a glucose moiety (Donald and
Shimabukuro 1980).

106
100
80
60
40
20
0100
80
60
40
20
0100
80
60
40
20
¡t23IclL9XÉ)
-I.laGI-o-Croñ
-.-.-JIGIoEcúLfr
-Ho.-I--€.-L-çr).-â
072
Time after application (h)
Figure 6.34-C Distribution of radioactivity in diclofop-methyl (A), diclofop acid @)
and other metabolites (C) up to 72 h after trearment with I ¡rL of 5 mM [14C]¿iclofop-
methyl in susceptible SAF 19 (O) and resistant NAS 4 (O) wild oat biotypes. Points are
the mean + standard error of three replicates.
0 u 48
B
c
-r.

107
I.--Ira-LGIliù¡
ol¡LG
t¡.l-¡Ic!o.lE6lú
A
susceptible wild oat{.
B
resistant wild oat ,F
0 5 10 15 20 2s
Elution time (min)
30 35
Figure 6.4.4'-8 HPLC elution profiles of extracts from susceptible SAF 19 (A) and
resistant NAS 4 (B) wild oat biotypes}4hafter exposure to [14C]diclofop-methyl dissolved
in a commercial herbicide formulation. ¡l4g¡¿tclofop acid eluted at the retention time of 26
min (*).

108
6.3.3 Effect of Tetcyclasis on Diclofop Metabolism
To test for involvement of cytochrome P450 enzymes in metabolism of diclofop in wild oat
biotype NAS 4, tetcyclasis, a broad-spectn¡m cytochrome P450 inhibitor @rear et al. 1991,
Mougin et al. 199L, Christopher 1993), was applied in combination with diclofop.
Tetcyclasis was applied24hbefore application of diclofop-methyl to wild oat plants. By
24 h following diclofop-methyl application, the susceptible biotype SAF 19 contained
4L.8Vo of the extactable radioactivity as diclofop acid in plants not treated with tetcyclasis
(Tabte 6.1). Pre-teatment with tetcyclasis did not affect metabolism of diclofop acid in this
biotype with4l.l7o of the extractable herbicide as diclofop acid. Conversely, control plants
of the resistant biotype NAS 4 had only 30.17o of the extractable radioactivity as diclofop
acid. Pre-treating these plants with tetcyclasis significantly reduced metabolism of diclofop
acid such that 39.l%o of the extracted radioactivity uras in this form 24 h after treatment
(Table 6.1). In wheat, which can rapidly metabolize diclofop acid to polar metabolites, the
level of diclofop acid in this tissue was unaffected by treatment with tetcyclasis.
An investigation of the HPLC profiles of the susceptible and resistant biotypes showed a
single obvious difference in metabolites (Figure 6.4A-8). The resistant biotype contained
considerably more metabolites eluting between 4 and 7 min than did the susceptible. These
metabolites are much reduced in the resistant biotype after treatment with tetcyclasis such
that there was little difference between susceptible and resistant biotypes in the amount of
radioactivity eluting in the first seven minutes (Figure 6.5). Treatment with tetcyclasis did
not affect the amounts of any other polar metabolites in either biotype. Although tetcyclasis
did not inhibit the overall rate of metabolism of diclofop acid in wheat plants, there was a
significant difference in the metabolites produced in wheat (Figure 6.64-8). In the
tetcyclasis-treated wheat plants, a reduction in the amount of metabolites eluting between 2
and 5 min and an increase in a metabolite with a retention time of 16 min were observed.

109
Table 6.1 The amount of radiolabel as ¡l4ç¡diclofop acid in the susceptible SAF 19
and resistant NAS 4 wild oatbioqpes at24hafter treatment with 5 mlvt ¡149¡¿¡"lofop-
methyl t 20 FM tetcyclasis. Data a¡e the mean I standa¡d deviation of five replicate
experiments.
TreaEnent Biotpe Diclofop acid(Vo)
Diclofop-methyl SAF 19
NAS 4
wheat
Diclofop-methyl + Tetcyclasis SAF 19
NAS 4
wheat
41.8 f 3.3
30.1 + 2.8
to.2 + 2.o
41.1 r 3.5
39.1 r 3.9
11.4 t 3.0

110
rl.A
susceptible wild oat
{qB
resistant wild oat
€.l-:.¡a-LeÉL-.lÊLcl
-.l.l
-IcËo
OIEcÉ
ú
10 15 20 25 30 35
Elution time (min)
Figure 6.54-8 HPLC elution profiles of ll4c]diclofop-methyl and metabolites in
extracts from susceptible SAF 19 (A) and resistant NAS 4 @) wild oat biotypes 24 h after
treatment with ¡l+gldiclofop-methyl. Prior to treatment with diclofop-methyl, the plants
had been exposed to 20 pM tetcyclasis for 24 h. ¡t+çrdt"lofop acid eluted at the retention
time of 26 min (*).
05

lll
A
*
_^
B
{.
r.lËra-LcËLt¡.!lÊLG
È.-a-!Ð
Iclc.-EÉB
ú
05 10 15 20 25
Elution time (min)30 35
Figure 6.64.8 HPLC elution profiles of ll4C]d'clofop-methyl and metabolites in
extracts from wheat cv. Halberd 24 h after exposure to ll4c]d'clofop-methyl (A) or
diclofop-methyl plus 20 ¡rM tetcyclasis (B). ¡t+g¡diclofop acid eluted at the retention time
of 26 min (*).

tt2
6.3.4 Effects of letcyclasis on Plant Growth
Application of 20 pM tetcyclasis to the nutrient solution of hydroponically-grown plants
partially inhibited diclofop acid metabolism in the resistant biotype NAS 4 (Table 6.1). To
test effects on whole plant $owth, tetcyclasis was applied to the roots of hydroponically-
grown plants in combination with diclofop methyl (topically applied to the axil) to determine
whether inhibition of diclofop acid metabolism leads to an increase in phytotoxic effects of
herbicide application. Three experiments were conducted, the first compared biotype
NAS 4 with wheat cv. Halberd which can rapidly metabolize diclofop (Table 6.1).
Diclofop-methyl, applied alone, had no effect on shoot length or shoot dry weight of
biotype NAS 4 (Table 6.2). In wheat, diclofop-methyl application reduced shoot length but
the shoot dry weight was not affected. Tetcyciasis (20 pM), applied alone via the nutrient
solution of hydroponically grown plants, decreased the shoot length of both species but
shoot dry weight was not significantly different from the control (Table 6.2). Tetcyclasis,
applied alone, had no effect on the length and dry weight of roots in either species.
Diclofop-methyl (10 mM), applied alone, had only a small effect on shoot growth in both
NAS 4 and wheat since they a¡e both resistant to the herbicide. Tetcyclasis, when applied
with herbicide, was able to increase the phytotoxic effect of diclofop-methyl on dry weight
and shoot length in wild oat biotype NAS 4 and on shoot length in wheat cv. Halberd,
however, shoot dry weight of wheat when treated with the combination of diclofop-methyl
and tetcyclasis was not significantly reduced. Although there w¿¡s no mortality caused by
the application of diclofop-methyl and tetcyclasis, the shoot length and dry weight of wild
oat biotype NAS 4 and shoot length of wheat were significantly reduced (Table 6.2). Thus,
tetcyclasis does increase the ph¡otoxicity of diclofop-methyl in wild oat biotype NAS 4.
The second experiment compared biotype NAS 4 with a susceptible biotype SAF 19 and
SAS I, a biotype with a highly resistant ta¡get enzyme ACCase, but no increase in diclofop-
methyl metabolism (Chapter 4). In this experiment (Table 6.3), diclofop-methyl alone had
no effect on biotypes NAS 4 and SAS 1 but caused a substantial reduction in shoot and root
dry weight of biotype SAF 19. Tetcyclasis alone reduced shoot length of all three biotypes

113
but root length of the three biotypes were r¡naffected. A combination of diclofop-methyl and
tetcyclasis had a synergistic effect on reduction of shoot and root length and shoot and root
dry weight of biotype NAS 4. Tetcyclasis in combination with diclofop-methyl also
reduced shoot dry weight in this experiment, however, this reduction uras not significantly
greater than treatment with tetcyclasis. In contrast, treatment with diclofop-methyl plus
tetcyclasis showed no synergistic effects on shoot and root dry weight in the susceptible
biotype. The inhibition of dry weight of shoot and root in the susceptible biotype when
treated with the combination of two chemicals was the result of diclofop-methyl alone.
Diclofop-methyl and tetcyclasis combined had a synergistic effect on shoot and root length
and shoot dry weight in biotype SAS l.
The third experiment examined the responses of susceptible biotype SAF 19 and resistant
biotype NAS 4 to different concentrations of diclofop-methyl when applied with tetcyclasis.
Diclofop-methyl applied at all concentrations did not affect total dry weight of plants of the
resistant biotype, however, treatments as low as 2.5 mIVl reduced the dry weight of the
susceptible biotype to about 40Vo of. the control (Figure 6.7). Diclofop-methyl applied at
l0 mM reduced dry weight of the susceptible SAF 19 to less than 20Vo of the control.
Application of tetcyclãsis in combination with diclofop-methyl did not decrease total dry
weight of susceptible SAF 19 over that acheived by diclofop-methyl applied alone. In
contrast, tetcyclasis applied in combination with diclofop-metþl had a synergistic effect on
total dry weight of the resistant NAS 4 biotype. At 10 mM diclofop-metþI, in the presence
of tetcyclasis, total dry weight was reducedby 4OVo compared to no reduction in the absence
of tetcyclasis @igure 6.7).
These in vitro and in vivo experiments demonstrate that enhanced diclofop metabolism
contributes to diclofop-methyl resistance in biotype NAS 4. Tetcyclasis did not increase
phytotoxicity of diclofop, as determined by plant dry weight, in wheat or susceptible
biotype SAF 19 where it does not decrease diclofop metabolism. However, biotype NAS 4
did not become fully susceptible upon treaünent with tetcyclasis as this biotype also contains
a diclofop-resistant ACCase.

t14
Table 6.2 Effects of diclofop-methyl and tetcyclasis alone and in combination on shoot
and root length (cm) and shoot and root dry maner (mg/planQ of wild oat biotype NAS 4
and wheat cv. Halberd grown in hydroponic culture. Values are the means of four
replicates.
Biotype Control Diclofopmethyl Tetcyclasis Diclofopmethyl +Tetcyclasis
Wild oat biotvoe NAS 4
Shoot length 27 a* 24a r8b 14c
Root length 14a 15a 13a 13a
Shoot dry weight lM a 94a 88a 58b
Root dry weight 44 a 45a 47a 34b
Wheat cv. Halberd
Shoot length 27a 22b 2tb l8c
Root length 13a 14a 12a 12a
Shoot dry weight 143 a I33 a ll0a 102 a
Root dry weight 33 a 3tb 4la 29b
*Values within a row followed with the same letter a¡e not significantly different at p <0.05
according to Duncan's multiple range test.

115
Table 63 Effects of diclofop-methyl and teæyclasis alone and in combination on shoot
and root lenglh (cm) and shoot and root dry matter (mg/plant) of susceptible SAF 19,
resistant SAS I and NAS 4 wild oat biotypes grown in hydroponic culture. Values are the
means of fotu replicates.
Biotpe Contol Diclofopmethyl Teûcyclasis Diclofopmethyl + Tetcyclasis
Susceptible biotvpe SAF 19
Shoot length
Root length
Shoot dry weight
Root dry weight
Resistant biotvoe SAS I
Shoot length
Root length
Shoot dry weight
Root dry weight
29 a* 13b
19a lla
96a 55b
49a 2tb
30a 28a
19a 19a
lO7 a 97a
46a 38b
3la 30a
2la 2Oa
109 a l0l ab
48a 4la
Resistant biotype NAS 4
Shoot length
Root lengfh
Shoot dry weight
Root dry weight
18 ab
17a
62 ab
45a
20b
19a
83a
47a
20b
2ta
84 ab
t2b
10a
40b
20b
14c
t7b
59b
43 ab
l6c
17b
65b
49a 30b
*Values within a ro\ry followed with the same letter are not signifrcantly different at p <0.05
according to Duncan's multiple range test.

116
120
-o¡rI-toICHo
èa
I--è0a-e)Þ
LE-cÉ
-oFr
80
40
050 10
Diclofop-methyl (mM)
Figure 6.7 Total dry weight of susceptible SAF 19 (O, O) and resistant NAS 4 (1, tr)
wild oat biotypes grown in hydroponic culture when treated with diclofop-methyl alone
(closed symbols) and in the presence (open symbols) of 20 ttM tetcyclasis (Tet). Diclofopr
tetcyclasis data are a%o of. the tetcyclasis alone to give lOÙVo values. Point symbols are the
mean t standard error of four replicates.
- Tet
+ Tet
+ Tet
- Tet

t17
6.3.5 Inhibition of ACCase Activity by Diclofop Acid in Single Lines of
Biotype NAS 4
Biotype NAS 4 has two mechanisms of resistance to diclofop-methyl, a less-sensitive
ACCase (Chapter 3) and enhanced metabolism of diclofop (Figure 6.3). To establish
whether both mechanistns are present within all individuals of the population, seed from ten
individual plants were collected and each family tested for the presence of a diclofop-
resistant ACCase. These lines were compared to biotype SAF 19, with a sensitive ACCase,
and biotype SAS 1, with a highly-resistant ACCase. As expected, ACCase from SAS 1
biotype exhibited 50-fold less sensitivity to diclofop inhibition compared to that of
susceptible SAF 19 biotype. This level of resistance is simila¡ to reported in Chapter 3
when ACCase from biotype SAS 1 was compared to that of susceptible biotype SAS 2.
Concentrations of diclofop acid giving 507o inhibition (I5g) of ACCase activity from the ten
families of biotype NAS 4 ranged from 1.2 to 3.0 pM. The I59 of ACCase from these NAS
4 lines were between 6 to 15 times greater than that of the susceptible (Table 6.5). In
previous studies, ACCase extracted from the whole population of NAS 4 biotype was found
to be six-fold more resistant to diclofop than the susceptible (Table 3.6). These small
differences of R/S values between the single lines and bulk population suggested that the
population of NAS 4 is homogeneous and all individuals contain a resistant ACCase. The
6- to l5-fold resistance of ACCase to diclofop in the single lines and in the bulk population
of NAS 4 biotype is probably insufficient to account for the high degree of resistance to
diclofop-methyl evident at the whole plant level. For example, biotype NAS 4 shows
greater resistance to diclofop-methyl than biotype SAS I despite possessing an ACCase 3-
to 8-fold less resistant. It is likely that resistance to diclofop-methyl in this biotype involves
two mechanisms, a mutant ACCase enzyme and enhanced metabolism to detoxify the
herbicide, and these two mechanisms of resistance co-exist in all individuals.

118
Table 6.5 Concentration of diclofop acid giving 507o inhibition of ACCase activity (Iso)
from the susceptible SAF 19, resistant SAS I and l0lines of resistant NAS 4 (#1410) wild
oat biotypes. Ratios are the value for the resistant divided by the value for the susceptible
biotype. ACCase activity was measured in partially purified extracts of the shoot
meristematic regions. Data are the mean t standard error of four replicates.
Biotype Iso 0¡¡¡) R/S Ratio
Susceptible SAF 19
Resistant SAS 1
Resistant NAS 4
#l
#2
#3
H
#5
#6
#7
#8
#9
#10
0.20 r 0.04
9.96 t 1.1
2.48 + 0.5
3.01 r 0.7
2.08 + 0.6
1.96 + 0.5
1.38 r 0.6
1.28 + 0.5
1.19 r 0.6
1.98 + 0.4
1.82 + 0.6
2.11 r 0.6
49.8
12.4
15.0
10.4
9.8
6.9
mean (#1+10)
6.4
5.9
9.9
9.1
10.5
1.93 + 0.18 9.6 + 2.6

119
6.3.6 Effect of Diclofop Acid on Plasma Membrane Potential
The ability of diclofop acid to depolarize plasma membrane potentials in etiolated coleoptile
cells of susceptible SAF 19 and resistant NAS 4 wild oat biotypes was examined. Plasma
membrane potentials of untreated coleoptile cells from susceptible and resistant biotypes
were between -110 to -l15 mV (Figure 6.8). Diclofop acid (50 pM) depolarized plasma
membrane potentials from both biotypes within 15 min to between -50 and -60 mV (Figure
6.3A-8). After removal of herbicide, plasma membrane potentials from both biotypes
stayed depolarized for at least 20 min. The resistant biotype NAS 4 was unable to repolarize
the plasma membrane potential following diclofop acid-induced depolarization.
6.4 Discussion
rWild oat biotype NAS 4 shows high level resistance to diclofop-methyl, but possesses only
a moderately-resistant ACCase (Chapter 3). Studies revealed that there were no differences
in uptake and translocation of diclofop-methyl between susceptible and resistant biotypes
(Figures 6.L and 6.2). Nor does this biotype possess a mechanism which allows rapid
repolarization of the plasma membrane potential (Figure 6.8). Biotype NAS 4 possesses an
ACCase with 6-fold resistance to diclofop acid (Chapter 3). In addition, this biotype
exhibits an increase in the rate of diclofop-methyl metabolism (Figure 6.34-C). A similar
result was reported for L. rigidum biotype SLR 3l which exhibited a l.5-fold increase in
metabolism of diclofop-methyl (Holtum et al. 1991). In vivo studies with whole plants
show that the level of diclofop-methyl resistance in NAS 4 biotype is reduced in the
presence of the cytochrome P450 inhibitor, tetcyclasis, whereas tetcyclasis has little effect
on the response of the susceptible biotype to diclofop-methyl (Table 6.3). The rate of
metabolism of diclofop acid in biotype NAS 4 decreased in the presence of tetcyclasis.
'When plants were treated with tetcyclasis 24 h prior to application of ¡l4c1diclofop-methyl,
metabolism of diclofop in the resistant biotype was reduced to the level of the susceptible
biotype (Table 6.1, Figures 6.5 and 6.7). These results suggest that the enhanced
metabolism of diclofop-methyl observed in biotype NAS 4 is probably due to a cytochrome

t20
-120
-E-cl.---I(¡)
-ofe)-tcl¡i
--IIq)|=À
.100
-E0
-60
-120
-100
-80
-60
0
00 10 20
Time (min)30
Figure 6.84-B The effect of 50 pM diclofop acid on plasma membrane potentials in
coleoptile cells of the susceptible SAF 19 (A) and resistant NAS 4 (B) biotypes. Arrows,
addition (+) and removal C) of diclofop acid (DA). The data shown is from a single
coleoptile and is representative of ten coleoptiles each of the susceptible and resistant
biotypes.
+DA
üA
DA
,I
- +DA
ü B
.DA
+
,I

tzt
P450 monooxygenase. Although wheat contains a P450 eni¿)me able to rapidly metabolize
diclofop-methyl (Zimmerlin and Drust 1990 and 1992) and tercyclasis was able to enhance
the phytotoxicity of chlorotoluron towa¡d wheat (Mougin et al. 1991), diclofop metabolism
in wheat cv. Halberd was not affected by tetcyclasis (Table 6.1). It may be that tetcyclasis
is not an effective inhibitor of diclofop metabolism in wheat or the concentration used was
not sufflrcient to inhibit the detoxification of diclofop in wheat. Specific herbicide-inhibition
interactions were evident in L. rigidum where tetcyclasis does increase chlorotoluron
phytotoúcity in biotype WLR 2 but does not affect the phytotoxicity of chlorsulfuron in
biotype SLR 3l (Christopher et al. 1994). Nevertheless, more work has to be done to
establish the direct involment of P450 enzymes in enhanced metabolism of diclofop-methyl
in biotype NAS 4.
It is likely that two mechanisms of resistance are present in each individual of the NAS 4
population (Table 6.4). The combined contributions of enhanced metabolism and mutant
ACCase enzyme to herbicide resistance may be required to explain the high degree of
resistance at the whole plant level. Hence, it is probable that both mechanisms contribute to
diclofop-methyl resistance in this biotype. Resistance to diclofop-methyl endowed by a
resistant ACCase tn A. sterilis and Z. multiforum is conferred by a single, nuclear encoded,
incompletely dominant gene (Barr et al.1992, Betts et al.1992). In case of biotype NAS 4
which has two mechanisms responsible for resistance, there must be more than one gene
involved. Definite proof of a number of genes conferring resistance in this biotype has yet
to be determined.
Resistant ACCase is a mechanism frequently found in species resistant to ACCase-inhibiting
herbicides, whereas non-target site resistance mechanisms, such as enhanced metabolism of
herbicide, a¡e seldom observed. However, a 1.5 fold increase in the rate of metabolism of
diclofop-methyl has been observed in one biotype of L. rigidum (Holtum et at. 1991) which
probably confer at least partial resistance in this biotype on its own. Where resistant weeds
have at least two mechanisms of resistance, target site-based resistance will most often be
the most significant mechanism (Devine and Shimabukuro 1994). Therefore, biotype

t22
NAS 4 not only has an insensitive target ACCase, albeit not sufficient to confer the high
level of resistance to diclofop-methyl at the whole plant, but it also has an increased capacity
to metabolize diclofop-methyl. Together, these two mechanisms enable the NAS 4 plants to
resist high rates of the herbicide. This is the first evidence for wild oat having two
mechanisms of resistance (target and non-target site based) to ACCase-inhibiting herbicides.
ln L. rigiduzr, biotype SLR 3l has three mechanisms of resistance, an enhanced metabolism
to detoxify diclofop-methyl, an alteration of plasma membrane, and resistant ACCase
(Häusler et al. 1991, Holn¡m et al. 1991, Tardif and Powles 1994). However, orùy a L27o
subset of this biotype contained the insensitive ACCase (Tardif and Powles 1994). In
contrast, it is likely that both mechanisms of resistance are present in all individuals of
biotype NAS 4.

t23
Chapter 7
Non-Target Site Mechanism Conferring Resistance to ACCase-
Inhibiting Herbicide in Biotype NAS 14
7.1 Introduction
As described in Chapter 3, resistant wild oat biotlpe NAS 14 has a low level of resistance to
ACCase-inhibiting herbicides and yet contains a sensitive target ACCase enzyme
(Chapter 3). Thus, resistance to herbicides in this biotype must involve mechanisms other
than changes at the,target site. For example, changes to the leaf cuticle may reduce herbicide
uptake by leaves, or herbicide may be prevented from reaching the target site by reduced
translocation, enhanced herbicide detoxification or increased sequestration.
Reduced absorption or translocation of ACCase-inhibiting herbicides have not been reported
to be mechanisms of herbicide resistance. Foliar absorption of diclofop-metþl occurred
readily in susceptible and resistant plants with little difference between them (Donald and
Shimabukuro 1980, Dahroug and Muller 1990, Holtum et al. 1991, Shimabukuro and
Hoffer 1991, Devine et rl. 1992). Once in the plant, translocation of radioactivity from
[1aC]diclofop-methyl out of the treated area is limited (Brezeanu et al. 1976, Boldt and
Putnam 1980, Hall et al. 1982, Jacobson and Shimabukuro 1982, Baker and Chamel 1990).
Although reduced uptake and ranslocation have not been reported to be major factors in the
selective action of diclofop-methyl and other APP herbicides in the earlier studies, they could
contribute to low-level resistance in biotype NAS 14.
To determine the mechanism conferring low level resistance to ACCase-inhibiting herbicides
in biotype NAS 14, studies of uptake, translocation and metabolism of diclofop-methyl were
conducted. The effect of diclofop acid on the plasma membrane potential was also
investigated.

124
7.2 Materials and Methods
7.2.! Uptake, Translocation and Metabotism of ll4C]pictofop-Methyl
The protocol for uptake, translocation and metabolism of herbicide in both susceptible
SAF 19 and resistant NAS 14 wild oat biotypes are described in Chapter 2.
7.2.2 Absorption of lr4c]Diclofop-Methyl to Leaf Cuticle
To determine the amount of herbicide absorbed into the leaf cuticle, a 1 pL drop of 5 mM
[1aC]diclofop-methyl was smeared on the adaxial surface of the second leaf of two-leaf
seedlings. External radioactivity was removed by washing treated leaves in 5 mL of
methanol: H2O: Triton X-100 (20:79.99:0.01 by volume) and blotted dry. The radiolabel
in the cuticle of treated leaf was subsequently removed by washing in 4 mL of chloroform
for 2 min (Tucker et al. 1994). Radioactivity in both the methanol and chloroform washes
was determined by LSS. Plant tissue was oxidized in the Biological Sample Oxidizer.
7.3 Results
7.3.1 Uptake and Metabolism of ll4c]Diclofop-Methyl
The absorption of llaC]diclofop-methyl of biotype NAS 14 was similar to that of the
susceptible SAF 19 (Figure 7.1). At 6 h following application,4T and45Vo of the applied
radioactivity had been absorbed by susceptible and resistant biotypes, respectively and
herbicidal absorption was at a ma¡cimum (80Vo) by 36 h after application in bothbiotypes.
ll4c]diclofop-methyl was metabolized at an equal rate in both susceptible and resistant
biotypes (Figure 7.2 A). Radioactivity rapidly accumulated in diclofop acid with about 55Vo
of radiolabel as diclofop acid by t h after treatment (Figure 7.28). The amount of
[laC]diclofop acid, the phytotoxic form, reached a maximum at 6 h after application and
contributed 81 and 86Vo of the total radioactivity in susceptible SAF 19 and resistant NAS 14
bioqpes, respectively. Diclofop acid was converted to more polar metabolites from 6 h after
application at an equal rate in both biotypes (Figure 7.2C).

t25
100
EO
60
40
20
Eq).--ÈÈGI
-clIo€Ê-io
èe
J.I.I-Icgo.-EcÉ
ú0
24 4E
Time after application (h)
72
Figure 7.1 Uptake of l14c]d'clofop-methyl in the susceptible SAF 19 (O) and resistant
NAS 14 (O) wild oat biotypes when applied to the leaf axil of the two-leaf stage seedlings
cultued in soil.
0

126
100
80
60attù¡H40Ëõ20-aI
ãoË 1ooft
:80èr
;60a-
840G,
€206lLr0; roooËBoÊ
.Ë 60
ô40
20
00 u48
Time after application (h)
72
Figure 7.2A-C Percentage of radioactivity extracted as diclofop-methyl (A), diclofop
acid (B) and turther metabolites (C) in the susceptible SAF 19 (O) and resistant NAS 14 (O)
wild oat biotypes from 0 to 72h after treatment. Each point is the mean of three replicate
experiments. Vertical bars represent the standa¡d erors and a¡e sometimes obscured by
symbols.
A
B
c

r27
7.3.2 Translocation of ll4C]nictofop-Methyl
Herbicide was absorbed into the leaves of the susceptible and resistant biotypes at the same
rate @igrue 7.1) and metabolism of diclofop in both biotypes was similar (Figure 7.zA-C).
One other possible mechanism of resistance is that the herbicide may be prevented from
reaching the target site. Translocation of tlaCldiclofop-methyl was examined when
herbicide was applied on the leaf axil. By 6 h after application, 9.5 and 12.8 7o of the
radioactivity absorbed was found in the meristematic zone of resistant and susceptible
biotypes, respectively and this decreasedto 5Vo by 96 h after treatnent (Figure 7.3A). More
than 807o of the radioactivity remained in the stem of both susceptible and resistant biotypes
over the time course of the experiment (Figure 7.38). More radioactivity was translocated to
the leaves of resistant NAS 14 than that of susceptible by 96 h after the ¡lag¡*clofop-methyl
had been applied to the leaf axil of two-leaf seedlings (Figure 7.3C). This may be a result of
the growth of the second leaf and third leaf of NAS 14 biotype being less affected by
herbicide application than the growth of the second leaf of the susceptible SAF 19 biotype
where the thftd leaf did not appear. The distribution of radioactivity in the stem and the root
were similar for both biotypes (Figure 7.3 B, D).
The small but nevertheless significant difference in the amount of radioactivity in the
meristem of the two biotypes (p<0.05) (Figure 7.3A) may be sufficient to explain the low
level of resistance in biotype NAS 14, however, more evidence is required to support this
conclusion.
7.3.3 Absorption of Herbicide to Leaf Cuticle
Although there are no differences in herbicide absorption into the leaves of the two biotypes,
more herbicide may be bound by the leaf cuticle of the resistant plants and therefore not
available for translocation to the meristem. To test this hypothesis, an experiment was
performed where herbicide was smeared onto the leaf surface and after harvest the leaf was
successively washed with water/methanol, to remove unabsorbed herbicide, and
chloroform, to remove the cuticular waxes. No significant differences in the amount of

t28
0061218z 4896
Time after application (h)
Figure 7.34.D Distribution of radioactivity from ¡l4çldiclofop-methyl in the shoot
meristematic region (A), stem (B), leaf (C) and root (D) in susceptible SAF 19 (O) and
resistant NAS 4 (O) when applied to the leaf axil of nvo-leaf stage seedlings.
20
10
100
50
0€),¡É
ÉË
-È¡l--GI!¡¡
o9CHo
Èa
È.-.!lttIÉtlo.-
rÉcË
ú
0
10
0
10
A
h
or9B

t29
[laC]diclofopmethyl retained on the leaf surface or in the cuticle of susceptible and resistant
biotypes was observed (Figure 7.4). Radioactivity from [l4C]dictofop-methyl was
distributed similarly in the external fractions of treated leaves. By 48 h after application,
4OVo of radioactivity was recovered in the \ilater/methanol wash (Figure 7.4A) and 57o of
radioactivity was extracted from the cuticle by chloroform @igure 7.48). The remainder of
the radioactivity was found within the leaf. These experiments indicate that passage of
diclofop-methyl through the cuticle does not appear to be impeded in the resistant biotype.
7.3.4 ACCase Activity fz Vivo
Previous experiments (Chapter 4, Figure 4.7) have shown that it is possible to treat Avena
seedlings with diclofop-methyl and then extract ACCase to determine the in vivo effect of
herbicide on ACCase activity. This technique was employed with biotype NAS 14 in which
a 1 ttJ- droplet of 5 mM diclofop-methyl was applied to the leaf a¡ril of 2-leaf seedlings. By
6,71 and 72 h following application, ACCase activity in the crude extract of meristematic
region from both biotypes was assayed. ACCase activity in both biotypes was inhibited
following application of the herbicide, however, the resistant biotype NAS 14 had more
extractable ACCase activity 6 h after herbicide application than did the susceptible biotype
SAF 19 (Figure 7.5). Some diclofop-methyl reached the meristems in both biotypes as
ACCase extracted from treated plants were inhibited by 45 and 35Vo in the susceptible and
resistant biotypes, respectively. As both biotypes contain sensitive ACCase as shown in
Table 3.10, ACCase activity from both biotypes remained inhibited 24 a¡d 72 h following
herbicide application. However, the greater inhibition of ACCase in vivo in the susceptible
biotype would suggest that more herbicide reaches the meristem in this biotype compared to
the susceptible. The result supports the conclusions from the translocation experiments in
that slightly less herbicide appears to be translocated to the meristematic regions and reach
the active site in the resistant biotype.

130
100
EO
60
40
20ELq)
oIq)L
CHo
èa
-.!lal
-c)cË
.-EGú
0
10
0100
80
60
40
20
06 12u
Time after application (h)
4E
Figure 7.4A-C The amount of [lag¡¿iclofop in methanol wash (A), chloroform wash
(B) and plant tissue (C) of susceptible SAF 19 (tr) and resistant NAS 14 (f) wild oat
biotypes following treatment with 5 mM ¡t+6¡¿iclofop-methyl. Data a¡e the mean of five
replicate experiments. Vertical bars represent the standard errors of the means.
A
B
TIc
I

131
u48Time after application (h)
72
Figure 7.5 Extractable ACCase activity from the meristematic region of susceptible (O)
SAF 19 and resistant (O) NAS 14 wild oat biotypes after treatment with I ttl. of 5 mM
diclofop-methyl to the leaf axil of twoJeaf stage seedlings. Data presented as percentage of
ACCase from untreated plants. Vertical ba¡s are the standard deviations of four replicates.
100
EO
60
40
20
-o¡rIHcI
tto
Èa
ìa-
.-rCJGI(¡)aÀ6tOU
00

132
7.3.5 Effect of Diclofop on Plasma Membrane Potential
The plasma membrane potential of coleoptile cells from resistant biotype NAS 14 was
depolarized by diclofop acid in the same manner as that of the susceptible biotype (Figure
7.64-B). In the presence of 50 pM diclofop acid, the electrochemical plasma membrane
potential in susceptible SAF 19 and resistant NAS 14 biotypes raprdly depolarized from -110
to -120 mV after a lag period of I to 5 min (Figure 7.6). Following herbicide removal, the
plasma membrane potential from both biotypes remained depolarized with a potential
between -50 and -60 mV. No recovery in polarity was observed in either biotypes for up to
60 min after herbicide removal (data not shown).
7.4 Discussion
The low level of diclofop resistance evident in biotype NAS 14 (Table 3.10) is not due to
possession of an insensitive ACCase, an alteration in the response of the plasma membrane
to diclofop acid, reduced herbicide absorption or an enhanced rate of herbicide metabolism
(Figures 7.1, 7.3 and 7.5). The only difference observed between these biotypes is a
reduction in herbicide content of the shoot meristematic zone following application of
diclofop-methyl to the leaf axil of two-leaf plants @gure 7 .2A). This difference in herbicide
distribution is reflected in measurements of ACCase activity in the shoot meristematic region
at 6 h following herbicide application, which showed ACCase was inhibitedby 45Vo in the
susceptible biotype and by 35Vo in the resistant biotype (Figure 7.5). This indicates that
herbicide might not reach the ACCase in the meristem as readily in the resistant biotype
compared to the susceptible.
The presence of diclofop-methyl in the shoot meristem is crucial to its herbicidal action. For
example, application of diclofop-methyl on or below the meristematic region caused death of
wild oat and barley whereas application to the leaves did not (Friesen et al. 1976).
Translocation of diclofop-methyl in plants is limited and usually less than SVo of ndioactivity
absorbed is translocated out of the treated area (Boldt and Putman 1980, Donald and
Shimabukuro 1980, Hall et al. 1982, Jacobson and Shimabukuro 1982, Dahroug and Muller

133
-100
-80
-60
-120
-140
-120
.100
-80
-Ha-6ta-
--to)9oÈ(Ð-lI(Ëli¡-Tte)¡ià
0
-60
00 10 20
Time (min)30
Figure 7.6^-B The effect of 50 ¡rM diclofop acid on plasma membrane potential of
coleoptile cells of the susceptible SAF 19 (A) and resistant NAS 14 (B) wild oat biotypes.
Arows, addition (+) and removal G) of diclofop acid (DA). The data shown is from a
single coleoptile and is representative of ten coleoptiles each of the susceptible and resistant
biotypes.
+DA
ü A
.DA
,
+DA
ü B
DA
)I I

t34
1990, Baker and Shamel 1990). However, translocation of diclofop-methyl to the shoot
meristem can be increased by placement of the herbicide. A greater amount of diclofop-
methyl and its metabolites were found in the meristematic zone when the herbicide was
placed closer to the leaf base of wild oat seedlings (Walter et al. 1980). In general, little of
the applied herbicide reaches the shoot meristem and therefore, small decreases in
translocation may have a large impact on plant suntival.
In conclusion, the small difference in translocation of diclofop between the susceptible and
resistant biotypes may be suffrcient to confer the low level of resistance to herbicides in the
NAS 14 biotype, however, at present this cannot be substantiated.

135
Chapter I
Investigations of Alterations of Membrane Resporæes to Diclofop
as Mechanism of Resistance in \ryild Oat
8.1 Introduction
As presented in the previous chapters, resistance to APP and CHD herbicides in the range of
biotypes of wild oat studied can involve a modified forrr of ACCase enzyme, enhanced rate
of herbicide metabolism or retarded translocation of herbicide. However, membrane
depolarization by diclofop has been proposed to be an alternative mechanism of resistance to
ACCase-inhibiting herbicides (Shimabukuro and Hoffer 1992). Initially, diclofop was
presumed to act as a protonophore in the plasma membrane (rWright and Shimabukuro
1987). Later, this hypothesis was revised by Shimabukuro and Hoffer (1992) who
suggested that the collapse of the proton motive force in response to diclofop acid is due to
the specific interaction of diclofop with a plasma membrane protein. APP and CHD
herbicides have the ability to depolarize plasma membrane potentials in a number of
susceptible and resistant species (Lucas et al. 1984, rWright and Shimabukuro t987,
DiTomaso et al. 1991, Häusler et al. 1991, Holtum et al. 1991, Shimabukuro and Hoffer
t992, Devine et al. L993, DiTomaso 1993, Dotray et al. 1993). However, in some
diclofop-resistant weed biotypes, repolarization of the plasma membrane potentials occurred
following removing herbicide from the treatment solution (Häusler et al. 1991, Holtum et al.
199I, Shimabukuro and Hoffer l992,Devine et al. 1993). Despite these studies, it is still
unclear how membrane repolarization following herbicide removal in the resistant biotypes is
related to resistance under field conditions since plasma membrane potentials from resistant
crops (e.g. wheat and pea) are also depolarized by diclofop acid (Wright and Shimabukuro
1987, DiTomaso et al. 1991, DiTomaso 1993). In addition, a full recovery of membrane
polarity has been observed in both susceptible corn and resistant pea root cells following
herbicide removal (DiTomaso et al. 1991).

136
A number of membrane-related phenomena have been investigated in resistantAvenafatua,
Loliutn rigidum and crop species. It has been found that diclofqp-induced depolarization
and subsequent repolarization of the membrane potential in susceptible and resistant I.
rigídumis pH dependent (DiTomaso 1993, Holtum et al. 1994). Differences in the ability of
susceptible and resistant L. rigidum and A. fatua roots to acidify the external media were
also noted (Häusler et al. 1991, Devine et al. 1993). The roots of the resistant L. rigidum
biotypes exhibited a decreased rate of acidification of the external solution relative to the
susceptible biotypes (Häusler et al. l99l).
In this chapter, the effects of diclofop acid on depolarization and repolarization of plasma
membrane potentials in coleoptiles of five resistant biotypes of wild oat were investigated.
In addition, other membrane-related experiments such as the effect of pH on recovery of the
plasma membrane potential following removal of diclofop acid and the ability of intact roots
of these biotypes to acidify the external solution were also conducted to ascertain whether an
altered membrane response is a mechanism of resistance in these resistant wild oat biotypes.
8.2 Materials and Methods
8.2.1 Effect of pH on Membrane Response to Diclofop Acid
Etiolated coleoptiles of susceptible and resistant wild oat biotypes were used. The
experiments \ryere conducted in the same manner as described in section 2.7. The basal
solution was replaced with 50 pM diclofop acid for 15 min. The herbicidal solution was
then replaced with the basal solution buffered by 2.5 mM MES-Tris (pH 5.6, 5.8, 6.0,6.2,
6.4,6.5). Five to six etiolated coleoptiles were used for each treatment.
E.2.2 Effect of Diclofop Acid on Acidification of External Solution
One day after root emergence, germinated seeds of the resistant (SAS 1, NAS 4, NAF 6,
NAF 14, SAF 14, NAF 8, NAS 6, SAF 34, V/AF 12) and susceptible (SAS 2, SAF 19)
biotypes of wild oat were placed on filter paper Ekwip@ R6 (Robert Bryce, NSW, Australia)
soaked with distilled water. Filter papers were rolled up, wrapped with plastic sheet and

r37
placed vertically in a 500-mI beaker. Plants were kept in the growth room under the same
conditions as described in section 2.2.2. For each biotype, roots of l0 intact wild oat
seedlings (300450 mg F'W) at the two-leaf stage were washed with distilled water before
being placed in a 10-rnl. beaker containing 4mL of basal medium (1 rnlvf KzSO¿ and I rnlvf
CaSO¿). Root-induced pH changes in solution urere measured by an Action mini pH
combination AglAgCl elecrode connected to a Beckman pH meter. Technical grade diclofop
acid was dissolved in acetone and diluted with the treatment solution to obtain a 50 pM
herbicide solution with<l%o acetone. After a I h treatnent, the basal solution was replaced
with the herbicide solution. After I h exposure to herbicide, the herbicide solution was
replaced with the basal solution. Changes in pH were recorded at room temperature.
Experiments were repeated three times and the data were pooled.
8.3 Results
8.3.1 Effect of Diclofop Acid on Plasma Membrane Potential
The response to diclofop acid of plasma membrane potentials from etiolated coleoptiles cells
of resistant and susceptible biotypes are shown in Figure 8.14-F. Initial membrane
potentials in coleoptile cells from susceptible and resistant wild oat were between -100 and
-120 mV. In the presence of 50 ttJvf diclofop acid, rapid depolarization of the membrane
potential occurred reaching a minimum potential of between -50 and -70 mV within 15 min.
Upon removal of herbicide, membrane potentials from all biotypes remained depolarized
(Figure 8.14-F). No recovery of plasma membrane potential was evident in any of these
resistant wild oat biotypes.
A correlation between recovery of the plasma membrane potential following diclofop acid-
induced depolarization and resistance to APP herbicides was reported in some resistant
biotypes of L. rigidurn (Häusler et al. 1991, Holtum et al. 1991, Shimabukuro and Hoffer
1992, DiTomaso 1993) andA. fatua (Devine et al. 1993). These resistant biotypes had the
ability to recover from diclofop acid induced depolarization whereas the susceptible biotypes
did not. To verify that plasma membrane potential could recover in a resistant L. rigidum

138
biotype and to check that (in the same system) resistant wild oat biotypes could not, the
experiments u,ere also conducted with L. rigidum. In this study, coleoptiles of resistant l.
rigidum biotype SLR 3l showed a recovery of membrane potentials whereas membrane
potentials remained depolarized in the susceptible biotype VIR 1 following removal of
diclofop acid from the bathing solution (Figure 8.2). This result confirms the observations
of Holtum et al. (1991) and provides additional evidence that this recovery phenomenon
does not (rccur in the resistant wild oat biotypes studied here.
8.3.2 Effects of pH on Plasma Membrane Repolarization
The experiments conducted in section 8.3.1 were performed in Higinbotham's solution
(Higinbotha¡n et al. 1970) at pH 5.8. The recovery of membrane polarity following removal
of ACCase-inhibiting herbicide has been reported in resistant biotypes of A. fatua (Devine et
al. 1993) and l,. rigidum (Häusler et al. 1991, Holn¡m et al. 1991, Shimabukuro and Hoffer
1992) at pH 5.7. However, a susceptible L. rigidum biotype also showed membrane
repolarization upon removal of herbicide when the basal solution was buffered at pH 6.0
(DiTomaso 1993). In this case, pH seems to be an important factor in membrane
repolarization. To determine whether pH has an effect on membrane response of wild oat
biotypes, the repolarization solution was buffered with 2.5 mM MES, at a range of pH from
5.6 to 6.5. Coleoptile cells of two susceptible and two resistant biotypes were examined.
At pH 5.6, none of the biotypes demonstrated any recovery of plasma membrane potential
following removal of the herbicide (Figure 8.3). As the pH increased partial repolarization
of the membrane potential was observed in all biotypes and at pH 6.5 full repolarization of
the membrane potential was observed in all biotypes. There were some slight differences
between biotypes with biotype NAF 8 showing repolarization of the membrane potential at a
lower pH than the other biotypes. This biotype had reached 50Vo repolarization at pH 6.0
compared with pH 6.2 f.or biotype SAF 19 and SAS 1 and at pH 6.3 for biotype SAS 2
(Figure 8.3).

139
-120
-80
-120
-100
-80
.60
-12.0
-120
.100
.80
.60
-100
0
0
-40
0---I¡-6t
--H(¡)+¡oÈ()-t(Ë¡r¡-It()tià
-120
-100
-80
.60
-120
-80
-60
-100
-80
-60
00
001020 300
Time (min)10 20 30
Figure 8.14.F The effect of 50 pM diclofop acid on plasma membrane potentials in
coleoptile cells of resistant biotypes NAF 8 (A), SAF 14 (B), SAF 34 (C), WAF 12 (D),
NAS 6 (E) and susceptible biotype SAF 19 (F). Anows, addition (+) and removal G) of
diclofop acid (DA). The data shown is from a single coleoptile and is representative of ten
coleoptiles each of the susceptible and resistant biotypes.
trrttll
+DA
B
DA
,Itrrlllll
+A
DA
7
rrttllll
+DA.DA
c
artrttlll
+DAü D
DA
+DA
.DA
Eü
,rrrttlt
+DA
.DA
Fü
aI

140
-120
-100 +DA
A
susceptible L rigidum
.DA
ü B
resistant L rigìdum+DA
10 20
Time (min)30
Figure 8.24-B The effect of 50 pM diclofop acid on plasma membrane potentials in
coleoptile cells of the susceptible VLR I (A) and resistant SLR 31 (B) ¿. rigidum. Arrows,
addition (+) and removal (-) of diclofop acid (DA). The data shown is from a single
coleoptile and is representative of five coleoptiles each of the susceptible and resistant
biotypes.
-It-cg.--Iq)IoÈÉ)-LcÉ¡rÊÅIIc)l!(a
-E0
.60
40
0
DA
100
-80
-60
.40
00

100
80
60
40
20
0
-Ho.-€N.-LG-oÈé)E
CHo
èa
Lé)
oCJê)ú
r4l
0 5.6 5.8 6.0 6.4 6.6
Figure E.3 Repolarization of plasma membrane potentials in cells of etiolated coleoptiles
of susceptible SAF 19 (tr) and SAS 2 (O) and resistant NAF 8 (I), SAS I (O) wild oat
biotypes as a function of pH of the bathing solution during the recovery phase.
Repolarization is presented as a percentage of the fuIl depolarization. Data are the mean tstandard error and calculated on steady-state potentials measured I h afrer the removal of 50
¡tM diclofop acid.
6.2
pH

142
From the results in Figure 8.3, it is likely that pH affects the ability of plasma membraries to
repolarize following removal of diclofop acid. To confirm this observation, single
coleoptiles of three resistant and nvo susceptible biotypes were bathed in 50 pM diclofop
acid for 15 min prior to placement in a herbicide-free solution buffered with 2.5 mM MES at
pH 6.5. After 20 min, the sa¡ne coleoptile was treated a second time with diclofop acid for
15 min and the herbicide was replaced with a buffered solution at pH 5.8. The results
indicated that an initial diclofop acid-induced depolarization of the membrane potential was
reversible in the herbicide-free solution buffered at pH 6.5 @igures 8.4 to 8.5). The second
exposure to diclofop acid caused a similar depolarization to the first, but the membrane
potential did not recover in a herbicide-free solution buffered at pH 5.8. Biotype NAF 8
showed a full recovery of membrane depolarization within 20 min when the herbicide
solution was replaced with solution buffered at pH 6.5 whereas in the other biotypes
recovery of membrane polarity was only 60to80Vo of the initial value.
8.3.3 Acidification of the External Solution by Roots of Intact Plants
The ability of roots of wild oat biotypes to acidify the external medium was examined by
placing ten wild oat plants at the two-leaf stage in an unbuffered solution. The pH of the
external solution was decreased by about I unit within I h (Figure 8.6). There were no
differences in the rate of acidification in the susceptible SAS 2 biotype compared to the
resistant SAS I biotype. The pH of the solution slowly decreased when the roots of
susceptible and resistant plants were exposed to 50 ¡tM diclofop acid, decreasing by about
0.2 pH units over I h. Following herbicide treatment, the roots were washed and
transferred to a herbicide-free solution. The roots of both biotypes continued to acidify the
extemal solution, however the rate of acidification was much reduced compared to the initial
acidification @gure 8.6).
Similar results were obtained from a range of other susceptible and resistant biotypes of wild
oat (Table 8.1). The roots of all biotypes tested readily acidified the pH of the external
solution which decreased by 2 to 2.5 pH units g-1 root FW over a I h period. Addition of

t43
-120
120
20 30 40 50 60 70 80
Time (min)
Figure 8.44-B Electrochemical potentials measured in coleoptile cells of the susceptible
wild oat biotypes SAF 19 (A) and SAS 2 (B). Arows, addition (+) and removal (-) of
diclofop acid (DA) from the bathing solution. Following removal of diclofop the solution
was buffered at either pH 6.5 or pH 5.8. The data shown is a single coleoptile and is
representative of five coleoptiles each of the two susceptible wild oat biotypes.
.100
.80
-60
-tT-GIa-
--té)IoÈq)-TcÉ¡r¡-:Ic)Hà
0
100
-80
-60
0100
+DA+
A
6.5
DA 5.8
DA
,rrrtttttllttttll
+DA
B+DA
pH 6.5pH 5.8
,+ DA
ltllttlttltlttlt

+DA
pH 5.8
+
A
6.5
+DA
pH 6.5
pH 5.8
.DA
B
+DA DA
c
5.86.5
t4
-t2x)
.100
-80
0
120
-120
20 30 40 50 60 70 80
Time (min)
Figure 8.54-C Electrochemical potentials measured in coleoptile cells of three resistant
wild oat biotypes NAS 4 (A), SAS I (B) and NAF 8 (C). Arrows, addition (+) and removal
C) of diclofop acid (DA) from the bathing solution. Following removal of diclofop acid the
solution was buffered at either pH 6.5 or pH 5.8. The data shown is a single coleoptile and
is representative of five coleoptiles each of the th¡ee resistant wild oat biotypes.
-60
100
-80
-60
0
-LI
-cla-€-I€)t¡oÈ(¡)tcÉL¡IIq)
-à
-100
-80
-60
0100

t45
diclofop acid to the solution resulted in a reduced rate of acidification for all biotypes in a
range of 0.3 to 0.6 pH units g-1 root F\M h-1. When roots were transferred to herbicide-
free solution, acidification of the external solution was about the sarne, 0.4 to 0.7 pH
units g-1 root F-\M ¡-1, as in the presence of diclofop acid. These results indicated that the
roots of susceptible and resistant biotypes had an equal ability to acidify the pH of external
solution and diclofop acid prevented the acidification of the external medium in all biotypes.
The roots of the susceptible L. rigidumbiotype rü,R 1 acidified the external medium atl.27
pH units g-1 root F'\M h-l Gable 8.1). In contrast, roots of the resistant biotype SLR 3l
acidified the external solution at a much slower rate of 0.16 pH units g:1 root FlV h-l. These
results a¡e similar to those reported previously (Häusler et al. 1991, DiTomaso 1993).
Addition of 50 pM diclofop acid reduced acidification in both biotypes and caused an
alkalization of the medium in biotype SLR 31. Upon removal of the herbicide, acidification
of the susceptible biotype VLR 1 remained inhibited whereas acidification resumed in the
resistant biotype SLR 31 to the same rate as that prior to teatment.
8.4 Discussion
Membrane repolarization following diclofop acid-induced depolarization was observed in the
resistant L. rigidum biotype SLR 3l but not in the susceptible biotype VLR I (Figure 8.2A-
B). Similarly, the ability of the diclofop-resistant biotypes to overcome the effects of
diclofop acid on plasma membrane depolarization were reported for resistant biotypes of
A. fatua (Devine et al. 1993) and L. rigidum (Häusler et al. 1991, Holtum et al. 1991,
Shimabukuro and Hoffer 1992, DiTomaso 1993). It has been suggested that this recovery
of membrane polarity following the removal of herbicide is associated with resistance to
ACCase-inhibiting herbicides (Häusler et al. 1991, Devine et al. 1993). This phenomenon
did not occur in the resistant wild oat biotypes examined here where the membrane remained
depolarized following removal of diclofop acid in both susceptible and resistant biotypes
(Chapters 4 to 7 and Figure 8.14-E). Clearly, repolarization of the plasma membrane

t46
7.0
l̂- oa
120
Time (min)
Asusceptible wild oat
Bresistant wild oat
180
6.0
HÊÈ 0
5.0
7.0
6.0
5.0
+DA
+DA
Al- pe
00 60 u0
Figure 8.6 Acidification of an unbuffered solution (1 mM CaSO4, I mM K2SOa) by the
roots of ten seedlings of susceptible SAS 2 (A) and resistant SAS 1 (B) wild oat biotypes.
Anows, addition (+) and removal (-) of 50 pM diclofop acid (DA). The data shown is a
representative of three measurements of the susceptible and resistant biotypes.

t47
Table E.l Acidification (A pH g-1 root fresh weight h-l) of an unbuffered solution (lmM
CaSO¿, knlvf KzSO¿)bytherootsof susceptibleandresistantbiotypesof wildoat. ThepH
was measured prior to addition of 50 ¡rM diclofop acid, after addition of the herbicide, and
following removal of the herbicide. Values are the mean of t standard deviation of th¡ee
mea$¡tements
Biotype  pH g-t root FW h-l
Control +Diclofopacid -Diclofopacid
Susceotible biowoes
SAS 2
SAF 19
Resistant biotvoes
SAS 1
NAF 6
NAS 4
NAS 14
NAF 8
NAS 6
SAF 14
SAF 34
rwAF 12
Inlium ripi.dum
VLR I (susceptible)
SLR 31 (resistant)
2.47 + 0.22 0.30 + 0.08 0.45 r 0.04
2.4t + 0.24 4.28 + 0.12 0.54 r 0.20
2.25 + O.t8 0.46 r 0.09 0.55 + 0.05
2.16 r 0.09 0.25 + 0.11 0.72 + o.tt
2.44 t 0.lo 0.67 r 0.08 0.45 r 0.14
2.47 + O.to o.42 + O.20 0.62 + 0.15
2.05 + O.t2 0.37 ! 0.14 0.54 t 0.14
2.35 + 0.t6 0.43 + 0.13 0.69 r 0.18
2.r3 + O.20 0.33 + 0.10 0.71 t 0.17
2.22 + 0.t8 0.50 r 0.12 0.58 + 0.17
2.14 + O.05 0.42 + 0.05 0.39 + 0.18
t.27 + O.22 0.08 + 0.01 0.2t + 0.14
0.16+0.19 +O.49t0.14 0.21 +. o.tg

148
potential following removal of the herbicide in these wild oat biotypes is not correlated with
resistrnce to ACCase-inhibiting herbicides.
The kinetics of depolarization and repolarization of plasma membrane potential depend on
the herbicide, the herbicide concentration, the biotype and the pH of the bathing solution
(Holtum et at. 1994). It is clear from this study and previous reports that pH is an important
determinant for the recovery of the plasma membrane potential following removal of
diclofop acid (DiTomaso 1993, Holtum et al. 1994). It was proposed that the differences in
membrane repolarization following removal of herbicide between susceptible, VLR 1, and
resistant, SLR 31, biotypes of L. rigidum were due to differences in acidification of the cell
wall by these biotypes (DiTomaso 1993). Therefore, strict control of pH by buffer should
eliminate the differences between these biotypes. Recovery of membrane polarity is
dependent on pH rather than buffering capacity as the resistant biotype SLR 3l will recover
from diclofop acid-induced depolarization in a solution buffered at pH 5.8 whereas the
susceptible biotype VLR 1 will not (data not shown). In this study the effects of pH on
recovery of the membrane potential following removal of diclofop acid were examined
(Figure 8.3) and it is also apparent that different biotypes display different pH profiles for
the recovery. Therefore, the differences observed in recovery of the plasma membrane
potential between resistant and susceptible biotypes of L. rigidum (Häusler et al. 1991,
Figure 8.2) and A. fatua (Devine et al. 1993) might be attributable to different pH profiles
for recovery between these biotypes. It has long been known that depolarization of the
membrane potential by diclofop acid is pH dependent (Lucas et al. 1984, Wright and
Shimabukuro 1987, DiTomaso 1993), however, this study demonsüates that recovery from
depolarization is also pH dependent (Figure 8.3). Plasma membrane potentials of coleoptile
cells from two susceptible and three resistant wild oat biotypes depolarized by diclofop acid
at pH 5.8 recover from depolarization upon herbicide removat if the pH is increased to pH
6.5, but not at pH 5.8 (Figures 8.4 and 8.5). Hence, depolarization of the plasma
membrane potential by ACCase-inhibiting herbicides is probably not lethal to plants and this

t49
phenomenon may be only transitory following field application of these herbicides
(DiTomaso et al. 1991, DiTomaso 1993).
Two hypotheses have been proposed to explain the diclofop acid-induced depolarization of
the plasma membrane potential. Either, that diclofop acid acts as a protonophore (Wright
and Shimabukuro 1987), or diclofop acid interacts with a plasma membrane protein to
induce an influx of protons (Shimabukuro and Hoffer 1992). Diclofop acid and other
ACCase-inhibiting herbicides are weak acids with a pKa of between 3.5 and 4.6 (Dotray et
al. 1993). At the pH prevalent in the cell wall, pH 5.5 (Sterling 1994), a higher proportion
of these herbicides will be in a protonated, lipid soluble form than at the pH of the
cytoplasm, pH 7.5. The herbicides will therefore tend to move from the apoplast to the
cytoplasm, lose a proton and become trapped in the cytoplasm. This will lead to an influx of
protons into the cell and a dissipation of the plasma membrane potential. Increasing the
external pH to 6.5 following removal of the herbicide will tend to mobilize some of the
protonated diclofop acid from the cell membrane to the external solution where it will be
deprotonated. Such a scenario could easily explain the response of repolarization to pH
observed in this study (Figure 8.3). Differences between biotypes in the pH required for
recovery of the plasma membrane potential may reflect differences in the amount of diclofop
acid which partitions into the plasma membrane or in the micro-environment surrounding the
plasma membrane.
If there is a mechanism(s) of resistance which give rise to the repolarization of plasma
membrane potential at pH 5.7 in some resistant biotypes of Z. rigidum (Häusler et al. 1991)
then it does not operate in the resistant Avena biotypes examined here. To date, the
molecular basis of repolarization as a resistance mechanism is unknown, but have been
postulated to involve sequestration of the herbicide (Holtum et aJ. 1994). Correlated with
this mechanism(s) in L. rigidurn biotypes is a reduced ability to acidify the external medium
(Häusler et al. 1991). For example, the roots of susceptible biotypes VLR I and SLR 2
decreased the pH of the external medium by 1.7 to 2.4 pH units, whereas the diclofop-
resistant biotypes SLR 31, WLR 96, VLR 6 and NLR 2 showed an acidification of less than

150
0.2 pH units (Häusler et al. 1991). DiTomaso (1993) reported that roots of the susceptible
biotype VLR I were able to acidify the external medium ^t4.5
times the rate of the resistant
biotype SLR 31. This has been proposed to explain both the membrane repolarization and
resistance in one biotype of. L. rigidum, SLR 31, (DiTomaso 1993). Briefly, this
hypothesis suggests that the reduced acidification leads to a higher pH in the apoplast. At
this higher pH, more of the herbicide is in the unprotonated form and unable to enter the cell.
Under such conditions, biotype SLR 3l should be resistant to all weak acid herbicides,
however, this is clearly not the case as most of the SLR 31 population is susceptible to
sethoxydim (Tardif and Powles 1994) and sulfometuron-methyl (Christopher et al. l99l).
In contrast to the situation observed with ¿. rigidum, there were no differences in
acidification of the external medium between resistant, UM-l, and susceptible, UM-5,
biotypes of. A. fatua which differed in thei¡ ability to recover from diclofop acid-induced
membrane depolarization (Devine et al. 1993). However, in this case the resistant biotype
was able to recommence acidification following treatment with diclofop acid, whereas the
susceptible biotype could not (Devine et al. 1993). Similar results were obtained here with
resistant and susceptible biotypes of Z. rigidum (Table 8.1). Addition of 50 ¡tM diclofop
acid to the medium bathing the roots reduced acidification by both biotypes, however, only
the resistant biotype SLR 3l was able to recommence acidification at pre-treatment rates
following removal of the herbicide. In contrast, none of the Avena biotypes examined here
were able to recommence acidification at the pre-treatment rates following removal of
diclofop acid (Table 8.1). A simple explanation for the diclofop acid-induced reduction in
acidification would be proton movement into the cells as the protonated form of diclofop
acid. Alternatively, diclofop acid may be having effects on membrane components which
reduce net proton efflux. The fact that susceptible biotypes of Z. rigidum (Table 8.1) andA.
føtuø (Devine et al. 1993) are unable to recommence acidification following treaünent with
diclofop acid supports the second rather than the first hypothesis. In common with the
membrane repolarization phenomenon, rapid re-establishment of acidification following
treatment with diclofop acid is not required for resistance to the ACCase-inhibiting
herbicides, even when active site changes are not involved (See Chapter 7).

151
In conclusion, the mechanism(s) which give rise to the repolarization of the plasma
membrane potential and the re-establishment of acidification by intact roots following
diclofop acid treatment in some resistant biotypes of L. rtgidum md A. fatua do not operate
in the resistantz4,v¿n¿ biotypes examined here. Clea¡ly, more work is required to understand
these phenomema, however, they are not always correlated with resistance to ACCase-
inhibiting herbicides.

152
Chapter 9
General Conclusions
This project has been conducted to elucidate the mechanisms endowing resistance to
ACCase-inhibiting herbicides in wild oat (Avena spp.) populations in Australia. Studies
with ten putative resistant wild oat biotypes, which had been exposed to different herbicide
treatments in the field, showed resistance to one or more APP or CHD herbicides. The
patterns of resistance to APP and CHD herbicides observed in wild oat could be classified
into three goups; 1) High to moderate resistance to APP herbicides and low-level resistance
to CHD herbicides, 2) Moderate resistance to both APP and CHD herbicides and 3) Low-
level resistance to both classes of herbicides. \When the response of the target site ACCase
to herbicides was examined, it proved in most biotypes to be resistant to at least one CHD or
APP herbicide. With a few exceptions the patterns of resistance at the ACCase level
mi¡rored resistance at the whole plant level (Chapter 3).
Four biotypes with different patterns of resistance were selected for further examination of
mechanisms of resistance. Biotypes SAS 1 and NAS 4 are highly resistant to APP
herbicides, biotype NAF 6 is moderately resistant to both APP and CHD herbicides, and
biotype NAS 14 is slightly resistant to both groups of herbicides. The results indicated that
resistance to ACCase-inhibiting herbicides can be conferred by both target site and non-
target site mechanisms (Chapters 4 to 7).
Target site-based resistance is by far the most common mechanism of herbicide resistance
documented. Almost all cases of PS tr-inhibitor resistance in weeds a¡e due to a single
change in the Dl protein of photosystem II such that the triazine herbicides no longer inhibit
photosystem tr (Gronwald 1994). Similarly, resistance to Als-inhibiting herbicides is
mostly the result of changes to the target site, ALS, which renders the enzyme resistant to
these herbicides (Saari et al. 1994). Target site resistance to the ACCase-inhibiting
herbicides is also common (Devine and Shimabukuro 1994). This mechanism has also been

153
widely observed in this study (Chapter 3). For both biotypes SAS I and NAF 6, a resistant
ACCase was the only mechanism observed (Chapters 4 and 5). Experiments examining
uptake, translocation and metabolism of ¡tagtd'clofop-methyl showed no differences
between resistant and susceptible biotypes. Likewise, no differences in uptake or
metabolism of ¡l+ç¡r"*oxydim were observed between susceptible SAS 2 and resistant
NAF 6 (Chapter 5). These two resistant biotypes were not identical. Biotype SAS 1 was
highly resistant to APP herbicides, but only slightly resistant to CHD herbicides, whereas
biotype NAF 6 was moderately resistant to both classes of herbicides. Frequently, target
site resistance to APP herbicides results in resistance to the CHD herbicides (Devine and
Shimabukuro 1994), however, this does not always prevail as has been observed in a
diclofop-methyl resistant biotype of L. multiþrum (Gronwald et al. 1992). V/ith the
exception of biotype NAF 6, none of the biotypes of wild oat with target site-based
resistance studied in this thesis have sEong resistance to the CHD herbicides (Chapter 3).
Clearly, there are a number of potential mutations within ACCase which can endow
resistance to ACCase-inhibiting herbicides. This has been observed in maize cell cultures
selected with sethoxydim or haloxyfop where th¡ee mutant ACCase alleles of ACCase
structural gene (Accl), Accl-S, AccI-HI and AccI-H2, have been identified (Marshall et al.
1992). The Accl-S exhibited high resistance to sethoxydim and haloxyfop. The AccI-Hl
allele exhibited resistance only to haloxyfop and the Accl-H2 showed high resistance to
haloxyfop and intermediate resistance to sethoxydim. A similar situation is likely in the wild
oat biotypes studied here with several possible different resistant alleles of ACCase, based
on responses of ACCase to herbicides (Chapter 3). Simila¡ly, resistance to AlS-inhibiting
herbicides can be conferred by many possible mutations within ALS in plants (reviewed in
Saa¡i et al. 1994). Mutations in the ALS gene conferring resistance have been related to 24
different mutations at 10 sites. In contrast, a single change ser 264 to gly in the D1 protein
of PS tr has been observed to provide target-site resistance to PS ll-inhibiting herbicides in
higher plants (Gronwald 1994). The large number of possible mutations within ALS, and
probably ACCase, may account in part for the relative rapid rate of development of
resistance to the ALS- and ACCase-inhibiting herbicides. Resistance to these herbicides can

154
appear in as little as 3 or 4 applications (Tardif et al. 1993, Devine and Shimabukuro 1994,
Maxwell and Mortimer 1994), whereas resistance to PS tr-inhibiting herbicides only appears
after l0 or more applications of herbicide (Ryan 1970, Burnet et al. 1991). Therefore, the
more rapid development of resistance to the ACCase and AlS-inhibiting herbicides might be
due to an increased nu¡rrber of viable mutations within the target sites.
Biotype NAS 4 has target site resistance to APP herbicides, however, in this case moderate
resistance at the enzyme level appears insufFlcient to explain the high-level resistance at the
whole plant level. This biotype contains a second mechanism of resistance, enhanced
metabolism of diclofop. The seeds used in these experiments were collected from a number
of plants so it is possible that different mechanisms of resistance were present in different
individuals of this population. This hypothesis was tested by examining the response of
ACCase to diclofop acid in the progeny from ten individual resistant plants (Chapter 6). The
result showed that ten lines of biotype NAS 4 exhibited about the same level of resistance at
the ACCase as the whole population (Chapter 3). This means that resistance at the ACCase
level is quite uniform within the NAS 4 population and therefore the two mechanisms of
resistance coexist within each individual. Enhanced metabolism of diclofop has been
observed in a biotype of Lolium rigidum which showed a 1.5 fold increased capacity to
metabolize diclofop (Holtum et al. 1991). In addition, enhanced metabolism has been
reported in biotypes of L. rigidurn resistant to AlS-inhibiting (Christopher et al. 1992) and
PS tr-inhibiting herbicides (Burnet et al. 1993a and 1993b). In these last cases, enhanced
metabolism due to cytochrome P450 monooxygenases has been implicated. Studies with the
cytochrome P450 inhibitor, tetcyclasis, revealed that this compound can both decrease
diclofop acid metabolism in biotype NAS 4 and increase diclofop-methyl phytotoxicity
(Chapter 6). Following pre-treatment with tetcyclasis, the amount of diclofop acid
remaining in biotype NAS 4 24hafter treatment with diclofop-methyl, was increased to the
same level as that found in the susceptible biotype SAF 19. Therefore, it is likely that
enhanced metabolism of diclofop acid in biotype NAS 4 is endowed by a cytochrome P450
monooxygenase.

155
Resistance as a result of a target site mutation and enhanced metabolism has been reported
for a tnazine-resistant biotype of Brachepodium distachyon '(Gressel et al. 1983), a
sulfonylurea-resistant biotype of L. rigidun (Cbristopher et al. 1992), a diclofop-methyl
resistant biotype of L. rigidurø (Preston et al. unpublished) and in small subsets of diclofop-
methyl resistant (Tardif and Powles 1994) and sulfonylurea-resistant (Burnet et al. 1994)
populations of Z. rigidum. In all of these cases, the target site resistance is the most
powerful resistance mechanism. Multiple mechanisms of resistance to the same herbicide
have been reported on a few occasions, most often in biotypes of L. rígidum. lt was noted
that herbicide treatment of a large, polymorphic population results in the survival of
individuals that possess one or more (different) mechanisms conferring the ability to
withstand the dose of herbicide applied (Powles and Matthews 1992). rù/ith obligate
outcrossing species like Z. rigidum, there is gene flow among the survivors resulting in
exchange of different resistance genes and their accumulation in the next generation.
However, dependent upon genetic variation, the size of a population exposed to herbicide
selection, and the ability and efficiency of cross-pollination, there can be enrichment of a
number of different resistance mechanisms (Powles and Matthews 1992). Although wild
oat is self-pollinated species, outcrossing can occur at rates up to l2vo (Imam and Allard
1965, Chancellor 1976, Bickelmann 1993). Hence, the genetic variability between and
within natural populations of wild oat can be maintained. It is, therefore, an indication of
biological diversity that two mechanisms of resistance, both contributing to resistance, have
appeared in a single biotype of wild oat.
Resistance to diclofop in biotype NAS 14 is not due to a change in the ACCase target site
nor is it due to enhanced herbicide detoxification. Biotype NAS 14 did demonstate reduced
translocation of radioactivity to the meristematic region from leaf-applied [t+C]diclofop-
methyl relative to the susceptible biotype. This result was confirmed by measuring the
activity of ACCase in the meristematic region of susceptible and resistant biotypes following
application of 5 mM diclofop-methyl to the leaf axil. As shown in Chapter 3, biotype NAS
14 has a herbicide-sensitive ACCase, however, the extractable ACCase activity in the
meristematic region was greater in this biotype following application of diclofop-methyl than

156
in the susceptible biotype SAF 19. This indicates that herbicide moves more slowly to the
meristematic region in the resistant biotype. It can be difficult to determine the mechanism of
resistance in a biotype that has a low level of resistance, however, the small difference in
translocation observed here may be sufficient to account for the low level of resistance
observed in this biotype. Reduced herbicide translocation is a mechanism of resistance in
some paraquat-resistant biotypes (reviewed in Preston 1994), however, this mechanism has
not been documented as a resistance mechanism in weeds for any other herbicides.
As summarised above, three mechanisms of resistance were found in resistant wild oat
biotypes (Chapters 3 to 7), however, the modified target site, ACCase, was the most
common phenomenon. The results in Chapters 4 to 7 may be used to indicate possible
resistance mechanisms operating in other resistant biotypes. Resistance in biotypes NAF 8,
NAS 2 and SAF 14 that are highly resistant to diclofop-metþl in the field but exhibit a
moderate-level of resistance at the enzyme level might involve more than one mechanism as
found in biotype NAS 4. However, these biotypes are less resistant than biotype NAS 4
and, therefore, the resistant ACCase may be the most important, or sole mechanism
operating. Biotype WAF 12 has low-level-resistance to diclofop-methyl like biotype NAS
14, however, ACCase from this biotype is sensitive to diclofop acid. This biotype might
have a similar non-target site mechanism conferring resistance. The obligate cross-pollinated
L. rigidum is capable of evolving multiple mechanisms to overcome herbicides (Holtum and
Powles 1991). As has been demonstrated here, the self-pollinated Avena species can also
exhibit diversity in resistance mechanisms.
Some resistant weeds have the ability to re-establish the plasma membrane potential
following treatment with diclofop acid (Häusler et al. 1991, Holtum et al. 1991, Devine et
al. 1993). This phenomenon was not observed in the ten wild oat biotypes studied here.
The recovery of membrane polarity is depended upon the pH of bathing solution
(Chapter 8). Both susceptible and resistant wild oat biotypes exhibited membrane
repolarization when the herbicide solution was replaced with the herbicide-free solution
buffered at pH 6.5. Although repolarization of plasma membrane potentials has been

t57
documented as a mechanism of resistance to ACCase-inhibiting herbicides in some resistant
biotypes of. A. fana and Z. rigidum, this phenomenon was not appeared in the resistant
biotypes of A. fatua and.á. sterilis studied here. However, it is still unclea¡ how the ability
to restore the membrane potential to its normal state is correlated with the selective action of
diclofop acid between susceptible and resistant plants. What is clear is that this rapid
repolarization of the plasma membrane potential is not a requirement for resistance to
ACCase-inhibiting herbicides.
The number of applications and classes of herbicides cannot be used to predict the patterns
and level of resistance to APP and"/or CHD herbicides in wild oat. For example, biotype
NAS 4 had been exposed to two applications of tralkoxydim but it is sensitive to CHD
herbicides at the whole plant and the enzyme level (Chapter 3). In contrast, biotype NAF 6
had never been exposed to CHD herbicides but is moderately resistant to CHD herbicides
(Chapter 3). Biotype NAS 14 had been treated with four applications of diclofop-methyl
and two of fluazifop-butyl, however, it has developed lowlevel resistance to both APP and
CHD herbicides whereas biotype SAS 1 with five applications of APP herbicides has high-
level resistance to APP herbicides.
Wild oat seeds can survive in the soil for up to nine years (Jones t976, Gwynne and Murray
1985, Miller and Nalewaja 1990). Therefore, once resistant wild oat are in a field, the
resistance genes can remain for a considerable period. There are few alternative herbicides
to diclofop-methyl, fenoxaprop-ethyl and talkoxydim, for the selective control of wild oat in
cereals. With the development of target site-based resistance to the ACCase-inhibiting
herbicides, these alternative herbicides will be relied on to control wild oat. Wild oat can
also develop resistance due to non-target site mechanisms such as enhanced metabolism, for
example biotype NAS 4, and decreased translocation, for example biotype NAS 14. Such
non-target site mechanisms, as has been documented for Z. rigidum and A. myosuroides
(Hall et al. 1994) can lead to cross and multiple resistance across chemical classes.
Enhanced metabolism, in particular, has been demonstrated as a mechanism endowing cross
and multiple resistance in these two species. Such a scenario appearing in wild oat may

158
mea¡r that herbicides will not be an effective means sf ço¡trslling this species in the future.
Already there are at least 15 populations of triallate-resistant wild oat exhibiting cross-
resistance to difenzoquat (O'Dovovan et al. 1994) and more cases of cross and multiple
resistance could be expected.

159
Literature Cited
Adkins, S.}V., Simpson, G.M. and Naylor, J.M. (1984a). The Physiological basis of seed
dormancy rnAvenafatuø. fr.. Action of nitrogeneous compounds. Physiol. Plant.
6O:227-233.
Adkins, S.W., Simpson, G.M. and Naylor, J.M. (1984b). The physiological basis of seed
dormancy rn Avenafatw.lY. Altemative respiration and nitogeneous compounds.
Physiol. Plant. 60: 234-238.
Adkins, S.'W., Naylor, J.M. and Simpson, G.M. (1984c). The physiological basis of seed
dormancy in Avena fatua. V. Action of ethanol and other organic compounds.
Physiol. Plant. 62: 18-24.
Agenbag, G.A. and DeVilliers, O.T. (1989). The effect of nitrogen fertilizers on
germination and seedling emergence of wild oat (l4,. fatua L.) seed in different soil
types. V/eed Res. 29: 239-245.
Aguero-Alvarado, R., Appleby, A.P. and Armstrong, D.J. (1991). Antagonism of
haloxyfop activity in tall fescue (Festuca arundinacea) by dicamba and bentazon.
Weed Sci.39: 1-5.
Alban, C., Baldet, P. and Douce, R. (1994). Localization and cahracterization of two
structurally different forms of acetyl-CoA carboxylase in young pea leaves, of
which one is sensitive to aryloxyphenoxypropionate herbicides. Biochem. J. 300:
557-565.
Al-Feel, W., Chirala, S.S. and Wakil, S.J. (1992). Cloning of the yeast FAS3 gene and
primary structure of yeast acetyl-coenzyme A carboxylase. Proc. Natl. Acad. Sci.
USA 89: 4534-4538.

160
Anderson, R.N. and Gronwald, J.W. (1987). Noncytoplasmic inheritance of atrazine
tolerance in velvetleaf (Abutílon theophrastí). Weed Sci. 35: 496498.
Asa¡e-Boamah, N.K. and Fletcher, R.A. (1983). Physiological and cytological effects of
BAS 9052 OH on corn (7za mays) seedlings. Weed Sci. 31: 49-55.
Ashton, F.M. and Monaco, T.J. (1991). Aryloxyphenoxys, benzoics, bipyridiliums and
pyrazoliums. .In Weed Science Principles and Practices, (F.M. Ashton and T.J.
Monaco, Eds.), A Wiley-Interscience Publication, NY, PP. 172-lg}.
Ashton, A.R., Jenkin, C.D.L. and Whitfeld, P.R. (1994). Molecular cloning of two
different cDNAs for maize acteyl CoA carboylase. Plant Mol. Biol. A:3549.
Baker, E.A. and Chamel, A.R. (1990). Herbicide penetration across isolated and intact leaf
cuticles. Pestic. Sci. 29: 187-196.
Ba¡r, 4.R., Mansooji, A.M., Holtum, J.A.M. and Powles, S.B. (1992). The inheritance
of herbicide resistance inAvena sterílis ssp.ludoviciana,biotype SAS l. Proc. lst
International Weed Contol Congress, Melbourne, Australia, pp. 7 0-7 5.
Ba¡rentine, W.L., Snipes, C.E. and Smeda, R.J. (1992). herbicide resistance confirmed in
johnsongrass biotypes. Miss. Agric. Forest. Exp. Stn. Res. Rep. 17: l-5.
Beckett, T.H., Stroller, E.'W. and Bode, L.E. (1992). Quizalofop and sethoxydim activity
as affected by adjuvants and ammonium fertilizers. rü/eed Sci. 40: 12-19.
Betts, K.J., Ehlke, N.J., Wyse, D.L., Gronwald, J.W. and Somers, D.A. (1992).
Mechanism of inheritance of diclofop resistance in Italian ryegrass (Lolium
multiþrum). V/eed Sci. 40: 184-189.
Bickelmann, U. (1993). On fatuiods and hybrids of Avena spp. and their significance for
seed production. I. Flowering behaviour of oats and origin of fatuoids and hybrids.
Plant Va¡ieties and Seeds 6:21-26.

161
Bishop, T., Powles, S.B. and Cornic, G. (1987). Mechanism of paraquat resistance in
Hordeum glaucuni.Il. Paraquat uptake and translocation.' Aust. J. Plant Physiol.
t4: 539-547.
Black, M. (1959). Dormancy studies in seed of Avena fatua. I. Possible role of
germination inhibitors. Can. J. Bot. 37:393402.
Boldt, L.D. and Barrett, M. (1991). Effects of diclofop and haloxyfop on lipid synthesis in
corn(7*amoys) and bean (Phaseolus vulgaris). rüVeed Sci. 39: 143-148.
Boldt, P.F. and Putnam, A.R. (1980). Selectivity mechanisms for foliar application of
diclofop-methyl. I. Retention, absorption, translocation and volatility. S/eed Sci.
28: 474477.
Boutsalis, P., Holtum, J.A.M. and Powles, S.B. (1990). Diclofop-methyl resistance in a
biotype of wild oat (Avena fatua L.). Proc. 9th Australian V/eed Conf., Adelaide,
Australia, pp. 216-219.
Boutsalis, P. and Powles, S.B. (1995a). Resistance of dicot weeds to acetolactate synthase
(Als)-inhibiting herbicides in Australia. T9Veed Res. in press.
Boutsalis, P. and Powles, S.B. (1995b). Inheritance and mechanism of resistance to
herbicides inhibiting acetolactate synthase in Sonchus oleraceus L. Theor. Appl.
Genet. in press.
Brezeanu, 4.G., Davis, D.G. and Shimabukuro, R.H. (1976). Ultrastructural effects and
translocation of metþl-2-(4-(2,4 -diclorophenoxy) phenoxy) propanoate in wheat
(Triticum aesticum) and wild oat(Avenafana). Can. J. Bot.54: 2038-2M8.
Browse, J., Roughan, P.G. and Slack, C.R. (1981). Light control of fatty acid synthesis
and diurnal fluctuations of fatty acid composition in leaves. Biochem. 1.196:347-
354.

t62
Browse, J. and Somerville, C. (1991). Glycerolipid synthesis: biochemistry and
regulation. Annu. Rev. Plant Physiol. Plant Mol. Biol. 42: 267-506.
Brun, V.S. (1965). The effects of fresh water storage on the germination of certain weed
seeds.'Weeds 31: 38-39.
Buhler, D.D., Swisher, B.A. and Burnside, O.C. (1985). Behavior of l4C haloxyfop-
methyl in intact plants and cell cultures. V/eed Sci. 33: 291-299.
Burnet, M.V/.M., Hildebran, O.8., Holtum, J.A.M. and Powles, S.B. (1991). Amitrole,
triazine, substituted urea and metribuzin resistance in a biotype of rigid ryegrass
(Inlium rigidum). Weed Sci. 39: 317-323.
Burnet, M.V/.M., Loveys, 8.R., Holtum, J.A.M. and Powles, S.B. (1993a). A
mechanism of chlorotoluron resistance in Lolium rigidum. Planta 190: 182-189.
Burnet, M.W.M., Loveys, 8.R., Holtum, J.A.M. and Powles, S.B. (1993b). Increased
detoxification is a mechanism of simazine resistance in l-olium rigidum. Pestic.
Biochem. Physiol. 46: 2O7 -2L8.
Burnet, M.W.M., Christopher, J.T., Holtum, J.A.M. and Powles, S.B. (1994).
Identification of two mechaisms of sulfonylurea resistance within one population of
rigid ryegrass (Lolium rigidum) using a selective germination medium. rù/eed Sci.
42:468-473.
Burton, J.D., Gronwald, J.W., Somers, D.4., Connelly, J.4., Gengenbach, B.G. and
Wyse, D.L. (1987). Inhibition of plant acetyl coenzyme A carboxylase by the
herbicides sethoxydim and haloxyfop. Biochem. Biophys. Res. Commun. 14E:
to39-t34r'..
Burton, J.D., Gronwald, J.W., Somers, D.4., Gengenbach, B.G. and Wyse, D.L.
(1989). Inhibition of corn acetyl-CoA carboxylase by cyclohexane-dione and
aryloxyphenoxypropionate herbicides. Pestic. Biochem. Physiol. 34: 7 6-85.

163
Burton, J.D., Gronwald, J.W., Keith, R.4., Somers, D.4., Gengenbach, B.G. and'lVyse,
D.L. (1991). Kinetics of inhibition of acetyl-coenzyme A carboxylase by
sethoxydim and haloxyfop. Pestic. Biochem. Physiol. 39: 100-109.
Cairns, A.L.P. and DeVilüers, O.T" (1986). Breaking dormancy of. AvenafatusL. seed by
treatment with ammonia. V/eed Res. 26: t9l-197.
Carrpbell, J.R. and Penner, D. (1985). Sethoxydim metabolism in monocotyledonous and
dicotyledonous plants. V/eed Sci. 33: 771-773.
Carr, J.C., Davies¡ L.G., Cobb, A.H. and Pallett, K.E. (1985). The metabolic activity of
fluazifop acid in excised apical meristem sections. Proc. British Crop Prot. Conf.
-Weeds: 155-L62.
Catanzaro, C.J., Burton, J.D. and Kroch, W.A. (1993). Graminicide resistance of acetyl-
CoA carboxylase from ornamental grasses. Pestic. Biochem. Physiol. 45: 147-
153.
Chancellor, R.J. (1976). Seed behaviour. /n V/ild Oats in lVorld Agriculture, (D.P.
Jones, Ed.), The Whitefriars Press, Ltd., London, pp. 65-87.
Chandrasena, N.R. and Sagar, G.R. (1987). Effect of fluazifop-butyl on the chlorophyll
content, fluorescence and chloroplast ultrastructure of Elymus repens (L.) Gould
leaves. Weed Res.27: lO3-112.
Chandrasena, N.R. and Sagar, G.R. (1989). Fluazifop toxicity to quackgrass (Agropyron
repens) as influenced by some application factors and site of application. Weed Sci.
37:79O-796.
Cha¡les, D.J., Hasegawa, P.M. and Cherry, J.H. (1986). Characterization of acetyl-CoA
carboxylase in the seed of two soybean genotypes. Phytochem. 25: 55-59.

tg
Chauvel, B. (1991). Polymorphisme génétique et sélection de la resistance aux urées
substitutées chez Alopecurus myosuroÍd¿s Huds. Ph.D. Thesis, University of
Paris-Orsay.
Cho, H., V/idholm, J.M. and Slife, F.W. (1986). Effects of haloxyfop on corn (Zea
mays) and soybean (Glycine max) cell suspension cultures. Weed Sci.34: 496-
501.
Christopher, J.T. (1993). Mechanisms of resistance to herbicide which inhibit acetolactate
synthase in annual ryegrass (Lolium rigidum). Ph.D. Thesis, University of
Adelaide, Austalia.
Christopher, J.T., Powles, S.B.and Holtum, J.A.M. (1992). Resistance to acetolactate
synthase-inhibiting herbicides in annual ryegfass (Inlium rigidum) involves at least
two mechanisms. Plant Physiol. 100: 1909-L9I3.
Christopher, J.T., Powles, S.8., Liljegren, D.R. and Holtum, J.A.M. (1991). Cross-
resistance to herbicides in annual ryegrass (Lolium rigidum).II. Chlorsulfuron
resistance involves a wheat-like detoxification system. Plant Physiol. 95: 1036-
to43.
Christopher, J.T., Preston, C. and Powles, S.B. (1994). Malathion antagonizes
metabolism-based chlorsulfuron resistance in l-olium rigidum. Pestic. Biochem.
Physiol. 49: 172-182.
Cotterman, J.C. and Sarri, L.L. (1992). Rapid metabolic inactivation is the basis for cross-
resistance to chlorsulfuron in diclofop-methyl-resistant rigid ryegrass (Lolium
rigidum) biotype SR4/84. Pestic. Biochem. Physiol. 43:182-192.
Coupland, D. (1994). Resistance to the auxin analog herbicides. /n Herbicide Resistance in
Plants: Biochemistry and Biology, (S.B. Powles and J.A.M. Holtum, Eds.), CRC
Press Inc., Boca Raton, EL, pp.l7l-214.

l6s
Coupland, D., Lutman, P.J.W. and Health, C. (1990). Uptake, translocation and
metabolism of mecoprop in a sensitive and a resistant biotype of Stellaria media.
Pestic. Biochem. Physiol. 36: 6l'67.
Dahroug, S. and Muller, F. (1990). Response of susceptible and tolerant plants to
diclofop-methyl. J. Plant Dis. Prot. 97: 154-167.
Devine, M.D., Maclsaac, S.4., Romano, M.L. and Hall, J.C. (1992). Investigation of the
mechanism of diclofop resistance in two biotypes of. Averufatua. Pesttc. Biochem.
Physiol. 42:88-96.
Devine, M.D., Marles, M.A.S. and Hall, L.M. (1991). Inhibition of acetolactate synthase
in susceptible and resistant biotypes of Stellaria media. Pestic. Sci. 31: 273-280.
Devine, M.D., Hall, J.C., Romano, M.L., Marles, M.A.S., Thompson, L.W. and
Shimabukuro, R.H. (1993). Diclofop and fenoxaprop resistance in wild oat is
associated with an altered effect on the plasma membrane electrogenic potential.
Pestic. Biochem. Physiol. 45: 167-177.
Devine, M.D. and Shimabukuro, R.H. (1994). Resistance to acetyl coenzyme A
carboxylase inhibiting herbicides. /z Herbicide Resistance in Plants: Biochemistry
and Biology, (S.8. Powles and J.A.M. Holtum, Eds.), CRC Press Inc., Boca
Raton, FL, pp. 14l-169.
Dicks, J.W., Slater, J.lW. and Bewick, D.W. (1985). PP 005- The R-enantiomer of
fluazifop-butyl. Proc. British Crop Prot. Conf. -V/eeds: 271-280.
DiTomaso, J.M., Brown, P.H., Stowe, 4.E., Linscott, D.L. and Kochian, L.V. (1991).
Effects of diclofop and diclofop-methyl on membrane potentials in roots of intact
oat, maize and pea seedlings. Plant Physiol. 95: 1063-1069.
DiTomaso, J.M. (1993). Membrane response to diclofop is pH dependent and is regulated
by the protonated form of the herbicide in roots of pea and resistant and susceptible
rigid ryegrass. Plant Physiol. 102: L33l-1336.

166
Donald, W. \V. and Shimabukuro, R. H. (1980). Selectivity of diclofop-methyl between
wheat and wild oat: growth and herbicide metabolism. Physiol. Plant. 49:459-
464.
Dotray, P.4., DiTomaso, J.M., Gronwald, J.}V., V/yse, D.L. and Kochian, L.V. (1993).
Effects of acetyl-coenzyme A carboxylase inhibitors on root cell transmembrane
electric potentials in graminicide-tolerant and -susceptible corn (kamays L.). Plant
Physiol. 103:919-924.
Duke, S.O. and Kenyon, W.H. (1988). Polycyclic alkanoic acids. fn Herbicides:
Chemistry, Degradation, Mode of Action. Vol. 3, (P.C. Kearney and D.D.
Kaufman, Eds.), Marcel-Dekker, NY, pp. 71-116.
Egli, M.4., Gengenbach,8.G., Gronwald, J.'W., Somers, D.A. and Wyse, D.L. (1993).
Cha¡acterization of maize acetyl coenzyme A carboxylase. Plant Physiol. 101:
499-506.
Evenson, K.J., Gronwald, J.rW. and Wyse, D.L. (1994.) Purification and characterization
of acetyl-coenzyme A carboxylase from diclofop-resistant and -suscepttble l-olium
multiþrum. Plant Physiol. 105: 671-680.
Finlayson, S. and Dennis, D.T. (1983). Acetyl-coenzyme A carboxylase from developing
endosperm of Ricinus communis. I. Isolation and characterization. Archv.
Biochem. Biophys. 255: 576-585.
Frear, D.S., Swanson, H.R. and Thalaker, F.W. (1991). Induced microsomal oxidation
of diclofop, triasulfuron, chlorsulfuron and linuron in wheat. Pestic. Biochem.
Physiol. 4t 274-287.
Friesen, H.4., O'Sullivan, P.A. and Vanden Born, Vf.H. (1976). HOE 23408, a new
selective herbicide for wild oats and green foxtail in wheat and barley. Can. J.
Plant Sci. 56: 567-578.

t67
Fuerst, E.P., Nakatani, H.Y., Dodge, 4.D., Penner, D. and Arntzen, C.J. (1985)
Paraquat resistance ln Conyza. Plant Phys iol. 77 : 984-989:
Gerwick, B.C. (1988). Potential mechanisms for bentazon antagonism with haloxyfop.
Weed Sci. 36: 286-290.
Genryick, B.C., Jackson, L.4., Handly, J., Gray, N.R. and Russell, J.W. (1988). Pre-
emergence and post-emergence activities of the (R) and (S) enantiomers of
haloxyfop. Weed Sci. 36: 453456.
Gressel, J., Regev, Y., Malkin, S. and Kleifeld, Y. (1983). Characterization of a s-
triazine-resistant biotype of Brachypodium distachyon V/eed Sci. 31: 450456.
Gronwald, J.W. (1991). Lipid biosynthesis inhibitors. V/eed Sci. 39: 43549.
Gronwald, J.rW. (1994). Resistance to photosystem tr inhibiting herbicides. /n Herbicide
Resistance in Plants: Biochemistry and Biology (S.B. Powles and J.A.M. Holtum,
Eds.) CRC Press Inc., Boca Raton, EL, pp.27-60.
Gronwald, J.W., Eberlein, C.V., Betts, K.J., Baerg, R.J., Ehlke, N.J. and Wyse, D.L.
(1992). Mechanism of diclofop resistance in an Italian ryegrass (Lolium
multiþrum Lam.) biotype. Pestic. Biochem. Physiol. 44: 126-139.
Gronwald, J.W., Pfister, K. and Steinback, K.E. (1989). Atrazine resistance in velvetleaf
(Abutilon theophrastd) due to enhanced atrazine detoxification. Pestic. Biochem.
Physiol. 34: 149-163.
Gwynne, D.C. and Murray, R.B. (1985). Grass weeds- wild oats. /n V/eed Biology and
Control in Agriculture and Horticulture, (D.C. Gwynne and R.B. Murray, Eds.),
Batsford Academic and Educational, London, pp. 99-1L2.
Hadfield, S.T., rù/ilson, 4., Kuet, S.F. and Mason, R. (1994). The metabolism of
tralkoxydim in field grown spring wheat and maize plant cell suspension culture.
Pestic. Sci. 40: 193-20[..

168
Hall, C., Edgington, L.V. and Switzer, C.M. (19S2). Translocation of different 2,4-D,
bentazon, diclofop or diclofop-methyl combinations in oat (Avena sativa) and
soybean (Glycine mtu\). lVeed Sci. 30: 676-682.
Hall, L.M., Holtum, J.A.M. and Powles, S.B. (1994). Mechanisms responsible for cross
resistance and multiple resistance. /n Herbicide Resistance in Plants: Biochemistry
and Biology, (S.8. Powles and J.A.M. Holtum, Eds.), CRC Press Inc., Boca
Raton, FL, pP. 243-26L.
Hall, L.M., Moss, S.R. and Powles, S.B. (1993). Towa¡ds an understanding of
resistance to APP herbicides in Alopecurus rnyosuroides. Froc. l4th Asian Pacific
Weed Science Society, Brisbane, Australia, pp. 299-301.
Harker, K.N. and Blackshaw, R.E. (1991). Influence of growth stage and broadleaf
herbicides on Ealkoxydim activity. rÙVeed Sci. 39: 650-659.
Harwood, J.L. (1983). Fatty acid metabolism. Annu. Rev. Plant Physiol. 39: 155-162.
Harwood, J.L. (1989). The properties and importance of acetyl-coenzyme A carboxylase
in plants. Proc. British Crop Prot. Conf.-'Weeds: 155-162.
Harwood, J.L., Ridley, S.M. and Walker, K.A. (1989). Herbicides inhibiting lipid
synthesis. In Herbicide and Plant Metabolism, (4.D. Dodge, Ed.), Cambridge
University Press, NY, pp. 73-96.
Harwood, J.L., Walker, K.4., Abulnaja, D. and Ridley, S.M. (1987). Herbicides
affecting lipid metabolism. Proc. British Crop Prot. Conf. -'Weeds: 159-169.
Häusler, R.E., Holtum, J.A.M. and Powles, S.B. (1991). Cross-resistance to herbicides
in annual ryegrass (I-olium rigidum).IV. Correlation between membrane effects
and resistance to graminicides. Plant Physiol. 97: 1035-1043.

169
Heap, I.M., Murray, B.G., Leoppsþ, H.A. and Morrison, I.N. (1993). Resistance to
aryloxyphenoxypropionate and cyclohexanedione herbicides in wild oat (Avena
latua). Weed Sci.41: 232-238.
Heap, J. and Knight, R. (1982). A population of ryegrass tolerant to the herbicide
diclofop-methyl. J. Aust. Agric. Sci.48: 156-157.
Heap, I.M. and Knight, R. (1986). The occurrence of herbicide cross resistance in a
populationof annualryegrass, Loliumrigídum, resistanttodiclofop-methyl. Aust.
J. Agric. Res.37: 149-156.
Heap, I.M. and Knight, R. (1990). Variation in herbicide cross-resistance among
population of annual ryegrass (Lolium rtgidum) resistant to diclofop-methyl. Aust.
J. Agric. Res. 41: L2l-128.
Hendley, P., Dicks, J.'W., Monaco, T.J., Slyfield, S.M., Tummon, O.J. and Ba¡rett, J.C.
(1985). Translocation and metabolism of pyridinyloxyphenoxy-propionate
herbicides in rhizomatous quackgrass (Agropyron repens). rJVeed Sci. 33: ll-24.
Higinbotham, N., Graves, J.S. and Davis, R.F. (1970). Evidence for an electrogenic ion
transport pump in cells of higher plants. J. Membr. Biol. 3: 2lO-222.
Hoagland, D.R. and Arnon, D.L (1938). The water-culture method for growing plants
without soil. Calif. Agric. Exp. Stn. Circ.347; l-32.
Hoerauff, R.A. and Shimabukuro, R.H. (1979). The response of resistant and susceptible
plants to diclofop-methyl. Weed Res. 19: 293-299.
Holt, J.S., Powles, S.B. and Holtum, J.A.M. (1993). Mechanisms and agronomic aspects
of herbicide resistance. Annu. Rev. Plant Physiol. Plant Mol. Biol. 44: 203-229.
Holtum, J.A.M. and Powles, S.B. (1991). Annual ryegrass: an abundance of resistance, a
plethora of mechanisms. Proc. Brighton Crop Prot. Conf. -'Weeds: l07l-1077.

t70
Holtum, J.A.M., Matthews, J.M., Häusler, R.E., Liljegren, D.A. and Powles, S.B.
(1991). Cross-resistance to herbicides in annual ryegrass (Loliutn rigidum).Itr. On
the mechanism of resistance to diclofop-methyl. Plant Physiol. 97: lO2Gl034.
Holtum, J.A.M., Häusler, R.E., Devine, M.D. and Powles, S.B. (1994). Recovery of
transmembrane potentials in plants resistant to aryloxyphenoxypropanoate
herbicide: a phenomenon awaiting explanation. Weed Sci. 42: 293-301.
Hosaka, H., Hideo, I., and ishikawa, H. (1984). Response of monocotyledons to BAS
9052 OH. Weed Sci.32: 28-32.
Hosaka, H., Inaba, H., Satoh, 4., Tanoue, T. (1987). Selectivity mechanisms of
sethoxydim between red fescue (Festuca rubra) and tall fescue (Festuca
arundinaceø). Weed Res. (Japan) 32 255-262.
Howard, J.L. and Ridley, S.M. (1990). Acetyl-CoA carboxylase: a rapid novel assay
procedure used in conjunction with the preparation of enzyme from maize leaves.
FrEBS Lett. 261: 26I-264.
Hutson, D.H. and Roberts, T.R. (1987). Wild oat herbicides. In Progress in Pesticide
Biochemistry and Toxicology. (D.H. Hutson and T.R. Roberts, Eds.), John Wiley
& Sons Ltd., NY, pp. t69-197.
Imam, A.G. and Allard, R.W. (1965). Population studies in predominantly self-pollinated
species. VI. Genetic variability between and within natural population of wild oat in
differing habitats in California. Genetics 51: 49-52.
Ishikawa, H., Yamada, S., Hosaka, H., Kawana, T., Okunuki, S. and Kohara, K.
(1985). Herbicidal properties of sethoxydim for the control of gramineous weeds.
J. Pestic. Sci. 10: 187-193.
Ishihara, K., Shiotani, H., Soeda, Y., Ono, S. (1988). Fate of the herbicide sethoxydim in
sugar beet. J. Pestic. Sci. 13: 231-237.

17t
Itoh, K. and Miyahara, M. (1984). Inheritance of paraquat resistance in Erigeron
phitadetphicus L. Weed Res. (Japan) 29: 3Ot-307 .
Islam, A.K.M.R. and Powles, S.B. (1988). Inheritance of resistance to paraquat in badey
grass Hordeum glaucum Steud. Weed Res. 28: 393'397.
Jacobson, A. and Shimabukuro, R.H. (1982). The absorption and translocation of
diclofop-methyl and amitrole in wheat and oat roots. Physiol. Plant. 54:3440.
Jasieniuk, M., Brtlé-Babel, A.L. and Morrison, I.N. (1994). Inheritance of trifluralin
resistance in green foxtail (Setaria viridis). V/eed Sci. 42: 123-127 .
Jensen, K.I.N. and Caseley, J.C. (1990). Antagonistic effects of 2,4-D amine and
bentazone on control of Avenafatuawithtralkoxydim. Weed Res. 30: 389-395.
Jordan, D.L., York, A.C. and Corbin, F.T. (1989). Effect of ammonium sulfate and
bentazon on sethoxydim absorption. V/eed Technol. 3:674-677.
Joseph, O.O., Hobbs, S.L.A. and Jana, S. (1990). Diclofop resistance in wild oat (Avena
fatua). Weed Sci. 38: 475479.
Kannangara, C.G. and Stumpf, P.K. (1972). Fat metabolism in higher plants. A
prokaryotic type acetyl CoA ca¡boxylase in spinach chloroplasts. Arch. Biochem.
Biophys. 152: 83-91.
Kell, J.J., Meggitt, W.F. and Penner, D. (1984). Absorption, translocation and activity of
fluazifop-butyl as influenced by plant growth stage and environment. V/eed Sci.
32: 143-149.
Kobek, K., Forcke, M., Litchtenthaler, H.K., Retzlaff, G. and Wurzer, B. (1988).
Inhibition of fatty acid biosynthesis in isolated chloroplasts by cycloxydim and
other cyclohexane-1, 3-diones. Physiol. Plant. 72: 492498.

172
Köcher, H., Kellner, H.M., Lotzsch, K. and Dorn, E. (1982). Mode of action and
metabolic fate of the herbicide fenoxaprop-ethyl, HOE 33171. Proc. British Crop
Prot. Conf. -Weeds: 341'347.
Konishi, T. and Sasaki, Y. (1994). Compartmentalization of wo forms of acetyl-CoA
carboxylase in plants and the origin of their tolerance toward herbicides. Proc.
Natl. Acad. Sci. USA 91: 3598-3601.
Leach, G.E., Kirkwood, R.C. and Ma¡shall, G. (1993). The basis of resistance displayed
to fluazifop-butyl by biotypes of. Elucine indica. Proc. Brighton Crop Prot. Conf.
-'Weeds: 2OI-207.
Lefsrud, C. and Hall, J.C. (1989). Basis for sensitivity differences among crabgrass, oat,
and wheat to fenoxaprop-ethyl. Pestic. Biochem. Physiol. 34:218-227.
Lewis, J. (1961). The influences of water level, soil depth and type on the survival of crop
and weed seeds. Proc. Int. Seed Test Assoc. 26: 68-85.
Li, S.J. and Cronan, J.E. (1992a). The gene encoding the biotin carboxylase subunit of
Escherichia coli acetyl-CoA carboxylase. J. Biol. Chem. 267:855-863.
Li, S.J. and Cronan, J.E. (1992b). The genes encoding the two carboxyltransferase
subunits of. Escherichia coli acetyl-CoA carboxylase. J. Biol. Chem. 267 16841-
16847.
Lichtenthaler, H.K. and Kobek, K. (1987). Inhibition by sethoxydim of pigment
accumulation and fatty acid biosynthesis in chloroplasts of Avena seedlings. Z.
Naturforsch 42c: 127 5 -127 9.
López-Casillas, F., Bai, D.H., Luo, X. Kong, I.S., Hermodson, M.A. and Kim, K.H.
(1988). Stn¡cture of the coding sequence of acetyl-coerøyme A carboxylase. Proc.
Natl. Acad. Sci. USA 85: 5784-5788.

t73
Lucas, \M.J., lVilson, C. and Wright, J.P. (1984). Perbutation of Chara plasmalemma
transport function by 214(2',4'-dichlorophenoxy)phenoxy'lpropionic acid. Plant
Physiol. 74:61-66.
Mansooji, 4.M., Holtum, J.A.M., Boutsalis, P., Matthews, J.M. and Powles, S.B.
(1992). Resistance to aryloxyphenoxypropionate herbicides in two wild oats
species (Avenafatua andAvena sterilis). lVeed Sci. 40: 599-605.
Mansooji, A.M. (1994). Herbicide resistance in wild oats, Avend spp. Ph.D. Thesis,
Universþ of Adelaide, Ausralia.
Mallory-Smith, C.4., Thill, D.C., Dial, M.J. and Zemet¡a, R.S. (1990). Inheritance of
sulfonylurea herbicide resistance tn l-actuca spp. V/eed Technol. 4:787-79O.
Marles, M.A.S., Devine, M.D. and Hall, J.C. (1993). Herbicide resistance in Setaria
viridis conferred by a less sensitive form of acetyl coenzyme A carboxylase. Pestic.
Biochem. Physiol. 46: 7 -L4.
Marshall, L.C., Somers, D.4., Dotray, P.D., Gengenbach, B.G., 'Wyse, D.L. and
Gronwald, J.W. (1992). Allelic mutations in acetyl-coenzyme A carboxylase
confer herbicide tolerance in maize. Theor. Appl. Genet. 83:435442.
Marorder, H.L., Prego, I.A. and Cairoli, M.A. (1987). Behaviour of l4C haloxyfop in
common bermudagrass (Cynodon dacryIon) stolons. Weed Sci. 35: 599-603.
Marshall, D.R. and Jain, S.K. (1970). Seed predation in the population dynamics of Avena
fatw andAvenabarbata. Ecology 51: 886-891.
Martin, R.J., McMillian, M.G. and Cook, J.B. (1988). Survey of farm management
practices of the northern wheat belt of New South Wales. Aus. J. Exp. Agric. 28:
499-509.

r74
Manwell, B.D. and Mortimer, A.M. (1994). Selection for herbicide resistance. In
Herbicide Resistance in Plants: Biochemistry and Biology, (S.8. Powles and
J.A.M. Holtum, Eds.), CRC Press Inc., Boca Raton, FL' pp' 1-26'
McAlister, F.M., Holtum, J.A.M. and Powles, S.B. (1995). Dinitroaniline herbicide
resistance in rigid ryegfass (Lolium rigidum). Iffeed Sci. 43: 55-62.
McFadden, J.J., Frear, D.S. and Mansager, E.R. (1989). Aryl hydroxylation of diclofop
by a cytochrome P-450 dependent monooxygenase from wheat. Pestic. Biochem.
Physiol. 34:92-100.
Medd, R.W. and Pandey, S. (1990). Estimating the cost of wild oats (Avena spp.) in the
Australian wheat indusry. Plant Prot. Quaterþ 5: t42'144.
Medd, R.W., McMillan, M.G. and Cook, A.S. (1992). Spray-topping of wild oats (Avena
spp.) in wheat with selective herbicides. Plant Prot. Quarterly 7:62'65.
Miller, S.D. and Nalewaja, J.D. (1990). Influence of burial depth on wild oats (Avena
fatua) seed longevity. V/eed Technol. 4:514-517.
Moss, S.R. (1990). Herbicide cross-resistance in slender foxtail (Alopecurus
myosuroides). Weed Sci. 38: 492496.
Mougin, C., Polge, N., Scalla, R. and Cabanne, F. (1991). Interactions of va¡ious
agrochemicals with cytochrome P450-dependent monoxygenases of wheat cells.
Pestic. Biochem. Physiol. 40: 1-11.
Mueller, T.C., William, W.'W. and Barrett, M. (1989). Antagonism of Johnsongrass
(Sorghum halepense) control with fenoxaprop, haloxyfop and sethoxydimby 2,4-
D. Weed Technol. 3: 86-89.
Mueller, T.C., Ba¡rett, M. and'Witt, W.W. (1990). A basis for the antagonistic effect of
2,4-D on haloxyfop-metþl toxicity to johnsongrass (Sorghum halepense). rWeed
Sci.38: 103-107.

175
Naka¡nura, Y. and Yamada, M. (1975). Fatty acid synthesis by spinach chloroplasts. I.
Properties of fatty acid synthesis from acetate. Plant Cell Physiol. 16z 139-149.
Nakamura, Y. and Yamada, M. (1979). The light-dependent step of de novo synthesis of
long chain fatty acids in spinach chloroplasts. Plant Sci. I-ett. 14:29I-295.
Naylor, J.M. and Simpson, G.M. (1961). Dormancy studies in seeds of Avena fatua.n.
A gibberellin-sensitive inhibitory mechanism in the embryo. Can. J. Bot. 39: 281-
295.
Nikolau, B.J. and Hawke, J.C; (1934). Purification and cha¡acterization of maize leaf
acetyl-coenzyme A carboxylase. Arch. Biochem. Biophys. 228:86-96.
O'Donavan, J.T., Sharma, M.P., Harker, K.N., Maurice, D., Baig, M. and Blackshaw,
R.E. (1994). Wild oat (Avena fatua) populations resistant to triallate are also
resistant to difenzoquat. Weed Sci. 42: 195-199.
Parker,'W.B., Marshall, L.C., Burton, J.D., Somers, D.4., Wyse, D.L., Gronwald, J.W.
and Gengenbach, B.G. (1990a). Dominant mutations causing alterations in acetyl
coenzyme A carboxylase confer tolerance to cyclohexanedione and
aryloxyphenoxypropionate herbicides in maize. Proc. Natl. Acad. Sci. USA 87:
7175-7r79.
Parker, Vf.B., Somers, D.4.,'Wyse, D.L., Keith, R.4., Burton, J.D., Gronwald, J.W.
and Gengenbach, B.G. (1990b), Selection and cha¡acterization of sethoxydim-
tolerant maize tissue cultures. Plant Physiol.92: t22O-1225.
Paterson, J.G. (1976). The distribution and ecology of wild oats (Avena spp.) in Western
Australia. J. Appl. Ecol. 13: 257-26r''
Paterson, J.G., Boyd, V/.J.R. and Goodchild, N.A. (1976). Effect of temperature and
depth of burial on the persistence of seed of. AvenafatuaL. in'Western Australia. J.
Appl. Ecol. 13: 841-847.

176
Peters, N.C.B. (1986). Factors affecting sssdling emergence of different strains of. Avena
fatw L. Weed Res. 26: 29-38.
Powles, S.B. (1986). Appearance of a biotype of the weed, Hordeum glaucum Steud.,
resistant to herbicide Paraquat. $[eed Res. 26: t67-172.
Powles, S.B. and Matthews, J.M. (1992). Multiple herbicide resistance in annual ryegrass
(Lotium rigidum): a driving force for the adoption of the integrated weed
mangement. /n Resistance '91: Acheivements and Developments in Combating
Pesticide Resistance, (I. Denholm, A.L. Devonshire and D.W. Hollomon, Eds.),
Elsevier, NY, pp. 75-87.
Powles, S.8., Tucker, E.S. and Morgan, T.R. (1989). A capeweed (Arctotheca calendula)
biotype in Australia resistant to bipyridyl herbicides. rWeed Sci. 37: 60-62.
Preston, C. (1994). Resistance to photosystem I inhibiting herbicides. In Herbicide
Resistance in Plants: Biochemistry and Biology (S.8. Powles and J.A.M. Holtum,
Eds.), CRC Press Inc., Boca Raton, FL, pp. 6l-82.
Preston, C., Holtum, J.A.M. and Powles, S.B. (1992). On the mechanism of resistance to
paraquat in Hordeum glaucum and I1. leporinum. Delayed inhibition of
photosynthetíc 02 evolution after paraquat application. Plant Physiol. 100: 630-
636.
Preston, C., Balachandran, S. and Powles, S.B. (L994). Investigation of mechanisms of
resistance to bipyridyl herbicides in Arthotheca calendula (L.) Levyns. Plant Cell
Environ. 17 : lll3-1123.
Purba, E. (1993). Factors influencing the development of resistance to the bipyridyl
herbicides in Australia. Ph.D. Thesis, University of Adelaide, Austalia.
Purba, E., Preston, C. and Powles, S.B. (1993a). Inheritance of bipyridyl herbicide
resistance in Anhotheca calendula and Hordeum leporinum. Theor. Appt. Genet.
87:598-602.

t77
Purba, E., Preston, C. and Powles, S.B. (1993b). Paraquat resistance in a biotype of
Vulpia bromoides (L.) S.F. Gray. lVeed Res. 33: 409413.
Quail, P.H. and Carter, O.G. (1968). Survival and seasonal germination of seeds of Avena
Íatua andÁ. ludoviciana. Aust. J. Agric. Res. 19: 721'729.
Ratterman, D.M. and Balke, N.E. (1988). Herbicide disruption of proton gradient
development and maintainance by plasmalemma and tonoplast vesicles from oat
root. Pectic. Biochem. Physiol. 3l:22I-236.
Ratterman, D.M. and Balke, N.E. (1989). Diclofop-methyl increases the proton
permeability of isolated oat root tonoplast. Plant Physiol. 91: 756-765.
Rendina, A.R. and Felts, J.M. (1988). Cyclohexanedione herbicides are selective and
potent inhibitors of acetyl-CoA carboxylase from grasses. Plant Physiol. 86: 983-
986.
Rendina, 4.R., Felts, J.M., Beaudoin, J.D., Craig-Kennard, 4.C., Look, L.L., Paraskos,
S.L. and Hangenah, J.A. (1988). Kinetic characterization, stereoselectivity, and
species selectivity of the inhibition of plant acetyl-CoA carboxylase by the
aryloxyphenoxypropionic acid herbicides. Arch. Biochem. Biophys. 265: 219-
225.
Rendina, A.R., Beaudoin, J.D., Craig-Kennard, A.C. and Breen, M.K. (1989). Kinetic
of inhibition of acetyl-coenzymeA carboxylase by aryloxyphenoxypropionate and
cyclohexanedione graminicides. Proc. British Crop Prot. Conf. -'Weeds: 163-172.
Rendina, 4.R., Craig-Kennard, 4.C., Beaudoin, J.D. and Breen, M.K. (1990).
Inhibition of acetyl-coenzyme A carboxylase by two classes of grass-selective
herbicides. J. Agric. Food Chem.38: 1282-87.
Ritter, R.J. (1989). The spread of triazine-resistant weeds. In Understanding Herbicide
Resistance in W'eeds. Sandoz Crop Prot. Corp., Illinois, pp.16-20.

178
Rgyneberg, T., Balke, N.E. and Lund-Høie, K. (1994). Cycloxydim absorption by
suspension-culn¡red velvetleaf (Abutilon theophrastí Medic.) cells. 'Weed Res. 34:
l-9.
Ryan, G.F. (1970). Resistance of corn¡non groundsel to simazine and atrazine. Weed Sci.
18: 614-616.
Saari, L.L., CotteÍnan, J.C. and Primiani, M.M. (1990). Mechanism of sulfonylurea
herbicide resistance in the broadleaf weed, Kochia scoparia. Plant Physiol.
93:55-61.
Saari, L.L., Cottennan, J.C., Smith, lV.F. and Primiani, M.M. (1992). Sulfonylurea
herbicide resistance in common chickweed, perennial rye$ass and Russian thistle.
Pestic. Biochem. Physiol. 42: 110-1 18.
Saari, L.L., Cottennan, J.C. and Thill, D.C. (1994). Resistance to acetolactate synthase
inhibiting herbicides. /n Herbicide Resistance in Plants: Biochemistry and Biology,
(S.8. Powles and J.A.M. Holtum, Eds.), CRC Press Inc., Boca Raton, FL, Pp.
83-139.
Sasaki, Y., Hakamada, K., Suama, Y., Nagano, Y., Furusawa, I., Matsuno, R. (1993).
Chloroplast-encoded protein as a subunit of acetyl-CoA carboxylase in pea plant.
J. Biol. Chem. 268: 25118-25123.
Schmitzer, P.R., Eilers, R.J. and Cséke, C. (1993). Lack of cross-resistance of
imazaquin-resistantX¿nthium strumoriurn acetolactate synthase to flumetsulam and
cilorimuron. Plant Physiol. 103: 281-283.
Secor, J. and Cséke, C. (1988). Inhibition of acetyl-CoA carboxylase activity by
haloxyfop and tralkoxydim. Plant Physiol. 86: L0-t2.
Secor, J., Cséke, C. and Owen, V/.J. (1989). The discovery of the selective inhibition of
acetyl coenzyme A carboxylase activity by two classes of graminicides. Proc.
British Crop Prot. Conf. -Weeds: 145-154.

t79
Shaaltiel, Y., Chua, N.H., Gepatein, S. and Gressel, J. (1988). Dominant pleiotropy
controls enzymes co-segragating with paraquat resistance'in Conyza bonariensis.
Theor. Appl.Genet. 75: 850-856.
Shimabukuro, R.H., Walsh, W.C. and Hoerauf, R.A. (1979). Metabolism and selectivity
of diclofop-methyl in wild oat and wheat. J. Agric. Food Chem.27:615-620.
Shimabukuro, R.H. (1990). Selectivity and mode of action of the Post emergence herbicide
diclofop-methyl. PGRSA Quarterly lt: 37 -54.
Shimabukuro, R.H. and Hoffer, B.L. (1991). Metabolism of diclofop-methyl in
susceptible and resistant biotypes of Lolium rigidum. Pestic. Biochem. Physiol.
39:251-260.
Shimabukuro, R.H. and Hoffer, B.L. (L992). Effect of diclofop on membrane potentials
of herbicide-resistant and -susceptible annual ryegrass root tips. Plant Physiol. 98:
l4t5-1422.
Shimabukuro, R.H., Walsh, V/.C. and Jacobson, J.A. (1987). Aryl-Ø-glucoside of
diclofop: A detoxification product in wheat shoots and wild oat suspension culture.
J. Agric. Food Chem.35:393-397.
Shoaf, A.R. and Carlson, W.C. (1986). Analytical techniques to measure sethoxydim and
breakdown products. Weed Sci. 34: 745-751.
Shoaf, A.R. and Carlson, W.C. (1992). Stability of sethoxydim and its degradation
products in solution, in soil, and on surfaces. V/eed Sci. 40: 384-389.
Shooler, 4.8., Bell, A.R., Nalewaja, J.D. (1972). Inheritance of siduron tolerance in
foxtail barley. 'Weed Sci. 20: t67-169.
Shorrosh, B.S., Dixon, A.D. and Ohlrogge, J.B. (1994). Molecular cloning,
cha¡acterization, and elicitation of acetyl-CoA carboxylase from alfafa. Proc. Natl.
Acad. Sci. USA 9l: 4323-4327.

180
Smeda, R.J., Barrentine, W.L. and Snipes, C.E. (1993). Johnsongrass (Sorghum
halepense L. Pers.) resistance to post emergence grass herbicides. V/SSA Abstract
33: 53.
Smeda, R.J. and Vaughn, K.C. (1994). Resistance to dinitroaniline herbicides. fn
Herbicide Resistance in Plants: Biochemistry and Biology, (S.8. Powles and
J.A.M. Holtum, Eds.), CRC Press Inc., Boca Raton, FL, pp. 215-228.
Smith, A.M. and Vanden Born, W.H. 1992. Ammonium sulfate increases efficacy of
sethoxydim through increased absorption and translocation. Weed Sci. 40: 351-
358.
Stanger, C.E. and Appleby, A.P. (1989). Italian ryegr¿¡ss (I-olium multiþrum ) accessions
tolerant to diclofop. V/eed Sci. 37: 350-352.
Sterling, T.M. (1994). Mechanisms of herbicide absorption across plant membranes and
accumulation in plant cells. Weed Sci. 42:263-276.
Stoltenberg, D.E., Gronwald, J.W., Wyse, D.L., Burton, J.D., Somers, D.A. and
Gengenbach, B.G. (1939). Effect of sethoxydim and haloxyfop on acetyl
coenzyme A carboxylase activity in Festuca species. Weed Sci. 37 : 512-516.
Stoltenberg, D.E. and Vfeiderholt, R.J. (1993). Giant foxtail (Setaria faberi Herrm.)
resistance to acetyl-CoA carboxylase inhibitors. WSSA Abstract 33: 183.
Stonebridge, W.C. (1981). Selective post-emergence grass weed control in broad-leaf
arable crops. Outl. Agric. 10: 385-392.
Swisher, B.A. and Corbin, F.T. (1982). Behavior of BAS-90520H in soybean and
Johnsongrass plant and cell cultures. \Veed Sci. 30: 640-650.
Takai, T., Yokohama, C., Wada, K. and Tanabe, T. (1988). Primary srtucture of chicken
liver acetyl CoA carboxylase deduced from cDNA sequence. J. Biol. Chem. 263:
2651-2657.

181
Tal, 4., Romano, M.L.Stephenson, G.R., Schwan, A.L. and Hall, J.C. (1993).
Glutathione conjugation: a detoxification pathway for fenoxaprop-ethyl in barley,
crabgrass, oat, and wheat. Pestic. Biochem. Physiol. 46: 190-199.
Tanaka, Y., Chisaka, H. and Saka, H. (1986). Movement of paraquat in resistant and
susceptible biotypes of Erigeron Philadelphiczs and E. canadensis. Physiol. Plant.
66: 605-608.
Tardif, F.J., Holtum, J.A.M. and Powles, S.B. (1993). Occurrence of a herbicide-
resistant acetyl-coenzyme A carboxylase mutant in annual ryegrass (Lolium
rigidum) selected by sethoxydim. Planta 190: 176-181.
Tardif, F.J. and Powles, S.B. (1993). Target-site-based resistance to herbicides inhibiting
acetyl-CoA carboxylase. Proc. Brighton Crop Prot. Conf. -Weeds: 533-540.
Ta¡dif, F.J. and Powles, S.B. (1994). Herbicide multiple-resistance in a Inlium rigidum
biotype is endowed by multiple mechanisms: isolation of a subset with resistant
acetyl-CoA carboxylase. Physiol. Plant. 9L: 488494.
Thomas, H. and Jones, D.D. (1976). Origin and identification of weed species of Avena.
/n V/ild Oats in lVorld Agriculture, (D.P. Jones, Ed.), Agricultural Resea¡ch
Council, London, pp. 1-18..
Thurston, J.M. (1951). Some experiments and field observations on the germination of
wild oats (A. fatua and A. ludoviciana) seeds in soil and the emergence of
seedlings. Ann. Appl. Biol.38: 812-832.
Thurston, J.M. (1959). A comparative study of the growth of wild oats ( Avenafatua L.
andAvena ludovicianø Dur.) and of cultivated cereals with varied nitrogen supply.
Ann. Appl. Biol. 47:7t6-793.
Thurston, J.M. (1966). Survival of seeds of wild oats (Avena fatua and A. Iudoviciana)
and cha¡lock (Sinøpsr,s ravensis) in soil under leys. rWeed Res. 6: 67-80.

t82
Todd, B.G. and Stobbe, E.H. (1980). The basis of the antagonistic effect of 2,4-D on
diclofop-methyl toxicity to wild oat (Avenafatua). lVeed Sci. 2E: 371-377.
Tritter, S.4., Holl, F.B. and Todd, B.G. (1987). Diclofop-methyl and diclofoP uptake in
oat (Avena sativa L.) protoplasts. Can. J. Plant Sci. 67: 2t5-223.
Tucker, E.S. and Powles, S.B. (1991). A biotype of hare barley (Hordeum leporinum)
resistant to paraquat and diquat. Weed Sci.39: 159-162.
Tucker, T.4., Langeland, K.A. and Corbin, F.T. (1994). Absorption and translocation of
t{C-imazapyr and l4C-glyphosate in alligatorweed (Altemanthera philoxeroides).
Weed Technol. 8:32-36.
Turnham, E. and Northcote, D.H. (1983). Changes in the activity of acetyl-CoA
carboxylase during rape-seed formation. Biochem. I . 212: 223 -229 .
Uchiyama, M., Washio, N., Ikai, T., Igarashi, H. and Suzuki, K. (1986). Stereospecific
responses to (R)-(+)-and (S)-(l-quizalofop-ethyl in tissues of several plants.
Pestic. Sci. 11: 459-467.
Watkins, F.B. (1971). Efffects of annual dressings of nitrogen fertilizer on wild oat
infestations. Weed Res. 11: 292-3OI.
Walker, K.4., Ridley, S.M., Lewis, T. and Harwood, J.L. (1989). Action of aryloxy-
phenoxy carboxylic acid on lipid metabolism. Rev. Weed Sci. 4: 7l-84.
'Walter, H., Koch, W. and Muller, F. (1980). EinfluB des Applikationsortes auf die
Penetration und Translokation von diclofop-methyl bei Flughafer (AvenafatuaL.).
Weed Res. 20: 325-331.
Weber, 4., Fischer, E., VonBraitz, H.S. and Lüttge, U. (1988). The effects of the
herbicide sethoxydim on transport processes in sensitive and tolerant grass species.
I. Effects on the electrical membrane potential and alanine uptake. Z. Naturforsch
43c:249-256.

183
Weber, A. and Lünge, U. (1988). The effects of the herbicide sethoxydim on transport
processes in sensitive and tolerant grass species. tr. Effects on membrane-bound
redox systems in plant cells. Z. Naturforsch. 43c: 257-263.
V/halley, R.D.B. and Burfrtt, J.M; (1972). Ecotypic variation ín A. fatuaL., A. sterilis
L.(A. Iudoviciana), and A. barbata Pott. in New South V/ales and southern
Queensland. Aust. J. Agric. Res.23: 799-810.
Wilson, B.J. (1972). Studies of the fate of Avena fatua seeds on cereal stubble, as
influenced by autumn treatment. hoc. British Crop Prot. Conf. -'Weeds: 242-247.
\I/ilson, R.G. (1988). Biology of weed seeds in soil. /n V/eed Management in
Agroecosystem: Ecological Approaches, (M.4. Altien and M. Liebman, Eds.),
CRC PRess, Boca Raton, FL, pp. 25-39.
V/ilson, B.J. and Cussans, G.rW. (1975). A study of the population dynamics of Avena
fatua L. as influenced by straw burning, seed shedding and cultivation. 'Weed Res.
15:249-258.
Wink, O., Dorn, E. and Beyermann, K. (1984). Metabolism of the herbicide Hoe 33171in
soybean. J. Agric. Food Chem.32 187-192.
Worthing, C.R. and Hance, R.J. (1991). The Pesticide Mannual: A World Compendium.
The British Crop Protection Council, Unwin Brothers Ltd., Old'Woking, Surrey,
UK, 1140p.
rWright, J.P. and Shimabukuro, R.H. (1987). Effects of diclofop and diclofop-methyl on
the membrane potentials of wheat and oat coleoptiles. Plant Physiol. 85: 188-193.
Zimmerlin, A. and Durst, F. (1990). Xenobiotic metabolism in plants: aryl hydroxylation
of diclofop by a cytochrome P-450 erxzyme from wheat. Phytochem.29: 1729-
t732.

184
Zimmerlin, A. and Durst, F. (1992). Aryl hydroxylation of the herbicide diclofop by a
wheat cytochfome P450 monoxygenase. Plant Physiol. 1ü): 874-881.

185
Appendix
The following l0 tables (Tables A.l-10) show the herbicide use histories of the ten resistant
biotlpes of wild oat.
Table 4.1 History of herbicide application of field from which wild oat biotype SAS 1
was collected
Year Crop Herbicide use
1981 wheat diclofop-methyl
1982 pasture none
1983 medic trifluralin, fluazifop-butyl
1984 rapeseed trifluralin, fluazifop-butyl
1985 wheat triallate, diclofop-methyl
1986 wheat diclofop-methyl
1987 clover trifluralin, fluazifop-butyl
1988 wheat glyphosate
1989 clover trifluralin, haloxyfop-ethoxyethyl (failed)

186
Table 4.2 History of herbicide application of field from which wild oat biotype NAS 4
was collected
Year Crop Herbicide use
1980 wheat diclofopmethyl
1981 wheat diclofopmethyl
1982 lupin nifluralin
1983 wheat diclofopmethyl
1984 wheat diclofopmethyl
1985 wheat diclofopmethyl
1986 pea trifluralin, fluazifop-butyl
1987 wheat fralkoxydim
1988 wheat talkoxydim
1989 canola trifluralin, haloxyfop-ethoxyethyl
1990 wheat fenoxaprop-ethyl (tailed)

187
Table 4.3 History of herbicide application of field from which wild oat biotype NAF 6
was collected
Year Crop Herbicide use
1988 pasture none
1989 wheat flamprop-methyl
1990 wheat fenoxaprop-ethyl
L99l wheat fenoxaprop-ethyl (failed)
Table 4.4 History of herbicide application of field from which wild oat biotype NAS 14
was collected
Year Crop Herbicide use
t982 wheat none
1983 wheat none
1984 wheat none
1985 lupin simazine, fluazifop-butyl
1986 wheat diclofop-methyl
1987 wheat diclofop-methyl
1988 lupin simazine, fluazifop-butyl
1989 wheat diclofopmethyl (failed)

188
Table AS History of herbicide application of field from which wild oat biotype NAF 8
was collected
Year Crop Herbicide use
1988 pasture
lupin
wheat
wheat
1989
1990
1991
Table 4.6 History of herbicide application of field from which wild oat biotype NAS 2
was collected
Year Crop Herbicide use
none
fluazifop-butyl
diclofopmethyl
diclofopmethyl (tailed)
1980
1981
1982
1983
1984
1985
1986
1987
1988
1989
wheat
wheat
lupins
wheat
wheat
wheat
pea
wheat
wheat
canola
wheat
diclofop-methyl
diclofop-methyl
tifluralin
diclofop-methyl
diclofop-methyl
diclofop-methyl
trifl uralin, fl uazifop-butyl
tralkoxydim
tralkoxydim
trifluralin, haloxyfop-ethoxyethyl
talkoxydim (failed)1990

r89
Table 4.7 History of herbicide apptication of field from which wild oat biotype NAS 6
was collected
Year Crop Herbicide use
1988 pasture none
1989 wheat flamprop-methyl
1990 wheat fenoxapropethyl
1991 wheat fenoxaprop-ethyl (tailed)
Table 4.8 History of herbicide application of field from which wild oat biotype SAF 14
was collected
Yea¡ Crop Herbicide use
1984 barley diclofop-methyl
1985 rapeseed diclofop-methyl
1986 wheat diclofopmethyl
L987 bean simazine, fluazifop-butyl
1988 wheat diclofop-methyl
1989 pea fluazifop-butyl, glyphosate
1990 wheat fenoxaprop-ethyl (failed)

190
Table 4.9 History of herbicide application of field from which wild oat biotype SAF 34
was collected
Year Crop Herbicide use
l98l pasture none
t982 titicale diclofopmethYl
1983 pasture none
1984 wheat diclofop-methyl
1985 pasture none
1986 wheat diclofop-methyl
t987 beans fluazifopbutyl
1988 wheat diclofop-methyl
1989 barley tralkoxydim
1990 wheat diclofop-methyl (failed)

191
Table 4.10 History of herbicide application of field from which wild oat biotype V/AF 12
was collected
Year Crop Herbicide use
1985 pastue none
1986 pasture none
1987 pasture none
1988 wheat tiallaæ
1989 wheat diclofopmethyl
1990 wheat diclofopmethyl
t99t wheat triasulfuron