longitudinal analysis of calorie restriction on rat taste bud morphology and expression of sweet...
TRANSCRIPT

Page 1 of 13
Journals of Gerontology: BIOLOGICAL SCIENCESCite journal as: J Gerontol A Biol Sci Med Scidoi:10.1093/gerona/glt129
Published by Oxford University Press on behalf of the Gerontological Society of America 2013.
Longitudinal Analysis of Calorie Restriction on Rat Taste Bud Morphology and Expression of
Sweet Taste Modulators
Huan Cai,1 Caitlin M. Daimon,1 Wei-na Cong,1 Rui Wang,1 Patrick Chirdon,1 Rafael de Cabo,2 Jean Sévigny,3,4 Stuart Maudsley,5,* and Bronwen Martin1,*
1Metabolism Unit, Laboratory of Clinical Investigation and 2Translational Gerontology Branch, National Institute on Aging, National Institutes of Health, Baltimore, Maryland.
3Centre de recherche en Rhumatologie et Immunologie, Centre de recherche du CHU de Québec, Canada.4Département de microbiologie-infectiologie et d’immunologie, Faculté de médecine, Université Laval, Québec, Canada.
5Receptor Pharmacology Unit, Laboratory of Neuroscience, National Institute on Aging, National Institutes of Health, Baltimore, Maryland.
*These authors contributed equally to this work.
Address correspondence to Bronwen Martin, PhD, Metabolism Unit, Laboratory of Clinical Investigation, National Institute on Aging, National Institutes of Health, 251 Bayview Boulevard, Suite 100, Baltimore, MD 21224. Email: [email protected]
Calorie restriction (CR) is a lifestyle intervention employed to reduce body weight and improve metabolic functions pri-marily via reduction of ingested carbohydrates and fats. Taste perception is highly related to functional metabolic status and body adiposity. We have previously shown that sweet taste perception diminishes with age; however, relatively little is known about the effects of various lengths of CR upon taste cell morphology and function. We investigated the effects of CR on taste bud morphology and expression of sweet taste–related modulators in 5-, 17-, and 30-month-old rats. In ad libitum (AL) and CR rats, we consistently found the following parameters altered significantly with advancing age: reduction of taste bud size and taste cell numbers per taste bud and reduced expression of sonic hedgehog, type 1 taste receptor 3 (T1r3), α-gustducin, and glucagon-like peptide-1 (GLP-1). In the oldest rats, CR affected a significant reduc-tion of tongue T1r3, GLP-1, and α-gustducin expression compared with age-matched AL rats. Leptin receptor immuno-positive cells were elevated in 17- and 30-month-old CR rats compared with age-matched AL rats. These alterations of sweet taste–related modulators, specifically during advanced aging, suggest that sweet taste perception may be altered in response to different lengths of CR.
Key Words: Taste buds—Sweet taste—Calorie restriction—T1r3 —Aging.
Received March 19, 2013; Accepted July 2, 2013
Decision Editor: David Le Couteur, PhD
TASTE perception begins with chemical activation of oral taste cells, which share many physical characteris-
tics with neurons (1). Upon activation by tastant molecules, transmitters are released by taste cells to activate associated gustatory nerve fibers. Taste cells are organized into onion-shaped structures known as taste buds, within three types of papillae in the tongue including the circumvallate, foliate, and fungiform papillae. Taste cells can be classified accord-ing to four types. Type I cells, the most abundant cells present in taste buds, are recognized as glial-like cells that maintain taste bud structure and terminate synaptic trans-mission. Nucleoside triphosphate diphosphohydrolase-2 that is strongly expressed in type I taste cells is a plasma membrane–bound nucleotidase that hydrolyzes extracellu-lar ATP, one of the multiple neurotransmitter in taste buds (2). Type II cells, also known as receptor cells, express many taste-related G protein–coupled receptors, which are responsible for sweet, bitter, and umami taste detection (3).
The sweet taste receptor is composed of the taste receptor type 1 member 2 (T1r2) and T1r3 heterodimer, whereas the umami receptor contains T1r1 and T1r3 (4). When the sweet taste receptor (T1r2–T1r3) is stimulated, phospholi-pase C-beta 2 (PLC-β2) is activated by βγ subunits of the G protein. The resulting inositol trisphosphate liberation yields intracellular calcium elevation, and action potentials are eventually evoked in the receptor cells (5). Type III cells are considered as presynaptic cells, which express synaptic proteins and show depolarization-dependent Ca2+ transients (6,7). A number of neuronal-like markers, including neural cell adhesion molecule, are expressed in these cells. Type IV cells are nonpolarized, undifferentiated basal cells (8). Taste bud cells that have an average life span of approxi-mately 10 days are constantly generated from these basal cells. Sonic hedgehog (Shh), which is required for cell growth and differentiation, is exclusively expressed in these basal cells (9).
by guest on June 16, 2016http://biom
edgerontology.oxfordjournals.org/D
ownloaded from

Page 2 of 13 CAI Et AL.
The gustatory system and taste perception are considered a gateway for controlling food intake and maintaining energy homeostasis. An increasing body of literature suggests that food preference and taste perception are tightly linked to body adiposity and metabolic state (10–12). For example, specific populations of African Americans that are prone to obesity have been shown to have an elevated desire for sweet diets (13). Overweight or obese individuals often perceive sweet tastes as less intense and may be more attracted to sweet-tasting agents (14). Furthermore, in animal studies, cholecystokinin type A receptor–deficient Otsuka Long-Evans Tokushima fatty rats display an increased preference for sucrose compared with lean control rats, and this effect was partially influenced by the orosensory stimulating effect of sucrose (15). The taste neurons in Otsuka Long-Evans Tokushima fatty rats possess augmented responses to higher sucrose concentrations (16). Similarly, high-fat diet-induced obese Sprague–Dawley rats were reported to shift sucrose and corn oil preferences toward higher concentrations com-pared with chow-fed lean controls (17). Conversely, food cravings, especially the craving of food high in sugar and/or fat, were revealed to be reduced in situations of calorie restriction (CR) and weight loss (18,19). Along with food preferences, taste perception has also been demonstrated to be related to body mass index and CR-induced weight loss. Esses and Herman (20) found that dieters rated 20% and 40% sucrose solutions less pleasant than nondieters both before and after an oral glucose load. However, perhaps due to the difference of metabolic status of the participants (eg, obese or nonobese) and terms of weight loss program, a recent study found that following 12 weeks of a weight loss program, obese Japanese women displayed improved sweet taste sensitivity (21). Despite being a simple lifestyle inter-vention, CR has been demonstrated to exert numerous bene-ficial effects on a wide variety of organ systems. Along with reducing body weight, CR has been shown to improve meta-bolic function, increase insulin sensitivity, extend life span, and delay the onset or reduce the emergence of age- associated diseases (22–29). CR can also induce neuroendocrine adaptations that may be involved in the antiaging-related effects of CR, such as reducing anabolic hormones, altering hormones regulating thermogenesis, and cellular metabo-lism as well as increasing hormones that suppress inflam-mation (30,31). At the present time, however, relatively little is known about taste bud morphology and the expression of taste modulating hormones in the taste cells of short- and long-term calorie-restricted rats. As increasing evidence suggests that taste perception, especially sweet taste per-ception, is highly related to functional metabolic status and body adiposity, we assessed whether CR affected taste bud morphology and the expression of sweet taste modulators in taste cells of 5-, 17-, and 30-month-old CR rats. In brief, our findings suggested that CR could enhance the age-driven reduction in expression of multiple sweet taste–related fac-tors in the tongue.
Materials and Methods
Animal and tissue ProcessingMale Fischer-344 rats used in this study were part of a
larger cohort of rats that were bred, reared, and housed in a vivarium maintained at the Gerontology Research Center, National Institute on Aging, National Institutes of Health, Baltimore, MD (32). All animal procedures and husbandry were approved by the Animal Care and Use Committee of the National Institute on Aging (Animal Protocol number 205-TJB). Rats were divided into two groups, one with ad libitum (AL) access to food and another receiving CR. As described previously, rats in the AL group were sup-plied with NIH-31/NIA standard rodent chow, whereas CR rats were fed with NIH-31/NIA fortified rodent chow (Harlan Teklad) (33). The macronutrient contents of these two chows are listed in Table 1. The CR paradigm was initiated at 2 months of age and a sequential 10% reduc-tion in food allocation was performed every 2 weeks until 40% CR was reached. Therefore, by 4 months of age, the animals were on 40% CR for the rest of their life. Animals of both AL and CR groups were euthanized at the age of 5, 17, and 30 months of age using isoflurane overdose and the tongues were collected from each animal. Excised tongues were fixed in 4% paraformaldehyde (Sigma Aldrich, St Louis, MO) for 1 hour and then cryoprotected with 20% sucrose in 0.1 M phosphate buffer overnight at 4°C. Serial sections (8–10 μm thickness) were cut from the tissues containing circumvallate papillae using a cry-ostat (HM 500M, MICRON, GmbH, Germany). In order to obtain a systematic appreciation without bias of the entire papillae, each papilla was sectioned and every 10th section was saved onto a slide. As taste buds are approxi-mately 80–100 μm in length, sampling every 10th section ensured that no two sections were from the same taste bud. Nonfasting glucose levels were measured using the EasyGluco blood glucose system (US Diagnostics, Inc., New York, NY). Nonfasting insulin levels were measured by commercial rat insulin ELISA Kit (Crystal Chem, Inc., Downers Grove, IL).
ImmunohistochemistryFollowing antigen retrieval with 1× citrate buffer
(Biogenex, San Ramon, CA) at 98°C for 20 min, immuno-fluorescence analyses were performed as described previ-ously (34). Cryostat sections were blocked in 5% bovine serum albumin (Sigma) and 0.1% Tween-20 in 1× Tris-buffered saline (pH 7.4) for 1 hour at room temperature, followed by incubation in a specific primary antibody in 1% bovine serum albumin and 0.1% Tween-20 in Tris-buffered saline (pH 7.4) overnight at 4°C. Sources and dilutions of the applied primary antibodies are listed in Table 2 (35,36). After washing, sections were incubated for 1 hour in fluo-rescent secondary antibodies (1:1,000 dilution; Invitrogen,
by guest on June 16, 2016http://biom
edgerontology.oxfordjournals.org/D
ownloaded from
ALtERAtIONS IN CALORIE-REStRICtED tAStE BuDS Page 3 of 13
Grand Island, NY) along with 4′,6-diamidino-2-phenylin-dole (1:5,000 dilution; Invitrogen) for nuclear staining. No fluorescent staining was observed in any sections when the primary antibodies were omitted.
Quantification of Immunoreactive taste CellsRat taste bud images were collected using an LSM-
710 confocal microscope (Carl Zeiss MicroImaging, Thornwood, NY) in single planes. As described previously, cells were scored as immunoreactive only if a nuclear pro-file was present within the cell (34,37). The total number of cells in the section was determined by counting the number of 4′,6-diamidino-2-phenylindole-stained nuclei present in each taste bud. Finally, the percentage of immunoreactive taste cells was calculated by dividing the number of immu-noreactive taste cells by the total number of the taste cells in each taste bud. Both image capture and data analysis were performed by trained researchers who were blind to the experimental and control conditions.
Quantification of taste Bud Size and taste Cell Numbers per taste Bud
Taste bud sizes were calculated according to previous methods (34). In brief, the perimeters of the taste bud from every 10th section were outlined and the corresponding area was computed by the Zeiss LSM image browser software. Simultaneously, 20 taste buds were randomly selected at different regions of each animal tongue to count the num-ber of cells in a single taste bud, where one nucleus corre-sponded to one cell on the section. Taste cell numbers were calculated by average taste cells per taste bud.
Statistical AnalysesAll data represent means ± SEM from at least three inde-
pendent experimental replicates. Error bars on graphs rep-resent the ±95% confidence interval. Two-way analysis of variance (ANOVA) with the Bonferroni post-test was per-formed by GraphPad Prism (version 5.0) as appropriate. To assess the effect of time in our study, one-way ANOVA repeated measures with the Bonferroni post-test were also used to analyze body weight, blood glucose, and insulin levels in AL and CR groups separately. The 5-month time point was regarded as the control in each diet paradigm. p < .05 was considered statistically significant throughout the study.
Results
Age- and CR-Related Alterations in Body Weight, Blood Glucose, and Insulin Levels
As expected, CR rats possessed a significantly lower body weight than the AL rats across all the ages (Figure 1A). Compared with 5-month-old AL rats, 17- and 30-month-old AL rats exhibited a significant increase in body weight (p < .01, 17-month-old AL vs 5-month-old AL; p < .01, 30-month-old AL vs 5-month-old AL with both one-way and two-way ANOVA). Statistical significance was con-firmed using one-way ANOVA repeated measures to assess
Table 1. Average Nutrient Composition of the Diets
Nutrient Composition UnitNIH-31/NIA
Fortified NIH-31/NIA
Protein % 18.74 18.42Fat % 4.41 4.47Fiber % 4.58 4.05Ash % 6.51 6.64Nitrogen-free extract % 55.04 55.91Gross energy kcal/g 3.95 4.02Digestible energy kcal/g 3.36 —Metabolizable energy kcal/g 3.07 —Linoleic acid % 1.79 —Moisture % — 10.51Amino acids Arginine % 1.1 1.06 Methionine % 0.36 0.39 Histidine % 0.42 0.41 Leucine % 1.5 1.61 Lysine % 0.96 0.95 Tryptophan % 0.22 0.24 Valine % 0.88 0.96 Cystine % 0.26 0.28 Isoleucine % 0.76 0.9 Threonine % 0.71 0.71 Pherylalanine % 1.53 0.92 +Tyrosine % — 0.7Minerals Calcium % 1.03 1.06 Phosphorus % 0.93 0.92 Sodium % 0.3 0.26 Chlorine % 0.48 0.42 Potassium % 0.59 0.59 Magnesium % 0.2 0.2 Iron mg/kg 336.41 300.2 Manganese mg/kg 156.01 152.8 Zinc mg/kg 48.41 50.4 Copper mg/kg 13.28 13.2 Iodine mg/kg 2.01 1.94 Cobalt mg/kg 0.53 0.53 Selenium mg/kg 0.3 —Vitamins Vitamin A IU/g 40.49 — Vitamin A3 IU/g — 30.73 Vitamin D3 IU/g 7 4.19 Vitamin E mg/kg 52.15 38.3 Choline mg/g 2.6 1.96 Niacin mg/kg 116.16 92.2 Pantothenic acid mg/kg 55.07 39.5 Pyridoxine mg/kg 13.16 10.2 Riboflavin mg/kg 11.04 7.8 Thiamine mg/kg 123.44 77.3 Menadione mg/kg 111.01 22 Folic acid mg/kg 2.13 1.7 Biotin mg/kg 0.38 0.13 Vitamin B12 mcg/kg 93.8 53 Vitamin C mg/g — — Carotene mg/kg — —
by guest on June 16, 2016http://biom
edgerontology.oxfordjournals.org/D
ownloaded from

Page 4 of 13 CAI Et AL.
the effect of time. The body weight of 17-month-old CR rats was significantly elevated compared with the 5-month-old CR rats, but 30-month-old CR rats possessed compa-rable mean body weight to 5-month-old CR rats (p < .01, 17-month-old CR vs 5-month-old CR with both one-way and two-way ANOVA). CR rats also showed significantly lower blood glucose and insulin levels when compared with AL rats at all age points (Figure 1B and C). Compared with 5-month-old AL rats, 17- and 30-month-old AL rats possessed a significantly progressive elevation in blood
glucose levels, whereas in CR rats, mean glucose levels were increased in 30-month-old rats compared with 5-month-old rats (p < .01, 17-month-old AL vs 5-month-old AL; p < .01, 30-month-old AL vs 5-month-old AL; p < .01, 30- month-old CR vs 5-month-old CR with both one-way and two-way ANOVA; Figure 1B). When compared with 5-month-old AL rats, the 17-month-old AL rats displayed a significant increase in insulin levels, whereas 30-month-old AL rats showed a significant decrease in insulin levels. However, mean insulin levels were only elevated in 30-month-old
Figure 1. Body weight, glucose, and insulin levels in 5-, 17-, and 30-month-old ad libitum (AL) and calorie restriction (CR) rats. (A) Mean body weight of 5-, 17-, and 30-month-old AL and CR rats. (B) Mean glucose levels of 5-, 17-, and 30-month-old AL and CR rats. (C) Mean insulin levels of 5-, 17-, and 30-month-old AL and CR rats. Values are expressed as means ± SEM. *p < .05; **p < .01.
Table 2. Primary Antibodies Used in Immunofluorescence Analyses
Antigen Host Vendor Dilution
Nucleoside triphosphate diphosphohydrolase-2
Rabbit Provided by Dr Sévigny, University of Laval, Quebec (http://www.ectonucleotidases-ab.com)
1:1,000
Phospholipase Cβ2 Rabbit Santa Cruz Biotechnology 1:200
Neural cell adhesion molecule
Rabbit Millipore 1:500
Sonic hedgehog Rabbit Santa Cruz Biotechnology 1:100Type 1 taste receptor 3 Goat Santa Cruz Biotechnology 1:100
α-Gustducin Rabbit Santa Cruz Biotechnology 1:200
Leptin receptor (Ob-Rb) Goat Abcam 1:500Glucagon-like peptide-1 Mouse USBiological 1:100
by guest on June 16, 2016http://biom
edgerontology.oxfordjournals.org/D
ownloaded from
ALtERAtIONS IN CALORIE-REStRICtED tAStE BuDS Page 5 of 13
CR rats compared with 5-month-old CR rats (p < .01, 17- month-old AL vs 5-month-old AL; p < .01, 30-month-old AL vs 5-month-old AL; p < .05, 30-month-old CR vs 5-month-old CR with both one-way and two-way ANOVA; Figure 1C). Hence, application of the CR paradigm was able to largely prevent the age-related increases in rat body weight as well as mean glucose and insulin levels, demonstrating its beneficial effects on metabolic health during aging.
Age-Related but Not CR-Related Alterations in taste Bud Size and Number of taste Cells per taste Bud
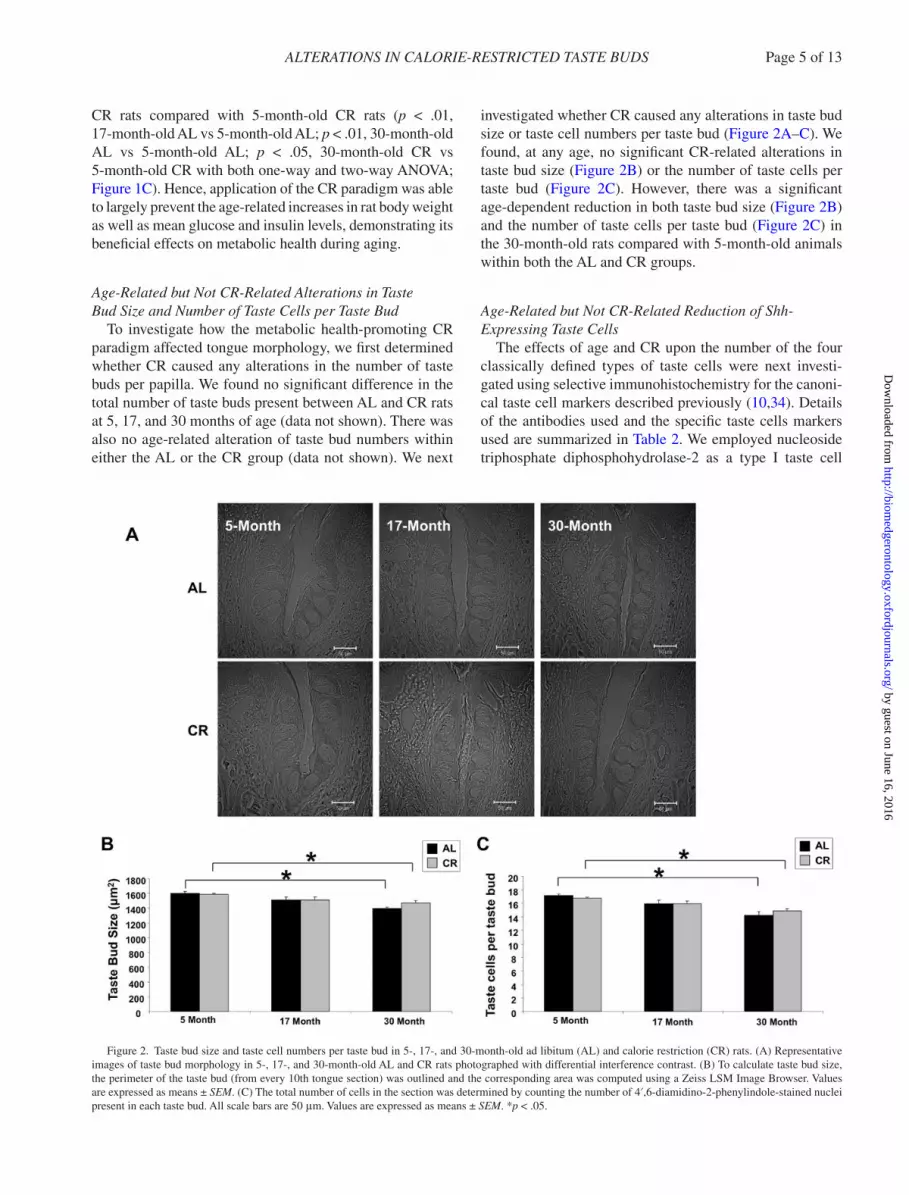
To investigate how the metabolic health-promoting CR paradigm affected tongue morphology, we first determined whether CR caused any alterations in the number of taste buds per papilla. We found no significant difference in the total number of taste buds present between AL and CR rats at 5, 17, and 30 months of age (data not shown). There was also no age-related alteration of taste bud numbers within either the AL or the CR group (data not shown). We next
investigated whether CR caused any alterations in taste bud size or taste cell numbers per taste bud (Figure 2A–C). We found, at any age, no significant CR-related alterations in taste bud size (Figure 2B) or the number of taste cells per taste bud (Figure 2C). However, there was a significant age-dependent reduction in both taste bud size (Figure 2B) and the number of taste cells per taste bud (Figure 2C) in the 30-month-old rats compared with 5-month-old animals within both the AL and CR groups.
Age-Related but Not CR-Related Reduction of Shh-Expressing taste Cells
The effects of age and CR upon the number of the four classically defined types of taste cells were next investi-gated using selective immunohistochemistry for the canoni-cal taste cell markers described previously (10,34). Details of the antibodies used and the specific taste cells markers used are summarized in Table 2. We employed nucleoside triphosphate diphosphohydrolase-2 as a type I taste cell
Figure 2. Taste bud size and taste cell numbers per taste bud in 5-, 17-, and 30-month-old ad libitum (AL) and calorie restriction (CR) rats. (A) Representative images of taste bud morphology in 5-, 17-, and 30-month-old AL and CR rats photographed with differential interference contrast. (B) To calculate taste bud size, the perimeter of the taste bud (from every 10th tongue section) was outlined and the corresponding area was computed using a Zeiss LSM Image Browser. Values are expressed as means ± SEM. (C) The total number of cells in the section was determined by counting the number of 4′,6-diamidino-2-phenylindole-stained nuclei present in each taste bud. All scale bars are 50 μm. Values are expressed as means ± SEM. *p < .05.
by guest on June 16, 2016http://biom
edgerontology.oxfordjournals.org/D
ownloaded from

Page 6 of 13 CAI Et AL.
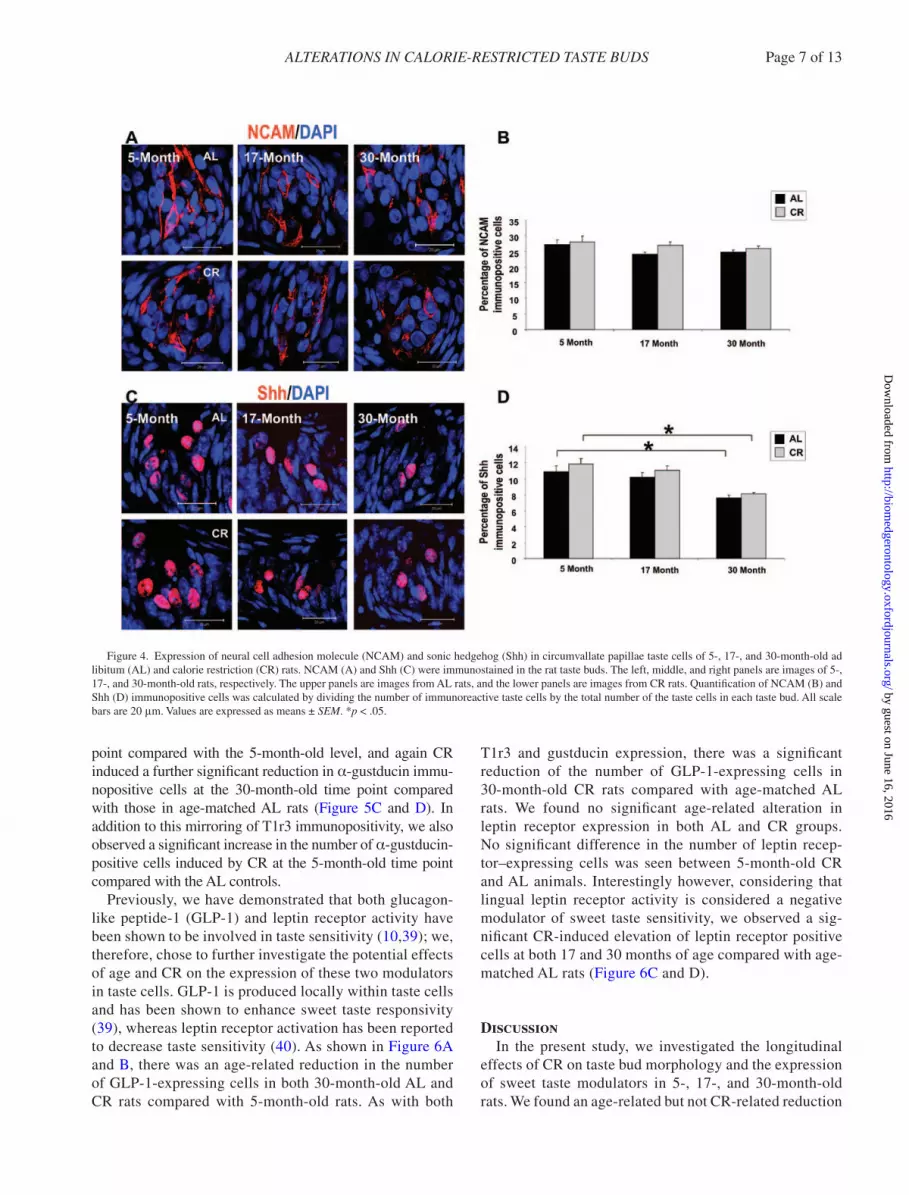
marker, PLC-β2 as a type II taste cell marker, neural cell adhesion molecule as a type III taste cell marker, and Shh as a type IV taste cell marker. There were no significant CR- or age-related alterations in nucleoside triphosphate diphosphohydrolase-2-, PLC-β2-, or neural cell adhesion molecule–expressing cells (Figures 3 and 4). In addition, there were no significant differences in Shh-expressing cells between 5-, 17-, and 30-month-old AL and CR rats. However, as shown in Figure 4D, there was a significant reduction of Shh-expressing cells in the 30-month-old ani-mals compared with the 5-month-old animals in both AL and CR groups.
Age- and CR-Related Alterations in Expression of the Sweet taste Regulators
As neither age nor CR particularly affected the total number of type I, II, or III cells, and only exerted small effects on type
IV (Shh-positive basal cells) numbers, we next investigated whether these various taste cells were able to express markers of the sweet taste transduction system. The T1r3 subunit, a critical component of the heterodimeric sweet taste receptor T1r2/T1r3, plays an important role in sweet taste responsiv-ity, for example, T1r3 knockout mice displayed diminished response to natural sugars and abolished response to artifi-cial sweeteners (3,4,38). Between the ages 5 and 17 months, minimal differences in the expression of T1r3 were observed (Figure 5A and B); however, a significant reduction in the number of T1r3 immunopositive cells was observed between the 5- and 30-month-old time points. In addition to this, at this most advanced age time point, CR affected an additional significant reduction of T1r3 immunopositive cells compared with the age-matched AL rats (Figure 5B). As expected, due to the strong mechanistic connection between T1r3 and α-gustducin, a similar significant reduction of α-gustducin immunopositive cells was only observed at the oldest time
Figure 3. Expression of nucleoside triphosphate diphosphohydrolase-2 (NTPDase2) and phospholipase C-beta 2 (PLC-β2) in circumvallate papillae taste cells of 5-, 17-, and 30-month-old ad libitum (AL) and calorie restriction (CR) rats. NTPDase2 (A) and PLC-β2 (C) were immunostained in the rat taste buds. The left, middle, and right panels are images of 5-, 17-, and 30-month-old rats, respectively. The upper panels are images from AL rats, and the lower panels are images from CR rats. Quantification of NTPDase2 (B) and PLC-β2 (D) immunopositive cells was calculated by dividing the number of immunoreactive taste cells by the total number of the taste cells in each taste bud. All scale bars are 20 μm. Values are expressed as means ± SEM.
by guest on June 16, 2016http://biom
edgerontology.oxfordjournals.org/D
ownloaded from
ALtERAtIONS IN CALORIE-REStRICtED tAStE BuDS Page 7 of 13
point compared with the 5-month-old level, and again CR induced a further significant reduction in α-gustducin immu-nopositive cells at the 30-month-old time point compared with those in age-matched AL rats (Figure 5C and D). In addition to this mirroring of T1r3 immunopositivity, we also observed a significant increase in the number of α-gustducin-positive cells induced by CR at the 5-month-old time point compared with the AL controls.
Previously, we have demonstrated that both glucagon-like peptide-1 (GLP-1) and leptin receptor activity have been shown to be involved in taste sensitivity (10,39); we, therefore, chose to further investigate the potential effects of age and CR on the expression of these two modulators in taste cells. GLP-1 is produced locally within taste cells and has been shown to enhance sweet taste responsivity (39), whereas leptin receptor activation has been reported to decrease taste sensitivity (40). As shown in Figure 6A and B, there was an age-related reduction in the number of GLP-1-expressing cells in both 30-month-old AL and CR rats compared with 5-month-old rats. As with both
T1r3 and gustducin expression, there was a significant reduction of the number of GLP-1-expressing cells in 30-month-old CR rats compared with age-matched AL rats. We found no significant age-related alteration in leptin receptor expression in both AL and CR groups. No significant difference in the number of leptin recep-tor–expressing cells was seen between 5-month-old CR and AL animals. Interestingly however, considering that lingual leptin receptor activity is considered a negative modulator of sweet taste sensitivity, we observed a sig-nificant CR-induced elevation of leptin receptor positive cells at both 17 and 30 months of age compared with age-matched AL rats (Figure 6C and D).
DiscussionIn the present study, we investigated the longitudinal
effects of CR on taste bud morphology and the expression of sweet taste modulators in 5-, 17-, and 30-month-old rats. We found an age-related but not CR-related reduction
Figure 4. Expression of neural cell adhesion molecule (NCAM) and sonic hedgehog (Shh) in circumvallate papillae taste cells of 5-, 17-, and 30-month-old ad libitum (AL) and calorie restriction (CR) rats. NCAM (A) and Shh (C) were immunostained in the rat taste buds. The left, middle, and right panels are images of 5-, 17-, and 30-month-old rats, respectively. The upper panels are images from AL rats, and the lower panels are images from CR rats. Quantification of NCAM (B) and Shh (D) immunopositive cells was calculated by dividing the number of immunoreactive taste cells by the total number of the taste cells in each taste bud. All scale bars are 20 μm. Values are expressed as means ± SEM. *p < .05.
by guest on June 16, 2016http://biom
edgerontology.oxfordjournals.org/D
ownloaded from

Page 8 of 13 CAI Et AL.
in both taste bud size and taste cell number per taste bud. Taste cells typically experience continuous turnover, with an individual life span of approximately 9–12 days (41). The epithelial cells surrounding the taste bud divide and some of the daughter cells enter the taste bud and slowly move into the center (42). A growing body of evidence, however, indicates that this dynamic maintenance of taste cells tends to deteriorate with advancing age (43–45). Aged mice demonstrate delayed cell renewal and a highly vacuolated cytoplasm in their taste buds compared with young adult mice (46). Reduced canonical Wnt signal-ing in older mice has been demonstrated to be involved in loss of taste sensitivity with aging (47). We have recently also demonstrated that a significant reduction of taste bud size and taste cell numbers per bud is found in older mice compared with younger control animals (34). Our observation of significant age-related reductions of Shh-expressing taste cells (Figure 4) may, therefore, be strongly associated with this observation of reduced taste bud size and taste cell numbers with age.
In addition, there are reports of several studies investi-gating papilla size, papilla numbers, and taste bud numbers in aged rodents and humans. A 30% decrease in total taste bud number in the circumvallate papillae has been reported in aged mice (48). However, in rats, Mistretta and Baum (49) found that both fungiform and circumvallate papillae were larger in 24-month-old Wistar-derived rats than that of 6-month-old rats and the circumvallate papillae in older rats were deeper and longer but with the same width. They also demonstrated no difference in taste bud number and size in aged rats. In our study, we observed a decreased taste bud size in 30-month-old Fischer-344 rats compared with the 5-month-old rats, which distinguishes our data from the results of the Mistretta and Baum’s study. This discrepancy may due to the difference of rat strain, age, and diverse methods for calculating taste bud size. Further investiga-tions from other independent research groups will likely help resolve these issues in the future. Currently, there is a limited literature base concerning the study of taste bud size in aged human circumvallate papillae. It has though been
Figure 5. Expression of T1r3 and α-gustducin in circumvallate papillae taste cells of 5-, 17-, and 30-month-old ad libitum (AL) and calorie restriction (CR) rats. T1r3 (A) and α-gustducin (C) were immunostained in the rat taste buds. The left, middle, and right panels are images of 5-, 17-, and 30-month-old rats, respectively. The upper panels are images from AL rats, and the lower panels are images from CR rats. Quantification of T1r3 (B) and α-gustducin (D) immunopositive cells was calculated by dividing the number of immunoreactive taste cells by the total number of the taste cells in each taste bud. All scale bars are 20 μm. Values are expressed as means ± SEM. *p < .05.
by guest on June 16, 2016http://biom
edgerontology.oxfordjournals.org/D
ownloaded from

ALtERAtIONS IN CALORIE-REStRICtED tAStE BuDS Page 9 of 13
reported that in aged human circumvallate papillae, the taste bud number decreased by about 50% after 74 years (50). In another study of circumvallate papillae in a Turkish popula-tion, the authors found no significant relationship between diameters of circumvallate papillae and age by direct naked eye observation using a wooden tongue depressor, a gauge, and a light source (51). Shimizu (52) demonstrated that for human circumvallate papillae, the taste bud and taste bud cell density in the older age groups were lower than that of the younger age groups. In general, the data of aged human circumvallate papillae are quite variable due to variations in methods, data analysis, freshness, and pathological degree of specimens. As with the variation in experimental data, further independent research into these age-related factors will likely resolve these inconsistencies.
A significant corpus of literature exists that demonstrates a strong link between the craving of foodstuffs high in sugar and body adiposity/energy homeostasis (10,12,53,54). As the sweet taste perceptive modality plays a crucial role in the desire for sugary foodstuffs, we investigated the effects
of CR, a well-known controller of adiposity/metabolic sta-tus, on sweet taste perception at a cellular level. We found that the number of T1r3-, α-gustducin-, and GLP-1-positive cells was significantly reduced, whereas the number of leptin receptor (implicated in attenuation of sweet taste responsivity) positive cells was increased in 30-month-old calorie-restricted rats compared with age-matched AL rats. Indicative of the coherent systematic taste cell alterations we observed in this study, it has been previously shown that T1r3 knockout mice possess diminished preference and peripheral nerve responses to sugars and umami compounds (3,38); α-gustducin knockout mice display decreased pref-erence and attenuated nerve response to sucrose and other artificial sweeteners (55); GLP-1 receptor knockout mice possess a significantly reduced sensitivity to sweet tastant molecules (39) and leptin receptor–deficient mice demon-strate enhanced neural responses and behavioral preferences to sweet stimuli (40). Therefore, in our current study, we have found that aging may mediate the previously observed reduction in sweet taste perception (34) via simultaneous
Figure 6. Expression of Leptin receptor and GLP-1 in circumvallate papillae taste cells (TCs) of 5-, 17-, and 30-month-old ad libitum (AL) and calorie restric-tion (CR) rats. GLP-1 (A) and leptin receptor (C) were immunostained in the rat taste buds. The left, middle, and right panels are images of 5-, 17-, and 30-month old rats, respectively. The upper panels are images from AL rats, and the lower panels are images from CR rats. Quantification of GLP-1 (B) and leptin receptor (D) immunopositive cells was calculated by dividing the number of immunoreactive taste cells by the total number of the taste cells in each taste bud. All scale bars are 20 μm. Values are expressed as means ± SEM. *p < .05.
by guest on June 16, 2016http://biom
edgerontology.oxfordjournals.org/D
ownloaded from

Page 10 of 13 CAI Et AL.
reductions in the expression of the T1r3, α-gustducin, and GLP-1. Since T1r3 is also the subunit of the heteromeric umami taste receptor, the umami taste perception may be potentially changed with CR. In addition, the alteration of T1r3 during the aging process might also be correlated to the evidence of changed monosodium glutamate taste perception in elderly participants compared with young participants (56,57). For each of these crucial sweet taste sensitivity loci, we found that CR induced an additional enhancement of these “default” age-related changes; hence, it appears in our experimental paradigm that CR is modu-lating the natural aging-induced attenuation of sweet taste perception. One potential hypothesis that may explain this observation is that naturally animals reduce their desire for carbohydrate-rich foodstuffs with age to prevent excessive age-related insulin resistance. Most animals and humans demonstrate an increase in insulin resistance and diabetic-related pathophysiology as they age. We have demonstrated previously that the presence of diabetes-like pathologies can induce compensatory changes in lingual sweet taste perception (10). These changes seem to occur coherently to modulate sweet taste perception, thereby potentially reduc-ing further sweet tastant ingestion that would further exac-erbate the diabetic pathology. In this current study, we have demonstrated that at the molecular level, a largely similar “program” of molecular changes also occurs with natural aging, that is, reductions in T1r3, α-gustducin, and GLP-1, with a concomitant elevation of leptin receptor expression. Thus, we contend that lingual reductions in T1r3, gust-ducin, and GLP-1, factors that promote and support sweet taste responsivity, are part of a natural “program” that pre-vents excessive sugar ingestion in older age, a state in which general insulin resistance/diabetes is commonplace. In this manner, CR accelerates this ameliorate “program,” that is, accentuating the divergence away from sugary, high-calo-rie foodstuffs in old age. In agreement with this, we also observed an increase in lingual leptin receptor expression that also serves to suppress excessive sweet taste responsiv-ity in old age. In our study, therefore, we do indeed see the positive effects of CR in an aging “program” setting.
Brought together, all of these molecular sequelae are likely to reduce sweet taste perception. For each of these indices, the addition of CR to the aging process further accelerates this potential ameliorative “program” opposing the inevitable age-related diabetic-related pathology, via an effective reduction in sweet taste perception (34). There is a considerable body of evidence showing that CR delays many diseases associated with advanced aging including cancer, diabetes, atherosclerosis, cardiovascular disease, and neurodegenerative diseases (23,58–60). For example, CR can activate SIRT3 to reduce oxidative stress and com-bat aging (61,62). CR also has been shown to downregu-late the insulin/IGF-1/mTOR pathways, which in turn can activate additional antiaging signaling systems (63–65). Furthermore, CR has been shown to enhance autophagy and
DNA repair systems, which decline during the aging pro-cess (66,67). Thus, one of the many aging-pathology ame-liorative mechanisms of CR may be through a concerted effect upon taste cell functional integrity.
There is some evidence that the potential CR-mediated sweet taste–depressive effect occurs in humans, for exam-ple, the craving of high-sugar food reportedly diminishes with CR in some human studies (18,19,68), whereas human CR dieters rated sucrose solutions less pleasant compared with nondieting participants (20). As a caveat, however, as with most complex lifestyle behavioral experiments, there are some findings inconsistent with this hypothesis. For example, high-fat diet-induced obese rats have dem-onstrated decreased preference to saccharin (69), whereas 6 weeks diet-restricted rats showed the opposite response (69) and short-term CR has been reported to associate with increased sweet and salty taste sensitivity (70). These dif-ferent observations may be related to different terms of the CR regimen. The CR regimen administered in our current study was initiated postpubescently to avoid any develop-mental effects, and by 4 months of age, the animals were on 40% CR for the rest of their life. Thus, 5-month-old CR rats can be considered as a model of short-term CR, whereas 30-month-old CR rats can be considered as a model of long-term CR. In the current study, the number of α-gustducin-positive cells was significantly elevated in 5-month-old CR rats compared with age-matched AL rats. There was also a nonsignificant trend for increases in T1r3- and GLP-1-expressing cells in 5-month-old CR rats compared with age-matched AL rats. α-Gustducin, which is primarily a taste-specific G protein alpha-subunit, can activate a taste-specific phosphodiesterase, lowering the cellular concentra-tion of cyclic nucleotides (71). Although α-gustducin, T1r3, GLP-1, and leptin receptors are all involved in sweet taste modulation, it is highly likely that the specific molecular mechanisms that each employ to facilitate sweet taste per-ception will be specific and distinct to each molecule. In 5-month-old CR rats, the animals were involved in 40% CR for 1 month, in contrast to animals that received the diet until 17 and 30 months of age, respectively. The evidence that both messenger RNA and protein levels of α-gustducin are altered in diabetic rat taste bud cells suggest that α-gustducin is closely correlated to the body’s metabolic state (72). Therefore, α-gustducin, the taste-specific G pro-tein alpha-subunit, might be more sensitive to acute calo-rie deprivation (1 month) other than other components (eg, T1r3, GLP-1, and leptin receptor) of the sweet taste percep-tive function. Therefore, this short-term diet paradigm may exert distinct functional actions upon α-gustducin, T1r3, GLP-1, and leptin receptor signaling modalities.
After stimulation of tastant G protein–coupled recep-tors by taste stimuli, two parallel streams of intracellular events can be triggered. α-Gustducin can activate a taste-specific phosphodiesterase, thus altering cellular concen-trations of cyclic nucleotides. PLC-β2 can be activated by
by guest on June 16, 2016http://biom
edgerontology.oxfordjournals.org/D
ownloaded from

ALtERAtIONS IN CALORIE-REStRICtED tAStE BuDS Page 11 of 13
the βγ subunit complex of G proteins (73), leading to the generation of intracellular messengers, which mobilize cal-cium and activate protein kinase C. PLC-β2 is expressed in a huge variety of cell and tissue types across the body. Although PLC-β2 is involved in transduction of multiple taste qualities including sweet, umami, and bitter tastes, it is also crucially involved in an enormous array of other non-tastant G protein–coupled receptor and other receptor-based signaling systems (74,75). In contrast to the near-ubiquitous nature of expression and multifactorial signaling activity of PLC-β2, the T1r3 taste receptor and α-gustducin pos-sess, in general, extremely limited expression profiles and are involved in highly specific actions, that is, tastant per-ception. In our study, we failed to observe any significant changes in PLC-β2 expression but instead we discovered significant age-specific diminution of the more specific fac-tors, T1r3 and α-gustducin. If indeed the molecular sweet taste perceptive changes we observed are part of some physiological age-related program, then the most selec-tive mechanism to alter this pathway would be to reduce the more taste-selective components of this systems, that is, T1r3 and α-gustducin, rather than alter levels of PLC-β2 that would affect far more signaling systems and therefore not achieve a selective age-related taste modification effect.
Interestingly, we found opposite alterations in the short-term and long-term CR rats. The number of T1r3-, α-gustducin-, and GLP-1-positive cells tended to reduce in 17-month-old CR rats compared with age-matched AL rats and these reductions were significant in the 30-month-old group. One potential explanation for this could be that with the initial onset of CR, the animals having no expectation of future chronic calorie deprivation respond by enhancing sweet taste to compensate for the “temporary” sweet defi-ciency. However, with the eventual chronic nature of the applied CR, the animals efficiently adapt to the long-term deficit of sweet tastants by attenuating the sensitivity of the seemingly “less environmentally important” sweet taste system.
As our study suggests, components of the sweet taste perception machinery, that is, the taste receptors and down-stream signaling elements, are tightly associated with energy homeostasis and metabolic status. For example, elevated taste cell α-gustducin has been observed in diabetic rats that possess increased body weight, blood glucose levels, and insulin levels (72). Also, our previous study found that vasoactive intestinal peptide knockout mice, which pos-sessed a diabetic-like phenotype, showed a greater prefer-ence for sweet compounds compared with wild-type mice. Additionally, vasoactive intestinal peptide knockout mice exhibited significant increases in taste cell GLP-1 expres-sion with a concomitant decrease in taste cell leptin receptor expression (10). Furthermore, alterations of taste modula-tors have also been demonstrated to be correlated with the type and term of diet regime, for example, gene transcript levels of T1r3 are significantly decreased in the high-fat
diet-induced obese rats (69). In that study, however, it was demonstrated that an applied short 6-week CR regimen increased expression levels of both T1r3 and α-gustducin (69), which was consistent with our observation of short-term CR, that is, the 5-month-old CR rats (Figure 5).
In this study, we have elucidated some potential molecu-lar mechanisms by which both CR and aging affect taste bud morphology and sweet taste functional integrity. These findings potentially reveal the presence of a natural age-related mechanistic basis for functional sweet taste modula-tion that we reported recently (34). Our data, therefore, have revealed a potentially novel locus for the beneficial actions of the CR lifestyle intervention. Further investigation into this potential aging–health–taste nexus may include meas-urement of taste sensitivities in animal models of advanced aging such as the SAMP8 mice (76) and animal models of metabolic dysfunction such as the N171-82Q Huntington’s disease mice (77) as well as potential investigations into the sweet taste reward systems in aging mice.
Funding
This work was supported entirely by the Intramural Research Program of the National Institute on Aging, National Institutes of Health (AG000916-01).
References 1. Chaudhari N, Roper SD. The cell biology of taste. J Cell Biol.
2010;190:285–296. 2. Bartel DL, Sullivan SL, Lavoie EG, Sévigny J, Finger TE. Nucleoside
triphosphate diphosphohydrolase-2 is the ecto-ATPase of type I cells in taste buds. J Comp Neurol. 2006;497:1–12.
3. Zhao GQ, Zhang Y, Hoon MA, et al. The receptors for mammalian sweet and umami taste. Cell. 2003;115:255–266.
4. Li X. T1R receptors mediate mammalian sweet and umami taste. Am J Clin Nutr. 2009;90:733S–737S.
5. Liu D, Liman ER. Intracellular Ca2+ and the phospholipid PIP2 regu-late the taste transduction ion channel TRPM5. Proc Natl Acad Sci uSA. 2003;100:15160–15165.
6. DeFazio RA, Dvoryanchikov G, Maruyama Y, et al. Separate popula-tions of receptor cells and presynaptic cells in mouse taste buds. J Neurosci. 2006;26:3971–3980.
7. Dvoryanchikov G, Tomchik SM, Chaudhari N. Biogenic amine synthe-sis and uptake in rodent taste buds. J Comp Neurol. 2007;505:302–313.
8. Farbman AI. Fine structure of the taste bud. J ultrastruct Res. 1965;12:328–350.
9. Miura H, Kusakabe Y, Sugiyama C, et al. Shh and Ptc are associ-ated with taste bud maintenance in the adult mouse. Mech Dev. 2001;106:143–145.
10. Martin B, Shin YK, White CM, et al. Vasoactive intestinal peptide-null mice demonstrate enhanced sweet taste preference, dysglyce-mia, and reduced taste bud leptin receptor expression. Diabetes. 2010;59:1143–1152.
11. Martin B, Maudsley S, White CM, Egan JM. Hormones in the naso-oropharynx: endocrine modulation of taste and smell. trends Endocrinol Metab. 2009;20:163–170.
12. Berthoud HR, Zheng H. Modulation of taste responsiveness and food preference by obesity and weight loss. Physiol Behav. 2012;107:527–532.
13. Schiffman SS, Graham BG, Sattely-Miller EA, Peterson-Dancy M. Elevated and sustained desire for sweet taste in African-Americans: a potential factor in the development of obesity. Nutrition. 2000;16:886–893.
by guest on June 16, 2016http://biom
edgerontology.oxfordjournals.org/D
ownloaded from

Page 12 of 13 CAI Et AL.
14. Sartor F, Donaldson LF, Markland DA, Loveday H, Jackson MJ, Kubis HP. Taste perception and implicit attitude toward sweet related to body mass index and soft drink supplementation. Appetite. 2011;57:237–246.
15. De Jonghe BC, Hajnal A, Covasa M. Increased oral and decreased intestinal sensitivity to sucrose in obese, prediabetic CCK-A recep-tor-deficient OLETF rats. Am J Physiol Regul Integr Comp Physiol. 2005;288:R292–R300.
16. Hajnal A, Norgren R, Kovacs P. Parabrachial coding of sapid sucrose: relevance to reward and obesity. Ann N Y Acad Sci. 2009;1170:347–364.
17. Shin AC, Townsend RL, Patterson LM, Berthoud HR. “Liking” and “wanting” of sweet and oily food stimuli as affected by high-fat diet-induced obesity, weight loss, leptin, and genetic predisposition. Am J Physiol Regul Integr Comp Physiol. 2011;301:R1267–R1280.
18. Martin CK, O’Neil PM, Pawlow L. Changes in food cravings dur-ing low-calorie and very-low-calorie diets. Obesity (Silver Spring). 2006;14:115–121.
19. Martin CK, Rosenbaum D, Han H, et al. Change in food cravings, food preferences, and appetite during a low-carbohydrate and low-fat diet. Obesity (Silver Spring). 2011;19:1963–1970.
20. Esses VM, Herman CP. Palatability of sucrose before and after glucose ingestion in dieters and nondieters. Physiol Behav. 1984;32:711–715.
21. Umabiki M, Tsuzaki K, Kotani K, et al. The improvement of sweet taste sensitivity with decrease in serum leptin levels during weight loss in obese females. tohoku J Exp Med. 2010;220:267–271.
22. Martin B, Mattson MP, Maudsley S. Caloric restriction and intermit-tent fasting: two potential diets for successful brain aging. Ageing Res Rev. 2006;5:332–353.
23. Anderson RM, Weindruch R. Metabolic reprogramming, caloric restriction and aging. trends Endocrinol Metab. 2010;21:134–141.
24. Johnson JB, Summer W, Cutler RG, et al. Alternate day calorie restric-tion improves clinical findings and reduces markers of oxidative stress and inflammation in overweight adults with moderate asthma. Free Radic Biol Med. 2007;42:665–674.
25. Harvie MN, Pegington M, Mattson MP, et al. The effects of inter-mittent or continuous energy restriction on weight loss and metabolic disease risk markers: a randomized trial in young overweight women. Int J Obes (Lond). 2010;35:714–727.
26. Stranahan AM, Martin B, Chadwick W, et al. Metabolic context regulates distinct hypothalamic transcriptional responses to antiaging interventions. Int J Endocrinol. 2012;2012:732975.
27. Martin B, Pearson M, Brenneman R, et al. Gonadal transcriptome alterations in response to dietary energy intake: sensing the reproduc-tive environment. PLoS One. 2009;4:e4146.
28. Martin B, Pearson M, Brenneman R, et al. Conserved and differential effects of dietary energy intake on the hippocampal transcriptomes of females and males. PLoS One. 2008;3:e2398.
29. Carlson O, Martin B, Stote KS, et al. Impact of reduced meal fre-quency without caloric restriction on glucose regulation in healthy, normal-weight middle-aged men and women. Metabolism. 2007;56:1729–1734.
30. Fontana L, Klein S. Aging, adiposity, and calorie restriction. JAMA. 2007;297:986–994.
31. Omodei D, Fontana L. Calorie restriction and prevention of age-asso-ciated chronic disease. FEBS Lett. 2011;585:1537–1542.
32. Hunt ND, Li GD, Zhu M, et al. Effect of calorie restriction and refeed-ing on skin wound healing in the rat. Age (Dordr). 2012;34:1453–1458.
33. Zhu M, de Cabo R, Anson RM, Ingram DK, Lane MA. Caloric restric-tion modulates insulin receptor signaling in liver and skeletal muscle of rat. Nutrition. 2005;21:378–388.
34. Shin YK, Cong WN, Cai H, et al. Age-related changes in mouse taste bud morphology, hormone expression, and taste responsivity. J Gerontol A Biol Sci Med Sci. 2012;67:336–344.
35. Sévigny J, Sundberg C, Braun N, et al. Differential catalytic prop-erties and vascular topography of murine nucleoside triphosphate
diphosphohydrolase 1 (NTPDase1) and NTPDase2 have implications for thromboregulation. Blood. 2002;99:2801–2809.
36. Heine P, Braun N, Sévigny J, Robson SC, Servos J, Zimmermann H. The C-terminal cysteine-rich region dictates specific catalytic proper-ties in chimeras of the ectonucleotidases NTPDase1 and NTPDase2. Eur J Biochem. 2001;268:364–373.
37. Shin YK, Martin B, Kim W, et al. Ghrelin is produced in taste cells and ghrelin receptor null mice show reduced taste responsivity to salty (NaCl) and sour (citric acid) tastants. PLoS One. 2010;5:e12729.
38. Damak S, Rong M, Yasumatsu K, et al. Detection of sweet and umami taste in the absence of taste receptor T1r3. Science. 2003;301:850–853.
39. Shin YK, Martin B, Golden E, et al. Modulation of taste sensitivity by GLP-1 signaling. J Neurochem. 2008;106:455–463.
40. Kawai K, Sugimoto K, Nakashima K, Miura H, Ninomiya Y. Leptin as a modulator of sweet taste sensitivities in mice. Proc Natl Acad Sci uSA. 2000;97:11044–11049.
41. Farbman AI. Renewal of taste bud cells in rat circumvallate papillae. Cell tissue Kinet. 1980;13:349–357.
42. Beidler LM, Smallman RL. Renewal of cells within taste buds. J Cell Biol. 1965;27:263–272.
43. Takeda M, Suzuki Y, Obara N, Nagai Y. Apoptosis in mouse taste buds after denervation. Cell tissue Res. 1996;286:55–62.
44. Imoscopi A, Inelmen EM, Sergi G, Miotto F, Manzato E. Taste loss in the elderly: epidemiology, causes and consequences. Aging Clin Exp Res. 2012;24:570–579.
45. Aliani M, Udenigwe CC, Girgih AT, Pownall TL, Bugera JL, Eskin MN. Zinc deficiency and taste perception in the elderly. Crit Rev Food Sci Nutr. 2013;53:245–250.
46. Fukunaga A. [Age-related changes in renewal of taste bud cells and expression of taste cell-specific proteins in mice]. Kokubyo Gakkai Zasshi. 2005;72:84–89.
47. Gaillard D, Barlow LA. Taste bud cells of adult mice are responsive to Wnt/β-catenin signaling: implications for the renewal of mature taste cells. Genesis. 2011;49:295–306.
48. Conger AD, Wells MA. Radiation and aging effect on taste structure and function. Radiat Res. 1969;37:31–49.
49. Mistretta CM, Baum BJ. Quantitative study of taste buds in fungiform and circumvallate papillae of young and aged rats. J Anat. 1984;138(Pt 2):323–332.
50. Arey LB, Tremaine MJ, Monzingo FL. The numerical and topographi-cal relations of taste buds to human circumvallate papillae throughout the life span. Anat Rec. 1935;64:9–25.
51. Mavi A, Ceyhan O. Bitter taste thresholds, numbers and diameters of circumvallate papillae and their relation with age in a Turkish popula-tion. Gerodontology. 1999;16:119–122.
52. Shimizu Y. A histomorphometric study of the age-related changes of the human taste buds in circumvallate papillae. Oral Med Pathol. 1997;2:17–24.
53. Miras AD, le Roux CW. Bariatric surgery and taste: novel mechanisms of weight loss. Curr Opin Gastroenterol. 2010;26:140–145.
54. Aguayo GA, Vaillant MT, Arendt C, Bachim S, Pull CB. Taste prefer-ence and psychopathology. Bull Soc Sci Med Grand Duche Luxemb. 2012:7–14.
55. Danilova V, Damak S, Margolskee RF, Hellekant G. Taste responses to sweet stimuli in alpha-gustducin knockout and wild-type mice. Chem Senses. 2006;31:573–580.
56. Schiffman SS, Sattely-Miller EA, Zimmerman IA, Graham BG, Erickson RP. Taste perception of monosodium glutamate (MSG) in foods in young and elderly subjects. Physiol Behav. 1994;56:265–275.
57. Mojet J, Heidema J, Christ-Hazelhof E. Taste perception with age: generic or specific losses in supra-threshold intensities of five taste qualities? Chem Senses. 2003;28:397–413.
58. Minor RK, Allard JS, Younts CM, Ward TM, de Cabo R. Dietary inter-ventions to extend life span and health span based on calorie restric-tion. J Gerontol A Biol Sci Med Sci. 2010;65:695–703.
by guest on June 16, 2016http://biom
edgerontology.oxfordjournals.org/D
ownloaded from

ALtERAtIONS IN CALORIE-REStRICtED tAStE BuDS Page 13 of 13
59. Ribarič S. Diet and aging. Oxid Med Cell Longev. 2012;2012:741468. 60. Higami Y, Pugh TD, Page GP, Allison DB, Prolla TA, Weindruch R.
Adipose tissue energy metabolism: altered gene expression profile of mice subjected to long-term caloric restriction. FASEB J. 2004;18:415–417.
61. Martin-Montalvo A, Cabo RD. Mitochondrial metabolic repro-gramming induced by calorie restriction. Antioxid Redox Signal. 2013;19:310–320.
62. Zhang D, Liu Y, Chen D. SIRT-ain relief from age-inducing stress. Aging (Albany NY). 2011;3:158–161.
63. Mercken EM, Crosby SD, Lamming DW, et al. Calorie restriction in humans inhibits the pi3k/akt pathway and induces a younger tran-scription profile. Aging Cell. 2013;12:645–651.
64. Salmon AB, Murakami S, Bartke A, Kopchick J, Yasumura K, Miller RA. Fibroblast cell lines from young adult mice of long-lived mutant strains are resistant to multiple forms of stress. Am J Physiol Endocrinol Metab. 2005;289:E23–E29.
65. Bartke A. Minireview: role of the growth hormone/insulin-like growth factor system in mammalian aging. Endocrinology. 2005;146:3718–3723.
66. Weraarchakul N, Strong R, Wood WG, Richardson A. The effect of aging and dietary restriction on DNA repair. Exp Cell Res. 1989;181:197–204.
67. Cuervo AM, Bergamini E, Brunk UT, Dröge W, Ffrench M, Terman A. Autophagy and aging: the importance of maintaining “clean” cells. Autophagy. 2005;1:131–140.
68. Harvey J, Wing RR, Mullen M. Effects on food cravings of a very low calorie diet or a balanced, low calorie diet. Appetite. 1993;21:105–115.
69. Chen K, Yan J, Suo Y, Li J, Wang Q, Lv B. Nutritional status alters sac-charin intake and sweet receptor mRNA expression in rat taste buds. Brain Res. 2010;1325:53–62.
70. Zverev YP. Effects of caloric deprivation and satiety on sensitivity of the gustatory system. BMC Neurosci. 2004;5:5.
71. McLaughlin SK, McKinnon PJ, Margolskee RF. Gustducin is a taste-cell-specific G protein closely related to the transducins. Nature. 1992;357:563–569.
72. Zhou LH, Liu XM, Feng XH, Han LO, Liu GD. Expression of alpha-gustducin in the circumvallate papillae of taste buds of diabetic rats. Acta Histochem. 2009;111:145–149.
73. Maudsley S, Davidson L, Pawson AJ, Chan R, López de Maturana R, Millar RP. Gonadotropin-releasing hormone (GnRH) antagonists promote proapoptotic signaling in peripheral reproductive tumor cells by activating a Galphai-coupling state of the type I GnRH receptor. Cancer Res. 2004;64:7533–7544.
74. Rhee SG. Regulation of phosphoinositide-specific phospholipase C. Annu Rev Biochem. 2001;70:281–312.
75. Barr AJ, Marjoram R, Xu J, Snyderman R. Phospholipase C-beta 2 interacts with mitogen-activated protein kinase kinase 3. Biochem Biophys Res Commun. 2002;293:647–652.
76. Morley JE, Armbrecht HJ, Farr SA, Kumar VB. The senescence accelerated mouse (SAMP8) as a model for oxidative stress and Alzheimer’s disease. Biochim Biophys Acta. 2012;1822:650–656.
77. Martin B, Chadwick W, Cong WN, et al. Euglycemic agent-mediated hypothalamic transcriptomic manipulation in the N171-82Q model of Huntington disease is related to their physiological efficacy. J Biol Chem. 2012;287:31766–31782.
by guest on June 16, 2016http://biom
edgerontology.oxfordjournals.org/D
ownloaded from