legacy effects of different land-use histories interact with current grazing patterns to determine...
TRANSCRIPT

Legacy Effects of Different Land-UseHistories Interact with CurrentGrazing Patterns to DetermineGrazing Lawn Soil Properties
Hugo Valls Fox,1,2,6* Olivier Bonnet,1 Joris P. G. M. Cromsigt,3,4
Herve Fritz,5 and Adrian M. Shrader1
1School of Life Sciences, University of KwaZulu-Natal, Private Bag X01, Scottsville 3209, South Africa; 2Biogeochimie et Ecologie desMilieux Continentaux, UMR 7618, Ecole Normale Superieure, 46 rue d’Ulm, 75230 Paris Cedex 05, France; 3Department of Wildlife,
Fish, and Environmental Studies, Swedish University of Agricultural Sciences, 901 83 Umea, Sweden; 4Centre for African Conser-
vation Ecology, Department of Zoology, Nelson Mandela Metropolitan University, PO Box 77000, Port Elizabeth, South Africa;5Laboratoire de Biometrie et Biologie Evolutive, UMR 5558, Bat. Mendel, Universite Lyon 1, 43 bd du 11 novembre 1918, 69622
Villeurbanne, France; 6Centre d’Ecologie Fonctionnelle et Evolutive, CEFE UMR 5175, CNRS - Universite de Montpellier - Universite
Paul-Valery Montpellier – EPHE, Campus du CNRS, 1919 route de Mende, 34293 Montpellier 5, France
ABSTRACT
Pastoralism and agriculture have affected range-
land ecosystems over the past millennia, including
many ecosystems that are currently protected as
reserves. However, the legacy of these land-use
practices on current ecosystem functioning remains
unclear. We studied legacy effects of former human
land use on soil physical and chemical properties in
a South African savanna. We did this by comparing
soil properties in grazing lawns (patches of short
grass maintained by the positive feedback between
grazing intensity and forage quality) with the sur-
rounding less grazed bunch grasslands within three
different human land-use history contexts: (i)
Abandoned bomas: permanent stone enclosures
where livestock were kept overnight, and dung and
urine accumulated for several years or decades. (ii)
Old fields: areas where vegetation was cleared, soil
tilled, and cultivated, but received little or no fer-
tilization. (iii) Natural grasslands: not cultivated but
grazed by livestock before the establishment of the
reserve and wildlife thereafter. Former human land
use rather than soil texture was the main deter-
minant of grazing lawn location. Moreover, lawn
soil properties also varied among land-use histories.
In all grazing lawns, soil nutrient concentrations
were higher than in adjacent grasslands but aban-
doned bomas contained three times more phos-
phorus, and twice as much nitrogen and carbon
than old fields and natural grassland lawns. In ad-
dition to past land use, soil texture influenced lawn
soil nutrients: Concentrations of phosphorus,
potassium, calcium, magnesium, total nitrogen,
and carbon in lawns were higher on clayey soils
than sandy soils, whereas phosphorus, C:N ratio,
and pH did not change with soil texture. Our study
confirms previous findings on the effect of human
land use on savanna heterogeneity, but also high-
lights how legacy effects may vary among different
historic land-use practices.
Key words: savanna; boma; nutrient cycling;
Ithala Game Reserve; herbivory; resource hotspots.
Received 11 June 2014; accepted 27 January 2015
Author contributions O. Bonnet, J. P. G. M. Cromsigt, and H. Fritz
came up with the initial idea; H. Valls Fox, A. M. Shrader, and O. Bonnet
designed the study; H. Valls Fox and O. Bonnet collected and analyzed
the data and all authors contributed to the manuscript.
*Corresponding author; e-mail: [email protected]
EcosystemsDOI: 10.1007/s10021-015-9857-x
� 2015 Springer Science+Business Media New York

INTRODUCTION
Pastoralism and cultivation have substantially
changed vegetation patterns and nutrient distri-
butions across African savannas over the past mil-
lennia (Prins 2000). Moreover, many areas
currently dedicated to wildlife conservation have
historically been affected by these practices (Ellis
and Ramankutty 2008). Several studies have
shown that former human land-use practices may
influence foraging patterns of wild savanna grazers
after agricultural activities have stopped, with po-
tentially large cascading effects on other parts of the
ecosystem (Blackmore and others 1990; Young and
others 1995; Donihue and others 2013). The legacy
of former pastoralism and cultivation may persist
for decades (McLauchlan 2007) or even millennia
(Cobo and others 2010). However, in this study, we
show that the legacy effects of livestock husbandry
and cultivation may differ considerably.
A typical practice of pastoralism in African sa-
vannas is the overnight containment of animals in
enclosed bomas (or corrals) to protect livestock
against predation. This practice leads to prolonged
accumulation of dung and urine in localized areas
from months to decades resulting in elevated soil
nutrient concentrations and organic matter content
for decades after boma abandonment (Augustine
2003). Bush clearing is also a central practice in the
creation of boma sites, creating open glades with a
specific herbaceous and woody vegetation com-
munity in an often denser savanna matrix
(Porensky and Veblen 2012). In comparison, the
legacy of cultivation differs substantially from that
of bomas. Native vegetation is removed during the
cultivation of fields. In contrast to bomas, cultiva-
tion often leads to prolonged nutrient and soil
organic matter loss caused by the replacement
of native vegetation by crops, biomass removal,
and especially tilling (Roberts and others 2003;
McLauchlan 2007).
After abandonment, wild grazers may select for
and maintain former boma sites as open, nutrient-
rich resource hotspots, or grazing lawns (Young
and others 1995). Grazing lawns are conspicuous
patches of short, mat-forming, grazing-tolerant
grass species resulting from repeated cropping by
herbivores (Figure 1; McNaughton 1984; Cromsigt
and Kuijper 2011). Continuous grazing in these
lawns keeps grass short and sustains high-quality
forage, generating a positive feedback between
herbivore consumption and the nutritional value of
the grass (McNaughton 1979, 1984; Bonnet and
others 2010). Localized dung and urine deposition
by herbivores strengthens this feedback by accel-
erating the rate of nutrient cycling in the top soil of
grazing lawns and facilitating sustained plant
growth (Ruess and McNaughton 1987), likely me-
diated by increasing microbial activity (Bardgett
and Wardle 2003; Sankaran and Augustine 2004;
de Vries and others 2012). The intense grazing also
shifts the grass community toward more grazing-
tolerant and palatable species with higher litter
quality which may further accelerate soil nutrient
cycling (Bardgett and Wardle 2003; Cromsigt and
Kuijper 2011). Although grazing lawns were ini-
tially described in the Serengeti plains in Tanzania
(McNaughton 1984) and not originally associated
with abandoned bomas, more recent studies sug-
gest that the short grass hotspots in former boma
sites are functionally similar to the lawns, as
originally formalized by McNaughton (Cromsigt
and Kuijper 2011).
In contrast, we know much less about the effects
of abandoned fields on the foraging patterns of wild
savanna grazers and to what extent these grazers
may maintain grazing lawns in old fields. The cre-
ation of open habitat through bush clearing and the
removal of tall grass in old fields create the neces-
sary starting conditions to initiate the development
of grazing lawns (Cromsigt and Olff 2008). Similar
to former boma sites, the open habitats in old fields
may attract herbivores by reducing predation risk
(Ford and others 2014), whereas the removal of tall
caespitose grasses facilitates colonization by higher
quality lawn grass species (Huisman and Olff 1998;
Cromsigt and Olff 2008; Novellie and Gaylard
2013). Once grazed upon, herbivores would be
Figure 1. An example of a typical grazing lawn from the
study area. During the growing season, the mat-forming
Cynodon dactylon grasses are cropped daily by resident
grazers. Note the sharp edge with the surrounding bunch
grass matrix (Sporobolus sp.).
H. Valls Fox and others

more likely to return to these sites comprising
younger, more nutritious forage (Bonnet and oth-
ers 2010), thus initiating a positive feedback known
as grazing optimization (de Mazancourt and others
1998). This process may then initiate a similar de-
velopment of grazing lawns on old fields as on
former boma sites. The contrasting effects of culti-
vation and bomas on nutrient and organic matter
build-up should, however, lead to grazing lawns
with quite contrasting post-agro-pastoral biogeo-
chemical signatures. Whereas lawn soils on former
boma sites should be enriched with organic matter
and less volatile nutrients such as phosphorous, we
predict that lawn soils on old fields do not show this
enrichment. In addition, we currently lack under-
standing of the spatial extent at which land use
affects soil parameters and resulting grazing lawns.
This is particularly relevant, because this scale may
differ between land-use types, with old fields
being significantly larger than boma sites (Lewu
and Assefa 2009).
Soil parent material likely modifies the legacy
effects of different land uses (McLauchlan 2007).
Theoretical (de Mazancourt and others 1998) and
empirical evidence (Ruess and McNaughton 1987)
suggests that the positive feedback between herbi-
vores, plants and soil biota characterizing grazing
lawns should only come into being in nutrient-rich
contexts (Bardgett and Wardle 2003). Several
studies indeed confirmed that nutrient-rich soil
patches favored the formation of grazing lawns
(Cromsigt and Olff 2008) and were associated with
their distribution and persistence (Scholes and
Walker 1993). A recent study discussed another
important aspect of parent material, namely, soil
texture as a potential driver of soil compaction
(Veldhuis and others 2014). This builds upon pre-
vious studies that showed that grazing tolerance in
grass species may in fact reflect adaptations to low
water availability (Coughenour 1985). Veldhuis
and others (2014) discuss how herbivore trampling
drives soil compaction of lawn soils, hereby
reducing water infiltration rates and favoring
the more drought-tolerant lawn grass species,
specifically on fine textured soils. Indeed, grazing
lawns were first described on fertile, fine textured
volcanic soils of the Serengeti plains (McNaughton
1984). However, other studies have shown that
resource hotspots derived from old bomas have
been described across a wide range of soil types,
including fine textured clayey soils (Veblen 2012),
loamy soils (Young and others 1995; Augustine
2003), and coarser sandy soils (Cech and others
2010; van der Waal and others 2011). We currently
lack studies that describe grazing lawn soil prop-
erties for different former land uses and with var-
iation in soil texture.
Our aim was to quantify the relative importance
of human land-use legacy and soil parent material
(especially texture) on the soil nutrient and carbon
status of grazing lawns, and to compare legacy ef-
fects among different land uses. We conducted our
study in Ithala Game Reserve, South Africa, which
provided us with a unique setting that combined
different past land-use practices with contrasting
soil geological origins within a relatively small area.
Moreover, as we were unsure about the spatial
extent of past land-use legacies and expected this
extent may differ between the larger old fields and
smaller bomas, we compared soil from within in-
tensively used grazing lawns with close as well as
far away tall grassland controls (10 m and 200 m
away from the lawn, respectively). To assess the
legacy effects of different land-use types, we com-
pared lawns found in abandoned bomas, old fields,
and natural grasslands. Natural grasslands are areas
comprising indigenous grasses and forbs that are
grazed by wildlife and livestock (Allen and others
2011). Based on the different impacts to soil nu-
trients and texture, we predicted that the properties
of grazing lawns would differ with past land use.
We expected large stocks of soil nutrients and car-
bon in former bomas to persist long after their
abandonment and attract herbivores to these
lawns. Grazing lawns in old fields would have
originated from the rapid onset of grazing opti-
mization after herbivores aggregated in the large
disturbed grasslands that emerged and not from
large stocks of soil nutrients and carbon. These
differences may then be further mediated by var-
iation in soil texture, where lawns should be pre-
dominantly found on fertile fine textured soils if
nutrient limitation drives grazing patterns.
MATERIALS AND METHODS
Study Area
We conducted the study in Ithala Game Reserve
(hereafter Ithala), KwaZulu-Natal, South Africa
(27�30¢S; 31�20¢E). The 30,000 ha reserve com-
prises a series of deep valleys running from Ngotshe
Mountain (alt. 1446 m) to the Pongola River (alt.
320 m). Soils are generally shallow and undiffer-
entiated ranging from coarse textured (<15% clay)
dystrophic regosols of granitic origin in the east to
more fine-grained (15–35% clay) and eutrophic
leptosols soils in the center and west (Figure 2A; van
Rooyen and van Rooyen 2008; Dewitte and others
2013). Climate is sub-tropical. From 1973 to 2010,
Legacy of Land Use on Grazing Lawn Soil Properties
annual rainfall ranged from 394 to 1164 mm
(mean = 763 mm, CV = 27%) and 82% of rainfall
occurred during the rainy season from October to
March. Mean daily temperatures range from 14 to
27�C in January during the rainy season to 4–21�Cin July during the cold dry season.
Ithala contains a wide variety of habitat types.
Open areas comprise 37% of the reserve. These
include the following: Natural grasslands which
comprise 19% of the reserve and are dominated by
the grass species Trachypogon spicatus, Tristachya
leucothrix, Hyparrhenia hirta, Senecio microglossus, and
Bewsia biflora; Old field grasslands that comprise 9%
of the reserve and are dominated by the grass
species Hyparrhenia hirta and Sporobolus africanus,
and 9% riparian vegetation dominated by the tree
Syzygium cordatum and sedge Cyperus sexangularis.
The most dominant habitats are the dense bush-
veld/thickets comprising 52% of the reserve, and
containing the tree species Acacia nilotica, Combre-
tum apiculatum, Euclea schimperi, Greyia sutherlandii,
and grasses such as Tristachya leucothrix, Melinis
nerviglumis, Eragrostis curvula, and Themeda triandra.
Woodlands comprise 11% of the reserve and con-
taining both trees (Nuxia oppositifolia, Dovyalis zey-
heri, Salix mucronata, Dalbergia armata, and
Combretum hereroense) and the grasses (Phragmites
australis, Imperata cylindrica, Paspalum urvillei, and
Paspalum dilatatum) (van Rooyen and van Rooyen
2008). Main grazers in the reserve include white
rhinoceros (Ceratotherium simum c. 41 ind.), plains
zebra (Equus quagga c. 1600 ind.), wildebeest
(Connochaetes taurinus c. 1044 ind.), warthog (Pha-
cochoerus aethiopicus c. 2000 ind.) and impala
(Aepyceros melampus c. 2200 ind.).
Until the establishment of the game reserve in
1972, Ithala was a patchwork of fields (now termed
old field grasslands) and natural grassland areas
interspersed by homesteads each containing en-
closed stone wall corrals (or bomas) where live-
stock were kept at night. Van Rooyen and Van
Rooyen (2008) identified 111 old field grassland
patches across Ithala, ranging from 0.2 to 250 ha
with a mean of 22 ha and median of 7.1 ha. These
old field grassland patches would consist of multi-
ple associated fields and the average size of indi-
vidual old fields in Ithala was therefore likely
around 1–2 ha and similar to current small-scale
farming in KwaZulu-Natal, South Africa (Lewu and
Assefa 2009). Hence, old fields were much larger
than bomas that were typically between 10 and
30 m radius. Although we do not have data on the
lifetime of such bomas for Ithala, we know from
similar historic and current practices elsewhere in
Africa that they are likely used for several decades
if not longer (Blackmore and others 1990; Young
and others 1995). Moreover, the fact that in Ithala
bomas were encircled with stonewalls and not
thorny branches suggests long-term use. Native
grazers were absent during this period, but subse-
quently reintroduced as the farms were abandoned
and the fenced reserve expanded (van Rooyen and
van Rooyen 2008). Ultimately, this varied land-use
history provided us with a unique opportunity to
compare grazing lawns found in areas with differ-
ent land-use histories within a single reserve.
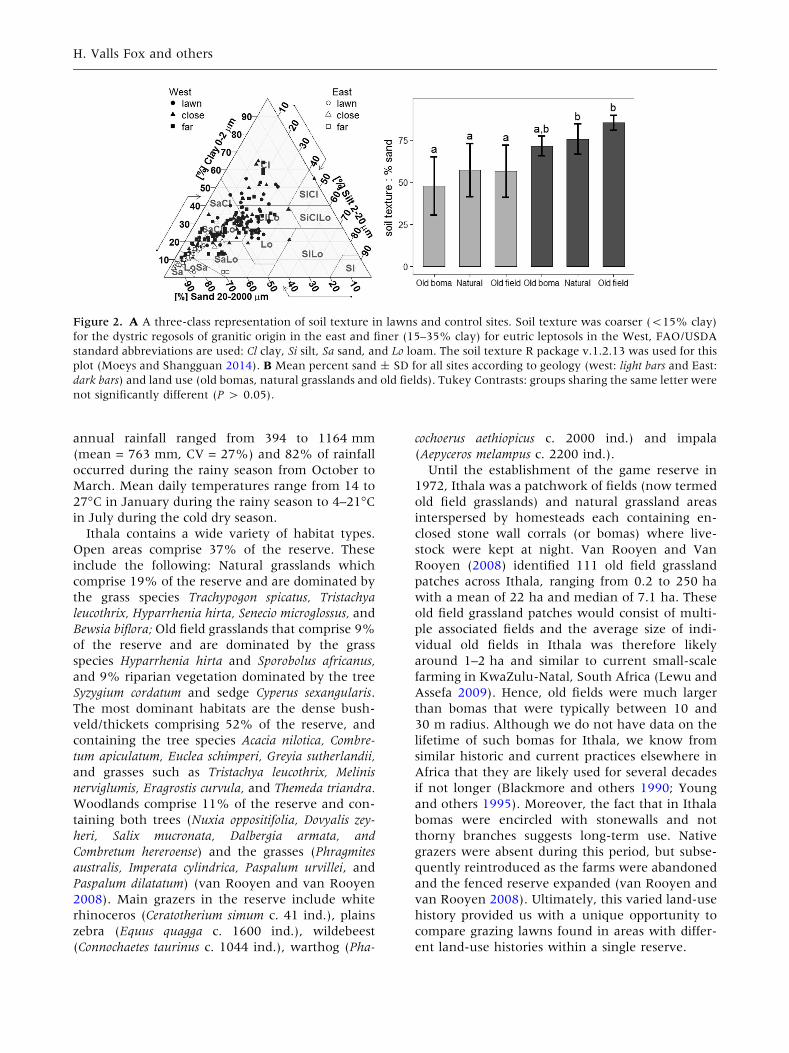
Figure 2. A A three-class representation of soil texture in lawns and control sites. Soil texture was coarser (<15% clay)
for the dystric regosols of granitic origin in the east and finer (15–35% clay) for eutric leptosols in the West, FAO/USDA
standard abbreviations are used: Cl clay, Si silt, Sa sand, and Lo loam. The soil texture R package v.1.2.13 was used for this
plot (Moeys and Shangguan 2014). B Mean percent sand ± SD for all sites according to geology (west: light bars and East:
dark bars) and land use (old bomas, natural grasslands and old fields). Tukey Contrasts: groups sharing the same letter were
not significantly different (P > 0.05).
H. Valls Fox and others
Grazing Lawn Selection According toLand-Use History
During the wet season, during which we sampled,
grazing lawns contrasted sharply with surrounding
taller bunch grassland (Figure 1) and mostly con-
sisted of short monospecific patches of Cynodon
dactylon (mean height = 4 cm). We know from
previous work in a similar savanna system near
Ithala that during the wet season grazing lawns
need to be intensely grazed, nearly daily, to remain
in such a short state (Cromsigt and Olff 2008;
Bonnet and others 2010). Moreover, from other
work by some of the co-authors, we know that in
Ithala white rhinoceros spend nearly all of their
foraging time in grazing lawns (Bonnet and Shra-
der unpublished data).
We first located grazing lawns using high-
resolution aerial photographs (1/30,000) from 2003
taken by the South African Department of Land
Affairs. Grazing lawn presence was then confirmed
on foot in 2009 and 2010. This mapping exercise
showed that within Ithala grazing lawns were
generally smaller than 0.5 ha and only covered
0.14% of the reserve. Grazing lawns were pre-
dominantly found in open old field grasslands
where herbivore densities were highest (pers. ob-
servation) and to a lesser extent natural grasslands
(Table 1). Former bomas were distinguished from
other grazing lawns in the field based on evidence
of human occupation. We limited our study to
boma lawns that were found within the remains of
rectangular stone walls. To differentiate between
old fields and natural grasslands sites, we used the
vegetation map generated by van Rooyen and van
Rooyen (2008). To generate their map, van Rooyen
and van Rooyen (2008) used both physical features
of the landscape (for example, terraced rows on
predominantly north-facing gentle slopes) and
present grass species composition to determine the
location of the old field grasslands across the re-
serve. Because most of Ithala has been subjected to
human land use, grazing lawns in natural grass-
lands were scarce; however, we often found they
were located close to natural nutrient hotspots such
as termite mounds or white rhinoceros dung mid-
dens.
To further confirm that our grazing lawn classi-
fications did indeed reflect different previous land-
use histories, we used (1/30,000) aerial pho-
tographs from 1969 to 1976 in order to confirm
that the lawns used in our study matched land use
at the time of the proclamation. We then confirmed
the classification was coherent with the oldest
series of photographs we were able to exploit dat-
ing from 1948; indicating these land uses had per-
sisted for at least two or three decades. Nonetheless,
it is possible that some of the grazing lawns in the
old field grasslands had been boma sites prior to
1948. Although bomas were also found in old field
grasslands, we restricted our choice of boma lawns
to those that clearly fell outside of old fields to
avoid possible confounding effects between these
two land-use histories. As a result, by selecting
these boma sites we reduced the possibility of a link
between old field lawns and boma lawns.
In total, we sampled 32 grazing lawns (13 boma
sites, 8 old field sites, and 11 natural grassland
sites). We only sampled grazing lawns larger than
50 m2 to limit edge effects. Each grazing lawn was
paired with two control grassland sites, one close
(10 m from the chosen lawn) and one far from the
lawn (200 m from any grazing lawn). We took a
close and far away control because we were unsure
of the spatial extent of the legacy of agricultural
land use, and because we expected that this extent
might differ between the larger old fields and the
smaller bomas. To limit confounding effects, both
controls were sampled on the same day as the
grazing lawn and were located in random direc-
tions from the grazing lawn, while remaining in the
same landscape unit (for example, past land use,
vegetation type, and hillside).
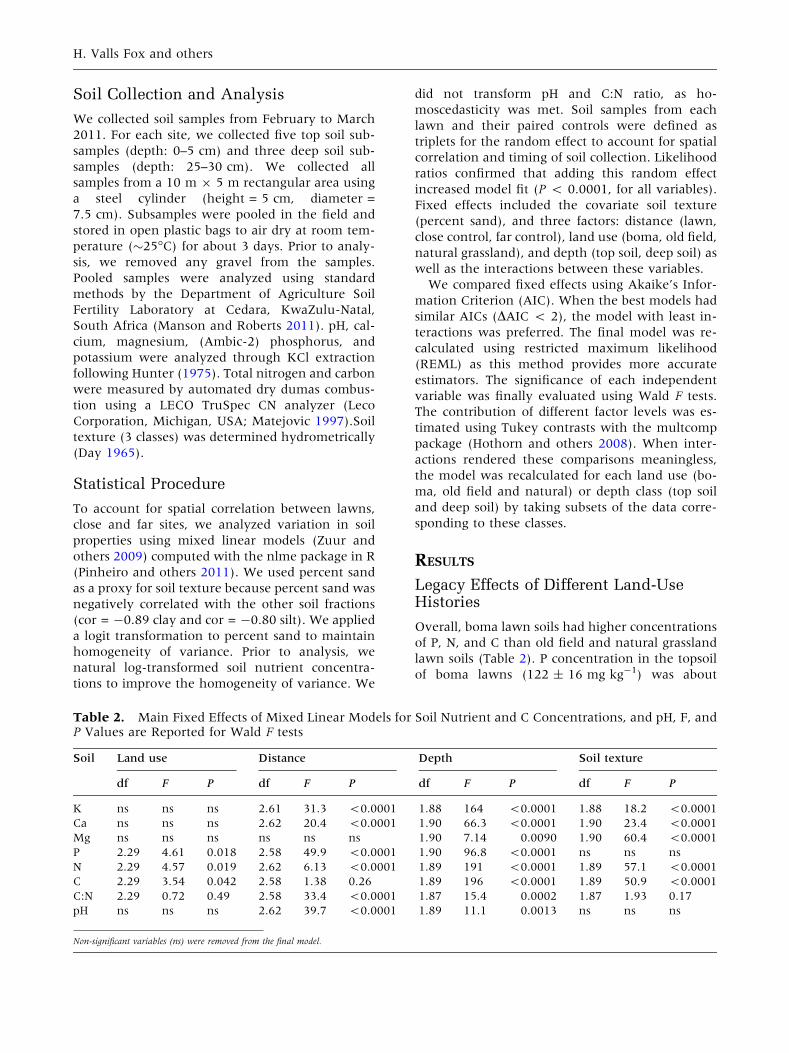
Table 1. Main Habitat Types in Ithala Game Reserve
Habitat type Surface area (ha) Surface area (%) Lawn surface area (ha) Grazing lawn cover (%)
Bushveld/woodland 18,581 63 12 0.06
Old field grassland 2535 9 18 0.71
Natural grasslands 5658 19 10 0.18
Riparian areas 2509 9 1 0.06
Total 292,831 100 41
The proportion of each habitat covered by lawns is expressed in m2 ha-1.1Built-up areas and cliffs inaccessible to grazers were excluded.
Legacy of Land Use on Grazing Lawn Soil Properties
Soil Collection and Analysis
We collected soil samples from February to March
2011. For each site, we collected five top soil sub-
samples (depth: 0–5 cm) and three deep soil sub-
samples (depth: 25–30 cm). We collected all
samples from a 10 m 9 5 m rectangular area using
a steel cylinder (height = 5 cm, diameter =
7.5 cm). Subsamples were pooled in the field and
stored in open plastic bags to air dry at room tem-
perature (�25�C) for about 3 days. Prior to analy-
sis, we removed any gravel from the samples.
Pooled samples were analyzed using standard
methods by the Department of Agriculture Soil
Fertility Laboratory at Cedara, KwaZulu-Natal,
South Africa (Manson and Roberts 2011). pH, cal-
cium, magnesium, (Ambic-2) phosphorus, and
potassium were analyzed through KCl extraction
following Hunter (1975). Total nitrogen and carbon
were measured by automated dry dumas combus-
tion using a LECO TruSpec CN analyzer (Leco
Corporation, Michigan, USA; Matejovic 1997).Soil
texture (3 classes) was determined hydrometrically
(Day 1965).
Statistical Procedure
To account for spatial correlation between lawns,
close and far sites, we analyzed variation in soil
properties using mixed linear models (Zuur and
others 2009) computed with the nlme package in R
(Pinheiro and others 2011). We used percent sand
as a proxy for soil texture because percent sand was
negatively correlated with the other soil fractions
(cor = -0.89 clay and cor = -0.80 silt). We applied
a logit transformation to percent sand to maintain
homogeneity of variance. Prior to analysis, we
natural log-transformed soil nutrient concentra-
tions to improve the homogeneity of variance. We
did not transform pH and C:N ratio, as ho-
moscedasticity was met. Soil samples from each
lawn and their paired controls were defined as
triplets for the random effect to account for spatial
correlation and timing of soil collection. Likelihood
ratios confirmed that adding this random effect
increased model fit (P < 0.0001, for all variables).
Fixed effects included the covariate soil texture
(percent sand), and three factors: distance (lawn,
close control, far control), land use (boma, old field,
natural grassland), and depth (top soil, deep soil) as
well as the interactions between these variables.
We compared fixed effects using Akaike’s Infor-
mation Criterion (AIC). When the best models had
similar AICs (DAIC < 2), the model with least in-
teractions was preferred. The final model was re-
calculated using restricted maximum likelihood
(REML) as this method provides more accurate
estimators. The significance of each independent
variable was finally evaluated using Wald F tests.
The contribution of different factor levels was es-
timated using Tukey contrasts with the multcomp
package (Hothorn and others 2008). When inter-
actions rendered these comparisons meaningless,
the model was recalculated for each land use (bo-
ma, old field and natural) or depth class (top soil
and deep soil) by taking subsets of the data corre-
sponding to these classes.
RESULTS
Legacy Effects of Different Land-UseHistories
Overall, boma lawn soils had higher concentrations
of P, N, and C than old field and natural grassland
lawn soils (Table 2). P concentration in the topsoil
of boma lawns (122 ± 16 mg kg-1) was about
Table 2. Main Fixed Effects of Mixed Linear Models for Soil Nutrient and C Concentrations, and pH, F, andP Values are Reported for Wald F tests
Soil Land use Distance Depth Soil texture
df F P df F P df F P df F P
K ns ns ns 2.61 31.3 <0.0001 1.88 164 <0.0001 1.88 18.2 <0.0001
Ca ns ns ns 2.62 20.4 <0.0001 1.90 66.3 <0.0001 1.90 23.4 <0.0001
Mg ns ns ns ns ns ns 1.90 7.14 0.0090 1.90 60.4 <0.0001
P 2.29 4.61 0.018 2.58 49.9 <0.0001 1.90 96.8 <0.0001 ns ns ns
N 2.29 4.57 0.019 2.62 6.13 <0.0001 1.89 191 <0.0001 1.89 57.1 <0.0001
C 2.29 3.54 0.042 2.58 1.38 0.26 1.89 196 <0.0001 1.89 50.9 <0.0001
C:N 2.29 0.72 0.49 2.58 33.4 <0.0001 1.87 15.4 0.0002 1.87 1.93 0.17
pH ns ns ns 2.62 39.7 <0.0001 1.89 11.1 0.0013 ns ns ns
Non-significant variables (ns) were removed from the final model.
H. Valls Fox and others

three times higher than in natural grassland lawns
(45 ± 14 mg kg-1) and old field lawns (39 ± 21
mg kg-1) (Figure 3D). Similarly, top soil N con-
centrations in boma lawns (4.7 ± 0.7 g kg-1) were
about twice the level of N concentrations in natural
grassland (2.4 ± 0.4 g kg-1) and old field lawns
(2.1 ± 0.5 g kg-1) (Figure 3E). This pattern also
held for total carbon (bomas: 48 ± 7 g kg-1, nat-
ural grasslands: 27 ± 4 g kg-1, old fields: 25 ± 6
g kg-1) (Figure 3F). In contrast, the C:N ratio was
lower in boma lawns (10.4 ± 0.3) compared to
lawns in natural grasslands (11.2 ± 0.4) and old
fields (11.9 ± 0.4) (Figure 3H; Table 2). We found
no effect of past land use on lawn soil properties in
terms of K, Ca, Mg, and pH (Tables 2, 3; Figure 3).
Grazing lawn soils had higher concentrations of
P, N, K, Ca, higher pH, and lower C:N ratio com-
pared to the close (10 m) and far (200 m) control
soils (Figure 3; Table 2). The only nutrient that did
not vary among lawn and control soils was Mg
(Tables 1, 2; Figure 3C). However, the spatial ex-
tent of the elevated soil nutrient concentrations
varied among nutrient type and across the different
land-use histories (Table 3). With regard to P, boma
lawns had concentrations that were ten times
higher than controls, whereas for lawns in old
fields and natural grasslands P concentrations were
only four times higher (Figure 3D). Concentrations
of C and N were significantly higher in boma lawns
compared to close controls (C dist: F2,24 = 4.0,
P = 0.031; N dist: F2,24 = 7.4, P = 0.003), but not
for lawns in old fields or natural grasslands: C (dist:
F2,36 = 2.5, P = 0.095) and N (dist: F2,36 = 3.2,
P = 0.054). This means that only boma lawns rep-
resented nutrient hotspots for N, P, and C com-
bined, whereas old field and natural grassland
lawns were only hotspots for P (albeit to a lesser
degree than boma lawns). Moreover, for some
nutrients, the patterns did not differ between the
different land-use histories. For example, top soil
(z = -0.21 and P = 1) and deep soil (z = 0.15 and
P = 0.42) concentrations of K did not differ be-
tween lawns and close sites, but these were more
than twice the K concentrations recorded at the far
sites across all land-use histories (Figure 3A). In the
case of Ca, concentrations in lawns of the different
land-use histories were 1.5 higher than close con-
trols (z = 3.3 and P = 0.003), which in turn were
1.2 times higher than far controls (z = -3.1 and
P = 0.006, Figure 3B).
Finally, with regard to soil depth, differences in
P, K, and C:N between top and deep soils were
more pronounced in the far control soils than in
the lawn soils of the different land-use histories
(Figure 3). In contrast, for Ca and Mg, patterns
varied between lawns and controls (see dis-
tance 9 depth in Table 2; Figure 3). Lastly, we
found that deep soil pH in the grazing lawns of the
different land-use histories was generally a full pH
unit higher than in the close controls (z = 8.03,
P < 0.001) (Table 2; Figure 3H), whereas the top
soil pH of the lawns was similar to close sites
(z = 1.64, P = 0.56). However, top and deep soils
pH did not differ for close (z = 0.79, P = 0.97) or far
away control sites (z = 1.85, P = 0.42) (Table 2;
Figure 3H).
The Influence of Soil Texture on GrazingLawn Soil Properties
Grazing lawns covered a wide range of soil textures
that were not associated with specific land-use
histories, but rather with the two distinct geological
areas in Ithala with coarse granitic soils in the East
and finer soils of volcanic origin in the West
(F1,28 = 73.7, P < 0.0001; Figure 2A). Soil texture
did not differ between grazing lawns and either
controls (F2,62 = 1.15, P = 0.32), nor did it differ
according to land use (Figure 2B). However, most
soil nutrient concentrations varied with soil tex-
ture. Clayey soils had higher concentrations of K,
Ca, Mg, total N, and C than sandy soils for lawns as
well as control sites (Tables 2, 3; Figure 4A–F). In
contrast, P, C:N, and pH were not influenced by soil
texture (Tables 2, 3; Figure 4D, G, H).
DISCUSSION
Former land use was the key factor explaining
grazing lawn location in Ithala. More than 40 years
after the reserve was established, three quarters of
grazing lawns were found on locations that were
substantially altered by former livestock husbandry
(bomas) or agriculture (old fields) (Table 1). Fur-
thermore, soil nutrient content in the grazing lawns
was influenced by both former land use and soil
texture. Specifically, P concentrations in the grazing
lawns found in abandoned bomas were three times
higher than in lawns found in old fields or natural
grasslands. In addition, N and C concentrations were
twice as high in boma lawns compared to the lawns
in the other locations (Figure 3). On clayey soils,
grazing lawns were hotspots for all measured soil
nutrients, whereas on sandy soils they only differed
from controls for P, C:N, and pH (Figure 4). By
collecting samples within and away from grazing
lawns, we found that soils from boma lawns con-
trasted more strongly with close controls than old
fields and natural grassland lawn soils, particularly
for N, P, and C.
Legacy of Land Use on Grazing Lawn Soil Properties
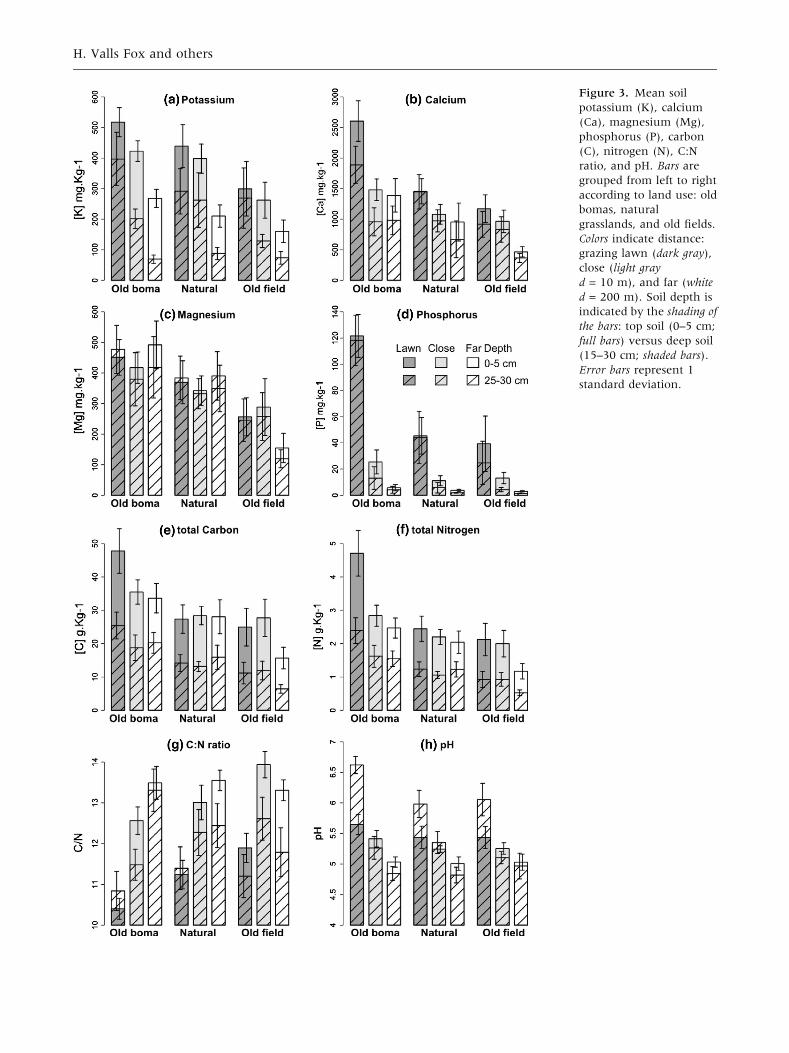
Figure 3. Mean soil
potassium (K), calcium
(Ca), magnesium (Mg),
phosphorus (P), carbon
(C), nitrogen (N), C:N
ratio, and pH. Bars are
grouped from left to right
according to land use: old
bomas, natural
grasslands, and old fields.
Colors indicate distance:
grazing lawn (dark gray),
close (light gray
d = 10 m), and far (white
d = 200 m). Soil depth is
indicated by the shading of
the bars: top soil (0–5 cm;
full bars) versus deep soil
(15–30 cm; shaded bars).
Error bars represent 1
standard deviation.
H. Valls Fox and others
Legacy Effects Differ Between Types ofFormer Human Activities
The sharp boundary that separates abandoned bo-
ma grazing lawns from control sites is consistent
with previous work from South Africa (Blackmore
and others 1990) and East Africa (Young and oth-
ers 1995; Augustine 2003; Cech and others 2010).
Even after 40 years of abandonment, the boma
sites in our study area were characterized by higher
soil organic content associated with a unique, thick
top soil horizon created by the accumulation of
manure. In contrast, grazing lawns found in nat-
ural grasslands and old fields exhibited remarkably
lower P, Ca, total N, and C than lawns found in
former bomas. All lawns had similarly elevated le-
vels of pH compared with control soils. These re-
sults thus suggest that the type of land use
influences grazing lawn characteristics.
Several studies point out that nutrient concen-
trations in bomas gradually decrease due to a ne-
gative nutrient balance (Young and others 1995;
Augustine 2003; Muchiru and others 2009). Ac-
cordingly, we cannot rule out that grazing lawns
found in old fields or natural grasslands do not
originate from even older bomas pre-dating our
1948 aerial pictures. However, as explained in the
methods, we avoided selecting old boma sites in, or
close to, old fields (for example, avoiding sites with
remains of stone walls or old grinding stones). The
fact that the practice in Ithala had been to encircle
bomas with stone walls, and not thorny bushes as
elsewhere in Africa (Young and others 1995) made
it possible for us to avoid even very old boma sites.
Thus, we reduced the possibility of an overlap be-
tween old field lawns and very old abandoned
bomas. In addition, because the fields we selected
had been cultivated for at least 2–3 decades, we
believe that even if some fields would have been
very old boma sites, legacy effects of these bomas
would have been strongly diluted through decades
of quite intense cultivation (including the creation
of terraces and annual plowing).
One could also argue that bomas and old fields
were originally restricted to different parts of the
landscape. For example, if bomas had been located
in good grazing areas for cattle, which might still be
very attractive for wild grazers, this may explain
the difference in legacy effect with old fields.
However, we did not find a correlation between
land-use history and soil texture (Figure 2), nor
with the other soil properties we measured (see far
controls Figure 3). Moreover, bomas in Ithala were
not necessarily directly linked to the best grazing
areas but instead found in the vicinity of home-
steads. These homesteads were on slopes and crests,
possibly to avoid risk of diseases prevalent in the
valleys where the good grazing lands are situated
(Feely 1980). Livestock were then herded to good
grazing areas during the day and brought back to
the boma during the night. Hence, we do not think
differences in the positioning of boma and old field
sites explain their difference in legacy effects.
Soil nutrient and carbon hotspots may contain a
substantial proportion of the total stocks of a given
ecosystem (Ostle and others 2009). This might be
the case for former bomas and grazing lawns de-
rived from them. Unfortunately, we lacked soil
bulk density measures to come to reliable estimates
of ecosystem-scale contributions of lawns to nu-
trient and carbon stocks. However, doing a rough
calculation, grazing lawns cover less than 0.2% of
Ithala (Table 1) and P concentrations in lawns were
on average eight times higher than controls. Based
on these estimates, the legacy of human land use
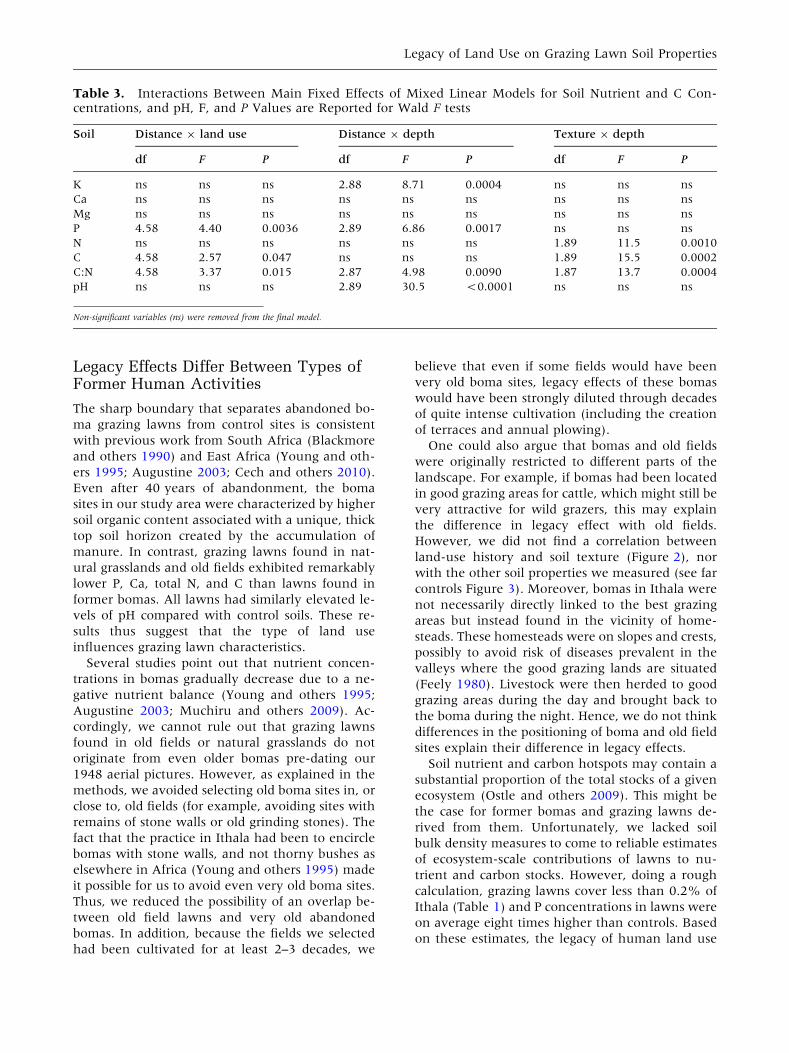
Table 3. Interactions Between Main Fixed Effects of Mixed Linear Models for Soil Nutrient and C Con-centrations, and pH, F, and P Values are Reported for Wald F tests
Soil Distance 9 land use Distance 9 depth Texture 9 depth
df F P df F P df F P
K ns ns ns 2.88 8.71 0.0004 ns ns ns
Ca ns ns ns ns ns ns ns ns ns
Mg ns ns ns ns ns ns ns ns ns
P 4.58 4.40 0.0036 2.89 6.86 0.0017 ns ns ns
N ns ns ns ns ns ns 1.89 11.5 0.0010
C 4.58 2.57 0.047 ns ns ns 1.89 15.5 0.0002
C:N 4.58 3.37 0.015 2.87 4.98 0.0090 1.87 13.7 0.0004
pH ns ns ns 2.89 30.5 <0.0001 ns ns ns
Non-significant variables (ns) were removed from the final model.
Legacy of Land Use on Grazing Lawn Soil Properties
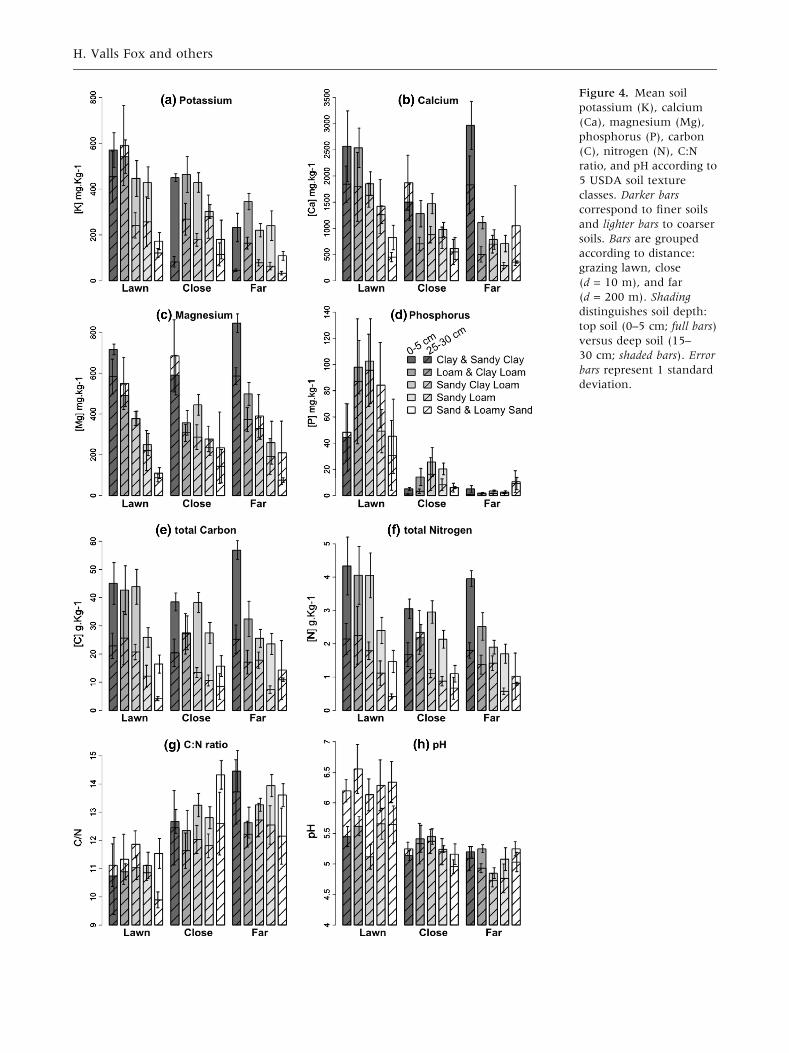
Figure 4. Mean soil
potassium (K), calcium
(Ca), magnesium (Mg),
phosphorus (P), carbon
(C), nitrogen (N), C:N
ratio, and pH according to
5 USDA soil texture
classes. Darker bars
correspond to finer soils
and lighter bars to coarser
soils. Bars are grouped
according to distance:
grazing lawn, close
(d = 10 m), and far
(d = 200 m). Shading
distinguishes soil depth:
top soil (0–5 cm; full bars)
versus deep soil (15–
30 cm; shaded bars). Error
bars represent 1 standard
deviation.
H. Valls Fox and others

accounts at most for 2% of the total nutrient and
carbon stocks in Ithala. Total estimates of soil nu-
trient stocks in all horizons are needed to assess the
human legacy by comparing former bomas and
grazing lawns to other nutrient hotspots such as
dung middens and termite mounds. Although the
contribution to total stocks may be small, lawns
likely are crucial nutrient hotspots for mammalian
grazers because they present concentrations of key
limiting nutrients, such as P, above the nutritional
requirement thresholds, in contrast to surrounding
bunch grassland (McNaughton and others 1988).
Grazing lawns can also be attractive for wild her-
bivores because these open habitats reduce preda-
tion risk (Ford and others 2014). Indeed, previous
studies demonstrated that lawns of a scale similar to
the ones in our study were preferred by a wide
range of grazer species (Cromsigt and Olff 2006).
Grazing Lawns as Hotspots: A Matter ofScale
We found that soil P, C, N, Ca, and pH declined and
C:N increased within a few meters of former boma
grazing lawns (Figure 3). Similar edge effects have
also been reported by other studies (Blackmore and
others 1990; Porensky 2011; Porensky and others
2013). Potassium was the only nutrient with si-
milar concentrations for lawn and close control
soils, although concentrations were still higher
than in the far control (also see Muchiru and others
2008; Veblen and Young 2010). In contrast, in old
field and natural grassland grazing lawns, only P
and Ca concentrations were clearly elevated in
comparison with control soil, although lawns did
not stand out for other nutrients. These results
indicate that abandoned bomas create more con-
trasting nutrient hotspots than the other land-use
types. It also shows that, although the spatial ex-
tent of old field impact is much larger than that of
bomas, the legacy of bomas is in fact stronger,
probably because the intensity of land use in bomas
is stronger and more concentrated than in old
fields. In contrast to our findings, recent studies in
Hluhluwe-iMfolozi Park, South Africa (Stock and
others 2010), Kruger National Park, South Africa,
and Serengeti National Park, Tanzania, (Arnold and
others 2014) found that soil nutrients did not differ
between lawn and bunch grass plots despite dif-
ferences in foliar nutrient concentrations. Howev-
er, it remains unclear whether the lawns described
in these studies are associated with former human
land use or originate from localized grazing by
native wild grazers (Waldram and others 2008).
Furthermore, in their study, Stock and others
(2010) sampled soil from under lawn and bunch
grass patches co-occurring within the same
40 9 40 m site. Hence, their controls could have
been situated within a lawn site. In contrast, both
our controls were clearly away from the lawns out
in the caespitose grassland. In addition, we in-
cluded a far control site at least 200 m away from
any mapped lawn to confirm our findings. In the
case of Arnold and others (2014), it seems they did
not necessarily compare grazing lawns with control
sites composed by a clearly distinctive plant com-
munity. At a fine scale, Arnold and others (2014)
selected grazing lawns based on grass height, not
grass species, and grass species composition did not
significantly differ between their grazing lawns and
control sites. As a result, they might have compared
intensely grazed with less intensely grazed areas
within lawn sites. In contrast, by selecting grazing
lawns dominated by the lawn grass Cynodon dactylon
and control grassland sites with clearly different
grass species compositions (that is, tall, caespitose
species), we compared clearly different grassland
communities and not grazing lawns in different
states of intensity of grazing.
Our results are consistent with the existence of
perennial, compositional, or stoloniferous, grazing
lawns (see Cromsigt and Olff 2008) that are main-
tained by grazers and develop a distinct enriched
soil and grazing-tolerant community (Blackmore
and others 1990; Young and others 1995; Augustine
2003; Muchiru and others 2008; Veblen and Young
2010). Browsers may also contribute to lawn
maintenance by depositing nutrient-rich feces,
while using the lawns as resting sites with relatively
low predation risk (Augustine 2004; Ford and oth-
ers 2014). These perennial lawns differ from grazed
patches that are temporarily grazed short (structural
or caespitose ‘lawns,’ see Cromsigt and Olff 2008) that
may occur when a disturbance such as fire, mow-
ing, trampling, or localized intense grazing keeps
the tall caespitose grass short (Archibald 2008;
Cromsigt and Olff 2008). Younger grass phenology
may attract grazers to these temporary ‘lawns’ by
providing high-quality forage. However, at this time
scale, soil properties and species composition would
not have changed and this may provide an alter-
native explanation to the apparently conflicting
results found by different studies.
Soil Texture: A Modifier of Land-Use andGrazing Effects
In Ithala, soil texture does not appear to govern
grazing lawn location (Figure 2). This confirms
previous findings that grazing lawns can be found
Legacy of Land Use on Grazing Lawn Soil Properties

on a range of soil textures ranging from clayey to
sandy soils whether they originate from bomas
(Young and others 1995; Augustine 2003; Cech
and others 2010; van der Waal and others 2011;
Veblen 2012) or from free-ranging herbivore
grazing (Cromsigt and Beest 2014). This suggests
that in our study soil texture varied at a larger
spatial scale than patterns remaining from past land
use or emerging from current grazing intensity.
This is further confirmed by the fact that soil tex-
ture did not vary among the land-use types in our
study (Figure 2). In Ithala, bomas were built and
fields were plowed on very diverse soil types where
grazing lawns were established locally by herbi-
vores. However, whereas the human legacy set the
template for today’s landscape heterogeneity, this
study does not undermine the importance of
edaphic factors such as soil texture as shown by
many other studies (Anderson and others 2007;
Veldhuis and others 2014). In our study, soil tex-
ture influenced which soil nutrients were elevated
in lawn soils (Figure 4). Whereas most of the soil
nutrient concentrations were higher in lawn soils
on clay, on sandy soils only P, C:N ratio, and pH
differed between lawns and controls. This may
indicate the importance of P:C:N stoichiometry and
pH in processes that govern grazing lawn persis-
tence.
N:P Stoichiometry, Potential Role of SoilBiota, and Grazing Lawn Persistence
N and P are major nutrients that shape many
grazed ecosystems (Cech and others 2010).
Whereas P may persist for decades (Augustine
2003) or even centuries (Blackmore and others
1990), N is much more volatile and its availability is
driven by short-term flows (Scholes and Walker
1993; de Mazancourt and others 1998; Coetsee and
others 2010). High concentrations of P may be
necessary for grazing optimization (Chapin and
McNaughton 1989) and P hotspots may be vital for
herbivores in dystrophic contexts (Augustine 2003,
2004; Verweij and others 2006; Treydte and others
2011). We found that P was the only nutrient that
was clearly elevated in grazing lawns for all land-
use histories and across a wide range of soil tex-
tures. This may indicate that P concentration is a
key process that governs grazing lawn persistence
(Cech and others 2010). The persistence of nutri-
ents in lawns is likely further mediated by soil biota
and the role of such biota perhaps varies among the
different land uses. Increased N inputs and higher
pH negatively impacts fungi (de Vries and others
2006), and lower C:N and higher pH in former
boma lawn soils thus likely resulted in a high mi-
crobial versus fungal biomass ratio (de Vries and
others 2006). Such higher microbial biomass may
in turn enhance nutrient cycling rates and facilitate
plant nutrient uptake (Bardgett and Wardle 2003).
Previous studies have stressed the importance of
such microbe-induced high rates of N cycling in
grazing lawns (McNaughton and others 1997;
Frank and others 2000) even in the absence of N
stocks (Coetsee and others 2010). Future studies
exploring soil biota composition across different
former land uses may reveal to what extent dif-
ferent microorganisms may alter the legacy of past
human land-use practices and drive grazing lawn
persistence.
CONCLUSION
In many protected areas, former human activity
has created nutrient hotspots across the landscape
that influences the grazing patterns of wild herbi-
vores. Grazing lawns illustrate this legacy due to
the strong interactions between herbivores, plants,
and the underlying soil. The legacy of bomas dif-
fered substantially from other land uses due the
persistence of large stocks of P, N, and C within
restricted areas. P limitation in surrounding grass-
lands may favor the maintenance of these grazing
lawns over decades or even centuries, whereas soil
organic matter may alter the role of soil biota in
nutrient cycling. Conversely, grazing lawns in old
field and natural grasslands were also soil nutrient
hotspots but did not have the typical higher soil
organic matter content found in boma top soil and
had less pronounced P stocks. Ultimately, our study
revealed how the legacy of different land-use his-
tories modifies the balance between soil properties
and grazers by shaping C:N:P stoichiometry in
grazing lawns. As such, we confirm that human
activities have a long-lasting legacy that shapes
landscapes and ecosystems over decades and
probably longer, including within so called pristine
protected areas.
ACKNOWLEDGMENTS
We would like to thank KZN wildlife for giving us
the opportunity to conduct our research in Ithala
Game reserve. We also thank our colleagues from
UKZN particularly Vincent Chaplot (IRD) for advice
on soil sample collection and analysis. We are
grateful to Jacques Gignoux and Simon Chamaille-
Jammes for their comments on earlier versions of
the manuscript. Funding for the project was pro-
vided to A.S. (UKZN) by the National Research
H. Valls Fox and others

Foundation (NRF) and to H.F. (LBBE) by the CNRS
INEE ‘‘Zone Atelier’’ grant, and the CNRS GDRI
Biodiversity Dynamics in Southern Africa. H.V.F. was
supported by the Ecole Normale Superieure as an
eleve fonctionnaire stagiaire. J.P.G.M.C. was sup-
ported by a Marie Curie Career Integration Grant
(PCIG10-GA-2011-304128) and by the Swedish
Thematic research program WILDLIFE & FORESTRY.
We thank two anonymous reviewers for their com-
ments which greatly improved a previous version of
this manuscript.
REFERENCES
Allen VG, Batello C, Berretta EJ, Hodgson J, Kothmann M, Li X,
McIvor J, Milne J, Morris C, Peeters A, Sanderson M. 2011.
An international terminology for grazing lands and grazing
animals. Grass Forage Sci 66:2–28.
Anderson TM, Ritchie ME, McNaughton SJ. 2007. Rainfall and
soils modify plant community response to grazing in Serengeti
National Park. Ecology 88:1191–201.
Archibald S. 2008. African grazing lawns—how fire, rainfall, and
grazer numbers interact to affect grass community states. J
Wildl Manag 72:492–501.
Arnold SG, Anderson TM, Holdo RM. 2014. Edaphic, nutritive,
and species assemblage differences between hotspots and
matrix vegetation: two African case studies. Biotropica
46:387–94.
Augustine DJ. 2003. Long-term, livestock-mediated redistribu-
tion of nitrogen and phosphorus in an East African savanna. J
Appl Ecol 40:137–49.
Augustine DJ. 2004. Influence of cattle management on habitat
selection by impala on central Kenyan rangeland. J Wildl
Manag 68:916–23.
Bardgett RD, Wardle DA. 2003. Herbivore-mediated linkages
between aboveground and belowground communities. Ecol-
ogy 84:2258–68.
Blackmore AC, Mentis MT, Scholes RJ. 1990. The origin and
extent of nutrient-enriched patches within a nutrient-poor
savanna in South Africa. J Biogeogr 17:463–70.
Bonnet O, Fritz H, Gignoux J, Meuret M. 2010. Challenges of
foraging on a high-quality but unpredictable food source: the
dynamics of grass production and consumption in savanna
grazing lawns. J Ecol 98:908–16.
Cech PG, Venterink HO, Edwards PJ. 2010. N and P cycling in
Tanzanian Humid Savanna: influence of herbivores, fire, and
N2-fixation. Ecosystems 13:1079–96.
Chapin FS, McNaughton SJ. 1989. Lack of compensatory growth
under phosphorus deficiency in grazing-adapted grasses from
the Serengeti Plains. Oecologia 79:551–7.
Cobo JG, Dercon G, Cadisch G. 2010. Nutrient balances in
African land use systems across different spatial scales: a re-
view of approaches, challenges and progress. Agric Ecosyst
Environ 136:1–15.
Coetsee C, Stock WD, Craine JM. 2010. Do grazers alter nitrogen
dynamics on grazing lawns in a South African savannah? Afr J
Ecol 49:62–9.
Coughenour MBB. 1985. Graminoid responses to grazing by
large herbivores: adaptations, exaptations, and interacting
processes. Ann Missouri Bot Gard 72:852–63.
Cromsigt J, Beest M. 2014. Restoration of a megaherbivore:
landscape level impacts of white rhinoceros in Kruger Na-
tional Park, South Africa. J Ecol 102:566–75.
Cromsigt JPGM, Kuijper DPJ. 2011. Revisiting the browsing
lawn concept: evolutionary Interactions or pruning herbi-
vores? Perspect Plant Ecol Evol Syst 13:207–15.
Cromsigt JPGM, Olff H. 2006. Resource partitioning among sa-
vanna grazers mediated by local heterogeneity: an ex-
perimental approach. Ecology 87:1532–41.
Cromsigt JPGM, Olff H. 2008. Dynamics of grazing lawn for-
mation: an experimental test of the role of scale-dependent
processes. Oikos 117:1444–52.
Day PR. 1965. Particle fractionation and particle-size analysis. In:
Black CA, Ed. Methods of soil analysis. Part 2 Agronomy.
Madison: American Society of Agronomy. p 1367–78.
De Mazancourt C, Loreau M, Abbadie L. 1998. Grazing opti-
mization and nutrient cycling: when do herbivores enhance
plant production? Ecology 79:2242–52.
De Vries FT, Hoffland E, van Eekeren N, Brussaard L, Bloem J.
2006. Fungal/bacterial ratios in grasslands with contrasting
nitrogen management. Soil Biol Biochem 38:2092–103.
De Vries FT, Bloem J, Quirk H, Stevens CJ, Bol R, Bardgett RD.
2012. Extensive management promotes plant and microbial
nitrogen retention in temperate grassland. PLoS ONE
7:e51201.
Dewitte O, Jones A, Spaargaren O, Breuning-Madsen H, Bros-
sard M, Dampha A, Deckers J, Gallali T, Hallett S, Jones R,
Kilasara M, Le Roux P, Micheli E, Montanarella L, Thiom-
biano L, Van Ranst E, Yemefack M, Zougmore R. 2013. Har-
monisation of the soil map of Africa at the continental scale.
Geoderma 211–212:138–53.
Donihue C, Porensky L, Foufopoulos J. 2013. Glade cascades:
indirect legacy effects of pastoralism enhance the abundance
and spatial structuring of arboreal fauna. Ecology 94:827–37.
Ellis EC, Ramankutty N. 2008. Putting people in the map: an-
thropogenic biomes of the world. Front Ecol Environ 6:439–
47.
Feely J. 1980. Did iron age man have a role in the history of
Zululand’s wilderness landscapes? S Afr J Sci 76:150–2.
Ford AT, Goheen JR, Otieno TO, Bidner L, Isbell LA, Palmer TM,
Ward D, Woodroffe R, Pringle RM. 2014. Large carnivores
make savanna tree communities less thorny. Science
346:346–9.
Frank D, Groffman P, Evans R. 2000. Ungulate stimulation of
nitrogen cycling and retention in Yellowstone Park grasslands.
Oecologia 123:116–21.
Hothorn T, Bretz F, Westfall P. 2008. Simultaneous inference in
general parametric models. Biom J 50:346–53.
Huisman J, Olff H. 1998. Competition and facilitation in multi-
species plant-herbivore systems of productive environments.
Ecol Lett 1:25–9.
Hunter AH. 1975. New techniques and equipment for routine
soil-plant analytical procedures. In: Borremiza A, Alvarado A,
Eds. Soil management in tropical America. Raleigh: N. C. State
University.
Lewu FB, Assefa Y. 2009. Farmers’ knowledge in the cropping
systems of Northern KwaZulu-Natal, South Africa: current
challenges and solution for sustainable future food produc-
tion. Afr J Agric Res 4:1148–53.
Manson AD, Roberts VG. 2011. Analytical methods used by the
soil fertility and analytical services section. KwaZulu-Natal
Department of Agriculture and Environmental Affairs. pp. 6.
Legacy of Land Use on Grazing Lawn Soil Properties

Matejovic I. 1997. Determination of carbon and nitrogen in
samples of various soils by the dry combustion. Commun Soil
Sci Plant Anal 28:1499–511.
McLauchlan K. 2007. The nature and longevity of agricultural
impacts on soil carbon and nutrients: a review. Ecosystems
9:1364–82.
McNaughton SJ. 1979. Grazing as an optimization process: grass-
ungulate relationships in the Serengeti. Am Nat 113:691–703.
McNaughton SJ. 1984. Grazing lawns: animals in herds, plant
form, and coevolution. Am Nat 124:863–86.
McNaughton SJ, Ruess RW, Seagle SW. 1988. Large mammals
and process dynamics in African ecosystems. Bioscience
38:794–800.
McNaughton SJ, Banyikwa FF, McNaughton MM. 1997. Pro-
motion of the cycling of diet-enhancing nutrients by African
Grazers. Science 278:1798–800.
Moeys J, Shangguan W. 2014. Soiltexture: functions for soil tex-
ture plot, classification and transformation. R package version
1.2.19. http://CRAN.R-project.org/package=soiltexture.
Muchiru AN, Western DJ, Reid RS. 2008. The role of abandoned
pastoral settlements in the dynamics of African large herbi-
vore communities. J Arid Environ 72:940–52. doi:10.1016/j.
jaridenv.2007.11.012.
Muchiru AN, Western D, Reid RS. 2009. The impact of aban-
doned pastoral settlements on plant and nutrient succession in
an African savanna ecosystem. J Arid Environ 73:322–31.
Novellie P, Gaylard A. 2013. Long-term stability of grazing lawns
in a small protected area, the Mountain Zebra National Park.
Koedoe 55(1):1–7. doi:10.4102/koedoe.v55i1.1108.
Ostle NJ, Levy PE, Evans CD, Smith P. 2009. UK land use and
soil carbon sequestration. Land Use Policy 26:S274–83.
Pinheiro J, Bates D, DebRoy S, Sarkar D, R Development Core
Team. 2011. nlme: linear and nonlinear mixed effects models.
R package version 3.1-104. Vienna: R Foundation for Statis-
tical Computing.
Porensky LMG. 2011. When edges meet: interacting edge effects
in an African savanna. J Ecol 99:926–34.
Porensky LM, Veblen KE. 2012. Grasses and browsers reinforce
landscape heterogeneity by excluding trees from ecosystem
hotspots. Oecologia 168:749–59.
Porensky LM, Bucher SF, Veblen KE, Treydte AC, Young TP.
2013. Megaherbivores and cattle alter edge effects around
ecosystem hotspots in an African savanna. J Arid Environ
96:55–63.
Prins HHT. 2000. Competition between wildlife and livestock in
Africa. In: Prins HHT, Grootenhuis JG, Dolan TT, Eds. Wildlife
conservation by sustainable use. Boston: Kluwer. p 52–80.
Roberts VG, Adey S, Manson AD. 2003. An investigation into
soil fertility in two resource-poor farming communities in
KwaZulu-Natal (South Africa). S Afr J Plant Soil 20:146–51.
Ruess RW, McNaughton SJ. 1987. Grazing and the dynamics of
nutrient and energy regulated microbial processes in the
Serengeti grasslands. Oikos 49:101–10.
Sankaran M, Augustine DJ. 2004. Large herbivores suppress
decomposer abundance in a semiarid grazing ecosystem.
Ecology 85:1052–61.
Scholes RJ, Walker BH. 1993. An African savanna: synthesis of
the Nylsvley study. Cambridge: Cambridge University Press.
Stock WD, Bond WJ, van de Vijver C. 2010. Herbivore and
nutrient control of lawn and bunch grass distributions in a
southern African savanna. Plant Ecol 206:15–27.
Treydte AC, van der Beek JGM, Perdok AA, van Wieren SE.
2011. Grazing ungulates select for grasses growing beneath
trees in African savannas. Mamm Biol 76:345–50.
Van der Waal C, Kool A, Meijer SS, Kohi E, Heitkonig IM, de
Boer WF, van Langevelde F, Grant RC, Peel MJS, Slotow R, de
Knegt HJ, Prins HHT, de Kroon H. 2011. Large herbivores may
alter vegetation structure of semi-arid savannas through soil
nutrient mediation. Oecologia 165:1095–107.
Van Rooyen N, van Rooyen MW. 2008. Vegetation of Ithala
Game Reserve. Pietermaritzburg: Ezemvelo KZNWildlife.
Veblen KE. 2012. Savanna glade hotspots: plant community
development and synergy with large herbivores. J Arid En-
viron 78:119–27. doi:10.1016/j.jaridenv.2011.10.016.
Veblen KE, Young TP. 2010. Contrasting effects of cattle and
wildlife on the vegetation development of a savanna land-
scape mosaic. J Ecol 98:993–1001.
Veldhuis MP, Howison RA, Fokkema RW, Tielens E, Olff H.
2014. A novel mechanism for grazing lawn formation: large
herbivore-induced modification of the plant-soil water bal-
ance. J Ecol 102:1506–17.
Verweij RJT, Verrelst J, Loth PE, Heitkonig IMA, Brunsting
AMH. 2006. Grazing lawns contribute to the subsistence of
mesoherbivores on dystrophic savannas. Oikos 114:108–16.
Waldram MS, Bond WJ, Stock WD. 2008. Ecological engineering
by a mega-grazer: white rhino impacts on a South African
savanna. Ecosystems 11:101–12.
Young TP, Patridge N, Macrae A. 1995. Long-term glades in
acacia bushland and their edge effects in Laikipia, Kenya. Ecol
Appl 5:97–108.
Zuur AF, Ieno EN, Walker NJ, Saveliev AA, Smith GM. 2009. 5
mixed effects modelling for nested data. In: Smith GM, Ed.
Mixed effects models and extensions in ecology with R. Ber-
lin: Springer. p 101–42.
H. Valls Fox and others