leishmania infantum secreted iron superoxide dismutase purification and its application to the...
TRANSCRIPT

LpL
SCD
ARR2A
KLIMICSEWOP
1
eLhcvnthw
0h
Comparative Immunology, Microbiology and Infectious Diseases 36 (2013) 499– 506
Contents lists available at SciVerse ScienceDirect
Comparative Immunology, Microbiologyand Infectious Diseases
j o ur na l ho me pag e: www.elsev ier .com/ locate /c imid
eishmania infantum secreted iron superoxide dismutaseurification and its application to the diagnosis of canineeishmaniasis
ilvia S. Longoni, Manuel Sánchez-Moreno, Jorge Emilio Rivera López,lotilde Marín ∗
epartamento de Parasitología, Facultad de Ciencias, Universidad de Granada, Severo Ochoa s/n, 18071 Granada, Spain
a r t i c l e i n f o
rticle history:eceived 12 June 2012eceived in revised form8 November 2012ccepted 3 May 2013
eywords:eishmania infantumon exchange chromatography
olecular sieve chromatographyron superoxide dismutaseanine Leishmaniasis
a b s t r a c t
Leishmania spp. are digenetic parasites whose infection occurs inside the mononuclearphagocitary system. The iron superoxide dismutase secreted (Fe-SODe) by promastigotes ofLeishmania spp. seems to plays an important role in the defense to environmental detoxifi-cation and neutralization of oxidative stress damage caused by reactive oxygen species(ROS) produced by macrophages during the infection. Parasites Fe-SODe is involved inestablishing the infection and manifestation of Leishmaniasis. Its high immunogenicitymakes it a useful molecular marker in diagnosing trypanosomatids infections. The aim ofthis study is demonstrate that purified Fe-SODe from Leishmania infantum is much moresensitive than un-purified Fe-SODe for diagnosis canine Leishmaniasis. We have purified aFe-SODe of L. infantum using an ion exchange and a molecular sieve chromatographies andits application in diagnosis of canine Leishmaniasis was tested. One hundred and forty-five
painLISAestern blot
xidative stressrotein purification
dogs’ sera from Andalusia Autonomous Community, Spain were tested by ELISA and West-ern blot and the antigen Fe-SODe purified is compared with two different antigens: thetotal parasites soluble lysate and the unpurified Fe-SODe. To validate the results obtainedusing the Fe-SODe purified we tasted 10 L. infantum infected dogs’ sera from Lombardy,Italy as positive control.
. Introduction
Leishmaniasis is a group of sand fly transmitted dis-ases caused by kinetoplastids belonging to the genuseishmania, 21 species of which have been identified asuman pathogens. These diseases have wide-ranging clini-al symptoms from self-healing cutaneous lesions to severeisceral disorders. In the year 2001, the World Health Orga-ization/Tropical Disease Research (WHO/TDR) recognized
his disease as a global health problem. In 2010 the WHOad estimate that around 12 million people are infectedorldwide and that a further 250 million are at risk of∗ Corresponding author. Tel.: +34 958 242369; fax: +34 958 243174.E-mail address: [email protected] (C. Marín).
147-9571/$ – see front matter © 2013 Elsevier Ltd. All rights reserved.ttp://dx.doi.org/10.1016/j.cimid.2013.05.004
© 2013 Elsevier Ltd. All rights reserved.
contracting the disease, and two million new infectionsoccurring every year (0.5 million of visceral Leishmaniasisand l.5 million of cutaneous Leishmaniasis). Leishmaniasisis endemic in 98 countries, or territories in all continentsexcept for Oceania and Antarctic. Visceral Leishmaniasiscauses an estimated over 50,000 deaths annually, a ratesurpassed among parasitic diseases only by malaria. How-ever, the majority of cases remain unreported, mainly dueto the fact that reporting is obligatory in only 33 countries.The reservoirs of this disease include both wild and domes-tic mammals.
Leishmania infantum is originated from Old World and
is endemic in all Mediterranean basin countries and it rep-resents a very big veterinarian problem due to the fact thatdogs are the most important reservoirs [1]. Just a few casesof human Leishmaniasis have been registered in European
icrobiol
500 S.S. Longoni et al. / Comparative Immunology, Mcountries, all of them in children or immune compromisedindividuals. Spain is considered an endemic country wherecanine Leishmaniasis prevalence varies between 3.7% and44.2%. Lowest prevalence was found in Pyrenean areas andin the northeast areas while the highest prevalence wasfound in the Mediterranean coast especially in the southern[2–5].
Leishmania spp. are digenetic parasites whose pro-gression into the host occurs inside the mononuclearphagocitary system of the immune system. Althoughit can invade many cell types, macrophages are selec-tively targeted by these microorganisms for replicationand for establish infection. When macrophages recog-nize Leishmania sp. they are activated to become so-called“effector cells”. After macrophages activation many cellu-lar processes start, including phagolysosomal degradationenzymes, oxidative burst generation, and nitric oxideproduction, all of these can phagocytosis and destroythe unwanted guests [6]. Promastigotes released dur-ing the bite of an infected sand fly are phagocytized bymacrophages. Similarly, amastigotes released after theburst of their host macrophage are rapidly phagocytizedby new macrophages.
During mitochondrial respiration, several reactive oxy-gen species (ROS) are formed, including superoxide radicals(O2
•−), hydrogen peroxide (H2O2) and hydroxyl radicals(•OH), and reactive nitrogen species (RNS), including nitric
oxide (NO•) [7,8]. O2
•− is produced by macrophages NAD-PHox in the phagocytosis phase of infection and theparasites form promastigotes. Superoxide Dismutase (SOD)dismute it in H2O2 and O2 in phagocytosis and intracellularamastigotes phases by both parasites forms, promastig-otes and amastigotes, which is convert in H2O by theparasitic antioxidant defense: T(SH)2 (Reductase Trypan-othione), TXN (Trypanoredoxin), PRX (Peroxiredoxin) andAPX (Ascorbate Peroxidase) [6,9] enzymes involved in thecommonly known as Thiol Redox Balance from whichdepends the parasites survival and the infection propaga-tion.
SODs are a group of antioxidant metalloenzymes. Theyplay an important role in the protection of normal cells,as well as, of a number of pathogens from reactive oxy-gen species (ROS) [10–12]. Classically, SODs have beenclassified into three types according to the metal cofactorbound at the active site: copper–zinc (Cu–Zn-SOD), man-ganese SODs (Mn-SOD), (both of which are present in mostprokaryotes and eukaryotes, including humans), and aniron SODs (Fe-SOD), detected in some prokaryotes, proto-zoans and chloroplasts of plants and algae. Fe-SOD playsa very important role in the Leishmania infection andestablishing of the disease [13–15]. Each Leishmania spp.presents different SODs isoforms. Among which there isa secreted isoforms (Fe-SODe), important in the estab-lishment of the parasites within the hosts. Many studieshave shown that the Fe-SODe has high immunogenicityand specificity, making it a useful and successful molecularmarker in the diagnosis of cutaneous and visceral canine
Leishmaniasis [16–18], feline Leishmaniasis [19], as wellas, mucocutaneous Leishmaniasis in human [20].In this study we have purified the Fe-SODe secreted byL. infantum promastigotes (SODe-Li) using two different
ogy and Infectious Diseases 36 (2013) 499– 506
chromatography types (ion exchange, QAE-Sephadex A-50column and molecular sieve, Sephadex G-100). Then wecompared, using two serodiagnostic techniques (ELISAand Western blot) different antigens: the total parasitessoluble lysate (H), the Fe-SODe-np (not purified) andthe SODe-Li (Fe-SODe purified) in 145 dogs’ sera fromAndalusia Autonomous Community (Spain).
The final aim of this study is to demonstrate that puri-fied antigen is better than the same antigen not purified.Employing a purified Fe-SODe the technique becomes moresensitive and specific.
2. Materials and methods
2.1. Parasite culture
Promastigotes of L. infantum (MCAN/ES/2001/UCM-10) were grown in axenic medium trypanosomes liquid(MTL) medium (Gibco®) supplemented with 10% heat-inactivated fetal bovine serum (FBS) at 26 ◦C in tissue-culture flasks until reaching an amount of approximately1 × 107 parasites/mL. Cells were collected at the logarith-mic growth phase by centrifugation (1500 × g for 10 minat room temperature). The pellet of cells (0.5–0.6 g wetweight/mL) was suspended in 25 mL of MTL’s medium notenriched with fetal bovine serum (FBS) and cultured at26 ◦C for 24 h.
2.2. Antigen preparation: extraction and purification ofFe-SODe
After 24 h, the promastigotes culture was centrifuged(1500 × g for 10 min) and the supernatant filtered(Minisart®, 20 �m). The filtered supernatant wassubjected to ice-cold ammonium sulphate precipitationat 35% salt concentration. Following centrifugation, theresultant supernatant was then treated with 85% ice coldammonium sulphate and the second precipitate collected.The resulting precipitate was finally dissolved in 2.5 mLof distilled water and desalted by chromatography inSephadex G-25 column (GE Healthcare Life Sciences®, PD10 column), previously equilibrated with 25 mL of distilledwater, bringing it to a final volume of 3.5 mL (FractionP85e or Fe-SODe-np) [15]. Degradation due to proteaseactivities present in the sample was minimized by addi-tion of 25 �L of protease inhibitor to the fraction P85e(CompleteMini, Roche®). Fraction P85e was applied to aQAE-Sephadex A-50 column (Sigma Immunochemicals®),30 cm × 2 cm, equilibrated with Buffer 1 (20 mM Potas-sium Phosphate, pH 7.4, containing 1 mM EDTA). Theadsorbed proteins were eluted with a linear gradient ofKCl (0–0.6 M). Fractions with a total volume of 2.5 mL wererecollected. Fractions exhibiting specific SOD activity werepooled (peak Q1e) and concentrated by ultrafiltration inMicrocon® filter tubes (Amicon®) at 11,200 × g for 30 min.The peak Q1e was applied to a Sephadex G-100 (Sigma
Immunochemicals®) molecular sieve chromatographycolumn (75 cm × 1.6 cm) equilibrated with Buffer 1 andeluted with 200 mL of the same buffer. The eluted fractions(4.5 mL each fraction) which showed SOD activity were
icrobiol
aa
2
aofc
2
psp
2
etpcaLwin(sSAaSsS
2
3plsAsFa
2
wTi(p(r
S.S. Longoni et al. / Comparative Immunology, M
gain collected (peak SODe-Li), concentrated (to 2 mg/mL)nd used for the assays described below.
.3. Protein determination
The protein content of the fractions P85e, the peaks Q1end SODe-Li, and all the fractions resulting from the elutionf the two columns were quantified using the Sigma Brad-ord test, which uses BSA as a standard (no traceability wasertified for the BSA standard) [21].
.4. Spectrometric enzyme assays
SOD activity was performed on the fraction P85e, theeaks Q1 and SODe-Li and the all the fractions eluted bypectrometric measurement of NBT-UV light oxidation asreviously described by Beyer and Fridovich [22].
.5. Determination of the molecular weight
Apparent molecular weight was calculated using thequation (y = −0.952x + 11.851) obtained from the linearrend line generated from the Standard Chromatogra-hy (Bovine Serum Albumine, 67 kDa; Albumin fromhicken egg white, 45 kDa;Carbonic Anhydrase, 30 kDa;nd Citocrome C, 12.4 kDa) (Sigma Immunochemicals, St.ouis) graphic (Microsoft Excel®, 2007). Molecular weightas confirmed by application of the SODe-Li fraction
nto a native gel electrophoresis in PhastGel Homoge-eous 12.5% as described in the Phast System manualGE Healthcare Life Sciences®). Molecular weights oftandard proteins were: Phosphorylase B (94 kDa), Bovineerum Albumin (67 kDa), Ovalbumin (43 kDa), Carbonicnhydrase (30 kDa), Soybean trypsin inhibitor (20.1 kDa)nd �-lactalbumin (14.4 kDa) (LMW, GE Healthcare Lifeciences®). The gels were stained for protein with high-ensitive 0.5% silver nitrate staining according to the Phastystem manual (GE Healthcare Life Sciences®).
.6. Determination of the isoelectric point
Isoelectro Focusing (IEF) in polyacrylamide Phast Gel-9 enabled us to determine the isoelectric points of theurified enzymes [23], using as markers: trypsinogen (9.3),
entil-lectin acidic band (8.8), horse-heart myoglobin (6.8),oybean trypsin inhibitor (4.6) and amyloglucosidase fromspergillus niger (3.6). The gels were stained with theilver–nitrate solution for proteins and with the Beyer andridovich method [22], in which enzyme activities appears achromatic bands in a blue background.
.7. Total parasites soluble lysate (fraction H)
L. infantum culture (in the exponential growth phase)as concentrated by centrifugation at 1500 × g for 10 min.
he pellet of the cells was washed twice and resuspended ince-cold sodium Tris/HCl ethylenediaminetetraacetic acid
EDTA) buffer (0.25 M sucrose, 25 mM Tris/HCl, 1 mM EDTA,H 7.8; buffer 2). Afterwards, the pellet was suspended0.5–0.6 g wet weight mL−1) in 3 mL of buffer 2 and dis-upted by 3 cycles of sonic disintegration, 30 s each at 60 W.ogy and Infectious Diseases 36 (2013) 499– 506 501
The sonicated homogenate was centrifuged at 1500 × g for10 min at 4 ◦C, and the pellet was washed three times withbuffer 2 for a total supernatant fraction of 9 mL. This frac-tion was centrifuged (2500 × g for 10 min at 4 ◦C) and thesupernatant (fraction H) was collected.
2.8. Serological test (ELISA)
The antigenic fractions: H, Fe-SODe-np (correspondingto fraction P85), and SODe-Li, at a total concentration of5 �g, 1.5 �g and, 0.5 �g, respectively, were used to sen-sitize polyethylene microtiter plates (Nunc, Dermark) incarbonate buffer (pH 8.2) for 2 h at 37 ◦C. Antigens notfixed in the plate were eliminated and washed 3 timeswith phosphate buffered saline (PBS), Tween 20® 0.05%(wash buffer). Free absorption sites where the antigen didnot come together were blocked for 2 h (37 ◦C) using ablocking solution (PBS-Tween 20® at 0.2%, bovine serumalbumine at 1%). After washing as before, the plate wasincubated for 45 min in the presence of dog serum at a 1:80dilution. After a further washing it was again incubatedfor 30 min (37 ◦C) with inmunoconjugate (Anti IgG withanti-dog peroxidase, Sigma Immunochemicals®), at a dilu-tion of 1:1000. The enzymatic, chromogenic reaction wasdone with OPD substrate (phenylenediamine dihydrochlo-ride, Sigma Immunochemicals®) and 10 �L of H2O2 at 30%for each 25 mL for 20 min in darkness. The reaction wasblocked by adding 50 �L of HCl 3N. Absorbency was readto 492 nm in a microplate reader (SunriseTM, TECAN). Allsamples were analyzed in triplicate in microtiter plates.The mean and standard deviation (SD) of the optical densityof negative-control sera (10 healthy dogs) were utilized tocalculate the cut-off value (mean + 3 × SD).
2.9. Western blot analysis
The Fe-SODe-np (corresponding to fraction P85) andSODe-Li antigens fraction (total concentration of 1.5 �gand 0.5 �g of protein) were merged into IEF 3-9 gels andsubsequently transferred to nitrocellulose for 30 min, asdescribed in the Phast System manual. The membrane wasblocked for 2 h at room temperature using 0.4% gelatineand 0.2% Tween 20 in PBS, followed by three washes in0.1% Tween 20 in PBS (PBS-T) and incubated for 2 h atroom temperature with dog sera at a dilution of 1:80.After being washed, the membrane was incubated for twomore hours at room temperature with the second antibody,anti-dog immunoglobulin G conjugate with peroxidase(Sigma Immunochemicals®; dilution 1:1000). The latterwas washed with diaminobenzidine substrate (0.5 mg/mLin a buffer of Tris/HCL 0.1 M, pH 7.4 that contains 1/5000H2O2 [10 v/v]) and it was added to the reaction, which wasstopped with a series of washes with distilled water.
2.10. Dogs population
A total of 145 dogs’ sera from Andalusia Autonomous
Community (76 from Granada, 17 from Cadiz, 9 from Jaen,5 from Malaga, 15 from Seville, 7 from Cordoba and 15from Huelva) were obtained between 2010 and 2011 fromveterinary clinics. Complete history is not available for all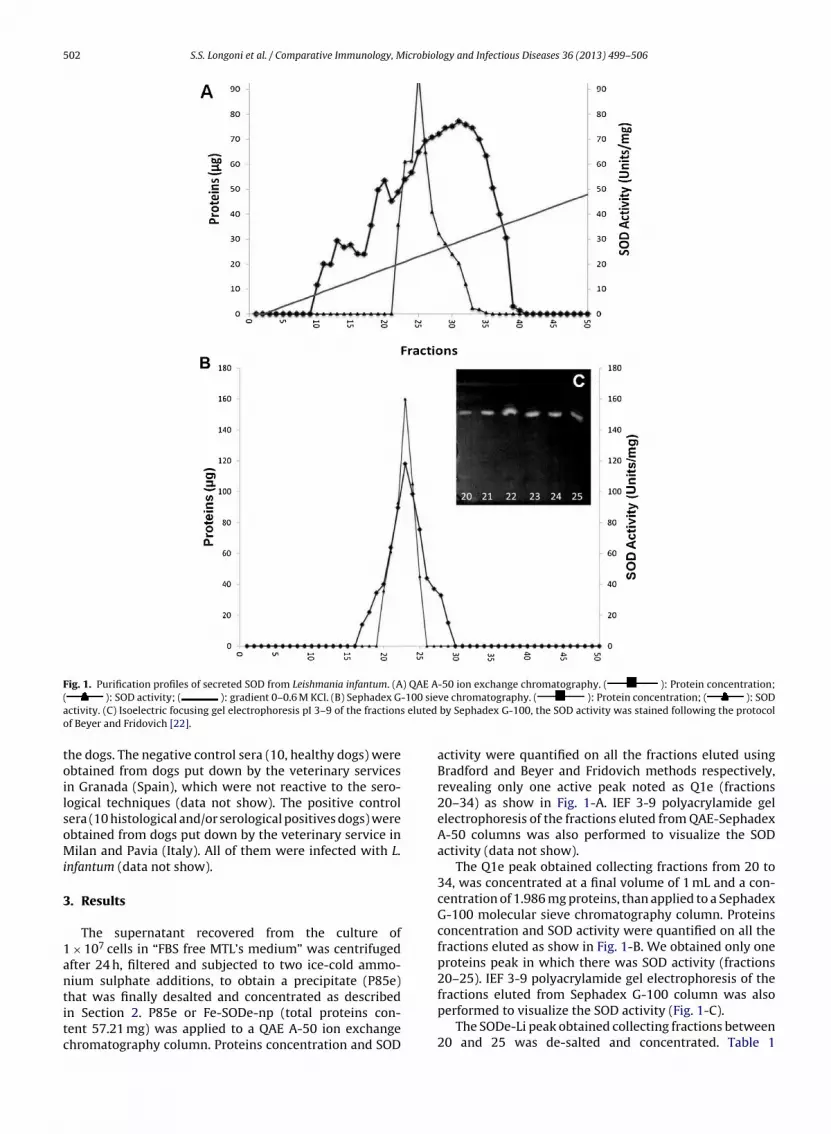
502 S.S. Longoni et al. / Comparative Immunology, Microbiology and Infectious Diseases 36 (2013) 499– 506
Fig. 1. Purification profiles of secreted SOD from Leishmania infantum. (A) QAE A-50 ion exchange chromatography. ( ): Protein concentration;-100 sies eluted
( ): SOD activity; ( ): gradient 0–0.6 M KCl. (B) Sephadex Gactivity. (C) Isoelectric focusing gel electrophoresis pI 3–9 of the fractionof Beyer and Fridovich [22].
the dogs. The negative control sera (10, healthy dogs) wereobtained from dogs put down by the veterinary servicesin Granada (Spain), which were not reactive to the sero-logical techniques (data not show). The positive controlsera (10 histological and/or serological positives dogs) wereobtained from dogs put down by the veterinary service inMilan and Pavia (Italy). All of them were infected with L.infantum (data not show).
3. Results
The supernatant recovered from the culture of1 × 107 cells in “FBS free MTL’s medium” was centrifugedafter 24 h, filtered and subjected to two ice-cold ammo-nium sulphate additions, to obtain a precipitate (P85e)
that was finally desalted and concentrated as describedin Section 2. P85e or Fe-SODe-np (total proteins con-tent 57.21 mg) was applied to a QAE A-50 ion exchangechromatography column. Proteins concentration and SODve chromatography. ( ): Protein concentration; ( ): SOD by Sephadex G-100, the SOD activity was stained following the protocol
activity were quantified on all the fractions eluted usingBradford and Beyer and Fridovich methods respectively,revealing only one active peak noted as Q1e (fractions20–34) as show in Fig. 1-A. IEF 3-9 polyacrylamide gelelectrophoresis of the fractions eluted from QAE-SephadexA-50 columns was also performed to visualize the SODactivity (data not show).
The Q1e peak obtained collecting fractions from 20 to34, was concentrated at a final volume of 1 mL and a con-centration of 1.986 mg proteins, than applied to a SephadexG-100 molecular sieve chromatography column. Proteinsconcentration and SOD activity were quantified on all thefractions eluted as show in Fig. 1-B. We obtained only oneproteins peak in which there was SOD activity (fractions20–25). IEF 3-9 polyacrylamide gel electrophoresis of the
fractions eluted from Sephadex G-100 column was alsoperformed to visualize the SOD activity (Fig. 1-C).The SODe-Li peak obtained collecting fractions between20 and 25 was de-salted and concentrated. Table 1
S.S. Longoni et al. / Comparative Immunology, Microbiology and Infectious Diseases 36 (2013) 499– 506 503
Table 1Leishmania infantum excreted superoxide dismutase purification summary.
Fraction Total protein (mg) Total activitya (Units) ± SD Specific activitya (U/mg ± SD) Purification Yield (%)
P85b 57.21 443.88 ± 21.98 7.76 ± 0.72 1 100Peak Q1b 1.986 436 ± 1.75 219.54 ± 0.389 28.29 98SODe-Li 0.387 339 ± 0.00 875.97 ± 0.00 112.88 76
“].
sp8
brSnmbb
EamE0fitsc
F10(
SD” is the standard deviation of the mean of four determinations.a SOD activity determined by the technique of Beyer and Fridovich [22b The total number of experiments being 4.
ummarizes the purification, indicating that SODe-Li wasurifies 112.88 times to a specific activity of about75.97 U/mg of protein.
The SODe-Li molecular weight (MW) was determinatey native homogeneous 12.5% electrophoresis analysisevealed according to the nitrate-staining method (Phast-ystem). The gel exhibited only one band of 25 kDa (dataot show). The SODe-Li isoelectric point (pI) was deter-ined carrying out an IEF 3-9 polyacrylamide gel followed
y SOD activity and silver nitrate staining. A single activityand was shown with a pI of 3.75 (data not show).
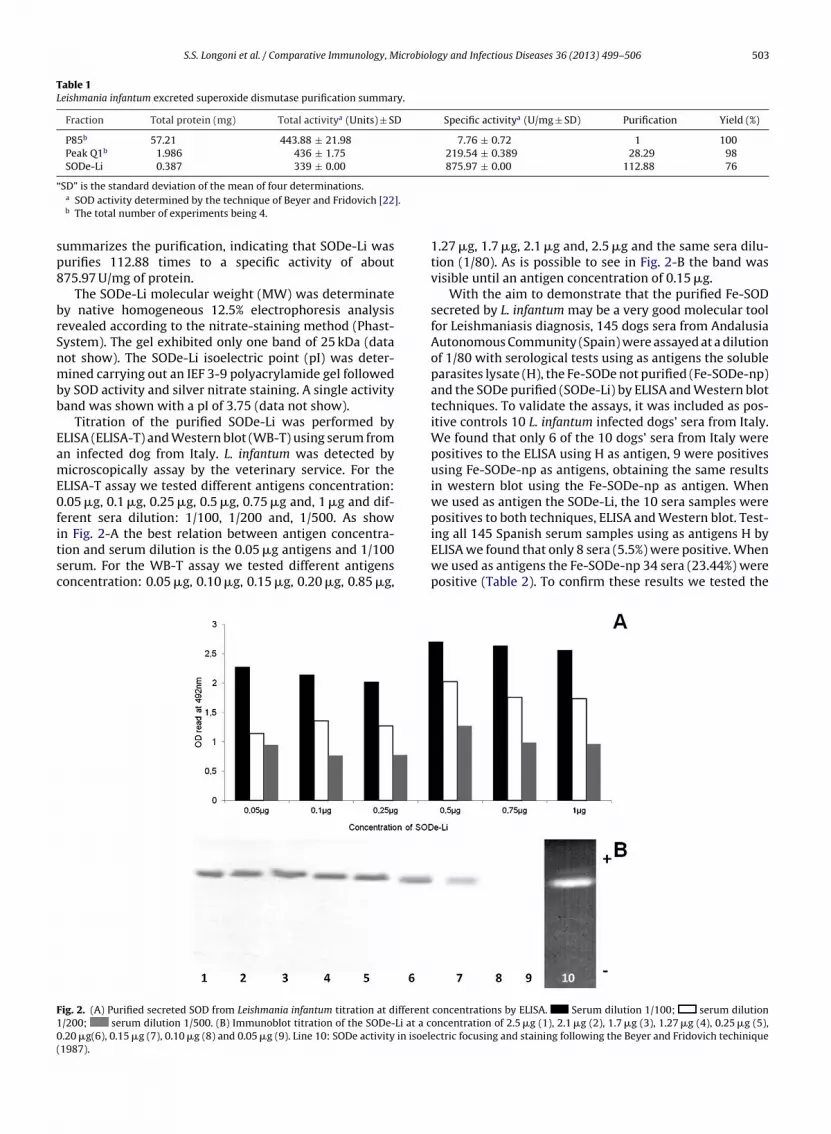
Titration of the purified SODe-Li was performed byLISA (ELISA-T) and Western blot (WB-T) using serum fromn infected dog from Italy. L. infantum was detected byicroscopically assay by the veterinary service. For the
LISA-T assay we tested different antigens concentration:.05 �g, 0.1 �g, 0.25 �g, 0.5 �g, 0.75 �g and, 1 �g and dif-erent sera dilution: 1/100, 1/200 and, 1/500. As show
n Fig. 2-A the best relation between antigen concentra-ion and serum dilution is the 0.05 �g antigens and 1/100erum. For the WB-T assay we tested different antigensoncentration: 0.05 �g, 0.10 �g, 0.15 �g, 0.20 �g, 0.85 �g,ig. 2. (A) Purified secreted SOD from Leishmania infantum titration at different
/200; serum dilution 1/500. (B) Immunoblot titration of the SODe-Li at a c.20 �g(6), 0.15 �g (7), 0.10 �g (8) and 0.05 �g (9). Line 10: SODe activity in isoele1987).
1.27 �g, 1.7 �g, 2.1 �g and, 2.5 �g and the same sera dilu-tion (1/80). As is possible to see in Fig. 2-B the band wasvisible until an antigen concentration of 0.15 �g.
With the aim to demonstrate that the purified Fe-SODsecreted by L. infantum may be a very good molecular toolfor Leishmaniasis diagnosis, 145 dogs sera from AndalusiaAutonomous Community (Spain) were assayed at a dilutionof 1/80 with serological tests using as antigens the solubleparasites lysate (H), the Fe-SODe not purified (Fe-SODe-np)and the SODe purified (SODe-Li) by ELISA and Western blottechniques. To validate the assays, it was included as pos-itive controls 10 L. infantum infected dogs’ sera from Italy.We found that only 6 of the 10 dogs’ sera from Italy werepositives to the ELISA using H as antigen, 9 were positivesusing Fe-SODe-np as antigens, obtaining the same resultsin western blot using the Fe-SODe-np as antigen. Whenwe used as antigen the SODe-Li, the 10 sera samples werepositives to both techniques, ELISA and Western blot. Test-
ing all 145 Spanish serum samples using as antigens H byELISA we found that only 8 sera (5.5%) were positive. Whenwe used as antigens the Fe-SODe-np 34 sera (23.44%) werepositive (Table 2). To confirm these results we tested theconcentrations by ELISA. Serum dilution 1/100; serum dilutiononcentration of 2.5 �g (1), 2.1 �g (2), 1.7 �g (3), 1.27 �g (4), 0.25 �g (5),ctric focusing and staining following the Beyer and Fridovich techinique

504 S.S. Longoni et al. / Comparative Immunology, Microbiology and Infectious Diseases 36 (2013) 499– 506
Table 2Relationship of positive dogs’ sera from Andalusia region (Spain) to Leishmania infantum by ELISA and Western blot (WB). Antigen fraction: total parasitessoluble lysate (H), iron superoxide dismutase excreted not purified (Fe-SODe-np), and iron superoxide dismutase excreted purified (SODe-Li).
H Fe-SODe-np SODe-Li H Fe-SODe-np SODe-Li
ELISA ELISA WB ELISA WB ELISA ELISA WB ELISA WB
4 − − − + + 64 − − − + +5 − − − + + 65 − + + + +6 − − − + + 66 − + − + +7 − − − + + 67 − + + + +8 − − − + + 68 − + + + +9 − − − + + 69 − + − + +
13 − − − + + 71 − + − + +15 − − − + + 72 − − − + +17 − − − + + 74 − − − + +18 − − − + + 77 − − − + +22 − + + + + 80 − − − + +23 − + − − − 82 − − − + +25 − + + + + 83 − + + + +29 − + + + + 85 − − − + +32 − + − − − 86 − − − + +33 − + + − − 87 − − − + +34 − + + − − 89 − − − + +37 − − − − − 93 + + − − −38 + + + + + 94 − − − + +42 − + + − − 95 − − − + +44 − + + + + 96 − − − + +47 − + + + + 97 − − − + +49 − + + + + 98 − + + − −50 − + + + + 100 − − − + +51 − + + + + 101 − − − + +52 − + + + + 108 − + + + +53 − + + + + 109 − − − + +54 − + + + + 111 − − − + +55 − − − + + 115 + − − − −56 − + + + + 116 − − − + +57 − + + + + 117 − − − + +59 − + − + + 120 − − − + +60 + + + + + 122 + − − − −61 − + + + + 125 + − − + +62 + + + + + 126 + − − − −
1
63 − + + + +ELISA positives sera by Western blot using as antigens theFe-SODe-np and we found that only 27 sera (18.62%) werepositives (Fig. 3-A), showing false positives results with theFe-SODe-np ELISA technique.
When we tested all the sera using as antigens SODe-Li by ELISA we found 61 sera (42.07%) samples positives(Table 2) and we found the same results testing the ELISApositives sera by Western blot techniques (Fig. 3-B) givinga 100% of correspondence between both techniques.
4. Discussion
Leishmaniasis diagnosis is still a problem, microscopicexamination is the most specific technique but the sen-sitivity is not good enough influenced by the laboratorytechnician expertise. The PCR can detect very slow para-sites presence in blood or bone marrow but is still anexpensive tool, available only in central hospital or ref-erence center. Different serological test are been used for
the diagnosis of Leishmania spp. infection (IFA, HAI, ELISA)but the absence of a good molecular marker made themnot high sensitive and specific [1]. Different antigens havebeen proposed in the last years, including antigen k39 from28 − − − + +8 34 27 61 61
L. chagasi, giving good results for visceral Leishmaniasis,a 32 kDa membrane antigen (P32) present in L. donovani,two hydrophilic antigens (k9 and k26) from L. chagasi, andantigen A2 and antigens rk26 and rK39 from L. infantum[24,25]. Of all of these, the antigen rK39 has shown greatsensitivity and a predictive value of the manifestations ofcutaneous Leishmaniasis. Different studies proposed theuse of a Fe-SODe for the diagnosis of visceral Leishmania-sis as well as cutaneous and mucocutaneous Leishmaniasis,in human, dogs and cats. Fe-SODe has been demonstratedto be parasites specific and no cross reaction is presentbetween different trypanosomatids [16,19,20]. The Fe-SODsecreted by the parasites, involved in the environmentaldetoxification and in the damage neutralization caused bythe ROS, seems to be the most important defense againstoxidative stress in these organisms lacking in catalase andperoxidase, the most common antioxidant complexes ineukaryotes.
In this work we report the purification of the Fe-SODe
secreted by L. infantum and its use in the canine Leishmania-sis diagnosis. A Fe-SODe was purified and characterized inPhytomonas sp. and Trypanosoma cruzi demonstrating thehigh immunogenicity of this enzyme [13,15]. Also a partial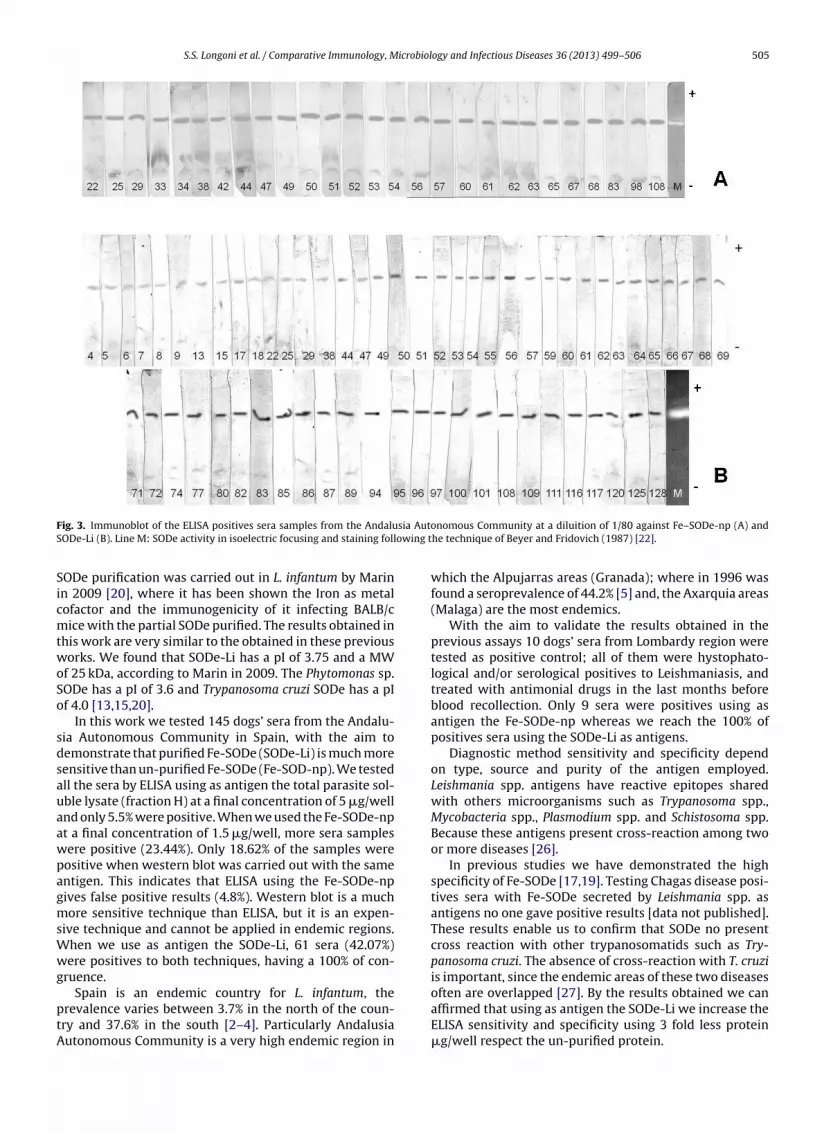
S.S. Longoni et al. / Comparative Immunology, Microbiology and Infectious Diseases 36 (2013) 499– 506 505
F sia AutoS owing t
SicmtwoSo
sdsauaawpagmsWwg
ptA
ig. 3. Immunoblot of the ELISA positives sera samples from the AndaluODe-Li (B). Line M: SODe activity in isoelectric focusing and staining foll
ODe purification was carried out in L. infantum by Marinn 2009 [20], where it has been shown the Iron as metalofactor and the immunogenicity of it infecting BALB/cice with the partial SODe purified. The results obtained in
his work are very similar to the obtained in these previousorks. We found that SODe-Li has a pI of 3.75 and a MW
f 25 kDa, according to Marin in 2009. The Phytomonas sp.ODe has a pI of 3.6 and Trypanosoma cruzi SODe has a pIf 4.0 [13,15,20].
In this work we tested 145 dogs’ sera from the Andalu-ia Autonomous Community in Spain, with the aim toemonstrate that purified Fe-SODe (SODe-Li) is much moreensitive than un-purified Fe-SODe (Fe-SOD-np). We testedll the sera by ELISA using as antigen the total parasite sol-ble lysate (fraction H) at a final concentration of 5 �g/wellnd only 5.5% were positive. When we used the Fe-SODe-npt a final concentration of 1.5 �g/well, more sera samplesere positive (23.44%). Only 18.62% of the samples wereositive when western blot was carried out with the samentigen. This indicates that ELISA using the Fe-SODe-npives false positive results (4.8%). Western blot is a muchore sensitive technique than ELISA, but it is an expen-
ive technique and cannot be applied in endemic regions.hen we use as antigen the SODe-Li, 61 sera (42.07%)ere positives to both techniques, having a 100% of con-
ruence.
Spain is an endemic country for L. infantum, therevalence varies between 3.7% in the north of the coun-ry and 37.6% in the south [2–4]. Particularly Andalusiautonomous Community is a very high endemic region in
nomous Community at a diluition of 1/80 against Fe–SODe-np (A) andhe technique of Beyer and Fridovich (1987) [22].
which the Alpujarras areas (Granada); where in 1996 wasfound a seroprevalence of 44.2% [5] and, the Axarquia areas(Malaga) are the most endemics.
With the aim to validate the results obtained in theprevious assays 10 dogs’ sera from Lombardy region weretested as positive control; all of them were hystophato-logical and/or serological positives to Leishmaniasis, andtreated with antimonial drugs in the last months beforeblood recollection. Only 9 sera were positives using asantigen the Fe-SODe-np whereas we reach the 100% ofpositives sera using the SODe-Li as antigens.
Diagnostic method sensitivity and specificity dependon type, source and purity of the antigen employed.Leishmania spp. antigens have reactive epitopes sharedwith others microorganisms such as Trypanosoma spp.,Mycobacteria spp., Plasmodium spp. and Schistosoma spp.Because these antigens present cross-reaction among twoor more diseases [26].
In previous studies we have demonstrated the highspecificity of Fe-SODe [17,19]. Testing Chagas disease posi-tives sera with Fe-SODe secreted by Leishmania spp. asantigens no one gave positive results [data not published].These results enable us to confirm that SODe no presentcross reaction with other trypanosomatids such as Try-panosoma cruzi. The absence of cross-reaction with T. cruziis important, since the endemic areas of these two diseases
often are overlapped [27]. By the results obtained we canaffirmed that using as antigen the SODe-Li we increase theELISA sensitivity and specificity using 3 fold less protein�g/well respect the un-purified protein.
icrobiol
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
506 S.S. Longoni et al. / Comparative Immunology, M
These are preliminary results that will lead to furtherrefinement of method. Therefore, this work could be con-sidered the first step in future research that will have theaim to understand better the complex parasite–host inter-actions, to clarify the high parasite resistance to drugs and,to improve the diagnostic test.
Acknowledgments
This work was supported by Spanish Ministry of Educa-tion (S.S. Longoni for a FPU grant).
The authors would like to thank the veterinaries fromAndalusia Autonomous Community who have given thesera samples and a special thank you to Dr. Claudia Paltaninfor the Italian dogs’ sera.
References
[1] WHO. Control of the leishmaniasis. Geneva: World Health Organiza-tion Technical Report Series; 2010.
[2] Chitimia L, Munoz-García CI, Sánchez-Velasco D, Lizana V, Del Río L,Murcia L, et al. Cryptic Leishmaniosis by Leishmania infantum, a fea-ture of canines only? A study of natural infection in wild rabbits,humans and dogs in southeastern Spain. Veterinary Parasitology2011;181(1):12–6.
[3] Cabezón O, Millán J, Gomis M, Dubey JP, Ferroglio E, Almería S. Kenneldogs as sentinels of Leishmania infantum, Toxoplasma gondii, andNeospora caninum in Majorca Island, Spain. Parasitology Research2010;107(6):1505–8.
[4] Millán J, Zanet S, Gomis M, Trisciuoglio A, Negre N, Ferroglio E. Aninvestigation into alternative reservoirs of canine leishmaniasis onthe endemic island of Mallorca (Spain). Transboundary and EmergingDiseases 2010;158(4):352–7.
[5] Acedo Sánchez C, Martín Sánchez J, Vélez Bernal ID, SanchísMarín MC, Louassini M, Maldonado JA, et al. Leishmaniasis eco-epidemiology in the Alpujarra region (Granada Province, southernSpain). International Journal for Parasitology 1996;26(3):303–10.
[6] van Assche T, Deschacht M, da Luz RA, Maes L, Cos P.Leishmania–macrophage interactions: insights into the redox biol-ogy. Free Radical Biology & Medicine 2011;51(2):337–51.
[7] Serarslan G, Yilmaz HR, Sögüt S. Serum antioxidant activities, malon-dialdehyde and nitric oxide levels in human cutaneous leishmaniasis.Clinical and Experimental Dermatology 2005;30(3):267–71.
[8] Bogdan C, Rollinghoff M, Diefenbach A. Reactive oxygen and reac-tive nitrogen intermediates in innate and specific immunity. CurrentOpinion in Immunology 2000;12:64–76.
[9] Piacenza L, Irigoín F, Alvarez MN, Peluffo G, Taylor MC, Kelly JM,et al. Mitochondrial superoxide radicals mediate programmed celldeath in Trypanosoma cruzi: cytoprotective action of mitochondrialiron superoxide dismutase overexpression. The Biochemical Journal2007;403(2):323–34.
10] Fridovich I. Superoxide dismutases: an adaptation to a paramagneticgas. Journal of Biological Chemistry 1989;264:7761–4.
11] McCord JM, Fridovich I. Superoxide dismutase: the first twenty years(1968–1988). Free Radical Biology and Medicine 1998;5:363–9.
12] Bannister JV, Bannister WH, Rottilio G. Aspects of the structure, func-tion and application of superoxide dismutase. Critical Reviews inBiochemistry 1987;22:111–80.
[
ogy and Infectious Diseases 36 (2013) 499– 506
13] Mateo H, Sánchez-Moreno M, Marín C. Enzyme-linked immunosor-bent assay with purified Trypanosoma cruzi secreted superoxidedismutase. Clinical Biochemistry 2010;43(15):1257–64.
14] Marín C, Rodríguez-González I, Sánchez-Moreno M. Identification ofsecreted iron superoxide dismutase for the diagnosis of Phytomonas.Memorias do Instituto Oswaldo Cruz 2006;101(6):649–54.
15] Marín C, Hitos AB, Rodríguez-González I, Dollet M, Sánchez-MorenoM. Phytomonas iron superoxide dismutase: a possible molecularmarker. FEMS Microbiology Letters 2004;234(1):69–74.
16] Arjona-Jiménez G, Villegas N, López-Céspedes A, Marín C, LongoniSS, Bolio-González ME, et al. Prevalence of antibodies against threespecies of Leishmania (L. mexicana, L. braziliensis, L. infantum) andpossible associated factors in dogs from Mérida, Yucatán, Mexico.Transactions of the Royal Society of Tropical Medicine and Hygiene2012;106(4):252–8.
17] Longoni SS, Marín C, Sauri-Arceo CH, López-Cespedes A, Rodríguez-Vivas RI, Villegas N, et al. An iron-superoxide dismutase antigen-based serological screening of dogs indicates their potential rolein the transmission of cutaneous leishmaniasis and trypanoso-miasis in Yucatan, Mexico. Vector-Borne and Zoonotic Diseases2011;11(7):815–21.
18] Marín C, Longoni SS, Mateo H, de Diego JA, Alunda JM, Minaya G,et al. The use of an secreted superoxide dismutase in an ELISA andWestern blotting for the diagnosis of Leishmania (Leishmania) infan-tum naturally infected dogs. Parasitology Research 2007;101(3):80–8.
19] Longoni SS, López-Cespedes A, Sánchez-Moreno M, Bolio-GonzalezME, Sauri-Arceo CH, Rodríguez-Vivas RI, et al. Detection ofdifferent Leishmania spp. and Trypanosoma cruzi antibod-ies in cats from the Yucatan Peninsula (Mexico) using aniron superoxide dismutase secreted as antigen. Compara-tive Immunology, Microbiology and Infectious Diseases 2012,http://dx.doi.org/10.1016/j.cimid.2012.04.003.
20] Marín C, Longoni SS, Urbano J, Minaya G, Mateo H, de Diego JA, et al.Enzyme-linked immunosorbent assay for superoxide dismutase-secreted antigen in diagnosis of sylvatic and Andean cutaneousleishmaniasis of Peru. American Journal of Tropical Medicine andHygiene 2009;80(1):55–60.
21] Bradford MM. A refined and sensitive method for the quantificationofmicroquantities of protein-dye binding. Analytical Biochemistry1976;72:248.
22] Beyer WF, Fridovich I. Assaying for superoxide dismutase activity:some large consequences of minor changes in conditions. AnalyticalBiochemistry 1987;161:559–66.
23] Bécuwe P, Gratepanche S, Fourmaux MN, van Beeumen J, SamynB, Mercereau-Puijalon O, et al. Characterization of iron-dependentendogenous superoxide dismutase of Plasmodium falciparum. Molec-ular and Biochemical Parasitology 1996;76:125–34.
24] Mohapatra TM, Singh DP, Sen MR, Bharti K, Sundar S. Compararativeevaluation of rK9, rK26 and rK39 antigens in the serodiagnosis ofIndian visceral leishmaniasis. The Journal of Infection in DevelopingCountries 2010;4:114–7.
25] Mizbani A, Taheri T, Zahedifard F, Taslimi Y, Azizi H, AzadmaneshK, et al. Recombinant Leishmania tarentolae expressing the A2virulence gene as a novel candidate vaccine against visceral leish-maniasis. Vaccine 2009;28:53–62.
26] Kar K. Serodiagnosis of leishmaniasis. Critical Reviews in Microbiol-ogy 1995;21:145–8.
27] Villagrán ME, Marín C, Rodríguez-Gonzalez I, de Diego JA, Sánchez-Moreno M. Use of an iron superoxide dismutase excreted byTrypanosoma cruzi in the diagnosis of Chagas disease. Seroprevalencein rural zones of the State of Queretaro, México. American Journal ofTropical Medicine and Hygiene 2005;73(3):510–6.