mcx193.pdf - oxford academic
TRANSCRIPT

Annals of Botany 122: 679–696, 2018doi: 10.1093/aob/mcx193,
© The Author(s) 2018. Published by Oxford University Press on behalf of the Annals of Botany Company. All rights reserved. For permissions, please e-mail: [email protected].
available online at www.academic.oup.com/aob
REVIEW: PART OF A SPECIAL ISSUE ON FUNCTIONAL–DEVELOPMENTAL PLANT CELL BIOLOGY
The intracellular and intercellular cross-talk during subsidiary cell formation in Zea mays: existing and novel components orchestrating cell polarization and
asymmetric division
P. Apostolakos1, P. Livanos1, E. Giannoutsou1, E. Panteris2 and B. Galatis1,*
1Department of Botany, Faculty of Biology, National and Kapodistrian University of Athens, Athens, Greece and 2Department of Botany, School of Biology, Aristotle University of Thessaloniki, Thessaloniki, Macedonia, Greece
*For correspondence. E-mail: [email protected]
Received: 7 September 2017 Returned for revision: 24 October 2017 Editorial decision: 20 November 2017 Accepted: 25 November 2017 Published electronically 15 January 2018
• Background Formation of stomatal complexes in Poaceae is the outcome of three asymmetric and one symmetric cell division occurring in particular leaf protodermal cells. In this definite sequence of cell division events, the generation of subsidiary cells is of particular importance and constitutes an attractive model for studying local intercellular stimulation. In brief, an induction stimulus emitted by the guard cell mother cells (GMCs) triggers a series of polarization events in their laterally adjacent protodermal cells. This signal determines the fate of the latter cells, forcing them to divide asymmetrically and become committed to subsidiary cell mother cells (SMCs).• Scope This article summarizes old and recent structural and molecular data mostly derived from Zea mays, focusing on the interplay between GMCs and SMCs, and on the unique polarization sequence occurring in both cell types. Recent evidence suggests that auxin operates as an inducer of SMC polarization/asymmetric division. The intercellular auxin transport is facilitated by the distribution of a specific transmembrane auxin carrier and requires reactive oxygen species (ROS). Interestingly, the local differentiation of the common cell wall between SMCs and GMCs is one of the earliest features of SMC polarization. Leucine-rich repeat receptor-like kinases, Rho-like plant GTPases as well as the SCAR/WAVE regulatory complex also participate in the perception of the morphogenetic stimulus and have been implicated in certain polarization events in SMCs. Moreover, the transduction of the auxin signal and its function are assisted by phosphatidylinositol-3-kinase and the products of the catalytic activity of phospholipases C and D.• Conclusion In the present review, the possible role(s) of each of the components in SMC polarization and asymmetric division are discussed, and an overall perspective on the mechanisms beyond these phenomena is provided.
Key words: Asymmetric division, auxin, morphogenesis, pectins, polarity, reactive oxygen species, stomatal complexes
INTRODUCTION
The four-celled stomatal complexes of Poaceae constitute an ideal system to investigate the establishment of cell polarity and subsequent asymmetric division in angiosperms. They ori-ginate from three asymmetric and one symmetric cell division (Fig. 1). The first asymmetric division generates the guard cell mother cell (GMC; Fig. 1A), a cell emitting a morphogenetic signal (Fig. 1B) that establishes polarity in the laterally adjacent subsidiary cell mother cells (SMCs; Fig. 1C). The latter divide asymmetrically to produce two tiny lens-shaped subsidiary cells (SCs) laterally to the GMC (Fig. 1D). Development of stomatal complexes is completed by the symmetric GMC division which produces the guard cells (Fig. 1E). Although the majority of protodermal cells divide transversely to the leaf axis, the SMCs and GMCs divide in a plane parallel to the leaf axis (review by Galatis and Apostolakos, 2004). This reorientation constitutes a major shift in poaceous protodermal differentiation.
SMC polarization and asymmetric division have been exten-sively discussed in a series of reviews (Pickett-Heaps et al.,
1999; Cleary, 2000; Gallagher and Smith, 2000; Galatis and Apostolakos, 2004; De Smet and Beeckman, 2011; Facette and Smith, 2012; Pilliteri and Torii, 2012; Li, 2013; Facette et al., 2015; Shao and Dong, 2016). The present article attempts to offer an up to date overview of the mechanisms underlying these phenomena, combining older data with recent findings which reveal new players involved in polarization and asym-metric division of Zea mays SMCs. It focuses on the interplay between GMCs and SMCs, and the unique sequence of struc-tural and molecular polarization events occurring in both cell types. Of particular interest are recent findings showing that this sequence seems to commence with local cell wall differen-tiation (Giannoutsou et al., 2016).
GMC SHAPING
The generation and differentiation of stomatal complexes in grasses proceeds in a linear sequence from the base towards the tip of the leaf. The newly formed GMCs of Z. mays
Dow
nloaded from https://academ
ic.oup.com/aob/article/122/5/679/4810445 by guest on 14 Septem
ber 2022

Apostolakos et al. — Subsidiary cell formation in maize stomatal complexes680
appear in a region close to the leaf base and display a rect-angular shape in surface view with their long axis oriented transversely to the stomatal row axis (Fig. 1A). They grow parallel to the stomatal row axis, tending to obtain a more or less square form (Fig. 1B, C). At this stage, the GMCs induce their lateral epidermal cells to function as SMCs. In general, GMCs of Z. mays with a width (dimension perpen-dicular to the stomatal row axis)/length (dimension parallel to the stomatal row axis) ratio of 0.8–1.4 (advanced GMCs) are capable of emitting an inductive stimulus towards the adjacent SMCs (Apostolakos et al., 2008). Quantification of these observations showed that 70 % of SMCs lying lat-erally to these GMCs have divided asymmetrically (Livanos et al., 2015). The GMCs displaying a width/length ratio >1.4 are located closer to the basal meristematic leaf region and are termed young GMCs (Apostolakos et al., 2008). Only 17 % of SMCs adjoining those GMCs divide asymmetric-ally (Livanos et al., 2015). During GMC morphogenesis, the length increases approx. 125 % (from 4.27 ± 0.1 μm in young GMCs to 9.62 ± 0.1 μm in advanced GMCs), while their width decreases approx. 34.60 % (from 13.46 ± 0.1 μm in young GMCs to 8.81 ± 0.1 μm in advanced GMCs; Giannoutsou et al., 2011). This decrease is the outcome of a microtubule (MT)-dependent morphogenetic mechanism that creates a median constriction in GMCs (Galatis, 1982; Galatis and Apostolakos, 2004). Consequently, the adjacent SMCs protrude locally towards the advanced GMCs (Galatis, 1982; Panteris et al., 2007; Apostolakos et al., 2008; Giannoutsou et al., 2011; Humphries et al., 2011). The SMC protrusion keeps pace with the local expansion and deformation of the common cell wall between GMCs and SMCs (Giannoutsou et al., 2016). The protruding region of the SMC is defined as the polar SMC site. These local cell rearrangements are co-ordinated with the highly controlled growth of the inter-vening cells of the stomatal row (Giannoutsou et al., 2011). They remind us of the local cell growth accompanying the extensive local rearrangements occurring among developing lobed pavement cells, which have been elegantly described by Armour et al., (2015).
THE NATURE OF THE INDUCTIVE GMC STIMULUS
Since the middle of the previous century, it has been suspected that the inductive stimulus establishing SMC polarization in grasses might be of hormonal nature (Bünning, 1956, 1957; Stebbins and Shah, 1960; Stebbins and Jain, 1960; Pickett-Heaps and Northcote, 1966). The chemical nature of the induct-ive stimulus is favoured by data derived from caffeine treatment on developing stomatal complexes of Triticum vulgare (Pickett-Heaps, 1969) and Z. mays (Apostolakos and Galatis, 1987). Caffeine interferes with the coalescence and/or consolidation of cell plate vesicles, resulting in the formation of incom-plete daughter cell walls during cell division (Bonsignore and Hepler, 1985). Consequently, a number of affected SCs were separated from the adjacent typical epidermal cells by curved cell walls exhibiting small or large perforations. These epider-mal cells normally do not divide; however, being separated from SCs by incomplete cell walls, they were induced to enter a new asymmetric division. This unexpected asymmetric div-ision of the distally lying nucleus supports the existence of a stimulus, which is chemical in nature, persisting in the GMCs and transported through the SC cell wall gaps into adjacent epi-dermal cells (Apostolakos and Galatis, 1987).
Recent data suggest that auxin (indole-3-acetic acid) operates as an inducer of polarization/asymmetric division in SMCs of Z. mays (Livanos et al., 2015, 2016). In particular, it has been found that in the Z. mays protoderm auxin accumulates in the contact regions of GMCs–SMCs in a time-dependent manner (Livanos et al., 2016). Thus, GMCs in this plant seem to func-tion as local auxin sources similarly to GMCs of the dicoty-ledonous Arabidopsis thaliana (Le et al., 2014; Zhang et al., 2014). Moreover, exogenous application of indole-3-acetic acid or its synthetic analogue 1-naphthaleneacetic acid promotes SMC polarization and asymmetric division, and triggers SC for-mation in protodermal areas closer to the leaf meristem (Livanos et al., 2015), whereas treatment with the specific auxin transport inhibitors 2,3,5-triiodobenzoic acid (TIBA) and 1-naphthoxy-acetic acid inhibits both the establishment of polarity and the asymmetric division in SMCs (Livanos et al., 2015).
Guard cell mother cell
SMC’s nucleus SMC’s nucleus (polar position) Actin patch MT-PPB Monopolar prophase MT-spindle Metaphase MT-spindle MT-phragmoplast/cell plate
Guard cells Intervening cell Subsidiary cell mother cell
SMC polar site
A B C D E
Subsidiary cell Inductive stimuli PIN1 carriers
Fig. 1. Diagram illustrating the development of Z. mays stomatal complexes.
Dow
nloaded from https://academ
ic.oup.com/aob/article/122/5/679/4810445 by guest on 14 Septem
ber 2022

Apostolakos et al. — Subsidiary cell formation in maize stomatal complexes 681
STRUCTURAL SMC POLARIZATION
Local cell wall differentiation at the polar SMC site
Interestingly, in Z. mays SMCs and GMCs, the matrix cell wall materials are asymmetrically distributed (Giannoutsou et al., 2016). In particular, the common cell wall between GMCs and SMCs was found to be enriched with: (a) highly esterified homogalacturonans (HGAs) carrying the JIM7-HGA epitope; (b) HGAs with a high degree of de-esterification having the JIM5-HGA epitope; (c) non-esterified HGAs displaying the 2F4-HGA epitope; and (d) arabinogalactan proteins (AGPs). Moreover, rhamnogalacturonans (RGAs) containing highly branched arabinans (LM6-RGA epitope) were preferentially deposited at the margins of the above cell wall. These matrix cell wall materials were absent from the rest of the SMC cell walls. Importantly, their accumulation in the common GMC–SMC cell wall commences at an initial GMC morphogenetic stage when the SMC nucleus lies far from the inducing GMC. Thus, the local cell wall matrix differentiation seems to consti-tute the first structural feature of the initiation of SMC polariza-tion detected so far. This distinct cell wall composition persists in SMCs adjacent to advanced GMCs and, after asymmetric div-ision, in the young SCs (Giannoutsou et al., 2016). In a similar direction, in various angiosperm cell types (Bosch and Hepler, 2005; Nguena-Ona et al., 2012; Wolf et al., 2012; Castro et al., 2013; Lamport et al., 2014), algae (Belanger and Quatrano, 2000) and fungi (Lesage and Bussey, 2006), the establishment of polarity is accompanied by cell wall modification(s) at the polar cell site.
Thus, the particular cell wall composition at the SMC–GMC contact site can be correlated to the pattern of its growth (Giannoutsou et al., 2016). Due to its local expansion, the com-mon SMC–GMC cell wall is likely to experience mechanical stress and deformation. The consistent presence of the JIM7-HGA epitope at the polar SMC site could facilitate its local expansion, since the esterified HGAs form cell wall matrix gels of low viscosity, which are required for cell wall expansion (Verhertbruggen and Knox, 2006; Wolf et al., 2012; Bidhendi and Geitmann, 2016).
Although molecules of highly de-esterified or non-esterified HGAs tightly interconnected with Ca2+ bridges form cell wall matrices of high viscosity offering stability to the cell wall (Verhertbruggen and Knox, 2006), in the cell wall regions where pectin molecules strongly linked with Ca2+ dominate, enzymes breaking down HGAs can be activated. Consequently, HGA turnover and remodelling of cell wall molecules accom-panied by cell wall expansion occurs (Verhertbruggen and Knox, 2006; Pelletier et al., 2010; Wolf and Greiner, 2012; Bidhendi and Geitmann, 2016). Therefore, the constant pres-ence of JIM5-HGA and 2F4-HGA epitopes at the SMC–GMC contact site may contribute to their expansion and may also fa-vour cell wall deformation at the polar SMC site (Giannoutsou et al., 2016). Data derived from atomic force microscopy ana-lysis in certain Arabidopsis thaliana cell types revealed that de-methylesterification of HGAs increases cell wall elasticity (Peaucelle et al., 2011, 2015; Braybrook and Peaucelle, 2013).
The AGPs residing on the plasmalemma interlink the cell wall with the cell interior and appear to participate in cell wall expansion, functioning as cell wall plasticizers (Knox, 2006;
Lamport et al., 2006, 2014; Nguema-Ona et al., 2012). In this direction, AGPs residing at the SMC–GMC contact site may be involved in cell wall expansion and deformation at the polar SMC site. Although RGAs generally favour cell wall soften-ing (Verhertbruggen and Knox, 2006; Bidhendi and Geitmann, 2016), the arabinans of their lateral branches are linked to cellulose microfibrils (Zykwinska et al., 2005) contributing to cross-linking of cell wall molecules (Verhertbruggen and Knox, 2006; Bidhendi and Geitmann, 2016). In this context, the presence of the LM6-RGA epitope in the marginal regions of the cell wall of the polar SMC site may promote the inter-linking of the cell wall molecules, providing resistance to de-formation (Giannoutsou et al., 2016). Finally, the JIM5-HGAs and the 2F4-HGAs as well as the AGPs might also participate in the transduction mechanism of the GMC stimulus towards the SMC (see ‘Plausible transduction mechanisms of the GMC stimulus toward the SMC’).
Polar migration and anchoring of the SMC nucleus
The migration of the SMC nucleus in the immediate prox-imity of the inducing GMC (Fig. 1C) is a process sine qua non for SMC asymmetric division (Stebbins and Shah, 1960) and is mediated by the cytoplasmic actin filament (AF) system (Kennard and Cleary, 1997; reviewed by Pickett-Heaps et al., 1999; Cleary, 2000; Smith, 2001; Galatis and Apostolakos, 2004). In fact, a detailed drug test study concerning pre-mitotic nuclear migration in Tradescantia virginiana pointed to the in-volvement of AFs rather than MTs in repositioning of SMC nu-clei towards the GMC after displacement (Kennard and Cleary, 1997). However, in maize SMCs, it has been described that the anchoring of the nucleus at its polar position, which is a prerequisite for the successful completion of the asymmetric division, is achieved by a unique MT ‘cage’ (Panteris et al., 2006). The latter ensures that the nucleus and subsequently the one pole of the future mitotic spindle will be localized ‘almost in contact’ with a median region of the common SMC–GMC cell wall and that the mitotic spindle axis will keep an align-ment more or less perpendicular to the above cell wall (Fig. 1C, D). When these protoplasmic presuppositions are fulfilled, the cortical zone defined by the pre-prophase band (PPB) interacts with the growing edges of the cell plate, causing its curved growth that results in the formation of a tiny lens-like SC lat-erally to the inducing GMC (Fig. 1D; Galatis and Apostolakos, 2004).
The displacement of the SMC nucleus results in variable cell plate arrangement patterns. As a result, atypical SCs, usually triangular in surface view, are generated (Galatis et al., 1983, 1984a, b). Some of these SCs seem to display a unique shape, different from the typical one: although the shape is lens-like at the external planes, it is significantly altered in median planes, assuming a triangular form (Galatis et al., 1984a). Among fac-tors preventing the proper SMC nuclear positioning are the small SMC size, an eccentric position of the GMC in relation to the SMC and a synchronous induction of the same SMC by two stimuli coming from different directions (Galatis et al., 1983, 1984a).
Aberrant SCs are also formed in discordia (Gallagher and Smith, 1999; Wright et al., 2009), brick (Gallagher and Smith,
Dow
nloaded from https://academ
ic.oup.com/aob/article/122/5/679/4810445 by guest on 14 Septem
ber 2022

Apostolakos et al. — Subsidiary cell formation in maize stomatal complexes682
2000) and panglosses (Gallagher and Smith, 2000; Cartwright et al., 2009; Humphries et al., 2011; Zhang et al., 2012) Z. mays mutants. At least for discordia mutants, the formation of these atypical SCs has been attributed to the defects in PPB organ-ization (Gallagher and Smith, 1999; Wright et al., 2009). For brk1 and pan1 mutants, among others, it has been proposed that SMC polarization is disrupted and consequently the SMC nu-cleus fails to align with the GMC (Gallagher and Smith, 2000). Nevertheless, the failure of the nucleus to be placed properly close to the inducing GMC might occur due to the smaller size of the SMCs of the above Z. mays mutants, compared with the SMCs of the wild type. Of course, this is only a speculation based on morphological observation, and it should be experi-mentally examined further in order to be confirmed. It has al-ready been described that in two Triticum species the inability of the nucleus to assume its proper polar position due to space limitation subsequently leads to a very inclined arrangement of the mitotic spindle axis in relation to the common SMC–GMC cell wall, resulting in the formation of atypical SCs (Galatis et al., 1984a).
Polar organization of the cytoskeleton
Actin filaments. The cortical AFs in Z. mays SMCs are aggre-gated in a well-defined AF patch lining the whole surface of the plasmalemma at the polar SMC site (Gallacher and Smith, 1999, 2000; Panteris et al., 2006, 2007; Facette and Smith, 2012; Facette et al., 2015; see Fig. 1C). The AF patch was ini-tially found in SMCs of the grass Secale cereale (Cho and Wick, 1990, 1991) and later in the SMCs of the monocotyledonon T. virginiana (Cleary, 1995; Cleary and Mathesius, 1996). In Z. mays, AF patch formation commences in interphase SMCs adjoining advanced GMCs, reaches maximum organization during mitosis and is bequeathed to the young SCs (Panteris et al., 2006, 2007; see Fig. 1C, D). In prophase SMCs, AFs pos-sibly originating from the AF patch region display a cup-like conformation on the nuclear surface facing the inducing GMC actually forming a prophase AF half-spindle (Panteris et al., 2006; Fig. 2). During mitosis, filaments of the AF cup enter the mitotic spindle (Panteris et al., 2006). A similar rearrangement of cortical AFs has been described in prophase/mitotic SMCs of S. cereale (Cho and Wick, 1990).
The AF patch has been considered as a basic element of SMC polarization (see reviews by Wick, 1991a; Cleary, 2000; Smith, 2001; Galatis and Apostolakos, 2004; Facette and Smith, 2012; Facette et al., 2015), and it serves as a useful marker of po-larity in SMCs (Cartwright et al., 2009; Humphries et al., 2011; Zhang et al., 2012). Cho and Wick (1990, 1991) suggested that it contributes to the positioning of the nucleus at the polar SMC site. Further, it has been proposed that, among other functions, it also facilitates the anchoring of the one pole of the mitotic spindle close to the inducing GMC (Cleary, 1995; Cleary and Mathesius, 1996; reviewed by Wick, 1991a; Cleary, 2000; Smith, 2001; Galatis and Apostolakos, 2004). However, there are data contradicting the above-mentioned view. In particular: (a) treatment of living Z. mays SMCs displaying polarized nu-clei with anti-actin drugs did not affect the position of the nu-cleus (Panteris et al., 2006); (b) exogenous auxin application
to SMCs adjacent to young Z. mays GMCs favoured the place-ment of the nucleus close to GMCs although these SMCs are devoid of an AF patch (Livanos et al., 2015); (c) in Z. mays SMCs adjoining advanced GMCs, an AF patch was assembled even when the nuclear polarization and the subsequent asym-metric division had been experimentally inhibited (Apostolakos et al., 2008; Livanos et al., 2015, 2016), (d) in Triticum tur-gidum SMCs, the nucleus migrates and is anchored close to the inducing GMC, although the majority of SMCs lack AF patches (Panteris et al., 2006, 2007); (e) in the thorough stud-ies of Cartwright et al. (2009), Humphries et al. (2011) and Zhang et al. (2012), in panglosses and rops mutants of Z. mays, quantification of AF patch formation and nuclear polarization showed that, in a population of mutant SMCs with polarized nuclei, the AF patch was delocalized or absent, whereas many SMCs with unpolarized nuclei had a normal AF patch; and (f) in T. virginiana, each GMC is flanked by four SMCs, two lateral and two terminal, which all display polarized nuclei (Cleary, 1995, 2000; Cleary and Mathesius, 1996), although well-organ-ized AF patches appear only in the lateral SMCs (Cleary and Mathesius, 1996).
Thus, the AF patch is likely to play a role irrelevant to posi-tioning of the nucleus in SMCs. It has been proposed that it might be implicated in polarized vesicle trafficking (Humphries et al., 2011), but its function is still unknown (Zhang et al., 2012). A quite reasonable explanation of its purpose is that it might protect the plasmalemma of the SMC polar site from the mechanical stress imposed on it by local SMC growth towards the GMC (Panteris et al., 2007). This hypothesis is supported by the following findings. (1) Although in seedlings supplied with exogenous auxin, SMCs adjacent to both young and advanced GMCs underwent division, AF patches were assembled only in those protruding towards advanced GMCs (Livanos et al., 2015). (2) In T. turgidum (Panteris et al., 2007) and T. virgini-ana (Cleary and Mathesius, 1996), distinct AF patches were detected only in SMCs protruding towards the inducing GMCs. (3) The AF patch is maintained in the young SCs (Fig. 1D, E),
GMC
SMC
Fig. 2. Diagram showing the organization of the MTs (green lines) and the AFs (red lines) in prophase SMCs of Z. mays (from Panteris et al., 2006).
Dow
nloaded from https://academ
ic.oup.com/aob/article/122/5/679/4810445 by guest on 14 Septem
ber 2022

Apostolakos et al. — Subsidiary cell formation in maize stomatal complexes 683
which still protrude towards the GMCs or young guard cells (Panteris et al., 2007). Such a role for the AF patch in SMCs is reminiscent of the fact that in animal cells the mechanically stressed plasma membrane induces the formation of AF arrays to reinforce its stressed regions (Frame and Sarelius, 2000; Ko and McCulloch, 2000). The distinct AF meshworks organized within outgrowing bulges of trichoblasts and at tips of young root hairs of Z. mays seem to display a similar activity (Baluška et al., 2000a).
Microtubules. In SMCs adjoining advanced GMCs, a micro-tubule pre-prophase band (MT-PPB) is assembled at the mar-gins of the site of SMC–GMC contact (Gallagher and Smith, 1999; Panteris et al., 2006, 2007; see Figs 1C and 2). The cor-tical division site defined by the PPB directs or attracts the growing ends of the SMC cell plate to curve and meet the pre-determined parent cell wall regions in a not yet fully under-stood mechanism, thus generating a small lens-shaped SC (see reviews by Wick, 1991b; Mineyuki, 1999; Smith, 2001; Galatis and Apostolakos, 2004; Paciorek and Bergmann, 2010; see Fig. 1D). It should be noted that the cortical SMC cytoplasm area adjacent to the GMC, where AF patch formation occurs, is devoid of MTs since interphase (Panteris et al., 2006).
The SMC MT-PPB reaches its highest degree of organization at prophase. At this stage, an unusual prophase MT half-spindle is assembled at the nuclear hemisphere distal to the inducing GMC (Panteris et al., 2006). Importantly, the plus ends of these MTs are connected to the MT-PPB, forming an MT cage (Figs 1C and 2), which secures the anchoring/stabilization of the nu-cleus and the one mitotic spindle pole almost in contact with the common SMC–GMC cell wall. This is strongly supported by data derived from the treatment of living prophase Z. mays SMCs with anti-MT drugs. In the affected SMCs, the prophase nucleus moves away from the inducing GMC (Panteris et al., 2006). Such an organization of the cytoskeleton in prophase clearly demonstrates the intense SMC polarization and has not been described so far in other plant cell types. SMC polariza-tion establishes conditions that trigger the divergent differenti-ation of the daughter nuclei of the SMC asymmetric division. The SC nucleus remains at a lag phase morphologically iden-tical to telophase, while in contrast the nucleus of the adjacent epidermal cell enters interphase rapidly. In fact, SC differenti-ation is delayed until completion of the symmetric GMC div-ision, so that all the cells of the young stomatal complex enter a synchronized route of differentiation (Galatis et al., 1983).
In angiosperms, the prophase MT spindle is assembled by MTs which are nucleated on the pre-prophase/prophase nuclear surface and assume bipolarity via motors and MT-associated proteins (Masoud et al., 2013; Li et al., 2015). In SMCs, the AF patch seems somehow to prevent the formation of a prophase MT half-spindle at the nuclear hemisphere proximal to the GMC, promoting instead a prophase AF half-spindle assembly (Panteris et al., 2006). This view is supported by observations made in SMCs of brick Z. mays leaves, which lack organized AF patches (Gallagher and Smith, 2000). In these SMCs, the prophase nucleus is globally surrounded by MTs, while the cor-tical cytoplasm adjacent to the common SMC–GMC cell wall is lined by MTs (Panteris et al., 2006).
Notably, in SMCs in which the asymmetric division was experimentally inhibited, the MT-PPB assembly was not affected (Apostolakos et al., 2008; Livanos et al., 2015, 2016).
This observation allows the hypothesis that the division plane of asymmetric SMC division is determined by a mechanism different from that inducing the nuclear division (see ‘Plausible transduction mechanisms of the GMC stimulus towards the SMC’).
Polar endoplasmic reticulum organization
Endoplasmic reticulum (ER) immunolabelling studies using antibodies recognizing residential luminar ER proteins carry-ing the C-terminal HDEL sequence, BiP proteins and calreti-culin revealed the presence of significant ER accumulations in the cortical cytoplasm of the polarized site of Z. mays SMCs protruding towards advanced GMCs. They tend to form an ER patch co-localizing with the AF patch, both extending along the whole surface of the common SMC–GMC cell wall (Giannoutsou et al., 2011). During pre-prophase/prophase, the ER patch is disorganized, whereas ER elements are gathered in the MT-PPB region forming an ER-PPB and at the region of the prophase MT half-spindle forming an ‘ER half prophase spindle’ (Giannoutsou et al., 2011, 2012). However, other authors following anti-BiP and anti-calreticulin application did not consistently trace ER patches in Z. mays SMCs (Cartwright et al., 2009). Further, in transgenic lines of Z. mays expressing the ER marker YFP–PDI (yellow fluorescent protein–protein di-sulphide isomerase), signal was only occasionally detected at the polar SMC site (Facette et al., 2015). These results cast doubt on the credibility of this ER patch as a real cytoplasmic entity. The following data of Giannoutsou et al., (2011, 2012) counter this doubt. (a) The ER patch followed a definite pattern of appear-ance. It was absent from the SMCs adjoining young GMCs, it appeared in SMCs adjacent to advanced GMCs, it disintegrated in mitotic and cytokinetic SMCs and reappeared in young SCs. (b) The T. turgidum SMCs lack an ER patch, except for those protruding towards the inducing GMCs. (c) The presence of an ER patch has been confirmed in specimens which have under-gone chemical fixation, as well as cryofixation. Thus, the possi-bility of this ER patch being an artefact might be reconsidered.
The co-localization and the close similarity in organization between an AF patch and an ER patch, the absence of an ER patch in SMCs treated with drugs interfering with AFs, the ab-sence of an ER patch from the majority of SMCs of T. turgidum which lack an organized AF patch and their presence in a few of them displaying an AF patch strongly indicate that ER patch organization seems to be controlled by the AF cytoskeleton (Giannoutsou et al., 2011). Additional evidence supporting this hypothesis is that the absence of an AF patch in 1-butanol-treated Z. mays SMCs (Apostolakos et al., 2008) is accompa-nied by inhibition of ER patch formation (Giannoutsou et al., 2011). In contrast, in 2-butanol- and neomycin-treated SMCs, exhibiting AF patches (Apostolakos et al., 2008), ER patch for-mation was not affected (Giannoutsou et al., 2011).
Notably, in pre-prophase/prophase SMCs, the AF patch reaches the maximum of its organization (Panteris et al., 2006, 2007), while the ER patch is disorganized and an ER-PPB and a prophase ER half-spindle are formed (Giannoutsou et al., 2011, 2012). At this cell cycle stage, ER organization is prob-ably controlled by the MT cytoskeleton and not by the AFs, since the pre-prophase/prophase ER organization is seriously
Dow
nloaded from https://academ
ic.oup.com/aob/article/122/5/679/4810445 by guest on 14 Septem
ber 2022

Apostolakos et al. — Subsidiary cell formation in maize stomatal complexes684
affected in MT-deprived oryzalin- and colchicine-treated SMCs (Giannoutsou et al., 2011). Consistent with this, there are con-vincing data showing that ER organization in interphase plant cells is controlled by AF-related myosins, while in dividing cells it is controlled by MT-related molecular motors (refer-ences cited in Giannoutsou et al., 2011, 2012).
In addition, tubulin acetylation seems to be a critical factor for the MT-mediated organization of the ER. When MTs con-sist of or contain acetylated tubulin, ER mitotic and cytokinetic arrays identical to those of MTs are assembled (Zachariadis et al., 2001, 2003; Giannoutsou et al., 2012). The acetylated MTs are not only stable but are also capable of interacting with a variety of molecules, among which are proteins involved in ER organization (Perdiz et al., 2011).
In many polarized cell types, cortical ER aggregations are gathered in the polar region and actively contribute to cell polar-ization by participating in localized protein synthesis, favouring local plasmalemma differentiation or establishing Ca2+ gradients (English et al., 2009; Žarský et al., 2009; Griffing et al., 2017). However, experimental evidence accumulated so far suggests that in SMCs the ER patch does not seem to play a primary role in polarization/asymmetric division (Giannoutsou et al., 2011) because: (a) an ER patch is not generally present in the polarized SMCs of T. turgidum; (b) ER patches were organized in the neomycin-treated SMCs of Z. mays in which the asym-metric division was inhibited (Apostolakos et al., 2008); and (c) ER patches were also found in SCs of Z. mays which did not show any structural polarity. The ER patch organized in SMCs is probably functionally related to mechanisms of cell wall expan-sion and deformation at their polar region (Giannoutsou et al., 2011). According to the above authors, AF and ER accumula-tions demarcate ‘cortical cytoplasmic domains’ activated below the locally expanding Z. mays stomatal cell wall regions, prob-ably via a mechanosensing mechanism, triggered by the locally stressed plasmalemma–cell wall continuum. The ‘cortical cyto-plasmic domains’ are comparable with the ‘cell periphery com-plexes (cages)’ described by Baluška et al., (2000a, b, 2001) that determine cell growth polarity and morphogenesis.
As has been mentioned previously, the common SMC–GMC cell wall is enriched with particular matrix materials, in which the AGPs are included. The ER patch may be implicated in the for-mation of the AGPs at the polar SMC site, since they are synthe-sized in the endomembrane system (Nguema-Ona et al., 2012). This localized ER accumulation which is closely associated with the plasmalemma might serve as a source of lipid molecules for the expanding plasmalemma (Giannoutsou et al., 2011; see also Pichler et al., 2001). Moreover, considering that the plasma-lemma at the polar SMC site recruits numerous proteins (see the next section), it could be proposed that the ER patch might con-tribute to the local plasmalemma differentiation in this region. ER participation in the local differentiation of the plasmalemma is a generally accepted view (Griffing et al., 2017).
MOLECULAR PLAYERS INVOLVED IN SMC POLARIZATION AND ASYMMETRIC CELL DIVISION
Auxin transporters
In Z. mays, auxin is polarly transported along the leaf axis (Tsiantis et al., 1999; Nelissen et al., 2012). PINFORMED
(PIN) efflux carriers and the AUXIN INSENSITIVE 1 influx carriers are among the protein molecules facilitating auxin transport (Nick, 2010; Forestan and Varotto, 2012; Balzan et al., 2014). PIN1 efflux carriers exhibit a polar distribution in Z. mays leaf protoderm (Livanos et al., 2015). In regions close to the basal leaf meristem, where young GMCs dominate, PIN1 carriers were predominantly localized in the plasmalemma lin-ing the transverse, i.e. perpendicular to the leaf axis, cell walls in all protodermal cell types, including the GMCs (Fig. 1A). In contrast, PIN1 carriers were scarce in the plasmalemma adja-cent to the lateral cell walls, parallel to the leaf axis.
At more differentiated protodermal regions, a marked shift in PIN1 carrier distribution occurred in advanced GMCs, as the plasmalemma adjacent to their lateral cell walls, in common with the SMCs, became selectively enriched in PIN1 carriers (Fig. 1B). At the stage at which the GMCs seemed to function as local auxin sources, PIN1 accumulation was more prominent along the lateral GMC cell walls than along the transverse walls (Livanos et al., 2015; see also Fig. 1C). PIN1 redistribution in the advanced GMCs probably allows preferential auxin trans-port in the apoplast between GMCs and SMCs, which accu-mulates auxin at the polar SMC site (Livanos et al., 2016). Considering that the inhibition of SMC polarization and the subsequent asymmetric divisions induced by TIBA is accompa-nied by disturbance of PIN1 carrier distribution in the affected advanced GMCs, it may be suggested that redistribution of PIN1 carriers should represent a crucial initial step in SMC po-larization (Livanos et al., 2015). TIBA disrupts the cycling and transport of PIN1 carriers and affects auxin transport by inter-fering with membrane trafficking processes (Geldner et al., 2001). Furthermore, treatment with the auxin influx inhibitor 1-naphthoxyacetic acid inhibited the establishment of polar-ization/asymmetric division in Z. mays SMCs (Livanos et al., 2015). 1-Naphthoxyacetic acid interferes with the cellular auxin uptake mediated by the AUXIN INSENSITIVE 1 influx auxin carriers (Tsuda et al., 2011). Thus, it can be predicted that the polar site of SMCs might also be enriched with the above auxin influx carriers.
Taken together, the above further support the view that auxin functions as an inductive stimulus emitted by the GMCs and transported, via the asymmetrically distributed auxin carriers, towards the SMCs.
Reactive oxygen species
The functional interplay between reactive oxygen species (ROS) and auxin is a common prerequisite for several develop-mental processes and mediates cell polarity establishment (Joo et al., 2001, 2005; Foreman et al., 2003; Pasternak et al., 2005; Coelho et al., 2008; Mittler et al., 2011; Sandalio et al., 2016). Recent data revealed that ROS are critical partners of auxin dur-ing establishment of polarity and manifestation of asymmetric division in Z. mays SMCs (Livanos et al., 2016).
In particular, application of ROS fluorescent probes revealed that in SMCs flanking advanced GMCs, ROS were mainly local-ized at the polarized SMC site including the apoplast between the SMC and the inducing GMC. ROS enrichment in the polar-ized SMC site was also confirmed by transmission electron mi-croscopy after staining with CeCl3. Moreover, H2O2 promoted SC formation at protodermal regions close to the leaf meristem
Dow
nloaded from https://academ
ic.oup.com/aob/article/122/5/679/4810445 by guest on 14 Septem
ber 2022

Apostolakos et al. — Subsidiary cell formation in maize stomatal complexes 685
and induced polarization and asymmetric division in a number of intervening cells of the stomatal row, thus leading to the for-mation of supernumerary SCs, i.e. stomatal complexes with three SCs. Importantly, H2O2 also interfered with the pattern of auxin distribution in Z. mays protoderm as significant auxin quantities were detected at the polar site of SMCs adjacent to young GMCs and at the cortical cytoplasm of intervening cells of the stomatal rows adjoining the GMCs. Modulation of ROS homeostasis in favour of oxidants or antioxidants resulted in in-hibition of polarization/asymmetric SMC division and averted the promotional effect of the exogenously supplied auxin on SC formation. In addition, H2O2 alleviated the inhibitory effects of TIBA on SC generation. Considering these data, it can be deduced that auxin and ROS co-operate in SMC polarization/asymmetric division (Livanos et al., 2016).
Rho-like GTPases of plants
Rho-like GTPases of plants (ROPs) are implicated in the es-tablishment of polarity and the promotion of polarized growth in various cell types (Nibau et al., 2006; Yang and Fu, 2007; Yang, 2008). The membrane-associated type I ROP2 and ROP9 are preferentially localized at the polar site of Z. mays SMCs adjoining advanced GMCs, and it has been proposed that both play a key role in SMC polarization and asymmetric division (Humphries et al., 2011). In many SMCs of rop mutants, AF patches were absent or disturbed, the polar movement of the nucleus was inhibited, while the alignment of the plane of cell division was disturbed, resulting in formation of atypical SCs (Humphries et al., 2011). The main function of ROPs at the polar SMC site seems to be related to AF patch organization via activation of proteins involved in F-actin stimulation, such as RIC4, components of the ARP2/3 complex and/or formins (Humphries et al., 2011; Facette et al., 2015). Furthermore, the data of Humphries et al. (2011) imply that ROPs interfere with SMC nuclear polarization. However, a closer examination of the published protodermal images of rop mutants favours the hypothesis that the size of SMCs is significantly smaller than that of wild-type plants. Although, to date, there are no quanti-fication data supporting this hypothesis, this may provide an al-ternative explanation for the inability of the nucleus to become polarized (see also ‘Polar migration and anchoring of the SMC nucleus’).
Leucine-rich repeat-receptor-like proteins
The leucine-rich repeat receptor-like kinases (LRR-RLKs) regulate many aspects of plant development and display an extracellular and an intracellular domain. The interaction of extrinsic ligand(s) with their extracellular domains regulates the activity of the intracellular domains, triggering downstream sig-nalling cascades (Llompart et al., 2003; Boudeau et al., 2006; Rajakulendran and Sicheri, 2010). In Arabidopsis, secreted pep-tides recognized by cell surface receptors with LRR domains have been implicated in stomatal patterning (Lee et al., 2012). The first gene identified to be implicated in stomatal pattern-ing was TOO MANY MOUTHS (TMM) which encodes an LRR-RLK in Arabidopsis (Nadeau and Sack, 2002). Subsequently,
three ERECTA family LRR-RLKs were shown to act dur-ing stomatal stem cell fate specification and differentiation (Shpak et al., 2005). In Z. mays, the preferential localization of PANGLOSS 1 (PAN1) and PANGLOSS 2 (PAN2) LRR-RLKs at the polar site of SMCs has been considered as an essential event of SMC polarization (Cartwright et al., 2009; Zhang et al., 2012). PAN2 at the SMC polar site regulates PAN1 accu-mulation at the same site (Zhang et al., 2012). PAN1, in turn, is involved in the recruitment of ROPs at the polar SMC site (Humphries et al., 2011), suggesting that ROPs are implicated in F-actin dynamics, thus interfering with the polar positioning of the nucleus (Humphries et al., 2011; see reviews by Facette and Smith, 2012; Facette et al., 2015). According to this view, PAN1 and PAN2 determinatively participate in the transduction of the GMC stimulus establishing SMC polarity and the mani-festation of SMC asymmetric division. Ιn pan Z. mays mutants, many SMCs lack AF patches and the nucleus is incapable of becoming polarized, while the orientation of the cell division plane is disturbed (Gallagher and Smith, 2000; Cartwright et al., 2009; Humphries et al., 2011; Zhang et al., 2012). However, the defects in nuclear polarization and AF patch formation are con-sidered as largely independent effects of pan mutations on SMC polarity (Cartwright et al., 2009). Thus, as already mentioned, the AF patch does not seem to contribute to polar SMC nuclear positioning but is related to SMC bulging towards the inducing GMC (see ‘Polar organization of the cytoskeleton’). On the other hand, the misalignment of the division plane of SMCs in pan mutants of Z. mays may be attributed to the relatively small SMC size, although this is just an unproven hypothesis. Furthermore, according to more recent findings, PAN receptors may play more and/or different roles apart from participating in the trans-duction of the GMC stimulus. According to Sutimantanapi et al. (2014), PAN1 is present in cell plates, although this observa-tion cannot be readily interpreted. In addition, it appears that PANs are strongly correlated with F-actin aggregations, essen-tial for stomatal complex development (Panteris et al., 2013; Sutimantanapi et al., 2014). In particular, similarly to the AF patch, which is bequeathed from SMCs to newly formed SCs, PAN1 was also found on the plasmalemma lining the common cell wall between the SC and GMC and on that between the SC and young guard cells (Sutimantanapi et al., 2014; see also Fig. 3A). Moreover, in differentiating SCs, PAN1 was localized in the plasmalemma beneath the junction of the cell wall laid down during the asymmetric SMC division with the parent cell walls (Sutimantanapi et al., 2014; see also Fig. 3B, C). In the above regions, particular cortical AF aggregations are organized (Panteris et al., 2007; Sutimantanapi et al., 2014). These data favour the view that PAN1 is somehow involved in the organiza-tion of cortical AF arrays.
PAN2, in addition to the polar SMC site, was localized along the plasmalemma of young and differentiating SCs as well as of the intervening cells of the stomatal rows. In particular, PAN2 is highly accumulated in the plasmalemma lining the SC cell wall regions shared with the intervening cells of the stomatal row (Sutimantanapi et al., 2014). These cell wall regions are lined by distinct AF aggregations (Panteris et al., 2007). Therefore, apart from any involvement in SMC polarization and asymmetric division, PAN1 and PAN2, via their influence on AF polymer-ization, may function in all aspects of stomatal morphogenesis, mediating polarized membrane trafficking (Sutimantanapi et al.,
Dow
nloaded from https://academ
ic.oup.com/aob/article/122/5/679/4810445 by guest on 14 Septem
ber 2022

Apostolakos et al. — Subsidiary cell formation in maize stomatal complexes686
2014). Last, but not least, the localization of PAN1 in Z. mays seedlings treated with the phospholipase modulators 1-butanol or neomycin does not support a crucial role for PAN1 in the mech-anism driving SMC induction for cell division and/or polariza-tion. Although such treatments inhibited the polar positioning of the nucleus and the subsequent asymmetric division of SMCs (Apostolakos et al., 2008), PAN1 recruitment to the polar SMC site was not affected (Fig. 3D–F).
SCAR–WAVE regulatory complex
The products of three Brick genes have been implicated in SC formation in Z. mays. In many SMCs of brk mutants, the polarization and cell division plane orientation were disturbed, while in others the asymmetric SMC division was inhibited (Gallagher and Smith, 2000; Frank and Smith, 2002; Frank et al., 2003). Brk1 encodes a small subunit of the SCAR/WAVE regulatory complex (SWRC) which is homologous to
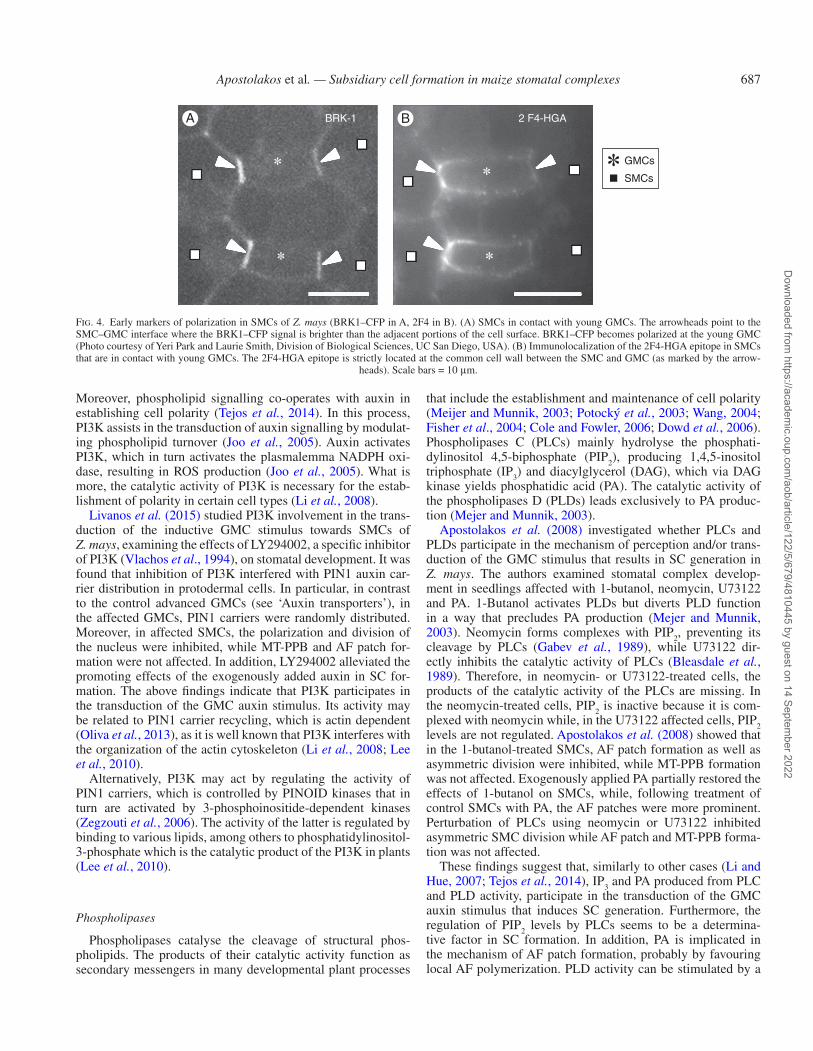
mammalian HSPC300 (Frank and Smith, 2002; Frank et al., 2003). The latter activates the ARP2/3 complex, hence control-ling AF polymerization (Miki et al., 1998; Machesky et al., 1999). In rice, homologues of SWRCs contribute to AF rear-rangements during morphogenesis of leaf epidermal cells (Zhou et al., 2016). In Z. mays SMCs, the SWRC displays a polar distribution, initially localized at the contact region of SMCs with young GMCs, persisting in the polar SMC site of polarized SMCs and bequeathed to young SCs (Facette et al., 2015; see also Fig. 4A). According to Facette et al. (2015), the SWRC regulates PAN2 recruitment at the polar SMC site, and thus is considered to be one of the first markers of establish-ment of polarity in SMCs of Z. mays.
Phosphatidylinositol-3-kinase (PI3K)
As has been previously described, ROS and auxin syner-gistically trigger the asymmetric division in Z. mays SMCs.
Control Control Control
GMCsSCs
Guard cellsSMCs
But-1 But-1 Neomycin
A B C
D E F
Fig. 3. Immunolocalization of PAN1 in developing Z. mays stomatal complexes of seedlings grown under physiological conditions (A–C) and in others treated with phospholipase modulators (D–F). The nuclei are shown in red. Immunodetection of PAN1 was performed following the procedure described in Cartwright et al. (2009). The anti-PAN1 antibody was kindly provided by Professor L. G. Smith (University of California San Diego, USA). (A) GMCs on both sides of which SCs are or will be formed. The arrowheads point to local PAN1 aggregations. (B and C) Young stomatal complexes in successive developmental stages. The arrowheads indicate local PAN1 aggregations in differentiating SCs. (D and E) SMCs treated with 1-butanol (But-1). The arrowheads point to PAN1 aggregation in the SMC polar site. Note that in all the SMCs the nucleus has not occupied its polar position. Treatment: 20 mm 1-butanol for 24 h. (F) Part of a stomatal row of
a seedling treated with neomycin. The arrowheads point to local PAN1 aggregations. Treatment: 100 mm neomycin for 24 h. Scale bars = 10 μm.
Dow
nloaded from https://academ
ic.oup.com/aob/article/122/5/679/4810445 by guest on 14 Septem
ber 2022
Apostolakos et al. — Subsidiary cell formation in maize stomatal complexes 687
Moreover, phospholipid signalling co-operates with auxin in establishing cell polarity (Tejos et al., 2014). In this process, PI3K assists in the transduction of auxin signalling by modulat-ing phospholipid turnover (Joo et al., 2005). Auxin activates PI3K, which in turn activates the plasmalemma NADPH oxi-dase, resulting in ROS production (Joo et al., 2005). What is more, the catalytic activity of PI3K is necessary for the estab-lishment of polarity in certain cell types (Li et al., 2008).
Livanos et al. (2015) studied PI3K involvement in the trans-duction of the inductive GMC stimulus towards SMCs of Z. mays, examining the effects of LY294002, a specific inhibitor of PI3K (Vlachos et al., 1994), on stomatal development. It was found that inhibition of PI3K interfered with PIN1 auxin car-rier distribution in protodermal cells. In particular, in contrast to the control advanced GMCs (see ‘Auxin transporters’), in the affected GMCs, PIN1 carriers were randomly distributed. Moreover, in affected SMCs, the polarization and division of the nucleus were inhibited, while MT-PPB and AF patch for-mation were not affected. In addition, LY294002 alleviated the promoting effects of the exogenously added auxin in SC for-mation. The above findings indicate that PI3K participates in the transduction of the GMC auxin stimulus. Its activity may be related to PIN1 carrier recycling, which is actin dependent (Oliva et al., 2013), as it is well known that PI3K interferes with the organization of the actin cytoskeleton (Li et al., 2008; Lee et al., 2010).
Alternatively, PI3K may act by regulating the activity of PIN1 carriers, which is controlled by PINOID kinases that in turn are activated by 3-phosphoinositide-dependent kinases (Zegzouti et al., 2006). The activity of the latter is regulated by binding to various lipids, among others to phosphatidylinositol-3-phosphate which is the catalytic product of the PI3K in plants (Lee et al., 2010).
Phospholipases
Phospholipases catalyse the cleavage of structural phos-pholipids. The products of their catalytic activity function as secondary messengers in many developmental plant processes
that include the establishment and maintenance of cell polarity (Meijer and Munnik, 2003; Potocký et al., 2003; Wang, 2004; Fisher et al., 2004; Cole and Fowler, 2006; Dowd et al., 2006). Phospholipases C (PLCs) mainly hydrolyse the phosphati-dylinositol 4,5-biphosphate (PIP2), producing 1,4,5-inositol triphosphate (IP3) and diacylglycerol (DAG), which via DAG kinase yields phosphatidic acid (PA). The catalytic activity of the phospholipases D (PLDs) leads exclusively to PA produc-tion (Mejer and Munnik, 2003).
Apostolakos et al. (2008) investigated whether PLCs and PLDs participate in the mechanism of perception and/or trans-duction of the GMC stimulus that results in SC generation in Z. mays. The authors examined stomatal complex develop-ment in seedlings affected with 1-butanol, neomycin, U73122 and PA. 1-Butanol activates PLDs but diverts PLD function in a way that precludes PA production (Mejer and Munnik, 2003). Neomycin forms complexes with PIP2, preventing its cleavage by PLCs (Gabev et al., 1989), while U73122 dir-ectly inhibits the catalytic activity of PLCs (Bleasdale et al., 1989). Therefore, in neomycin- or U73122-treated cells, the products of the catalytic activity of the PLCs are missing. In the neomycin-treated cells, PIP2 is inactive because it is com-plexed with neomycin while, in the U73122 affected cells, PIP2 levels are not regulated. Apostolakos et al. (2008) showed that in the 1-butanol-treated SMCs, AF patch formation as well as asymmetric division were inhibited, while MT-PPB formation was not affected. Exogenously applied PA partially restored the effects of 1-butanol on SMCs, while, following treatment of control SMCs with PA, the AF patches were more prominent. Perturbation of PLCs using neomycin or U73122 inhibited asymmetric SMC division while AF patch and MT-PPB forma-tion was not affected.
These findings suggest that, similarly to other cases (Li and Hue, 2007; Tejos et al., 2014), IP3 and PA produced from PLC and PLD activity, participate in the transduction of the GMC auxin stimulus that induces SC generation. Furthermore, the regulation of PIP2 levels by PLCs seems to be a determina-tive factor in SC formation. In addition, PA is implicated in the mechanism of AF patch formation, probably by favouring local AF polymerization. PLD activity can be stimulated by a
BRK-1 2 F4-HGAA B
GMCs
SMCs
Fig. 4. Early markers of polarization in SMCs of Z. mays (BRK1–CFP in A, 2F4 in B). (A) SMCs in contact with young GMCs. The arrowheads point to the SMC–GMC interface where the BRK1–CFP signal is brighter than the adjacent portions of the cell surface. BRK1–CFP becomes polarized at the young GMC (Photo courtesy of Yeri Park and Laurie Smith, Division of Biological Sciences, UC San Diego, USA). (B) Immunolocalization of the 2F4-HGA epitope in SMCs that are in contact with young GMCs. The 2F4-HGA epitope is strictly located at the common cell wall between the SMC and GMC (as marked by the arrow-
heads). Scale bars = 10 μm.
Dow
nloaded from https://academ
ic.oup.com/aob/article/122/5/679/4810445 by guest on 14 Septem
ber 2022

Apostolakos et al. — Subsidiary cell formation in maize stomatal complexes688
large number of cell surface receptors, at least in animal cells (Oude Weernik et al., 2007). Therefore, we may conclude that cell surface receptors and their interaction with PLDs respond to the particular conditions established on the SMC polar sur-face region. Nevertheless, whether in SMCs the activated PLDs display a polar accumulation, as hypothesized, remains to be investigated. Finally, the findings described above also suggest that in SMCs the nuclear division and determination of the div-ision plane may be independent phenomena, probably induced by different signals emitted by the GMCs.
PLAUSIBLE TRANSDUCTION MECHANISMS OF THE GMC STIMULUS TOWARD THE SMC
A detailed proteomic analysis from successive developmen-tal leaf stages in Z. mays revealed a significant enrichment of auxin-related proteins in leaf regions close to the leaf meristem (Facette et al., 2013). Therefore, it is likely that the first step during SMC induction probably includes auxin transport in the apoplast between GMCs and SMCs. This makes sense if we also consider that, in advanced GMCs, the PIN1 efflux carriers are redistributed and a large number of them are recruited to the plasmalemma lining the lateral GMC cell walls (see ‘Auxin transporters’). The recycling process of the PIN1 auxin carriers
depends on AFs. In fact, auxin-driven generation of cortical AFs has been implicated in the inhibition of PIN1 endocyto-sis which, in turn, leads to PIN polarization (Nagawa et al., 2012; Oliva et al., 2013). Therefore, it seems to be more than a coincidence that in protodermal leaf areas enriched with young GMCs where the PIN1 carriers were localized in the plasma-lemma lining the transverse cell walls, AFs mostly lined the same cell walls (Fig. 5A, B). In contrast, in protodermal areas where advanced GMCs dominated, PIN1 efflux carriers mark the whole lateral cell wall faces (see ‘Auxin transporters’) and AFs line the whole surface of the same GMC cell walls (Panteris et al., 2007; see also Fig. 5C). Hence, it is reason-able to correlate this AF reorganization with the redistribution of PIN1 carriers, which mediate auxin transport to the GMC/SMC apoplast (Fig. 6). PI3K and phospholipases seem also to be directly or indirectly involved in the recycling and/or the activation of PIN1 carriers (Fig. 6). Considering that inhibition of PI3K activity interferes with the redistribution of PIN1 car-riers in GMCs (Livanos et al., 2015) and that the arrest of PA production through PLDs disturbs cortical AF redistribution in GMCs (Fig. 5D; cf. Fig. 5C), it may be assumed that PI3K and PA are involved in auxin transport in the GMC/SMC apoplast. To confirm this hypothesis accurately, it would be interesting to examine PIN1 distribution in protodermal regions treated with phospholipase modulators. Then, similarly to other cases
Control Control
Control But-1
A B
C D
GMCs
SMCs
Fig. 5. AF organization in Z. mays GMCs and SMCs. AF staining was performed according to Panteris et al. (2006, 2007). (A and B) Optical sections passing through the surface (A) and the middle (B) of a young GMC. The AFs mainly line the transverse cell walls (arrowheads). (C) Optical section passing through the middle of an advanced GMC. Intense AF aggregations line the whole surface of the lateral cell walls of the GMC (arrowheads). The arrow points to the AF patch of the neighbouring SMC. (D) Advanced GMC and SMC treated with 20 mm 1-butanol (But-1) for 24 h. The AF organization in them is disturbed compared with
the respective cell types of the untreated seedlings (compare with C). Scale bars = 10 μm.
Dow
nloaded from https://academ
ic.oup.com/aob/article/122/5/679/4810445 by guest on 14 Septem
ber 2022

Apostolakos et al. — Subsidiary cell formation in maize stomatal complexes 689
(Joo et al., 2001, 2005; Foreman et al., 2003; Potocký et al., 2007; Ivanchenko et al., 2013; Ma et al., 2014; Sandalio et al., 2016), auxin accumulating in the apoplast between GMC and SMC is likely to induce ROS production, by PI3K-mediated activation of NADPH oxidases (Livanos et al., 2016; see Fig. 6). Subsequently, auxin and ROS may trigger three pos-sibly independent, yet synchronous, processes in the SMC: (1) local SMC growth towards the inducing GMC and AF patch formation; (2) nuclear polarization and division; and (3) deter-mination of the plane of the asymmetric division.
Local SMC growth towards the inducing GMC and AF patch formation
An emerging significant factor mediating local SMC growth is the preferential deposition of de-esterified HGAs (2F4 and JIM5 epitopes) and AGPs in the cell wall lining the polar SMC site (see ‘Local cell wall differentiation at the polar SMC site’; see also Fig. 7). These cell wall constituents probably allow the expansion and deformation of the common cell wall between the SMC and GMC, which accompany the local growth of the former cell towards the latter. HGA de-methyl esterification is mediated by pectin de-methylesterases (PMEs; Verhertbruggen and Knox, 2006; Wolf et al., 2012; Müller et al., 2013), while PME activity is regulated by auxin (Bryan and Newcomb, 1954; see also Braybrook and Peaucelle, 2013; Xiong et al., 2015). Moreover, Xiong et al. (2015) showed that H2O2 induces PME activation that controls HGA de-esterification and increases cell wall expansion in roots. Relying on these findings, it may be presumed that ROS in the apoplast between GMCs and SMCs act downstream of auxin to modulate, among others, the activity of PMEs at the SMC polar site (Fig. 7). In a similar direction, in the shoot apical meristem of A. thaliana, the polar distribu-tion of PIN1 auxin efflux carriers is also controlled by HGA
de-esterification (Braybrook and Peaucelle, 2013). Therefore, it may be hypothesized that in the common SMC–GMC cell wall, auxin, ROS and PME function in a feedback loop system that reinforces auxin flow from the GMC to the apoplast of the polar SMC site, promoting local HGA de-esterification (Fig. 7).
The 2F4-HGA epitope and the SWRC are early features of SMC polarization, as both of them have been localized at the polar site of SMCs, adjacent to young GMCs (Facette et al., 2015; Giannoutsou et al., 2016; see also Fig. 4A, B). Consequently, the de-esterified HGAs at the polar SMC site may be involved in the recruitment of the SWRC in the plasma-lemma of this region (Fig. 7). Evidence favouring this hypoth-esis is that the extracellular domain of the cell wall-associated kinases binds tightly to pectins, apparently with a pronounced preference for the Ca2+ egg box pectin conformations (Wolf et al., 2012; Kohorn, 2016; Wolf, 2017), which mainly de-velop between molecules of the 2F4-HGA epitope (Liners et al., 1989). However, although it is already known that cell wall-associated kinases are implicated in downstream signal transduction (Wolf et al., 2012; Kohorn, 2016; Wolf, 2017), their impact on SWRC accumulation at the polar site of SMCs remains to be investigated.
Consequently, the SWRC recruits PAN2 at the polar SMC site, which in turn recruits PAN1 at the same region. PAN1 physically interacts with ROPs, favouring their polar accu-mulation and activation. The activated ROPs interact with the SWRC, leading to SWRC-mediated activation of the ARP2/3 complex (Facette et al., 2015). The ARP2/3 complex promotes AF polymerization, which results in AF patch assembly (Facette et al., 2015; see Fig. 7). The assembly of the AF patch which is thought to act protectively against plasmalemma damage (see ‘Polar organization of the cytoskeleton’) is implicated in the assembly of the ER patch at the polar SMC site (Giannoutsou et al., 2011), while the emergence of AGPs at the polar SMC site might be related to this localized ER accumulation (Fig. 7).
ICS
R
PLDsPIN1
PI3K
PI3K PLCs
Auxin
Intermediate cell of stomatal rowICSR
NADPH oxidase
GMC
Apoplast
SMC
AFsIC
SR
PLDs
IAA IAA
IAA
IAA
IAA
IAA
IAA
IAAIAA
IAA
IAAIAAIAA
ROS
PLCsPIN1
ROS
ROS
Fig. 6. Diagram illustrating the possible mechanism of auxin and ROS accumulation in the apoplast between the SMC and inducing GMC. Briefly, in the advanced GMC, PIN1 auxin carriers are redistributed presumably in a cortical AF-dependent manner, whereas PI3K and phospholipases are thought to be directly or indir-ectly involved in the activation of PIN1 carriers. Afterwards, auxin is transported from the GMC to the apoplast between the GMC and SMC. The auxin accumu-
lation in the apoplast is likely to induce, in turn, ROS production catalysed by NADPH oxidases, mediated by PI3K.
Dow
nloaded from https://academ
ic.oup.com/aob/article/122/5/679/4810445 by guest on 14 Septem
ber 2022

Apostolakos et al. — Subsidiary cell formation in maize stomatal complexes690
PAN1 and PAN2 are recruited to the polar SMC site, binding hypothetical apoplast ligands, and this binding may promote or retain their polarization or regulate their function (Facette et al., 2015). These hypothetical ligands might be the de-esterified HGAs or oligogalacturonide molecules derived from the break-down of HGAa (Fig. 7). However, although RLKs have been implicated in cell wall binding and cell wall-related signalling, it is still unclear whether structural cell wall components are actual ligands of these proteins (Wolf et al., 2012; Wolf, 2017). Finally, considering that the activity of PLDs is related to AF patch formation (see ‘Phospholipases’), it may be suggested that phospholipases facilitate the recruitment of molecules related to AF patch formation, regulating the plasmalemma lipid content at the polar SMC site (Fig. 7).
Nuclear polarization and division
Auxin accumulated in the apoplast between the GMC and SMC seems to be transferred to the cytoplasm of the polar SMC site (Fig. 8). Considering that PIN1 auxin carrier distribution seems to be controlled by de-esterified HGAs (Braybrook and
Peaucelle, 2013), it may be assumed that the de-esterified HGAs localized in the cell wall of the polar SMC site (Giannoutsou et al., 2016) are involved in the recruitment of the auxin influx carriers to the plasmalemma of this region (Fig. 8). Afterwards, auxin might activate PI3K and subsequently ROS production via NADPH oxidase activation at the cytoplasm of the SMC polar site (Livanos et al., 2016; see also Fig. 8). Local ROS pro-duction may also be mediated by oligogalacturonide molecules derived from HGA breakdown. It is known that oligogalacturo-nides are generated in cell wall regions, where highly de-ester-ified and non-esterified HGAs dominate and that they function as signalling molecules, which among others are implicated in ROS production (Spiro et al., 1998; Seifert and Blankopf, 2010; Wolf et al., 2012; Wolf, 2017). Therefore, it is tempting to suggest that the 2F4- and JIM5-HGA epitopes, preferentially localized in the cell wall of the SMC polar site, are the source of local oligogalacturonide production, which in turn contrib-utes to local ROS accumulation (Giannoutsou et al., 2016). In this way, the cell wall may be related to the transduction of the auxin stimulus in the SMC interior (Fig. 8).
Similarly to other polarized cell types (Foreman et al., 2003; Potocký et al., 2007; Coelho et al., 2008; Duan et al., 2010;
ICS
R PI3K
PLCsPLDs
PLDs
ROS
ROSPIN1 PIN1
ROS
ROS
PI3KPLCs
PME
AFs
AFs
Apoplast
GMC
SMC
ICS
R
Esterified HGAs
AGPs??
?
Auxin
Intermediate cell of stomatal rowICSR
NADPH oxidase
Calcium ions
De-esterified homogalacturonans
Polarsite
ER
ROPSC
AR
/W
AV
E
SC
AR
/W
AV
E
PA
N2
PA
N1
Oligogalacturonides or ‘egg boxes’
IAA
IAA IAA IAA
IAA
IAA
IAA
IAA
ARP
2/3
IAA
Fig. 7. Diagrammatic representation of the possible steps of the SMC protrusion towards the inducing GMC and the formation of the AF-patch. (1) ROS and auxin accumulation in the apoplast between the GMC and SMC might modulate the activity of pectin methylesterases which catalyse the de-esterification of HGAs. (2) The latter de-esterified HGAs are localized exclusively at the common cell wall between the two cells and contribute to its deformation. Consequently, SMC grows locally towards the GMC. (3) It is also likely that the de-esterified HGAs at the cell wall in the proximity of the polar SMC site are involved in ROS generation as well as in the recruitment of the SWRC in the SMC plasmalemma lining this region. (4) SWRC recruits PAN2 at the polar SMC site, which in turn recruits PAN1. (5) PAN1 facilitates local accumulation and activation of ROPs. (6) Activated ROPs interact with the SWRC, triggering the activation of the ARP2/3 complex which leads in the end to the formation of the AF patch. (7) The presence of the latter is implicated in the assembly of a respective ER patch at the polar SMC site.
Dow
nloaded from https://academ
ic.oup.com/aob/article/122/5/679/4810445 by guest on 14 Septem
ber 2022

Apostolakos et al. — Subsidiary cell formation in maize stomatal complexes 691
De Smet and Beeckman, 2011; Wu et al., 2011; Nakamura et al., 2012), via the mechanisms described above, the cyto-plasm of the polar SMC site becomes enriched with auxin and ROS (Livanos et al., 2015, 2016). Furthermore, the AGPs local-ized at the polar SMC site (Giannoutsou et al., 2016) probably contribute to the enrichment of the polar SMC cytoplasm by higher Ca2+ concentrations (Fig. 8). AGPs bind to Ca2+ more strongly than to pectins, becoming dominating apoplastic Ca2+ stores (Lamport and Varnai, 2013). Auxin activates plasma-lemma H+ ATPases that transport H+ in the apoplast, decreasing its pH. Low pH triggers the release of Ca2+ from the AGPs. They enter the cytoplasm though Ca2+ channels, increasing the cytosolic Ca2+ concentrations. In this way, auxin establishes a cytosolic Ca2+ gradient (Lamport et al., 2014). Therefore, the transduction of the GMC auxin stimulus in SMCs may be accompanied by the establishment of local Ca2+ oscillations (Fig. 8).
Afterwards, as described earlier, the nucleus migrates to-wards the inducing GMC via the actomyosin system (Fig. 8; see also Kennard and Cleary 1997). Recent data showed that
auxin alone or in co-operation with ROS controls cytoplasmic AF systems in SMCs, mediating the polar nuclear movement (Livanos et al., 2015, 2016). It might be suggested that Ca2+ in the polar SMC site also plays a role in this process, e.g. modu-lating the function of the actomyosin system (see Hepler, 2005).
In addition, mechanisms that trigger the nuclear division of the SMC are presumably activated simultaneously with the polar movement of the nucleus (Fig. 8). In plants, auxin, alone or in collaboration with ROS, controls the cell cycle (Feher et al., 2008; Considine and Foyer, 2014). Therefore, in SMCs, auxin, via ROS production, might be implicated in nuclear div-ision (Livanos et al., 2015, 2016), influencing the expression of auxin-responsive genes, as occurs in other cases (Wu et al., 2011; Ma et al., 2014).
Mitogen-activated protein kinase (MAPK) cascades facili-tate the transduction of hormonal and ROS stimuli towards the nucleus (Hirt, 2000; Fernadez-Marcos et al., 2013). A simi-lar signalling pathway may also function in Z. mays SMCs (Fig. 8). A p38-like MAPK participates in the mechanism sensing ROS changes in root tips (Livanos et al., 2014). This
ICSRPI3K
PI3K
PLDs
PLDs
PLCs
PLCs
PIN1
ROS
ROS
ROS
ROS
PIN1
AUX1AUX1
AREAuxin-induced
gene expression ?
Apoplast
AFs
SMC
GMC
AGPs
Hypothetical MAPKsignalling cascade
Induction ofnuclear division
Nucleus
?
ICSR
Auxin
Auxin response elements
Intermediate cell of stomatal rowICSR
ARE
NADPH oxidase
Calcium ions
De-esterified homogalacturonans
Oligogalacturonides or “egg boxes”
IAA
IAAIAA IAA IAA
IAA
IAA
IAA
IAA
IAAIAA
IAAIAA
IAA
IAA
Fig. 8. Diagram depicting the possible mechanism of the induction of nuclear division of the SMC. Auxin accumulated in the apoplast between the GMC and SMC seems to be transferred to the cytoplasm of the polar SMC site. The presence of auxin leads to the accumulation of ROS in the SMC cortical cytoplasm, presumably via the activation of PI3K. Oligogalacturonide molecules originating from de-esterified HGA breakdown at the cell wall in the vicinity of the polar SMC site might also stimulate ROS production. Further, the AGPs localized at the same site probably contribute to the accumulation of Ca2+. Higher calcium concentrations, auxin and ROS modulating the function of the actomyosin system mediate the polar migration of the nucleus close to the inducing GMC. Auxin, ROS and phospholipases at the polar SMC site might be involved in the induction of nuclear division in multiple ways, i.e. triggering the activation of MAPK
cascades or the expression of auxin-responsive genes.
Dow
nloaded from https://academ
ic.oup.com/aob/article/122/5/679/4810445 by guest on 14 Septem
ber 2022

Apostolakos et al. — Subsidiary cell formation in maize stomatal complexes692
MAPK is activated by PA (Komis et al., 2004). Considering that the inhibition of PLC and PLD activity is accompanied by inhibition of SMC division (see ‘Phospholipases’), it could be interesting to examine the probable participation of ROS-sensing kinases along with phospholipid turnover in the signal-ling cascades that induce SMC nuclear division.
The transport of the inducing stimulus seems to be a highly controlled directional movement. The daughter cell wall of the SMC asymmetric division prevents the transfer of the GMC stimulus to the neighbouring epidermal cells (see ‘The na-ture of the inductive GMC stimulus’), although the plasma-lemma lining this cell wall displays PIN1 auxin transporters (Sutimantanapi et al., 2014). The young SC cell wall, in con-trast to the common GMC–SMC wall, is not enriched in de-esterified and non-esterified HGAs (Giannoutsou et al., 2016). This difference might explain why the transfer of the auxin stimulus towards the adjacent protodermal cell is prevented, while the possibility that the PIN1 auxin carriers on the sur-faces of cell wall generated by the asymmetric SMC division are not activated cannot be excluded. In contrast, the transfer of the inductive stimulus of the asymmetric SMC division towards the lateral epidermal cells proceeds when the SC cell wall is perforated (Apostolakos and Galatis, 1987).
Determination of the SMC division plane
It has been suggested that the determination of the asym-metric SMC division plane, as it is expressed by the MT-PPB, is defined by mechanical stresses imposed by the GMC on the SMC (Green et al., 1970; Galatis and Apostolakos, 2004;
Panteris et al., 2007; Apostolakos et al., 2008). Recently, it was found that in Z. mays SMCs displaying polarized nuclei, the 2F4-HGA epitope is not restricted to the common SMC–GMC cell wall, but it is also deposited in the proximal cell wall regions adjacent to the cortical cytoplasm, where the MT-PPB is assembled (Fig. 9), and the future cell plate will meet the parent cell walls (Giannoutsou et al., 2016). This indicates, for the first time, that cell wall matrix actively participates in the determination of the plane of cell division.
The cell wall at the polar SMC site expands and simultane-ously seems to deform since it displays large quantities of de-esterified HGAs. As a result, the SMC locally grows towards the GMC while the neighbouring SMC cell wall regions that lack de-esterified HGAs do not expand (Giannoutsou et al., 2016). Therefore, tensile forces developing on the protruding SMC region, which might react in the opposite way, could lead to the generation of intensive mechanical stresses at the con-tact region between expanding and non-expanding SMC wall regions (Fig. 9). In other words, the local cell wall matrix dif-ferentiation in the protruding SMC region imposes an aniso-tropic pattern of growth, generating mechanical stresses on the surface of the growing SMC region. Taking into account the distribution of mechanical stresses in the developing shoot api-cal meristem dome (see the references in Landrein and Hamant, 2013; Nick, 2013), it may be suggested that intense mechanical stresses prevail at the margins of the expanding and deforming SMC wall areas (Fig. 9).
It is well established that cortical MTs sense the mechanical stresses and respond by their grouping at the regions of max-imum stress, while as a rule they are absent from the regions of minimum stress (Landrein and Hamant, 2013; Nick, 2013;
ICSR GMC
MAPs?ROP6?
SMC
Calcium ions
De-esterified homogalacturonans
Intermediate cell of stomatal row
MTs
ROS
Katanin?
MT disappearance
MT bundling(MT-PPB)
MT bundling(MT-PPB)
ROS
Maximummechanical
stress
Maximummechanical
stress
Expandingcell wall region
Expandingcell wall region
Stretch mechanical tensionNon-expandingcell wall region
Non-expandingcell wall region
Compressonforce
Compressonforce
MAPs?ROP6?
ICSR
ICSR
Apoplast
Fig. 9. Diagram showing the possible mechanism of MT-PPB assembly in SMCs. The division plane is delineated by the MT-PPB at the cortical cytoplasm lining the contacts between expanding and non-expanding SMC cell wall regions. Maximum mechanical stress generated at these regions triggers the reorganization of
cortical MTs in these sites and their bundling, leading to the generation of the MT-PPB.
Dow
nloaded from https://academ
ic.oup.com/aob/article/122/5/679/4810445 by guest on 14 Septem
ber 2022

Apostolakos et al. — Subsidiary cell formation in maize stomatal complexes 693
Sampathkumar et al., 2014; Chen et al., 2016). This could pro-vide an explanation for the absence of MTs from the dome of the polar SMC site and their grouping at its margins, where the MT-PPB is formed (Fig. 9). The latter, polarizing its cyto-plasmic site, seems to mark accurately the junction sites of the cell plate of the asymmetric division with the parent cell walls (Galatis and Apostolakos, 2004; Facette and Smith, 2012; Pillitteri and Torri, 2012). This hypothesis may also explain MT-PPB formation in SMCs, in which the asymmetric division has been experimentally inhibited (Apostolakos et al., 2008; Livanos et al., 2015, 2016), as in these SMCs local growth to-wards the inducing GMCs is not affected.
Regarding the molecular mechanism leading to MT ‘dis-appearance’ from the dome of the SMC protrusion, as well as to that orchestrating MT-PPB formation at its margins, nothing is certain so far. In other cell systems, MTs function as sensors of mechanical stress in co-operation with various molecules, such as katanin, CLASP and other MT-associated proteins, ROP6 and its effector RIC1, as well as Ca2+ gradients and ROS (Landrein and Hamant, 2013; Nick, 2013; Sampathkumar et al., 2014; Chen et al., 2016). The presence of the above molecules in the SMC polar region remains to be investigated (Fig. 9).
CONCLUSIONS, OUTLOOK AND FUTURE DIRECTIONS
This review attempts to summarize the existing knowledge on polarization and asymmetric division that lead to the generation of the SCs in the stomatal complexes of Z. mays. Although there has been remarkable progress in understanding many aspects of these phenomena, it becomes clear that further work is needed to elucidate the mechanisms underlying these processes. Therefore, it would be interesting to search for the presence and function of novel molecules localized in the SMC polar site and investigate putative MAPKs that accomplish the transduction of GMC-initiated stimulation. Presumably, the analysis and com-parison between the transcriptional profiles of GMCs and SMCs would also reveal more participants. In addition, it would be of special interest to investigate the transcriptional responses that orchestrate the transduction and the perception of the auxin stimulus, which operates this complicated intercellular cross-talk and leads to the generation of SCs. In this direction, we would like to draw readers’ attention to recently published work by Raissig et al. (2016, 2017) describing a network of transcrip-tion factors governing the development and patterning of sto-matal complexes of the grass Brachypodium distachyon.
ACKNOWLEDGEMENTS
We are grateful to the reviewers for their critical and valu-able comments, and we would like to thank Professor Laurie G. Smith and Yeri Park for providing the image shown in Fig. 4A.
LITERATURE CITED
Apostolakos P, Galatis B. 1987. Induction, polarity and spatial control of cytokinesis in some abnormal subsidiary cell mother cells of Zea mays. Protoplasma 140: 26–42.
Apostolakos P, Panteris E, Galatis B. 2008. The involvement of phospholi-pases C and D in the asymmetric division of subsidiary cell mother cells of Zea mays. Cell Motility and the Cytoskeleton 65: 863–875.
Armour WJ, Barton DA, Law AML, Overall RL. 2015. Differential growth in periclinal and anticlinal walls during lobe formation in Arabidopsis cotyledon pavement cells. The Plant Cell 27: 2484–2500.
Baluška F, Salaj J, Mathur J, et al. 2000a. Root hair formation: F-actin dependent tip growth is initiated by local assembly of profile-supported F-actin meshworks accumulated within expansin-enriched bulges. Developmental Biology 227: 618–632.
Baluška F, Volkmann D, Barlow PW. 2000b. Actin-based domains of the ‘cell periphery complex’ and their associations with polarized ‘cell bod-ies’ in higher plants. Plant Biology 2: 253–267.
Baluška F, Volkmann D, Barlow PW. 2001. Motile plant cell body: a ‘bug’ within a ‘cage’. Trends in Plant Science 6: 104–111.
Balzan S, Gurmukh SJ, Carraro N. 2014. The role of auxin transporters in monocots development. Frontiers in Plant Science 5: 393.
Belanger KD, Quatrano RS. 2000. Polarity: the role of localized secretion. Current Opinion in Plant Biology 3: 67–72.
Bidhendi AJ, Geitmann A. 2016. Relating the mechanics of the primary plant cell wall to morphogenesis. Journal of Experimental Botany 67: 449–461.
Bleasdale JE, Bundy GL, Bunting S, et al. 1989. Inhibition of phospho-lipase C dependent processes by U-73122. Advances in Prostaglandin, Thromboxane and Leukotriene Research 19: 590–593.
Bonsignore CL, Hepler PK. 1985. Caffeine inhibition on cytokinesis: dynam-ics of cell plate formation–deformation in vivo. Protoplasma 129: 28–35.
Bosch M, Hepler PK. 2005. Pectin methylesterases and pectin dynamics in pollen tubes. The Plant Cell 17: 3219–3226.
Boudeau J, Miranda-Saavedra D, Barton GJ, Alessi DR. 2006. Emerging roles of pseudokinases. Trends in Cell Biology 16: 443–452.
Braybrook SA, Peaucelle A. 2013. Mechano-chemical aspects of organ for-mation in Arabidopsis thaliana: the relationship between auxin and pectin. PLoS One 8: e57813.
Bryan WH, Newcomb KH. 1954. Stimulation of pectin methylesterase activ-ity of cultured tobacco pith by indoleacetic acid. Physiologia Plantarum 7: 290–297.
Bünning E. 1956. General processes of differentiation. In: Milthorpe FL, ed. The growth of leaves. London: Butterworths Scientific Publications, 18–30.
Bünning E. 1957. Polarität und inäquale teilung pflanzlichen protoplasten. Protoplasmatologia 8: 1–86.
Cartwright HN, Humphries JA, Smith LG. 2009. PAN1: a receptor-like pro-tein that promotes polarization of an asymmetric cell division in maize. Science 323: 649–651.
Castro AJ, Suarez C, Zienkiewicz K, Alche JdD, Zienkiewicz A, Rodriguez-Garcia MI. 2013. Electrophoretic profiling and immunocytochemical detection of pectins and arabinogalactan proteins in olive pollen during germination and pollen tube growth. Annals of Botany 112: 503–513.
Chen X, Wu S, Liu Z, Friml J. 2016. Environmental and endogenous control of cortical microtubule orientation. Trends in Cell Biology 26: 409–419.
Cho S-O, Wick SM. 1990. Distribution and function of actin in the developing stomata complex of the winter rye (Secale cereale cv. Puma). Protoplasma 157: 154–164.
Cho S-O, Wick SM. 1991. Actin in developing stomatal complex of the win-ter rye: a comparison of actin antibodies and Rh–phalloidin labeling of control and CB-treated tissues. Cell Motility and the Cytoskeleton 19: 25–36.
Cleary AL. 1995. F-actin redistributions at the division site in living Tradescantia stomatal complexes as revealed by microinjection of rhoda-mine–phalloidin. Protoplasma 185: 152–165.
Cleary AL. 2000. Actin in formation of stomatal complexes. In: Staiger C, Baluska F, Volkmann D, Barlow P, eds. Actin: a dynamic framework for multiple plant cell functions. Dordrecht, The Netherlands: Kluwer Academic Publishers, 411–426.
Cleary AL, Mathesius U. 1996. Rearrangements of F-actin during stoma-togenesis visualized by confocal microscopy in fixed and permeabilised Tradescantia leaf epidermis. Botanica Acta 109: 15–24.
Coelho SMB, Brownlee C, Bothwell JHF. 2008. A tip-high, Ca2+-
interdependent, reactive oxygen species gradient is associated with polar-ized growth in Fucus serratus zygotes. Planta 227: 1037–1046.
Cole R, Fowler J. 2006. Polarized growth: maintaining focus on the tip. Current Opinion in Plant Biology 9: 579–588.
Dow
nloaded from https://academ
ic.oup.com/aob/article/122/5/679/4810445 by guest on 14 Septem
ber 2022

Apostolakos et al. — Subsidiary cell formation in maize stomatal complexes694
Considine MJ, Foyer CH. 2014. Redox regulation of plant development. Antioxidants and Redox Signaling 21: 1305–1326.
De Smet I, Beeckman T. 2011. Asymmetric cell division in land plants and algae: the driving force for differentiation. Nature Reviews. Molecular Cell Biology 12: 177–188.
Dowd PE, Coursol S, Skirpan AL, Kao T-H, Gilroy S. 2006. Petunia phospholipase C1 is involved in pollen tube growth. The Plant Cell 18: 1438–1453.
Duan Q, Kita D, Li C, Cheung AY, Wu H-M. 2010. FERONIA receptor-like kinase regulates RHO GTPase signaling of root hair development. Proceedings of the National Academy of Sciences, USA 107: 17821–17826.
English AR, Zurek N, Voeltz GK. 2009. Peripheral ER structure and function. Current Opinion in Cell Biology 21: 596–602.
Facette MR, Smith LG. 2012. Division polarity in developing stomata. Current Opinion in Plant Biology 15: 585–592.
Facette MR, Shen Z, Björnsdóttir FR, Briggs SP, Smith LG. 2013. Parallel proteomic and phosphoproteomic analyses of successive stages of maize leaf development. The Plant Cell 25: 2798–2812.
Facette MR, Park Y, Sutimantanapi D, et al. 2015. The SCAR/WAVE com-plex polarizes PAN receptors and promotes division asymmetry in maize. Nature Plants 1: 14024.
Fehér A, Ötvös K, Pasternak TP, Szandtner AP. 2008. The involvement of reactive oxygen species (ROS) in the cell cycle activation (G0-to-G1 tran-sition) of plant cells. Plant Signaling and Behavior 3: 823–826.
Fernández-Marcos M, Sanz L, Lewis DR, Muday GK, Lorenzo O. 2013. Control of auxin transport by reactive oxygen and nitrogen species. In: Chen R, Baluška F, eds. Polar auxin transport. Heidelberg: Springer, 103–118.
Fischer V, Men S, Grebe M. 2004. Lipid function in plant cell polarity. Current Opinion in Plant Biology 7: 670–676.
Foreman J, Demidchik V, Bothwell JHF, et al. 2003. Reactive oxygen spe-cies produced by NADPH oxidase regulate plant cell growth. Nature 422: 442–446.
Forestan C, Varotto S. 2012. The role of PIN auxin efflux carriers in polar auxin transport and accumulation and their effect on shaping maize devel-opment. Molecular Plant 5: 787–798.
Frame MD, Sarelius IH. 2000. Flow-induced cytoskeletal changes in endothe-lial cells growing on curved surfaces. Microcirculation 7: 419–427.
Frank MJ, Smith LG. 2002. A small, novel protein highly conserved in plants and animals promotes the polarized growth and division of maize leaf epi-dermal cells. Current Biology 12: 849–853.
Frank MJ, Cartwright HN, Smith LG. 2003. Three Brick genes have distinct functions in a common pathway promoting polarized cell division and cell morphogenesis in the maize leaf epidermis. Development 130: 753–762.
Gabev E, Kasianowicz J, Abbott T, McLaughlin S. 1989. Binding of neo-mycin to phosphatidylinositol 4,5-bisphosphate (PIP2). Biochimica et Biophysica Acta 979: 105–112.
Galatis B. 1982. The organization of microtubules in guard cell mother cells of Zea mays. Canadian Journal of Botany 60: 1148–1166.
Galatis B, Apostolakos P. 2004. The role of the cytoskeleton in the morphogen-esis and function of stomatal complexes. New Phytologist 161: 613–639.
Galatis B, Apostolakos P, Katsaros C. 1983. Synchronous organization of two preprophase microtubule bands and final cell plate arrangement in subsidiary cell mother cells of some Triticum species. Protoplasma 117: 24–39.
Galatis B, Apostolakos P, Katsaros C. 1984a. Positional inconsistency between preprophase microtubule band and final cell plate arrangement during subsidiary cell and hair cell formation in two Triticum species. Canadian Journal of Botany 62: 343–359.
Galatis B, Apostolakos P, Katsaros C. 1984b. Experimental studies on the function of the cortical cytoplasmic zone of the preprophase microtubule band. Protoplasma 122: 11–26.
Gallagher K, Smith LG. 1999. Discordia mutations specifically misorient asymmetric cell divisions during development of maize leaf epidermis. Development 126: 4623–4633.
Gallagher K, Smith LG. 2000. Roles for polarity and nuclear determinants in specifying daughter cell fates after an asymmetric cell division in the maize leaf. Current Biology 10: 1229–1232.
Geldner N, Friml J, Stierhof Y-D, Jürgens G, Palme K. 2001. Auxin transport inhibitors block PIN1 cycling and vesicle trafficking. Nature 413: 425–428.
Giannoutsou E, Apostolakos P, Galatis B. 2011. Actin filament-organized local cortical endoplasmic reticulum aggregations in developing stomatal complexes of grasses. Protoplasma 248: 373–390.
Giannoutsou E, Galatis B, Zachariadis M, Apostolakos P. 2012. Formation of an endoplasmic reticulum ring associated with acetylated microtubules in the angiosperm preprophase band. Cytoskeleton 69: 252–265.
Giannoutsou E, Apostolakos P, Galatis B. 2016. Spatio-temporal diversifica-tion of the cell wall matrix materials in the developing stomatal complexes of Zea mays. Planta 244: 1125–1143.
Green PB, Erickson RO, Richmond PA. 1970. On the physical basis of cell morphogenesis. Annals of the New York Academy of Sciences 175: 712–731.
Griffing LR, Lin C, Perico C, White RR, Sparkes I. 2017. Plant ER geom-etry and dynamics: biophysical and cytoskeletal control during growth and biotic response. Protoplasma 254: 43–56.
Hepler PK. 2005. Calcium: a central regulator of plant growth and develop-ment. The Plant Cell 17: 2142–2155.
Hirt H. 2000. Connecting oxidative stress, auxin, and cell cycle regulation through a plant mitogen-activated protein kinase pathway. Proceedings of the National Academy of Sciences, USA 97: 2405–2407.
Humphries JA, Vejlupkova Z, Luo A, et al. 2011. ROP GTPases act with the receptor-like protein PAN1 to polarize asymmetric cell division in maize. The Plant Cell 23: 2273–2284.
Ivanchenko MG, den Os D, Monshausen GB, Dubrovsky JG, Bednárová A, Krishnan N. 2013. Auxin increases the hydrogen peroxide (H2O2) concentration in tomato (Solanum lycopersicum) root tips while inhibiting root growth. Annals of Botany 112: 1107–1116.
Joo JH, Bae YS, Lee JS. 2001. Role of auxin-induced reactive oxygen species in root gravitropism. Plant Physiology 126: 1055–1060.
Joo JH, Yoo HJ, Hwang I, Lee JS, Nam KH, Bae YS. 2005. Auxin-induced reactive oxygen species production requires the activation of phosphati-dylinositol 3-kinase. FEBS Letters 579: 1243–1248.
Kennard JL, Cleary AL. 1997. Pre-mitotic nuclear migration in subsidiary mother cells of Tradescantia occurs in G1 of the cell cycle and requires F-actin. Cell Motility and the Cytoskeleton 36: 55–67.
Knox JP. 2006. Up against the wall: arabinogalactan-protein dynamics at cell surfaces. New Phytologist 169: 443–445.
Ko KS, McCulloch CA. 2000. Partners in protection: interdependence of cyto-skeleton and plasma membrane in adaptations to applied forces. Journal of Membrane Biology 174: 85–95.
Kohorn BD. 2016. Cell wall-associated kinases and pectin perception. Journal of Experimental Botany 67: 489–494.
Komis G, Apostolakos P, Gaitanaki C, Galatis B. 2004. Hyperosmotically induced accumulation of a phosphorylated p38-like MAPK involved in protoplast volume regulation of plasmolyzed wheat root cells. FEBS Letters 573: 168–174.
Lamport DTA, Varnai P. 2013. Periplasmic arabinogalactan glycoproteins act as a calcium capacitor that regulates plant growth and development. New Phytologist 197: 58–64.
Lamport DTA, Kieliszewski MJ, Showalter AM. 2006. Salt stress upregu-lates periplasmic arabinogalactan proteins: using salt stress to analyze AGP function. New Phytologist 169: 479–492.
Lamport DTA, Varnai P, Seal CE. 2014. Back to the future with the AGP–Ca2+ flux capacitor. Annals of Botany 114: 1069–1085.
Landrein B, Hamant O. 2013. How mechanical stress controls microtubule behavior and morphogenesis in plants: history, experiments and revisited theories. The Plant Journal 75: 324–338.
Le J, Liu X-G, Yang K-Z, et al. 2014. Auxin transport and activity regulate stomatal patterning and development. Nature Communications 5: 3090.
Lee SJ, Kuroha T, Hnilova M, et al. 2012. Direct interaction of ligand–recep-tor pairs specifying stomatal patterning. Genes and Development 26: 126–136.
Lee Y, Munnik T, Lee Y. 2010. Plant phosphatidylinositol 3-kinase. In: Munnik T, ed. Lipid signaling in plants. Heidelberg: Springer-Verlag, 95–106.
Lesage G, Bussey H. 2006. Cell wall assembly in Saccharomyces cerevisiae. Microbiology and Molecular Biology Reviews 70: 317–343.
Li G, Xue H-W. 2007. Arabidopsis PLDζ2 regulates vesicle trafficking and is required for auxin response. The Plant Cell 19: 281–295.
Li L, Saga N, Mikami K. 2008. Phosphatidylinositol 3-kinase activity and asymmetrical accumulation of F-actin are necessary for establishment of cell polarity in the early development of monospores from the marine red alga Porphyra yezoensis. Journal of Experimental Botany 59: 3575–3586.
Li R. 2013. The art of choreographing asymmetric cell division. Developmental Cell 25: 439–450.
Dow
nloaded from https://academ
ic.oup.com/aob/article/122/5/679/4810445 by guest on 14 Septem
ber 2022

Apostolakos et al. — Subsidiary cell formation in maize stomatal complexes 695
Li S, Sun T, Ren H. 2015. The functions of the cytoskeleton and associated proteins during mitosis and cytokinesis in plant cells. Frontiers in Plant Science 6: 282.
Liners F, Letesson JJ, Didembourg C, Van Cutsem P. 1989. Monoclonal antibodies against pectin: recognition of a conformation induced by cal-cium. Plant Physiology 91: 1419–1424.
Livanos P, Galatis B, Gaitanaki C, Apostolakos P. 2014. Phosphorylation of a p38-like MAPK is involved in sensing cellular redox state and drives atypical tubulin polymer assembly in angiosperms. Plant, Cell and Environment 37: 1130–1143.
Livanos P, Giannoutsou E, Apostolakos P, Galatis B. 2015. Auxin as an inducer of asymmetrical division generating the subsidiary cells in stoma-tal complexes of Zea mays. Plant Signaling and Behavior 10: e984531.
Livanos P, Galatis B, Apostolakos P. 2016. Deliberate ROS production and auxin synergistically trigger the asymmetrical division generating the sub-sidiary cells in Zea mays stomatal complexes. Protoplasma 253: 1081–1099.
Llompart B, Castells E, Rio A, et al. 2003. The direct activation of MIK, a germinal center kinase (GCK)-like kinase, by MARK, a maize atypical receptor kinase, suggests a new mechanism for signaling through kinase-dead receptors. Journal of Biological Chemistry 278: 48105–48111.
Ma F, Wang L, Li J, et al. 2014. Interaction between HY1 and H2O2 in auxin-induced lateral root formation in Arabidopsis. Plant Molecular Biology 85: 49–61.
Machesky LM, Mullins RD, Higgs HN, et al. 1999. Scar, a WASp-related protein, activates nucleation of actin filaments by the Arp2/3 complex. Proceedings of the National Academy of Sciences, USA 96: 3739–3744.
Masoud K, Herzog E, Chabouté ME, Schmit AC. 2013. Microtubule nucle-ation and establishment of the mitotic spindle in vascular plant cells. The Plant Journal 75: 245–257.
Meijer HJ, Munnik T. 2003. Phospholipid-based signaling in plants. Annual Review of Plant Biology 54: 265–306.
Miki H, Suetsugu S, Takenawa T. 1998. WAVE, a novel WASP-family pro-tein involved in actin reorganization induced by Rac. EMBO Journal 17: 6932–6941.
Mineyuki Y. 1999. The preprophase band of microtubules: its function as a cytoki-netic apparatus in higher plants. International Review of Cytology 187: 1–49.
Mittler R, Vanderauwera S, Suzuki N, et al. 2011. ROS signaling: the new wave? Trends in Plant Science 16: 300–309.
Müller K, Levesque-Tremblay G, Bartels S, et al. 2013. Demethylesterification of cell wall pectins in Arabidopsis plays a role in seed germination. Plant Physiology 161: 305–316.
Nadeau JA, Sack FD. 2002. Control of stomatal distribution on the Arabidopsis leaf surface. Science 296: 1697–1700.
Nagawa S, Xu T, Lin D, et al. 2012. ROP GTPase-dependent actin micro-filaments promote PIN1 polarization by localized inhibition of clathrin-dependent endocytosis. PLoS Biology 10: e1001299.
Nakamura M, Kiefer CS, Grebe M. 2012. Planar polarity, tissue polarity and planar morphogenesis in plants. Current Opinion in Plant Biology 15: 593–600.
Nelissen H, Rymen B, Jikumaru Y, et al. 2012. A local maximum in gibber-ellin levels regulates maize leaf growth by spatial control of cell division. Current Biology 22: 1183–1187.
Nguema-Ona E, Coimbra S, Vicre-Gibouin M, Mollet JC, Driouich A. 2012. Arabinogalactan proteins in root and pollen-tube cells: distribution and functional aspects. Annals of Botany 110: 383–404.
Nibau C, Wu HM, Cheung AY. 2006. RAC/ROP GTPases: ‘hubs’ for signal inte-gration and diversification in plants. Trends in Plant Science 11: 309–315.
Nick P. 2010. Probing the actin–auxin oscillator. Plant Signaling and Behavior 5: 94–98.
Nick P. 2013. Microtubules, signalling and abiotic stress. The Plant Journal 75: 309–323.
Oliva M, Vernoux T, Traas J. 2013. From auxin transport to patterning. In: Chen R, Baluška F, eds. Polar auxin transport. Heidelberg: Springer-Verlag, 259–279.
Oude Weernink PA, López de Jesús M, Schmidt M. 2007. Phospholipase D signaling: orchestration by PIP2 and small GTPases. Naunyn Schmiedeberg’s Archives of Pharmacology 374: 399–411.
Paciorek T, Bergmann DC. 2010. The secret to life is being different: asym-metric divisions in plant development. Current Opinion in Plant Biology 13: 661–669.
Panteris E, Apostolakos P, Galatis B. 2006. Cytoskeletal asymmetry in Zea mays subsidiary cell mother cells: a monopolar prophase microtubule half-spindle anchors the nucleus to its polar position. Cell Motility and the Cytoskeleton 63: 696–709.
Panteris E, Galatis B, Quader H, Apostolakos P. 2007. Cortical actin fila-ment organization in developing and functioning stomatal complexes of Zea mays and Triticum turgidum. Cell Motility and the Cytoskeleton 64: 531–548.
Panteris E, Galatis B, Apostolakos P. 2013. Examination of the possible role of PAN1 protein in the ontogenesis of Zea mays stomatal complexes. In: Gkelis S, Karousou R, Kokkini S, Panteris E. eds. Program and Abstracts. 13th Panhellenic Scientific Conference, Hellenic Botanical Society, Thessaloniki, 118.
Pasternak T, Potters G, Gaubergs R, Jansen MAK. 2005. Complementary interactions between oxidative stress and auxins control plant growth responses at plant, organ, and cellular level. Journal of Experimental Botany 56: 1991–2001.
Peaucelle A, Braybrook SA, Le Guillou L, Bron E, Kuhlemeier C, Höfte H. 2011. Pectin-induced changes in cell wall mechanics underlie organ initiation in Arabidopsis. Current Biology 21: 1720–1726.
Peaucelle A, Wightman R, Höfte H. 2015. The control of growth sym-metry breaking in the Arabidopsis hypocotyl. Current Biology 25: 1746–1752.
Pelletier S, Van Orden J, Wolf S, et al. 2010. A role for pectin de-methyl-esterification in a developmentally regulated growth acceleration in dark-grown Arabidopsis hypocotyls. New Phytologist 188: 726–739.
Perdiz D, Mackeh R, Poüs C, Baillet A. 2011. The ins and outs of tubu-lin acetylation: more than just a post-translational modification? Cell Signaling 23: 763–771.
Pickett-Heaps JD. 1969. Preprophase microtubule bands in some abnormal mitotic cells of wheat. Journal of Cell Science 4: 397–420.
Pickett-Heaps JD, Northcote DH. 1966. Cell division in the formation of the stomatal complex of the young leaves of wheat. Journal of Cell Science 1: 121–128.
Pickett-Heaps JD, Gunning BES, Brown RC, Lemmon BE, Cleary AL. 1999. The cytoplast concept in dividing plant cells: cytoplasmic domains and the evolution of spatially organized cell division. American Journal of Botany 86: 153–172.
Pichler H, Gaigg B, Hrastnik C, et al. 2001. A subfraction of the yeast endo-plasmic reticulum associates with the plasma membrane and has a high capacity to synthesize lipids. European Journal of Biochemistry 268: 2351–2361.
Pillitteri LJ, Torii KU. 2012. Mechanisms of stomatal development. Annual Review of Plant Biology 63: 591–614.
Potocký M, Eliáš M, Profotová B, Novotná Z, Valentová O, Žárský V. 2003. Phosphatidic acid produced by phospholipase D is required for tobacco pollen tube growth. Planta 217: 122–130.
Potocký Μ, Jones Μ, Bezvoda R, Smirnoff N, Žárský V. 2007. Reactive oxygen species produced by NADPH oxidase are involved in pollen tube growth. New Phytologist 174: 742–751.
Raissig MT, Abzash E, Bettadapur A, Vogel JP, Bergmann DC. 2016. Grasses use an alternative wired bHLH transcription factor network to establish stomatal identity. Proceedings of the National Academy of Sciences, USA 113: 8326–8331.
Raissig MT, Matos JL, Gil MXA, et al. 2017. Mobile MUTE specifies sub-sidiary cells to build physiologically improved grass stomata. Science 355: 1215–1218.
Rajakulendran T, Sicheri F. 2010. Allosteric protein kinase regulation by pseudokinases: insights from STRAD. Science Signaling 3: pe8.
Sandalio LM, Rodriquez-Serrano M, Romero-Puertas MC. 2016. Leaf epi-nasty and auxin: a biochemical and molecular overview. Plant Science 253: 187–193.
Sampathkumar A, Krupinski P, Wightman R, et al. 2014. Subcellular and supracellular mechanical stress prescribes cytoskeleton behavior in Arabidopsis cotyledon pavement cells. eLife 3: e01967.
Seifert GJ, Blaukopf C. 2010. Irritable walls: the plant extracellular matrix and signaling. Plant Physiology 153: 467–478
Shao W, Dong J. 2016. Polarity in plant asymmetric cell division: division orientation and cell fate differentiation. Developmental Biology 419: 121–131.
Shpak ED, McAbee JM, Pillitteri LJ, Torii KU. 2005. Stomatal patterning and differentiation by synergistic interactions of receptor kinases. Science 309: 290–293.
Smith LG. 2001. Plant cell division: building walls in the right places. Nature Reviews 2: 33–39.
Spiro MD, Ridley BL, Eberhard S, et al. 1998. Biological activity of reduc-ing-end-derivatized oligogalacturonides in Tobacco tissue cultures. Plant Physiology 116: 1289–1298.
Dow
nloaded from https://academ
ic.oup.com/aob/article/122/5/679/4810445 by guest on 14 Septem
ber 2022

Apostolakos et al. — Subsidiary cell formation in maize stomatal complexes696
Stebbins LG, Jain SK. 1960. Developmental studies of cell differentiation in the epidermis of monocotyledons. I. Allium, Rhoeo and Commelina. Developmental Biology 2: 409–426.
Stebbins LG, Shah SS. 1960. Developmental studies on the cell differentiation in the epidermis of monocotyledons. II. Cytological features of stomatal development in the Graminae. Developmental Biology 2: 477–500.
Sutimantanapi D, Pater D, Smith LG. 2014. Divergent roles for maize PAN1 and PAN2 receptor-like proteins in cytokinesis and cell morphogenesis. Plant Physiology 164: 1905–1917.
Tejos R, Sauer M, Vanneste S, et al. 2014. Bipolar plasma membrane dis-tribution of phosphoinositides and their requirement for auxin-mediated cell polarity and patterning in Arabidopsis. The Plant Cell 26: 2114–2128.
Tsiantis M, Brown MIN, Skibinski G, Langdale JA. 1999. Disruption of auxin transport is associated with aberrant leaf development in maize. Plant Physiology 121: 1163–1168.
Tsuda E, Yang H, Nishimura T, et al. 2011. Alkoxy-auxins are selective inhibitors of auxin transport mediated by PIN, ABCB, and AUX1 trans-porters. Journal of Biological Chemistry 286: 2354–2364.
Verhertbruggen Y, Knox JP. 2006. Pectic polysaccharides and expanding cell walls. In: Verbelen JP, Vissenberg K, eds. The expanding cell. Berlin, Heidelberg: Springer-Verlag, 139–158.
Vlahos CJ, Matter WF, Hui KY, Brown RF. 1994. A specific inhibitor of phos-phatidylinositol 3-kinase, 2-(4-morpholinyl)-8-phenyl-4H-1-benzopyran-4-one (LY294002). Journal of Biological Chemistry 269: 5241–5248.
Wang X. 2004. Lipid signaling. Current Opinion in Plant Biology 7: 329–336.Wick SM. 1991a. Spatial aspects of cytokinesis in plant cells. Current Opinion
in Cell Biology 3: 253–260.Wick SM. 1991b. The preprophase band. In: Lloyd CW, ed. The cytoskeletal
basis of plant growth and form. London: Academic Press, 231–244.Wolf S. 2017. Plant cell wall signalling and receptor-like kinases. Biochemical
Journal 474: 471–492.Wolf S, Greiner S. 2012. Growth control by cell wall pectins. Protoplasma 249
(Suppl 2): S169–S175.Wolf S, Hématy K, Höfte H. 2012. Growth control and cell wall signaling in
plants. Annual Review of Plant Biology 63: 381–407.Wright AJ, Gallagher K, Smith LG. 2009. Discordia1 and alternative discordia1
function redundantly at the cortical division site to promote preprophase band formation and orient division planes in maize. The Plant Cell 21: 234–247.
Wu Η, Hazak O, Cheung AY, Yalovsky S. 2011. RAC/ROP GTPases and auxin signaling. The Plant Cell 23: 1208–1218.
Xiong J, Yang Y, Fu G, Tao L. 2015. Novel roles of hydrogen peroxide (H2O2) in regulating pectin synthesis and demethylesterification in the cell wall of rice (Oryza sativa) root tips. New Phytologist 206: 118–126.
Yang Z. 2008. Cell polarity signaling in Arabidopsis. Annual Review of Cell and Developmental Biology 24: 551–575.
Yang Z, Fu Y. 2007. ROP/RAC GTPase signaling. Current Opinion in Plant Biology 10: 490–494.
Žarský V, Cvrckova F, Potocky M, Hala M. 2009. Exocytosis and cell polarity in plants – exocyst and recycling domains. New Phytologist 183: 255–272.
Zachariadis M, Quader H, Galatis B, Apostolakos P. 2001. Endoplasmic reticulum preprophase band in dividing root-tip cells of Pinus brutia. Planta 213: 824–827.
Zachariadis M, Quader H, Galatis B, Apostolakos P. 2003. Organization of the endoplasmic reticulum in dividing cells of the gymnosperms Pinus brutia and Pinus nigra and of the pterophyte Asplenium nidus. Cell Biology International 27: 31–40.
Zegzouti H, Anthony RG, Jahchan N, Bögre L, Christensen SK. 2006. Phosphorylation and activation of PINOID by the phospholipid signal-ing kinase 3-phosphoinositide-dependent protein kinase 1 (PDK1) in Arabidopsis. Proceedings of the National Academy of Sciences, USA 103: 6404–6409.
Zhang J-Y, He S-B, Li L, Yang H-Q. 2014. Auxin inhibits stomatal devel-opment through MONOPTEROS repression of a mobile peptide gene STOMAGEN in mesophyll. Proceedings of the National Academy of Sciences, USA 111: E3015–E3023.
Zhang X, Facette M, Humphries JA, et al. 2012. Identification of PAN2 by quantitative proteomics as a leucine-rich repeat-receptor-like kinase acting upstream of PAN1 to polarize cell division in maize. The Plant Cell 24: 4577–4589.
Zhou W, Wang Y, Wu Z, et al. 2016. Homologs of SCAR/WAVE complex components are required for epidermal cell morphogenesis in rice. Journal of Experimental Botany 67: 4311–4323.
Zykwinska AW, Ralet MC, Garnier CD, Thibault JF. 2005. Evidence for in vitro binding of pectin side chains to cellulose. Plant Physiology 139: 397–407.
Dow
nloaded from https://academ
ic.oup.com/aob/article/122/5/679/4810445 by guest on 14 Septem
ber 2022